







Bahasa
Halaman
Hukum


Hy drobiolo gia lz 8;' t+g-tl o, zooz.P.H. Nienhuis & R.D. Gulati (eds), Ecological Restoration of Aquatic andSemïAquatic Ecosystems in the Netherlands (NW Europe).
@ 20O2 Kluwer Academic Publishers. Printed in the Netherlands.
149
Restoration of brook valley meadows in the Netherlands
A.P. Grootjansl, J.P. Bakkerl, A.J.M. Jansen2 & R.H. Kemmers3rlnboratory of Plant Ecology, [Jniversity of Groningen, P.O. Box ]4, 9750 AA Haren, The NetherlandsE-mail : A. P. Gro otj ans @ biol. ru g.nl2wlrto, Water Company Overijsel, P.O. Box 10.005, 8000 GA Zwolle, The Netherlands3Alterra, P.O. Box 25, 6700 AC, Wageningen, The Netherlands
Key words: fen meadows, hydrology, Red List species, restoration management, rewetting, sod cutting
Abstract
Until recently, restoration measures in Dutch brook valley meadows consisted of re-introducing traditional man-
agement techniques, such as mowing without fertilisation and low-intensity grazing. In the Netherlands, addit-ional
measures, such as rewetting and sod cutting, are now carried out on a large scale to combat negative influences
of drainage and acidifying influences by atmospheric deposition. An analysis of successful and unsuccessful pro-jects shows that restoration of brook valley meadows is most successful if traditional management techniques are
applied in recently abandoned flelds that had not been drained or fertilised. Large-scale topsoil removal in formeragricultural fields that had been used intensively for several decades is often unsuccessful since seed banks are
depleted, while hydrological conditions and seed dispersal mechanisms are sub-optimal. In areas with an organic
topsoil, long-term drainage had often led to irreversible changes in chemical and physical properties of the soil.
Successful sites were all characterised by a regular discharge of calcareous groundwater provided by local orregional hydrological systems, and, where not very long ago, populations of target species existed. On mineralsoils, in particular, sod removal in established nature reserves was a successful measure to increase the number ofendangered fen meadow species. It is argued that attempts to restore species-rich meadows should be avoided on
former agricultural fields, where pedological processes have led to almost irreversible changes in the soil proflleand where soil seed banks have been completely depleted. From a soil conservation point of view, such areas
should be exploited as eutrophic wetlands that are regularly flooded.
Introduction
Fen meadows or litter meadows are nutrient-poorgrasslands which can be derived from groundwaterfed mires after modest drainage (Ellenberg, 1978),but can also occur on groundwater fed mineral soils.Such meadows may produce 1-4 tons dry weight ha-lyr-l lKlapp, 1965). Hay meadows, usually derivedfrom eutrophic marshes in flood plains, may produce4-8 tons ha-1 yr-1. The most species-rich fen mead-ows are classified as Cirsio-Molinietum and harbour a
large number of species that have been listed on the'Red List' of highly endangered species. Examplesare Cirsium dissectum, Carex dioica, C. pulicaris, C.
hostiana, Viola persicifulia, Parnassia palustris andorchid species, such as Gymnadenia conopsea, Dac-tylorhiza incarnata. Hay meadows are usually less
species-rich and have less Red List species. Target
species here are: Caltha palustris, Crepis paludosa,Dactylorhiza majalis, Juncus acutiflorus, Lychnis floscuculi, Carex aquatilis, Pedicularis palustris (see
Appendix 1). In Western Europe, such species-richmeadows have become increasingly rare as a con-sequence of changes in agricultural practices. Es-timates from the United Kingdom are that 95-987oof species-rich hay meadows that were present be-fore 1940, have been lost due to intensiflcation ofagricultural exploitation or due to abandonment and
subsequent development of forest (Garcia, 1992). Inthe Netherlands, a similar process has occurred. Mostspecies-rich meadows that exist today are managedby nature conservation organisations or have been re-stored after intensive agricultural use (Bakker & Oltr,1995). The establishment of nature reseryes, however,

150
does not necessarily mean that endangered species are
protected. Even in existing nature reseryes many spe-
cies are still threatened by extinction. Runhaar et al.(1999) estimated, for instance, that at least 50% ofall groundwater dependent ecosystems in the Nether-lands have been moderately to strongly affected byhydrological changes. Funds have become availablesince the 1990, in particular, to counteract the effectsof atmospheric N-deposition and desiccation in naturereserves (Runhaar, 1999). Not all restoration projects,
however, were successful, and little information is
available to identify the abiotic or biotic causes forthese failures. In order to learn from the mistakes thathave been made we have selected 12 projects that hadbeen monitored in a systematic way for at least 8 years.
In the present paper, we will evaluate the successes
and failures of these restoration projects and use thenumber of re-established Red List species and char-
acteristic species of target communities to judge theirsuccess.
The actual analysis is preceded by a short re-
view on the eco-hydrological functioning of brookvalleys, the human impact on nutrient cycling, seed
dispersal and technical measures to restore deterior-ated meadows. After the assessment of successful andless successful projects, we will present a conceptualmodel for the restoration of wet meadows.
Structure and hydrological functioning of brookvalleys
Vegetation differentiation in brook valleys reflectsgradients in soil fertility and hydrology, which in turnreflect properties of the landscape system surroundingthe valley. If the impact of man (drainage, fertilisa-tion) on these landscape properties has not been verystrong, vegetation differentiation within semi-naturalcommunities is an expression of the trophic gradients
in the soil. Such trophic gradients are very stable as
long as regional and local hydrological systems are
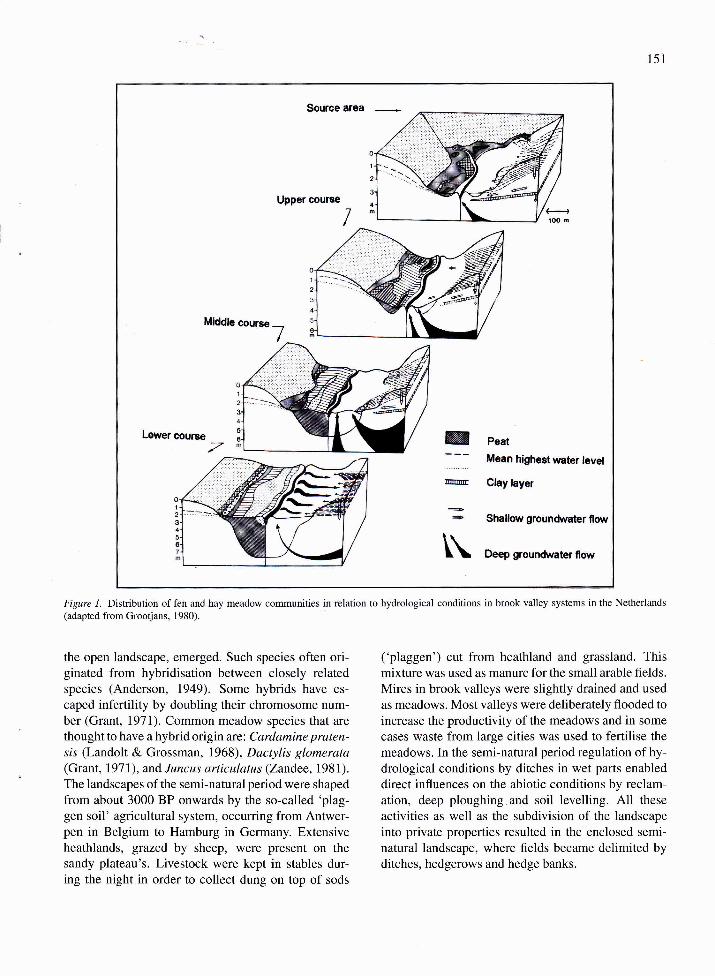
not disturbed. Grootjans (1980) described the spa-
tial variation in wet plant communities along heightgradients in Dutch brook valleys. Along the valleysslopes nutrient-poor heathland communities (Ericiontetralicis) were present (Fig. 1). In some areas wheresemi-pervious clay layers were present in the subsoil,wet heathlands and bogs covered extensive areas onthe higher plateaus. Shallow groundwater flows ori-ginating from these heathlands are generally poor indissolved minerals since they passed through decalci-
fied cover sands. Groundwater originating from deeper
aquifers has passed through calcaÍeous deposits and,
therefore, is rich in calcium and hydrocarbonate. Up-stream areas influenced by calcareous groundwaterusually have a better soil fertility than the acid heath-
lands and support more productive meadow types(Junco-Molinion and Calthion palustris). The most
productive meadow communities occur in downstream
areas, which are regularly flooded with stream watercontaining silt and clay particles, which increase soilfertility significantly. Reed marshes (Phragmition) and
flood meadows (Calthion palustris and Magnocari-cion) are most common here.
Human impact
During the development of the cuÍïent landscape ofNW-Europe, we may discern three periods: the nat-
ural period, the semi-natural period and the culturalperiod (Bakker & Londo, 1998). The natural periodis characterised by the dominance of communities,landscapes and processes without any noticeable hu-
man impact. The major patterns in the landscape were
largely determined by geological and hydrologicalfactors. Grazing and browsing took place by indigen-ous herbivores. The mires in the brook valleys were
lormed in eroded melt water valleys during wet peri-ods about 6000-3000 BP (Fig. 2). Most peat depositsare older than the existing bed of the small rivers,
Rivers may have shifted position several times. Large
aeolian dunes sometimes blocked the course of a river,
which then forced its way through sandy ridges to f,ndits way to another mire. These ancient mires consisted
mostly of sedge peat underlain by remnants of formerA/nus woods.
The first agricultural invasion took place about7000 BP followed by a second one around 4600 BP.
These people grew arable crops in a shifting cultiv-ation system after the clearance of primeval forest.For the greater part, livestock gradually replaced in-digenous large herbivores. In medieval times, degrad-ation and destruction of primeval forests continuedand large oligotrophic bogs. mesotrophic fens and
eutrophic reed swamps were drained, reclaimed and
in some places the peat was completely removed forfuel (Louwe Kooijmans, 1974). AlÍhough the openlandscape was new, the majority of the species that in-vaded grasslands and heathlands were already present
in open parts of forests, in fringes along streams,
or in fens and bogs. Also new species, adapted to
151
§ource arsa
Upper course
Lower couree7 Y
ffi
Peat
Mean higheet water lavel
Chy layer
Shallow grounórater lor
N Deepgroundrrateríow
Figure 1. Distribution of fen and hay meadow communities in relation to hydrological conditions in brook valley systems in the Netherlands(adapted from Grootjans, 1980).
the open landscape, emerged. Such species often on-ginated from hybridisation between closely relatedspecies (Anderson, 1949). Some hybrids have es-
caped infertility by doubling their chromosome num-ber (Grant, 1971). Common meadow species that arethought to have a hybrid origin are: Cardamine praten-sis (Landolt & Grossman, 1968), Dactylis glomerata(Grant, 197 I), and Juncus articulatus (Zandee, I 98 1 ).The landscapes of the semi-natural period were shapedfrom about 3000 BP onwards by the so-called 'plag-gen soil' agricultural system, occuÍïing from Antwer-pen in Belgium to Hamburg in Germany. Extensiveheathlands, grazed by sheep, were present on thesandy plateau's. Livestock were kept in stables dur-ing the night in order to collect dung on top of sods
('plaggen') cut from heathland and grassland. Thismixture was used as manure for the small arable flelds.Mires in brook valleys were slightly drained and used
as meadows. Most valleys were deliberately flooded toincrease the productivity of the meadows and in some
cases waste from large cities was used to fertilise the
meadows. In the semi-natural period regulation of hy-drological conditions by ditches in wet parts enableddirect influences on the abiotic conditions by reclam-ation, deep ploughing and soil levelling. All these
activities as well as the subdivision of the landscapeinto private properties resulted in the enclosed semi-natural landscape, where flelds became delimited byditches, hedgerows and hedge banks.

152
ffi=mEa3
Fens
Clay layer
Drainage system
Groundwaler ílow line
Strongly dec,omposad pèat
Eutrophic mires and hay meadows
Figure 2. Changes in time in a brook valley landscape due to changed agricultural practices. First, natural mires, such as bogs and fens werechanged into semi-natural fen meadows. This was followed by ecological degradation due to intensive drainage and fertilisation (adapted fromGrootjans & van Diggelen, 1995).
The transition from the semi-natural to the presentcultural period in NW-Europe was caused by the intro-duction of artiflcial fertiliser. The large-scale reclama-tion and subsequent fertilisation of common grasslandand heathland occurred after 1920 when mechanisa-tion in agriculture started. It resulted in the develop-ment of the cultural landscape, in which not only thevegetation but also the flora and fauna became heavilyinfluenced by man. Indigenous species were eradic-ated by herbicides and non-indigenous species wereintroduced. These landscapes represent the culturalperiod.
These changes in land use had strong repercussionsfor brook valley systems. The large-scale reclamationof heathlands also led to many short cuts in the hydro-logical cycle, which increased flooding of meadowsin the middle- and lower courses of the brook valleysafter heavy rainfall. This in turn led to large-scale in-terference with the hydrological conditions in the early1960s. It often resulted in the complete disappear-ance of all natural watercourses and in deep drainageof all peat soils. This resulted in subsidence of thepeat, increased mineralisation, eutrophication of sur-face water, replacement of calcareous groundwater byrainwater, and subsequent acidification of the topsoil(van Wirdum, l99l; Vy'assen et al., 1996). All afore-mentioned processes resulted in a dramatic decreaseof species of the former semi-natural landscape. Many
plant species became endangered and were restrictedto marginal environments in a fragmented landscape.
Nutrient cycling
Pedogenetic processes in brook valleys are mainlycontrolled by hydrological conditions. Both quant-itative and qualitative aspects of the hydrology areconditioning site factors, for instance by controllingdecomposition and humification processes in the or-ganic topsoil. Consequently, the balance between in-put of litter and these processes determines whetherpeat or humus is formed. This balance between litterinput and consequent decomposition or humificationis displayed in the humus form of a site (Klinka et al.,1981), consisting of a sequence of soil horizons. Themorphology of such horizons is often very informa-tive for the type of nutrient cycling that is going on inthe soil (Kemmers et al., 1995). Fibric humus formsconsist of poorly decomposed fibric plant remainsindicating accumulation of nutrients, slow nutrientcycling and poor N-availability. Potassium, however,is easily available from rapid cytolysis. Very wet siteswith fibric humus forms are usually N-limited andrespond to drainage with increased above-ground pro-duction (Grootjans et al., 1985). Humic humus formsconsist of organic matter that is already decomposedand humifled. Such humus forms have a rapid nutrient
153
I
cycling and the availability of nutrients is high. How-ever, the productivity of sites with humic soil horizonsmay gradually decrease due to continued removal ofbiomass by haymaking. Very old fen meadows, forinstance, have developed P-limitation due to long-term removal of phosphorus by mowing (Eglotr, I986;Koerselman et al., 1990; Pegtel et a1.,1996). Even po-tassium may become in short supply (Kapfer, 1988),particularly after deep drainage (de Mars eÍ al., 1996:'
van Duren etal., 1997a,b; van Duren & Pegtel, 1999).Such meadows respond to drainage with a decreased
above-ground biomass (Grootjans et al., 1986). Po-tassium deficiency favours grasses while inhibitingthe development of leguminosae seedlings and otherherbs (Mengel & Kirkby, 1982;Pegtel,1983). If thepotassium level drops below 0.5loK dry weight, fertil-isation with K is necessary to enable establishment oftarget species, while (co-) limitation of P and N stim-ulates the establishment of many fen meadow species(Biewer, 1997).
Mesic humus forms have an intermediate characterand usually occur in wet sites influenced by base-richsurface or groundwater.
During the 'natural' period, typical species of fenmeadows were probably only present along a smallfringe of brook valleys where humic-soil rypes coulddevelop (Fig. 3). During the 'semi-natural' period,most of the natural fens were slightly drained andsites with humic topsoils developed, causing Junco-Molinion species to spread in the new meadows.Simultaneously more productive Calthion meadowsdeveloped from Magnocaricion mires with fibric top-soils. which changed into mesic sites.
At present Junco-Molinion, Parvocaricion andCalthion meadows are associated with humics, mesicsand fibrics, respectively (Kemmers et al., 1995). Fenmeadows (Junc o - M o linion ) developed where humicspersisted from the natural period to present. These
low productive meadows are the most species-rich and
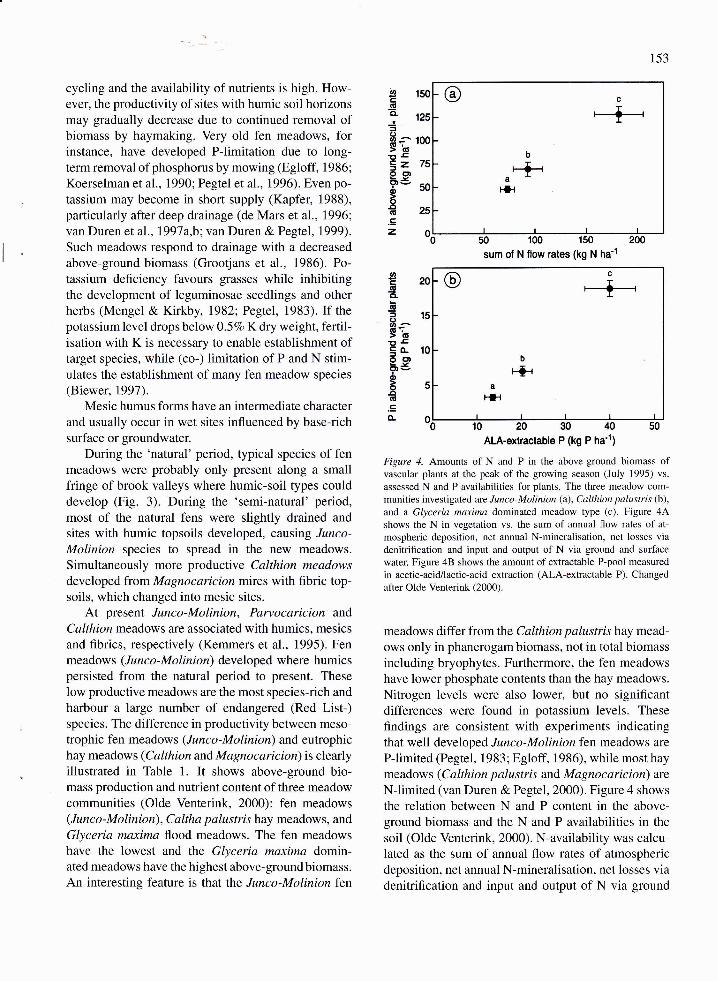
harbour a ).arge number of endangered (Red List-)species. The difference in productivity between meso-trophic fen meadows (Junco-Molinion) and eutrophichay meadows (C althion and M a gno c aric io n) is clearlyillustrated in Thble 1. It shows above-ground bio-mass production and nutrient content of three meadowcommunities (Olde Venterink, 2000): fen meadows(Junco-Molinion), Caltha palustris hay meadows, and
Glyceria maxima flood meadows. The fen meadowshave the lowest and the Glyceria maxima domin-ated meadows have the highest above-ground biomass.An interesting feature is that the Junco-Molinion fert
í) 100 150 200
sum oÍ N ílow rates (kg N ha'1
%' 10 20 30 40 50
ALA-extractable P (kg P ha{)
Figure 4. Amounts of N and P in the above-ground biomass oÍvascular plants at the peak of the growing season (July 1995) vs.
assessed N and P availabilities for plants. The three meadow com-munities investigated are Junco-Molinion (a), Calthion palustris (b),
and a Glyceria maxima dominated meadow type (c). Figure 4Ashows the N in vegetation vs. the sum of annual flow rates of at-
mospheric deposition, net annual N-mineralisation, net losses viadenitrification and input and output of N via ground and surface
water. Figure 48 shows the amount of extractable P-pool measured
in acetic-acid/lactic-acid extraction (AlA-extractable P). Changed
after Olde Venterink (2000).
meadows differ from the Calthion palustris hay mead-ows only in phanerogam biomass, not in total biomassincluding bryophytes. Furthermore, the fen meadowshave lower phosphate contents than the hay meadows.Nitrogen levels were also lower, but no significantdifferences were found in potassium levels. These
flndings are consistent with experiments indicatingthat well developed Junco-Molinion fen meadows are
P-limited (Pegtel, 1983; Egloff, 1986), while most haymeadows (Calthion palustris and Magnocaricion) are
N-limited (van Duren & Pegtel, 2000). Figure 4 shows
the relation between N and P content in the above-ground biomass and the N and P availabilities in the
soil (Olde Venterink, 2000). N-availability was calcu-lated as the sum of annual flow rates of atmosphericdeposition, net annual N-mineralisation, net losses viadenitriflcation and input and output of N via ground
1
t25
50
25
oó
20
6c§CL
l.=o3a>qE5
=zOEDErooo-o(,.Ez
15
(É
È10C"v
oɧCL
-goo6!,c3oP6)oo6.Eo
@ =j-b
r{_,a
raí
@ --{-----
b
FiIra
r-G

154
Í = Or =organiclibÍicm = Om= organic mesic
h - Oh = organic humicA = endorganic mineralL = litter
C - mineralsoil
-Y- watertable
Figure 3. Typical brook valley system with mesoEophic fen meadows on the valley margins influenced by discharge ofcalcareous groundwater,bordered by sedge communities which are flooded by surface water. Pedological changes in the organic topsoil are shown in three transects,indicating the soil transformation a.fter drainage during the semi-natural period and the cultural period.
and surface water (Fig. 4A). Figure 48 shows theextractable P-pool measured in acetic-acid/lactic-acidextraction (AlA-extractable P). A close relationshipexists between N and P availability in the topsoil(10 cm) and N and P content at peak standing crop.Olde Venterink (2000) found that most of the avail-able N and K was stored in above ground biomass(757o and 577o, resp.). Storage of P in the above-groundbiomass was less. Mowing, therefore, removesrelatively more N and K then P when fertilisation isstopped. However, since nutrient pools in the soil aregenerally much higher than plant available N and K(Giisewell, 1997) the soil is not easily depleted. Asmentioned earlier, P-depletion can occur in fen mead-ows that have been mown for a long time withoutfertilisation (Koerselman et al., 1990). Olde Venterink(2000) also found that the availability of both P and Kwas primarily determined by internal nutrient cycling(labile pools, mineralisation), although in flood mead-ows almost 2OVo of the K originated from floodwater.Atmospheric N-deposition was the most important Nsource in low productive meadows, while N frommineralisation was more important in high productivemeadows.
The above mentioned studies indicate that pro-ductivity and consequently also species compositionin hay meadows is primarily determined by internalnutrient cycling within the ecosystem, which is gov-erned by the groundwater fluctuation pattern and ioniccomposition of the groundwater. In low productivefen meadows, atmospheric deposition of N and S
may result in eutrophication and increased alkalin-ity (Lamers et al., 1998). The latter might triggeran increased P-availability and a shift from P to N-limitation. Many present-day fen meadows are alsoinfluenced by drainage from surrounding agriculturalareas, and acidify rapidly. Their topsoil is develop-ing fibric characteristics indicating nutrient accumu-lation and decreased productivity. Pedologically thesesoils irreversibly change towards podzolic systems, inwhich iron is leached by a process offerrolyses. Suchsoils can not easily be restored by hydrological meas-ures intended to restore the original high base status(Faulkner& Richardson, 1989; Kemmers et al., 2000).
Junco-MolinionCailcion nigrae

155
Table l. Above-ground biomass and nutrient contents of phanerogams and mosses of three meadow communities, measured in July 1995):
Junco-Molinion, Calthion palustris and Glyceria maxima dominated flood meadow (after Olde Venterink, 2000). SigniÍicant differences
(p<0.05) between plant communities are indicated with different characters
Plant community Above ground. _4.
blomass (g m -)N(kg ha-l;
P
(kg ha-1)
K(kg ha-l)
Molinia coe rulea meadow
Phanerogams
Mosses
C altha palustris meadow
Phanerogams
Mosses
Glyceria maxima meadow
Phanerogams
Mosses
8
8
'7
7
9
I
352c
t3tp
47gb
24q
9lsa3r
47c
19p
6gb
5s
l32q
1r
3d
2P
7c
lp
18',
0r
3ob
12p
32b
2q
l5a0'
Seed set, seed banks and seed dispersal
Haymaking is a dominant feature of meadows and
the period of cutting depends on the productivity ofthe ecosystem. In general, the least fertile meadow
types (fen- or litter meadows) are mown late, whilethe more productive hay meadows are cut early inthe summer, and they may be cut twice. Nutrient-poor fen meadows are usually mown in September
and many characteristic species set seed in late sum-
mer. Most hay meadow species set seed between Mayand July. Mowing occurs in June/July. Some product-
ive Magnocaricionmeadows are cut twice (eady June
and late August). Ter Borg (1972) showed that some
species have adapted the period of seed set to the mow-
ing period. She found genetic differentiation withinnitianthus angustifulius with respect to the period ofseed set, Early flowering populations were found inrelatively productive grasslands mown in May. Inter-mediate flowering populations produced seed before
the flrst cut in July, while a late flowering population,
found in a large sedge meadow that was mown twicea year, flowered after the flrst cut and produced ripe
seeds before the second cut.The period of mowing is important for seed dis-
persal in yet another way. Nowadays species-rich
meadows are generally mown by machines that go
from one fleld to another on the same day. As an exper-
iment, Strijkstra et al. (1997) cleaned different parts
of the mower between two flelds in order to estimate
seed transport. The mower has a horizontal safety skirtcovering the mowing disk, which traps a lot of seeds.
When the machine leaves the field, the part covered
by the safety skirt is put vertically, and all the seeds
drop onto the soil. The skirt is, therefore, only im-portant for seed transport within fields. The skid disk
of the mower, at soil level, remains in the same po-
sition and is, therefore, important for seed transport
between fields. Strijkstra et al. (1991) estimated that
transport by haymaking machinery could amount to
over 1 000000 seeds during haymaking.Seeds that are not removed from the field by
mowers drop onto the soil. The seeds that are not pred-
ated or do not decay become incotporated into the soil
seeds bank. Seeds in the seed bank can be classifled
according to their estimated longevity. Species onlypresent in the established vegetation and not in the
seed bank are referred to as having a transient or short-
lived seed bank. The same holds for species that onlyhave viable seeds in the topsoil, Species with many vi-able seeds in deeper soil layers are classifled as having
a long-term persistent seed bank, since it is assumed
that it takes time to become buried in the deeper soil
layers (Bekker et al., 1998). Van Diggelen (1998) used
the soil seed data bank of Thompson et al. (1997)
and found that in comparison to natural communities,meadow communities seem to consist mainly of spe-
cies with a longer-lived seed bank. Fifty seven percent
of the characteristic meadow species show indications
of still having viable seeds after 5 years and 307o
even after 20 years. These results suggest that mead-
ows can be easily restored from the soil seed bank
after they have been lost, This was, however, true forcommon hay meadow species, not for the rare ones.
Field studies showed that most Red List species offen meadow s (J unc o - M ol ini on; Maas & Schopp- Guth,

156
1995) hay meadows (Calthion palustris; Biewer &Poschlod, 1995) and flood plain meadows (Magnoca'
ricion; McDonald et al., 1996) have short-lived seed
banks.
Restoration of species-rich meadows: why andhow?
From the beginning of the 20th century onwards, ateas
have been acquired by private organisations for land-
scape and nature conservation purposes, more recently
also by the State.
The reason for the maintenance or restoration ofbrook valley systems is their typical semi-natural char-
acter; a complex historical matrix superimposed on a
very complex geological matrix. The occurrence of a
large variety of environmental gradients ranks brook
valley systems in the top of species-rich environments
in the Netherlands.
M ow in g without fe rtilis ation
The most common technique to restore species-rich
meadows is to re-introduce a mowing regime in formeragricultural flelds or abandoned meadows without ad-
ditional input of fertiliser (Bakker & Olff, 1995:'
Muller et al., 1998). Grassland communities that have
high species richness achieve a maximum standing
cróp, including litter, of between 4 and 6 tons ha-l(Grime, 1979). Both fertilised grasslands (because ofnutrient input) and abandoned grasslands (because oflitter accumulation) have a much higher above-ground
standing crop. Therefore, it is anticipated that a re-
duction of the maximum above-ground biomass willtrigger an increase of species richness, provided thatthe target species are indeed present in the soil seed
bank, or can be dispersed from the actual species
pool. Grasslands under a restoration management, in-deed, showed a decrease of above-ground standing
crop, after the cessation of fertiliser application (Olff& Bakker, 1991).
In order to investigate how the result of the restor-
ation management depends on the fertilisation history,fields have been selected with similar soil and hydro-logical conditions, but with different fertilisation his-
tories. All fields were cut once a year in late July/earlyAugust, Fertiliser application ceased in different years
in rhe selected fields (Fig. 5): 1985 (field B), 1967
(field F) and 1945 (fleld D), respectively. Differencesbetween flelds are assumed to have been established
/:\ FIELD\9@
@
@
Figure 5. Estimated cumulative N input (a), (fertiliser application
and atmospheric deposition since 1947), cumulative N off-take by
mowing (b) and cumulative N balance (c) (difference between input
and off-take) in three grasslands differing in time under restora-
tion management (not fertilised for 45 (D), 20 (F) and 2 (B) years,
respectively, changed after Bakker & Oltr, 1995)'
since then. The species composition of the flelds was
assessed in 1994. Poa trivialis and lnlium perenne
dominated in field B, Rhinanthus angustifulius in f,eld
R and Juncus acutiflorus in field D' Only field Dhad many target species. Taking 1945 as a starting
point, when the input of artiflcial fertiliser was low, an
estimation can be made of the cumulative input of ni-trogen (including atmospheric deposition) and output
via cuttings. The difference between input and out-
put estimates enables a cumulative nitrogen balance(CNB) to be calculated for different fields (Bakker &Olff, 1995). It is obvious that field D shows a negative
CNB from the beginning of restoration management.
LE ooooia
=o..E 4000zo
E zooo5E=o
3000
2000
'1000
(íEËnvoY(íoz0)
(d
=Efo
(uECDvooc(§Eozo.=(d
=E:o
@@@
@
@

Field F reached the tuming point from positive to neg-ative CNB in 1980, whereas f,eld B still has manyyears to go to achieve a negative CNB. Differences inCNB coincided well with differences in above-groundstanding crop in July and accounted better for the num-ber of species per plot than did the number of yearswithout fertiliser application (Bakker & OIff, 1995).Concluding, it is important to consider the fertilisationhistory in restoration management.
The low yields of hay obtained from unfertilisedspecies-rich meadows are uneconomical for farmersto harvest without receiving financial compensationfrom public sources. These meadows need special-ised nature management using equipment especiallydesigned to mow extensive wet areas with little man-power.
Grazing in weï meadows
The introduction of extensive grazing systems, whichis promoted in many European countries to sustain orrestore species-rich grasslands (Pfadenhauer & Groot-jans, 1999), is not applied on a regular basis in Dutchwet meadow ecosystems. Most often it is combinedwith a mowing regime; in particular young cattle maygraze wet meadows in late summer after the meadowshave been mown. Light cattle breeds are sometimesused in large eutrophic wetlands on mineral soils.Small brook valleys with wet organic soils are con-sidered unsuitable for cattle grazhg by the Dutchnature conservation organisations. Cattle prefer dryplaces within a wetland leading to a rapid growth oftall herbs in the wet areas (Bakker & Grootjans, 1991;
Bakker, 1998). Similar effects of grazing were ob-served by Kaiser (1995) in eutrophic wet meadows inthe former German Democratic Republic where a res-
toration management by extensive grazing (stockingrate of 1 animal ha-1) was applied in an intensivelyused agricultural field of 37 ha. After four years ofgrazing(1991-1995), ruderal species, such as Cirsiumarvense, were spreading in dry and moist areas, whiletall grasses, such as Phalaris arundinacea, Elymus re-pens and Agrostis stolonifura became dominant in thewet parts of the meadows.
Grazing in very low productive Cirsio-Molinietumfen meadows may cause health problems for cattle,particularly in autumn, when Molinia caerulea rcal-locates nutrients from the leafs and stores them in thebasal nodes ofthe tussock to be used again for the nextseason. Fen meadows dominated by Molinia caeruleabecome very unsuitable for grazing in autumn, and the
151
health of live stock may suffer severely (Tallowin &Smith, 1996).
Rewetting
Rewetting often requires more than just raising localwater levels in nature reserves. If, for example, wa-ter levels in the surroundings have been lowered such
measures may lead to acidification of the top layersince only precipitation water will be stored withinthe reserve (Grootjans eÍ al., 19961, van der Hoek& Kemmers, 1998). Sometimes the closing of deep
ditches to promote seepage of calcareous groundwa-ter must be followed by re-installing shallow ditches
to prevent inundation with acid precipitation water.Restoration of the whole hydrological system is some-
times necessary (van Diggelen et al,, 1994), for in-stance, by eliminating drainage systems or closingdown groundwater abstraction facilities. But often thisis not an option. Technical measures, such as isolat-ing the effects of drainage by placing artiflcial barriersagainst groundwater flow, is possible but is very costly.New techniques, e.g. deep well infiltration to produce
drinking water without lowering water tables is avail-able but application is mostly restricted to dune areas
(Janssen & Salman, 1995). Rewetting by initiatingflooding of surface water is also a method to restore
hay meadows. Although originally artificial floodingwith surface water was applied to increase the pro-ductivity of the hay meadows (Burny, 1999),it is nowapplied in nature reserves to prevent desiccation and
acidification of the meadows (Spieksma et al., 1994).In nutrient-poor ecosystems, however, flooding withnutrient-rich or sulphate-rich surface water may leadto severe eutrophication (Lamers et al., 1998). This iswhy helophyte filters or Iarge ditch systems are oftenused to purify the water (Meuleman, 1990; van Durenet al., 1998). On one occasion, groundwater from deep
aquifers was pumped to the surface to mimic formerflooding practises (van der Hoek & Kemmers, 1998).
Sod cutting
Sod cutting used to be a traditional measure to in-crease the soil fertility in small arable flelds by tak-ing sods from the common heathlands, which weremixed with dung. Later, nature conservation appliedsod cutting on a large scale in Dutch nature reserves
in order to combat grass encroachment in heathlandvegetation. Sod cutting is now also applied in fenmeadow reserves to counteract the effects of acidific-

158
ation and atmospheric nitrogen deposition (Jansen &Roelofs, 1996). The most drastic impoverishment can
be achieved by complete removal of the nutrient-richtopsoil. This measure is often applied in former agri-cultural fields now destined to become nature reserves.
In some cases the topsoil is removed up to a depth of10-40 cm (van Diggelen et al., 1997; Jansen, 2000).As a disadvantage it must be mentioned that sod cutsites often have obtained a low pH buffer capacity due
to removal of the (organic) cation exchange complex.Such soils are inclined to rapid acidiflcation ifthey are
not buffered by base-rich groundwater (Jansen, 2000).In addition to this radical depletion of nutrients, un-desirable competitive elements are removed, althoughthey may redevelop from the soil seed bank when sod
cutting was too shallow (van Diggelen et al., 1997).Inextreme cases, topsoil removal eliminates practicallythe whole soil seed bank (Klooker et al.,1999).
Successful projects
In the Netherlands, a restoration project is considereda success if many Red List species or target species,
which were common in meadows that existed halfa century ago, re-establish. In a sense, successfulrestoration is like constructing a fleld museum for pre-
serving a living part of a lost cultural heritage. Thisidea of success can very different in other countries(Pfadenhauer & Grootjans, 1999) and even in otherareas ofthe Netherlands, where natural processes canform new ecosystems little disturbed by man.
In the following, we will use the number of targetspecies that has established since the restoration meas-
ures to judge the success of restoration projects. Thelist of target species (see Appendix 1) shows officiallyrecognised Red List species and characteristic species
of wet meadow ecosystems (Schaminée et al., 1996).Fen and hay meadow sites where recent restoration
projects have been carried out are shown in Figure 6.
Projects discussed in the present paper are indicated inblack.
Mowing without fertilisation in wet meadow reseryes('Drentse Aa'valley)
The Drentse Aa brook valley reserve is the best pre-served valley system in the northern part of the Nether-lands. This landscape reserve was established in 1965
when it was decided that 3500 ha (of the 30000 hacatchment area) would eventually be acquired by the
Figure 6. Areas where restoration measuÍes have been applied
recently in existing hay- and fen meadow reserves in the Neth-erlands (changed after van der Linden et al., 1996). Study areas
discussed in the present paper are indicated in black. l=Taarlo,2=Anloo, 3=Eexterveld, 4=de Reitma, 5=de BaÍen, 6=Punthuizen,7=Stroothuizen, 8=Groenet 9=Lemselermaten, l0=Veenkampen,1 l=Grootzandbrink, 12=Wyldlannen.
years
Figure 7. Increase of target species monitored in three peÍmanentplots during 25 years in two brook valley meadows in the Drentse Aaarea. The meadow near the village of Taarlo was fertilised only littlebefore abandonment followed by restoration management, while the
meadow near Anloo had been fertilised for several decades before amowing regime without fertilisation was installed.
State and managed by the State Forestry Service in or-der to preserve and restore the semi-natural landscape.The hydrology of the reserve is, however, heavily in-fluenced by agricultural drainage systems along the
Éo.ÍloooooE"(U
De Barten
a target meadow (rèwetted)
^ rewetting+sod cutting
6
years
Figure 8. Changes in the number of target species in the hay
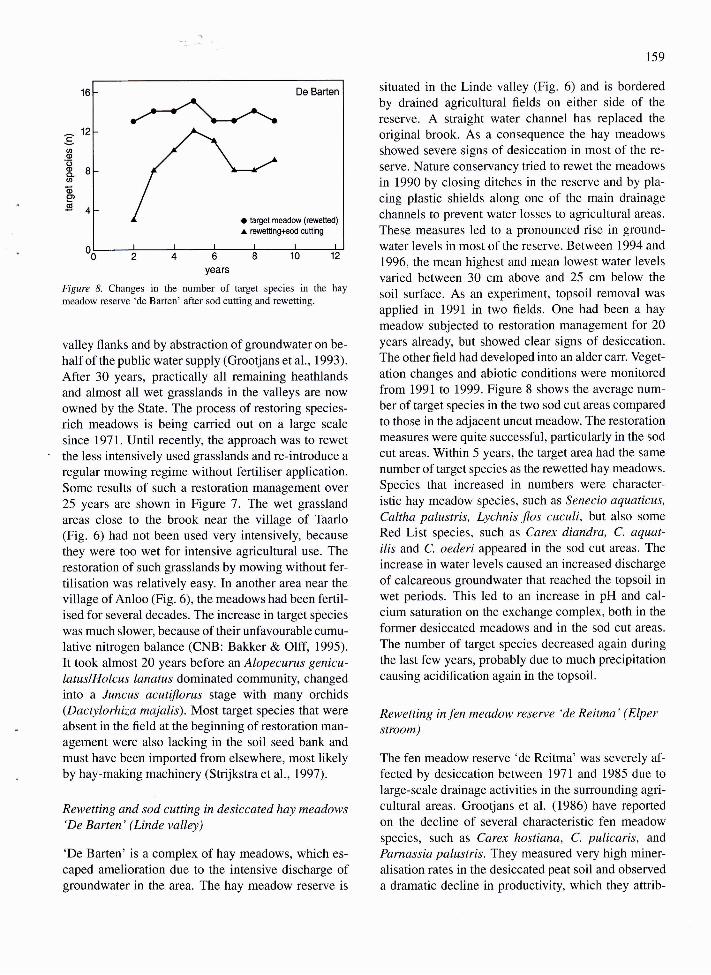
meadow reserve 'de Barten' after sod cutting and rewetting.
valley flanks and by abstraction of groundwater on be-half of the public water supply (Grootjans et al., 1993).After 30 years, practically all remaining heathlandsand almost all wet grasslands in the valleys are nowowned by the State. The process of restoring species-
rich meadows is being carried out on a large scale
since 1971. Until recently, the approach was to rewet
the less intensively used grasslands and re-introduce a
regular mowing regime without fertiliser application.Some results of such a restoration management over25 years are shown in Figure 7. The wet grassland
areas close to the brook near the village of Tharlo(Fig. 6) had not been used very intensively, because
they were too wet for intensive agricultural use. The
restoration of such grasslands by mowing without fer-tilisation was relatively easy. In another area near thevillage of Anloo (Fig. 6), the meadows had been fertil-ised for several decades. The increase in target species
was much slower, because of theirunfavourable cumu-Iative nitrogen balance (CNB: Bakker & Olff, 1995).It took almost 20 years before an Alopecurus genicu-
latus / Holcus lanatus dominated community, changedinto a Juncus acutifiorus stage with many orchids(Dactylorhiza majalis). Most target species that wereabsent in the field at the beginning of restoration man-agement were also lacking in the soil seed bank andmust have been imported from elsewhere, most likelyby hay-making machinery (Strij kstra et al., 1997 ).
Rewefiing and sod cutting in desiccated hay meadows'De Barten' (Linde valley)
'De Barten' is a complex of hay meadows, which es-
caped amelioration due to the intensive discharge ofgroundwater in the area. The hay meadow reserve is
159
situated in the Linde valley (Fig. 6) and is borderedby drained agricultural flelds on either side of the
reserve. A straight water channel has replaced the
original brook. As a consequence the hay meadows
showed severe signs of desiccation in most of the re-
serve. Nature conservancy tried to rewet the meadows
in 1990 by closing ditches in the reserve and by pla-cing plastic shields along one of the main drainage
channels to preyent water losses to agricultural areas.
These measures led to a pronounced rise in ground-water levels in most of the reserve. Between 1994 and1996, the mean highest and mean lowest water levels
varied between 30 cm above and 25 cm below the
soil surface. As an experiment, topsoil removal was
applied in 1991 in two fields. One had been a hay
meadow subjected to restoration management for 20years already, but showed clear signs of desiccation.The other fleld had developed into an alder carr. Veget-
ation changes and abiotic conditions were monitoredfrom 1991 to 1999. Figure 8 shows the average num-ber of target species in the two sod cut areas compared
to those in the adjacent uncut meadow. The restorationmeasures were quite successful, particularly in the sod
cut areas. Within 5 years, the target area had the same
number of target species as the rewetted hay meadows.
Species that increased in numbers were character-istic hay meadow species, such as Senecio aquaticus,
Caltha palustris, Lychnis flos cuculi, but also some
Red List species, such as Carex diandra, C. aquat-ilis and C. oederi appeared in the sod cut areas. The
increase in water levels caused an increased discharge
of calcareous groundwater that reached the topsoil inwet periods. This led to an increase in pH and cal-cium saturation on the exchange complex, both in the
former desiccated meadows and in the sod cut areas.
The number of target species decreased again duringthe last few years, probably due to much precipitationcausing acidiflcation again in the topsoil.
Rewetïing in fen meadow reserve 'de Reitma' (Elperstroom)
The fen meadow reserve 'de Reitma' was severely af-fected by desiccation between 1971 and 1985 due tolarge-scale drainage activities in the surrounding agri-cultural areas. Grootjans et al. (1986) have reported
on the decline of several characteristic fen meadowspecies, such as Carex hostiana, C. pulicaris, andParnassia palustris. They measured very high miner-alisation rates in the desiccated peat soil and observeda dramatic decline in productivity, which they attrib-
16
^12go
.o)og8ooP§4
r0

À PunthuizenJ \-{fStoothuizen
^-.--'"J+Ff
3 o mowino8 t sod cuiing
160
-05101s2025303540years
Figure 9. Changes in the number of target species after res-toration observed in two fen meadow reserves (Punthuizen andStroothuizen).
uted to flxation of phosphorus by iron complexes.Most of the drainage ditches around the reserve wereclosed again in 1981, leading to a considerable in-crease in groundwater discharge to the reserve. Thefen meadows in 'de Reitma' were almost completelyrestored (Zeeman, 1986). Despite almost 10 years ofsevere desiccation, all target species, except Pamassiapalustris, returned, although some in smaller numbers(C. hostiana).
Rewetting and sod cutting in a heathland andfenme adow re s e rv e (' P unthuiTen' )
'Punthuizen', situated in the north-eastern part ofTwente, is one of the best preserved heathlands and fenmeadows in the eastern part of the Netherlands (Fig.6). The reserve ofc. 80 ha. is characterised by a mosaicof strongly podzolised elevated parts and depressionswhere some organic material has accumulated. Thereserve was established in the 1960s to preserve relicsof the former extensive heathlands, fen meadows andsoft water pools. The traditional management of mow-ing without fertilisation and occasional sod cutting hasbeen continued since the 1960s by the State ForestryService. However, the reserve is surrounded by in-tensively used agricultural flelds, and its hydrology isheavily influenced by drainage activities. This has ledto a sharp decrease in typical fen meadow species.
Sod cutting along the height gradient resulted inthe successful restoration of species-rich fen mead-ows with many Red List-species (Fig. 9). Lycopo-dium inundatum, Rhynchospora and Drosera speciesre-appeared on acid sites, while Cicendia filiformis,Juncus tenageia and Radiola linoides were found on
Lemselermaden
Groener
Eexterveld
6
years
Figure 10. Changes in the number oftarget species after removingthe eutrophic topsoil in three former agricultural lields (Groener,
Lemselermaten and Eexterveld).
base rich sites. Sometimes, species of initial phases
of fen meadows, e.g. Pinguicula vulgaris and Saginanodosa, appeared within I year after sod cutting, butthey did not establish a stable population,
The successful restoration of basiphilous fenmeadows was almost entirely due to successful re-construction of the original hydrological system, inwhich the presence of pools during wet periods iscrucial in sustaining the required site conditions (pH,base-status) of these plant communities (Jansen et al.,
200 I ).
Rewetting and topsoil removql in former arable fields('Stroothuizen' and' Lemselermaten' )
The former agricultural f,eld 'Groener' (c, 7 ha) bor-ders the old fen meadow reserve 'Stroothuizen', wheremany typical fen meadow species are still present.This intensively used arable land was reclaimed fromheathlands some 40-60 years ago and acquired in199l.It had been completely levelled after reclama-tion from heathland in the 1930s. Before 1991, comwas grown here and huge amounts of manure andpesticides were used. During the winter of 199311994,the original topography was restored by removingthe eutrophic topsoil. These measures were carriedout so that the former modest topography of pools,long and narrow slacks and ridges was restored and
connected to the topography of the adjacent reserveStroothuizen. Thus, the entire area could be floodedagain. Several additional hydrological measures werecarried out to further rewet the area: filling up deepditches and drain tubes, which resulting in a local in-crease in groundwater discharge and consequently a
25
tro.o(toCLo0)P(U
U'o(,oCLoo)CD(U
15
10
1210-0

161
higher base-saturation in the topsoil at such sites. Thecombination of hydrological measures and topsoil re-moval has resulted in the colonisation by many newspecies (Fig. 10), such as Littorella uniflora, Pilulariaglobulifura, Sagina nodosa and Carex echinata. Sev-eral endangered moss species also established, such
as Phaeoceros carolinus and Anthoceros caucasicus,which was last found in the Netherlands in 1926. Mostof these species still occur and are spreading. Some
of the soft-water species have disappeared, due to thedry year 1996.IÍ is astonishing that so many pioneerspecies were still present in the seed bank. Some ofthese species, such as Samolus valerandi, Luroniumnat(tns, and Littorella uniflora were unknown for thispart of Stroothuizen.
Topsoil removal and rewetting was also appliedin a former agricultural grassland which was situatedadjacent to an existing fen meadow reserve (Lemsel-ermaten). This grassland had been used as agricul-tural grassland for 40 years, the last decades ratherintensively. This restoration technique was very suc-cessful, since many Red List species re-appeared (Fig.10). High water tables prevailed in the sod cut pafisand base-rich groundwater discharged from a largegroundwater system. This deep groundwater, how-ever, is now rich in SO]-, while originally SO]-concentrations were very low These higher sulphateconcentrations might cause eutrophication in the rootzone in the future (Lamers et al., 1998). It remainsdoubtful, therefore, whether the restoration measures
in this part of the meadow reserve will result in a
sustainable restoration of the fen meadows.Sod cutting of a degraded fen meadow in the
Lemselermaten did not result in the re-establishmentof a species-rich fen meadow. Sod cutting has ledto prolonged stagnation, which probably has caused
NHf -rich conditions, which are harmful for many fenmeadow species (De Graaf et al., 1998).
Unsuccessful projects
Tharlose diep (' Drentse Aa' valley)
The wet meadows along the Taarlose Diep are part ofthe hay meadow reserve Drentse Aa, where just beforethe establishment of the reserve deep drainage ditcheswere constructed to drain adjoining agricultural areas.
Furthermore, this area of the Drentse Aa is partly in-fluenced by groundwater abstraction on behalf of thepublic water supply (Grootjans et al., 1993). The activ-
Taarlo, drained
{VCo.ÍtooCLo(Dc»(§
4
0ö"0s1015202530years
Figure 11. Changes in target species monitored in permanent plots
during 25 years in hay meadows along the brook valley (Taarlose
Diep) in the Drentse Aa area. Plots were situated in fields influencedby drainage and in flelds with an undisturbed hydrology.
o mowinga mowing+rewetting
À mowing+sod cutting
^ mowing+rewetting+sod cutting
Veenkampen
ffi
o24681012years
Figure 12. Changes in the number of target species in the experi-mental restoration project 'Veenkampen', where former species richmeadows are being restored under different hydrological regimes.
ities had led to local desiccation of meadows subjectedto a restoration management since 1972. Analysis ofpermanentplots permitted an evaluation on the species
level over a period of 25 years. Some plots were situ-ated in areas affectedby drainage ditches, while otherswere unaffected. The plots were lying close together inflelds that had the same management history (Bakker,
1989). After 25 years, practically all target species ofthe drained Calthion meadows had disappeared (Fig.11).
These results indicate that, despite a restorationmanagement of 23 years, no real progress has been
made with respect to target communities, because ofsub-optimal hydrological conditions.
12
c;8o'6g6@
g4(o
2
0

162
Table 2. Changes in nutrient conditions in the topsoil of the hay meadow reserve Veenkampen after two different hydrological restorationmeasures (after van der Hoek & Kemmers, 1998). Values are given in percentages (relative to the control where no hydrological restoration
measures were taken)
CompaÍment 1986 r989 1997t991 t993
Exch. Ca
Org. Matter
N-tot
P{ot
P-exch.
Conservation
Inigation
Conservation
Irrigation
Conservation
Irrigation
Conservation
Irrigation
Conservation
Irrigation
100
r00
t00
100
r00
100
r00
100
100
r00
l498
9l102
105
rt2105
87
t69
69
103
86
89
t07
104
105
t2086
132
56
95
96
98
105
95
t01
108
86
135
46
89
91
91
106
95
t07
108
95
t4l46
Rewetting and sod cutting in eutrophicatedfenme adow s (' Ve enkamp en' )
The 'Veenkampen' is situated in the Gelderse Vallei inthe lower Rhine area (Fig. 6), which once was fam-ous for its wealth of species-rich fen meadows. Onlysmall remnants have remained. The Veenkampen isa complex of former agricultural fields where the re-generation of species-rich meadows has been studiedin experiments since 1986. Mowing without fertilisa-tion has been applied in part of the complex. Theother pafis have been additionally rewetted and sod
cut. Vegetation response on changed geochemical andhydrological conditions has been studied for morethan a decade (Berendse et al., 1994; Oomes et al.,1996, 1991; van der Hoek & Kemmers, 1998). Re-wetting was carried out by conserving precipitationwater within some fields and by subsurface irrigationof calcium-poor groundwater (Ca-content=10 mg l-1)from an artesian well, supplying groundwater froma conflned aquifer at 50 m below the surface. Thesoils of the experimental fields consisted of peat witha shallow clayey top layer. They have been seriouslyaffected by 40 years of intensive agricultural use. Inor-ganic phosphate accumulation had occurred and couldnot be reduced to its original levels by the restorationmeasures applied (Table 2). The study clearly showedthat rewetting by conserving precipitation water in thefleld actually increased the P-availability for the ve-getation, even after 10 years. Subsurface irrigationwith calcium-poor groundwater from an artesian welldecreased P-availability considerably after 5 years,
but could not prevent further acidification of the to-player. The decrease in P-availability was caused by
the dissolution of iron-phosphates under anaerobic
conditions and subsequent leaching of P to the sub-
soil. The conservation ofprecipitation water appeared
to be an inadequate measure to restore species-richgrasslands. Almost no target species re-appeared inthe experimental fields. Fields that were sod cut to
drastically remove the high nutrient stocks were more
successful in establishing populations of target spe-
cies (Fig. l2), bÉ most of the species were pioneerspecies, which probably emerged from long-persistentseed banks, even after 40 years.
Rewetting and sod cutting in eutrophicatedfenmeadows ('Stroothuizen' )
After the nature reserve 'Stroothuizen' (c. 40 ha.)
was founded in the 1960s, the manager decided to
construct a dike along its western border in order tomaintain high water levels. As a result, many de-
pressions were inundated with base-poor precipitationwater for at least 4 months. Calcareous groundwatercould no longer reach the surface due to the pres-
ence of drainage ditches and drainage pipes in adjacentarable fields. This led to inf,ltration of precipitationwater and subsequent acidif,cation and eutrophication.
To stop the decline of mesotrophic fen meadowspecies, sod cutting was applied in large parts ofthe reserve. This measure was successful in restoringspecies-rich dwarf-rush communities, wet heathlandsand small-sedge marshes, but not in restoring fenmeadows (Fig. 9). Whereas the restored wet heath-lands are now situated at the upper part of the heightgradient, the small-sedge communities occur in the
lower part. The failed restoration of the fen meadow

vegetation might be due to increased P-levels in thetopsoil because of prolonged inundation leading todissolution of Fe-P and Al-P minerals under anaerobicconditions (Boeye er"al., 199'7; Jansen, 2000).
Rewetting without sod cutting in the degeneratedfen meadow led to very incomplete recovery comparedto the original fen meadow that existed some decades
ago (Jansen, 2000). This might be due to irreversiblechanges in the structure of the organic topsoil layer.Long-term drainage probably had led to iron depletion(de Mars, 1996).
Acidifie d fen me adows (' Wyldlannen' )
The fen meadow complex 'Wyldlannen' consists ofc. 60 ha of degenerated and species-poor Cirsio-Molinietum vegetation in which Agrostis caninaand Phalaris arundinacea are dominating on clay-peat soil. Characteristic species of the community,e.g. Cirsium dissectum, Carex panicea and Moliniacaerulea, are still present, although in low numbers.The meadows were inundated by stagnating precipita-tion water for several months during winter and earlyspring. Calcareous groundwater could no longer reachthe nature reserve, since the surrounding polder areas
had been deeply drained for several decades. These
deep drainage practices had caused a severe loweringof the peat surface in all polder areas, leaving the fenmeadow reserve as the highest area in the landscape.
Attempts to restore the fen meadows started in1986 when the water levels in the ditches were raised.This led to some rewetting in the summer, but not toan increase of characteristic fen meadow species. Anold practice to flood the meadow with surface waterwas resumed in l99l to stop the acidification. A smallwetland (helophyte filter) of c. t ha was used to reducenutrient availability in the surface water. Sod cuttingwas applied to a small area of 0.5 ha to investigate iftopsoil removal of c. 20 cm would produce a less acidi-fied environment for fen meadow species. Sod cutting,however, exposed a former Sphagnum-peat, with quitedifferent chemo-physical characteristics.
At some distance from the degraded fen meadowcomplex, a small area ('Ulekrite') exists where a well-developed Cirsio-Molinietum st1ll harbours species,e.g. Pedicularis palustris, Carex hostiana, C. flacca,C. echinata and C. lasiocarpa.
Van Duren et al. (1998) attempted to restore thefen meadow by means of rewetting, sod cutting, lim-ing and introduction of target species. An evaluationwas done in the framework of a monitoring project
Wyldlannen
o mowinga reÍerencea sod cutting
6
years
Figure 13. Changes in the number of target species in the fenmeadow reserve 'Wyldlannen' after carrying out Íestoration meas-
ures, such as sod cutting and re-flooding with purilied surface
water.
carried out at three sites: a degenerated, a sod cutand a reference site. Groundwater levels in the de-
generated and the sod cut sites differed slightly fromthat of the reference site but were within the range
of Cirsio-Molinietum requirements as known from theliterature. Topsoil removal considerably had reducedmacro-nutrient contents and caused a pronounced P-
deficiency. No increase in soil pH was observedimmediately after the topsoil removal. A slight in-crease in soil pH was found, however, after 5 years
in the sod cut site, indicating a slight recovery fromacidiflcation. Liming after topsoil removal had littleeffect on the soil pH after 6 months but stimulatedthe growth of established species as Agrostis canina.Few viable seeds of characteristic fen meadow species
were present in the seed banks of both degenerated
and well-developed fen meadow sites and the species
composition after topsoil removal closely reflected the
seed bank of the degenerated fen meadow. Establish-ment of characteristic fen meadow species was poor,
even after 9 years (Fig. 13). The introduced species
survived during the first year in both the degeneratedand the sod cut sites. Liming stimulated the growthof two of the introduced species. In the second yeaq
however, all introduced species showed considerablegrowth reduction and the positive effect of liming dis-appeared. After 2 years, none of the introduced species
survived in the sod cut site, indicating that the envir-onmental conditions were still unfavourable. It wasconcluded that the restoration measures failed so far.apparently due the continuation of deep drainage in thesurrounding agricultural areas. Restoration of base-
rich conditions is further hampered by the very low
163
16
^12Éo.Í9o&8o0)o)§4
r64
iron contents of the Sphagnum-peat (Kemmers et a1.,
2000).
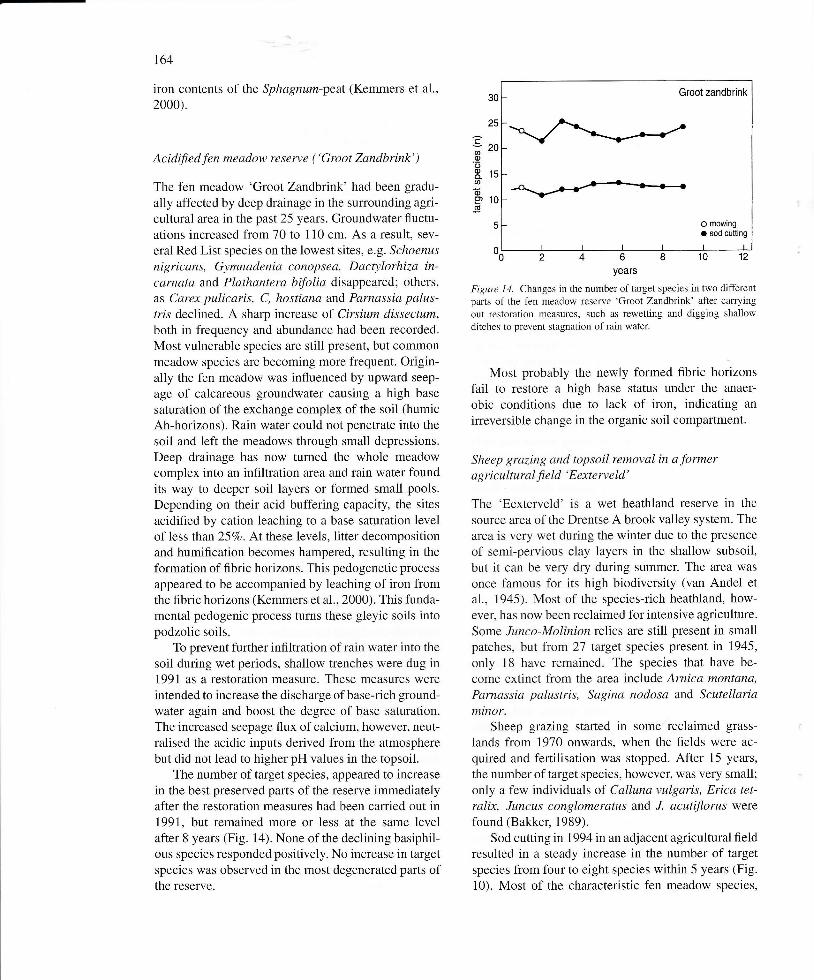
Acidified fen meadow reserve ( 'Groot Zandbrink' )
The fen meadow 'Groot Zandbrink' had been gradu-
ally affected by deep drainage in the sur:rounding agri-cultural area in the past 25 years. Groundwater fluctu-ations increased from 70 to 110 cm. As a result, sev-
eral Red List species on the lowest sites, e.g. Schoenus
nigricans, Gymnadenia conopsea, Dactylorhiza in-caruata arrd Plathantera bfolia disappeared; others,as Carex pulicaris, C, hostiana and Parnassia palus-rrls declined. A sharp increase of Cirsium dissectum,both in frequency and abundance had been recorded.
Most vulnerable species are still present, but commonmeadow species are becoming more frequent. Origin-ally the fen meadow was influenced by upward seep-
age of calcareous groundwater causing a high base
saturation of the exchange complex of the soil (humicAh-horizons). Rain water could not penetrate into the
soil and left the meadows through small depressions.Deep drainage has now turned the whole meadowcomplex into an infiltration area and rain water foundits way to deeper soil layers or formed small pools.
Depending on their acid buffering capacity, the sites
acidified by cation leaching to a base saturation levelof less than 257o. AÍ these levels, litter decompositionand humiflcation becomes hampered, resulting in theformation of flbric horizons. This pedogenetic process
appeared to be accompanied by leaching of iron fromthe fibric horizons (Kemmers et al., 2000). This funda-mental pedogenic process turns these gleyic soils intopodzolic soils.
To prevent further inflltration of rain water into the
soil during wet periods, shallow trenches were dug in1991 as a restoration measure. These measures wereintended to increase the discharge ofbase-rich ground-water again and boost the degree of base saturation.The increased seepage flux of calcium, however, neut-ralised the acidic inputs derived from the atmospherebut did not lead to higher pH values in the topsoil.
The number of taÍget species, appeared to increasein the best preserved parts of the reserve immediatelyafter the restoration measures had been carried out in1991, but remained more or less at the same levelafter 8 years (Fig. 1a). None of the declining basiphil-ous species respondedpositively. No increase in targetspecies was observed in the most degenerated parts ofthe reserve.
0-024681012years
Figure 14. Changes in the number of target species in two differentparts of the fen meadow reserve 'Groot Zandbrink' after carrying
out restoÍation measures, such as rewetting and digging shallowditches to prevent stagnation of rain water.
Most probably the newly formed flbric horizonsfail to restore a high base status under the anaer-
obic conditions due to lack of iron, indicating an
irreversible change in the organic soil compartment.
Sheep grazing and topsoil removal in a formeragricultural field' E ext erv eld'
The 'Eexterveld' is a wet heathland reserve in the
source area of the Drentse A brook valley system. The
area is very wet during the winter due to the presence
of semi-pervious clay layers in the shallow subsoil,
but it can be very dry during summer. The area was
once famous for its high biodiversity (van Andel et
al., 1945). Most of the species-rich heathland, how-ever, has now been reclaimed for intensive agriculture.Some Junco-Molinion relics are still present in smallpatches, but from 27 target species present in 1945,
only 18 have remained. The species that have be-
come extinct from the area include Arnica montana,
Pamassia palustris, Sagina nodosa and Scutellariaminor-
Sheep grazing started in some reclaimed grass-
lands from 1970 onwards, when the flelds were ac-
quired and fertilisation was stopped. After 15 years,
the number of target species, however, was very small;only a few individuals of Callunavulgaris, Erica teÍ-
ralix, Juncus conglomeratus and J. acutiflorus werefound (Bakker, 1989).
Sod cutting ir 1994 in an adjacent agricultural fieldresulted in a steady increase in the number of target
species from four to eight species within 5 years (Fig.10). Most of the characteristic fen meadow species,
c;20o'6815o()P10(§
5
Groot zandbrink
o mowinga sod cutting

165
such as Cirsium dissectum, Carex hostiana, however,
did not re-appear. Many species that emerged aftertopsoil removal (Juncus bufonius, Rorippa palustris,Alopecurus geniculatus and Holcus lanatus, were also
dominant in the soil seed bank (Klooker et al., 1999).
Other species (Juncus effusus, Ranunculus repens)
probably recovered from root fragments that were notremoved. Erica tetralix was the only wet heathland
species still present is the soil seed bank. This spe-
cies was not only found in the topsoil, but also at
depths between 20 and 50 cm. Species that could reach
the target area by wind dispersal were practically the
same species as the ones present in the seed bank(Alopecurus geniculatus, Poa annua, Juncus bufoniusand Sagina procumbens. The only target species thatwas found in the local seed rain was Viola palustris.
Environmental conditions, immediately after topsoilremoval were not favourable for acidophilous species.
The pH remained relatively high (c. 5.5) and also the
phosphorus levels remained high. These high values
still reflect the effect of intensive fertilisation beforer 995.
Evaluation
The case studies discussed above show that the timethat has elapsed since the degradation ofa fen meadow
is critical for the restoration prospects of a site. Some-
times restoration is achieved with relatively little ef-fort, while in other cases, restoration fails completelydespite large costs on restoration measures.
Conceptual model of fen meadow restoration
A conceptual model of fen meadow restoration sum-
marising successes and failures of restoration meas-
ures is presented in Figure 15. In most species-rich
meadow reserves in the Netherlands, a traditionalmowing regime without fertilisation was continuedor re-instated by state or private nature conservation
organisations. A success story is, forinstance, the cre-
ation of the largest meadow reserye in the Netherlandsin the Drentse Aa catchment. Most of such meadows
responded very well to a restoration management (Fig.15: 1) after a short period of abandonment at time ft.Parts affected by drainage, however, responded wellin the beginning, but lost many target species aftersome 10 years of restoration management (2; example:'Taarlo drained'). Rewetting and/or sod cutting wasquite successful in cases, where the process of de-
Tl T2 T3 f4 T5
time ------>
Figure 15. Conceptual model of occuffence of target species in
fen and wet hay meadows under restoration management. Continu-
ation of the traditional management (mowing without fertilisation)in meadow reserves can not not always prevent the extinction ofmany endangered species negative influences from surrouldingagricultural areas. Restoration measures, e.g. rewetting and sod
cutting are less effective after long-term exposure to these influ-ences. Resuming a traditional management after a short period
of abandonment is often very successful with respect to estab-
lishing of target species. If restoration management is resumed
shortly after cessation of agricultural use, the restoration success
is usually high compared to situations where intensive fertilisationhas taken place for a long time. M=mowing without fertilisation,
W=(re)wetting, S=sod-cutting. The numbers correspond to the case
studies mentioned in the text.
siccation had proceeded only for a few decades (3;
examples; 'Barten', 'Reitma'). When restoration man-
agement starts in meadows, which had been subjected
to intensive fertilisation for 10-30 years (4), the in-crease of target species (starting at T2), depends on
the amount of nutrients accumulated during former
fertiliser application (Bakker & Olfl 1995; example'Anloo'). It may take 5-15 years (Ts-fa), before the
nutrient output has reduced the nutrients stocks to a
level in which the productivity of dominant species
no longerprevents the establishment oftarget species.
Even more target species established in former agri-cultural areas on mineral soils after top soil removal,but only when former hydrological conditions were
restored, seed banks had not been depleted and where
dispersal mechanisms (flooding) from adjacent nature
reserves were effective (Jansen et a1., 2000; examples
'stroothuizen','Punthuizen','Lemselermaten' and
'Groener').Resuming the traditional management (grazing,
mowing without fertilisation) in grasslands that have
been fertilised intensively in former days, will notlead to rapid success without rewetting and topsoil
1
a.ooooaoo(É

166
removal. This is clearly illustrated for the 'Veen-
kampen" experiment (Oomes et al., 1996) wheremowing without fertilisation was very unsuccessful,even after l0 years (7), although several target species
were present in the seed bank. Mowing in combina-tion with rewetting was much more successful (6) andcombined with sod cutting it has lead to re-appearanceof many Red List species with a long persistent seed
bank (5). These species had survived in the soil seed
bank up to 40 years (Oomes et al., 1996). Under ap-
propriate site conditions, therefore, activation of seed
banks can be beneflcial for the survival of endangeredplant species, since the rapid population expansionsafter restoration measures raise the opportunity toproduce a new seed banks.
When the degradation, for instance by acidifica-tion, had affected large sections of the soil proflle,sod cutting was not very successful in increasing thenumber of target species again (8; examples 'Wyld-lannen'). Sod cutting in former agricultural fields,drained and fertilised for a long time ('Eexterveld'), isusually also ineffective since (i) soil degradation has
initiated irreversible changes in the topsoil, (ii) seed
banks are depleted and (iii) dispersal mechanisms are
ineffective (van Diggelen, 1998).
Mechanisms of soil degradation afier long termdrainage
Application of a restoration management in areas thatare influenced by drainage or abstraction of ground-water may yield some success in the beginning, butdepending on the degree of disturbance, it may take5-25 years before the target species disappear again(Bakker, 1989; Grootians et al., 1996). This retardedresponse of plant communities is probably the reason
why the failure of many restoration projects is onlydetected after 10-20 years of restoration manage-ment. Based on hydrological calculations, Runhaar(1999) has estimated that c. 75Vo of the mesotrophicfen meadows (Cirsio -Molinietum, Caricetum ni grae),covering 1500 ha, and 5O7o of the more eutrophichay meadows (Calthion palustris), covering 3500ha, have been negatively affected by hydrologicalchanges. This probably implies that without additionalhydrological measures a reduced success of restora-tion management may be expected in roughly 60la ofall cases.
Several case studies have shown that rewettingdoes not always succeed in 'repairing' fen meadows,which have been affected by drainage for a long time.Indications exist that acidiflcation leads to ferrolysisand loss of iron from the topsoil (Kemmers et al.,
2000). Lack of Fe3+ appears to block an increase inbase saturation after rewetting, despite high calciumlevels in the groundwater. If, under anaerobic condi-tions, both oxygen and iron are absent to facilitate the
exchanges of protons for base-cations on the exchange
complex of the soil, an increase in base-saturation is
not expected and the soil would remain acidic (van
Bremen, 1987; Mulder et al., 1989). We hypothise thatiron plays a key role in the restoration of acidifled fenmeadows. Depending on the iron content in the rootzone, fen meadows can be restored only by hydrolo-gical measures aimed to stimulate upward discharge ofgroundwater or by additional measures as sod cuttingwhich not only remove high nutrient stocks, but also
iron-depleted soil horizons.
Other constraints.for restoration success
Under the present conditions in the Central European
cultural landscape, many characteristic meadow spe-
cies are hardly capable of seed dispersal over large
distances (Bakker et al., 1996; Poschlod et al.,1996;Bakker & Berendse, 1999). Regular flooding withsurface wateq which probably is the most import-ant dispersal agent for many mire species (Danvid &Nilsson, 1997) is nowadays largely prevented. Fur-thermore, donor areas for diaspores have become veryscarce (Poschlod & Bonn, 1998) or can not be reached
by flood water. Hence, even with regular flooding, ad-
equate seed transport to many restoration sites mightbe very restricted. Biological constraints are, there-fore, most severe in restoration areas which have been
intensively used by modern agriculture, where the
eutrophic top layer have been completely stripped,
where atmospheric N-deposition is high (>15-20 kgN ha-l yr-l; Bakker & Berendse, 1999) and where
no species-rich meadows are situated close by (van
Diggelen et al., 1997).Other biological constraints for successful restor-
ation could be the absence of decomposer organ-isms and mycosymbionts, particularly in renderingthe restored system viable in the long term (Fenner,
1985; Turner & Friese, 1998). Furthermore, earth-worm activity has a substantial impact on the soil seed
bank dynamics and hence on the possibility of species
establishment in species-rich grasslands (Willems &Huijsmans, 1994). Mychorchizal fungi enhance the Nuptake of orchids, and could be essential for the sur-
vival of young orchids (Dijk & Eck, 1995). Also, the
presence or absence of speciflc mycorrhizal symbiontsmay be of more importance for the establishment oforchid species than their dispersal capacities. Orchidshave very small and light seeds, that can be dis-

persed easily by wind. So, the limited re-establishment
of most characteristic orchid species in restored fen
meadows after topsoil removal might be explained bythe absence of speciflc mycorrhizal symbionts.
Restoration prospects
Summarising, we can conclude that the most success-
ful projects are on sites that have been least affected
by intensive agriculture and drainage. Thus, the res-
toration projects should be initiated preferably in areas
which have not been affected by drainage and stillhave some relics of meadow species in the vegetation(Giisewell, 1997). In such cases, plant communit-ies similar to target (reference) communities can be
restored relatively easily, although a complete restor-
ation may be impossible (Jansen et al., 2000). Some
authors even object to defining reference states. Parker
& Pickett (1977), for instance, state that the condi-tions of any particular site are the result of a historicalunique combination of processes for that location, im-plying that ideal reference states do not exist (see also
Tallis, 1991). The use ofreferences is, indeed, a veryacute problem in target areas where intensive fertilisa-tion has taken place for a long time, soil degradation
has occurred, and where practically all target spe-
cies have disappeared in both the existing vegetation
and the seed bank. If hydrological conditions cannotbe restored, the new species assemblages that appear
will deviate considerably from old references. In such
cases the goal of traditional references is probably an
illusion. In very degraded brook valleys, for instance,
we would have to remove the degraded topsoil and
introduce meadow species, mycorrhizal fungi, nitri-fying bacteria and take all other (expensive) measures
to stafi an unknown ecosystem anew. In such cases,
it would be much cheaper not to formulate botanicalgoals, but concentrate on environmental goals, such
as soil conservation. Severely degraded peat soils, forinstance, may be effectively conserved if they are
managed as eutrophic wetlands, which are regularlyflooded. Such eutrophic marshes could harbour an in-teresting fauna and could eventually develop into fens
again, when the large nutrient availability is reducedagain by natural biotic or abiotic processes.
Acknowledgements
The monitoring of vegetation changes in the reserves
Barten, Punthuizen, Stroothuizen, Lemselermaten,Groener, Grootzandbrink and Wyldlannen was fln-anced by the Ministry of Agriculture, Nature and
t67
Fisheries in the framework of the Survival Plan ofNature (OBN). Thies Oomes of Plant Research Inter-national (Wageningen) supplied vegetation data of the
reserve Veenkampen. Henk Everts, Latzi Fresco and
Yzaakde Vries helped to proces§ permanent plot data.
Dick Visser and Erwin Adema prepared the figures.
Their help is gratefully acknowledged. Two anonym-ous reviewers are also acknowledged for their positive
comments.
References
Anderson,8.,1949. Hybridisation ofthe habitat. Evolution 2: 1-9.
Bakker, J. P., 1989. Nature Management by Grazing and Cutting.
Kluwet Academic Publishers, Dordrecht.Bakker, J. P. & F. Berendse, 1999. Constraints in the restoÍation
of ecological diversity in grassland and heathland communities.
Trends Ecol. Evol. 14:63-68.Bakker, J. P., 1998. The impact of grazing on plant communit-
ies. In Wallis De Vries, M. F., J. P. Bakker & S. E. van
Wieren (eds), Grazing and Conservation Management. KluwerAcademic Publishers, Dordrecht: 131-184.
Bakker, J. P. & A. P. Grootjans, 1991. Potential for vegetation re-
generation in the middle course of the Drentsche A Brook valley
lthe Netherlands.l. Verh. Gesell. Ökol. 20: 249 -263.
Bakker, J. P. & H. Olff, 1995. Nutrient dynamics during restorationof fen meadows by haymaking without fertiliser application. InWheeler, B. D., S. C. Shaw, W. J. Foyt & R. A. Robertson (eds),
Restoration of Temperate Wetlands. John Wiley, Chichester:
143-166.Bakker, J. P. & G. Londo, 1998. Grazing for conservation man-
agement in historical perspective. In Wallis De Vries, M. F." J.
P. Bakker & S. E. van Wieren (eds), Grazing and Conservation
Management. Kluwer Academic Publishers, Dordrecht: 23-54.
Bakker, J. P., P. Poschlod, R. J. Strijkstra, R. M. Bekker &K. Thompson, 1996. Seed bank dynamics and seed dispersal:
important topics in restoration ecology. Acta bot. Neerl. 45:
46t490.Bekker, R. M., J. H. J. Schaminee, J. P. Bakker & K. Thompson,
1998. Seed bank characteristics of Dutch Plant communities.
Acta bot. neerl.47: 15-26.Biewer, H., 1997. Regeneration aÍenreicher Feuchtwiesen. Veröff.
PAÓ (Karlsruhe) 24: ll-323.Biewer, H. & P. Poschlod, 1997. Wiedervemàssung und Wieder-
herstellung artenreicher Feuchtwiesen im geplannten Naturs-
chutzgebiet Stidliches Federseeried. Veröff. PAÖ 1Karlsruhe) 12:
25t-269.Boeye, D. & R. Verheyen, 1994. The relation between vegetation
and soil chemistry gradients in a groundwater discharge fen. J.
Veg. Sci. 5: 553-560.Boeye, D. & B. Verhagen, 1997. Plaggen in een beekbegeleidend
laagveen. De Levende Natuur 98: 273-211 (with English sum-
mary).Burny, J., 1999. Bijdrage tot de historische ecologie van de
Limburgse Kempen (1910-1950). Natuurhistorisch genootschap
Limburg.Danvid, M. & C. Nilsson, 1997. Seed floating ability and distribu-
tion of alpine plants along a northern Schwedish river. J. Veg.
Sci.. 8: 2'7 l-27 6.
De Graaf, M. C. C., P. J. M. Verbeek, R. Bobbink & J. G. M.Roelofs, 1998b. Restoration of species-rich dry heaths: the im-portance of appropriate soil conditions. Acta bot. neerl. 4'7:
89-tt2.

168
De Mars, H., M. J. Wassen & W. H. M. Peeters, 1996. Theeffect of drainage and management on peat chemistry and nutri-ent deficiency in the former Jegrzenia floodplain (N.E. Poland).Vegetatio 126:59J2.
Dijk, E. & N. D. Eck, 1995. Effects of mycorrhizal fungi on in vitronitrogen response of some Dutch indigenous orchid species. Can.
J. Bot.73: 1203-1211.Egloff, T. B., 1983. Der Phosphor als primair limitierender Niihr-
stoff in Streuwiesen (Molinion); Diingungsexperiment im un-teren Reustal. Ber. Geobot. Inst. ETH Stiftung Riibel, Zirich 59:I 19-148.
Ellenberg, H., 1978. Vegetation Mitteleuropas mit den Alpen.Ulmer. Everts, F. H. & N. P. J. De Vries, 1991. The vegetationdevelopment in stream valley systems; a landscape ecologicalstudy of Drenthian stÍeam valleys. Ph.D. thesis, Univ. of Gronin-gen, Historische Uitgeverij Groningen (in Dutch with Englishsummary).
Faulkner, S. P. & C. J. Richardson, 1989. Physical and chemicalcharacteristics of freshwater wetland soils. In Hammer, D. A.(ed.), Constructed Wetlands foÍ Wastewater Treatment. Lewis,Publ., Chelsea, Michigan: 4lJ 2.
Garcia, A., 1992. Conserving the species-rich meadows of Europe.
Agric. Ecosyst. Envir. 40: 219-232.Gough, R. H. & R. H. Marss, 1990. A comparison of soil fertility
between semi-natural and agricultural plant communities: im-plications for the creation of species-rich grassland on abandoned
agricultural land. Biol. Cons. 5l : 83-96.Grant, V., 1971. Plant Speciation, Columbia Univ. Press.
Grime, J. P.,1979. Plant Strategies and Vegetation Processes. Wiley,Chichester.
Grootjans, A. P., 1980. Distribution of plant communities alongrivulets in relation to hydrology and management. In Wilmanns,O. & R. Tiixen (eds), Epharmonie, Berichte der Int. Symp. der IV.F.V. Cramer Verlag, Vaduz: 143-165.
Groodans, A. P. & R. van Diggelen, 1995. Assessing the restora-
tion prospects of degraded fens. In Wheeler, B. D., S. C. Shaw,
W. J. Fo)'t & R. A. Robertson (eds), Restoration of Temperate
Wetlands. Wiley, Chichester: 73-90.Grootjans, A. P., P. C. Schipper & H. J. van der Windt, 1985. In-
fluence of drainage on N-mineralisation and vegetation responsein wet meadows.l: Calthion palustris stands. Oecol. Plant. 6:403417.
Grootjans, A. P., P. C. Schipper & H. J. van der Windt, 1986. In-fluence of drainage on N-mineralisation and vegetation response
in wet meadows.fÍ: Cirsio-Molinietum stands. Oecol. Plant. 7:3-t4.
Grootjans, A. P., G. J. R. Allersma & C. Kik, 1987. Hybridisationof the habitat in disturbed hay meadows. In van Andel, J., J.
P. Bakker & R. W. Snaydon (eds), Disturbance in Grasslands.Geobotany l0:.67-:77.
Grootjans, A. P., L. F. M. Fresco, C. C. de Leeuw & P. C. Schipper,1996. Degeneration of species-rich Calthion palusrns hay mead-ows; some considerations on the community concept. J. Veg. Sci.7:185-194.
Grootjans, A. P., W. Bijkerk, F. H. Everts, M. Jongman, M. Sa-
lomons & M. E. Tolman, 1997. Monitoring van effectgerichtemaatregelen tegen verzuring 1994-1996. Lab. Plant Ecology,Haren/Bureau Everts & De Vries, Groningen.
Grootjans, A. P., R. van Diggelen, F. H. Everts, P. C. Schipper,J. Streefkerk, N. P. J. De Vries & A. Wierda, 1993. Linkingecological patterns to hydrological conditions on various spatialscales, a case study of small valleys. In Vos, C. C. & P. Opdam(eds), Landscape Ecology of a Stressed Environment. Chapmanand Hall, London: 60-76.
Giisewell, S., 1997. Evaluation and management of fen meadows
invaded by common reed (Phragmites australis). Ph.D. thesis,
ETH Ziirich.Hobbs, R. J. & D. A. Norton, 1996. Toward a conceptual framework
foÍ restoration ecology. Rest. Ecol. 4:93-110.Jansen, A. J. M., 2000. Hydrology and restoration of wet heathland
and fen meadow communities. Ph.D. thesis, Univ. of Groningen.Jansen, A. J. M. & J. G. M. Roelofs, 1996. Restoration of Cirsio-
Molinietum wet meadows by sod cutting. Ecol. Engen. T: 279-298.
Jansen, A. J. M., M. C. C. de Graaf & J. G. M. Roelofs, 1996.
The Íestoration of species-rich heath land communities in the
Netherlands. Vegetatio 126: 73-88.Jansen, A. J. M., A. P. Groodans & M. H. Jailing, 2000. Hydrology
of Dutch Cirsio-Molinietun meadows: Prospects of Íestoration.Appl. Veg. Sci. 3: 51-64.
Jansen, A. J. M., A. Th. W. Eysink & C. Maas, 2001. Hydrologicalprocesses in a Cirsio-Molínietum weÍ meadow: implications forrestoration. Ecol. Engin. 11 : 3347.
Janssen, M. P. J. M. & A. H. P M. Salman, 1995. A national strategy
for dune conservation in the Netherlands. In Salman A. H. P. M.,H. Berends & M. Bonazountas (eds), Coastal Management and
Habitat Conservation. EUCC, Leiden: 215-230Kaiser, T., 1995. Vegetationskundliche Untersuchungen auf re-
liefiertem Niedermoor bei Extensivweide. Zeitschrift fiir Kultur-technik und Landentwicklung 36: 175-177.
Kapfer, A., 1988. Versuche zur Renaturierung gediingten Feucht-griinlands - Aushagerung und Vegetationsentwicklung. Diss.Bot.120: l-144.
Kemmers, R. H., P. C. Jansen & P Mekkink, 1995. Humus formprofiles and ecohydrological system analysis. In Schouten et
al. (eds), Scenario Studies for the Rural Environment. KluwerAcademic Publishers, Dordrecht: 489494.
Kemmers, R. H., P. C. Jansen & S. P. J. Van Delft, 2000. De regulatievan de basentoestand in kwelafhankelijke schraalgraslanden en
laagvenen. Report Ministry of Agriculture, Nature and Fisher-ies/Alterra, Wageningen.
Klinka, K., R. N. Green, R. N. Trowbridge & L. E. Lowe,1981. Taxonomic classiflcation of humus forms in ecosystems
of British Columbia, Ministry of Forests, U.S.A.Klötzli, F., 1991. Möglichkeiten und erste Ergebnisse mitteleur-
opàischer Renaturierungen. Verh. Ges. Ökol. 20: 229-242.Koerselman, W., S. A. Bakker & M. Blom, 1990. Nitrogen,
phosphorus and potassium mass balances for two small fens
surrounded by heavily fertilised pastures. J. Ecol. 70: 428442.Koerselman, W. & J. T. A Verhoeven, 1995. Eutrophication of fen
ecosystems: external and internal nutrient sources and restora-
tion strategies. In Wheeler, B. D., S. C. Shaw, W. J. Foyt & R.
A. Robertson (eds), Restoration of Temperate Wetlands. Wiley,Chichester: 91-l 12.
Lamers, L. P. M., H. B. M. Tomassen & J. G. M. Roelofs, 1998.
Sulphate-induced eutrophication and phytotoxicity in freshwaterwetlands. Envir Sci. Technol. 32:199-205.
Landolt, E. & F. Grossman, 1968. Zur vermutlich hybridogenenEntstehung von Wiesenpflanzen des schweizerischen Mittel-landes. Verh. Schweiz. Naturf. Ges. 1968: 114-117.
Louwe Kooijmans, L. P., 1974. The Rhine/lVleuse delta. Ph.D.Thesis, Univ. of Leiden.
Mengel, K. & E. A. Kirkby, 1982. Principles of Plant Nutrition, 3
ed. International Potas Institute, Bern, Switserland.Meuleman, A. F. M., 1999. Performance of treatment wetlands.
Ph.D. Thesis, Univ. of Utrecht.Mulder, J., N. van Breemen & H. C. Eiick, 1989. Depletion ol
soil aluminium by acid deposition and implications for acidneutÍalisation. Nature 337: 247 -249.

Muller, S., T. Dutoit, D. Alard & F. Grevilliot, 1998. Restoration and
rehabilitation of species-rich grassland ecosystems in France; a
review Rest. Ecol. 6: 94-101.Olde Venterink, H. 2000. Nitrogen, phosphorus and potassium flows
controlling plant productivity and species richness; eutrophica-
tion and nature management in fens and meadows. PhD Thesis,
Uffecht UniveÍsity.Oltr, H. & J. P. Bakker, 1991. Long term dynamics of standing crop,
vegetation composition and species richness after cessation offertiliser application to hay-fields. J. Appl. Ecol.28: 104G-1052.
Olff, H., D. M. Pegtel, J. Van Groenendael & J. P. Bakker, 1994.
Germination strategies during grassland succession. J. Ecol. 82:
69-17.Oomes, M. J. M., H. Oltr& H. J. Altena, 1996. Effects of vegetation
management and raising the water table on nutrient dynamics
and vegetation change in a wet grassland. J. Appl. Ecol. 33: 516-588.
Oomes, M. J. M., P. J. Kuikman & F. H. H. Jakobs, 1997. Nitrogen
availability and uptake in mesocosms at two water levels and twowater qualities. Plant Soil L92:249-259.
Parker, V. T. & S. T. A. Pickett, 1997. Restoration as an ecosys-
tem process: implications ofthe modem ecological paradigm. InUrbanska, K. M., N, R. Webb & P. J. Edwards (eds), Restoration
Ecology and Sustainable Development. Cambridge Univ. Press:
17-32.Patrick, W. H. Jr. & R. A. Khalid, 1974. Phosphate release and
sorption by soils and sediments: effect of aeÍobic and anaerobic
conditions. Science 186: 53-55.Pegtel, D. M., 1983. Ecological aspects of nutrient deÍicient wet
grassland (Cirsio-Molinietun). Verhandl. Ges. Okol I0: 217-228.
Pegtel, D. M., J. P. Bakker, G. L. Verwey & L. F. M. Fresco, 1996.
N, K and P deficiency in chronosequential cut summer-dry grass-
lands on gley podsol after cessation of fertiliser application. Plant
Soil 178: l2l-131.Pfadenhauer, J. & A. P. Grootjans, 1999. Wetland restoration in
Central Europe: aims and methods. Appl. Veg. Sci. 2: 95-106.
Poschlod, P., J. P Bakker, S. Bonn & S. Fischer, 1996. Dispersal ofplants in fragmented landscapes; changes of dispersal problems
in the actual and historical man-made landscape. In Settle, J.,
C. R. Margules, P Poschlod & K. Henley (eds), Species Sur-
vival in Fragmented Landscapes. Kluwer Academic Publishers,
Dordrecht: 123-127.Poschlod, P. & S. Bonn, 1998. Changing dispersal processes in the
central European landscape since the last ice age: an explana-
tion for the actual decrease of plant species richness in differenthabitats? Acta bot. Neei. 47:2744.
Runhaar, 1., 1999. Impact of hydrological changes on nature con-
servation areas in the Netherlands. Ph.D. thesis, Univ. ofLeiden.Runhaar, J., C. R. Van Gool & C. L. G. Groen, 1996. Impact
of hydrological changes on nature conservation areas in the
Netherlands. Biol. Cons. 7 6: 269-27 6.
Putman, R. J., A. D. Fowler & S. Tout, 1991. Patterns of use ofancient grassland by cattle and horse and effects on vegetationalcomposition and stÍucture. Biol. Cons. 56: 329-347.
Schaminée, J. H. J., A. H. F StorteldeÍ & E. J. Weeda, 1996. Devegetatie van Nederland part 3. Opulus Press, Uppsala,/Leiden.
StrUkstra, R. J., G. L. Verweij & J. P. Bakker, 1997. Seed dispersal
by mowing machinery in a Dutch brook valley system. Acta bot.Neerl.46: 307401.
Tallis, J. H., 1991. Plant Community History. Chapman & Hall.Tallowin, J. R. B. & R. E. N. Smith, 1996. Management op-
tions to conserve a Cirsio-Molinietum and integrate its use intoproductive livestock systems. Asp. appl. Biol. 44: 203-210.
t69
Ter Borg, S. J.,1972. Variability of Rhinanthus serotinus in relation
to the environment. Ph.D. thesis, Univ. of Groningen.
Thompson, K., J. P. Bakker & R. M. Bekker, 1997. The Soil
Seed Banks of North West Europe: Methodology, Density and
Longevity. Cambridge Univ. Press.
Tirrner, S. D. & C. F. Friese, 1998. PlanrMycorrhizal community
dynamics associated with a moisture gradient within a rehabilit-
ated prairie fen. Rest. Ecol. 6: 44-51.Van Andel, M., T. van Andel & T. Waterbolk, 1945. Bodem &
plantengroei in het dal van het Anderense Diep. Kruipnieuws 7:
1-48.Van Breemen, N., 1987. Effects ofredox processes on soil acidity.
Neth. J. agric. §ci,.35,2'75-279.Van der Hoek, D. & R. H. Kemmers, 1998. The effectiveness of
rewetting. The impact of rewetting measures on key processes
in the topsoil of the nature restoration project 'De Veenkampen'.
Landschap 15:. 2ll-224 (in Dutch with English summary).
Van der Linden, M. et aI. 1996. Herstel van natte en vochtige
ecosystemen. Report 9-1, National Research Programme on
Desiccation (in Dutch, with English summary)'
Van Diggelen, R., 1998. Moving gradients: assessing restoration
prospects of degraded brook valleys. Ph.D. thesis, Univ. ofGroningen.
Van Diggelen, R., A. P Grootjans & R. Burkunk, 1994. Assessing
restoration perspectives of disturbed brook valleys: the Gorecht
area, the Netherlands. Rest. Ecol. 2:87-96.Van Diggelen, R., J. P. Bakker & J. Klooker, 1997. Topsoil re-
moval: new hope for threatened plant species? In Cooper, A.& J. Power (eds), Species Dispersal and Land Use Processes.
Proceedings Sixth Annual IALE (U.K.) Conference, Coleraine,
9-1 lth September 1997:257-263.Van Duren, I. C. & D. M. Pegtel, 2000. Nutrient limitation in wet,
drained and rewetted fen meadows: Evaluation of methods and
results. Plant So1l220: 3547 .
Van Duren, I., D. Boeye & A. P. Grootjans, 1997a. Nutrientlimitation in an extant and drained poor fen: implications forrestoration. Plant Ecol. 133: 9l-100.
Van Duren, I. C., D. M. Pegtel, B. A. Aerts & J. A. Inberg, 1997b.
Nutrient supply in undrained and drained Calthion meadows. J.
Veg. Sci 8: 829-838.Van Duren, I. C., R. J. Strijkstra, A. P Grootjans, G. N. J. Ter Heerdt
& D. M. Pegtel, 1998. A multidisciplinary evaluation of restor-
ation measures in a degraded fen meadow (Cirsio-Molinietum).
Appl. Veg. Sci. 1: 115-130.Van Wirdum, G., 1991. Vegetation and hydrology of floating
rich-fens. Ph.D. thesis, University of Amsterdam. Datawyse,
Maastricht.Wassen, M. J., R. van Diggelen, J. T. Verhoeven & L. Wolejko,
1996. A comparison of fens in natural and artificial landscapes.
Vegetatio 126:5-26.Weeda, E. J., R. van der Meijden & P. A. Bakker, 1990. Rode Lijst
van de in Nederland verdwenen en bedreigde planten over de
periode I 980-1990. Gorteria 16: 2-26.Young, P., 1996. The 'new' science of wetland restoration. Envir.
Sci. Technol. 30: 292A-2964.Zandee, M., 1981. Contribution to the biosystematics of two cy-
anogenic polyploid complexes (Juncus articulatus and Holcus
moLlus).Ph.D. thesis, Univ. of Leiden.Zeeman, W P. C., 1986. Application in land, natuÍe and water
management; the Reitma case study. Proc. & Inf. CHO-TNO 34:
|7-126.
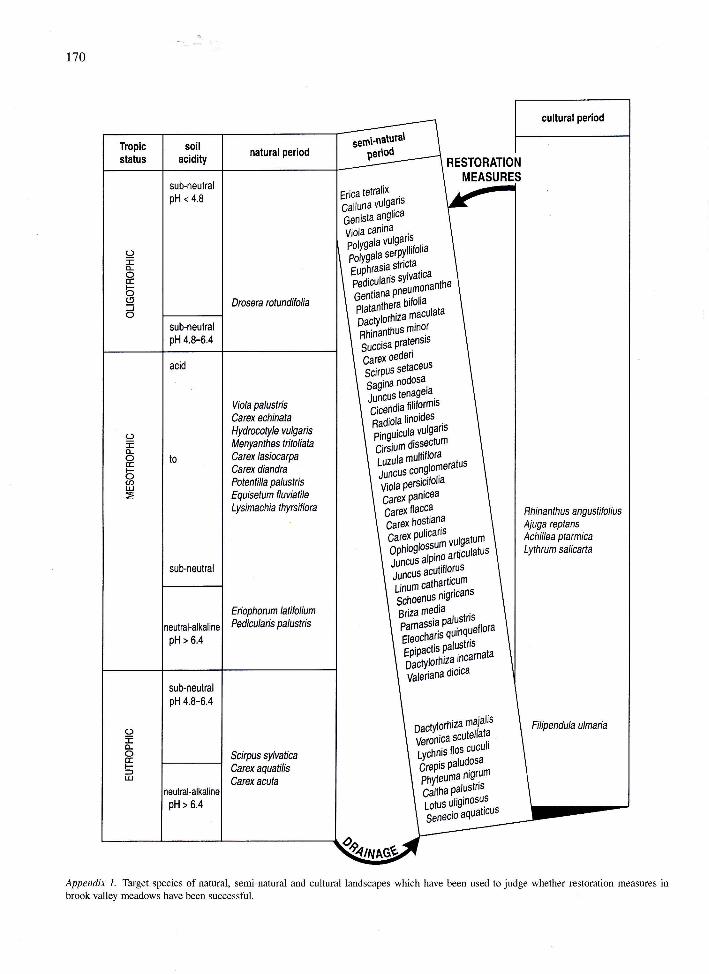
t'70
cultural period
semi'naturalPeriotl -
Tropicstatus
soilacidity
natural period
Rhi n anth us ang ustiÍol iu s
Ajuga reptans
Achillea ptarmica
Lythrum salicatta
Filipendula ulmaria
RESTORATIOMEASURE
oto-oTEFo(,Jo
sub-neutralpH < 4.8
Drosera rotundifolia
T*l,i:N::
,Yffi7;;ii,,,,'^t',HÍilil'í"i;
"i:xií:lïiï:',ï',ijÏilfí#;[srt.it, Pt't'n
\?:fi,#l'
\',;,ï1i,1;*\ Radiola li'o
\ Pinguicula t
\ cirsium dts
\ Luzula mu
\ Junt" to\ vida pets
\ cr.,uP'\ carexíla\ carex ht
\ carex P
\ oPniog\ Juncus
\ Jrnt"\ Linum
\ stho'\ aiza\ Pa,n
\ ato\ rpir\ oar\ vat
a,
ica
onanthe
ia
oulatar0I
sis
?us I
u\enlrrmis \des \dgaris \sedum \ilflora \rglomeratus \icilolia \icea \cca \)stiana I
ifïf;lll:urr,\: acutiÍlorus I
calharlicunrnus nigricans
media
ilïÍJílïïffi*,,
"?lli,!,1',xï^i,^^,^iianadioica
,*?l'#i;i:i:
d;##ÏÍ:íj,t.
Lt'fí.{xï;ïj,:,i;,,
sub-neutralpH 4.8-6.4
(J
=o-oCEl-c)U)uJ
=
acid
to
sub-neutral
Viola palustris
Carex echinata
Hydrocotyle vulgaris
Menyanthes triÍoliata
Carex lasiocarpa
Carex diandra
Potentiila palustris
Equisetun fluviatile
Ly si m a c h i a thy r s it I o ra
Eriophorun latiÍoliunPedicularis palustris
pH > 6.4
o=ÍLoÉ.FfLrJ
sub-neutralpH 4.8-6.4
Scirpus sylvatica
Carex aquatilis
Carex acutara, rtral.allzalina
pH > 6.4
Appendix 7. Target species of natural, semi-natural and cultural landscapes which have been used to judge whether restoration measures inbrook valley meadows have been successful.
Top Related
Copyright © 2022 FDOKUMEN

