







Bahasa
Halaman
Hukum


http://het.sagepub.com/Human & Experimental Toxicology
http://het.sagepub.com/content/30/8/860The online version of this article can be found at:
DOI: 10.1177/0960327110382130
2011 30: 860 originally published online 27 August 2010Hum Exp ToxicolMA Siddiqui, MP Kashyap, AA Al-Khedhairy, J. Musarrat, VK Khanna, S. Yadav and AB Pant
in PC12 cells-estradiol against co-exposure of 4-hydroxynonenal and 6-hydroxydopamineβProtective potential of 17
Published by:
http://www.sagepublications.com
can be found at:Human & Experimental ToxicologyAdditional services and information for
http://het.sagepub.com/cgi/alertsEmail Alerts:
http://het.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://het.sagepub.com/content/30/8/860.refs.htmlCitations:
What is This?
- Aug 27, 2010 OnlineFirst Version of Record
- Jul 19, 2011Version of Record >>
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

Protective potential of 17β-estradiolagainst co-exposure of 4-hydroxynonenaland 6-hydroxydopamine in PC12 cells
MA Siddiqui1, MP Kashyap2, AA Al-Khedhairy1,J Musarrat1, VK Khanna2, S Yadav2 and AB Pant2
Abstract4-hydroxynonenal (4-HNE) and 6-hydroxydopamine (6-OHDA)-mediated damage in dopaminergic neurons iswell documented. Protective potential of steroidal hormone (17b-estradiol) has also been suggested.However, therapeutic potential of such promising hormone is hampered due to complex brain anatomy andphysiology. Thus, the present investigations were studied to suggest the applicability of dopamine expressingPC12 cells as in vitro tool to screen the pharmacological potential of 17b-estradiol against 4-HNE and6-OHDA. MTT assay was conducted for cytotoxicity assessment of both 4-HNE (1 mM to 50 mM) and6-OHDA (10�4 to 10�7 M). Non-cytotoxic concentrations, that is, 4-HNE (1 mM) and 6-OHDA (10�6 M) wereselected to study the synergetic/additive responses. PC12 cells were found to be more vulnerable towardsco-exposure of individual exposure of 4-HNE and 6-OHDA, even at non-cytotoxic concentrations. Then, cellswere subjected to pre-treatment (24 hours) of 17b-estradiol (1 mM), followed by a permutation of combina-tions of both 4-HNE and 6-OHDA. Pretreatment of 17b-estradiol was found to be significantly effective againstthe cytotoxic responses of 4-HNE and 6-OHDA, when the damage was at lower level. However, 17b-estradiolwas found to be ineffective against higher concentrations. Physiological-specific responses of PC12 cells against4-HNE/6-OHDA and 17b-estradiol suggest its applicability as first tier of screening tool.
KeywordsPC12 cells, 4-HNE, 6-OHDA, 17b-estradiol, cytotoxicity
Introduction
Lipid peroxidation is well documented as one among
the key factors involved in the etiology of various
neurodegenerative disorders including stroke,1
Alzheimer’s disease,2 and Parkinson’s disease.3 The
higher concentrations of 4-hydroxynonenal, an unsa-
turated aldehydic product of o-6 polyunsaturated
fatty acids, produced during toxic insults is known
to cause neurodegeneration and neurotoxicity in
variety of cells.4-7 The role of 6-hydroxydopamine
(6-OHDA), a hydroxylated form of dopamine, in the
pathogenesis of Parkinson’s disease is well estab-
lished.8,9 It is a widely used chemical to investigate
the neurotoxic effects on dopaminergic neurons and
usually thought to cross cell membrane through dopa-
mine uptake transporters to inhibit mitochondrial
respiration and to generate intracellular reactive
oxygen species.10-12
The neuroprotective potential of estrogens, primarily
known as female sex hormones, is well documented.13
Early estrogen therapy has been suggested as one
among the strategy to reduce the risk of neurodegenera-
tive disorders such as Alzheimer’s disease,14,15
ischemic stroke,16 and Parkinson’s disease.17 Neuropro-
tective role of estrogens has been reported in a number
of experimental models of acute cerebral ischemia.18
1 Department of Zoology, College of Science, King SaudUniversity, Riyadh, Saudi Arabia2 In Vitro Toxicology Laboratory, Indian Institute of ToxicologyResearch, Lucknow, India
Corresponding author:A B Pant, In Vitro Toxicology Laboratory, Indian Institute ofToxicology Research, PO Box: 80, MG Marg, Lucknow-226001,UP, IndiaEmail: [email protected]
Human and Experimental Toxicology30(8) 860–869
ª The Author(s) 2010Reprints and permission:
sagepub.co.uk/journalsPermissions.navDOI: 10.1177/0960327110382130
het.sagepub.com
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

Estrogens-mediated neuroprotection has also been
demonstrated as an age-independent phenomenon in
female rats.19 Variety of neuronal cell types have been
employed to study the neuroprotective potential of
estrogens against number of toxic insults mainly includ-
ing H2O2,20 serum deprivation,21 oxygen-glucose depri-
vation,22 iron,23 amyloid peptide,24 excitotoxicity,25
and mitochondrial toxins such as 3-nitropropionic
acid,26 sodium azide,27 etc. 17b-Estradiol, a weak natu-
ral estrogen, has been recently evaluated for its protec-
tive potential through mitochondrial function against
ROS and H2O2 insult in cultured neuronal cell line.20,28
Thus, the present investigations were aimed to
study the protective potential of 17b-estradiol in
PC12 cells receiving experimental co-exposure of two
known neurotoxicants viz., 6-OHDA and 4-HNE,
which may be useful to understand the vulnerability
of brain cells in Parkinson’s patients.
Materials and methods
PC12 cells (a rat pheochromocytoma cell line) were
originally procured from National Centre for Cell
Sciences, Pune, India, and have been maintained at
In Vitro Toxicology Laboratory, Indian Institute of
Toxicology Research, Lucknow, India, as per the
standard protocol provided by the supplier. Briefly,
cells were cultured in Nutrient Mixture F-12 (Ham),
supplemented with 2.5% fetal bovine serum (FBS),
15% horse serum (HS), 0.2% sodium bicarbonate
and antibiotic, and antimycotic solution (100�,
1 mL/100 mL of medium, Invitrogen, Life Technol-
ogies, Carlsbad, CA, USA). Cells were grown in 5%CO2 – 95% atmosphere in high humidity at 37�C.
Each batch of cells was assessed for physiological
characteristics using established markers7,29 and cell
viability by trypan blue dye exclusion test30 prior to
experiments, and batches showing more than 95%cell viability were used in the present study. Cells
of passage number between 18 and 25 were used
in the present study.
Nutrient mixture F-12 Hams culture medium, anti-
biotics, fetal bovine, and horse serum were purchased
from Gibco BRL, Grand Island, NY, USA. Culture
wares and other plastic consumables were procured
from Nunc, Denmark. 4-hydroxy-trans-2-nonenal (4-
HNE) was generously gifted by Dr Sanjay Srivastava,
Associate Professor, Department of Cardiology,
University of Louisville, KY. All other specified che-
micals and reagents were purchased from Sigma
Chemical Company Pvt Ltd St Louis, Missouri, USA.
PC12 cells were exposed to various concentrations
of either of 4-HNE (1 mM to 50 mM) or 6-OHDA
(10�4 to 10�7 M) for a period up to 24 hours. To study
the prophylactic protection, cells were pretreated for
24 hours with 17b-estradiol at physiological concen-
tration (1 mM), then subjected to receive a permuta-
tion combination of both 4-HNE (1 mM to 50 mM)
and 6-OHDA (10�4 to 10�7 M) for 24 hours. Percent-
age loss in cell viability was assessed by MTT assay.
Percentage cell viability was assessed using the
3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazo-
lium bromide (MTT) assay as described.7 Briefly,
cells (1 � 104) were allowed to adhere for 24 hours
under high humid environment in 5% CO2 at 37�Cin poly-L-lysine�coated 96-well culture plates. The
medium was then replaced with the medium contain-
ing either of 4-HNE (1 mM to 50 mM) or 6-OHDA
(10�4 to 10�7) or combination of both 4-HNE and
6-OHDA, for a period up to 24 hours. After the
respective exposure, MTT (5 mg/mL of stock in PBS)
was added (10 mL/well in 100 mL of cell suspension),
and plates were incubated for 4 hours. At the end of
incubation period, the reaction mixture was carefully
taken out and 200 mL of dimethyl sulfoxide (DMSO)
was added to each well by pippeting up and down sev-
eral times until the content gets homogenized. The
plates were kept on rocker shaker for 10 min at room
temperature and then read at 550 nm using Multiwell
Microplate Reader (Synergy HT, USA and Bio-Tek,
Bio-Tek Instruments, Winooski, VT, USA). Untreated
sets were also run under identical conditions and
served as basal control.
Statistical analysis
The results are expressed as mean and standard
error of means (SEM) for six wells of at least three
experiments, as indicated in the figures. One-way
ANOVA and Student’s t test were employed to
detect differences between the groups of treated
and control. p < 0.05 was taken to indicate signif-
icant differences.
Results
Data on cytotoxic responses of 4-HNE and/or 6-OHDA
and prophylactic protection of 17b-estradiol in cul-
tured PC12 cells are summarized in Figures 1�8.
A concentration-dependent decrease in percentage
cell viability was recorded following experimental
exposure of 4-HNE in PC12 cells. Maximum reduc-
tion in mitochondrial activity, that is, 50% + 2.7%
Siddiqui MA et al. 861
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from
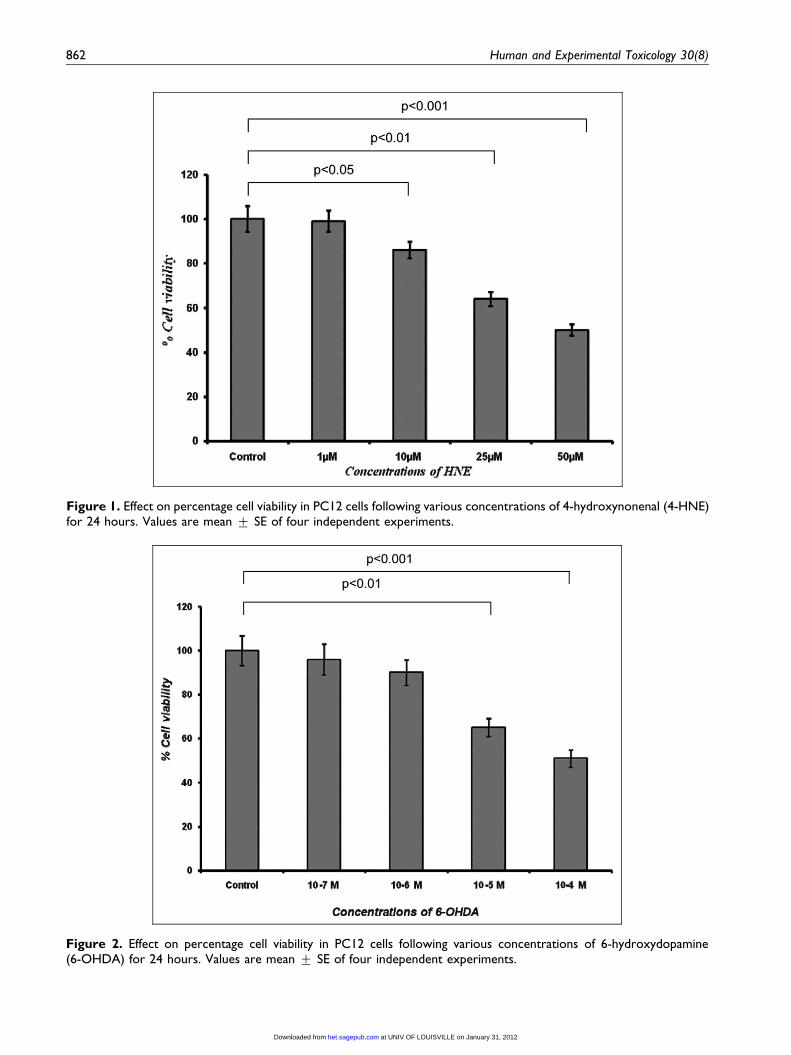
Figure 1. Effect on percentage cell viability in PC12 cells following various concentrations of 4-hydroxynonenal (4-HNE)for 24 hours. Values are mean + SE of four independent experiments.
Figure 2. Effect on percentage cell viability in PC12 cells following various concentrations of 6-hydroxydopamine(6-OHDA) for 24 hours. Values are mean + SE of four independent experiments.
862 Human and Experimental Toxicology 30(8)
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

of control was recorded with 50 mM of 4-HNE expo-
sure, followed by 25 mM (36% + 2.4%) and 10 mM
(16% + 0.7%), respectively. In general, the effect of
1 mM HNE was found to be insignificant on mito-
chondrial activity. Exposure (24 hours) of 4-HNE at
25 and 50 mM was found to be cytotoxic, whereas,
0
20
40
60
80
100
120
Control 1μM HNE 17β-Estradiol +1μM HNE
1μM HNE + 10-6 M 6-OHDA
17β-Estradiol +1μM HNE + 10-
6 M 6-OHDA
Concentrations
% C
ell v
iabi
lity
p < 0.001
p < 0.01
Figure 3. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 1 mM of4-hydroxynonenal (4-HNE) and 10�6 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments.
Figure 4. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 10 mM of4-hydroxynonenal (4-HNE) and 10�6 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments.
Siddiqui MA et al. 863
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from
10 mM could be considered as cytostatic and 1 mM
non-cytotoxic (Figure 1). 6-OHDA was able to pose
statistically significant reduction in mitochondrial
activity at 10�4 and 10�5 M concentrations. Whereas,
lower concentrations of 6-OHDA (10�6 and 10�7 M)
used in the study could not cause significant detri-
mental effects in PC12 cells, thus considered as
non-cytotoxic concentrations (Figure 2).
Data on the protective potential of 17b-estradiol fol-
lowing experimental co-exposure of 4-HNE (1 mM)
and 6-OHDA (10�6 M) are summarized in Figure 3.
Statistically highly significant (p < 0.001), that is,
30% + 1.9% reduction in the percentage cell viability
was recorded. Interestingly, pretreatment of 17b-
estradiol (1 mM) was found to be significantly effec-
tive to protect the cells against the co-exposure of
4-HNE and 6-OHDA (Figure 3).
Although, the protective response of 17b-estradiol
was statistically significant in the cells co-exposed
to 4-HNE (10 mM) and 6-OHDA (10�6 M), the mag-
nitude was less pronounced when compared with the
cells exposed to non-cytotoxic concentrations of both
4-HNE and 6-OHDA (Figures 3 and 4). Severity of
damage got more significant and prominent with the
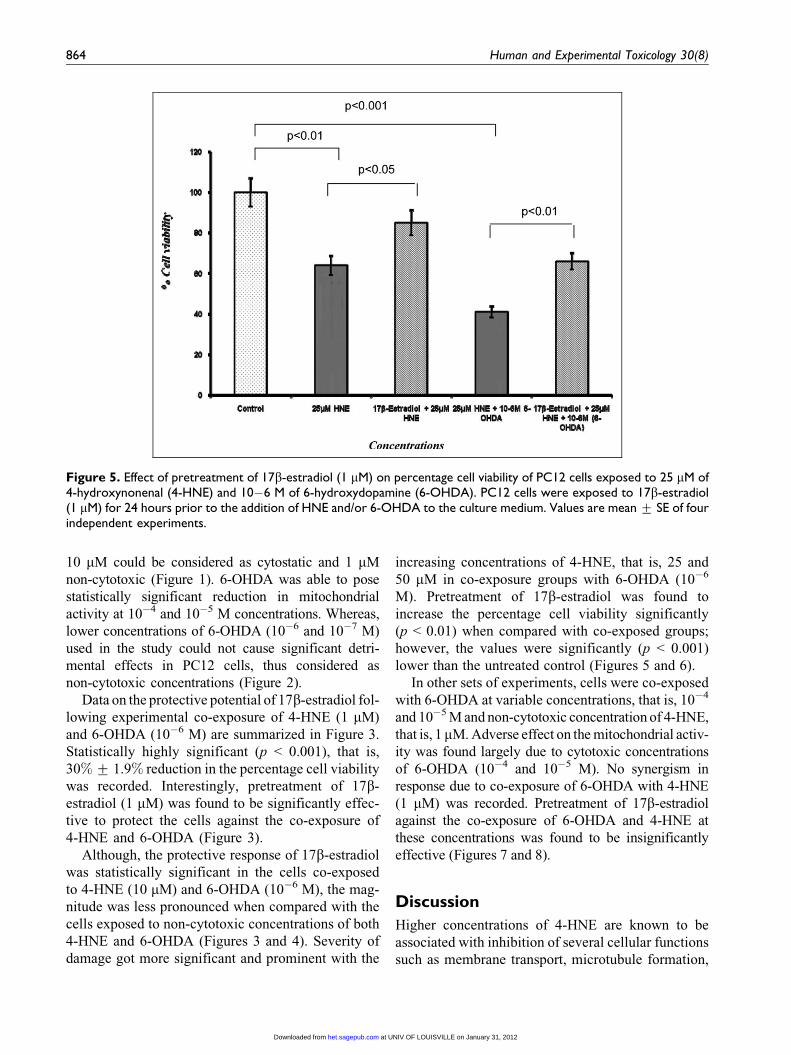
increasing concentrations of 4-HNE, that is, 25 and
50 mM in co-exposure groups with 6-OHDA (10�6
M). Pretreatment of 17b-estradiol was found to
increase the percentage cell viability significantly
(p < 0.01) when compared with co-exposed groups;
however, the values were significantly (p < 0.001)
lower than the untreated control (Figures 5 and 6).
In other sets of experiments, cells were co-exposed
with 6-OHDA at variable concentrations, that is, 10�4
and 10�5 M and non-cytotoxic concentration of 4-HNE,
that is, 1mM. Adverse effect on the mitochondrial activ-
ity was found largely due to cytotoxic concentrations
of 6-OHDA (10�4 and 10�5 M). No synergism in
response due to co-exposure of 6-OHDA with 4-HNE
(1 mM) was recorded. Pretreatment of 17b-estradiol
against the co-exposure of 6-OHDA and 4-HNE at
these concentrations was found to be insignificantly
effective (Figures 7 and 8).
Discussion
Higher concentrations of 4-HNE are known to be
associated with inhibition of several cellular functions
such as membrane transport, microtubule formation,
Figure 5. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 25 mM of4-hydroxynonenal (4-HNE) and 10�6 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments.
864 Human and Experimental Toxicology 30(8)
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from
0
20
40
60
80
100
120
Control 50 μM HNE 17 β-Estradiol +50μM HNE
50μM HNE + 10-6 M 6-OHDA
17β-Estradiol + 50μM HNE + 10-
6 M 6-OHDA
% C
ell v
iabi
lity
Concentrations
P < 0.01
P < 0.01
P < 0.05
P < 0.05
Figure 6. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 50 mM of4-hydroxynonenal (4-HNE) and 10�6 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments.
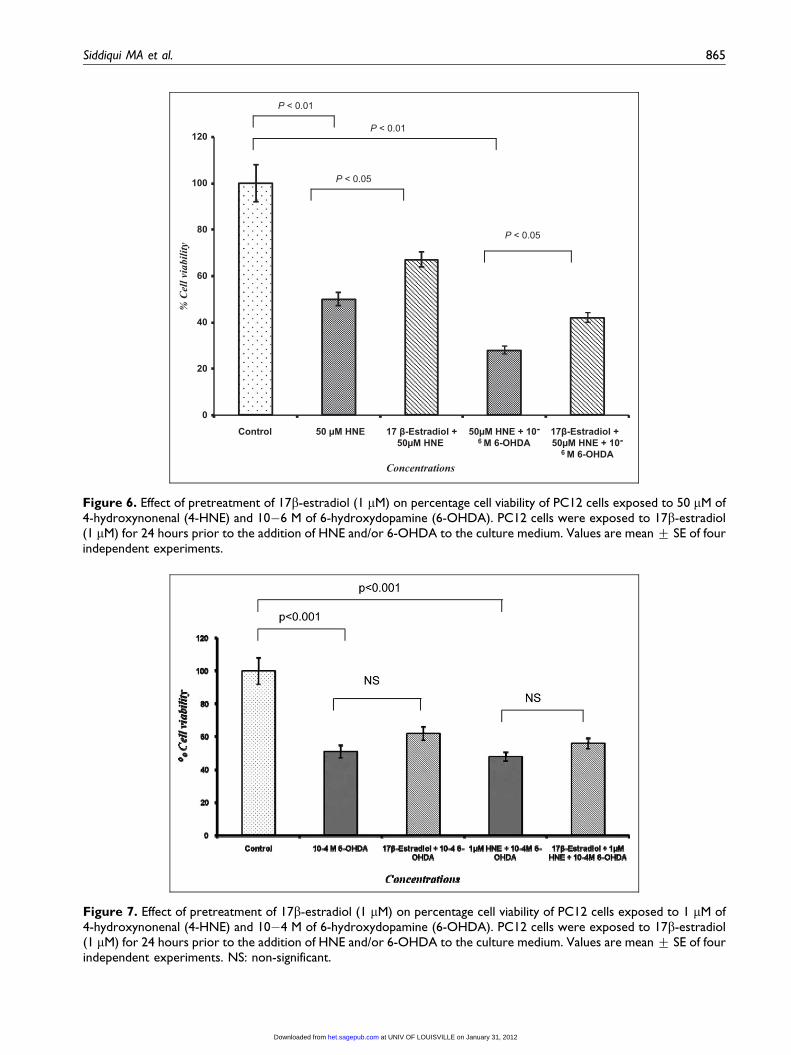
Figure 7. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 1 mM of4-hydroxynonenal (4-HNE) and 10�4 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments. NS: non-significant.
Siddiqui MA et al. 865
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

and mitochondrial respiration.6,7 Cytotoxic responses
of 4-HNE have been reported in variety of cell sys-
tems including HepG2 cells,30 V79 cells,31 cerebellar
granule neurons,5 and primary cultures of normal
human osteoblasts.32 In the present investigations,
results are consistent with earlier studies and have
shown significant cytotoxic responses of 4-HNE
at 25 and 50 mM concentrations in PC12 cells. The
physiological stress was recorded even at lower
concentration (10 mM) of 4-HNE. Lower concen-
trations have also been reported to exert cytotoxi-
city by inducing signaling pathway for the
production of cytokines linked with cellular injury
and apoptosis.33 Possible involvement of oxidative
stress either by generating reactive oxygen and
nitrogen species or by the stress activated transcrip-
tion factors such as NF-kappa B, AP-1, and P53
has also been reported.34,35 Mitochondrial catabo-
lism involves in MTT assay, as 4-HNE or its meta-
bolites are reactive in nature and may interact
either directly to the proteins36 or oxidative stress
signaling pathways.37 HNE at 1 mM was found to
be ineffective in reducing the percentage cell viabi-
lity in PC12 cells. It might be correlated with the
earlier studies demonstrating the normal physiolo-
gical concentrations of 4-HNE in the plasma and
erythrocytes of human and experimental animals
ranged between 1 and 10 mM.38
Higher concentrations (10�4 and 10�5 M) of
6-OHDA were found be cytotoxic to PC12 cells
under our experimental conditions; whereas, 10�6
M and lower concentration did not cause any sig-
nificant decrease in percentage cell viability. Our
findings are consistent with earlier reports showing
6-OHDA induced mitochondrial abnormalities in
the pathogenesis of experimental model of Parkin-
son’s disease.39 6-OHDA is known to enter through
both dopaminergic and noradrenergic neurons and
induce damages to the catecholaminergic pathways
of both peripheral and central nervous systems.40
Therefore, it is one among the widely accepted
neurotoxicants to develop in vitro and in vivo
experimental models of Parkinson’s disease.10,41
6-OHDA-induced apoptotic cell death, mitochon-
drial dysfunction-mediated release of cytochrome
c, and the activation of caspase 3 have also been
reported in PC12 cells.11,42
Data indicates that 4-HNE (1 mM) has statistically
significant synergistic response to 6-OHDA (10�6 M)
cytotoxicity, even at non-toxic concentrations, where
independent exposure of either of 4-HNE or 6-OHDA
does not cause any significant cytotoxicity in PC12 cells
Figure 8. Effect of pretreatment of 17b-estradiol (1 mM) on percentage cell viability of PC12 cells exposed to 1 mM of4-hydroxynonenal (4-HNE) and 10�5 M of 6-hydroxydopamine (6-OHDA). PC12 cells were exposed to 17b-estradiol(1 mM) for 24 hours prior to the addition of HNE and/or 6-OHDA to the culture medium. Values are mean + SE of fourindependent experiments. NS: non-significant.
866 Human and Experimental Toxicology 30(8)
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

(Figure 3). The magnitude of this synergism was found
to be increased further with increased concentrations of
4-HNE. Thus, suggestive that co-exposure of 4-HNE
and 6-OHDA makes cellular system more vulnerable
under experimental conditions. This might be due to
comparatively faster 4-HNE-induced cellular changes,
which could lead an early physiological impairment in
the cells and this compromised state of cells made them
more vulnerable towards the 6-OHDA exposure. How-
ever, no synergism could be apparent in cells co-
exposed to non-cytotoxic concentration of 4-HNE
(1 mM) and toxic doses of 6-OHDA (10�4 and 10�5
M). As lower concentrations of 4-HNE are reported to
metabolize very fast and converted into non-toxic or
low-toxic metabolites in to short span of time,43,44 thus,
the loss of percentage cell viability was largely due to
6-OHDA when observed at 24 hours of exposure.
A pretreatment of 24 hours of 17b-estradiol at
physiological concentration (1 mM) was found to
be protective against damages induced at lower
concentrations by 4-HNE and 6-OHDA in PC12
cells. Our data are consistent with the earlier stud-
ies showing that 17b-estradiol exposure stabilizes
mitochondrial potential against oxidative stress26
and mutant presenilin-1.45 Mounting evidence sug-
gests that estrogens, acting as mitochondrial ener-
gizers by targeting mitochondrial sites to inhibit
opening of permeability transition pores (20), inhi-
bit the mitochondrial calcium uniport,46 increase
mitochondrial-specific proteins expression,47 cause
recovery of ATP production,20 and up-regulate the
antiapoptotic protein Bcl-2.48-50 Our findings could
also be supported by earlier findings demonstrating
the protective potential of 17b-estradiol in SK-N-SH,
a human neuroblastoma, by reducing lipid peroxida-
tion and 4-HNE production.20
Our data suggest that PC12 cells are vulnerable
to independent exposure of 4-HNE and 6-OHDA at
higher concentrations and co-exposure of both makes
cellular system more vulnerable even at non-cytotoxic
concentrations. Pretreatment of 17b-estradiol at phy-
siological concentration was found to be protective
against lower concentrations of both independent
and co-exposure of 4-HNE and 6-OHDA in PC12
cells.
Acknowledgment
The authors are grateful to Director, Indian Institute of
Toxicology Research, Lucknow, for his keen interest in the
present work. The technical laboratory assistance by Mr.
Rajesh Misra is also acknowledged.
Funding
Financial support for this study was obtained from Council of
Scientific & Industrial Research, New Delhi, India (SIP-08).
References
1. Adibhatla RM, Hatcher JF. Lipid oxidation and perox-
idation in CNS Health and disease: from molecular
mechanisms to therapeutic opportunities. Antiox Redox
Signal 2010; 12: 125–169.
2. Dmitriev LF. The involvement of lipid radical cycles
and the adenine nucleotide translocator in neurodegen-
erative diseases. J Alzheimer’s Dis 2007; 11: 183–190.
3. Liang LP, Huang J, Fulton R, Day BJ, and Patel M. An
orally active catalytic metalloporphyrin protects against
1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine neuro-
toxicity in vivo. J Neurosci 2007; 27: 4326–4333.
4. Piga R, Saito Y, Yoshida Y, and Niki E. Cytotoxic
effects of various stressors on PC12 cells: involvement
of oxidative stress and effect of antioxidants. Neuro-
toxicology 2007; 28: 67–75.
5. Arakawa M, Ishimura A, Arai Y, Kawabe K, Suzuki S,
Ishige K, et al. N-Acetylcysteine and ebselen but not
nifedipine protected cerebellar granule neurons against
4-hydroxynonenal-induced neuronal death. Neurosci
Res 2007; 57: 220–229.
6. Abdul HM, Butterfield DA. Involvement of PI3K/PKG/
ERK1/2 signaling pathways in cortical neurons to trig-
ger protection by co-treatment of acetyl-L-carnitine and
alpha-lipoic acid against HNE-mediated oxidative
stress and neurotoxicity: implications for Alzheimer’s
disease. Free Radic Biol Med 2007; 42: 371–384.
7. Siddiqui MA, Singh G, Kashyap MP, Khanna VK,
Yadav S, Chandra D, et al. Influence of cytotoxic doses
of 4-hydroxynonenal on selected neurotransmitter
receptors in PC-12 cells. Toxicol In Vitro 2008; 22:
1681–1688.
8. Mazzio EA, Reams RR, and Soliman KF. The role of
oxidative stress, impaired glycolysis and mitochon-
drial respiratory redox failure in the cytotoxic effects
of 6-hydroxydopamine in vitro. Brain Res 2004;
1004: 29–44.
9. Inden M, Kitamura Y, Kondo J, Hayashi K, Yana-
gida T, Takata K, et al. Serofendic acid prevents 6-
hydroxydopamine-induced nigral neurodegeneration
and drug-induced rotational asymmetry in hemi-
parkinsonian rats. J Neurochem 2005; 95: 950–961.
10. Blum D, Torch S, Lambeng N, Nissou M, Benabid AL,
Sadoul R, et al. Molecular pathways involved in the
neurotoxicity of 6-OHDA, dopamine and MPTP: con-
tribution to the apoptotic theory in Parkinson’s disease.
Prog Neurobiol 2001; 65: 135–172.
Siddiqui MA et al. 867
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

11. Hanrott K, Gudmunsen L, O’Neill MJ, and Wonnacott S.
6-hydroxydopamine-induced apoptosis is mediated via
extracellular auto-oxidation and caspase 3-dependent
activation of protein kinase C delta. J Biol Chem 2006;
281: 5373–5382.
12. Wang T, Zhang QJ, Liu J, Wu ZH, and Wang S. Firing
activity of locus coeruleus noradrenergic neurons
increases in a rodent model of Parkinsonism. Neurosci
Bull 2009; 25: 15–20.
13. Kulkarni J, de Castella A, Fitzgerald PB, Gurvich CT,
Bailey M, Bartholomeusz C, et al. Estrogen in severe
mental illness: a potential new treatment approach.
Arch Gen Psychiatry 2008; 65: 955–960.
14. Bagger YZ, Tanko LB, Alexandersen P, Qin G, and
Christiansen C. Early postmenopausal hormone ther-
apy may prevent cognitive impairment later in life.
Menopause 2005; 12: 12–17.
15. McClean J, Nunez JL. 17alpha-Estradiol is neuropro-
tective in male and female rats in a model of early
brain injury. Exp Neurol 2008; 210: 41–50.
16. Huppmann S, Romer S, Altmann R, Obladen M, and
Berns M. 17beta-estradiol attenuates hyperoxia-
induced apoptosis in mouse C8-D1A cell line. J Neu-
rosci Res 2008; 86: 3420–3426.
17. Morissette M, Di Paolo T. Effect of estradiol on striatal
dopamine activity of female hemiparkinsonian mon-
keys. J Neurosci Res 2008; 87: 1634–1644.
18. Cheng MY, Sun G, Jin M, Zhao H, Steinberg GK, and
Sapolsky RM. Blocking glucocorticoid and enhan-
cing estrogenic genomic signaling protects against
cerebral ischemia. J Cereb Blood Flow Metab 2009;
29: 130–136.
19. Wise PM, Dubal DB, Wilson ME, Rau SW, Bottner M,
and Rosewell KL. Estradiol is a protective factor in the
adult and aging brain: understanding of mechanisms
derived from in vivo and in vitro studies. Brain Res
Brain Res Rev 2001; 37: 313–319.
20. Wang X, Dykens JA, Perez E, Liu R, Yang S, Covey
DF, et al. Neuroprotective effects of 17beta-estradiol
and nonfeminizing estrogens against H2O2 toxicity in
human neuroblastoma SK-N-SH cells. Mol Pharmacol
2006; 70: 395–404.
21. Saengsirisuwan V, Pongseeda S, Prasannarong M,
Vichaiwong K, and Toskulkao C. Modulation of insu-
lin resistance in ovariectomized rats by endurance
exercise training and estrogen replacement. Metabo-
lism 2009; 58: 38–47.
22. Zhang H, Xie M, Schools GP, Feustel PF, Wang W,
Lei T, et al. Tamoxifen mediated estrogen receptor
activation protects against early impairment of hippo-
campal neuron excitability in an oxygen/glucose
deprivation brain slice ischemia model. Brain Res
2009; 1247:196–211.
23. Zhao Y, Hu I, and Jin W. Transformation of oxidation
products and reduction of estrogenic activity of
17beta-estradiol by a heterogeneous photo-Fenton
reaction. Environ Sci Technol 2008; 42: 5277–5284.
24. Hruska Z, Dohanich GP. The effects of chronic estra-
diol treatment on working memory deficits induced
by combined infusion of beta-amyloid (1–42) and ibo-
tenic acid. Horm Behav 2007; 52: 297–306.
25. Perrella J, Bhavnani BR. Protection of cortical cells by
equine estrogens against glutamate-induced excito-
toxicity is mediated through a calcium independent
mechanism. BMC Neurosci 2005; 6: 34.
26. Wang J, Green PS, and Simpkins JW. Estradiol protects
against ATP depletion, mitochondrial membrane poten-
tial decline and the generation of reactive oxygen species
induced by 3-nitroproprionic acid in SK-N-SH human
neuroblastoma cells. J Neurochem 2001; 77: 804–811.
27. Regan RF, Guo Y. Estrogens attenuate neuronal injury
due to hemoglobin, chemical hypoxia, and excitatory
amino acids in murine cortical cultures. Brain Res
1997; 764: 133–140.
28. Zhang P, Li HF, Tian ZF, Qiu XQ, Wu JX, and Jia ZJ.
Effects of phytoestrogens and 17beta-estradiol on
vasoconstriction elicited by reactive oxygen species.
Pharmazie 2007; 62: 378–381.
29. Galbiati F, Volonte D, Gil O, Zanazzi G, Salzer JL.,
Sargiacomo M, et al. Expression of caveolin-1 and -2
in differentiating PC12 cells and dorsal root ganglion
neurons: caveolin-2 is up-regulated in response to cell
injury. Proc Natl Acad Sci U S A 1998; 95: 10257–
10262.
30. Pant AB, Agarwal AK, Sharma VP, and Seth PK. In
vitro cytotoxicity evaluation of plastic biomedical
devices. Hum Exp Toxicol 2001; 20: 412–417.
31. Li D, Hinshelwood A, Gardner R, McGarvie G, and
Ellis EM. Mouse aldo-keto reductase AKR7A5 pro-
tects V79 cells against 4-hydroxynonenal-induced
apoptosis. Toxicology 2006; 226: 172–180.
32. Borovic S, Cipak A, Meinitzer A, Kejla Z, Perovic D,
Waeg G, et al. Differential sensitivity to
4-hydroxynonenal for normal and malignant mesench-
ymal cells. Redox Rep 2007; 12: 50–54.
33. Shi Q, Vaillancourt F, Cote V, Fahmi H, Lavigne P,
Afif H, et al. Alterations of metabolic activity in
human osteoarthritic osteoblasts by lipid peroxidation
end product 4-hydroxynonenal. Arthritis Res Ther
2006; 8: 159.
34. Chung FL, Pan J, Choudhury S, Roy R, Hu W, and
Tang MS. Formation of trans-4-hydroxy-2-nonenal-
868 Human and Experimental Toxicology 30(8)
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from

and other enal-derived cyclic DNA adducts from
omega-3 and omega-6 polyunsaturated fatty acids and
their roles in DNA repair and human p53 gene muta-
tion. Mutat Res 2005; 531: 25–36.
35. Siddiqui MA, Singh G, Khanna VK, Kashyap MP,
Yadav S, Chandra D, et al. Oxidative stress mediated
cellular responses in 4-hydoxynonenal exposed PC12
cells. J Ecophysiol Occup Health 2007; 7: 97–109.
36. VChoudhary S, Xiao T, Srivastava S, Zhang W, Chan
LL, Vergara LA, et al. Toxicity and detoxification of
lipid-derived aldehydes in cultured retinal pigmented
epithelial cells. Toxicol Appl Pharmacol 2005; 204:
122–134.
37. Dwivedi S, Sharma A, Patrick B, Sharma R, and
Awasthi YC. Role of 4-hydroxynonenal and its meta-
bolites in signaling. Redox Rep 2007; 12: 4–10.
38. Srivastava S, Dixit BL, Cai J, Sharma S, Hurst HE,
Bhatnagar A, et al. Metabolism of lipid peroxidation
product, 4-hydroxynonenal (HNE) in rat erythrocytes:
role of aldose reductase. Free Radic Biol Med 2000;
29: 642–651.
39. Seth K, Agrawal AK, Aziz MH, Ahmad A, Shukla Y,
Mathur N, et al. Induced expression of early response
genes/oxidative injury in rat pheochromocytoma
(PC12) cell line by 6-hydroxydopamine: implication for
Parkinson’s disease. Neurosci Lett 2002; 330: 89–93.
40. Bove J, Prou D, Perier C, and Przedborski S. Toxin-
induced models of Parkinson’s disease. NeuroRx
2005; 2: 484–494.
41. Chaturvedi RK, Shukla S, Seth K, Chauhan S, Sinha C,
Shukla Y, et al. Neuroprotective and neurorescue
effect of black tea extract in 6-hydroxydopamine-
lesioned rat model of Parkinson’s disease. Neurobiol
Dis 2006; 22: 421–434.
42. Eminel S, Klettne A, Roemer L, Herdegen T, and
Waetzig V. JNK2 translocates to the mitochondria and
mediates cytochrome c release in PC12 cells in
response to 6-hydroxydopamine. J Biol Chem 2004;
279: 55385–55392.
43. Finkelstein EI, Ruben J, Koot CW, Hristova M, and
van der Vliet A. Regulation of constitutive neutrophil
apoptosis by the alpha,beta-unsaturated aldehydes
acrolein and 4-hydroxynonenal. Am J Physiol Lung
Cell Mol Physiol 2005; 289: 1019–1028.
44. Duarte AI, Santos MS, Oliveira CR, and Rego AC.
Insulin neuroprotection against oxidative stress in cor-
tical neurons–involvement of uric acid and glutathione
antioxidant defenses. Free Radic Biol Med 2005; 39:
876–889.
45. Mattson MP, Robinson N, and Guo Q. Estrogens stabi-
lize mitochondrial function and protect neural cells
against the pro-apoptotic action of mutant presenilin-
1. Neuroreport 1997; 8: 3817–3821.
46. Carmen D, Lobaton LV, Hernandez-SanMiguel E,
SantoDomingo J, Moreno A, Montero M, et al. Modu-
lation of mitochondrial Ca2þ uptake by estrogen recep-
tor agonists and antagonists. Br J Pharmacol 2005;
145: 862–871.
47. Razmara A, Duckles SP, Krause DN, and Procaccio V.
Estrogen suppresses brain mitochondrial oxidative stress
in female and male rats. Brain Res 2007; 1176: 71–81.
48. Nilsen J, Diaz BR. Mechanism of estrogen-mediated
neuroprotection: regulation of mitochondrial calcium
and Bcl-2 expression. Proc Natl Acad Sci U S A
2003; 100: 2482–2487.
49. Burris TP, Krishnan V. Estrogen: a mitochondrial
energizer that keeps on going. Mol Pharmacol 2005;
68: 956–958.
50. Stirone C, Duckles SP, Krause DN, and Procaccio V.
Estrogen increases mitochondrial efficiency and
reduces oxidative stress in cerebral blood vessels. Mol
Pharmacol 2005; 68: 959–965.
Siddiqui MA et al. 869
at UNIV OF LOUISVILLE on January 31, 2012het.sagepub.comDownloaded from
Top Related
Copyright © 2022 FDOKUMEN

