







Bahasa
Halaman
Hukum


Phylogenetic analysis of the genus Thricops Rondani(Diptera: Muscidae) based on molecular andmorphological characters
JADE SAVAGE 1 , TERRY A . WHEELER 1 and BR IAN M. WIEGMANN2
1Department of Natural Resource Sciences, McGill University, Macdonald Campus, Ste-Anne-de-Bellevue, Quebec, Canada
and 2Department of Entomology, North Carolina State University, Raleigh, North Carolina, U.S.A.
Abstract. The muscid genus Thricops Rondani comprises forty-four species andtwo subspecies restricted to the northern hemisphere. A species-level phylogeneticanalysis of Thricops was conducted using forty-four morphological characters,426 bp of the nuclear gene white and 523 bp spanning the 50 end of the cytochromec oxidase subunit I (COI), the tRNA leucine gene (L2 region) and the 30 end of thecytochrome c oxidase subunit II (COII). Thirty-nine species and two subspecies ofThricops were included in the analysis. Two species of Azelia Robineau-Desvoidyand one species of Hydrotaea Robineau-Desvoidy were used as outgroups. Mor-phological characters were coded for all included species, the mitochondrial genefragment (COIþ II) was sequenced for a subset of seventeen species of Thricops andthree outgroup species, and white for twelve of those seventeen Thricops species andtwo outgroup species. Six separate maximum parsimony analyses were performed onthree taxon sets of different sizes (n¼ 14, n¼ 20, n¼ 44). Results from the partitionhomogeneity test indicated no significant incongruence between data partitions, andfour combined maximum parsimony analyses were conducted (DNAþmorphologyfor n¼ 14; COIþ IIþmorphology for n¼ 20; DNAþmorphology for n¼ 20;DNAþ morphology for n¼ 44). The relative contribution of each data partitionto individual nodes was assessed using partitioned Bremer support. Strict consensustrees resulting from the unweighted analyses of each dataset are presented. Combi-nation of datasets increased resolution for the small taxon set (n¼ 14), but not forthe larger ones (n¼ 20, n¼ 44), most probably due to increasing amounts of missingdata in the larger taxon sets. Results from both individual and combined analyses ofthe smaller taxon sets (n¼ 14, n¼ 20) provided support for the monophyly ofThricops and a complete division of the genus into two monophyletic subgroups.The strict consensus cladograms resulting from the analysis of the morphologicaldata alone and the combined data for the large taxa set (n¼ 44) both supported themonophyly of the genus, but placed the species Thricops foveolatus (Zetterstedt) andThricops bukowskii (Ringdahl) at the base of the ingroup, in a polytomy with arelatively well-resolved branch comprising all remaining species of the genus. Thebasal position of these two species, included in the morphological taxon set butabsent in the others, illustrates the potential pitfalls of taxon sampling and missingdata in phylogenetic analyses. The synonymy of Alloeostylus with Thricops asproposed by previous authors was supported by our results. Relative contributionsof different data partitions is discussed, with the mitochondrial sequence generallyproviding finer resolution and better branch support than white.
Correspondence: Jade Savage, Department of Natural Resource
Sciences, McGill University, Macdonald Campus, Ste-Anne-de-
Bellevue, Quebec, Canada, H9X 3V9. E-mail: [email protected]
Systematic Entomology (2004) 29, 395–414
# 2004 The Royal Entomological Society 395

Introduction
Thricops Rondani is a morphologically diverse genus of the
family Muscidae. Several anthophilous muscid flies are
considered to be major pollinators of open blossoms in
arctic and subarctic ecosystems (Pont, 1993), and species
of Thricops can often be found in large numbers feeding on
the pollen and nectar of a variety of flowering plants. In
northern Sweden, Thricops was shown to be among the
most important flower visitors above the timberline (Elberling
& Olesen, 1999). The world fauna of Thricops was revised
recently (Savage, 2003) and now includes forty-four species
and two subspecies. All species are restricted to the northern
hemisphere, wheremost favour arctic or alpine habitats.Most
species are endemic to either the Palaearctic or Nearctic
regions, with a few known exclusively from the Oriental
region. Two species are found in all three realms and seven
have a Holarctic distribution.
In spite of the broad interspecific spectrum of colour, size
and external structures displayed by members of Thricops,
all species can be separated easily from related groups based
on the combination of the following characters: posterior
surface of hind coxa setulose, vein A1 extending more than
halfway to the wing margin, the presence of a strong poster-
odorsal bristle on the apical third of the hind tibia (calcar)
and the presence of more than one pair of frontal bristles in
the male.
Thricops was erected by Rondani (1856), who designated
Anthomyza hirtula Zetterstedt as the type species. No
hypothesis of phylogenetic relationships exists withinThricops,
but Schnabl (1888a, b, c, 1889) attempted to organize parts of
the Palaearctic fauna into a variety of genera and subgenera.
Schnabl (1888a) erected the genus Alloeostylus for a
new species which was distinguished mainly by the peculiar
genitalia with elongated surstyli. Soon, however, Schnabl
(1888b) transferred more species to Alloeostylus, including
some with short surstyli such as those seen in Thricops as
well as most other azeliine genera. As more species were
added to Alloeostylus, eventually no character other than the
presence or absence of a posteroventral bristle on the midtibia
was left to separate Alloeostylus from Thricops.
Most of Schnabl’s later generic and subgeneric divisions
of Thricops (see Savage, 2003 for the complete synonymy)
were ignored or modified by most subsequent authors
(Malloch, 1921; Ringdahl, 1947; Hennig, 1962, 1965; Huckett,
1965a, b; Skidmore, 1985), except for the recognition of
Alloeostylus. Skidmore (1985) suggested that Alloeostylus
should be maintained as a genus distinct from Thricops,
based on larval and pupal characters of six species. Adult
morphology, however, suggests otherwise (Hennig, 1962),
and the presence of a posteroventral bristle as the single
character to separate Alloeostylus from Thricops has
left many unconvinced (Malloch, 1921; Hennig, 1962,
1965; Huckett, 1965a). The aberrant Palaearctic species
T. bukowskii (Ringdahl), for example, has elongated surstyli,
similar to those seen in the type species of Alloeostylus, but no
posteroventral bristle on the midtibia. Males of the Nearctic
species T. tarsalis (Walker) have the bristle, but it is absent
occasionally in females, and a few specimens ofT. rufisquamus
ssp. rufisquamus (Schnabl), a striking species placed tradition-
ally in Alloeostylus, lack a posteroventral bristle on the mid-
tibia of both sexes. Consequently, the use of this bristle as the
single character supporting the monophyly ofAlloeostylus has
not been accepted by all researchers, and the genus was even-
tually synonymized with Thricops (Pont, 1986a). It should be
noted, however, that the synonymy was made in a catalogue
and therefore was not based on a phylogenetic analysis.
In view of these problems in classification, the primary
objective of the current study was to carry out a phylogen-
etic analysis of the species of Thricops sensu Pont (1986a)
in order to test the monophyly of Thricops and Alloeostylus.
A growing trend among systematists over the last decade
has been to combine morphological and molecular data in a
single ‘total evidence’ analysis. When datasets are not sig-
nificantly incongruent with one another, this approach will
generally increase phylogenetic accuracy (Bull et al., 1993;
Chippindale & Wiens, 1994; Kluge, 1998). However, many
species of Diptera are rarely collected or known only from
old type series. Therefore, it is impossible to acquire mater-
ial suitable for DNA extraction for some taxa. Technical
difficulties related to the amplification and sequencing of
DNA as well as the relatively high cost of gathering mole-
cular characters are additional problems limiting the num-
ber of species included in phylogenetic analyses. To cope
with these issues, many scientists use the exemplar
approach, in which a sample of species is selected to repre-
sent the major lineages of a higher taxon. This was the case
here: almost all species of Thricops were included in the
morphological dataset, but less than half could be included
in the molecular datasets.
Three character sets were used: morphological characters
of adult flies; mitochondrial DNA sequence data from a
fragment spanning the end of the cytochrome c oxidase
subunit I (COI), the tRNA leucine gene (L2 region), and a
small section of the cytochrome c oxidase subunit II (COII);
and nuclear DNA sequence data from a fragment of the
nuclear gene white. The use of three character sets also allowed
us to examine congruence among these data sources and to
compare the relative contribution of the three character sets to
the overall phylogeny. We also compared phylogeny estimates
based on a small matrix of exemplar species with one based
on a larger matrix containing nearly all described species.
Materials and methods
Morphological data
Thirty-nine species and two subspecies of Thricops were
included in the morphological analysis (Table 1). Thricops
flavidus Xue, T. jiyaoi Feng and T. tuberculatus Deng, Mao
& Feng were excluded because no specimens were available
396 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

for examination and data from the literature were not
detailed enough to allow coding of most of the characters.
The generic placements of T. jiyaoi and T. tuberculatus
appear to be correct based on original descriptions, but
T. flavidus obviously does not belong to Thricops (see
Savage, 2003). Almost 60% of morphological characters
used in this study pertain exclusively to structures of
males, resulting in large proportions of missing data for
T. angelorum Savage and T. ineptus (Stein), described only
from females. Several studies (Novacek, 1992a, b; Wilkinson
& Benton, 1995; Wilkinson, 1995) have demonstrated that
highly incomplete character matrices tend to reduce the over-
all accuracy of estimated trees. Preliminary analyses includ-
ing the two incompletely scored Thricops species resulted in
high numbers of conflicting equally parsimonious trees and a
substantial decrease in resolution in a consensus tree. The
uncertainty added to the matrix by just these two taxa thus
substantially obscures the phylogenetic signal in male
Table 1. Taxa included in the morphological analysis. See Savage (2003) for the locations of the type material and a detailed list of specimens
examined.
Species Distribution Material examined
Ingroup
Thricops aculeipes (Zetterstedt) Palaearctic Lectotype
Thricops aduncus Savage Nearctic Holotype
Thricops albibasalis (Zetterstedt) Holarctic Holotype
Thricops beckeri (Pokorny) Palaearctic Nontypes
Thricops bukowskii (Ringdahl) Palaearctic Syntypes (4)
Thricops calcaratus (Porchinskiy) Palaearctic Nontypes
Thricops coquilletti (Malloch) Holarctic Holotype
Thricops culminum (Pokorny) Palaearctic Nontypes
Thricops cunctans (Meigen) Palaearctic Nontypes
Thricops dianae Savage Nearctic Holotype
Thricops diaphanus (Wiedemann) Holarctic, Oriental Lectotype
Thricops fimbriatus (Coquillett) Nearctic Holotype
Thricops foveolatus (Zetterstedt) Palaearctic Holotype
Thricops furcatus (Stein) Holarctic Nontypes
Thricops genarum (Zetterstedt) Palaearctic Holotype
Thricops hakusanus (Shinonaga & Kano) Palaearctic Nontypes
Thricops hirtulus (Zetterstedt) Holarctic Neotype
Thricops innocuus (Zetterstedt) Holarctic Lectotype
Thricops lividiventris lividiventris (Zetterstedt) Holarctic Lectotype
Thricops lividiventris plumbeus (Hennig) Palaearctic Paratypes (4)
Thricops longipes (Zetterstedt) Palaearctic Lectotype
Thricops nepalensis (Pont) Oriental Holotype
Thricops nigriabdominalis Savage Palaearctic Holotype
Thricops nigrifrons (Robineau-Desvoidy) Palaearctic Nontypes
Thricops nigritellus (Zetterstedt) Palaearctic Lectotype
Thricops ponti Savage Palaearctic Holotype
Thricops rostratus (Meade) Palaearctic Holotype
Thricops rufisquamus rufisquamus (Schnabl) Holarctic Nontypes
Thricops rufisquamus himalayensis (Pont) Oriental Holotype
Thricops semicinereus (Wiedemann) Palaearctic Lectotype
Thricops separ (Zetterstedt) Palaearctic Lectotype
Thricops septentrionalis (Stein) Nearctic Nontypes
Thricops simplex (Wiedemann) Palaearctic Nontypes
Thricops spiniger (Stein) Holarctic Nontypes
Thricops sudeticus (Schnabl) Palaearctic Nontypes
Thricops tarsalis (Walker) Nearctic Syntype (1)
Thricops tatricus Gregor Palaearctic Paratypes (5)
Thricops thudamensis Shinonaga Palaearctic Holotype
Thricops vaderi Savage Palaearctic Holotype
Thricops villicrus (Coquillett) Nearctic Holotype
Thricops villosus (Hendel) Palaearctic Nontypes
Outgroups
Azelia cilipes (Haliday) Holarctic Nontypes
Azelia gibbera (Meigen) Holarctic Nontypes
Hydrotaea spinifemorata Huckett Nearctic Nontypes
Phylogenetic analysis of Thricops 397
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

characters. For this reason, the two species were excluded
from all analyses presented here.
All morphological characters included in the data matrix
derive from external and genitalic structures of adults. All
characters were treated as unordered and multistate charac-
ters as nonadditive. Five cases of polymorphism in terminal
taxa were treated as such, adding extra steps to the overall
tree length for each instance of polymorphism. The complete
set of morphological characters and states is provided in
Appendix 1 and the data matrix is shown in Appendix 2.
Outgroups
Species from two other azeliine genera, Hydrotaea and
Azelia, were chosen as outgroup taxa. Azelia is considered
the sister group of Thricops (Hennig, 1965; Skidmore, 1985;
Savage & Wheeler, in press) and Hydrotaea was included to
root the cladograms. The only male Hydrotaea specimen
available for this study was identified by the senior author
as Hydrotaea spinifemorata Huckett. However, because the
holotype (and only known male specimen) is apparently lost
(R. J. Gagne, pers. comm.), the precise identity of the speci-
men used in the molecular analysis could not be confirmed.
Nevertheless, the modified fore femur of the specimen leaves
no doubt concerning its generic assignment. Outgroups are
used only to polarize characters, and as long as generic
assignment of an outgroup exemplar is accurate and the
chosen specimen is not aberrant or highly autapomorphic,
the exact species identification, although preferable, is not
crucial to the outcome of the analysis.
Notes were taken on the external morphology of the
specimen before DNA extraction, but only the legs were
saved as vouchers. Therefore, the genitalia of this specimen
were not available for the coding of characters. However,
based on the presence of a basal spine on the hind femur,
the specimen could be assigned to the Hydrotaea armipes spe-
cies group (¼Hydrotaea occulta species group sensu Hennig,
1962) (see Pont, 1986b for an explanation of the synonymy of
Hydrotaea occulta (Meigen) with Hydrotaea armipes (Fallen)).
The armipes species group was recently shown to occupy a
basal position within Hydrotaea (Savage & Wheeler, in press).
The genitalia of most other members of this group were exam-
ined and found to be very homogeneous. Therefore, genitalic
characters for the exemplar ofHydrotaea used here were coded
according to the structures seen within the armipes species
group rather than the specimen itself.
Molecular data – DNA extraction, amplification and
sequencing
The COIþ II gene region and white gene fragment were
obtained for only a subset of the study taxa due to a limited
number of adequately preserved specimens available to the
project (COIþ II: seventeen species; white: twelve species;
Table 2). Where possible, the exemplar species were chosen
to cover the breadth of morphological variation found in
the group. All specimens used for DNA extraction were
collected alive and transferred directly to 95% ethanol.
Parts of the original specimen, or a second conspecific speci-
men (identification by the senior author) from the same
collection site, were preserved as vouchers, deposited in
the North Carolina State University Insect Collection
(Raleigh, North Carolina, U.S.A.) and stored at �80 �C.For T. tarsalis, a dry specimen was used in the extraction
and the voucher was deposited in the Canadian National
Collection of Insects, Ottawa, Ontario, Canada.
Genomic DNA was extracted from a single fly using
a standard protocol involving homogenization in lysis
buffer, proteinase K incubation, phenol/chloroform extrac-
tion and isopropanol precipitation of nucleic acids (Hillis
et al., 1996; Stahls & Nyblom, 2000). A Chelex1-based
extraction procedure was used to obtain amplifiable DNA
from a dry specimen of T. tarsalis. In this procedure, a few
muscle fibres from the thorax of the specimen were used
following Skevington & Yeates (2000) as modified from
Walsh et al. (1991). All genomic DNA samples were stored
at �80 �C.Two gene fragments were amplified: COIþ II, a region
spanning the 30 end of the mitochondrial gene COI, the L2
region and the first half of the COII gene; and a portion of
white, a nuclear gene involved in eye pigmentation (primers
in Table 3). Polymerase chain reaction (PCR) amplification
reactions followed standard protocols (Kocher et al., 1989)
with TaKaRa Ex TaqTM (Mirus Corp., Madison, Wisconsin,
U.S.A.). The amplification programme for the mitochondrial
fragment was as follows: one cycle of 95 �C (5min), 93 �C(20 s), 50 �C (40 s), 72 �C (1min); thirty-three cycles of 93 �C(20 s), 50 �C (40 s), 72 �C (1min); one cycle of 72 �C (5min).
For white, the amplification programme followed: one cycle
of 94 �C (5min); thirty-five cycles of 94 �C (1min), 47 �C(1min), 72 �C (1min 30 s). The resulting PCR products
were either used directly in the sequencing reaction or were
gel purified prior to cycle sequencing.
Sequences were obtained by dye terminator cycle sequen-
cing using the ABI Prism dRhodamine Terminator Cycle
Sequencing Kit (PE Biosystems, Warrington, U.K.) and run
on the Applied Biosystems automated DNA sequencer
(ABI Prism 377). Sequence confirmation was accomplished
by comparing complementary DNA strands. Editing
nucleotide sequences, contig assembly, and consensus
sequence calculation were performed using the software
program SEQUENCHER (Gene Codes Corp., Ann Arbor,
Michigan, U.S.A.). The reverse strand of the white gene
failed to satisfactorily sequence for T. diaphanus (Wiede-
mann), T. longipes (Zetterstedt), T. spiniger (Stein) and
T. semicinereus (Zetterstedt). However, the high quality of
the forward sequence for these taxa and confirmation by
protein translation allowed their inclusion in the analysis.
Data analysis
After editing the sequences, �192 bp from the 30 end of
COI, the L2 region (�73 bp) and �258 bp from the 50 end of
398 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414
COII were aligned for the analysis. For the white gene,
�426 bp were aligned. Alignment was achieved manually
requiring few indels and was confirmed by translation of
protein coding regions. Nucleotide sequence alignments
may be obtained upon request from the senior author or
from TreeBase (http://www.treebase.org).
For both genes, third positions were included in the analy-
sis. The distribution of parsimony informative sites between
codon position differed between the mitochondrial and the
nuclear genes. ForCOIþ II, 50.8–57.9% of parsimony informa-
tive characters were found at the third position, whereas this
value rose to 89.4% for white. The 30 end of the L2 region
contained a small number of variable sites requiring inser-
tion of gaps in the alignment. Gaps were treated as missing
data in the analyses reported below. Parallel analyses run
with these seven nucleotide positions excluded did not
change any of the reported tree topologies.
Ten different analyses were performed on three taxon sets
of different sizes (n¼ 14, n¼ 20, n¼ 44).
The first set of analyses included only taxa for which all
three data partitions were available (Table 2). This taxon set
(n¼ 14) included twelve exemplar species of Thricops sensu
lato and two outgroup species, Azelia gibbera (Meigen) and
Hydrotaea spinifemorata. All character partitions (COIþ II,
white, morphology) were analysed separately and then in
combination.
Table 2. Taxa used for DNA extraction, collecting data, voucher type and GenBank accession numbers.
Accession number
Taxon Collecting data Voucher type COIþ II white
Ingroup
Thricops aculeipes Sweden: Mt. Nuolja. 5.viii.2001. J. Savage Conspecific specimen AY367142 –
Thricops albibasalis U.S.A.: NH, Mount Washington. 16.vii.2001. J. Savage Hind legs, abdomen AY367139 AY367159
Thricops cunctans Sweden: Abisko Ntl. Pk. 6.viii.2001. J. Savage Conspecific specimen AY367143 –
Thricops diaphanus Sweden: Jebrenjokk. 9.viii.2001. J. Savage Conspecific specimen AY367144 AY367162
Thricops furcatus Sweden: Mt. Nuolja. 5.viii.2001. J. Savage Conspecific specimen AY367141 AY367161
Thricops genarum Sweden: Jebrenjokk. 9.viii.2001. J. Savage Conspecific specimen AY367140 AY367160
Thricops hirtulus U.S.A.: NH, Mount Washington. 16.vii.2001. J. Savage Conspecific specimen AY367145 AY367163
Thricops innocuus Sweden: Abisko Ntl. Pk. 30.vii.2001. J. Savage Conspecific specimen AY367146 AY367164
Thricops lividiventris Sweden: Abisko Ntl. Pk. 6.viii.2001. J. Savage Conspecific specimen AY367147 –
Thricops longipes Sweden: Jebrenjokk. 7.viii.2001. J. Savage Conspecific specimen AY367148 AY367165
Thricops nigrifrons Sweden: Jebrenjokk.7.viii.2001. J. Savage Conspecific specimen AY367149 AY367166
Thricops nigritellus Sweden: Jebrenjokk.7.viii.2001. J. Savage Conspecific specimen AY367150 AY367167
Thricops semicinereus Sweden: Jebrenjokk. 9.viii.2001. J. Savage Conspecific specimen AY367151 AY367168
Thricops separ Sweden: Abisko Ntl. Pk. 30.vii.2001. J. Savage Conspecific specimen AY367155 –
Thricops septentrionalis U.S.A.: NH. Mt. Washington. 16.vii.2001. J. Savage Conspecific specimen AY367152 AY367169
Thricops spiniger U.S.A.: NH. Mount Washington. 16.vii.2001. J. Savage Conspecific specimen AY367153 AY367170
Thricops tarsalis Canada: MB. Along Souris river. 9.vi.1993. B. Gallaway Dry specimen AY367154 –
Outgroups
Azelia cilipes U.S.A.: VT, Camel’s hump trail. 26.viii.2001. S.E. Brooks Hind legs, abdomen AY367157 –
Azelia gibbera Sweden: Abisko Ntl. Pk. 8.viii.2001. J. Savage Conspecific specimen AY367156 AY367171
Hydrotaea spinifemorata U.S.A.: NC, GSMNP; Ravensford, nr. Cherokee,
9–16.vi.2001. B.Cassel
Legs AY367158 AY367172
COI þ II¼ cytochrome c oxidase subunits I and II.
Table 3. The primers used to amplify mitochondrial and nuclear genes. The direction of the primers is provided by S or F (sense or forward)
and A or R (antisense or reverse).
Gene region Primer name and sequence Reference
COI þ COII S2792lep: ATACCTCGACGTTATTCAGA Brown et al. (1994)
A3389lep: TCATAAGTTCARTATCATTG Brown et al. (1999)
white white-1F: TGYGCNTATGTNCARCARGAYGA Baker et al. (2001)
white-2R: ACYTGNACRTAAAARTCNGCNGG Baker et al. (2001)
white-3R: ACATAR AARTCNGCNGGRTTRTARTT J. K. Moulton, pers. comm.
TMwhite 1F: TGTAAAACGACGGCCAGT þ white-1F J. K. Moulton, pers. comm.
TMwhite 2R: CAGGAAACAGCTATGACC þ white-2R J. K. Moulton, pers. comm.
COIþII¼ cytochrome c oxidase subunits I and II.
Phylogenetic analysis of Thricops 399
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

The second set of analyses (n¼ 20) included five more
ingroup species (Table 2) and an additional outgroup
species, Azelia cilipes (Haliday). No white sequence could
be obtained for these additional species, thus only the
morphology and the mitochondrial gene were analysed
separately. Two combined analyses were conducted for
this taxon set, one including COIþ II and morphology,
and another including the incomplete white character set.
The third set of analyses (n¼ 44) included twenty more
species and two subspecies of Thricops and the same three
outgroup taxa as in the previous analyses (Table 1). Only
morphological data were available for these additional taxa.
The large morphological matrix was analysed separately
and a final combined analysis was performed including all
available data.
All phylogenetic analyses were conducted under the par-
simony criterion, using PAUP* version 4.0b10 (Swofford,
2002). Unweighted parsimony analyses were performed on
the morphology and molecular datasets separately. For
smaller datasets (n¼ 14, n¼ 20), the branch and bound
algorithm was used (addition sequence¼ furthest). Heuristic
searches were performed on the larger datasets (1000
random addition sequences, tree bisection-reconnection
(TBR) swapping, collapse branch if maximum length¼ 0).
Tree statistics are presented in Table 4, uncorrected pairwise
sequence divergence in Table 5, average nucleotide composi-
tion and percentage of parsimony informative characters per
codon position in Table 6.
The incongruence length difference test (Farris et al.,
1995a, b), implemented in PAUP* as the partition homo-
geneity test, was used to assess the degree of incongruity
between various partitions (P< 0.05 indicates significant
incongruence between data partitions). The results of the
partition homogeneity test (Table 7) indicated no signifi-
cant incongruence between pairwise comparisons of all
data partitions of the same size. Based on these results,
four combined maximum parsimony analyses were con-
ducted (DNAþmorphology for n¼ 14; COIþ IIþmorph-
ology when n¼ 20; DNAþmorphology for n¼ 20;
DNAþmorphology when n¼ 44) using the same search
parameters as those used for individual datasets of the same
size.
Evidential support for different clades in all individual
and combined analyses was assessed using nonparametric
bootstrap and Bremer support (BS) indices (Bremer, 1994).
For datasets including twenty or less species, we performed
100 bootstrap replicates excluding uninformative characters
and using the same parameters as in the parsimony
analyses, whereas for the larger datasets (n¼ 44), we set
the maximum number of trees to be retained at each repeti-
tion to 2000. BS and partitioned Bremer support (PBS), a
measure used to evaluate the relative contribution of each
partition to the overall BS at a particular node (Baker &
DeSalle, 1997), were calculated with TREEROT v.2c
(Sorenson, 1999).
Results and discussion
The results are presented for each taxon set, from the
smallest to the largest, to demonstrate progressively the
effects of missing data and taxon sampling on the outcome
of each analysis.
Small taxon set (n¼ 14)
COIþ II. Analysis of the 523 bp of mitochondrial DNA
for this small exemplar dataset yielded five equally parsi-
monious trees. The strict consensus tree (Fig. 1A) supported
the monophyly of Thricops sensu lato, and a well-supported
division of the ingroup into two major clades. One of these
clades (BS¼ 8, bootstrap¼ 100) contained only species pre-
viously placed in Alloeostylus, and was named the diaphanus
group. The other clade (BS¼ 7, bootstrap¼ 97) contained
only species belonging to Thricops sensu stricto, and was
named the semicinereus group. The diaphanus group was
fully resolved, with strong support for each node, whereas
the semicinereus group showed poor basal resolution and
support, with the exception of the sister-group relationship
Table 4. Tree statistics for all separate and combined analyses.
Data partitions Total PI EPT Length CI RI
COIþ II (n¼ 14) 523 67 5 178 0.664 0.753
white (n¼ 14) 426 45 48 171 0.708 0.770
Morphology (n¼ 14) 44 33 4 63 0.672 0.768
Combined small (n¼ 14) 993 145 1 415 0.672 0.754
COIþ II (n¼ 20) 523 95 12 246 0.613 0.750
Morphology (n¼ 20) 44 36 28 69 0.652 0.832
Combined COIþ II and morphology (n¼ 20) 567 131 22 320 0.611 0.765
Combined medium (n¼ 20) 993 176 34 493 0.632 0.762
Morphology (n¼ 44) 44 44 6 117 0.513 0.848
Combined large (n¼ 44) 993 184 46 980 540 0.596 0.793
Total¼ total number of characters; PI¼ parsimony informative characters; EPT¼ number of equally parsimonious trees; length¼ length of the mostparsimonious tree(s); CI¼ consistency index; RI¼ retention index; COI¼ cytochrome c oxidase subunit I; COII¼ cytochrome c oxidase subunit II.
400 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414
between T. septentrionalis (Stein) and T. spiniger. Uncor-
rected pairwise divergence (Table 5) for species in the diapha-
nus group (3.2–6.0%) was considerably higher than in the
semicinereus group (0.4–4.2%).
White. The analysis of the white fragment (426 bp)
generated forty-eight most parsimonious trees (strict
consensus tree in Fig. 1B). Basal relationships were the
same as those supported by the mitochondrial sequence
but with lower branch support values, and the range of
uncorrected pairwise sequence divergence (Table 5) among
species of the diaphanus group was also higher (1.9–6.1%)
for this gene than among species of the semicinereus group
(1.0–3.5%). Within the diaphanus group, white supported a
sister-group relationship between T. genarum (Zetterstedt)
and T. furcatus (Stein), which was in conflict with the
positions of T. genarum and T. furcatus supported by the
mitochondrial sequence (Fig. 1A). The nuclear gene failed
to resolve relationships within the T. semicinereus group,
with the exception of the strongly supported sister-
group relationship between T. septentrionalis and
T. spiniger.
Morphology. The analysis of morphological characters
for this dataset resulted in four most parsimonious trees
(strict consensus tree in Fig. 1C). The deep branch relation-
ships were the same as those in Fig. 1(A) and (B), but with
low support for the monophyly of the semicinereus group
(BS¼ 1, bootstrap 54). Within the diaphanus group,
T. diaphanus stood as the sister group to an unresolved
polytomy including T. genarum, T. albibasalis (Zetterstedt)
and T. furcatus. As with the other datasets, there was poor
basal resolution within the semicinereus group, with the
exception of a well-supported sister-group relationship
between T. septentrionalis and T. spiniger.
Combined data. The combination and analysis of all
three datasets generated a single most parsimonious tree
(Fig. 2). This tree was better resolved and had much higher
branch support for the basal nodes than any of the previous
ones but showed some conflict at some of the internal and
terminal nodes. Within the diaphanus group, PBS showed
that the sister-group relationship between T. albibasalis and
T. genarum was supported strongly by COIþ II, but in
conflict with white. Mild conflict was seen within the semi-
cinereus group. The individual contribution of both mole-
cular partitions to most internal nodes within the
semicinereus group was either low or mildly conflicting
with other partitions and most of the support for those
branches could be attributed to the morphological dataset.
Although T. nigrifrons (Robineau-Desvoidy) and
T. longipes are extremely similar in both external and internal
morphology, both molecular datasets provided only weak
support for a sister-group relationship between these two
species. By contrast, T. spiniger and T. septentrionalis,
another pair of very similar species, did receive strong
support from all partitions. Evidence from the individual
and combined analyses of the smaller taxon set indicated
that Alloeostylus could regain full generic status and be
treated as the sister genus to Thricops.
Table 5. Uncorrected pairwise sequence divergence between species for small (n¼ 14) and medium (n¼ 20) taxon sets.
Taxa COIþ II (tRNA included) COI only L2 only COII only white
Small taxon set (n¼ 14)
Across all taxa (n¼ 14) 0.4–13.1% 0.0–15.1% 0.0–10.2% 0.4–13.6% 1.0–20.0%Within ingroup (n¼ 12) 0.4–11.2% 0.0–12.5% 0.0–10.2% 0.4–12.0% 1.0–9.1%T. diaphanus group (n¼ 4) 3.2–6.0% 3.3–6.8% 0.0–5.6% 3.1–7.0% 1.9–6.1%T. semicinereus group (n¼ 8) 0.4–4.2% 0.0–5.3% 0.0–2.9% 0.4–3.3% 1.0–3.5%
Medium taxon set (n¼ 20)
Across all taxa (n¼ 20) 0.4–13.1% 0.0–15.3% 0.0–10.2% 0.4–13.6% –
Within ingroup (n¼ 17) 0.4–11.8% 0.0–14.1% 0.0–10.2% 0.4–12.0% –
T. diaphanus group (n¼ 6) 3.2–8.6% 3.4–9.7% 0.0–7.0% 3.1–8.9% –
T. semicinereus group (n¼ 11) 0.4–5.2% 0.0–8.9% 0.0–3.0% 0.4–4.7% –
COI¼ cytochrome c oxidase subunit I; COII¼ cytochrome c oxidase subunit II.
Table 6. Average nucleotide composition and percentage of parsimony informative (PI) characters per codon position for small (n¼ 14) and
medium (n¼ 20) datasets.
Gene and taxon set Average proportion of A :C :G :T % of PI characters per codon position
COIþ II small (n¼ 14) 34 : 12 : 10 : 44 39.3, 9.9, 50.8
COIþ II medium (n¼ 20) 34 : 12 : 10 : 44 33.0, 9.1, 57.9
white small (n¼ 14) 25 : 23 : 25 : 27 8.5, 2.1, 89.4
COI þ II¼ cytochrome c oxidase subunits I and II.
Phylogenetic analysis of Thricops 401
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

Medium taxon set (n¼ 20)
COIþ II. Analysis of the mitochondrial fragment for this
taxon set yielded twelve equally parsimonious trees (strict
consensus tree in Fig. 3A). When compared with the analy-
sis of COIþ II for the smaller dataset (Fig. 1A), this con-
sensus tree still showed good support for the basal branches
(BS¼ 6, bootstrap¼ 99 for the diaphanus group; BS¼ 5,
bootstrap¼ 96 for the semicinereus group) as well as
increased resolution within the semicinereus group, but
with low support for almost all branches.
Morphology. The analysis of morphological characters
generated twenty-eight equally parsimonious trees (strict
consensus tree in Fig. 3B). The addition of taxa to this
dataset did not improve the internal resolution of the two
major clades but slightly increased branch support for the
diaphanus and semicinereus groups. There were several dif-
ferences between the trees seen in Fig. 3(A) and B, especially
within the semicinereus group. Although most nodes in
Fig. 3A were weakly supported, the placement of T. separ
and T. longipes was quite different between the two trees.
Combined COIþ II and morphology. The combination of
these two datasets yielded twenty-two equally parsimonious
trees (strict consensus tree in Fig. 4A). This consensus tree
showed strong support for the monophyly of Thricops, as
well as the division of the group into two separate clades
(BS¼ 10, bootstrap¼ 100 for the diaphanus group; BS¼ 9,
bootstrap¼ 99 for the semicinereus group). Basal relation-
ships within the diaphanus group were unresolved, but all
resolved nodes within this group were well supported.
Within the semicinereus group, T. aculeipes stood as the
sister group to the other species, but this node was weakly
supported. The basal relationships of the remaining species
were unresolved, but there was relatively good support for
the placement of T. separ as the sister group to T. spiniger
and T. septentrionalis, and good support for a sister-group
relationship between T. longipes and T. nigrifrons. PBS
showed very little conflict in this combined analysis, with
weak disagreement seen only in two branches of the semi-
cinereus group.
Combined DNA and morphology. The strict consensus
tree presented in Fig. 4(B) resulted from the analysis of all
species with at least one molecular dataset. This combined
analysis yielded thirty-four equally parsimonious trees. The
Table 7. The results of the partition homogeneity test.
Partition comparison P value
COIþ II (n¼ 14) and white (n¼ 14) 0.76
COIþ II (n¼ 14) and morphology (n¼ 14) 0.47
Morphology (n¼ 14) and white (n¼ 14) 1.0
COIþ II (n¼ 20) and morphology (n¼ 20) 0.21
COI þ II¼ cytochrome c oxidase subunits I and II.
T. albibasalisT. genarumT. furcatusT. diaphanusT. hirtulusT. innocuusT. longipesT. septentrionalisT. spinigerT. nigrifronsT. semicinereusT. nigritellusA. gibberaH. spinifemorata
diaphanusgroup
semicinereusgroup
9897
72
100100
96
3
3
98
7
2
A
11
1
T. albibasalisT. genarumT. furcatusT. diaphanusT. hirtulusT. innocuusT. longipesT. nigrifronsT. nigritellusT. semicinereusT. septentrionalisT. spiniger A. gibberaH. spinifemorata
A. gibbera
T. albibasalisT. genarumT. furcatusT. diaphanusT. hirtulusT. innocuusT. septentrionalisT. spinigerT. longipesT. nigrifronsT. nigritellusT. semicinereus
H. spinifemorata
54
1
832
99
82
2
3
C
1 98
4
76
1
93
3
90
4
987 57
180
2
B
653
Fig. 1. A–C, Results of individual analyses of all three data
partitions for the reduced taxon set (n¼ 14). A, Maximum
parsimony strict consensus of five most parsimonious trees for
cytochrome c oxidase subunits I and II; B, maximum parsimony
strict consensus of forty-eight most parsimonious trees for white; C,
maximum parsimony strict consensus of four most parsimonious
trees for morphology. The values above the branches refer to
bootstrap values (from 100 replicates). The values below the
branches refer to Bremer support.
402 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

consensus tree resulting from this analysis was almost iden-
tical to Fig. 4(A), with the exception that T. aculeipes was
included in the basal polytomy of the semicinereus group.
When compared with the combined analysis of COIþ II
and morphology (Table 4), the addition of the incomplete
white dataset (missing data for T. lividiventris, T. tarsalis, T.
aculeipes, T. culminum, T. separ, and A. cilipes) has had little
impact. It resulted in a small increase in the number of
equally parsimonious trees, a slight loss of basal resolution
in the semicinereus group and some mild conflict in the PBS
at a number of nodes.
Large taxon set (n¼ 44)
Morphology. The analysis of the large morphological
matrix (n¼ 44) generated six equally parsimonious trees.
The strict consensus tree (Fig. 5A) was well resolved but
with relatively low branch support for most nodes, as
expected from a dataset with the same number of characters
as taxa. There is topological similarity with Figs 1–4, with
all species formerly placed in Alloeostylus forming a mono-
phyletic group. Also present was a branch comprising
almost all species of Thricops sensu stricto. However, the
major contrast with previous topologies was that two spe-
cies of Thricops sensu stricto, T. foveolatus (Zetterstedt) and
T. bukowskii, formed a basal polytomy with the branch
comprising the semicinereus þ diaphanus groups.
This has some implications for the diaphanus group, in
that in spite of its monophyly, this branch constitutes a
derived clade nested within Thricops sensu stricto, rather
than a true sister group and thus Alloeostylus cannot be
treated as valid without rendering Thricops sensu stricto
paraphyletic.
One of the six equally parsimonious trees generated by
the analysis of the large morphological matrix is shown in
Fig. 6 with character changes plotted on the tree. The
monophyly of Thricops was supported by six apomorphic
character states, two of which are uniquely derived synapo-
morphies. As predicted by Michelsen (1978), the presence
of flared lateral sclerites (character 31: 1) is unique to Thri-
cops.
The basal placement of T. foveolatus is not surprising, as
it has retained many of the ground plan character states
present in both Hydrotaea and Azelia. However, the basal
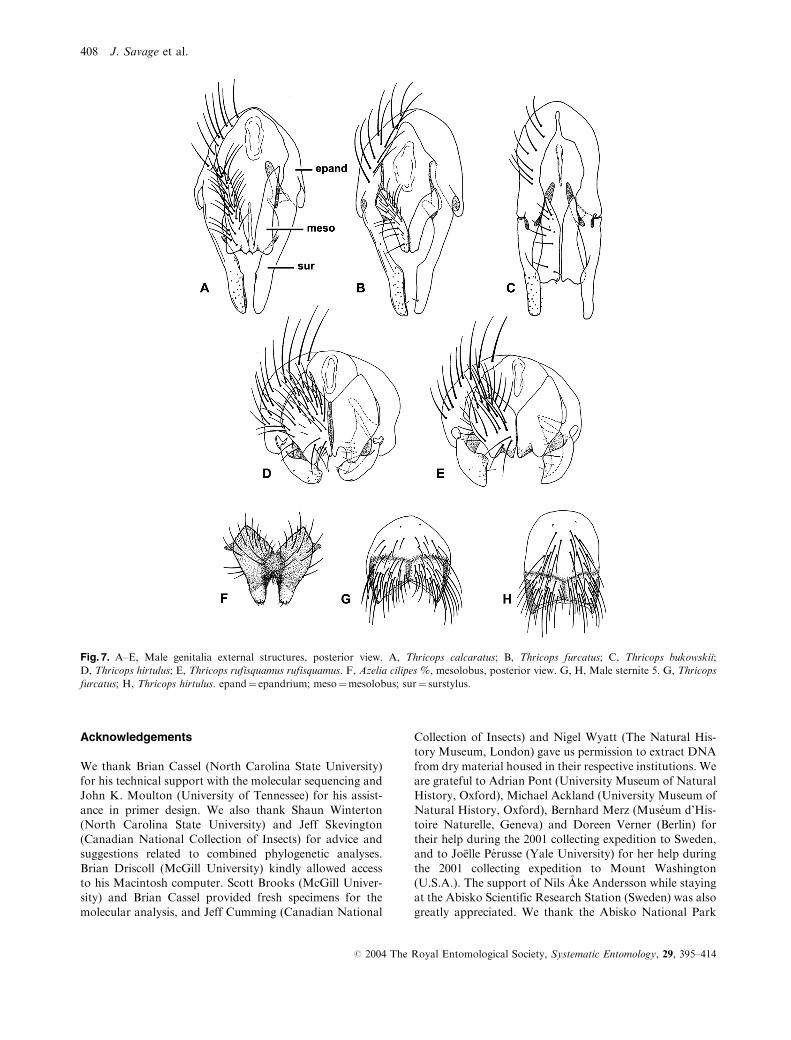
position of T. bukowskii is puzzling. Thricops bukowskii is
a peculiar species, with the genitalia superficially resembling
those of an apical clade of the diaphanus species group
(Fig. 7C). However, the resemblance stops at the elongated
surstyli, as the other genitalic structures of this species differ
from any other in the genus (Fig. 8C). The mesolobus
(Fig. 7C) is very flat, completely cleft medially and equipped
with very few setae. Internally (Fig. 8C), the basiphallus of
T. bukowskii is extremely well sclerotized and forms a long
tube in lateral view. The distiphallus abruptly expands from
an apical constriction of the basiphallus, rather than gradu-
ally expanding from base to apex. The lateral sclerites are
almost rectangular in lateral view and very well sclerotized
dorsally. Long narrow pregonites are found in some species
of the diaphanus group (i.e. Fig. 8B), but those of
T. bukowskii differ by the very narrow base and the rounded
apex. Finally, the postgonite has a bulbous base and a
strongly tapered apex, rather than a flat base and a rounded
apex. These differences have often made it hard to homo-
logize structures between T. bukowskii and other species,
and on two occasions (the shape of the mesolobus and the
shape of the postgonite), an additional character state had
to be included to properly describe the structures observed
in this taxon. The placement of this highly autapomorphic
diaphanusgroup
A. gibbera
T. albibasalis
T. genarum
T. furcatus
T. diaphanus
T. hirtulus
T. nigritellus
T. innocuus
T. septentrionalis
T. spiniger
T. longipes
T. nigrifrons
T. semicinereus
H. spinifemorata
semicinereusgroup
3(0.3, 1.1, 1.6)
14(6, 5, 3)
1(–0.4, 0.4, 1)
1(–0.6, 0.6, 1)
9(4, 3, 2)
19(9, 7.5, 2.5)
2(3, –1, 0)
78100
100
15(8, 4, 3)
93
1(–0.4, 0.4, 1) 72
100
11(4, 3, 4)100 77
4(0.4, 0.7, 2.9)
Fig. 2. The single most parsimonious tree resulting from the maximum parsimony combined analysis for the reduced taxon set (n¼ 14). The
values above the branches refer to bootstrap values (from 100 replicates). The values below the branches refer to Bremer support and
partitioned Bremer support (cytochrome c oxidase subunits I and II, white, morphology).
Phylogenetic analysis of Thricops 403
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

species should be considered doubtful until more data are
obtained. Unfortunately, T. bukowskii is rare and attempts
to amplify DNA from dry material in our possession failed.
All other ingroup species were included in a large clade
supported by a well-developed prealar bristle (7: 1) and the
uniquely derived presence of a long epiphallus (35: 1). This
clade was then divided into the diaphanus and semicinereus
species groups.
The diaphanus species group was well supported, and the
presence (at least in the male) of a posteroventral bristle on
the midtibia (18: 1) is a uniquely derived synapomorphy for
this group, although that bristle is occasionally absent in
aberrant specimens of T. r. rufisquamus. The presence of
long surstyli (28: 1), originally a diagnostic character of
Alloeostylus, supported a large derived clade of eleven spe-
cies. This character state was also present in T. bukowskii,
but obviously as the result of homoplasy. Another genitalic
character, the presence of a V-shaped mesolobus (30: 1),
also supported this group.
The semicinereus species group was supported by three
characters. None of these was uniquely derived, but the
shape of the pregonite (36: 1) and the attachment of the
epandrial arms (39: 0) are character states which arose
only once within the ingroup. Thricops beckeri (Pokorny),
a species with a dichoptic male, stood as sister to the
remaining species. Most basal relationships within the semi-
cinereus group presented in this cladogram should be treated
with caution, as most nodes are supported by characters
which are highly variable within Thricops as well as other
azeliine genera.
Combined data. The analysis of all available data resulted
in 46 980 equally parsimonious trees (strict consensus tree in
Fig. 5B). This high number can be explained mostly by
excessive missing data as well as mild conflict within the
semicinereus group. As mentioned earlier, highly incomplete
taxa are often recognized to have a negative impact on
overall phylogenetic accuracy. Incomplete character sets,
however, appear to be more forgiving when analysed in
combination with at least one other complete dataset
(Wiens, 1998). Based on simulated datasets, Wiens (1998)
demonstrated that the addition or combination of incom-
plete character sets is generally more likely to increase or
have little impact than decrease phylogenetic accuracy,
especially if the added dataset contains less than 50% miss-
ing data. The large combined dataset studied here had more
than 50% missing data, accounting for the large number of
equally parsimonious trees.
The strict consensus tree resulting from the combined
analysis of all available data summarizes our current view
of Thricops relationships. The basal relationships are iden-
tical to those of Fig. 5(A), with T. foveolatus and
T. bukowskii placed as the basal species. Among the remain-
ing species, the diaphanus clade was by far the best sup-
ported and better resolved. Low support for the basal
branches of this lineage suggests that these relationships
should be viewed with caution. Unfortunately, the most
basal clade including T. r. rufisquamus was the only major
branch of the diaphanus group for which no molecular data
were available. However, based on morphology, we are
confident that this group does hold a basal position within
the diaphanus group. The terminal clade of eleven species
containing T. sudeticus (Schnabl) had low branch support,
but similarities in genitalic structures within this group
provide evidence strongly supporting its monophyly.
Branch support for the semicinereus group in Fig. 5(B) is
low (bootstrap¼ 63) but slightly higher than on Fig. 5(A).
In spite of this low value, we consider this group to be valid,
especially as it was well supported in all other combined
analyses where missing data had less impact on branch
3
1
4
2
8278
T. albibasalisT. genarumT. furcatusT. lividiventrisT. diaphanusT. tarsalisT. aculeipesT. cunctansT. nigritellusT. hirtulusT. innocuusT. longipesT. nigrifronsT. semicinereusT. septentrionalisT. spiniger T. separ A. gibberaA. cilipesH. spinifemorata
973
88
1
381
95
89B985
156
100
107
291
10099
A
14
2
6
T. albibasalisT. genarumT. furcatusT. lividiventrisT. diaphanusT. tarsalisT. aculeipesT. cunctansT. hirtulusT. innocuusT. longipesT. septentrionalisT. spiniger T. nigrifronsT. semicinereusT. separ T. nigritellusA. gibberaA. cilipesH. spinifemorata
diaphanusgroup
semicinereusgroup
67
965 1
39411
1 11
100
Fig. 3. A, B, The results of individual analyses of cytochrome c
oxidase subunits I and II and morphology for the medium taxon
set (n¼ 20). A, Maximum parsimony strict consensus of twelve
most parsimonious trees for cytochrome c oxidase subunits I and
II; B, maximum parsimony strict consensus of twenty-eight most
parsimonious trees for morphology. The values above the branches
refer to bootstrap values (from 100 replicates). The values below
the branches refer to Bremer support.
404 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

support values. The semicinereus group lost most basal
resolution in this combined analysis. This was to be
expected, as no individual dataset provided well-supported
relationships for these nodes.
Relative contributions of different character partitions
The mitochondrial sequence seems, overall, to have per-
formed better, providing finer resolution and better branch
support than white. For the same number of taxa (n¼ 14),
the COIþ II dataset had twenty-twomore parsimony informa-
tive characters (33%) than the white dataset. Overall,
COIþ II was only slightly more variable than white
(12.8% of parsimony informative sites over the total length
of the fragment for COIþ II vs 10.6% for white) but the
fragment used in the analysis of the mitochondrial gene was
longer by 97 bp.
Combination of characters
Combined datasets generated a better resolved tree for
the small taxon set (Fig. 2), but not if more species were
added (Fig. 4B). In the case of our study, 30% of missing
data in the white dataset was enough to generate more
equally parsimonious trees and a slight loss of resolution
in the consensus tree of the combined analysis when com-
pared with the trees generated by the complete COIþ II and
morphology (Fig. 3A, B), or by the combination of these
last two datasets (Fig. 4A). This is not surprising; Wiens
(1998) suggested that as much as 50% of missing data in
one character set of a combined analysis is unlikely to
T. albibasalis
T. genarum
T. furcatus
T. lividiventrisT. tarsalis
T. diaphanus
T. aculeipes
T. cunctans
T. hirtulus
T. innocuus
T. semicinereus
T. septentrionalis
T. spiniger
T. separ
T. longipes
T. nigrifrons
T. nigritellus
A. gibbera
A. cilipes
H. spinifemorata
diaphanusgroup
semicinereusgroup
(0.5, –0.2, 2.7)
(0.3, 0.7, 0)
3
42
1
(–0.4, –0.5, 2.9)(4, 0, 0)
7699
83
(2, –1, 0)
15
1
(10, 0, 5)
68
100
T. albibasalis
T. genarum
T. furcatus
T. lividiventrisT. tarsalis
T. diaphanus
T. aculeipes
T. cunctans
T. semicinereus
T. hirtulus
T. innocuusT. septentrionalis
T. spiniger
T. separ
T. longipes
T. nigrifronsT. nigritellus
A. gibbera
A. cilipes
H. spinifemorata
11
9(7.8, 0, 1.2)
100
16(15.3, –0.5, 1.2)
8
95
B(8, 0, 3)
11(7, 0, 4)
(2.9, 2.9, 2.2)
(14.3,, 2.7)
4(4, 0)2
(–0.7, 2.7)
3(2.4, 0.6)
17
6498
81
100
A2
(2, 0)
15(10, 5)
82
1009(7, 2)
100
10(6, 4)
100
(2.5, 2.5)5
88
9(8.3, 0.7)
99
1(0.9, 0.1)
100
100
100
Fig. 4. A, B, The results of combined analyses for the medium taxon set (n¼ 20). A, Maximum parsimony strict consensus of twenty-two
most parsimonious trees resulting from the combined analysis of cytochrome c oxidase subunits I and II and morphology; B, maximum
parsimony strict consensus of thirty-four most parsimonious trees resulting from the combined analysis of all taxa with data available for at
least one molecular data partition. The values above the branches refer to bootstrap values (from 100 replicates). The values below the
branches refer to Bremer support and partitioned Bremer support (cytochrome c oxidase subunits I and II, white, morphology).
Phylogenetic analysis of Thricops 405
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

decrease phylogenetic accuracy, but that study was based
on simulated datasets free of homoplasy and with the
branch length held constant across all branches of a tree.
Overall, the total evidence approach has allowed us to
increase our confidence in those nodes which were sup-
ported by all datasets, while also enabling us to determine
what parts of the tree were supported by each data parti-
tion. Based on the PBS values shown in Fig. 2, both genes
were generally more useful in resolving the basal nodes,
whereas most terminal resolution derived from morph-
ological characters.
Effects of taxon sampling
Zwickl & Hillis (2002) argued that an increase in taxon
sampling would generally result in more accurate phylo-
genetic hypotheses. This seems to be the case here, at least
T. genarumT. hakusanusT. furcatusT. l. lividiventrisT. l. plumbeusT. tatricusT. simplexT. calcaratusT. sudeticusT. pontiT. diaphanusT. nepalensisT. tarsalisT. aduncusT. r. himalayensisT. r. rufisquamusT. thudamensisT. aculeipesT. hirtulusT. culminumT. vaderiT. innocuusT. septentrionalisT. spinigerT. separT. fimbriatusT. villicrusT. longipesT. nigrifronsT. dianaeT. cunctansT. semicinereusT. nigriabdominalisT. rostratusT. coquillettiT. nigritellusT. villosusT. beckeriT. bukowskiiT. foveolatus
H. spinifemorata
A. cilipesA. gibbera
T. genarumT. hakusanusT. furcatusT. l. lividiventrisT. l. plumbeusT. tatricusT. simplexT. calcaratusT. sudeticusT. pontiT. diaphanusT. nepalensisT. tarsalisT. aduncusT. r. himalayensisT. r. rufisquamusT. thudamensisT. aculeipesT. cunctansT. hirtulusT. culminumT. innocuusT. septentrionalisT. spinigerT. separT. fimbriatusT. villicrusT. longipesT. nigrifronsT. nigritellusT. semicinereusT. nigriabdominalisT. coquillettiT. dianaeT. rostratusT. vaderiT. villosusT. beckeriT. bukowskiiT. foveolatusA. cilipesA. gibberaH. spinifemorata
A
diaphanusgroup
semicinereusgroup
1
1
1
4
1
21
1
21
3
32
1
11
1
1
21
21
21
1
1
1
1
1
1
2
3
1
62
99
57
6991
66
91
81
7288
60
5990
58
85
76
59
77
93
68
94
70
52
7091
78
99
75 8883
66
84
67
7080
91
63
88
99
56
T. albibasalis T. albibasalisB
Fig. 5. A, B, The results of an individual analysis of morphology and a combined analysis of all data for the large taxon set (n¼ 44). A,
Maximum parsimony strict consensus tree of six most parsimonious trees resulting from the analysis of morphology; B, maximum parsimony
strict consensus of 46 980 most parsimonious trees resulting from the combined analysis of all available data. The values above the branches
refer to bootstrap values (from 100 replicates). The values below the branches refer to Bremer support.
406 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

with the morphological data. The large taxon set generated
a consensus tree (Fig. 5A) which was better resolved than
those generated by the smaller datasets (Figs 1C, 3B).
Whereas all analyses based on the exemplar approach indi-
cated that Alloeostylus could regain its generic status, it is
now apparent that some taxa (T. foveolatus and T. bukowskii)
left out of the exemplar taxon set were critical. This
illustrates a disadvantage of the exemplar approach, in
which one is never certain to have sampled accurately the
full ‘tree diameter’ of a taxonomic group until a phylo-
geny is actually produced.
Conclusions
Our data support the monophyly of Thricops sensu Pont
(1986a). The synonymy of Alloeostylus with Thricops by Pont
(1986a) is also supported, as all species formerly included in
Alloeostylus belong to a derived clade nested within Thricops.
Although the data analysed here provided some import-
ant insights into the species-level relationships of Thricops,
additional data would help us to refine our phylogenetic
hypothesis. First, the sequences of COIþ II and white for
T. foveolatus and T. bukowskii would allow us to test the
basal relationships presented here. The inclusion of an
exemplar species of the T. r. rufisquamus branch might
also provide additional support to the basal relationships
of the diaphanus group. Finally, additional nucleotide data
from the mitochondrial genome or from more rapidly
evolving gene regions from the nuclear genome should
allow us to resolve relationships more fully within the semi-
cinereus group.
All sources of data used in this study proved useful
in understanding the basal relationships within the
genus Thricops. Because of the high congruence for all
datasets at the basal nodes, we are confident that the basal
placement of T. foveolatus (and possibly T. bukowskii),
even if supported only by morphological characters, is
reliable.
-
15:124:1
------
--
---
--
-
---
3:15:19:210:040:1
*
4:16:128:130:333:236:341:1
9:110:118:134:1
*
*
38:1
3:117:143:144:1
23:15:0
20:130:237:1
**
3:121:140:1
14:123:1
*8:019:1
15:1
5:&6:1
1:1 *
*
11:120:140:2
36:139:041:1
20:124:1
1:1
3:16:125:0
25:1
27:136:1
*
16:2*
42:1
16:1
25:0
7:0
*
*
1:1
7:0
24:0
8:041:0
T. d
iaph
anus
T. n
epal
ensi
s
T. a
lbib
asal
is
T. h
akus
anus
T. g
enar
um
T. fu
rcat
us
T. l.
livi
dive
ntris
T. l.
plu
mbe
us
T. ta
tric
us
T. s
impl
ex
T. c
alca
ratu
s
T. s
udet
icus
T. p
onti
T. a
cule
ipes
T. h
irtu
lus
T. c
ulm
inum
T. v
ader
i
T. d
iana
e
T. c
unct
ans
T. lo
ngip
es
T. n
igrif
rons
T. b
ecke
ri
A. g
ibbe
ra
A. c
ilipe
s
H. s
pini
fem
orat
a
T. ta
rsal
is
T. b
ukow
skii
T. fo
veol
atus
T. in
nocu
us
T. s
epte
ntrio
nalis
T. s
pini
ger
T. s
epar
T. fi
mbr
iatu
s
T. v
illic
rus
T. s
emic
iner
eus
T. n
igria
bdom
inal
is
T. r
ostr
atus
T. c
oqui
lletti
T. n
igrit
ellu
s
T. v
illos
us
T. a
dunc
us
T. r.
him
alay
ensi
s
T. r.
ruf
isqu
amus
T. th
udam
ensi
s1:129:133:236:240:1
*
*
*
*
14:124:1
* 12:124:1
8:&26:&
20:1
4:125:0
20:022:125:0
*
27:1
7:135:1
6:132:133:2
5:028:130:1*
3:143:1
15:1
8:025:0
**
13:114:2
25:0
4:16:1
*
*
6:114:330:433:234:1
*
26:1
5:18:125:131:133:139:1
**
2:1* -
-----
---
-
- --
--
------
--
-
--
--
- -
---
----
-
-
---
-
---
---
------ -
--
----
-------
---
--
----
-
-----
--
---
--
--
---
*
8:0-
*
*
*
25:& 6:&-
Fig. 6. One of six most parsimonious cladograms for the large morphological matrix. Uniquely derived character states are marked with an
asterisk, polymorphisms are denoted by ‘&’.
Phylogenetic analysis of Thricops 407
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414
Acknowledgements
We thank Brian Cassel (North Carolina State University)
for his technical support with the molecular sequencing and
John K. Moulton (University of Tennessee) for his assist-
ance in primer design. We also thank Shaun Winterton
(North Carolina State University) and Jeff Skevington
(Canadian National Collection of Insects) for advice and
suggestions related to combined phylogenetic analyses.
Brian Driscoll (McGill University) kindly allowed access
to his Macintosh computer. Scott Brooks (McGill Univer-
sity) and Brian Cassel provided fresh specimens for the
molecular analysis, and Jeff Cumming (Canadian National
Collection of Insects) and Nigel Wyatt (The Natural His-
tory Museum, London) gave us permission to extract DNA
from dry material housed in their respective institutions. We
are grateful to Adrian Pont (University Museum of Natural
History, Oxford), Michael Ackland (University Museum of
Natural History, Oxford), Bernhard Merz (Museum d’His-
toire Naturelle, Geneva) and Doreen Verner (Berlin) for
their help during the 2001 collecting expedition to Sweden,
and to Joelle Perusse (Yale University) for her help during
the 2001 collecting expedition to Mount Washington
(U.S.A.). The support of Nils Ake Andersson while staying
at the Abisko Scientific Research Station (Sweden) was also
greatly appreciated. We thank the Abisko National Park
Fig. 7. A–E, Male genitalia external structures, posterior view. A, Thricops calcaratus; B, Thricops furcatus; C, Thricops bukowskii;
D, Thricops hirtulus; E, Thricops rufisquamus rufisquamus. F, Azelia cilipes %, mesolobus, posterior view. G, H, Male sternite 5. G, Thricops
furcatus; H, Thricops hirtulus. epand¼ epandrium; meso¼mesolobus; sur¼ surstylus.
408 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

for permission to collect specimens. We thank Insect
Systematics and Evolution Supplements for permission to
reproduce Figs 1B, 2E, 3B, F, G, 4C, 10A, C, 11A, C,
16G, 19A, C, D, G, 22A, D, G, H, 36A, C, E, 41H and
43G from Savage (2003) . Figure 11, p. 1120 fromManual of
Nearctic Diptera, Volume 2, 1987, Agriculture and Agri-
Food Canada, was reproduced here with the permission of
the Minister of Public Works & Government Services
Canada, 2003. Laboratory space was provided at the
North Carolina State University, the Lyman Entomological
Museum and the Abisko Scientific Research Station.
Financial support was provided by NSERC Postgraduate
Scholarship (Canada), bourses d’etudes FQRNT (Quebec),
The Dipterology Fund, Abisko Scientific Research Station,
research grants to Terry A. Wheeler from NSERC, and
Brian M. Wiegmann from the U.S. National Science
Foundation.
References
Baker, R.H. & DeSalle, R. (1997) Multiple sources of character
information and the phylogeny of Hawaiian drosophilids.
Systematic Biology, 46, 654–673.Baker, R.H., Wilkinson, G.S. & DeSalle, R. (2001) Phylogenetic
utility of different types of molecular data used to infer
evolutionary relationships among stalk-eyed flies (Diopsidae).
Systematic Biology, 50, 87–105.Bremer, K. (1994) Branch support and tree stability. Cladistics, 6,
369–372.Brown, B., Emberson, R.M. & Paterson, A.M. (1999) Phylogeny of
‘Oxycanus’ lineages of hepialid moths from New Zealand
inferred from sequence variation in the mtDNA COI and II
gene regions. Molecular Phylogenetics and Evolution, 13,
463–473.Brown, J.M., Pellmyr, O., Thompson, J.N. & Harrison, R.G.
(1994) Phylogeny of Greya (Lepidoptera: Prodoxidae), based on
nucleotide sequence variation in mitochondrial cytochrome
Fig. 8. A–E, Male genitalia internal structures, lateral view. A, Thricops hirtulus; B, Thricops rufisquamus rufisquamus; C, Thricops bukowskii;
D, Thricops furcatus; E, Thricops calcaratus. d inf¼dorsal infold; epiph¼ epiphallus; hypd¼hypandrium; lat scl¼ lateral sclerite;
phap¼phallapodeme; preg¼ pregonite.
Phylogenetic analysis of Thricops 409
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

oxidase I and II: congruence with morphological data.
Molecular Biology and Evolution, 11, 128–141.Bull, J.J., Huelsenbeck, J.P., Cunningham, C.W., Swofford, D.L. &
Waddel, P.J. (1993) Partitioning and combining data in
phylogenetic analyses. Systematic Biology, 42, 384–397.Chippindale, P.T. & Wiens, J.J. (1994) Weighting, partitioning and
combining characters in phylogenetic analyses. Systematic
Biology, 43, 278–287.Elberling, H. & Olesen, J.M. (1999) The structure of a high latitude
plant-flower visitor system: the dominance of flies. Ecography,
22, 314–323.Farris, J.S., Kallersjo, M., Kluge, A.G. & Bult, C. (1995a)
Constructing a significance test for incongruence. Systematic
Biology, 44, 570–572.Farris, J.S., Kallersjo, M., Kluge, A.G. & Bult, C. (1995b) Testing
significance of incongruence. Cladistics, 10, 315–319.Hennig, W. (1962) 63b. Muscidae. Die Fliegen der Palaearktischen
Region 7 (2) (ed. by E. Lindner), p. 625. Schweizerbart’sche,
Stuttgart.Hennig, W. (1965) Vorarbeiten zu einem phylogenetischen System
der Muscidae (Diptera: Cyclorrhapha). Stuttgarter Beitrage zur
Naturkunde, 141, 1–100.Hillis, D.M., Mable, B.K., Larson, A., Davis, S.K. & Zimmer, E.A.
(eds) (1996) Chapter 9. Nucleic acids IV: sequencing and
cloning. Molecular Systematics, 2nd edn, pp. 321–381. Sinauer
Associates, Sunderland, Massachusetts.Huckett, H.C. (1965a) The Muscidae of northern Canada, Alaska,
and Greenland (Diptera). Memoirs of the Entomological Society
of Canada, 42, 1–369.Huckett, H.C. (1965b) Family Muscidae. A Catalog of Diptera of
America North of Mexico (ed. by A. Stone, C. W. Sabrowsky,
W. W. Wirth, R. H. Foote and J. R. Coulson), pp. 869–914.
Agriculture Handbook no. 276. United States Department of
Agriculture, Washington, DC.Kluge, A.G. (1998) Total evidence or taxonomic congruence:
cladistic or consensus classification. Cladistics, 14, 151–158.Kocher, T.D., Thomas, W.K., Meyer, A., Edwards, S.V., Paabo, S.,
Villablanca, F.X. & Wilson, A.C. (1989) Dynamics of
mitochondrial DNA evolution in animals: amplification and
sequencing with conserved primers. Proceedings of the National
Academy of Sciences USA, 86, 6196–6200.Malloch, J.R. (1921) A synopsis of the anthomyiid genus
Trichopticus Rondani (Diptera). Canadian Entomologist, 52
[1920], 271–274.Michelsen, V. (1978) Cryptophyra lundbecki n. gen. et n. sp. from
north Europe, with some notes on allied hydrotaeine genera
(Diptera: Muscidae). Entomologica Scandinavica, 9, 85–92.Novacek, M.J. (1992a) Fossils as critical data for phylogeny.
Extinction and Phylogeny (ed. by M. J. Novacek and Q. D.
Wheeler), pp. 46–88. Colombia University Press, New York.Novacek, M.J. (1992b) Fossils, topologies, missing data, and the
higher level phylogeny of eutherian mammals. Systematic
Biology, 41, 58–73.Pont, A.C. (1986a) Family Muscidae. Catalogue of Palaearctic
Diptera, Vol. 11. Scathophagidae–Hypodermatidae (ed. by
A. Soos and L. Papp), pp. 57–215. Academiai Kiado, Budapest.Pont, A.C. (1986b) A revision of the Fanniidae and Muscidae
described by J. W. Meigen (Insect: Diptera). Annalen des
Naturhistorischen Museums in Wien, 87B, 197–253.
Pont, A.C. (1993) Observation on anthophilous Muscidae and
other Diptera (Insects) in Abisko National Park, Sweden.
Journal of Natural History, 27, 631–643.Ringdahl, O. (1947) Bestimmungstabelle der mir bekannten
europaischen und nordamerikanischen Arten von Alloeostylus
und Lasiops (Dipt: Musc.). Opuscula Entomologica, 12, 90–95.Rondani, C. (1856) Dipterologiae Italicae Prodromus. Vol. I.
Genera Italica ordinis Dipterorum ordinatim disposita et distincta
et in familias et stirpes aggregata. A. Stoschi, Parma.Savage, J. (2003) Revision of the genus Thricops Rondani (Diptera:
Muscidae). Insect Systematics and Evolution Supplement, 61,
1–143.Savage, J. & Wheeler, T.A. (in press) Phylogeny of the Azeliini
(Diptera: Muscida). Studia Dipterologica, in press.Schnabl, J. (1888a) Alloeostylus, nov. gen. Anthomyidarum.
Entomologisches Nachrichten Berlin, 14, 49–50.Schnabl, J. (1888b) Nachtrag zur Gattung Alloeostylus Schnabl.
Entomologisches Nachrichten Berlin, 14, 82–83.Schnabl, J. (1888c) Hera, nov. gen. Anthomyidarum, (0Hra,
Mutter der Gracien). [Aricia, pt. auctt. Yetodesia, pt. auctt.
Hyetodesia, pt. auctt. Lasiops, pt. Mg. Trichophthicus, pt.
Mydaea, pt.]. Entomologisches Nachrichten Berlin, 14, 113–120.Schnabl, J. (1889) Contributions a la faune dipterologique. Trudy
Russkago Entomologicheskago Obshchestva [¼ Horae Societatis
Entomologicae Rossicae], 23, 313–347.Skevington, J.H. & Yeates, D.K. (2000) Phylogeny of the
Syrphoidea (Diptera) inferred from mtDNA sequences and
morphology with particular reference to classification of the
Pipunculidae (Diptera). Molecular Phylogenetics and Evolution,
16, 212–224.Skidmore, P. (1985) The biology of the Muscidae of the world.
Series Entomologica, Vol. 29. Dr W. Junk, The Hague.Sorenson, M.D. (1999) Treerot, Version 2. Boston University,
Boston.Stahls, G. & Nyblom, K. (2000) Phylogenetic analysis of the genus
Cheilosia (Diptera, Syrphidae) using mitochondrial COI
sequence data. Molecular Phylogenetics and Evolution, 15,
235–241.Swofford, D.L. (2002) PAUP*: Phylogenetic Analysis Using
Parsimony (* and Other Methods), Version 4.0b10. Sinauer,
Sunderland, Massachusetts.Walsh, P.S., Metzger, D.A. & Higuchi, R. (1991) Chelex1100 as a
medium for simple extraction of DNA for PCR-based typing
from forensic material. Biotechniques, 10, 506–513.Wiens, J.J. (1998) Does adding characters with missing data
increase or decrease phylogenetic accuracy? Systematic Biology,
47, 625–640.Wilkinson, M. (1995) Coping with abundant missing entries in
phylogenetic inference using parsimony. Systematic Biology, 44,
501–514.Wilkinson, M. & Benton, M.J. (1995) Missing data and rhynch-
osaur phylogeny. Historical Biology, 10, 137–150.Zwickl, D.J. & Hillis, D.M. (2002) Increased taxon sampling
greatly reduces phylogenetic error. Systematic Biology, 51,
588–598.
Accepted 14 December 2003
410 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

Appendix 1
List of characters and character states for morphological
data.
1. Eye (male): (0) bare; (1) setulose.
2. Gena: (0) very narrow, with lower eye margin strongly
extending ventrally below level of vibrissae (Fig. 9B); (1)
relatively broad, with lower eye margin not or only
weakly extending ventrally below level of vibrissae
(Fig. 9A).
3. Interfrontal bristles (female): (0) present; (1) absent.
4. Aristal pubescence: (0) short; (1) plumose.
5. Presutural acrostichal bristles: (0) present; (1) absent.
6. Dorsocentral bristles: (0) 2þ 4; (1) 2þ 3.
7. Prealar bristles: (0) weak,<0.5� length of second
notopleural bristle; (1) strong,>0.75� length of second
notopleural bristle.
8. Lower katepisternal bristle: (0) absent or very weak; (1)
present.
9. Metathoracic spiracle: (0) small and round (Fig. 9D); (1)
large and triangular (Fig. 9C); (2) small and triangular
(Fig. 9E).
10. Setulae on metathoracic spiracle: (0) simple (Fig. 9D, E);
(1) branched (Fig. 9C).
11. Scutellum colour: (0) dark; (1) mostly to completely
yellow.
12. Basal half of midfemur (male): (0) not swollen; (1)
swollen (Fig. 9F).
13. Apical third of midfemur (male): (0) without dense
brush of hairs on apical third; (1) with dense brush of
hairs on apical third.
14. Posteroventral surface of midfemur (male): (0) with
hairs only; (1) with one to two rows of recurved
spinules on basal half; (2) with a row of very long and
Fig. 9. A, B, Head. A, Thricops vaderi ?; B, Hydrotaea anxia ? (source: Manual of Nearctic Diptera, Vol. 2, 1987, p. 1120, fig. 11). C–E,
Posterior thoracic spiracle. C, Thricops diaphanus/; D, Thricops vaderi /; E, Thricops furcatus/. F, Thricops spiniger ?, midfemur, anterior
view; G, Thricops semicinereus ?, fore tarsomeres 2–5, anterior view; H, Thricops diaphanus ?, hind tibia, anterior view; I, Thricops
rufisquamus rufisquamus ?, fore tibia, posterior view. hlt¼halter; ktg¼ katatergite; p spr¼posterior spiracle.
Phylogenetic analysis of Thricops 411
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

strong straight bristles; (3) bare except for a single
strong spine in middle.
15. Fore tibia (female): (0) without posteroventral bristle;
(1) with one to two posteroventral bristles.
16. Apical third of fore tibia (male): (0) without strong
spines on apical third; (1) with one row of six to eight
strong posteroventral spines; (2) with two strong
posterior and one to three strong posteroventral spines.
17. Posteroventral surface of fore tibia (male): (0) without
short flat spines; (1) with two rows of short flat spines
(Fig. 9I).
18. Midtibia (at least in male): (0) without one to two
posteroventral bristles; (1) with one to two poster-
oventral bristles.
19. Posterodorsal surface of hind tibia (male): (0) without a
fringe of long bristles; (1) with a fringe of six to eight
long bristles.
20. Ventral surface of hind tibia (male): (0) without apical
or preapical spur; (1) with apical or preapical spur
(Fig. 9H).
21. Apicoventral surface of hind tibia (male): (0) without a
fringe of spinules; (1) with a fringe of short spinules.
22. Fore tarsomere 4 (male): (0) without a pad of poster-
oventral spinules; (1) with a pad of posteroventral
spinules (Fig. 9G).
23. Fore tarsomere 5 (male): (0) flat and rounded; (1)
pointed and projecting dorsally (Fig. 9G).
24. Midtarsomeres 3–5 (male): (0) without a fringe of pale
posteroventral setulae; (1) with a distinct fringe of pale
posteroventral setulae.
25. Halter colour (male): (0) yellow; (1) dark.
26. Sternite 1 (male): (0) bare; (1) haired.
27. Length of sternite 5 (male): (0) short, <1.3� as long as
wide (Fig. 7G); (1) long, >1.4� as long as wide
(Fig. 7H).
28. Shape of surstyli (male): (0) short and bent medially
(Fig. 7D, E); (1) long and projecting ventrally (Fig.
7A–C).
29. Shape of apical portion of surstyli in posterior view
(male): (0) not very broad; (1) very broad with apex
flat medially (Fig. 7E).
30. Shape of mesolobus (male): (0) broad and flat, about as
high as long (Fig. 7D, E); (1) V-shaped (Fig. 7B); (2)
rectangular and strongly cleft medially (Fig. 7A); (3)
rectangular, split but uncleft (Fig. 7C); (4) broad with
raised central area, about as high as long (Fig. 7F).
31. Lateral sclerites of distiphallus (male): (0) narrow or
weakly developed; (1) well sclerotized and flared
apically (Fig. 8A–E).
32. Lateral sclerites of distiphallus (male): (0) fused
dorsally with one another on basal half only,
not forming a tube; (1) fused dorsally with one
another along most of the dorsal margin, forming a
long tube.
33. Dorsal infold of distiphallus (dorsal sclerite of Savage,
2003) (male): (0) large and very strongly sclerotized
with long black spines and weakly invaginated; (1)
large, with or without spinules and deeply invaginated;
(2) reduced and membranous, without spinules and
weakly invaginated.
34. Dorsal infold of distiphallus (male): (0) partially to
completely split longitudinally; (1) completely fused.
35. Length of epiphallus (male): (0) <0.5� length of
phallapodeme (Fig. 8C); (1) >0.75�length of phallapo-
deme (Fig. 8B).
36. Shape of pregonite (male): (0) broad and short, with
apex blunt (Fig. 8D); (1) broad at least on basal two
thirds and with apex projecting laterally (Fig. 8A); (2)
long and narrow, with apex bent ventrally at a 90�
angle (Fig. 8B); (3) long and narrow, with apex rounded
(Fig. 8C).
37. Apex of pregonite (male): (0) mostly well sclerotized,
never jagged; (1) strongly desclerotized and jagged
(Fig. 8E).
38. Length of hypandrial arms (male): (0) short; (1) very
long (Fig. 8D).
39. Attachment of hypandrial arms (male): (0) attached to
surstyli before apex; (1) attached to surstyli at apex.
40. Tergite 8 (female): (0) split in two relatively parallel rods
(Fig. 10B, C); (1) connected medially and M-shaped
(Fig. 10A); (2) connected anteriorly and posteriorly,
forming a hollow rectangle (Fig. 10D).
41. Spines of tergite 8 (female): (0) straight (Fig. 10A); (1)
weakly to strongly recurved (Fig. 10B).
42. Sternite 8 (female): (0) divided into two mostly parallel
rods (Fig. 10F); (1) fused posteriorly and V-shaped
(Fig. 10G).
43. Tergite 10 (female): (0) bare (Fig. 10A, B, D); (1)
haired at least on apical half (Fig. 10C).
44. Cercus (female): (0) bare or with setulae only on apical
half; (1) densely setulose over most of the ventral
surface (Fig. 10E).
412 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414

Fig. 10. A–D, Female genitalia, dorsal view. A, Thricops furcatus; B, Thricops hirtulus; C, Thricops tarsalis; D, Thricops diaphanus. E,
Thricops rufisquamus rufisquamus &, cercus, ventral view. F, G, Female genitalia, ventral view. F, Thricops hirtulus; G, Thricops spiniger.
cerc¼ cercus; tg 8¼ tergite 8; tg 10¼ tergite 10; st 8¼ sternite 8; st 10¼ sternite 10.
Phylogenetic analysis of Thricops 413
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414
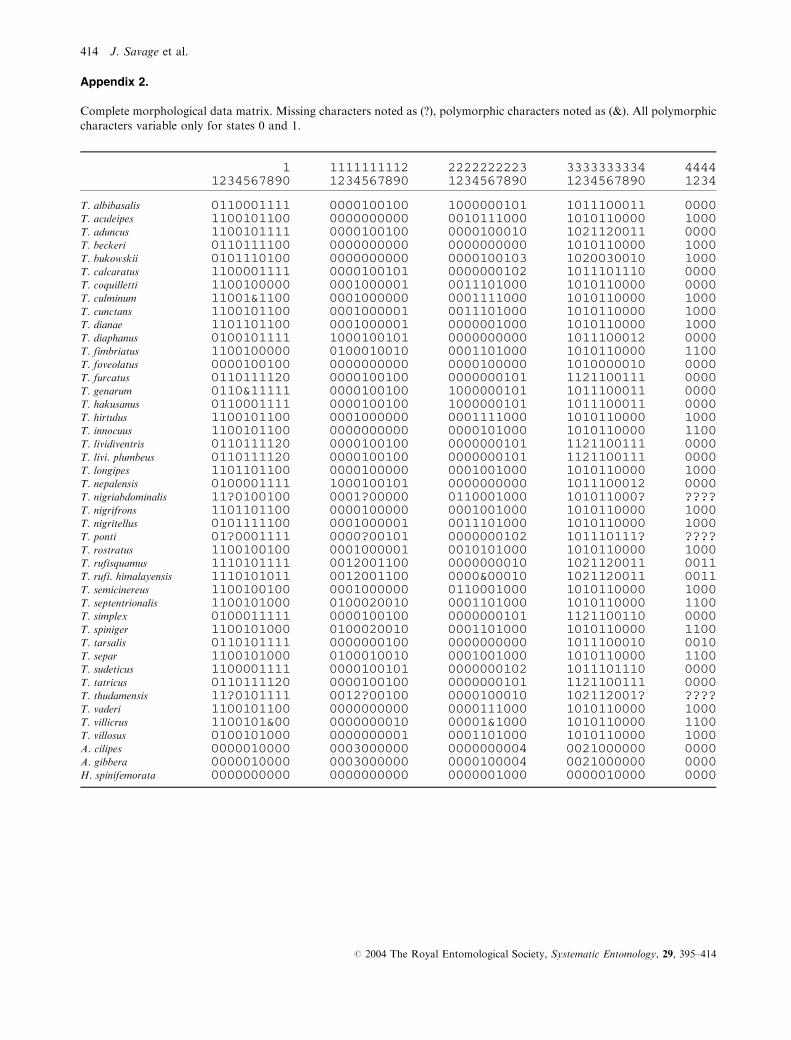
Appendix 2.
Complete morphological data matrix. Missing characters noted as (?), polymorphic characters noted as (&). All polymorphic
characters variable only for states 0 and 1.
12345678911234567890
11111111121234567890
22222222231234567890
33333333341234567890
44441234
T. albibasalis 0110001111 0000100100 1000000101 1011100011 0000T. aculeipes 1100101100 0000000000 0010111000 1010110000 1000T. aduncus 1100101111 0000100100 0000100010 1021120011 0000T. beckeri 0110111100 0000000000 0000000000 1010110000 1000T. bukowskii 0101110100 0000000000 0000100103 1020030010 1000T. calcaratus 1100001111 0000100101 0000000102 1011101110 0000T. coquilletti 1100100000 0001000001 0011101000 1010110000 0000T. culminum 11001&1100 0001000000 0001111000 1010110000 1000T. cunctans 1100101100 0001000001 0011101000 1010110000 1000T. dianae 1101101100 0001000001 0000001000 1010110000 1000T. diaphanus 0100101111 1000100101 0000000000 1011100012 0000T. fimbriatus 1100100000 0100010010 0001101000 1010110000 1100T. foveolatus 0000100100 0000000000 0000100000 1010000010 0000T. furcatus 0110111120 0000100100 0000000101 1121100111 0000T. genarum 0110&11111 0000100100 1000000101 1011100011 0000T. hakusanus 0110001111 0000100100 1000000101 1011100011 0000T. hirtulus 1100101100 0001000000 0001111000 1010110000 1000T. innocuus 1100101100 0000000000 0000101000 1010110000 1100T. lividiventris 0110111120 0000100100 0000000101 1121100111 0000T. livi. plumbeus 0110111120 0000100100 0000000101 1121100111 0000T. longipes 1101101100 0000100000 0001001000 1010110000 1000T. nepalensis 0100001111 1000100101 0000000000 1011100012 0000T. nigriabdominalis 11?0100100 0001?00000 0110001000 101011000? ????T. nigrifrons 1101101100 0000100000 0001001000 1010110000 1000T. nigritellus 0101111100 0001000001 0011101000 1010110000 1000T. ponti 01?0001111 0000?00101 0000000102 101110111? ????T. rostratus 1100100100 0001000001 0010101000 1010110000 1000T. rufisquamus 1110101111 0012001100 0000000010 1021120011 0011T. rufi. himalayensis 1110101011 0012001100 0000&00010 1021120011 0011T. semicinereus 1100100100 0001000000 0110001000 1010110000 1000T. septentrionalis 1100101000 0100020010 0001101000 1010110000 1100T. simplex 0100011111 0000100100 0000000101 1121100110 0000T. spiniger 1100101000 0100020010 0001101000 1010110000 1100T. tarsalis 0110101111 0000000100 0000000000 1011100010 0010T. separ 1100101000 0100010010 0001001000 1010110000 1100T. sudeticus 1100001111 0000100101 0000000102 1011101110 0000T. tatricus 0110111120 0000100100 0000000101 1121100111 0000T. thudamensis 11?0101111 0012?00100 0000100010 102112001? ????T. vaderi 1100101100 0000000000 0000111000 1010110000 1000T. villicrus 1100101&00 0000000010 00001&1000 1010110000 1100T. villosus 0100101000 0000000001 0001101000 1010110000 1000A. cilipes 0000010000 0003000000 0000000004 0021000000 0000A. gibbera 0000010000 0003000000 0000100004 0021000000 0000H. spinifemorata 0000000000 0000000000 0000001000 0000010000 0000
414 J. Savage et al.
# 2004 The Royal Entomological Society, Systematic Entomology, 29, 395–414
Top Related
Copyright © 2022 FDOKUMEN

