







Bahasa
Halaman
Hukum


Article
Non-canonical NF-kB Anta
gonizes STING Sensor-Mediated DNA Sensing in RadiotherapyGraphical Abstract
Highlights
d Canonical NF-kB pathway is required for radiation-induced
anti-tumor immunity
d Non-canonical NF-kB deficiency promotes anti-tumor
immunity after radiotherapy
d Non-canonical NF-kB pathway inhibits radiation-induced
STING-mediated type I IFNs
d Inhibiting non-canonical NF-kB pathway potentiates the
therapeutic effect of radiation
Hou et al., 2018, Immunity 49, 490–503September 18, 2018 ª 2018 Elsevier Inc.https://doi.org/10.1016/j.immuni.2018.07.008
Authors
Yuzhu Hou, Hua Liang, Enyu Rao, ...,
Ainhoa Arina, Ralph R. Weichselbaum,
Yang-Xin Fu
[email protected] (R.R.W.),[email protected](Y.-X.F.)
In Brief
It is known that the NF-kB pathway plays
a crucial role in supporting tumor
initiation, progression, and the
radioresistance of tumor cells. Hou et al.
demonstrate that the deficiency of non-
canonical NF-kB, but not canonical
NF-kB, promotes radiation-induced anti-
tumor immunity by regulating the STING-
mediated type I IFN expression.

Immunity
Article
Non-canonical NF-kB Antagonizes STINGSensor-Mediated DNA Sensing in RadiotherapyYuzhu Hou,1 Hua Liang,1 Enyu Rao,1,2 Wenxin Zheng,1 Xiaona Huang,1 Liufu Deng,1,3 Yuan Zhang,1 Xinshuang Yu,4
Meng Xu,1 Helena Mauceri,1 Ainhoa Arina,1 Ralph R. Weichselbaum,1,* and Yang-Xin Fu5,6,*1Ludwig Center for Metastasis Research, Department of Radiation and Cellular Oncology, The University of Chicago, Chicago, IL 60637, USA2Cancer Institute, Xuzhou Medical University, Xuzhou, Jiangsu, China3Shanghai Institute of Immunology; Department of Immunology and Microbiology, Shanghai Jiao Tong University School of Medicine, China4Department of Radiation Oncology, Shandong Provincial Qianfoshan Hospital, Shandong University, Jinan, China5Department of Pathology, University of Texas Southwestern Medical Center, Dallas, TX 75235-9072, USA6Lead Contact*Correspondence: [email protected] (R.R.W.), [email protected] (Y.-X.F.)
https://doi.org/10.1016/j.immuni.2018.07.008
SUMMARY
The NF-kB pathway plays a crucial role in supportingtumor initiation, progression, and radioresistance oftumor cells. However, the role of the NF-kB pathwayin radiation-induced anti-tumor host immunity re-mains unclear. Here we demonstrated that inhibitingthe canonical NF-kB pathway dampened the thera-peutic effect of ionizing radiation (IR), whereas non-canonical NF-kB deficiency promoted IR-inducedanti-tumor immunity. Mechanistic studies revealedthat non-canonical NF-kB signaling in dendritic cells(DCs) was activated by the STING sensor-dependentDNA-sensing pathway. By suppressing recruit-ment of the transcription factor RelA onto the Ifnbpromoter, activation of the non-canonical NF-kBpathway resulted in decreased type I IFN expression.Administration of a specific inhibitor of the non-ca-nonical NF-kB pathway enhanced the anti-tumor ef-fect of IR in murine models. These findings reveal thepotentially interactive roles for canonical and non-canonical NF-kB pathways in IR-induced STING-IFN production and provide an alternative strategyto improve cancer radiotherapy.
INTRODUCTION
Radiotherapy (RT) is widely used as a primary treatmentmodality
for cancer, and the induction of damage to the tumor or to tumor
stroma was long thought to be its major mode of action (Prise
and O’Sullivan, 2009). Recently, however, increasing evidence
demonstrates that ionizing radiation (IR) induces tumor-specific
immunity (Reits et al., 2006) and that anti-tumor immunity is
required for the full anti-tumor effect of RT (Lee et al., 2009). It
is now recognized that the infiltration and priming of CD8+
T cells play a critical role in the efficacy of RT (Lee et al., 2009;
Lugade et al., 2005; Takeshima et al., 2010). DCs have been re-
ported to be activated by local IR and required for CD8+ T cell
activation and contribute to tumor control by RT (Gupta et al.,
490 Immunity 49, 490–503, September 18, 2018 ª 2018 Elsevier Inc.
2012). These antitumor immune responses have been found to
be highly dependent on type I interferon (IFN) signaling (Burnette
et al., 2011). Type I IFNs promote dendritic cell (DC) function by
stimulating their capacity to process and present antigens as
well as to promote DC migration toward lymph nodes (Gardner
and Ruffell, 2016; Zitvogel et al., 2015). Stimulator of interferon
genes (STING) is an endoplasmic-reticulum-associated protein
that activates transcription of the type I IFN gene, via a STING-
TBK kinase-IRF3 transcription factor-NF-kB signal transduction
pathway (Weichselbaum et al., 2017). Our previous studies
demonstrated that STING-dependent cytosolic DNA sensing
was required for IR-induced anti-tumor immunity by regulating
type I IFN expression (Deng et al., 2014b). However, emerging
research also demonstrates that STING-induced IFN production
by IR also promotes innate immune suppression by recruiting
monocytic myeloid-derived suppressor cells (MDSCs) into the
tumor microenvironment (Liang et al., 2017). Thus, IR not only
kills tumor cells directly, but also promotes innate immunity as
well as adaptive immune responses via the STING-mediated
DNA-sensing pathway. These immune responses induced by
IR have important implications for control of localized cancers,
as well as control of metastatic disease through the immune-
mediated abscopal effect. Therefore, elucidating the interaction
of IR and the immune systemmay have amajor impact on cancer
treatment.
The mammalian NF-kB family of transcription factors,
including RelA, c-Rel, RelB, NF-kB1 (p50 and its precursor
p105), and NF-kB2 (p52 and its precursor p100), play a central
role in the immune system. The most well-studied activation
pathway is the canonical NF-kBpathway, whichmainly impinges
upon RelA-p50 and c-Rel-p50 heterodimers (Hayden and
Ghosh, 2008; Sun, 2012; Vallabhapurapu and Karin, 2009). The
activation of the non-canonical NF-kB pathway pivots on activa-
tion of RelB-p52 heterodimers in response to a subset of TNF
family members, including CD40L, LTab, BAFF, RANKL, and
TWEAK. In the presence of these ligands, cIAP1 and 2 degrades
the TRAF3 ubiquitin ligase and NIK kinase stabilization is
achieved. NIK then phosphorylates IKKa and IKKa phosphory-
lates p100, which is important for the subsequent ubiquitination
and partial degradation of p100 by the proteasome to form p52.
RelB-p52 heterodimers then translocate into the nucleus to
regulate non-canonical NF-kB target genes (Sun, 2012).

Figure 1. Host Canonical NF-kB Is Required for Anti-tumor Immunity Induced by Ionizing Radiation
(A–E) B6 mice were inoculated s.c. with MC38 cells (A, C, D, and E) or B16-SIY cells (B) on day 0. On day 10, tumors received one dose of 20 Gy ionizing radiation
(IR) and 6 mg/kg JSH-23 was given by gavage daily starting 1 day before radiation for 14 days.
(A and B) Tumor growth was monitored after radiation.
(C) Seven days after IR, tumors were removed and digested. The frequency of CD8+ T cells in tumor was analyzed by flow cytometry.
(D and E) Four days after IR, tumors were removed and digested. The expressions of I-Ab (D) and CD80 (E) on CD45+CD11c+ DCs were analyzed by flow
cytometry.
(F) BMDCs were co-cultured with irradiated or non-irradiated MC38-SIY cells in presence or absence of 25 mM JSH-23. After 6–8 hr, CD11c+ DCs were isolated
and co-cultured with CD8+ T cells from naive 2C mice for 3 days and the cross-priming activity of BMDCs were analyzed by ELISPOT assays.
(legend continued on next page)
Immunity 49, 490–503, September 18, 2018 491

The canonical NF-kB signaling pathway has been extensively
studied in the context of cancer initiation, progression, and
response to RT and chemotherapy (chemo) in both mice and hu-
mans (Erstad and Cusack, 2013; Perkins, 2012; Shishodia and
Aggarwal, 2004). However, the involvement of the non-canonical
NF-kB pathway in cancer biology, and in response to RT, is less
well defined. Given that treatment with RT induces NF-kB activa-
tion in certain cancer cells and that the activation of NF-kB is
associated with the development of radioresistance (Erstad
and Cusack, 2013), NF-kB pathways have been considered as
a clinical target to counteract radio- and chemo-resistance.
However, the role of the interaction of canonical and non-canon-
ical NF-kBpathways in IR-induced anti-tumor immunity is largely
unknown. An understanding the role andmechanism of NF-kB in
RT may be of great value in designing therapeutic strategies that
take advantage of host response in the context of RT.
In this study, we demonstrated that the canonical NF-kB
pathway was required for IR-induced anti-tumor immune re-
sponses, suggesting that it is not a good therapeutic target to
enhance the effect of IR. In contrast, the non-canonical NF-kB
pathway negatively regulated RT-induced anti-tumor immunity
through STING-dependent DC activation. We demonstrated
that irradiated tumor cells activated the non-canonical NF-kB
pathway in DCs in a STING-dependent manner and that the
activated non-canonical NF-kB pathway controlled the anti-
tumor functions of DCs and CD8+ T cells by regulating type I
IFN expression. Importantly, inhibition of non-canonical NF-kB
signaling with p52-RelB nuclear translocation blockade
enhanced the therapeutic effect of RT. Thus, our findings
demonstrate that the non-canonical NF-kB pathway inhibits
IR-induced innate immune sensing and the resulting anti-tumor
immunity. These findings provide a fresh direction for developing
therapeutic strategies targeting NF-kB pathways.
RESULTS
Host Canonical NF-kB Signaling Is Required for IR-Induced Anti-tumor ImmunityTo determine whether inhibition of the NF-kB pathway promotes
the therapeutic effect of IR, we investigated tumor growth after
IR in the presence of JSH-23, a canonical NF-kB pathway inhib-
itor. JSH-23 abrogated the anti-tumor effect of IR, which sug-
gested that the canonical NF-kB pathway signaling was required
for the therapeutic effect of IR (Figures 1A and 1B). Analysis of
tumor-infiltrating immune cells profile revealed that the fre-
quency of CD8+ T cells increased in irradiated tumors and
decreased in the presence of JSH-23 (Figure 1C). Tumor-infil-
trating DCs showed lower MHC-II and CD80 expression in
JSH-23-treated mice than control mice (Figures 1D and 1E),
which suggested that JSH-23 might alter the antigen-presenting
capacity of DCs. Furthermore, our results here indicated that ca-
nonical NF-kB pathway inhibition dampened the cross-priming
capacity of DCs after RT (Figure 1F). Next, we employed
(G and H) Relaf/f mice and Itgax-cre Relaf/f mice were inoculated with MC38 cells (
IR. Tumor growth was monitored after radiation.
Representative data are shown from three experiments (one experiment for B an
*p < 0.05, **p < 0.01, and ***p < 0.001.
492 Immunity 49, 490–503, September 18, 2018
DC-specific RelA-deficient mice Itgax-cre Relaf/f and monitored
the tumor growth after IR. The results showed that IR controlled
tumor growth in control Relaf/f mice but not in Itgax-cre Relaf/f
mice, which suggested that RelA deficiency in DCs dampened
the anti-tumor effect of IR (Figures 1G and 1H).
Although canonical NF-kB signaling is documented to facili-
tate radioresistance in tumor cells, our results demonstrated
that inhibiting canonical NF-kB dampened the therapeutic effect
of RT by regulating IR-induced anti-tumor immunity. The canon-
ical NF-kB pathway signaling in DCs was required for the mani-
festation of IR-induced therapeutic effects.
Host Non-canonical NF-kB Signaling DeficiencyEnhances the Therapeutic Effect of Ionizing RadiationTo investigate the involvement of the non-canonical NF-kB
pathway in host response to IR, we employed Itgax-cre Relbf/f
(DC-deficient) or Lyz2-cre (myeloid-deficient) mice which
showed no symptoms of autoimmune disease or premature
death found in whole body-deficient mice. Using Relbf/f mice
as controls, we monitored MC38 tumor growth after a single IR
dose (20 Gy). The results showed that the anti-tumor effect of
RT was enhanced in Itgax-cre Relbf/f mice compared with that
of Relbf/f mice (Figures 2A and S1A). A similar result was
observed using a B16-SIY tumor model (Figures S1B and
S1C), which suggested that the role of RelB in the anti-tumor ef-
fect of RT is not restricted to a specific tumor histology. In
contrast, the tumor growth in Lyz2-cre Relbf/f mice was compa-
rable with control mice following RT (Figures S1D and S1E),
which suggested that RelB in DCs, but not myeloid cells (mono-
cytes, mature macrophages, and granulocytes), plays an inhibi-
tory role in IR-induced tumor control.
RelB is promiscuous in choosing binding partners depending
on cell type and stimuli. Instead of its conventional partner,
p52, it was reported that RelB could also promote DC activation
as a RelB-p50 dimer regulated by the canonical NF-kB pathway
(Shih et al., 2012). To determinewhether RelB in DCs controls the
therapeutic effect of RT through the non-canonical NF-kB
pathway, we employed NF-kB2-deficient (Nfkb2�/�) mice on a
Rag1�/� background. This genetic cross allowed us to focus
on the role of NF-kB2 in DCs and rule out the effect of intrinsic
defects after depletion of NF-kB2 in B cells (Caamano et al.,
1998). T cells were reconstituted by adoptively transferring
wild-type CD3+ T cells into a Rag1�/� background. The resulting
tumor growth showed that NF-kB2 deficiency led to enhanced IR
response compared with Nfkb2+/+ mice (Figures 2B and S1F).
Together, these data demonstrated that the host non-canonical
NF-kB pathway negatively regulates the therapeutic efficiency
of RT.
The Non-canonical NF-kB Pathway NegativelyRegulates IR-Induced Anti-tumor ImmunityTo determine whether IR induces stronger immune responses in
mice with deficient non-canonical NF-kB signaling, we analyzed
G) or B16-SIY cells (H) on day 0. On day 10, tumors received one dose of 20 Gy
d H) conducted with 4–5 mice per group. Data are represented as mean ± SD;

Figure 2. Non-canonical NF-kB Deficiency
Enhances Adaptive Immunity via Augmenting
DC Priming Function after IR
(A) Relbf/f mice and Itgax-cre Relbf/f mice were
inoculated with MC38 cells on day 0. On day 10,
tumors locally received one dose of 20 Gy IR. Tumor
growth was monitored after radiation.
(B) Nfkb2+/+ and Nfkb2�/� mice, bred to Rag1�/�
background, were inoculated with MC38 cells on
day 0. On day 10, tumors locally received one dose
of 20 Gy IR and the mice were transferred i.v. with
1 3 107 CD3+ T cells isolated from naive WT mice.
Tumor growth was monitored after radiation.
(C) MC38-SIY tumor model were established in
Relbf/f mice and Itgax-cre Relbf/f mice and treated
with IR as described in (A). On day 4 post IR, tumors
were removed and digested. The CD11c+ DCs were
sorted and then co-cultured with CD8+ T cells from
naive 2C mice for 3 days and the cross-priming ac-
tivity of tumor infiltrating DCs were analyzed by
ELISPOT assays.
(D) BMDCs from Relbf/f mice and Itgax-cre Relbf/f
mice were used for co-culture with irradiated or non-
irradiated MC38-SIY cells. Subsequently cross-
priming activity of purified BMDCs was analyzed by
ELISPOT assays as described in Figure 1F.
(E–G) MC38 tumor model were established in Relbf/f
mice and Itgax-cre Relbf/f mice and treated with IR as
described in (A).
(E) On day 7 post IR, tumors were removed and the
frequency of infiltrating CD8+ T cells was analyzed by
flow cytometry.
(F) On day 7 post IR, tumor-draining lymph nodes
(TDLNs) were removed and digested. Tumor anti-
gen-specific CD8+ T cell function was measured by
ELISPOT assays by co-culturing purified TDLNCD8+
cells with IFN-g-treated MC38 tumor cells.
(G) 200 mg anti-CD8 mAb was administered i.p. in
tumor bearing mice every 3 days for a total of three
times starting from the day of radiation. Tumor
growth was monitored after radiation.
(H) Nfkb2+/+ and Nfkb2�/�mice were inoculated with
MC38 cells and treated with IR and T cell transfer as
described in (B). 200 mg anti-CD8 mAb was admin-
istered as described in (G). Tumor growth was
monitored after radiation.
Representative data are shown from three experi-
ments (one experiment for G) conducted with 4–5
mice per group. Data are represented as mean ± SD;
*p < 0.05, **p < 0.01, and ***p < 0.001. Please also
see Figures S1 and S2.
phenotypes and functions of DCs and CD8+ T cells in NF-kB2-
and Relb-deficient mice after RT. The percentage of CD11c+ tu-
mor-infiltrating DCs in Nfkb2�/� mice was higher than that of
Nfkb2+/+ mice after IR (Figure S2A). NF-kB2-deficient DCs ex-
pressed higher MHC class I and II proteins and the co-stimula-
tory molecule CD80 (Figure S2B), which suggested more mature
and higher cross-priming ability inNfkb2�/�mice. In an ELISPOT
cross-priming assay, DCs sorted from tumors grown in Itgax-
cre Relbf/f and Nfkb2�/� mice showed increased priming func-
tion compared with control (Figures 2C and S2C). To rule out
possible influences from other cell types, the antigen-specific
cross-priming ability of bone marrow-derived DCs (BMDCs)
were examined. Increased IFN-g+ in wells containing RelB- or
NF-kB2-deficient BMDCs (Figures 2D and S2D) suggested that
non-canonical NF-kB signaling deficiency intrinsically promoted
the antigen processing and presenting capacities of DCs.
The frequency and function of CD8+ T cells in tumors grown in
Itgax-cre Relbf/f and Nfkb2�/� mice also increased compared to
those of WT mice after IR. IR enhanced the accumulation of
CD45+ hematopoietic cells and CD8+ T cells among CD45+ cells
in MC38 tumors grown in WTmice, and to a greater extent, in tu-
mors grown in Nfkb2�/� and RelB-deficient mice (Figures S2E,
2E, and S2F). By ELISPOT assay, we observed that IR induced
greater CD8+ T cell responses against tumor-specific antigen
in the TDLN of Itgax-cre Relbf/f and Nfkb2�/� mice compared
with control mice (Figures 2F and S2G). To examine whether
Immunity 49, 490–503, September 18, 2018 493

increased accumulation and enhanced function of CD8+ T cells
are relevant to the therapeutic efficacy of RT, we investigated tu-
mor growth inRelbf/f and Itgax-cre Relbf/f mice in the presence of
a CD8+ T cell depletion antibody. The enhanced anti-tumor ef-
fect of IR in Itgax-cre Relbf/f mice and Nfkb2�/� mice was abro-
gated when T cells were not present (Figures 2G and 2H). Taken
together, our results demonstrate that non-canonical NF-kB
pathway signaling is a negative regulator of anti-tumor immune
responses in the context of IR. Deficiency of non-canonical
NF-kB signaling in DCs promotes DC function in T cell priming,
which in turn leads to an enhanced therapeutic effect of IR.
Non-canonical NF-kB Signaling Deficiency in DCsPromotes IFN-b Expression after IRWe previously demonstrated that the induction of type I IFNs in
DCs by IR is essential for functional CD8+ T cell-mediated anti-
tumor immunity (Burnette et al., 2011; Deng et al., 2014b). To
test whether type I IFNs are required for increased anti-tumor
effect in mice deficient in non-canonical NF-kB signaling, we
blocked type I IFN signaling following IRwith an antibody against
IFNAR1. The result showed that the increased anti-tumor effect
in Nfkb2�/� and Itgax-cre Relbf/f mice was abrogated by admin-
istration of IFNAR1 antibody (Figures 3A and 3B). To test whether
the non-canonical NF-kB pathway modulates the anti-tumor
functions of DCs and CD8+ T cells following IR by regulating
type I IFN expression, wemeasured concentrations of IFN-b pro-
tein in tumors. The induction of IFN-b in tumors was increased
in Nfkb2�/� mice after IR (Figure 3C). We also found that
the expression of CXCL10, a type I IFN-stimulated gene, was
increased inNfkb2�/�mice after IR (Figure 3D). To further assess
whether the non-canonical NF-kB pathway regulates IFN-b
expression in DCs, we measured IFN-b production by BMDCs
purified after co-culturing with control or irradiated tumor cells.
The amount of secreted IFN-b protein was increased in NF-
kB2-deficient BMDCs stimulated with irradiated tumor cells (Fig-
ure 3E). We also demonstrated that IFN-b production induced by
irradiated tumor cells was increased in BMDCs derived from
Itgaqx-cre Relbf/f mice compared to that from Relbf/f control
mice (Figure 3F). Using type I IFN production as readout, we
demonstrated that the non-canonical NF-kB pathway is a nega-
tive regulator of DC function following IR.
Non-canonical NF-kB Pathway Activation in DCs IsRegulated by the STING-TBK1 Axis after IRThe cGAS-STING pathway plays a critical role in anti-tumor im-
munity by regulating type I IFN expression (Woo et al., 2014),
and our previous studies indicate that STING drives IR-induced
anti-tumor immunity by stimulating type I IFN expression (Deng
et al., 2014b). Therefore, we investigated whether the non-ca-
nonical NF-kB pathway controls STING-mediated type I IFN
expression. To address this question, we utilized 5, 6 dimethyl-
xanthenone-4-acetic acid (DMXAA), which is a direct ligand for
murine STING and induces type I IFN expression in a STING-
dependent manner (Corrales et al., 2015; Deng et al., 2014b).
While the production of IFN-b by BMDCs was increased by
stimulation with DMXAA, IFN-b concentrations were even
greater in NF-kB2- and RelB-deficient BMDCs compared with
WT controls (Figures 3G and 3H). These data suggested that
the non-canonical NF-kB pathway is downstream of STING
494 Immunity 49, 490–503, September 18, 2018
and therefore able to negate STING action in regulating IR-
induced type I IFN expression.
Next, we investigated the activation of STING and non-canon-
ical NF-kB pathways in BMDCs after co-culture with irradiated
tumor cells. We found that irradiated tumor cells enhanced the
phosphorylation of TBK1 and IRF3 and the activation of non-ca-
nonical NF-kB signaling in the BMDCs of WT mice (Figures 4A,
4B, S3A, and S3B). Furthermore, we found that the enhanced
activation of non-canonical NF-kB after co-culture with irradi-
ated tumor cells was abolished in STING-deficient BMDCs
(Figures 4C and S3C). These results suggested that STING
activation is required for IR-induced activation of non-canonical
NF-kB signaling in BMDCs. To determine whether STING activa-
tion is sufficient to activate non-canonical NF-kB, DMXAA was
utilized to stimulate WT- and STING-deficient BMDCs. DMXAA
treatment activated nuclear translocation of RelB in a STING-
dependent manner (Figures 4D and S3D). As anticipated,
TBK1 was also activated by DMXAA in a STING-dependent
manner (Figures 4D and S3D). When TBK1 is deficient,
DMXAA-induced activation of non-canonical NF-kB was abro-
gated (Figures 4E and S3E). These results suggested that
TBK1 is a key regulator in STING-induced activation of the
non-canonical NF-kB pathway.
To understand whether DNA from irradiated tumor cells is able
to trigger the activation of non-canonical NF-kB signaling in DCs
through the STING-mediated DNA-sensing pathway in tumor
microenvironment, we labeled tumor DNA with EdU before inoc-
ulation and detected the up-take of tumor DNA and the activa-
tion of STING and non-canonical NF-kB pathways in DCs. As
shown in Figure 4F, IR not only increased the up-take of tumor
DNA in DCs in vivo, but also enhanced phosphorylation of
TBK1 and p100 in WT DCs. In contrast, although DCs from
STING-deficient mice showed comparable up-take of tumor
DNA, the enhanced phosphorylation of TBK1 and p100 was
abrogated. These data suggested that IR-induced up-take of
tumor DNA in DCs subsequently activated TBK1 and non-ca-
nonical NF-kB signaling through the STING pathway.
To dissect which component in the non-canonical NF-kB
pathway is the target of STING-TBK1, we examined the activa-
tion of up-stream components of the non-canonical NF-kB
pathway in BMDCs after stimulation with DMXAA. The degrada-
tion of TRAF2 and 3, the accumulation of NIK, and the phosphor-
ylation of IKKawere not altered by DMXAA (Figures 4G and S3F).
However, the phosphorylation of p100 (NF-kB2) was increased
by DMXAA, suggesting that p100 could be the primary target
of STING regulation. By using NF-kB2-deficient BMDCs, we
found that p100-p52 was required for nuclear translocation of
RelB during STING activation (Figures 4H and S3G). By using
BMDCs derived from Chukf/f (IKKa) or Itgax-cre Chukf/f mice,
our results revealed that activation of non-canonical NF-kB in
IKKa-deficient BMDCs by DMXXA was comparable to that in
WT cells (Figures 4I and S3H), suggesting that IKKa is dispens-
able in STING activation of RelB translocation. Furthermore,
we observed direct binding between TBK1 and p100 in BMDCs
stimulated with DMXAA by using co-IP assay (Figures 4J and
S3I). In vitro kinase assays showed that WT TBK1, but not
TBK1 kinase loss-of-function mutants (including K38A domi-
nant-negative mutation or kinase domain deletion), phosphory-
lated p100, which suggested that p100 is a substrate of TBK1

Figure 3. Non-canonical NF-kB Deficiency in DCs Promote IFN-b Expression after Irradiation
(A andB)MC38 tumormodel were established inNfkb2+/+ andNfkb2�/�mice (A) orRelbf/f and Itgax-cre Relbf/f (B) mice and treatedwith IR as described in Figures
2B and 2A. 200 mg anti-IFNAR1 was administered intratumorally on days 0 and 2 after radiation. Tumor growth was monitored after radiation.
(C and D) Tumors were removed 3 days after radiation and homogenized in PBS with protease inhibitor. ELISA assay was performed to measure IFN-b (C) and
CXCL10 (D).
(E and F) BMDCs derived from Nfkb2+/+ and Nfkb2�/� mice (E) or Relbf/f and Itgax-cre Relbf/f mice (F) were co-cultured with irradiated or non-irradiated MC38
cells. The purified CD11c+ cells were incubated for additional 3 days and supernatants were collected to measure IFN-b by ELISA assay.
(G and H) BMDCs derived from Nfkb2+/+ and Nfkb2�/� mice (G) or Relbf/f and Itgax-cre Relbf/f mice (H) were stimulated with 50 mg/mL DMXAA for 24 hr. The
amount of IFN-b in supernatants was measured by ELISA.
Representative data are shown from three experiments (one experiment for B) conducted with 4–5 (A, B) or 3–4 (C–H) mice per group. Data are represented as
mean ± SD; **p < 0.01 and ***p < 0.001.
Immunity 49, 490–503, September 18, 2018 495

(legend on next page)
496 Immunity 49, 490–503, September 18, 2018

(Figure 4K). Taken together, these data suggest that irradiated
tumor cells or a STING agonist can activate the STING pathway
in DCs, which subsequently activates non-canonical NF-kB
signaling in a TBK1-dependent manner.
Non-canonical NF-kB Signaling Regulates IFN-bExpression through Inhibition of RelA Binding to Ifnb
Promoter in DCsIn antiviral immunity, Ifnb expression requires recruitment of a
number of transcription factors, including IRF3 and p50-RelA
heterodimer, binding to the positive regulatory domains of
the Ifnb promoter (Jin et al., 2014; Wang et al., 2010). In order
to elucidate the mechanism by which non-canonical NF-kB
signaling suppresses IR-induced Ifnb expression, we examined
the activation of IRF3 and canonical NF-kB in BMDCs after
co-culturing with irradiated tumor cells. The immunoblotting
demonstrated that irradiated MC38 cells promoted the nuclear
translocation of both IRF3 and RelA in BMDCs (Figures 5A and
S4A). Similarly, the STING agonist, DMXAA, also enhanced the
phosphorylation of IkBa and the nuclear translocation of RelA
in a time-dependent manner (Figures 5B and S4B). Furthermore,
the activation of IRF3 and canonical NF-kB signaling in DCs was
both STING and TBK1 dependent, as indicated by diminished
activation in Tmem173- or Tbk1-deficient cells (Figures 5C, 5D,
S4C, and S4D). The activation of these components are required
for IR-induced Ifnb expression in BMDCs when co-cultured with
irradiated tumor cells or stimulated by a STING agonist; as in
cells deficient in Tbk1 (Figures 5E and S4E), Irf3 (Figures 5F
and S4E), or Rela (Figures 5G and S4G), IFN-b production was
abrogated. Inhibition of the canonical NF-kB pathway by JSH-
23 also showed similar results (Figures 5H and S4H). These
data suggest that both the TBK1-IRF3 and canonical NF-kB
pathways are required for IR-induced Ifnb expression in DCs.
To understand how non-canonical NF-kB signaling negatively
regulates type I IFN induction, we examined the effect of non-
canonical NF-kB deficiency on STING-induced activation of
signaling and transcriptional factors involved in Ifnb expression.
The loss of NF-kB2 did not affect DMXAA-induced activation of
Figure 4. Irradiated Tumor Cells Promote Non-canonical NF-kB Activa
(A and B) Immuno-blotting analysis of indicated phosphorylated (p-) and total prot
purified BMDCs after co-cultured with irradiated or non-irradiated MC38 cells fo
(C) Immuno-blotting analysis of indicated phosphorylated (p-) and total protei
Tmem173�/� BMDCs after co-cultured with irradiated or non-irradiated MC38 c
(D) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein i
BMDCs stimulated with 50 mg/mL DMXAA for indicated time.
(E) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein
phages after stimulated with 50 mg/mL DMXAA for indicated time points.
(F) MC38 cells were labeled with EdU and inoculated into mice and the tumor bum
digested into single-cell suspensions. Cells were fixed, permeabilized, and staine
p-p100+ cells in live CD45+CD11c+ DCs were analyzed by flow cytometry.
(G) Immuno-blotting analysis of indicated protein in whole-cell lysates of BMDCs
(H) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Nfk
indicated time points.
(I) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of BMD
50 mg/mL DMXAA for indicated time points.
(J) WT BMDCs were stimulated with 50 mg/mL DMXAA for indicated time in pre
followed by detecting p100 or TBK1 associated TBK1 or p100 by immuno-blotti
(K) In vitro kinase assay of TBK1 WT, K38A (Lys38 mutated to Ala), and KDD (k
detected to reflect the kinetics of the kinase reaction.
Representative data are shown from three experiments (one experiment for K). D
either the TBK1-IRF3 or canonical NF-kB pathways, suggesting
that the NF-kB2 pathway does not directly regulate the STING-
TBK1-IRF3 axis (Figures 5I and S4I). Next we asked whether
non-canonical NF-kB signaling suppressed type I IFN expres-
sion by inhibiting STING-induced transcriptional factors binding
to the Ifnb promoter by Chromatin immunoprecipitation (ChIP)
assays. The results showed that the binding of RelB to the Ifnb
promoter was increased by DMXAA treatment in WT BMDCs,
but not in NF-kB2-deficient cells (Figure 5J). Additionally, the
binding of RelA to the Ifnb promoter was increased by DMXAA
treatment in WT BMDCs, which was further elevated markedly
in NF-kB2-deficient cells (Figure 5K). In contrast, the DMXAA-
induced binding of IRF3 to the Ifnb promoter was comparable
between WT and NF-kB2-deficient BMDCs (Figure 5L). These
data suggest that non-canonical NF-kB signaling negatively reg-
ulates IR-induced type I IFN production by inhibiting binding of
RelA to the Ifnb promoter without having an effect on the binding
ability of IRF3.
To further explore the relationship between canonical and
non-canonical NF-kB signaling in regulating the anti-tumor effect
of IR, we monitored the tumor growth in Itgax-cre Relbf/f mice in
the presence of JSH-23. The result showed that the enhanced
anti-tumor effect in Itgax-cre Relbf/f mice was abrogated by
JSH-23, which suggested that canonical NF-kB was required
for an enhanced anti-tumor effect in RelB-deficient mice (Figures
2A and S4J). Furthermore, canonical NF-kB signaling in DCs
contributed to the anti-tumor effect of IR by inducing IFN-b, since
administration of exogenous IFN-b restored the anti-tumor effect
of IR in Itgax-cre Relaf/f mice (Figures 1G and S4K).
Inhibition of Non-canonical NF-kB Signaling CanPromote IR-Induced Anti-tumor ImmunityTo further investigate the role of non-canonical NF-kB signaling
in the context of the clinical response of human cancer patients,
we analyzed overall survival of patients receiving RT for glioblas-
toma (GBM) in a publicly available database (The Cancer
Genome Atlas [TCGA]); we found that patients with low Nfkb2
expression had better overall survival after RT (Figure S5A). A
tion in DCs in a STING-TBK1-Dependent Manner
ein in whole-cell lysates (A) or cytoplasmic (CE) and nuclear (NE) extracts (B) of
r indicated time.
n in cytoplasmic (CE) and nuclear (NE) extracts of purified Tmem17+/+ and
ells for 1 hr.
n cytoplasmic (CE) and nuclear (NE) extracts of Tmem17+/+ and Tmem173�/�
in cytoplasmic (CE) and nuclear (NE) extracts of Tbk1+/+ and Tbk1�/� macro-
ps were irradiated with 20 Gy. The next day, tumor bumps were harvested and
d with indicated antibodies. The frequency of EdU+, EdU+ p-TBK1+, and EdU+
after being stimulated with 50 mg/mL DMXAA for indicated time points.
b2+/+ and Nfkb2�/� BMDCs after being stimulated with 50 mg/mL DMXAA for
Cs derived from Chukf/f and Itgax-cre Chukf/f mice after being stimulated with
sence of 20 mM MG132. Whole-cell lysates were subjected to p100 or TBK1
ng.
inase domain depletion) on substrate p100. The rate of ADP production was
ata are represented as mean ± SD; ***p < 0.001. Please also see Figure S3.
Immunity 49, 490–503, September 18, 2018 497

(legend on next page)
498 Immunity 49, 490–503, September 18, 2018

similar result was obtained when the overall survival was
analyzed based on the Relb expression (Figure S5B). These re-
sults suggest that non-canonical NF-kB signaling diminishes
the therapeutic effect of IR not only in pre-clinical tumor models,
but also in patient clinical response.
We hypothesize that the inhibition of the non-canonical NF-kB
pathway may be used as a translational strategy to enhance
RT-induced anti-tumor effects. To address this question, we em-
ployed a competitive inhibitor (SN52) of NF-kB2 (Xu et al., 2008),
which inhibits the nuclear translocation of p52-RelB hetero-
dimers. SN52, but not SN52Mu (a mutant control for SN52), in-
hibited DMXAA-induced nuclear translocation of RelB in BMDCs
(Figure S5C). In contrast, treatment with SN52 did not change
the activation of canonical NF-kB signaling. Furthermore, we
found that nuclear translocation of RelB was increased in DCs
isolated from irradiated tumors, and this activation was abol-
ished by SN52 intratumoral (i.t.) treatment (Figure S5D). Consis-
tent with the results of NF-kB2-deficient cells, inhibition of non-
canonical NF-kB by SN52 increased Ifnb expression in BMDCs
stimulated with irradiated tumor cells or STING agonist (Figures
6A and S5E). These results demonstrate that SN52 can effec-
tively recapitulate the phenotype of non-canonical NF-kB dele-
tion, resulting in alleviation of the negative regulation on IR- or
STING-induced IFN-b production. To test the efficacy of SN52
in vivo, we administrated SN52Mu and SN52 i.t. immediately
after IR. The combination of SN52 and IR enhanced anti-tumor
immune functions of both DCs (Figure 6B) and CD8+ T cells (Fig-
ure 6C) and subsequently reduced tumor burden (Figures 6D and
S5F) more effectively compared with IR alone, suggesting that
non-canonical NF-kB inhibition can potentiate the anti-tumor ef-
fect of IR. We next tested whether the combination of IR and
SN52 produced an abscopal effect by using a contralateral
tumor model, in which the primary tumors were treated with IR
and/or SN52 and the secondary tumors did not receive any treat-
ment. A growth delay was observed in the IR+SN52 group but
not in groups receiving either treatment alone (Figure 6E). These
results suggested that non-canonical NF-kB inhibition also pro-
moted systemic anti-tumor immunity induced by IR. To further
Figure 5. Non-canonical NF-kB Regulates IFN-b Expression through In
(A) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of puri
indicated time.
(B) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in c
DMXAA for indicated time points.
(C) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Tmem
time points.
(D) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Tbk1
time points.
(E and F) ELISA assay was performed tomeasure IFN-b produced by Tbk1+/+ and
irradiated or non-irradiated MC38 cells.
(G) BMDCs were induced with bone marrow cells from WT and Relaf/f mice whic
measure IFN-b produced by TAT-Cre pretreated WT and Relaf/f BMDCs after be
(H) ELISA assay was performed to measure IFN-b produced by BMDCs after b
presence of 25 mM JSH-23.
(I) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in
stimulated with 50 mg/mL DMXAA for indicated time points.
(J–L)Nfkb2+/+ andNfkb2�/�BMDCswere stimulated with 50 mg/mLDMXAA for 2 h
RelB (J), RelA (K), and IRF3 (L) to the Ifnb promoter. Input DNA was used as an i
control.
Representative data are shown from three experiments with 3–5 duplicated sam
0.001. Please also see Figure S4.
test whether SN52 promoted the therapeutic effect of IR through
the STING-IFNs axis, we investigated tumor growth in STING-
deficient mice (Figure 6F) and in the presence of IFNAR1-block-
ing antibody (Figure 6G) following treatment with IR and/or SN52.
The results indicated that STING and type I IFNs were required
for SN52-enhanced anti-tumor effects of RT.
Although SN52 promoted the therapeutic effect of IR by regu-
lating type I IFNs, the tumors that received combination treat-
ment were not completely eliminated. Sustained IFN-I signaling
induces immunosuppressivemechanisms, including the expres-
sion of both PD-L1 on DCs and other myeloid cells and PD-1 on
T cells, which result in CD8+ T cell exhaustion (Garcia-Diaz et al.,
2017; Tang et al., 2018). Therefore, we treated tumor-bearing
mice with anti-PD-L1 after SN52 and IR treatment. PD-L1
blockade enhanced the therapeutic effect of the SN52+IR com-
bination (Figure 6H) and led to tumor rejection (Figure S5G). To
address whether this combination treatment resulted in genera-
tion of prolonged protective T cell immunity, tumor-free mice
were rechallenged with higher dose of MC38 tumor cells on
the opposite flank. No palpable tumors were detected on the
treated mice after a few weeks (Figure 6I). Together, combining
IR with manipulation of the STING-IFN pathway and checkpoint
inhibition for better innate priming and alleviation of immunosup-
pression may present a fresh avenue for cancer therapy.
DISCUSSION
Radiation induces both innate and adaptive anti-tumor immune
responses, and the axis of DCs and CTLs plays a crucial role
in this process (Gupta et al., 2012; Lee et al., 2009). Type I
IFNs induced by IR are essential for both cross-presentation ca-
pacity of dendritic cells and anti-tumoral function of CD8+ T cells
(Burnette et al., 2011; Zitvogel et al., 2015). Our previous work
established that the cGAS- and STING-dependent cytosolic
DNA-sensing pathways in DCs are required for type I IFN induc-
tion after IR (Deng et al., 2014b). In this report, we identify a
previously unknown downstream control mechanism of the
DNA-sensing pathway in DCs that has particular relevance to
hibition of RelA Binding to the Ifnb Promoter in DCs
fied BMDCs after co-cultured with irradiated or non-irradiated MC38 cells for
ytoplasmic (CE) and nuclear (NE) extracts of BMDCs stimulated with 50 mg/mL
17+/+ and Tmem173�/�BMDCs stimulated with 50 mg/mL DMXAA for indicated
+/+ and Tbk1�/� macrophages stimulated with 50 mg/mL DMXAA for indicated
Tbk1�/�macrophages (E) or Irf3+/+ and Irf3�/�BMDCs (E) after co-culturedwith
h were treated with 50 mg/mL TAT-Cre for 2 hr. ELISA assay was performed to
ing co-cultured with irradiated or non-irradiated MC38.
eing co-cultured with irradiated or non-irradiated MC38 cells in absence or
cytoplasmic (CE) and nuclear (NE) extracts of Nfkb2+/+ and Nfkb2�/� BMDCs
r. ChIP assayswere performed and quantified by qPCR to detect the binding of
nternal control and the data are shown as the relative fold increased over IgG
ples in each group. Data are represented as mean ± SD; **p < 0.01 and ***p <
Immunity 49, 490–503, September 18, 2018 499

Figure 6. Inhibition of Non-canonical NF-kB Can Promote Radiation-Induced Anti-tumor Immunity
(A) WT BMDCs were pretreated with 40 mg/mL N52Mu or SN52 for 30 min and then co-cultured with irradiated or non-irradiated MC38 cells. ELISA assay was
performed to measure IFN-b produced by purified BMDCs as described in Figure 3E.
(B and C) MC38-SIY tumors (B) and MC38 tumors (C) established in B6 mice were treated locally with one dose of 20 Gy IR. 40 mg SN52Mu or SN52 was
administered i.t. in tumor-bearing mice on day �1, day 1, and day 3 of radiation.
(B) On day 4 post IR, tumors were removed and the cross-priming ability of DCs was evaluated by counting IFN-g+ spots as described in Figure 2C.
(C) On day 7 post IR, TDLNs were removed and tumor antigen-specific CD8+ T cell function was measured by ELISPOT assays as described in Figure 2F.
(D) MC38 tumors established in B6mice were treated with one dose of 20 Gy IR. 40 mg SN52Mu or SN52was administered i.t. every 2 days for a total of four times
staring from 1 day before radiation. Tumor growth was monitored after radiation.
(legend continued on next page)
500 Immunity 49, 490–503, September 18, 2018

cancer therapy. Our data demonstrate that the innate DNA-
sensing pathway not only exerts immunostimulatory functions
via IFN production, but also activates the non-canonical NF-kB
pathway, which in turn negatively regulates type I IFN induction
after IR. These results describe an alternative mechanism of
STING regulation and indicates that targeting the non-canonical
NF-kB pathway enhanced anti-tumor effects of IR.
We found that irradiated tumor cells stimulated the activation
of the non-canonical NF-kB pathway in DCs in a STING-TBK1-
dependent manner. Activation of the STING pathway was both
necessary and sufficient for the activation of the non-canonical
NF-kB pathway. The stabilization of NIK and phosphorylation
of IKKa are well known as central early steps in activation of
non-canonical NF-kB pathway (Vallabhapurapu and Karin,
2009). Jin et al. (2012) found that TBK1 controls IgA class
switching in B cells by negatively regulating phosphorylation
and subsequent degradation of NIK. Cytosolic DNA can activate
the non-canonical NF-kB pathway in a STING-dependent and
TBK1-independent manner in MEFs or human tumor cells (Abe
and Barber, 2014; Bakhoum et al., 2018). However, our data
demonstrated that DMXAA, a STING agonist, did not increase
the accumulation of NIK or phosphorylation of IKKa in the pro-
cess of activating p100 phosphorylation and nuclear transloca-
tion of p52-RelB heterodimers. Rather, our results suggest that
in response to IR-induced damage, host immune cells may pref-
erentially use the cGAS-STING DNA-sensing pathway, instead
of NIK-IKKa, to activate the non-canonical NF-kB pathway.
Understanding this alternative regulation would be of value for
choosing or designing inhibitors of the non-canonical NF-kB
pathway to enhance the therapeutic effect of IR as well as ther-
apies in virus infection and auto-immune diseases. It is intriguing
that activation of the STING pathway induced both the canonical
(triggering production of IFN) and the non-canonical (inhibiting
IFN production) NF-kB pathways in DCs. This may represent
an intricate and complex regulation of IFN production respond-
ing to differential stimuli.
A growing body of evidence shows that the canonical NF-kB
pathway plays a crucial role in supporting tumor initiation and
progression via mechanisms including stimulating cell prolifera-
tion, inhibiting apoptosis (Beg and Baltimore, 1996), increasing
metastasis (Wang et al., 1999), and angiogenesis (Koch et al.,
1992). Radiation induces constitutive canonical NF-kB activation
in certain cancers (Brach et al., 1991; Chen et al., 2002), and acti-
vation of NF-kB is associated with the development of radiore-
sistance. Given this evidence, a large number of inhibitors of
canonical NF-kB pathway are undergoing development and
(E) B6 mice were injected s.c. with 1 3 106 MC38 cells on left flank on day 0 (prim
tumor). On day 10, primary tumors were treated with IR and SN52Mu or SN52 as d
inoculation.
(F) MC38 tumors established in Tmem17+/+ and Tmem173�/� mice were treated
every 2 days for a total of four times staring from 1 day before radiation. Tumor
(G)MC38 tumors established in B6mice were treated with one dose of 20 Gy IR. 4
staring from 1 day before radiation. All mice were treated with 200 mg anti-IFNAR
(H) MC38 tumors in B6 mice were established and treated with IR and SN52 as de
were given by i.p. every 3 days for a total of four times. Tumor growth was moni
(I) Thirty days after tumor eradication, tumor-free mice from anti-PD-L1+SN52+IR
flank. Tumor growth on naive and tumor-free mice was monitored.
Representative data are shown from three experiments (one experiment for F and
*p < 0.05, **p < 0.01, and ***p < 0.001. Please also see Figure S5.
some have been tested in clinical trials. However, the anti-tumor
efficacy and toxicity of these inhibitors are not promising. In our
present study, an inhibitor of the canonical NF-kB pathway
decreased the therapeutic effect of IR by suppressing IR-
induced anti-tumor immune response. Specifically, canonical
NF-kB, together with IRF3, is required for the induction of
type I IFN in DCs stimulated by irradiated tumor cells through
STING-TBK1 signaling. Taken together, the canonical NF-kB
pathwaymay not be an ideal target for promoting the therapeutic
effect of IR because of its differential roles in inducing tumor ra-
dioresistance and its requirement for IR-induced anti-tumor im-
munity. Similar to the canonical NF-kB pathway, non-canonical
NF-kB is also known as a tumor-promoting factor due to its
role in regulating the expression of BIM, BMF (Vallabhapurapu
et al., 2015), and BCL2 (Wang et al., 2007) in tumor cells. In addi-
tion, non-canonical NF-kB signaling in tumor cells is reported to
be activated by IR and mediates radioresistance by regulating
the expression of MnSOD (Josson et al., 2006), Survivin, and
BCL2 (Mineva et al., 2009). However, we found that, in contrast
to canonical NF-kB, the non-canonical NF-kB pathway in DCs
negatively regulates IR-induced anti-tumor immunity by control-
ling type I IFN expression. Consistently, we found that non-
canonical NF-kB inhibition, but not canonical NF-kB inhibition,
promoted tumor regression induced by local IR. We have previ-
ously shown that, in some tumors, intrinsic radiobiological prop-
erties of the tumor cells are less important than the immune
microenvironments (Liang et al., 2013) and, in this context, inhib-
iting the non-canonical NF-kB pathway may be particularly
important. Our findings reveal a molecular mechanism of IR-
mediated anti-tumor immunity through an alternative model of
STING regulation and provide insight into therapeutic strategies
targeting the NF-kB pathway in cancer immunotherapy.
STAR+METHODS
Detailed methods are provided in the online version of this paper
and include the following:
d KEY RESOURCES TABLE
d CONTACT FOR REAGENT AND RESOURCE SHARING
d EXPERIMENTAL MODEL AND SUBJECT DETAILS
ary
esc
loca
grow
0 mg
1 a
scri
tore
tre
G) c
B In vivo Animal Studies
B Cell Lines
d METHOD DETAILS
B Tumor Growth and Treatments
B Flow Cytometry
tumor) and another 1 3 106 MC38 cells on left flank on day 5 (secondary
ribed in (D). The growth of secondary tumors was monitored after 7 days of
lly with one dose of 20 Gy IR. 40 mg SN52Mu or SN52 was administered i.t.
th was monitored after radiation.
SN52Mu or SN52was administered i.t. every 2 days for a total of four times
s described in Figure 3A. Tumor growth was monitored after radiation.
bed in (D). On the day of IR, 200 mg anti-PD-L1 (10F.9G2) or isotype control
d after IR.
atment group were rechallenged with 2 3 106 MC38 cells on the opposite
onductedwith 4–6mice in each group. Data are represented asmean ± SD;
Immunity 49, 490–503, September 18, 2018 501

502
B In Vitro Culture and Function Assay of BMDCs
B Measurement of IFNg-Secreting CD8+ T Cells by ELI-
SPOT Assay
B ELISA
B Western Blot Analysis
B Chromatin Immunoprecipitation Assays
B Immunoprecipitation Assays
B In vitro Kinase Assays
d QUANTIFICATION AND STATISTICAL ANALYSIS
SUPPLEMENTAL INFORMATION
Supplemental Information includes five figures and can be found with this
article online at https://doi.org/10.1016/j.immuni.2018.07.008.
ACKNOWLEDGMENTS
The authors would like to thank Amy K. Huser for editing assistance and Ro-
lando Torres for assistance in animal studies. This research was supported
in part by Mr. & Mrs. Vincent Foglia, The Chicago Tumor Institute, an endow-
ment from the Ludwig Cancer Research Foundation to R.R.W., and NIH grant
NCI-R21 CA195075 to R.R.W. This work was in part supported by the NIH
grant CA134563 to Y.-X.F. and by Cancer Prevention and Research Institute
of Texas grant RR150072 to Y.-X.F. Flow cytometry cost was covered in
part by University of Chicago Cancer Center Support Grant (P30CA014599).
AUTHOR CONTRIBUTIONS
Y.H. designed research studies, conducted experiments, and analyzed data.
Y.H. and H.L. wrote the manuscript. W.Z. analyzed data; L.D., X.H., E.R.,
M.X., Y.Z., X.Y., A.A., and H.M. conducted experiments. Y.-X.F. and R.R.W.
provided guidance for the research.
DECLARATION OF INTERESTS
The authors declare no competing interests.
Received: September 29, 2017
Revised: January 23, 2018
Accepted: July 17, 2018
Published: August 28, 2018
REFERENCES
Abe, T., and Barber, G.N. (2014). Cytosolic-DNA-mediated, STING-dependent
proinflammatory gene induction necessitates canonical NF-kB activation
through TBK1. J. Virol. 88, 5328–5341.
Bakhoum, S.F., Ngo, B., Laughney, A.M., Cavallo, J.A., Murphy, C.J., Ly, P.,
Shah, P., Sriram, R.K., Watkins, T.B.K., Taunk, N.K., et al. (2018).
Chromosomal instability drives metastasis through a cytosolic DNA response.
Nature 553, 467–472.
Beg, A.A., and Baltimore, D. (1996). An essential role for NF-kappaB in pre-
venting TNF-alpha-induced cell death. Science 274, 782–784.
Brach, M.A., Hass, R., Sherman, M.L., Gunji, H., Weichselbaum, R., and Kufe,
D. (1991). Ionizing radiation induces expression and binding activity of the nu-
clear factor kappa B. J. Clin. Invest. 88, 691–695.
Burnette, B.C., Liang, H., Lee, Y., Chlewicki, L., Khodarev, N.N.,
Weichselbaum, R.R., Fu, Y.X., and Auh, S.L. (2011). The efficacy of radio-
therapy relies upon induction of type i interferon-dependent innate and adap-
tive immunity. Cancer Res. 71, 2488–2496.
Caamano, J.H., Rizzo, C.A., Durham, S.K., Barton, D.S., Raventos-Suarez, C.,
Snapper, C.M., and Bravo, R. (1998). Nuclear factor (NF)-kappa B2 (p100/p52)
is required for normal splenic microarchitecture and B cell-mediated immune
responses. J. Exp. Med. 187, 185–196.
Immunity 49, 490–503, September 18, 2018
Chen, X., Shen, B., Xia, L., Khaletzkiy, A., Chu, D., Wong, J.Y., and Li, J.J.
(2002). Activation of nuclear factor kappaB in radioresistance of TP53-inactive
human keratinocytes. Cancer Res. 62, 1213–1221.
Corrales, L., Glickman, L.H., McWhirter, S.M., Kanne, D.B., Sivick, K.E.,
Katibah, G.E., Woo, S.R., Lemmens, E., Banda, T., Leong, J.J., et al. (2015).
Direct activation of STING in the tumor microenvironment leads to potent
and systemic tumor regression and immunity. Cell Rep. 11, 1018–1030.
Deng, L., Liang, H., Burnette, B., Beckett, M., Darga, T., Weichselbaum, R.R.,
and Fu, Y.X. (2014a). Irradiation and anti-PD-L1 treatment synergistically pro-
mote antitumor immunity in mice. J. Clin. Invest. 124, 687–695.
Deng, L., Liang, H., Xu, M., Yang, X., Burnette, B., Arina, A., Li, X.D., Mauceri,
H., Beckett, M., Darga, T., et al. (2014b). STING-dependent cytosolic DNA
sensing promotes radiation-induced type I interferon-dependent antitumor im-
munity in immunogenic tumors. Immunity 41, 843–852.
Erstad, D.J., and Cusack, J.C., Jr. (2013). Targeting the NF-kBpathway in can-
cer therapy. Surg. Oncol. Clin. N. Am. 22, 705–746.
Garcia-Diaz, A., Shin, D.S., Moreno, B.H., Saco, J., Escuin-Ordinas, H.,
Rodriguez, G.A., Zaretsky, J.M., Sun, L., Hugo, W., Wang, X., et al. (2017).
Interferon receptor signaling pathways regulating PD-L1 and PD-L2 expres-
sion. Cell Rep. 19, 1189–1201.
Gardner, A., and Ruffell, B. (2016). Dendritic cells and cancer immunity. Trends
Immunol. 37, 855–865.
Gupta, A., Probst, H.C., Vuong, V., Landshammer, A., Muth, S., Yagita, H.,
Schwendener, R., Pruschy, M., Knuth, A., and van den Broek, M. (2012).
Radiotherapy promotes tumor-specific effector CD8+ T cells via dendritic
cell activation. J. Immunol. 189, 558–566.
Hayden, M.S., and Ghosh, S. (2008). Shared principles in NF-kappaB
signaling. Cell 132, 344–362.
Hou, Y., Lin, H., Zhu, L., Liu, Z., Hu, F., Shi, J., Yang, T., Shi, X., Guo, H., Tan, X.,
et al. (2014). The inhibitory effect of IFN-g on protease HTRA1 expression in
rheumatoid arthritis. J. Immunol. 193, 130–138.
Jin, J., Xiao, Y., Chang, J.H., Yu, J., Hu, H., Starr, R., Brittain, G.C., Chang, M.,
Cheng, X., and Sun, S.C. (2012). The kinase TBK1 controls IgA class switching
by negatively regulating noncanonical NF-kB signaling. Nat. Immunol. 13,
1101–1109.
Jin, J., Hu, H., Li, H.S., Yu, J., Xiao, Y., Brittain, G.C., Zou, Q., Cheng, X.,
Mallette, F.A., Watowich, S.S., and Sun, S.C. (2014). Noncanonical NF-kB
pathway controls the production of type I interferons in antiviral innate immu-
nity. Immunity 40, 342–354.
Josson, S., Xu, Y., Fang, F., Dhar, S.K., St Clair, D.K., and St Clair, W.H. (2006).
RelB regulates manganese superoxide dismutase gene and resistance to
ionizing radiation of prostate cancer cells. Oncogene 25, 1554–1559.
Koch, A.E., Polverini, P.J., Kunkel, S.L., Harlow, L.A., DiPietro, L.A., Elner,
V.M., Elner, S.G., and Strieter, R.M. (1992). Interleukin-8 as a macrophage-
derived mediator of angiogenesis. Science 258, 1798–1801.
Lee, Y., Auh, S.L., Wang, Y., Burnette, B., Wang, Y., Meng, Y., Beckett, M.,
Sharma, R., Chin, R., Tu, T., et al. (2009). Therapeutic effects of ablative radi-
ation on local tumor require CD8+ T cells: changing strategies for cancer treat-
ment. Blood 114, 589–595.
Liang, H., Deng, L., Chmura, S., Burnette, B., Liadis, N., Darga, T., Beckett,
M.A., Lingen, M.W., Witt, M., Weichselbaum, R.R., and Fu, Y.X. (2013).
Radiation-induced equilibrium is a balance between tumor cell proliferation
and T cell-mediated killing. J. Immunol. 190, 5874–5881.
Liang, H., Deng, L., Hou, Y., Meng, X., Huang, X., Rao, E., Zheng, W., Mauceri,
H., Mack, M., Xu, M., et al. (2017). Host STING-dependent MDSCmobilization
drives extrinsic radiation resistance. Nat. Commun. 8, 1736–1746.
Lugade, A.A., Moran, J.P., Gerber, S.A., Rose, R.C., Frelinger, J.G., and Lord,
E.M. (2005). Local radiation therapy of B16 melanoma tumors increases the
generation of tumor antigen-specific effector cells that traffic to the tumor.
J. Immunol. 174, 7516–7523.
Mineva, N.D., Wang, X., Yang, S., Ying, H., Xiao, Z.X., Holick, M.F., and
Sonenshein, G.E. (2009). Inhibition of RelB by 1,25-dihydroxyvitamin D3 pro-
motes sensitivity of breast cancer cells to radiation. J. Cell. Physiol. 220,
593–599.

Nelson, J.D., Denisenko, O., and Bomsztyk, K. (2006). Protocol for the fast
chromatin immunoprecipitation (ChIP) method. Nat. Protoc. 1, 179–185.
Perkins, N.D. (2012). The diverse and complex roles of NF-kB subunits in can-
cer. Nat. Rev. Cancer 12, 121–132.
Prise, K.M., and O’Sullivan, J.M. (2009). Radiation-induced bystander signal-
ling in cancer therapy. Nat. Rev. Cancer 9, 351–360.
Reits, E.A., Hodge, J.W., Herberts, C.A., Groothuis, T.A., Chakraborty, M.,
Wansley, E.K., Camphausen, K., Luiten, R.M., de Ru, A.H., Neijssen, J.,
et al. (2006). Radiation modulates the peptide repertoire, enhancesMHC class
I expression, and induces successful antitumor immunotherapy. J. Exp. Med.
203, 1259–1271.
Shih, V.F., Davis-Turak, J., Macal, M., Huang, J.Q., Ponomarenko, J., Kearns,
J.D., Yu, T., Fagerlund, R., Asagiri, M., Zuniga, E.I., and Hoffmann, A. (2012).
Control of RelB during dendritic cell activation integrates canonical and nonca-
nonical NF-kB pathways. Nat. Immunol. 13, 1162–1170.
Shishodia, S., and Aggarwal, B.B. (2004). Nuclear factor-kappaB: a friend or a
foe in cancer? Biochem. Pharmacol. 68, 1071–1080.
Sun, S.C. (2012). The noncanonical NF-kB pathway. Immunol. Rev. 246,
125–140.
Takeshima, T., Chamoto, K., Wakita, D., Ohkuri, T., Togashi, Y., Shirato, H.,
Kitamura, H., and Nishimura, T. (2010). Local radiation therapy inhibits tumor
growth through the generation of tumor-specific CTL: its potentiation by com-
bination with Th1 cell therapy. Cancer Res. 70, 2697–2706.
Tang, H., Liang, Y., Anders, R.A., Taube, J.M., Qiu, X., Mulgaonkar, A., Liu, X.,
Harrington, S.M., Guo, J., Xin, Y., et al. (2018). PD-L1 on host cells is essential
for PD-L1 blockade-mediated tumor regression. J. Clin. Invest. 128, 580–588.
Vallabhapurapu, S., and Karin, M. (2009). Regulation and function of
NF-kappaB transcription factors in the immune system. Annu. Rev.
Immunol. 27, 693–733.
Vallabhapurapu, S.D., Noothi, S.K., Pullum, D.A., Lawrie, C.H., Pallapati, R.,
Potluri, V., Kuntzen, C., Khan, S., Plas, D.R., Orlowski, R.Z., et al. (2015).
Transcriptional repression by the HDAC4-RelB-p52 complex regulates multi-
ple myeloma survival and growth. Nat. Commun. 6, 8428.
Wang,W.,Abbruzzese,J.L., Evans,D.B., andChiao,P.J. (1999).Overexpression
of urokinase-type plasminogen activator in pancreatic adenocarcinoma is
regulated by constitutively activated RelA. Oncogene 18, 4554–4563.
Wang, X., Belguise, K., Kersual, N., Kirsch, K.H., Mineva, N.D., Galtier, F.,
Chalbos, D., and Sonenshein, G.E. (2007). Oestrogen signalling inhibits inva-
sive phenotype by repressing RelB and its target BCL2. Nat. Cell Biol. 9,
470–478.
Wang, J., Basagoudanavar, S.H., Wang, X., Hopewell, E., Albrecht, R., Garcıa-
Sastre, A., Balachandran, S., and Beg, A.A. (2010). NF-kappa B RelA subunit is
crucial for early IFN-beta expression and resistance to RNA virus replication.
J. Immunol. 185, 1720–1729.
Weichselbaum, R.R., Liang, H., Deng, L., and Fu, Y.X. (2017). Radiotherapy
and immunotherapy: a beneficial liaison? Nat. Rev. Clin. Oncol. 14, 365–379.
Woo, S.R., Fuertes, M.B., Corrales, L., Spranger, S., Furdyna, M.J., Leung,
M.Y., Duggan, R., Wang, Y., Barber, G.N., Fitzgerald, K.A., et al. (2014).
STING-dependent cytosolic DNA sensing mediates innate immune recogni-
tion of immunogenic tumors. Immunity 41, 830–842.
Xu, Y., Fang, F., St Clair, D.K., Sompol, P., Josson, S., and St Clair, W.H.
(2008). SN52, a novel nuclear factor-kappaB inhibitor, blocks nuclear import
of RelB:p52 dimer and sensitizes prostate cancer cells to ionizing radiation.
Mol. Cancer Ther. 7, 2367–2376.
Zitvogel, L., Galluzzi, L., Kepp, O., Smyth, M.J., and Kroemer, G. (2015). Type I
interferons in anticancer immunity. Nat. Rev. Immunol. 15, 405–414.
Immunity 49, 490–503, September 18, 2018 503
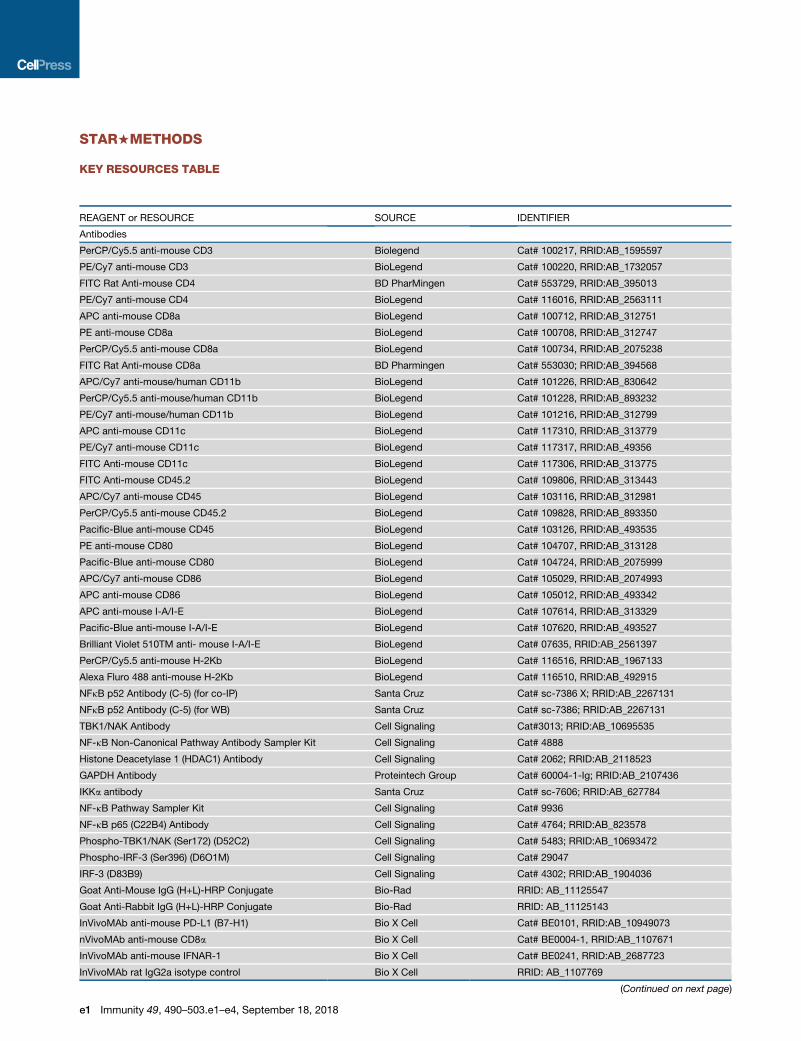
STAR+METHODS
KEY RESOURCES TABLE
REAGENT or RESOURCE SOURCE IDENTIFIER
Antibodies
PerCP/Cy5.5 anti-mouse CD3 Biolegend Cat# 100217, RRID:AB_1595597
PE/Cy7 anti-mouse CD3 BioLegend Cat# 100220, RRID:AB_1732057
FITC Rat Anti-mouse CD4 BD PharMingen Cat# 553729, RRID:AB_395013
PE/Cy7 anti-mouse CD4 BioLegend Cat# 116016, RRID:AB_2563111
APC anti-mouse CD8a BioLegend Cat# 100712, RRID:AB_312751
PE anti-mouse CD8a BioLegend Cat# 100708, RRID:AB_312747
PerCP/Cy5.5 anti-mouse CD8a BioLegend Cat# 100734, RRID:AB_2075238
FITC Rat Anti-mouse CD8a BD Pharmingen Cat# 553030; RRID:AB_394568
APC/Cy7 anti-mouse/human CD11b BioLegend Cat# 101226, RRID:AB_830642
PerCP/Cy5.5 anti-mouse/human CD11b BioLegend Cat# 101228, RRID:AB_893232
PE/Cy7 anti-mouse/human CD11b BioLegend Cat# 101216, RRID:AB_312799
APC anti-mouse CD11c BioLegend Cat# 117310, RRID:AB_313779
PE/Cy7 anti-mouse CD11c BioLegend Cat# 117317, RRID:AB_49356
FITC Anti-mouse CD11c BioLegend Cat# 117306, RRID:AB_313775
FITC Anti-mouse CD45.2 BioLegend Cat# 109806, RRID:AB_313443
APC/Cy7 anti-mouse CD45 BioLegend Cat# 103116, RRID:AB_312981
PerCP/Cy5.5 anti-mouse CD45.2 BioLegend Cat# 109828, RRID:AB_893350
Pacific-Blue anti-mouse CD45 BioLegend Cat# 103126, RRID:AB_493535
PE anti-mouse CD80 BioLegend Cat# 104707, RRID:AB_313128
Pacific-Blue anti-mouse CD80 BioLegend Cat# 104724, RRID:AB_2075999
APC/Cy7 anti-mouse CD86 BioLegend Cat# 105029, RRID:AB_2074993
APC anti-mouse CD86 BioLegend Cat# 105012, RRID:AB_493342
APC anti-mouse I-A/I-E BioLegend Cat# 107614, RRID:AB_313329
Pacific-Blue anti-mouse I-A/I-E BioLegend Cat# 107620, RRID:AB_493527
Brilliant Violet 510TM anti- mouse I-A/I-E BioLegend Cat# 07635, RRID:AB_2561397
PerCP/Cy5.5 anti-mouse H-2Kb BioLegend Cat# 116516, RRID:AB_1967133
Alexa Fluro 488 anti-mouse H-2Kb BioLegend Cat# 116510, RRID:AB_492915
NFkB p52 Antibody (C-5) (for co-IP) Santa Cruz Cat# sc-7386 X; RRID:AB_2267131
NFkB p52 Antibody (C-5) (for WB) Santa Cruz Cat# sc-7386; RRID:AB_2267131
TBK1/NAK Antibody Cell Signaling Cat#3013; RRID:AB_10695535
NF-kB Non-Canonical Pathway Antibody Sampler Kit Cell Signaling Cat# 4888
Histone Deacetylase 1 (HDAC1) Antibody Cell Signaling Cat# 2062; RRID:AB_2118523
GAPDH Antibody Proteintech Group Cat# 60004-1-Ig; RRID:AB_2107436
IKKa antibody Santa Cruz Cat# sc-7606; RRID:AB_627784
NF-kB Pathway Sampler Kit Cell Signaling Cat# 9936
NF-kB p65 (C22B4) Antibody Cell Signaling Cat# 4764; RRID:AB_823578
Phospho-TBK1/NAK (Ser172) (D52C2) Cell Signaling Cat# 5483; RRID:AB_10693472
Phospho-IRF-3 (Ser396) (D6O1M) Cell Signaling Cat# 29047
IRF-3 (D83B9) Cell Signaling Cat# 4302; RRID:AB_1904036
Goat Anti-Mouse IgG (H+L)-HRP Conjugate Bio-Rad RRID: AB_11125547
Goat Anti-Rabbit IgG (H+L)-HRP Conjugate Bio-Rad RRID: AB_11125143
InVivoMAb anti-mouse PD-L1 (B7-H1) Bio X Cell Cat# BE0101, RRID:AB_10949073
nVivoMAb anti-mouse CD8a Bio X Cell Cat# BE0004-1, RRID:AB_1107671
InVivoMAb anti-mouse IFNAR-1 Bio X Cell Cat# BE0241, RRID:AB_2687723
InVivoMAb rat IgG2a isotype control Bio X Cell RRID: AB_1107769
(Continued on next page)
e1 Immunity 49, 490–503.e1–e4, September 18, 2018

Continued
REAGENT or RESOURCE SOURCE IDENTIFIER
Bacterial and Virus Strains
IFN-b-expressing adenovirus (Ad-IFN-b) Byron Burnette
(Burnette et al., 2011)
N/A
Chemicals, Peptides, and Recombinant Proteins
Recombinant Mouse GM-CSF Protein R&D systems 415-ML-010
Cre Recombinase, TAT-Cre Excellgen EG-1001
JSH-23 Selleckchem S7351
ALBUMIN, BOVINE AMRESCO 0332-100G
DMXAA SIGMA D5817-25MG
2030-cGAMP Invivogen tlrl-cga23
MG-132 Selleckchem S2619
Critical Commercial Assays
EasySep Mouse CD8a Positive Selection Kit STEM CELL 18753
EasySep Mouse CD11c Positive Selection Kit II STEM CELL 18780
NE-PER Nuclear and Cytoplasmic Extraction Reagents Thermo Scientific 78833
VeriKine-HS Mouse Interferon Beta Serum ELISA Kit PBL assay science 42410-2
Magna ChIPTM A/G Millipore 17-10085
EasySep Mouse T Cell Enrichment Kit STEM CELL 19751
Pierce Crosslink Magnetic IP/Co-IP Kit Thermo Scientific 88805
Mouse CXCL10/IP-10/CRG-2 Quantikine ELISA Kit R&D Systems MCX100
HA Tag IP/Co-IP Kit Thermo Scientific 26180
Dynabeads His-Tag Isolation & Pulldown Invitrogen 10103D
Universal Kinase Activity Kit R&D systems EA004
Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry
Assay Kit
Thermo Scientific C10635
Experimental Models: Cell Lines
MC38 Fu Lab (Deng et al., 2014b)
MC38-SIY Fu Lab (Deng et al., 2014b)
B16-SIY Fu Lab PMCID: PMC3927846
Experimental Models: Organisms/Strains
Mouse: C57BL/6J (WT) Jackson Laboratory RRID:IMSR_JAX:000664
Mouse: B6.129S1-Relatm1Ukl/J (Relaf/f) Jackson Laboratory RRID:IMSR_JAX:024342
Mouse: B6.Cg-Tg(Itgax-cre)1-1Reiz/J Jackson Laboratory RRID:IMSR_JAX:008068
Mouse: B6.129P2-Lyz2tm1(cre)Ifo/J (Lyz2-Cre) Jackson Laboratory RRID:IMSR_JAX:004781
Mouse: Nfkb2�/� Ulrich Siebenlist https://doi.org/10.1084/jem.187.2.147
Mouse: Relbf/f Falk Weih https://doi.org/10.4049/jimmunol.167.4.1909
Mouse: Irf3�/� T. Taniguchi PMID: 12821121
Mouse: Tmem173�/� Glen N. Barber https://doi.org/10.1038/nature08476
Mouse: Chukf/f Yinling Hu https://doi.org/10.1016/j.ccr.2008.07.017
Oligonucleotides
Primer: Ifnb promoter forward: 50- ATTCCTCTGAGGCAGAAAGGACCA-30
IDT N/A
Primer: Ifnb promoter forward: 50- GCAA
GATGAGGCAAAGGCTGTCAA �30IDT N/A
Software and Algorithms
Graphpad Prism 6 GraphPad Software RRID:SCR_002798
ImageJ NIH RRID: SCR_003070
FlowJo v10.3 FlowJo RRID:SCR_008520
ImmunoSpot 5.0 ImmunoSpot RRID:SCR_011082
(Continued on next page)
Immunity 49, 490–503.e1–e4, September 18, 2018 e2

Continued
REAGENT or RESOURCE SOURCE IDENTIFIER
Other
Peptide: SN52: AAVALLPAVLLALLAPVQRKRRKALP GenScript N/A
Peptide: SN52Mu: AAVALLPAVLLALLAPVQRNGRKALP GenScript N/A
Vector: pcDNA3.1-6His-TBK1 WT GenScript Cat# SC1200 U7841DD090_1
Vector: pcDNA3.1-6His-TBK1 K38A GenScript Cat# SC1200M U7841DD090_2
Vector: pcDNA3.1-6His-TBK1 KDD GenScript Cat# SC1200M U7841DD090_3
Vector: pcDNA3.1-HA-p100 GenScript Cat# SC1200 U7841DD090_4
CONTACT FOR REAGENT AND RESOURCE SHARING
Further information and requests for resources and reagents should be directed to and fulfilled by the Lead Contact, Dr. Yang-Xin Fu
EXPERIMENTAL MODEL AND SUBJECT DETAILS
In vivo Animal StudiesC57BL/6J wild-type (WT), Itgax-cre+-Tg, Relaf/f mice were purchased from Jackson Laboratory. Nfkb2�/� mice were provided by
Dr. Ulrich Siebenlist, NIAID, NIH. Chukf/f mice were kindly provided by Dr. Yinling Hu, NCI, NIH. Relbf/f mice were kindly provided
by Dr. Falk Weih, Forschungszentrum Karlsruhe, Germany. Irf3�/� mice were kindly provided by T. Taniguchi of University of Tokyo.
Tmem173�/� mice were kindly provided by Dr. Glen N. Barber of University of Miami School of Medicine and backcrossed to B6
background for 8 times in our lab. All experimental groups included randomly chosen female littermates of ages around 8 weeks
and of the same strain. All themiceweremaintained and used in accordance to the animal experimental guidelines set by the Institute
of Animal Care and Use Committee of the University of Chicago.
Cell LinesSingle-cell suspensions of bone marrow cells were obtained from C57BL/6J, Nfkb2�/�, Itgax-cre Relbf/f, Itgax-cre Chukf/f,
Tmem173�/�, and Irf3�/� mice. The cells were cultured in RPMI-1640 medium containing 10% FBS (DENVILLE), supplemented
with 20 ng/ml GM-CSF. Fresh media with GM-CSF was added into culture on day 3. BMDCs were harvested for stimulation assay
on day 7. Tbk1+/+ and Tbk1�/�macrophages were kindly provided by Dr. Leticia Corrales of University of Chicago. MC38, MC38-SIY
and B16-SIY tumor cell lines were kindly provided by Dr. Xuanming Yang of University of Chicago and grown in DMEMmedium con-
taining 10% FBS, at 37�C and 5% CO2.
METHOD DETAILS
Tumor Growth and Treatments1 3 106 MC38, MC38-SIY or B16-SIY tumor cells were subcutaneously injected into the flank of mice. Tumors were measured and
irradiated at 20 Gy as described in (Deng et al., 2014a). To reconstitute T cell deficiency in Rag1�/� background, 1 3 107 wild-type
CD3+ T cells were adoptively transferred into Rag1�/�Nfkb2�/�and Rag1�/� mice immediately prior to irradiation. For type I IFN
blockade experiments, 200 mg anti-IFNAR1 mAb was intratumorally injected on day 0 and 2 after radiation. For CD8+ T cell depletion
experiments, 200 mg anti-CD8 mAb was delivered 4 times by i.p. injection every 3 days starting 1 day before radiation. For JSH-23
treatment experiments, 6 mg/kg JSH-23 was given by gavage daily starting 1 day before IR for totally 14 days. For SN52 treatment
experiments, 40 mg SN52 was administered for 4 times by i.t. injection every 2 days starting 1 day before radiation and same amount
of SN52Mu was used as control. For anti-PD-L1 treatment experiments, 200 mg anti-PD-L1 (10F.9G2) or isotype control were given
by i.p. every three days for a total of four times starting on the day of IR.
Flow CytometryTumor tissues were cut into small pieces and digested by 1 mg/ml collagenase IV (Sigma) and 0.2 mg/ml DNase I (Sigma) for 1 hr at
37�C. Suspension cells were blocked with anti-FcR (2.4G2, BioXcell) and then stained with fluorescence-labeled antibodies against
CD11c, I-Ab, H-2Kb, CD80, CD86, CD8 andCD45.2. Flow cytometry were performed on BD LSRFortessa at core facility of University
of Chicago and data were analyzed with FlowJo software. For up-taking of tumor DNA test, MC38 tumor cells were labeled with
10 mM EdU (Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry Assay Kit) overnight and inoculated into mice (4 3 106 cells/mouse).
The next day, tumors were harvested and EdU and phoshpo-TKB1 or -p100 was analyzed by flow cytometry.
e3 Immunity 49, 490–503.e1–e4, September 18, 2018

In Vitro Culture and Function Assay of BMDCs83 106 MC38-SIY cells were plated into 10 cm cell culture dishes overnight, and then pretreated with 40 Gy and incubated for 12 hr.
BMDCs were added and co-cultured with MC38-SIY cells at the ratio of 1:1 in the presence of fresh GM-CSF for an additional 6–8 hr.
Subsequently purified CD11c+ cells with EasySep Mouse CD11c Positive Selection Kit II (STEMCELL) were incubated with isolated
CD8+ T cells from naive 2C mice for 3 days (Figure S3A). For IFN-b detection, 13 106 cells/ml purified CD11c+ cells from co-culture
were seeded into 96-well plates and the supernatants were harvested after 3 day incubation (Figure S3B).
Measurement of IFNg-Secreting CD8+ T Cells by ELISPOT Assay23 104 purified CD11c+ cells were incubated with CD8+ T cells isolated from naive 2C mice (EasySep Mouse CD8a Positive Selec-
tion Kit (STEMCELL)) for 3 days at the ratio of 1:10. For tumor-specific CD8+ T cells functional assay,MC38 tumor cells were exposed
to 20 ng/ml murine IFNg for 24 hr prior to plating with purified CD8+ T from DLN. 23 105 CD8+ T cells were incubated with MC38 at
the ratio of 10:1 for 48 hr. ELISPOT assays were performed to detect the cytokine spots of IFN-g according to product protocol
(Millipore).
ELISATumor tissues were homogenized in PBS with protease inhibitor followed by addition of Triton X-100. Cell culture supernatants were
obtained from isolated CD11c+ cells after 48 hr-incubation with fresh GM-CSF. The concentration of IFN-b and CXCL10 was
measured with VeriKine-HS Mouse Interferon Beta Serum ELISA Kit (PBL Assay Science) and mouse CXCL10 Quantikine ELISA
kit (R&D) in accordance with the manufacturer’s instructions, respectively.
Western Blot AnalysisWhole-cell protein was extracted with Triton X-100 buffer (150 mM sodium chloride, 50 mM Tris, 1% Triton X-100; pH 8.0) with pro-
teinase inhibitors and phosphatase inhibitors (Thermo Scientific). Cytoplasmic (CE) and nuclear (NE) protein were extracted with
NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific). Immuno-blotting analyses were performed as previously
described (Hou et al., 2014). The amount of loaded protein was normalized to GAPDHmAb (Proteintech Group) or HDAC1 mAb (Cell
Signaling).
Chromatin Immunoprecipitation AssaysBMDCs (2 3 107) were stimulated with 50 mg/ml DMXAA for 1 hr, flowed by fixation with 1% formaldehyde and sonication as previ-
ously described (Nelson et al., 2006). Lysates were subjected to immunoprecipitation with the indicated antibodies or control IgG,
and the precipitated DNA was then purified by Magna ChIP A/G Chromatin Immunoprecipitation Kit (Millipore) and quantified by
qPCR via a pair of primers that amplify the PRDIII-II region of the Ifnb promoter (forward: 50- ATTCCTCTGAGGCAGAAAG
GACCA-30; reverse: 50- GCAAGATGAGGCAAAGGCTGTCAA�30). Input DNAwas used as an internal control and the data are shown
as the relative fold increased over IgG control.
Immunoprecipitation AssaysWT BMDCs (2 3 107) were treated with 20 mM MG132 for 15 min followed by stimulating of 50 mg/ml DMXAA for indicated time.
Cell lysates were subjected to immunoprecipitation with 2 mg indicated antibodies by using Pierce Crosslink Magnetic Co-IP Kit
(ThermoFisher). The protein level in eluted supernatant were detected by using indicated antibodies with Western Blot.
In vitro Kinase AssayspCDNA3.1 vectors containing His-tagged TBK1WT (wild-type), TBK1 K38A (Lys38 mutated to Ala), TBK1 KDD (kinase domain dele-
tion) and HA-tagged wild-type p100 were purchased from GenScript. Recombinant proteins were expressed in HEK293 cells and
purified with His-tag and HA-tag isolation kits from Invitrogen and Thermo Scientific. In vitro kinase activities were analyzed by
reading the generation of ADP with Universal Kinase Activity Kit from R&D Systems.
QUANTIFICATION AND STATISTICAL ANALYSIS
Analyses were performed using GraphPad Prism software 6. Data were analyzed by one-way ANOVA with Tukey’s Multiple Com-
parison Test or Student’s t test. P values < 0.05 were considered statistically significant.
Immunity 49, 490–503.e1–e4, September 18, 2018 e4

Immunity, Volume 49
Supplemental Information
Non-canonical NF-kB Antagonizes STING
Sensor-Mediated DNA Sensing in Radiotherapy
Yuzhu Hou, Hua Liang, Enyu Rao, Wenxin Zheng, Xiaona Huang, Liufu Deng, YuanZhang, Xinshuang Yu, Meng Xu, Helena Mauceri, Ainhoa Arina, Ralph R.Weichselbaum, and Yang-Xin Fu

C
F*
A*
0
200
400
600
800
1000
1200
0 10 13 17 20 24 27 31
Tumor Volum
e (m
m3)
Days after tumor inoculation
B
20Gy
**
**
0
500
1000
1500
2000
2500
0 11 16 19 22 25 29 34
Tumor Volum
e (m
m3 )
Days after tumor inoculation
N.S.**D
20Gy
N.S.
*E
Suppl Figure 1 (Related to Figure 2). Host non-canonical NF-κB deficiency enhances therapeutic effect of ionizing radiation(A) Age- and sex-matched Relbf/f mice and Itgax-cre Relbf/f mice were inoculated s.c. with 1 × 106 MC38 cells on day 0. On day 10, tumors locally received one dose of 20Gy IR. Survival of tumor bearing mice was monitored.(B-C) Age- and sex-matched Relbf/f mice and Itgax-cre Relbf/f mice were inoculated s.c. with 1 × 106 B16-SIY cells on day 0. On day 10, tumors locally received one dose of 20Gy IR. Tumor growth (B) and survival of tumor bearing mice (C) were monitored.(D-E) Age- and sex-matched Relbf/f mice and Lyz2-cre Relbf/f mice were inoculated s.c. with 1 × 106 MC38 cells on day 0. On day 10, tumors locally received one dose of 20Gy ionizing radiation. Tumor growth (D) and survival of tumor bearing mice (E) were monitored.(F) Age- and sex-matched Nfkb2+/+ and Nfkb2-/- mice, bred to Rag1-/- background, were inoculated s.c. with 1 × 106 MC38 cells on day 0. On day 10, tumors locally received one dose of 20Gy ionizing radiation (IR) and the mice were transferred i.v. with 1 × 107 CD3+ T cells isolated from naive WT mice. Survival of tumor bearing mice was monitored.Representative data are shown from three experiments (two experiments for D and E) conducted with 4-6 mice in each group. Data are represented as mean ± SD. ∗p < 0.05, ∗∗p < 0.01.
*
Relbf/fRelbf/f+IRItgax‐cre Relbf/fItgax‐cre Relbf/f+IR
Relbf/fRelbf/f+IRItgax‐cre Relbf/fItgax‐cre Relbf/f+IR
Relbf/fRelbf/f+IRLyz2‐cre Relbf/fLyz2‐cre Relbf/f+IR
Relbf/fRelbf/f+IRItgax‐cre Relbf/fItgax‐cre Relbf/f+IR
Relbf/fRelbf/f+IRLyz2‐cre Relbf/fLyz2‐cre Relbf/f+IR
Rag1‐/‐Nfkb2‐/‐+IR
Rag1‐/‐Nfkb2+/++IRRag1‐/‐Nfkb2‐/‐
Rag1‐/‐Nfkb2+/+

D
0
10
20
30
CD45
+cells in
live cell (%) Control
IR
*****
A
****
***
0
1000
2000
3000
4000
5000
I‐Abin DCs (M
FI)
ControlIR
0
1000
2000
3000
4000
5000
6000
H2Kb
in DCs (M
FI)
ControlIR
B
****
0
1
2
3
4
5
CD11
c+ in
live cells (%
) ControlIR
GF
0
3000
6000
9000
12000
CD80
in DCs (M
FI)
ControlIR
***
Suppl Figure 2 (Related to Figure 2). Nfkb2 deficient mice showed higher anti-tumor immunity following IR (A-B and E-G) Age- and sex-matched Nfkb2+/+ and Nfkb2-/- mice were inoculated MC38 cells and tumors locally received local irradiation and T cells transfer as described in Figure 2B. (A-B) Four days after radiation, tumors were removed and digested into single-cell suspensions. The proportion of CD11c+ infiltrating DCs (A) and MHC class I, II and expression of co-stimulating molecules CD80 (B) on DCs were analyzed by flowcytometry.(C) MC38-SIY tumor model were established in Nfkb2+/+ and Nfkb2-/- mice and treated with IR as described in Figure 2B. On day 4 post IR, tumors were removed and digested into single cell suspension. The CD11c+ DCs were sorted and then co-cultured with isolated CD8+ T cells from naive 2C mice for 3 days and the cross-priming activity of tumor infiltrating DCs were analyzed by ELISPOT assays. The cross-priming ability of DCs was evaluated by counting IFN-γ+ spots.(D) BMDCs from Nfkb2+/+ and Nfkb2-/- mice were used for co-culture with irradiated or non-irradiated MC38-SIY cells. Subsequently cross-priming activity of purified BMDCs was analyzed by ELISPOT assays as described in Figure 2C.(E-F) Seven days after IR, tumors were removed and digested into single-cell. The proportion of infiltrating CD45+ hematopoietic cells (E) and CD8+ T cells (F) were analyzed by flowcytometry. (G) MC38 tumor model were established in Nfkb2+/+ and Nfkb2-/- mice and treated with IR as described in Figure 2B. On day 7 post IR, tumor draining lymph nodes (TDLNs) were removed and digested into single-cell suspensions. Tumor antigen-specific CD8+ T cell function was measured by ELISPOT assays by co-culturing purified TDLN CD8+ cells with IFN-γ-treated MC38 tumor cells.
0
30
60
90
120
IFNγ+
spots p
er 2x105
CD8+
T
ControlIR
****
0
200
400
600
800
IFNγ+
spots p
er 2x105
CD8+
T cells
ControlIR
****
0
1
2
3
4
CD8+
T in
live cells (%
)
ControlIR
*****
E
0
200
400
600
IFNγ+
spots p
er 2x105
CD8+
T cells
ControlIR
*****
C
Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐
Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐ Nfkb2+/+ Nfkb2‐/‐

**
0
2
4
6
8
10
Normalized
p‐TBK
1
0
2
4
6
8
10
Normalized
p‐IR
F3
A B
02468
1012
Normalized
RelB
0
2
4
6
8
10
Normalized
p‐p10
0
C
02468
10
Control
MC3
8MC3
8+IR
Control
MC3
8MC3
8+IRNormalized
p‐p10
0
02468
10
Control
MC3
8MC3
8+IR
Control
MC3
8MC3
8+IRN
ormalized
RelB
D E F
0
2
4
6
8
0h 1h 6h 0h 1h 6hNormalized
RelB
02468
10
0h 1h 6h 0h 1h 6hNormalized
p‐TBK
1
02468
10
0h 1h 6h 0h 1h 6hNormalized
p‐p10
0
0123456
0h 1h 6h 0h 1h 6hNormalized
RelB
0123456
Normalized
p‐p10
0
G
02468
1012
0h 1h 6h 0h 1h 6hNormalized
RelB
H I
01234
0h 1h 6h 0h 1h 6hNormalized
RelB
0
1
2
2
3
Normalized
TBK
1
0
1
2
3Normalized
p10
0
(C) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in cytoplasmic (CE) and nuclear (NE) extracts of purified Tmem17+/+ and Tmem173-/- BMDCs after co-cultured with irradiated or non-irradiated MC38 cells for 1 hour.(D) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in cytoplasmic (CE) and nuclear (NE) extracts of Tmem17+/+ and Tmem173-/- BMDCs stimulated with 50 μg/ml DMXAA for indicated time. (E) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in cytoplasmic (CE) and nuclear (NE) extracts of Tbk1+/+ and Tbk1-/- macrophages after stimulated with 50 μg/ml DMXAA for indicated time points. (F) Immuno-blotting analysis of indicated protein in whole-cell lysates of BMDCs after stimulated with 50 μg/ml DMXAA for indicated time points. (G) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Nfkb2+/+ and Nfkb2-/- BMDCs after stimulated with 50 μg/ml DMXAA for indicated time points. (H) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of BMDCs derived from Chukf/f and Itgax-cre Chukf/f mice after stimulated with 50 μg/ml DMXAA for indicated time points. (I) WT BMDCs were stimulated with 50 μg/ml DMXAA for indicated time in presence of proteasome inhibitor MG132 (20 µM). Whole cell lysates were subjected to p100 or TBK1 followed by detecting p100 or TBK1 associated TBK1 or p100 by immno-blotting.
Tmem173‐/‐Tmem173+/+ Tmem173‐/‐Tmem173+/+
Tmem173‐/‐Tmem173+/+ Tmem173‐/‐Tmem173+/+ Tbk1‐/‐Tbk1+/+ Tbk1‐/‐Tbk1+/+ Nfkb2‐/‐Nfkb2+/+
***
***
*****
** ** ** *** **
** ********** ******
**
****
N.S.**
** **Suppl Figure 3 (Related to Figure 4). Irradiated tumor cells promote non-canonical NF-κB activation in DCs in a STING-TBK1 dependent manner Phosphorylation or expression level of indicated protein was normalized against inner control. The graphs show densitometric quantification of immune-blotting, which are presented as relative changes against control group. Data are represented as mean ± SD. ∗∗p < 0.01 and ∗∗∗p < 0.001.(A-B) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in whole-cell lysates and nuclear (NE) extracts of purified BMDCs after co-cultured with irradiated or non-irradiated MC38 cells for indicated time.
Chukf/f Itgax‐cre Chukf/f

A B C
01223456
Normalized
IRF3
01234567
Normalized
RelA
01223456
0h0.5h 1h 2h 6h 12hN
ormalized
p‐Ik
Bα
01223456
0h0.5h 1h 2h 6h 12h
Normalized
RelA
02468
10
0h 1h 6h 0h 1h 6h
Normalized
RelA
0
1
2
3
4
0h 1h 6h 0h 1h 6h
Normalized
IRF3
0123456
0h 1h 6h 0h 1h 6h
Normalized
RelA
012345
0h 1h 6h 0h 1h 6h
Normalized
IRF3
DTmem173‐/‐Tmem173+/+ Tmem173‐/‐Tmem173+/+
Tbk1‐/‐Tbk1+/+ Tbk1‐/‐Tbk1+/+
Suppl Figure 4 (Related to Figure 5). Canonical NF-κB was required for STING mediated IFN-β production and IR-induced anti-tumor immunity(A-D, I) Phosphorylation or expression level of indicated protein was normalized against inner control. The graphs show densitometric quantification of immune-blotting, which are presented as relative changes against control group. Data are represented as mean ± SD. ∗∗p < 0.01 and ∗∗∗p < 0.001.(A) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of purified BMDCs after co-cultured with irradiated or non-irradiated MC38 cells for indicated time.(B) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in cytoplasmic (CE) and nuclear (NE) extracts of BMDCs stimulated with 50 μg/ml DMXAA for indicated time points.(C) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Tmem17+/+ and Tmem173-/-BMDCs stimulated with 50 μg/ml DMXAA for indicated time points. (D) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of Tbk1+/+ and Tbk1-/- macrophages stimulated with 50 μg/ml DMXAA for indicated time points. (E-F) ELISA assay was performed to measure IFN-β produced by WT and Tbk1-/- macrophages (E) or WT and Irf3-/- BMDCs (F) after stimulated with 50 μg/ml DMXAA for 24 h.(G) BMDCs were induced with bone marrow cells from WT and Relaf/f mice which were treated with 50 μg/ml TAT-Cre for 2 h. ELISA assay was performed to measure IFN-β produced by TAT-Cre pretreated WT and Relaf/f BMDCs after stimulated with 50 μg/ml DMXAA for 24 h. (H) ELISA assay was performed to measure IFN-β produced by BMDCs after stimulated with 50 μg/ml DMXAA for 24 h in absence or presence of 25 μM JSH-23.(I) Immuno-blotting analysis of indicated phosphorylated (p-) and total protein in cytoplasmic (CE) and nuclear (NE) extracts of Nfkb2+/+ and Nfkb2-/- BMDCs stimulated with 50 μg/ml DMXAA for indicated time points.(J) MC38 tumors established in Relbf/f mice and Itgax-cre Relbf/f mice were treated locally with one dose of 20Gy radiation or untreated. 6 mg/kg JSH-23 was given by gavage daily starting 1 day before radiation for totally 14 days. Tumor growth was monitored after radiation.(K) MC38 tumors established in Relaf/f mice and Itgax-cre Relaf/f mice were treated locally with one dose of 20Gy radiation or untreated. 2x10^10 Ad-IFN-β vp was given by i.t. on day 1 and day 3 post radiation. Tumor growth was monitored after radiation.
***
****
*****
****** **
*** ****
E
0
10
20
30
40
50
Irf3+/+ Irf3‐/‐
IFN‐β (p
g/ml)
ControlDMXAA
0
10
20
30
Tbk1+/+ Tbk1‐/‐
IFN‐β (p
g/ml)
ControlDMXAA
F G
0
50
100
150
Medium JSH‐23
IFN‐β (p
g/ml)
ControlDMXAA
0
50
100
150
IFN‐β (p
g/ml)
ControlDMXAA
H
*** *** ******
02468
10
0h 1h 6h 0h 1h 6h
Normalized
p‐TBK
1
012345
0h 1h 6h 0h 1h 6hNormalized
IRF3
01234
0h 1h 6h 0h 1h 6hNormalized
RelA
Nfkb2‐/‐Nfkb2+/+ Nfkb2‐/‐Nfkb2+/+ Nfkb2‐/‐Nfkb2+/+
0300600900
120015001800
0 10 12 14 17 20 26
Tumor Volum
e (m
m3 )
Days after tumor inoculation
N.S.I
0
400
800
1200
1600
0 10 12 14 17 26 30
Tumor Volum
e (m
m3 )
Days after tumor inoculation
K **
**
20Gy 20Gy
JItgax‐cre Relbf/f+JSH‐23
Relaf/fWT
Itgax‐cre Relbf/f+IR+JSH‐23
Relbf/f+JSH‐23Relbf/f+IR+JSH‐23
Itgax‐cre Relaf/f+IR+Ad‐IFN‐β
Relaf/f+Ad‐IFN‐βRelaf/f+IR+Ad‐IFN‐βItgax‐cre Relaf/f+Ad‐IFN‐β

Log‐rank P = 1.8e‐9
A
Log‐rank P = 2.5e‐8
B
Suppl Figure 5 (Related to Figure 6). Inhibition of non-canonical NF-κB can promote radiation-induced anti-tumor immunity(A-B) Overall survival analysis of glioblastoma patients with either high or low mRNA expression for Nfkb2 (A) or Relb (B). All patients received radiotherapy. Normalized Gene expression (RNAseq) and corresponding clinical data of patients with glioblastoma (GBM) were obtained from The Cancer Genome Atlas (TCGA) through the UCSC Cancer Genomics Browser.(C) Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of BMDCs pretreated with 40 μg/ml N52Mu or SN52 for 30 min and then stimulated with 50 μg/ml DMXAA for additional 1 h. (D) MC38 tumors established in B6 mice were treated locally with one dose of 20Gy radiation or untreated. 40 μg N52Mu or SN52 was administered i.t. in tumor-bearing mice on day -1, day 1 and day 3 of radiation. On day 4, tumors were removed and digested into single-cell suspensions and CD11c+ DCs and CD11c- non-DCs were sorted. Immuno-blotting analysis of indicated protein in nuclear (NE) extracts of purified DCs and non-DCs was performed.(E) WT BMDCs were pretreated with 40 μg/ml N52Mu or SN52 for 30 min and then stimulated with 50 μg/ml DMXAA for 24 h. ELISA assay was performed to measure IFN-β produced by BMDCs.(F) B16-SIY tumors established in B6 mice were treated locally with one dose of 20Gy radiation or untreated. 40 μg SN52Mu or SN52 was administered i.t. in tumor-bearing mice every 2 days for a total of four times staring from one day before radiation. Tumor growth was monitored after radiation.(G) MC38 tumors established in B6 mice were treated locally with one dose of 20Gy radiation or untreated. 40 μg SN52Mu or SN52 was administered i.t. in tumor-bearing mice every 2 days for a total of four times staring from one day before radiation. On the day of IR, 200 μg anti-PD-L1 (10F.9G2) or isotype control were given by i.p. every three days for a total of four times. Tumor rejection was monitored.
0 3 6 12 24 0 3 6 12 24
HDAC1
Time after IR (h)
RelB
SN52Mu
HDAC1
RelB
SN52
DCsNE
Non‐DCsNE
‐ + ‐ + ‐ +
HDAC1
RelA
DMXAARelB
NE
C D
0
10
20
30
40
50
60
SN52Mu SN52
IFN‐β (p
g/ml)
Control
DMXAA
*****
E
0
200
400
600
800
1000
1200
1400
0 10 12 14 17 20 26
Tumor Volum
e (m
m3 )
Days after tumor inoculation
SN52MuSN52Mu+IRSN52SN52+IR
F
****
G
20Gy
**
**
Top Related
Copyright © 2022 FDOKUMEN

