














Bahasa
Halaman
Hukum


Redox activity of AIF
1
Mo: 10498
NADH-oxidase activity of mitochondrialapoptosis-inducing factor (AIF)
M. Dolores Miramar1, Paola Costantini2, Luigi Ravagnan2,
Ligia M. Saraiva3, Delphine Haouzi2, Greg Brothers4,
Josef M. Penninger4, M. Luisa Peleato1*,
Guido Kroemer2*, and Santos A. Susin2*
1 Departamento de Bioquímica y Biología Molecular y Celular. Universidad de Zaragoza,
pza San Francisco s/n. 50009 Zaragoza, Spain
2 Centre National de la Recherche Scientifique, UMR1599, Institut Gustave Roussy, 39
rue Camille Desmoulins, F-94805 Villejuif, France;
3 Instituto de Tecnología Química y Biológica. Universidade Nova de Lisboa, rua da
Quinta Grande, 6. Apartado 127. 2780 Oeiras, Portugal;
4 The Amgen Institute and Ontario Cancer Institute, Department of Medical Biophysics
and Immunology, University of Toronto, 620 University Avenue, Suite 706, Toronto,
Ontario M5G 2C1, Canada.
Correspondence to: Dr. Guido KroemerCNRS-UMR 1599Institut Gustave RoussyPavillon de Recherche I39, rue Camille-DesmoulinsF-94805 VillejuifFranceTel. 33-1-42 11 60 46Fax 33-1-42 11 60 47e-mail: [email protected]
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on February 13, 2001 as Manuscript M010498200 by guest on O
ctober 8, 2016http://w
ww
.jbc.org/D
ownloaded from

Redox activity of AIF
2
Running title: Redox activity of AIF
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
3
Summary
Apoptosis-inducing factor (AIF) is a mitochondrial flavoprotein which translocates to the
nucleus during apoptosis and causes chromatin condensation and large scale DNA
fragmentation. Here we report the biochemical characterization of AIF's redox activity.
Natural AIF purified from mitochondria and recombinant AIF purified from bacteria
(AIF∆1-120) exhibit NADH oxidase activity while superoxide anion (O2-) is formed.
AIF∆1-120 is a monomer of 57 kDa containing 1 mol of non covalently bound FAD per
mol of protein. ApoAIF∆1-120, which lacks FAD, has no NADH oxidase activity.
However, native AIF∆1-120, apoAIF∆1-120, and the reconstituted (FAD-containing)
holoAIF∆1-120 protein exhibit a similar apoptosis-inducing potential when microinjected
into the cytoplasm of intact cells. Inhibition of the redox function, by external addition of
superoxide dismutase or covalent derivatization of FAD with diphenyleneiodonium, failed
to affect the apoptogenic function of AIF∆1-120 assessed on purified nuclei in a cell-free
system. Conversely, blockade of the apoptogenic function of AIF∆1-120 with the thiol
reagent para-chloromercuriphenylsulfonic acid did not affect its NADH oxidase activity.
Altogether, these data indicate that AIF has a marked oxidoreductase activity which can be
dissociated from its apoptosis-inducing function.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
4
Introduction
Mitochondria are considered as central players in apoptosis of mammalian cells (1-
3). Early during the apoptotic process, the outer mitochondrial membrane becomes
permeabilized, and mitochondria release soluble proteins normally confined to the
intermembrane space (4). Such apoptogenic proteins include the caspase activator
cytochrome c (5), procaspases 2, 3 and 9 (6-8), the IAP inhibitor Smac/DIABLO (9, 10),
as well as AIF (11). In contrast to cytochrome c and Smac/DIABLO, AIF is a caspase-
independent death effector, which translocates via the cytosol to the nucleus, where it
causes chromatin condensation and large-scale (50 kbp) DNA fragmentation (12, 13).
Neutralization of the AIF protein by microinjection of a specific antibody into the
cytoplasm of intact cells has revealed AIF to be rate-limiting for apoptotic chromatin
condensation and, in some cases, for mitochondrial membrane permeabilization (11, 14,
15). Conversely, microinjection of AIF may cause full-blown apoptosis with nuclear
condensation, dissipation of the mitochondrial transmembrane potential, release of
cytochrome c, and exposure of phosphatidylserine on the outer plasma membrane leaflet
(11, 14, 15).
The AIF precursor protein (612 aa) contains an N-terminal (first 100 aa) mitochondrial
localization sequence (MLS). The protein is synthesized in cytoplasmic ribosomes and
imported into the mitochondrial intermembrane space, where the MLS is cleaved off (11).
The C-terminal domain of AIF (last 485 aa) shares significant homology with
oxidoreductases from other vertebrates (X. laevis), non-vertebrate animals (C. elegans,
D. melanogaster), plants, fungi, eubacteria, and archaebacteria (16). The mature AIF
protein purifies as a flavoprotein, both from mitochondria and from E.coli used to
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
5
produce recombinant AIF (11). This fact prompted us to investigate the putative electron
transfer (redox) function of AIF, in relation to its apoptogenic activity. Indeed, apoptosis
is accompanied by a general shift of the redox balance characterized by a depletion of
NADH, NADPH, glutathione, as well as by an increase of free radicals, including
superoxide anion, lipid peroxidation products (such as 4-hydroxynonenal), and oxidative
damage of membranes and DNA (17). In several paradigms of apoptosis, culture in
anoxic conditions, treatments with cell-permeable antioxidants, or overexpression of anti-
oxidant enzymes (such as superoxide dismutase, glutathione peroxidase, catalase, the
thioredoxin system) have profound inhibitory effects on cell death (18-20). It appears that
the respiratory chain is a prime source for the generation of oxygen radicals (presumably
derived from uncoupling and/or interruption of electron transfer due to the release of
cytochrome c) (21). Moreover, mitochondrial targeting of anti-oxidant enzymes is
particularly efficient in blocking apoptosis in several models (19, 22).
Here we report the detailed biochemical characterization of the redox function of AIF,
which turns out to be an FAD-containing oxidase capable of oxidizing NAD(P)H while
generating superoxide anion. Interestingly, the electron transfer function of AIF can be
dissociated from its apoptogenic activity, both in cell-free systems and in intact cells.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
6
Experimental procedures
Cells and culture conditions. HeLa cells were cultured in DMEM supplemented with 2
mM L-glutamine, 1 mM Pyruvate, 100 mM Hepes, 100 U/ml penicillin/streptomycin,
and 10% decomplemented FCS (Life Technologies). These cells were used for the
purification of nuclei and cell free system experiments, as described (23). Rat-1 fibroblast
cells were cultured as above and were used in microinjection experiments.
Recombinant AIF proteins. AIF deletion mutant (∆1-120) and AIF deletion mutant (∆1-
351) (11) were expressed from a Novagen pET32 expression vector and purified from E.
coli. The proteins were stored at -80°C in 50 mM Hepes, pH 7.9, 100 mM NaCl, 2 mM
EDTA, 1 mM DTT and 10% glycerol.
Mass spectroscopy, HPLC and elemental analysis. Molecular mass was measured by
means of a MALDI system from Applied Biosystems at the Services Cientifico-Técnicos,
Barcelona University. Sinapinic acid was used as matrix and bovine serum albumin
(BSA) was used as standard protein. For the experimental analysis, AIF∆1-120 (1.8
mg/ml) was mixed with sinapinic acid (10 mg/ml in H2O:CH3CN 1:1 + 0.3% TFA),
using a 1:6 ratio. Gel filtration on a Superose 12 (Amersham Pharmacia Biotech) was
also used for molecular weight determination. The protein was eluted using 0.3 ml/min of
50 mM Tris-HCl pH 8, 2 mM EDTA, 1 mM DTT and 200 mM NaCl. Semi-quantitative
detection of metals was performed in a Perkin Elmer ELAN 6000 system and semi-
quantitative determination of calcium was performed in a multicanal Thermo Jarred Ash
61E Polyscan following standard protocols.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
7
SDS-PAGE. Proteins were run in a Phast System from Amersham Pharmacia Biotech,
following the manufacturer's instructions.
Free thiols' content of AIF∆1-120 preparations was determined by using a 50-fold molar
excess of Ellman´s reagent, 5.5´-dithiobis-(2-nitrobenzoate-(Nbs2) (DTNB) (24). The
total thiol content was confirmed in the presence of 8 M urea and excess NaBH4 (25).
AIF was quantified spectrophotometrically on the basis of the extinction coefficient
calculated in this work.
Absorption spectrometry studies were carried out using a Kontron Uvikon 860
spectrophotometer. The molar extinction coefficient of bound FAD at 450 nm was
determined based on the absorption changes detected after releasing the bound FAD from
the enzyme by heating (5 minutes at 90°C) in Tris-HCl 50 mM pH 8. An extinction
coefficient of 11.3 mM-1 cm-1 at 450 nm was assumed for the free FAD (26). Reductive
titrations were performed under anaerobic conditions at 25°C. The anaerobic enzyme
sample, in 50 mM Tris-HCl pH 8 was prepared in an anaerobiosis cuvette by sequential
air evacuation and re-equilibration with oxygen-free argon. Anaerobic NAD(P)H was
prepared identically, introduced by means of the titration syringe. Identical amounts of
NAD(P)H were added to the reference cuvette.
HPLC for identification and quantification of FAD was performed using a C18 Vydac
column. A linear gradient 0-100% of ammonium acetate 0.1 M pH 6 and methanol in 40
minutes was performed using 1ml/min flow rate. FMN, riboflavin and FAD were used as
standards. 1 mg of AIF∆1-120 in 1 ml 50 mM Tris-HCl pH 8, maintained in the dark,
was heated for 10 min at 90°C. After centrifugation, aliquots of the supernatant were
injected into the HPLC, and flavins were detected at 445 nm.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
8
Phosphorylated residues were determined by dot-blot analysis using mouse monoclonal
antiphosphoserine, antiphosphothreonine and antiphospho-tyrosine antibodies (Sigma)
and an antimouse IgG alkaline phosphatase conjugate (Sigma). The method used was
based on the procedure previously described (27), using PVDF membranes (Immobilon-
P from Millipore).
Visible redox titrations of AIF were performed under anaerobic conditions. Appropriated
mediators that covered a potential range from +11 mV to - 450 mV (1mM) (28) were
added to the protein solution (10 µM) in 50 mM Bis-Tris for pH 6.5 or 100 mM Tris-HCl
for pHs 7.5 and 9. Injection of small volumes of air-free sodium dithionite allowed the
reduction of the protein. A Crison 2002 digital potentiometer was used and spectra were
recorded on a Shimadzu UV-260 spectrophotometer. The reduction potential was
determined by following the absorbance changes at 450 nm.
Potential involvement of cysteinyl redox centers in electron transfers was tested at 25°C,
under anaerobic conditions, following the NADH-DTNB oxidoreductase assay described
by Ohnisni, K. et al. (29). Briefly, 18 µM AIF∆1-120 in 50 mM sodium phosphate pH 7
containing 0.5 mM EDTA was mixed with 0.5 mM NADH, 0.02% BSA. The reaction
was started adding 0.4 mM DTNB and the nitrothiobenzoate anion production was
monitored at 412 nm using an extinction coefficient of 13.6 mM-1 cm-1.
Apoprotein preparation and holoprotein reconstitution. AIF∆1-120 apoprotein was
prepared following the protocol described by Chapman, S.K. and Reid, G.A. (30) by
exhaustive dialysis against 0.1 M Hepes, 2.5 M CaCl2, 1mM DTT, 0.1mM EDTA, 0.1M
guanidine chloride, 17% glycerol (v/v) at pH 7.5, concentration on Centricon 30 K
(Amicon) membranes, and a second round of dialysis against 50 mM Hepes, 100 mM
NaCl, 2mM EDTA, 1mM DTT, 10% glycerol at pH 7.9. The yield of this preparation
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
9
was ~15%. Reconstitution of the holoprotein was performed by incubation with 1000
times molar excess of FAD, acetone precipitation, and repeated ultracentrifugation on
Centricon 30 K membranes to remove non-bound FAD.
Determination of AIF redox activities. Initial velocity studies of the NAD(P)H-oxidase
activity of the flavoprotein followed assay procedures described previously (31). Briefly,
NAD(P)H oxidase activity was measured at 25°C in a total volume of 0.5 ml containing
0.25 mM NAD(P)H in air-saturated 50 mM Tris-HCl pH 8 buffer. The reaction was
initiated by the addition of the enzyme and was followed by the decrease in absorbance at
340 nm. Steady-state kinetic data were obtained by varying NADH concentration. One
unit of activity is defined as the amount of protein required to catalyse the conversion of 1
µmol of NAD(P)H to NAD(P)+ per min at 25°C. NBT reduction and
monodehydroascorbate reductase activity (32, 33), superoxide formed in the reaction of
AIF∆1-120 with oxygen (34) and hydrogen peroxide production (35) were quantified as
described. This latter reaction was carried out coupled with NADH oxidase in a mixture
containing 1mM sodium phosphate buffer pH 6.9, 2.34 mg/ml phenol, 1mg/ml 4-
aminoantipyrine and 0.02 units of horseradish peroxidase in a total volume of 0.5 ml.
The absorbance was measured at 505 nm, and the concentration of H2O2 was calculated
from a calibration curve (36). DCPIP (85 µM) or ferricyanide (2mM) reduction were
assayed as described (37). SOD inhibition was measured by adding the indicated units of
enzyme to the reaction mixtures. The electron transfer between NADH-AIF-
ferredoxin/adrenodoxin-cytochrome c was assayed as described (37), using AIF∆1-120
instead of ferredoxin-NADP+ reductase and adrenodoxin instead of ferredoxin.
AIF western blot. Supernatants obtained from mitochondria undergoing permeability
transition were subjected to a 10% SDS-PAGE and transferred (100 V, 75 min at room
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
10
temperature) to a nitrocellulose membrane. AIF immunoblot analysis was performed
using a rabbit antiserum generated against a mixture of three peptides derived from the
mouse AIF amino acids 151-200 (11).
In situ detection of 2, 2´-Di-p-nitrophenyl-5-5´-diphenyl-3,3´[3-3´-dimetoxy-4-
4´difenilen] tetrazolium chloride (NBT) reduction on native-PAGE was done using the
reaction mixture described by Pez-Huertas, E. L. et al. (32). Briefly, samples obtained
from mitochondria undergoing permeability transition were loaded onto a 10% native-
PAGE. The gel was incubated 20 min in the dark with 2 mM NBT solution. Then, 1 mM
NADH was added to reduce NBT and the reaction was stopped with water after the
appearance of the blue band.
Cell-free systems of nuclear apoptosis. Purified HeLa cell nuclei (103/µl) were exposed
90 min at 37°C to AIF∆1-120 preincubated (15 min at 37°C) with or without NADH,
NADPH, para-chloromercuriphenylsulfonic acid, superoxide dismutase, or
diphenyleneiodonium. For the standard assessment of chromatin condensation, nuclei
were stained with Hoechst 33342 (2 µM, 15 min, room temperature) and analyzed by
fluorescence microscopy (Leica DM IRB). DNA content was determined by staining with
propidium iodide (PI; 10 µg/ml) followed by analysis in a FACS Vantage (Becton-
Dickinson). A minimum of 2500 events were scored.
Microinjection. Rat-1 fibroblasts were microinjected using a computer-controlled
microinjector (pressure 150 hPa; 3 sec; Eppendorf) with buffer only, 7.5 µM of AIF∆1-
120, apoAIF∆1-120, apoAIF∆1-120, FAD-reconstituted holoprotein, and AIF∆1-351.
After microinjection, cells were cultured at 37°C for 180 min and stained with the ∆Ψm-
sensitive dye CMXRos (100 nM, 15 min) and the DNA-intercalating dye Hoechst 33342
(1.5 µM, 15 min) (11). Microinjected viable cells (100 per session, three independent
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
11
sessions of injections) were identified by inclusion of 0.25% (w:v) FITC-dextran (green
fluorescence) in the injectate. Only the blue and the red fluorescence was recorded.
All chemical reagents used in this work were purchased from Sigma.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
12
Results
Properties of recombinant AIF protein. Recombinant AIF∆1-120 was found to elute as a
single peak on a gel filtration column, with a calculated molecular weight of 57 kDa,
which corresponds to its theoretical molecular mass (57046), indicating that, at near-to
physiological salt concentrations (200 mM NaCl), AIF∆1-120 is a monomer. This result
was confirmed by mass spectroscopy analysis (not shown). It is in contrast with its
apparent molecular weight determined by SDS-PAGE (67.5 kDa, about 10 kDa more
than expected), a migration behaviour reported for other proteins such as ferredoxins
(38). AIF∆1-120 does not contain significant amounts of metals including Ca2+, Co2+,
Cu2+, Mg2+, Fe2+, Se2+ and Zn2+. However, it contains phosphogroups among serine
and threonine residues, as revealed by immunoblotting (not shown). AIF∆1-120 has 3
cysteins in its sequence, and it was found to contain 3.2 accessible thiol groups per
molecule, as determined by derivatization with the Ellman's reagent. Total thiol content
was confirmed after unfolding in 8 M urea. This suggests that none of the three cysteins
contained in the AIF amino acid sequence engages in disulfide links. The flavin moiety of
AIF∆1-120 is non-covalently bound and was identified by HPLC as FAD (not shown).
FAD is the only identified prosthetic group present in AIF∆1-120, at a molar ratio of 1:1.
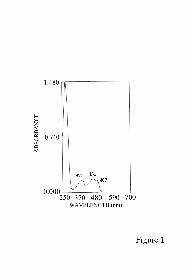
The absorption spectrum of AIF (Fig. 1) shows the typical features of an oxidized FAD
flavoprotein, with the visible maximum at 378 nm and 450 nm and a shoulder at 467 nm.
The ratio A270nm/A450nm was 7 in pure preparations, and the extinction coefficient for
oxidized AIF at 450 nm was calculated to be 12.12 mM-1 cm-1.
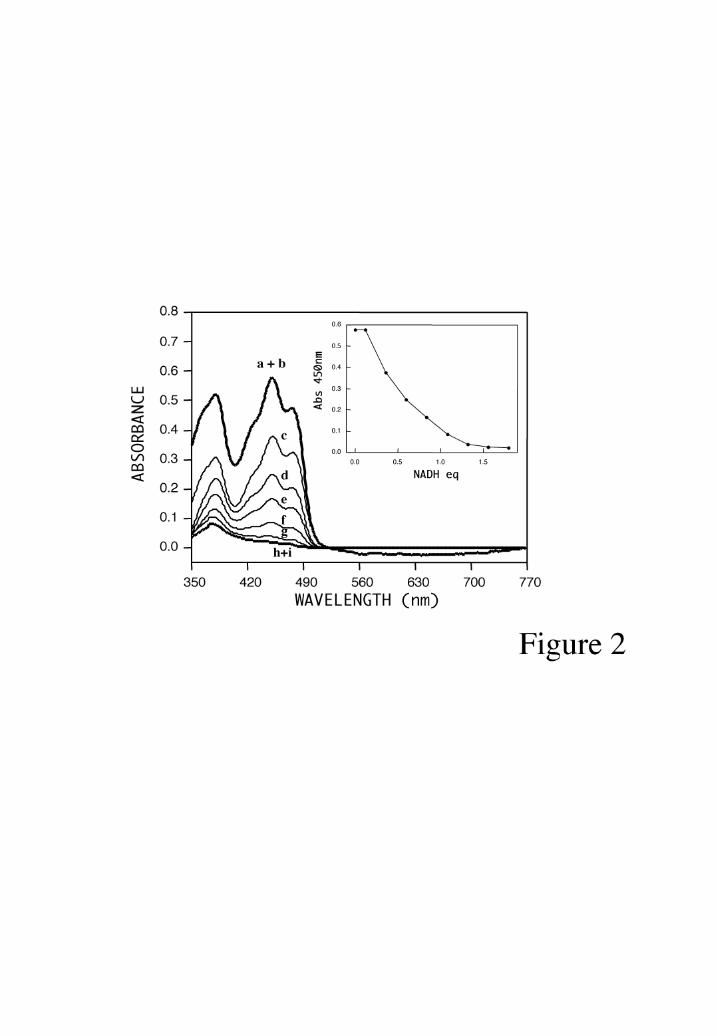
Electron transfer reactions from NADH and NADPH to AIF. The addition of an
equimolar amount of NADH to AIF∆1-120 in anaerobic conditions leads to complete
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
13
flavin reduction, without intermediate semiquinone formation (Fig. 2). Similar titration
curves were obtained using NADPH as reductant, and in both cases the spectral changes
are similar to the reduction of AIF∆1-120 with dithionite (not shown). The reduced form
was stable over several hours, and admission of air to the sample did not lead to the
immediate appearance of the oxidized AIF spectrum. Upon addition of an increasing
molar excess of NADH or NADPH over AIF∆1-120, the appearance of long wavelength
absorbance bands was observed (Fig. 3). These long-wavelength absorbances of the
reduced enzyme were stable at 25°C for hours, even upon exposure to air. They exhibit
the blue-green and green color described for other electron transfer complexes (31). By
analogy to other enzymes (31), these long-wavelength absorptions are likely to
correspond to charge transfer complexes between the reduced FAD and tightly bound
NAD+ or NADP+.
Redox potential of AIF. The spectral titration of AIF∆1-120 with dithionite revealed that
the redox potential of AIF is strongly influenced by the pH (Fig. 4). Assuming a two-
electron reduction step, midpoint redox potentials were determined to be -264 mV+15 mV
at pH 6.5 (Fig. 4A) , -308 mV + 15 mV at pH 7.5 (Fig. 4B), and -373 mV + 15 mV at
pH 9.0 (Fig. 4C). Neither semiquinone formation nor long-wavelength absorbing bands
were detected upon reduction by dithionite. A plot of AIF's redox potential
(FAD/FADH2) versus pH has a slope of -44 mV (not shown). This slope deviates from
that expected for a two-electron reduction involving two protons (58 mV/pH unit) or one
proton (29 mV/pH unit). The deviation from theoretical values indicates the possible
presence of other dissociable groups whose pKa values are linked to the redox state of the
enzyme. Eventhough titrations with dithionite and NAD(P)H showed an uptake of two
electrons, it was considered important to discard the involvement of cysteinyl residues as
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
14
active redox acceptors: in the NADH:DTNB oxidoreductase assay no involvement of
cysteinyl residues in redox transferences was detected (not shown).
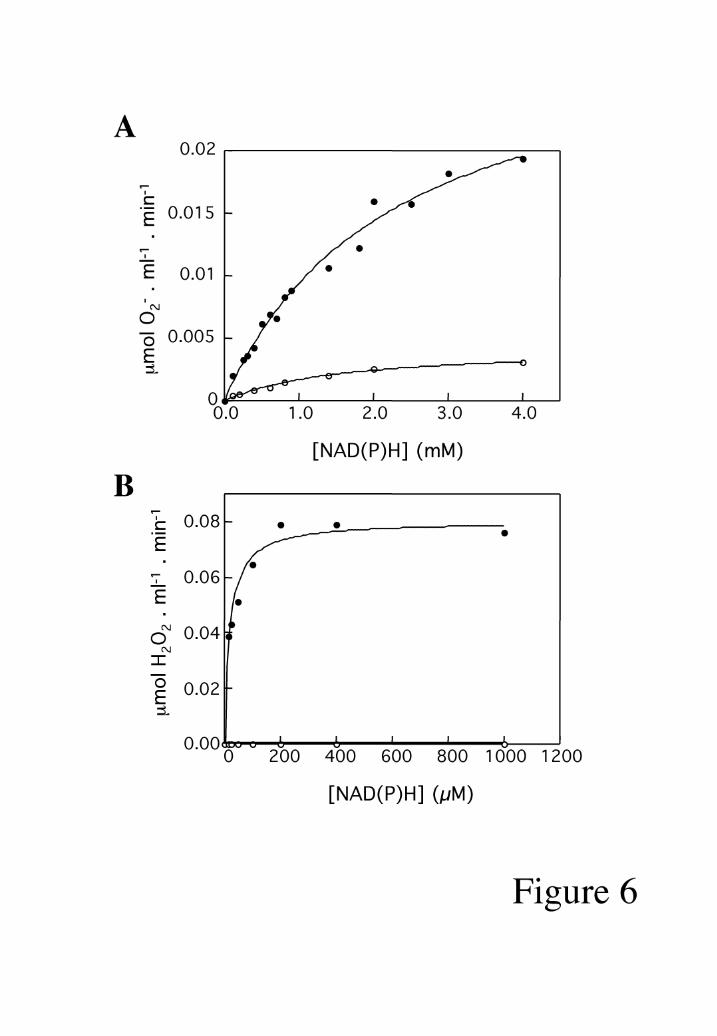
AIF redox activities. AIF∆1-120 was found to exhibit NADH- and NADPH-oxidase
activities (Fig. 5A). NAD(P)H oxidation in presence of AIF was followed measuring
initial rates of ∆A340 nm. The apparent Km for NADH was calculated as 99.4+10 µM
and the turnover number 2.09 min-1. When NADPH was used as electron donor, the
apparent Km was 52.9+ 12 µM and the turnover number 2.8 min-1 (Table 1). These
kinetic parameters are very similar to previous values described for other superoxide
forming NADH oxidases (39), and the steady-state kinetic data may be interpreted taking
into account the possible formation of relatively stable charge-transfer complexes.
Addition of exogenous FAD did not stimulate the NADH oxidase activity of AIF∆1-120
(not shown), in contrast to several NADH oxidases from bacteria (29, 35, 40-42). When
oxidizing NADH or NADPH, AIF catalyzed the reduction of the tetrazolium salt NBT
(Fig. 5B). This was due to the AIF(FADH2)-mediated reduction of O2 to O2- (Fig. 6A).
Accordingly, the reduction of NBT was completely abolished by the O2- scavenger
superoxide dismutase (SOD, 40 U/ml). Hydrogen peroxide formation resulting from
superoxide production was also measured (Fig. 6B). AIF did not exhibit any peroxidase
activity (not shown).
AIF∆1-120 was also found to possess a monodehydroascorbate reductase activity with a
specific activity of 8.8 U/mg and a kcat of 0.505 min-1, a feature that is common to
several AIF's homologous described in plants (16). The AIF-monodehydroascorbate
reductase activity was inhibited by SOD, indicating that the reaction occurs via superoxide
radicals (not shown). Cytochrome c reductase activity was also measured, with a Km of
0.46 mM for NADH and a kcat of 21.76 min-1 (Fig. 7A). Again, SOD inhibited the
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
15
cytochrome c reductase activity with an IC50 of ~10 U/ml, indicating that the AIF-
catalyzed cytochrome c reduction occurs via O2- radicals (Fig. 7B). When ferrodoxin
was used as mediator (NADH-AIF-ferredoxin-cytochrome c) it did not modify the
cytochrome c reduction rate (not shown). In contrast, when adrenodoxin was used as
mediator (NADH-AIF-adrenodoxin-cytochrome c), cytochrome c reduction was
increased (Fig. 7C). The kinetic parameters calculated from Figure 7C gave an apparent
Km for adrenodoxin of 1.08 µM with a kcat of 18.58 min-1.
AIF∆1-120 was found to transfer electrons at low rate from NADH (but not from
NADPH) to DCPIP or ferricyanide, two typical electron acceptors of NAD(P)H
dehydrogenases. From NADH, a two electron donor, AIF∆1-120 was able to reduce
DCPIP, a two electron acceptor, with a kcat of 21.6 min-1 and a calculated Km for the
NADH of 3.4 + 0.4 mM (Table 1). SOD did not affect the electron transfer rate indicating
that the diaphorase activity of AIF is not mediated by O2-. When the diaphorase activity
was assayed using a one-electron acceptor, ferricyanide, the catalytic efficiency for
NADH was reduced by half approximately.
In summary, AIF exhibits NADH oxidase activity and is able to transfer one electron (to
molecular oxygen and ferricyanide) or two electrons (to DCPIP), as summarized in Table
1.
AIF is the dominant NADH oxidase released through the mitochondrial outer
membrane. Purified mitochondria can be induced to undergo the so-called permeability
transition, a manipulation that leads to osmotic swelling of the matrix and physical rupture
of the outer membrane, causing the release of soluble intermembrane proteins (4). Upon
induction of permeability transition with Ca2+ or arsenite, immunodetectable AIF was
found in the mitochondrial supernatant. The release of AIF was inhibited by cyclosporin
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
16
A, a specific inhibitor of the permeability transition pore (Fig. 8). Separation of proteins
via native PAGE, followed by the in situ detection of an NADH oxidase activity causing
the reduction of NBT, yielded one single blue band (Fig. 8A). This band also reacted
with a specific anti-AIF antiserum and co-migrated with recombinant AIF∆1-120 (Fig.
8B). Altogether, these data confirm that natural (mitochondrial) AIF possesses a NADH
oxidase/NBT reductase activity and indicate that AIF is the quantitatively most important
NADH oxidase/NBT reductase contained in the mitochondrial intermembrane space.
Apoptotic versus oxidoreductase activity of AIF. The FAD moiety of AIF∆1-120 can be
removed by dialysis in the presence of guanidine chloride (a manipulation which
generates apoAIF∆1-120) and may be reconstituted by external addition of FAD (a
manipulation which generates reconstituted holoAIF∆1-120). ApoAIF lacked any
detectable NADH oxidase-NBT reductase activity, whereas the reconstituted holoAIF had
a NADH oxidase-NBT reductase activity undistinguishable from native AIF∆1-120 (Fig.
9A). Unmanipulated AIF∆1-120, apoAIF∆1-120, and reconstituted holoAIF∆1-120 were
micro-injected into the cytoplasm of Rat-1 fibroblasts. All three protein preparations
induced a similar level of nuclear chromatin condensation, as well as a dissipation of the
mitochondrial transmembrane potential (Fig. 9B, C). A deletion mutant of AIF∆1-351,
which lacks part of the oxidoreductase domain, yielded negative results in this system
(Fig. 9C). The nuclear effects of AIF were recapitulated in a cell-free system, in which
AIF∆1-120 was added to purified HeLa nuclei. In such a system, AIF∆1-120 caused
marked peripheral chromatin condensation (Fig. 10A) as well as a loss in DNA content
(Fig. 10B). Addition of NADH or NADPH failed to enhance the apoptogenic activity of
AIF∆1-120 (Fig. 10C). Moreover, inhibition of the oxidoreductase activity by external
addition of SOD or diphenyleneiodonium, an inhibitor of flavonoid-containing enzymes
covalently reacting with FAD (43-45), failed to modify the apoptogenic activity of AIF
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
17
(Fig. 10D). In contrast, addition of para-chloromercuriphenylsulfonic acid, a thiol-
reactive agent, did abolish the apoptogenic activity of AIF∆1-120, yet did not affect its
NTB reductase activity (Fig. 10D).
Altogether, these data show that the apoptogenic and oxidase functions of AIF can be
dissociated.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
18
Discussion
The results from this work indicate that AIF has a marked NADH oxidase activity.
According to the classification by Massey (46), AIF may belong to the electron-
transferase class of NADH reductases, because it reacts rapidly with oxygen, forming
O2- and the flavoprotein neutral radical as products. No semiquinone intermediate could
be detected although such a neutral radical is expected to be produced after the transfer of
a single electron. In each of the activities described, the reaction could be initiated by the
reduction of AIF by the two electron donors NAD(P)H, in the absence of exogenous
electron acceptor. The reduced form of AIF reacted with one electron (molecular oxygen
and ferricyanide) or two electron (DCPIP) acceptors. Kinetic analysis of the AIF steady-
state data are rather difficult to interpret given the formation of relatively stable charge
transfer complexes.
Several NADH oxidases from bacterial sources have been isolated and characterized (31,
35). The putative role of those enzymes is to maintain the cellular redox balance under
aerobic conditions, by converting NADH to NAD+ (42, 47). Also, several poorly
characterized superoxide (O2-) forming NADH oxidases have been detected in animal
cells, namely in heart mitochondria (48), liver mitochondria (49), microsomes from
bovine myocytes (50), and endothelial cells (51). However, AIF is not similar to any of
the previously described NADH oxidases. First it is monomeric, whereas bacterial
NADH oxidases are usually dimeric or tetrameric (52). Second, AIF transfers electrons
without the involvement of cysteinyl groups, whereas other NADH oxidases rely on a
redox-active disulphide center constituted by two vicinal cysteine residues (35, 42).
Third, AIF oxidizes NADH via a mechanism that does not require the addition of
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
19
exogenous FAD whereas several NADH oxidases from bacteria do require FAD (29, 35,
40-42). Fourth, in contrast to several NADH oxidases (42), AIF does not function as a
hydrogen peroxide scavenger.
AIF protein is present in the mitochondria of all mouse tissues that have been assessed,
and has also been found in a panel of 60 human cancer cell lines (53), suggesting that
AIF may fulfill important metabolic functions. However, based on the present data, it is
difficult to understand what the physiological function of AIF in normal (non-apoptotic)
conditions may be. Since AIF is the only NADH oxidase detected in the intermembrane
space, it is tempting to speculate that AIF accounts for the mitochondrial superoxide anion
or hydrogen peroxide-generating NADH oxidase activity (54, 55), which is lost from
mitochondria, once cells have been induced to die (55). Clearly, an NADH oxidase
activity causing the collateral generation of superoxide anion radicals would be of no
advantage for the cell. It thus may be speculated that the true, yet-to-be-discovered
substrates of the AIF oxidoreductase compete for endogenous NADH and/or that AIF is
normally inactivated by local inhibitory factors within the intermembrane space. If AIF
acted as a superoxide-generating NADH reductase outside of mitochondria, after its
apoptotic release, what might be the contribution of this enzymatic activity to the apoptotic
process? Apoptosis is notoriously associated with a massive depletion of NADH/NADPH
(56), as well as an increase in the generation of superoxide anions (57), at both
mitochondrial and extramitochondrial localizations (17). Furthermore, it should be
mentioned that, in isolated mitochondria, the permeability transition pore complex
(PTPC) is tightly regulated by the oxidation-reduction state of the pyridine nucleotide
pool, with oxidation causing an increase in the pore opening probability (58). Although
these changes in the redox potential may be explained by a variety of factors, including
uncoupling/blockade of the respiratory chain (21) and activation of PARP (59), it will be
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
20
interesting to study the contribution of AIF to this process, for instance in embryonic
stem cells in which the AIF gene is ablated.
It appears that the known apoptogenic functions of AIF and its novel oxidoreductase
activity can be dissociated from each other, based on three arguments. First, removal of
the prosthetic FAD group (which obviously abolishes the oxidoreductase function of
AIF) does not curtail the apoptogenic effects of AIF on mitochondria and nuclei of
microinjected cells. Second, addition of NADH, addition of SOD, or covalent
derivatization of FAD failed to modulate the capacity of AIF to induce nuclear apoptosis
in a cell-free system. Third, inhibition of the apoptogenic effect of AIF by means of para-
chloromercuriphenylsulfonic acid failed to affect its NADH oxidase activity. These data
are similar to those obtained for cytochrome c in the sense that the apoptogenic activity of
cytochrome c does not depend on its redox status. Exchange of the Fe2+ by Co2+ within
the heme prosthetic group of cytochrome c (a manipulation which abolishes the electron
transfer function of heme) fails to alter its caspase-activatory functions (60), whereas
certain amino acid substitutions which do not affect its redox function do abrogate
cytochrome c-mediated caspase activation (60, 61). In conclusion, both cytochrome c and
AIF thus appear to be bifunctional molecules with clearly dissociable redox and
apoptogenic activities.
Acknowledgments
We would like to thank Dr. Rita Bernhartd (Universität des Saarlandes,
Saarbrucken, Germany) for kindly providing adrenodoxin.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
21
References
1. Green, D. R., and Kroemer, G. (1998) Trends Cell Biol. 8, 267-271.
2. Green, D. R., and Reed, J. C. (1998) Science 281, 1309-1312.
3. Kroemer, G., and Reed, J. C. (2000) Nat. Med. 6, 513-519.
4. Patterson, S., Spahr, C. S., Daugas, E., Susin, S. A., Irinopoulos, T., Koehler, C., and Kroemer, G.
(2000) Cell Death Differ. 7, 137-144.
5. Budijardjo, I., Oliver, H., Lutter, M., Luo, X., and Wang, X. (1999) Annu. Rev. Cell Dev. Biol. 15,
269-290.
6. Mancini, M., Nicholson, D. W., Roy, S., Thornberry, N. A., Peterson, E. P., Casciola-Rosen, L. A.,
and Rosen, A. (1998) J. Cell Biol. 140, 1485-1495.
7. Krajewski, S., Krajewska, M., Ellerby, L. M., Welsh, K., Xie, Z. H., Deveraux, Q. L., Salvesen, G.
S., Bredesen, D. E., Rosenthal, R. E., Fiskum, G., and Reed, J. C. (1999) Proc. Natl. Acad. Sci.
USA 96, 5752-5757.
8. Susin, S. A., Lorenzo, H. K., Zamzami, N., Marzo, I., Larochette, N., Alzari, P. M., and Kroemer,
G. (1999) J. Exp. Med. 189, 381-394.
9. Du, C., Fang, M., Li, Y., Li, L., and Wang, X. (2000) Cell 102, 33-42.
10. Verhagen, A. M., Ekert, P. G., Pakusch, M., Silke, J., Connolly, L. M., Reid, G. E., Moritz, R.
L., Simpson, R. J., and Vaux, D. L. (2000) Cell 102, 43-53.
11. Susin, S. A., Lorenzo, H. K., Zamzami, N., Marzo, I., Snow, B. E., Brothers, G. M., Mangion, J.,
Jacotot, E., Costantini, P., Loeffler, M., Larochette, N., Goodlett, D. R., Aebersold, R., Siderovski,
D. P., Penninger, J. M., and Kroemer, G. (1999) Nature 397, 441-446.
12. Daugas, E., Susin, S. A., Zamzami, N., Ferri, K., Irinopoulos, T., Larochette, N., Prevost, M. C.,
Leber, B., Andrews, D., Penninger, J., and Kroemer, G. (2000) FASEB J. 14, 729-739.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
22
13. Susin, S. A., Daugas, E., Ravagnan, L., Samejima, K., Zamzami, N., Loeffler, M., Costantini, P.,
Ferri, K. F., Irinopoulou, T., Prévost, M.-C., Brothers, G., Mak, T. W., Penninger, J., Earnshaw,
W. C., and Kroemer, G. (2000) J. Exp. Med. 192, 577-585.
14. Susin, S. A., Larochette, N., Geuskens, M., and Kroemer, G. (2000) Meth. Enzymol. 322, 205-208.
15. Ferri, K. F., Jacotot, E., Blanco, J., Esté, J. A., Zamzami, A., Susin, S. A., Brothers, G., Reed, J.
C., Penninger, J. M., and Kroemer, G. (2000) J. Exp. Med. 192, 1081-1092.
16. Lorenzo, H. K., Susin, S. A., Penninger, J., and Kroemer, G. (1999) Cell Death Differ. 6, 516-524.
17. Kroemer, G., Petit, P. X., Zamzami, N., Vayssière, J.-L., and Mignotte, B. (1995) FASEB J. 9,
1277-1287.
18. Zhang, P., Liu, B., Kang, S. W., Seo, M. S., Rhee, S. G., and Obeid, L. M. (1997) J. Biol. Chem.
272, 30615-30619.
19. Bai, J., and Cederbaum, A. I. (2000) J. Biol. Chem. 275, 19241-19249.
20. Kampranis, S. C., Damianova, R., Atallah, M., Toby, G., Kondi, G., Tsichlis, P. N., and Makris,
A. M. (2000) J. Biol Chem. 275, 29207-29216.
21. Cai, J., and Jones, D. P. (1998) J. Biol. Chem. 273, 11401-11404.
22. Coulter, C. V., Kelos, G. F., Lin, T. K., Smith, R. A., and Murphy, M. P. (2000) Free Radic.
Biol. Med. 15, 1547-1554.
23. Susin, S. A., Zamzami, N., Larochette, N., Dallaporta, B., Marzo, I., Brenner, C., Hirsch, T., Petit,
P. X., Geuskents, M., and Kroemer, G. (1997) Exp. Cell Res. 236, 397-403.
24. Ellman, G. L. (1959) Arch. Biochem. Biophys. 82, 70-77.
25. Cavallini, D., Graziani, M. T., and Drupe, S. (1966) Nature (London) 212, 294-295.
26. Whitby, L. G. (1953) Biochem. J. 54, 437-442.
27. Ternynck, T.H. and Avrameas, S. (1989). Tecnicas de inmunologia: Tecnicas inmunoenzimaticas.
Grupo Editorial Iberoamericana, Mexico.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
23
28. Bes, M. T., Parisini, E., Inda, L. A., Saraiva, L. R., Peleato, M. L., and Sheldrick, G. (1999)
Structure 7, 1201-1213.
29. Ohnisni, K., Niimura, Y., Yokoyama, K., Hidaka, M., Masaki, H., Uchimura, T., Suzuki, K.,
Uozumi, T., Kozaki, M., Komagata, K., and Nishimo, T. (1994) J. Biol. Chem. 269, 31418-31423.
30. Zanetti, G., Cidaria, D., and Curti, B. (1982). Eur. J. Biochem. 126, 453-458.
31. Ahmed, S. A., and Claiborne, A. (1989) J. Biol. Chem. 264, 19856-19863.
32. Pez-Huertas, E. L., Corpas, F. J., Sandalio, L. M., and Del Roeo, L. A. (1999) Biochem. J. 337,
531-536.
33. Lumper, L., Schneider, W., and H.J., S. (1967) Biol Chem.Joppe-Seyler 348, 323-328.
34. McCord, J. M., and Fridowich, I. (1969) J.Biol. Chem 244, 6049-6055.
35. Arcari, P., Masullo, L., Masullo, M., Cantazano, F., and Bocchini, V. (2000) J. Bio. Chem. 275,
895-890.
36. Green, M. J., and Hill, A. O. (1984) Methods in Enzymology 105, 15-16.
37. Sancho, J., Peleato, M. L., Gomez-Moreno, C., and Edmondson, D. E. (1988) Arch. Biochem.
Biophys. 260, 200-207.
38. Böhme, H., and Schrautemeier, S. (1987) Biochim. Biophys.Acta 981, 1-7.
39. Glass, G. A., DeLisle, D. M., P., D., Gabig, T. G., Magee, B. H., Marker, M., and Babior, B. M.
(1986) J. Biol. Chem. 261, 13247-13251.
40. Niimura, Y., Yokoyama, K., Ohnisni, K., and Massey, V. (1994) Biosci.Biothecnol. Biochem. 58,
2310-2311.
41. Niimura, Y., Poole, L. B., and Massey, V. (1995) J. Biol. Chem. 270, 25645-25650.
42. Toomey, D., and Mayhew, S. G. (1998) Eur. J. Biochem. 251, 935-945.
43. O'Donnel, V. B., Tew, D. G., Jones, O. T. G., and England, P. J. (1993) Biochem. J. 290, 41-49.
44. Majander, A., Finel, M., and Wikstrom, M. (1994) J. Biol. Chem. 269, 21037-21042.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
24
45. Coves, J., Lebrun, C., Gervasi, G., Dalbon, P., and Fontecave, M. (1999) Biochem. J. 342, 465-
472.
46. Massey, V. (1994) J. Biol. Chem. 269, 22459-22462.
47. Niimura, Y., Ohnisni, K., Yarita, Y., Hidaka, M., Masaki, H., Uchimura, T., Suzuki, H., Kozaki,
M., and Uozumi, T. (1993) J. Bacteriol 175, 7945-7950.
48. Nohl, H. (1987) Eur. J. Biochem. 169, 585-591.
49. Linke, B., Henke, W., and Gerber, G. (1993) Ren. Physiol. Biochem. 16, 244-248.
50. Mohazzab, K. M., Kaminsky, P. M., and Wollin, M. S. (1997) Circulation 96, 614-620.
51. Zalba, G., Beaumont, F. J., San José, G., Fortuno, A., Fortuno, M. A., Etayo, J. C., and Diez, J.
(2000) Hypertension 35, 1055-1061.
52. Jarasch, E. D., Grund, C., Bruder, G., Heid, H. W., Keenan, T. W., and Franke, W. W. (1981) Cell
25, 67-82.
53. Daugas, E., Nochy, D., Ravagnan, L., Loeffler, M., Susin, S. A., Zamzami, N., and Kroemer, G.
(2000) FEBS Lett. 476, 118-123.
54. Linke, B., Henke, W., and Gerber, G. (1993) Renal Physiol. Biochem. 16, 44-248.
55. Vandeplassche, G., Hermans, C., Thone, F., and Borgers, M. (1989) J. Mol. Cell. Cardiol. 21, 383-
392.
56. Poot, M., and Pierce, R. H. (1999) Cytometry 35, 311-317.
57. Zamzami, N., Marchetti, P., Castedo, M., Decaudin, D., Macho, A., Hirsch, T., Susin, S. A., Petit,
P. X., Mignotte, B., and Kroemer, G. (1995) J. Exp. Med. 182, 367-377.
58. Costantini, P., Chernyak, B. V., Petronilli, V., and Bernardi, P. (1996) J. Biol. Chem. 271, 6746-
6751.
59. Ha, H. C., and Snyder, S. H. (2000) Neurobiol. Dis. 7, 225-239.
60. Kluck, R. M., Martin, S. J., Hoffman, B. M., Zhou, J. S., Green, D. R., and Newmeyer, D. D.
(1997) EMBO J. 16, 4639-4649.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
25
61. Kluck, R. M., Ellerby, L. M., Ellerby, H. M., Naiem, S., Yaffe, M. P., Margoliash, E., Bredesen,
D., Mauk, A. G., Sherman, F., and Newmeyer, D. D. (2000) J. Biol. Chem. 275, 16127-16133.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
26
Footnotes
# This work has been supported by a special grant of the Ligue Nationale contre le Cancer, as well as by
grants from Agence Nationale de Recherches sur le Sida, Fondation pour la Recherche Medicale (FRM)
and European Union (to GK). M.D.M. receives a short fellowship from Caja de Ahorros de la Inmaculada
(CAI). P.C. receives a Postdoctoral fellowship from FRM, L.R. receives a PhD fellowship from the
French Ministry of Science & Technology.
* M.L.P., G.K., and S.A.S. share senior co-authorship.
1 The abbreviations used are: AIF, apoptosis inducing factor; Cyt-c, cytochrome c; ∆Ψm, mitochondrial
transmembrane potential; DPI, diphenyleneiodonium; DTNB, 5.5´-dithiobis-(2-nitrobenzoate-(Nbs2) ;
NBT, 2,2´-Di-p-nitrophenyl-5-5´-diphenyl-3,3´[3-3´-dimetoxy-4-4´difenilen] tetrazolium chloride;
PCMPS, para-chloromercuriphenylsulfonic acid; PT, permeability transition; PTPC, permeability
transition pore complex; SMAC, second mitochondria-derived activator of caspases; SOD, superoxide
dismutase.
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
27
Figure legends
Fig. 1: Absorption spectrum of recombinant AIF∆1-120. The concentration used was 13
µM.
Fig. 2: NADH titration of AIF∆1-120. The top line shows the spectrum of the oxidized
enzyme (47 µM) before (a) and after addition of 0.12 (b), 0.35 (c), 0.59 (d), 0.83 (e),
1.07 (f), 1.31 (g), 1.57 (h), and 1.79 (i) equivalents of NADH/FAD. The inset shows
the absorbance at 450 nm versus added NADH equivalents.
Fig. 3: Anaerobic titration of AIF∆1-120 with an excess of NAD(P)H. Long wavelength
absorbance changes observed in oxidized AIF∆1-120 (22 µM) without (a) or after
addition of NADH (b: molar excess NADH/AIF was 3:1 and c: molar excess was 6:1,
with maxima at 637 nm and 774 nm, respectively) (A). Same as A using NADPH instead
of NADH, with appearance of maxima at 740 nm (b, molar excess NADPH/AIF, 3:1)
and 769 nm (c, molar excess 6:1) (B).
Fig. 4: pH effect on the redox potential of AIF∆1-120. Titration curves obtained
following absorbance variations at 450 nm of oxidized AIF∆1-120 (10 µM) upon
dithionite addition. Solid curves calculated from a dielectronic Nernst equation of -264
mV at pH 6.5 (A), -308 mV at pH 7.5 (B) and -373 mV at pH 9.0 (C).
Fig. 5: NAD(P)H oxidation by AIF∆1-120. NADH (•) and NADPH (o) oxidation
measured following absorbance variation at 340 nm, after addition of different amounts
of NAD(P)H. AIF was added at a concentration of 3 µM (A). AIF∆1-120 induced NBT
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
28
reduction with NADH (•) or NADPH (o) as electron donors. AIF was added at a
concentration of 95 nM (B).
Fig. 6: NADH (•) and NADPH (o) oxidase activities of AIF result in generation of
superoxide anion (A) and hydrogen peroxide (B). AIF concentration was 95 nM in A and
3 µM in B.
Fig. 7: Cytochrome c reduction mediated by superoxide anion, using NADH as electron
donor (A). Specific inhibition of superoxide anion-mediated Cyt c reduction by SOD (B).
Adrenodoxin enhances Cyt c reduction (C). AIF concentrations were 72 nM in A and B,
and 90 nM in C. NADH concentration was 3.4 mM in B and C.
Fig. 8: AIF is the dominant NADH-oxidase released from mitochondria undergoing
permeability transition. AIF release induced by 100 µM Ca2+ and 2 mM sodium arsenite
measured by in situ NBT detection (A). AIF detection by immunoblot of mitochondrial
intermembrane proteins (B). As a control of permeability transition induction,
mitochondria were pretreated with 1 µM of cyclosporine A to inhibit the release of the
intermembrane mitochondrial proteins.
Fig. 9: Apoptotic activity of apoAIF∆1-120 and reconstituted holoAIF∆1-120 in a cell
free system and in microinjection assays. Spectral characteristics of apoAIF and
reconstituted holoAIF are shown in A. The inset shows the recovery of the NADH-
oxidase activity of apoAIF after FAD reconstitution measured by in situ NBT detection
(as in Fig. 8A). Rat-1 cells were microinjected with the indicated protein (7.5 µM of
AIF∆1-120, apoAIF∆1-120 and reconstituted holoAIF∆1-120), and stained with Hoechst
33342 (blue fluorescence) and the ∆Ψm-sensitive dye CMXRos (red fluorescence).
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
29
Representative phenotypes obtained 3 h after injection are shown in (B). Quantification of
nuclear apoptosis and ∆Ψm reduction induced by microinjection (100-150 Rat-1 cells per
session) of AIF∆1-120, apoAIF∆1-120, reconstituted holoAIF∆1-120 or AIF∆1-351
(determined as in B) (C; X +/- SE of three experiments).
Fig. 10: Independence between the redox and apoptotic activities of AIF. Apoptotic
nuclear features induced by AIF∆1-120 or AIF∆1-120+NADH in a cell-free system.
HeLa nuclei were exposed 120 min to 100 µg/ml of AIF∆1-120 and/or 2 mM NADH
followed by staining with Hoechst 33342 (A). In addition, nuclei were stained with
propidium iodide and the percentage of hypoploid nuclei was measured by flow
cytometry (B). Dose-response of the apoptogenic effect of AIF∆1-120 in the presence or
absence of 2 mM NADH or NADPH measured as in B (C). Comparison of AIF redox
and apoptotic activity obtained after pretreatment of AIF∆1-120 (100 µg/ml, 15 min at
37°C) with NADH (2 mM), NADPH (2 mM), SOD (40 U/ml), diphenyleneiodonium
(DPI, 250 µM), or PCMPS (30 µM) ( D; X +/-SE of five experiments).
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Redox activity of AIF
30
Table 1.- Steady-state kinetic parameters of AIF 1-120.
NAD(P)H OXIDASE ACTIVITY
Specific activity (U/mg) kcat (min-1) Km (µM) kcat/Km (µM-1 min-1)
NADH 0.036 2.09 99.4 + 10 0.021
NADPH 0.098 2.84 52.9 + 12 0.054
NBT REDUCTASE ACTIVITY
Specific activity (U/mg) kcat (min-1) Km (mM) kcat/Km (mM-1 min-1)
NADH 38.733 2244.2 2.2 + 0.3 1020
NADPH 0.383 22.1 1.5 + 0.1 14.8
SUPEROXIDE PRODUCTION
Specific activity (U/mg) kcat (min-1) Km (mM) kcat/Km (mM-1 min-1)
NADH 5.559 322.1 2.2 + 0.2 146.4
NADPH 0.79 45.6 1.5 + 0.1 30.4
PEROXIDE PRODUCTION
Specific activity (U/mg) kcat (min-1) Km (µM) kcat/Km (mM-1 min-1)
NADH 0.462 26.7 19 + 3 1.4
NADPH n.d. n.d. n.d.
DIAPHORASE ACTIVITY (DCPIP)
Specific activity (U/mg) kcat (min-1) Km (mM) kcat/Km (mM-1 min-1)
NADH 0.375 21.6 3.4 + 0.4 6.3
NADPH n.d. n.d. n.d.
n.d.: non detected
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Santos A. SusinHaouzi, Greg Brothers, Josef M. Penninger, M. Luisa Peleato, Guido Kroemer and
M. Dolores Miramar, Paola Costantini, Luigi Ravagnan, Ligia M. Saraiva, DelphineNADH-oxidase activity of mitochondrialapoptosis-inducing factor (AIF)
published online February 13, 2001J. Biol. Chem.
10.1074/jbc.M010498200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/early/2001/02/13/jbc.M010498200.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on October 8, 2016
http://ww
w.jbc.org/
Dow
nloaded from
Top Related
Copyright © 2022 FDOKUMEN

