







Bahasa
Halaman
Hukum


www.elsevier.com/locate/chemgeo
Chemical Geology 21
Mg–silica precipitation in fossilization mechanisms of sand tufa
endolithic microbial community, Mono Lake (California)
Virginia Souza-Egipsya,*, Jacek Wierzchosb, Carmen Ascasoc, Kenneth H. Nealsona
aDepartment of Earth Sciences, University of Southern California, 3651 Trousdale Parkway, Los Angeles, CA 90089-0740, USAbServei de Microscopia Electronica, Universitat de Lleida, Rovira Roure 44, Lleida 25198, Spain
cCentro Ciencias Medioambientales, CSIC, Serrano 115 bis, Madrid 28006, Spain
Received 15 April 2004; received in revised form 5 November 2004; accepted 16 December 2004
Abstract
The detection of Mg–silica associated with endolithic biofilms of cyanobacteria and bacterial communities living in lake
shore sand tufa formations in Mono Lake (California) has been observed using scanning electron microscopy with
backscattered electron imaging (SEM–BSE) and energy-dispersive X-ray spectroscopy (EDS). Mg accumulated in cell walls of
living cyanobacteria is interpreted to promote Mg-silicification with structural preservation while non-cyanobacterial
prokaryotes are not fossilized. However, extracellular polymeric substances around the living communities were prominent
as nucleation sites for authigenic mineral precipitation: a Mg–silicate served to stabilize the biofilm remains. Further diagenesis
of these textures resulted in occlusion of the cytoplasm pore spaces by carbonate precipitates. This precipitate damaged the
microfossil structures but the Mg–silica fabric was still present. The paleontological significance of Mg–silica precipitates as a
possible biosignature of the former presence of endolithic biofilms is addressed.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Cyanobacteria; Silica precipitation; Biomineralization; Tufa
1. Introduction
Two different approaches have been identified in
the study of the fossilization mechanisms of micro-
organisms and the study of the Earth’s earliest life
forms (Konhauser et al., 2003). In one side, laboratory
0009-2541/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.chemgeo.2004.12.004
* Corresponding author. Present address: Centro de Astrobiologıa
CSIC-INTA, Ctra. Torrejon a Ajalvir Km 4, Torrejon de Ardoz
28850, Madrid, Spain. Tel.: +34 91 520 64 08; fax: +34 91 520 1074.
E-mail address: [email protected] (V. Souza-Egipsy).
studies using metabolic different microorganisms will
describe the preservation potential of specific micro-
organisms and the mechanisms of fossilization under
controlled conditions. And on the other side, the study
of the fossilization mechanisms of natural microbial
communities will provide the description of specific
fossilization mechanisms depending on the environ-
ment under study. These fossilization mechanisms
may depend also on the post-fossilization alteration
processes responsible for the final structure of the
geological deposits.
7 (2005) 77–87

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–8778
Current knowledge of fossils of microorganisms,
as traces of microbial activity on ancient Earth, tends
to be scarce in comparison to the wealth of
information available for present living microbial
biofilm and mat-building communities. The study of
such modern mat systems has led to the ability to
relate the basic physiological activities to structural
components and fabrics (Gerdes et al., 1993).
However, the physiological changes that occur upon
death and decay are much less well known. This lack
of knowledge of lithification and microfossil forma-
tion on the various components of diverse mat
communities is a major impediment bridging the
gap between the modern ecosystems and fossil
records (Walter et al., 1992). One of the steps needed
to reach such an understanding is the study of the
geobiological processes as they occur. We thus
present in this paper an in situ approach to the study
of the mineralogical changes related with the decay,
fossilization, and diagenic processes in an endolithic
microbial community.
The endolithic mode of life includes several differ-
ent ecological niches: chasmoendoliths and cryptoen-
doliths occupy preexisting fissures and structural
cavities in the rocks, whereas euendoliths penetrate
the rock by etching (Golubic et al., 1981). The
morphological description of the community structure
of microorganisms colonizing the inside of lithic
materials, and solid interfaces has presented enormous
technical challenges; challenges that have often dis-
couraged scientific involvement and delayed our
understanding of these complex systems. Recently,
new approaches based on scanning electron micro-
scopy operated in backscattered electron mode (SEM–
BSE) combined with an auxiliary X-ray energy-
dispersive spectroscopy (EDS) has been implemented
for the study of lithobiontic microbial communities
(Wierzchos and Ascaso, 1994, 2001; Ascaso and
Wierzchos, 1994). This technique permits the chem-
ical characterization of mineral features, and the
observation of the ultrastructural characteristic of the
cells without the need to remove them from their
microenvironment. Indeed, this in situ microscopic
method permits both the description of the living
community and the associated mineralization pro-
cesses, as well as the study of the mineralogical
changes associated with diagenetic events. This allows
one to interpret the microfossil structures from extant
microbial communities in the context of biominerali-
zation processes occurring in living and dying micro-
bial ecosystems, according to taphonomic principles as
suggested by Golubic and Seong-Joo (1999).
In this study we have applied such approaches to
the knowledge of the ultrastructure of the endolithic
communities from the shores of an alkaline salt lake at
California. Mono Lake is famous for the tufa towers
(Council and Benneth, 1993; Scholl and Taft, 1964)
but tufa deposits also occur as pavements of calcite
impregnate sands and silts beneath and adjacent to the
shoreline in areas saturated with brine (Rieger, 1992).
In this article we report the results of studies of sand
tufa microbial endolithic community dynamic pro-
cesses in situ. It is our long-term goal to be able to
recognize signs of past life in geological materials.
Through the description of the living and degradation
sequences of modern cyanobacteria and bacteria
endolithic communities it should be possible to
reconstruct the information lost during the selective
process of preservation, fossilization, and diagenesis
in the geological record.
2. Materials and method
Pieces of sand tufa from Navy Beach (southern
shore lake) were cut with a chisel and collected in
clean and sterile plastic bags. The samples were kept
cold (5 8C) and in the dark until processing. The
samples were processed for scanning electron micro-
scopy following the method published previously
(Ascaso and Wierzchos, 1994; Wierzchos and
Ascaso, 1994) with the following modifications:
they were chemically fixed with 2.5% glutaraldehyde
in filter-sterilized (0.2 Am) lake water. All samples
were then rinsed in filter sterilized lake water, and
post-fixed in 2% osmium tetroxide. During dehy-
dration with an ethanol series, samples were con-
trasted with saturated 70% ethanol uranyl acetate.
The samples were embedded in LR-White embed-
ding medium. Once polymerized the blocks were cut
transversally using a diamond saw, fine polished and
examined under a scanning electron microscope with
backscattered electron (BSE) detector plus an auxil-
iary X-ray energy-dispersive spectroscopy (EDS)
microanalytical system. Transverse sections of the
polished surfaces of the rocks were examined using a

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–87 79
DSM 940 A Zeiss SEM equipped with a BSE
detector and a Link ISIS microanalytical EDS
system. Prior to observation the samples were carbon
coated with a Polaron CA508 evaporation PSC
equipment. EDS operating conditions were as
follows: 08 tilt angle, 358 take-off angle (EDS), 15
kV acceleration potential, 10 or 25 (EDS) mm
working distance, and 1–5 nA specimen current.
For EDS semiquantitative point analysis following
internal standards were used: quartz for O and Si,
magnesium oxide for Mg, alumina for Al, and
wollastonite for Ca, respectively.
3. Results
The living sand tufa endolithic communities at the
shores of the lake were present in the sediment
covered with a crust of salts (Fig. 1a). Some of these
sediments are currently exposed due to the decrease in
the level of the lake (Fig. 1b) and it was possible to
observe that the precipitation of carbonates induces
Fig. 1. (a) General view of the sand tufa formation on the shores of the lake
of the lake level.
the preservation of the upper part of the sediment and
some columnar structures while other areas were
easily weathered. The detail image of the sand tufa
indicates that this crytoendolithic community is
confined to an exact area below the surface and has
a constant thickness of 1–2 mm (lines in Fig. 2a). The
sediment grains below the living community were
loose, while in the upper part they were attached to
each other through the carbonate precipitate (white
square in Fig. 2b). Fig. 2c is an image obtained with
the scanning electron microscope (SEM) operating in
backscattered electron (BSE) emission mode. The
BSE signal is strongly dependent on the mean atomic
number of the target (Joy, 1991) so that the image not
only reveals different contrasts of inorganic particles
but also allows the identification of microorganisms
by staining with heavy metals. The grey areas marked
with the stars are areas were the biofilm is present
between the brilliant sand particles (SP). In some
areas a precipitate was observed between the sand
particles and the biofilm (arrows Fig. 2c). The biofilm
in the area marked with a white square is magnified in
. (b) General view of the exposed sand tufa formations after receding

Fig. 2. (a) Optical image of the endolithic sand tufa community. Area between lines indicates situation of the endolithic green community. Width
of the image is 2 cm. (b) SEM–BSE general view of the sand tufa. White square area magnified in panel c. Bar=1 mm. (c) SEM–BSE image of
the polished cross-section. Stars indicate distribution of the biofilm. White square indicates living biofilm area magnified in panel d. Black
square indicates magnified area in Fig. 6a. Arrows indicate calcium carbonate precipitate around sand particles (SP). Bar=500 Am. (d) Detailed
image of the aspect of the biofilm. Living cells appear brilliant due to the presence of uranyl acetate and osmium tetroxide fixatives (L),
decaying cells exhibit a less brilliant interior (D), fossils remains as cell walls (F). Bar=20 Am. (e) Detailed image of the living cyanobacteria
(cb) and the bacteria cells (b) among them. Bar=10 Am. (f) Silica digital mapping distribution in panel d. (g) Magnesium digital mapping
distribution in panel d.
V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–8780
Fig. 2d. The living biofilm is composed of unicellular
and filamentous cyanobacteria (cb) and bacterial (b)
communities. The living cyanobacterial cells (L)
exhibit a brilliant interior, whereas the decaying ones
(D) show much less contrasted cytoplasm, and it was
collapsed and detached from the cell walls. A detail
image of the biofilm shows the relation between
bacteria cells (b in Fig. 2e) and living cyanobacteria
cells (cb in Fig. 2e). Only the remains of the
cyanobacteria cell walls can be recognized fossilized
in the biofilm (F in Fig. 2d) due to the accumulation
of silica–magnesium precipitate. The EDS and digital
mappings of silica and magnesium were similar (Fig.
2f and g) in the area shown in Fig. 2d and showed a
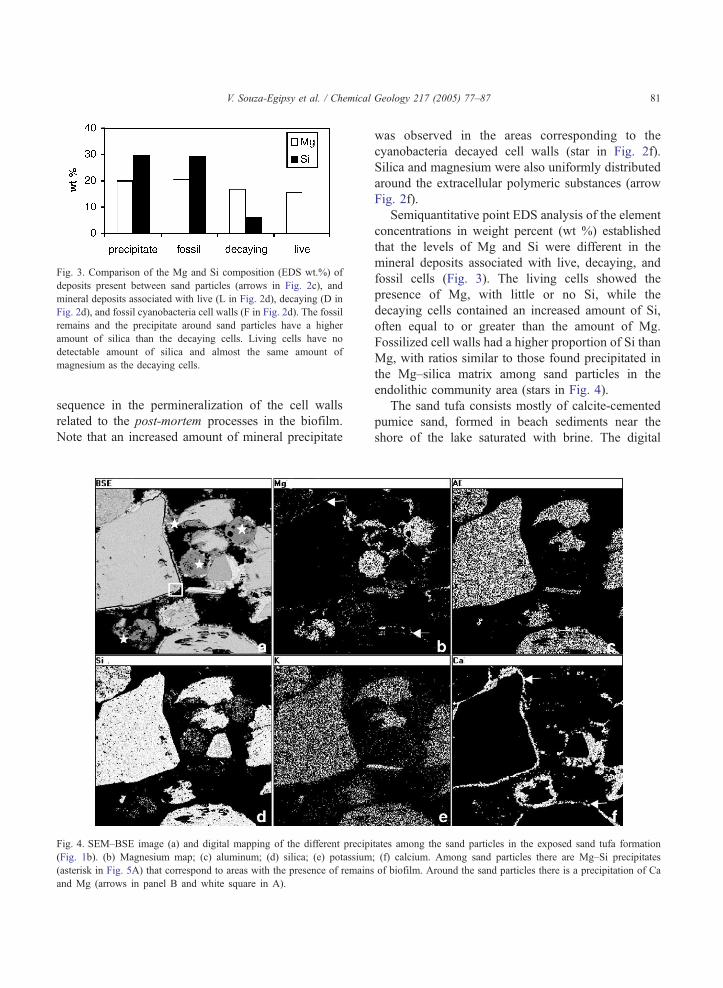
Fig. 3. Comparison of the Mg and Si composition (EDS wt.%) of
deposits present between sand particles (arrows in Fig. 2c), and
mineral deposits associated with live (L in Fig. 2d), decaying (D in
Fig. 2d), and fossil cyanobacteria cell walls (F in Fig. 2d). The fossil
remains and the precipitate around sand particles have a higher
amount of silica than the decaying cells. Living cells have no
detectable amount of silica and almost the same amount of
magnesium as the decaying cells.
V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–87 81
sequence in the permineralization of the cell walls
related to the post-mortem processes in the biofilm.
Note that an increased amount of mineral precipitate
Fig. 4. SEM–BSE image (a) and digital mapping of the different precipi
(Fig. 1b). (b) Magnesium map; (c) aluminum; (d) silica; (e) potassium
(asterisk in Fig. 5A) that correspond to areas with the presence of remains
and Mg (arrows in panel B and white square in A).
was observed in the areas corresponding to the
cyanobacteria decayed cell walls (star in Fig. 2f).
Silica and magnesium were also uniformly distributed
around the extracellular polymeric substances (arrow
Fig. 2f).
Semiquantitative point EDS analysis of the element
concentrations in weight percent (wt %) established
that the levels of Mg and Si were different in the
mineral deposits associated with live, decaying, and
fossil cells (Fig. 3). The living cells showed the
presence of Mg, with little or no Si, while the
decaying cells contained an increased amount of Si,
often equal to or greater than the amount of Mg.
Fossilized cell walls had a higher proportion of Si than
Mg, with ratios similar to those found precipitated in
the Mg–silica matrix among sand particles in the
endolithic community area (stars in Fig. 4).
The sand tufa consists mostly of calcite-cemented
pumice sand, formed in beach sediments near the
shore of the lake saturated with brine. The digital
tates among the sand particles in the exposed sand tufa formation
; (f) calcium. Among sand particles there are Mg–Si precipitates
of biofilm. Around the sand particles there is a precipitation of Ca

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–8782
mapping of the area near the surface also showed
other precipitates among the sand particles (stars Fig.
4a) formed of Mg–silica (Fig. 4b, d). The coatings
around the sand particles were formed from a mix of
magnesium and calcium (arrows Fig. 4b, f). A
detailed study of these coatings (square Fig. 4a) with
Linescan distribution of Ca and Mg (Fig. 5) showed a
banded pattern indicating different episodic precip-
itation reflecting differences in the solution around the
sediments. The low-magnesium calcite precipitate
was present around most of the sand particles and
around Mg–silica areas formerly colonized by living
biofilm.
Fig. 5. SEM–BSE magnified image of the area marked with a white square
the transect indicate in the image. The precipitation around the sand partic
pattern in the image and compositional analysis.
In some zones in the endolithic microbial com-
munity, life, and/or decaying cells were not observed
(Fig. 6a corresponds to magnified black square from
Fig. 2c). The size of the voids in the Mg–silica
precipitate can be related to the space occupied by the
cytoplasm of the cyanobacteria. Since the voids are
observed to be empty, permineralization of the interior
of the cells does not occur, only the cell walls had
been effectively fossilized and were present inside a
matrix of Mg–silica precipitate. The presence of the
matrix surrounding the cell wall remains indicates that
the bacterial extracellular polymers were effective at
binding ions from solution and serving as nucleation
in Fig. 4a with Linescan of Ca (black line) and Mg (grey line) around
les has a variable proportion of Mg and Ca as shown by the banding
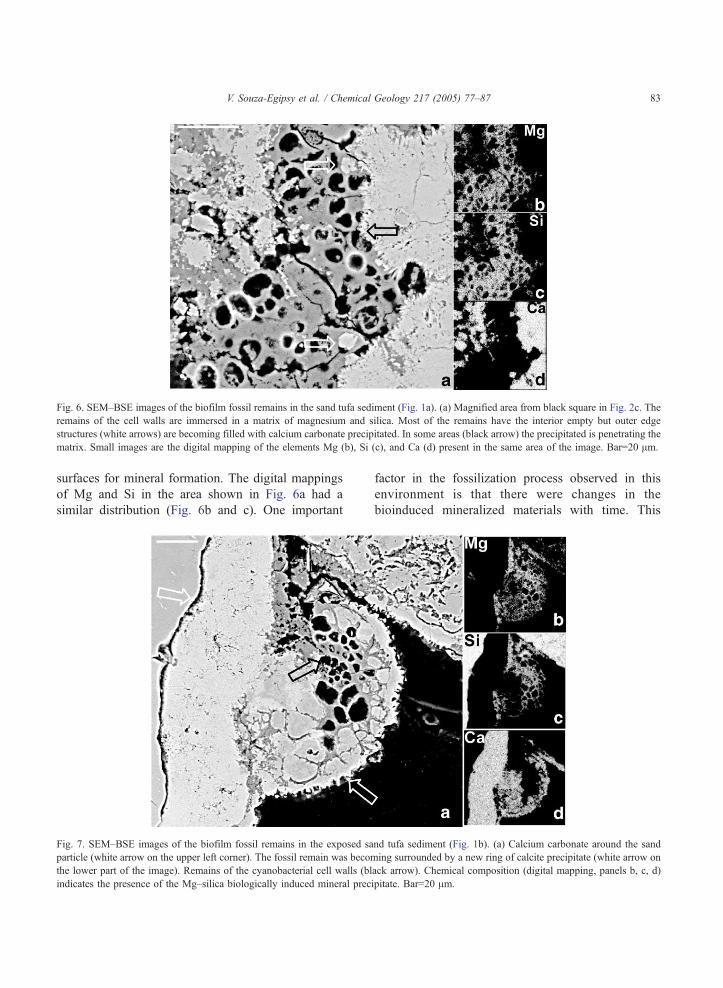
Fig. 6. SEM–BSE images of the biofilm fossil remains in the sand tufa sediment (Fig. 1a). (a) Magnified area from black square in Fig. 2c. The
remains of the cell walls are immersed in a matrix of magnesium and silica. Most of the remains have the interior empty but outer edge
structures (white arrows) are becoming filled with calcium carbonate precipitated. In some areas (black arrow) the precipitated is penetrating the
matrix. Small images are the digital mapping of the elements Mg (b), Si (c), and Ca (d) present in the same area of the image. Bar=20 Am.
V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–87 83
surfaces for mineral formation. The digital mappings
of Mg and Si in the area shown in Fig. 6a had a
similar distribution (Fig. 6b and c). One important
Fig. 7. SEM–BSE images of the biofilm fossil remains in the exposed sa
particle (white arrow on the upper left corner). The fossil remain was becom
the lower part of the image). Remains of the cyanobacterial cell walls (bl
indicates the presence of the Mg–silica biologically induced mineral prec
factor in the fossilization process observed in this
environment is that there were changes in the
bioinduced mineralized materials with time. This
nd tufa sediment (Fig. 1b). (a) Calcium carbonate around the sand
ing surrounded by a new ring of calcite precipitate (white arrow on
ack arrow). Chemical composition (digital mapping, panels b, c, d)
ipitate. Bar=20 Am.

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–8784
aspect is related with the physico-chemical conditions
of the environment, as there is a close contact with the
surrounding lake water and ions present. Silicification
of the cyanobacteria cell walls does not seem to
permanently preserve the microbial remnants; after
the cytoplasmic decay, the area became infiltrated
with calcium carbonate. Fig. 6 shows how part of the
empty cytoplasmatic spaces were starting to be filled
with a calcium carbonate precipitate (white arrows in
Fig. 6a) and was in other areas the growth of the new
carbonate crystals were displacing the Mg–silica
matrix (black arrow in Fig. 6a).
The calcite precipitation is a periodic process that
affects the surface of the sand particles and
permineralized biofilm remains (Fig. 4). On older
formations such as the exposed shore sediments
(Fig. 1b), we found some fossil microbialites. In
Fig. 7a there was a ring of calcium carbonate
around the sand particle (white arrow on the upper
left corner), and over this there was a fossil remain
of the biofilm that was becoming surrounded by a
new ring of calcite precipitate (white arrow on the
lower part of the image). Between these two
coatings, in the Mg–silica precipitate, it was possible
to distinguish the remains of the cyanobacterial cell
walls (black arrow Fig. 7a). However, the deposit of
calcium carbonate around the remains of the fossil
biofilm area was inducing the damage of the fossil
remains. In some areas the cell wall structure was
not present anymore, although the chemical compo-
sition (digital mapping in Fig. 7b, c) indicates the
presence of the Mg–silica biologically induced
mineral precipitate.
4. Discussion
The adsorption of magnesium in dead or decaying
cell structures and organic matter is an important
factor to take into account in the preservation potential
of the microorganism and extracellular polymers at
Mono Lake. Our observations indicate a biological
influence in the presence of Mg–silica precipitates.
There is a correlation between areas of biofilm
presence and Mg adsorption and finally Mg–silica
complexation. With this in situ approach it was
possible to see that the Mg2+ adsorption process was
related to decay and post-mortem processes at the
cyanobacteria cell walls. It has been demonstrated that
bacteria and cyanobacteria have the ability to bind and
to complex metallic cations (Beveridge and Fyfe,
1985; Beveridge, 1989; Schultze-Lam et al., 1993;
Benning et al., 2004). The possible inhibition of the
autocatalytic degradion activity of the cyanobacteria
by magnesium can be explained by data which show
that autolysis tend to be inhibited by other metal
cations (Ferris et al., 1988). Differences in the size and
cell wall structure between cyanobacteria and other
bacteria during the process of fossilization might
explain the different affinity to Mg2+ and thus the
absence of visual remains of smaller bacteria in the
sediment. Recently, Mg–Si permineralization of cell
walls was reported as a possible explanation for the
preservation of green algae remains in subfossil
microbialites of Lake Satonda (Arp et al., 2003).
Schultze-Lam et al. (1996) noted that Gram-positive
bacteria have a higher metal-binding capacity than
Gram-negative bacteria and this aspect has implica-
tions in the fossilization potential as shown in
experimental silicification studies (Westall et al.,
1995; Toporski et al., 2002) and in geothermal hot
springs (Schultze-Lam et al., 1995; Jones and Renaut,
1997; Konhauser et al., 2001). The extent to which the
cells were degraded by their own autocatalytic
enzymes effectively determined whether intact struc-
tures conforming to the original morphology can be
silicified and preserved in the sediments.
Mono Lake is a high pH alkaline lake where at pH
above 9 the solubility of amorphous silica rises sharply
due to the formation of silicate ion (Govett, 1961):
SiðOHÞ4 þ ðOHÞ�YSiOðOHÞ�3 þ H2O
Proposed silicification models at neutral pH ranges
(Leo and Barghoorn, 1976) consider polysilicic acid
as the agent in petrification by means of the attach-
ment of silica molecules to available groups; further
polymerization will lead to the formation of a
precipitate. Recent laboratory experiments demonstra-
ted that neutral undersaturated solutions of Si dis-
played low affinity and did not nucleate onto
cyanobacteria surfaces (Yee et al., 2003). However,
experiments conducted with ferrihydrite-coated cya-
nobacteria significantly increased the rate of Si
removal from the solutions (Yee et al., 2003). In our
field site the pH induces the formation of silica ions

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–87 85
that are available to react with the decaying cyano-
bacteria with Mg2+ onto the surfaces. A similar
process should happen into the extracellular poly-
meric substances around decaying biofilms. Recent
studies on the formation of subfossil microstromatolic
crusts suggested that a primary carbonate mineral may
have been replaced by the amorphous Mg–Si layers
present in the microbialites (Arp et al., 2003).
Although in our samples low-Mg calcite was present
around the sand particles, it was absent from the area
where the biofilm was present. Therefore, in high-pH
alkaline water–sand interface, Mg–silica precipitation
is induced on the surface of the cyanobacteria and
extracellular polymeric substances during the decay-
ing processes of the biofilm.
The particular distribution of Mg–silica precipita-
tion on the sediment indicates the biological influence
in the formation. Chemical Mg–hydroxysilicate pre-
cipitation is likely to occur in carbonate-containing
siliceous sediments because the CaCO3 dissolution
helps to maintain pH values near 8 (Kent and Kastner,
1985), a hydrated amorphous Mg–Si phase was
reported to form within sediments in other alkaline
lakes (Spencer et al., 1985). However in the sand tufa
formations it is not homogeneously distributed. This
precipitate only occurs in the area where the living or
decaying biofilm was present. In the search of the first
fossil record of microorganisms that lived on Earth,
and in the search of life or its fossil record on Mars, it
might be possible to find minerals formed by, or in
association with microorganisms. Previous studies
have proved the present of oxyhydroxides precipitates
associated with living bacteria in Antarctic sandstones
(Wierzchos et al., 2003). When the bacteria died, the
biogenecity of these precipitates was assessed through
the special distribution and morphological features
presented in contrast to the surrounding minerals. This
was an example of how bacteria can template mineral
crystallization, resulting in biosignatures of evidence
of past life in the sediments.
In contrast with the heterogeneity in the distribu-
tion of the Mg–silica precipitates, the distribution of
low magnesium calcite coatings around the sediment
particles presents a homogenous distribution. The
banding pattern, shown with the linescan analysis,
indicates variations in the solutions during time.
Variation in Ca2+ and Mg2+ content in the coatings
around sand particles is probably the result of
pronounced seasonality with stronger evaporation
during summer in front of conditions in the winter
rainy season. Recently, Eggins et al. (2004) have
shown a similar banding in the tests of Orbulina
universa, a foraminifera. They explain the bandings
as a consequence of the day–night variations in pH
due to the activity of the algal symbionts. The same
process can be happening around the area where the
biofilm is present in the sand tufa. The activity of the
biofilm can induce microenvironmental variations in
the pore water around the grains resulting in differ-
ences of Mg/Ca precipitation.
During diagenesis, physico-chemical alteration of
the fossil remains occurred. Similar processes were
also observed by Oehler (1976) during the exper-
imental fossilization of cyanobacteria filaments in
synthetic chert. In those experiments, morphological
alterations of the fossil remains occurred due to quartz
crystallization, as radial expansion of growing quartz
spherulites displaced the cyanobacterial-mineralized
material. In the sand tufa formation, the continuous
precipitation of calcite also provokes the alteration of
the fossil remains. In some areas of the sand tufa, the
Mg–Si precipitates show no evidence for a former
presence of cyanobacteria or bacteria dominated
biofilms. However, the shape and spatial distribution
of these precipitates indicates the presence of micro-
bial communities in the sand tufa. The in situ study of
the diagenetic changes that induce physico-chemical
alterations of microfossils have a clear importance in
the recognition of the fossil remains of biofilms in
geological material.
5. Conclusions
The microenvironment around the endolithic com-
munities during the fossilization process influences
the fidelity of microbial preservation and also has an
increasing importance in paleoecological reconstruc-
tions of the microbial habitat. In Mono Lake sand tufa
formations the precipitation of calcite is a process that
creates coatings around the sand particles but calci-
fication is not happening around the living and
decaying microorganisms while they are alive. Post-
mortem events lead to the accumulation of Mg–silica
in the biofilm area, influencing the preservation and
formation of fossil remains. However, the preserva-

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–8786
tion potential of these structures is finally controlled
by the continuous precipitation of calcite in the
environment. The biosignature of the former presence
of an endolithic biofilm in sand tufa formations will
be the presence of Mg–silica precipitate between the
calcite and sand particles.
The study of in situ microbe–mineral interactions
revealed the transition between accumulations of
elements in the living biofilm to post-mortem
nucleation sites and the influence of the environ-
mental conditions in the preservation of the biosigna-
tures in the geological record.
Acknowledgements
We are grateful to Alicia Thompson (Center for
Electron Microscopy) and Dr. Frank Corsetti (Depart-
ment of Earth Sciences) at University of Southern
California for their assistance and support. Thanks are
due to Dr. Rafael Rodriguez Ochoa for helpful
comments and discussions. Dr. H. Sun kindly guided
in the collection of the samples. V. S-E was supported
by a post-doctoral grant Secretarıa de Estado de
Educacion y Universidades (Spain). The Spanish
MCyT supported the present research under project
REN2003-07366-CO2-02. [LW]
References
Arp, G., Reimer, A., Reitner, J., 2003. Microbialite formation in
seawater of increased alkalinity, Satonda Crater Lake, Indonesia.
J. Sediment. Res. 73 (1), 105–127.
Ascaso, C., Wierzchos, J., 1994. Structural aspects of the lichen–
rock interface using back-scattered electron imaging. Bot. Acta
107, 251–256.
Benning, L.G., Phoenix, V.R., Yee, N., Konhauser, K.O., 2004. The
dynamics of cyanobacterial silicification: an infrared micro-
spectroscopic investigation. Geochim. Cosmochim. Acta 68,
743–757.
Beveridge, T.J., 1989. Role of cellular design in bacterial metal
accumulation and mineralization. Annu. Rev. Microbiol. 43,
147–171.
Beveridge, T.J., Fyfe, W.S., 1985. Metal fixation by bacterial cell
walls. Can. J. Earth Sci. 22, 1893–1898.
Council, T.C., Benneth, P.C., 1993. Geochemistry of Ikaite
formation at Mono Lake, California: implications for the origin
of tufa mounds. Geology 21, 971–974.
Eggins, S.M., Sadekov, A., De Deckker, P., 2004. Modulation and
daily banding of Mg/Ca in Orbulina universa tests by simbiont
photosynthesis and respiration: a complication for seawater
thermometry. Earth Planet. Sci. Lett. 225, 411–419.
Ferris, F.G., Fyfe, W.S., Beveridge, T.J., 1988. Metallic ion binding
by Bacillus subtilis: implications for the fossilization of micro-
organism. Geology 16, 149–152.
Gerdes, G., Claes, M., Dunajtschik-Piewak, K., Krumbein, W.E.,
Reineck, H.E., 1993. Contribution of microbial mats to
sedimentary surface structures. Facies 29, 61–74.
Golubic, S., Seong-Joo, L., 1999. Early cyanobacteria fossil record:
preservation, palaeoenvironments and identification. Eur. J.
Phycol. 34, 339–348.
Golubic, S., Friedmann, I., Schneider, J., 1981. The lithobiontic
ecological niche, with special reference to microorganisms.
J. Sediment. Petrol. 51, 475–478.
Govett, G.J.S., 1961. Critical factors in the colorimetric determi-
nation of silica. Anal. Commun. 25, 69–80.
Jones, B., Renaut, R.W., 1997. Formation of silica oncoids around
geysers and hot springs at el Tatio northern Chile. Sedimentol-
ogy 44, 287–304.
Joy, D.C., 1991. An introduction to Monte Carlo simulation.
Scanning Microsc. 5, 329–337.
Kent, D.B., Kastner, M., 1985. Mg2+ removal in the system Mg2+
-amorphous SiO2–H2O by adsorption and Mg–hydroxylicate
precipitation. Geochim. Cosmochim. Acta 49, 1123–1136.
Konhauser, K.O., Phoenix, V.R., Bottrell, S.H., Adams, D.G., Head,
I.M., 2001. Microbial–silica interactions in Icelandic hot spring
sinter: possible analogues for some Precambrian siliceous
stromatolites. Sedimentology 48, 415–433.
Konhauser, K.O., Jones, B., Reysenbach, A.-L., Renault, R.W.,
2003. Hot spring sinters: key to understanding Earth’s earliest
life forms. Can. J. Earth Sci. 40, 1713–1724.
Leo, R.F., Barghoorn, E.S., 1976. Silicification of wood. Bot. Mus.
Leaf., Harv. Univ. 25, 1–47.
Oehler, J.H., 1976. Experimental studies in Precambrian paleonto-
logy: structural and chemical changes in blue green algae during
simulated fossilization in synthetic chert. Geol. Soc. Amer. Bull.
87, 117–129.
Rieger, T., 1992. Calcareous tufa formations, Searles Lake and
Mono Lake. Calif. Geol. 45, 99–109 (July/August).
Scholl, D.W., Taft, W.H., 1964. Algae, contributors to the formation
of calcareous tufa, Mono Lake, California. J. Sediment. Petrol.
34, 309–319.
Schultze-Lam, S., Thompson, J.L., Beveridge, T.J., 1993. Metal ion
immobilization by bacterial surfaces in fresh water environ-
ments. Water Pollut. Res. J. Can. 28, 51–81.
Schultze-Lam, S., Ferris, F.G., Konhauser, K.O., Wiese, R.G., 1995.
In situ silicification of an Icelandic hot spring microbial mat:
implications for microfossil formation. Can. J. Earth Sci. 32,
2021–2026.
Schultze-Lam, S., Fortin, D., Davis, B.S., Beveridge, T.J., 1996.
Mineralization of bacterial surfaces. Chem. Geol. 132, 171–181.
Spencer, R.J., Eugster, H.P., Jones, B.F., 1985. Geochemistry of
Great Salt Lake, Utah: II. Pleistocene–Holocene evolution.
Geochim. Cosmochim. Acta 49, 739–747.
Toporski, J.K.W., Steele, A., Westall, F., Thomas-Keprta, K.L.,
McKay, D.S., 2002. The simulated silicification of bacteria—
new clues to the modes and timing of bacteria preservation and

V. Souza-Egipsy et al. / Chemical Geology 217 (2005) 77–87 87
implications for the search for extraterrestrial microfossils.
Astrobiology 2, 1–26.
Walter, M.R., Bauld, J., Des Marais, D.J., Schopf, W.J., 1992. A
general comparison of microbial mats and microbial stroma-
tolites: bridging the gap between the modern and the fossil.
In: Schopf, W.J., Klein, C. (Eds.), The Proterozoic Biosphere:
A Multidisciplinary Approach. Cambridge University Press,
New York, pp. 335–338.
Westall, F., Boni, L., Guerzoni, M.E., 1995. The experimental
silicification of microorganisms. Palaeontology 38, 495–528.
Wierzchos, J., Ascaso, C., 1994. Application of back-scattered
electron imaging to the study of the lichen–rock interface.
J. Microsc. 175, 54–59.
Wierzchos, J., Ascaso, C., 2001. Life, decay and fossilisation of
endolithic microorganisms from the Ross Desert Antarctica:
suggestion for in situ further research. Polar Biol. 24,
863–868.
Wierzchos, J., Ascaso, C., Sancho, L.G., Green, A., 2003. Iron-rich
diagenetic minerals are biomarkers of microbial activity in
Antarctic rocks. Geomicrobiol. J. 20 (1), 15–24.
Yee, N., Phoenix, V.R., Konhauser, K.O., Benning, L.G., Ferris,
F.G., 2003. The effect of cyanobacteria on silica precipitation at
neutral pH: implications for bacterial silicification in geothermal
hot springs. Chem. Geol. 199, 83–90.
Top Related
Copyright © 2022 FDOKUMEN

