







Bahasa
Halaman
Hukum


Chemosphere 61 (2005) 323–331
www.elsevier.com/locate/chemosphere
Levels of PCDDs and PCDFs in Korean river sedimentsand their detection by biomarkers
Keon Sang Ryoo a,*, Seong-Oon Ko a, Yong Pyo Hong a, Jong-Ha Choi a,Sunghwan Cho b, Yonggyun Kim b, Yeon Jae Bae c
a Department of Applied Chemistry, Andong National University, Andong 760-749, Koreab School of Bioresource Sciences, Andong National University, Andong 760-749, Korea
c Department of Biology, Seoul Women�s University, Seoul 139-774, Korea
Received 11 October 2004; received in revised form 3 February 2005; accepted 24 February 2005
Available online 21 April 2005
Abstract
Polychlorinated dibenzo-p-dioxins (PCDDs) and dibenzofurans (PCDFs) are a group of toxic halogenated aryl
hydrocarbons inducing various physiological disorders against biological organisms. Here, we investigated their levels
in sediment samples taken from 12 different rivers in Korea. The levels of PCDD/PCDFs in sediment samples were
expressed as concentrations and international TEQ values. Among 17 PCDD/PCDFs selected as target compounds
in this study, the 1,2,3,4,7,8-HxCDD and OCDD were found in all river sediments with significant variation in various
congener profiles of PCDD/PCDFs in sediments. PCDD/PCDFs could be monitored by sensitive biomarkers using
insect immune system. Out of 12 river sediment samples, the biomarkers reported four spots (up, middle, and down
Singil sites and Ansan) as putative contamination areas. When comparing both chemical and biological monitoring
results, two methods agreed three spots of Singil as contamination areas (above 10 ppt levels) as well as six river sed-
iment samples as relatively less-contaminated areas, but differed in the results in Ansan and Miho, probably due to re-
lative non-specificity of biomarkers. Despite some disparity between bio- and chemical monitoring results, the
biomarkers can be recommended as a device warning the contamination of dioxins in the environment because of a
fast and inexpensive detection method.
� 2005 Elsevier Ltd. All rights reserved.
Keywords: PCDDs; PCDFs; Biomarker; Spodoptera exigua; Sediment; River; Korea
1. Introduction
The presence of polychlorinated dibenzo-p-dioxins
(PCDDs) and polychlorinated dibenzofurans (PCDFs)
0045-6535/$ - see front matter � 2005 Elsevier Ltd. All rights reserv
doi:10.1016/j.chemosphere.2005.02.093
* Corresponding author. Tel.: +82 54 820 5453; fax: +82 54
823 1628.
E-mail address: [email protected] (K.S. Ryoo).
in the environment is strongly related with a number of
sources including waste combustion and metal industry,
as well as their formation in the manufacture of various
chlorinated chemicals (Safe, 1990; Alcock and Jones,
1996; Fiedler, 1999). Due to low water solubility and vol-
atility of PCDDs/PCDFs, they undergo so long-range
atmospheric transport and can be deposited into aquatic
systems, especially the sediments as a sink, where they act
as a long-term source of release of these pollutants into
the aquatic food chain (Gobas and Mackay, 1987;
ed.

324 K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331
Connolly and Pedersen, 1988; Thomann, 1989). More-
over, their chemical stability affords significant biomag-
nification through a food chain (Kang et al., 2002).
These chemicals are a group of highly toxic, extre-
mely anthropogenic compounds that give a potential
health hazard to humans and other organisms. In verte-
brates, PCDDs/PCDFs cause a broad spectrum of phys-
iological disorders including weight loss, reproductive
failure, thymic atrophy, hepatotoxicity, several types of
dermal lesion, and immunotoxicity (Safe, 1990; Mocar-
elli et al., 1996; Luebke et al., 2001). The molecular
mechanism of the toxicities has been illustrated with a
TCDD model binding to aryl hydrocarbon (Ah) recep-
tor via induction of 7-ethoxyresorufin-O-deethylase
(EROD). However, though invertebrates have their
own Ah receptors, the TCDD toxicity is limited proba-
bly due to lack of specific binding of the receptors to the
toxic ligand (Butler et al., 2001). The resistance of the
aquatic invertebrates against PCDDs/PCDFs allows
them to accumulate relatively high concentrations,
which may be transferred to sensitive vertebrates via
food webs (West et al., 1997).
To assess the environmental quality, many countries
have monitored the level of PCDDs/PCDFs in various
environmental samples including soil and sediment. In
Korea, the reported levels of PCDDs/PCDFs have been
so limited to certain environmental samples such as ambi-
ent air, food, and incinerator ash (Kim et al., 2001a,b; Im
et al., 2004). Few studies concerning the PCDDs/PCDFs
levels in river sediments have been reported in Korea. In
this paper, we report and discuss the data on the
PCDD/PCDF levels found in sediment samples collected
at 12 different river sites. It also analyzed the toxic effects
of the dioxin residues on insect immune system,whichwas
used for developing sensitive biomarkers.
2. Materials and methods
2.1. Reagents
Two standard solutions containing 17 unlabeled
PCDD/PCDF compounds and 18 isotopically labeled13C12- and 37Cl4-PCDD/PCDF compounds in nonane
were supplied by Cambridge Isotope Laboratories, Inc.
(Andover, MA, USA). All solvents (hexane, dichloro-
methane, acetone, iso-octane, toluene) used in this study
were HPLC grade (J.T. Baker, Deventer, Netherlands).
Anhydrous sodium sulfate (Merck, Darmstadt, Ger-
many) was used after heating overnight at 120 �C. Silicagel and alumina (Aldrich, WI, USA), before use, were
Soxhlet extracted with dichloromethane and then acti-
vated in a foil covered glass container at 150 �C for
24 h. Concentrated sulfuric acid 95–97% (Merck) were
analytical grade reagent.
2.2. Sampling
Field collection was carried out at 12 Korean rivers
(Fig. 1) in October 2003, representing heavy industrial
area (R1–R3), moderately industrial area (R4–R7), near
capital city (R8–R11), and a well reserved reference area
(R12). R1–R3 are the sampling spots from upstream to
downstream near multi-chemical factories and were ex-
pected to accumulate toxic compounds gradually from
R1 to R3. R4–R11 are the places mixed with some fac-
tories and civilian habitats, in which R4–R7 comprised
relatively more factories, but R8–R11 did more civilian
habitats near Seoul with an increasing polluted gradi-
ent from R8 to R11. R12 is located far from chemical
factories and civilian habitats so that it seems to be
hardly contaminated. The upper 0–20 cm layer of sed-
iment was sampled, and then immediately homogenized
and sieved (sieve fraction: �2 mm). In laboratory, all
samples were air-dried at 40 �C for 24 h, ground in
an agitate ball mill. The samples were kept under the
refrigeration at 4 �C until their analysis. The prepared
samples were used for both chemical residual analysis
and biomarker analysis. The PCDD/PCDFs analysis
in samples was performed using the isotope dilution
technique, designated by EPA Method 1613 (high res-
olution GC/MS method for the determination of tet-
ra-octa chlorinated dioxins and furans). About 10 g
sample was weighed and spiked with isotopically la-
beled 37Cl4-2,3,7,8-TCDD as an internal standard be-
fore extraction. Along with each series of samples, a
complete procedure blank was also performed to check
the purity of solvents, reagents, material, etc. The
blank was spiked with an internal standard mentioned
above and analyzed exactly in the same way as the
samples.
2.3. Extraction and clean-up
Extraction was carried out by soxhlet for 24 h with
toluene. After extraction, a total of 15 13C12-labelled
PCDD/PCDF congeners, shown in Table 1, as internal
standards were added to the extract. The extract was
placed in a multi-layer silica gel column including
anhydrous Na2SO4, neutral silica gel, acid modified
silica gel, and NaOH impregnated silica gel, and then
eluted with 100 ml of n-hexane. Afterwards, the extract
was purified in a basic alumina column, by eluting with
100 ml of hexane, followed by 30 ml hexane/dichloro-
methane (1:1 v/v). A hexane/dichloromethane fraction
was fractioned with an activated carbon column con-
taining Celite and then concentrated in a Kudena-
Danish apparatus under a gentle stream of nitrogen
to 1 ml. To test the recovery, the syringe standards
such as 13C12-1,2,3,4-TCDD and 13C12-1,2,3,7,8,9-
HxCDD were added to the final concentrate before
GC/MS analysis.
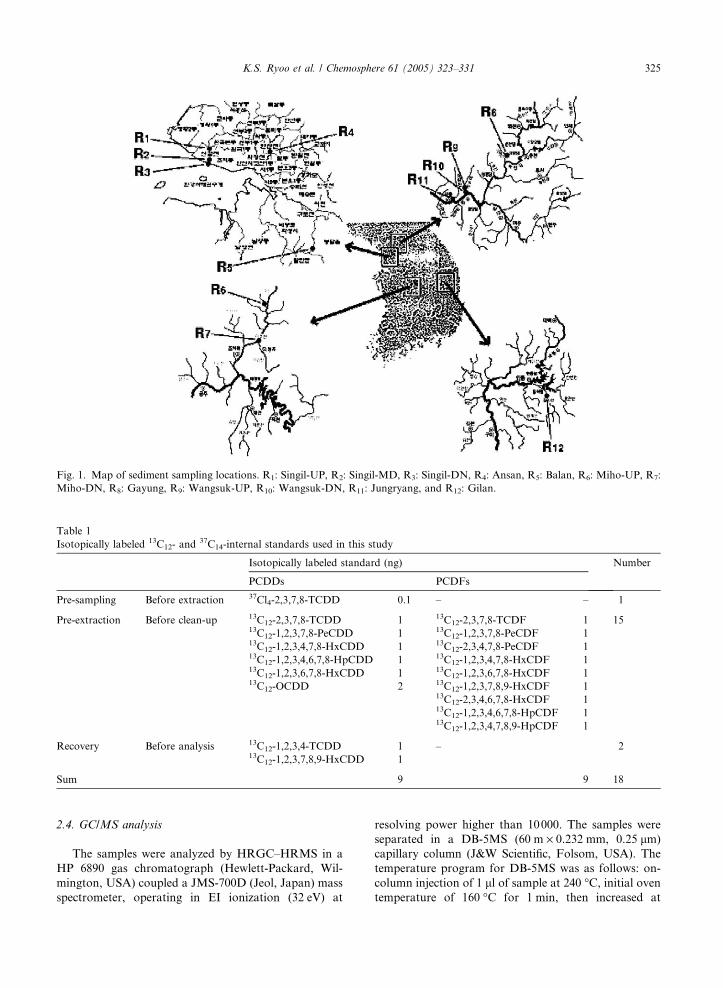
Fig. 1. Map of sediment sampling locations. R1: Singil-UP, R2: Singil-MD, R3: Singil-DN, R4: Ansan, R5: Balan, R6: Miho-UP, R7:
Miho-DN, R8: Gayung, R9: Wangsuk-UP, R10: Wangsuk-DN, R11: Jungryang, and R12: Gilan.
Table 1
Isotopically labeled 13C12- and37C14-internal standards used in this study
Isotopically labeled standard (ng) Number
PCDDs PCDFs
Pre-sampling Before extraction 37Cl4-2,3,7,8-TCDD 0.1 – – 1
Pre-extraction Before clean-up 13C12-2,3,7,8-TCDD 1 13C12-2,3,7,8-TCDF 1 1513C12-1,2,3,7,8-PeCDD 1 13C12-1,2,3,7,8-PeCDF 113C12-1,2,3,4,7,8-HxCDD 1 13C12-2,3,4,7,8-PeCDF 113C12-1,2,3,4,6,7,8-HpCDD 1 13C12-1,2,3,4,7,8-HxCDF 113C12-1,2,3,6,7,8-HxCDD 1 13C12-1,2,3,6,7,8-HxCDF 113C12-OCDD 2 13C12-1,2,3,7,8,9-HxCDF 1
13C12-2,3,4,6,7,8-HxCDF 113C12-1,2,3,4,6,7,8-HpCDF 113C12-1,2,3,4,7,8,9-HpCDF 1
Recovery Before analysis 13C12-1,2,3,4-TCDD 1 – 213C12-1,2,3,7,8,9-HxCDD 1
Sum 9 9 18
K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331 325
2.4. GC/MS analysis
The samples were analyzed by HRGC–HRMS in a
HP 6890 gas chromatograph (Hewlett-Packard, Wil-
mington, USA) coupled a JMS-700D (Jeol, Japan) mass
spectrometer, operating in EI ionization (32 eV) at
resolving power higher than 10000. The samples were
separated in a DB-5MS (60 m · 0.232 mm, 0.25 lm)
capillary column (J&W Scientific, Folsom, USA). The
temperature program for DB-5MS was as follows: on-
column injection of 1 ll of sample at 240 �C, initial oventemperature of 160 �C for 1 min, then increased at

326 K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331
20 �C/min to 200 �C for 2 min, then increased at 5 �C/min to 220 �C for 15 min, then increased at 5.0 �C to
235 �C for 5 min, and finally at 3 �C to 310 �C for
3 min hold-up. Helium was used as the carrier gas.
The results were expressed as concentration in pg/g
and in pg TEQ/g in order to evaluate the toxicity of sam-
ple due to dioxins. To calculate TEQ, the international
toxicity equivalent factors (ITEF) were used (Van Zorge
et al., 1989).
2.5. Biomarkers detecting dioxins
Insect immune responses were modified as different
bioassays, which were used to monitor dioxin residues.
The fifth instar larvae of the beet armyworm, Spodoptera
exigua, was used as a test insect and reared on an artifi-
cial diet of Gho et al. (1990).
For hemocyte nodulation assay by the method of
Park and Kim (2000), 105 cells of the heat-killed
(10 min at 95 �C) bacteria, Escherichia coli, along with
10 ll test solution were injected into the larval hemocoel
by 10 ll microsyringe (Hamilton, Nevada) and then
incubated for 12 h at 25 �C. After the treated larvae were
dissected by opening hemoceol, melanized and dark
nodules were counted under a stereomicroscope at 50·magnification. Even though their sizes were varied
(100–800 lm at diameter), the nodules were distinct
enough to find out their locations.
For an assay of activation of prophenoloxidase
(pPO), PO activity was measured by the method of Park
and Kim (2003). Hemolymph (10 ll) of the fifth instar
larvae was pre-incubated with 2 ll of a test solution at
25 �C for 10 min, and then added by 988 ll of 50 mM
phosphate saline containing 1 lg of laminarin and
0.01 mM L-3,4-dihydroxyphenylalanine (Sigma, St.
Louis, MO). The increased absorbance was measured
at 495 nm with 5 min interval. PO activity was expressed
as absorbance change per 5 min.
For phospholipase A2 (PLA2) assay, an indirect assay
was devised by Park and Kim (2003). Briefly, two identi-
cal hemolymph samples (�A� and �B�) were prepared. �A�sample included 10 ll of hemolymph, 1 ll of arachidonicacid (5,8,11,14-eicosatetraenoic acid, AA, Sigma) stock,
and 1 ll of a test PLA2 inhibitor. �B� sample included
10 ll of hemolymph, 1 ll of 50% ethanol, and 1 ll ofthe test PLA2 inhibitor. To these 12 ll reaction mixtures,
after 10 min, 988 ll of PO substrate solution was added.
PO activity was then measured by the method just des-
cribed. Comparing these three PO activities, PLA2
inhibitor was recognized in the fraction that the reduced
PO activity in the �B� sample was significantly elevated by
the addition of AA as in the �A� sample.
Apoptosis assay of insect immunocompetent cells
(hemocytes) followed the method of Cho and Kim
(2004). Briefly, the larval hemolymph was collected
and centrifuged at 200g for 10 min at 4 �C. After resus-
pending the precipitated hemocytes with Grace�s insect
cell culture medium (Sigma), 10 ll of hemocyte suspen-
sion (containing over 100 cells) was loaded on the glass
coverslip and incubated in a wet chamber at 25 �C for
30 min. The supernatant was replaced with 10 ll of testsolution and further incubated for 2 h. Apoptotic cells
were recognized by membrane blebbings or apoptotic
vesicles and counted under a phase contrast microscope
at 200· magnification.
3. Results
3.1. Levels of PCDD/PCDFs in the river sediment
The concentrations of PCDD/PCDFs in each sedi-
ment sample collected from 12 river sites, designated by
R1–R12, are summarized in Table 2, in which toxic equiv-
alent factors (TEQs) levels are also presented after con-
version to the international 2,3,7,8-TetraCDD toxicity
equivalents (ITEQ). Total concentration and TEQ level
of PCDD/PCDFs at each sampling site ranged from
0.88 to 369.21 pg/g and from 0.07 to 23.93 pg TEQ/g
dry weight sediment, respectively. There was a high cor-
relation (r = 0.9995; P < 0.0001) between total concen-
tration and TEQ values of PCDD/PCDFs. The
1,2,3,4,7,8-HxCDD and OCDD were found in all sedi-
ment samples, though OCDF was the most heavily accu-
mulated when the detection levels of all sediment samples
were combined. The OCDFwas the secondmost popular
congener, with the exception of the R4 and R12. Except
extreme level of R3, relative contamination levels can
be divided by a threshold level of 10 pg/g of total
PCDD/PCDFs concentration into the contaminated
(R1, R2, R3, R6, and R7) and the relatively less-contami-
nated (R4, R5, R8, R9, R10, R11, and R12) sediments.
3.2. Biomonitoring PCDD/PCDFs
The effect of PCDD/PCDFs on insect immune system
was analyzed using TCDD as a representative com-
pound and S. exigua as a model insect. When TCDD
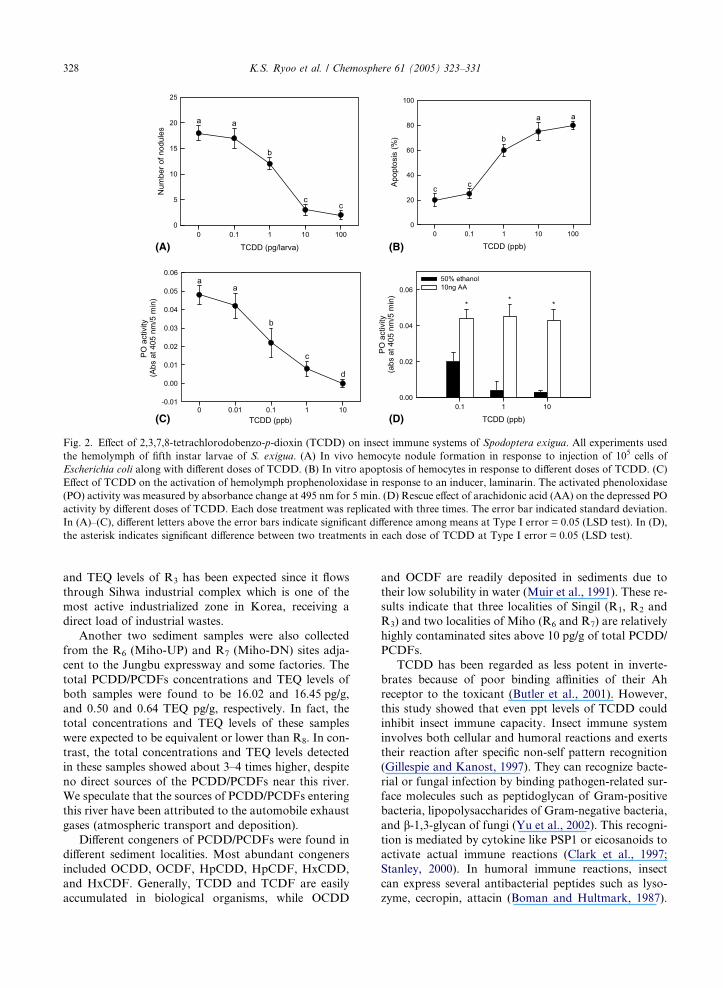
was injected into the larvae, it inhibited the hemocyte
(insect immunocyte) capacity of nodule formation
against the bacteria in a dose-dependent manner (Fig.
2A). Only 1 pg was enough to significantly inhibit the
nodule formation. Nodule formation of hemocytes is
mediated by the activation of prophenoloxidase (Dula-
ray and Lackie, 1985). When the activated phenoloxi-
dase activity was measured after incubation with
different doses of TCDD, 0.1 ppb of TCDD exhibited
significant inhibition in in vitro hemolymph assay (Fig.
2B). Considering a positive or functional correlation
between PLA2 activity and phenoloxidase activity in
S. exigua (Park and Kim, 2003), PLA2 can be targeted
by TCDD. The depressed phenoloxidase activity in the
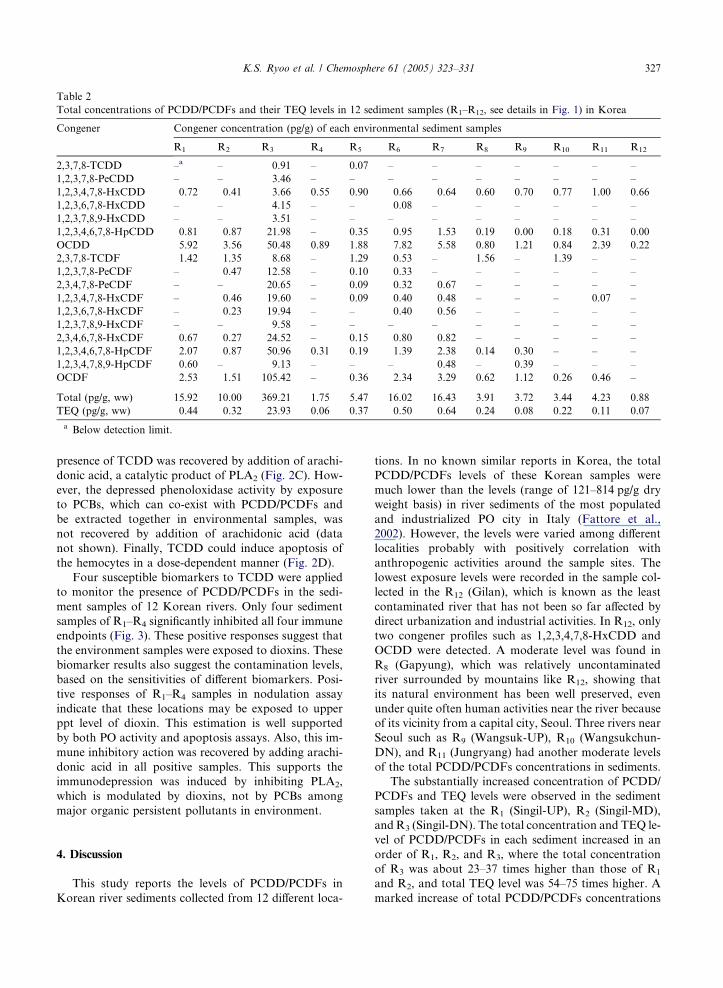
Table 2
Total concentrations of PCDD/PCDFs and their TEQ levels in 12 sediment samples (R1–R12, see details in Fig. 1) in Korea
Congener Congener concentration (pg/g) of each environmental sediment samples
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10 R11 R12
2,3,7,8-TCDD –a – 0.91 – 0.07 – – – – – – –
1,2,3,7,8-PeCDD – – 3.46 – – – – – – – – –
1,2,3,4,7,8-HxCDD 0.72 0.41 3.66 0.55 0.90 0.66 0.64 0.60 0.70 0.77 1.00 0.66
1,2,3,6,7,8-HxCDD – – 4.15 – – 0.08 – – – – – –
1,2,3,7,8,9-HxCDD – – 3.51 – – – – – – – – –
1,2,3,4,6,7,8-HpCDD 0.81 0.87 21.98 – 0.35 0.95 1.53 0.19 0.00 0.18 0.31 0.00
OCDD 5.92 3.56 50.48 0.89 1.88 7.82 5.58 0.80 1.21 0.84 2.39 0.22
2,3,7,8-TCDF 1.42 1.35 8.68 – 1.29 0.53 – 1.56 – 1.39 – –
1,2,3,7,8-PeCDF – 0.47 12.58 – 0.10 0.33 – – – – – –
2,3,4,7,8-PeCDF – – 20.65 – 0.09 0.32 0.67 – – – – –
1,2,3,4,7,8-HxCDF – 0.46 19.60 – 0.09 0.40 0.48 – – – 0.07 –
1,2,3,6,7,8-HxCDF – 0.23 19.94 – – 0.40 0.56 – – – – –
1,2,3,7,8,9-HxCDF – – 9.58 – – – – – – – – –
2,3,4,6,7,8-HxCDF 0.67 0.27 24.52 – 0.15 0.80 0.82 – – – – –
1,2,3,4,6,7,8-HpCDF 2.07 0.87 50.96 0.31 0.19 1.39 2.38 0.14 0.30 – – –
1,2,3,4,7,8,9-HpCDF 0.60 – 9.13 – – – 0.48 – 0.39 – – –
OCDF 2.53 1.51 105.42 – 0.36 2.34 3.29 0.62 1.12 0.26 0.46 –
Total (pg/g, ww) 15.92 10.00 369.21 1.75 5.47 16.02 16.43 3.91 3.72 3.44 4.23 0.88
TEQ (pg/g, ww) 0.44 0.32 23.93 0.06 0.37 0.50 0.64 0.24 0.08 0.22 0.11 0.07
a Below detection limit.
K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331 327
presence of TCDD was recovered by addition of arachi-
donic acid, a catalytic product of PLA2 (Fig. 2C). How-
ever, the depressed phenoloxidase activity by exposure
to PCBs, which can co-exist with PCDD/PCDFs and
be extracted together in environmental samples, was
not recovered by addition of arachidonic acid (data
not shown). Finally, TCDD could induce apoptosis of
the hemocytes in a dose-dependent manner (Fig. 2D).
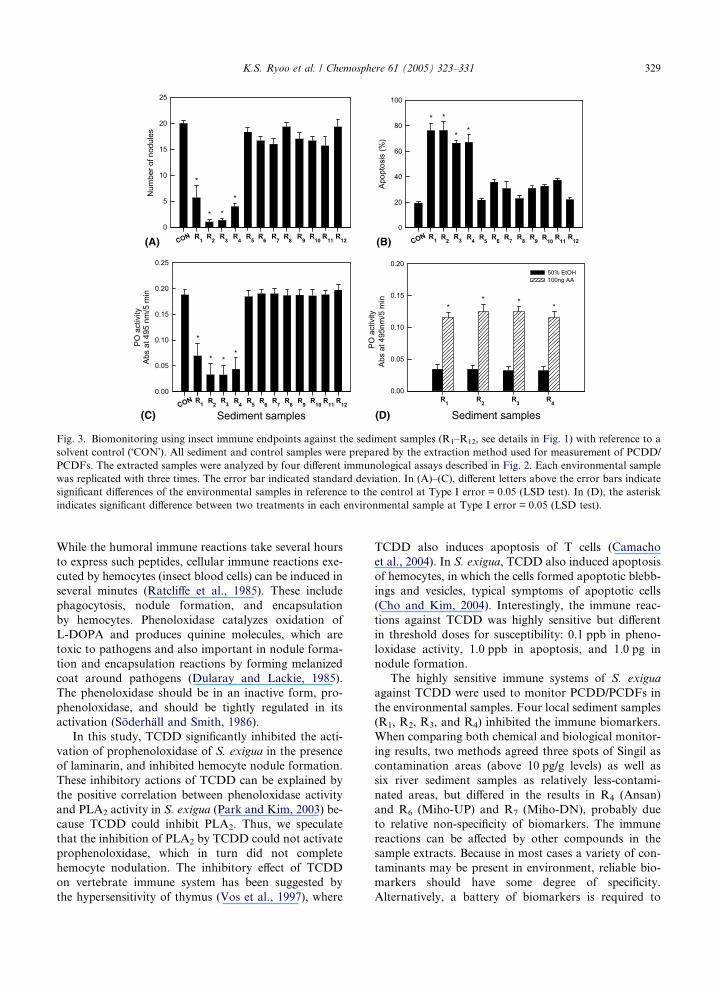
Four susceptible biomarkers to TCDD were applied
to monitor the presence of PCDD/PCDFs in the sedi-
ment samples of 12 Korean rivers. Only four sediment
samples of R1–R4 significantly inhibited all four immune
endpoints (Fig. 3). These positive responses suggest that
the environment samples were exposed to dioxins. These
biomarker results also suggest the contamination levels,
based on the sensitivities of different biomarkers. Posi-
tive responses of R1–R4 samples in nodulation assay
indicate that these locations may be exposed to upper
ppt level of dioxin. This estimation is well supported
by both PO activity and apoptosis assays. Also, this im-
mune inhibitory action was recovered by adding arachi-
donic acid in all positive samples. This supports the
immunodepression was induced by inhibiting PLA2,
which is modulated by dioxins, not by PCBs among
major organic persistent pollutants in environment.
4. Discussion
This study reports the levels of PCDD/PCDFs in
Korean river sediments collected from 12 different loca-
tions. In no known similar reports in Korea, the total
PCDD/PCDFs levels of these Korean samples were
much lower than the levels (range of 121–814 pg/g dry
weight basis) in river sediments of the most populated
and industrialized PO city in Italy (Fattore et al.,
2002). However, the levels were varied among different
localities probably with positively correlation with
anthropogenic activities around the sample sites. The
lowest exposure levels were recorded in the sample col-
lected in the R12 (Gilan), which is known as the least
contaminated river that has not been so far affected by
direct urbanization and industrial activities. In R12, only
two congener profiles such as 1,2,3,4,7,8-HxCDD and
OCDD were detected. A moderate level was found in
R8 (Gapyung), which was relatively uncontaminated
river surrounded by mountains like R12, showing that
its natural environment has been well preserved, even
under quite often human activities near the river because
of its vicinity from a capital city, Seoul. Three rivers near
Seoul such as R9 (Wangsuk-UP), R10 (Wangsukchun-
DN), and R11 (Jungryang) had another moderate levels
of the total PCDD/PCDFs concentrations in sediments.
The substantially increased concentration of PCDD/
PCDFs and TEQ levels were observed in the sediment
samples taken at the R1 (Singil-UP), R2 (Singil-MD),
and R3 (Singil-DN). The total concentration and TEQ le-
vel of PCDD/PCDFs in each sediment increased in an
order of R1, R2, and R3, where the total concentration
of R3 was about 23–37 times higher than those of R1
and R2, and total TEQ level was 54–75 times higher. A
marked increase of total PCDD/PCDFs concentrations
TCDD (ppb)0 0.01 0.1 1 10
PO a
ctiv
ity
(Abs
at 4
05 n
m/5
min
)
-0.01
0.00
0.01
0.02
0.03
0.04
0.05
0.06a
a
b
c
d
TCDD (pg/larva)0 0.1 1 10 100
Num
ber o
f nod
ules
0
5
10
15
20
25
aa
b
cc
TCDD (ppb)0.1 1 10
PO a
ctiv
ity(a
bs a
t 405
nm
/5 m
in)
0.00
0.02
0.04
0.0650% ethanol10ng AA
* * *
(A)
(D)
(B)
(C)
TCDD (ppb)0 0.1 1 10 100
Apop
tosi
s (%
)
0
20
40
60
80
100
aa
b
cc
Fig. 2. Effect of 2,3,7,8-tetrachlorodobenzo-p-dioxin (TCDD) on insect immune systems of Spodoptera exigua. All experiments used
the hemolymph of fifth instar larvae of S. exigua. (A) In vivo hemocyte nodule formation in response to injection of 105 cells of
Escherichia coli along with different doses of TCDD. (B) In vitro apoptosis of hemocytes in response to different doses of TCDD. (C)
Effect of TCDD on the activation of hemolymph prophenoloxidase in response to an inducer, laminarin. The activated phenoloxidase
(PO) activity was measured by absorbance change at 495 nm for 5 min. (D) Rescue effect of arachidonic acid (AA) on the depressed PO
activity by different doses of TCDD. Each dose treatment was replicated with three times. The error bar indicated standard deviation.
In (A)–(C), different letters above the error bars indicate significant difference among means at Type I error = 0.05 (LSD test). In (D),
the asterisk indicates significant difference between two treatments in each dose of TCDD at Type I error = 0.05 (LSD test).
328 K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331
and TEQ levels of R3 has been expected since it flows
through Sihwa industrial complex which is one of the
most active industrialized zone in Korea, receiving a
direct load of industrial wastes.
Another two sediment samples were also collected
from the R6 (Miho-UP) and R7 (Miho-DN) sites adja-
cent to the Jungbu expressway and some factories. The
total PCDD/PCDFs concentrations and TEQ levels of
both samples were found to be 16.02 and 16.45 pg/g,
and 0.50 and 0.64 TEQ pg/g, respectively. In fact, the
total concentrations and TEQ levels of these samples
were expected to be equivalent or lower than R8. In con-
trast, the total concentrations and TEQ levels detected
in these samples showed about 3–4 times higher, despite
no direct sources of the PCDD/PCDFs near this river.
We speculate that the sources of PCDD/PCDFs entering
this river have been attributed to the automobile exhaust
gases (atmospheric transport and deposition).
Different congeners of PCDD/PCDFs were found in
different sediment localities. Most abundant congeners
included OCDD, OCDF, HpCDD, HpCDF, HxCDD,
and HxCDF. Generally, TCDD and TCDF are easily
accumulated in biological organisms, while OCDD
and OCDF are readily deposited in sediments due to
their low solubility in water (Muir et al., 1991). These re-
sults indicate that three localities of Singil (R1, R2 and
R3) and two localities of Miho (R6 and R7) are relatively
highly contaminated sites above 10 pg/g of total PCDD/
PCDFs.
TCDD has been regarded as less potent in inverte-
brates because of poor binding affinities of their Ah
receptor to the toxicant (Butler et al., 2001). However,
this study showed that even ppt levels of TCDD could
inhibit insect immune capacity. Insect immune system
involves both cellular and humoral reactions and exerts
their reaction after specific non-self pattern recognition
(Gillespie and Kanost, 1997). They can recognize bacte-
rial or fungal infection by binding pathogen-related sur-
face molecules such as peptidoglycan of Gram-positive
bacteria, lipopolysaccharides of Gram-negative bacteria,
and b-1,3-glycan of fungi (Yu et al., 2002). This recogni-
tion is mediated by cytokine like PSP1 or eicosanoids to
activate actual immune reactions (Clark et al., 1997;
Stanley, 2000). In humoral immune reactions, insect
can express several antibacterial peptides such as lyso-
zyme, cecropin, attacin (Boman and Hultmark, 1987).
Sediment samples
PO a
ctiv
ity
Abs
at 4
95nm
/5 m
in0.00
0.05
0.10
0.15
0.2050% EtOH100ng AA
*
R1 R2 R3 R4
* **
PO a
ctiv
ityAb
s at
495
nm
/5 m
in
0.00
0.05
0.10
0.15
0.20
0.25
CON R1 R2 R3 R4 R5 R6 R7 R8 R9 R10 R11 R12
*
* **
Sediment samples(C) (D)
Apop
tosi
s (%
)
0
20
40
60
80
100
* *
* *
CON R1 R2 R3 R4 R5 R6 R7 R8 R9 R10 R11 R12
Num
ber o
f nod
ules
0
5
10
15
20
25
*
* *
*
CON R1 R2 R3 R4 R5 R6 R7 R8 R9 R10 R11 R12(A) (B)
Fig. 3. Biomonitoring using insect immune endpoints against the sediment samples (R1–R12, see details in Fig. 1) with reference to a
solvent control (�CON�). All sediment and control samples were prepared by the extraction method used for measurement of PCDD/
PCDFs. The extracted samples were analyzed by four different immunological assays described in Fig. 2. Each environmental sample
was replicated with three times. The error bar indicated standard deviation. In (A)–(C), different letters above the error bars indicate
significant differences of the environmental samples in reference to the control at Type I error = 0.05 (LSD test). In (D), the asterisk
indicates significant difference between two treatments in each environmental sample at Type I error = 0.05 (LSD test).
K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331 329
While the humoral immune reactions take several hours
to express such peptides, cellular immune reactions exe-
cuted by hemocytes (insect blood cells) can be induced in
several minutes (Ratcliffe et al., 1985). These include
phagocytosis, nodule formation, and encapsulation
by hemocytes. Phenoloxidase catalyzes oxidation of
L-DOPA and produces quinine molecules, which are
toxic to pathogens and also important in nodule forma-
tion and encapsulation reactions by forming melanized
coat around pathogens (Dularay and Lackie, 1985).
The phenoloxidase should be in an inactive form, pro-
phenoloxidase, and should be tightly regulated in its
activation (Soderhall and Smith, 1986).
In this study, TCDD significantly inhibited the acti-
vation of prophenoloxidase of S. exigua in the presence
of laminarin, and inhibited hemocyte nodule formation.
These inhibitory actions of TCDD can be explained by
the positive correlation between phenoloxidase activity
and PLA2 activity in S. exigua (Park and Kim, 2003) be-
cause TCDD could inhibit PLA2. Thus, we speculate
that the inhibition of PLA2 by TCDD could not activate
prophenoloxidase, which in turn did not complete
hemocyte nodulation. The inhibitory effect of TCDD
on vertebrate immune system has been suggested by
the hypersensitivity of thymus (Vos et al., 1997), where
TCDD also induces apoptosis of T cells (Camacho
et al., 2004). In S. exigua, TCDD also induced apoptosis
of hemocytes, in which the cells formed apoptotic blebb-
ings and vesicles, typical symptoms of apoptotic cells
(Cho and Kim, 2004). Interestingly, the immune reac-
tions against TCDD was highly sensitive but different
in threshold doses for susceptibility: 0.1 ppb in pheno-
loxidase activity, 1.0 ppb in apoptosis, and 1.0 pg in
nodule formation.
The highly sensitive immune systems of S. exigua
against TCDD were used to monitor PCDD/PCDFs in
the environmental samples. Four local sediment samples
(R1, R2, R3, and R4) inhibited the immune biomarkers.
When comparing both chemical and biological monitor-
ing results, two methods agreed three spots of Singil as
contamination areas (above 10 pg/g levels) as well as
six river sediment samples as relatively less-contami-
nated areas, but differed in the results in R4 (Ansan)
and R6 (Miho-UP) and R7 (Miho-DN), probably due
to relative non-specificity of biomarkers. The immune
reactions can be affected by other compounds in the
sample extracts. Because in most cases a variety of con-
taminants may be present in environment, reliable bio-
markers should have some degree of specificity.
Alternatively, a battery of biomarkers is required to

330 K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331
cross-check different classes of contaminants (Melancon,
1995). Our current test using the immune biomakers
suggests that heavy metals also give significant inhibi-
tory actions on insect immune systems (unpublished
data). So, to clarify somewhat vagueness in this study,
various environmental compounds should be tested in
their inhibitory actions against the insect immune sys-
tem. Despite some disparity between bio- and chemical
monitoring results, the biomarkers can be recommended
as a device warning the contamination of dioxins in the
environment because of a fast and inexpensive detection
method.
Acknowledgement
This study was funded as Ecotechnopia 21 project by
Korea Institute of Environmental Science and Technol-
ogy (KIEST). This support is gratefully acknowledged.
References
Alcock, R.E., Jones, K.C., 1996. Dioxins in the environment: a
review of trend data. Environ. Sci. Technol. 30, 3133–3143.
Boman, H.G., Hultmark, D., 1987. Cell-free immunity in
insects. Ann. Rev. Microbiol. 41, 103–126.
Butler, R.A., Kelley, M.L., Powell, W.H., Hahn, M.E., Van
Beneden, R.J., 2001. An aryl hydrocarbon receptor (AHR)
homologue from the soft-shell clam, Mya arenaria: evidence
that invertebrate AHR homologues lack 2,3,7,8-tetrachlo-
rodibenzo-p-dioxin and b-naphthoflavone binding. Gene
278, 223–234.
Camacho, I.A., Magarkatti, M., Nagarkatti, P.S., 2004.
Evidence for induction of apoptosis in T cells from murine
fetal thymus following perinatal exposure to 2,3,7,8-tetra-
chlorodibenzo-p-dioxin (TCDD). Toxicol. Sci. 78, 96–106.
Cho, S., Kim, Y., 2004. Hemocyte apoptosis induced by
entomopathogenic bacteria, Xenorhabdus and Photorhabdus,
in Bombyx mori. J. Asia-Pacific Entomol. 7, 195–200.
Clark, K.D., Pech, L.L., Strand, M.R., 1997. Isolation and
identification of a plasmatocyte spreading peptide from
hemolymph of the lepidopteran insect Pseudoplusia inclu-
dens. J. Biol. Chem. 272, 23440–23447.
Connolly, J.P., Pedersen, C., 1988. A thermodynamically based
evaluation of organic chemical accumulation in aquatic
organisms. Environ. Sci. Technol. 22, 120–127.
Dularay, B., Lackie, A.M., 1985. Haemocytic encapsulation
and the prophenoloxidase-activation pathway in the locust
Schistocerca gregaria Forsk. Insect Biochem. 15, 827–834.
Fattore, E., Vigano, L., Mariani, G., Guzzi, A., Benfenati, E.,
Fanelli, R., 2002. Polychlorinated dibenzo-p-dioxins and
dibenzofurans in River Po sediments. Chemosphere 49,
749–754.
Fiedler, H., 1999. National and regional dioxin and furan.
Organohal. Compd. 41, 473–476.
Gho, H.G., Lee, S.G., Lee, B.P., Choi, K.M., Kim, J.H., 1990.
Simple mass-rearing of beet armyworm, Spodoptera exigua
(Hubner) (Lepidoptera: Noctuidae), on an artificial diet.
J. Korean Appl. Entomol. 29, 180–183.
Gillespie, J.P., Kanost, M.R., 1997. Biological mediators of
insect immunity. Ann. Rev. Entomol. 29, 180–183.
Gobas, F.A.P.C., Mackay, D., 1987. Dynamics of hydrophobic
organic chemical bioconcentration in fish. Environ. Toxicol.
Chem. 6, 495–504.
Im, S.H., Karl, D.S., John, P.S., Chang, Y.S., Matsuda, M.,
Wakimoto, T., 2004. Concentration and accumulation
profiles of polychlorinated dibenzo-p-dioxins and dib-
enzofurans in aquatic tissues, and ambient air from South
Korea. Chemosphere 55, 1293–1438.
Kang, Y.S., Yamamuro, M., Masunaga, S., Nakanishi, J.,
2002. Specific biomagnification of polychlorinated dibenzo-
p-dioxins and dibenzofurans in tufted ducks (Aythya fulig-
ula), common cormorants (Phalacrocorax carbo) and their
prey from Lake Shinji, Japan. Chemosphere 46, 1373–1382.
Kim, Y., Lee, S.Y., Kim, M., Kim, S.D., 2001a. The survey of
PCDDs and PCDFs in ambient air of the urban and
industrial sires in Korea, 1998–99. Chemosphere 43, 501–
506.
Kim, Y., Yang, S.H., Kim, M., Shin, D.C., 2001b. PCDD and
PCDF exposures in workers and controls living near an
industrial waste incinerator. Chemosphere 43, 985–987.
Luebke, R.W., Copeland, C.B., Daniels, M., Lambert, A.L.,
Gilmour, M.I., 2001. Suppression of allergic immune
responses to house dust mite (HDM) in rats exposed to
2,3,7,8-TCDD. Toxicol. Sci. 62, 71–79.
Melancon, M.J., 1995. Bioindicators used in aquatic and
terrestrial monitoring. In: Hoffman, D.J., Rattner, B.A.,
Burton Jr., G.A., Cairns Jr., J. (Eds.), Handbook of
Ecotoxicology. Lewis Publishers, Boca Raton, FL, pp. 220–
240.
Mocarelli, P., Brambilla, P., Gerthoux, P.M., Patterson,
D.G.J., Needham, L.L., 1996. Change in sex ratio with
exposure to dioxin. Lancet 348, 409.
Muir, D.C.G., Lawrence, S., Holoka, M., Fairchild, W.L.,
Segstro, M.D., Webster, G.R.B., Servos, M.R., 1991.
Partitioning of polychlorinated dioxins and furans between
water, sediments and biota in lake mesocosms. Chemo-
sphere 25, 119–124.
Park, Y., Kim, Y., 2000. Eicosanoids rescue Spodoptera exigua
infected with Xenorhabdus nematophilus, the symbiotic
bacteria to the entomopathogenic nematode Steinernema
carpocapsae. J. Insect Physiol. 11, 1469–1476.
Park, Y., Kim, Y., 2003. Xenorhabdus nematophilus inhibits
p-bromophenacyl bromide (BPB)-sensitive PLA2 of Spo-
doptera exigua. Arch. Insect Biochem. Physiol. 54, 134–142.
Ratcliffe, N.A., Rowley, A.F., Fitzgerald, S.W., Rhodes, C.P.,
1985. Invertebrate immunity—basic concepts and recent
advances. Int. Rev. Cytol. 97, 183–350.
Safe, S., 1990. Polychlorinated biphenyls (PCBs), dibenzo-p-
dioxins (PCDDs), dibenzofurans (PCDFs), and related
compounds: environmental and mechanistic considerations
which support the development of toxic equivalency factors
(TEFs). Toxicology 21, 51–87.
Soderhall, K., Smith, V.J., 1986. Prophenoloxidase-activating
cascade as a recognition and defense system in arthropods.
In: Gupta, A.P. (Ed.), Hemocytic and Humoral Immunity
in Arthropods. Wiley, New York, pp. 251–286.

K.S. Ryoo et al. / Chemosphere 61 (2005) 323–331 331
Stanley, D.W., 2000. Eicosanoids in Invertebrate Signal
Transduction Systems. Princeton University Press, Prince-
ton, NJ.
Thomann, R.V., 1989. Bioaccumulation model of organic
chemical distribution in aquatic food chains. Environ. Sci.
Technol. 23, 699–707.
Van Zorge, J.A., Van Wijnen, J.H., Theelen, R.M.C., Olie, K.,
Vanden Berg, M., 1989. Assessment of the toxicity of
mixtures of halogenated dibenzo-p-dioxins and dibenzofu-
rans by use of toxicity equivalency factors TEF. Chemo-
sphere 19, 1881–1895.
Vos, J.G., De Heer, C., Van Loveren, H., 1997. Immunotoxic
effects of TCDD and toxic equivalency factors. Teratog.
Carcinog. Mutagen. 17, 275–284.
West, C.W., Ankley, G.T., Nichols, J.W., Elonen, G.E., Nessa,
D.E., 1997. Toxicity and bioaccumulation of 2,3,7,8-tetra-
chlorodibenzo-p-dioxin in long-term tests with the freshwater
benthic invertebrates Chironomus tentans and Lumbriculus
variegates. Environ. Toxicol. Chem. 16, 1287–1294.
Yu, X.-Q., Zhu, Y.-F., Ma, C., Fabrick, J.A., Kanost, M.R.,
2002. Pattern recognition proteins in Manduca sexta
plasma. Insect Biochem. Mol. Biol. 32, 1287–1293.
Top Related
Copyright © 2022 FDOKUMEN

