







Bahasa
Halaman
Hukum


LAPORAN HASIL PENELITIAN
PENGEMBANGAN TEKNIK PENGAWETAN Pediococcus acidilactici
U318 SEBAGAI KULTUR STARTER FERMENTASI URUTAN
Oleh
IR. NYOMAN SEMADI ANTARA, MP., Ph.D. ANAK AGUNG MADE DEWI ANGGRENI, S.TP., M.Si.
Dibiayai oleh Direktorat Jendral Pendidikan Tinggi, Kementerian Pendidikan Nasional, sesuai dengan SP2HP No.: 175/SP2HP/PL/Dit.litabmas/IV/2011,
tanggal 14 April 2011
JURUSAN TEKNOLOGI INDUSTRI PERTANIAN FAKULTAS TEKNOLOGI PERTANIAN
UNIVERSITAS UDAYANA 2011

i
HALAMAN PENGESAHAN LAPORAN AKHIR
1. Judul Penelitian : Pengembangan Teknik Pengawetan Pediococcus acidilactici U318 sebagai Kultur Starter Fermentasi Urutan 2. Ketua Peneliti
a. Nama Lengkap : Prof. Ir. Nyoman Semadi Antara, MP., Ph.D.
b. Jenis Kelamin : Laki-laki
c. NIP : 19610923 198702 1 001
d. Pangkat/Golongan : Pembina Tk.I / IVb
e. Jabatan Fungsional : Guru Besar
f. Fakultas/Jurusan : Teknologi Pertanian / Teknologi Industri Pertanian
g. Perguruan Tinggi : Universitas Udayana
h. Pusat Penelitian : -
3. Jumlah Tim Peneliti : 2 orang
4. Lokasi Penelitian : Laboratorium Bioindustri, FTP – Unud
5. Kerja Sama dengan Institusi Lain: Tidak Ada
6. Masa Penelitian : 8 bulan
7. Biaya yang Diperlukan : Rp 31.000.000,00 (Tiga puluh satu juta rupiah)
Denpasar, 24 Nopember 2011
Mengetahui, Ketua Peneliti, Dekan Prof. Dr. Ir. G.P. Ganda Putra, MP. Prof. Ir. Nyoman S. Antara, MP., Ph.D. NIP.: 19620930 198803 1 001 NIP.: 19610923 198702 1 001
Mengetahui Ketua Lembaga Penelitian dan Pengabdian Kepada Masyarakat
Universitas Udayana
Prof. Dr. Ir. I Ketut Satriawan, MT. NIP.: 19640717 198903 1 001

ii
PRAKATA
Penelitian mengenai Pengembangan Teknik Pengawetan Pediococcus
acidilactici U318 sebagai Kultur Starter Fermentasi Urutan dapat terselesaikan
dengan baik, walaupun ada beberapa perubahan yang dilakukan karena adanya
perubahan besaran dana yang dialokasikan untuk penelitian ini. Selain itu,
karakteristik pektin dan karagenan yang berbeda dengan maizena juga
merupakan penyebab dilakukan perubahan terhadap konsentrasi dari bahan
tersebut yang digunakan sebagai bahan proteksi pada teknik pengawetan BAL
yang dikembangkan.
Penelitian ini dibiayai dari skema dana Penelitian Fundamental. Untuk itu
kami mengucapkan terima kasih kepada Kementerian Pendidikan dan
Kebudayaan yang telah mengalokasikan anggaran untuk jalannya penelitian ini,
sehingga penelitian ini dapat diselesaikan dengan baik sesuai dengan waktu
yang dijadwalkan. Kami juga mengucapkan terima kasih kepada Dekan Fakultas
Teknologi Pertanian, Unud yang telah memfasilitasi kelancaran pelaksanaan
penelitian ini.
Kami sangat berharap agar hasil penelitian ini dapat berkontribusi untuk
perkembangan ilmu dan teknologi terutama dibidang pengembangan teknik
pengawetan kultur starter untuk keperluan proses fermentasi.
Denpasar, Nopember 2011
Penulis

iii
DAFTAR ISI
HALAMAN PENGESAHAN ............................................................................. i PRAKATA ....................................................................................................... ii DAFTAR TABEL ............................................................................................. iii DAFTAR GAMBAR ......................................................................................... iv DAFTAR LAMPIRAN ...................................................................................... vi I. PENDAHULUAN ......................................................................................... 1 II. TINJAUAN PUSTAKA ................................................................................ 2 III. TUJUAN DAN MANFAAT PENELITIAN .................................................... 7 IV. METODE PENELITIAN ............................................................................. 10 V. HASIL DAN PEMBAHASAN ...................................................................... 11 VI. KESIMPULAN DAN SARAN ..................................................................... 16 DAFTAR PUSTAKA ........................................................................................ 17 LAMPIRAN ...................................................................................................... 19

iv
DAFTAR TABEL
Tabel 1. Nilai rata-rata total P. acidilactici U318 (cfu/g) sebelum dikeringkan 10
Tabel 2. Nilai rata-rata total P. acidilactici U318 (cfu/g) setelah dikeringkan 10
Tabel 3. Nilai rata-rata total mikroba (cfu/g) pada bubuk inokulum sebelum dikeringkan ……………………………………………………………. 12
Tabel 4. Nilai rata-rata total mikroba (cfu/g) pada bubuk inokulum setelah dikeringkan ……………………………………………………………. 12

v
DAFTAR GAMBAR
Gambar 1. P. acidilactici U318 pada bubuk inokulum maizena 5% …….. 11
Gambar 2. Total bakteri asam laktat () dan Enterobacteriaceae () pada
urutan dengan perlakuan penambahan lemak ........................ 13

vi
DAFTAR LAMPIRAN
Lampiran 1. Buram Publikasi
Lampiran 2. Instrumen Penelitian
Lampiran 3. Personalia Tenaga Peneliti

1
I. PENDAHULUAN
Urutan merupakan salah satu produk makanan asli Indonesia dari Bali.
Urutan mempunyai karakteristik yang berbeda dengan sosis terfermentasi lainnya
karena penggunaan jenis bumbu yang berbeda dalam formulasinya. Urutan dibuat
dari campuran potongan kecil daging dan lemak yang dicampur dengan bumbu
(bawang putih, kunyit, kencur, cabai, lengkuas, dan merica), garam, dan gula,
kemudian dimasukkan ke dalam usus yang sudah dibersihkan, dan dikeringkan di
bawah sinar matahari selama 2-5 hari. Secara tradisional, fermentasi terjadi secara
alami pada saat pengeringan.
Kegagalan fermentasi dalam pembuatan urutan, menyebabkan menurunnya
minat masyarakat Bali untuk memproduksinya. Masyarakat lebih banyak
memproduksi urutan tanpa fermentasi. Hal ini terjadi karena fermentasi terjadi
secara alami dan tidak terkontrol, sehingga sering terjadi pertumbuhan
mikroorganisme yang tidak diinginkan yang menyebabkan mutu menjadi tidak
konsisten. Selain itu banyaknya beredar sosis terfermentasi, seperti salami,
paperoni, bologna, dll., di pasaran untuk pasokan hotel/restoran dan keperluan
lainnya menyebabkan produk lokal tidak berkembang. Dilain pihak urutan
merupakan produk daging terfermentasi lokal yang mempunyai potensi untuk
bersaing dengan produk sosis terfermentasi dari Eropa.
Bakteri Asam Laktat (BAL) merupakan bakteri yang dominan tumbuh
selama proses fermentasi urutan yang dibuat secara tradisional. Penggunaan
kultur starter BAL dalam proses pembuatan urutan telah dibuktikan dapat
mengatasi kegagalan fermentasi (Aryanta, 1996; Aryanta, 1998; Antara et al.,
2004). Untuk mengatasi kegagalan fermentasi, penyediaan kultur BAL dalam
bentuk bubuk kultur starter sangat diperlukan.
Proses produksi bubuk kultur starter melalui proses pengeringan dengan
menggunakan suhu tinggi. Di lain pihak, sel BAL sangat sensitif terhadap panas,
sehingga dengan proses pengeringan suhu tinggi akan menyebabkan tingkat
kematian sel yang tinggi. Untuk mengatasi permasalahan tersebut perlu
dikembangkan teknik pengawetan sel BAL dengan melindungi sel BAL selama
proses pengeringan.

2
II. TINJAUAN PUSTAKA
Secara umum, sosis terfermentasi didefinisikan sebagai campuran daging
dan lemak dicampur dengan garam, gula, bumbu-bumbu, dan nitrat dan/atau nitrit,
yang kemudian dimasukkan ke dalam casing, dan selanjutnya dilakukan
fermentasi yang dilanjutkan dengan proses pengeringan atau pematangan
(Bukenhukes, 1993; Hugas dan Monfort, 1997; Caplice dan Fltzgerald, 1999). Di
negara Eropah produksi sosis terfermentasi melalui tiga tahapan proses, yaitu
formulasi, fermentasi, dan pengeringan/pematangan. Pada tahap formulasi, semua
campuran bahan disiapkan untuk dimasukkan ke dalam casing. Selama fermentasi
reaksi-reaksi mikrobiologis dihasilkan secara simultan dan mempengaruhi satu
dengan yang lainnya, yaitu pembentukkan nitrit oksida oleh bakteri pereduksi
nitrat dan nitrit, dan penurunan pH akibat terbentuknya asam laktat. Pada tahap
pengeringan yang juga proses pematangan terjadi perkembangan citarasa (flavor),
aroma, dan tekstur dari sosis (Ordonez et al., 1999). Di Eropah proses fermentasi
sosis dilakukan pada suhu di bawah 26oC (Lucke, 1998), sedangkan di Amerika
Serikat kebanyakan sosis terfermentasi dilakukan fermentasi pada suhu 30oC
sampai 45oC dalam jangka waktu yang pendek (sekitar 24 jam).
Di Indonesia, khususnya di Bali, produksi sosis terfermentasi dilakukan
dalam dua tahapan, yaitu tahap formulasi dan fermentasi/pengeringan. Proses
fermentasi secara tradisional terjadi pada saat pengeringan di bawah sinar
matahari (pada siang hari) dan di atas tungku tradisional (pada malam hari).
(Antara, 2004).
Pada fermentasi sosis, BAL yang berasal dari bahan baku dan lingkungan
bertanggung jawab untuk produksi asam laktat dan menurunkan pH. Komunitas
awal mikroba sangat bervariasi hampir sama dengan yang ditemukan pada daging
segar, seperti lactobacilli, enterobacteria, Psudomonas, Bacillus, Achromobacter,
Flavobacterium, dan lain-lain, dan juga kapang dan khamir (Ordonez et al., 1999).
Pada tahap fermentasi, selanjutnya dominasi BAL sangat penting perannya untuk
menghambat bakteri pembusuk dan patogen. Lb. sake dan Lb. curvatus
merupakan species BAL dominan pada salami (Naple type) (Hugas dan Monfort,
1997; Coppola et al., 2000), tetapi pada urutan Lb. plantarum, P. acidilactici, dan
Lb. farciminis merupakan BAL dominan selama fermentasi dengan suksesi

3
pertumbuhan tertentu (Antara et al., 2002). Sampai saat ini urutan masih
diproduksi secara tradisional dengan fermentasi yang alami. Holzapfel (2002)
menyatakan bahwa fermentasi alami memerlukan waktu yang lebih lama dan
resiko kegagalan yang tinggi. Proses ini dapat diperpendek dengan menginokulasi
sebagian bahan yang telah terfermentasi dengan baik (back-slopping) atau
menggunakan kultur starter yang terseleksi (Ordonez et al., 1999; Gariga et al.,
1996).
BAL sangat penting perannya dalam makanan terfermentasi karena
memberikan keuntungan seperti dapat memperbaiki karakteristik citarasa dan
berperan sebagai pengawet (biopreservative) pada makanan. Sebagai kultur starter,
BAL sangat luas digunakan pada proses-proses fermentasi, seperti fermentasi susu
(Fitzsimons et al., 1999; Parente et al., 1997), sayuran (Sanchez et al., 2000;
Kalac et al., 2000), minuman beralkohol (Patarata et al., 1994; Pattison et al.,
1998), dan daging ( Hammes dan Hertel, 1998). Dan juga sudah terbukti bahwa
mutu sosis yang lebih baik dicapai dengan menggunakan kultur starter (Gariga et
al., 1996; Stahnke, 1995; Erkkila dan Petaja, 2000; Erkkila et al., 2001). Sudah
dinyatakan bahwa urutan mempunyai karakteristik yang berbeda dengan jenis
sosis terfermentasi lainnya, dan kenyataan bahwa penggunaan bumbu yang lebih
beragam selalu dilakukan pada proses pembuatan urutan, sehingga
pengembangan kultur starter yang spesifik untuk urutan sangat dibutuhkan.
Kultur starter didefinisikan sebagai preparasi mikrobia dalam jumlah yang
banyak dari paling sedikit satu jenis mikrobia yang ditambahkan ke dalam bahan
baku untuk menghasilkan makanan terfermentasi dengan memacu proses
fermentasinya (Leroy dan De Vuyst, 2003). Kelompok BAL sudah lama
diaplikasikan pada makanan dan minuman terfermentasi (Ray, 1992; Wood, 1997).
Pertumbuhan yang cepat dan penurunan pH akibat asam laktat yang dihasilkan
menghasilkan makanan terfermentasi yang aman dan bermutu. Pada preparasi
sosis, P. acidilactici, L. plantarum dan Staphylococcus carnosus diinokulasi pada
fermentasi sosis (Mayra-Makinen dan Bigret, 1998).
BAL umumnya termasuk ke dalam kelompok mesofilik, dan dapat tumbuh
pada kisaran suhu dari 5oC sampai 45oC. Bakteri ini tumbuh pada kisaran pH 4,0
– 4,5, beberapa aktif pada pH 9,6 dan beberapa pada pH 3,2. Kelompok bakteri ini

4
umumnya mempunyai aktivitas proteolitik dan lipolitik yang lemah dan
membutuhkan asam amino, basa purin dan pirimidin, dan vitamin B untuk
tumbuh (Jay, 1996). BAL memproduksi asam laktat dari heksosa dan tidak
mempunyai fungsi heme yang berkaitan dengan rantai transport electron dan
fungsi siklus Krebs. Jalur metabolisme heksosa dibagi menjadi dua kelompok
yaitu homofermentatif dan heterofermentatif. Kelompok BAL homofermentative
menghasilkan asam laktat yang utama dari fermentasi glukosa. Homofermenter
menggunakan jalur Embeden-Meyerhof-Parnas untuk menghasilkan dua mol
asam laktat per mol glukosa dan menghasilkan energi sekitar dua kalinya
dibandingkan dengan heterofermentatif. Kelompok BAL heterofermentatif
memproduksi asam laktat, CO2 dan etanol dari glukosa dalam jumlah yang sama
melalui jalur fosfoketolase.
Urutan merupakan salah satu produk makanan asli Indonesia (Bali). Di Bali
urutan biasanya dibuat untuk merayakan hari raya Galungan, hari suci bagi
masyarakat Hindu di Bali. Urutan mempunyai karakteristik yang berbeda dengan
sosis terfermentasi lainnya karena penggunaan jenis bumbu yang berbeda dalam
formulasinya. Urutan dibuat dari campuran potongan kecil daging dan lemak
yang dicampur dengan bumbu (bawang putih, kunyit, kencur, cabai, laos, dan
merica), garam, dan gula. Daging yang biasanya digunakan adalah daging
babi. Campuran adonan ini kemudian dimasukkan ke dalam usus yang sudah
dibersihkan, dan dikeringkan di bawah sinar matahari selama 2-5 hari. Fermentasi
terjadi secara alami pada saat pengeringan. Penggunaan banyak jenis bumbu dan
fermentasi pada suhu yang tinggi berhubungan dengan karakteristik BAL yang
terlibat di dalam proses fermentasi (Antara, 2004)
Mutu yang tidak konsisten dan resiko kegagalan yang tinggi merupakan
masalah yang umum pada makanan terfermentasi yang diproduksi secara
tradisional. Resiko kegagalan dalam produksi urutan sangat tinggi yang ditandai
dengan aroma yang tidak sedap dari produk. Untuk memperbaiki mutu urutan,
penggunaan kultur starter dalam produksi urutan perlu dikembangkan.
Penggunaan kultur indigenus akan memberikan keuntungan terutama proses
fermentasi yang terkontrol dan produk akhir dapat distandarisasi.
Penelitian sebelumnya menunjukkan bahwa BAL mendominasi mikrobia

5
yang berperan dalam proses fermentasi urutan (Antara et al., 2002). Aryanta
(1996) melaporkan bahwa secara penotipik genus Lactobacillus dan Pediococcus
berperan dalam proses fermentasi urutan. Dengan karakteristik penotipik untuk
mengidentifikasi BAL mempunyai keterbatasan untuk mendapatkan hasil yang
akurat sampai pada tingkat spesies ataupun strain. Antara et al. (2002)
menggunakan kombinasi metode penotipik dan analisis sekuen dari 16S rDNA
diperoleh hasil yang lebih akurat mengenai BAL yang terlibat di dalam proses
fermentasi urutan secara alami. Dari identifikasi tersebut diperoleh tiga strain Lb.
plantarum, empat strain Lb. farciminis, dan dua strain yang merupakan obligat
heterofermentative yaitu Lb. fermentum dan Lb. hilgardii. Ada dua spesies dari
Pediococcus yang teridentifikasi yaitu P. acidilactici (dua strain) dan P.
pentosaceus (dua strain). Dari semua spesies yang teridentifikasi, ternyata ada tiga
spesies yang mendominasi fermentasi dengan suksesi tertentu. Ketiga spesies
tersebut adalah Lb. plantarum, P. acidilactici, dan Lb. farciminis.
Hasil skrining terhadap BAL penghasil bakteriosin menunjukkan bahwa
strain P. acidilactici U318 dapat menghasilkan bakteriosin dengan spektrum
penghambatan yang luas (Antara, 2000). Keberadaan strain penghasil
bakteriosin berperan nyata terhadap suksesi pertumbuhan BAL dalam urutan
selama fermentasi (Antara 2004). Hasil karakterisasi semua strain BAL telah
terpilih tiga strain yang dapat digunakan sebagai kultur starter tunggal maupun
ganda. Sebagai kultur starter tunggal Lb. plantarum U201, P. acidilactici U318
dan P. pentosaceus U208 sudah dicoba dalam produksi urutan (Antara et al.,
2002). Ketiga strain ini yang merupakan indigenus strain dipilih sebagai kultur
starter karena keberadaannya di dalam urutan selama fermentasi secara alami. Lb.
plantarum U201 merupakan BAL yang mendominasi pertumbuhan pada awal
fermentasi dan mempunyai peran untuk menurunkan pH yang cepat pada awal
fermentasi. P. acidilactici U318 merupakan strain BAL penghasil bakteriosin yang
dapat mengontrol fermentasi. Walaupun strain P. pentosaceus keberadaannya
tidak dominan, namun dapat tumbuh dengan baik pada suhu 30oC – 45oC yang
merupakan suhu fermentasi urutan.
Penggunaan Lb. plantarum U201 dan P. acidilactici U318 sebagai kultur
starter tunggal dan ganda dalam produksi urutan juga sudah diteliti secara intensif.

6
Dari hasil penelitian ini penggunaan kedua strain bersama sebagai kultur starter
ganda memperlihatkan hasil yang baik, Lb. plantarum U201 berperan untuk
menurunkan pH dalam waktu yang cepat pada awal fermentasi. Penurunan pH ini
sangat penting untuk menghambat bakteri pembusuk dan patogen. P. acidilactici
U318 yang merupakan strain penghasil bakteriosin dapat mengontrol fermentasi,
yaitu dengan menghambat pertumbuhan Lb. plantarum sehingga penurunan pH
dapat dicegah tidak terlalu rendah. Selain itu keberadaan P. acidilactici U318
dapat menghambat micrococci secara sempurna pada akhir fermentasi (Antara et
al., 2004).
Penggunaan kultur starter dalam proses produksi urutan dapat mencegah
resiko kegagalan dan dihasilkan mutu urutan yang konsisten. Walaupun demikian,
penyiapan kultur starter memerlukan ketrampilan dan kondisi yang aseptis. Hal
ini terlalu rumit dilakukan untuk skala produksi yang lebih besar. Untuk itu
pengembangan kultur starter yang siap pakai sangat penting dilakukan, sehingga
penggunaan kultur starter dalam bentuk kultur starter siap pakai dapat dilakukan
dengan lebih mudah pada skala industri. Pada penelitian sebelumnya mengenai
proses pembuatan bubuk kultur starter, proses pengeringan merupakan proses
yang sangat kritis karena terjadi penurunan viabilitas yang nyata dan kontaminasi
mikroorganisme lain. Penggunaan oven vakum dengan suhu rendah merupakan
proses pengeringan yang baik digunakan dalam pengeringan kultur starter (Antara
et al., 2006). Penelitian selanjutnya menunjukkan bahwa penggunaan pengisi
berbasis karbohidrat (pati) tidak memberikan pengawetan yang nyata terhadap
viabilitas P. acidilactici U318 (Trisna et al., 2008), sehingga diperlukan penelitian
lanjutan untuk mendapatkan teknik pengawetan yang tepat terhadap sel P.
acideilactici U318 dengan hasil viabilitas yang tinggi dan dapat disimpan dalam
jangka waktu yang lama.

7
III. TUJUAN DAN MANFAAT PENELITIAN
Tujuan Penelitian
Tujuan penelitian yang dilakukan adalah untuk memperoleh teknik pengawetan
terhadap sel Pediococcus acidilactici U318 dalam pembuatan bubuk kultur starter,
sehingga diperoleh bubuk kultur starter dengan viabilitas sel yang tinggi.
Manfaat Penelitian
Luaran penelitian ini adalah teknik pengawetan baketri asam laktat, khususnya
Pediococcus acidilactici U318, yang bermanfaat untuk proses pembuatan bubuk
inokulum dengan teknologi sederhana dan dapat dimanfaatkan untuk
mengembangkan proses produksi bubuk inokulum. Lebih lanjut, bubuk inokulum
akan bermanfaat bagi masyarakat yang berminat untuk memproduksi urutan, dan
kegagalan fermentasi dapat ditanggulangi.

8
IV. METODE PENELITIAN
Strain Bakteri Asam Laktat
Strain BAL digunakan dalam penelitian ini adalah P. acidilactici U318. Strain
tersebut diperoleh dari koleksi UPT Laboratorium Terpadu Biosain dan
Bioteknologi, Universitas Udayana.
Media dan Bahan
Media: glukosa, ekstrak khamir, ekstrak daging, pepton, natrium asetat (tri-hidrat),
MgSO4, MnSO4, FeSO4, NaCl, Tween 80, agar, MRS broth, Plate Count Agar,
Violet Red Bile Dxtrose Agar.
Bahan: tepung maizena, pectin, dan karagenan (tepung agar)
Alat
Inkubator, oven vacum, laminair flow, sentrifugasi, mikroskop, dan alat-alat gelas.
Persiapan Kultur
Strain BAL diperbanyak dengan menggunakan media sintetik, yaitu GYP broth
yang terdiri dari glukosa 10 g/l, ekstrak yeast 10 g/l, larutan garam 5 ml/l
(MgSO4.7H2O 40 mg/ml, MnSO4.4H2O 2 mg/ml, FeSO4.7H2O 2 mg/ml, dan
NaCl 2 mg/ml), Tween 80 0,5 mg/l. Masing-masing kultur ditumbuhkan pada
suhu 30oC dan ditentukan fase logaritmiknya untuk menentukan waktu panen sel.
Pada saat fase logaritmik sel bakteri dipanen dengan sentrifugasi pada 3500 rpm
selama 20 menit. Sel masing-masing strain dicuci dengan menggunakan larutan
fisiologis (larutan NaCl 0,85%) steril dan disuspensikan kembali dengan larutan
yang sama untuk mendapatkan jumlah sel sekitar 109 sel /ml.
Produksi Kultur starter
Kultur dari strain BAL yang sudah dipersiapkan di dalam larutan fisiologis
ditambahkan 10% susu skim dan 0,1% sodium glutamat. Cairan kultur selanjutnya
ditambahkan bahan pelindung sesuai dengan perlakuan dan dilakukan
pengeringan di dalam oven vakum pada suhu 45oC sehingga diperoleh bubuk
kultur starter.
Rancangan Percobaan
Percobaan dilaksanakan dalam 2 tahap:
Tahap I: Percobaan Pembuatan Bubuk Inokulum
Percobaan yang dirancang dengan rancangan acak kelompok (RAK) dan pola

9
percobaan faktorial yang terdiri dari dua faktor. Faktor pertama adalah jenis
pelindung yang terdiri dari 3 aras (tepung maizena, tepung pektin, dan tepung
agar/karagenan), dan faktor kedua adalah konsentrasi bahan pelindung yang
terdiri dari 4 aras (0%, 5%, dan 10%), sehingga dari kedua faktor tersebut
diperoleh 9 kombinasi percobaan. Masing-masing percobaan dilakukan dalam tiga
kelompok, sehingga akan dilakukan 27 unit percobaan.
Data yang diperoleh dinalisis dengan analisis variance (sidik ragam) untuk
menentukan pengaruh masing-masing perlakuan dan interaksinya. Apabila hasil
sidik ragam menunjukkan pengaruh yang nyata, maka dilanjutkan dengan uji t
untuk menentukan perbedaan antar aras perlakuan ataupun interaksi perlakuan.
Tahap II: Uji Bubuk Inokulum Pada Adonan Urutan
Pada tahapan ini, bubuk inokulum yang memperlihatkan viabilitas BAL terbaik
diuji kemampuan tumbuhnya pada media urutan. Media urutan merupakan media
yang sesuai dengan formulasi urutan dan tidak disterilkan. Media urutan
ditempatkan dalam petri steril dan diinokulasi dengan bubuk inokulum terbaik
dari percobaan Tahap I. Setelah fermentasi dilakukan análisis terhadap
pertumbuhan BAL dan karakteristik lainnya. Pada percobaan Tahap II dirancang
percobaan dengan memvariasi lemak yang ditambahkan ke dalam formulasi
lemak.
Variabel yang Diamati
Pada percobaan Tahap I dianalisis variabel yang merupakan indikator viabilitas
kultur starter. Variabel yang diamati adalah total bakteri asam laktat dan total
mikroba di dalam bubuk kultur starter.
Pada percobaan Tahap II dianalisis kemampuan BAL tumbuh dalam media urutan.
Kemampuan BAL tumbuh dalam media urutan dilakukan dengan cara
menumbuhkan kultur starter (bubuk kultur starter) dalam cawan petri steril yang
sudah diisi adonan urutan. Selain BAL, juga dilakukan análisis terhadap
Enterobacteriaceae dan karakteristik adonan setelah dilakukan fermentasi.
10
V. HASIL DAN PEMBAHASAN
Percobaan Tahap I:
Total Pediococcus acidilactici U318 Pada Bubuk Inokulum
Hasil analisis keragaman menunjukkan bahwa perlakuan jenis (karagenan, pektin,
dan maizena) dan konsentrasi bahan proteksi serta interaksinya berpengaruh
sangat nyata (P<0,01) terhadap viabilitas total P. acidilactici U318 sebelum dan
setelah dikeringkan. Ini membuktikan bahwa P. acidilactici U318 dapat bertahan
hidup diketiga jenis bahan proteksi dengan konsentrasi yang berbeda. Nilai
rata-rata total P. acidilactici U318 sebelum dan sesudah dikeringkan dapat
dilihat pada Tabel 2 dan 3.
Tabel 1. Nilai rata-rata total P. acidilactici U318 (cfu/g) sebelum dikeringkan
Jenis Bahan Proteksi
Konsentrasi bahan proteksi (%) 0 5 10
Karagenan 3,39 x 1010 a 1,95 x 109 b 4,44 x 109 b Pektin 4,57 x 1010 a 2,58 x 1010 a 3,42 x 109 b
Maizena 3,13 x 1010 a 2,87 x 1010 a 5,23 X 109 b Keterangan : Huruf yang sama dibelakang rata-rata menunjukkan perbedaan yang tidak nyata, (P>0,05).
Tabel 2. Nilai rata-rata total P. acidilactici U318 (cfu/g) setelah dikeringkan
Jenis Bahan Proteksi
Konsentrasi bahan proteksi (%) 0 5 10
Karagenan 2,78 x 109 cd 9,30 x 108 d 3,85 x 109 b Pektin 3,78 x 109 bc 3,07 x 109 c 3,22 x 109 c
Maizena 1,16 x 1010 a 1,26 x 1010 a 4,46 x 109 b Keterangan : Huruf yang sama dibelakang rata-rata menunjukan perbedaan yang tidak nyata, (P>0,05).
Hasil dari penelitian yang telah dilakukan menunjukkan bahwa rata-rata total P.
acidilactici U318 mengalami penurunan pada masing - masing perlakuan
setelah proses pengeringan, hal tersebut mungkin disebabkan oleh masing-masing
bahan proteksi memililiki karakteristik polimer yang berbeda. Data yang
diperoleh memperlihatkan perlakuan bahan proteksi maizena memberikan

11
perlindungan yang paling baik setelah proses pengeringan berlangsung, dan pada
perlakuan maizena 5 % memberikan perlindungan terbaik yaitu sebesar 1,26 x
1010 cfu/g.
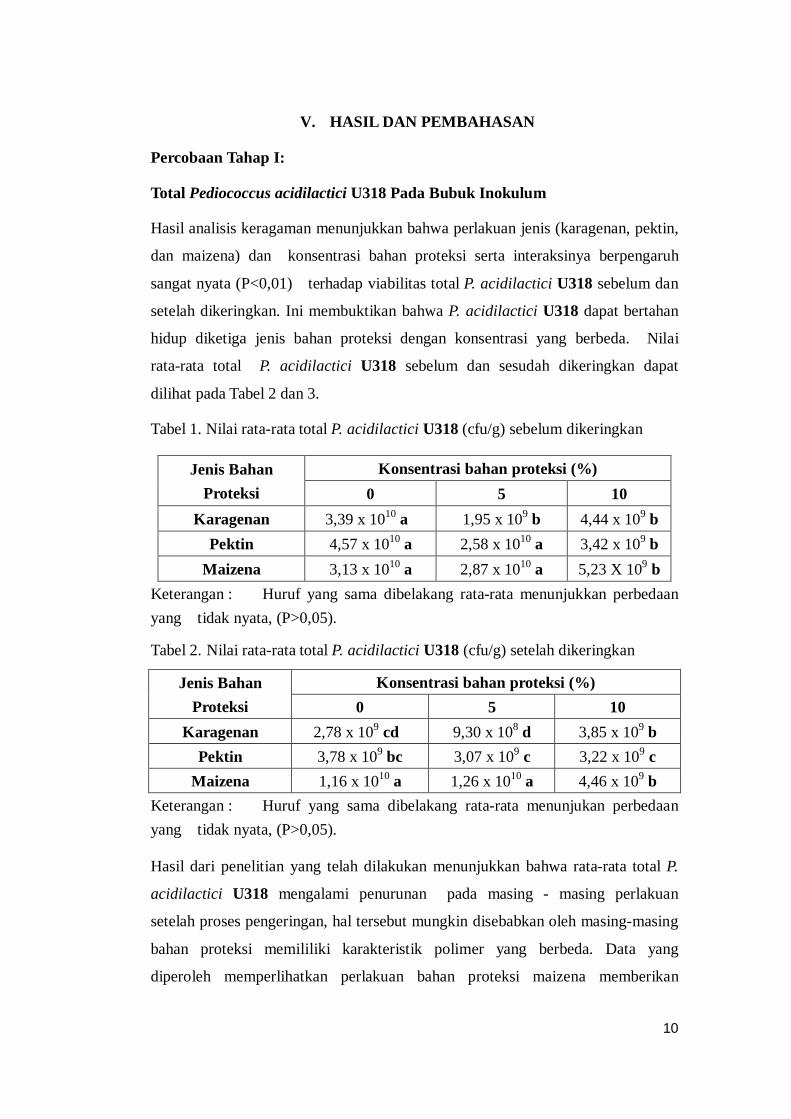
Pengamatan mikroskopis Pediocroccus acidilactici U318
Pengamatan secara mikroskopis P. acidilactici U318 menggunakan metode
Pewarnaan Gram, pada bubuk inokulum setelah pengeringan. Dari metode
pewarnaan gram tersebut diperoleh sembilan buah prefarat sesuai dengan
jumlah kombinasi perlakuan, yang selanjutnya kesembilan prefarat tersebut
diamati dan diambil gambarnya menggunakan mikroskop dengan pembesaran
1000X. Dari prefarat yang telah dibuat maka akan dihasilkan sembilan foto
mikroskopis dari masing-masing perlakuan. Pada Gambar 1 ini disajikan foto
mikroskopis P. acidilactici U318 .
Gambar 1 . P. acidilactici U318 pada bubuk inokulum maizena 5%
Total Mikroba
Hasil analisis keragaman menunjukkan bahwa perlakuan jenis (karagenan, pektin,
dan maizena) dan konsentrasi bahan proteksi serta interaksinya berpengaruh
sangat nyata (P<0,01) terhadap total mikroba sebelum dan setelah dikeringkan.
Hal tersebut membuktikan bahwa mikroba dapat hidup pada ketiga bahan proteksi
dengan konsentrasi yang berbeda, yang disebabkan karena pada prinsipnya semua
bakteri/mikroba membutuhkan zat organik seperti protein, lemak, karbohidrat air

12
serta beberapa zat anorganik seperti fosfor, kalsium, besi dan magnesium
(Wibowo dan Ristanto, 1988). Data nilai rata-rata total mikroba pada bubuk
inokulum sebelum dan setelah dikeringkan dapat dilihat pada Tabel dan Tabel
3 dan 4.
Tabel 3. Nilai rata-rata total mikroba (cfu/g) pada bubuk inokulum sebelum dikeringkan.
Jenis Bahan Proteksi
Konsentrasi bahan proteksi (%) 0 5 10
Karagenan 2,46 x 1011 ef 3,64 x 1011 f 3,45 x 1011 cd Pektin 2,46 x 1011 b 4,88 x 1011 c 5,12 x 1010 d
Maizena 2,46 x 1011 a 5,47 x 1010 b 3,93 x 1010 c
Tabel 4. Nilai rata-rata total mikroba (cfu/g) pada bubuk inokulum setelah dikeringkan.
Jenis Bahan Proteksi
Konsentrasi bahan proteksi (%) 0 5 10
Karagenan 8,43 x 1010 b 2,06 x 1010 c 1,83 x 1010 c Pektin 3,66 x 1010 d 4,63 x 1010 c 3,37 x 1010 c
Maizena 4,27 x 1010 a 3,65 x 1010 c 2,75 x 1010 cd
Percobaan Tahap II:
Total Bakteri Asam Laktat dan Enterobacteriaceae
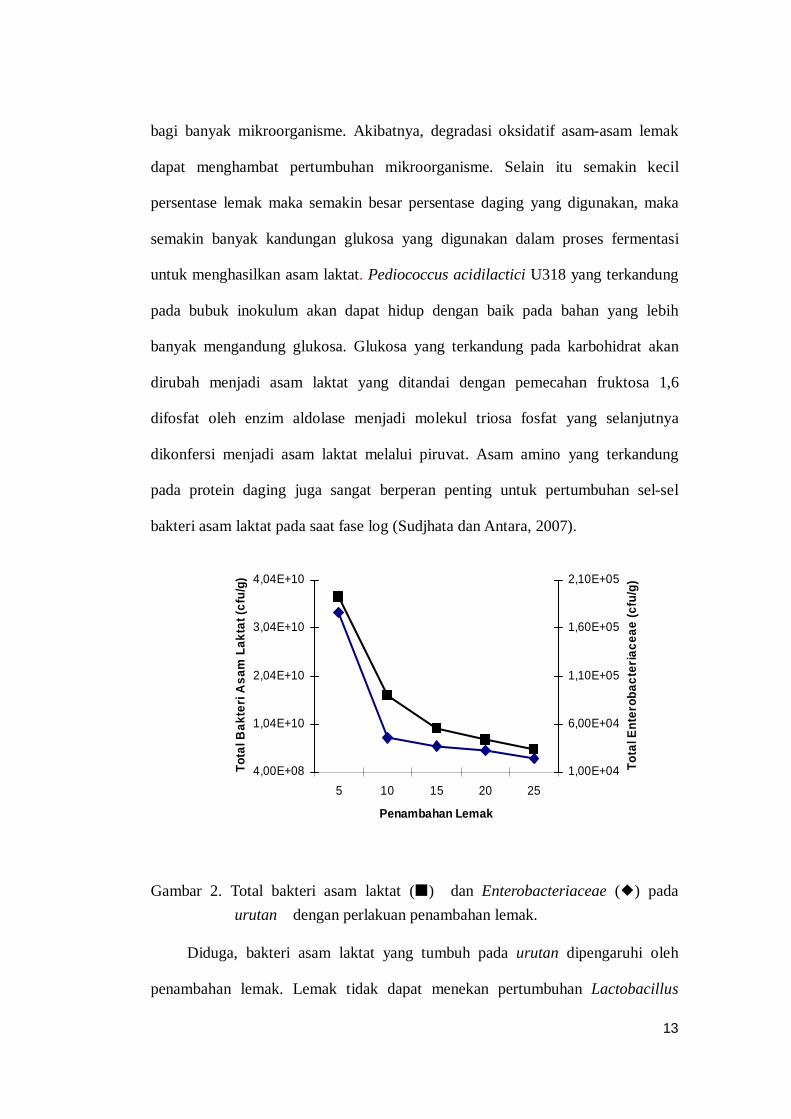
Hasil penelitian menunjukkan bahwa penambahan lemak pada urutan dapat
menurunkan jumlah populasi mikroorganisme. Kisaran populasi bakteri asam
laktat pada urutan adalah 5,2x109cfu/g sampai 3,7x1010cfu/g. Populasi bakteri
asam laktat ini mengalami penurunan dari 3,7x1010cfu/g (penambahan lemak 5%)
menjadi 5,2x109cfu/g (penambahan lemak 25%) (Gambar 2).
Pada saat fermentasi, lemak mengalami proses hidrolisis yang menghasilkan
asam-asam lemak. Menurut Leochowich (1971) dan Judge et al., (1989) dalam
Soeparno (1992), peroksida yang dihasilkan dari asam-asam lemak bersifat toksik
13
bagi banyak mikroorganisme. Akibatnya, degradasi oksidatif asam-asam lemak
dapat menghambat pertumbuhan mikroorganisme. Selain itu semakin kecil
persentase lemak maka semakin besar persentase daging yang digunakan, maka
semakin banyak kandungan glukosa yang digunakan dalam proses fermentasi
untuk menghasilkan asam laktat. Pediococcus acidilactici U318 yang terkandung
pada bubuk inokulum akan dapat hidup dengan baik pada bahan yang lebih
banyak mengandung glukosa. Glukosa yang terkandung pada karbohidrat akan
dirubah menjadi asam laktat yang ditandai dengan pemecahan fruktosa 1,6
difosfat oleh enzim aldolase menjadi molekul triosa fosfat yang selanjutnya
dikonfersi menjadi asam laktat melalui piruvat. Asam amino yang terkandung
pada protein daging juga sangat berperan penting untuk pertumbuhan sel-sel
bakteri asam laktat pada saat fase log (Sudjhata dan Antara, 2007).
Gambar 2. Total bakteri asam laktat () dan Enterobacteriaceae () pada urutan dengan perlakuan penambahan lemak.
Diduga, bakteri asam laktat yang tumbuh pada urutan dipengaruhi oleh
penambahan lemak. Lemak tidak dapat menekan pertumbuhan Lactobacillus
4,00E+08
1,04E+10
2,04E+10
3,04E+10
4,04E+10
5 10 15 20 25
Penambahan Lemak
Tota
l Bak
teri
Asa
m L
akta
t (cf
u/g)
1,00E+04
6,00E+04
1,10E+05
1,60E+05
2,10E+05
Tota
l Ent
erob
acte
riace
ae (c
fu/g
)

14
plantarum pada awal fermentasi sehingga bakteri inilah yang mendominasi
pertumbuhan BAL pada urutan. Antara (2004) menyebutkan Lactobacillus
plantarum lebih cepat menghasilkan asam dibanding dengan Pediococcus
acidilactici, sehingga menyebabkan penurunan pH. pH yang semakin rendah akan
menghambat pertumbuhan Enterobacteriaceae, sehingga pertumbuhan
Enterobacteriaceae pada urutan mengalami penurunan yaitu dari 1,77x105 cfu/g
(penambahan lemak 5%) menjadi 2,45x104 cfu/g (penambahan 25%) (Gambar 2).
Total Asam, pH, dan aw
Nilai total asam pada urutan berkisar antara 2,19% sampai 2,47%. Nilai
total asam pada urutan mengalami peningkatan dari 2,19% (penambahan lemak
5%) menjadi 2,47% (penambahan lemak 25%) (Gambar 3). Semakin banyak
kandungan karbohidrat dalam produk maka semakin banyak asam laktat yang
dihasilkan pada saat terfermentasi oleh bakteri asam laktat. Tetapi pada penelitian
ini terdapat perbedaan pada formulasi daging dan lemak yang digunakan yang
dapat berpengaruh pada total asam yang dihasilkan. Selain diduga dipengaruhi
oleh pertumbuhan Lactobacillus plantarum yang mendominasi pada urutan, juga
dipengaruhi oleh populasi Enterobacteriaceae. Pada saat fermentasi bakteri ini
dapat memetabolisme asam-asam amino secara enzimatik (deaminasi oksidatif
dan reduktif) menjadi ammonia, asam keto dan asam lemak (Soeparno ,1992).
Terdeaminasinya senyawa-senyawa amino organik ini menghasilkan
senyawa-senyawa basa yang dapat menetralkan asam sehingga pH cenderung
tinggi. Pada perlakuan penambahan lemak 5% lebih banyak mengandung protein
sehingga terjadi peningkatan pH yang lebih tinggi. Kisaran nilai pH urutan adalah
4,39 sampai 4,54. Nilai pH urutan ini mengalami penurunan dari perlakuan

15
penambahan lemak 5% yaitu sebesar 4,54 menjadi 4,39 pada perlakuan
penambahan lemak 25%.
Aktivitas air atau water activity (aw) dinyatakan sebagai jumlah air bebas
yang dapat digunakan mikroba untuk pertumbuhannya (Winarno, 1981).
Nilai aw urutan hasil penelitian berkisar antara 0,89–0,93. Bakteri asam
laktat Pediococcus acidilactici membutuhkan aw minimum 0,91-0,95 untuk
pertumbuhannya pada produk daging (Fenema, 1985).
Perlakuan pada penelitian ini menggunakan jumlah daging dan
lemak yang berbeda-beda. Dari hasil penelitian dapat dilihat bahwa nilai aw
urutan cenderung mengalami penurunan seiring dengan bertambahnya
lemak dan berkurangnya jumlah daging yang ditambahkan pada bahan. Aw
urutan mengalami penurunan dari 0,93 (penambahan lemak 5%) menjadi
0,89 (penambahan lemak 25%) (Gambar 3). Lemak jenuh yang terkandung
pada lemak babi akan pecah menjadi asam-asam lemak jenuh pada saat
fermentasi. Asam-asam lemak jenuh cenderung memiliki titik lebur yang
relatife tinggi sehingga aw yang terkandung pada lemak lebih tinggi dari pada
aw pada daging. Selain dipengaruhi oleh lemak, aw pada urutan juga
dipengaruhi oleh daging. Pada saat fermentasi terjadi denaturasi protein
sehingga air bebas pada molekul protein jumlahnya akan menurun. Selain itu
penurunan aw juga dipengaruhi oleh RH lingkungan yang rendah sehingga
terjadi penguapan air (Fenema, 1985).

16
VI. KESIMPULAN DAN SARAN
Kesimpulan
Dari hasil penelitian dan pembahasan dapat disimpulkan bahwa:
1. Polimer karbohidrat dapat digunakan sebagai bahan proteksi P. Acidilactici
U318 dengan viabilitas lebih besar dari 109 cfu/g. Penggunaan polimer
pati (maizena) merupakan polimer yang lebih baik digunakan untuk
pengawetan BAL dibandingkan dengan pektin dan karagenan.
2. Bubuk inokulum dengan tepung maizena sebagai bahan proteksi dapat
tumbuh dengan baik pada media urutan, namun semakin tinggi lemak
yang terkandung dalam formulasi media urutan pertumbuhan BAL
semakin rendah.
Saran
Kendala penggunaan pektin maupun karagenan sebagai bahan proteksi pada
proses produksi bubuk inokulum adalah pada saat pengeringan, yaitu tekstur yang
lengket dan tekstur menjadi keras setelah proses pengeringan. Untuk itu
disarankan untuk dilakukan percobaan lanjutan mengenai penggunaan campuran
maizena dan pektin maupun karagenan sebagai bahan proteksi BAL.

17
DAFTAR PUSTAKA
Antara, N.S. 2000. Purification using simple method adsorption-desorption and partial sequencing of bacteriocin produced by Pediococcus acidilacciti 3110. Scientific Meeting of Indonesia Society for Microbiology, PIT PERMI, June 27-28, 2000. Denpasar.
Antara, N.S., Sujaya,, I N., Yokota, A., Asano, K., Aryanta, W.R. and Tomita, F. 2002. Identification and succession of lactic acid bacteria during fermentation of urutan, a Balinese indigenous fermented sausage. World J. Microbiol. Bitechnol. 18: 255-262.
Antara, N.S., Sujaya,, I N., Yokota, A., Asano, K., Aryanta, W.R. and Tomita, F. 2002. The effects of indigenous lactic acid bacteria as single starter culture on the quality of “urutan”. Annual Meeting of Japan Society for Lactic Acid Bacteria. August 23-25, 2002. Tokyo. Japan.
Antara, N.S. 2004. Isolation and Identification of Indigenous Lactic Acid Bacteria, Their role and Application in Production of Urutan, a Balinese Fermented Sausage. A Thesis Submit for The Requirement of Ph.D. Degree in Agriculture Garaduate School of Agriculture, Hokkaido University.
Antara, N.S., Sujaya,, I N., Yokota, A., Asano, K., Aryanta, W.R. and Tomita, F. 2004. Effects of indigenous starter culture on the microbial and physico-chemical characteristic of urutan, a Balinese fermented sausage. J. Biosci. Bioeng. 98(2): 92-98.
Antara, N.S., Sujaya, I N., dan Ramona, Y. 2007. Pengembangan Kultur starter bakteri Asam Laktat dan Metode Produksi Untuk Meningkatkan Mutu Urutan (Sosis Bali Terfermentasi). Laporan Penelitian Hibah Bersaing. Fakultas Teknologi Pertanian, Unud.
Aryanta, W.R. 1996. Characteristic of Balinese traditional fermented sausage. J. Ilmu dan Teknologi Pangan. 1: 74-77.
Bacus, J.N. 1984. Utilization of Microorganism in Meat Processing. A Handbook for Meat Plant Operator. John Wiley and Son Inc. New York.
Buckle, K.A., R.A. Edwards, G.H Fleet dan M. Wooton. 1987. Ilmu Pangan. Penerjemah Hari Purnomo dan Adiono. Universitas Indonesia. Press Jakarta.
Bukenhuskes, H.J. 1993. Selection criteria for lactic acid bacteria to be used as starter cultures for various food commodities. FEMS Microbiol. Rev. 12: 253-272.
Caplice, A. and Fitzgerald, G.F. 1999. Food fermentation: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 50: 131-149.
Castano, A., Garcia Fonta, M.C., Fresno, J.M., Tornadijo, M.E. and Carballo, J. 2002. Survival of Enterobacteriaceae during processing of chorizo de cebolla, Spanish fermented sausage. Food Contrl. 13: 107-115.
Coppola, S., Mauriello, G., Aponte, M., Moschetti, G. and Villani, F. 2000. Microbial succession during ripening of Napples-type salami, a Shoutern Italian fermented sausage. Meat Sci. 56: 321-329.
Erkkila, S. and Petaja, E. 2000. Screening of commercial meat starter cultures at low pH and in the presence of bile salts for potential probiotic use. Meat Sci. 55: 297-300.

18
Erkkila, S., Petaja, E., Eerola, S., Lilleberg, L., Mattila-Sandholm, T. and Suihko, M.L. 2001. Flavour profiles of dry sausages fermented by selected novel meat starter cultures. Meat Sci. 58: 111-116.
Fitzsimons, N.A., Cogan, T.M., Condon, S. and Beresford, T. 1999 Phenotypic and genotypic characterization of non-starter lactic acid bacteria in mature cheddar cheese. Appl. and Environ. Microbiol. 65: 3418-3426.
Gariga, M., Hugas, M., Gou, P., Aymerich, M.T., Arnau, J. and Monfort, J.M. 1996. Technological and sensorial evaluation of Lactobacillus strains as starter cultures in fermented sausages. Int. J. Food Microbiol., 32: 173-183.
Hammes, W.P. and Hertel, C. 1998 New development in meat starter cultures. Meat Sci. 49 (Suppl. 1): S125-S138.
Hansen, E.B. 2002. Commercial bacterial starter cultures for fermented foods of the future. Int. J. Food Microbiol. 78:119-131.
Holzapfel, W.H. 2002. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 75: 197-212.
Hugas, M. and Monfort, J.M. 1997. Bacterial starter cultures for meat fermentation. Food Chemist. 59: 547-554.
Jay, J.M. 1996. Modern Food Microbiology, 5th Edition. Chapman and Hall. New York.
Kalac, P., Spicka, J., Krizek, M. and Pelikanova, T. 2000 The effects of lactic acid bacteria inoculants on biogenic amines formation in sauerkraut. Food Chemist.70: 355-359.
Lucke, F.K. 1998. Fermented sausages. In: B.J.B. Wood (Ed.). Microbiology of Fermented Food – Volume 2. 2nd Edition. pp. 441-483. Blakie Academic and Professional. London.
Ordonez, J.A., Hiero, E.M., Bruna, J.M. and de la Hoz, L. 1999. Changes in the components of dry-fermented sausages during ripening. Crit. Rev. Food Sci. Nutr. 39: 329-367.
Papadima, S.N. and Bloukas, J.G. 1999. Effect of fat level and storage conditions on quality characteristics of traditional Greek sausages. Meat Sci. 51: 103-113.
Parente, E., Rota, M.A., Ricciardi, A. and Clementi, F. 1997 Characterization of natural starter cultures used in the manufacture of Pasta Filata cheese in Basilicata (Southern Italy). Int. Dairy J. 7: 775-783.
Ray, B. 1992. The Need For food Biopreservation. In B. Ray and M. Daeschel (Eds.). Food Biopreservative of Microbial Origin, pp. 1-23. CRC Press. Bocaraton, Florida.
Sanchez, I., Palop, L. and Ballesteros, C. 2000 Biochemical characterization of lactic acid bacteria isolated from spontaneous fermentation of almagro eggplants. Int. J. Food Microbiol. 59: 355-359.
Stahnke, L.H. 1995. Dried sausages fermented with Staphylocpccus xylosus at different temperatures and with different ingredient levels – Part II. Volatile components. Meat Sci. 41: 193-209.
Wood, B.J.B. 1997. Microbiology of Fermented Foods. Balcky Academic and Professional, London.

19
LAMPIRAN
Top Related
Copyright © 2022 FDOKUMEN

