







Bahasa
Halaman
Hukum


Interactions of intrathecally administered ziconotide, a selective blocker ofneuronal N-type voltage-sensitive calcium channels, with morphine on
nociception in rats
Yong-Xiang Wang*, Da Gao, Mark Pettus, Cora Phillips, S. Scott Bowersox
Department of Pharmacology, Elan Pharmaceuticals, 3760 Haven Avenue, Menlo Park, CA 94025, USA
Received 13 February 1999; received in revised form 16 June 1999; accepted 13 August 1999
Abstract
Ziconotide is a selective, potent and reversible blocker of neuronal N-type voltage-sensitive calcium channels (VSCCs). Morphine is an
agonist of m-opioid receptors and inhibits N-type VSCC channels via a G-protein coupling mechanism. Both agents are antinociceptive when
they are administered intrathecally (spinally). The present study investigated the acute and chronic (7-day) interactions of intrathecally
administered ziconotide and morphine on nociception in several animal models of pain. In the acute study, intrathecal bolus injections of
morphine and ziconotide alone produced dose-dependent inhibition of formalin-induced tonic ¯inch responses and withdrawal responses to
paw pressure. The combination of ziconotide and morphine produced an additive inhibition of formalin-induced tonic ¯inch responses and a
signi®cant leftward shift of the morphine dose-response curve in the paw pressure test. After chronic (7-day) intrathecal infusion, ziconotide
enhanced morphine analgesia in the formalin test. In contrast, chronic intrathecal morphine infusion produced tolerance to analgesia, but did
not affect ziconotide antinociception. Antinociception produced by ziconotide alone was the same as that observed when the compound was
co-administered with morphine to morphine-tolerant rats. In the hot-plate and tail immersion tests, chronic intrathecal infusion of morphine
lead to rapid tolerance whereas ziconotide produced sustained analgesia with no loss of potency throughout the infusion period. Although
ziconotide in combination with morphine produced an apparent synergistic analgesic effects during the initial phase of continuous infusion, it
did not prevent morphine tolerance to analgesia. These results demonstrate that (1) acute intrathecal administrations of ziconotide and
morphine produce additive or synergistic analgesic effects; (2) chronic intrathecal morphine infusion results in tolerance to analgesia but does
not produce cross-tolerance to ziconotide; (3) chronic intrathecal ziconotide administration produces neither tolerance nor cross-tolerance to
morphine analgesia; (4) intrathecal ziconotide does not prevent or reverse morphine tolerance. q 2000 International Association for the
Study of Pain. Published by Elsevier Science B.V.
Keywords: Ziconotide; Voltage-sensitivity calcium channels (VSCCs); Blocker; Morphine; m-Opioid receptors; Interaction; Addition; Synergistism; Noci-
ception; Tolerance; Spinal cord; Intrathecal route
1. Introduction
Several sub-types of voltage-sensitive calcium channels
(VSCCs) (e.g. L-, N-, T-, P, Q-, and O-type) have been
identi®ed in the nervous system and other tissues (see
reviews by Olivera et al., 1994; Miljanich and Ramachan-
dran, 1995). De®ned as high af®nity binding sites for v -
conotoxin GVIA (Olivera et al., 1987), N-type calcium
channels are located in presynaptic nerve terminals where
they permit the calcium in¯ux necessary for transmitter
release from both central and peripheral nerves (see review
by Miljanich and Ramachandran, 1995). Ziconotide (SNX-
111), a 25-amino acid polypeptide, is the synthetic version
of the naturally occurring v -conotoxin MVIIA (see review
by Miljanich and Ramachandran, 1995). Ziconotide selec-
tively and reversibly binds to v -conotoxin GVIA binding
sites (N-type calcium channels) with an extremely high
potency (Kd < 8 pM) (Kristipati et al., 1994; Wang et al.,
1998), blocks calcium currents mediated by N-type VSCCs
in IMR-32 human neuroblastoma cells and other prepara-
tions (see Wang et al., 1998), and inhibits transmitter release
from peripheral and central neurons (Gaur et al., 1994;
Wang et al., 1998). Ziconotide or v -conotoxin MVIIA is
antinociceptive in animal models of acute, persistent and
Pain 84 (2000) 271±281
0304-3959/00/$20.00 q 2000 International Association for the Study of Pain. Published by Elsevier Science B.V.
PII: S0304-3959(99)00214-6
www.elsevier.nl/locate/pain
* Corresponding author. Tel.: 11-650-833-1231; fax: 11-650-614-
1002.
E-mail address: [email protected] (Y.-X. Wang)

neuropathic pain when it is administered intrathecally
(Chaplan et al., 1994; Malmberg and Yaksh, 1994, 1995;
Bowersox et al., 1996; Yamamoto and Sakashita, 1998), or
by other routes (Xiao and Bennett, 1995; White and
Cousins, 1998). Ziconotide is currently under clinical inves-
tigation for the treatment of acute and chronic pain
syndromes via an intrathecal route (Brose et al., 1997; Pres-
ley et al., 1998).
Morphine is a conventionally prescribed narcotic and a
spinal analgesic agent (Abram and Yaksh, 1993). It binds
mainly to m-opioid receptors coupled to GTP-binding
proteins (G-proteins) that initiate a variety of downstream
cellular effects, including reductions in intracellular cAMP
levels, increased potassium channel activity and decreased
calcium channel activity (Bourinet et al., 1996; see review
by Basbaum, 1996). These activities hyperpolarize neurons,
which results in inhibition of spike ®ring and decreased
neurotransmitter release (see review by Basbaum, 1996).
Recently, m-opioid receptors have been shown to be
coupled to the pore-forming a 1 subunits of N-type VSCCs
via G-proteins. Activation of m-opioid receptors leads to
inhibition of N-type calcium currents (Seward et al., 1991;
Bourinet et al., 1996; Toth et al., 1996; Zhang et al., 1996;
De Waard et al., 1997). Therefore, inhibition of N-type
calcium currents may be one of the mechanisms by which
morphine produces analgesia. Repeated administration
(including intrathecal administration) of morphine can
result in tolerance to analgesia, but the mechanism is not
well understood. It has been proposed that tolerance is due
to the uncoupling of opioid receptors and G-proteins
(Kennedy and Henderson, 1991, 1992; Tao et al., 1993;
see reviews by Basbaum, 1995, 1996). G-protein-coupled
effectors, such as calcium and potassium channel activities,
and cAMP levels are not altered in morphine tolerance
status (see reviews by Basbaum, 1995, 1996). However,
an increase in v -conotoxin GVIA binding sites (presumably
N-type VSCCs) has been observed in mouse brains after
chronic morphine treatment (Suematsu et al., 1993). In
another study, v -conotoxin GVIA partially inhibited KCl-
induced calcium in¯ux in synaptosomes prepared from
naive rats but not from morphine-tolerant rats (Welch and
Olson, 1991).
As both ziconotide and morphine inhibit N-type VSCCs
and as both agents may be given intrathecally to produce
analgesia at the same or different times, it is important to
understand possible interactions between morphine and
ziconotide with respect to nociception and tolerance to
analgesia. The speci®c aims of the present study were to
determine whether (1) acute intrathecal administration of
ziconotide and morphine affect each other's antinociceptive
properties, (2) chronic (7-day) intrathecal morphine or zico-
notide infusion alter ziconotide or morphine analgesia,
respectively, and (3) intrathecal ziconotide treatment
prevents or reverses morphine tolerance to analgesia. Preli-
minary ®ndings have been published previously in abstract
form (Wang et al., 1997).
2. Materials and methods
2.1. Animals
Male, Sprague±Dawley rats weighing between 220 and
280 g (Simonson Laboratories, Gilroy, CA) were acclimated
to the laboratory environment for 5±7 days before entering
the study. While in the home cage environment, the animals
were allowed free access to water and a commercial rat diet.
Room temperature was maintained at 20±238C and room
illumination was on a 12/12 h light/dark cycle (07:00/
19:00). All experiments were carried out with the approval
of the Animal Care and Use Committee of Elan Pharmaceu-
ticals.
2.2. Drugs
Ziconotide tri¯uoroacetic acid (Elan Pharmaceuticals,
Menlo Park, CA) and morphine sulfate pentahydrate
(Sigma Chemical Co., St. Louis, MO) were dissolved and
diluted in sterile, preservative free 0.9% NaCl solution
(Phoenix Scienti®c, Inc., St. Joseph, MO). For the acute
study with the formalin test, three test article solutions
were prepared: ziconotide alone, morphine alone, and the
combination of ziconotide and morphine in a ratio of 1:10.
All three test article solutions were then diluted in a ratio of
1:3 or 1:3.33.
2.3. Spinal catheterization procedures
The animals were implanted with polyethylene catheters
(PE-10: 0.23 mm i.d. and 0.61 mm o.d., and with volume of
approximately 7 ml (Clay Adams, Parsippany, NJ)) ®lled
with heparinized (25 IU/ml) saline under halothane anesthe-
sia as described by Yaksh and Rudy (1976). The catheter
measured 8.5 cm in length and terminated at the lumbar
enlargement of the spinal cord. The external end of the
catheter was anchored to the adjacent muscle tissue where
it emerged from the cisterna magna with one stay suture. In
single treatment studies including the bolus injection of test
articles in the paw pressure test and continuous infusion of
test articles in the hot-plate and tail immersion tests (see
Experiment 3.1 and 3.3), compounds were administered
approximately 7 days after catheterization. In the second
experiment of 3.2 where chronic morphine infusion
followed by bolus injection of ziconotide, the catheter was
connected to an osmotic minipump (Alzet, Model 2001,
Alza Co., Palo Alto, CA). The minipump containing test
article solution was implanted subcutaneously between the
scapulae. Seven days after implantation, the minipump was
removed under halothane anesthesia, the catheter was
¯ushed with saline, and used for bolus injection of zicono-
tide 3 h later. In the ®rst experiment of 3.2 where chronic
intrathecal infusion of ziconotide followed by bolus injec-
tion of morphine, a `PE-5' catheter (0.006 `i.d. and 0.14'
o.d., Spectranetics, Colorado Springs, CO) connected with
PE-10 tubing was implanted, together with another PE-10
Y.-X. Wang et al. / Pain 84 (2000) 271±281272

tubing, into the spinal cord as described above for intrathe-
cal bolus injection.
2.4. Formalin test
Saline containing 5% formalin was injected subcuta-
neously into the right dorsal hindpaw. The rat was immedi-
ately placed in a 23 £ 35 £ 19 cm polycarbonate box
positioned in front of a mirror for behavioral observations.
Nociceptive behavior was quanti®ed by counting the
number of paw ¯exions in 1-min epochs. Measurements
were taken at 10-min intervals beginning immediately
after formalin injection and ending 90 min later.
2.5. Paw pressure test
The rat was placed in a cone restrainer that allowed its
feet to hang free. The mechanical nociceptive threshold for
the ¯exion re¯ex elicited by stimulation of the dorsal
surface of the right hindpaw was quanti®ed using an Ugo
Basile analgesymeter (Biological Research Apparatus,
Varese, Italy). This device generates a mechanical force,
with a dome-shaped plunger placing directly on the dorsum
of the rat's hindpaw, that increases linearly with time. The
mechanical nociceptive threshold was de®ned as the force
(in g) at which the rat withdrew its paw. The cut-off force
was 750 g.
2.6. Hot-plate test
The animal was placed on a hot-plate (Hot Plate Analge-
sia Meter, Model 39D, IITC, Inc., Woodland, CA) main-
tained at 52.58C (52.2±52.88C). The thermal nociceptive
threshold was de®ned as the time required to elicit either
a hindpaw lick or a jump. The cut-off time was 60 s.
2.7. Tail immersion test
The animal was placed in a cone restrainer and the end of
the tail (5 cm) was placed in a 508C water bath (49.5±
50.58C). The thermal nociceptive threshold was de®ned as
the time required to elicit a ¯ick of the tail. The cut-off time
was 30 s. As the hot-plate and tail immersion tests were
performed in the same session, the tail immersion test was
followed by the hot-plate test.
2.8. Data analysis
Nociceptive thresholds were expressed as means ^ SEM.
For the hot-plate and tail immersion tests, data were
converted to percent maximum possible effect (% MPE)
by the following formula: % MPE � (post-treatment
value 2 pre-treatment value)/( cut-off value 2 pre-treat-
ment value) £ 100. Antinociceptive ED50 values were calcu-
lated from dose-response curves of morphine, ziconotide, or
the combination of morphine and ziconotide by a program
(Drug Interaction Toolbox: Version 1.32, Tallarida and
McCary, 1998) based on the Graded Dose Response Method
of Tallarida and Murray (1987). The program also calcu-
lated the regression lines for individual dose-response
curves, theoretic additive curves, and combination (mixture)
curves according to the methods described by Tallarida et
al. (1997). Interactions between morphine and ziconotide
were analyzed by comparing the composite additive line
vs. the mixture regression line (F-test) and the theoretical
additive ED50 vs. the mixture ED50 (t-test). As this drug
interaction analysis approach is relatively new, the drug
interaction was also analyzed by the conventional isobolo-
graphical analysis (Tallarida et al., 1989). Statistical signif-
icance was also determined by repeated measures analysis
of variance (ANOVA) followed by post-hoc two-tailed t-
tests, and one-factor ANOVA via using a statistical analysis
program (StatView, Abacus Concepts, Berkeley, CA). P #0:05 was considered statistically signi®cant.
3. Results
3.1. Acute interaction of ziconotide and morphine in the
formalin test
To study the acute interactions of ziconotide and
morphine, dose-response curves for ziconotide, morphine,
and the combination of ziconotide and morphine were
measured. The ®rst ®ve groups of rats (n � 8 in each
group) received intrathecal bolus injections of 10 ml saline
or ziconotide (0.03, 0.1, 0.3, or 1 mg). The second six groups
of rats (n � 8 in each group) received intrathecal bolus
injections of 10 ml saline or morphine (0.1, 0.3, 1, 3 or 10
mg). The last ®ve groups of rats (n � 8 in each group)
received intrathecal bolus injections of 10 ml saline or
four doses of the combination of ziconotide and morphine
in a ®xed dose ratio of 1:10. The dose combinations were as
follows: 0.01 mg ziconotide 1 0:1 mg morphine, 0.03 mg
ziconotide 1 0:3 mg morphine, 0.1 mg ziconotide 1 1 mg
morphine, and 0.3 mg ziconotide 1 3 mg morphine. All
rats were treated 10 min before initiating the formalin test.
Subcutaneous injection of formalin in control rats receiv-
ing intrathecal saline produced a characteristic bi-phasic
¯inch response consisting of an initial, rapidly decaying
acute phase (within 10 min after formalin injection)
followed by a slowly rising and long-lived (10±90 min)
tonic phase. Intrathecal ziconotide up to 1 mg did not inhibit
¯inch responses in the acute phase, but produced a dose-
dependent inhibition of formalin-induced ¯inch responses in
the tonic phase (Fig. 1A). The areas under the ¯inch
response curve from 10 to 90 min (AUC10±90s) were calcu-
lated and plotted in Fig. 2A. The ED50 for ziconotide analge-
sia was 0:11 ^ 0:5 mg. Intrathecal bolus injection of
morphine in doses up to 10 mg did not signi®cantly inhibit
formalin-induced ¯inch responses in the acute phase, but
blocked the tonic phase responses in a dose-dependent
manner (Fig. 1B). The AUC10±90s were calculated and
plotted in Fig. 2A. The ED50 for morphine analgesia was
Y.-X. Wang et al. / Pain 84 (2000) 271±281 273
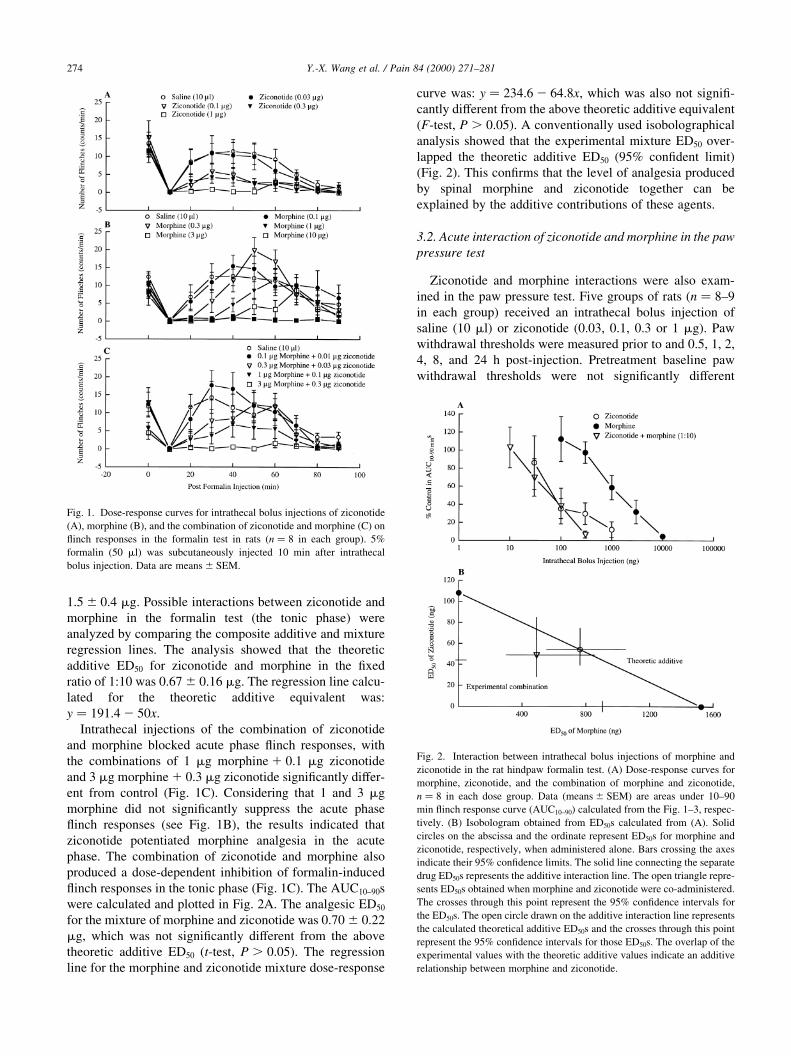
1:5 ^ 0:4 mg. Possible interactions between ziconotide and
morphine in the formalin test (the tonic phase) were
analyzed by comparing the composite additive and mixture
regression lines. The analysis showed that the theoretic
additive ED50 for ziconotide and morphine in the ®xed
ratio of 1:10 was 0:67 ^ 0:16 mg. The regression line calcu-
lated for the theoretic additive equivalent was:
y � 191:4 2 50x.
Intrathecal injections of the combination of ziconotide
and morphine blocked acute phase ¯inch responses, with
the combinations of 1 mg morphine 1 0:1 mg ziconotide
and 3 mg morphine 1 0:3 mg ziconotide signi®cantly differ-
ent from control (Fig. 1C). Considering that 1 and 3 mg
morphine did not signi®cantly suppress the acute phase
¯inch responses (see Fig. 1B), the results indicated that
ziconotide potentiated morphine analgesia in the acute
phase. The combination of ziconotide and morphine also
produced a dose-dependent inhibition of formalin-induced
¯inch responses in the tonic phase (Fig. 1C). The AUC10±90s
were calculated and plotted in Fig. 2A. The analgesic ED50
for the mixture of morphine and ziconotide was 0:70 ^ 0:22
mg, which was not signi®cantly different from the above
theoretic additive ED50 (t-test, P . 0:05). The regression
line for the morphine and ziconotide mixture dose-response
curve was: y � 234:6 2 64:8x, which was also not signi®-
cantly different from the above theoretic additive equivalent
(F-test, P . 0:05). A conventionally used isobolographical
analysis showed that the experimental mixture ED50 over-
lapped the theoretic additive ED50 (95% con®dent limit)
(Fig. 2). This con®rms that the level of analgesia produced
by spinal morphine and ziconotide together can be
explained by the additive contributions of these agents.
3.2. Acute interaction of ziconotide and morphine in the paw
pressure test
Ziconotide and morphine interactions were also exam-
ined in the paw pressure test. Five groups of rats (n � 8±9
in each group) received an intrathecal bolus injection of
saline (10 ml) or ziconotide (0.03, 0.1, 0.3 or 1 mg). Paw
withdrawal thresholds were measured prior to and 0.5, 1, 2,
4, 8, and 24 h post-injection. Pretreatment baseline paw
withdrawal thresholds were not signi®cantly different
Y.-X. Wang et al. / Pain 84 (2000) 271±281274
Fig. 2. Interaction between intrathecal bolus injections of morphine and
ziconotide in the rat hindpaw formalin test. (A) Dose-response curves for
morphine, ziconotide, and the combination of morphine and ziconotide,
n � 8 in each dose group. Data (means ^ SEM) are areas under 10±90
min ¯inch response curve (AUC10±90) calculated from the Fig. 1±3, respec-
tively. (B) Isobologram obtained from ED50s calculated from (A). Solid
circles on the abscissa and the ordinate represent ED50s for morphine and
ziconotide, respectively, when administered alone. Bars crossing the axes
indicate their 95% con®dence limits. The solid line connecting the separate
drug ED50s represents the additive interaction line. The open triangle repre-
sents ED50s obtained when morphine and ziconotide were co-administered.
The crosses through this point represent the 95% con®dence intervals for
the ED50s. The open circle drawn on the additive interaction line represents
the calculated theoretical additive ED50s and the crosses through this point
represent the 95% con®dence intervals for those ED50s. The overlap of the
experimental values with the theoretic additive values indicate an additive
relationship between morphine and ziconotide.
Fig. 1. Dose-response curves for intrathecal bolus injections of ziconotide
(A), morphine (B), and the combination of ziconotide and morphine (C) on
¯inch responses in the formalin test in rats (n � 8 in each group). 5%
formalin (50 ml) was subcutaneously injected 10 min after intrathecal
bolus injection. Data are means ^ SEM.

from among the ®ve groups of rats. Saline did not signi®-
cantly change paw withdrawal thresholds. Ziconotide
caused a long-lived and reversible inhibition of pressure-
induced paw withdrawal responses; the peak effect was
observed 1 h post-injection and was maintained for at
least 8 h before returning to pre-treatment levels within 24
h (Fig. 3A). The antinociceptive effect of ziconotide was
dose-dependent with an ED50 of 0.6 mg (95% con®dence
interval: 0.3±1.3 mg). Fig. 3B shows the dose-response
curve when the paw withdrawal thresholds 1 h post injection
were plotted.
Six groups of rats (n � 8 in each group except for 20 mg
morphine group where n � 4) received an intrathecal bolus
injection of saline (10 ml), morphine (0.5, 2 and 20 mg), and
the combinations of ziconotide (0.1 mg) and morphine (0.5
and 20 mg). Paw withdrawal thresholds were measured prior
to and 0.25, 0.5, 1, 2, and 4 h post-injection. Pretreatment
baseline paw withdrawal thresholds were not signi®cantly
different among the six groups of rats. Morphine increased
the mechanical thresholds in a time-dependent manner.
Ziconotide potentiated the antinociceptive effects of
morphine at both 0.5 and 2 mg (Fig. 4A). Fig. 4B shows
the dose-response curves for morphine in the absence or
presence of ziconotide when mechanical nociceptive thresh-
olds 1 h post injection were plotted. Morphine produced
dose-dependent analgesia. Ziconotide signi®cantly shifted
the analgesic dose-response curve for morphine to the left.
The ED50s for morphine analgesia in the absence and
presence of ziconotide (0.1 mg, which by itself did not
signi®cantly increased mechanical threshold, see Fig. 3)
were 1.3 (95% con®dence interval: 0.5±3.9 mg) and 0.2
mg (95% con®dence interval: 0.02±0.3 mg), respectively,
and were signi®cantly different from each other (P , 0:05).
3.3. Effects of chronic intrathecal ziconotide infusion on
morphine analgesia in the formalin test
To test whether chronic (7-day) intrathecal ziconotide
affected morphine analgesia in the formalin test, 44 rats
were in four groups (n � 10±12 in each group) and each
group received two treatments. They were (1) saline 1saline; (2) ziconotide 1 saline; (3) saline 1 morphine; and
(4) ziconotide 1 morphine. The ®rst treatment was contin-
uous, constant-rate, intrathecal infusion of saline (1 ml/h) or
ziconotide (0.1 mg/h) for 7 days; the second treatment was
intrathecal bolus injection of saline (10 ml) or morphine (1
mg) using different sets of intrathecal tubings after 7-days of
Y.-X. Wang et al. / Pain 84 (2000) 271±281 275
Fig. 3. Time-course (A) and dose-response curve (B) of the antinociceptive
effect of ziconotide (0.03±1 mg) administered intrathecally in the paw
pressure test in rats (n � 8±9 in each group). Data are means ^ SEM.
Data in B were pressure nociceptive thresholds at 1 h after intrathecal
bolus injection from A. ED50 of ziconotide analgesia was 0.6 mg (95%
con®dence interval: 0.3±1.3 mg).
Fig. 4. Time-course (A) and dose-response curve (B) of the antinociceptive
effect of morphine (0.5±20 mg) administered intrathecally, with or without
ziconotide, (0.1 mg) in the paw pressure test in rats (n � 8±9 in each group
except for 20 mg morphine where n � 4). Data are means ^ SEM. Data in
B were pressure nociceptive thresholds at 1 h after intrathecal bolus injec-
tion from A. ED50s of morphine analgesia in the absence and presence of
ziconotide were 1.3 mg (95% con®dence interval: 0.5±3.9 mg) and 0.2 mg
(95% con®dence interval: 0.02±0.3 mg), respectively.
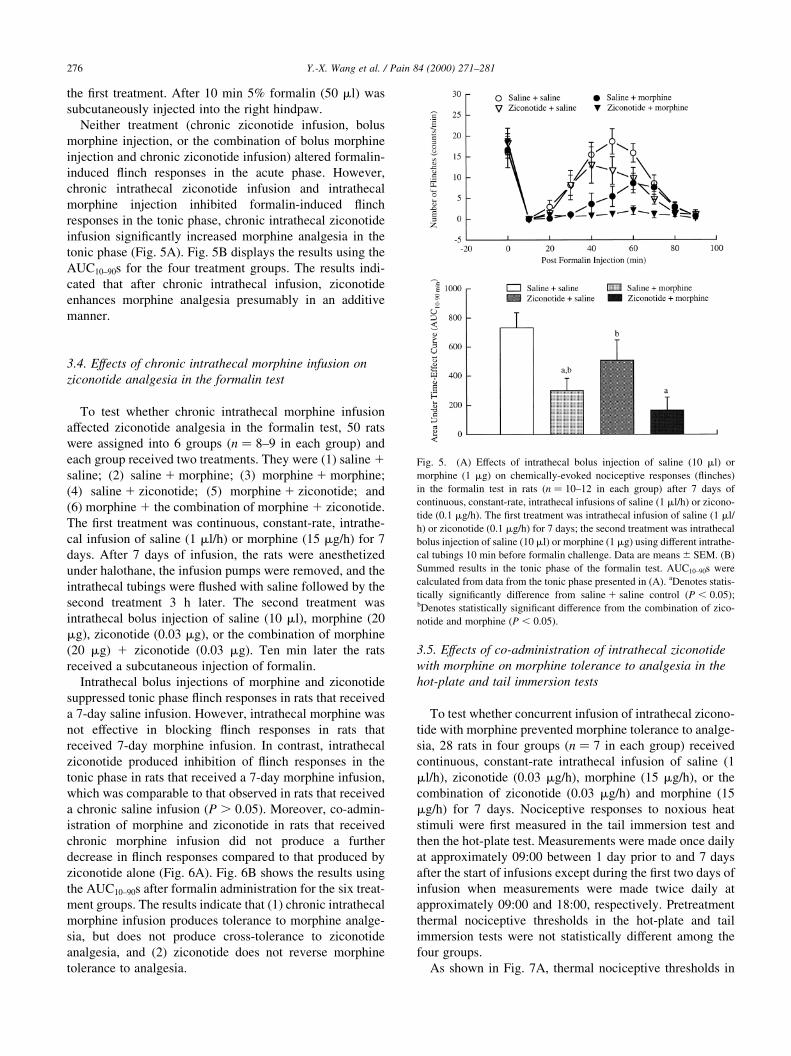
the ®rst treatment. After 10 min 5% formalin (50 ml) was
subcutaneously injected into the right hindpaw.
Neither treatment (chronic ziconotide infusion, bolus
morphine injection, or the combination of bolus morphine
injection and chronic ziconotide infusion) altered formalin-
induced ¯inch responses in the acute phase. However,
chronic intrathecal ziconotide infusion and intrathecal
morphine injection inhibited formalin-induced ¯inch
responses in the tonic phase, chronic intrathecal ziconotide
infusion signi®cantly increased morphine analgesia in the
tonic phase (Fig. 5A). Fig. 5B displays the results using the
AUC10±90s for the four treatment groups. The results indi-
cated that after chronic intrathecal infusion, ziconotide
enhances morphine analgesia presumably in an additive
manner.
3.4. Effects of chronic intrathecal morphine infusion on
ziconotide analgesia in the formalin test
To test whether chronic intrathecal morphine infusion
affected ziconotide analgesia in the formalin test, 50 rats
were assigned into 6 groups (n � 8±9 in each group) and
each group received two treatments. They were (1) saline 1saline; (2) saline 1 morphine; (3) morphine 1 morphine;
(4) saline 1 ziconotide; (5) morphine 1 ziconotide; and
(6) morphine 1 the combination of morphine 1 ziconotide.
The ®rst treatment was continuous, constant-rate, intrathe-
cal infusion of saline (1 ml/h) or morphine (15 mg/h) for 7
days. After 7 days of infusion, the rats were anesthetized
under halothane, the infusion pumps were removed, and the
intrathecal tubings were ¯ushed with saline followed by the
second treatment 3 h later. The second treatment was
intrathecal bolus injection of saline (10 ml), morphine (20
mg), ziconotide (0.03 mg), or the combination of morphine
(20 mg) 1 ziconotide (0.03 mg). Ten min later the rats
received a subcutaneous injection of formalin.
Intrathecal bolus injections of morphine and ziconotide
suppressed tonic phase ¯inch responses in rats that received
a 7-day saline infusion. However, intrathecal morphine was
not effective in blocking ¯inch responses in rats that
received 7-day morphine infusion. In contrast, intrathecal
ziconotide produced inhibition of ¯inch responses in the
tonic phase in rats that received a 7-day morphine infusion,
which was comparable to that observed in rats that received
a chronic saline infusion (P . 0:05). Moreover, co-admin-
istration of morphine and ziconotide in rats that received
chronic morphine infusion did not produce a further
decrease in ¯inch responses compared to that produced by
ziconotide alone (Fig. 6A). Fig. 6B shows the results using
the AUC10±90s after formalin administration for the six treat-
ment groups. The results indicate that (1) chronic intrathecal
morphine infusion produces tolerance to morphine analge-
sia, but does not produce cross-tolerance to ziconotide
analgesia, and (2) ziconotide does not reverse morphine
tolerance to analgesia.
3.5. Effects of co-administration of intrathecal ziconotide
with morphine on morphine tolerance to analgesia in the
hot-plate and tail immersion tests
To test whether concurrent infusion of intrathecal zicono-
tide with morphine prevented morphine tolerance to analge-
sia, 28 rats in four groups (n � 7 in each group) received
continuous, constant-rate intrathecal infusion of saline (1
ml/h), ziconotide (0.03 mg/h), morphine (15 mg/h), or the
combination of ziconotide (0.03 mg/h) and morphine (15
mg/h) for 7 days. Nociceptive responses to noxious heat
stimuli were ®rst measured in the tail immersion test and
then the hot-plate test. Measurements were made once daily
at approximately 09:00 between 1 day prior to and 7 days
after the start of infusions except during the ®rst two days of
infusion when measurements were made twice daily at
approximately 09:00 and 18:00, respectively. Pretreatment
thermal nociceptive thresholds in the hot-plate and tail
immersion tests were not statistically different among the
four groups.
As shown in Fig. 7A, thermal nociceptive thresholds in
Y.-X. Wang et al. / Pain 84 (2000) 271±281276
Fig. 5. (A) Effects of intrathecal bolus injection of saline (10 ml) or
morphine (1 mg) on chemically-evoked nociceptive responses (¯inches)
in the formalin test in rats (n � 10±12 in each group) after 7 days of
continuous, constant-rate, intrathecal infusions of saline (1 ml/h) or zicono-
tide (0.1 mg/h). The ®rst treatment was intrathecal infusion of saline (1 ml/
h) or ziconotide (0.1 mg/h) for 7 days; the second treatment was intrathecal
bolus injection of saline (10 ml) or morphine (1 mg) using different intrathe-
cal tubings 10 min before formalin challenge. Data are means ^ SEM. (B)
Summed results in the tonic phase of the formalin test. AUC10±90s were
calculated from data from the tonic phase presented in (A). aDenotes statis-
tically signi®cantly difference from saline 1 saline control (P , 0:05);bDenotes statistically signi®cant difference from the combination of zico-
notide and morphine (P , 0:05).
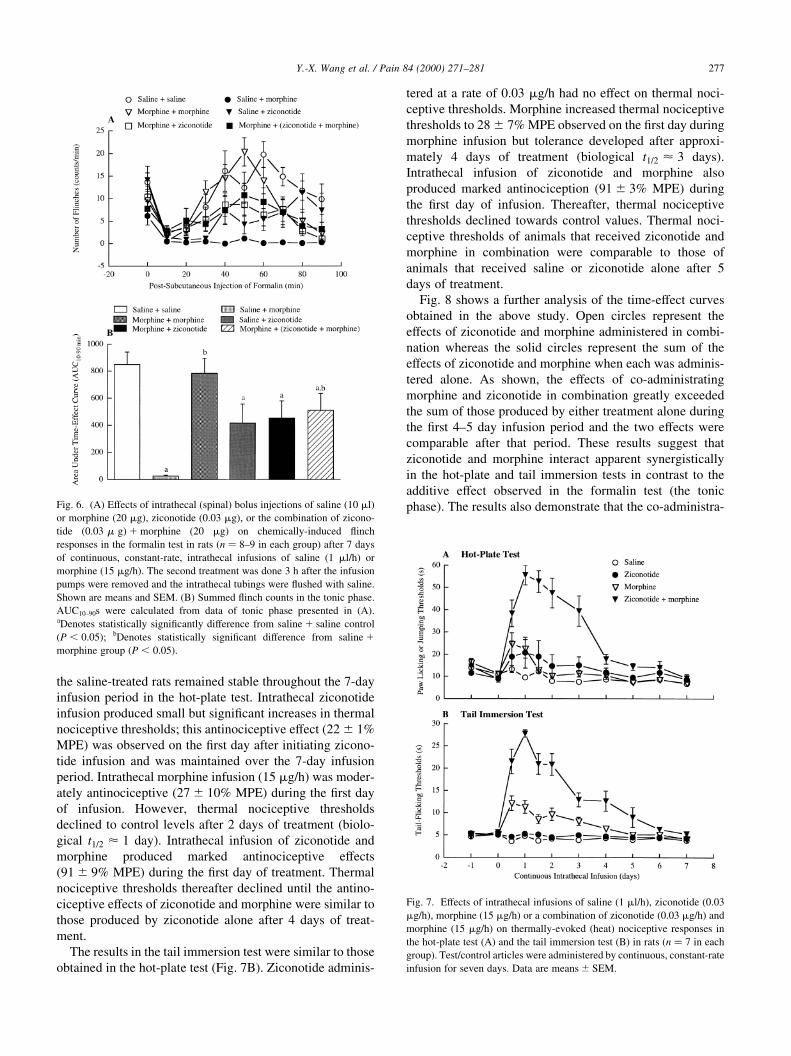
the saline-treated rats remained stable throughout the 7-day
infusion period in the hot-plate test. Intrathecal ziconotide
infusion produced small but signi®cant increases in thermal
nociceptive thresholds; this antinociceptive effect (22 ^ 1%
MPE) was observed on the ®rst day after initiating zicono-
tide infusion and was maintained over the 7-day infusion
period. Intrathecal morphine infusion (15 mg/h) was moder-
ately antinociceptive (27 ^ 10% MPE) during the ®rst day
of infusion. However, thermal nociceptive thresholds
declined to control levels after 2 days of treatment (biolo-
gical t1=2 < 1 day). Intrathecal infusion of ziconotide and
morphine produced marked antinociceptive effects
(91 ^ 9% MPE) during the ®rst day of treatment. Thermal
nociceptive thresholds thereafter declined until the antino-
ciceptive effects of ziconotide and morphine were similar to
those produced by ziconotide alone after 4 days of treat-
ment.
The results in the tail immersion test were similar to those
obtained in the hot-plate test (Fig. 7B). Ziconotide adminis-
tered at a rate of 0.03 mg/h had no effect on thermal noci-
ceptive thresholds. Morphine increased thermal nociceptive
thresholds to 28 ^ 7% MPE observed on the ®rst day during
morphine infusion but tolerance developed after approxi-
mately 4 days of treatment (biological t1=2 < 3 days).
Intrathecal infusion of ziconotide and morphine also
produced marked antinociception (91 ^ 3% MPE) during
the ®rst day of infusion. Thereafter, thermal nociceptive
thresholds declined towards control values. Thermal noci-
ceptive thresholds of animals that received ziconotide and
morphine in combination were comparable to those of
animals that received saline or ziconotide alone after 5
days of treatment.
Fig. 8 shows a further analysis of the time-effect curves
obtained in the above study. Open circles represent the
effects of ziconotide and morphine administered in combi-
nation whereas the solid circles represent the sum of the
effects of ziconotide and morphine when each was adminis-
tered alone. As shown, the effects of co-administrating
morphine and ziconotide in combination greatly exceeded
the sum of those produced by either treatment alone during
the ®rst 4±5 day infusion period and the two effects were
comparable after that period. These results suggest that
ziconotide and morphine interact apparent synergistically
in the hot-plate and tail immersion tests in contrast to the
additive effect observed in the formalin test (the tonic
phase). The results also demonstrate that the co-administra-
Y.-X. Wang et al. / Pain 84 (2000) 271±281 277
Fig. 6. (A) Effects of intrathecal (spinal) bolus injections of saline (10 ml)
or morphine (20 mg), ziconotide (0.03 mg), or the combination of zicono-
tide (0:03 m g�1 morphine (20 mg) on chemically-induced ¯inch
responses in the formalin test in rats (n � 8±9 in each group) after 7 days
of continuous, constant-rate, intrathecal infusions of saline (1 ml/h) or
morphine (15 mg/h). The second treatment was done 3 h after the infusion
pumps were removed and the intrathecal tubings were ¯ushed with saline.
Shown are means and SEM. (B) Summed ¯inch counts in the tonic phase.
AUC10±90s were calculated from data of tonic phase presented in (A).aDenotes statistically signi®cantly difference from saline 1 saline control
(P , 0:05); bDenotes statistically signi®cant difference from saline 1
morphine group (P , 0:05).
Fig. 7. Effects of intrathecal infusions of saline (1 ml/h), ziconotide (0.03
mg/h), morphine (15 mg/h) or a combination of ziconotide (0.03 mg/h) and
morphine (15 mg/h) on thermally-evoked (heat) nociceptive responses in
the hot-plate test (A) and the tail immersion test (B) in rats (n � 7 in each
group). Test/control articles were administered by continuous, constant-rate
infusion for seven days. Data are means ^ SEM.
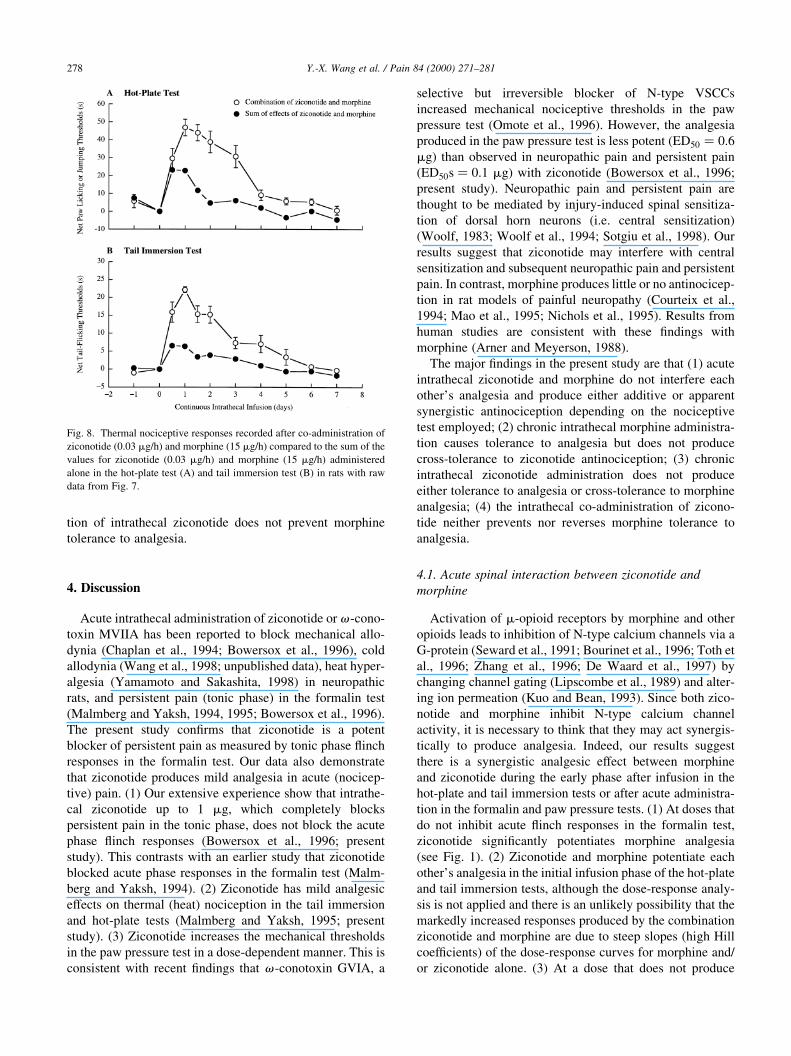
tion of intrathecal ziconotide does not prevent morphine
tolerance to analgesia.
4. Discussion
Acute intrathecal administration of ziconotide or v -cono-
toxin MVIIA has been reported to block mechanical allo-
dynia (Chaplan et al., 1994; Bowersox et al., 1996), cold
allodynia (Wang et al., 1998; unpublished data), heat hyper-
algesia (Yamamoto and Sakashita, 1998) in neuropathic
rats, and persistent pain (tonic phase) in the formalin test
(Malmberg and Yaksh, 1994, 1995; Bowersox et al., 1996).
The present study con®rms that ziconotide is a potent
blocker of persistent pain as measured by tonic phase ¯inch
responses in the formalin test. Our data also demonstrate
that ziconotide produces mild analgesia in acute (nocicep-
tive) pain. (1) Our extensive experience show that intrathe-
cal ziconotide up to 1 mg, which completely blocks
persistent pain in the tonic phase, does not block the acute
phase ¯inch responses (Bowersox et al., 1996; present
study). This contrasts with an earlier study that ziconotide
blocked acute phase responses in the formalin test (Malm-
berg and Yaksh, 1994). (2) Ziconotide has mild analgesic
effects on thermal (heat) nociception in the tail immersion
and hot-plate tests (Malmberg and Yaksh, 1995; present
study). (3) Ziconotide increases the mechanical thresholds
in the paw pressure test in a dose-dependent manner. This is
consistent with recent ®ndings that v -conotoxin GVIA, a
selective but irreversible blocker of N-type VSCCs
increased mechanical nociceptive thresholds in the paw
pressure test (Omote et al., 1996). However, the analgesia
produced in the paw pressure test is less potent (ED50 � 0:6
mg) than observed in neuropathic pain and persistent pain
(ED50s � 0:1 mg) with ziconotide (Bowersox et al., 1996;
present study). Neuropathic pain and persistent pain are
thought to be mediated by injury-induced spinal sensitiza-
tion of dorsal horn neurons (i.e. central sensitization)
(Woolf, 1983; Woolf et al., 1994; Sotgiu et al., 1998). Our
results suggest that ziconotide may interfere with central
sensitization and subsequent neuropathic pain and persistent
pain. In contrast, morphine produces little or no antinocicep-
tion in rat models of painful neuropathy (Courteix et al.,
1994; Mao et al., 1995; Nichols et al., 1995). Results from
human studies are consistent with these ®ndings with
morphine (Arner and Meyerson, 1988).
The major ®ndings in the present study are that (1) acute
intrathecal ziconotide and morphine do not interfere each
other's analgesia and produce either additive or apparent
synergistic antinociception depending on the nociceptive
test employed; (2) chronic intrathecal morphine administra-
tion causes tolerance to analgesia but does not produce
cross-tolerance to ziconotide antinociception; (3) chronic
intrathecal ziconotide administration does not produce
either tolerance to analgesia or cross-tolerance to morphine
analgesia; (4) the intrathecal co-administration of zicono-
tide neither prevents nor reverses morphine tolerance to
analgesia.
4.1. Acute spinal interaction between ziconotide and
morphine
Activation of m-opioid receptors by morphine and other
opioids leads to inhibition of N-type calcium channels via a
G-protein (Seward et al., 1991; Bourinet et al., 1996; Toth et
al., 1996; Zhang et al., 1996; De Waard et al., 1997) by
changing channel gating (Lipscombe et al., 1989) and alter-
ing ion permeation (Kuo and Bean, 1993). Since both zico-
notide and morphine inhibit N-type calcium channel
activity, it is necessary to think that they may act synergis-
tically to produce analgesia. Indeed, our results suggest
there is a synergistic analgesic effect between morphine
and ziconotide during the early phase after infusion in the
hot-plate and tail immersion tests or after acute administra-
tion in the formalin and paw pressure tests. (1) At doses that
do not inhibit acute ¯inch responses in the formalin test,
ziconotide signi®cantly potentiates morphine analgesia
(see Fig. 1). (2) Ziconotide and morphine potentiate each
other's analgesia in the initial infusion phase of the hot-plate
and tail immersion tests, although the dose-response analy-
sis is not applied and there is an unlikely possibility that the
markedly increased responses produced by the combination
ziconotide and morphine are due to steep slopes (high Hill
coef®cients) of the dose-response curves for morphine and/
or ziconotide alone. (3) At a dose that does not produce
Y.-X. Wang et al. / Pain 84 (2000) 271±281278
Fig. 8. Thermal nociceptive responses recorded after co-administration of
ziconotide (0.03 mg/h) and morphine (15 mg/h) compared to the sum of the
values for ziconotide (0.03 mg/h) and morphine (15 mg/h) administered
alone in the hot-plate test (A) and tail immersion test (B) in rats with raw
data from Fig. 7.

signi®cant analgesia by itself, ziconotide potentiates
morphine antinociception in the paw pressure test. These
synergistic effects on thermal and mechanical nociception
are supported by recent ®ndings that v -conotoxin GVIA,
another blocker of N-type VSCCs, has a naloxone-reversi-
ble synergistic effect with morphine in the tail-¯ick and paw
pressure tests by using isobolographical analysis (Omote et
al., 1996), and that v -conotoxin GVIA left-shifts morphine
analgesia dose-response curve in the tail-¯ick test (Wei et
al., 1996).
In contrast, there is an additive rather than synergistic
relationship between ziconotide and morphine analgesia in
the tonic phase of the formalin test. This result was obtained
using to different methods of drug interaction analysis: i.e.
comparing the composite additive and mixture regression
line approach and isobolographical analysis approach. Simi-
lar results were obtained with spinal ziconotide and the
selective a 2-adrenoceptor agonist clonidine, or the selective
GABA-B receptor agonist baclofen, in the tonic phase
responses in the formalin test (Wang et al., 1998; unpub-
lished data). The mechanisms for the difference are not
known.
4.2. Intrathecal morphine and ziconotide do not produce
cross-tolerance to each other's analgesia
A signi®cant increase of v -conotoxin GVIA binding sites
in brain was observed after chronic morphine treatment
(Suematsu et al., 1993), and v-conotoxin GVIA was
found to inhibit KCl-induced calcium in¯ux in synapto-
somes prepared from naive rats but not from morphine-
tolerant rats (Welch and Olson, 1991). These results suggest
that ziconotide antinociception might be altered in
morphine-tolerant rats. Our results indicate that the antino-
ciceptive effect of was the same in morphine-tolerant rats as
in naive rats. The inability of morphine to produce cross-
tolerance to ziconotide antinociception suggests that the
electrophysiological properties and function of N-type
calcium channels remain the same after morphine tolerance.
Indeed, chronic exposure to morphine reduced morphine-
induced N-type calcium currents but does not change the
electrophysiological properties of N-type calcium channels
(Kennedy and Henderson, 1991, 1992). It has been proposed
that morphine tolerance develops as opioid receptors are
uncoupled from their G-proteins (rather than reduction of
m-opioid receptors) and consequently from their effectors
such as N-type VSCCs (Kennedy and Henderson, 1991,
1992; Tao et al., 1993; see reviews by Basbaum, 1995,
1996). Our results with ziconotide in morphine-tolerant
rats are consistent with this model.
That ziconotide maintains the same antinociception ef®-
cacy in morphine tolerant animals makes it a unique anti-
nociceptive agent. Morphine is known to produce cross-
tolerance to other opioids and many other analgesic agents.
For example, cross-tolerance between intrathecal morphine
and a 2-adrenoceptor agonists have been observed (Solomon
and Gebhart, 1988; Roerig, 1995). Therefore, ziconotide
would not be expected to have reduced analgesic ef®cacy
when used in morphine-tolerant patients. This is an impor-
tant clinical implication as ziconotide is under investigation
for the treatment of chronic pain and many of these patients
have developed morphine tolerance to analgesia or
morphine-resistant (Brose et al., 1997; Presley et al., 1998).
One of the advantages of ziconotide is that long-term
administration does not produce tolerance to antinocicep-
tion in the formalin test (Malmberg and Yaksh, 1995;
Bowersox et al., 1996), hot-plate test (Malmberg and
Yaksh, 1995), and in neuropathic rats (Bowersox et al.,
1996; Wang et al., 1998, unpublished data). The present
study supports these ®ndings by showing that the antinoci-
ceptive response to ziconotide in the hot-plate test is main-
tained during the period of infusion. In addition, ziconotide
potentiates morphine analgesia after 7 days of infusion at
the same level as found with acute treatment. Therefore,
ziconotide produces neither tolerance to analgesia nor
cross-tolerance to morphine analgesia. Previous reports on
the effect of pretreatment with v -conotoxin GVIA on
morphine analgesia are con¯icting. It was reported that
pretreatment (24 h earlier) with intracerebroventricular
administration of v -conotoxin GVIA potentiated morphine
analgesia (Spampinato et al., 1994) but the same treatment
was reported to block morphine analgesia (Basilico et al.,
1992).
4.3. Intrathecal concurrent ziconotide does not prevent or
reverse spinal morphine tolerance to analgesia
Co-intrathecal infusion of ziconotide and morphine
produce synergistic antinociceptive effects in the hot-plate
and tail immersion tests during the initial phase of infusion.
However, this markedly increased effect disappears as the
rats develop tolerance to morphine analgesia. In addition,
co-intrathecal bolus injection of ziconotide and morphine
produce the same antinociception as ziconotide alone in
established morphine-tolerant rats. These results indicate
that ziconotide does not prevent or reverse morphine toler-
ance to analgesia. In contrast, it has been shown that NMDA
receptor antagonists such as MK-801 block morphine toler-
ance to analgesia (Trujillo and Akil, 1991; Gutstein and
Trujillo, 1993) in the spinal cord (Gutstein and Trujillo,
1993). NMDA receptors antagonists are known to block
voltage-dependent calcium currents including N-type
currents (Church et al., 1994; Biton et al., 1994). Our results
with ziconotide suggest that inhibition of spinal N-type
calcium channels is not involved in the effect of NMDA
receptor antagonists on morphine tolerance.
Acknowledgements
The research was supported by funds from Elan Pharma-
ceuticals. We thank Dr Sandra Roerig at Louisiana State
University and Mr Jeffrey McCary at Temple University
Y.-X. Wang et al. / Pain 84 (2000) 271±281 279

for their kind help with drug interaction analysis. We also
thank Dr Bill Hopkins at Elan Pharmaceuticals for his care-
ful review of the manuscript.
References
Abram SE, Yaksh TL. Morphine, but not inhalation anesthesia, blocks post-
injury facilitation. The role of preemptive suppression of afferent trans-
mision. Anesthesiology 1993;78:713±721.
Arner S, Meyerson BA. Lack of analgesic effect of opioids on neuropathic
and idiopathic forms of pain. Pain 1988;33:11±23.
Basbaum AI. Insights into the development of opioid tolerance. Pain
1995;61:349±352.
Basbaum AI. Memories of pain. Sci Med 1996;3:22±31.
Basilico L, Parolaro D, Rubino T, Gori E, Giagnoni G. In¯uence of v-
conotoxin on morphine analgesia and withdrawal syndrome in rats. Eur
J Pharmacol 1992;218:75±81.
Biton B, Granger P, Carreau A, Depoortere H, Scatton B, Avenet P. The
NMDA receptor antagonist eliprodil (SL 82.0715) blocks voltage-oper-
ated Ca21 channels in rat cultured cortical neurons. Eur J Pharmacol
1994;257:297±301.
Bourinet E, Soong TW, Stea A, Snutch TP. Determinants of the G protein-
dependent opioid modulation of neuronal calcium channels. Proc Natl
Acad Sci USA 1996;93:1486±1491.
Bowersox SS, Gadbois T, Singh T, Pettus M, Wang YX, Luther RR. Selec-
tive N-type neuronal voltage-sensitive calcium channel blocker, zico-
notide, produces spinal antinociception in rat models of acute, persistent
and neuropathic pain. J Pharmacol Exp Ther 1996;279:1243±1249.
Brose WG, Gutlove DP, Luther RR, Bowersox SS, McGuire D. Use of
intrathecal ziconotide, a novel, N-type, voltage-sensitive, calcium chan-
nel blocker, in the management of intractable brachial plexus avulsion
pain. Clin J Pain 1997;13:256±257.
Chaplan SR, Pogrel JW, Yaksh TL. Role of voltage-dependent calcium
channel subtypes in experimental tactile allodynia. J Pharmacol Exp
Ther 1994;269:1117±1123.
Church J, Fletcher EJ, Abdel-Hamid K, MacDonald JF. Loperamide blocks
high-voltage-activated calcium channels and N-methyl-d-aspartate-
evoked responses in rat and mouse cultured hippocampal pyramidal
neurons. Mol Pharmacol 1994;45:747±757.
Courteix C, Bardin M, Chantelauze C, Lavarenne J, Eschalier A. Study of
the sensitivity of the diabetes-induced pain model in rats to a range of
analgesics. Pain 1994;57:153±160.
De Waard M, Liu H, Walker D, Scott VE, Gurnett CA, Campbell KP.
Direct binding of G-protein betagamma complex to voltage-dependent
calcium channels. Nature 1997;385:446±450.
Gaur S, Newcomb R, Rivnay B, Bell JR, Yamashiro D, Ramachandran J,
Miljanich GP. Calcium channel antagonist peptides de®ne several
components of transmitter release in hippocampus. Neuropharmacol-
ogy 1994;33:1211±1219.
Gutstein HB, Trujillo KA. MK-801 inhibits the development of morphine
tolerance at spinal sites. Brain Res 1993;626:332±334.
Kennedy C, Henderson G. Mu-opioid receptor inhibition of calcium
current: development of homologous tolerance in single SH-SY5Y
cells after chronic exposure to morphine in vitro. Mol Pharmacol
1991;40:1000±1005.
Kennedy C, Henderson G. Chronic exposure to morphine does not induce
dependence at the level of the calcium channel current in human SH-
SY5Y cells. Neuroscience 1992;49:937±944.
Kristipati R, Nadasdi L, Tarczy-Hornoch K, Lau K, Miljanich GP, Rama-
chandran J, Bell JR. Characterization of the binding of omega-conopep-
tides to different classes of non-L-type neuronal calcium channels. Mol
Cell Neurosci 1994;5:219±228.
Kuo CC, Bean BP. G±protein modulation of ion permeation through N±
type calcium channels. Nature 1993;365:258±262.
Lipscombe D, Kongsamut S, Tsien RW. Alpha-adrenergic inhibition of
sympathetic neurotransmitter release mediated by modulation of N-
type calcium-channel gating. Nature 1989;340:639±642.
Malmberg AB, Yaksh TL. Voltage-sensitive calcium channels in spinal
nociceptive processing: blockade of N- and P-type channels inhibits
formalin-induced nociception. J Neurosci 1994;14:4882±4890.
Malmberg AB, Yaksh TL. Effect of continuous intrathecal infusion of
omega-conopeptides N-type calcium-channel blockers, on behavior
and antinociception in the formalin and hot-plate tests in rats. Pain
1995;60:83±90.
Mao J, Price DD, Mayer DJ. Experimental mononeuropathy reduces the
antinociceptive effects of morphine: implications for common intracel-
lular mechanisms involved in morphine tolerance and neuropathic pain.
Pain 1995;61:353±364.
Miljanich GP, Ramachandran J. Antagonists of neuronal calcium channels:
structure, function, and therapeutic implications. Annu Rev Pharmacol
Toxicol 1995;35:707±734.
Nichols ML, Bian D, Ossipov MH, Lai J, Porreca F. Regulation of
morphine antiallodynic ef®cacy by cholecystokinin in a model of neuro-
pathic pain in rats. J Pharmacol Exp Ther 1995;275:1339±1345.
Olivera BM, Cruz LJ, de Santos V, LeCheminant GW, Grif®n D, Zeikus R,
McIntosh JM, Galyean R, Varga J, et al. Neuronal calcium channel
antagonists. Discrimination between calcium channel subtypes using
omega-conotoxin from Conus magus venom. Biochemistry
1987;26:2086±2090.
Olivera BM, Miljanich GP, Ramachandran J, Adams ME. Calcium channel
diversity and neurotransmitter release: the v-conotoxins and v-agatox-
ins. Annu Rev Biochem 1994;63:823±867.
Omote K, Kawamata M, Satoh O, Iwasaki H. Spinal antinociceptive action
of an N-type voltage-dependent calcium channel blocker and the syner-
gistic interaction with morphine. Anesthesiology 1996;84:636±643.
Presley R, Charapata S, Ferrar-Brechner T, Yearwood T, Staats P, Wallace
MS, Ordia J, Gaeta R, Follett KA. Chronic, opioid-resistant, neuro-
pathic pain: marked analgesic ef®cacy of intrathecal ziconotide. Am
Pain Soc Abstr. 1998;17:697.
Roerig SC. Decreased spinal morphine/clonidine antinociceptive syner-
gism in morphine-tolerant mice. Life Sci. 1995;56:PL115±PL122.
Seward E, Hammond C, Henderson G. Mu-opioid-receptor-mediated inhi-
bition of the N-type calcium-channel current. Proc R Soc Lond B Biol
Sci 1991;244:129±135.
Solomon RE, Gebhart GF. Intrathecal morphine and clonidine: antinoci-
ceptive tolerance and cross-tolerance and effects on blood pressure. J
Pharmacol Exp Ther 1988;245:444±454.
Sotgiu ML, Biella G, Firmi L, Pasqualucci V. Topical axonal transport
blocker vincristine prevents nerve injury-induced spinal neuron sensi-
tization in rats. Neurotrauma 1998;15:1077±1082.
Spampinato S, Speroni E, Govoni P, Pistacchio E, Romagnoli C, Murari G,
Ferri S. Effect of omega-conotoxin and verapamil on antinociceptive,
behavioural and thermoregulatory responses to opioids in the rat. Eur J
Pharmacol 1994;254:229±238.
Suematsu M, Ohnishi T, Shinno E, Maeda S, Matsumoto K, Sakuda M,
Saito K. Effect of prolonged administration of clonidine on [3H]PN 200-
110 and [125I]omega conotoxin binding in mouse brain. Neurosci Lett
1993;163:193±196.
Tallarida RJ, Murray RB. Manual of pharmacologic calculations, New
York: Springer, 1987. pp. 26±31.
Tallarida RJ, Porreca F, Cowan A. Statistical analysis of drug-drug and site-
site interactions with isobolograms. Life Sci 1989;45:947±961.
Tallarida RJ, Stone Jr DJ, Raffa RB. Ef®cient designs for studying syner-
gistic drug combinations. Life Sci 1997;61:417±425.
Tao PL, Lee CR, Law PY, Loh HH. The interaction of the mu-opioid
receptor and G protein is altered after chronic morphine treatment in
rats. Naunyn-Schmiedebergs Arch Pharmacol 1993;348:504±508.
Toth PT, Shekter LR, Ma GH, Philipson LH, Miller RJ. Selective G-protein
regulation of neuronal calcium channels. J Neurosci 1996;16:4617±
4624.
Trujillo KA, Akil H. Inhibition of morphine tolerance and dependence by
the NMDA receptor antagonist MK-801. Science 1991;251:85±87.
Y.-X. Wang et al. / Pain 84 (2000) 271±281280

Wang YX, Pettus M,Luther RR, Bowersox SS.Effects of the selective N-type
calcium channel blocker, ziconotide, on the development of tolerance to
spinal morphine analgesia in rats. Neurosci Soc Abstr 1997;23:1121.
Wang YX, Bezprozvannaya S, Bowersox SS, Nadasdi L, Miljanich G,
Mezo G, Silva D, Tarczy-Hornoch K, Luther RR. Peripheral versus
central potencies of N-type voltage-sensitive calcium channel blockers.
Naunyn-Schmiedebergs Arch Pharmacol 1998;357:159±168.
Wei ZY, Karim F, Roerig SC. Spinal morphine/clonidine antinociceptive
synergism: involvement of G proteins and N-type voltage-dependent
calcium channels. J Pharmacol Exp Ther 1996;278:1392±1407.
Welch SP, Olson KG. Opiate tolerance-induced modulation of free intra-
cellular calcium in synaptosomes. Life Sci 1991;48:1853±1861.
White DM, Cousins MJ. Effect of subcutaneous administration of calcium
channel blockers on nerve injury-induced hyperalgesia. Brain Res
1998;801:50±58.
Woolf CJ, Shortland P, Sivilotti LG. Sensitization of high mechanothres-
hold super®cial dorsal horn and ¯exor motor neurones following
chemosensitive primary afferent activation. Pain 1994;58:141±155.
Woolf CJ. Evidence for a central component of post-injury pain hypersen-
sitivity. Nature 1983;306:686±688.
Xiao WH, Bennett GJ. Synthetic omega-conopeptides applied to the site of
nerve injury suppress neuropathic pains in rats. J Pharmacol Exp Ther
1995;274:666±672.
Yaksh TL, Rudy TA. Chronic cauterization of the spinal subarachnoid
space. Physiol Behav 1976;17:1031±1036.
Yamamoto T, Sakashita Y. Differential effects of intrathecally administered
N- and P-type voltage-sensitive calcium channel blockers upon two
models of experimental mononeuropathy in the rat. Brain Res
1998;794:329±332.
Yamamoto T, Sakashita Y. Differential effects of intrathecally administered
N- and P-type voltage-sensitive calcium channel blockers upon two
models of experimental mononeuropathy in the rat. Brain Res
1998;794:329±332.
Zhang JF, Ellinor PT, Aldrich RW, Tsien RW. Multiple structural elements
in voltage-dependent Ca21 channels support their inhibition by G
proteins. Neuron 1996;17:991±1003.
Y.-X. Wang et al. / Pain 84 (2000) 271±281 281
Top Related
Copyright © 2022 FDOKUMEN

