







Bahasa
Halaman
Hukum


Current Biotica 5(1): 1-16
1
In vitro clonal propagation of Jatropha curcas (L.) using nodal explants and
assessment of genetic fidelity through RAPD markers
Subramanyam Koona1* Subramanyam Kondeti2, Muralidhararao Doulathabadh
3 and
Rajasekhar Pinnamaneni4
1,4,*
Department of Biotechnology, Sreenidhi Institute of Science and Technology, JNTU,
Yamnampet, Ghatkesar, R.R. District, Hyderabad – 501 301, India 2Plant Molecular Biology Unit, Department of Biotechnology, Bharathidasan University,
Tiruchirappalli – 620 024, Tamilnadu, India 3Department of Biotechnology, Sri Krishnadevaraya University, Anantapur – 515 003, A. P., India
*E-mail: [email protected]
ABSTRACT
An efficient micropropagation protocol was developed for Jatropha curcas (L.) using
nodal explants, and its genetic fidelity and was assessed using RAPD markers. Auxillary shoot
buds were induced from seven–month–old nodal explants on MS medium supplemented with
Kn, BAP, and TDZ along with AdS. A maximum number of shoot buds (6.2 ± 0.56) were
induced on MS medium fortified with 21.6 µM BAP and 54.2 µM AdS. Shoot multiplication
from the auxiliary buds was significantly enhanced (about 32.0 ± 0.01) on MS medium
containing 0.4 µM IBA, 4.4 µM Kn, 21.6 µM BAP and 27.1 µM AdS. About 54% rooting
was achieved on MS medium supplemented with 1.0 µM IBA. The well–rooted plantlets were
established in green house with 91% survival frequency. A total of 15 RAPD primers
produced in 925 distinct and reproducible bands and confirmed the genetic fidelity of the
regenerated Jatropha curcas plants.
KEYWORDS : Auxillary shoot buds, Clonal propagation, Genetic fidelity, Jatropha curcas,
RAPD
ABBREVIATIONS
RAPD Randomly amplified polymorphism of DNA
MS Murashige and Skoog
Kn Kinetin
BAP Benzylamino purine
TDZ Thidiazuron
AdS Adenine sulphate
IBA Indole-3-butyric acid

Current Biotica 5(1): 1-16
2
INTRODUCTION
Jatropha curcas (L.) species is a
perennial, monoecious shrub or a small tree,
native to America, widely distributed in
tropics. J. curcas, a member of
Euphorbiaceae family, can grow well under
any unfavorable agro climatic conditions,
because of its low moisture demands,
fertilizer requirements and tolerance to high
temperature (Tiwari et al. 1994). It is found
throughout the tropics and known by nearly
200 names, which indicate its significance
(Koushik et al. 2007). Recently, the
economic importance of J. curcas has been
increasing because of its oil used as a fuel,
biofuel and is being looked at as an
important alternative fuel in the overall
energy security worldwide. The oil is also a
rich source of hydrocarbon (27.0–48.5% of
seed oil) (Martin and Mayeux 1985). It
promises for its use as an oil crop for
biodiesel (Henning 1998). The main
advantages of using biodiesel are its
renewability, better quality exhaust gas
emission, and biodegradability. It does not
contribute to a rise in the level of carbon
dioxide in the atmosphere (Korbitz 1999;
Beet et al. 2002; Sims 2001). Extracts from
the plant are known for their medicinal
properties and their effects on a wide array
of organisms including insects, pests, and
nematodes (Jain and Trivedi 1997). It is a
multipurpose tree species, which fits for
agro forestry and other afforestation
programmes (Wood and Boorely 1991).
Conventional agriculture uses seeds
and cuttings for its propagation. But the
seeds are heterozygous in nature, and the
cuttings are seasonal. Moreover, it is
reported that vegetative cuttings are not
deep-rooted and are easily uprooted as they
do not form a taproot system. Seed set has
been reported to be low in vegetatively
propagated plants (Sujatha et al. 2006).
Tissue culture studies were
undertaken in different species of Jatropha.
Morphogenesis from an endosperm tissue
has been reported in J. panduraefolia
(Srivastava 1971; Srivastava and Johri
1974). High frequency regeneration from
various explants of J. integerrima has been
reported (Sujatha and Dhingra 1993).
Using different explants, plant
regeneration protocols have also been
described in J. curcas (Sujatha et al. 2006;
Qin et al. 2004; Rajore and Batra 2005;
Sujatha and Mukta 1996), but multiplication
rate was low for field applications. Nodal
meristems are an important source tissue for
micropropagation (Mukul Manjari et al.
2007) and plants raised from these are
comparatively more resistant to genetic
variation (Pierik 1991). Keeping in mind the
economical importance of Jatropha curcas,
critical analysis of the earlier protocols
necessitated formulating a well-documented,
reproducible, in vitro micropropagation
protocol.
The genetic integrity of
micropropagated plants can be determined
with the use of various techniques. Various
authors have found RAPD technique useful
in examining genetic fidelity of tissue
culture clones. With the use of RAPD
markers, clonal fidelity of micropropagated
plants has been determined in Pinus
thunbergii (Goto et al. 1998), Lilium
(Varshney et al. 2001), and Tylophora
indica (Jayanthi and Mandal 2001). Anna
and Ewa (2004) have confirmed the
suitability of RAPD technique for
determining the genetic fidelity in two
micro-propagated Drosera species, D.
anglica and D. binata. The present study has
been taken up to develop an efficient micro
propagation protocol using nodal explants
and assess its genetic fidelity using RAPD
markers.

Current Biotica 5(1): 1-16
3
MATERIALS AND METHODS
Plant material
Jatropha curcas (L.) seeds were
procured from the National Bureau of Plant
Genetic Resources (NBPGR), New Delhi,
India. The seeds were germinated in sterile
vermiculate mixture at 25–30°C in light and
later transferred to field. Nodal explants
were collected from these plants to
standardize the micropropagation protocol.
Nodal explants of 2–3 cm long, collected
from the seven–month–old plants were
immersed in 1% systemic fungicide
“Bavistin” (w/v) (BASF India Ltd) for 3 h,
they were surface sterilized by soaking in
0.1% mercuric chloride for 5 min, and
finally rinsed three times with sterile double
distilled water to remove remnants of
mercuric chloride.
Induction of shoots
For auxillary shoot bud induction,
about 1 cm long nodal explants were
inoculated onto Murashige and Skoog’s
(MS) medium (Murashige and Skoog 1962)
supplemented with 54.2 µM AdS in
combination with cytokinins viz., 2.2–
36.6µM Kn, 2.1–34.6 µM BAP, and 2.2–
36.4µM TDZ (shoot induction medium)
separately. The pH of the medium was
adjusted to 5.7 prior to autoclaving at 121°C
for 15 min. Cultures were incubated at
25±1°C under 16 h photo period provided
by cool white fluorescent lamps (Philips,
New Delhi) for 4 weeks. Shoot induction
was recorded after 4 weeks of culture in
each treatment which was replicated thrice
that consisted of 10 nodal explants each.
Shoot multiplication and elongation
After 4 weeks of culturing, the
induced auxillary shoot buds were separated
from nodal explants, transferred onto MS
medium fortified with 27.1 µM AdS in
combination with 2.1–21.6 µM BAP alone
or in combination with 0.2–1.6 µM IBA and
2.2–22.6 µM Kn (shoot multiplication
medium) for efficient shoot multiplication.
After 3 weeks of shoot multiplication, the
shoot clusters were sub cultured into the
same medium for elongation. The cultures
were maintained at 25 ±1°C under 16 h
photoperiod for another 3 weeks.
Root initiation and elongation
The elongated shoots were separated
from the shoot clusters and placed in rooting
medium consisting of MS medium
supplemented with 0.5–10 µM IBA for 3
weeks under 16 h photo period at 25 ±1ºC
temperature. The root elongation was
achieved on only MS basal medium in 2
weeks.
Hardening and field transfer
The plantlets with well–developed
roots were gently removed from the culture
flasks and thoroughly washed in sterile
double distilled water. The plantlets were
then transferred into pots containing a
mixture of soil and vermiculate in the ratio
of 1:1 and covered with clear polythene bags
to maintain high humidity. After 3 weeks of
hardening, the plantlets were transferred to
big earthen pots.
Statistical analysis
The experiments were carried out in
a completely randomized design. Data were
analyzed by analysis of variance (ANOVA)
to detect significant differences between
means at P ≤ 0.05. Means differing
significantly were compared using Duncan’s
Multiple Range Test (DMRT).

Current Biotica 5(1): 1-16
4
Assessment of genetic fidelity
The total genomic DNA was isolated
from the leaves of mother plant and ten
randomly selected tissue culture-raised J.
curcas plants by following modified Cetyl
Tri Methyl Ammonium Bromide (CTAB)
method (Doyle and Doyle 1990). The
isolated genomic DNA concentration and
quality were cheecked by using the
spectrophotometer (BL198 ELICO, INDIA)
at 260 nm. Fifteen RAPD primers were used
for the analysis of genetic fidelity. The
RAPD amplification were performed using
20 µl PCR containing 50 ng of template
DNA, 1 x PCR Buffer, 2.5 mM MgCl2
(Bangalore Genei Pvt. Ltd., Bangalore), 200
µM dNTP mix (Genetix, New Delhi, India),
0.5 µM of RAPD primer (OPERON
Technologies, Inc., USA), 0.2 U of Taq
DNA polymerase (Fermentas Pvt. Ltd).
The PCR was performed at an initial
denaturation at 94ºC for 5 min followed by
35 cycles of 1 min denaturation at 94ºC, 1
min annealing at 36ºC, and 2 minutes
extension at 72ºC with final extension at
72ºC for 5 min using thermal cycler (UVI
Gene technologies Pvt. Ltd. INDIA). The
PCR products along with the λ DNA ECoR I
– Hind III double digest were separated on
1.4% agarose gel in 1X TBE buffer
containing ethidium bromide at 150 V and
photographed under ultraviolet light using
Gel Doc 2000 (Bio - RAD).
Data analysis
RAPD markers for parent plant and
regenerates were visually scored for the
presence (1) and absence (0) for each
primer. By comparing the banding patterns
of parent and regenerates for a specific
primer, specific bands were identified, and
faint or unclear bands were not considered.
Similarity matrices were generated by
percent similarity, and dendrogram was
constructed by using the Unweighed Pair
Group method with arithmetic Average
(UPGMA) with the MVSP 3.0 (Multi
Variate Statistical Analysis; [Kovach
Computing Services, 1987-1998]) to show
genetic relationships as revealed by the
percent similarity.
RESULTS
The nodal explants cultured on MS
medium supplemented with 54.2 µM AdS
with different cytokinins (Kn, TDZ, and
BAP) showed varied response (Table. 1).
The better response (6.2 ± 0.56
shoots per nodal explant) was obtained in
the presence of 54.2 µM AdS, 21.6 µM BAP
and was found to be significantly (P ≤ 0.05
level) higher than shoots induced per nodal
explant in other concentrations of cytokinins
(Kn, TDZ) used in the present study (Table
1, Fig. 1 C).
Four-week-old induced shoot buds
on MS medium supplemented with 54.2 µM
AdS and 21.6 µM BAP were transferred
onto MS basal medium fortified with 27.1
µM AdS with different cytokinins (BAP,
IBA, and Kn) singly or in combination
showed varied response (Table 2). In the
present study, transfer of shoot buds onto
MS medium with lower concentration of
AdS (27.1 µM) and with same concentration
of BAP (21.6 µM) produced 10.2 ± 0.05
shoots per nodal explant with average shoot
length of 1.5 ± 0.08 cm. Below or above the
21.6 µM BAP, shoot number and shoot
length gradually reduced (Table. 2). In
order to increase the shoot number, we
analyzed the effect of IBA and Kn in
combination with 21.6 µM BAP. A 0.4 µM
IBA and 4.4 µM Kn along with 21.6 µM
BAP produced maximum number of shoots
(32.0 ± 0.01 shoots per nodal explant) with
avarage shoot length of 2.1 ± 0.01 cm
(Table 2, Fig. 1 D and E ).

Current Biotica 5(1): 1-16
5
When approximately 2.0 cm length
elongated shoots were inoculated onto MS
medium with lower concentrations of IBA
(0.5–1.0 µM), 54% of the inoculated shoots
were induced roots with average number of
5.6 ± 0.04 roots per shoot at 1.0 µM
concentration within three weeks (Table 3,
Fig. 1 F). Below or above 1.0 µM IBA,
percentage of response to root formation
drastically reduced. A distinct tap root
system developed with slender and white
secondary roots; this was considered
important for hardening and field transfer
(Fig. 1 G). Significant increase in root
length occurred on transfer to MS basal
medium for another 2 weeks. The well–
rooted plantlets were separated from the
medium, thoroughly washed with distilled
water, dipped for 1 h in 0.1% (w/v) bavistin
(systemic fungicide), transplanted to plastic
pots containing a mixture of soil and
vermiculate in equal ratio and covered with
polythene bags to maintain high humidity.
The plantlets were irrigated with tap water
as and when required. Polythene covers
were completely withdrawn after 3 weeks of
hardening. Plants were then transferred to
potted soil for further growth (Fig. 1 H). The
plants ranged from 86.0 ± 0.12 to 102.0 ±
0.21 cm in height after 10 months (Fig. 1 I).
RAPD pattern of 10 randomly
selected micropropagules obtained from
nodal explants were compared with mother
plant. Of the fifteen primers (Table 4)
screened, 14 primers yielded clear,
reproducible bands. The number of bands
for each primer varied from three in OPA-06
to nine in OPB-01. Each primer produced
amplification products in the size range 0.2
kb in OPB- 04 to 2.0 kb in OPA-01. The 14
tested primers yielded totally 925 scorable
bands (number of propagules x number of
screened markers) with an average of six
bands per primer. The entire tested primers
produced monomorphic pattern across all
the shoots, as examples, the patterns
obtained for primers OPA-01 and OPA-06
and OPA-07 are shown in (Fig. 2 A–C).
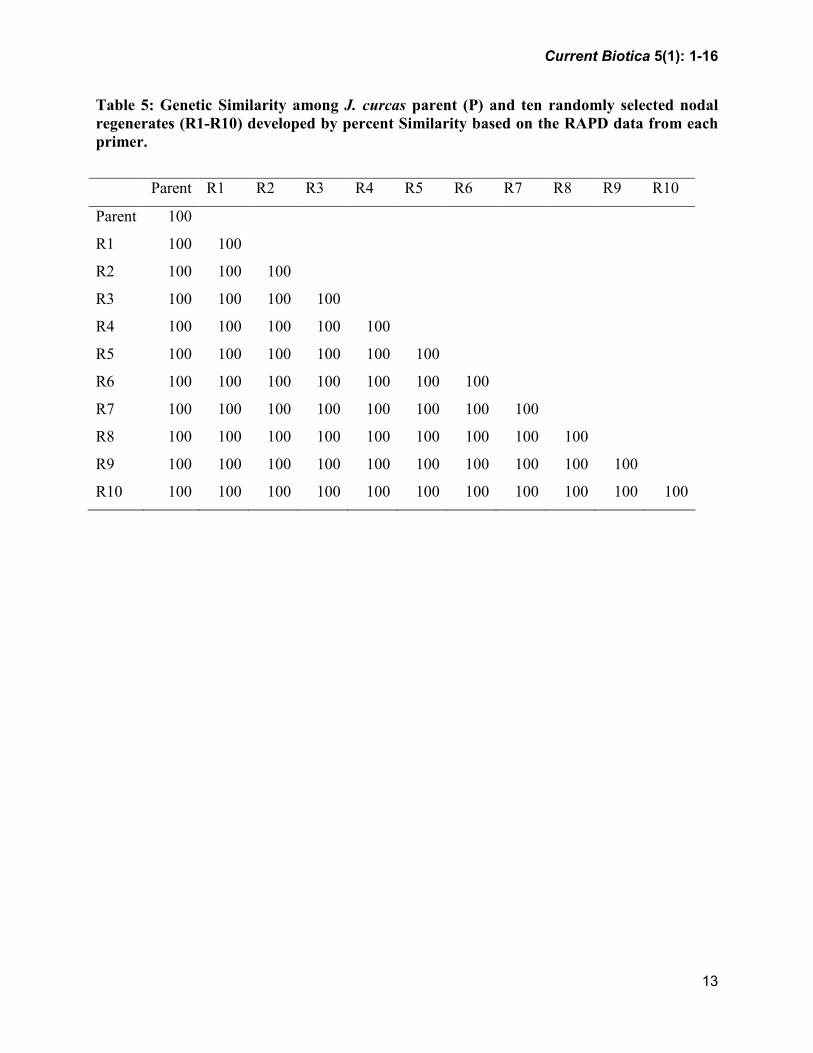
UPGMA Dendrogram revealed that
regenerates and mother plant grouped in
single cluster with 100% similarity. Genetic
Similarity was also found to be 100%
between the mother plant and regenerates,
and between the regenerates (Table 5;
Fig. 3), confirming the genetic uniformity of
the micro propagated plant material.
DISCUSSION
The best shoot induction response
(6.2 ± 0.56 shoots per nodal explant) was
obtained in the presence of 54.2 µM adenine
sulphate, 21.6 µM BAP. At the same
concentration of 21.6 µM BA, a lower
initiation (1.5 ± 0.2 shoot buds per explant;
2.2 ± 0.6 shoots per nodal explant) and
shoot bud proliferation were reported
(Rajore and Batra 2005). This differential
response may be attributed to the specific
age and physiological condition of the donor
plant from which the nodal explants were
excised. Nodes cultured on medium with
different concentrations of Kn showed lower
induction and proliferation of axillary shoot-
buds. Though TDZ is known to induce
cytokinin–like effects in a number of plant
species, particularly woody (Barrueto et al.
1999) as well as herbaceous crop species,
the present study showed inefficient effect
of TDZ on shoot induction, similar findings
were also reported in Jatropha curcas
(Sujatha et al. 2006).
The present study revealed that
shoots cultured on medium with
combinations of BA and Kn did not
proliferate further or showed slow response.
On the other hand, transferring the cultures
from a set having higher concentration of
BAP (21.6 µM) and 54.2 µM adenine

Current Biotica 5(1): 1-16
6
sulphate to a set with lower concentration of
AdS (27.1 µM), in combination with 21.6
µM BAP, 0.4 µM IBA, and 4.4 µM Kn, led
to a significantly higher nodal shoot
multiplication (32.0 ± 0.01 shoots per nodal
explant) within the next 3 weeks, which was
not recorded previously with this
concentrations. This was possibly due to a
combined effect of different growth
regulators along with other additives.
Lowering of growth regulators in
micropropagation studies to achieve higher
rate of multiplication has been reported in
Holarrhena antidysenterica (Kumar and
Sharma 2005). However, it has been
observed in Jatropha that it requires higher
concentration of only one type of cytokinin
(BAP) for induction phase and favors lower
concentration of another type of cytokinins
(IBA and Kn) along with other additives for
escalation and proliferation of shoot
cultures. In a recent report, Sujatha et al.
(2006) indicated similar effect in J. curcas
with BA and TDZ, but the authors obtained
lesser number of shoots (2.0 ± 0.8 to 12.3 ±
1.7) with comparatively higher amount of
cytokinin.
The genetic integrity of
micropropagated plants can be determined
with the RAPD as it has the advantage of
ease, quick performance, relatively low cost
and requires very little plant material
(Rafalski et al. 1993). The results presented
in this study demonstrate that the RAPD
technique proved to be effective in
generating reproducible results useful in
assessment of genetic fidelity of
micropropagated propagules in Jatropha
cultivars.
CONCLUSION
A simple, efficient and high fidelity
protocol for mass propagation of Jatropha
curcas from nodal explants has been
established. Using this protocol, it is
possible to produce viable, uniform and
healthy plants with maximum survival rate
for the proposed in vitro germplasm. The
protocol should also provide an efficient
means for large-scale cultivation and in vitro
multiplication of J. curcas. It is evident that
the RAPD technique proved to be effective
in generating reproducible results that are
useful in assessment of genetic stability of in
vitro propagated propagules in J. curcas.
ACKNOWLEDGMENTS
Authors are thankful to the National Bureau
of Plant Genetic Resources (NBPGR), New
Delhi, India, Jawaharlal Nehru
Technological University, Anantapur, Sri
Krishnadevaraya University, Anantapur, and
Gandhi Krishi Vignana Kendra (GKVK),
Sreedher Bhat’s Research laboratory (SBL),
Bangalore, for facilitating the research work.
REFERENCES
Anna K, Ewa L (2004) Application of
RAPD in the determination of genetic
fidelity in micropropagated Drosera
plantlets. In Vitro Cellular and
Developmental Biology plant 40,
592-595
Barrueto LP, Machado ACMG, Carvalheira
SBRC, Brasileiro ACM (1999) Plant
regeneration from seedling explants
of Eucalyptus grandis × Europhylla.
Plant Cell Tissue and Organ Culture
56, 17–23
Beet T, Grant T, David W, Harry W (2002)
Fuel cycle green house emissions
from alternative fuels in Australian
heavy vehicles. Atmospheric
environment 36, 753–763

Current Biotica 5(1): 1-16
7
Doyle JJ, Doyle JL (1987) A rapid DNA
isolation procedure for small
quantities of fresh leaf tissue.
Phytochemical Bulletin 19, 11-15
Goto S, Thakur RC, Ishii K (1998)
Determination of genetic stability in
long term micropropagated shoots of
Pinus thunbergii Parl. Using RAPD
markers. Plant Cell Reports 18, 193-
197
Henning R (1998) Use of Jatropha curcas –
household perspective and its
contribution to rural employment
creation, in Proceedings of the
regional workshop on the ‘‘potential
of Jatropha curcas in rural
development and environmental
protection’’ May 13–15, (Harare,
Zimbabwe).
Jain C, Trivedi PC (1997) Nimaticidal
activity of certain plants against root-
knot nematode, Meloidogyne
incognita infecting chick pea. Annals
of Plant Protection Science 5, 171-
174
Jayanthi M, Mandal PK (2001) Plant
regeneration through somatic
embryogenesis and RAPD analysis of
regenerated plants in Tylophora
indica (Burm. F. Merrrii.). In Vitro
Cellular Developmental Biology Plant
37, 576-580
Korbitz W (1999) Biodiesel production in
Europe and American encouraging
prospect. Renew Energy 16: 1078–
1083
Kumar R, Sharma K, Agrawal V (2005) In
vitro clonal propagation of
Holarrhena antidysenterica (L.)
(Wall.) through nodal explants of
mature trees. In Vitro Cellular
Developmental Biology Plant 41,
137–144
Koushik N, Krishnan K, Shshil K, Nutan K
Roy S (2007) Biomass and Bioenergy
31, 497-502
Martin G, Mayeux A (1985) Curcas oil
(Jatropha curcas L.): a possible fuel.
Agricultural Tropics 9, 73–75
Mukul Manjari D, Priyanka M, Timir B
(2007) In vitro clonal propagation of
biodiesel plant (Jatropha curcas L.).
Current Science 10, 1438-1442
Murashige T, Skoog F (1962) A revised
medium for rapid growth and
bioassays with tobacco tissue
cultures. Physiology of Plants 15,
473–479
Pierik RLM (1991) Commercial aspects of
micropropagation. In: Prakash, JK,
Pierik RLM, (Eds.), Horticulture –
New Technologies and Applications
Dordercht, The Netherlands.
Qin W, Wei-Da L, Yi L, Shu-Lin P, Ying
XU, Lin T, Fang C (2004) Plant
regeneration from epicotyl explants of
Jatropha curcas. Journal of Plant
Physiologyand Molecular Biology
30, 475–478
Rafalski A, Tingey S, Williams JGK (1993)
Random amplified polymorphic DNA
(RAPD) markers. In: Gelvin SB,
Schilperoort RMS, Verma DPS
(Eds.), Plant molecular biology
manual suppl 6 Dordrecht: Kluwer
Academic Publishers, pp 1-9
Rajore S, Batra A (2005) Efficient plant
regeneration via shoot tip explant in

Current Biotica 5(1): 1-16
8
Jatropha curcas. Journal of Plant
Biochemistry and Biotechnology 14,
73–75
Sims REH (2001) Bioenergy – a renewable
carbon sink. Renewable Energy 22,
31–37
Srivastava PS (1971) In vitro induction of
triploid roots and shoots from mature
endosperm of Jatropha
panduraefolia. Z Pflanzenphysiololy
66, 93–96
Srivastava PS, Johri BM (1974)
Morphogenesis in mature endosperm
cultures of Jatropha panduraefolia.
Beitr Biol Planz 50, 255–268
Subramanyam K, Muralidhararao D,
Devanna N (2009) Novel molecular
approach for optimization of DNA
isolation and PCR protocol for RAPD
analysis and genetic diversity
assessment of Jatropha curcas
(Euphorbiaceae). Current Biotica 3,
1-13.
Subramanyam K, Muralidhararao D,
Devanna N (2009) Genetic diversity
assessment of wild and cultivated
varieties of Jatropha curcas (L.) in
India by RAPD analysis. Afr Journal
of Biotechnology 8, 1900-1910
Sujatha M, Dhingra M (1993) Rapid plant
regeneration from various explants of
Jatropha integerrima. Plant Cell
Tissue and Organ Culture 35, 293–
296
Sujatha M Makkar HPS, Becker K (2006)
Shoot bud proliferation from axillary
nodes and leaf sections of non-toxic
Jatropha curcas L. Plant Growth
Regulations 47, 83–90
Sujatha M, Mukta N (1996) Morphogenesis
and plant regeneration from tissue
cultures of Jatropha curcas. Plant
Cell Tissue and Organ Culture 44,
135–141
Tiwari DN (1994) Brouchur on Jatropha.
Dehra Dun: ICFRI.
Varshney A, Lakshmikumaran M,
Srivastava PS, Dhawan V (2001)
Establishment of genetic fidelity of in
vitro raised Lilium bulbets through
RAPD markers. In Vitro Cellular and
Developmental Biology Plant 37,
227-231
Wood PJ, Burely J (1991) A tree for all
reasons: the introduction and
evaluation of Multipurpous trees for
agroforestry. Kenya:ICRAF, Nairobi;
58

Current Biotica 5(1): 1-16
9
Table 1: Effect of different types of medium and concentrations of cytokinins on shoot
induction of J. curcas
MS basal medium
+ 54.2 µM AdS +
Conc. of
cytokinin (µM)
Average number
of shoots (9> 0.5 cm)
developed per nodal
Average length of
shoots developed
(cm ± SE)
Kn
TDZ
BAP
2.2
4.4
11.3
22.6
36.6
2.2
4.4
11.4
22.7
36.4
2.1
4.2
10.8
21.6
34.6
2.2 ± 0.17b
2.0 ± 0.10b
2.4 ± 0.12b
3.5 ± 0.35c
1.0 ± 0.01a
3.0 ± 0.02c
1.2 ± 0.04a
1.0 ± 0.05a
1.0 ± 0.02a
1.0 ± 0.01a
1.0 ± 0.01a
2.0 ± 0.13b
2.7 ± 0.23bc
6.2 ± 0.56f
5.3 ± 0.40e
0.8 ± 0.02a
0.8 ± 0.04a
0.8 ± 0.06a
0.9 ± 0.07a
0.8 ± 0.02a
0.8 ± 0.03a
0.9 ± 0.06a
0.9 ± 0.02a
0.8 ± 0.03a
0.9 ± 0.02a
0.8 ± 0.02a
0.8 ± 0.05a
0.9 ± 0.03a
2.0 ± 0.18b
1.4 ± 0.12ab
Data were recorded after 4 weeks of culture. Each treatment was repeated thrice and each
replicate consisted of 10 nodal explants. Means having different letters as superscripts are
significantly different from each other (P ≤ 0.05) according to DMRT.

Current Biotica 5(1): 1-16
10
Table 2: Effect of growth regulators on shoot multiplication and elongation from nodal
explants of J. curcas
Shoot multiplication and elongation
medium* with growth regulators (µM)
BAP IBA Kn
Number of
shoots per explants
Length of shoots
(cm)
2.1
4.2
10.8
21.6
34.6
21.6
21.6
21.6
21.6
21.6
21.6
21.6
21.6
-
-
-
-
-
0.2
0.4
0.8
1.6
0.4
0.4
0.4
0.4
-
-
-
-
-
-
-
-
-
2.2
4.4
11.3
22.6
2.0 ± 0.01a
4.0 ± 0.03b
8.0 ± 0.03a
10.2 ± 0.05c
7.0 ± 0.05e
18.2 ± 0.01a
22.1 ± 0.09a
12.0 ± 0.03b
8.5 ± 0.02c
26.3 ± 0.03g
32.0 ± 0.01f
21.2 ± 0.02e
15.1 ± 0.01d
0.8 ± 0.02a
0.8 ± 0.05a
1.0 ± 0.05a
1.5 ± 0.08b
1.2 ± 0.04ab
1.8. ± 0.01a
2.0 ± 0.03a
1.7 ± 0.01a
1.1 ± 0.04b
1.3. ± 0.01a
2.1 ± 0.01a
1.7 ± 0.02a
1.1 ± 0.03b
*MS basal medium + 27.1 µM AdS along with different plant growth regulators. Data were
recorded after 3 weeks of culture. Each treatment was repeated thrice and each replicate
consisted of 10 nodal explants. Means having different letters as superscripts are significantly
different from each other (P ≤ 0.05) according to DMRT.

Current Biotica 5(1): 1-16
11
Table 3: Effect of IBA on initiation of roots in excised shoots of J. curcas
MS + IBA
(µM)
Percentage of
root induction
Number of roots
per shoot
Length of roots
(cm ± SE)
0.5
1.0
2.5
5.0
10.0
15
54
20
16
10
1.0 ± 0.06a
5.6 ± 0.04b
4.1 ± 0.02a
2.4 ± 0.06a
2.1 ± 0.01a
2.8 ± 0.09a
5.9 ± 0.10b
3.8 ± 0.06a
2.9 ± 0.04a
2.4 ± 0.02a
Data were recorded after 3 weeks of culture. Each treatment was repeated thrice and each
replicate consisted of 20 shoots. Means having different letters as superscripts are significantly
different from each other (P ≤ 0.05) according to DMRT.

Current Biotica 5(1): 1-16
12
Table 4: Number of amplification products generated from each RAPD primers in the
analysis of genetic fidelity of in vitro propagated J. curcas plants.
S.No Primer Sequence (5’-3’) Number of
generated bands
1
2
3
4
5
6
7
8
9
10
11
12
13
14
OPA-01
OPA-02
OPA-03
OPA-04
OPA-05
OPA-06
OPA-07
OPA-08
OPA-09
OPA-10
OPB-01
OPB-02
OPB-04
OPB-05
CAGGCCCTTC
TGCCGAGCTG
AGTCAGCCAC
AATCGGGCTG
AGGGGTCTTG
GGTCCCTGAC
GAAACGGGTG
GTGACGTAGG
GGGTAACGCC
GTGATCGCAG
GTTTCGCTCC
TGATCCCTGG
GGACTGGAGT
TGCGCCCTTC
07
05
05
06
08
03
06
05
05
08
09
08
06
04
Current Biotica 5(1): 1-16
13
Table 5: Genetic Similarity among J. curcas parent (P) and ten randomly selected nodal
regenerates (R1-R10) developed by percent Similarity based on the RAPD data from each
primer.
Parent R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
Parent 100
R1 100 100
R2 100 100 100
R3 100 100 100 100
R4 100 100 100 100 100
R5 100 100 100 100 100 100
R6 100 100 100 100 100 100 100
R7 100 100 100 100 100 100 100 100
R8 100 100 100 100 100 100 100 100 100
R9 100 100 100 100 100 100 100 100 100 100
R10 100 100 100 100 100 100 100 100 100 100 100

Current Biotica 5(1): 1-16
14
Fig. 1: In vitro clonal propagation of Jatropha curcas (L.) nodal explants. A. Nodal explant from
the seven-month old mother plant. B. Nodal explant after the first week of culture on shoot
induction medium (MS medium containing 54.2 µM AdS and 21.6 µM BAP). C. Auxillary shoot
bud induction from nodal explant after 2 weeks of culture on shoot induction medium (MS
medium containing 54.2 µM AdS and 21.6 µM BAP). D. Shoot multiplication after 3 weeks of
culturing on shoot multiplication medium (MS medium containing 27.1 µM AdS, 21.6 µM BAP,
0.4 µM IBA, and 4.4 µM Kn). E. Elongated shoots on shoot multiplication medium (MS
medium containing 27.1 µM AdS, 21.6 µM BAP, 0.4 µM IBA, and 4.4 µM Kn) after 6 weeks of
culturing. F. Root initiation from the elongated shoots on rooting medium (MS medium
containing 1.0 µm IBA) after 3 weeks of culturing. G. In vitro roots with slender and white tap
root system. H. Jatropha curcas (L.) plants after 10 weeks of hardening. I. Ten month-old
micropropagated Jatropha curcas (L.) plant in big earthen pot under natural sun light

Current Biotica 5(1): 1-16
15
Fig.2: Agarose gel electrophoresis and RAPD banding pattern of Jatropha curcas (L.) mother
and regenerated lines generated with primer OPA-01 (A), OPA-06 (B), and OPA-07 (C). Lane
P: Mother plant; Lanes 1 – 10: Regenerated plant lines; Lane M: λ DNA EcoR I and Hind III
double digest

Current Biotica 5(1): 1-16
16
Fig.3: Dendrogram generated from the RAPD data obtained from the parent (mother
plant) and nodal explant derived regenerated plant lines (R1-R10) based on percent
similarity
[MS received: 20-4-2011;
MS accepted: 03-6-2011]
Top Related
Copyright © 2022 FDOKUMEN

