







Bahasa
Halaman
Hukum


Review
Human ankyrins and their contribution to disease biology:An update
NIDHI SHARMA, KAMLESH BHAM and SABYASACHI SENAPATI*
Immunogenomics Laboratory, Department of Human Genetics and Molecular Medicine,Central University of Punjab, Bathinda, Punjab 151401, India
*Corresponding author (Email, [email protected])
MS received 28 June 2020; accepted 16 October 2020
Ankyrins (Ank) are ubiquitously expressed proteins that play a critical role in the integrity of cytoskeleton andcellular signalling. Their presence in metazoans and evolutionary conserved protein primary sequence indicatestheir functional significance. Tissue-specific isoforms and an array of transcript variants make this protein oneof the indispensable cellular components. Membrane-binding domains consist of ankyrin repeats that bind withseveral functional membrane proteins that enable maintaining cellular integrity. Cytosolic ankyrins help incellular signal transduction. Linkage studies and recent genome-wide association studies uncovered thepathogenic roles of ankyrins (ankyrin-R, ankyrin-B and ankyrin-G) in several diseases, such as hereditaryspherocytosis, long QT syndrome, intellectual disability, and CRASH syndrome, among several others.Identification of Ank3 in celiac disease may potentially explain the link between neuronal health and immunity.It is thus warranted to investigate the role of neuronal factors in immune diseases and vice versa. In this review,we briefly discussed the contribution of ankyrin genes to human diseases.
Keywords. Ankyrins; cellular adhesion; cellular integrity; immunity; neuronal health
1. Introduction
In multicellular organisms, the plasma membrane playsa key role in maintaining the functional organization oftissues and cellular cytoskeleton. The integrity of thecellular cytoskeleton is crucial in maintaining the shapeand specific function of cells. The property of theplasma membrane depends on various factors, includ-ing membrane-spanning proteins and intracellularstructural protein complexes. Adaptor proteins are oneof these specific membrane-spanning proteins whichorganize the protein networks, link membrane proteinswith the cytoskeleton, carry out signalling processesand maintain electrogenic properties (Rubtsov andLopin 2000). Ankyrins are one of such modularadaptor proteins, which mediate linkage of integralmembrane proteins to spectrin cytoskeleton (Cunha
and Mohler 2009). Different ankyrins can bind withvarious membrane proteins located in the particularareas of the plasma membrane. Through these inter-actions, ankyrins perform a wide range of molecularfunctions including membrane transport, cellularadhesion, cellular signalling and maintenance ofcytoskeleton structures (figures 1 and 2).A 33-amino-acid sequence known as ankyrin repeats
was first described in the Swi6p/cdc10p, a yeast cellcycle regulator and NOTCH, a Drosophila signallingprotein (Breeden and Nasmyth 1987). A similarsequence was later identified in the cytoskeleton pro-tein ankyrin which comprises 24 copies of such repeats(Lux et al. 1990). Ankyrin repeats are present in all thekingdoms such as archea, bacteria, and eukaryotes aswell as in various viral genomes. There has been sig-nificant variation in the number of ankyrin repeats
Electronic supplementary material: The online version of this article (https://doi.org/10.1007/s12038-020-00117-3) containssupplementary material, which is available to authorized users.
http://www.ias.ac.in/jbiosci
J Biosci (2020) 45:146 � Indian Academy of SciencesDOI: 10.1007/s12038-020-00117-3 ( 0123456789().,-volV)( 0123456789().,-volV)

present in a single protein (Mosavi et al. 2004; Li et al.2006). Tumour suppressor protein (p15, p16, p18,p19), a regulator of tumour suppressor protein p53, i.e.53BP2 and signalling protein Notch contains ankyrinrepeats (Mosavi et al. 2004). Various genetic and epi-genetic alterations like methylation, point mutation,deletion have been detected in genes encoding variousankyrin repeat proteins such as p16 and p18 indicatesthat any disruption in the functioning of ankyrin repeatsmay lead to serious health issues (Ortega et al. 2002).Ankyrin proteins are also present in plants, loss ofwhich leads to chlorotic phenotype (Zhang et al. 1992).Due to lack of enzymatic activity ankyrin repeats needto collaborate with other domains to find out the defi-nite functions of ankyrin protein in plants. Based on itsdiverse domains ankyrin protein in plants is classifiedinto 13 subgroups or clusters.
These proteins evolved early in metazoans, havinginvertebrate orthologs of ankyrins, which shows a highdegree of similarity in structural and functional aspectsunder different cellular adaptation conditions (Mohleret al. 2002). Recent developmental biology studies onmodel organisms uncovered an orthologous relation-ship between mammalian ankyrins (Ank1, Ank2, andAnk3) with unc-44 in Caenorhabditis elegans andDank1, Dank2 in Drosophila melanogaster (Lambertet al. 1990; Otto et al. 1991; Dubreuil and Yu 1994;Kordeli et al. 1995; Otsuka et al. 1995; Peters et al.1995; Bouley et al. 2000; Mohler et al. 2002).Over three decades ago ankyrin was first character-
ized as an adaptor protein in red blood cells (RBCs),which play a critical role in the formation of proteincomplexes consisting of ion channels/transporters, celladhesion molecules, signalling proteins and
Figure 1. Different interactions of ankyrin proteins in human. MBD: Membrane binding domain; SBD: Spectrin bindingdomain; DD: Death domain; CTD: C-terminal domain.
146 Page 2 of 16 Nidhi Sharma et al.
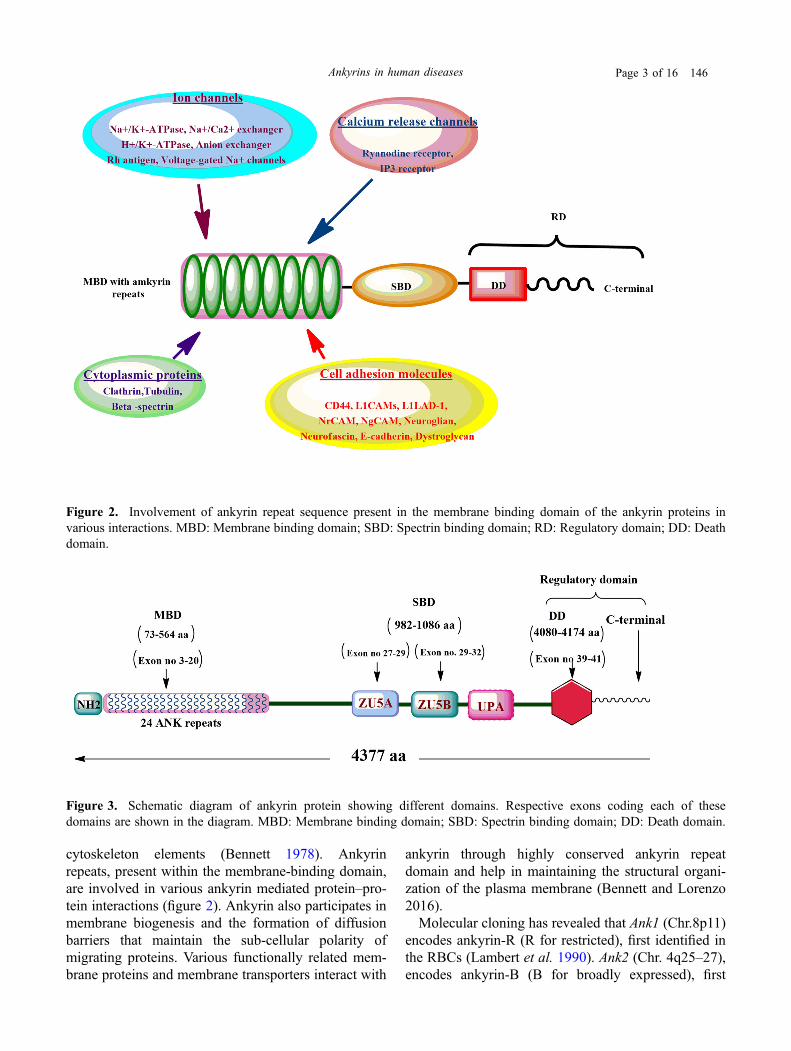
cytoskeleton elements (Bennett 1978). Ankyrinrepeats, present within the membrane-binding domain,are involved in various ankyrin mediated protein–pro-tein interactions (figure 2). Ankyrin also participates inmembrane biogenesis and the formation of diffusionbarriers that maintain the sub-cellular polarity ofmigrating proteins. Various functionally related mem-brane proteins and membrane transporters interact with
ankyrin through highly conserved ankyrin repeatdomain and help in maintaining the structural organi-zation of the plasma membrane (Bennett and Lorenzo2016).Molecular cloning has revealed that Ank1 (Chr.8p11)
encodes ankyrin-R (R for restricted), first identified inthe RBCs (Lambert et al. 1990). Ank2 (Chr. 4q25–27),encodes ankyrin-B (B for broadly expressed), first
Figure 3. Schematic diagram of ankyrin protein showing different domains. Respective exons coding each of thesedomains are shown in the diagram. MBD: Membrane binding domain; SBD: Spectrin binding domain; DD: Death domain.
Figure 2. Involvement of ankyrin repeat sequence present in the membrane binding domain of the ankyrin proteins invarious interactions. MBD: Membrane binding domain; SBD: Spectrin binding domain; RD: Regulatory domain; DD: Deathdomain.
Ankyrins in human diseases Page 3 of 16 146

identified in the brain (Otto et al. 1991) and Ank3(Chr.10q21) encodes ankyrin-G (G for giant or gen-eral), first characterized in the node of Ranvier andepithelial tissues (Kordeli et al. 1995). Structurallyankyrins have three distinct functional domainsnamely, membrane-binding domain (MBD, comprisesof 24 consecutive ankyrin repeats) (Davis and Bennett1990; Michaely et al. 2002); spectrin binding domain(SBD, comprises of ZU51 and ZU52) (Bennett 1978;Wang et al. 2012); and regulatory domain (comprisesof C-terminal domain and death domain) (Hall and
Bennett 1987; Wang et al. 2014) assembled throughlinker amino acid chains (figure 3). These specializeddomains facilitate ankyrins to interact with variouscytoskeleton and membrane proteins (El Refaey andMohler 2017) and help in cellular stability.Functional domains such as death domain (DD),
ZU51 and ZU52 have considerable amino acidsequence variability across different ankyrins (fig-ure 4), which may indicate specific functions attributedto different ankyrins. Ankyrin-B death domain (DD2)has 60% sequence homology with DD of ankyrin-G,
Figure 4. Amino acid sequence homology (a–c) and phylogenetic relationships (d–f) between three functional domains(death domain, ZU51 and ZU52) respectively present in ankyrin proteins. Sequence similarities between different domains ofankyrin proteins were studied using ‘‘MEGA-X’’ software. Amino acid sequences of different domains of ankyrin wereretrieved from UniProt database and details are given in supplementary Table 1.
146 Page 4 of 16 Nidhi Sharma et al.
while DD of ankyrin-R showed 55.2% homology. Inthe case of ZU51, ankyrin-B showed 58.2% sequencesimilarity with ankyrin-R, while ankyrin-G showed69.2%, and in the case of ZU52, ankyrin-B share63.9% sequence similarity with ankyrin-R whileankyrin-G shared 65.3% sequence similarity withankyrin-R (figure 4a–c). The phylogenetic relationshipshows similar patterns across ankyrins for ZU51 andZU52 domains. A similar branching pattern is seen inthe cladogram of these three domains among differentankyrins (figure 4d–f). These evidences indicate thefunctional stratification of each of these ankyrins. Thesource of sequence information about these domains ofankyrin is given in supplementary Table 1.Considering the organizational diversity of different
ankyrins present in different tissues, the major focus ofthis review article is to give an insight into their con-tribution to human health and diseases. Ankyrins havebeen implicated in several pathophysiological condi-tions such as ankyrin-B syndrome, cardiac arrhythmia,sick sinus syndrome, hereditary spherocytosis, etc.(Mohler and Bennett 2005; Bennett and Healy 2008;Hashemi et al. 2009; Cunha and Mohler 2009). Therole of Ank3 in the nervous system is well establishedand its common sequence variations are known to beassociated with neuronal dysfunction that leads tobipolar disorder, autism spectrum disorders,schizophrenia, amyotrophic lateral sclerosis (Cassidyet al. 2014). Ankyrin-L1CAM interaction in peripheralneurons explains the function of ankyrins in neuro-logical disorders such as CRASH syndrome (Bennettand Healy 2008). Structure complexities and functional
role of ankyrins in the nervous system, RBC and car-diac muscles had been investigated in depth. However,their interactions with key cellular regulatory proteinsin determining complex tissue homeostasis have notbeen illustrated well. Elaborate investigations onankyrins are warranted to understand the diseasepathology and target them for disease treatment. Herewe presented a focused update on ankyrin researchaiming their contribution to human disease.
2. Human ankyrins: structure and types
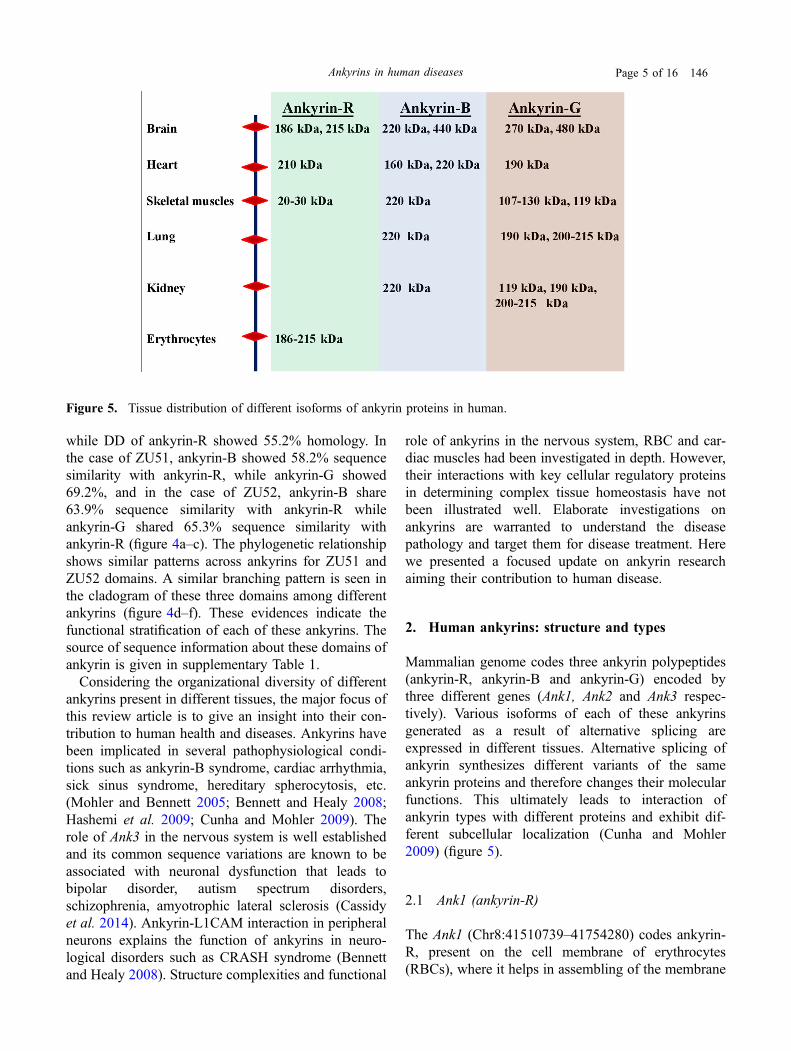
Mammalian genome codes three ankyrin polypeptides(ankyrin-R, ankyrin-B and ankyrin-G) encoded bythree different genes (Ank1, Ank2 and Ank3 respec-tively). Various isoforms of each of these ankyrinsgenerated as a result of alternative splicing areexpressed in different tissues. Alternative splicing ofankyrin synthesizes different variants of the sameankyrin proteins and therefore changes their molecularfunctions. This ultimately leads to interaction ofankyrin types with different proteins and exhibit dif-ferent subcellular localization (Cunha and Mohler2009) (figure 5).
2.1 Ank1 (ankyrin-R)
The Ank1 (Chr8:41510739–41754280) codes ankyrin-R, present on the cell membrane of erythrocytes(RBCs), where it helps in assembling of the membrane
Figure 5. Tissue distribution of different isoforms of ankyrin proteins in human.
Ankyrins in human diseases Page 5 of 16 146
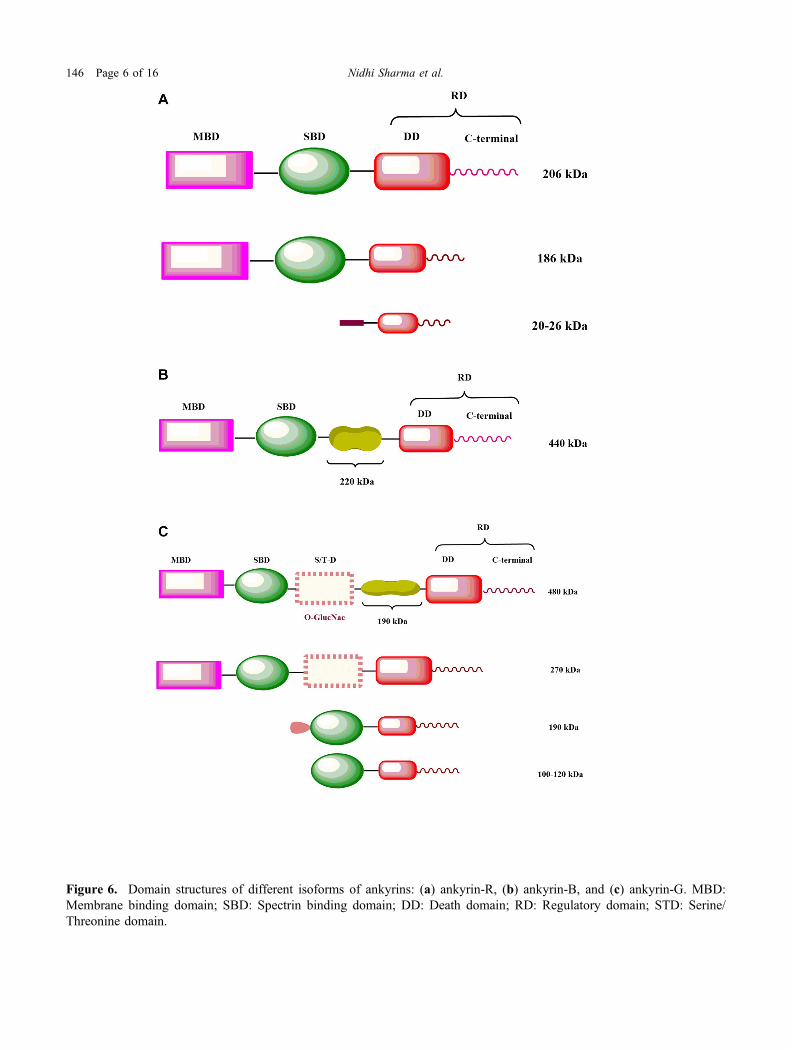
Figure 6. Domain structures of different isoforms of ankyrins: (a) ankyrin-R, (b) ankyrin-B, and (c) ankyrin-G. MBD:Membrane binding domain; SBD: Spectrin binding domain; DD: Death domain; RD: Regulatory domain; STD: Serine/Threonine domain.
146 Page 6 of 16 Nidhi Sharma et al.

proteins. This specific assembly helps in maintainingstructural flexibility and functional stability. Duringblood circulation, erythrocytes experience shear forcesand ankyrin-R enables RBCs to resist these forces. Itlinks the membrane skeleton to the Cl-/HCO3
- anionexchanger. Ankyrin-R is also present in the macro-phages, muscles, endothelial cells, Purkinje cells of thecerebellum, neurons of the hippocampus and spinalcord. Various isoforms generated as a result of alter-native splicing are expressed in the erythrocytes,skeletal and cardiac muscles. 186 kDa isoform, formedas a result of the deletion of 162 aa from the regulatorydomain and smaller isoforms of 20–26 kDa is presentin the Z and M line of skeletal muscle fibre. Smallisoforms carry a small C-terminal region and are defi-cient in membrane and spectrin binding domains. TheN-terminal has a distinct hydrophobic structure of 72amino acids long with a membrane extending helix(Rubtsov and Lopina 2000; Hashemi et al. 2009)(figure 6a).
2.2 Ank2 (ankyrin-B)
Isoforms of ankyrin-B generated by the alternativesplicing of Ank2 (Chr4: 113739265–114304896) areexpressed in the brain, thymus and skeletal muscles.The 440-kDa isoform, having a rod-shaped domain of220-kDa between the c-terminal and spectrin bindingdomain (SBD), present in the unmyelinated axons anddendrites in the neonatal brain. While the 220 kDaisoform having similar domain structure as that ofankyrin-R is present in the neural cell bodies, dendrites,and glial cells of the adult brain respectively (Rubtsovand Lopina 2000) (figure 6b).Ankyrin-B helps in organizing the cytoskeleton and
links certain proteins to the cell membrane. Along withthis, it helps in cell migration, cell growth and prolif-eration. It is mainly functionally active in the brain andcardiac muscles, where it interacts with ion channelsand ion transporters across the cell membrane. AnkyrinB is uniformly expressed in the sinoatrial node, ven-tricles and atria (Cunha and Mohler 2009). In cardiacmuscles, it maintains heart rhythm by controlling theflow of ions through ion channels and ion transporters,which in turn generates electrical signals to regulateheartbeat. It also plays an important role in the sig-naling pathway of cardiomyocytes by acting as abinding partner of B56a in the ventricular myocytes,which is the regulatory subunit of protein phosphatase2A (PP2A). PP2A regulates various transporters andion channels such as Na/K ATPase, L-type Ca2?
channels, IP3 receptors and ryanodine receptors. In thebrain, along with ankyrin-G, it regulates axonalmovements (DeGrande et al. 2012; Little et al. 2015).
2.3 Ank3 (ankyrin-G)
Various isoforms of Ank-3 (Chr10: 61786056–62493248) expressed in the brain, kidney, skeletal aswell as cardiac muscles and epithelial tissues. Threeisoforms of ankyrin-G express in the brain. 190 kDaisoform expressed mainly in the unmyelinated axonsand 270 and 480 kDa get expressed in the initial seg-ments of myelinated axons and nodes of Ranvierrespectively (Kordeli et al. 1995; Peters et al. 1995). Afragment of 46 kDa rich in serine and threonine isincorporate between the terminal part and SBD of270–480 kDa isoform of ankyrin-G. Glycosylation ofthis fragment appears to be the reason behind thepeculiar presence of these isoforms. 270 kDa isoformis lacking a stretch of 190 kDa. 119 kDa isoform ofankyrin-G interconnected with Golgi is present in theepithelium and the muscles. Another 190 kDa isoformof ankyrin-G is present in the kidney and contains adistinctive fragment of 31 amino acids at its N-terminaldomain (Devarajan et al. 1996; Hoock et al. 1997).There are four isoforms of ankyrin-G that are present inthe kidney, i.e. 215 and 200 kDa (in the proximal cells)as well as 170 and 120 kDa (in the distal tubular cellsof the inner medulla). Smaller isoforms of ankyrin-Gi.e. 100–120 kDa are localized in the lysosomes andsarcoplasmic reticulum (Doctor et al. 1998). The smallisoforms, which are expressed in the colon, kidney, andtestes, are deficient in the membrane-binding domainand have a small regulatory domain of 29 amino acids(Hoock et al. 1997) (figure 6c).The expression of Ank3 is ubiquitous and it plays an
important role in neural health. The excitability ofneurons is guided by an ankyrin-G dependent subcel-lular gradient of neurofascin in Purkinje neurons(Jenkins and Bennett 2001). It also helps in synapto-genesis, synaptic stability and maintaining neuromus-cular junction (Musarella et al. 2006). There arevarious mechanisms by which ankyrin-G impacts theneural circuit. One of them is by targeting Na?/K?
channels to axon initial segment (AIS) and nodes ofRanvier in neurons of the central nervous system.Ankyrin-G plays a crucial role in the initial nodeassembly, as many key nodal proteins have an ankyrin-G binding site. Thus, any mutation in ankyrin-Gdestabilizes the assembly of these protein complexes atthe nodes of Ranvier. Dense cluster of ion channels,
Ankyrins in human diseases Page 7 of 16 146

cell adhesion molecules and interaction betweenankyrin-G and spectrin at nodes of Ranvier and AIS arevital for the production of action potential (Zhang andBennett 1998; Saifetiarova et al. 2017).
3. Ankyrin-associated human diseases
All forms of ankyrin proteins (ankyrin-R, ankyrin-B,and ankyrin-G) expressed in different tissues are vitalfor normal tissue functioning. Any disruption in thestructural domains of ankyrins disturbs the bindingefficiency with various associated proteins, which maylater affect molecular processes and thus interrupt thecellular homeostasis that ultimately leads to variousdiseases.Ankyrin-B is associated with various forms of
acquired and congenital cardiovascular diseases (ElRefaey and Mohler 2017). The continuous expressionof ankyrin-B in the various parts of the cardiac region(sinoatrial node, atria, ventricles) causes cardiac dis-orders i.e. delayed conduction/conduction block, sinusarrhythmia, idiopathic ventricular fibrillation, brady-cardia and catecholaminergic polymorphic ventriculartachycardia (Cunha and Mohler 2009).Various ankyrin associated diseases/syndromes are
discussed below:
3.1 Ankyrin-B syndrome/Type 4 long QT syndrome(LQT4)
Ankyrin-B syndrome includes Type 4 long QT syn-drome, cardiac arrhythmia and sinoatrial node auto-maticity (Mohler et al. 2003. 2007). Ankyrin-B-syndrome, formerly labelled as LQT4 (also known asLQTS4) is the result of a heterozygous loss-of-functionmutation in Ank2. It disturbs Ca2? homeostasis due todecreased L-type Ca2? current (ICa.L), Na?- Ca2?
exchange current (INCX), expression of IP3 receptor andNa?-K? ATPase. Mutation in the Ank2 gene is linkedwith both sinus bradycardia as well as prolonged cor-rected QT interval (QTC) (Wilders and Verkerk 2018).Repolarization of the heart gets affected in LQT4
condition (Hashemi et al. 2009), which results inbradycardia, that can cause fainting or sudden death ofthe subject. In 1995 Schott and colleagues first identifiedLQT4 in a French cohort having atrial fibrillation, sinusnode dysfunction and irregular T-wave (Schott et al.1995). A missense mutation (c.4274A[G, Exon 36),which causes the substitution of glycine to glutamic acidat amino acid 1425 (E1425G) in the spectrin-bindingdomain of ankyrin-B was identified in patients only(Mohler and Bennett 2005; Mohler 2006; Hashemi et al.2009; El Refaey and Mohler 2017; Koenig and Mohler2017). Ank-2 E1425G mutation in human and anankyrin-B ± mice exhibits various common cardiacdisorders, like episodes of atrioventricular dissociation,prolonged QT interval, severe bradycardia, heart ratevariability, catecholamine-induced ventricular arrhyth-mias, and sudden death (Hashemi et al. 2009; Koenigand Mohler 2017). Additionally, ankyrin-B ± mice andankyrin-B syndrome patients exhibit sudden cardiacdeath in response to exercise and catecholamine injec-tion, syncope and polymorphic ventricular tachycardia.The loss-of-function variants of Ank2 (G1406C,R1450W, L1503V, T1404I, T1552N, V1777M, E1425G,V1516D, R1788W) have been classified into three clas-ses (Table 1) on the basis of in vitro and clinical phe-notypes exhibited by the protein (Hashemi et al. 2009).A study by Swayne et al. on the Gitxsan population
has identified a novel variant of Ank2 i.e. Ank2c.1937C[T which causes p.Ser646Phe non-synony-mous substitution and linked with cardiac abnormali-ties as well as LQT4 syndrome. This loss-of-functionmutation modifies the tertiary structure of MBD, whichultimately disturbs the factors responsible for its sta-bility and localization (Koenig and Mohler 2017;
Table 1. The loss-of-function variants of Ank2 associated with several clinical phenotypes
S.No. Variants SNP Class Phenotype
1. G1406C, R1450W, L1503V rs34591340rs762281932rs45570339
Ist It remains asymptomatic and exhibits mild in vitro as wellas clinical phenotypes as compare to the wild-type neonatalcardiomyocytes
2. T1404I, T1552N, V1777M rs142534126rs45608232rs199473314
IInd Associated with serious human ventricular phenotypes andshow loss-of-function alleles in case of primary myocytes
3. E1425G, V1516D, R1788W rs72544141rs66785829rs121912706
IIIrd Most severe variants are associated with third class anddisplay most severe in vitro as well as clinical phenotype
146 Page 8 of 16 Nidhi Sharma et al.

Swayne et al. 2017). Another non-synonymous sub-stitution (p.R990Q) in ankyrin-B disrupts the Ankrin-Band bII-spectrin interaction and leads to severearrhythmia. A study of this variant by Smith et al. hasrevealed that bII-spectrin regulates the localization ofankyrin-B, Ryanodine receptor 2 (RyR2) and Na/Caexchanger (NCX) (Smith et al. 2015).
3.2 Cardiac arrhythmia
Cardiac arrhythmia initially known as long QT syn-drome is the result of loss-of-function mutation(p.Q1283H) in ankyrin-B which increases the risk ofsudden cardiac death (Zhu et al. 2018). This variant islocated in the ZU5 region on Ank2. Studies on ankyrin-B (±) cardiomyocytes have documented that the sud-den death due to arrhythmia is possibly because ofirregular heartbeat caused due to increased calciumtransient (Mohler et al. 2007).Ankyrin-B is also associated with abnormal distri-
bution and function of membrane proteins i.e. Na/Caexchanger (NCX), Na/K ATPase (NKA) and InsP3receptors. In the adult and neonatal cardiomyocytes,ankyrin-B is localized in the transverse (T) tubule/Z-line and M-line respectively. Decreased expression ofankyrin-B in the ankyrin-B (±) cardiomyocytes resultsin reduced function and irregular localization of thesemembrane proteins which indicates that ankyrin-B mayplay a central role in arrhythmia and the electricalremodelling (Mohler 2006; Hashemi et al. 2009; ElRefaey and Mohler 2017).
3.3 Ankyrin-B and sinoatrial node automaticity(SND)
Sinoatrial node automaticity also known as sick sinussyndrome affects elderly people with a rapid increase infrequency with age. Its rate of occurrence is 1 in 600individuals aged above 65 years and accounts for 50%of pacemaker implantations in the USA (Bernstein andParsonnet 1992). A family-based study identified Ank2mutation (E1425G) to have absolute penetrance.Mutation was not found among unaffected members(Le Scouarnec et al. 2008). A relatively low penetrantbut strongly linked Ank2 haplotype was identified in anindependent study comprising 44 members of a family.A very high LOD score for marker D4S1616 wasobserved (Le Scouarnec et al. 2008). Mutation in theAnk2 leads to decreased expression of ankyrin-B,which is associated with the familial form of normal
SND and may appear at a younger age than normalsporadic SND (Hashemi et al. 2009).
3.4 Ankyrin-B in arrhythmogenic cardiomyopathy
A recent study by Robert et al. has revealed that thereis a direct interaction between Ankyrin-B and b-cate-nin. Heterogeneous pattern of expression of b-cateninin the hearts of human probands as well as Ank2-cKO(cardioselective knockout) mice, indicated disturbedhomeostasis of b-catenin, which causes structuralremodeling in arrhythmogenic cardiomyopathy. Thus,ankyrin-B may indirectly affect b-catenin by affectingits phosphorylation through ankyrin-B-PP2A interac-tion (Roberts et al. 2019).
3.5 Ank3 and early-life stress (ELS)
Luoni et al. have reported the association betweenearly-life stress (ELS) and the methylation of Ank3which may play a critical role in the susceptibility ofpsychiatric disorders. Reduced DNA methylation (atthe 1st intron) and two differentially methylatedregions downstream at the 24th intron of Ank3 havebeen observed in the adult prefrontal cortex (PFC) ofthe prenatal stressed (PNS) rats as compared to thecontrol rats. However, the same genomic region washypermethylated in the hippocampus of male ratsexposed to PNS when compared to the control one.The reduced transcript level of Ank3 in the PNS ratswas reported due to differential methylation. mRNAlevel was stable before the infant stage while there is anabrupt increase in adults indicating Ank3 has its role inpostnatal neurodevelopment. Contrary to these changesfrom weaning to adulthood is affected in the ratsexposed to PNS while there is marked reduction in theexpression of Ank3 at adulthood. It has been shownthat PNS affects the subcellular distribution of ankyrin-G and decreases its protein level in the membranecompartment. Besides this, there is a marked increaseof ankyrin-G interaction with PSD95 (postsynapticdensity protein 95) which may change the functionalintegrity of this domain (Luoni et al. 2016).
3.6 Hereditary spherocytosis
Ankyrin-R is majorly implicated in hereditary sphero-cytosis, a rare disorder affecting RBCs. Hereditaryspherocytosis is hemolytic anemia which is
Ankyrins in human diseases Page 9 of 16 146

characterized by increased hemolysis due to change inthe shape of RBCs from biconcave to spherical onewhich may result in fragmentation of RBCs in the highshear environment of the vascular system. In rareconditions, some neurological problems were alsofound to be associated with hereditary spherocytosis,e.g. a normoblastosis murine model exhibit motorataxia along with severe hemolytic anemia (Cunha andMohler 2009; Bennett and Healy 2008). Randon et al.have identified a few cases of hereditary spherocytosisthat were caused by de novo mutation of Ank1 whilestudying three Italian families showing an apparentlyrecessive pattern of hereditary spherocytosis. Threenovel frameshift mutations in the Ank1 were reported.These mutations include allele Bari (1361delG,Exon12), Napoli II (2883delC, Exon26) and Anzio(3032delCA, Exon26) (Randon et al. 1997).A study by Clarkson et al. has shown that the
association between ankyrin-R and bIII-spectrin iscrucial for regulating the activity of sodium channel inthe neurons. Thus any impairment in this interaction isassociated with human ataxia and motor defects such asbroad-based gait and loss of balance in individuals withhereditary spherocytosis. bIII-spectrin also regulatesthe ankyrin-R throughout the plasma membrane ofPurkinje cells of dendrites and any alteration in thismay lead to abnormal dendrite morphology and humanataxia (Clarkson et al. 2014).
3.7 Alzheimer’s disease
Epigenome-wide association study (EWAS) on Alz-heimer’s disease (AD) has identified the significantenrichment of DNA methylation at Ank1 in the braintissue taken from AD patients. Reduced expression ofAnk2 (ortholog of human Ank1) in Drosophila is thereason behind memory impairment like in humans.Overexpression of mutant amyloid precursor protein(APP) leads to AD in human. Reduced locomotion andshortening of life span have been observed in the ani-mals with decreased expression of Ank2 or overex-pression of APP (Higham et al. 2019). A studyconducted by Lunnon et al. on three independent cohortshas identified that cortex-specific hypermethylation onCpG island of Ank1 is linked with Alzheimer’s diseaseneuropathology (Lunnon et al. 2014). Ank3 expression(Participates in brain plasticity and neural transmission)found to be reduced in Alzheimer’s disease, has beenrescued using riluzole treatment. Riluzole, a glutamatemodulator has also been approved to cure amyotrophiclateral sclerosis (Pereira et al. 2017).
4. Ankyrin interacting partners and impacton human health
4.1 L1CAM–ankyrin interaction
CAMs (cell adhesion molecules) are transmembranereceptors that belong to the subfamily ofimmunoglobulin. These molecules comprise six Ig-likedomains, five fibronectin-like repeats, a transmembraneregion and a cytoplasmic domain (Kamiguchi andLemmon 1997). This family consists of structurallyrelated proteins such as L1, NrCAM, Neurofascin,Neuroglian (in D. melanogaster), NgCAM and CD44.L1CAMs plays role in the processes of nervous systemdevelopment such as neurite fasciculation, neuriteoutgrowth and inner neuronal adhesion (Davis et al.1993).Ankyrin-B and G are major forms expressed in the
nervous system. The specific binding of L1-moleculewith ankyrin takes place through the C-terminal of itscytoplasmic domain. This ankyrin binding site of L1-molecule is very specific and does not have anyhomology with other ankyrin-proteins (Rubtsov andLopina 2000). The key amino acid residues for thisspecific binding lies within a 26-residue-long stretchhaving a sequence EDGSFIGQY, which is very specificto all family members from C.elegens to mammals(Bennett and Baines 2001). This interaction facilitatesthe physiological assembly of membrane proteinsincluding transmembrane adhesion molecules, spectrinskeleton and voltage-gated Na? channels at the initialsegment of axon (Jenkins and Bennett 2001). Twohighly conserved motifs LADY (Leu-Ala-Asp-Tyr) andFIGQY (Phe-Ile-Gly-Gln-Tyr) are necessary for inter-action between L1CAM and ankyrin (Tuvia et al.1997; Garver et al. 1997). This interaction increases thehomophilic adhesive activity of L1CAM which reducesits mobility within the neuronal plasma membrane(Kamiguchi and Lemmon 1997). This immobilizationof the L1-molecule in the neuronal membrane isrequired for axon maintenance. L1 also elevates cyclic-AMP levels in neurons via ankyrin-B and mediatesCa?2 dependent interactions (Ooashi and Kamiguchi2009). The phosphorylated state of tyrosine in the L1FIGQY motif causes the release of L1 from ankyrins.Whereas in the dephosphorylated state ankyrins bindsto L1. These reactions are catalysed by tyrosine kinaseenzyme. Tyrosine phosphorylation abolishes ankyrinbinding and also increases L1 lateral mobility andneurite growth. Ankyrins link both L1 and ion channelproteins by coupling them to the spectrin-actincytoskeleton. Spectrin-actin network is linked to
146 Page 10 of 16 Nidhi Sharma et al.

voltage-gated sodium channels, voltage-gated potas-sium channel subunits KCNQ2 and KCNQ3 and L1(Devaux 2010).Mutations in L1CAM leads to intellectual disability
and other neurological symptoms referred to asCRASH syndrome (corpus callosum hypoplasia,retardation, aphasia, spastic paraplegia, and hydro-cephalus) (Fransen et al. 1997). After binding of L1-type proteins with ankyrin, latter guide the proteins tobehave either in an ankyrin-independent manner(neurite outgrowth and axonal pathfinding) or ankyrin-dependent manner (L1’s role at axon initial segments(AIS), paranodal regions, synapses and in dendrites)(Hortsch et al. 2009).
4.2 Ankyrin-G and Nav1.5
Brugada syndrome, an autosomal dominant syndromecharacterized by the elevation of ST-segment alongwith inversion of T-wave and blockage of right bundlebranch (Antzelevitch et al. 2005). Sudden death atnight because of ventricular fibrillation is most com-mon in the affected individual. Around 30% of Bru-gada syndrome is due to the variation in SCN5A, whichencodes Nav1.5 that interacts with ankyrin-G. BothNav1.5 and ankyrin-G (190 kDa) are present at theT-tubule membrane and intercalated disc (Antzelevitch2006; Antzelevitch and Patocskai 2016). Brugadasyndrome mutation E1053K, present in the 9 aminoacid long stretch containing ankyrin binding motif ofNav1.5. By disrupting the association between ankyrin-G and Nav1.5, this mutation disturbs the ankyrin-G-dependent sodium channel which ultimately reducesthe sodium channel current density (INa) (Mohler andBennett 2005; Cunha and Mohler 2009).In neurons, the crosstalk between ankyrin-G and
voltage-gated sodium channel (Nav) is well docu-mented (Mohler et al. 2004). Nav1.5, the primaryvoltage-gated sodium channel in the heart that isrequired for the rapid initial depolarization of the actionpotential of ventricular cardiomyocytes (Bennett andHealy 2008). In rat, the ventricular myocytes lackingankyrin-G has shown decreased expression and local-ization of Nav1.5 and sodium channel current density(INa) (Cui et al. 2002; Mohler et al. 2004). In neurons,ankyrin-G plays a vital role in the functioning of Navchannel where both found co-localize at the node ofRanvier, axon initial segment and neuromuscularjunction. In mice, lacking cerebellar specific ankyrin-Gexhibits marked reduction in Nav channel in the nodeof Ranvier, axon initial segment, and abnormal
distribution of neurofascin, b4-spectrin as well asirregular action potential and ataxia (Mohler 2006;Bennett and Healy 2008; Hashemi et al. 2009; ElRefaey and Mohler 2017).
4.3 Putative role of ankyrins in cross-diseaseassociation
Several genome-wide association studies (GWAS) haveidentified genes expressed in neuronal tissues that areassociated with autoimmune diseases (Lam et al.2017). Recent high-throughput genetics studies identi-fied the association of Ank3 with several neurologicalconditions and celiac disease (Ferreira et al. 2008;Athanasiu et al. 2010; Chen et al. 2013; Xie et al.2014; Senapati et al. 2015, 2016). Celiac disease is anautoimmune disease affecting the duodenum of sus-ceptible individuals, who are exposed to ‘gluten’ anti-gen (Tye-Din et al. 2018). These findings highlightedpossible cross-talk between neuronal factors andimmune health for a disease co-occurrence. Ankyrin-Gexpressed in the mammalian small intestine and acts asan anchoring protein between the epithelial membraneand cytoskeletal proteins (Peters et al. 1995; Cadwellet al. 2015). It provides a direct link between E-cad-herin and cytoskeleton (b-2-spectrin) by recruiting b-2-spectrin to E-cadherin (Kizhatil et al. 2007). E-cadherinbinds to the N-terminal of ankyrin-G which furtherlinks E-cadherin to the b-2-spectrin (figure 7). Thusboth ankyrin-G and b-2-spectrin are critical for thelocalization of E-cadherin and the co-existence of thesethree proteins controls the membrane motility andrequisite for the cell–cell contact (Kizhatil and Bennett2004; Kizhatil et al. 2007; Bennett and Healy 2009;Kumar et al. 2011; Cadwell et al. 2015) (figure 7).Na–K-ATPase is the fundamental protein positioned
in the basolateral membrane of the mammalianepithelium cells which supports cell motility andNa ? dependent co-transportation over the basolateralmembrane (Rodriguez-Boulan and Nelson 1989;Geering 2008). Its efficient and smooth functioning iscrucial for the effective absorption of nutrients andultimately for the overall intestinal health. Ankyrin–spectrin skeleton is mandatory for the appropriateworking of Na–K-ATPase and any change in thisskeleton can affect the performance of Na–K-ATPase(Nelson and Veshnock 1987; Rodriguez-Boulan andNelson 1989; Devarajan et al. 1994; Saha et al. 2015).It has been documented that the decreased expressionof ankyrin in the villus cells of chronically inflamedintestine results in the depolarization of ankyrin-G
Ankyrins in human diseases Page 11 of 16 146

protein (Saha et al. 2015; Klemens et al. 2016).Depolarization of ankyrin-G results in the alteredlocation of Na–K-ATPase which ultimately limits theactivity of Na–K-ATPase in the chronic inflammatorycondition of the intestine. Thus, ankyrin-G plays asignificant role in maintaining the proper position andfunctioning of Na–K-ATPase in chronic intestinalconditions (Saha et al. 2015).It is thus evident that ankyrin-G is critical for the
epithelial membrane integrity and its disturbed func-tional network with spectrin and E-cadherin may alterthe structure anatomy of small intestinal epithelial cells,which further may affect its physiological function andcould initiate disruption of epithelial membrane integ-rity prerequisite for celiac disease.
5. Conclusion
Ankyrins, the anchoring proteins, acts as a connectinglink between the membrane proteins and cytoskeletonelements. The modular architecture of ankyrins enablesthem to bind with the different signaling molecules,cytoskeleton molecules, membrane proteins, etc. Dif-ferent forms of ankyrin (ankyrin-B, ankyrin-G andankyrin-R) and their different isoforms are localized indifferent tissues where they interact with various
associated proteins to maintain cellular structure andfunctional integrity and contribute to the normal func-tioning of the body. Specific genetic variations areknown to alter the functions of ankyrins and contributeto the disease biology. Disruption in the structuralarchitecture of ankyrins disturbs the structural andfunctional integrity of the plasma membrane throughits associated proteins (channels, pumps, signalingproteins), cytoskeleton elements, etc.The involvement of ankyrins and its associated
proteins in various inherited disorders support theimportance of ankyrins in disease biology. The futurechallenge is to elucidate the role of ankyrins in variousneurological and cardiac disorders. The other areawhich needs to be explored is how ankyrins maintainthe membrane integrity, disturbance of which leads tothe collapse of cellular homeostasis. However, sys-tematic molecular studies are warranted to demonstratethe specific roles of ankyrins in human diseases.
Acknowledgements
We acknowledge funding support from DST-SERB,New Delhi, India (ECR/2016/001660), UGC-BSR(F.30/2014(BSR)) and Women Scientist Scheme-A(WOS-A) of Department of Science and Technology
Figure 7. Ankyrin, spectrin and E-cadherin network in the epithelial cells.
146 Page 12 of 16 Nidhi Sharma et al.

(DST), New Delhi, India (SR/WOS-A/LS-333/2018(G). DST-FIST support (SR/FST/LS-I/2017/49-C)to Department of Human Genetics & Molecular Med-icine is acknowledge.
References
Antzelevitch C 2006 Brugada syndrome. Pacing Clin.Electrophysiol. 29 1130
Antzelevitch C and Patocskai B 2016 Brugada syndrome:clinical, genetic, molecular, cellular, and ionic aspects. -Curr. Probl. Cardiol. 41 7–57
Antzelevitch C, Brugada P, Borggrefe M, Brugada J,Brugada R, Corrado D and Shimizu W 2005 Brugadasyndrome: report of the second consensus conference:endorsed by the Heart Rhythm Society and the EuropeanHeart Rhythm Association. Circulation 111 659–670
Athanasiu L, Mattingsdal M, Kahler AK, Brown A,Gustafsson O, Agartz I and Pietilainen OP 2010 Genevariants associated with schizophrenia in a Norwegiangenome-wide study are replicated in a large Europeancohort. J. Psychiatr. Res. 44 748–753
Bennett V 1978 Purification of an active proteolyticfragment of the membrane attachment site for humanerythrocyte spectrin. Chem. Biodivers. 253 2292–2299
Bennett V and Baines AJ 2001 Spectrin and ankyrin-basedpathways: metazoan inventions for integrating cells intotissues. Physiol. Rev. 81 1353–1392
Bennett V and Healy J 2008 Organizing the fluid membranebilayer: diseases linked to spectrin and ankyrin. TrendsMol. Med. 14 28–36
Bennett V and Healy J 2009 Membrane domains based onankyrin and spectrin associated with cell–cell interac-tions. Cold Spring Harb. Perspect. Biol. 1 a003012
Bernstein AD and Parsonnet V 1992 Survey of cardiacpacing in the United States in 1989. Am J. Cardiol. 69331–338
Bouley M, Tian MZ, Paisley K, Shen YC, Malhotra JD andHortsch M 2000 The L1-type cell adhesion moleculeneuroglian influences the stability of neural ankyrin in theDrosophila embryo but not its axonal localization. J.Neurosci. 20 4515–4523
Breeden L and Nasmyth K 1987 Similarity between cell-cycle genes of budding yeast and fission yeast and theNotch gene of Drosophila. Nature 329 651–654
Cadwell CM, Jenkins PM, Bennett V, Kowalczyk A 2015Ankyrin-G inhibits endocytosis of cadherin dimers. J.Biol. Chem. 291 691–704
Cassidy C, Buchy L, Bodnar M, Dell’Elce J, Choudhry Z,Fathalli F and Iyer S 2014 Association of a risk allele ofANK3 with cognitive performance and cortical thicknessin patients with first-episode psychosis. J. PsychiatryNeurosci. 39 31
Chen DT, Jiang X, Akula N, Shugart YY, Wendland JR,Steele CJ and Cheng A 2013 Genome-wide associationstudy meta-analysis of European and Asian-ancestrysamples identifies three novel loci associated with bipolardisorder. Mol. Psychiatry 18 195
Clarkson YL, Perkins EM, Cairncross CJ, Lyndon AR,Skehel PA, and Jackson M 2014 b-III spectrin underpinsankyrin R function in purkinje cell dendritic trees: Proteincomplex critical for sodium channel activity is impairedby SCA5-associated mutations. Hum. Mol. Genet. 233875–3882
Cui Y, Golob J, Kelleher E, Ye Z, Pardoll D and Cheng L2002 Targeting transgene expression to antigen-present-ing cells derived from lentivirus-transduced engraftinghuman hematopoietic stem/progenitor cells. Blood 99399–408.
Cunha SR and Mohler PJ 2009 Ankyrin protein networks inmembrane formation and stabilization. J Cell Mol Med.13 4364–4376
Davis JQ, McLaughlin T and Bennett V 1993 Ankyrin-binding proteins related to nervous system cell adhesionmolecules: candidates to provide transmembrane andintercellular connections in adult brain. J. Cell Biol. 121121–133
Davis LH and Bennett V 1990 Mapping the binding sites ofhuman erythrocyte ankyrin for the anion exchanger andspectrin. Int. J. Biol. Chem. 265 10589–10596
DeGrande S, Nixon D, Koval O, Curran JW, Wright P, WangQ and Anderson ME 2012 CaMKII inhibition rescuesproarrhythmic phenotypes in the model of humanankyrin-B syndrome. Heart Rhythm. 9 2034–2041
Devarajan P, Scaramuzzino DA and Morrow JS 1994Ankyrin binds to two distinct cytoplasmic domains ofNa, K-ATPase alpha subunit. Proc. Natl. Acad. Sci. USA91 2965–2969
Devarajan P, Stabach PR, Mann AS, Ardito T, Kashgarian Mand Morrow JS 1996 Identification of a small cytoplasmicankyrin (AnkG119) in the kidney and muscle that bindsbeta I sigma spectrin and associates with the Golgiapparatus. J. Cell Biol. 133 819–830
Devaux JJ 2010 The C-terminal domain of bIV-spectrin iscrucial for KCNQ2 aggregation and excitability at nodesof Ranvier. J. Physiol. 588 4719–4730
Doctor RB, Chen J, Peters LL, Lux SE and Mandel LJ 1998Distribution of epithelial ankyrin (Ank3) spliceoforms inrenal proximal and distal tubules. Am. J. Physiol. RenalPhysiol. 274 F129–F138
Dubreuil RR and Yu J 1994 Ankyrin and beta-spectrinaccumulate independently of alpha-spectrin in Drosophi-la. PNAS 91 10285–10289
El Refaey MM and Mohler PJ 2017 Ankyrins and spectrinsin cardiovascular biology and disease. Front. Physiol. 8852
Ferreira MA, O’Donovan MC, Meng YA, Jones IR,Ruderfer DM, Jones L and Smoller JW 2008
Ankyrins in human diseases Page 13 of 16 146

Collaborative genome-wide association analysis supportsa role for ANK3 and CACNA1C in bipolar disorder. Nat.Genet. 40 1056
Fransen E, Van Camp G, Vits L and Willems PJ 1997 L1-associated diseases: clinical geneticists divide, moleculargeneticists unite. Hum. Mol. Genet. 6 1625–1632
Garver TD, Ren Q, Tuvia S and Bennett V 1997 Tyrosinephosphorylation at a site highly conserved in the L1family of cell adhesion molecules abolishes ankyrinbinding and increases lateral mobility of neurofascin. J.Cell Biol. 137 703–714.
Geering K 2008 Functional roles of Na, K-ATPase sub-units. Curr. Opin. Nephrol. Hypertens. 17 526–532
Hall TG and Bennett VANN 1987 Regulatory domains oferythrocyte ankyrin. J. Biol. Chem. 262 10537–10545.
Hashemi SM, Hund TJ, Mohler PJ 2009 Cardiac ankyrins inhealth and disease. J. Mol. Cell Cardiol. 47 203–209
Higham, JP, Malik BR, Buhl E, Dawson JM, Ogier AS,Lunnon K and Hodge JJ 2019 Alzheimer’s diseaseassociated genes Ankyrin and Tau cause shortenedlifespan and memory loss in Drosophila. Front. CellNeurosci. https://orcid.org/10.3389/fncel.2019.00260
Hoock TC, Peters LL and Lux SE 1997 Isoforms of ankyrin-3 that lack the NH2-terminal repeats associate with mousemacrophage lysosomes. J. Cell Biol. 136 1059–1070
Hortsch M, Nagaraj K, Godenschwege TA 2009 Theinteraction between L1-type proteins and ankyrins-amaster switch for L1-type CAM function. Cell Mol. Biol.Lett. 14 57
Jenkins SM and Bennett V 2001 Ankyrin-G coordinatesassembly of the spectrin-based membrane skeleton,voltage-gated sodium channels, and L1 CAMs at Purkinjeneuron initial segments. J. Cell. Biol. 155 739–746
Kamiguchi H and Lemmon V 1997 Neural cell adhesionmolecule L1: signaling pathways and growth conemotility. J. Neurosci. Res. 49 1–8
Kizhatil K, and Bennett V (2004) Lateral membranebiogenesis in human bronchial epithelial cells requires190 kDa ankyrin-G. J. Biol. Chem. 279 16706–16714
Kizhatil K, Davis JQ, Davis L, Hoffman J, Hogan BL andBennett V 2007 Ankyrin-G is a molecular partner ofE-cadherin in epithelial cells and early embryos. J. Biol.Chem. 282 26552–26561
Klemens CA, Edinger RS, Kightlinger L, Liu X andButterworth MB 2016 Ankyrin G expression regulatesapical delivery of the epithelial sodium channel(ENaC). J. Biol. Chem. 292 375–385
Koenig SN and Mohler PJ 2017 The evolving role ofankyrin-B in cardiovascular disease. Heart Rhythm 141884–1889
Kordeli E, Lambert S and Bennett V 1995 Ankyrin A newankyrin gene with neural-specific isoforms localized at theaxonal initial segment and node of Ranvier. J. Biol.Chem. 270 2352–2359
Kumar S, Park SH, Cieply B, Schupp J, Killiam E, RimmDL and Frisch SM 2011 A pathway for the control ofanoikis-sensitivity by E-cadherin and EMT. Mol. Cell.Biol. 31 4036–4051
Lam M, Trampush JW, Yu J, Knowles E, Davies G, LiewaldDC and Christoforou A 2017 Large-scale cognitiveGWAS meta-analysis reveals tissue-specific neuralexpression and potential nootropic drug targets. CellRep. 21 2597–2613
Lambert S, Yu H, Prchal JT, Lawler J, Ruff P, Speicher Dand Palek J 1990 cDNA sequence for human erythrocyteankyrin. Proc. Natl. Acad. Sci. USA 87 1730–1734
Le Scouarnec S, Bhasin N, Vieyres C, Hund TJ, Cunha SR,Koval O and Song LS 2008 Dysfunction in ankyrin-B-dependent ion channel and transporter targeting causeshuman sinus node disease. Proc. Natl. Acad. Sci. USA105 15617–15622
Li J, Mahajan A, Tsai MD 2006 Ankyrin repeat: a uniquemotif mediating protein-protein interactions. Biochemistry45 15168–15178
Little SC, Curran J, Makara MA, Kline CF, Ho HT, Xu Zand Carnes CA 2015 Protein phosphatase 2A regulatorysubunit B56a limits phosphatase activity in the heart. Sci.Signal. 8 ra72–ra72
Lunnon K, Smith R, Hannon E, De Jager PL, Srivastava G,Volta M and Condliffe D 2014 Methylomic profilingimplicates cortical deregulation of ANK1 in Alzheimer’sdisease. Nat. Neurosci. 17 1164
Luoni A, Massart R, Nieratschker V, Nemoda Z, Blasi G,Gilles M and Rizzo G 2016 Ankyrin-3 as a molecularmarker of early-life stress and vulnerability to psychiatricdisorders. Transl. Psychiatry 6 e943
Lux SE, John KM and Bennett V 1990 Analysis of cDNAfor human erythrocyte ankyrin indicates a repeatedstructure with homology to tissue-differentiation andcell-cycle control proteins. Nature 344 36–42
Michaely P, Tomchick DR, Machius M and Anderson RG2002 Crystal structure of a 12 ANK repeat stack fromhuman ankyrinR. EMBO J. 21 6387–6396
Mohler PJ 2006 Ankyrins and human disease: what theelectrophysiologist should know. J. Cardiovasc. Electro-physiol. 17 1153–1159
Mohler PJ and Bennett V 2005 Ankyrin-based cardiacarrhythmias: a new class of channelopathies due to loss ofcellular targeting. Curr. Opin. Psychiatry 20 189–193
Mohler PJ, Gramolini AO and Bennett V 2002 Ankyrins. J.Cell. Sci. 115 1565–1566
Mohler PJ, Healy JA, Xue H, Puca AA, Kline CF,Allingham RR and Bennett V 2007 Ankyrin-B syndrome:enhanced cardiac function balanced by risk of cardiacdeath and premature senescence. PLoS ONE 2 e1051
Mohler PJ, Rivolta I, Napolitano C, LeMaillet G, Lambert S,Priori SG and Bennett V 2004 Nav1. 5 E1053K mutationcausing Brugada syndrome blocks binding to ankyrin-G
146 Page 14 of 16 Nidhi Sharma et al.

and expression of Nav1. 5 on the surface of cardiomy-ocytes. Proc. Natl. Acad. Sci. USA 101 17533–17538
Mohler PJ, Schott JJ, Gramolini AO, Dilly KW, GuatimosimS, Song LS and Lederer WJ 2003 Ankyrin-B mutationcauses type 4 long-QT cardiac arrhythmia and suddencardiac death. Nature 421 634–639
Mosavi LK, Cammett TJ, Desrosiers DC and Peng ZY 2004The ankyrin repeat as molecular architecture for proteinrecognition. Protein Sci. 13 1435–1448
Musarella M, Alcaraz G, Caillol G, Boudier JL, Couraud Fand Autillo-Touati A 2006 Expression of Nav1. 6 sodiumchannels by Schwann cells at neuromuscular junctions:role in the motor endplate disease phenotype. Glia 5313–23
Nelson WJ and Veshnock PJ 1987 Ankyrin binding to(Na ?? K ?) ATPase and implications for the organi-zation of membrane domains in polarized cells. Nature328 533
Ooashi N and Kamiguchi H 2009 The cell adhesionmolecule L1 controls growth cone navigation viaankyrinB-dependent modulation of cyclic AMP. J. Neu-rosci. 63 224–226
Ortega S, Malumbres M and Barbacid M 2002 CyclinD-dependent kinases, INK4 inhibitors and cancer. Bio-chim. Biophys. Acta Rev. Cancer. 1602 73–87
Otsuka AJ, Franco R, Yang B, Shim KH, Tang LZ, ZhangYY and Wheaton VI 1995 An ankyrin-related gene (unc-44) is necessary for proper axonal guidance in Caenorhab-ditis elegans. J. Cell. Biol. 129 1081–1092
Otto E, Kunimoto M, McLaughlin T and Bennett V 1991Isolation and characterization of cDNAs encoding humanbrain ankyrins reveal a family of alternatively splicedgenes. J. Cell. Physiol. 114 241–253
Pereira AC, Gray JD, Kogan JF, Davidson RL, Rubin TG,Okamoto M and McEwen BS 2017 Age and Alzheimer’sdisease gene expression profiles reversed by the glutamatemodulator riluzole. Mol. Psychiatry. 22 296
Peters LL, John KM, Lu FM, Eicher EM, Higgins A,Yialamas M and Lux SE 1995 Ank3 (epithelial ankyrin),a widely distributed new member of the ankyrin genefamily and the major ankyrin in kidney, is expressed inalternatively spliced forms, including forms that lack therepeat domain. J. Cell. Biol. 130 313–330
Randon J, del Giudice EM, Bozon M, Perrotta S, De VivoM, Iolascon A and Morle L 1997 Frequent de novomutations of the ANK1 gene mimic a recessive mode oftransmission in hereditary spherocytosis: three newANK1 variants: ankyrins Bari, Napoli II and Anzio. Br.J. Haematol. 96 500–506
Roberts JD, Murphy NP, Hamilton RM, Lubbers ER, JamesCA, Kline CF and El-Refaey M 2019 Ankyrin-Bdysfunction predisposes to arrhythmogenic cardiomyopa-thy and is amenable to therapy. J. Clin. Invest. 1293171–3184
Rodriguez-Boulan E and Nelson WJ 1989 Morphogenesis ofthe polarized epithelial cell phenotype. Science 245718–725
Rubtsov AM and Lopina OD 2000 Ankyrins. FEBSLett. 482 1–5
Saha P, Manoharan P, Arthur S, Sundaram S, Kekuda R,Sundaram U 2015 Molecular mechanism of regulation ofvillus cell Na-K-ATPase in the chronically inflamedmammalian small intestine. Biochim. Biophys. Acta1848 702–711
Saifetiarova J, Taylor AM and Bhat MA 2017 Early and lateloss of the cytoskeletal scaffolding protein, ankyrin Greveals its role in maturation and maintenance of nodes ofRanvier in myelinated axons. J. Neurosci. 37 2524–2538
Schott JJ, Charpentier F, Peltier S, Foley P, Drouin E,Bouhour JB and Le Marec H 1995 Mapping of a gene forlong QT syndrome to chromosome 4q25–27. Am. J. Hum.Genet. 57 1114
Senapati S, Gutierrez-Achury J, Sood A, Midha V, Szperl A,Romanos J and Wijmenga C 2015 Evaluation of Euro-pean coeliac disease risk variants in a north Indianpopulation. BMC Med. Genomics 23 530
Senapati S, Sood A, Midha V, Sood N, Sharma S, Kumar Land Thelma BK 2016 Shared and unique common geneticdeterminants between pediatric and adult celiac dis-ease. BMC Med Genomics. 9 44
Smith SA, Sturm AC, Curran J, Kline CF, Little SC, BonillaIM and Webb TR 2015 Dysfunction in the bII spectrin–dependent cytoskeleton underlies human arrhythmia. Cir-culation 131 695–708
Swayne LA, Murphy NP, Asuri S, Chen L, Xu X, McIntoshS and Sanatani S 2017 Novel variant in the ANK2membrane-binding domain is associated with Ankyrin-Bsyndrome and structural heart disease in a first nationspopulation with a high rate of long QT syndrome. Circ.Cardiovasc. Interv. 10 e001537
Tuvia S, Garver TD and Bennett V 1997 The phosphory-lation state of the FIGQY tyrosine of neurofascindetermines ankyrin-binding activity and patterns of cellsegregation. Proc. Natl. Acad. Sci. USA India 9412957–12962
Tye-Din JA, Galipeau HJ and Agardh D 2018 Celiacdisease: a review of current concepts in pathogenesis,prevention, and novel therapies. Front. Pediatr. 6 350
Wang C, Wei Z, Chen K, Ye F, Yu C, Bennett V and ZhangM 2014 Structural basis of diverse membrane targetrecognitions by ankyrins. Elife 3 e04353
Wang C, Yu C, Ye F, Wei Z and Zhang M 2012 Structure ofthe ZU5-ZU5-UPA-DD tandem of ankyrin-B revealsinteraction surfaces necessary for ankyrin function. Proc.Natl. Acad. Sci. USA 109 4822–4827
Wilders R and Verkerk AO 2018 Long QT syndrome andsinus bradycardia-a mini review. Front Cardiovasc.Med. 5 106
Ankyrins in human diseases Page 15 of 16 146

Xie T, Deng L, Mei P, Zhou Y, Wang B, Zhang J and Xu R2014 A genome-wide association study combining path-way analysis for typical sporadic amyotrophic lateralsclerosis in Chinese Han populations. Neurobiol. Aging35 1778-e9.
Zhang H, Scheirer DC, Fowle WH and Goodman HM 1992Expression of antisense or sense RNA of an ankyrinrepeat-containing gene blocks chloroplast differentiationin arabidopsis. Plant Cell. 4 1575–1588
Zhang X and Bennett V 1998 Restriction of 480/270-kDankyrin G to axon proximal segments requires multipleankyrin G-specific domains. J. Cell. Bio. 1421571–1581
Zhu W, Wang C, Hu J, Wan R, Yu J, Xie J and Chen L 2018Ankyrin-B Q1283H variant linked to arrhythmias via lossof local protein phosphatase 2A Activity Causes Ryan-odine Receptor Hyperphosphorylation. Circulation 1382682–2697
Corresponding editor: KUNDAN SENGUPTA
146 Page 16 of 16 Nidhi Sharma et al.
Top Related
Copyright © 2022 FDOKUMEN

