







Bahasa
Halaman
Hukum


ORIGINALARTICLE
Historical biogeography of an Indo-Pacificpasserine bird family (Pachycephalidae):different colonization patterns in theIndonesian and Melanesian archipelagos
Knud A. Jønsson1,2*, Rauri C. K. Bowie2, Robert G. Moyle3, Les Christidis4,5,
Janette A. Norman5,6, Brett W. Benz3 and Jon Fjeldsa1
1Vertebrate Department, Zoological Museum,
University of Copenhagen, Universitetsparken
15, DK-2100 Copenhagen Ø, Denmark,2Museum of Vertebrate Zoology and
Department of Integrative Biology, 3101 Valley
Life Science Building, University of California,
Berkeley, CA 94720-3160, USA, 3Natural
History Museum and Biodiversity Research
Center and Department of Ecology and
Evolutionary Biology, University of Kansas, KS
66045-7561, USA, 4Division of Research and
Collections, Australian Museum, 6 College
Street, Sydney, NSW 2010, Australia,5Department of Genetics, University of
Melbourne, Parkville, Vic. 3052, Australia,6Sciences Department, Museum Victoria, GPO
Box 666, Melbourne, Vic. 3001, Australia
*Correspondence: Knud A. Jønsson, Vertebrate
Department, Zoological Museum, University of
Copenhagen, Universitetsparken 15, DK-2100
Copenhagen Ø, Denmark.
E-mail: [email protected]
ABSTRACT
Aim We use molecular-based phylogenetic methods and ancestral area
reconstructions to examine the systematic relationships and biogeographical
history of the Indo-Pacific passerine bird family Pachycephalidae (whistlers).
Analysed within an explicit spatiotemporal framework, we elucidate distinct
patterns of diversification across the Melanesian and Indonesian archipelagos and
explore whether these results may be explained by regional palaeogeological
events. We further assess the significance of upstream colonization and its role in
species accumulation within the region.
Location The Indo-Pacific region, with an emphasis on the archipelagos on
either side of the Australo-Papuan continent.
Methods We used three nuclear and two mitochondrial markers to construct a
molecular phylogenetic hypothesis of the Pachycephalidae by analysing 35 of the
49 species known to belong to the family. The programs diva and MrBayes were
used to reconstruct ancestral area relationships and to examine biogeographical
relationships across the family, and beast was implemented to assess the timing
of dispersal events.
Results We constructed a molecular phylogenetic hypothesis for the
Pachycephalidae and estimated divergence times and ancestral area relationships.
Different colonization patterns are apparent for the Pachycephalidae in the
Indonesian and the Melanesian archipelagos. The Indonesian archipelago was
colonized numerous times, whereas one or two colonizations of the Melanesian
archipelagos account for the entire diversity of that region. After initial
colonization of the Melanesian archipelagos some whistler species recolonized
Australia and may have commenced a second round of colonization into
Melanesia.
Main conclusions The contrasting dispersal patterns of whistlers in
archipelagos on either side of the Australo-Papuan continent are congruent
with the arrangement and history of islands in each of the regions and
demonstrate that knowledge of palaeogeography is important for an
understanding of evolutionary patterns in archipelagos. We also highlight that
recolonization of continents from islands may be more common than has
previously been assumed.
Keywords
Colonization, dispersal, divergence, Melanesia, Pachycephalidae, Pacific,
Passeriformes, Pleistocene, Pliocene, Wallacea.
Journal of Biogeography (J. Biogeogr.) (2010) 37, 245–257
ª 2009 Blackwell Publishing Ltd www.blackwellpublishing.com/jbi 245doi:10.1111/j.1365-2699.2009.02220.x

INTRODUCTION
Islands have traditionally been regarded as evolutionary dead
ends, where populations that are established through dispersal
from biologically diverse continental biotas rarely contribute
to broader regional species generation via secondary coloni-
zation events beyond the island system (Darwin, 1859;
Wallace, 1876; MacArthur & Wilson, 1963, 1967). Conse-
quently, island biodiversity has been viewed in the context of
dynamic interactions between colonization and extinction,
with the direction and rate of these phenomena influenced by
population size, productivity, vagility, relative species diversity
of source and sink communities, and habitat conditions
(MacArthur & Wilson, 1963, 1967; Diamond, 1977).
Recent advances in the understanding of biodiversity
dynamics in island systems have shed new light on the
interface of these mechanisms (colonization, speciation,
extinction) and the role they may play in shaping patterns of
biodiversity in island environments (reviewed in Whittaker &
Fernandez-Palacios, 2007; Emerson & Gillespie, 2008; Gillespie
et al., 2008). Combined with further novel insight from
detailed molecular-based phylogenies examined within a
geographical framework, evidence of numerous secondary
colonization events (from islands back to the mainland) is now
challenging island biogeography dogma, suggesting that island
systems may in fact be recognized as a potential source for the
build-up of continental biodiversity (Filardi & Moyle, 2005;
Bellemain & Ricklefs, 2008). As such, understanding the
evolutionary histories of island biotas will probably be integral
to interpreting broader regional biogeographical patterns and
processes, and is thus one of the major challenges in modern
macroecology and biogeography (Jetz et al., 2004; Storch et al.,
2006; Fjeldsa et al., 2007; Bellemain & Ricklefs, 2008).
The Indo-Pacific region represents a particularly rich model
system for such work, as knowledge and understanding of the
geological history in this complex region has recently seen
significant advances (Hall, 1998, 2002; Hall & Holloway, 1998)
and few molecular-based studies have investigated distribu-
tions across the entire Indo-Pacific region (except Moyle et al.,
2009). Using molecular, palaeogeological and biogeographical
evidence, we aim to investigate biogeographical patterns and
the timing of diversification in the avian family Pachycepha-
lidae (whistlers), which includes 49 species distributed across
the Indo-Pacific realm (Gill & Wright, 2006). Within this
region, major archipelagos are found on either side of the
Australo-Papuan continent, with mobile oceanic island arcs to
the east and a mosaic of microplates and young volcanics
compressed in Wallacea to the west. The geological history of
each subregion is complex with respect to their composite
origins, each requiring a synoptic timeline of their respective
orogeny to set the geographical stage for analysis of diversi-
fication in the Pachycephalidae.
The present-day centre of pachycephalid diversity lies in
New Guinea’s extensive hill forests (Fig. 1), with a secondary
centre of diversity on the Australian continent (Boles, 2007).
The shared geological history of these regions began in the late
Mesozoic with the collision of the Pacific and Australian plates;
however, initial formation of the New Guinea orogen via
mountain uplift and terrane accretion along the northern
margin of the Australian continental shelf is thought to have
begun more recently in the late Oligocene (Hamilton, 1979;
Audley-Charles, 1991; Pigram & Symonds, 1991). Consider-
able uncertainty surrounds the timing of accretion history as
well as the rates of subsequent uplift across the 32 distinct
terranes that comprise the New Guinea orogen; however, a
general consensus drawing on diverse lines of evidence suggests
that the salient topographic features of the extensive central
dividing ranges and the Papuan peninsula were in place by the
Pliocene. Accreted North Coast terranes of oceanic affinity as
well as the Australian continental fragments that form New
Guinea’s Birds Head region may have completed docking more
recently, in the late Pliocene (Audley-Charles, 1991; Pigram &
Symonds, 1991; Hall, 1998, 2002).
To the east of the Australo-Papuan continent, the Melane-
sian arcs comprising the Solomons, New Britain, New Ireland,
Tonga and Fiji are thought to have rifted from the eastern
Figure 1 Worldmap representation of the
present distribution of members of the
Pachycephalidae. Each 1� square represents
110 km · 110 km at the equator. The
maximum number of species in a square
(warmest colour) is 13.
K. A. Jønsson et al.
246 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

Australian margin in the mid-Palaeogene, migrating north-east
into the Pacific before progressing back towards the Australian
margin; however, an alternative hypothesis suggests that these
arcs may have originated within the Pacific and subsequently
migrated to their present position (Yan & Kroenke, 1993; Hall,
2002). Regardless, their present-day close proximity to New
Guinea appears to be a gradual development initiated in the
mid-Miocene.
By comparison, the geological history of the Indonesian
archipelago and surrounding island systems to the north and
west of New Guinea’s Birds Head region is considerably more
complex, with a mixture of old and young terranes juxtaposed
in close proximity (Hall, 1998, 2002). The Philippine–Izu–
Bonin archipelago extends south to the Molucca Sea, where
ophiolitic terranes of the Halmahera Arc meet Gondwanan
continental fragments from the Australian Craton along the
Seram Trough, collectively forming the Moluccas. The Sangihe
Arc lies to the west of the Molucca Sea, and although its
collision with the Halmahera Arc initiated in the late Pliocene,
the Sangihe volcanics probably date to the mid-Miocene given
their affinity with north Sulawesi (Hall, 2002). Reconstructing
the evolutionary history of the Banda Sea region is problematic
owing to the diversity of the terranes present and the
potentially large and recent displacements involved (Hall,
2002). The Lesser Sunda Islands were formed by the emergence
of the Banda arc, which consists of an inner volcanic arc
(Wetar west to Bali) that has been active since the Miocene,
and an outer non-volcanic arc (Sumba, Timor and Tanimbar)
that has formed more recently in the Quaternary (Hall, 1998,
2002). Although improved isotopic, palaeomagnetic and GPS
tectonic data have enabled more rigorous plate tectonic models
in recent years, thus providing a clearer understanding of the
geographical history across the Indo-Pacific region, the
spatiotemporal uncertainty inherent in these data warrants
caution in applying such information to explicitly dated
biogeographical analyses.
Herein, we investigate the phylogenetic relationships of the
Pachycephalidae, examining the patterns and timing of
whistler diversification across the Indo-Pacific to: (1) shed
light on the phylogenetic relationships and general biogeo-
graphical patterns within the Pachycephalidae, (2) test whether
branching patterns and timing of major radiations coincide
with regional palaeogeographical histories, and (3) examine
the role of dispersal in lineage accumulation between the
distinctive Indonesian and Melanesian archipelagos and the
Australo-Papuan continent, in order to highlight potential
colonization events and assess how this phenomenon may have
affected the evolutionary history of the Pachycephalidae.
MATERIALS AND METHODS
Taxon sampling
Recent molecular studies have revealed that several species
historically assigned to the Pachycephalidae represent inde-
pendent lineages outside of the family, or belong to other bird
families altogether. Aleadryas rufinucha, Oreoica gutturalis and
Pitohui cristatus form a small assemblage outside of, but still
somewhat close to, the Pachycephalidae (Jønsson et al., 2008b;
Norman et al., 2009). Hylocitrea bonensis was surprisingly
found to be within a different parvorder (Jønsson et al., 2008a)
and is now robustly placed within the Bombycillidae (Spellman
et al., 2008). Rhagologus leucostigma, Pachycare flavogrisea and
Eulacstoma nigropectus are also known to not be closely related
to the Pachycephalidae but rather form part of the broader
radiation of corvoids (Jønsson et al., 2008b; Norman et al.,
2009, in press). Thus, the circumscription of the Pachyceph-
alidae needs to be redefined in order to properly infer
biogeographical patterns.
Based on our current knowledge, the avian family Pachy-
cephalidae encompasses all taxa assigned to the genera
Pachycephala and Colluricincla as well as Pitohui ferrugineus,
Pitohui nigrescens and the monotypic Coracornis raveni. With
this circumscription in mind, our sampling includes 35 of the
49 species (Table 1) known to belong to the family (Gill &
Wright, 2006). The lack of complete sampling reflects the
absence of modern ornithological field expeditions for much of
this region. As such, DNA extraction from museum study
skins was performed for 15 key taxa to help remedy this
sampling deficiency.
Although lacking genetic samples from nearly 30% of the taxa
in the family may appear to severely limit the biogeographical
conclusions that can be drawn, our sampling includes represen-
tatives of all major species groups identified in previous
taxonomic studies from all major biogeographical regions within
the family’s range. Three species (Pachycephala fulvotincta,
Pachycephala mentalis and Pachycephala graeffii) have previously
been considered subspecies of the Pachycephala pectoralis com-
plex(Galbraith,1956),whichcomprises66namedtaxa.Although
this complex appears to represent a species group rather than a
single species, the uniformity in morphological characters and
ecology give reason to believe that they all are relatively closely
related. Six additional species that we lack have restricted
distributions in New Guinea (Pachycephala aurea, Pachycephala
monacha, Pachycephala meyeri and Colluricincla umbrina), the
Philippines (Pachycephala homeyeri, which is considered closely
related to other Philippine species) and Australia (Pachycephala
rufogularis, which is probably closely related to Pachycephala
inornata). Although we lack Pachycephala johni, several checklists
(e.g. Dickinson, 2003) consider this taxon to be a subspecies of
Pachycephala griseonota. The remaining three missing taxa are
Pachycephala flavifrons of Samoa, Pachycephala jacquinoti of
Tonga and Pachycephala implicata of the Solomon Islands, which
are often considered to be part of a superspecies complex that
includes other Pacific whistlers and these taxa are sometimes
treated as subspecies of Pachycephala pectoralis (Boles, 2007).
Laboratory procedures
Three nuclear gene regions – myoglobin intron-2 (Myo2),
ornithine decarboxylase (ODC) introns 6 to 7, and glyceral-
dehyde-3-phosphodehydrogenase (GAPDH) intron-11 – and
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 247ª 2009 Blackwell Publishing Ltd
two mitochondrial markers – NADH dehydrogenase subunit
2 (ND2) and subunit 3 (ND3) – were sequenced to
estimate phylogenetic relationships among members of the
Pachycephalidae. For each gene and taxon, multiple sequence
fragments were obtained by sequencing with various primer
combinations (see below).
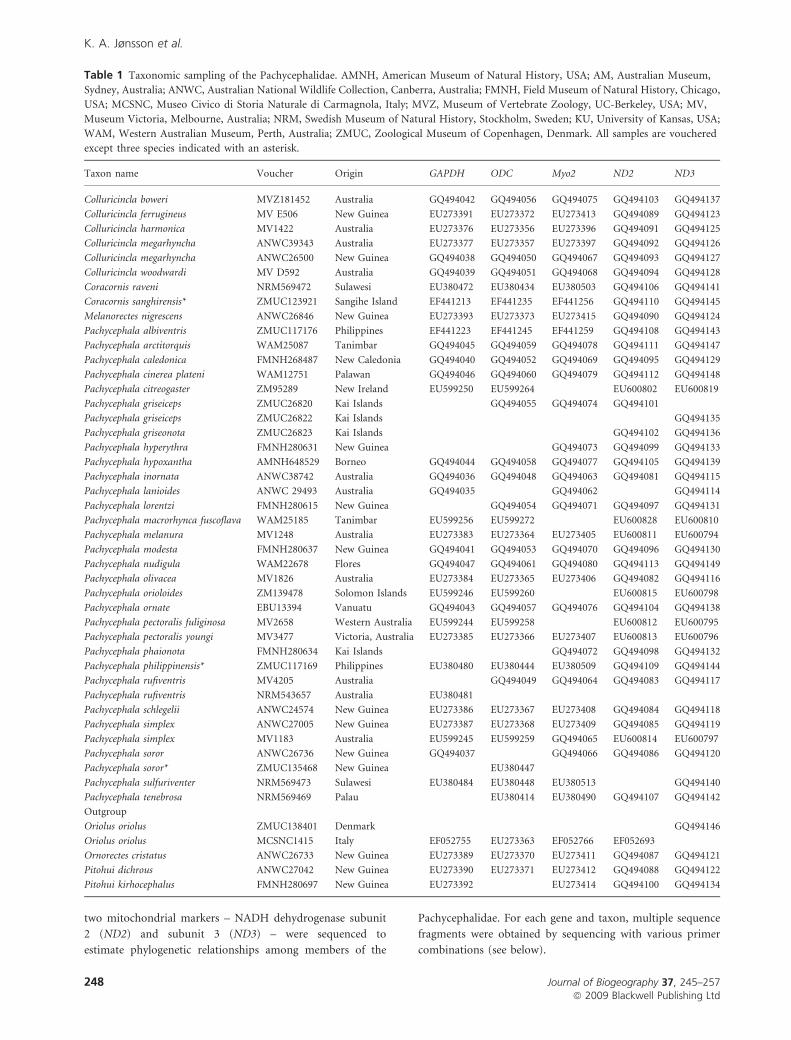
Table 1 Taxonomic sampling of the Pachycephalidae. AMNH, American Museum of Natural History, USA; AM, Australian Museum,
Sydney, Australia; ANWC, Australian National Wildlife Collection, Canberra, Australia; FMNH, Field Museum of Natural History, Chicago,
USA; MCSNC, Museo Civico di Storia Naturale di Carmagnola, Italy; MVZ, Museum of Vertebrate Zoology, UC-Berkeley, USA; MV,
Museum Victoria, Melbourne, Australia; NRM, Swedish Museum of Natural History, Stockholm, Sweden; KU, University of Kansas, USA;
WAM, Western Australian Museum, Perth, Australia; ZMUC, Zoological Museum of Copenhagen, Denmark. All samples are vouchered
except three species indicated with an asterisk.
Taxon name Voucher Origin GAPDH ODC Myo2 ND2 ND3
Colluricincla boweri MVZ181452 Australia GQ494042 GQ494056 GQ494075 GQ494103 GQ494137
Colluricincla ferrugineus MV E506 New Guinea EU273391 EU273372 EU273413 GQ494089 GQ494123
Colluricincla harmonica MV1422 Australia EU273376 EU273356 EU273396 GQ494091 GQ494125
Colluricincla megarhyncha ANWC39343 Australia EU273377 EU273357 EU273397 GQ494092 GQ494126
Colluricincla megarhyncha ANWC26500 New Guinea GQ494038 GQ494050 GQ494067 GQ494093 GQ494127
Colluricincla woodwardi MV D592 Australia GQ494039 GQ494051 GQ494068 GQ494094 GQ494128
Coracornis raveni NRM569472 Sulawesi EU380472 EU380434 EU380503 GQ494106 GQ494141
Coracornis sanghirensis* ZMUC123921 Sangihe Island EF441213 EF441235 EF441256 GQ494110 GQ494145
Melanorectes nigrescens ANWC26846 New Guinea EU273393 EU273373 EU273415 GQ494090 GQ494124
Pachycephala albiventris ZMUC117176 Philippines EF441223 EF441245 EF441259 GQ494108 GQ494143
Pachycephala arctitorquis WAM25087 Tanimbar GQ494045 GQ494059 GQ494078 GQ494111 GQ494147
Pachycephala caledonica FMNH268487 New Caledonia GQ494040 GQ494052 GQ494069 GQ494095 GQ494129
Pachycephala cinerea plateni WAM12751 Palawan GQ494046 GQ494060 GQ494079 GQ494112 GQ494148
Pachycephala citreogaster ZM95289 New Ireland EU599250 EU599264 EU600802 EU600819
Pachycephala griseiceps ZMUC26820 Kai Islands GQ494055 GQ494074 GQ494101
Pachycephala griseiceps ZMUC26822 Kai Islands GQ494135
Pachycephala griseonota ZMUC26823 Kai Islands GQ494102 GQ494136
Pachycephala hyperythra FMNH280631 New Guinea GQ494073 GQ494099 GQ494133
Pachycephala hypoxantha AMNH648529 Borneo GQ494044 GQ494058 GQ494077 GQ494105 GQ494139
Pachycephala inornata ANWC38742 Australia GQ494036 GQ494048 GQ494063 GQ494081 GQ494115
Pachycephala lanioides ANWC 29493 Australia GQ494035 GQ494062 GQ494114
Pachycephala lorentzi FMNH280615 New Guinea GQ494054 GQ494071 GQ494097 GQ494131
Pachycephala macrorhynca fuscoflava WAM25185 Tanimbar EU599256 EU599272 EU600828 EU600810
Pachycephala melanura MV1248 Australia EU273383 EU273364 EU273405 EU600811 EU600794
Pachycephala modesta FMNH280637 New Guinea GQ494041 GQ494053 GQ494070 GQ494096 GQ494130
Pachycephala nudigula WAM22678 Flores GQ494047 GQ494061 GQ494080 GQ494113 GQ494149
Pachycephala olivacea MV1826 Australia EU273384 EU273365 EU273406 GQ494082 GQ494116
Pachycephala orioloides ZM139478 Solomon Islands EU599246 EU599260 EU600815 EU600798
Pachycephala ornate EBU13394 Vanuatu GQ494043 GQ494057 GQ494076 GQ494104 GQ494138
Pachycephala pectoralis fuliginosa MV2658 Western Australia EU599244 EU599258 EU600812 EU600795
Pachycephala pectoralis youngi MV3477 Victoria, Australia EU273385 EU273366 EU273407 EU600813 EU600796
Pachycephala phaionota FMNH280634 Kai Islands GQ494072 GQ494098 GQ494132
Pachycephala philippinensis* ZMUC117169 Philippines EU380480 EU380444 EU380509 GQ494109 GQ494144
Pachycephala rufiventris MV4205 Australia GQ494049 GQ494064 GQ494083 GQ494117
Pachycephala rufiventris NRM543657 Australia EU380481
Pachycephala schlegelii ANWC24574 New Guinea EU273386 EU273367 EU273408 GQ494084 GQ494118
Pachycephala simplex ANWC27005 New Guinea EU273387 EU273368 EU273409 GQ494085 GQ494119
Pachycephala simplex MV1183 Australia EU599245 EU599259 GQ494065 EU600814 EU600797
Pachycephala soror ANWC26736 New Guinea GQ494037 GQ494066 GQ494086 GQ494120
Pachycephala soror* ZMUC135468 New Guinea EU380447
Pachycephala sulfuriventer NRM569473 Sulawesi EU380484 EU380448 EU380513 GQ494140
Pachycephala tenebrosa NRM569469 Palau EU380414 EU380490 GQ494107 GQ494142
Outgroup
Oriolus oriolus ZMUC138401 Denmark GQ494146
Oriolus oriolus MCSNC1415 Italy EF052755 EU273363 EF052766 EF052693
Ornorectes cristatus ANWC26733 New Guinea EU273389 EU273370 EU273411 GQ494087 GQ494121
Pitohui dichrous ANWC27042 New Guinea EU273390 EU273371 EU273412 GQ494088 GQ494122
Pitohui kirhocephalus FMNH280697 New Guinea EU273392 EU273414 GQ494100 GQ494134
K. A. Jønsson et al.
248 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

Primer pairs used for amplification were: Lmet (Hackett,
1996)/H6312 (Cicero & Johnson, 2001) for ND2; ND3-
L10755/ND3-H11151 (Chesser, 1999) for ND3; Myo2 (Slade
et al., 1993)/Myo-cora2R (Jønsson et al., 2008a) and Myo-
coraF1 (Jønsson et al., 2008a)/Myo3F (Heslewood et al., 1998)
for Myo2; OD6/OD8 (Allen & Omland, 2003) for ODC; and
G3P13/G3P14b (Fjeldsa et al., 2003) for GAPDH.
Thermocycling conditions included a hot start at 95 �C, an
initial denaturation step at 95 �C for 5 min, followed by 32
cycles at 95 �C for 40 s, 54–63 �C for 40 s, and 72 �C for 60 s
and completion by a final extension at 72 �C for 8 min. One
microlitre of polymerase chain reaction (PCR) products was
electrophoresed on a 1.5% agarose gel and visualized under
UV light with ethidium bromide to check for correct fragment
size and to control for the specificity of the amplifications. The
PCR products were purified using ExoSap enzymes (exonu-
clease and shrimp alkaline phosphatase). Purified PCR prod-
ucts were cycle-sequenced using Big Dye terminator chemistry
(ABI; Applied Biosystems, Foster City, CA, USA) in both
directions with the same primers used for PCR amplifications,
with the exception of the primer G3P13 which was replaced by
G3PintL1 (Fjeldsa et al., 2003), and run on an automated ABI
3730 DNA sequencer. Corresponding laboratory procedures
for the sequencing of ancient DNA from study skins are
detailed in Irestedt et al. (2006). Additional internal primers
for tissues taken from museum skins are specified in Jønsson
et al. (2008a) (for Myo2 and GAPDH), Jønsson et al. (2008c)
(for ND2) and in Irestedt et al. (2006) (for ODC). Sequences
were assembled with SeqMan II (DNASTAR Inc., Madison,
WI, USA). Nucleotides that could not be determined with
certainty (mostly heterozygous sites in nuclear loci) were
coded with the appropriate IUPAC code. GenBank accession
numbers are provided in Table 1.
Sequence alignment and phylogenetic analyses
Sequences were aligned using MegAlign (DNASTAR Inc.)
followed by a few manual adjustments. Coding genes (ND2
and ND3) were checked for the presence of stop codons or
insertion/deletion events that would have disrupted the
reading frame. All alignments are available from the first
author upon request. We used Bayesian inference (BI) (e.g.
Holder & Lewis, 2003; Huelsenbeck & Ronquist, 2003), as
implemented in MrBayes 3.1.2 (Huelsenbeck et al., 2001;
Ronquist & Huelsenbeck, 2003), to estimate phylogenetic
relationships. Substitution models were determined with
MrModeltest 2.0 (Nylander, 2004), using the Akaike
information criterion (AIC) (Akaike, 1973; Posada & Buckley,
2004). Model parameters were permitted to vary between the
seven partitions [GAPDH, ODC, Myo2, first, second and
third mitochondrial DNA (mtDNA) codon positions and
transfer RNA (tRNA)] in the mixed-model Bayesian analyses
(Ronquist & Huelsenbeck, 2003; Nylander, 2004). In all
MrBayes analyses, four Metropolis-coupled Markov chain
Monte Carlo (MCMC) simulations, one cold and three
heated, were run for 5–15 million iterations with trees
sampled every 1000 iterations. The burn-in was graphically
estimated using awty (Wilgenbusch et al., 2004; Nylander
et al., 2008) and this program was also used to graphically
assess whether the MCMC analysis had run for long enough,
such that tree topologies were being sampled in proportion
to their true posterior probability distribution. Two inde-
pendent Bayesian runs initiated from random starting trees
were performed for each data set, and the log-likelihood
values and posterior probabilities of nodes from each run
were compared to aid in assessing stationarity.
Maximum likelihood (ML) analyses were performed using
garli 0.95 (Zwickl, 2006). Five independent analyses (15 mil-
lion generations for the combined analysis and 5 million
generations for the individual partitions) were performed and
nodal support was evaluated with 100 nonparametric boot-
strap pseudoreplications.
Dating analyses
Establishing a general timeframe for evolution of a group can
allow one to reject hypotheses about factors that influenced
the process of diversification. Unfortunately, the fossil record
provides no calibration points within or close to the
Pachycephalidae, so we used two alternative approaches to
estimate divergence times in the group. First, we assigned
island ages in the region to nodes on the phylogeny and then
used those calibrated nodes to estimate the ages of other
nodes. Second, we applied to our data an ND2 divergence
rate calculated for another group of island-dwelling passe-
rines. Each of these methods includes numerous caveats and
assumptions (see below), but our interest was in seeing if two
independent methods returned similar results, thus providing
some degree of support for the general timeframe of
diversification.
The following geological events are relevant as calibration
points for the dating analysis. Islands in the Sangihe Arc were
formed by volcanism beginning in the early Miocene (Hall,
2002). However, it is only at the southern end of the arc, in the
north arm of Sulawesi, and at the north end of the arc, in
Mindanao, that this has led to the formation of a substantial
subaerial landmass. The intervening section has remained
submerged or has been intermittently emergent for the last
20 million years (Myr). For Sangihe Island, an emergence age
of the order of 2–3 Myr is likely to be realistic (Jezek et al.,
1981; Morrice et al., 1983). Tanimbar has been subaerial for a
shorter time, as this low island (< 300 m) is covered with early
Pleistocene marine deposits, which indicate that it may have
risen above sea level only within the last 1 Myr. Furthermore,
Quaternary reefs occur up to 200 m a.s.l., also supporting a
recent period of rapid emergence (de Smet et al., 1989;
Charlton et al., 1991).
We used the age of Sangihe Island north of Sulawesi to date
the split between Coracornis raveni and Coracornis sanghirensis
at 2.5 ± 0.5 million years ago (Ma) and the age of Tanimbar
to date the split between Pachycephala macrorhynca and
Pachycephala pectoralis/melanura at 0.8 ± 0.2 Ma.
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 249ª 2009 Blackwell Publishing Ltd

A concern when using island ages to infer ages of speciation
events is that one must have accounted for all taxa in the
region and all possible land areas in the region. If some taxa are
not sampled, the island age might be applied to the incorrect
node because the appropriate one does not exist in the
phylogeny. Likewise, the colonization source must be unam-
biguous, which is complicated by the possibility of recently
submerged islands in dynamic regions such as the Indonesian
archipelago. This is a problem of missing islands rather than
missing taxa, and is impossible to remedy. We did not satisfy
either of these conditions, and so to further corroborate the
island divergence dates we applied to our data a rate of 0.028
substitutions per site per lineage per Myr for ND2 (corrected
pairwise distances), which is derived from Galapagos mock-
ingbirds (Drovetski et al., 2004). This divergence rate yields
ages similar to the above-mentioned nodes: 2.9 Ma for the
Coracornis raveni/sanghirensis split and 0.8 Ma for the Pachy-
cephala macrorhynca/pectoralis split.
We used beast version 1.4.6 (Drummond et al., 2002, 2006;
Drummond & Rambaut, 2007) to estimate divergence times
within the Pachycephalidae. We assigned the best fitting
model, as estimated by MrModeltest 2.0 (Nylander, 2004) to
each of the partitions. We ran three separate analyses using one
or both calibration points and found that the main effect of
using only one calibration point is to enlarge the 95% highest
posterior density (HPD) interval either distally or basally. We
assumed a Yule speciation process for the tree prior and an
uncorrelated lognormal distribution for the molecular clock
model (Ho, 2007). We used default prior distributions for all
other parameters and ran MCMC chains for 50 million
generations.
Establishing ancestral areas
Ancestral areas of clades were estimated with dispersal–
vicariance analysis (Ronquist, 1997) using the computer
program diva version 1.1 (Ronquist, 1996). Five geographical
regions were recognized: A, Australia/New Guinea; B, Philip-
pines; C, Wallacea; D, the Pacific; E, Asia (the area west of
Huxley’s modification of Wallace’s Line); and F, Palau.
Maxarea values were set to 2 (assuming that the ancestral
members of the extant Pachycephalidae radiation had the same
ability to disperse), and the analysis was carried out several
times to explore the effect of changing the cost settings (co-
divergence = 0–5, duplication = 0–5, sorting = 0–5, switch-
ing = 0–5). None of these changes altered the outcome of the
analysis, suggesting a robust result.
In addition, ancestral distributions were reconstructed by
performing a series of separate constrained BI analyses
(Huelsenbeck & Bollback, 2001; Ronquist, 2004) for each
node on the 50% majority-rule consensus tree derived from
analysis of the full five gene dataset. To do this we included the
distribution data as a separate partition and assumed the
evolutionary rate to be proportional to that of the other data;
other models and settings were the same as for the phylo-
genetic analyses.
RESULTS
Alignment and phylogenetic analyses
The concatenated alignment consisted of 3069 bp (660 parsi-
mony-informative sites). Amplification of DNA extracted from
museum study skins was not successful for all loci. Thus partial
or whole sequence fragments are missing for some taxa. The
total GAPDH alignment was 317 bp (30 parsimony-informa-
tive sites). We sequenced between 266 and 300 bp from fresh
samples, and from skin samples managed to obtain 197 bp for
Pitohui kirhocephalus, 210 bp for Pachycephala modesta,
178 bp for Colluricincla boweri and 189 bp for Pachycephala
hypoxantha. The total alignment of ODC intron 6 and 7 was
609 bp (43 parsimony-informative sites). We obtained
between 540 and 607 bp from fresh samples and obtained
ODC intron-6 sequences (249 bp) from skin samples of
Pachycephala modesta, Pachycephala lorentzi and Pachycephala
griseiceps. The total alignment of Myo2 was 707 bp (31
parsimony-informative sites). We sequenced between 674 and
707 bp from fresh samples, and from the museum study skins
obtained 499 bp from Pachycephala phaionota and 379 bp from
Colluricincla boweri. The ND2 alignment was 1041 bp (411
parsimony-informative sites). We obtained between 844 and
1041 bp from fresh samples and from skin samples we obtained
549 bp for Pachycephala griseonota, 567 bp for Colluricincla
tenebrosa, 580 bp for Pachycephala phaionota, 790 bp for
Pachycephala hypoxantha and 796 bp for Pachycephala lorentzi.
For ND3 and flanking tRNAs we sequenced 395 bp (145
parsimony-informative sites). However, we only obtained
318 bp for Colluricincla tenebrosa, 367 bp for Pachycephala
schlegelii and 391 bp for Pachycephala sulfuriventer.
Analyses performed on the concatenated data set (seven
partitions: GAPDH, ODC, Myo2, first, second and third
mtDNA codon positions and tRNA ML: )ln = 19,835.74, BI
harmonic mean )ln = 18,829.30) (Fig. 2) and on the individ-
ual partitions (GAPDH: AIC: GTR+C, ML: )ln = )1049.00, BI
harmonic mean )ln = 1191.85; ODC: AIC: HKY+C, ML:
)ln = )1943.28, BI harmonic mean )ln = 2166.67; Myo2:
AIC: K80+I, ML: )ln = )1740.65, BI harmonic mean
)ln = 1941.36; ND2: AIC: GTR+I+C, ML: )ln = )10075.14,
BI harmonic mean )ln = 9808.10; ND3: AIC: GTR+I+C, ML:
)ln = –3519.44, BI harmonic mean )ln = )3466.35) (see
Appendices S1–S5 in Supporting Information) yielded 50%
majority-rule consensus trees that were topologically congru-
ent for well-supported nodes (posterior probability > 0.95 and
bootstrap values > 70). Although the nuclear gene trees
(GAPDH, ODC and Myo2) contained few well-supported
clades, they did provide support for the partition between the
two main genera, Pachycephala and Colluricincla. The mito-
chondrial gene trees were more resolved, and as expected for
linked markers were congruent for well-supported nodes. The
only difference was within the basal part of Pachycephala,
where some discrepancies occurred. The differences are
supported only by Bayesian posterior probabilities in the
ND3 topology; likelihood bootstrap resampling supported
K. A. Jønsson et al.
250 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

none of the discrepancies. This is probably the result of the
small size of the ND3 gene (351 bp) and relatively few
informative sites relative to the ND2 gene (1041 bp). Scores of
the best likelihood trees were within 0.05 likelihood units of
the best tree recovered in each of the other four garli runs,
suggesting that the five runs had converged. With the
individual data sets combined, the ML tree topology was
almost completely congruent with the BI topology.
The 14 species not included in the study could potentially
come out anywhere in the phylogeny, but as discussed above
we have broad geographical and taxonomic sampling. As a
result it is unlikely to alter the main biogeographical patterns
recovered, i.e. many dispersal events into the Indonesian
archipelago and few into the Melanesian archipelago. To the
east, however, it is possible that we have underestimated the
complexity of the pattern, and it is conceivable that multiple
dispersal events took place from New Guinea into the Pacific.
Dating analyses
Results from the beast dating analyses (Fig. 3) place the origin
of the Pachycephalidae assemblage in the early Pliocene, and
the origin of the genus Pachycephala was estimated to date
back to the late Pliocene. Wide confidence intervals on all of
Figure 2 The 50% majority-rule consensus tree of the Pachycephalidae obtained from the Bayesian analysis of the combined dataset
(GAPDH, ODC, Myo2, ND2 and ND3). Above the branch is the posterior probability (only values above 0.90 are shown, asterisks indicate
1.00 posterior probabilities). Below the branch is the maximum likelihood bootstrap value (only values above 70% are shown) from 100
pseudoreplicates. Letters at nodes indicate putative ancestral areas: A, Australia/New Guinea; B, the Philippines; C, Wallacea; D, Pacific;
E, Asia (including Palawan); F, Palau. To the left of the nodes is first indicated the ancestral area, according to the Bayesian ancestral area
reconstruction (either indicated as a large letter, which indicates an ancestral area probability ‡ 0.99, or as a two or three small letters
indicating ancestral area probability fractions). Following the Bayesian ancestral areas are the ancestral areas recovered from the diva
analysis. Letters to the right of the taxon names indicate present distributions and thus coding for the ancestral area analyses. An asterisk
in front of a taxon name indicates a change of genus name according to the discussion. The inset map indicates the regions demarcated
for the ancestral area analyses.
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 251ª 2009 Blackwell Publishing Ltd
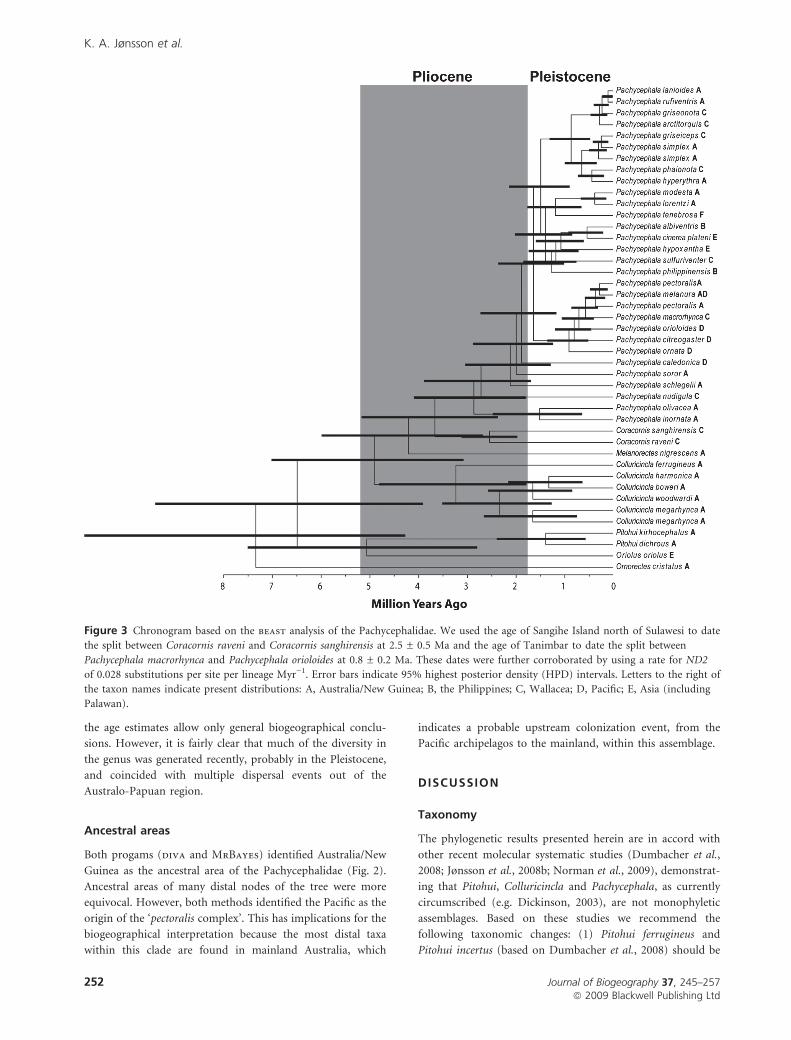
the age estimates allow only general biogeographical conclu-
sions. However, it is fairly clear that much of the diversity in
the genus was generated recently, probably in the Pleistocene,
and coincided with multiple dispersal events out of the
Australo-Papuan region.
Ancestral areas
Both progams (diva and MrBayes) identified Australia/New
Guinea as the ancestral area of the Pachycephalidae (Fig. 2).
Ancestral areas of many distal nodes of the tree were more
equivocal. However, both methods identified the Pacific as the
origin of the ‘pectoralis complex’. This has implications for the
biogeographical interpretation because the most distal taxa
within this clade are found in mainland Australia, which
indicates a probable upstream colonization event, from the
Pacific archipelagos to the mainland, within this assemblage.
DISCUSSION
Taxonomy
The phylogenetic results presented herein are in accord with
other recent molecular systematic studies (Dumbacher et al.,
2008; Jønsson et al., 2008b; Norman et al., 2009), demonstrat-
ing that Pitohui, Colluricincla and Pachycephala, as currently
circumscribed (e.g. Dickinson, 2003), are not monophyletic
assemblages. Based on these studies we recommend the
following taxonomic changes: (1) Pitohui ferrugineus and
Pitohui incertus (based on Dumbacher et al., 2008) should be
Figure 3 Chronogram based on the beast analysis of the Pachycephalidae. We used the age of Sangihe Island north of Sulawesi to date
the split between Coracornis raveni and Coracornis sanghirensis at 2.5 ± 0.5 Ma and the age of Tanimbar to date the split between
Pachycephala macrorhynca and Pachycephala orioloides at 0.8 ± 0.2 Ma. These dates were further corroborated by using a rate for ND2
of 0.028 substitutions per site per lineage Myr)1. Error bars indicate 95% highest posterior density (HPD) intervals. Letters to the right of
the taxon names indicate present distributions: A, Australia/New Guinea; B, the Philippines; C, Wallacea; D, Pacific; E, Asia (including
Palawan).
K. A. Jønsson et al.
252 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

included in Colluricincla; (2) Colluricincla sanghirensis and
Coracornis raveni are sister taxa and as C. sanghirensis is not
part of Colluricincla, but is instead more closely related to
Coracornis, we recommend its placement in this genus; (3) the
highly divergent Pitohui nigrescens should be separated gener-
ically, with the available name Melanorectes Sharpe, 1877
applied; (4) Pitohui cristatus has recently been separated into
Ornorectes Iredale, 1956 (Norman et al., 2009); as a conse-
quence, the only remaining members of Pitohui are P. kirho-
cephalus and Pitohui dichrous, for which the former is the type
species; and (5) Colluricincla tenebrosa from Palau should be
placed into Pachycephala (also suggested by Norman et al.,
2009).
Origins and biogeography
Although modern molecular-based biogeographical studies on
mammals (e.g. Steppan et al., 2003; Heaney et al., 2005; Jansa
et al., 2006), amphibians (e.g. Evans et al., 2003) and birds
(e.g. Filardi & Moyle, 2005; Cibois et al., 2007; Irestedt et al.,
2008; Outlaw & Voelker, 2008) have been carried out in
subregions within the Indo-Pacific, only one study (also a
family of passerine birds by Moyle et al., 2009) has investigated
biogeographical patterns of terrestrial vertebrates across the
entire Indo-Pacific region. As such, the present study provides
an important contribution towards a better understanding of
biogeographical patterns and processes that have shaped this
diverse region.
Based on the results of our ancestral area analyses and
patterns of present-day pachycephalid species richness, the
family clearly arose within the Australo-Papuan region
(Fig. 2). Although the presence of several New Guinea humid
forest species at the base of the phylogeny suggests a Papuan
origin of the family, it is important to note that much of
Australia’s humid forest diversity has been lost as a conse-
quence of aridification beginning in the mid-Miocene. The
distribution and branching patterns within Colluricincla are
equivocal with respect to clarifying the origins of the genus,
whereas the basal placement of Pachycephala inornata and
Pachycephala olivacea indicate a possible Australian origin of
Pachycephala, before the genus diversified within and from
New Guinea. Repeated Quaternary interchange between these
landmasses has been facilitated by the New Guinea–Australia
land bridge, present during periods of climate-mediated sea
fluctuations, which have differed by as much as 120 m
(Bintanja et al., 2005). This connectivity is reflected in the
broad shared distribution of several pachycephalid taxa as well
as close sister relationships between taxa in these regions.
Several processes have traditionally been proposed to
explain New Guinea’s high avian diversity. Striking patterns
of elevational replacement along the central cordillera and
outlying ranges involving abrupt elevational transitions
between species led Diamond (1972) to identify ‘montane
speciation’ as one of the most important mechanisms
promoting diversification in the New Guinean avifauna. At
lower elevations (< 1800 m a.s.l.), where habitat complexity is
at its greatest (Paijmans, 1976; Coates, 1985), high levels of
niche partitioning involving fine-scale differences in habitat
preference, vertical zonation within the forest stratum and/or
feeding ecology have also been identified in speciation
processes across the island (Diamond, 1972; Coates, 1985,
1990). New Guinea’s whistler diversity is principally composed
of lowland endemics and mid-elevation generalists, with
relatively few species present in the highlands (2000 m a.s.l.
and above). Given that the New Guinea highlands had largely
taken shape by the Pliocene, significantly pre-dating the
majority of Pachycephala diversification, more recent climatic
factors may have played a role in diversification of the genus.
Pleistocene glacial cycles have significantly impacted climate
and habitat change in New Guinea and adjacent regions. This
includes local aridification in southern lowland areas and
increased rainfall in montane areas, with significant cooling at
higher elevations (McAlpine et al., 1983). Pleistocene fluctu-
ations caused vegetational zones to be lowered and com-
pressed, with a corresponding increase in the area of montane
grasslands and lowland savanna (Hope et al., 2004). The
timing of these phenomena coincides with the major radiation
of the forest-associated Pachycephala species according to our
time estimates (Fig. 3). The genus Colluricincla, which is
adapted to drier, more open habitats, does not show the same
rapid radiation as observed for the forest-adapted taxa.
Timing and patterns of diversification
The distribution of the family provides a unique opportunity
to compare dispersal patterns in two archipelagos that display
distinctly different geological histories and distributions of
current land area west and east of the presumed area of origin
in New Guinea. Members of Pachycephalidae colonized the
Indonesian archipelago at least seven times (Fig. 4). All of
these species (with the exception of the widespread Pachycep-
hala cinerea) are restricted to one or a few adjacent islands and
several are restricted to highland primary forest. The close
proximity of islands throughout the Indonesian archipelago
(since the Miocene–Pliocene transition when the Lesser Sunda
Islands developed; Hall, 2002) seems to present easily acces-
sible stepping-stones for colonization throughout the Pliocene
and Pleistocene.
The phylogenetic results present a very different scenario for
dispersal into the Pacific (Fig. 4). Our study suggests one or two
independent dispersal events. One species colonized New
Caledonia in the late Pliocene and another lineage colonized
the entire Melanesian arc (including the Bismarcks, the
Solomons and Vanuatu) during the Pleistocene, before back-
colonizing mainland Australia. Island arcs are well known as
dispersal corridors (Polhemus, 1996; Irestedt et al., 2008) and
their importance for dispersal and diversification is supported
by these results. The Pacific ‘pectoralis’ complex probably
colonized the Melanesian arc as this chain of islands drifted
westwards towards New Guinea in the Pliocene. The colonizing
population then rapidly dispersed throughout the chain of
islands from New Guinea via the Bismarcks, and the Solomons
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 253ª 2009 Blackwell Publishing Ltd

to Vanuatu. Sparse taxon sampling, short internodes and lack of
branch support limit inferences about the exact sequence of
colonization, but underscore the rapid diversification. Reverse,
or ‘upstream’, colonization events have recently received
renewed attention (Filardi & Moyle, 2005; Bellemain & Ricklefs,
2008) and here we document another case of this hitherto
largely overlooked phenomenon. Jønsson et al. (2008c) recently
showed that Pachycephala melanura has colonized small islands
in the immediate vicinity of larger islands that hold populations
of P. pectoralis. This pattern extends as far east as the eastern
Bismarck archipelago. At the time it was not possible to
determine the origin of the clade. The present data set raises the
possibility that the lineage that back-colonized Australia
commenced a new round of colonization into the Bismarck
archipelago, exemplified by P. melanura.
Our study demonstrates that the dispersal and diversifica-
tion of a lineage in an archipelago can vary significantly
according to the tectonic history of the area. The Indonesian
and Melanesian radiations discussed here are closely related
and are thus expected to have similar dispersal potentials. Yet
the diversification process proceeded differently in the two
areas. Thus, our study emphasizes the importance of integrat-
ing knowledge of palaeogeology and plate tectonics for a given
region with modern phylogenetic and molecular clock dating
methods in order to appropriately describe and interpret
biogeographical patterns. Our findings also underscore the fact
that islands may be very important in the speciation and
diversification process and in shaping continental biotas,
contrary to traditional biogeographical paradigms.
ACKNOWLEDGEMENTS
We thank the following institutions for kindly providing access
to fresh tissue and/or toe-pads: American Museum of Natural
History, USA; Australian Museum, Sydney, Australia; Austra-
lian National Wildlife Collection, Canberra, Australia; Field
Museum of Natural History, Chicago, USA; Museo Civico di
Storia Naturale di Carmagnola, Italy; Museum of Vertebrate
Zoology, UC-Berkeley, USA; Museum Victoria, Melbourne,
Australia; Swedish Museum of Natural History, Stockholm,
Sweden; University of Kansas, USA; Western Australian
Museum, Perth, Australia; and the Zoological Museum of
Figure 4 Map indicating the dispersal and colonization routes of members of the Pachycephalidae in the Indonesian/Philippine archi-
pelago to the west of New Guinea and the Melanesian Archipelago to the east. Solid red lines indicate dispersal into the Pacific (note,
however, that support values are low for these relationships). Solid green lines indicate dispersal into Wallacea and beyond. The solid blue
line indicates dispersal to the Philippines. Solid black lines indicate dispersal within the Australo-Papuan region. Dotted lines indicate
putative dispersal routes (purple, the colonization of Palau from either New Guinea or the Philippines; red, the colonization of New
Caledonia either by long-distance ocean dispersal or via the Melanesian arc). Each terminal arrow indicates one colonization event of one
species. Yellow dotted lines indicate the boundaries of Wallacea; Wallace’s Line to the west and Lydekker’s Line to the east.
K. A. Jønsson et al.
254 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

Copenhagen, Denmark. K.A.J. would also like to acknowledge
the support from the Australian Museum Postgraduate Awards
2006–07. We would also like to thank Jim Patton and Craig
Moritz for valuable comments on earlier drafts. Finally, we
thank two anonymous referees and the editor, Lawrence
Heaney, for constructive criticism that helped improve the
manuscript markedly.
REFERENCES
Akaike, H. (1973) Information theory as an extension of
the maximum likelihood principle. Second International
Symposium on Information Theory (ed. by B.N. Petrov and
F. Csaki), pp. 276–281. Akademiai Kiado, Budapest.
Allen, E.S. & Omland, K.E. (2003) Novel intron phylogeny
(ODC) supports plumage convergence in orioles (Icterus).
The Auk, 120, 961–969.
Audley-Charles, M.G. (1991) Tectonics of the New Guinea area.
Annual Review of Earth and Planetary Sciences, 19, 17–41.
Bellemain, E. & Ricklefs, R.E. (2008) Are islands the end of the
colonization road? Trends in Ecology and Evolution, 23, 461–
468.
Bintanja, R., van de Wal, S.W. & Oerlemans, J. (2005) Mod-
elled atmospheric temperatures and global sea levels over the
past million years. Nature, 437, 125–128.
Boles, W.E. (2007) Family Pachycephalidae (whistlers).
Handbook of the birds of the world, Vol. 12 (ed. by J. del
Hoyo, A. Elliott and D.A. Christie), pp. 374–437. Lynx
Edicions, Barcelona.
Charlton, T.R., de Smet, M.E.M., Samdra, H. & Kaye, S.J.
(1991) The stratigraphic and structural evolution of the
Tanimbar islands, eastern Indonesia. Journal of Southeast
Asian Earth Sciences, 6, 343–358.
Chesser, R.T. (1999) Molecular systematics of the rhinocryptid
genus Pteroptochos. Condor, 101, 439–446.
Cibois, A., Thibault, J.-C. & Pasquet, E. (2007) Uniform
phenotype conceals double colonization by reed-warblers of
a remote Pacific archipelago. Journal of Biogeography, 34,
1150–1166.
Cicero, C. & Johnson, N.K. (2001) Higher level phylogeny of
vireos (Aves: Vireonidae) based on sequences of multiple
mtDNA genes. Molecular Phylogenetics and Evolution, 20,
27–40.
Coates, B.J. (1985) The birds of Papua New Guinea, Vol. I.
Dove Publications, Hong Kong.
Coates, B.J. (1990) The birds of Papua New Guinea, Vol. II.
Dove Publications, Hong Kong.
Darwin, C. (1859) On the origin of species by means of natural
selection. Murray, London.
Diamond, J.M. (1972) Avifauna of the Eastern Highlands of
New Guinea. Nuttall Ornithological Club, Cambridge, MA.
Diamond, J.M. (1977) Continental and insular speciation in
Pacific island birds. Systematic Zoology, 26, 263–268.
Dickinson, E.C. (2003) The Howard and Moore complete
checklist of the birds of the world, 3rd edn. Princeton Uni-
versity Press, Princeton, NJ.
Drovetski, S.V., Zink, R.M., Rohwer, S., Fadeev, I.V., Nesterov,
E.V., Karagodin, I., Koblik, E.A. & Redkin, Y.A. (2004)
Complex biogeographic history of a Holarctic passerine.
Proceedings of the Royal Society B: Biological Sciences, 271,
545–551.
Drummond, A.J. & Rambaut, A. (2007) BEAST v1.4.7. Avail-
able at: http://beast.bio.ed.ac.uk/.
Drummond, A.J., Nicholls, G.K., Rodrigo, A.G. & Solomon,
W. (2002) Estimating mutation parameters, population
history and genealogy simultaneously from temporally
spaced sequence data. Genetics, 161, 1307–1320.
Drummond, A.J., Ho, S.Y.W., Phillips, M.J. & Rambaut, A.
(2006) Relaxed phylogenetics and dating with confidence.
PLoS Biology, 4, e88.
Dumbacher, J.P., Deiner, K., Thompson, L. & Fleischer, R.C.
(2008) Phylogeny of the avian genus Pitohui and the evo-
lution of toxicity in birds. Molecular Phylogenetics and
Evolution, 49, 774–781.
Emerson, B.C. & Gillespie, R.G. (2008) Phylogenetic analysis of
community assembly and structure over space and time.
Trends in Ecology and Evolution, 23, 619–630.
Evans, B.J., Brown, R.M., McGuire, J.A., Supriatna, J.,
Andayani, N., Diesmos, A., Iskandar, D., Melnick, D.J. &
Cannatella, D.C. (2003) Phylogenetics of fanged frogs:
testing biogeographical hypotheses at the interface of the
Asian and Australian faunal zones. Systematic Biology, 52,
794–819.
Filardi, C.E. & Moyle, R.G. (2005) Single origin of a pan-
Pacific bird group and upstream colonization of Australasia.
Nature, 438, 216–219.
Fjeldsa, J., Zuccon, D., Irestedt, M., Johansson, U.S. & Ericson,
P.G.P. (2003) Sapayoa aenigma: a New World representative
of Old World suboscines. Proceedings of the Royal Society B:
Biological Sciences, 270, S238–S241.
Fjeldsa, J., Johansson, U.S., Lokugalappatti, L.G.S. & Bowie,
R.C.K. (2007) Diversification of African greenbuls in space
and time: linking ecological and historical processes. Journal
of Ornithology, 148, 359–367.
Galbraith, I.C.J. (1956) Variation, relationships and evolution
in the Pachycephala pectoralis superspecies (Aves, Musci-
capidae). Bulletin of the British Museum of Natural History,
4, 131–222.
Gill, F. & Wright, M. (2006) Birds of the world – recommended
English names. Princeton University Press, Princeton, NJ.
Gillespie, R.G., Claridge, E.M. & Roderick, G.K. (2008) Bio-
diversity dynamics in isolated island communities: interac-
tion between natural and human-mediated processes.
Molecular Ecology, 17, 45–57.
Hackett, S.J. (1996) Molecular phylogenetics and biogeography
of tanagers in the genus Ramphocelus (Aves). Molecular
Phylogenetics and Evolution, 5, 368–382.
Hall, R. (1998) Biogeographic implications of the Tertiary
palaeogeographic evolution of Sulawesi and Borneo. Bio-
geography and geological evolution of SE Asia (ed. by R. Hall
and J.D. Holloway), pp. 133–163. Backhuys Publishers,
Leiden.
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 255ª 2009 Blackwell Publishing Ltd

Hall, R. (2002) Cenozoic geological and plate tectonic evolu-
tion of SE Asia and the SW Pacific: computer-based
reconstructions, model and animations. Journal of Asian
Earth Sciences, 20, 353–431.
Hall, R. & Holloway, J.D. (1998) Biogeography and geological
evolution of SE Asia. Backhuys Publishers, Leiden.
Hamilton, W.B. (1979) Tectonics of the Indonesian region. US
Geological Survey Professional Paper 1078. US Government
Printing Office, Washington, DC.
Heaney, L.R., Walsh, J.S. & Peterson, A.T. (2005) The roles of
geological history and colonization abilities in genetic
differentiation between mammalian populations in the
Philippine archipelago. Journal of Biogeography, 32, 229–247.
Heslewood, M.M., Elphinstone, M.S., Tidemann, S.C. &
Baverstock, P.R. (1998) Myoglobin intron variation in the
Gouldian finch Erythrura gouldiae assessed by temperature
gradient gel electrophoresis. Electrophoresis, 19, 142–151.
Ho, S.Y.W. (2007) Calibrating molecular estimates of substi-
tution rates and divergence times in birds. Journal of Avian
Biology, 38, 409–414.
Holder, M.T. & Lewis, P.O. (2003) Phylogeny estimation:
traditional and Bayesian approaches. Nature Reviews
Genetics, 4, 275–284.
Hope, G., Kershaw, A.P., van der Kaars, S., Xiangjun, S., Liew,
P.-M., Heusser, L.E., Takahara, H., McGlone, M., Miyoshi,
N. & Moss, P.T. (2004) History of vegetation and habitat
change in the Austral-Asian region. Quaternary Interna-
tional, 118–119, 103–126.
Huelsenbeck, J.P. & Bollback, J.P. (2001) Empirical and hier-
archical Bayesian estimation of ancestral states. Systematic
Biology, 50, 351–366.
Huelsenbeck, J.P. & Ronquist, F. (2003) MrBayes: a program
for the Bayesian inference of phylogeny, Version 3.1.2. Avail-
able at: http://mrbayes.scs.fsu.edu/index.php.
Huelsenbeck, J.P., Ronquist, F. & Hall, B. (2001) MrBayes:
Bayesian inference of phylogeny. Bioinformatics, 17, 754–755.
Iredale, T. (1956) Birds of New Guinea, Vol. 2. Georgian
House, Melbourne.
Irestedt, M., Ohlson, J.I., Zuccon, D., Kallersjo, M. & Ericson,
P.G.P. (2006) Nuclear DNA from old collections of avian study
skins reveals the evolutionary history of the Old World
suboscines (Aves, Passeriformes). Zoologica Scripta, 35, 567–
580.
Irestedt, M., Fuchs, J., Jønsson, K.A., Ohlson, J.I., Pasquet, E. &
Ericson, P.G.P. (2008) The systematic affinity of the enig-
matic Lamprolia victoriae (Aves: Passeriformes) – an
example of avian dispersal between New Guinea and Fiji
over Miocene intermittent landbridges. Molecular Phylo-
genetics and Evolution, 48, 1218–1222.
Jansa, S.A., Barker, F.K. & Heaney, L.R. (2006) The pattern and
timing of diversification of Philippine endemic rodents:
evidence from mitochondrial and nuclear gene sequences.
Systematic Biology, 55, 73–88.
Jetz, W., Rahbek, C. & Colwell, R.K. (2004) The coincidence of
rarity and richness and the potential signature of history in
centers of endemism. Ecology Letters, 7, 1189–1191.
Jezek, P.A., Whitford, D.J. & Gill, J.B. (1981) Geochemistry of
recent lavas from the Sangihe-Sulawesi arc, Indonesia. The
geology and tectonics of eastern Indonesia (ed. by A.J. Barber
and S. Wiryosujono), pp. 383–389. Special Publication.
Geological Research and Development Centre, Bandung.
Jønsson, K.A., Irestedt, M., Fuchs, J., Ericson, P.G.P., Christi-
dis, L., Bowie, R.C.K., Norman, J.A., Pasquet, E. & Fjeldsa, J.
(2008a) Explosive avian radiations and multi-directional
dispersal across Wallacea: evidence from the Campephagi-
dae and other Crown Corvida (Aves). Molecular Phyloge-
netics and Evolution, 47, 221–236.
Jønsson, K.A., Bowie, R.C.K., Norman, J.A., Christidis, L. &
Fjeldsa, J. (2008b) Polyphyletic origin of toxic Pitohui birds
suggests widespread occurrence of toxicity in corvoid birds.
Biology Letters, 4, 71–74.
Jønsson, K.A., Bowie, R.C.K., Moyle, R.G., Christidis, L.,
Filardi, C.E., Norman, J.A. & Fjeldsa, J. (2008c) Molecular
phylogenetics and diversification within one of the most
geographically variable bird species complexes (Pachycep-
hala pectoralis/melanura). Journal of Avian Biology, 39, 473–
478.
MacArthur, R.H. & Wilson, E.O. (1963) An equilibrium theory
of insular zoogeography. Evolution, 17, 373–387.
MacArthur, R.H. & Wilson, E.O. (1967) The theory of island
biogeography. Princeton University Press, NJ.
McAlpine, J.R., Keig, G. & Falls, R. (1983) Climate of Papua
New Guinea. Commonwealth Scientific Industrial Research
Organisation, Canberra.
Morrice, M.G., Jezek, P.A., Gill, J.B., Whitford, D.J. &
Monoarfa, M. (1983) An introduction to the Sangihe Arc:
volcanism accompanying arc–arc collision in the Molucca
Sea, Indonesia. Journal of Volcanology and Geothermal
Research, 19, 135–165.
Moyle, R.G., Filardi, C.E., Smith, C.E. & Diamond, J. (2009)
Explosive Pleistocene diversification and hemispheric
expansion of a ‘great speciator’. Proceedings of the National
Academy of Sciences USA, 106, 1863–1868.
Norman, J.A., Ericson, P.G.P., Jønsson, K.A., Fjeldsa, J. &
Christidis, L. (2009) A multi-gene phylogeny reveals novel
relationships for aberrant genera of Australo-Papuan core
Corvoidea and polyphyly of the Pachycephalidae and
Psophodidae (Aves: Passeriformes). Molecular Phylogenetics
and Evolution, 52, 488–497.
Norman, J.A., Boles, W.E. & Christidis, L. (in press) Rela-
tionships of the New Guinean songbird genera Amalocichla
and Pachycare based on mitochondrial and nuclear DNA
sequences. Journal of Avian Biology.
Nylander, J.A.A. (2004) MrModeltest2. Available at: http://
www.abc.se/~nylander/.
Nylander, J.A.A., Wilgenbusch, J.C., Warren, D.L. & Swofford,
D.L. (2008) AWTY (are we there yet): a system for graphical
exploration of MCMC convergence in Bayesian phylo-
genetics. Bioinformatics, 24, 581–583.
Outlaw, D.C. & Voelker, G. (2008) Pliocene climatic change in
insular Southeast Asia as an engine of diversification in
Ficedula flycatchers. Journal of Biogeography, 35, 739–752.
K. A. Jønsson et al.
256 Journal of Biogeography 37, 245–257ª 2009 Blackwell Publishing Ltd

Paijmans, K. (1976) New Guinea vegetation. Elsevier Scientific
Publications, Amsterdam.
Pigram, C.J. & Symonds, P.A. (1991) A review of the timing of
the major tectonic events in the New Guinea Orogen.
Journal Southeast Asian Earth Science, 6, 307–318.
Polhemus, D.A. (1996) Island arcs, and their influence on
Indo-Pacific biogeography. The origin and evolution of
Pacific island biotas, New Guinea to eastern Polynesia:
patterns and processes (ed. by A. Keast and S.E. Miller),
pp. 51–66. Academic Publishing, Amsterdam.
Posada, D. & Buckley, T.R. (2004) Model selection and model
averaging in phylogenetics: advantages of Akaike informa-
tion criterion and Bayesian approaches over likelihood ratio
tests. Systematic Biology, 53, 793–808.
Ronquist, F. (1996) DIVA version 1.1. Computer program
and manual available by anonymous FTP from Uppsala
University. Available at: ftp.uu.se or ftp.systbot.uu.se.
Ronquist, F. (1997) Dispersal–vicariance analysis: a new
approach to the quantification of historical biogeography.
Systematic Biology, 46, 195–203.
Ronquist, F. (2004) Bayesian inference of character evolution.
Trends in Ecology and Evolution, 19, 475–481.
Ronquist, F. & Huelsenbeck, J.P. (2003) MrBayes 3: Bayesian
phylogenetic inference under mixed models. Bioinformatics,
19, 1572–1574.
Slade, R.W., Moritz, C., Heideman, A. & Hale, P.T. (1993)
Rapid assessment of single-copy nuclear DNA variation in
diverse species. Molecular Ecology, 2, 359–373.
de Smet, M.E.M., Charlton, T.R., Kaye, S., Faeni, H., Troelstra,
S.R. & van Marle, L.J. (1989) Late Cenozoic history of the
island of Yamdena, Tanimbar Archipelago, Eastern Indo-
nesia. Benthic foraminifera from the Banda Arc Region,
Indonesia, and their paleobathymetric significance for geologic
interpretations of the Late Cenozoic sedimentary record (ed. by
L.J. van Marle), pp. 145–162. Free University Press, The
Hague, Netherlands.
Spellman, G.M., Cibois, A., Moyle, R.G., Winker, K. & Barker,
F.K. (2008) Clarifying the systematics of an enigmatic avian
lineage: what is a bombycillid? Molecular Phylogenetics and
Evolution, 49, 1036–1040.
Steppan, S.J., Zawadski, C. & Heaney, L.R. (2003) Molecular
phylogeny of the endemic rodent Apomys (Muridae) and
the dynamics of diversification in an oceanic archipelago.
Biological Journal of the Linnean Society, 80, 699–715.
Storch, D., Davies, D., Dajicek, R.G., Orme, C.D.L., Olson, V.,
Thomas, G.H., Ding, T.-S., Rasmussen, P.C., Ridgely, R.S.,
Bennett, P.M., Blackburn, T.M., Owens, I.P.F. & Gaston,
K.J. (2006) Energy, range dynamics and global species
richness patterns: reconciling mid-domain effects and
environmental determinants of avian diversity. Ecology
Letters, 9, 1308–1320.
Wallace, A.R. (1876) The geographical distribution of animals.
Harper, New York.
Whittaker, R.J. & Fernandez-Palacios, J.M. (2007) Island
biogeography: ecology, evolution, and conservation, 2nd edn.
Oxford University Press, Oxford.
Wilgenbusch, J.C., Warren, D.L. & Swofford, D.L. (2004)
AWTY: a system for graphical exploration of MCMC
convergence in Bayesian phylogenetic inference. Available at:
http://ceb.csit.fsu.edu/awty.
Yan, C.Y. & Kroenke, L.W. (1993) A plate tectonic recon-
struction of the southwest Pacific, 0–100 Ma. Proceedings of
the Ocean Drilling Program Scientific Research, 130, 697–709.
Zwickl, D.J. (2006) Genetic algorithm approaches for the
phylogenetic analysis of large biological sequence datasets
under the maximum likelihood criterion. PhD Dissertation,
University of Texas at Austin, Austin, TX.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Appendix S1 The 50% majority-rule consensus tree
obtained from the Bayesian analysis of GAPDH.
Appendix S2 The 50% majority-rule consensus tree
obtained from the Bayesian analysis of ODC.
Appendix S3 The 50% majority-rule consensus tree
obtained from the Bayesian analysis of Myo2.
Appendix S4 The 50% majority-rule consensus tree
obtained from the Bayesian analysis of ND2.
Appendix S5 The 50% majority-rule consensus tree
obtained from the Bayesian analysis of ND3.
As a service to our authors and readers, this journal provides
supporting information supplied by the authors. Such mate-
rials are peer-reviewed and may be re-organized for online
delivery, but are not copy-edited or typeset. Technical support
issues arising from supporting information (other than
missing files) should be addressed to the authors.
BIOSKETCHES
Knud A. Jønsson is a PhD student at the Zoological
Museum, University of Copenhagen, Denmark. His main
interests include systematics and historical biogeography of
passerine birds with a particular emphasis on the Indo-Pacific
region.
Jon Fjeldsa is a professor at the Zoological Museum,
University of Copenhagen, Denmark. He has broad interests
in phylogeny and systematics of birds as well as a general
interest in several other aspects of ornithology, ecology and
conservation.
Author contributions: K.A.J. and J.F. conceived the ideas;
K.A.J. collected and analysed the data. All authors took part in
the writing process.
Editor: Lawrence Heaney
Phylogeny and biogeography of the Pachycephalidae
Journal of Biogeography 37, 245–257 257ª 2009 Blackwell Publishing Ltd
Top Related
Copyright © 2022 FDOKUMEN

