







Bahasa
Halaman
Hukum


Copyright 2003 by the Genetics Society of America
Green Fluorescent Protein Tagging Drosophila Proteins at Their Native GenomicLoci With Small P Elements
Peter J. Clyne,1 Jennie S. Brotman,1 Sean T. Sweeney and Graeme Davis2
Department of Biochemistry and Biophysics, University of California, San Francisco, California 94143-0448
Manuscript received March 31, 2003Accepted for publication August 11, 2003
ABSTRACTWe describe a technique to tag Drosophila proteins with GFP at their native genomic loci. This technique
uses a new, small P transposable element (the Wee-P) that is composed primarily of the green fluorescentprotein (GFP) sequence flanked by consensus splice acceptor and splice donor sequences. We demonstratethat insertion of the Wee-P can generate GFP fusions with native proteins. We further demonstrate thatGFP-tagged proteins have correct subcellular localization and can be expressed at near-normal levels. Wehave used the Wee-P to tag genes with a wide variety of functions, including transmembrane proteins. Agenetic analysis of 12 representative fusion lines demonstrates that loss-of-function phenotypes are notcaused by the Wee-P insertion. This technology allows the generation of GFP-tagged reagents on a genome-wide scale with diverse potential applications.
THE advent of green fluorescent protein (GFP) as intact fusion proteins for studies of subcellular localiza-tion in a living cell. This GFP trap is the first describeda means to visualize proteins in living cells has be-
gun a revolution in many fields of cellular biology that has little apparent alteration of protein function.The protein-trapping technology that we describe here(Chalfie et al. 1994). Three major drawbacks currently
hamper experiments using GFP tagging. First, proteins is similar to recent work from the Chia laboratory(Morin et al. 2001). A fundamental difference betweenof interest, once tagged, must generally be overexpressed
and this overexpression can generate novel phenotypes these approaches resides in the construction of the mo-bile P element. To make the Wee-P less mutagenic thanthat disrupt the processes being studied. Second, over-
expression will often disrupt the fine architecture of standard P elements, we have used the FLP-FRT systemto remove the mini-w� gene so that the resulting Wee-Pprotein localization. Finally, to tag a protein, a full-
length cDNA must be available. element is small in size and lacks a second transcriptionunit. Our genetic data suggest that the Wee-P rarelyWe have developed a GFP trap in Drosophila that is
capable of generating full-length GFP fusion proteins generates loss-of-function phenotypes when homozy-gous or when placed in trans to deficiency chromosomesthroughout the genome. This GFP trap is based on the
mobile genetic P element in Drosophila in which a GFP that uncover the insertion location. This allows theWee-P to be visualized as a homozygous element, therebysequence is flanked by splice acceptor and splice donor
sequences. This enables GFP to be spliced onto the tagging a larger portion of the endogenous transcript,which can be essential for visualizing proteins of lownative gene transcripts and take advantage of the endog-
enous splicing apparatus. The expression of GFP fusion abundance. In addition, this may allow the Wee-P pro-tein fusions to be used as reporters for changes in pro-proteins generated in this manner is controlled by en-
dogenous genomic regulatory sequences. As a result, tein abundance during development or in response toexperimental manipulation.the correct spatial and temporal expression patterns
are achieved and the resulting fusion proteins are notexpressed at elevated levels.
MATERIALS AND METHODSSeveral transposable-element-based trapping meth-odologies have been recently pursued. Most gene traps Construction of the Wee-P element: The Wee-P element
consists of a GFP sequence flanked by splice donor and spliceand enhancer traps in Drosophila have been designedacceptor sites, a mini-white gene flanked by FRT sites, anto facilitate gene disruption (Bier et al. 1989; Wilsonampicillin resistance gene and origin of bacterial replicationet al. 1989; Lukacsovich et al. 2001). In contrast, the(pUC segment), and P-element inverted repeat ends. To gen-
GFP trap that we describe here is designed to generate erate this transposon, we first made an intermediate vectorcontaining the following three elements: (1) a pUC segmentand P-element ends cut from pCaSpeR-4, (2) a mini-whitegene flanked by FRT sites cut from FLP-IMP (Keller et al.1These authors contributed equally to this work.2002), and (3) a short linker sequence. The GFP sequence2Corresponding author: Department of Biochemistry and Biophysics,was PCR amplified from pEGFP tubulin (CLONTECH, PaloUniversity of California, 513 Parnassus Ave., HSE 901, San Francisco,
CA 94143-0448. E-mail: [email protected] Alto, CA) using the primers JB NotI GFP-2 (TTAATTTACTAT
Genetics 165: 1433–1441 (November 2003)

1434 P. J. Clyne et al.
GCGGCCGCTTTACTTAATTACAGGGTCTATATAAGCAGA Molecular analysis of P-element insertions and Western blot-ting: The insertion site of the Wee-P element in generatedGCTGG) and JB-GFP BamHI (ATTCCAAGTTCTGCAGATA
AAGAATTACTTACCTCTCGAGATCTGAGTCCGGAC) and lines was determined using inverse PCR, as described by theBerkeley Drosophila Genome Project (http://www.fruitfly.inserted into this intermediate vector. The most conserved
Drosophila splice acceptor site, CAG, and splice donor site, org/about/methods/inverse.pcr.html). PCR amplificationwas performed using first-round primers IPCR-4 (CAATCAGTAAGT (Mount et al. 1992), were inserted at the 5� end
and 3� end, respectively, of the GFP sequence. To increase TATCGCTGTCTCAC) and IPCR-6 (GATTAACCCTTAGCthe likelihood that the cell’s splicing machinery will recognize ATGTCC) and second-round nested primers IPCR-7 (ACTthese artificially inserted splice sites, these splice sites were ATTCCTTTCACTCGCAC) and IPCR-9 (ACCTCTCGAGATboth positioned adjacent to 10- to 12-bp AT-rich regions char- CTGAGTCC). Western blots were done with standard SDS-acteristic of intronic DNA (Mount et al. 1992). The GFP se- PAGE gels according to standard protocols.quence also contains its own ATG codon for translation initia-tion. The pUC segment of the vector was used for subcloningpurposes only and does not integrate into the genomic DNA
RESULTSof the fly. The mini-white gene was used as a marker fortransformation of the injected vector and subsequently re- Construction and expression of the Wee-P: To gener-moved using heat-shock FLP-induced FRT recombination, as ate new reagents for live visualization of proteins wedescribed previously (Golic and Lindquist 1989). Therefore,
developed a P-element-based strategy for tagging pro-the resulting P element transposed to create independentWee-P-element insertions is �1.9 kb in length. teins with GFP at their native genomic loci. This ap-
Genetics: y, w, P[hsFLP12, ry�]; Sco/CyO used for FRT recom- proach takes advantage of the endogenous splicing ap-bination of the mini-white gene from the original Wee-P ele- paratus utilized by the majority of genes in Drosophila:ment vector was obtained from the Bloomington Stock Center.
�82% of the genes in Drosophila have introns and, ofThe stock containing the transposase enzyme (w; Cyo/Sp; ry,those, there are an average of 2.5 introns per geneDr � 2-3/TM6) was kindly provided by John Roote. The initial
en masse crosses to this transposase source are with silent, (Long et al. 1995; Deutsch and Long 1999; M. Long,starter Wee-P elements (ST) that are in w� backgrounds. w/Y; personal communication). There is also evidence thatWee-P ST/�; Dr �2-3/� were crossed en masse to virgin w fe- exons are frequently modular, encoding distinct motifsmales derived from a stock of w/Y P(hs-hid) (a gift from the
within a gene (Long et al. 1995). Therefore, genes withMatt Scott lab) that were heat shocked to yield high numbersexons may tolerate the insertion of a new motif such asof virgin females. Crosses were done in large cages on apple
or grape juice plates that held �300 parental flies. Each cage GFP without significant alterations to their function.was given a unique number and screened for several weeks To create a small P element that would act as a GFPuntil the yield of progeny was low. Care was taken to minimize trap and hop frequently, we took a two-step strategy.duplicated work due to the effects of premeiotic clusters by
First, we designed a P element containing the GFP se-noting down in detail the initial GFP expression pattern andquence as well as a scorable marker (mini-white) thatcage number of each GFP-positive animal selected. Each GFP-
expressing line typically had three separate Wee-P insertion would allow us to identify transgenic animals harboringevents, suggesting a high frequency of transposition (the ab- the P-GFP element (Figure 1A). Second, we excisedsence of a scorable marker independent of the GFP precludes the mini-white gene using the FLP recombinase systemthe determination of the transposition frequency using tradi-
(Figure 1B; Golic and Lindquist 1989). After remov-tional markers such as variegated eye color). To determineing the mini-white gene from the P-GFP element, thewhich Wee-P insertion causes the GFP expression we out-
crossed each line two generations to a y w stock, selecting on total size of the P element was �1.9 kb (compared tothe GFP expression pattern. After two generations we typically standard mini-white-containing P elements that areobserve only a single band by inverse PCR (IPCR) originating 10–15 kb; see Bier et al. 1989; Wilson et al. 1989). Wefrom a single Wee-P insertion. Infrequently, two bands persist
reasoned that a smaller P element would be less muta-after outcrossing. In this case, both bands are sequenced andgenic and increase the likelihood that a trapped proteinonly one is generally capable of generating a fusion event.
The Df(3R)�356 stock was a generous gift of the Zinsmaier would be expressed at endogenous levels by the nativelaboratory. The following lines (provided by Bloomington genomic transcriptional regulatory sequences. We referStock Center) were used for genetic analysis: (kis1cn1bw1sp1/ to this P element as the “Wee-P” due to its small size.SM6a), (y, w; P{w[�mC] � lacW}Hsc 70-4 L3929/TM3 Ser),
As P elements show a tendency to insert near or in the(Df(3R)P13), and (l(2)k10423).5� untranslated region (UTR) of genes, we chose toScreening techniques: Embryos were aged until late em-
bryogenesis and dechorionated using 50% bleach for 2–3 min keep the ATG of GFP in the Wee-P so that 5� splicedfollowed by a brief rinse through a sieve in water. The embryos GFP fusions would be expressed (Liao et al. 2000).were then placed in a small dish of heptane and spread with Therefore, we also placed consensus translation initia-the heptane on microscope slides with pipettors. Because they
tion sequences just 5� to the ATG (Cavener and Raysank in the heptane, the embryos formed single layers on1991). Although sequences associated with an initialthe microscope slides. Five to ten seconds after the heptane
evaporated, we placed halocarbon oil over the embryos and ATG were placed in the Wee-P, we deliberately chosethen screened visually for GFP on a GFP dissection microscope not to include any promoter sequences so that GFPusing a �10 compound objective (Zeiss). We also screened expression would accurately reflect the expression offirst instar larvae by rinsing them off of plates with PBT (1�
the trapped protein. As introns occur in three phases, wePBS, 0.1% Tween) into a sieve; we then rinsed them withconstructed three different Wee-P elements appropriatewater, placed them in a petri dish with a thin layer of PBT,
and screened visually with a GFP dissecting microscope. for each intron phase (Figure 1C).

1435Tagging Genes at Their Native Loci
Screening with the Wee-P for GFP-tagged proteins: independent Wee-P insertions, which complicated thecompletion of the IPCR step. We therefore did twoFor our initial screen we crossed Wee-PST0 (a second
chromosome phase 0 Wee-P insertion that, due to its rounds of outcrossing on the basis of GFP expressionto clean the chromosomes. IPCR on these animals gen-location, does not form a fusion protein) to a stock of
�2-3, Dr/TM6b (Figure 1D). Males heterozygous for erally yielded single clear bands. We then performedsequence analysis of these single bands to determineWee-PST0 and �2-3, Dr were then crossed to y w animals.
Embryos were aged until late embryogenesis to allow the site and orientation of the putative fusion event. Ina number of cases sequence analysis of RT-PCR productstime for zygotic protein translation before dechoriona-
tion and visualization on microscope slides. Embryos was performed to confirm the exact nature of the fusionevent (Table 1).were plated as a monolayer on standard microscope
slides and visualized using a fluorescent dissection mi- Approximately 60% of all GFP-positive animals repre-sent in-phase fusion events. Of the remaining 40% ofcroscope equipped with a �10 compound objective.
Each slide contained �2000 embryos and was screened animals that are selected on the basis of GFP expression,a large percentage are due to a hot spot for Wee-Pin 30 min. Using this method GFP� embryos were found
at a frequency of �1/400 animals. insertions in the lola locus. In these animals, the GFPamino acid sequence plus a few additional nonsenseEach embryo identified as having GFP expression was
placed in an individual vial. Adults that emerged were amino acid residues are translated after the Wee-P exonfuses with a 5� UTR exon of the lola locus that containsthen crossed to y w. In some cases the �2-3, Dr chromo-
some had to be selected against. Initially, we performed multiple stop codons. Other nonfusion events includecases where the Wee-P element has landed in the 5�IPCR directly on the progeny of the initial GFP� animal.
However, these animals typically had three or more UTR of a gene whose first intron is not in the samephase as the Wee-P element.
Identity and subcellular localization of GFP fusionevents: To date we have analyzed �100 lines that expressGFP. We have characterized a subset of these lines inmolecular and genetic detail to assess the properties ofWee-P fusion events. A diverse array of proteins havebeen tagged with GFP using the Wee-P element, includ-
Figure 1.—Construction and genetic crosses with theWee-P. (A) The first step in constructing the Wee-P was togenerate a P element that had a mini-white gene for scoringthe initial injections. Flanking this mini-white were two FRTsites for eventual excision. Intron sequences and splice ac-ceptor and splice donor sites flank the ORF of GFP. The mini-white, GFP, and flanking intron sequences are shown to scale.The FRT sites and 31-bp inverted repeats are enlarged forsimplicity in viewing. (B) Following excision of the mini-whiteby FLP recombinase, the remaining P element is only 1.9 kblong and is composed essentially of the GFP ORF and flankingintron and splicing sequences. Scale as in A. (C) Introns caninsert in any one of three positions relative to coding sequence.We have constructed three distinct Wee-P elements appro-priate for each of the three intron phases. To compensate forthe phases, we inserted nucleotide sequences (green) thatwould be as minimally disruptive as possible to the codingsequence. For example, in our phase 2 construct, just beforethe “gtaagt” splice donor sequence, we placed a “CT.” Wechose CT because it will encode for an extra leucine, a com-mon amino acid with a small and poorly reactive side chain.(D) Our first cross is designed to combine a transposase source(�2-3) with a starter Wee-P element (Wee-PST) that, due to itsinsertion location, does not form a fusion protein. In thesecond cross, males with the Wee-PST and the �2-3 are com-bined with y w virgins. These y w virgins are collected from astock that has a “hs-hid” element on the Y chromosome, which,following heat shock, allows for rapid collection of thousandsof virgins. Once a GFP� embryo or larva has been isolated,it is outcrossed two times to clean up the chromosomes andthen IPCR is performed on the line.

1436 P. J. Clyne et al.
TABLE 1
Summary of selected Wee-P fusions
Line Gene Location Function Predicted fusion No.a Localization
Wee-P1 hsc70-4 88E8-9 Chaperone N terminalb 3 Matches publishedWee-P2 kismet 21B3-4 Chromotin remodeling Internalb 2 Matches publishedWee-P3 ciboulot 4B1 Cytoskeleton N terminalb 1 Matches publishedWee-P4 sec61 26D4-7 ER channel Internalb 1 Matches publishedWee-P5 larp 98C3 Unknown N terminalb 1 CytosolicWee-P6 belle 85A5-8 RNA helicase 1, N terminal; 1, internal 2 Matches publishedWee-P20 CG9894 23A7-B1 Unknown Internal 5 See textWee-P25 hrb98DE 98D3-4 RNA binding N-terminal truncation 2 Matches publishedWee-P26 mhc 36A8-9 Muscle cytoskeleton Internal 1 Matches publishedWee-P27 tropomyosinI 88E9-12 Muscle cytoskeleton Internal 1 Matches publishedWee-P28 yip6 ND Ribosomal protein N terminal 1 Matches publishedWee-P59 pabp2 44A7-8 RNA polyadenylation Internal 1 Matches publishedWee-P114 calmodulin 48F8-10 Calcium binding N terminal 6 Matches publishedWee-P118 cp1 50D2-3 Cystein protease Internal 1 Matches publishedWee-P123 GH03576 67F4-68A7 Unknown N terminal 1 CytosolicWee-P153 topoisomerase1 13B6 DNA maintenance Internal 1 Matches publishedWee-P157 CG7970 65A5 Peroxisomal membrane N terminal 1 CytosolicWee-P283 ATPalpha 93B Na pump N terminal 2 Plasma membraneWee-P338 Pyk 94A10 Pyruvate kinase N terminal 1 CytosolicWee-P345 Karybeta3 82D Nuclear protein import Internal 1 Cytosolic
For an explanation of “Predicted fusion,” see text.a The number of separate Wee-P insertions in a given gene.b Lines for which RT-PCR has confirmed the predicted fusion and the presence of wild-type transcript.
ing transmembrane proteins as well as cytoplasmic and an endoplasmic reticulum (ER) resident protein (Fig-ure 2F). Finally, in the Wee-P:GFP fusion with the puta-nuclear proteins (Figures 2–4; Table 1). The GFP fusion
events can occur N-terminally as well as within the open tive chromatin remodeling protein Kismet, GFP is local-ized to the nucleus as predicted, where it is observedreading frame (ORF). N-terminal fusions are derived
from a variety of insertion sites including upstream of in puncta (Figure 2, G and H; Daubresse et al. 1999).Molecular analysis of Wee-P fusion events: We exam-the 5� UTR, within an exon of the 5� UTR, and within
introns bound by 5� UTR exons. Internal fusion events ined a subset of Wee-P fusions by Western blots usinganti-GFP antibodies. In all the fusions analyzed we foundare derived from insertions in introns bound by coding
exons and constitute about half of the fusions (Table 1). bands of the correct predicted size (Figure 3A). Further-more, these Western blots indicate that there is gener-For internal fusion events the average size of the in-
tron into which Wee-P elements have been observed to ally no evidence of altered protein stability or degrada-tion. It is also important to determine how much of theinsert is 2.9 kb (SD � 4.1 kb). The smallest intron into
which a Wee-P has inserted and generated a fusion is 183 native transcript includes the GFP sequence. A pre-viously published antibody to the Cib protein workedbp. In comparison, the average intron size for fusions
discovered with the protein trap transposon is �8 kb well on Westerns and demonstrates that a majority ofthe protein made from the Wee-P3:Cib locus includes(Morin et al. 2001), suggesting that the shorter Wee-P
transposon may have a significant advantage in trapping GFP (Figure 3B; Boquet et al. 2000). However, in thisinstance, there is a reduction in the total protein expres-shorter introns.
An analysis of subcellular localization for Wee-P fu- sion level. To address this question further we per-formed RT-PCR on five different Wee-P lines and foundsions demonstrates that the fusion proteins are correctly
localized in each insertion line analyzed to date. For that in each case, in addition to transcripts that includethe GFP, wild-type transcripts are also formed (Table 1).example, the Wee-P114:GFP fusion with calmodulin is
widely expressed and enriched in neuronal tissues (Fig- Analysis of the Wee-P1:GFP:Hsc70-4 fusion: To illus-trate in more detail the functioning of the Wee-P, weure 2A). In a second example, a fusion with Ciboulot
(a G-actin-binding protein) is distributed in several dif- present a detailed examination of the Wee-P1:GFP:Hsc70-4 fusion (Figure 4). This Wee-P inserted in anferent tissues, such as the central nervous system, the
chordotonal organs, and the trachea (Figure 2, C–E). intron of Hsc70-4 (Figure 4A), a broadly expressed andessential gene that plays important roles as a molecularThe subcellular localization of Wee-P3:GFP:Cib within
each of these cell types is cytoplasmic, as predicted for chaperone (Carbajal et al. 1993) and in catalyzing theuncoating of clathrin-coated vesicles (Newmyer anda G-actin-binding protein. A fusion with the transmem-
brane protein Sec61 shows localization consistent with Schmid 2001). Hsc70-4 has also been proposed to play
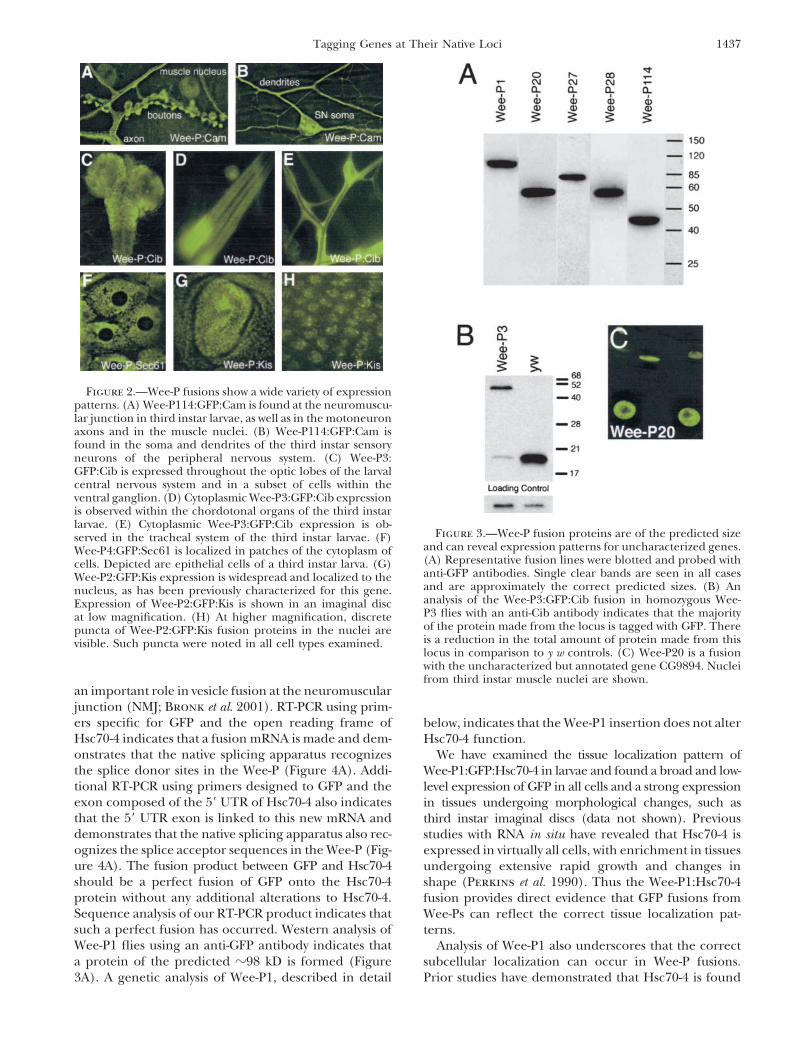
1437Tagging Genes at Their Native Loci
Figure 2.—Wee-P fusions show a wide variety of expressionpatterns. (A) Wee-P114:GFP:Cam is found at the neuromuscu-lar junction in third instar larvae, as well as in the motoneuronaxons and in the muscle nuclei. (B) Wee-P114:GFP:Cam isfound in the soma and dendrites of the third instar sensoryneurons of the peripheral nervous system. (C) Wee-P3:GFP:Cib is expressed throughout the optic lobes of the larvalcentral nervous system and in a subset of cells within theventral ganglion. (D) Cytoplasmic Wee-P3:GFP:Cib expressionis observed within the chordotonal organs of the third instarlarvae. (E) Cytoplasmic Wee-P3:GFP:Cib expression is ob-
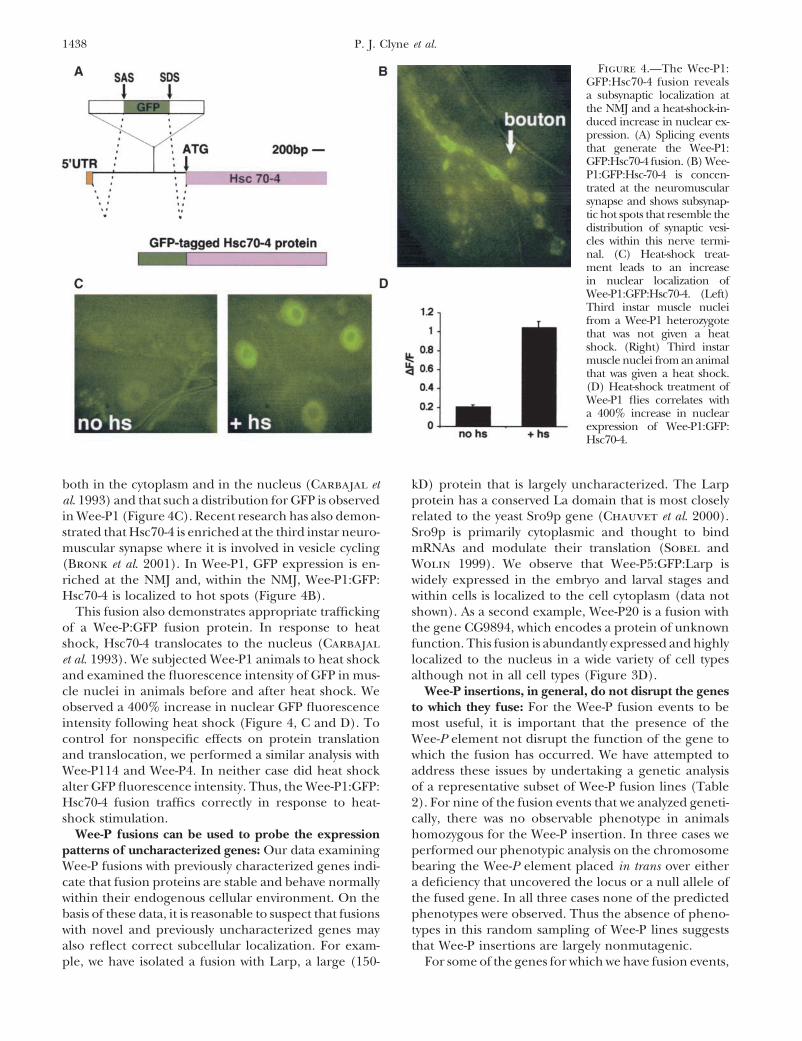
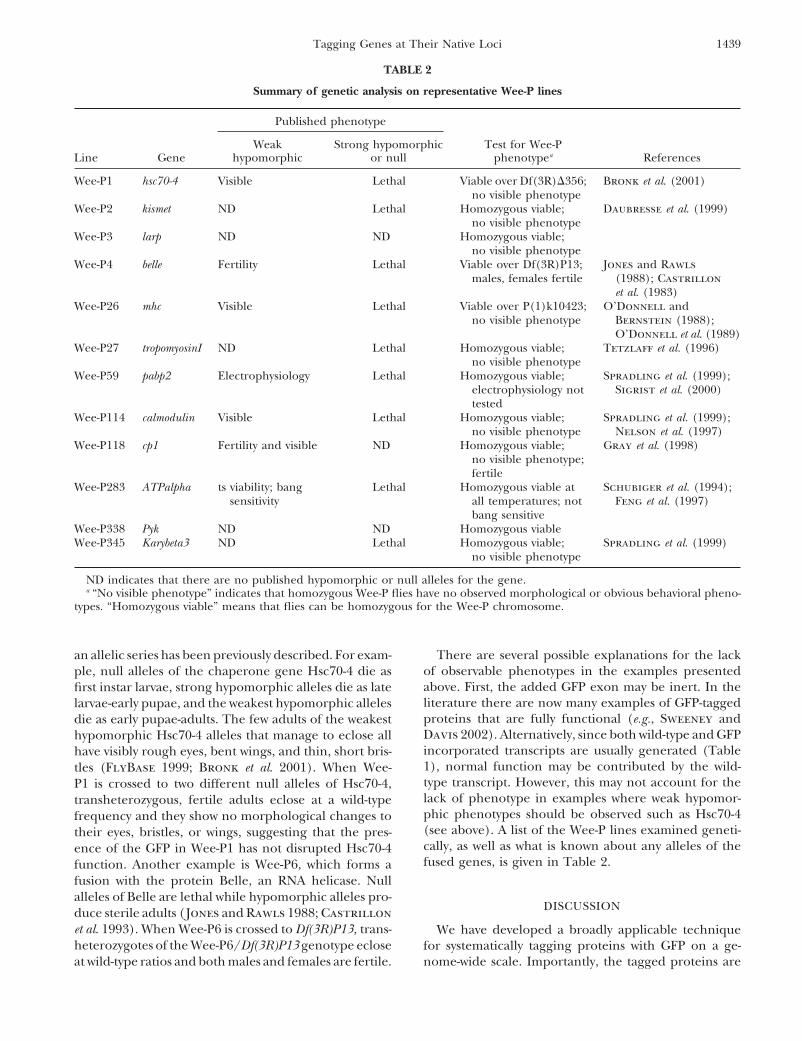
Figure 3.—Wee-P fusion proteins are of the predicted sizeserved in the tracheal system of the third instar larvae. (F)and can reveal expression patterns for uncharacterized genes.Wee-P4:GFP:Sec61 is localized in patches of the cytoplasm of(A) Representative fusion lines were blotted and probed withcells. Depicted are epithelial cells of a third instar larva. (G)anti-GFP antibodies. Single clear bands are seen in all casesWee-P2:GFP:Kis expression is widespread and localized to theand are approximately the correct predicted sizes. (B) Annucleus, as has been previously characterized for this gene.analysis of the Wee-P3:GFP:Cib fusion in homozygous Wee-Expression of Wee-P2:GFP:Kis is shown in an imaginal discP3 flies with an anti-Cib antibody indicates that the majorityat low magnification. (H) At higher magnification, discreteof the protein made from the locus is tagged with GFP. Therepuncta of Wee-P2:GFP:Kis fusion proteins in the nuclei areis a reduction in the total amount of protein made from thisvisible. Such puncta were noted in all cell types examined.locus in comparison to y w controls. (C) Wee-P20 is a fusionwith the uncharacterized but annotated gene CG9894. Nucleifrom third instar muscle nuclei are shown.
an important role in vesicle fusion at the neuromuscularjunction (NMJ; Bronk et al. 2001). RT-PCR using prim-ers specific for GFP and the open reading frame of below, indicates that the Wee-P1 insertion does not alterHsc70-4 indicates that a fusion mRNA is made and dem- Hsc70-4 function.onstrates that the native splicing apparatus recognizes We have examined the tissue localization pattern ofthe splice donor sites in the Wee-P (Figure 4A). Addi- Wee-P1:GFP:Hsc70-4 in larvae and found a broad and low-tional RT-PCR using primers designed to GFP and the level expression of GFP in all cells and a strong expressionexon composed of the 5� UTR of Hsc70-4 also indicates in tissues undergoing morphological changes, such asthat the 5� UTR exon is linked to this new mRNA and third instar imaginal discs (data not shown). Previousdemonstrates that the native splicing apparatus also rec- studies with RNA in situ have revealed that Hsc70-4 isognizes the splice acceptor sequences in the Wee-P (Fig- expressed in virtually all cells, with enrichment in tissuesure 4A). The fusion product between GFP and Hsc70-4 undergoing extensive rapid growth and changes inshould be a perfect fusion of GFP onto the Hsc70-4 shape (Perkins et al. 1990). Thus the Wee-P1:Hsc70-4protein without any additional alterations to Hsc70-4. fusion provides direct evidence that GFP fusions fromSequence analysis of our RT-PCR product indicates that Wee-Ps can reflect the correct tissue localization pat-such a perfect fusion has occurred. Western analysis of terns.Wee-P1 flies using an anti-GFP antibody indicates that Analysis of Wee-P1 also underscores that the correcta protein of the predicted �98 kD is formed (Figure subcellular localization can occur in Wee-P fusions.
Prior studies have demonstrated that Hsc70-4 is found3A). A genetic analysis of Wee-P1, described in detail
1438 P. J. Clyne et al.
Figure 4.—The Wee-P1:GFP:Hsc70-4 fusion revealsa subsynaptic localization atthe NMJ and a heat-shock-in-duced increase in nuclear ex-pression. (A) Splicing eventsthat generate the Wee-P1:GFP:Hsc70-4 fusion. (B) Wee-P1:GFP:Hsc-70-4 is concen-trated at the neuromuscularsynapse and shows subsynap-tic hot spots that resemble thedistribution of synaptic vesi-cles within this nerve termi-nal. (C) Heat-shock treat-ment leads to an increasein nuclear localization ofWee-P1:GFP:Hsc70-4. (Left)Third instar muscle nucleifrom a Wee-P1 heterozygotethat was not given a heatshock. (Right) Third instarmuscle nuclei from an animalthat was given a heat shock.(D) Heat-shock treatment ofWee-P1 flies correlates witha 400% increase in nuclearexpression of Wee-P1:GFP:Hsc70-4.
both in the cytoplasm and in the nucleus (Carbajal et kD) protein that is largely uncharacterized. The Larpprotein has a conserved La domain that is most closelyal. 1993) and that such a distribution for GFP is observed
in Wee-P1 (Figure 4C). Recent research has also demon- related to the yeast Sro9p gene (Chauvet et al. 2000).Sro9p is primarily cytoplasmic and thought to bindstrated that Hsc70-4 is enriched at the third instar neuro-
muscular synapse where it is involved in vesicle cycling mRNAs and modulate their translation (Sobel andWolin 1999). We observe that Wee-P5:GFP:Larp is(Bronk et al. 2001). In Wee-P1, GFP expression is en-
riched at the NMJ and, within the NMJ, Wee-P1:GFP: widely expressed in the embryo and larval stages andwithin cells is localized to the cell cytoplasm (data notHsc70-4 is localized to hot spots (Figure 4B).
This fusion also demonstrates appropriate trafficking shown). As a second example, Wee-P20 is a fusion withthe gene CG9894, which encodes a protein of unknownof a Wee-P:GFP fusion protein. In response to heat
shock, Hsc70-4 translocates to the nucleus (Carbajal function. This fusion is abundantly expressed and highlylocalized to the nucleus in a wide variety of cell typeset al. 1993). We subjected Wee-P1 animals to heat shock
and examined the fluorescence intensity of GFP in mus- although not in all cell types (Figure 3D).Wee-P insertions, in general, do not disrupt the genescle nuclei in animals before and after heat shock. We
observed a 400% increase in nuclear GFP fluorescence to which they fuse: For the Wee-P fusion events to bemost useful, it is important that the presence of theintensity following heat shock (Figure 4, C and D). To
control for nonspecific effects on protein translation Wee-P element not disrupt the function of the gene towhich the fusion has occurred. We have attempted toand translocation, we performed a similar analysis with
Wee-P114 and Wee-P4. In neither case did heat shock address these issues by undertaking a genetic analysisof a representative subset of Wee-P fusion lines (Tablealter GFP fluorescence intensity. Thus, the Wee-P1:GFP:
Hsc70-4 fusion traffics correctly in response to heat- 2). For nine of the fusion events that we analyzed geneti-cally, there was no observable phenotype in animalsshock stimulation.
Wee-P fusions can be used to probe the expression homozygous for the Wee-P insertion. In three cases weperformed our phenotypic analysis on the chromosomepatterns of uncharacterized genes: Our data examining
Wee-P fusions with previously characterized genes indi- bearing the Wee-P element placed in trans over eithera deficiency that uncovered the locus or a null allele ofcate that fusion proteins are stable and behave normally
within their endogenous cellular environment. On the the fused gene. In all three cases none of the predictedphenotypes were observed. Thus the absence of pheno-basis of these data, it is reasonable to suspect that fusions
with novel and previously uncharacterized genes may types in this random sampling of Wee-P lines suggeststhat Wee-P insertions are largely nonmutagenic.also reflect correct subcellular localization. For exam-
ple, we have isolated a fusion with Larp, a large (150- For some of the genes for which we have fusion events,
1439Tagging Genes at Their Native Loci
TABLE 2
Summary of genetic analysis on representative Wee-P lines
Published phenotype
Weak Strong hypomorphic Test for Wee-PLine Gene hypomorphic or null phenotypea References
Wee-P1 hsc70-4 Visible Lethal Viable over Df(3R)�356; Bronk et al. (2001)no visible phenotype
Wee-P2 kismet ND Lethal Homozygous viable; Daubresse et al. (1999)no visible phenotype
Wee-P3 larp ND ND Homozygous viable;no visible phenotype
Wee-P4 belle Fertility Lethal Viable over Df(3R)P13; Jones and Rawlsmales, females fertile (1988); Castrillon
et al. (1983)Wee-P26 mhc Visible Lethal Viable over P(1)k10423; O’Donnell and
no visible phenotype Bernstein (1988);O’Donnell et al. (1989)
Wee-P27 tropomyosinI ND Lethal Homozygous viable; Tetzlaff et al. (1996)no visible phenotype
Wee-P59 pabp2 Electrophysiology Lethal Homozygous viable; Spradling et al. (1999);electrophysiology not Sigrist et al. (2000)tested
Wee-P114 calmodulin Visible Lethal Homozygous viable; Spradling et al. (1999);no visible phenotype Nelson et al. (1997)
Wee-P118 cp1 Fertility and visible ND Homozygous viable; Gray et al. (1998)no visible phenotype;fertile
Wee-P283 ATPalpha ts viability; bang Lethal Homozygous viable at Schubiger et al. (1994);sensitivity all temperatures; not Feng et al. (1997)
bang sensitiveWee-P338 Pyk ND ND Homozygous viableWee-P345 Karybeta3 ND Lethal Homozygous viable; Spradling et al. (1999)
no visible phenotype
ND indicates that there are no published hypomorphic or null alleles for the gene.a “No visible phenotype” indicates that homozygous Wee-P flies have no observed morphological or obvious behavioral pheno-
types. “Homozygous viable” means that flies can be homozygous for the Wee-P chromosome.
an allelic series has been previously described. For exam- There are several possible explanations for the lackof observable phenotypes in the examples presentedple, null alleles of the chaperone gene Hsc70-4 die asabove. First, the added GFP exon may be inert. In thefirst instar larvae, strong hypomorphic alleles die as lateliterature there are now many examples of GFP-taggedlarvae-early pupae, and the weakest hypomorphic allelesproteins that are fully functional (e.g., Sweeney anddie as early pupae-adults. The few adults of the weakestDavis 2002). Alternatively, since both wild-type and GFPhypomorphic Hsc70-4 alleles that manage to eclose allincorporated transcripts are usually generated (Tablehave visibly rough eyes, bent wings, and thin, short bris-1), normal function may be contributed by the wild-tles (FlyBase 1999; Bronk et al. 2001). When Wee-type transcript. However, this may not account for theP1 is crossed to two different null alleles of Hsc70-4,lack of phenotype in examples where weak hypomor-transheterozygous, fertile adults eclose at a wild-typephic phenotypes should be observed such as Hsc70-4frequency and they show no morphological changes to(see above). A list of the Wee-P lines examined geneti-their eyes, bristles, or wings, suggesting that the pres-cally, as well as what is known about any alleles of theence of the GFP in Wee-P1 has not disrupted Hsc70-4fused genes, is given in Table 2.function. Another example is Wee-P6, which forms a
fusion with the protein Belle, an RNA helicase. Nullalleles of Belle are lethal while hypomorphic alleles pro-
DISCUSSIONduce sterile adults (Jones and Rawls 1988; Castrillonet al. 1993). When Wee-P6 is crossed to Df(3R)P13, trans- We have developed a broadly applicable techniqueheterozygotes of the Wee-P6/Df(3R)P13 genotype eclose for systematically tagging proteins with GFP on a ge-
nome-wide scale. Importantly, the tagged proteins areat wild-type ratios and both males and females are fertile.

1440 P. J. Clyne et al.
governed by their endogenous transcriptional and trans- and Wee-P1:GFP:Hsc70-4 in Figure 2 and Figure 4, re-lational controls. Fusion proteins generated with this spectively).technology show the predicted tissue-level expression Several important considerations must be taken intopattern and the correct subcellular localization allowing account when using Wee-P fusions. Most important ismicrodomains of protein localization to be visualized the question of the retention of wild-type protein func-in a living organism. We anticipate that this technology tion in the fused lines. Our phenotypic analysis of 12will enable a new generation of experiments to be pur- of the Wee-P lines indicated no alteration to functionsued in Drosophila. It may prove possible to use these and suggests that Wee-P insertions are largely nonmuta-reagents to study protein localization, trafficking, turn- genic. However, we anticipate that some fusion eventsover, concentration, and translation within an in vivo will alter protein function, mostly in those cases wheregenetic system without the caveat of protein overexpres- the GFP disrupts a critical domain of the protein. Al-sion that often precludes reliable experimental interpre- though exons are frequently modular, encoding distincttation. motifs within a gene (Long et al. 1995), there are in-
Wee-P technology is most significant if a tagged pro- stances to the contrary.tein retains all of the features of tissue expression and Our goal is to generate as many GFP-tagged proteinssubcellular localization for the native protein, and if the as possible with Wee-P elements. We anticipate makingfusion protein functions normally. Our analysis of Wee- these reagents freely available so that other workersP1:GFP:Hsc70-4 and other fusion lines demonstrates can perform similar screens for genes in their area ofthat Wee-Ps can accurately reflect the expression pat- interest, and we are constructing a website to facilitateterns of the genes they tag (Figures 2 and 4). In addition, widespread use of this technology.our findings that none of the representative 12 Wee-P
We thank G. Ruiz and S. Gandhi for helpful assistance. P.J.C. waslines analyzed have phenotypes suggests that Wee-Ps supported by a postdoctoral fellowship from the Medical Investigationare generally not mutagenic. Thus, Wee-P technology of Neurodevelopmental Disorders Institute at the University of Califor-
nia, Davis. S.T.S. is a Wellcome Prize Traveling Fellow (058327/Z/should allow for rapid and accurate assessments of the99/Z). This work was supported by a Burroughs Wellcome Youngtemporal and spatial patterns of genes.Investigator Award, a Merck Scholar Award, and a National InstitutesAlthough �40% of the GFP-expressing lines collectedof Health grant (44908-32374) to G.W.D.
in the initial screen are not true fusions, in the greatmajority of cases this fact can be discovered after justthe IPCR step. This is because the insertion site of the
LITERATURE CITEDWee-P element can immediately illustrate if a true pro-tein-trapping event has occurred or if some other event, Bier, E., H. Vaessin, S. Shepherd, K. Lee, K. McCall et al., 1989
Searching for pattern and mutation in the Drosophila genomesuch as fusion with a UTR exon, has occurred. Thus,with a P-lacZ vector. Genes Dev. 3 (9): 1273–1287.determining whether or not a GFP-expressing line is a
Boquet, I., R. Boujemaa, M. F. Carlier and T. Preat, 2000 Cibou-true fusion can be rapidly determined without a time- lot regulates actin assembly during Drosophila brain metamor-
phosis. Cell 102 (6): 797–808.consuming molecular analysis.Bronk, P., J. J. Wenniger, K. Dawson-Scully, X. Guo, S. Hong etA database of GFP-tagged proteins generated with
al., 2001 Drosophila Hsc70–4 is critical for neurotransmitterWee-P technology can have many potential applications. exocytosis in vivo. Neuron 30: 475–488.First, GFP-tagged proteins can reveal new patterns of Carbajal, E. M., J. F. Beaulieu, L. M. Nicole and R. M. Tanguay,
1993 Intramitochondrial localization of the main 70-kDa heat-gene expression for novel genes, as exemplified byshock cognate protein in Drosophila cells. Exp. Cell Res. 207Wee-P20:CG9894, as well as previously characterized (2): 300–309.
genes, as exemplified by the tracheal and chordotonal Castrillon, D. H., P. Gonczy, S. Alexander, R. Rawson, C. G.Eberhart et al., 1993 Toward a molecular genetic analysis ofexpression of WeeP:Cib (Figures 2 and 3). Second,spermatogenesis in Drosophila melanogaster: characterization ofthese reagents allow for subcellular protein localization male-sterile mutants generated by single P element mutagenesis.
to be characterized in detail in the living animal. Third, Genetics 135: 489–505.Cavener, D. R., and S. C. Ray, 1991 Eukaryotic start and stop transla-fusion events can tag specific splice variants of a gene,
tion sites. Nucleic Acids Res. 19 (12): 3185–3192.allowing for detailed analysis that is otherwise difficult. Chalfie, M., Y. Tu, G. Euskirchen, W. W. Ward and D. C. Prasher,Such isoform-specific tagging has occurred in our two 1994 Green fluorescent protein as a marker for gene expres-
sion. Science 263: 802–805.Wee-P:GFP:Kis lines, in which only the larger isoformChauvet, S., C. Maurel-Zaffran, R. Miassod, N. Jullien, J. Pradelhas been tagged. Fourth, the Wee-P will tag proteins
et al., 2000 dlarp, a new candidate Hox target in Drosophilaregardless of their size, enabling fusion events with large whose orthologue in mouse is expressed at sites of epithelium/
mesenchymal interactions. Dev. Dyn. 218: 401–413.proteins to be generated, which are otherwise difficultDaubresse, G., R. Deuring, L. Moore, O. Papoulas, I. Zakrajsekreagents to create. Our Wee-P:GFP:Kis lines, which have
et al., 1999 The Drosophila kismet gene is related to chromatin-tagged a 17-kb open reading frame, are also good exam- remodeling factors and is required for both segmentation and
segment identity. Development 126 (6): 1175–1187.ples of this point. Finally, patterns of GFP expressionDeutsch, M., and M. Long, 1999 Intron-exon structures of eukaryo-in specific cell types can be reagents to study develop-
tic model organisms. Nucleic Acids Res. 27 (15): 3219–3228.mental processes such as axon guidance, synaptic plas- Feng, Y., L. Huynh, K. Takeyasu and D. M. Fambrough, 1997 The
Drosophila Na,K-ATPase a-subunit gene: gene structure, pro-ticity, or tracheal development (see Wee-P3:GFP:Cib

1441Tagging Genes at Their Native Loci
moter function and analysis of a cold-sensitive recessive-lethal O’Donnell, P. T., and S. I. Bernstein, 1988 Molecular and ultra-mutation. Genes Funct. 1 (2): 99–117. structural defects in a Drosophila myosin heavy chain mutant:
FlyBase, 1999 The FlyBase database of the Drosophila genome differential effects on muscle function produced by similar thickprojects and community literature. Nucleic Acids Res. 27: 85–88 filament abnormalities. J. Cell Biol. 107 (6/1): 2601–2612.(http://flybase.bio.indiana.edu/). O’Donnell, P. T., V. L. Collier, K. Mogami and S. I. Bernstein,
Golic, K. G., and S. Lindquist, 1989 The FLP recombinase of yeast 1989 Ultrastructural and molecular analyses of homozygous-catalyzes site-specific recombination in the Drosophila genome. viable Drosophila melanogaster muscle mutants indicate thereCell 50: 499–509. is a complex pattern of myosin heavy-chain isoform distribution.
Gray, Y. H., J. A. Sved, C. R. Preston and W. R. Engels, 1998 Struc- Genes Dev. 3: 1233–1246.ture and associated mutational effects of the cysteine proteinase Perkins, L. A., J. S. Doctor, K. Zhang, L. Stinson, N. Perrimon et(CP1) gene of Drosophila melanogaster. Insect Mol. Biol. 7 (3): al., 1990 Molecular and developmental characterization of the291–293. heat shock cognate 4 gene of Drosophila melanogaster. Mol.
Jones, W. K., and J. M. Rawls, 1988 Genetic and molecular mapping Cell. Biol. 10 (6): 3232–3238.of chromosome region 85A in Drosophila melanogaster. Genetics Schubiger, M., Y. Feng, D. M. Fambrough and J. Palka, 1994 A120: 733–742. mutation of the Drosophila sodium pump �subunit gene results
Keller, A., S. T. Sweeney, T. Zars, C. J. O’Kane and M. Heisenberg, in bang-sensitive paralysis. Neuron 12 (2): 373–381.2002 Targeted expression of tetanus neurotoxin interferes with Sigrist, S. J., P. R. Thiel, D. F. Reiff, P. E. D. Lachance, P. Laskobehavioral responses to sensory input in Drosophila. J. Neurobiol. et al., 2000 Postsynaptic translation affects the efficacy and mor-50 (3): 221–233.
phology of neuromuscular junctions. Nature 405 (6790): 1062–Liao, G. C., E. J. Rehm and G. M. Rubin, 2000 Insertion site prefer-1065.ences of the P transposable element in Drosophila melanogaster.
Sobel, S. G., and S. L. Wolin, 1999 Two yeast La motif-containingProc. Natl. Acad. Sci. USA 97: 3347–3351.proteins are RNA-binding proteins that associate with polyribo-Long, M., C. Rosenberg and W. Gilbert, 1995 Intron phase corre-somes. Mol. Biol. Cell 10: 3849–3862.lations and the evolution of the intron/exon structure of genes.
Spradling, A. C., D. Stern, A. Beaton, E. J. Rhem, T. Laverty et al.,Proc. Natl. Acad. Sci. USA 92 (26): 12495–12499.1999 The Berkeley Drosophila Genome Project gene disruptionLukacsovich, T., Z. Asztalos, W. Awano, K. Baba, S. Kondo etproject: single P-element insertions mutating 25% of vital Dro-al., 2001 Dual-tagging gene trap of novel genes in Drosophilasophila genes. Genetics 153: 135–177.melanogaster. Genetics 157: 727–742.
Sweeney, S. T., and G. W. Davis, 2002 Unrestricted synaptic growthMorin, X., R. Daneman, M. Zavortink and W. Chia, 2001 A pro-in spinster-a late endosomal protein implicated in TGF-beta-medi-tein trap strategy to detect GFP-tagged proteins expressed fromated synaptic growth regulation. Neuron 36: 403–416.their endogenous loci in Drosophila. Proc. Natl. Acad. Sci. USA
Tetzlaff, M. T., H. Jaeckle and M. J. Pankratz, 1996 Lack of98 (26): 15050–15055.Drosophila cytoskeletal tropomyosin affects head morphogenesisMount, S. M., C. Burks, G. Hertz, G. D. Stormo, O. White et al.,
1992 Splicing signals in Drosophila: intron size, information and the accumulation of oskar mRNA required for germ cellcontent. Nucleic Acids Res. 20 (16): 4255–4262. formation. EMBO J. 15 (6): 1247–1254.
Nelson, H. B., R. G. Heiman, C. Bolduc, G. E. Kovalick, P. Whitley Wilson, C., R. K. Pearson, H. J. Bellen, C. J. O’Kane, U. Gross-et al., 1997 Calmodulin point mutations affect Drosophila devel- niklaus et al., 1989 P-element-mediated enhancer detection: anopment and behavior. Genetics 147: 1783–1798. efficient method for isolating and characterizing developmentally
Newmyer, S. L., and S. L. Schmid, 2001 Dominant-interfering regulated genes in Drosophila. Genes Dev. 3 (9): 1301–1313.Hsc70 mutants disrupt multiple stages of the clathrin-coated vesi-cle cycle in vivo. J. Cell Biol. 152 (3): 607–620. Communicating editor: K. Anderson

Top Related
Copyright © 2022 FDOKUMEN

