







Bahasa
Halaman
Hukum


Genomic clustering of cyanogenic glucoside biosyntheticgenes aids their identification in Lotus japonicus and suggeststhe repeated evolution of this chemical defence pathway
Adam M. Takos1, Camilla Knudsen1, Daniela Lai1, Rubini Kannangara1, Lisbeth Mikkelsen1, Mohammed S. Motawia1,
Carl E. Olsen2, Shusei Sato3, Satoshi Tabata3, Kirsten Jørgensen1, Birger L. Møller1 and Fred Rook1,*
1Plant Biochemistry Laboratory, Department of Plant Biology and Biotechnology, University of Copenhagen,
1871 Frederiksberg, Denmark,2Department of Basic Sciences and Environment, University of Copenhagen, 1871 Frederiksberg, Denmark, and3Kazusa DNA Research Insititute, Kisarazu, Japan
Received 11 May 2011; revised 20 June 2011; accepted 21 June 2011.*For correspondence (fax +45 35333300; e-mail [email protected]).
SUMMARY
Cyanogenic glucosides are amino acid-derived defence compounds found in a large number of vascular plants.
Their hydrolysis by specific b-glucosidases following tissue damage results in the release of hydrogen cyanide.
The cyanogenesis deficient1 (cyd1) mutant of Lotus japonicus carries a partial deletion of the CYP79D3 gene,
which encodes a cytochrome P450 enzyme that is responsible for the first step in cyanogenic glucoside
biosynthesis. The genomic region surrounding CYP79D3 contains genes encoding the CYP736A2 protein and
the UDP-glycosyltransferase UGT85K3. In combination with CYP79D3, these genes encode the enzymes that
constitute the entire pathway for cyanogenic glucoside biosynthesis. The biosynthetic genes for cyanogenic
glucoside biosynthesis are also co-localized in cassava (Manihot esculenta) and sorghum (Sorghum bicolor),
but the three gene clusters show no other similarities. Although the individual enzymes encoded by the
biosynthetic genes in these three plant species are related, they are not necessarily orthologous. The
independent evolution of cyanogenic glucoside biosynthesis in several higher plant lineages by the repeated
recruitment of members from similar gene families, such as the CYP79s, is a likely scenario.
Keywords: cyanogenic glucosides, gene clustering, cytochrome P450, Lotus japonicus, Manihot esculenta,
Sorghum bicolor.
INTRODUCTION
Plants have evolved a large spectrum of chemical defence
compounds, and while some are produced by only a limited
number of related species, others, such as cyanogenic glu-
cosides, are more widely distributed. Over 60 different cya-
nogenic glucosides, amino acid-derived a-hydroxynitrile
glucoside compounds, are known from over 2600 plant
species ranging from ferns to angiosperms, including a
disproportionately large number of crops (Seigler, 1975;
Conn, 1980; Jones, 1998). Upon tissue disruption, cyano-
genic glucosides are degraded by specific b-glucosidases,
resulting in the release of hydrogen cyanide, which provides
a defence mechanism against generalist herbivores (Morant
et al., 2008). Cyanogenesis is often regarded as an evolu-
tionarily ancient plant defence mechanism with a single
evolutionary origin implied due to the fact that all presently
identified enzymes for the first committed step, the conver-
sion of an amino acid into an oxime, are cytochrome P450s
belonging to the CYP79 family (for reviews, see Bak et al.,
2006; Bjarnholt and Møller, 2008).
The first genes encoding enzymes for cyanogenic gluco-
side biosynthesis were identified in Sorghum bicolor using
biochemical approaches (Koch et al., 1995; Bak et al., 1998a;
Jones et al., 1999), which provided the basis for the identi-
fication of genes in other plant species by sequence homo-
logy. Sorghum produces the aromatic cyanogenic glucoside
dhurrin, derived from tyrosine, and its biosynthesis requires
three sequential steps involving two cytochrome P450
enzymes and an UDP-glucosyltransferase. The first cyto-
chrome P450, CYP79A1, converts tyrosine into an oxime
intermediate, which is converted by the second cytochrome
ª 2011 The Authors 1The Plant Journal ª 2011 Blackwell Publishing Ltd
The Plant Journal (2011) doi: 10.1111/j.1365-313X.2011.04685.x

P450, CYP71E1, into a hydroxynitrile, which is then stabilized
by glucosylation catalysed by the UDP-glucosyltransferase
UGT85B1, resulting in dhurrin formation (Figure 1).
Similarly, in other plant species, members of the CYP79
family have been shown to catalyse the first step in
cyanogenic glucoside biosynthesis. In cassava (Manihot
esculenta), the cyanogenic glucosides linamarin and lot-
australin are synthesized from the amino acids valine and
isoleucine, respectively, involving the activity of CYP79D1
and CYP79D2 (Andersen et al., 2000). The model legume
Lotus japonicus contains two genes, CYP79D3 and
CYP79D4, that encode cytochrome P450 enzymes involved
in the production of linamarin and lotaustralin and related
non-cyanogenic b- and c-hydroxynitrile glucosides called
rhodiocyanosides (Forslund et al., 2004). Rhodiocyanoside
biosynthesis is thought to diverge from cyanogenic gluco-
side biosynthesis at the level of the hydroxynitrile interme-
diate, and natural variation for the presence or absence of
rhodiocyanosides involves a genetic locus that we refer to
here as Rho (Figure 1) (Bjarnholt et al., 2008). In white clover
(Trifolium repens), an adaptive defence polymorphism
controlling the presence or absence of cyanogenic gluco-
sides in natural populations is conferred by variation at the
Ac locus. Ac encodes the CYP79D15 enzyme, which cataly-
ses the first step in lotaustralin and linamarin biosynthesis
(Olsen et al., 2008). The production of oximes from amino
acids by members of the CYP79 family is also part of the
biosynthesis of non-cyanogenic defence compounds such
as glucosinolates and camalexin (Bak et al., 1998b; Hull
et al., 2000; Reintanz et al., 2001; Glawischnig et al., 2004).
Functional homologues of the sorghum CYP71E1 and
UGT85B1 enzymes are more difficult to identify because
they are members of large protein families and lack
conserved sequence motifs indicating their specific meta-
bolic function, and only recently have they been identified in
cassava (Jørgensen et al., 2011; Kannangara et al., 2011).
The legume Lotus japonicus is used as a genetic model to
study cyanogenesis, and we recently reported a high-
throughput genetic screen for cyanogenesis deficient
mutants (Takos et al., 2010). Here we identify the cyd1 locus
as encoding the CYP79D3 gene, providing genetic evidence
that CYP79D3 is the enzyme that produces cyanogenic
Figure 1. The hydroxynitrile glucoside biosynthetic pathways in S. bicolor and L. japonicus.
In S. bicolor, the synthesis of dhurrin from tyrosine involves CYP79A1 (Koch et al., 1995), CYP71E1 (Bak et al., 1998a) and UGT85B1 (Jones et al., 1999). L. japonicus
produces the cyanogenic glucosides linamarin (from valine) and lotaustralin (from isoleucine), as well as non-cyanogenic hydroxynitrile glucosides called
rhodiocyanoside A and D (from isoleucine). CYP79D3 is the predominant enzyme in leaves; CYP79D4 is expressed in the root (Forslund et al., 2004). The isoleucine-
derived 2-methylbutanal oxime intermediate is used in the synthesis of lotaustralin and rhodiocyanosides A and D. CYP736A2 is involved in the production of
cyanogenic glucosides, while rhodiocyanoside production depends on the product encoded by the Rho locus. The UDP-glucosyltransferases identified belong to the
UGT85 family.
2 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

glucosides in leaves of mature Lotus plants. The genomic
region containing CYP79D3 contains at least five loci encod-
ing enzymes involved in hydroxynitrile glucoside biosynthe-
sis. These include the CYP736A2 and UGT85K3 genes, which
complete the cyanogenic glucoside biosynthetic pathway,
and the genetic locus responsible for natural variation in
rhodiocyanoside biosynthesis (Rho). Clustering of genes
involved in cyanogenic glucoside biosynthesis was also
observed in cassava and sorghum, but the three genomic
regions are very diverse in structure. Based on the available
plant genome sequence data, it is a likely possibility that the
recruitment of these sets of genes for cyanogenic glucoside
synthesis represent independent events.
RESULTS
The cyanogenesis deficient1 mutant is defective in CYP79D3
A screen for mutants defective in cyanogenesis (Takos et al.,
2010) identified cyd1 as an acyanogenic mutant that, under
the screening conditions, did not contain hydroxynitrile
glucosides (either cyanogenic glucosides or rhodiocyano-
sides) in leaves. As synthesis of all hydroxynitrile glucosides
in the cyd1 mutant was severely affected, a defect in an early
step of the biosynthetic pathway, possibly in the formation of
the oxime from the amino acid precursors, was suggested
(Figure 1). Two paralogous cytochrome P450 genes,
CYP79D3 and CYP79D4, had been shown previously to
convert valine and isoleucine to the corresponding oximes,
and of these CYP79D3 is predominantly expressed in leaves
(Forslund et al., 2004). To determine the chromosomal
position of the cyd1 locus, the mutant was crossed with
L. japonicus cv. Gifu B-129 (Handberg and Stougaard, 1992).
Analysis of F2 plants for the acyanogenic phenotype dem-
onstrated close linkage of the recessive cyd1 locus with the
SSLP marker TM1185 on genomic contig CM0241 of chro-
mosome 3. In the L. japonicus genome sequence (Sato et al.,
2008; http://www.kazusa.or.jp/lotus), both CYP79D3 and
CYP79D4 are localized on contig CM0241 and estimated to be
240 kb apart. The sequences of the CYP79D3 (CM0241.700)
and CYP79D4 (CM0241.310) genes were determined in the
cyd1 mutant, and while both exons of the CYP79D4 gene
were intact, it was not possible to amplify the second exon of
CYP79D3 from the cyd1 mutant.
The Lotus genome database contains a 1926 bp fragment,
LjSGA_030566, that contains a sequence that is 98% iden-
tical at the nucleotide level to part of the CYP79D3 intron and
the entire second exon. LjSGA_030566 is thought to be a
pseudo-gene lacking the first exon, and its precise chromo-
somal location is presently unknown. A number of single
nucleotide differences, and most notably a 32 bp insertion in
the LjSGA_030566 sequence, distinguish it from CYP79D3.
Primers spanning this insertion site amplified two bands
from genomic DNA of the wild-type MG-20 (191 and 223 bp),
but only the larger LjSGA_030566-derived band was ampli-
fied from the cyd1 mutant (Figure 2a). Lack of a CYP79D3
transcript in the cyd1 mutant also indicated that the gene
was defective (Figure 2b).
To confirm that loss of a functional CYP79D3 gene was
responsible for the lack of cyanogenesis in the cyd1 mutant,
the gene was re-introduced by transforming the cyd1 mutant
with the CYP79D3 coding region under the control of the 35S
CaMV promoter. Cyanogenesis was restored in multiple
independent transformed lines as shown for line 5 in
Figure 2c. Complementation of the cyd1 phenotype was
further analysed by LC-MS of leaf extracts, and showed not
only restored biosynthesis of lotaustralin and linamarin, but
also of the non-cyanogenic hydroxynitrile glucosides rhod-
iocyanoside A and D (Figure 2d).
Contig CM0241 contains a UGT85 that is involved
in cyanogenic glucoside biosynthesis
The first UDP-glucosyltransferase identified as glucosylating
hydroxynitriles was SbUGT85B1, which is involved in
dhurrin production in sorghum (Jones et al., 1999). In
L. japonicus, the CM0241 contig contains a member of an-
other UGT85 sub-family (gene CM0241.610) that has been
assigned the name UGT85K3 following the recommenda-
tions for nomenclature of Mackenzie et al. (1997). A second
gene, UGT85K2 (T45I18.30), encodes a protein that is 86%
identical at the amino acid level to UGT85K3. UGT85K2 is
located further down on chromosome 3 at approximately
31.6 cM (approximately 18 cM from CM0241). Both
UGT85K2 and UGT85K3 show 41% amino acid identity with
SbUGT85B1, and 56% identity with UGT85K4 and UGT85K5,
which are involved in cyanogenic glucoside production in
cassava (Kannangara et al., 2011).
The coding regions of UGT85K2 and UGT85K3 were
amplified as cDNAs from MG-20 and expressed in Escher-
ichia coli. To test the ability of UGT85K2 and UGT85K3 to
glucosylate the different hydroxynitriles, E. coli extracts
were incubated with 20 mM of the putative acceptors and
3.3 lM UDP-[14C]-glucose as donor. The reaction products
formed were visualized on TLC plates and verified by LC-MS
analysis. Both UGT85K2 and UGT85K3 were able to glu-
cosylate acetone cyanohydrin (the linamarin aglycone), and
2-hydroxy-2-methylbutyronitrile (the lotaustralin aglycone)
(Figure 3).
The CYP736A2 gene completes a functional biosynthetic
pathway for cyanogenic glucoside biosynthesis in Lotus
japonicus
In sorghum, the second step in cyanogenic glucoside bio-
synthesis is mediated by CYP71E1 (Bak et al., 1998a), and a
functional homologue, CYP71E7, was recently characterized
in cassava (Jørgensen et al., 2011). CYP71E7 was identified
using CYP71E1 as the query in a BLASTp search of the cas-
sava genome sequence, and the proteins show 49% identity.
Using either CYP71E1 or CYP71E7 to query the Lotus
Cyanogenic glucoside biosynthetic gene clusters 3
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

genome sequence identified members of the CYP83E sub-
family as the most similar, with identity scores of approx-
imately 40%. Using CYP71E7 as the query, the highest
identity score (42%) was obtained for CYP83E4 (CM1089.70),
and no member of the CYP71E sub-family was identified in
the Lotus genome. However, the CM0241 contig contains a
member of another cytochrome P450 gene family named
CYP736A2 (CM0241.850), as well as three closely related
pseudo-genes (CM0241.540, CM0241.880 and CM0241.940).
The CYP736A2 amino acid sequence shows approximately
35% identity with CYP71E1 and CYP71E7. The CYP71, CYP83
and CYP736 families all belong to the CYP71 clan, which
contains over 50 CYP families (Nelson and Werck-Reichhart,
2011). Within the CYP71 clan, the three families are part of
the same sub-branch formed by seven CYP families. Their
phylogenetic relationship is shown in Figure 4. The genome
sequences of sorghum, cassava and Lotus were queried
using the CYP71E1, CYP71E7, CYP736A2 and CYP83E4
sequences. The sequences with the highest homology
scores from each BLASTp search were collected, and a
phylogenetic tree was constructed using the MEGA5
package (Figure 4) (Tamura et al., 2011). Additional protein
sequences of members characteristic for the relevant CYP71
clan families, mostly from other plant species, were included
in the analysis. Families just outside this sub-branch of the
CYP71 clan are represented by CYP76A3, CYP80A1,
CYP92A1 and CYP786A1, and anchor the tree.
As shown in Figure 4, the CYP71 and CYP83 families are
most closely related to each other, and group with the
CYP726 and CYP99 families. These four families are now
Figure 2. The cyd1 mutant is defective in CYP79D3.
(a) PCR amplification showing the 32 bp insertion polymorphism between
CYP79D3 and LjSGA_030566 using MG-20 and cyd1 genomic DNA as
template.
(b) Absence of the CYP79D3 transcript in the cyd1 mutant was determined by
quantitative RT-PCR.
(c) Restoration of cyanogenesis in leaves of the cyd1 mutant transformed with
a 35S:CYP79D3 construct was detected using HCN-sensitive Feigl–Anger
paper.
(d) Extracted ion chromatogram of the MG-20 parental line, the cyd1 mutant,
and the cyd1 mutant complemented with the 35S:CYP79D3 construct. Apical
leaves of 5-month-old tissue culture-grown plants were analysed. Extracted
ion peaks are for sodium adducts: linamarin (m/z 270, cyan); rhodiocyanoside
A and D (m/z 282, red); lotaustralin (m/z 284, blue).
(a) (b)
Figure 3. Glucosylation of hydroxynitrile substrates by UGT85K2 and
UGT85K3.
(a) Activity of E. coli-expressed UGT85K2 and UGT85K3 on acetone cyano-
hydrin (AC) and 2-hydroxy-2-methylbutyronitrile (HBN) substrates using UDP-
[14C]-glucose (G) as donor and analysed by TLC.
(b) Unlabelled standard for linamarin (lin) and lotaustralin (lot) run on the
same plate and visualized following charring by H2SO4.
4 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x
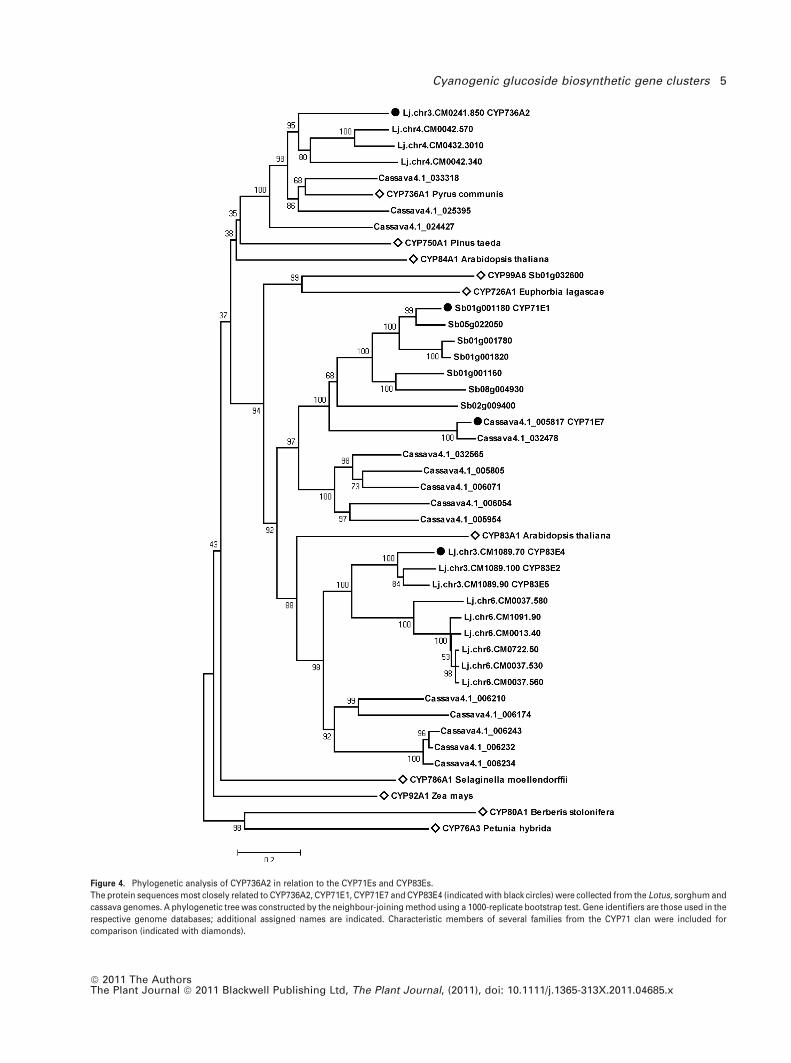
Figure 4. Phylogenetic analysis of CYP736A2 in relation to the CYP71Es and CYP83Es.
The protein sequences most closely related to CYP736A2, CYP71E1, CYP71E7 and CYP83E4 (indicated with black circles) were collected from the Lotus, sorghum and
cassava genomes. A phylogenetic tree was constructed by the neighbour-joining method using a 1000-replicate bootstrap test. Gene identifiers are those used in the
respective genome databases; additional assigned names are indicated. Characteristic members of several families from the CYP71 clan were included for
comparison (indicated with diamonds).
Cyanogenic glucoside biosynthetic gene clusters 5
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x
considered to have merged due to expansion of the CYP71
family as more sequences have become available (Nelson
and Werck-Reichhart, 2011). The CYP736 family containing
the Lotus CYP736A2 gene is not found in monocots, and
consequently no sorghum homologues were identified. The
CYP736 family is most closely related to the CYP84 and
CYP750 families, and is clearly distinct from the CYP71E sub-
family (Figure 4).
The genomic co-localization of CYP79D3 and UGT85K3
with CYP736A2 suggested that the latter might encode the
second step of the cyanogenic glucoside pathway. To test
this possibility, all three genes were cloned into the
pJAM1502 vector under the control of the CaMV 35S
promoter, and their activity was tested by Agrobacterium-
mediated transient expression in Nicotiana benthamiana.
Testing the L. japonicus genes separately and in various
combinations showed that only the triple combination of
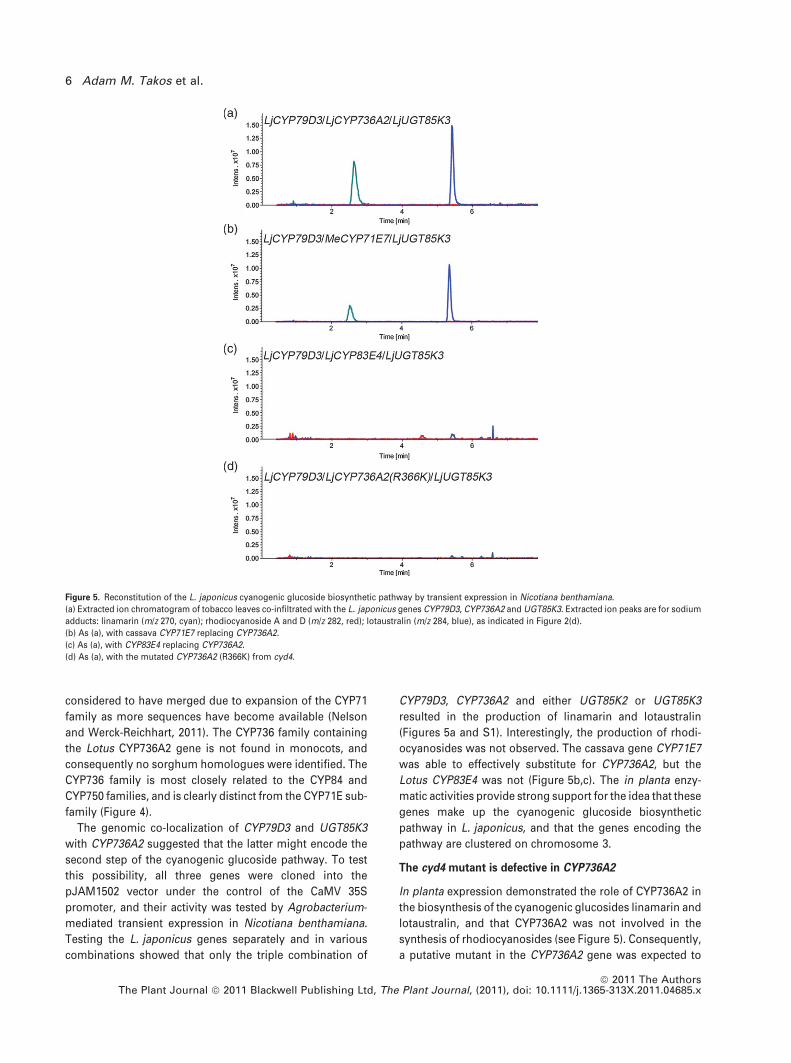
CYP79D3, CYP736A2 and either UGT85K2 or UGT85K3
resulted in the production of linamarin and lotaustralin
(Figures 5a and S1). Interestingly, the production of rhodi-
ocyanosides was not observed. The cassava gene CYP71E7
was able to effectively substitute for CYP736A2, but the
Lotus CYP83E4 was not (Figure 5b,c). The in planta enzy-
matic activities provide strong support for the idea that these
genes make up the cyanogenic glucoside biosynthetic
pathway in L. japonicus, and that the genes encoding the
pathway are clustered on chromosome 3.
The cyd4 mutant is defective in CYP736A2
In planta expression demonstrated the role of CYP736A2 in
the biosynthesis of the cyanogenic glucosides linamarin and
lotaustralin, and that CYP736A2 was not involved in the
synthesis of rhodiocyanosides (see Figure 5). Consequently,
a putative mutant in the CYP736A2 gene was expected to
Figure 5. Reconstitution of the L. japonicus cyanogenic glucoside biosynthetic pathway by transient expression in Nicotiana benthamiana.
(a) Extracted ion chromatogram of tobacco leaves co-infiltrated with the L. japonicus genes CYP79D3, CYP736A2 and UGT85K3. Extracted ion peaks are for sodium
adducts: linamarin (m/z 270, cyan); rhodiocyanoside A and D (m/z 282, red); lotaustralin (m/z 284, blue), as indicated in Figure 2(d).
(b) As (a), with cassava CYP71E7 replacing CYP736A2.
(c) As (a), with CYP83E4 replacing CYP736A2.
(d) As (a), with the mutated CYP736A2 (R366K) from cyd4.
6 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

show no or reduced levels of cyanogenic glucosides but
wild-type levels of rhodiocyanosides, resulting from the
presence of an intact Rho gene (Figure 1). One of the lines
obtained in the genetic screen for cyanogenesis-deficient
mutants, referred to here as cyd4, displayed the predicted
metabolic profile, with a >60% reduction in lotaustralin and
wild-type levels of rhodiocyanosides (Figure 6). Sequencing
of the CYP736A2 gene in the cyd4 mutant showed a G fi A
nucleotide substitution, changing the arginine at position
366 to a lysine. R366 in CYP736A2 is the invariant arginine
residue of the EXXR motif that is present in virtually all
cytochrome P450 proteins, and is essential for formation of
the tertiary structure (Hasemann et al., 1995; Rupasinghe
et al., 2006). When the mutated version of CYP736A2 was
cloned as a cDNA from cyd4 and transiently expressed in
Nicotiana benthamiana in combination with CYP79D3 and
UGT85K3, no production of either linamarin or lotaustralin
was observed (Figure 5d). These data provide genetic sup-
port for the role of CYP736A2 in the biosynthesis of cyano-
genic glucosides in L. japonicus.
Expression of cyanogenic glucoside biosynthetic genes in
relation to leaf age
Quantitative real-time PCR was used to examine the expres-
sion profile of CYP736A2, UGT85K2 and UGT85K3 in com-
parison with that of CYP79D3. Characteristically, CYP79D3
expression is related to leaf age, with the highest expression
observed in the apical leaves (Figure 7) (Forslund et al.,
2004). CYP736A2 expression showed a similar expression
profile to that observed for CYP79D3, with transcript levels
decreasing in older leaves. The UGT85K2 and UGT85K3 gene
expression levels did not change with leaf age. Although the
genes are not strictly co-regulated, the overlap in expression
pattern of CYP79D3, CYP736A2, UGT85K2 and UGT85K3
supports the view that these genes encode the enzymes of
the pathway for cyanogenic glucoside biosynthesis in
L. japonicus.
Natural variation in rhodiocyanoside biosynthesis is genet-
ically linked
We previously investigated natural variation in hydroxynit-
rile glucoside content in L. japonicus accessions, and
showed that absence of the non-cyanogenic rhodiocyano-
sides A and D in the MG-74 accession (see Figure 1) was due
to a single recessive genetic locus named rho (Bjarnholt
et al., 2008). The map position of rho was initially established
using 30 re-selected rho F2 lines from a MG-74 · MG-20
cross, and all showed the MG-74 genotype at marker
TM1185 on chromosome 3. The mapping population was
expanded to 350 F2 lines by initially selecting recombinants
between markers TM0436 (at 10.5 cM) and TM0403 (at
24 cM). These two markers were polymorphic between MG-
20 and MG-74 and flanked the region containing rho. The
recombinant F2 lines, and, where necessary, their segregat-
ing F3 progeny, were analysed for rhodiocyanoside pro-
duction, and genotyped using additional markers. This
established that rho was localized between markers TM1185
on contig CM0241 (at 13.3 cM) and TM0450 (at 15.3 cM). The
genomic sequence in this region contains most of contig
CM0241 but is discontinuous between CM0241 and BAC
clone TM0450, and the identity of the Rho gene remains to
be established.
Clustering of genes involved in cyanogenic glucoside
biosynthesis in the Lotus, cassava and sorghum genomes
The clustering of the genes for biosynthesis of cyanogenic
glucosides and related hydroxynitrile glucosides in the
genome of L. japonicus prompted the analysis of additional
plant genomes of cyanogenic species. Such an analysis is
possible in cassava and sorghum due to recent progress in
the availability of genome sequences (Paterson et al., 2009;
Cassava Genome Project 2010, http://www.phytozome.net/
cassava) and identification of CYP71 and UGT85 homo-
logues involved in cyanogenic glucoside biosynthesis in
cassava (Jørgensen et al., 2011; Kannangara et al., 2011).
In the sorghum genome sequence (available at http://
www.phytozome.net/sorghum), the three genes previously
identified as encoding the cyanogenic glucoside biosynthe-
tic pathway are clustered within a 104 kb region localized at
the top of chromosome 1 (Figure 8). Figure 8 also shows an
uncharacterized member of the CYP71E sub-family,
Figure 6. The cyd4 mutant defective in CYP736A2 contains reduced levels of
cyanogenic glucosides.
(a) Extracted ion chromatogram of the cyd4 mutant. Extracted ion peaks are
for sodium adducts: linamarin (m/z 270, cyan); rhodiocyanoside A and D (m/z
282, red); lotaustralin (m/z 284, blue), as indicated in Figure 2(d).
(b) Quantitative comparison of hydroxynitrile glucoside content in leaves of
MG-20 wild-type and cyd4. Values are the means of five biological repli-
cates � SE.
Cyanogenic glucoside biosynthetic gene clusters 7
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

Sb01g001160, located to the left of CYP71E1 (Sb01g001180),
that has 61% amino acid identity to CYP71E1 (see also
Figure 4). The region generally shows low gene density, but
also contains an alcohol dehydrogenase (Sb01g001170) and
two putative transposases of the hAT element superfamily.
Clustering of the biosynthetic genes for cyanogenic glu-
coside metabolism was also observed in cassava; a 100 kb
region of the 4.6 Mb Scaffold08265 (genome draft version
Cassava4.1; http://www.phytozome.net/cassava) is shown in
Figure 8. In cassava, two cytochrome P450 genes of the
Figure 7. Expression of the L. japonicus cyanogenic glucoside biosynthetic genes in relation to leaf age, determined by quantitative RT-PCR.
Values are relative to the expression level in the 3rd leaf, and are the means of three biological replicates � SE.
(a) Expression of CYP79D3.
(b) Expression of CYP736A2.
(c) Expression of UGT85K3.
(d) Expression of UGT85K2.
Figure 8. Schematic representation of the clustering of cyanogenic glucoside biosynthetic genes in the genomes of L. japonicus, S. bicolor and M. esculenta.
Functional genes are presented by arrows indicating their orientation. Confirmed genes in cyanogenic glucoside biosynthesis are labelled above each bar, with
CYP79 genes in pink, CYP71E and CYP736 genes in green, and UGT85 genes in blue. The three CYP736A2-like pseudo-genes are indicated below the L. japonicus
bar, as is the additional CYP71 in S. bicolor. The Rho locus is within 2 cM of the CM0241 contig. The remaining genes are numbered and annotated as follows: (1)
acireductone dioxygenase, (2) nucleic acid binding, OB-fold, (3,4) hypothetical proteins, (5) ribonuclease H, (6) short-chain dehydrogenase/reductase, (7,8) putative
transposases, (9) leucine-rich repeat receptor-like protein kinase, (10,11) a/b-fold hydrolases, (12) hypothetical protein, (13–15) putative hydroxynitrile lyases, (16)
hypothetical protein.
8 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

CYP79 family, CYP79D1 and CYP79D2, are known, and the
CYP79D2 gene (cassava4.1_005079) is part of the gene
cluster, which also contains the two CYP71E paralogues
(Andersen et al., 2000; Jørgensen et al., 2011) and the UDP-
glucosyltransferase genes UGT85K4 and UGT85K5 (Kann-
angara et al., 2011). The CYP79D1 gene (cassava4.1_005059)
is not part of the gene cluster and is localized on Scaf-
fold08167, which is 356 kb long. The CYP79D1 and CYP79D2
genes are each adjacent to a highly similar leucine-rich repeat
receptor-like protein kinase gene (>90% identity), suggesting
a duplication event. In contrast to the gene clusters in Lotus
and sorghum, the one in cassava is relatively gene-rich. Of
the additional genes in the region, a set of three genes with
homology to hydroxynitrile lyases is noteworthy, as these
may play a role in cyanogenic glucoside catabolism (cas
sava4.1_027709, cassava4.1_031085 and cassava4.1_014254,
numbered 13–15 in Figure 8).
The region of the cyanogenic glucoside biosynthetic gene
cluster in Lotus is the most extended in terms of overall size.
Only a limited number of functional genes are present,
including a putative aci-reductone dioxygenase (CM0241.-
600) and a retrotransposon-related ribonuclease H domain-
containing protein (CM0241.860), while additional partial
gene fragments related to retroelements are also present
(not indicated in Figure 8). Although clustering of genes for
cyanogenic glucoside biosynthesis is observed in all three
species, no other points of similarity could be identified, and
the regions differ substantially in size, gene density, the
identity of additional genes and the presence of transposon-
related sequences.
DISCUSSION
CYP79D3 as shared first enzyme in the biosynthesis of
hydroxynitrile glucosides
Cyanogenesis is the release of toxic HCN following tis-
sue damage and hydrolysis of cyanogenic glucosides, and
results from the inherent instability of the a-hydroxynitri-
le aglycones. In L. japonicus, the isoleucine-derived a-
hydroxynitrile glucoside lotaustralin co-occurs with the
non-cyanogenic compounds rhodiocyanoside A (a c-hy-
droxynitrile glucoside) and rhodiocyanoside D (a b-hy-
droxynitrile glucoside) (Forslund et al., 2004). We previously
reported natural variation in rhodiocyanoside content in
L. japonicus, and a diversification of the hydroxynitrile
glucoside biosynthetic pathway at the level of the nitrile
intermediate was suggested based on biochemical consid-
erations (Bjarnholt et al., 2008). The present study of the cyd1
mutant provides genetic evidence for the idea that the first
enzymatic step is shared. The cyd1 mutant was previously
identified in a leaf-based screen for cyanogenesis-deficient
mutants, and metabolite analysis showed that it contains
neither cyanogenic glucosides nor the related rhodiocyano-
sides (Takos et al., 2010). This suggested a defect early in the
biosynthetic pathway,possiblyat the formationof the oximes
from the amino acid precursors valine and isoleucine. The
map position of the cyd1 mutant and the absence of the sec-
ond exon indicate that cyd1 contains a defect in the CYP79D3
gene encoding a leaf-expressed oxime-producing cyto-
chrome P450 enzyme. Complementation of the cyd1 mutant
with a functional CYP79D3 gene restored the biosynthesis of
all hydroxynitrile glucosides.
Genomic clustering identified the CYP736A2 and UGT85K3
genes
The genomic region with the genes encoding CYP79D3 and
its paralogue CYP79D4 also contained candidate genes
for the remaining two steps in cyanogenic glucoside bio-
synthesis in L. japonicus: the UDP-glucosyltransferase
UGT85K3 and the cytochrome P450 CYP736A2. Both
UGT85K3 and the closely related UGT85K2 were able to
glucosylate the hydroxynitrile aglycones of linamarin and
lotaustralin in vitro. No close homologue of the sorghum
CYP71E1 or cassava CYP71E7 was identified in the Lotus
genome sequence, and the candidacy of CYP736A2 was
suggested by the observed genomic co-localization. The
activity of CYP736A2 was demonstrated by reconstituting
the Lotus biosynthetic pathway using transient co-expres-
sion in N. benthamiana. The triple combination of CYP79D3,
CYP736A2 and UGT85K3 produced the cyanogenic gluco-
sides linamarin and lotaustralin in tobacco. The cyd4 mutant
provided genetic evidence for the role of CYP736A2 in cya-
nogenic glucoside biosynthesis. The CYP736A2 protein in
the cyd4 mutant is defective in the invariant arginine of the
EXXR motif (Hasemann et al., 1995), and its substitution by a
lysine is likely to result in improper protein folding, thus
explaining the observed lack of enzymatic activity. The cyd4
mutant showed a large reduction in cyanogenic glucoside
content but wild-type levels of rhodiocyanosides, indicating
that CYP736A2 is specific for cyanogenic glucoside biosyn-
thesis. The metabolic profile of the cyd4 mutant most likely
reflects the activity of the Rho enzyme, primarily involved in
producing rhodiocyanosides but also capable of some cya-
nogenic glucoside production, although the possible exis-
tence of a third oxime-metabolizing enzyme cannot be
excluded. Consistent with the idea that the Rho enzyme does
not exclusively produces rhodiocyanosides is the observa-
tion that none of the available cyd mutants or any of eighty
natural Lotus accessions produce rhodiocyanosides in the
absence of lotaustralin. Identification and molecular char-
acterization of the Rho gene is required to confirm this.
Gene clustering in cyanogenic glucoside biosynthesis
Our work in L. japonicus demonstrated that the biosynthetic
genes for cyanogenic glucoside metabolism were co-local-
ized in the genome. Additional gene clusters containing the
genes for the biosynthesis of cyanogenic glucosides were
observed in sorghum and cassava. The regions differed
Cyanogenic glucoside biosynthetic gene clusters 9
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

considerably in size and structure, with only the presence of
the cyanogenic glucoside biosynthetic genes in common.
The Lotus cluster is 355 kb long when including CYP79D4
but not considering the Rho locus. The region containing
CYP79D3, CYP736A2 and UGT85K3 is 160 kb long, larger
than the 104 kb region containing the corresponding genes
in sorghum, or the 83 kb region observed in cassava. The
cluster in cassava is relatively gene-dense and contains a
total of 13 genes, including two CYP71E and two UGT85K
genes. The diverse genomic structure of the three gene
clusters suggests they have independent origins.
While gene clusters originating from repeated tandem
duplication events are common in eukaryotes, the genetic
and physical clustering of non-homologous genes that are
part of the same biosynthetic pathway is rare. In plants, such
clustering of non-homologous genes has only been
described for a number of chemical defence compounds,
mostly terpenoids (reviewed by Chu et al., 2011). Genes for
benzoxazinoid biosynthesis in maize (Zea mays) are
co-localized within a 6 cM region on chromosome 4 (Frey
et al., 1997), genetic loci for avenacin biosynthesis in oat
(Avena sativa) are clustered within 3.6 cM of the b-amyrin
synthase gene AsbAS1 encoding the first committed
enzyme of the pathway (Qi et al., 2004), and a 168 kb region
containing genes for the biosynthesis of momilactones has
been identified in rice (Oryza sativa) (Shimura et al., 2007).
Rice also contains a second diterpenoid biosynthetic gene
cluster that is multi-functional, containing genes for the
production of two distinct sets of phytoalexins (Swamina-
than et al., 2009). Arabidopsis contains a gene cluster for
synthesis of the triterpene thalianol (Field and Osbourn,
2008).
It is presently unclear how metabolic gene clusters arise
and why gene clustering is observed for some plant
secondary metabolic pathways, for example the biosynthe-
sis of terpenoids and cyanogenic glucosides, but not for
others, such as anthocyanin or glucosinolate biosynthesis.
Several selective advantages that may drive and maintain
the clustering of non-homologous genes in the biosynthesis
of plant defence compounds have been proposed (Gierl and
Frey, 2001; Qi et al., 2004; Chu et al., 2011). Clustering would
allow inheritance of the defensive trait as a single functional
unit, and in polymorphic populations this advantage could
drive the clustering of genes following their functional
evolution. Similarly, clustering of the genes of a biosynthetic
pathway with toxic reaction intermediates would reduce the
likelihood of its disruption by segregation. Clustering may
also allow coordinated expression of all genes in the
pathway, potentially by regulation at the chromatin level.
These proposed selective advantages for gene clusters are
not mutually exclusive, and may each contribute to estab-
lishing or maintaining this genomic organization. For exam-
ple, a two-stage selection process was suggested for cluster
formation in prokaryotes, where the proximity of genes
resulting from selection driving genes together would
establish the conditions for a separate selection for co-
regulation and operon assembly (Martin and McInerney,
2009).
When evaluating these selective advantages in relation to
gene clustering in the cyanogenic glucoside biosynthetic
pathway, the toxicity of intermediates is the most convinc-
ing amongst them. Cyanogenesis is a two-component
system, and inheritance as a single defence unit would
require the b-glucosidase to be co-localized with the bio-
synthetic genes. In L. japonicus, some linkage is observed
between LjBGD2 (Takos et al., 2010; T33P07.150) and the
biosynthetic genes, they are approximately 20 cM apart.
However in sorghum, the b-glucosidase responsible for the
hydrolysis of dhurrin is localized on chromosome 8
(Sb08g007570) while the biosynthetic genes are localized
on chromosome 1. Operon-like gene clusters that facilitate
coordinated expression of genes at the chromatin level have
been suggested (Qi et al., 2004; Field and Osbourn, 2008). In
L. japonicus, the CYP79D3 and CYP79D4 genes are differen-
tially expressed, while UGT85K2 and UGT85K3 expression
levels do not follow leaf age, arguing against co-ordination
of gene expression driving the clustering of genes in
cyanogenic glucoside biosynthesis. In contrast, the toxicity
of the intermediates in the biosynthesis of cyanogenic
glucosides has been experimentally demonstrated for the
dhurrin biosynthetic pathway. Introduction of the sorghum
genes CYP79A1 and CYP71E1 in Arabidopsis resulted in
stunted plants with pleiotropic phenotypes, which were
alleviated by the subsequent introduction of UGT85B1,
resulting in dhurrin production and restoration of plant
growth (Kristensen et al., 2005).
The CYP79 family and the repeated evolution of cyanogenic
glucoside biosynthesis
The complete set of genes required for the biosynthesis of
cyanogenic glucosides has now been identified in three
plant species: the monocot S. bicolor and the dicots
M. esculenta and L. japonicus. The genes encoding the
different enzymes in these species are related but not nec-
essarily orthologous. This suggests that cyanogenic gluco-
side biosynthesis has evolved independently in several
higher plant lineages by recruiting members from similar
gene families such as the CYP79s.
Members of the CYP79 family characteristically produce
oximes from amino acids in the biosynthetic pathways of
several chemical plant defence compounds, and besides
cyanogenic glucosides these include glucosinolates and
camalexin (Bak et al., 1998b; Hull et al., 2000; Reintanz et al.,
2001; Glawischnig et al., 2004). Presently available sequence
data show the first appearance of the CYP79 family in
angiosperms, as is the case for the CYP71 family (Nelson and
Werck-Reichhart, 2011). Because of the high substrate
specificity of the oxime-producing CYP79 enzyme, it is
10 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

regarded as the most selective step in determining which
cyanogenic glucoside is produced in a certain plant species
(Bjarnholt and Møller, 2008). A small CYP79 gene family is
present in higher plant genomes, including non-cyanogenic
species such as soybean (Glycine max) and poplar (Populus
trichocarpa). The biological role of most of these CYP79
genes, including additional CYP79 genes in cyanogenic
species, remains to be established. Phylogenetic analysis
shows that the CYP79 enzymes involved in cyanogenic
glucoside biosynthesis are more closely related to other
CYP79s in the same plant species or lineage than to CYP79s
involved in cyanogenic glucoside biosynthesis in more
distant plant species (Figure S2). For instance, CYP79D3
and CYP79D4 from L. japonicus are part of a legume-specific
clade, and only CYP79D15, which produces cyanogenic
glucosides in white clover (T. repens), is likely to be orthol-
ogous (>90% identity).
The independent evolution of a similar function from
homologous genes is referred to as ‘repeated evolution’,
and is thought to represent the majority of convergent
evolution events in plant secondary metabolism (Cseke
et al., 1998; Pichersky and Lewinsohn, 2011). For example, a
polyphyletic origin was proposed for the evolution of the
pyrrolizidine alkaloid-mediated defence system in angio-
sperms (Reimann et al., 2004). The first enzyme in this
pathway, homospermidine synthase, was independently
recruited from deoxyhypusine synthase at least four times
by loss of its ability to bind its protein substrate eIF5A. The
first enzyme in the synthesis of benzoxazinoid pesticides,
BX1, and the indole-3-glycerol phosphate lyase enzyme
(IGL) both produce indole, as an intermediate and as a
volatile defence compound, respectively, and are thought to
have originated from independent gene duplications of the
a-subunit of tryptophan synthase (Frey et al., 2000).
The ability to produce oximes from specific amino acids
seems to be the overall characteristic of the CYP79 family. It
has been suggested that the glucosinolate defence pathway,
which has evolved independently on at least two occasions,
originated from the cyanogenic glucoside pathway (Rodman
et al., 1998), but it can equally be said that both indepen-
dently evolved from the predisposition of members of the
CYP79 family to produce oximes from amino acids and the
need to convert the oximes into more stable and less
reactive compounds.
Broad substrate specificity facilitated the repeated evolu-
tion of cyanogenic glucoside biosynthesis
In contrast with the CYP79s, members of the CYP71 and UGT
families often have diverse and broad substrate specificities,
and it has been argued that enzymatic versatility enables a
limited number of glucosyltransferase genes to produce the
large number of glucosides found in plants (Jones et al.,
1999; Hansen et al., 2003; Schwab, 2003; Bjarnholt and
Møller, 2008; Jørgensen et al., 2011). Broad substrate spec-
ificity or promiscuity of enzymes provide an immediately
accessible starting point for the evolution of new functions
(Khersonsky and Tawfik, 2010), and would facilitate the
repeated evolution of cyanogenic glucoside biosynthesis
following the emergence of a more specific signature
enzyme, here for example a CYP79. It has also been
suggested that the emergence of a signature enzyme by
duplication and functional divergence could ‘seed’ the
formation of a metabolic gene cluster (Chu et al., 2011).
The independent evolution of cyanogenic glucoside
biosynthesis is most clearly demonstrated by the identifi-
cation of CYP736A2 in L. japonicus. Although it could be
argued that L. japonicus is exceptional in not possessing a
CYP71E orthologue, the primary basis for identifying
cyanogenic glucoside biosynthetic genes in plant species
to date has been their homology to the original biochem-
ically identified sorghum genes. Like the CYP79 and CYP71
families, CYP736s belong to the wider CYP71 clan. The
CYP736 family shows an early emergence in plant evolu-
tion and is first seen in liverworts, but is also absent from
several plant lineages including monocots (Nelson and
Werck-Reichhart, 2011). Recent reports connect the CYP736
family with defence responses against microbial patho-
gens in grape (Vitis vinifera) (Cheng et al., 2010), and using
microarray data, with nematode and Phytophthora infec-
tion and with nodulation in soybean (Guttikonda et al.,
2010).
Convergent evolution of cyanogenic glucoside biosynthesis
in plants and insects
In what is referred to as the ‘chemical arms race’ between
plants and herbivores, the same solution may recur fre-
quently, particularly for biosynthetic pathways involving a
relatively small number of enzymes. While cyanogenic glu-
cosides provide plants with a defence mechanism against
generalist herbivores, some specialized herbivores have
co-evolved with their host plants. The larvae of the Burnet
moth (Zygaena filipendulae) feed on Lotus corniculatus and
sequester the cyanogenic glucosides linamarin and lot-
australin from their food plants for use in their own defence
against predators (Zagrobelny et al., 2007). They are also
able to synthesize both compounds de novo when the sup-
ply from their food is insufficient. By convergent evolution,
the Burnet moth has obtained a biosynthetic pathway with
identical reaction intermediates but involving insect ver-
sions of the two cytochrome P450s, called CYP405A2 and
CYP332A3, and a UGT gene named UGT33A1 (Jensen et al.,
2011). This example from insects supports the idea of inde-
pendent recruitment of genes for cyanogenic glucoside
synthesis in plants. Further insights into the evolution of
cyanogenic defence pathways in plants can be obtained by
the identification of the biosynthetic genes in a diverse range
of species, including non-angiosperms, which seem to lack
members of the CYP79 family (Nelson and Werck-Reichhart,
Cyanogenic glucoside biosynthetic gene clusters 11
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

2011). Gene clustering, if a general feature, may aid this
identification.
EXPERIMENTAL PROCEDURES
Plant materials and growth conditions
Lotus japonicus cv. MG-20 (Legume Base, Miyazaki University, Ja-pan) and the cyd1 mutant (Takos et al., 2010) were grown on half-strength Murashige & Skoog basal salt mix containing 1% w/v su-crose in 1% w/v agar. Plants were germinated and grown at 24�Cwith a 16 h light cycle unless otherwise indicated. Nicotianabenthamiana plants were germinated from seed and grown in soilin a greenhouse at 22�C.
Chemicals
UDP-[14C]-glucose (Perkin Elmer NEN Radiochemicals, http://www.perkinelmer.com/) was dried under nitrogen gas to removeethanol and dissolved in water. The substrates acetone cyanohy-drin and 2-hydroxy-2-methylbutyronitrile were synthesized in ourlab by M.S.M. Lotaustralin was purified by preparative HPLC(Bjarnholt et al., 2008). Linamarin was purchased from AG Scientific(http://www.agscientific.com/). Amygdalin was purchased fromSigma-Aldrich (http://www.sigmaaldrich.com/).
RNA extraction and cDNA synthesis
Total RNA was prepared from 100 mg of plant tissue using anRNeasy plant mini kit (Qiagen, http://www.qiagen.com/) withon-column DNase I digestion. For cDNA synthesis, 1–2 lg of totalRNA was reverse-transcribed by SuperScript III reverse transcrip-tase (Invitrogen, http://www.invitrogen.com/) in a reaction primedusing 50 lM oligo(dT)20 or using an iScript cDNA synthesis kit (Bio-Rad, http://www.bio-rad.com/), which contains MMLV-derived re-verse transcriptase and is primed using a blend of oligo(dT) andrandom hexamer primers.
Genetic markers and genome analysis
The L. japonicus genome sequence (Sato et al., 2008) and markerinformation is available at http://www.kazusa.or.jp/lotus/index.html.Genome assembly build 2.5 was used in our analysis. Primersflanking the 32 bp insertion in LjSGA_030566 and distinguishing itfrom CYP79D3 were 5¢-GGAGAAAGATCAGAATCAATGAC-3¢ and 5¢-GATTCGCTGTGCAATGAGCTGAC -3¢. The polymorphism betweenthe PCR products was visualized on a 4% agarose gel. The S. bicolorgenome sequence (version 1.0 release) and the genome sequence ofM. esculenta (version 4.1) are both available from http://www.phy-tozome.net. Named cytochrome P450 sequences are available fromthe cytochrome P450 homepage at http://drnelson.uthsc.edu/cyto-chromeP450.html.
Preparation of constructs
The cDNA sequences of genes were amplified by PCR fromL. japonicus MG-20 cDNA prepared from RNA extracted from youngleaf tissue, and from cassava cv. TME12 cDNA. The PCR productswere cloned by Gateway recombination into the entry vector pDO-NOR207 (Invitrogen). Expression constructs were prepared by lin-earizing the pDONOR207 entry clones by restriction digest, andcloned by Gateway recombination into either the S-tag vectorpJAM1786 (similar to pJAM1784 but with reading frame C.1) forexpression in bacteria or pJAM1502 (Luo et al., 2007) for both stableand transient expression in plants. All the PCR primers containedthe attB1 and attB2 Gateway cloning sites, and the sequences aregiven in Table S1.
Complementation of the cyd1 mutant with CaMV35S:Lj-
CYP79D3
The CYP79D3 cDNA sequence under the control of the CaMV 35Spromoter/terminator elements in pJAM1502 was transferred intoAgrobacterium tumefaciens (AGL1) by electroporation. Transfor-mation of the cyd1 mutant with this construct was based on amethod using hypocotyl explants as previously described (Lombariet al., 2005).
Cyanogenesis detection and metabolite profiling
Cyanogenesis was visualized using Feigl–Anger paper (Feigl andAnger, 1966), which was prepared by wetting Whatman 3MM paper(http://www.whatman.com/) in a 5 g L)1 chloroform solution ofcopper ethylacetoacetate (Alfa Aesar, http://www.alfa.com/) and4,4¢-methylenebis(N,N-dimethylaniline) (Sigma-Aldrich). The driedpaper was stored at 4�C until use. Plant tissue was disrupted by afreeze–thaw cycle, and HCN release was visualized as describedpreviously (Takos et al., 2010).
For metabolite profiling of hydroxynitrile glucosides, plantmaterial was weighed and extracted in 300 ll of 85% v/v methanolspiked with amygdalin to 200 lM to correct for volume losses duringsample preparation. The extraction was performed by boiling in awater bath for 3 min and subsequent cooling of the sample on ice.The extracts were filtered through 45 lm Ultrafree-MC DuraporePVDF filters (Millipore, http://www.millipore.com/) and diluted 1:10in water. Extracts were stored in glass vials at 4�C prior to analysis.Analytical LC-MS was performed using an Agilent 1100 Series LC(Agilent Technologies, http://www.agilent.com) coupled to a BrukerHCT-Ultra ion trap mass spectrometer (Bruker Daltonics, http://www.bdal.com). A Zorbax SB-C18 column (Agilent, 2.1 mmid · 50 mm, 1.8 lm) was used, with chromatography conditionsas described previously (Takos et al., 2010). The mass spectrometerwas run in positive electrospray mode.
Quantitative real-time PCR
Transcript levels of genes were measured by real-time PCR using aSYBR Green I-based method on a Rotor-Gene Q cycler (Qiagen).The PCR reaction conditions were as previously described (Takoset al., 2010). For each primer pair, a single PCR product wasobtained from each cDNA as verified by gel electrophoresis andmelt curve analysis. Normalized gene expression of target geneswas obtained from the difference in cycle threshold of target genesand the reference gene, taking into account primer efficiencies cal-culated from standard curves of diluted PCR product using Q-Genesoftware (Muller et al., 2002). The L. japonicus RNA polymerase IIlarge subunit (AV777095) was used as a reference gene, and thesequences of the real-time PCR primers are given in Table S1.
Glycosyltransferase assays
Clones of the E. coli XJa autolysis strain (Zymo Research, http://www.zymoresearch.com) containing the cDNA sequences ofLjUGT85K3 and LjUGT85K2 in pJAM1786 were grown at 28�C inLuria broth containing ampicillin and 3 mM L-arabinose. Expressionwas induced after 5 h by addition of 4 mM IPTG, and incubation wascontinued overnight. Cells were harvested by centrifugation for 10min at 4�C, and briefly washed with 100 mM Tris/HCl, 2 mM EDTA,pH 7.5. Pellets were resuspended in 100 mM Tris/HCl pH 7.5, 2 mM
EDTA, 5 mM DTT, containing Complete protease inhibitor cocktail(Roche, http://www.roche.com). The resuspension was frozen inaliquots of 400 ll in liquid nitrogen, and then thawed at 37�C for5 min to lyse cells. The crude extract was clarified by centrifugation
12 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

at 20,000 · g for 20 min at 4�C, and the supernatant used for gly-cosylation assays. All steps were performed at 4�C. The amount ofrecombinant S-tagged enzyme was quantified using a FRETWorksS-Tag assay kit (Novagen, http://www.merck-chemicals.com/).
The glycosylation assay was preformed as previously described(Hansen et al., 2003) in assay mixtures of 20 ll containing 0.1 lgrecombinant S-tagged enzyme, 100 mM Tris/HCl pH 7.5, an initialconcentration of 20 mM of putative substrates (acetonecyanohydrin or 2-hydroxy-2-methylbutyronitrile) and 3.3 lM UDP-[14C]-glucose. Assay mixtures were incubated at 30�C for 60 min,and the reaction was terminated by addition of 2 ll 10% acetic acid.Glycosylation products were separated by TLC on Silica Gel 60 F254plates (Merck, http://www.merck-chemicals.com/) using a mobilephase of ethyl acetate:acetone:dichloromethane:methanol:water(40:30:12:10:8 by volume). Radiolabelled compounds were visual-ized by exposure of PhosphorImager screens (Molecular Dynamics,http://www.moleculardynamics.com/) for 4 days. Pictures weredeveloped using a Storm 860 scanner and Storm scanner controlsoftware version 5.03 (Amersham Biosciences, http://www.gelifes-ciences.com/). In addition to the assay mixtures, 5 lg of unlabelledlinamarin and lotaustralin were spotted on the same TLC plate. Afterseparation, the linamarin and lotaustralin lanes were cut from theTLC plate and the hydroxynitrile glucosides were visualized bydipping in 20% v/v H2SO4 in methanol to char the sugars.
Transient expression in leaves of N. benthamiana plants
Overnight cultures of A. tumefaciens (AGL1) containing expressionconstructs of cDNAs under the control of CaMV 35S promoter/ter-minator elements in pJAM1502 and the gene-silencing inhibitorprotein p19 (Voinnet et al., 2003) were grown. The cells were har-vested and resuspended to an OD600 of 2.0 in 10 mM MES, 10 mM
MgCl2 and 100 lM acetosyringone. After 4 h incubation at ambienttemperature, the A. tumefaciens was used to infiltrate leaves of3-week-old N. benthamiana plants. A. tumefaciens strains contain-ing expression constructs of plant cDNAs and the strain containingthe expression construct for the p19-encoding gene were co-infil-trated. After 4–5 days, leaf discs (1 cm diameter) were cut frominfiltrated leaves and extracted in 85% v/v methanol for metaboliteanalysis as described above.
ACKNOWLEDGEMENTS
This work was financially supported by the Danmark Grundforsk-ningsfond (Danish National Research Foundation) under the ‘NielsBohr Visiting Professorship’ program. B.L.M. acknowledgesfinancial support from the Villum Research Center ‘Pro-ActivePlants’ and from the Danish Council for Independent Research/Technology and Production Sciences. C.K. acknowledges a PhDstipend granted by the Faculty of Life Sciences, University ofCopenhagen, Denmark.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. LC-MS profiles of transient expression in N. benthami-ana.Figure S2. Phylogenetic tree of the CYP79 family.Table S1. PCR primer sequences.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Andersen, M.D., Busk, P.K., Svendsen, I. and Møller, B.L. (2000) Cytochromes
P-450 from cassava (Manihot esculenta Crantz) catalyzing the first steps in
the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin. J.
Biol. Chem. 275, 1966–1975.
Bak, S., Kahn, R.A., Nielsen, H.L., Møller, B.L. and Halkier, B.A. (1998a) Cloning
of three A-type cytochromes P450, CYP71E1, CYP98, and CYP99 from
Sorghum bicolor (L.) Moench by a PCR approach and identification by
expression in Escherichia coli of CYP71E1 as a multifunctional cytochrome
P450 in the biosynthesis of the cyanogenic glucoside dhurrin. Plant Mol.
Biol. 36, 393–405.
Bak, S., Nielsen, H.L. and Halkier, B.A. (1998b) The presence of CYP79
homologues in glucosinolate-producing plants shows evolutionary con-
servation of the enzymes in the conversion of amino acid to aldoxime in the
biosynthesis of cyanogenic glucosides and glucosinolates. Plant Mol. Biol.
38, 725–734.
Bak, S., Paquette, S.M., Morant, M. et al. (2006) Cyanogenic glucosides: a
case study for evolution and application of cytochromes P450. Phytochem.
Rev. 5, 309–329.
Bjarnholt, N. and Møller, B.L. (2008) Hydroxynitrile glucosides. Phytochem-
istry, 69, 1947–1961.
Bjarnholt, N., Rook, F., Motawia, M.S., Cornett, C., Jørgensen, C., Olsen, C.E.,
Jaroszewski, J.W., Bak, S. and Møller, B.L. (2008) Diversification of an
ancient theme: hydroxynitrile glucosides. Phytochemistry, 69, 1507–1516.
Cheng, D.W., Lin, H., Takahashi, Y., Walker, M.A., Civerolo, E.L. and Stenger,
D.C. (2010) Transcriptional regulation of the grape cytochrome P450
monooxygenase gene CYP736B expression in response to Xylella fastidi-
osa infection. BMC Plant Biol. 10, 135.
Chu, H.Y., Wegel, E. and Osbourn, A. (2011) From hormones to secondary
metabolism: the emergence of metabolic gene clusters in plant. Plant J. 66,
66–79.
Conn, E.E. (1980) Cyanogenic compounds. Annu. Rev. Plant Physiol. 31, 433–
451.
Cseke, L., Dudareva, N. and Pichersky, E. (1998) Structure and evolution of
linalool synthase. Mol. Biol. Evol. 15, 1491–1498.
Feigl, F. and Anger, V. (1966) Replacement of benzidine by copper ethylace-
toacetate and tetra base as spot-test reagent for hydrogen cyanide and
cyanogen. Analyst, 91, 282–284.
Field, B. and Osbourn, A.E. (2008) Metabolic diversification – independent
assembly of operon-like gene clusters in different plants. Science, 320, 543–
547.
Forslund, K., Morant, M., Jørgensen, B., Olsen, C.E., Asamizu, E., Sato, S.,
Tabata, S. and Bak, S. (2004) Biosynthesis of the nitrile glucosides rhodi-
ocyanoside A and D and the cyanogenic glucosides lotaustralin and
linamarin in Lotus japonicus. Plant Physiol. 135, 71–84.
Frey, M., Chomet, P., Glawischnig, E. et al. (1997) Analysis of a chemical plant
defense mechanism in grasses. Science, 277, 696–699.
Frey, M., Stettner, C., Pare, P.W., Schmelz, E.A., Tumlinson, J.H. and Gierl, A.
(2000) An herbivore elicitor activates the gene for indole emission in maize.
Proc. Natl Acad. Sci. USA, 97, 14801–14806.
Gierl, A. and Frey, M. (2001) Evolution of benzoxazinone biosynthesis and
indole production in maize. Planta, 213, 493–498.
Glawischnig, E., Hansen, B.G., Olsen, C.E. and Halkier, B.A. (2004) Camalexin
is synthesized from indole-3-acetaldoxime, a key branching point between
primary and secondary metabolism in Arabidopsis. Proc. Natl Acad. Sci.
USA, 101, 8245–8250.
Guttikonda, S.K., Trupti, J., Bisht, N.C., Chen, H., An, Y.-Q.C., Pandey, S., Xu,
D. and Yu, O. (2010) Whole genome co-expression analysis of soybean
cytochrome P450 genes identifies nodulation-specific P450 monooxygen-
ases. BMC Plant Biol. 10, 243.
Handberg, K. and Stougaard, J. (1992) Lotus japonicus, an autogamous,
diploid legume species for classical and molecular genetics. Plant J. 2, 487–
496.
Hansen, K.S., Kristensen, C., Tattersall, D.B., Jones, P.R., Olsen, C.E., Bak, S.
and Møller, B.L. (2003) The in vitro substrate regiospecificity of re-
combinant UGT85B1, the cyanohydrin glucosyltransferase from Sorghum
bicolor. Phytochemistry, 64, 143–151.
Hasemann, C.A., Kurumbail, R.G., Boddupalli, S.S., Peterson, J.A. and Deis-
enhofer, J. (1995) Structure and function of cytochromes P450: a compar-
ative analysis of three crystal structures. Structure, 2, 41–62.
Cyanogenic glucoside biosynthetic gene clusters 13
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x

Hull, A.K., Vij, R. and Celenza, J.L. (2000) Arabidopsis cytochrome P450s that
catalyze the first step of tryptophan-dependent indole-3-acetic acid bio-
synthesis. Proc. Natl Acad. Sci. USA, 97, 2379–2384.
Jensen, N.B., Zagrobelny, M., Hjernø, K., Olsen, C.E., Houghton-Larsen, J.,
Borch, J., Møller, B.L. and Bak, S. (2011) Convergent evolution in biosyn-
thesis of cyanogenic defence compounds in plants and insects. Nat.
Commun. 2, 273.
Jones, D.A. (1998) Why are so many food plants cyanogenic? Phytochemistry,
47, 155–162.
Jones, P.R., Møller, B.L. and Høj, P.B. (1999) The UDP-glucose: p-hydroxy-
mandelonitrile-O-glucosyltransferase that catalyzes the last step in
synthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor. J. Biol.
Chem. 274, 35483–35491.
Jørgensen, K., Morant, A.V., Morant, M., Jensen, N.B., Olsen, C.E., Kann-
angara, R., Motawia, M.S., Møller, B.L. and Bak, S. (2011) Biosynthesis of
the cyanogenic glucosides linamarin and lotaustralin in cassava (Manihot
esculenta Crantz): isolation, biochemical characterization and expression
pattern of CYP71E7, the oxime-metabolizing cytochrome P450 enzyme.
Plant Physiol. 155, 282–292.
Kannangara, R., Motawia, M.S., Hansen, N.K.K., Paquette, S.M., Olsen, C.E.,
Møller, B.L. and Jørgensen, K. (2011) Characterization and expression
profile of two UDP-glucosyltransferases, UGT85K4 and UGT85K5, catalyz-
ing the last step in cyanogenic glucoside biosynthesis in cassava. Plant J.
doi: 10.1111/j.1365-313X.2011.04695.x.
Khersonsky, O. and Tawfik, D.S. (2010) Enzyme promiscuity: a mechanistic
and evolutionary perspective. Annu. Rev. Biochem. 79, 471–505.
Koch, B.M., Sibbesen, O., Halkier, B.A., Svendsen, I. and Møller, B.L. (1995)
The primary sequence of cytochrome P450tyr, the multifunctional
N-hydroxylase catalyzing the conversion of L-tyrosine to p-hydroxyphe-
nylacetaldehyde oxime in the biosynthesis of the cyanogenic glucoside
dhurrin in Sorghum bicolor (L.) Moench. Arch. Biochem. Biophys. 323, 177–
186.
Kristensen, C., Morant, M., Olsen, C.E., Ekstrøm, C.T., Galbraith, D.W.,
Møller, B.L. and Bak, S. (2005) Metabolic engineering of dhurrin in
transgenic Arabidopsis plants with marginal inadvertent effects on the
metabolome and transcriptome. Proc. Natl Acad. Sci. USA, 102, 1179–
1784.
Lombari, P., Ercolano, E., El Alaoui, H. and Chiurazzi, M. (2005) Agrobacte-
rium-mediated in vitro transformation. In Lotus japonicus Handbook
(Marquez, A.J., ed.). Dordrecht, The Netherlands: Springer, pp. 251–259.
Luo, J., Nishiyama, Y., Fuell, C. et al. (2007) Convergent evolution in the BAHD
family of acyl transferases: identification and characterization of anthocy-
anin acyl transferases from Arabidopsis thaliana. Plant J. 50, 678–695.
Mackenzie, P.I., Owens, I.S., Burchell, B. et al. (1997) The UDP glycosyl-
transferase gene superfamily: recommended nomenclature update based
on evolutionary divergence. Pharmacogenetics, 7, 255–269.
Martin, F.J. and McInerney, J.O. (2009) Recurring cluster and operon
assembly for phenylacetate degradation genes. BMC Evol. Biol. 9, 36.
Morant, A.V., Jørgensen, K., Jørgensen, C., Paquette, S.M., Sanchez-Perez,
R., Møller, B.L. and Bak, S. (2008) b-Glucosidases as detonators of plant
chemical defense. Phytochemistry, 69, 1795–1813.
Muller, P.Y., Janovjak, H., Miserez, A.R. and Dobbie, Z. (2002) Processing of
gene expression data generated by quantitative real-time RT-PCR. Bio-
Techniques, 32, 2–7.
Nelson, D. and Werck-Reichhart, D. (2011) A P450-centric view of plant evo-
lution. Plant J. 66, 194–211.
Olsen, K.M., Hsu, S.-C. and Small, L.L. (2008) Evidence on the molecular basis
of the Ac/ac adaptive cyanogenesis polymorphism in white clover (Trifo-
lium repens L.). Genetics, 179, 517–526.
Paterson, A.H., Bowers, J.E., Bruggmann, R. et al. (2009) The Sorghum
bicolor genome and the diversification of grasses. Nature, 457, 551–556.
Pichersky, E. and Lewinsohn, E. (2011) Convergent evolution in plant spe-
cialized metabolism. Annu. Rev. Plant Biol. 62, 549–566.
Qi, X., Bakht, S., Leggett, M., Maxwell, C., Melton, R. and Osbourn, A. (2004)
A gene cluster for secondary metabolism in oat: implications for the evo-
lution of metabolic diversity in plants. Proc. Natl Acad. Sci. USA, 101, 8233–
8238.
Reimann, A., Nurhayati, N., Backenkohler, A. and Ober, D. (2004) Repeated
evolution of the pyrrolizidine alkaloid-mediated defense system in separate
angiosperm lineages. Plant Cell, 16, 2772–2784.
Reintanz, B., Lehnen, M., Reichelt, M., Gershenzon, J., Kowalczyk, M.,
Sandberg, G., Godde, M., Uhl, R. and Palme, K. (2001) Bus, a bushy Ara-
bidopsis CYP79F1 knockout mutant with abolished synthesis of short-chain
aliphatic glucosinolates. Plant Cell, 13, 351–367.
Rodman, J.E., Soltis, P.S., Soltis, D.E., Sytsma, K.J. and Karol, K.G. (1998)
Parallel evolution of glucosinolate biosynthesis inferred from congruent
nuclear and plastid gene phylogenies. Am. J. Bot. 85, 997–1006.
Rupasinghe, S., Schuler, M.A., Kagawa, N., Yuan, H., Lei, L., Zhao, B.,
Kelly, S.L., Waterman, M.R. and Lamb, D.C. (2006) The cytochrome P450
gene family CYP157 does not contain EXXR in the K-helix reducing the
absolute conserved P450 residues to a single cysteine. FEBS Lett. 580,
6338–6342.
Sato, S., Nakamura, Y., Kaneko, T. et al. (2008) Genome structure of the
legume, Lotus japonicus. DNA Res. 15, 227–239.
Schwab, W. (2003) Metabolome diversity: too few genes, too many metabo-
lites? Phytochemistry, 62, 837–849.
Seigler, D.S. (1975) Isolation and characterization of naturally occurring cya-
nogenic compounds. Phytochemistry, 14, 9–29.
Shimura, K., Okada, A., Okada, K. et al. (2007) Identification of a biosynthetic
gene cluster in rice for momilactones. J. Biol. Chem. 282, 34013–34018.
Swaminathan, S., Morrone, D., Wang, Q., Fulton, D.B. and Peters, R.J. (2009)
CYP76M7 is an ent-cassadiene C11a-hydroxylase defining a second mul-
tifunctional diterpenoid biosynthetic gene cluster in rice. Plant Cell, 21,
3315–3325.
Takos, A., Lai, D., Mikkelsen, L., Hachem, M.A., Shelton, D., Motawia, M.S.,
Olsen, C.E., Wang, T.L., Martin, C. and Rook, F. (2010) Genetic screening
identifies cyanogenesis-deficient mutants of Lotus japonicus and reveals
enzymatic specificity in hydroxynitrile glucoside metabolism. Plant Cell, 22,
1605–1619.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. and Kumar, S.
(2011) MEGA5: molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods. Mol.
Biol. Evol. doi: 10.1093/molbev/msr121.
Voinnet, O., Rivas, S., Mestre, P. and Baulcombe, D. (2003) An enhanced
transient expression system in plants based on suppression of gene
silencing by the p19 protein of tomato bushy stunt virus. Plant J. 33, 949–
956.
Zagrobelny, M., Bak, S., Ekstrøm, C.T., Olsen, C.E. and Møller, B.L. (2007) The
cyanogenic glucoside composition of Zygaena filipendulae (Lepidoptera:
Zygaenidae) as effected by feeding on wild-type and transgenic lotus
populations with variable cyanogenic glucoside profiles. Insect Biochem.
Mol. Biol. 37, 10–18.
14 Adam M. Takos et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), doi: 10.1111/j.1365-313X.2011.04685.x
Top Related
Copyright © 2022 FDOKUMEN

