







Bahasa
Halaman
Hukum


UNIVERSIDAD DE GUAYAQUIL
FACULTAD DE CIENCIAS NATURALES
ESCUELA DE BIOLOGÍA
TESIS PREVIA A LA OBTENCIÓN DEL TÍTULO DE BIÓLOGO
“ESPECTRO TRÓFICO DE Alopias pelagicus NAKAMURA 1935
(CHONDRICHTHYES: ALOPIIDAE) EN SANTA ROSA DE SALINAS,
GUAYAS, DURANTE MAYO - DICIEMBRE DEL 2004”
MARCOS DOUGLAS CALLE MORÁN
GUAYAQUIL – ECUADOR
2006

UNIVERSIDAD DE GUAYAQUIL
FACULTAD DE CIENCIAS NATURALES
ESCUELA DE BIOLOGÍA
TESIS PREVIA A LA OBTENCIÓN DEL TÍTULO DE BIÓLOGO
“ESPECTRO TRÓFICO DE Alopias pelagicus NAKAMURA 1935
(CHONDRICHTHYES: ALOPIIDAE) EN SANTA ROSA DE SALINAS,
GUAYAS, DURANTE MAYO - DICIEMBRE DEL 2004”
MARCOS DOUGLAS CALLE MORÁN
GUAYAQUIL – ECUADOR
2006

© Derechos del Autor Marcos D. Calle Morán
2006

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
iii
DIRECTOR DE TESIS
______________________________
Blgo. Iván E. Zambrano Alcívar

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
iv
CO - DIRECTOR DE TESIS
______________________________
Dr. Felipe Galván Magaña CICIMAR- IPN, La Paz, Baja California Sur, MÉXICO
Doctor en Ecología Marina Profesor e Investigador Principal
ASESOR CIENTÍFICO DE TESIS
______________________________
Dr. Philippe Béarez Museo Nacional de Historia Natural, Paris, FRANCIA
Doctor en Ictiología General y Aplicada Investigador

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
v
UNIVERSIDAD DE GUAYAQUIL
FACULTAD DE CIENCIAS NATURALES
ESCUELA DE BIOLOGÍA
HOJA DE APROBACIÓN DE TESIS
“ESPECTRO TRÓFICO DE Alopias pelagicus, NAKAMURA 1935 (CHONDRICHTHYES: ALOPIIDAE) EN SANTA ROSA DE SALINAS, GUAYAS, DURANTE MAYO- DICIEMBRE DEL 2004”
Presidente del Tribunal ______________________________
Primer Miembro del Tribunal ______________________________
Segundo Miembro del Tribunal ______________________________
Secretario de la Facultad ______________________________

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
vi
Dedicatoria íntima:
A Jesús, My Lord, centro de mi existencia; fuente perenne de la vida eterna;
puerto, camino y morada; tú Señor eres mi herencia, mi tesoro y mi esperanza,
eres la luz que me iluminas, sol de mi vida y mi corazón…
A María Santísima, mi Madre del Cielo , toda mi esperanza eres tú, eres tú,
como lluvia fresca entre mis manos, como fuerte risa eres tú, eres tú, así, así,
eres tú…
A Estania Elizabeth Ormaza Rosado
Al Dr. Roberto D. Jiménez Santistevan

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
vii
In memoriam:
A Marlon Alberto Chávez (+) y a Darío Pínela G. (+). Amigos desde la infancia, que ya están en el Cielo y a quienes de manera especial dedico esta tesis… lo mejor, siempre para y por ustedes…

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
viii
AGRADECIMIENTOS
A la Verdadera, Excelsa y Eterna Trinidad, Padre, Hijo y Espíritu Santo, por permitirme hacer realidad este sueño de toda la vida… A mi Patrem Omnipotenten, Dios Padre por el don de la Vida. Gracias por haberme llamado a la existencia… A mi Jesucristum Hominis Salvatore, Dios Hijo, por la Redención. Gracias por todo lo has hecho por nosotros… A mi Spiritum Paraclitum , Dios Espíritu Santo, por la Santificación. Gracias por la santificación… A mi Regina Caelis, la Virgen María, por ser mi Madre, Maestra y Amiga. Gracias por tanto amor, cariño y amparo…gracias por haberme acompañado a lo largo de toda mi vida y porque nunca me has fallado… Al Santo Cura de Ars, San Juan María Vianney, por ser Nuestro Patrono. Gracias por ser un ejemplo de vida… gracias por luchar contra todo para darnos todo… A Santo Tomás de Aquino, mi Maestro, por darme ejemplo, a quien admiro por tanta sabiduría… A la Doncella de Orleáns, Santa Juana de Arco, por tanta valentía, dedicación y amor demostrado, gracias por tanta fidelidad… A mi Evangelista preferido, San Marcos. Gracias por prestarme su nombre… por permitirnos conocer la vida de Jesús, a través de sus escritos… A los sacerdotes: Rvdo. P. Nelson Cevallos (+), Rvdo. P. Oscar Uribe, Rvdo. P. Miguel Camats, Rvdo. P. Mariano Correa, Rvdo. P. Agustín Mezcua, Rvdo. P. Eduardo Brito, Rvdo. P. Marcos Pérez y a Su Excma. Mons. Juan Ignacio Larrea Holguín, por haberme acogido en el Seminario Menor “César Antonio Mosquera Corral”, en donde aprendí que la vida es diferente a lo que el mundo piensa, es un regalo de Dios y de la cual se puede obtener todo lo bueno que en ella hay; por tantas enseñanzas, por la Fe, por el amor, por cuidarme siempre, por todo… la mejor época de mi existencia está allí… Al Rvdo. P. Byrone Tomalá, por haberme apoyado en los dos últimos años de mi carrera universitaria, por todo el ejemplo impartido, por su amistad, gracias mil… A mi madre, María Morán Sánchez , por ser padre y madre a la vez. Gracias por todos los cuidados, cariño y comprensión…

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
ix
A mis queridísimos hermanos: Beatriz, Xavier, Jessica, Orlando, Patricia, Rocío y Luis, gracias por todos aquellos momentos juntos… A Estania Ormaza, por la amistad y el cariño brindado, por todos los consejos dados, por ser mi Evangelium vitae, gracias por todo… A mis mejores amigos de toda la vida, Miguel Alvear, William Morocho y Vicente Villacís . Gracias por aquellos momentos inolvidables que pasamos en el Seminario Menor, ustedes saben que siempre los llevo conmigo. Ustedes me han enseñado que la amistad sobrepasa la muerte y que la vida es sólo es un soplo en la eternidad, no los considero amigos sino mis hermanos. A Carlos Saldaña, Maurilio Ortega, Yurman Àviles, Antonio Navas y a todos los seminaristas, por haber compartido la mejor parte de sus vidas conmigo… A Sandra Armijos, Walter Guillén, Geovanny Zambrano, Paola Rivadeneira y Fidel Egas mis mejores amigos de la universidad y a todos mis compañeros. Me equivoqué pensaba, que no iba a ser amistades tan profundas como las vuestras pero la realidad afortunadamente es otra. Fueron un ejemplo de vida, sin vuestro apoyo creo que no hubiera terminado la carrera, se los confieso… En especial a Walter por la colaboración en las fotografías de los organismos presas. A Lida Ramírez, Lorena Sánchez y Verónica Plaza mis mejores amigas por escucharme siempre, por haber compartido tantas cosas juntos, vuestra amistad vale más que cualquier tesoro… A Kelly Arteaga por las sugerencias y de manera especial por la colaboración en la edición de las fotografías… A Claudia Castro por la amistad y en forma exclusiva por la preparación de las diapositivas para la sustentación de mi tesis… A Adreanna Pankow por la corrección del resumen en inglés… A Ruth Muñiz, Vivian Valenzuela, Sonia Graham, Christian Naranjo e Irina Muñoz, por la amistad compartida, por todas las enseñanzas tanto en lo personal como en lo profesional, por todo lo vivido, por el inmenso cariño, aprendí casi todo de ustedes. Entendí que la vida es un mar de aprendizaje. Gracias por aquel empujón final que me ayudó a culminar este trabajo. Al Dr. Felipe Galván Magaña, del Centro Interdisciplinario de Ciencias Marinas (CICIMAR) e Instituto Politécnico Nacional (IPN), Baja California Sur, México. Por haberme iniciado en ésta disciplina, por el apoyo impartido desde el comienzo hasta el final de este hermoso sueño llamado TESIS. Un millón de gracias…

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
x
Al Dr. Philippe Béarez , del Museo Nacional de Historia Natural (MNHN), Paris, Francia. Por haber confiado en mí, por la ayuda, por las críticas a este trabajo y por todo… Al Blgo. Fernando Aguilar, del Instituto Nacional de Pesca, un especial agradecimiento por haberme facilitado la mandíbula y fotos del tiburón zorro común de México, así mismo por bibliografía, comunicaciones personales y por el apoyo brindado para la culminación de este trabajo. Gracias por la revisión del borrador del mismo… A la Blga. Mrna. Patricia Zarate de la Fundación Científica Charles Darwin de Galápagos, por las comunicaciones personales dadas para la presente investigación… A mi científico predilecto, el Dr. Roberto Jiménez, mi Padre, Maestro y Amigo. Gracias porque lo admire desde siempre y me ha dado la oportunidad de formarme bajo su tutela… que suerte la mía… gracias por todo Don Roberto… un Dios le pague… por confiar y apoyar siempre a la juventud investigadora… A mi querida Facultad de Ciencias Naturales, a mis apreciados profesores, a todos, gracias por la entrega, por el entusiasmo, por el ejemplo, por el profesionalismo, por el carisma y por la gran paciencia que me tuvieron siempre. En especial a la Blga. Ruth Chóez , Dr. Luis Muñíz, Blga. Mireya Cadena, Blga. Betty Salvatierra, Dra. María Luzuriaga y a la Dra. Matilde Cornejo. Sólo Dios sabrá recompensar vuestro trabajo… A Héctor Erazo y Nataly Bolaño, amigos y compañeros de trabajo, gracias por la amistad, el profesionalismo y toda la ayuda… En especial a Nataly por la colaboración en las fotografías de Santa Rosa de Salinas... Al Blgo. Iván Zambrano, Director de esta tesis, gracias por todos los consejos, sugerencias y correcciones brindadas para la realización del presente trabajo. Gracias por haberme animado a lo largo de esta carrera y por todo el apoyo brindado… un inmenso Dios le pague Don Iván…

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xi
RESUMEN
Se presenta el espectro trófico del Tiburón Zorro Pelágico Alopias pelagicus
Nakamura 1935. Un total de 110 ejemplares fueron analizados, provenientes
de la pesca artesanal en el puerto pesquero de Santa Rosa de Salinas,
Guayas. Todos los especimenes fueron muestreados de mayo a diciembre del
2004, los fines de semana.
El objetivo del estudio radica en la determinación del espectro trófico de
Alopias pelagicus mediante la identificación de las presas consumidas y la
relación existente entre la dieta y el sexo, la madurez sexual y las tallas de los
individuos.
Se tomaron los principales datos morfométricos de los especimenes y seguido
se colectaron los estómagos, cuyos contenidos se analizaron identificando las
especies consumidas hasta el mínimo taxón posible, para luego relacionarlas
con el sexo, la madurez sexual y las tallas. Se establecieron medidas
principales para los estados de madurez. Se obtuvieron las presas principales
mediante el Índice de Importancia Relativa (IIR), la diversidad de presas por
medio del Índice Shannon y Weaver (H’) y la uniformidad de las mismas con el
Índice de Pielou (E), la amplitud del nicho trófico de la especie mediante el
Índice de Levin (Bi) y las posibles diferencias tróficas mediante el Índice de
Morisita y Horn (Cλ).
De los 110 ejemplares, 74 eran hembras y 36 machos. Se presentaron 45
hembras sexualmente maduras de 258 – 365 cm LT (incluyendo 7 grávidas) y
29 inmaduras de 152 – 257 cm LT. Así mismo, 20 machos maduros de 244 -
310 cm. LT y 16 inmaduros de 176 - 243 cm LT. La variación mensual de los
individuos desembarcados, determinó que durante mayo y diciembre se
registraron el mayor número de dicha especie, así como también el mayor
número de hembras grávidas; mientras que de junio a julio no hubo ejemplares.
El 73% (80 estómagos) contenían alimentos, esto es, que presentaban por lo
menos un componente alimenticio y el 27% (30) vacíos, literalmente. La mitad
de los estómagos analizados (50.90%) estaban vacíos, en cuanto a proporción

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xii
de llenado (0= 0%) y de igual forma el 50% de las presas presentaban un
avanzado Estado de Digestión (ED) 4 y el 40% un ED 3.
El espectro trófico de Alopias pelagicus estuvo compuesto por 24
componentes alimenticios, siendo las presas principales de acuerdo al Índice
de Importancia Relativa (IIR): Sthenoteuthis oualaniensis (IIR= 2 071,91),
Merluccius gayi (IIR=1 569,02) y Benthosema panamense (IIR= 1 086,97).
La alimentación por sexos, estados de madurez sexual y tallas estuvo
dominada por M. gayi, S. oualaniensis, B. panamense, Dosidicus gigas y
Ommastrephes bartramii.
La diversidad de presas consumidas, según el Índice de Shannon y Weaver
(H’), fue bien baja para todas las categorías debido a la preferencia por tres
presas principales (S. oualaniensis, M. gayi y B. panamense). En forma
general el índice global fue de H’= 1,61 y E= 0,51; para los sexos, la mayor fue
para los machos (H’= 1,68 y E= 0,60) sobre las hembras (H’= 1,47 y E= 0,52).
Para los estados de madurez sexual y tallas, la mayor perteneció a las
hembras sexualmente inmaduras de 152 - 257 cm LT (H’= 1,98 y E= 0,79)
sobre las hembras sexualmente maduras de 258 - 365 cm LT (H’= 1,09 y
E= 0,49). Mientras que los machos sexualmente maduros de 244 - 360 cm LT
presentaron mayor diversidad (H’= 1,72 y E= 0,69) que los machos inmaduros
de 176 - 243 cm LT (H’= 1,26 y E= 0,53). A las hembras inmaduras
correspondieron la mayor diversidad y para las hembras maduras la menor
entre todas las siete categorías.
La amplitud del nicho trófico, según el Índice de Levin (Bi), para todas las
categorías fue bien baja, siendo así, que el índice general fue de Bi= 0,11 lo
que confirma una dieta dominada por tres presas de un total de 24
componentes en la alimentación de A. pelagicus, por lo que se trata de un
depredador especialista. Para los sexos, el mayor fue para los machos
(Bi= 0,17) sobre las hembras (Bi= 0,12). Para los estados de madurez sexual,
el mayor perteneció a las hembras inmaduras (Bi= 0,31) sobre las maduras
(Bi= 0,14). Los machos maduros obtuvieron el mayor valor (Bi= 0,32) que los

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xiii
inmaduros (Bi= 0,12). A los machos maduros correspondieron el mayor valor
del índice y para el general el menor valor.
La superposición de dieta, según el Índice Morisita – Horn (Cλ), demostró
valores aceptables para las tres categorías confrontadas, lo cual confirmó el
traslapamiento trófico (similaridad de componentes en sus dietas). El mayor
valor fue para los machos y hembras (Cλ= 0,79) lo que indica un
traslapamiento mayor, esto es, alta similitud en los elementos de la dieta de
A. pelagicus para ambos sexos; seguido de los machos maduros e inmaduros
(Cλ= 0,68), esto es, alta similitud. Finalmente, el menor valor fue para las
hembras maduras e inmaduras (Cλ= 0,42) lo que indica un traslapamiento
medio, es decir, similitud media en los elementos alimenticios.
La variación mensual de la dieta de A. pelagicus estableció que durante mayo
se alimentó de S. oualaniensis , D. gigas, O. bartramii y
Loliolopsis diomedeae y de octubre a diciembre lo hizo de M. gayi y
B. panamense.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xiv
ABSTRACT
The trophic spectre of Pelagic Thresher Shark Alopias pelagicus Nakamura
1935 is presented. A total of 110 exemplaries were analyzed coming from
artisan fishing in the Santa Rosa de Salinas fishing port, Guayas. The
specimens were sampled from May to December 2004, all weekends.
The objective of this study is based on the determination of the trophic spectre
of Alopias pelagicus through the identification of eaten preys and the
relationship between diet and sex, sexual maturity, and their lengths.
Main morphometric data of specimens were taken, next the stomachs were
collected. Then, the stomach contents were analyzed identifying the prey
species until the minimum taxon possible and they were related with the sex,
sexual maturity, and their lengths. Main preys were obtained by means of
Relative Importance Index (RII), the diversity through the Shannon and Weaver
Index (H’) and the uniformity of the prey species with the Pielou Index (E), the
breadth of trophic niche of the species with the help of Levin Index (Bi), and the
possible differences with the Morisita - Horn Index (Cλ), too.
From 110 exemplaries, 74 were females and 36 males. There were 45 sexually
mature females of 258 – 365 cm TL (including 7 pregnant ones) and 29
inmature ones of 152 – 257 cm TL. There were also, 20 sexually mature males
of 244 – 310 cm. TL and 16 inmature ones of 176 – 243 cm TL. The monthly
variation of landed individuals determined that during May and December the
highest number of this species and the highest number of pregnant females
was recorded. While, from June to July there were not any exemplary. 73% (80
stomachs) contained foods, that is, they had at least one feeding component
and 27% (30) were literally empty. Half of the studied stomachs (50.90%) were
empty, in relationship to fullness ratio (0= 0%) and 50% of preys had a digestive
state (DS) 4 and 40% in a DS 3.
The trophic spectre of Alopias pelagicus was composed by 24 feeding
components, being the main preys, according to Relative Importance Index

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xv
(RII): Sthenoteuthis oualaniensis (RII= 2 071,91), Merluccius gayi
(RII= 1 569,02) and Benthosema panamense (RII= 1 086,97). The feeding by
sexes, sexual maturity, and lengths was dominated by M. gayi,
S. oualaniensis, B. panamense, Dosidicus gigas and
Ommastrephes bartramii.
The diversity of eaten preys, in agreement with Shannon and Weaver Index
(H’), was very low for all categories due to preference by tree main preys
(S. oualaniensis, M. gayi & B. panamense). In general, the global Index was
H’= 1,61 & E= 0,51; for sexes, the highest value was for males (H’= 1,68 &
E= 0,60) over females (H’= 1,47 y E= 0,52). For sexual maturity and their
lengths, the highest value belonged to sexually inmature females 152 - 257 cm
TL (H’= 1,98 & E= 0,79) over the mature ones of 258 - 365 cm TL (H’= 1,09 &
E= 0,49). While sexually matures males of 244 - 360 cm TL presented a higher
diversity (H’= 1,72 & E= 0,69) than inmatures ones of 176 - 243 cm TL
(H’= 1,26 & E= 0,53). Inmatures females had highest diversity and mature
females had the lowest diversity of all seven categories.
Also, the breadth of trophic niche, according to Levin Index (Bi) for all
categories was very low. The general Index was Bi= 0,11 which confirms a diet
dominated by three of the 24 elements in the feeding of A. pelagicus, for this
reason this species is a specialist predator. For sexes, the highest value was
found in males (Bi= 0,17) over females (Bi= 0,12). For sexual maturity and their
lengths, the highest value belonged to sexually inmature females (Bi= 0,31)
over mature ones (Bi= 0,14). Mature males obtained a higher value (Bi= 0,32)
than inmature ones (Bi= 0,12). Inmature males had the highest value and the
general category had the lowest value overall between all seven categories.
The superposition of diet, in agreement with Morisita – Horn Index (Cλ) showed
acceptable values for three confronted categories, which confirmed a trophic
overlaping (similarity of components in their diets). The highest value belonged
for males and females (Cλ= 0,79), which indicates a high overlaping, that is,
high similarity in the dietary elements of A. pelagicus for both sexes; next of
mature and inmature males (Cλ= 0,68), that means a high similarity, too.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xvi
Finally, the lowest value was for mature and inmature females (Cλ= 0,42), that
indicates a half overlaping, that is, medium similarity in the feeding elements.
The monthly variation in the diet of A. pelagicus diet determined during May
included S. oualaniensis, D. gigas, O. bartramii y Loliolopsis diomedeae
and from October to December included M. gayi and B. panamense.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xvii
LISTA DE TABLAS, FIGURAS Y ANEXOS
LISTA DE TABLAS
Tablas Págs.
Tabla 1. Espectro trófico de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004 expresado en valores absolutos y porcentuales del método numérico (N y
%N), frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de
importancia relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa
Principal, S: Secundaria y C: Circunstancial........................................................46
Tabla 2. Espectro trófico de A. pelagicus hembras en Santa Rosa de Salinas durante mayo –
diciembre del 2004 en Santa Rosa de Salinas expresado en valores absolutos y
porcentuales del método numérico (N y %N), frecuencia de ocurrencia (FO y %FO),
gravimétrico (P y %PO), índice de importancia relativa (IIR) y Categoría de la
especie presa (Categ), donde P: Presa Principal, S: Secundaria y C: Circunstancial.
........................................................................................................................49
Tabla 3. Espectro trófico de A. pelagicus machos en Santa Rosa de Salinas durante mayo –
diciembre del 2004 expresado en valores absolutos y porcentuales del método
numérico (N y %N), frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO),
índice de importancia relativa (IIR) y Categoría de la especie presa (Categ), donde P:
Presa Principal, S: Secundaria y C: Circunstancial. .............................................52
Tabla 4. Espectro trófico de hembras sexualmente maduras de 258 - 365 cm. LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal, S:
Secundaria y C: Circunstancial...........................................................................55
Tabla 5. Espectro trófico de hembras sexualmente inmaduras de 152 - 257 cm LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal, S:
Secundaria y C: Circunstancial. .........................................................................58
Tabla 6. Espectro trófico de machos sexualmente maduros de 244 - 310 cm. LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xviii
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal, S:
Secundaria y C: Circunstancial. .........................................................................61
Tabla 7. Espectro trófico de machos sexualmente inmaduros de 176 - 243 cm LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal, S:
Secundaria y C: Circunstancial. .........................................................................64
Tabla 8. Valores de Diversidad y Equitabilidad para las diferentes categorías de A. pelagicus,
en Santa Rosa de Salinas durante mayo – diciembre del 2004. ............................65
Tabla 9. Valores del Índice de Levin para las diferentes categorías de A. pelagicus, en Santa
Rosa de Salinas durante mayo – diciembre del 2004. .........................................65
Tabla 10. Valores de Superposición de dietas para las categorías confrontadas del tiburón
zorro pelágico A. pelagicus, en Santa Rosa de Salinas durante mayo – diciembre
del 2004. ...........................................................................................................66

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xix
LISTA DE FIGURAS
Figuras Págs.
Fig. 1. Morfología externa del tiburón zorro (Compagno, Krupp y Schneider, 1995) ............. 13
Fig. 2. Tiburón zorro pelágico Alopias pelagicus hembra de 305 cm (LT) en el puerto de Santa
Rosa de Salinas (Marzo, 2005) ...............................................................................15
Fig. 3. Esquema de ciertas características morfológicas de A. pelagicus (Compagno et al.,
1995). ....................................................................................................................16
Fig. 4. Mandíbula de A. pelagicus mostrada en forma panorámica (Noviembre, 2005).
Detalles ampliados de zonas marcadas con flechas anaranjadas en figuras 5 y 6……16
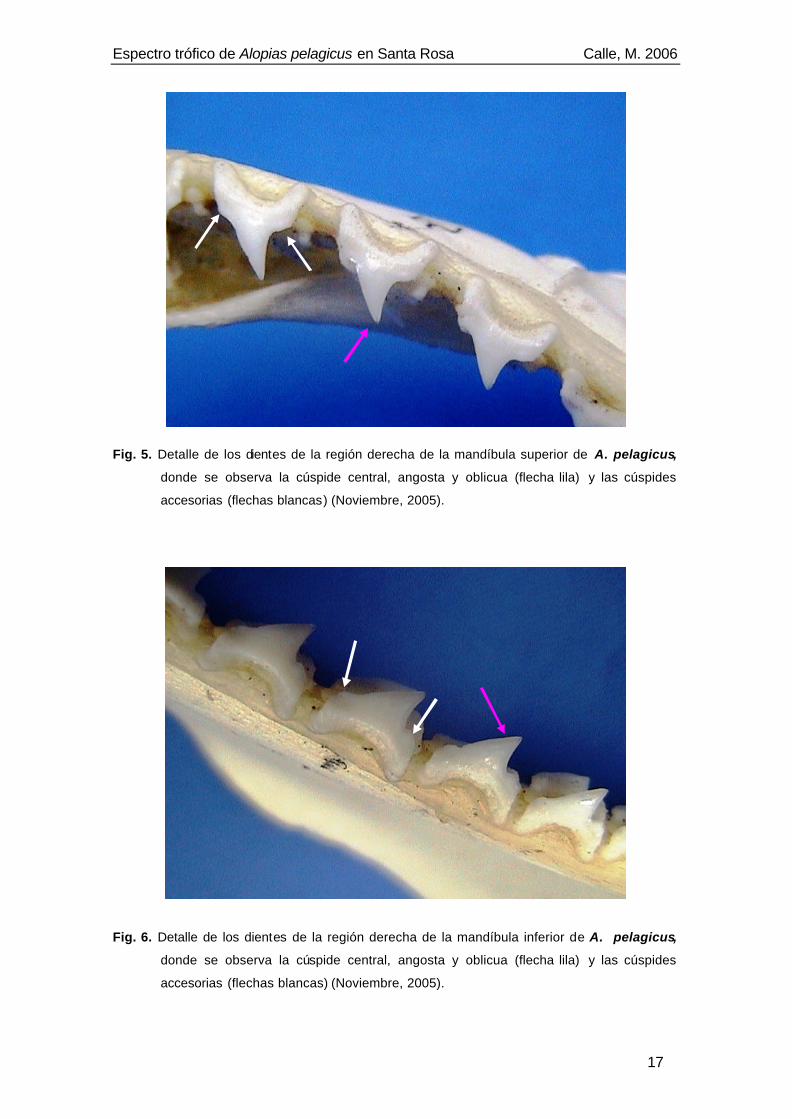
Fig. 5. Detalle de los dientes de la región derecha de la mandíbula superior de A. pelagicus,
donde se observa la cúspide central, angosta y oblicua (flecha lila) y las cúspides
accesorias (flechas blancas) (Noviembre, 2005). .....................................................17
Fig. 6. Detalle de los dientes de la región derecha de la mandíbula inferior de A. pelagicus,
donde se observa la cúspide central, angosta y oblicua (flecha lila) y las cúspides
accesorias (flechas blancas) (Noviembre, 2005) .......................................................17
Fig. 7. Vista lateral de A. pelagicus donde se aprecia la disposición de las aletas y
parcialmente la longitud de la enorme cola (Marzo, 2005). .....................................18
Fig. 8. Dos crías de A. pelagicus sobre el cuerpo de una hembra adulta, en Santa Rosa de
Salinas. Arriba: Macho de 55.5 cm (LT) y abajo: Hembra 57.2 cm (LT). (Marzo, 2005)
.............................................................................................................................19
Fig. 9. Distribución de A. pelagicus para el Pacífico Oriental................................................19 Fig. 10. Ubicación Geográfica del Puerto de Santa Rosa, en la provincia del Guayas (Solís,
1998). .................................................................................................................26
Fig. 11. Principales términos empleados en el muestreo. (Aleta escapular = Aleta pectoral)
(Montero, 2000). ..................................................................................................28
Fig. 12. Número de Individuos por sexo de A. pelagicus en Santa Rosa de Salinas durante
mayo – diciembre del 2004, (a) en número y (b) en porcentajes. ...............................36

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xx
Fig. 13. Variación mensual de individuos de A. pelagicus en Santa Rosa de Salinas durante
mayo – diciembre del 2004, (a) en número y (b) en porcentajes. ...............................37
Fig. 14. Curva Acumulativa de Especies presas de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004 .........................................................................38
Fig. 15. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 .............................................................38
Fig. 16. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para hembras y (b) para machos. .....39
Fig. 17. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para hembras sexualmente maduras y
(b) para inmaduras .................................................................................................39
Fig. 18. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para machos sexualmente maduros y
(b) para inmaduros .................................................................................................40
Fig. 19. Porcentajes de llenado estomacales de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004 (a) para organismos en general, (b) organismos en
general por meses (c) hembras por meses y (d) machos por meses..........................41
Fig. 20. Valores porcentuales del Método Numérico de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004. ........................................................................44
Fig. 21. Valores porcentuales del Método de Frecuencia de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004. ............................................................44
Fig. 22. Valores porcentuales del Método Gravimétrico de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004. ............................................................45
Fig. 23. Valores del Índice de Importancia Relativa (IIR) de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004. ............................................................45
Fig. 24. Valores porcentuales del Método Numérico para A. pelagicus hembras en Santa Rosa
de Salinas durante mayo – diciembre del 2004. ....................................................47
Fig. 25. Valores porcentuales del Método de Frecuencia para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ..............................................47

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxi
Fig. 26. Valores porcentuales del Método Gravimétrico para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ............................................48
Fig. 27. Valores del Índice de Importancia Relativa (IIR) para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ............................................48
Fig. 28. Valores porcentuales del Método Numérico para A. pelagicus machos en Santa Rosa
de Salinas durante mayo – diciembre del 2004. .......................................................50
Fig. 29. Valores porcentuales del Método de Frecuencia para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ...............................................50
Fig. 30. Valores porcentuales del Método Gravimétrico para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ............................................51
Fig. 31. Valores del Índice de Importancia Relativa (IIR) para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004. ............................................51
Fig. 32. Valores porcentuales del Método Numérico para hembras sexualmente maduras de
258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. .............................................................................................53
Fig. 33. Valores porcentuales del Método de Frecuencia para hembras sexualmente maduras
de 258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ............................................................................................53
Fig. 34. Valores porcentuales del Método Gravimétrico para hembras sexualmente maduras de
258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................54
Fig. 35. Valores del Índice de Importancia Relativa (IIR) para hembras sexualmente maduras
de 258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................54
Fig. 36. Valores porcentuales del Método Numérico para hembras sexualmente inmaduras de
152 - 257 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................56
Fig. 37. Valores porcentuales del Método de Frecuencia para hembras sexualmente inmaduras
de 152 - 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................56

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxii
Fig. 38. Valores porcentuales del Método Gravimétrico para hembras sexualmente inmaduras
de 152 - 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................57
Fig. 39. Valores del Índice de Importancia Relativa (IIR) para hembras sexualmente inmaduras
de 152 – 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................57
Fig. 40. Valores porcentuales del Método Numérico para machos sexualmente maduros de 244
- 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del
2004. .....................................................................................................................59
Fig. 41. Valores porcentuales del Método de Frecuencia para machos sexualmente maduros
de 244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ...............................................................................................59
Fig. 42. Valores porcentuales del Método Gravimétrico para machos sexualmente maduros de
244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004. ................................................................................................................60
Fig. 43. Valores del Índice de Importancia Relativa (IIR) para machos sexualmente maduros de
244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004. ................................................................................................................60
Fig. 44. Valores porcentuales del Método Numérico para machos sexualmente inmaduros de
176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004. ...............................................................................................................62
Fig. 45. Valores porcentuales del Método de Frecuencia para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ................................................................................................62
Fig. 46. Valores porcentuales del Método Gravimétrico para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ..............................................................................................63
Fig. 47. Valores del Índice de Importancia Relativa (IIR) para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004. ...............................................................................................63

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxiii
Fig. 48. Dosidicus gigas, (a y c) picos superiores de 35.4 mm y 12.0 mm (LT); (b) pico inferior
de 18.8 mm LT. La pigmentación de la pared difiere en los distintos estadíos del
organismo. .............................................................................................................68
Fig. 49. Sthenoteuthis oualaniensis, (a) pico superior de 32.3 mm LT y (b) pico inferior de
23.1 mm LT. ..........................................................................................................68
Fig. 50. Ommastrephes bartramii, (a) pico superior de 18.8 mm LT; (b) pico inferior de 24.8
mm LT; (c) vista dorsal del superior donde se observa línea pigmentada que recorre la
mitad de la cresta. .................................................................................................69
Fig. 51. Octopoteuthis sicula, (a) pico superior de 14.1 mm LT; (b) pico inferior de 13.5 mm
LT. ........................................................................................................................69
Fig. 52. Loliolopsis diomedeae, (a-c) picos superiores de 6.0 y 5.9 mm LT; (b) pico inferior
de 4.7 mm LT. ........................................................................................................70
Fig. 53. Histioteuthis sp, (a) pico superior de 12.0 mm LT y (b) pico inferior de 13.3 mm LT .…
.............................................................................................................................70
Fig. 54. Histioteuthis sp, pico inferior de 15.5 mm LT. .........................................................71 Fig. 55. Onychoteuthis banksii, pico inferior de 11.0 mm LT. .............................................71
Fig. 56. Mastigoteuthis dentata, de longitudes progresivas; (a, c, e) picos superiores de 3.2
mm, 5.6 mm y 6.0 mm LT y (b, d, f) picos inferiores de 4.8 mm, 5.2 mm y 6.7 mm LT.
.............................................................................................................................72
Fig. 57. Mastigoteuthis dentata, de longitudes progresivas; (a – e) picos superiores de 5.7
mm, 8.4 mm, 8.6 mm, 9.3 mm y 12.0 mm LT. ..........................................................73
Fig. 58. Morfotipo A (Patrón A), pico superior de 13.3 mm LT. ............................................74 Fig. 59. Morfotipo B (Patrón B), pico superior de 20.2 mm LT. ............................................74
Fig. 60. Morfotipo C (Patrón C), (a y b) pico superior de 5.9 mm (LT) ..................................74 Fig. 61. Merluccius gayi , (a) arriba: esqueleto de 125.2 mm LT; abajo: de 90.9 mm LT; (b)
detalle de las vértebras abdominales que muestra las parapófisis replegadas en forma
de tubo propio de la especie; (c) detalle de vértebras caudales; (d) otolito de 12.4 mm
(LT) y (e) variaciones de la forma del mismo producto de la digestión. ......................75

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxiv
Fig. 62. Benthosema panamense, (a) esqueleto de 18.2 mm LT; (b) otolito de 1.8 mm LT
observado bajo estereoscopio en 12x y (c) variaciones en la morfología del mismo
como producto de la digestión. ................................................................................76
Fig. 63. Larimus sp, (a) esqueleto de 63.9 mm LT; (b) otolito de 7.2 mm LT y (c) variaciones
en la morfología del mismo por desgaste de la digestión. ........................................76
Fig. 64. Ophichthus sp, (a) esqueleto de 254.0 mm LT; (b) detalle de las expansiones de los
arcos hemales; (c) detalle de las vértebras caudales (Vista lateral) y (d) esqueletos
de 20.0 mm y 12.0 mm LT....................................................................................77
Fig. 65. Mustelus sp, (a) macho de 225 mm LT; (b) detalle de la boca de una hembra de 220
mm LT bajo estereoscopio en 12x; (c) vista lateral la misma y (d) arriba: hembra de 220
mm LT, centro: macho de 225 mm LT y abajo: hembra de 232 mm LT. ....................78
Fig. 66. Thunnus sp, (a) vista lateral del cráneo de 59.8 mm (Long. craneal) y (b - c) vista
dorsal del mismo. ...................................................................................................79
Fig. 67. Coryphaena hippurus, vértebras caudales de. 36.2 mm (LT). .................................79
Fig. 68. Morfotipo 1 (Patrón 1), (a) esqueleto de 22.2 mm LT; (b) de 19.7 mm LT; (c) de 29.5
mm LT y (d) comparación de magnitudes de los tres anteriores. ..............................80
Fig. 69. Morfotipo 2 (Patrón 2), (a) esqueleto de 39.3 mm (LT) y (b) detalle de las vértebras
.........................................................................................................................................80
Fig. 70. Morfotipo 3 (Patrón 3), (a) esqueleto de 50.9 mm (LT) y (b-c) detalle de las vértebras
abdominales ..........................................................................................................81
Fig. 71. Gasterópodos de (a) 1 mm LT y (b) de 3.2 mm LT presentes en el estómago de un
macho inmaduro de A. pelagicus de 190 cm LT observados bajo estereoscopio en 12 x.
Su consumo incidental obedece probablemente a que estos organismos pelágicos
estaban cercanas a la presa objetivo en el momento de la alimentación del organismo.
.............................................................................................................................81

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxv
LISTA DE ANEXOS
Anexos.-
Anexo 1. Certificado de partipación de tesistas Bolaño, Calle y Erazo en el proyecto Biología
de los Elasmobranquios (CICIMAR) e Investigaciones Biológicas de la Fauna
Incidental (CIAT).
Anexo 2. Certificado de la especie Alopias pelagicus para aguas ecuatorianas, extendido
como carta de respaldo para la tesis en mención, por del Dr. Felipe Galván del IPN –
CICIMAR de La Paz, Baja California del Sur, México.
Anexo 3. Registro de la presencia del Tiburón Zorro Alopias pelagicus en Aguas
Ecuatorianas. Villón y Aguilar, 2005 (con permiso de Aguilar).
Anexo 4. Certificado de la especie Alopias pelagicus para aguas ecuatorianas, extendido
como carta de respaldo para la tesis en mención, por del Dr. George H. Burgess del
Museo de Historia Natural de Florida de la Universidad de Florida, Gainsville, Florida,
EE.UU.
Anexo 5. Tabla de campo para los muestreos durante mayo – diciembre del 2004.
Anexo 6. Datos principales de campo para Alopias pelagicus muestreados durante mayo –
diciembre del 2004.
Anexo 7. Diferencias morfológicas entre el tiburón zorro pelágico Alopias pelagicus y el
tiburón zorro común Alopias vulpinus.
Anexo 8. Tabla de registro de laboratorio Anexo 9. Vista panorámica de Santa Rosa de Salinas (Marzo 3 del 2005)
Anexo 10. Crías de Alopias pelagicus (a) machos de 22.5 cm (arriba) y 23 cm (LT) (abajo)
sobre el dorso de una hembra adulta de 290 cm. (LT) y (b) gonopterigios de 3 cm
(LG) (Marzo 3 del 2005).
Anexo 11. Proceso de evisceración de Alopias pelagicus (a) corte de cabeza; (b) corte de
región ventral; (c) aparato digestivo; (d) aletas y (e) aleta pectoral de 40 cm. LA
(Marzo 3 del 2005).
Anexo 12. Tiburón zorro de anteojos Alopias superciliosus (a) ejemplares desembarcados;
(b) vista frontal donde se observa el surco encefálico y los ojos grandes en un

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxvi
organismo de 300 cm LT; (c) vista lateral y la disposición de sus aletas; (d) vista
lateral; (e) gonopterigio de 25 cm. (LG) y (f) apertura del rifiodón (Marzo 3 del 2005).
Anexo 13. Tiburón puntas negras Carcharhinus limbatus de 255 cm LT.
Anexo 14. Tiburón tinto Isurus oxyrhinchus de 250 cm LT (con permiso de Erazo, 2005).
Anexo 15. Boca de Tiburón tinto Isurus oxyrhinchus de 250 cm LT (Erazo, 2005).
Anexo 16. Tiburón azul Prionace glauca de 270 cm LT (Erazo, 2005).
Anexo 17. Tiburón tollo Mustelus sp. macho de 60 cm LT.
Anexo 18. Detalle de la cabeza de Dorado Coryphaena hippurus macho de 140 cm LT.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
xxvii
ÍNDICE
Temas Págs.
CARÁTULA.................................................................................................................ii
DEDICATORIA ...........................................................................................................vi
AGRADECIMIENTOS ...............................................................................................viii
RESUMEN ..................................................................................................................xi
ABSTRACT.................................................................................................................xiv
LISTA DE TABLAS, FIGURAS Y ANEXOS...........................................................xvii
LISTA DE TABLAS....................................................................................................xvii
LISTA DE FIGURAS..................................................................................................xiv
LISTA DE ANEXOS...................................................................................................xxv
ÍNDICE...................................................................................................................... xxvii
1. INTRODUCCIÓN......................................................................................................1
2. ANTECEDENTES ....................................................................................................4
3. MARCO CONCEPTUAL........................................................................................12
3.1. Aspectos biológicos de la familia Alopiidae ....................................................12
3.2. Aspectos biológicos de la especie Alopias pelagicus ...................................14
3.3. Contenido estomacal..........................................................................................20
3.4. Digestión...............................................................................................................20
4. MARCO LEGAL......................................................................................................22
5. HIPÓTESIS..............................................................................................................24
6. OBJETIVOS ...........................................................................................................25
7. ÁREA DE ESTUDIO ..............................................................................................26
8. METODOLOGÍA .....................................................................................................28
9. RESULTADOS........................................................................................................36
10. DISCUSIONES.....................................................................................................82
11. CONCLUSIONES.................................................................................................89
12. RECOMENDACIONES .......................................................................................91
13. LITERATURA CITADA........................................................................................92
14. GLOSARIO
15. ANEXOS

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
1
1.- INTRODUCCIÓN
Los tiburones son uno de los grupos más abundantes de depredadores en el
mar, que habitan todos los océanos del mundo (Gruber, 1977; Gruber y
Myrberg, 1977), sin embargo la información biológica con respecto a sus dietas
es escasa (Cortés, 1999). Los tiburones se consideran depredadores por
excelencia, y comprenden entre sus presas una gran variedad de organismos,
desde crustáceos planctónicos e invertebrados del fondo, cefalópodos, peces
óseos de diversas tallas, otros tiburones y rayas, hasta mamíferos marinos y
otros grandes vertebrados (Compagno, 1995). El tipo de alimentación que más
se aparta de la del resto de los tiburones es la de los filtradores de plancton
que corresponde al tiburón ballena (Rhincodon typus), al tiburón peregrino
(Cetorhinus maximus) y al tiburón boquiancho (Megachasma pelagios)
(Stevens, 1992).
Los tiburones están encargados de cumplir un importante rol en la red
alimenticia oceánica a través de su historia evolutiva. Mientras es ampliamente
reconocido que muchas especies existentes de tiburones están entre los
depredadores cúspide de la cadena en las comunidades marinas,
sorpresivamente poca información cuantitativa se encuentra disponible de sus
dietas (Brooks y Dodson, 1965). Gracias a sus hábitos actúan como
carroñeros ayudando a eliminar de las aguas los animales muertos,
impidiéndose así la propagación de enfermedades y fortaleciendo la
composición genética de las poblaciones de presas. Como depredadores
ayudan a mantener la cadena alimenticia marina (Galván, Nienhuis y Klimley,
1989). Si los tiburones son removidos de la cadena trófica, se causará un
efecto en cadena que afectará la estabilidad del ecosistema marino (Pauly,
Christensen, Dalsgaard, Froese y Torres, 1998).
Debido a la creciente demanda internacional de aletas de tiburones y otros
productos derivados se está conduciendo a un enorme incremento en la pesca
de éstos animales alrededor del mundo. Los tiburones son particularmente
vulnerables porque son especies relativamente longevas y de baja proporción
reproductiva. Algunos tiburones no alcanzan la madurez sexual hasta edades

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
2
tardías y producen un pequeño número de crías. Por ende, las poblaciones de
tiburones son lentas en responder cuando son agotadas por sobrepesca
(Trejo, 2005).
El tiburón zorro pelágico, zorro, rabón o coludo Alopias pelagicus es una
especie típica costera y oceánica con una gran cola, la cual abarca casi la
mitad de su longitud total; de coloración azul grisácea a gris en el dorso y
blanca en la superficie ventral. Es una especie muy activa y un nadador veloz.
Se alimenta de peces y calamares que reúne con las ondas producidas por su
cola al nadar alrededor de los mismos y luego los golpea para consumir a
aquellos que quedan aturdidos o muertos.
La mayoría de los autores sostienen la importancia de los tiburones zorros,
especialmente del tiburón zorro pelágico Alopias pelagicus dentro de la
pesquería nacional debido al número y volumen que aportan éstos dentro de
las capturas de pelágicos mayores en la pesca artesanal.
Un aspecto básico en la biología de cualquier especie es el conocimiento de su
alimentación, ya que es un factor que determina adaptaciones anatómicas,
fisiológicas y etológicas. El estudio de la alimentación de forma integral aporta
información biológica básica que permite conocer las vías del flujo energético
en las comunidades y permite comprender las interacciones que se establecen
entre las especies, como la depredación y la competencia (Aburto, 1997).
Los estudios sobre taxonomía y biología de los tiburones en nuestro país son
escasos y a nivel regional sólo se conocen trabajos sobre la alimentación del
tiburón zorro pelágico (A. pelagicus) y del tiburón zorro de anteojos
(Alopias superciliosus), así mismo de los hábitos alimenticios de dos
especies de tiburones martillos: la cachona (Sphyrna zygaena) y de la
cachona ondulada (Sphyrna lewini) y del tiburón mico
(Carcharhinus falciformis) en la Playa de Tarqui, Manta (Manabí) durante el
2003 y 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
3
El presente trabajo permite el conocimiento del espectro trófico del tiburón
zorro pelágico Alopias pelagicus mediante la identificación de las especies
presas consumidas y la relación existente entre éstas y el sexo, la madurez
sexual y las tallas de los organismos, para de ésta manera poder entender la
dinámica energética que se suscitan en la cadena alimenticia marina y la
relación depredador - presa. De igual manera podrá servir de base para la
implementación de un manejo sostenido y sustentable para la pesquería de
ésta especie.
Cabe resaltar que este estudio forma parte del proyecto internacional “Red de
Alimentación Oceánica de Pelágicos Mayores del Pacífico Oriental” financiado
por la Universidad de Hawaii (EE.UU.), con la colaboración de la Comisión
Interamericana del Atún Tropical (CIAT), Centro Interdisciplinario de Ciencias
Marinas (CICIMAR) del Instituto Politécnico Nacional (IPN) (México) bajo la
dirección del Dr. Felipe Galván y del Dr. Roberth Olson. Además, que cuenta
con la ayuda de la Universidad Laica “Eloy Alfaro” de Manta y de la Universidad
de Guayaquil (Anexo1).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
4
2. ANTECEDENTES
Los estudios realizados en nuestro país se basan en trabajos de taxonomía,
diagnósticos de pesquerías, desembarques y utilización del tiburón realizados
por el Instituto Nacional de Pesca (INP). Es menester mencionar que la especie
en estudio se la citaba como Alopias vulpinus o Alopias sp. debido a una
identificación incorrecta, hasta que finalmente Villón y Aguilar (2005) reportan,
a nivel nacional, al tiburón zorro pelágico o tiburón rabón Alopias pelagicus
para nuestras costas.
Los estudios taxonómicos de la fauna ictiológica en el Ecuador, empezaron con
Orcés (1959) en su documento de contribuciones al conocimiento de los peces
marinos del Ecuador donde hizo una primera revisión de los peces, basado en
colecciones conservadas en Quito.
Cobo y Massay (1969) presentaron una lista de peces marinos del Ecuador,
donde no se reportó a la familia Alopiidae para nuestras aguas.
Massay (1983) en una revisión de la lista de peces marinos del Ecuador reportó
la presencia de dos especies de tiburones zorros A. vulpinus y
Alopias superciliosus para aguas ecuatorianas.
Béarez (1996) en una lista de peces marinos del Ecuador Continental describió
la ictiofauna pacífica en los puertos de Salango y Puerto López, Manabí,
encontrando una especie nueva y 14 nuevos registros lo que facilitó un
conocimiento más detallado de las especies de peces que se hallan en el
sector.
Massay y Massay (1999) en un reporte sobre los peces marinos del Ecuador,
reafirmaron la presencia de las dos especies de tiburones zorros marinos
(A. vulpinus y A. superciliosus).
Jiménez y Béarez (2004) en su trabajo sobre los peces marinos del Ecuador
continental, presentaron una clave para la identificación de familias y una lista

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
5
de especies de peces donde registraron a A. pelagicus. Además de una breve
taxonomía, descripción, distribución, hábitat y otros aspectos de los principales
peces comerciales, dentro de los cuales cita solamente al tiburón zorro de
anteojos (A. superciliosus) dentro de la familia Alopiidae.
Los trabajos relacionados a los diagnósticos de pesquerías y desembarques
han sido numerosos. Entre los que podemos citar a los siguientes:
Herdson, Rodríguez y Martínez (1985) en una investigación realizada sobre los
desembarques artesanales en las costas del Ecuador y sus capturas en el año
1982, manifiestan que éstos fueron entre 15 000 y 20 000 t, dentro de los
cuales los tiburones representaron el 10%. Sólo cita la presencia de
A. superciliosus en dichas capturas.
Martínez, Cobeña y Domínguez (1997), en un estudio realizado sobre las
pesquerías artesanales en Puerto López (Manabí) durante 1996 registraron
seis familias de tiburones (12 especies) y estimaron que el volumen total
desembarcado fue de 1 348 t, correspondiendo a los tiburones 125 t (9%); la
familia Alopiidae, ocupó el primer lugar, en volumen, seguido de las familias
Carcharhinidae, Lamnidae, Sphyrnidae y Triakidae.
Solís (1998) en el diagnóstico de la actividad pesquera artesanal en el puerto
de Santa Rosa, Guayas citó una lista de especies de tiburones desembarcados
en este puerto. Así mismo sostuvo que el desembarque total estimado para el
segundo semestre de 1997 fue de 1 122,5 t y que los tiburones representaban
el 5,5 % (61,66 t).
Martínez (1998) presentó un informe de estudios para la FAO sobre el manejo
de las pesquerías en Ecuador, donde describió la pesquería de las 38 especies
de tiburones que habitan en esta área y la exportación de cada una de sus
partes a diversos países, sobre todo a Japón y hace énfasis en los organismos
de control encargados de regular la explotación de este recurso a nivel
industrial como lo es el INP y la Subsecretaria de Recursos Pesqueros (SRP).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
6
Marín de López, Arriaga y Ormaza (1999) llevaron a cabo un trabajo sobre las
estadísticas de los desembarques pesqueros en Ecuador 1985 – 1997, en
donde establecieron las capturas para la flota industrial (cerquera – palangrera
– arrastrera) de los tiburones en general; durante 1985 – 1989 y 1996 – 1997,
donde se incrementa la captura de 151,3 a 1 547,5 TM (toneladas métricas); no
hubo registros desde 1990 hasta 1995. Mientras que para la flota artesanal, los
desembarques de 1985 a 1997 se presentan de 572,2 a 4 885,9 TM,
representando el tiburón rabón (Alopias spp.) casi el 50% de las capturas.
Revelo y Herrera (1999) en un estudio sobre los desembarques de la pesca
artesanal en ocho puertos de la costa continental ecuatoriana durante 1998,
estimaron que el desembarque total de tiburones fue de ca. 980 t, y que la
familia Alopiidae presentó mayor desembarque (586,3 t), seguida de las
familias Carcharhinidae, Lamnidae, Sphyrnidae y Triakidae.
Peralta (1999) en un documento sobre los desembarques de la pesca artesanal
en ocho puertos de la costa continental ecuatoriana durante el primer trimestre
de 1999, estimó que el desembarque total de tiburones fue de ca., 401,4 t,
siendo la familia Alopiidae (tiburón rabón, A. vulpinus) la más abundante
(292,9 t), seguido de la familia Carcharhinidae (72,3 t). Luego, las familias
Lamnidae, Sphyrnidae y Triakidae.
Dora (1999) en un estudio sobre los desembarques de la pesca artesanal en
ocho puertos de la costa continental durante el segundo trimestre de 1999,
determinó que el desembarque total de tiburones fue de ca., 222,9 t, y que la
familia Alopiidae presentó mayor abundancia (96,6 t), seguida de las familias
Carcharhinidae, Lamnidae, Sphyrnidae y Triakidae.
Alcívar y López (2001) realizaron un trabajo acerca de la pesca artesanal y
comercialización de tiburones zorros (Alopias superciliosus y
Alopias vulpinus) en Manta, haciendo énfasis en los individuos capturados,
su utilización, y otros aspectos pesqueros relacionados.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
7
Zarate (2002) en un trabajo de tiburones presentó un listado de 29 especies en
Galápagos, de las cuales cita a A. vulpinus, A. pelagicus y A. superciliosus
junto con otras especies explotadas. Además cita al tiburón blanco
Carcharodon carcharias como parte de los tiburones que habitan en las
aguas de la Reserva Marina de Galápagos.
Mora (2005) en su trabajo sobre el recurso tiburón en la pesca artesanal de la
costa continental ecuatoriana estimó en el 2004 un desembarque de tiburones
de 1 225,33 t, en donde A. pelagicus fue la especie mayormente capturada
por la flota artesanal con 487,16 t.
Las investigaciones biológicas se reportan sólo para la provincia de Manabí.
Baigorrí y Polo (2004) realizaron el primer estudio a nivel nacional sobre el
espectro trófico de dos especies de tiburón zorro A. pelagicus y
A. superciliosus en la playa de Tarqui, Manta, durante junio – diciembre del
2003, donde determinaron que los mayores componentes de la dieta
alimenticia para A. pelagicus eran principalmente el calamar gigante
(Dosidicus gigas) y la sardina luminosa (Benthosema panamense), mientras
que A. superciliosus presentaba mayor preferencia por peces
(Larimus argenteus, Merluccius gayi y B. panamense).
Castañeda y Sandoval (2004) realizaron una investigación sobre los hábitos
alimenticios de dos especies de tiburones martillos: Sphyrna zygaena y
Sphyrna lewini; en la playa de Tarqui, Manta, durante junio – diciembre del
2003.
Estupiñán y Cedeño (2005) también estudiaron los hábitos alimenticios del
tiburón mico Carcharhinus falciformis, de S. lewini y S. zygaena en la playa
de Tarqui, Manta, durante el 2004.
Debido a la incorrecta identificación de ésta especie desde hace 23 años
siempre ha sido citada como A. vulpinus o Alopias sp., por lo que es
necesario hacer hincapié en la verdadera identificación de la misma, siendo así

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
8
A. vulpinus es en realidad el tiburón zorro pelágico A. pelagicus que
comúnmente es desembarcado en nuestros puertos pesqueros.
Estupiñán (Com. pers. 2006) sostuvo que Galván identificó por primera vez a
A. pelagicus para aguas ecuatorianas durante una charla en Manta en el
2002. El siguiente año, Galván (Com. pers. 2003) en una conferencia sobre
biología básica de tiburones realizada en el Instituto Nacional de Pesca en
Guayaquil (27 de agosto del 2003) reportó nuevamente la presencia del
Tiburón Zorro Pelágico A. pelagicus, para nuestras aguas y advierte que dicha
especie se la estaba confundiendo con el Tiburón Zorro Común A. vulpinus.
Además citó que ésta última no presentaba distribución para nuestro país ya
que es propia de aguas templadas y frías y que se lo reconoce fácilmente por
ciertas características externas como la presencia de una banda blanca por
ambos lados del tronco, la misma que sobrepasa la altura de las aletas
pectorales.
Baigorrí y Polo (2004) reportaron correctamente a la especie A. pelagicus en
su tesis de grado.
Jiménez y Béarez (2004) registraron a A. pelagicus en su lista de especies de
peces marinos continentales.
Galván (Com. pers. 2005) en un certificado de la especie A. pelagicus
existente para aguas ecuatorianas, extendido el 11 de abril del 2005, que sirvió
de carta de respaldo para el trabajo en mención, verifica nuevamente a la
especie (Anexo 2).
Villón y Aguilar (2005) en un informe técnico acerca del registro de la presencia
de tiburón zorro (A. pelagicus) en aguas ecuatorianas reportaron oficialmente
la existencia de ésta especie para aguas ecuatorianas (Anexo 3).
Burgess (Com. pers. 2006) en un certificado de la especie A. pelagicus para
aguas ecuatorianas, extendido el 07 de marzo del 2006, como otra carta de

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
9
respaldo para el presente estudio, la certifica y ratifica nuevamente como una
especie común en nuestra región (Anexo 4).
Las investigaciones biológicas realizadas a nivel mundial, para ésta especie y
las otras dos especies de la Familia Alopiidae están enmarcados en diversos
temas de aspecto biológico, como morfología, crecimiento, alimentación,
reproducción y genética, también se incluye un trabajo de pesquería.
Castro y De Lachica (1973) en su documento sobre nuevos registros de peces
marinos en la costa del Pacífico mexicano realizaron el primer registro de
A. pelagicus con base en dos embriones de una hembra, en las cercanías de
las Islas Marías, México en el Pacífico Oriental.
Villavicencio, Estrada y Downton (1997), en el primer registro de A. pelagicus
en el Golfo de California, México reportan por primera vez la presencia de ésta
especie para dicha zona.
En cuanto a los aspectos de edad y crecimiento se encuentra el trabajo de Liu,
Chiang y Chen (1998), en Taiwán, donde estimaron la edad (10 -12,5 años) y el
tamaño (270 - 350 cm LT) en que los organismos alcanzan la talla de
reclutamiento en A. superciliosus.
Las investigaciones relacionadas con la ecología trófica de la especie citan a
Moteki, Arai, Tsuchiya y Okamoto (2001) en un estudio sobre la composición
de las presas en la dieta de los peces pelágicos grandes en el Océano Pacífico
Tropical Oriental, sostuvieron que los Paralepididos de por lo menos cuatro
especies comprendieron el 52,0% del número de peces consumidos por
A. pelagicus y que aparecieron con la frecuencia más alta (65,0%). Aunque los
Fosíctidos (Polymetme sp.) tuvieron el segundo lugar numéricamente (25,0%),
la frecuencia fue menor (5,0%). Estos organismos consumieron pequeños
peces mesopelágicos tales como Paralepididos, Fosíctidos y Gempílidos,
mientras que peces asociados al mar de los Sargazos no se encontraron en
dichos estómagos.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
10
Preti y Smith (2001) presentaron un estudio acerca de los hábitos alimenticios
del tiburón zorro común (A. vulpinus) muestreados en las pesquerías de
California 1998 - 1999, donde determinaron la dieta del tiburón zorro
A. vulpinus de 165 individuos mediante medidas de frecuencia de ocurrencia
(FO) y la determinación del índice de importancia relativa (IRI), identificando
todas las presas consumidas hasta el menor taxón posible, hallando que la
presa más importante fueron la anchoveta (Engraulis mordax) y de los
invertebrados el crustáceo (Pleuroncodes planipes) fue el más importante en
número.
Seitz (2003) afirmó que como todos los zorros, A. pelagicus se alimenta de
peces, especialmente de arenques (Familia Clupeidae), peces voladores
(Familia Exocoetidae) y macarelas (Familia Scombridae). Así mismo también
ingiere calamares pelágicos.
Entre los estudios de reproducción se cita a Otake y Mizue (1981) quienes
realizaron un estudio sobre la evidencia directa de la oofagia que se presenta
en embriones de A. pelagicus llegando a la conclusión que es un mecanismo
especial de nutrición el cual solo había sido reportado por un solo autor.
Moreno y Morón (1992) y Chen (1997) describieron aspectos reproductivos y
algunas características morfológicas de A. superciliosus capturados en el
Mediterráneo occidental y el Atlántico nororiental, respectivamente encontrando
una talla promedio de madurez para las hembras entre 332 - 341 cm LT y para
los machos de 270 - 288 cm LT.
Trejo (2005) en un estudio sobre filogeografía global (estructuración de
poblaciones mundiales) de tiburones zorros (Alopias spp.) basados en datos
de secuencia de ADN mitocondrial de un total de 149 muestras de tejidos
obtenidos de las costas Este y Oeste de los Estados Unidos, Golfo de
California, Hawaii, Golfo de México, Clipperton, Guatemala, Ecuador, Francia,
Sudáfrica, Taiwán, Indonesia, Nueva Caledonia y Nueva Zelanda estableció
que a pesar de su gran potencial de dispersión, el flujo genético entre éstas
poblaciones es limitado, pero los patrones filogeográficos difieren entre

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
11
especies. El flujo genético para A. pelagicus es limitado a través del Océano
Pacífico, pero es extenso entre sitios tanto para el Oriental y Occidental.
En cuanto a las pesquerías, Williams (1997) en un estudio realizado sobre las
capturas de tiburones y especies relacionadas en las pesquerías de atunes del
Océano Pacífico Tropical Oriental y Central, resaltó la presencia significativa de
A. pelagicus y A. superciliosus.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
12
3. MARCO CONCEPTUAL
3.1. ASPECTOS BIOLÓGICOS DE LA FAMILIA ALOPIIDAE
Los tiburones zorros tienen un cuerpo cilíndrico y moderadamente robusto, una
cabeza corta de longitud muy inferior a la del tronco; hocico moderadamente
largo, aguzado y cónico, no prolongado, ni aplanado o laminar, la boca es
pequeña y arqueada en posición ventral (Fig. 1); tienen dientes pequeños o
más o menos grandes de forma laminar, similares en ambas mandíbulas, los
dientes anteriores de la mandíbula superior son un poco más grandes que los
dientes laterales y separados de éstos por una hilera de pequeños dientes
intermedios; los ojos pueden ser de tamaño mediano a muy grande, aberturas
branquiales cortas confinadas a los lados de la cabeza, los últimos dos pares
situados por encima de las bases de las aletas pectorales, la primera aleta
dorsal es grande, alta, vertical y angulada, mientras que la segunda aleta
dorsal y anal son diminutas, bajas y de bases pivotantes (girables), la base de
la aleta anal esta situada por detrás de la segunda dorsal; las aletas pectorales
son muy largas y angostas (más largas que la cabeza en adultos), las aletas
pélvicas muy largas aproximadamente de igual longitud que la primera dorsal;
aleta caudal no semilunar, su lóbulo dorsal muy alargado casi tan largo como el
resto del animal, el lóbulo ventral corto, pero fuerte. (Compagno, 1984;
Compagno, Krupp y Schneider, 1995).
Los zorros son nadadores activos y muy resistentes que viven en aguas
costeras y oceánicas desde la superficie hasta grandes profundidades, se
encuentran en todos los mares tropicales, subtropicales, templados y fríos. Al
parecer, son muy especializados en sus hábitos alimenticios, atacan a
pequeños y grandes cardúmenes de peces y calamares, nadando alrededor de
ellos en círculos cada vez más estrechos y golpeando el agua con su larga
cola hasta concentrarlos en una pequeña superficie. Los golpes de la aleta
caudal también tienen el efecto de aturdir y matar las presas, pero en el curso
de esa acción, los zorros suelen ser capturados por su cola en el anzuelo
de palangres flotantes (Compagno, 1984; Compagno et al, 1995).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
13
Fig. 1. Morfología externa del tiburón zorro (Compagno, Krupp y Schneider, 1995)
Las tres especies de esta familia (A. pelagicus, A. superciliosus y
A. vulpinus) tienen áreas de distribución y hábitat superpuestos, pero las
diferencias en su morfología, hábitos alimenticios y distribución espacial, hacen
pensar que logran reducir la competencia ínterespecífica a través de una cierta
subdivisión del hábitat y de las presas que en él habitan (Compagno, 1984;
Compagno et al., 1995).
Las tres especies son nadadoras activas y vigorosas. El desarrollo de la cola,
casi tan largo como el resto del cuerpo, no merma la velocidad o la eficacia
depredadora. Los zorros marinos hacen servir su larga cola para aturdir a los
peces; las vértebras del extremo de la cola tienen las apófisis ventrales y
dorsales alargadas de forma que constituyen una verdadera maza (Stevens,
1992). Los zorros marinos se alimentan de presas pequeñas: peces pelágicos y
de fondo que formen bancos, pulpos y crustáceos (Compagno, 1992).
Los zorros representan un componente importante en las capturas de tiburones
en aguas oceánicas. Su carne es de muy buena calidad y se comercializa en
fresco, congelada, ahumada, salada y desecada. Sus aletas son utilizadas para
las sopas orientales, el hígado para la extracción de vitaminas y la piel para la
producción de cuero. Existen importantes pesquerías pelágicas de zorros con
palangres flotantes en el noreste del Océano Indico, en el Océano Pacífico
central y en el Océano Atlántico nororiental. Hace algunos años, se inició una
importante pesquería pelágica de tiburones zorros con redes de enmalle en el
Pacífico Oriental, frente a las costas de California, pero esta pesquería

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
14
recientemente declinó, probablemente debido a sobrepesca. Esta familia
comprende un solo género con 3 especies (Compagno, 1984; Compagno et al.,
1995).
3.2. ASPECTOS BIOLÓGICOS DE LA ESPECIE Alopias pelagicus
Clasificación Taxonómica (Compagno 1984):
REINO: ANIMALIA
PHYLUM: CHORDATA
SUBPHYLUM: VERTEBRATA
SUPERCLASE: PISCES
CLASE: CHONDRICHTHYES
SUBCLASE: ELASMOBRANCHII
ORDEN: LAMNIFORMES
FAMILIA: ALOPIIDAE
GÉNERO: Alopias
ESPECIE: pelagicus
N. CIENTÍFICO: Alopias pelagicus, Nakamura 1935
N. VULGAR: Tiburón zorro pelágico
Nombres Vernáculos: FAO: Es – Zorro pelágico, zorro marino; Fr – Renard
pélagique; In – Pelagic Thresher Shark (Compagno, 1984; Compagno et al.,
1995). Al - Pazifischer Fuchshai (Godknecht, 2004).
Caracteres distintivos: Es una especie de gran talla, en la cabeza tiene cinco
aberturas branquiales de tamaño mediano, las últimas dos se encuentran
encima de las bases de las aletas de pectorales, posee un surco horizontal
poco evidente a cada lado de la nuca, el cual empieza a nivel de la boca hasta
las aletas pectorales; no tiene barbillones nasales o surcos oro-nasales, el
hocico es largo y cónico; el perfil de la frente casi recto y el espacio ínter-
orbital fuertemente convexo (Compagno, 1984; Compagno et al., 1995) (Fig. 2).
La cabeza es angosta con ojos sin párpados nictitantes, estos son de tamaño
mediano en adultos y subadultos, pero muy grandes en ejemplares jóvenes,

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
15
boca larga y semicircular situada debajo de los ojos (Fig. 2) tiene dientes
pequeños de bordes cortantes, dispuestos en más de 29 hileras en cada
mandíbula, con una sola cúspide angosta, casi vertical u oblicua vista
distalmente y a menudo con una cúspide accesoria distal (Compagno, 1984
Compagno et al., 1995) (Figs. 3 - 6).
Fig. 2. Tiburón zorro pelágico Alopias pelagicus hembra de 305 cm (LT) en el puerto de Santa
Rosa de Salinas (Marzo, 2005)
Los dientes anteriores no son muy grandes, los de la mandíbula superior están
separados de los dientes laterales por dientes intermedios más pequeños
(Fig. 5). Poseen dos aletas dorsales, la primera en posición equidistante de las
bases de las aletas pectorales y pélvicas aunque se encuentra un poco más
cerca de las pectorales, la segunda aleta dorsal es diminuta y esta situada
delante de la pequeña aleta anal; las aletas pectorales son angostas, largas y
casi rectas, no falciformes y de ápices anchos; el lóbulo dorsal de la aleta
caudal muy largo y en forma de cinta, su longitud es aproximadamente igual al
resto del animal y el lóbulo ventral corto, pero robusto; los Alópidos tienen en
la parte dorsal un color que varía entre azul o gris con reflejos plateados; el
vientre es blanco, sin embargo el color blanco del vientre no se extiende por
30 cm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
16
encima de las bases de las aletas pectorales (Compagno, 1984 ; Compagno et
al., 1995) (Fig. 7).
Fig. 3. Esquema de ciertas características morfológicas de A. pelagicus (Compagno et al.,
1995).
Fig. 4. Mandíbula de A. pelagicus mostrada en forma panorámica (Noviembre, 2005).
Detalles ampliados de zonas marcadas con flechas anaranjadas en figuras 5 y 6.
Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
17
Fig. 5. Detalle de los dientes de la región derecha de la mandíbula superior de A. pelagicus,
donde se observa la cúspide central, angosta y oblicua (flecha lila) y las cúspides
accesorias (flechas blancas) (Noviembre, 2005).
Fig. 6. Detalle de los dientes de la región derecha de la mandíbula inferior de A. pelagicus,
donde se observa la cúspide central, angosta y oblicua (flecha lila) y las cúspides
accesorias (flechas blancas) (Noviembre, 2005).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
18
Fig. 7. Vista lateral de A. pelagicus donde se aprecia la disposición de las aletas y
parcialmente la longitud de la enorme cola (Marzo, 2005).
Talla Máxima: Por lo menos 3,3 m LT (hembras adultas); la talla de madurez
para hembras oscila entre 2,8 – 2,9 m LT y para machos entre 2,6 – 2,7 m LT
(Compagno, 1984; Compagno et al., 1995).
Hábitat y Biología: Es una especie circumtropical, primordialmente oceánica y
epipelágica, pero a veces capturada cerca de la costa entre la superficie y por
lo menos hasta 152 m de profundidad. Es un nadador activo y resistente, pero
su biología es poco conocida. Es un organismo ovovivíparo, por lo menos con
dos embriones (Fig. 8), tal como otras especies de Alopias, parece ser un
caníbal uterino. Probablemente se alimenta de peces pequeños y calamares
(Compagno, 1984; Compagno et al., 1995).
Pesca y utilización: Esta especie ha sido fuertemente explotada en la
pesquería de palangres en el Océano Indico noroccidental (principalmente por
Rusia), pero también se pesca en el Océano Pacifico central y probablemente,
en otras zonas. La carne se comercializa para el consumo humano, el hígado
para la extracción de vitaminas, la piel para la fabricación de cuero y las aletas
para la preparación de sopas (Compagno, 1984; Compagno et al., 1995).
40 cm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
19
Fig. 8. Dos crías de A. pelagicus sobre el cuerpo de una hembra adulta, en Santa Rosa de
Salinas. Arriba: Macho de 55,5 cm (LT) y abajo: Hembra 57,2 cm (LT) (Marzo, 2005).
Distribución: En contraste, a la distribución mundial de A. vulpinus y
A. superliciosus, Alopias pelagicus sólo se encuentra en los Océanos
Pacífico, tanto Occidental como Oriental, e Índico (Trejo, 2005) (Fig. 9).
Fig. 9. Distribución de A. pelagicus en Pacífico Oriental (Compagno et al, 1995).
10 cm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
20
3.3. CONTENIDO ESTOMACAL
El análisis de la alimentación puede realizarse mediante estudios de contenido
estomacal y coprológico (estudio de la materia fecal), haciendo observaciones
directas de lo que consume el pez dependiendo del objetivo de la investigación
se utilizará un método específico. Para el estudio del contenido estomacal, hay
una gran variedad de métodos, algunos se mencionan a continuación (Galván
et al, 1989):
Se han considerado dos formas principales para determinar los niveles tróficos
de organismos marinos:
a) Los estudios de composición de dietas que usan las proporciones relativas
de los distintos tipos de presa y su nivel trófico respectivo (Galván Com. pers.,
2004).
b) El análisis de isótopos estables, el cual provee estimaciones del alimento
asimilado basado en las mediciones de isótopos estables de carbono y
nitrógeno en los tejidos de los consumidores marinos (Galván Com. pers.,
2004).
3.4. DIGESTIÓN
Cualquiera que sea el método de captura, la presa llega al estómago donde es
atacada por los jugos gástricos. Los productos de la digestión se absorben en
el intestino, que en los tiburones contienen la llamada válvula espiral, un
órgano en forma parecida a la de la forma de una escalera de caracol. Esta
disposición proporciona una superficie da absorción máxima en un pequeño
espacio, dejando sitio para un estómago y un hígado mayores y en las
hembras de especies vivíparas para el desarrollo de los embriones (Stevens,
1992).
El examen del contenido estomacal demuestra que pocos tiburones tienen el
estómago lleno. Un estudio sobre el tiburón tinto (Isurus oxyrhinchus) reveló

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
21
que la capacidad media del estómago de esta especie era del 10% del peso
del cuerpo, mientras que la cantidad de comida que contenía era sólo, por
término medio, el 2,6% del peso corporal. Un tiburón tinto de 63 kg tendría que
comer 2 kg de alimento al día, once veces y medio su propio peso al año. Las
necesidades mínimas de mantenimiento (la cantidad de alimento necesaria
para mantener el peso, sin que haya ningún crecimiento) de un tiburón galano
(Negaprion brevirostris) joven es de 20 600 calorías o 16,5 g de comida al
día. Para un tiburón de 1 kg esto equivale al 1,7% de su por día (Stevens,
1992).
En cautiverio un tiburón consume entre 3 y un 14% de su peso semanalmente
o entre el 0,4 y el 2% al día. Estos individuos pueden dejar de comer durante
varios meses, en cuyo tiempo viven presumiblemente a costa de las reservas
que tienen en el hígado. Los tiburones blancos (C. carcharias) pueden ingerir
grandes cantidades de alimento de una sola vez y se ha calculado que una
sola comida podría satisfacer los requerimientos del animal durante dos meses.
Pero, parece ser que la mayor parte de los tiburones comen cada dos o tres
días de un 3% a un 5% de su peso (Stevens, 1992).
Estudios realizados con tiburones galano (N. brevirostris), tiburones trozo
(Carcharhinus plumbeus) y los tiburones azules (Prionace glauca) han
demostrado que la digestión inicial es bastante rápida, unas 24 horas, pero que
el alimento tarda dos o tres días en ser evacuado por completo. El proceso es
más largo cuando el agua está más fría: la mielga (Squalus acanthias), por
ejemplo, tarda cinco días en digerir por completo los arenques cuando la
temperatura es de 100C. Los Lámnidos además de tener una temperatura
muscular elevada, tienen un sistema intercambiador de calor en los vasos que
irrigan las vísceras. El hecho de tener el intestino más caliente les permite
digerir el alimento más rápidamente (Stevens, 1992).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
22
4. MARCO LEGAL
4.1. EL SECTOR PESQUERO ECUATORIANO
La flota pesquera ecuatoriana está conformada por dos sectores: el industrial y
el artesanal. En el sector pesquero industrial operan las flotas: cerquera
atunera, cerquera costera, arrastrera camaronera y la palangrera asociada,
mientras que el sector pesquero artesanal está compuesto por varios tipos de
embarcaciones que van desde las balsas, canoas de madera, botes de fibras
de vidrio, balandras y barcos; éstas dos últimas embarcaciones son usadas
como barcos “nodrizas” que llevan de 3 a 15 botes de fibra de vidrio,
remolcando hasta las zonas de pesca (Aguilar, Chalén, Villón, Solís y Gaibor,
2005)
El sector pesquero ecuatoriano está regulado por las siguientes instituciones:
El Ministerio de Comercio Exterior, Industria, Pesca y Competitividad (MICIP),
la Subsecretaría de Recursos Pesqueros (SRP), el Consejo Nacional de
Desarrollo Pesquero (CNDP), la Dirección General de Pesca (DGP) y el
Instituto Nacional de Pesca (INP) (Aguilar et al., 2005).
4.2. LEGISLACIONES PARA LA PESCA DEL TIBURÓN EN EL ECUADOR.
A continuación se resumen los Acuerdos y Decretos relacionados con la
protección y manejo de los tiburones en el Ecuador: (Aguilar et al., 2005)
El Acuerdo No. 151 trata sobre la Reserva de la Pesca Artesanal y la Zona de
Amortiguamiento dentro de Galápagos donde se prohíbe la pesca del tiburón
en dicha zona, así como el transporte y comercio para el continente de sus
aletas.
El Acuerdo No. 097 habla de las regulaciones para la captura y
comercialización del tiburón donde se prohíbe extraer las aletas de este
recurso y devolver el cuerpo no aprovechado al mar (aleteo), por lo que su

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
23
carne debe ser utilizada íntegramente. Se debe enviar un reporte sobre la
captura del organismo y el destino de su carne a la DGP para llevar un control.
El Acuerdo No. 036 se enfoca en las regulaciones para la comercialización de
aletas del tiburón donde los exportadores y las exportaciones de aletas de
tiburones provenientes de la pesca incidental en el mar continental deben estar
autorizados por la DGP. Además, de las sanciones a los infractores de la ley.
El Decreto No. 2130 es una modificación del acuerdo No. 036, donde se
prohíbe la exportación o comercialización de las aletas aunque éstas
provengan de la captura incidental, las sanciones; la prohibición de pesca
dirigida de tiburones, la importación de los artes de pesca para este fin y el
aleteo.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
24
5. HIPÓTESIS
La alimentación en los tiburones se relaciona directamente con la edad,
crecimiento, sexo y madurez sexual de éstos organismos, de lo que se deriva
la composición cualitativa y cuantitativa de sus dietas, cuyo análisis conlleva al
conocimiento de su espectro trófico.
Ho: La determinación de la dieta del tiburón zorro pelágico Alopias pelagicus
a través de la identificación de las especies presas consumidas permitirá
establecer similitudes tróficas; así mismo la abundancia de sus componentes y
la nutrición generalista de la especie.
Ha: La determinación de la dieta del tiburón zorro pelágico Alopias pelagicus
a través de la identificación de las especies presas consumidas permitirá
establecer diferencias tróficas; así mismo la escasez de sus componentes y la
nutrición específica de la especie.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
25
6. OBJETIVOS
6.1. OBJETIVO GENERAL:
Determinar el espectro trófico de la especie Alopias pelagicus Nakamura
1935 en Santa Rosa de Salinas, durante Mayo - Diciembre del 2004 a través
de la identificación de las especies presas consumidas y su relación con el
sexo, madurez sexual y tallas.
6.2. OBJETIVOS ESPECÍFICOS:
a) Identificar las especies presas de la dieta del organismo hasta el mínimo
taxón posible.
b) Determinar la importancia de los componentes alimenticios por medio del
Índice de Importancia Relativa (IIR).
c) Determinar la diversidad de presas de la dieta por medio del Índice de
Shannon y Weaver (H’), así como la diversidad de las mismas con el Índice de
Pielou (E).
d) Determinar la amplitud del nicho trófico de la especie mediante el Índice de
Levin (Bi).
e) Determinar diferencias en el tipo de alimento por sexo, madurez sexual y
tallas con el Índice de Morista - Horn (Cλ).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
26
7. ÁREA DE ESTUDIO
El puerto pesquero de Santa Rosa, localizado en la Parroquia de Santa Rosa
del cantón Salinas, Provincia del Guayas (Lat. 02º 11´ 10´´ S; Long. 80º 58´
11´´ W) (Fig. 10), al oeste de la provincia del Guayas a 144 Km. de Guayaquil y
a 5 Km. de Salinas es una comunidad que depende económicamente de la
actividad pesquera (Solís, 1998). En la actualidad, Santa Rosa, continúa siendo
uno de los principales puertos artesanales donde son desembarcados
importantes volúmenes de especies de alto valor comercial, las que son
destinadas para el mercado interno y de exportación (Revelo y Guzmán, 1997).
Así como por las oportunidades de trabajo y de ingresos económicos que se
generan mensualmente en este lugar (Solís, 1998).
El clima es determinado por la corriente fría de Humboldt y la cálida de El
Niño, su clima es tropical, la temperatura promedio es de 250 C (Solís, 1998).
Fig. 10. Ubicación Geográfica del Puerto de Santa Rosa, en la provincia del Guayas (Solís,
1998).
La población pesquera está conformada por los pescadores artesanales
activos, los comerciantes, los evisceradores y ayudantes. Se contabilizó entre
600 y 1 000 pescadores y 180 comerciantes (Solís, 1998).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
27
La actividad pesquera está dirigida a la extracción de varios recursos, entre
ellos los tiburones. Las principales familias son: Carcharhinidae, Squatinidae,
Triakidae, Sphyrnidae y Alopiidae (Solís, 1998).
Los artes de pesca empleados son la red de enmalle superficial, palangre
superficial, espinel de fondo, línea de mano de media agua y de fondo. Se
registró una flota pesquera de 608 embarcaciones, de las cuales
mayoritariamente se emplean la fibra de vidrio con motor fuera de borda (Solís,
1998).
Los caladeros (lugares) de pesca se encuentran ubicados en el Golfo de
Guayaquil y principalmente en la Isla Santa Clara. El tiempo que se requiere
para llegar a los caladeros depende del recurso que va a ser explotado. Para la
captura de los tiburones está ligada a la pesca de peces pelágicos grandes
como los atunes y picudos, ya que son capturados por los mismos artes de
pesca; por consiguiente, se requiere entre una y seis horas de recorrido para
llegar a las zonas de pesca. Con respecto a las salidas de pesca, por lo
general, tienen como duración uno y tres días (Solís, 1998).
Las faenas de pesca para la captura de los diferentes recursos son realizadas
en diversas épocas del año, dependiendo de los cambios estacionales, de la
abundancia y de la disponibilidad de los recursos hidrobiológicos (Solís, 1998).
En cuanto a los costos de operación, se determinó que el combustible
constituye el mayor rubro sobre los costos de operación por salida de pesca
entre el 75% y el 97%. Entre los limitantes que afectan a la actividad pesquera
está la falta de organización de los pescadores, falta de regulaciones que
controlen los precios de los recursos desembarcados, los elevados costos de
operación, la falta de seguridad en las faenas de pesca, etc. (Solís, 1998).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
28
8. METODOLOGIA
8.1. ACTIVIDADES DE MUESTREO (FASE DE MUESTREO)
Los muestreos se realizaron de mayo a diciembre del 2004, en el Puerto
Pesquero de Santa Rosa de Salinas, donde se tomaron muestras dos veces
por semana, durante los fines de semana.
Los datos se registraron en tablas de campos (Anexo 5) donde se hizo constar
la longitud total del tiburón (LT), cuya medida se tomó desde el extremo de la
cabeza hasta el final del lóbulo superior de la aleta caudal. De igual forma la
longitud precaudal (LP), desde el extremo de la cabeza hasta la foseta
precaudal (Fig. 11). Para el sexo en los machos se tomó la medida de los
gonopterigios desde su base al extremo distal del mismo (LG). Se observó el
grado de calcificación, rotación, abertura del rifiodón y presencia de semen. En
el caso de las hembras se determinó si existían marcas de copulación en el
cuerpo (MC); así mismo se observó si presentaban la abertura cloacal abierta o
cerrada, para conocer si éstas ya habían copulado o no. Para las hembras
grávidas (en gestación), se registró el número de embriones, identificando el
sexo y talla de cada uno. Se registró también la localidad de colecta, método
de colecta, y tipo de carnada utilizada (Anexo 5).
Fig. 11. Principales términos empleados en el muestreo (Aleta escapular = Aleta pectoral)
(Montero, 2000).
Para la extracción del estómago primero se realizó un corte ventral al tiburón
que va desde la cloaca hasta el centro de las aletas pectorales (realizado por

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
29
pescadores). Seguido, se cortó el estómago desde la región posterior del
esófago hasta la región anterior del intestino, para separar el estómago, al cual
se le calculó la proporción de llenado, tomando valores de Laevastu (1971):
0 = vacío; 1 = 25% lleno; 2 = 50% lleno; 3 = 75% lleno; y 4 = 100% lleno
(Anexo 5). Seguido, el contenido estomacal se colocó en un cedazo y se
procedió a lavarlo con agua de mar para eliminar los jugos gástricos y por
último se almacenó el contenido en bolsas de plástico herméticas marca Ziploc,
las cuales eran puestas en hielo para su traslado y posterior congelamiento,
previo a su revisión en el Laboratorio de Ecología Marina de la Facultad de
Ciencias Naturales de la Universidad de Guayaquil. En caso contrario, el
contenido estomacal se fija con formol (10%) para su posterior revisión (Galván
et al., 1989).
8.2. ACTIVIDADES DE LABORATORIO (FASE DE LABORATORIO)
El contenido estomacal de cada organismo fue colocado en bandejas y se
procedió a separar las presas por grupos taxonómicos para medir y pesar cada
uno de ellos, preservándolos en alcohol al 70% para su identificación hasta el
nivel más bajo posible que permita el grado de digestión en que se encuentren
los diversos organismos según la escala siguiente (Galván et al., 1989):
Estado #1: Individuos que presentan todas las características morfológicas
completas que lo hacen fácilmente identificable.
Estado #2: Individuos sin piel, sin ojos y músculos al descubierto
Estado #3: Individuos sin cabeza algunas partes del cuerpo presente y
esqueleto axial.
Estado #4: Presencia únicamente de otolitos y picos de calamar.
Para los peces, la determinación taxonómica se realizó por medio del esqueleto
axial y otolitos. Para el conteo de vértebras se utilizó trabajos de Clothier (1950),
Clothier y Baxter (1969) y, para los otolitos la clave de García - Godos (2001),
mientras que para aquellos peces que presentaron un estado de digestión

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
30
mínima se empleó las claves de Chirichigno (1980) y Fischer, Krupp, Schneider,
Sommer, Carpenter y Niem, (1995).
En los cefalópodos debido a la rápida digestión de las partes blandas de su
cuerpo la única estructura reconocible es el aparato mandibular comúnmente
conocido como "pico", que por estar compuesta de quitina, resiste más a la
digestión, lo cual permite su identificación, por lo que para este grupo en
particular se utilizó las claves para identificación de “picos” de cefalópodos de
Clarke (1962 y 1986), Wolff (1982 y 1984); Ingrid, Iverson y Pinkas (1971).
8.3. ACTIVIDADES DE GABINETE (FASE DE GABINETE)
8.3.1. Características morfométricas
Los datos morfométricos tomados durante la fase de campo y de laboratorio de
cada individuo se organizaron en una hoja de Excel con el fin de crear una
base de datos para un manejo óptimo de la información.
8.3.2. Relaciones tróficas
Se procedió a separar los organismos según tres aspectos, para la posterior
aplicación de los Índices Tróficos y Ecológicos, con el fin de obtener mejores
comparaciones. Estos aspectos fueron los siguientes:
a) Sexo
a.1) Hembras
a.2) Machos
b) Madurez Sexual y Tallas
b.1) Hembras Sexualmente Maduras de 258 - 365 cm de Longitud Total (LT).
b.2) Hembras Sexualmente Inmaduras de 152 – 257cm LT.
b.3) Machos Sexualmente Maduros de 244 – 310 cm. LT.
b.4) Machos Sexualmente Inmaduros de 176 – 243 cm. LT.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
31
8.3.2.1.Índices Tróficos
Para establecer el número de estómagos representativos para caracterizar el
espectro trófico de A. pelagicus se utilizó la metodología propuesta por
Hoffman 1978 (In: Bocanegra, Abitia y Galván, 2000) la cual consiste en
graficar en el eje horizontal el número de estómagos analizados contra número
acumulado de especies presa encontradas en dichos estómagos.
8.3.2.1.1. Método Numérico
El porcentaje en número consiste en separar los componentes de cada
muestra de estómago, en categorías de alimento, compilando el número total
de individuos de cada una y expresándolo como un porcentaje del número total
de individuos por categoría de alimento . Se calcula mediante la siguiente
fórmula (Hyslop, 1980):
%N = (Ni/NT)*100
Donde:
%N = Porcentaje en número
Ni = Número total de individuos de cada categoría o especie
NT = Número total de individuos de todas las especies
8.3.2.1.2. Método de Frecuencia de Ocurrencia
Determina de acuerdo con Peláez (1997), refleja la ocurrencia con la que se
encuentra cada componente alimenticio, el cual es expresado como porcentaje
de la totalidad de estómagos con alimento. Se compila el número total de
tractos (estómagos) que contuvieron a cada componente y se expresa como un
porcentaje del total de tractos con alimento. Se calcula mediante la siguiente
fórmula Peláez (1997):
Fa = (Na / NT) * 100
Donde:

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
32
Fa = frecuencia de ocurrencia
Na = número de tractos que tienen el mismo componente
NT = número de tractos con alimento.
8.3.2.1.3. Método Gravimétrico
Se toma en cuenta el peso de los contenidos estomacales, se suman los pesos
de cada componente alimenticio de los diferentes estómagos, el cual se
expresa como el porcentaje de la suma total de todos los componentes
alimenticios, para todos los estómagos. Se calcula mediante la siguiente
fórmula Peláez (1997):
%P = (Pi/PT)*100
Donde:
%P = Porcentaje en peso
Pi = Peso de una especie presa en las muestras
PT = Peso total de las diferentes presas encontradas en las muestras
8.3.2.1.4. Índice de Importancia Relativa (IIR)
Este índice propuesto por Pinkas, Oliphant e Ivirson (1971) es una herramienta
muy utilizada en estudios de alimentación ya que tiene la capacidad de mostrar
los componentes de mayor importancia dentro del espectro trófico, indicando
cuales alimentos son preferenciales, secundarios y circunstanciales, según la
escala propuesta por Duarte y Von Schiller (1997):
De 0 a 20: Presas circunstanciales (PC)
De 21 a 200: Presas secundarias (PS)
De 201 a 20 000: Presas principales (PP)
Permite confrontar las diferencias interespecíficas en un mismo grupo
taxonómico ya que incorpora la biomasa, cantidad y aparición de cada presa en
una sola medición (Peláez, 1997):

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
33
IIR = (%P + % N) * %FO
Donde:
IIR = Índice de Importancia Relativa
%P = Porcentaje en peso
%N = Porcentaje en número
%FO= Porcentaje de frecuencia de ocurrencia
8.3.2.2. Índices Ecológicos
8.3.2.2.1. Diversidad de presas
Se utilizó el Índice de Shannon y Weaver (1963). Este índice presenta un
intervalo de valores que va de 0 a 6, valores menores a 3 indican una dieta
poco diversa, que es dominada por pocas especies, en cambio valores
mayores a 3, indican una dieta dominada por varias especies (Vandermeer,
1981 In: Bocanegra et al., 2000). Se calculó con la siguiente fórmula:
H’ = - (∑ Pj ln Pj)
Donde:
H´= Índice de Shannon - Weaver
S = Número total de especies presas identificadas
Pj = Número de i especies, expresadas como una proporción de la suma de Pj
por todas las especies presas
Se utilizó logaritmo natural (ln) para la obtención de la diversidad, de ahí que
sus valores están en unidades Belnats.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
34
8.3.2.2.2. Uniformidad de presas
De igual manera se empleó el Índice de Equitabilidad de Pielou (1966) el cual
mostró si hubo uniformidad entre las especies identificadas o se presentó
dominancia de una o varias especies. Presenta una escala de 0 a 1, valores
menores a 0.7 muestran dominancia y valores mayores a 0.7 determinan
uniformidad. Se calculó con la siguiente fórmula:
.´´
máxHH
E =
Donde:
E = Índice de equitabilidad;
H’ = Índice de Shannon-Weaver y
H’ máx. = Diversidad máxima posible.
8.3.2.2.3. Amplitud del nicho trófico
Así mismo se calculó la amplitud del nicho trófico utilizando el Índice de Levin,
el cual propone que esta puede ser estimada por medio de la cuantificación de
la distribución de los depredadores con respecto a sus presas (Krebs, 1985)
de acuerdo a la técnica propuesta por Labropoulou y Eleftheriou (1997):
Bi = 1/n-1{(1/ΣJP2ij)-1}
Donde:
Bi= Índice de Levin para el predador J.
JPij = proporción de la presa j en la dieta del depredador i, y
n = número de categorías o especies presas.
Los valores de este índice están comprendidos de 0 a 1, valores menores a 0,6
indican que la dieta está dominada por pocas presas, por lo tanto sería un

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
35
depredador especialista y valores mayores a 0.6 indican dietas de
depredadores generalistas.
8.3.2.2.4. Superposición de dieta o traslapamiento trófico
Se realizó análisis de traslapamiento trófico entre sexos, madurez sexual y
tallas con el fin de comparar probables diferencias en el tipo de alimento de
éstos. Se utilizó el índice de Morisita y Horn (Morisita, 1959; Horn 1966; Smith y
Zaret, 1982):
Cλ= 2 Σ (Pxi * Pyi) / (Σ Pxi 2 + Σ Pyi 2 )
Donde:
Cλ = Índice de Morisita y Horn entre sexo o talla de x, y entre sexo o talla de y; Pxi = Proporción de presa i del total de presas usados por el sexo o talla del
depredador x;
Pyi = Proporción de presa i del total de presas usados por el sexo o talla del
depredador y,
n = Número total de presas.
Los valores de este índice van de 0 a 1 por lo tanto si llega a 0, significa que no
hay elementos en común, valores mayores a 0,6 indican traslapamiento o
similitud y cuando los valores llegan a 1 todos los elementos se encuentran
iguales en la dieta de ambos depredadores indicando un traslapamiento total.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
36
9. RESULTADOS
9. 1. NÚMERO DE INDIVIDUOS
Se muestrearon 110 individuos de A. pelagicus, durante los fines de semana
de mayo a diciembre del 2004 en Santa Rosa de Salinas, de los cuales 74 eran
hembras (67%) y 36 machos (33%) (Fig. 12). Se observó una gran variación
mensual, ya que el mes de diciembre presentó el mayor número con 39
individuos (35%), seguido de mayo con 36 (33%) y octubre 22 (20%). Durante
los meses de junio y julio no se registraron individuos (Fig. 13) (Anexo 5).
Número de Individuos por Sexo
36
74
MachosHembras
(a)
Porcentajes de Individuos por Sexo
33%
67%
MachosHembras
(b) Fig. 12. Número de Individuos por sexo de A. pelagicus en Santa Rosa de Salinas durante
mayo – diciembre del 2004, (a) en número y (b) en porcentajes.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
37
No. de Individuos
36
0 05
2
22
6
39
0
10
20
30
40
50
May Jun Jul Ago Sep Oct Nov Dic
Meses
Núm
ero
(a)
Porcentajes Mensuales de Individuos
33
0 05
2
20
5
35
05
10152025303540
May Jun Jul Ago Sep Oct Nov Dic
Meses
Po
rcen
taje
s
(b) Fig. 13. Variación mensual de individuos de A. pelagicus en Santa Rosa de Salinas durante
mayo – diciembre del 2004, (a) en número y (b) en porcentajes.
9. 2. NÚMERO DE ESTÓMAGOS
El número de estómagos analizados fue suficiente para caracterizar el espectro
trófico de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004, de acuerdo a la curva acumulativa de especies presas, ya que ésta
alcanzó un punto de estabilización a los 74 estómagos (67,3%). Además según
Galván (Com. pers., 2004) afirma que el número de estómagos necesario para
un estudio trófico de tiburones debe estar entre 80 y 90 muestras (Baigorrí y
Polo, 2004) (Fig. 14).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
38
Al establecer el número de individuos muestreados se determina lógicamente
el número de estómagos, por ende, fueron 110. En relación, al número de
estómagos (tractos) con alimento, el mes de diciembre presentó el mayor
número con 30 tractos con alimento y 9 vacíos, seguido de mayo con 23 y 13
vacíos y octubre 19 y 3. Durante los meses de junio y julio no se registraron
individuos (Fig. 15).
Curva Acumulativa de Especies Presas
0
5
10
15
20
25
0 20 40 60 80 100 120
No. Estómagos Analizados
No
. Esp
ecie
s P
resa
s
Fig. 14. Curva Acumulativa de Especies presas de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004.
No. de Estómagos (Con Alimentos y Vacíos)
23
0 03
0
19
5
30
0 02 2 3
1
9
13
0
5
10
15
20
2530
35
May Jun Jul Ago Sep Oct Nov Dic
Meses
Nú
mer
o
Estóm. con alimentosEstóm. vacíos
Fig. 15. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004.
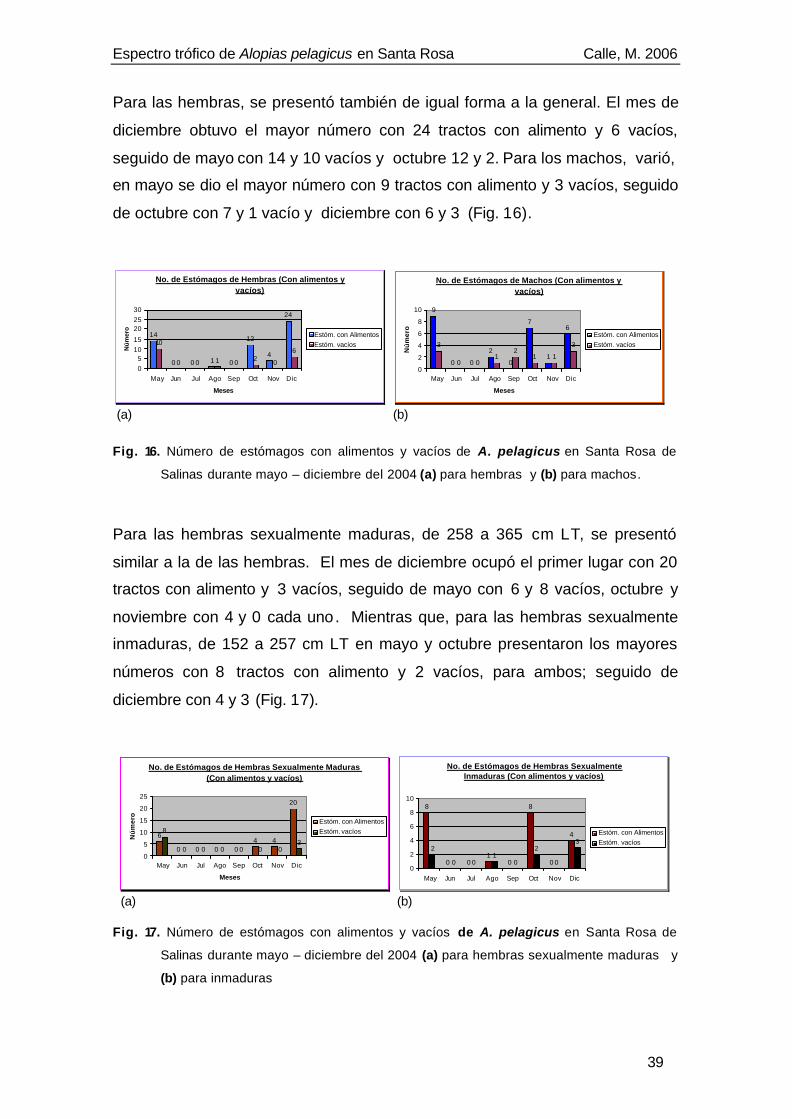
Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
39
Para las hembras, se presentó también de igual forma a la general. El mes de
diciembre obtuvo el mayor número con 24 tractos con alimento y 6 vacíos,
seguido de mayo con 14 y 10 vacíos y octubre 12 y 2. Para los machos, varió,
en mayo se dio el mayor número con 9 tractos con alimento y 3 vacíos, seguido
de octubre con 7 y 1 vacío y diciembre con 6 y 3 (Fig. 16).
No. de Estómagos de Hembras (Con alimentos y vacíos)
14
0 0 1 0
12
4
24
10
0 0 1 0 2 0
6
05
1015
202530
May Jun Jul Ago Sep Oct Nov Dic
Meses
Núm
ero
Estóm. con AlimentosEstóm. vacíos
No. de Estómagos de Machos (Con alimentos y vacíos)
9
0 0
2
0
7
1
6
3
0 01
21 1
3
0
2
4
6
8
10
May Jun Jul Ago Sep Oct Nov Dic
Meses
Nú
mer
o
Estóm. con AlimentosEstóm. vacíos
(a) (b)
Fig. 16. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para hembras y (b) para machos.
Para las hembras sexualmente maduras, de 258 a 365 cm LT, se presentó
similar a la de las hembras. El mes de diciembre ocupó el primer lugar con 20
tractos con alimento y 3 vacíos, seguido de mayo con 6 y 8 vacíos, octubre y
noviembre con 4 y 0 cada uno. Mientras que, para las hembras sexualmente
inmaduras, de 152 a 257 cm LT en mayo y octubre presentaron los mayores
números con 8 tractos con alimento y 2 vacíos, para ambos; seguido de
diciembre con 4 y 3 (Fig. 17).
No. de Estómagos de Hembras Sexualmente Maduras (Con alimentos y vacíos)
6
0 0 0 04 4
20
8
0 0 0 0 0 03
0
5
10
15
20
25
May Jun Jul Ago Sep Oct Nov Dic
Meses
Nú
mer
o
Estóm. con AlimentosEstóm. vacíos
No. de Estómagos de Hembras Sexualmente Inmaduras (Con alimentos y vacíos)
8
0 01
0
8
0
4
2
0 01
0
2
0
3
0
2
4
6
8
10
May Jun Jul Ago Sep Oct Nov Dic
Estóm. con AlimentosEstóm. vacíos
(a) (b)
Fig. 17. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para hembras sexualmente maduras y
(b) para inmaduras

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
40
Para los machos sexualmente maduros, de 244 a 310 cm LT, mayo ocupó el
primer lugar con 7 tractos con alimento y 2 vacíos, seguido de diciembre con 5
y 2 vacíos. Mientras que, para los machos sexualmente inmaduros, de 176 a
243 cm. LT, octubre presentó 5 tractos con alimento y 1 vacíos, seguido de
mayo y mayo y agosto con 2 y 1 ambos (Fig. 18).
No. de Estómagos de Machos Sexualmente Maduros (Con alimentos y vacíos)
7
0 0 0 0
21
5
2
0 0 0 0 01
2
0
2
4
6
8
May Jun Jul Ago Sep Oct Nov Dic
Meses
Nú
mer
o
Estóm. con AlimentosEstóm. vacíos
No. de Estómagos de Machos Sexualmente Inmaduros (Con alimentos y vacíos)
2
0 0
2
0
5
011
0 01
21
01
0123456
May Jun Jul Ago Sep Oct Nov Dic
MesesN
úmer
o
Estóm. con Alimentos
Estóm. vacíos
(a) (b)
Fig. 18. Número de estómagos con alimentos y vacíos de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004 (a) para machos sexualmente maduros y
(b) para inmaduros
9. 3. PORCENTAJES DE LLENADO
Una gran parte de los estómagos muestreados se encontraban vacíos (0)
50,9%, seguido de los estómagos con un 25% lleno (1) 21,82%; luego, con un
50% lleno (2) el 20%; con un 100% lleno (4) 5,45% y finalmente, con un 75%
lleno (3) 1,82% (Fig. 19a).
La variación mensual determinó para diciembre los mayores porcentajes tanto
para el 0% (vacío) con 36% y como para el 25% de llenado con el 50%, mayo
para el 50% de llenado con el 45%, octubre y diciembre para el 75% de llenado
con el 50%. Finalmente, octubre obtuvo para el 100% de llenado el 50% (Fig.
19b).
Para las hembras, diciembre presentó el mayor porcentaje para el 0% de
llenado con el 41% y también obtuvo para el 25% de llenado, el 59%; seguido
de mayo con el 50% para el 50% de llenado; no se registró ningún organismo
con el 75% de llenado y para el 100% de llenado, octubre con el 100% (Fig.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
41
19c). Para los machos, mayo encabezó los porcentajes para el 0% de llenado
con el 29%; de igual manera, mayo para el 25% de llenado con el 57%; octubre
para el 50% de llenado con el 50%; octubre y diciembre para el 75% de llenado
con el 50% ambos y, por último mayo, octubre, noviembre y diciembre con el
25% cada uno para el 100% de llenado (Fig. 19d).
Porcentajes de Llenado
50,9
21,82 20
1,82 5,45
0102030405060
Vacío 25% Lleno 50% Lleno 75% Lleno 100% Lleno
Escalas de llenado
Por
cent
ajes
Porcentajes de Llenado
0
10
20
30
40
50
60
May Jun Jul Ago Sep Oct Nov Dic
MesesP
orc
enta
jes 0 (0%)
1 (25%)2 (50%)3 (75%)
4 (100%)
(a) (b)
Porcentajes de Llenado para Hembras
0
20
40
60
80
100
120
May Jun Jul Ago Sep Oct Nov Dic
Meses
Po
rcen
taje
s 0 (0%)1 (25%)2 (50%)
3 (75%)4 (100%)
Porcentajes de Llenado para Machos
0
10
20
30
40
50
60
May Jun Jul Ago Sep Oct Nov Dic
Meses
Po
rcen
taje
s 0 (25%)1 (25%)2 (50%)3 (75%)4 (100%)
(c) (d) Fig. 19. Porcentajes de llenado estomacales de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004 (a) para organismos en general, (b) organismos en
general por meses (c) hembras por meses y (d) machos por meses.
9.4. ÍNDICES TRÓFICOS
9.4.1. General
La lista taxonómica de especies presas consumidas por A. pelagicus se
detalla a continuación. Dicha lista presenta entre paréntesis () el nombre del
autor de la especie y entre corchetes [ ] códigos con los que se representa a la
especie en los gráficos subsecuentes.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
42
Phylum Mollusca (según Markaida y Hochberg, documento en preparación)
Clase Gasteropoda
Clase Cephalopoda
Orden Teuthoidea
Familia Ommastrephidae
Dosidicus gigas (Orbigny, 1835) [D. gigas]
Sthenoteuthis oualaniensis (Lesson, 1830) [S. ouala]
Ommastrephes bartramii (LeSueur, 1821) [O. bartra]
Familia Octopoteuthidae
Octopoteuthis sicula (Grimpe, 1922) [O. sicu]
Familia Enoploteuthidae
Abraliopsis affinis (Pfeffer, 1912) [A. affin]
Ancistrocheirus lessuerii (Orbigny, 1839) [A. lessu]
Familia Mastigoteuthidae
Mastigoteuthis dentata (Hoyle, 1904) [M. denta]
Familia Loliginidae
Loliolopsis diomedeae (Hoyle, 1904) [L. diome]
Familia Histiotheuthidae
Histioteuthis sp. [Hist sp.]
Patrón A (Morfotipo A) [Patr A]
Patrón B (Morfotipo B) [Patr B]
Patrón C (Morfotipo C) [Patr C]
Phylum Chordata (según Nelson 1994)
Clase Choncrichthyes
Orden Carcharhiniformes
Familia Triakiidae
Mustelus sp. [Must sp.]
Clase Osteichthyes
Orden Anguiliformes
Familia Ophichthidae
Ophichthus sp. [Ophi sp.]

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
43
Orden Myctophiformes
Familia Myctophidae
Benthosema panamense (Taning, 1932) [B. panam]
Orden Gadiformes
Familia Merlucciidae
Merluccius gayi (Guichenot, 1848) [M. gayi]
Orden Perciformes
Familia Coryphaenidae
Coryphaena hippurus (Linnaeus, 1758) [C. hippu]
Familia Scianidae
Larimus sp. [Lari sp.]
Familia Scombridae
Thunnus sp. [Thun sp.]
Patrón 1 (Morfotipo 1) [Patr 1]
Patrón 2 (Morfotipo 2) [Patr 2]
Patrón 3 (Morfotipo 3) [Patr 3]
Se determinó 928 organismos consumidos por A. pelagicus durante mayo –
diciembre del 2004, con un peso de 86 977,88 g, pertenecientes a 24 especies
presas agrupadas en cuatro clases: Osteictios con 623 organismos (74,68%),
Cefalópodos con 224 (24,14%), Gasterópodos con 8 (0,86%) y Condrictios con
3 (0,32%). De acuerdo al Método Numérico se estableció que la Merluza del
Pacífico Merluccius gayi presentó el mayor número con 393 organismos
(42,35%), seguido de la Sardina luminosa Benthosema panamense con 288
individuos (31,03%) y del calamar Sthenoteuthis oualaniensis con 77 (8,30%)
(Fig. 20). El Método de Frecuencia determinó la mayor ocurrencia para M. gayi
y B. panamense con 28 veces (35%) para ambos, seguido del calamar
Ommastrephes bartramii con 24 ocasiones (30%) (Fig. 21). El Método
gravimétrico determinó que S. oualaniensis presentó el primer lugar en cuanto
a peso con 55 464,88 g (63,77%), seguido del calamar gigante Dosidicus
gigas con 15 916,89 g (18,30%) y luego, O. bartramii con 9 675,20 g

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
44
(11,12%) (Fig. 22). Finalmente, el Índice de Importancia Relativa (IIR)
estableció como presas principales a S. oualaniensis con 2 071, 91; M. gayi
con 1 569,02 y B. panamense con 1 086,97 (Fig. 23) (Tabla 1).
Método Numérico (General)
4,74
8,305,39
0,75 0,11 0,11 0,97 0,11 0,11 0,11 0,11 0,11
42,35
31,03
0,65 0,22 0,32 0,11 0,11 0,11 0,11 0,86
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
M. d
enta
O. b
anks
A.
affin
Pat
r A
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp
Mus
t sp
.
Thun
sp.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Po
rcen
taje
s
Fig. 20. Valores porcentuales del Método Numérico de A. pelagicus en Santa Rosa de Salinas
durante mayo – diciembre del 2004.
Método de Frecuencia (General)
2 0 , 0 0
28,753 0 , 0 0
6 , 2 5
1,25 1,25
5 , 0 0
1,25 1,25 1,25 1,25 1,25
3 5 , 0 0 3 5 , 0 0
5 , 0 0
2,501,25 1,25 1,25 1,25 1,25 1,25
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
D. g
igas
S. o
uala
O.
bartr
a
L. d
iom
e
His
t sp.
O. s
icu
A. l
essu
M. d
enta
O. b
anks
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Lari
sp.
Oph
i sp
Mus
t sp
.
Thun
sp.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 21. Valores porcentuales del Método de Frecuencia de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
45
Método Gravimétrico (General)
18,30
63,77
11,12
2 , 7 00,00 0,00 0,00 0,74 0,00 0,00 0 , 0 0 0,00
2,480,02 0,00 0,01 0,07 0,01 0,01 0 , 0 0 0,00 0,00
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A. l
essu
M.
dent
a
O.
bank
s
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp
Mus
t sp.
Thu
n sp
.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Po
rcen
taje
s
Fig. 22. Valores porcentuales del Método Gravimétrico de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004.
Índice de Importancia Relativa (General)
460,83
2071,91
495,35
21,58 0,14 0,13 4 , 8 5 1,05 0,14 0,13 0,13 0,13
1569,02
1086,97
3 , 2 4 0,55 0,49 0,15 0,14 0,13 0,13 1,080,00
500,00
1000,00
1500,00
2000,00
2500,00
D. g
igas
S. o
uala
O. b
artr
a
L. d
iom
e
His
t sp.
O. s
icu
A. l
essu
M. d
enta
O. b
anks
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M.
gayi
B.
pana
m
Lari
sp.
Oph
i sp
Mus
t sp.
Thu
n sp
.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 23. Valores del Índice de Importancia Relativa (IIR) de A. pelagicus en Santa Rosa de
Salinas durante mayo – diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
46
Tabla 1. Espectro trófico de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004 expresado en valores absolutos y porcentuales del método numérico (N y
%N), frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de
importancia relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa
Principal, S: Secundaria y C: Circunstancial.
Especies N % N FO % FO P % P IIR Categ D. gigas 44 4,74 16 20,00 15916,89 18,30 460,83 P S. oualaniensis 77 8,30 23 28,75 55464,88 63,77 2071,91 P O. bartramii 50 5,39 24 30,00 9675,20 11,12 495,35 P L. diomedeae 30 3,23 9 11,25 672,85 0,77 45,07 S Histioteuthis sp. 7 0,75 5 6,25 2347,53 2,70 21,58 S O. sicula 1 0,11 1 1,25 0,52 0,00 0,14 C A. lessuerii 1 0,11 1 1,25 0,00 0,00 0,13 C M. dentata 9 0,97 4 5,00 0,09 0,00 4,85 C O. banksii 1 0,11 1 1,25 639,45 0,74 1,05 C A. affinis 1 0,11 1 1,25 3,14 0,00 0,14 C Patrón A 1 0,11 1 1,25 0,00 0,00 0,13 C Patrón B 1 0,11 1 1,25 0,00 0,00 0,13 C Patrón C 1 0,11 1 1,25 0,00 0,00 0,13 C M. gayi 393 42,35 28 35,00 2156,94 2,48 1569,02 P B. panamense 288 31,03 28 35,00 18,89 0,02 1086,97 P Larimus sp. 6 0,65 4 5,00 1,35 0,00 3,24 C Ophichthus sp. 2 0,22 2 2,50 5,10 0,01 0,55 C Mustelus sp. 3 0,32 1 1,25 59,80 0,07 0,49 C Thunnus sp. 1 0,11 1 1,25 10,00 0,01 0,15 C C. hippurus 1 0,11 1 1,25 5,00 0,01 0,14 C Patrón 1 1 0,11 1 1,25 0,10 0,00 0,13 C Patrón 2 1 0,11 1 1,25 0,10 0,00 0,13 C Gasterópodos 8 0,86 1 1,25 0,05 0,00 1,08 C Suma 928 100,00 156 195 86977,88 100,00
9.4.2. Por Sexo
9.4.2.1. Hembras
Para las hembras el Método Numérico determinó el mayor porcentaje para
M. gayi con 319 organismos lo que representa el 51,95%; seguido, de
B. panamense con 146 (23,78%) y S. oualaniensis con 41 (6,68%) (Fig. 24).
Mientras, el Método de Frecuencia, estableció similares resultados al anterior,
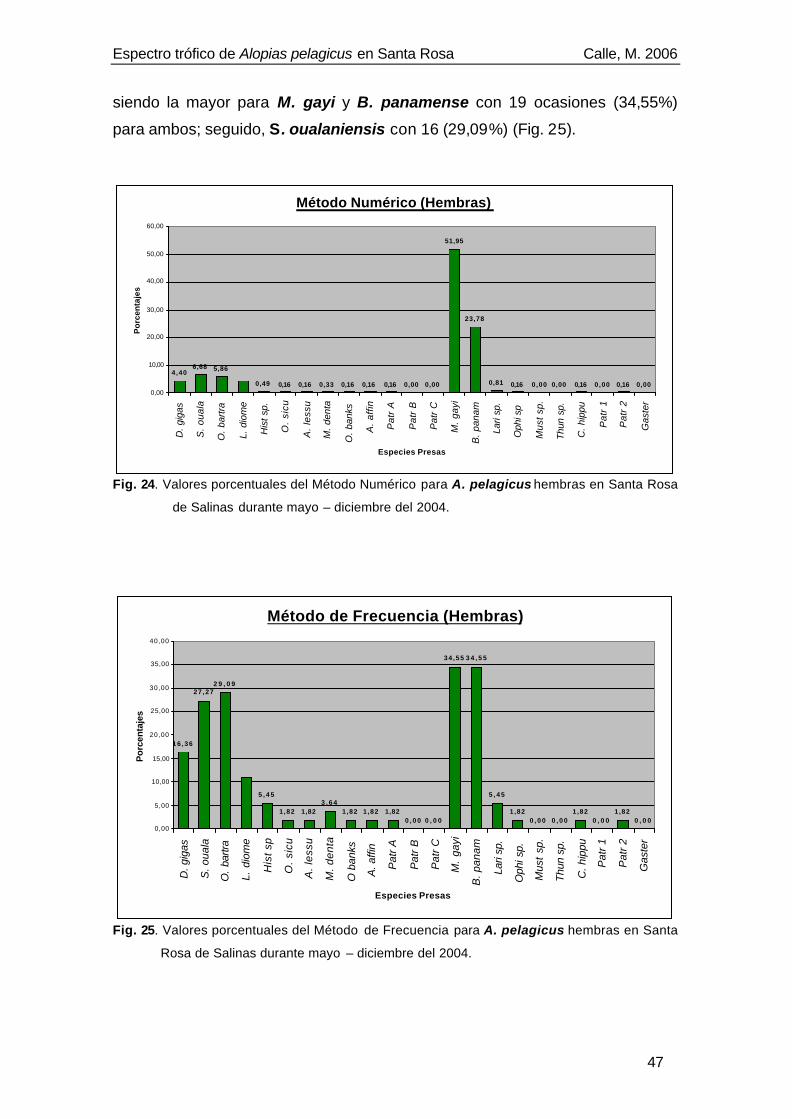
Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
47
siendo la mayor para M. gayi y B. panamense con 19 ocasiones (34,55%)
para ambos; seguido, S. oualaniensis con 16 (29,09%) (Fig. 25).
Método Numérico (Hembras)
4,406,68 5,86
0,49 0,16 0,16 0,33 0,16 0,16 0,16 0,00 0,00
51,95
23,78
0,81 0,16 0,00 0,00 0,16 0,00 0,16 0,000,00
10,00
20,00
30,00
40,00
50,00
60,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
M. d
enta
O. b
anks
A.
affin
Pat
r A
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp
Mus
t sp
.
Thun
sp.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Po
rcen
taje
s
Fig. 24. Valores porcentuales del Método Numérico para A. pelagicus hembras en Santa Rosa
de Salinas durante mayo – diciembre del 2004.
Método de Frecuencia (Hembras)
16 ,36
27 ,272 9 , 0 9
5 , 4 5
1,82 1,823 , 6 4
1,82 1,82 1,820 , 0 0 0 , 0 0
34 ,55 3 4 , 5 5
5 , 4 5
1,820 , 0 0 0 , 0 0
1,820 , 0 0
1,820 , 0 0
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
D. g
igas
S. o
uala
O.
bartr
a
L. d
iom
e
His
t sp
O. s
icu
A. l
essu
M. d
enta
O b
anks
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Lari
sp.
Oph
i sp.
Mus
t sp
.
Thun
sp.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 25. Valores porcentuales del Método de Frecuencia para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004.
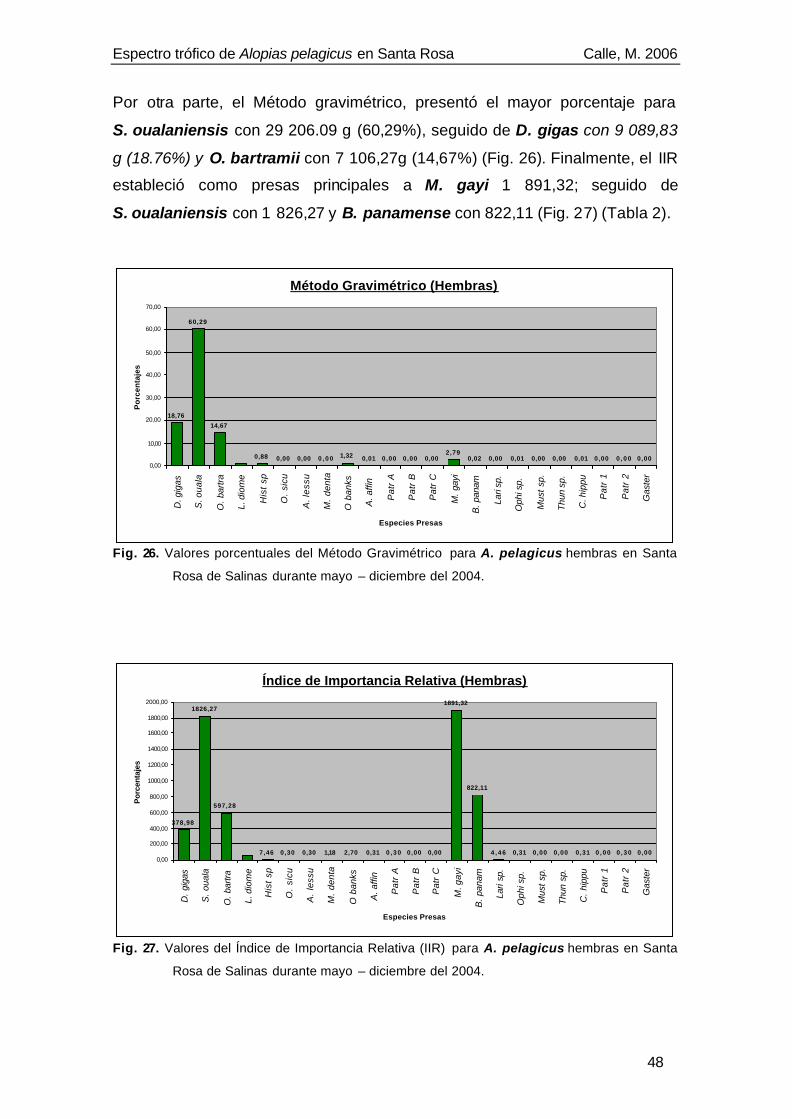
Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
48
Por otra parte, el Método gravimétrico, presentó el mayor porcentaje para
S. oualaniensis con 29 206.09 g (60,29%), seguido de D. gigas con 9 089,83
g (18.76%) y O. bartramii con 7 106,27g (14,67%) (Fig. 26). Finalmente, el IIR
estableció como presas principales a M. gayi 1 891,32; seguido de
S. oualaniensis con 1 826,27 y B. panamense con 822,11 (Fig. 27) (Tabla 2).
Método Gravimétrico (Hembras)
18,76
60,29
14,67
0,88 0,00 0,00 0 , 0 0 1,32 0,01 0,00 0,00 0,002,79
0,02 0,00 0,01 0,00 0,00 0,01 0,00 0 , 0 0 0,000,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp
O.
sicu
A. l
essu
M.
dent
a
O b
anks
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Mus
t sp.
Thu
n sp
.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Po
rcen
taje
s
Fig. 26. Valores porcentuales del Método Gravimétrico para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004.
Índice de Importancia Relativa (Hembras)
378,98
1826,27
597,28
7,46 0,30 0,30 1,18 2,70 0,31 0 , 3 0 0,00 0,00
1891,32
822,11
4 , 4 6 0,31 0,00 0,00 0,31 0 , 0 0 0 , 3 0 0,000,00
200,00
400,00
600,00
800,00
1000,00
1200,00
1400,00
1600,00
1800,00
2000,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp
O.
sicu
A.
less
u
M.
dent
a
O b
anks
A. a
ffin
Pat
r A
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Mus
t sp
.
Thun
sp.
C. h
ippu
Pat
r 1
Pat
r 2
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 27. Valores del Índice de Importancia Relativa (IIR) para A. pelagicus hembras en Santa
Rosa de Salinas durante mayo – diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
49
Tabla 2. Espectro trófico de A. pelagicus hembras en Santa Rosa de Salinas durante mayo –
diciembre del 2004 en Santa Rosa de Salinas expresado en valores absolutos y
porcentuales del método numérico (N y %N), frecuencia de ocurrencia (FO y %FO),
gravimétrico (P y %PO), índice de importancia relativa (IIR) y Categoría de la
especie presa (Categ), donde P: Presa Principal, S: Secundaria y C: Circunstancial.
Especies N % N FO % FO P % P IIR Categ D. gigas 27 4,40 9 16,36 9089,83 18,76 378,98 P S. oualaniensis 41 6,68 15 27,27 29206,09 60,29 1826,27 P O. bartramii 36 5,86 16 29,09 7106,27 14,67 597,28 P L. diomedeae 27 4,40 6 10,91 600,38 1,24 61,49 S Histioteuthis sp. 3 0,49 3 5,45 425,95 0,88 7,46 C O. sicula 1 0,16 1 1,82 0,52 0,00 0,30 C A. lessueri 1 0,16 1 1,82 0,00 0,00 0,30 C M. dentata 2 0,33 2 3,64 0,00 0,00 1,18 C O. banksii 1 0,16 1 1,82 639,45 1,32 2,70 C A. affinis 1 0,16 1 1,82 3,14 0,01 0,31 C Patrón A 1 0,16 1 1,82 0,00 0,00 0,30 C M. gayi 319 51,95 19 34,55 1353,74 2,79 1891,32 P B. panamense 146 23,78 19 34,55 9,38 0,02 822,11 P Larimus sp. 5 0,81 3 5,45 1,20 0,00 4,46 C Ophichthus sp. 1 0,16 1 1,82 5,00 0,01 0,31 C C. hippurus 1 0,16 1 1,82 5,00 0,01 0,31 C Patrón 2 1 0,16 1 1,82 0,10 0,00 0,30 C Suma 614 100,00 100 48446,04 100,00 9.4.2.1. Machos
Por otra parte, para los machos el Método Numérico, estableció el mayor
porcentaje para B. panamense con 142 especimenes (45,22%), seguido de
M. gayi con 74 (23,57%) y S. oualaniensis con 36 (11,46%) (Fig. 28).
Mientras, el Método Gravimétrico determinó mayor predomino de M. gayi y
B. panamense con 9 veces (36%) para ambos; seguido de S. oualaniensis y
O. bartramii con 8 (32%) (Fig. 29).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
50
Método Numérico (Machos)
5 , 4 1
11,46
4 , 4 6
1 , 2 7 2 , 2 30 , 3 2 0 , 3 2
23,57
4 5 , 2 2
0 , 3 2 0 , 3 2 0 , 9 6 0 , 3 2 0 , 3 22 , 5 5
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
50,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
M. d
enta
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Mus
t sp.
Thun
sp.
Pat
r 1
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 28. Valores porcentuales del Método Numérico para A. pelagicus machos en Santa Rosa
de Salinas durante mayo – diciembre del 2004.
Método de Frecuencia (Machos)
2 8
3 2 3 2
8 8
4 4
3 6 3 6
4 4 4 4 4 4
0
5
10
15
2 0
25
3 0
35
4 0
D. g
igas
S. o
uala
O.
bartr
a
L. d
iom
e
His
t sp.
M. d
enta
Pat
r B
Pat
r C
M.
gayi
B.
pana
m
Lari
sp.
Oph
i sp.
Mus
t sp.
Thu
n sp
.
Pat
r 1
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 29. Valores porcentuales del Método de Frecuencia para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
51
El método gravimétrico proporcionó mayor porcentaje para S. oualaniensis
con 26 258,29 g (68,15%), seguido de D. gigas con 6 827,06 (17,72%) y de
O. bartramii con 2 568,93 (6,67%) (Fig. 30). Luego, el IIR presentó
S. oualaniensis con 2 547,62; seguido de B. panamense con 1 628,91 y
M. gayi con 923,45 (Fig. 31) (Tabla 3).
Método Gravimétrico (Machos)
17,72
68,15
6 , 6 7 4 , 9 9
0 , 0 0 0 ,00 0 ,002 ,08
0 ,02 0 , 0 0 0 , 0 0 0,16 0 , 0 3 0 , 0 0 0 , 0 00,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
M. d
enta
Pat
r B
Pat
r C
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Mus
t sp
.
Thun
sp.
Pat
r 1
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 30. Valores porcentuales del Método Gravimétrico para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004.
Índice de Importancia Relativa (Machos)
6 4 7 , 7 0
2547,62
3 5 6 , 0 2
50 ,09 17,84 1,27 1,27
9 2 3 , 4 5
1 6 2 8 , 9 1
1,28 1,27 4 , 4 4 1,38 1,27 10,190,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
D. g
igas
S. o
uala
O. b
artr
a
L. d
iom
e
His
t sp.
M. d
enta
Pat
r B
Pat
r C
M.
gayi
B.
pana
m
Lari
sp.
Oph
i sp.
Mus
t sp
.
Thun
sp.
Pat
r 1
Gas
ter
Especies Presas
Po
rcen
taje
s
Fig. 31. Valores del Índice de Importancia Relativa (IIR) para A. pelagicus machos en Santa
Rosa de Salinas durante mayo – diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
52
Tabla 3. Espectro trófico de A. pelagicus machos en Santa Rosa de Salinas durante mayo –
diciembre del 2004 expresado en valores absolutos y porcentuales del método
numérico (N y %N), frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO),
índice de importancia relativa (IIR) y Categoría de la especie presa (Categ), donde
P: Presa Principal, S: Secundaria y C: Circunstancial.
Especies N % N FO % FO P % P IIR Categ
D. gigas 17 5,41 7 28 6827,06 17,72 647,70 P S. oualaniensis 36 11,46 8 32 26258,79 68,15 2547,62 P O. bartramii 14 4,46 8 32 2568,93 6,67 356,02 P L. diomedeae 3 0,96 3 12 72,47 0,19 13,72 C Histioteuthis sp. 4 1,27 2 8 1921,58 4,99 50,09 S M. dentata 7 2,23 2 8 0,09 0,00 17,84 C Patrón B 1 0,32 1 4 0,00 0,00 1,27 C Patrón C 1 0,32 1 4 0,00 0,00 1,27 C M. gayi 74 23,57 9 36 803,20 2,08 923,45 P B. panamense 142 45,22 9 36 9,51 0,02 1628,91 P Larimus sp. 1 0,32 1 4 0,15 0,00 1,28 C Ophichthus sp. 1 0,32 1 4 0,10 0,00 1,27 C Mustelus sp. 3 0,96 1 4 59,80 0,16 4,44 C Thunnus sp. 1 0,32 1 4 10,00 0,03 1,38 C Patrón 1 1 0,32 1 4 0,10 0,00 1,27 C Gasterópodos 8 2,55 1 4 0,05 0,00 10,19 C Suma 314 100,00 56 38531,84 100,00 9.4.3. Por Madurez Sexual y Tallas
9.4.3.1. Hembras Sexualmente Maduras
Para las Hembras Sexualmente Maduras de 258 - 365 cm. LT el Método
Numérico demostró que el mayor porcentaje fue para M. gayi con 307
especimenes (65,18%), seguido de B. panamense con 98 (20,81%), y
O. bartramii con 23 (4,88%) (Fig. 32). Luego, el Método de Frecuencia
determinó mayores porcentajes para M. gayi con 17 ocurrencias (50%),
seguido de B. panamense con 15 ocasiones (44,12%); S. oualaniensis y
O. bartramii ambas con 8 veces (23,53%) (Fig. 33).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
53
Método Numérico (Hembras Sexualmente Maduras)
4,03 3,40 4 , 8 8
0,42
65,18
20,81
0,21 0,210,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
D. gigas S. oual O. bartra L. diome M. denta M. gayi B. panam Lari sp. C. hippu
Especies Presas
Por
cent
ajes
Fig. 32. Valores porcentuales del Método Numérico para hembras sexualmente maduras de
258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Método de Frecuencia (Hembras Sexualmente Maduras)
14,71
23 ,53 23 ,53
5,88
50,00
44,12
2,94 2,94
0,00
10,00
20,00
30,00
40,00
50,00
60,00
D. gigas S. oual O. bartra L. diome M. denta M. gayi B. panam Lari sp. C. hippu
Especies Presas
Por
cent
ajes
Fig. 33. Valores porcentuales del Método de Frecuencia para hembras sexualmente maduras
de 258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Para el Método Gravimétrico, se estableció que el mayor porcentaje
correspondió para S. oualaniensis con 12 118,04 g (49,90%), seguido de
D. gigas con 6 859,21 g (28,25%) y O. bartramii 3 978,82 g (16,38%)
(Fig. 34). Finalmente, para el IIR, las presas principales fueron M. gayi con 3

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
54
509,63, seguido de S. oualaniensis con 1254.06 y B. panamense con
919.14 (Fig. 35) (Tabla 4).
Método Gravimétrico (Hembras Sexualmente Maduras)
28,25
49 ,90
16,38
0,00
5,01
0,03 0,00 0,020,00
10,00
20,00
30,00
40,00
50,00
60,00
D. gigas S. oual O. bartra L. diome M. denta M. gayi B. panam Lari sp. C. hippu
Especies Presas
Por
cent
ajes
Fig. 34. Valores porcentuales del Método Gravimétrico para hembras sexualmente maduras de
258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Índice de Importancia Relativa (Hembras Sexualmente Maduras)
474,70
1254,06
500,41
2 , 5 0
3509,63
919,14
0,63 0,690,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
3500,00
4000,00
D. gigas S. oual O. bartra L. diome M. denta M. gayi B. panam Lari sp. C. hippu
Especies Presas
Po
rcen
taje
s
Fig. 35. Valores del Índice de Importancia Relativa (IIR) para hembras sexualmente maduras
de 258 - 365 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
55
Tabla 4. Espectro trófico de hembras sexualmente maduras de 258 - 365 cm. LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal,
S: Secundaria y C: Circunstancial.
Especies N %N FO % FO P % P IIR Categ D. gigas 19 4,03 5 14,71 6859,21 28,25 474,70 P S. oualaniensis 16 3,40 8 23,53 12118,04 49,90 1254,06 P O. bartramii 23 4,88 8 23,53 3978,82 16,38 500,41 P L. diomedeae 4 0,85 4 11,76 99,47 0,41 14,81 C M. dentata 2 0,42 2 5,88 0,00 0,00 2,50 C M. gayi 307 65,18 17 50,00 1217,17 5,01 3509,63 P B. panamense 98 20,81 15 44,12 6,57 0,03 919,14 P Larimus sp. 1 0,21 1 2,94 0,15 0,00 0,63 C C. hippurus 1 0,21 1 2,94 5,00 0,02 0,69 C Suma 471 100,00 61 24284,43 100, 0 9.4.3.2. Hembras Sexualmente Inmaduras
Para las Hembras Sexualmente Inmaduras de 152 – 257 cm. LT el Método
Numérico determinó que el mayor porcentaje lo obtuvo B. panamense con 48
individuos (33,57%), seguido de S. oualaniensis con 25 organismos (17,48%)
y del calamar Loliolopsis diomedeae con 23 (16,08%) (Fig. 36). El Método de
Frecuencia determinó el mayor porcentaje para O. bartramii con 8 ocurrencias
(38,10%), seguido de S. oualaniensis con 7 ocasiones (33,33%),
B. panamense y D. gigas con 4 veces (19,05%) (Fig. 37).
El método gravimétrico presentó el mayor porcentaje para S. oualaniensis con
17 088,05 g (72,66%), seguido de O. bartramii con 3 127,44 g (13,30%) y
D. gigas con 2 230,62 g (9,48%) (Fig. 38). El IIR estableció como presas
principales a S. oualaniensis con 3 004,63, O. bartramii con 852,89 y
B. panamense con 639,59 (Fig. 39) (Tabla 5).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
56
Método Numérico (Hembras Sexualmente Inmaduras)
5,59
17,48
9 , 0 9
2,100 , 7 0 0,70 0,70 0 , 7 0 0 , 7 0
8 , 3 9
33 ,57
2 , 8 00,70 0 , 7 0
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
O. b
anks
A. a
ffin
Pat
r A
M. g
ayi
B.
pana
m
Lari
sp.
Oph
i sp.
Pat
r 2
Especies Presas
Por
cent
ajes
Fig. 36. Valores porcentuales del Método Numérico para hembras sexualmente inmaduras de
152 - 257 cm. LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Método de Frecuencia (Hembras Sexualmente Inmaduras)
1 9 , 0 5
33 ,33
38,10
1 4 , 2 9
4 , 7 6 4,76 4,76 4 , 7 6 4,76
9,52
19,05
9,52
4,76 4 , 7 6
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
O. b
anks
A. a
ffin
Pat
r A
M. g
ayi
B.
pana
m
Lari
sp.
Oph
i sp.
Pat
r 2
Especies Presas
Po
rcen
taje
s
Fig. 37. Valores porcentuales del Método de Frecuencia para hembras sexualmente inmaduras
de 152 - 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
57
Método Gravimétrico (Hembras Sexualmente Inmaduras)
9 , 4 8
72 ,66
13,30
1,810 , 0 0 0 , 0 0 0 , 0 0 0 , 0 0 0 , 0 0 0,58 0 , 0 1 0 , 0 0 0 , 0 2 0 , 0 0
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
O. b
anks
A. a
ffin
Pat
r A
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Pat
r 2
Especies Presas
Po
rcen
taje
s
Fig. 38. Valores porcentuales del Método Gravimétrico para hembras sexualmente inmaduras
de 152 - 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Índice de Importancia Relativa (Hembras Sexualmente Inmaduras)
2 8 7 , 2 1
3 0 0 4 , 6 3
852 ,89
55,84 3 , 3 4 3 , 3 3 3 , 3 3 3 , 3 3 3 , 3 385,45
6 3 9 , 5 9
2 6 , 6 8 3 , 4 3 3 , 3 30,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
3500,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
O.
sicu
A.
less
u
O. b
anks
A. a
ffin
Pat
r A
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Pat
r 2
Especies Presas
Por
cent
ajes
Fig. 39. Valores del Índice de Importancia Relativa (IIR) para hembras sexualmente inmaduras
de 152 – 257 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
58
Tabla 5. Espectro trófico de hembras sexualmente inmaduras de 152 - 257 cm LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal,
S: Secundaria y C: Circunstancial.
Especies N %N FO % FO P % P IIR Categ D. gigas 8 5,59 4 19,05 2230,62 9,48 287,21 P S. oualaniensis 25 17,48 7 33,33 17088,05 72,66 3004,63 P O. bartramii 13 9,09 8 38,10 3127,44 13,30 852,89 P L. diomedeae 23 16,08 2 9,52 500,91 2,13 173,46 S Histioteuthis sp. 3 2,10 3 14,29 425,95 1,81 55,84 S O. sicula 1 0,70 1 4,76 0,52 0,00 3,34 C A. lessueri 1 0,70 1 4,76 0,00 0,00 3,33 C O. banksii 1 0,70 1 4,76 0,00 0,00 3,33 C A. affinis 1 0,70 1 4,76 0,00 0,00 3,33 C Patrón A 1 0,70 1 4,76 0,00 0,00 3,33 C M. gayi 12 8,39 2 9,52 136,57 0,58 85,45 S B. panamense 48 33,57 4 19,05 2,81 0,01 639,59 P Larimus sp. 4 2,80 2 9,52 1,05 0,00 26,68 S Ophicthus sp. 1 0,70 1 4,76 5,00 0,02 3,43 C Patrón 2 1 0,70 1 4,76 0,10 0,00 3,33 C Suma 143 100,00 39 23519,02 100,00 9.4.3.3. Machos Sexualmente Maduros
Para los Machos Sexualmente Maduros de 244 – 310 cm. LT el Método
Numérico determinó que el mayor porcentaje fue para M. gayi con 50
individuos (30,86%), seguido de B. panamense con 43 especimenes (26,54%)
y S. oualaniensis con 33 (20,37%) (Fig. 40). Mientras que para el método
gravimétrico, el primer lugar lo obtuvo D. gigas y S. oualaniensis ambos con 6
ocasiones (40%), seguido de B. panamense con 5 (33,33%), O. bartramii y
M. gayi con 4 (26,67%), para ambos (Fig. 41).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
59
Método Numérico (Machos Sexualmente Maduros)
9 , 8 8
2 0 , 3 7
6,17
0 , 6 2 0 , 6 2 0 , 6 2
30 ,86
26 ,54
1 , 8 50 , 6 2 0 , 6 2
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
D. g
igas
S. o
uala
O. b
artra
His
t sp.
M.
dent
a
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Mus
t sp.
Thun
sp.
Pat
r 1
Especies Presas
Po
rcen
taje
s
Fig. 40. Valores porcentuales del Método Numérico para machos sexualmente maduros de 244
- 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del
2004.
Método de Frecuencia (Machos Sexualmente Maduros)
4 0 , 0 0 40 ,00
2 6 , 6 7
6 , 6 7 6 , 6 7 6 , 6 7
2 6 , 6 7
3 3 , 3 3
6 , 6 7 6 , 6 7 6 , 6 7
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
D. g
igas
S. o
uala
O. b
artra
His
t sp.
M.
dent
a
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Mus
t sp.
Thun
sp.
Pat
r 1
Especies Presas
Por
cent
ajes
Fig. 41. Valores porcentuales del Método de Frecuencia para machos sexualmente maduros
de 244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
El método gravimétrico, determinó que el mayor porcentaje fue para
S. oualaniensis con 22 990,17g (67,07%), seguido de D. gigas con 6 708,45g
(19,57%) y de O. bartramii con 22 219,68g (6,48%) (Fig.42). Luego, el IIR

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
60
estableció como presas principales a S. oualaniensis con 3 497,42; seguido
de D. gigas con 1 177,84 y M. gayi con 874,08 (Fig.43) (Tabla 6).
Método Gravimétrico (Machos Sexualmente Maduros)
19,57
67 ,07
6 , 4 8
0 , 0 0 0 , 0 0 0 , 0 01,91
0 , 0 1 0,17 0 , 0 3 0 , 0 00,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
D. g
igas
S. o
uala
O. b
artra
His
t sp.
M.
dent
a
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Mus
t sp.
Thun
sp.
Pat
r 1
Especies Presas
Por
cent
ajes
Fig. 42. Valores porcentuales del Método Gravimétrico para machos sexualmente maduros de
244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004.
Índice de Importancia Relativa (Machos Sexualmente Maduros)
1177,84
3497 ,42
337 ,28
4,12 4,12 4,12
8 7 4 , 0 8 8 8 5 , 0 6
13,51 4 , 3 1 4,120,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
3500,00
4000,00
D. g
igas
S. o
uala
O. b
artra
His
t sp.
M.
dent
a
Pat
r B
Pat
r C
M.
gayi
B. p
anam
Mus
t sp.
Thun
sp.
Pat
r 1
Especies Presas
Por
cent
ajes
Fig. 43. Valores del Índice de Importancia Relativa (IIR) para machos sexualmente maduros de
244 - 310 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
61
Tabla 6. Espectro trófico de machos sexualmente maduros de 244 - 310 cm. LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal,
S: Secundaria y C: Circunstancial.
Especies N % N FO % FO P % P IIR Categ D. gigas 16 9,88 6 40,00 6708,45 19,57 1177,84 P S. oualaniensis 33 20,37 6 40,00 22990,17 67,07 3497,42 P O. bartramii 10 6,17 4 26,67 2219,68 6,48 337,28 P Histioteuthis sp. 2 1,23 1 6,67 1633,16 4,76 39,99 S M. dentata 1 0,62 1 6,67 0,06 0,00 4,12 C Patrón B 1 0,62 1 6,67 0,00 0,00 4,12 C Patrón C 1 0,62 1 6,67 0,00 0,00 4,12 C M. gayi 50 30,86 4 26,67 656,00 1,91 874,08 P B. panamense 43 26,54 5 33,33 2,97 0,01 885,06 P Mustelus sp. 3 1,85 1 6,67 59,80 0,17 13,51 C Thunnus sp. 1 0,62 1 6,67 10,00 0,03 4,31 C Patrón 1 1 0,62 1 6,67 0,10 0,00 4,12 C Suma 162 100,00 32 34280,39 100,00 9.4.3.4. Machos Sexualmente Inmaduros
Para los Machos Sexualmente Inmaduros de 176 - 243 cm LT el Método
Numérico determinó que el mayor porcentaje fue para B. panamense con 99
individuos (65,13%), seguido de M. gayi con 24 organismos (15,79%) (Fig.44).
El método de frecuencia estableció el mayor porcentaje para M. gayi con 5
ocurrencias (50%), seguido de B. panamense y O. bartramii con 4 ocasiones
(40%) para ambos y L. diomedeae con 3 (30%) (Fig. 45).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
62
Método Numérico (Machos Sexualmente Inmaduros)
0,66 1,97 2,63 1,323,95
15,79
65,13
0,66 0,66
5,26
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
D. g
igas
S.
oual
a
O. b
artra
L. d
iom
e
His
t sp.
M. d
enta
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 44. Valores porcentuales del Método Numérico para machos sexualmente inmaduros de
176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre
del 2004.
Método de Frecuencia (Machos Sexualmente Inmaduros)
10
20
40
10 10
50
40
10 10 10
0
10
20
30
40
50
60
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
M. d
enta
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 45. Valores porcentuales del Método de Frecuencia para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
El método gravimétrico determinó el primer lugar para S. oualaniensis con
3 268,62g (76.88%), seguido de O. bartramii con 349,25g (8,21%) y del
calamar Histioteuthis sp. con 288,42g (6,78%) (Fig. 46). El IIR presentó como
presas principales a B. panamense con 2 611,42; seguido de
S. oualaniensis con 1 577,13 y M. gayi con 962,59 (Fig. 47).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
63
Método Gravimétrico (Machos Sexualmente Inmaduros)
2,79
76,88
8,21 6,78
0,003,46
0,15 0,00 0,00 0,000,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
M.
dent
a
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Gas
ter
Especies Presas
Po
rcen
taje
s
C
Fig. 46. Valores porcentuales del Método Gravimétrico para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.
Índice de Importancia Relativa (Machos Sexualmente Inmaduros)
34,48
1577,13
433,86
81,00 39,48
962,59
2611,42
6,61 6,60 52,64
0,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
D. g
igas
S. o
uala
O. b
artra
L. d
iom
e
His
t sp.
M. d
enta
M. g
ayi
B. p
anam
Lari
sp.
Oph
i sp.
Gas
ter
Especies Presas
Por
cent
ajes
Fig. 47. Valores del Índice de Importancia Relativa (IIR) para machos sexualmente inmaduros
de 176 - 243 cm LT de A. pelagicus en Santa Rosa de Salinas durante mayo –
diciembre del 2004.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
64
Tabla 7. Espectro trófico de machos sexualmente inmaduros de 176 - 243 cm LT de
A. pelagicus en Santa Rosa de Salinas durante mayo – diciembre del 2004
expresado en valores absolutos y porcentuales del método numérico (N y %N),
frecuencia de ocurrencia (FO y %FO), gravimétrico (P y %PO), índice de importancia
relativa (IIR) y Categoría de la especie presa (Categ), donde P: Presa Principal,
S: Secundaria y C: Circunstancial.
Especies N % N FO % FO P % P IIR Categ D. gigas 1 0,66 1 10 118,61 2,79 34,48 S S. oualaniensis 3 1,97 2 20 3268,62 76,88 1577,13 P O. bartramii 4 2,63 4 40 349,25 8,21 433,86 P L. diomedeae 3 1,97 3 30 72,47 1,70 110,35 S Histioteuthis sp. 2 1,32 1 10 288,42 6,78 81,00 S M. dentata 6 3,95 1 10 0,03 0,00 39,48 S M. gayi 24 15,79 5 50 147,20 3,46 962,59 P B. panamense 99 65,13 4 40 6,54 0,15 2611,42 P Larimus sp. 1 0,66 1 10 0,15 0,00 6,61 C Ophicthus sp. 1 0,66 1 10 0,10 0,00 6,60 C Gasterópodos 8 5,26 1 10 0,05 0,00 52,64 S
Suma 152 100,00 24 4251,44 100,00
9.5. ÍNDICES ECOLÓGICOS
9.5.1. Diversidad y Equitabilidad de las Especies Presas
Los Valores de diversidad de especies presas consumidas para todas las
categorías fue bien baja, siendo así, que el índice general fue de H’= 1,61 con
una equitabilidad de E= 0,51 lo que confirma la dominancia por tres presas
principales de 24 componentes alimenticios en la dieta de A. pelagicus. Para
los sexos, la mayor fue para los machos, H’= 1,68 (E= 0,60) sobre las hembras
de H’= 1,47 (E= 0,52). En cuanto a los estados de madurez sexual, la mayor
perteneció a las hembras sexualmente inmaduras de 152 - 257 cm. con
H’= 1,98 (E= 0,79) mientras que, para las hembras sexualmente maduras de
258 - 365 cm. fue de H’= 1,09 (E= 0,49). Los machos sexualmente maduros de
244 - 360 cm. obtuvieron mayor diversidad con H’= 1,72 (E= 0,69) que los
machos inmaduros de 176 - 243 cm con H’= 1,26 (E= 0,53). A las hembras
inmaduras correspondieron la mayor diversidad y para las hembras maduras el

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
65
menor valor, entre todas las siete categorías, debido a la preferencia por pocas
presas (Tabla 8).
Tabla 8. Valores de Diversidad y Equitabilidad para las diferentes categorías de A. pelagicus,
en Santa Rosa de Salinas durante mayo – diciembre del 2004.
Categorías Diversidad Equitabilidad General 1,61 0,51 Hembras 1,47 0,52 Machos 1,68 0,60 Hembras Maduras de 258 - 365 cm. 1,09 0,49 Hembras Inmaduras de 152 - 257 cm. 1,98 0,73 Machos Maduros de 244 - 360 cm. 1,72 0,69 Machos Inmaduros de 176 - 243 cm. 1,26 0,53
9.5.2. Amplitud del Nicho Trófico
Los Valores del Índice de Levin para todas las categorías fue bien baja, siendo
así, que el índice general fue de Bi= 0,11 lo que confirma una dieta dominada
por tres presas de un total de 24 componentes en la alimentación de
A. pelagicus, por lo que se trata de un depredador especialista. Para los
sexos, el mayor fue para los machos Bi= 0,17 sobre las hembras de Bi= 0,12.
Para los estados de madurez sexual, el mayor perteneció a las hembras
sexualmente inmaduras de 152 - 257 cm. con Bi= 0,31 mientras que, para las
hembras sexualmente maduras de 258 - 365 cm. fue de Bi= 0,14. Los machos
sexualmente maduros de 244 - 360 cm. obtuvieron el mayor valor con Bi= 0,32
que los machos inmaduros de 176 - 243 cm. con Bi= 0,12. A los machos
maduros perteneció el mayor valor del índice y para el general el menor valor
(Tabla 9).
Tabla 9. Valores del Índice de Levin para las diferentes categorías de A. pelagicus, en Santa
Rosa de Salinas durante mayo – diciembre del 2004.
Categorías Valores General 0,11 Hembras 0,12 Machos 0,17 Hembras Maduras de 258 - 365 cm 0,14 Hembras Inmaduras de 152 - 257 cm 0,31 Machos Maduros de 244 - 360 cm 0,32 Machos Inmaduros de 176 - 243 cm 0,12

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
66
9.5.3. Superposición de Dietas o Traslapamiento trófico
Los valores de superposición de dietas fueron bastante aceptables para las tres
categorías confrontadas. El mayor valor fue para los machos y hembras de
Cλ= 0,79 lo que indica un traslapamiento mayor, esto es, mayor similitud en los
elementos de la dieta de A. pelagicus; seguido de los machos maduros e
inmaduros con Cλ= 0,68, esto es, mayor similitud. Finalmente, el menor valor
fue para las hembras maduras e inmaduras con Cλ= 0,42 lo que indica un
traslapamiento medio, es decir, similitud media en los elementos alimenticios
(Tabla 10).
Tabla 10. Valores de Superposición de dietas para las categorías confrontadas del tiburón
zorro pelágico A. pelagicus, en Santa Rosa de Salinas durante mayo – diciembre
del 2004.
Categorías Valores
Hembras Vs. Machos 0,79 Hembras Maduras Vs. Inmaduras 0,42 Machos Maduros Vs. Inmaduros 0,68
9.6. VARIACIÓN MENSUAL DE LAS ESPECIES PRESAS
Se presentó una gran diferencia entre las especies presas consumidas durante
los meses muestreados. En mayo se presentó un predominio de
S. oualaniensis, D. gigas, O. bartramii y L. diomedeae, respectivamente, los
cuales iban disminuyendo conforme transcurría el tiempo. En junio y julio no se
obtuvo ejemplares, por ende, se prescinde de dicha información, lo cual se
deba probablemente a la migración de A. pelagicus hacia zonas oceánicas
debido a que éstos son capturados en zonas costeras adyacentes al Golfo de
Guayaquil, según los pescadores. Mientras que para agosto empezaron a
declinar las poblaciones de los cuatro calamares citados anteriormente y
M. gayi y B. panamense comenzaron a aparecer. Luego, en septiembre, se
muestreó en una sola ocasión por lo que se obtuvieron dos individuos sin
contenido estomacal (0% de llenado), por lo que no hay información disponible
de ese mes. Para octubre, permanecieron las poblaciones de
S. oualaniensis; por su parte B. panamense predomina sobre M. gayi que

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
67
empieza a entrar en apogeo. Durante noviembre, declinan todas las
poblaciones de calamares. Finalmente en Diciembre, persistió una pequeña
población de O. bartramii y M. gayi predomina con una gran diferencia sobre
B. panamense.
9.7. ESTADOS DE DIGESTIÓN DE PRESAS
La mayoría de las especies presas consumidas se presentaron en estados de
digestión avanzados, siendo así el 50% de las presas estaban completamente
digeridas [Estado de digestión (ED)= 4]; seguido de un 40% en ED= 3, un 8%
en ED= 2 y un 2% en ED= 1. Esto se justifica debido al tiempo necesario
(alrededor de 14 horas) y a la gran distancia que debían recorrer para que los
pescadores desembarquen éstos individuos, lapso suficiente para que los jugos
gástricos del organismo, ya muerto, digieran la mayor parte de sus presas. De
acuerdo a los pescadores del sector, el itinerario de la faena de pesca era, a
groso modo, el siguiente: 13H00, salida del puerto en botes de fibras de vidrio;
16H00, llegada al caladero (zona pesquera de pelágicos mayores en el Golfo
de Guayaquil); 17H00 a 20H00, colocación de los artes de pesca
(preferentemente, palangres y redes); 04H00, recogida de los artes de pesca y
con ello el embarque de la pesca; 07H00, salida del caladero; 10H00, llegada al
puerto y desembarque de la pesca. Es menester, manifestar que si el caso lo
requería la pesca se prolongaba a tres o cuatro días.
9.8. ESPECIES PRESAS DE CALAMARES CONSUMIDOS POR Alopias
pelagicus
La medida tomada en consideración para la descripción de las especies de
calamares fue la Longitud Rostral LR (desde el rostro hasta el ángulo
mandibular), sin embargo, para la presentación de las figuras se empleó la
Longitud Total LT (desde el rostro hasta el extremo inferior de la pared). Las
presas fueron las siguientes:
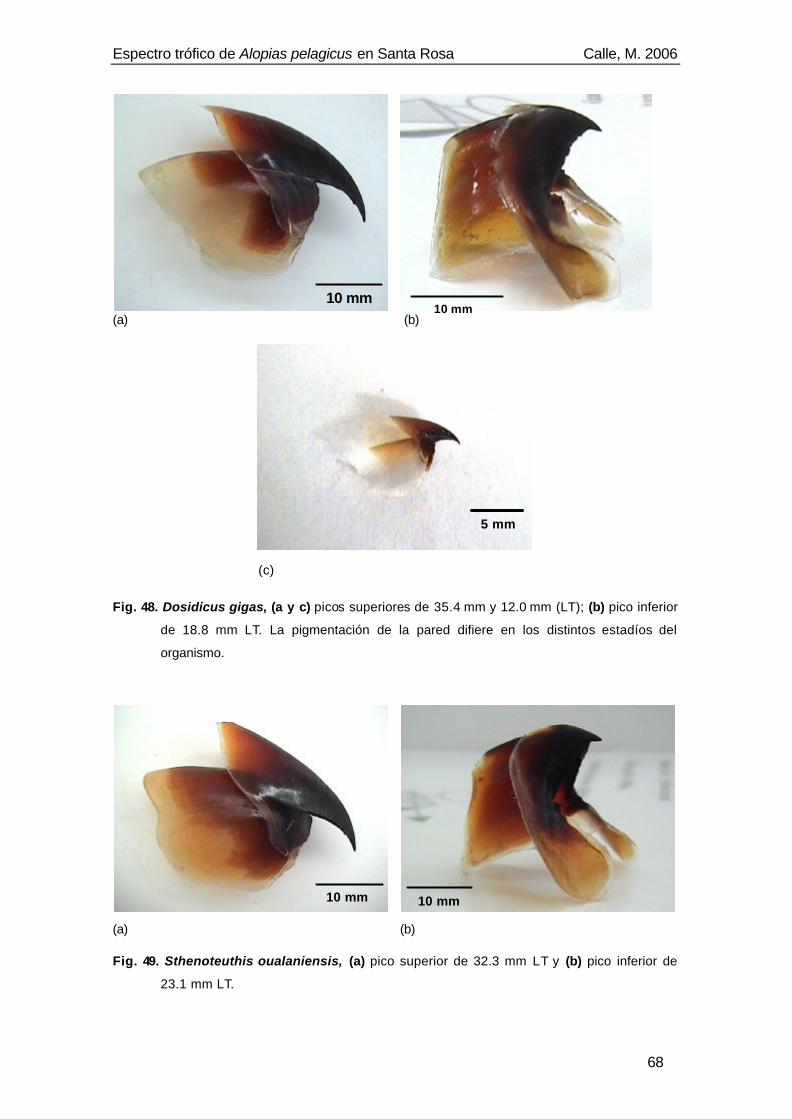
Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
68
(a) (b)
(c)
Fig. 48. Dosidicus gigas, (a y c) picos superiores de 35.4 mm y 12.0 mm (LT); (b) pico inferior
de 18.8 mm LT. La pigmentación de la pared difiere en los distintos estadíos del
organismo.
(a) (b) Fig. 49. Sthenoteuthis oualaniensis, (a) pico superior de 32.3 mm LT y (b) pico inferior de
23.1 mm LT.
10 mm 10 mm
5 mm
10 mm 10 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
69
(b)
(c) (a) (b)
(c)
Fig. 50. Ommastrephes bartramii, (a) pico superior de 18.8 mm LT; (b) pico inferior de 24.8
mm LT; (c) vista dorsal del superior donde se observa línea pigmentada que recorre la
mitad de la cresta.
(a) (b) Fig. 51. Octopoteuthis sicula, (a) pico superior de 14.1 mm LT; (b) pico inferior de 13.5 mm
LT.
5 mm
5 mm
5 mm
5 mm
6 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
70
(a) (b) (c) Fig. 52. Loliolopsis diomedeae, (a-c) picos superiores de 6.0 y 5.9 mm LT; (b) pico inferior
de 4.7 mm LT.
(a) (b)
Fig. 53. Histioteuthis sp, (a) pico superior de 12.0 mm LT y (b) pico inferior de 13.3 mm LT.
2 mm 3 mm
3 mm
5 mm 5 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
71
Fig. 54. Histioteuthis sp, pico inferior de 15.5 mm LT.
Fig. 55. Onychoteuthis banksii, pico inferior de 11.0 mm LT.
5 mm
5 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
72
(a) (b) (c) (d)
(e) (f) Fig. 56. Mastigoteuthis dentata, de longitudes progresivas; (a, c, e) picos superiores de 3.2
mm, 5.6 mm y 6.0 mm LT y (b, d, f) picos inferiores de 4.8 mm, 5.2 mm y 6.7 mm LT.
2.4 mm 4.8 mm
5.6 mm 5.2 mm
6 mm 6.7 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
73
(a) (b) (c) (d)
(e) Fig. 57. Mastigoteuthis dentata, de longitudes progresivas; (a – e) picos superiores de 5.7
mm, 8.4 mm, 8.6 mm, 9.3 mm y 12.0 mm LT.
5.7 mm 5 mm
4 mm 5 mm
5 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
74
Fig. 58. Morfotipo A (Patrón A), pico superior de 13.3 mm LT.
Fig. 59. Morfotipo B (Patrón B), pico superior de 20.2 mm LT.
(a) (b)
Fig. 60. Morfotipo C (Patrón C), (a y b) pico superior de 5.9 mm (LT)
5 mm
10 mm
2 mm 2 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
75
9.9. ESPECIES PRESAS DE PECES CONSUMIDOS POR Alopias pelagicus
(a) (b)
(c) (d)
(e) Fig. 61. Merluccius gayi , (a) arriba: esqueleto de 125.2 mm LT; abajo: de 90.9 mm LT; (b)
detalle de las vértebras abdominales que muestra las parapófisis replegadas en forma
de tubo propio de la especie; (c) detalle de vértebras caudales; (d) otolito de 12.4 mm
(LT) y (e) variaciones de la forma del mismo producto de la digestión.
50 mm
3 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
76
(a) (b)
(c) Fig. 62. Benthosema panamense, (a) esqueleto de 18.2 mm LT; (b) otolito de 1.8 mm LT
observado bajo estereoscopio en 12x y (c) variaciones en la morfología del mismo
como producto de la digestión.
(a)
(b) (c) Fig. 63. Larimus sp, (a) esqueleto de 63.9 mm LT; (b) otolito de 7.2 mm LT y (c) variaciones
en la morfología del mismo por desgaste de la digestión.
10 mm
1 mm
20 mm
7.2 mm 7.2 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
77
(a)
(b)
(c)
(d) Fig. 64. Ophichthus sp, (a) esqueleto de 254.0 mm LT; (b) detalle de las expansiones de los
arcos hemales; (c) detalle de las vértebras caudales (Vista lateral) y (d) esqueletos
de 20.0 mm y 12.0 mm LT.
20 mm
2 cm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
78
(a)
(b) (c)
(d) Fig. 65. Mustelus sp, (a) macho de 225 mm LT; (b) detalle de la boca de una hembra de 220
mm LT bajo estereoscopio en 12x; (c) vista lateral la misma y (d) arriba: hembra de 220
mm LT, centro: macho de 225 mm LT y abajo: hembra de 232 mm LT.
20 mm
30 mm
30 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
79
(a) (b)
(c) Fig. 66. Thunnus sp, (a) vista lateral del cráneo de 59.8 mm (Long. craneal) y (b - c) vista
dorsal del mismo.
Fig. 67. Coryphaena hippurus, vértebras caudales de. 36.2 mm (LT).
10 mm 10 mm
10 mm
5 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
80
(a) (b)
(c) (d) Fig. 68. Morfotipo 1 (Patrón 1), (a) esqueleto de 22.2 mm LT; (b) de 19.7 mm LT; (c) de 29.5
mm LT y (d) comparación de magnitudes de los tres anteriores.
(a) (b) Fig. 69. Morfotipo 2 (Patrón 2), (a) esqueleto de 39.3 mm (LT) y (b) detalle de las vértebras
5 mm 5 mm
5 mm
8 mm
5 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
81
(a) (b) (c) Fig. 70. Morfotipo 3 (Patrón 3), (a) esqueleto de 50.9 mm (LT) y (b-c) detalle de las vértebras
abdominales
9.10. GASTERÓPODOS CONSUMIDOS INCIDENTALMENTE POR Alopias
pelagicus
(a) (b) Fig. 71. Gasterópodos de (a) 1 mm LT y (b) de 3.2 mm LT presentes en el estómago de un
macho inmaduro de A. pelagicus de 190 cm LT observados bajo estereoscopio en 12 x.
Su consumo incidental obedece probablemente a que estos organismos pelágicos
estaban cercanas a la presa objetivo en el momento de la alimentación del predador.
1 mm 2 mm
10 mm
3 mm
3 mm

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
82
10. DISCUSIONES
Durante éstos últimos años la especie A. pelagicus estuvo sujeta a diversas
discusiones en cuanto a su identificación. Compagno et al. (1995) reporta la
distribución de ésta especie para aguas de México, Costa Rica, Panamá y las
Islas Galápagos. Según Estupiñán (Com. pers. 2006) Galván identifica por
primera vez a A. pelagicus para aguas ecuatorianas en Manta en el 2002. El
siguiente año, Galván (Com. pers. 2003) identifica nuevamente a A. pelagicus
en Guayaquil y advierte que ésta se encontraba mal identificada como
A. vulpinus. Baigorrí y Polo (2004) reportan por primera vez a la especie en
su tesis de grado. Jiménez y Béarez (2004) la registran en su lista de especies
de peces continentales. Galván (Com. pers. 2005) la certifica nuevamente en
abril como respaldo para el trabajo en mención. Villón y Aguilar (2005) reportan
A. pelagicus a nivel nacional durante octubre, siendo la especie confirmada
por el Dr. George Burgess del Museo de Historia Natural de Florida. Burgess
(Com. pers. 2006) en un certificado extendido también como respaldo para el
presente estudio certifica y ratifica a A. pelagicus como parte de nuestra
ictiofauna. Por lo citado anteriormente, no cabe duda alguna, que A. pelagicus
ha existido en aguas ecuatorianas y que ésta era confundida con la especie
A. vulpinus.
El número de organismos muestreados de A. pelagicus en el presente estudio
fue de 110, producto de un muestreo de 2 días por semana durante 8 meses
(mayo – diciembre del 2004), esto es, alrededor de 64 días de trabajo. Baigorrí
y Polo (2004) obtienen 111 durante junio – diciembre del 2003 en Manta
muestreando un promedio de 5 días por semana durante 7 meses junio –
diciembre del 2003, aproximadamente 140 días; lo que indica probablemente
un mayor número de desembarques de la especie en Santa Rosa de Salinas.
La distribución mensual en el número de estómagos de A. pelagicus
presentaron los valores más altos para diciembre y mayo, mientras que en
junio y julio no se presentó ningún organismo en el puerto de Santa Rosa de
Salinas. Baigorrí y Polo (2004) obtienen los más altos para septiembre y
octubre y, los más bajos para julio y diciembre; aparentemente las poblaciones

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
83
que migran en diciembre lo hacen desde Manta hasta Santa Rosa, y la otra
gran parte lo realiza sin que se pueda conocer su destino.
La mitad de los estómagos analizados de A. pelagicus (50,90%) estaban
vacíos, en cuanto a proporción de llenado (0= 0%) y el 50% de las presas
presentaban avanzados estados de digestión (ED) 4 y el 40% un ED 3.
Baigorrí y Polo (2004) sostienen que la mayoría de los estómagos muestreados
se encuentran entre un 25% y 50% llenos y el 70% de las presas consumidas
están en estado de digestión 3, presentando más organismos con mayores
porcentajes de llenado estomacal y confirma los avanzados estados de
digestión de las presas.
El 73% de los estómagos estudiados de A. pelagicus (80) contenían
alimentos, es decir, presentaban por lo menos un componente alimenticio y el
27% (30) vacíos, literalmente. Baigorrí y Polo (2004) presentan el 82% (91
estómagos) con alimentos y el 18% (20) vacíos, esto es, mayor porcentaje de
estómagos con alimentos.
Alrededor de 14 horas transcurrieron desde que A. pelagicus fue atrapado
hasta ser llevado al puerto de Santa Rosa de Salinas, tiempo suficiente para
que los jugos gástricos actúen en la descomposición y digestión de las presas
aún después de muerto el predador, motivo por el cual la mayor parte de las
presas se encuentran en un estado de digestión avanzado. Baigorrí y Polo
(2004), afirman que alrededor de 10 a 12 horas transcurren hasta ser llevados
a la playa de Tarqui, lo que confirma los avanzados estados de digestión.
Un total de 24 componentes alimenticios (presas) en la dieta de A. pelagicus
se presentaron agrupados en 4 categorías principales: 1 gasterópodo, 13
cefalópodos, 1 condrictio y 9 osteictios. Baigorrí y Polo (2004) también
encuentran 24 elementos en 3 categorías: 1 crustáceo, 7 cefalópodos y 16
teleósteos; esto indica que ingirieron la mitad del número de cefalópodos y casi
el doble de peces.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
84
El número de organismos consumidos por A. pelagicus fue de 928 con un
peso de 86 977,88 g, entre los cuales: los osteictios predominaron con
74,68%, cefalópodos con el 24,14%, los gasterópodos con el 0,86% y los
condrictios con (0,32%). Baigorrí y Polo (2004) cuantifican 1 374 organismos
que representan 89 002,3 g de los cuales los cefalópodos representan el
94,8%, los peces el 5,1% y los crustáceos el 0,1%, esto indica que
consumieron más organismos los cuales aportaron menor volumen alimenticio.
El IIR determinó tres componentes principales en la dieta: S. oualaniensis con
2 071,9; M. gayi con 1 569, 02 y B. panamense con 1 086,97. Baigorrí y Polo
(2004) encuentran de igual manera, tres componentes principales: D. gigas
(IIR=5 864), seguido por B. panamense (IIR=2 693) y S. oualaniensis
(IIR=262); presentándose 2 de las 3 presas principales halladas en los
organismos de Santa Rosa.
En cuanto a la biología y hábitat de las principales presas,
Sthenoteuthis oualaniensis es una especie pelágica que se alimenta
activamente durante la noche encontrándose frecuentemente en cercanías a la
plataforma continental en pequeños grupos, aunque en el día puede hallarse
en aguas con poco oxígeno (350 – 400 m) (Markaida y Sosa-Nishizaki, 2003).
Al parecer A. pelagicus se alimentó por la noche dirigiéndose hasta la
plataforma para la captura de esta presa, la cual es el primer componente
principal en su dieta.
Merluccius gayi, lo más probable Merluccius gayi peruanus por su
distribución (de 50 S a 14 0 S), es una especie mesopelágica que se encuentra
desde aguas pocos profundas de la plataforma (50 m) hasta la parte superior
del talud (por debajo de los 500 m) y su principal período de desove va de
Agosto a Marzo (de 40 S a 8 0 S) (Cohen, Inada, Iwamoto & Scialabba, 1990) lo
cual justifica que A. pelagicus se haya alimentado mayoritariamente de esta
especie de octubre a diciembre, no así el resto del año; siendo
Merluccius gayi la segunda presa principal en su dieta.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
85
Benthosema panamense, es una especie mesopelágica que puede
encontrarse sobre las plataformas continentales e insulares. Se extienden
desde la superficie (de noche) hasta por debajo de los 2 000 m de profundidad,
efectuando migraciones verticales desde profundidades mayores a 200 m en
donde permanecen de día llegando hasta la capa superior del agua en la noche
(Paxtón, Lavenberg y Sommer, In: Allen et al., 1995). Estudios realizados en
hábitos alimenticios del calamar gigante D. gigas por Nesis (1970), Bennet
(1978) y Markaida y Sosa-Nishizaki (2003), revelan que B. panamense es su
presa principal, lo cual permite suponer que A. pelagicus pudo dirigirse tanto a
D. gigas como a B. panamense. Esta última especie es la tercera presa
principal en su dieta.
Dosidicus gigas, es una especie abundante con amplia distribución en el
Pacífico Oriental que realiza importantes migraciones hacia Perú y Chile donde
se alimenta en invierno y verano (Zinder, 1998 In: Cabrera, 2003). Se desplaza
en grupos grandes y vive en aguas oceánicas profundas, se acerca a la costa
cuando realizan migraciones verticales con fines alimenticios o reproductivos
que dependen del estado de madurez de los organismos, así como de la hora
del día. Los juveniles realizan migraciones hacía la superficie durante la noche
para alimentarse mientras que los adultos migran pero se mantienen entre los
10 y 35 m, ambos grupos migran hacia las profundidades (2 000 m) haciéndose
menos vulnerables a los depredadores (Markaida y Sosa-Nishizaki, 2003), por
lo que A. pelagicus pudo haberse alimentado de los juveniles cuando realizan
migraciones hacia la superficie durante la noche o de los adultos a esa
profundidad antes de que empiecen las migraciones de descenso, ya que de
acuerdo a Compagno et al. (1995) este predador es epipelágico, se encuentra
frecuentemente cerca de la costa y alrededor de la plataforma continental hasta
los 152 m de profundidad.
Ommastrephes bartramii, es una especie oceánica que se encuentra desde
la superficie hasta unos 1 500 m de profundidad. A través de todo el verano y el
otoño se observan densos cardúmenes que se alimentan en aguas
superficiales, después migran a aguas más profundas, donde se dispersan
durante el invierno y la primavera. Realizan migraciones verticales diarias para

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
86
su alimentación entre aguas subsuperficiales (de noche) y más profundas (de
día) (Roper, Sweeney & Hochberg, 1995) lo que justifica que dicha especie
haya sido el tercer componente principal de la dieta de A. pelagicus de mayo a
agosto (época seca = verano).
Loliolopsis diomedeae, es una especie nerítica que frecuentemente forma
grandes cardúmenes en el Golfo de Panamá, a pesar de su distribución desde
el golfo de California hasta Perú (Roper et al., 1995); al parecer la especie
estuvo presente sólo para inicios de mayo lo cual justifica que dicha especie
haya sido el cuarto componente principal, de manera estacional, en la dieta de
A. pelagicus.
El calamar pelágico D. gigas conjuntamente con las especies O. bartramii,
S. oualaniensis y Thysanoteuthis rombus forman el grupo de especies
identificadas en aguas ecuatorianas (Pacheco, Bermeo, Pozo, Aguirre y
Oviedo, 1996) así como Loliolopsis diomedeae (Roper et al., 1995); lo que
confirma la presencia de las especies presas encontradas dentro de la
alimentación de A. pelagicus.
El resto de los componentes alimenticios de A. pelagicus se encuentran en
número, ambiente y profundidad disponibles para la alimentación del
organismo en estudio. Además, se confirmó los hábitos alimenticios nocturnos
de A. pelagicus según la biología y hábitat de los cuatro componentes
principales de su dieta.
La variación mensual en la alimentación de A. pelagicus registró diferencias
bien marcadas, por lo que en mayo se alimentó de S. oualaniensis, D. gigas,
Ommastrephes bartramii y Loliolopsis diomedeae y de octubre a diciembre
lo hizo de B. panamense y M. gayi; indicando una especificidad temporal que
está directamente relacionada con la disponibilidad del recurso. Baigorrí y Polo
(2004) presentan a D. gigas, B. panamense y S. oualaniensis como presas
preferidas durante todo el año con una variación en julio, lo que indica una
especificad bastante alta.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
87
De los 110 estómagos colectados de A. pelagicus, 45 eran de hembras
sexualmente maduras (incluidas 7 grávidas) y 29 de inmaduras; 20 de machos
maduros y 16 de inmaduros. Baigorrí y Polo (2004) sostiene que de los 111
estómagos, 53 pertenecen a individuos maduros (36 hembras y 17 machos),
32 inmaduros (22 hembras y 10 machos) y 26 de hembras grávidas,
presentando mayor número de hembras maduras y el menor de machos
maduros.
La alimentación por sexos, estados de madurez sexual y tallas de
A. pelagicus, según el IIR, se conforma principalmente por M. gayi, seguido
de S. oualaniensis, B. panamense, D. gigas y O. bartramii. Baigorrí y Polo
(2004) establecen como presas principales a D. gigas, B. panamense,
S. oualaniensis, Auxis thazard, M. gayi y Sardinops sagax, lo que difiere al
ser D. gigas el componente más importante.
Los valores de diversidad de presas consumidas para todas las categorías de
A. pelagicus fueron bien bajos, lo que confirma una dieta poco diversa
dominada por tres presas principales: S. oualaniensis, M. gayi y
B. panamense. De igual manera, Baigorrí y Polo (2004) presentan, en su
estudio, una dieta poco diversa debido a que está se encuentra dominada por
tres especies presas: D. gigas, B. panamense y S. oualaniensis, lo que
reafirma la baja diversidad.
La amplitud del nicho trófico determinó, para todas las categorías, a
A. pelagicus como un depredador especialista. Baigorrí y Polo (2004), de igual
manera, también indica esta característica esencial de esta especie.
El traslapamiento trófico en A. pelagicus se presentó mayor tanto para sexos
como para machos maduros e inmaduros, lo cual confirmó que no existe
diferencias en sus elementos de dieta. Sin embargo, para las hembras
maduras e inmaduras estableció traslapamiento medio, lo que sugirió similitud
media en sus dietas. Baigorrí y Polo (2004) establecen traslapamiento entre
sexos y estados de madurez lo que indica que consumen los mismos recursos
alimenticios principalmente: D. gigas y B. panamense independientemente de

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
88
la proporción, mostrando que no se presenta una segregación por alimento
debido al sexo o estado de madurez. Se presentó traslapamiento tanto entre
sexos, estados de madurez y tallas, tanto para los individuos colectados en
Manta como en Santa Rosa de Salinas.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
89
11. CONCLUSIONES
1. El espectro trófico de Alopias pelagicus en Santa Rosa de Salinas durante
mayo – diciembre del 2004 estuvo compuesto por 24 componentes
alimenticios, entre ellos: Dosidicus gigas, Sthenoteuthis oualaniensis,
Ommastrephes bartramii, Loliolopsis diomedeae, Histioteuthis sp.,
Octopoteuthis sicula, Ancistrocheirus lessueri, Mastigoteuthis dentata,
Onycoteuthis banksii, Abraliopsis affinis, Morfotipo A, Morfotipo B, Morfotipo
C, Merluccius gayi, Benthosema panamense, Larimus sp., Ophichthus sp.,
Mustelus sp., Thunnus sp., Coryphaena hippurus, Morfotipo 1, Morfotipo 2,
Morfotipo 3 y Gasterópodos.
2. Las especies presas principales consumidas por Alopias pelagicus, según
el Índice de Importancia Relativa fueron: Sthenoteuthis oualaniensis,
Merluccius gayi y Benthosema panamense.
3. La alimentación por sexos, estados de madurez sexual y tallas de
Alopias pelagicus estuvo dominada por Merluccius gayi, Sthenoteuthis
oualaniensis, Benthosema panamense, Dosidicus gigas y Ommastrephes
bartramii.
4. La diversidad de presas consumidas por Alopias pelagicus, según el
Índice de Shannon y Weaver, tanto en forma general como para sexos,
estados de madurez y tallas, fue bien baja, ya que a pesar de su alto
número de componentes alimenticios (24), la dieta estuvo dominada por
tres especies.
5. La amplitud del nicho trófico, de acuerdo al Índice de Levin, para sexos y los
estados de madurez y tallas, determinó a Alopias pelagicus como un
depredador especialista, debido a su preferencia por tres especies.
6. La superposición de dieta, según el Índice de Morisita-Horn, estableció
traslapamiento entre sexos, estados de madurez sexual y tallas, lo cual
confirmó que no existen diferencias en los elementos de la dieta de
Alopias pelagicus.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
90
7. La variación mensual de la dieta de Alopias pelagicus estableció que
durante mayo se alimentó de Sthenoteuthis oualaniensis, Dosidicus gigas,
Ommastrephes bartramii y Loliolopsis diomedeae y de octubre a diciembre
lo hizo de B. panamense y Merluccius gayi.
8. La variación mensual de los individuos de Alopias pelagicus analizados
en el puerto de Santa Rosa de Salinas determinó que durante mayo y
diciembre se registró el mayor número de dicha especie, así como el
número de hembras grávidas. Mientras que, de junio a julio no se
registraron organismos.
9. La mitad de los estómagos analizados de Alopias pelagicus estaban
vacíos, en cuanto a proporción de llenado (0= 0%) y de igual forma, la
mayoría de las presas presentaban avanzados estados de digestión.
10. La mayoría de los estómagos estudiados de Alopias pelagicus (80 de
110) contenían alimentos, es decir, presentaban por lo menos un
componente alimenticio, y los 30 restantes estaban vacíos literalmente.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
91
12. RECOMENDACIONES
1. La prioridad por el conocimiento de qué especies de peces, especialmente
de tiburones, rayas y batoideos y, los registros de sus desembarques en los
principales puertos pesqueros es fundamental para estudios de estructuras
poblacionales. De lo que se deriva la necesidad de hacer estudios
nacionales realmente serios con personal altamente calificado que conlleve
a una verdadera evaluación de nuestros recursos ícticos, con el fin de evitar
la suposición de especies que nunca han existido para nuestras aguas y el
de la sobrepesca para aquellos que si tenemos.
2. El estudio de la biología básica de los tiburones es indispensable debido a
que cumplen un importante rol para las cadenas alimenticias marinas y es
un componente sumamente importante en las pesquerías de nuestro país.
Por ello se hace énfasis en la necesidad de continuar con este tipo de
estudios que permitan determinar la biología y ecología de los tiburones.
3. Los estudios reproductivos de los tiburones son prioritarios debido a que se
conoce que la mayoría son especies longevas que tardan muchos años en
alcanzar la madurez sexual y tienen un bajo nivel reproductivo. Con dicho
conocimiento, se podrá establecer leyes que regulen la pesca en ciertas
zonas y épocas de apareamiento.
4. Las pesquerías de las diversas especies marinas permiten un equilibrio, si
bien es cierto no tan efectivo, en las poblaciones de los depredadores y de
las presas. De lo que se desprende, la necesidad del empleo de leyes y
técnicas que permitan una pesca sostenida y sustentable que eviten efectos
negativos sobre las poblaciones de predadores y, por ende, una ruptura en
la cadena trófica.
5. La realización de los trabajos de investigación científica sean estas tesis de
grado, de maestrías y de doctorados, conllevan a un conocimiento público
sobre la biología de los recursos naturales. Por ello es necesario, la
publicación de los mismos en las revistas de ésta índole y también en la
Internet.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
92
13. LITERATURA CITADA
ABURTO, O. 1997. Conducta alimenticia del ángel rey Holocanthus passer.
Baja California Sur, México. Tesis de Licenciatura. Universidad
Autónoma de Baja California Sur. pg. 7-23.
AGUILAR, F., X. CHALÉN, C. VILLÓN, P. SOLÍS y N. GAIBOR. 2005. Plan de
acción nacional para la conservación y ordenación de los tiburones en el
Ecuador (PAT-Ecuador). Guayaquil, Ecuador. Instituto Nacional de
Pesca (INP). 26 p.
ALCÍVAR, V. y F. LÓPEZ. 2001. Pesca artesanal y comercialización de
tiburones zorros Alopias superciliosus (Lowe 1839) y Alopias vulpinus
(Nakamura 1935) en el Puerto de Manta, Provincia de Manabí. Manta,
Ecuador. Tesis de grado. Univ. Laica “Eloy Alfaro” de Manabí. pg. 12-
24.
BAIGORRÍ, A. y C. POLO. 2004. Espectro trófico de dos especies de tiburón
zorro, Alopias pelagicus y Alopias superciliosus en la playa de Tarqui,
Manta, Ecuador. Bogotá, Colombia. Universidad de Bogotá “Jorge
Tadeo Lozano”. 133 p.
BÉAREZ, P. 1996. Lista de los peces marinos del Ecuador Continental. Revista
de Biología Tropical. 44 (2). pg. 731-741.
BENNET, B. 1978. Underwater observations of the squid Illex illebrosus in the
Newfoundland in shore waters. Canada. Fisheries and Environment
Canada. Report 833. pp. 12-19.
BOCANEGRA, N., L. ABITIA, y F. GALVÁN, 2000. Espectro trófico de la
berrugata californiana Menticirrus undulatus de Laguna Ojo de Liebre
Baja California Sur, México. Baja California Sur, México. Ciencias
Marinas. Instituto Politécnico Nacional (IPN), Centro Interdisciplinario de
Ciencias Marinas (CICIMAR). 26 (4). pg. 659-675.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
93
BROOKS, J. & S. DODSON. 1965. Predation, body size, and composition of
plankton. Science 150. pg. 28–35.
CABRERA, A. 2003. Hábitos alimenticios del Tiburón piloto
Carcharhinus falciformis (Bibron 1839) en la Costa Occidental de Baja
California Sur. Baja California Sur, México. Tesis de Maestría. IPN,
CICIMAR. 91 p.
CABRERA, M. 2004. Biología reproductiva del Tiburón azul Prionace glauca
(Linnaeus 1758) en la Costa Occidental de Baja California Sur. Baja
California Sur, México. Tesis de Maestría. IPN, CICIMAR. 67 p.
CARRIER, J., J. MUSICK & M. HEITHAUS. 2004. Biology of the sharks and
their relatives. USA. CRC Press. 596 p.
CASTAÑEDA, D. y A. SANDOVAL. 2004. Hábitos alimentarios de dos especies
de tiburones martillos: Sphyrna lewini (Griffith y Smith, 1838) y
Sphyrna zygaena (Linnaeus, 1758) en el Pacífico Ecuatoriano. Bogotá,
Colombia. Univ. de Bogotá “Jorge Tadeo Lozano”. 83 p.
CASTRO, J. y A. DE LACHICA. 1973. Nuevos registros de peces marinos en la
costa del Pacífico mexicano. Rev. Soc. Mex. Hist. Nat. 34. pg. 147-181.
CLARKE, M. 1962. The identification of cephalopod beaks and their relationship
between beak size and total body weight. United Kingdom. British
Museum of Natural History. Bull. 8 (10). pp. 422-480.
CLARKE, M. 1986. A Handbook for the Identification of Cephalopod Beaks.
Plymouth, United Kingdom. Clarendon Press. Marine Biological
Association of the United Kingdom. 55 p.
CLOTHIER, C. 1950. A Key to Some Southern California Fishes Based on
Vertebral Characters. California, USA. Depart. of Natural Res. Division of
Fish and Game. Bureau of Marine Fisheries. Fishery Bulletin 79. 83 p.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
94
CLOTHIER, C. y J. BAXTER. 1969. Vertebral Characters of Some Californian
Fishes with Notes on other Eastern Pacific Species. California, USA. The
Resources Agency. Department of Fish and Game. Marine Resources
Operations. 256 p.
COBO, M. y S. MASSAY. 1969. Lista de Peces Marinos del Ecuador.
Guayaquil, Ecuador. INP. Bol. Cient. y Téc. 2 (1). 68 p.
COHEN, D., T. INADA, T. IWAMOTO & N. SCIALABBA, 1990.
FAO Species catalogue. Gadiform fishes of the World (Order
Gadiformes). An annotated and illustrated catalogue of cods, hakes,
grenadiers and other Gadiform fishes known to date. Rome, Italy. FAO
Fisheries Synopsis. 10 (125). pp. 336-337.
COMPAGNO, L. 1984. Species Catalogue the Shark of the World. An
annotated and illustrated catalogue of shark species known to date.
Rome, Italy. FAO. 4. (1-2). pp. 228-231.
_______________. 1992. Tipos de Tiburones. Barcelona, España. Tiburones.
Gasca & Asociados. Colección Materia Viva. 2a. Edic. pg. 18, 28.
_______________. 2001. FAO Species catalogue the sharks of the world. An
annotated and illustrated catalogue of shark species known to date.
Rome, Italy. FAO. 2. 600-820 p.
COMPAGNO, L., M. DANDO & S. FOWLER. 2005. A field guide to the sharks
of the World. United Kingdom. Collins Publisher Ltd. pp. 179 -180.
COMPAGNO, L., F. KRUPP y W. SCHNEIDER. 1995. Tiburones. Roma, Italia.
Guía FAO para la Identificación de Especies para los Fines de la Pesca.
Pacífico Centro Oriental (Fischer, Krupp, Schneider, Sommer, Carpenter y
Niem, eds.). FAO. 2 Vertebrados (1). pg. 648-661.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
95
CORTÉS, E. 1999. Standardized diet compositions and trophic levels of sharks.
ICES Journal of Marine Sciences (56). pp. 707-717.
CHEN, L. 1997. Reproductive biology of the bigeye thresher shark, Alopias
superciliosus (Lowe 1839) (Choncrichthyes: Alopiidae) in the
northwestern Pacific. Ichthyologist Research. 44 (3) pp. 227-235.
CHIRICHIGNO, N. 1980. Clave para identificar los peces marinos del Perú.
Lima, Perú. Instituto del Mar del Perú (IMARPE). Informe (44). 388 p.
DORA, L. 1999. Desembarques de la pesca artesanal en ocho puertos de la
costa continental ecuatoriana durante el primer trimestre de 1999.
Guayaquil, Ecuador. INP. Bol. Inf. Téc. pg. 29, 33.
DUARTE, C. y D. VON SCHILLER. 1997. Comunidad de peces demersales del
Golfo de Salamanca (Caribe Colombiano): estructura espacio-temporal y
caracterización trófica con énfasis en los hábitos alimentarios de
Lutjanus analis (Cuvier, 1828), Lutjanis sinagris (Linneaus, 1758),
Balistes capricus (Gmelin, 1788) y Balistes vetula (Linneaus, 1758).
Santa Marta, Colombia. Universidad de Bogotá “Jorge Tadeo Lozano”.
Tesis de grado.
ESTUPIÑÁN, C. y L. CEDEÑO. 2005. Estudio de los hábitos alimenticios
mediante el análisis del contenido estomacal del tollo mico Carcharhinus
falciformis (Bibron, 1839) y los tiburones martillo Sphyrna lewini (Griffith
& Smith 1834) y Sphyrna zygaena (Linnaeus 1758) en el área de Manta
– Ecuador. Manta, Ecuador. Universidad Laica “Eloy Alfaro” de Manabí.
Tesis de grado. 100 p.
FISCHER, W., F. KRUPP, W. SCHNEIDER, C. SOMMER, K. CARPENTER y V.
NIEM. 1995. Guía FAO para la Identificación de Especies para los Fines
de la Pesca. Pacífico Centro Oriental. Roma, Italia. 2-3 Vertebrados (1-
2). pg. 799-1813.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
96
GALVÁN, F., H. NIENHUIS & P. KLIMLEY. 1989. Seasonal abundance and
feeding habits of shark of the lower Gulf of California, Mexico. California,
USA. California Fish & Game 75. pp. 74-84.
GARCÍA-GODOS, I. 2001. Patrones morfológicos del otolito sagitta de algunos
peces óseos del mar peruano. Callao, Perú. Instituto del Mar del Perú
(IMARPE). 20 (1-2). pg. 4-83.
GODSIL, H. & R. BYERS. 1944. A systematic study of the Pacific Tunas.
California, USA. Department of Natural Resources. Division of Fish and
Games. Fish Bulletin (60). pp. 118-122.
GRUBER, S. 1977. The visual system of sharks: Adaptations and capability.
Amer. Zool. (17). pp. 453 – 469.
GRUBER, S. y A. MYRBERG. 1977. Approaches to the study of the behavior of
sharks. Amer. Zool. 17. pp. 471 – 486.
HARVEY, C. & J. KITCHEL. 2000. A stable isotope evaluation of structure and
spatial heterogeneity of a Lake Superior food web. Canada. J. Fish and
Aqua. Sci. 57. pp. 1395-1403.
HERDSON, D., W. RODRÍGUEZ y J. MARTÍNEZ. 1985. Las pesquerías
artesanales da la costa del Ecuador y sus capturas en el año 1982.
Guayaquil, Ecuador. INP. Bol. Cient. y Técn. 7 (4). pp. 35 – 50.
HORN, H. 1966. Measurement of overlap in comparative ecological studies.
Am. Nat. (100). pp. 419-424.
HYSLOP, E. 1980. Stomach contents analysis – a review of methods and their
application. J. Fishery Biology. (17). pp. 411-429.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
97
INGRID, L., K. IVERSON & L. PINKAS. 1971. A pictorial guide to beaks of
certain eastern pacific cephalopods. California, USA. Marine Resources
Region California. Department of Fish and Game. Fish. Bull. 152. pp. 7-35.
JIMÉNEZ, P. y P. BÉAREZ. 2004. Peces marinos del Ecuador continental/
Marine fishes of continental Ecuador. Quito, Ecuador.
SIMBIOE/NAZCA/IFEA. Tomo 1-2. 130 & 330 p.
KREBS, C. 1985. Ecología: Estudio de la distribución y la abundancia. México.
2a. Edición. pg. 753.
KREBS, C. 1999. Ecological Methodology. England. Addison Wesley Lonman,
Inc. 620 p.
LABROPOULOU, M. & A. ELEFTHERIOU. 1997. The foraging ecology of two
pairs of congeneric demersal fish species: importance of morphological
characteristics in prey selection. Fish. Biol. 50. pp. 324 - 340.
LAEVASTU, T. 1971. Manual de métodos de Biología Pesquera. Zaragoza,
España. Editorial Acribia y FAO. pg. 216 – 222.
LIU, K., P. CHIANG & C. CHEN. 1998. Age and growth estimates of the bigeye
thresher shark, Alopias superciliosus, in north-eastern waters. Keelung,
Taiwan. Department of Fishery. Taiwan Ocean Science National
University. Fishery Bulletin. 96. pp. 482 - 491.
MARÍN DE LOPEZ, C., L. ARRIAGA y F. ORMAZA. 1999. Estadísticas de los
desembarques pesqueros en Ecuador 1985 -1997. Guayaquil, Ecuador.
INP. 151 p.
MARKAIDA, U. & O. SOSA-NISHIZAKI. 2003. Food and feeding habits of jumbo
squid Dosidicus gigas (Cephalopoda: Ommastrephiidae) from the Gulf of
California, Mexico. United Kingdom. Journal of the Marine Biological
Association of the United Kingdom. 83. 16 p.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
98
MARTINEZ, J., M. COBEÑA y C. DOMINGUEZ. 1997. Las Pesquerías
Artesanales en Puerto López (Provincia de Manabí) durante 1996.
Guayaquil, Ecuador. Área de Pesca Artesanal del programa de
Cooperación Técnica para la Pesca Unión Europea – VECEP ALA
92/43. Inf. Téc. PESCART 03/97. pg. 1, 10 – 11.
MASSAY, S. 1983. Lista de peces marinos del Ecuador. Guayaquil, Ecuador.
INP. Bol. Cient. y Téc. 6 (1). pg. 2, 23.
MASSAY, S. y J. MASSAY. 1999. Peces marinos del Ecuador. Guayaquil,
Ecuador. INP. Bol. Cient. y Téc. 17 (9). pg. 3, 54.
MORA, E. 2005. El recurso tiburón en la pesca artesanal de la costa continental
ecuatoriana. Guayaquil, Ecuador. Instituto Nacional de Pesca. 12 p.
MORENO, J. & J. MORON. 1992. Reproductive biology of the bigeye thresher
shark, Alopias superciliosus (Lowe 1839). Fresh Water Res. 43. pp. 77-
86.
MORISITA, M. 1959. Measuring interspecific association and similarity between
communities. Kyushu, Japan. Ser. Ed. (Biol.) Mem. Faculty Science,
Kyushu University. 65 p.
MOTEKI, M., M. ARAI, K. TSUCHIYA & H. OKAMOTO. 2001. Composition of
piscine prey in the diet of large pelagic fish in the eastern tropical Pacific
Oceanic Fisheries Science. 67 (6). pp. 1063.
ORCÉS, G. 1959. Contribuciones al conocimiento de los peces marinos del
Ecuador. Quito, Ecuador. Anales de la Universidad Central de Quito
87(348). pg. 109 -160.
OTAKE, T. y K. MIZUE. 1981. Direct evidence for oophagy in thresher shark
Alopias pelagicus. Journal Ichthyology. 28. pp. 171-172.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
99
PACHECO, L., J. BERMEO, J. POZO, T. AGUIRRE y M. OVIEDO. 1996.
Proyecto: Exploración y Evaluación de los desembarques de los
calamares gigantes (Dosidicus gigas) en aguas continentales costeras
ecuatorianas, para establecer la factibilidad de pesca a nivel artesanal
(Segunda etapa, 1996). Guayaquil, Ecuador. INP. pg. 3 .
PAULY, D., V. CHRISTENSEN, J. DALSGAARD, R. FROESE & F. TORRES
Jr. 1998. Fishing Down Marine Food Webs. Science. 279. pg. 860 - 863.
PELÁEZ, M. 1997. Hábitos alimenticios de la cabrilla sardinera Mycteroperca
rosacea Streets 1877 (Pisces: Serranidae) en la Bahía de La Paz BCS, y
las zonas adyacentes. Baja California Sur, México. Tesis de
Licenciatura. Universidad Autónoma de Baja California Sur. pg. 62.
PERALTA, M. 1999. Desembarques de la pesca artesanal en ocho puertos de
la costa continental ecuatoriana durante el primer trimestre de 1999.
Guayaquil, Ecuador. INP. Informe Técnico. pg. 6,10.
PIELOU, E. 1966. The measurement of diversity in different types of biological
collection. 13. pp. 131-144.
PINKAS, L., S. OLIPHANT & I. IVERSON. 1971. Food habits of albacore,
bluefin tuna, and bonito in California waters. Fish. Bulletin. 152. pp. 105.
PRETI, A. & S. SMITH. 2001. Feeding habits of the common thresher shark
(Alopias vulpinus) sampled from the California - based drift net fishery,
1998 - 1999. California, USA. California Department of Fish and Game.
CALCOFI Rep. 42. pp. 145-152.
REVELO, W. y M. GUZMÁN, 1997. Estudio sobre la gestión de la pesca
artesanal en regiones pobres: La pesca artesanal en Santa Rosa de
Salinas – Provincia del Guayas. Guayaquil, Ecuador. FAO. 44 p.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
100
REVELO, W. y M. HERRERA, 1999. Desembarques de la pesca artesanal en
ocho puertos de la costa continental ecuatoriana durante 1998.
Guayaquil, Ecuador. INP. Inf. Técn. pg. 23.
ROPER, C., M. SWEENEY & F. HOCHBERG. 1995. Cefalópodos. Roma, Italia.
Guía FAO para la Identificación de Especies para los Fines de la Pesca.
Pacífico Centro Oriental (Fischer, Krupp, Schneider, Sommer, Carpenter y
Niem, eds.). FAO. Invertebrados (3). pg. 325-335.
SHANNON, E. & C. WEAVER. 1963. The mathematical theory of
communication. Illinois, USA. University of Illinois Press Urbana. 119 p.
SMITH, P. y M. ZARET. 1982. Bias in estimating niche overlap. Ecology. 63
(5). pp. 1248-1253.
SOLÍS, P. 1998. Diagnóstico de la actividad pesquera artesanal en el puerto de
Santa Rosa, Provincia del Guayas. Guayaquil, Ecuador. INP. Bol. Cient.
y Téc. 16 (1). 55 p.
STEVENS, J. 1992. Biología de los Tiburones. Barcelona, España. Tiburones.
Gasca & Asociados. Colección Materia Viva. 2a. edición. pg. 53, 69 – 70.
VILLÓN, C. y F. AGUILAR. 2005. Registro de la Presencia de Tiburón Zorro
(Alopias pelagicus) en Aguas Ecuatorianas. Guayaquil, Ecuador. INP.
Informe técnico, IRBA 2 -86-02. 4 p.
WILLIAMS, P. 1997. Shark and related species catch in tuna fisheries of the
Tropical Western and Central Pacific Ocean. New Caledonia. Oceanic
Fisheries Programme South Pacific Commission. FAO in preparation for
the FAO Technical Working Group meeting on the Conservation and
Management of Sharks, held in Tokyo, Japan, 23rd–27th April 1998. 21
p.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
101
WOLFF, G. 1982. A beak key for eight eastern tropical Pacific cephalopods
species, with relationship between their beak dimension and size. Fish
Bulletin. 80 (2) pp. 357-370.
WOLFF, G. 1984. Identification and estimation of size from beaks of eighteen
species of cephalopods from the Pacific Ocean. NOAA Technical Report
NMFS. 17. 50 p.
ZARATE, P. 2002. Tiburones. Galápagos, Ecuador. Reserva Marina de
Galápagos. Línea Base de la Biodiversidad (Danulat y Edgar, eds.).
Fundación Charles Darwin, Servicio Parque Nacional Galápagos. pp.
374-375.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
102
REFERENCIAS ELECTRÓNICAS
GODKNECHT, A. 2004. Alopias pelagicus. On line: http://www.<shark.ch/cgi-
bin/Sharks/spec_conv.pl?E+Alopias.pelagicus.com> [2005].
SEITZ, J. 2003. Pelagic Thresher. Florida, U.S.A. In: Biological Profiles.
Dickinson Hall Ichthyology Dept. Florida Museum of Natural History.
University of Florida. Gainesville. On line: http://www.<flmnh.ufl.edu/
fish/Gallery/Descript/PelagicThresher/PelagicThresher.html> [2004].
TREJO, T. 2005. Global Phylogeography of Thresher Sharks (Alopias spp.)
Inferred from Mitochondrial DNA Control Region Sequences. California,
USA. Masters of Science Thesis. Moss Landing Marine Laboratories,
California State University, Monterey Bay. On line:
http ://< ichthy.mlml.calstate.edu/ebert_abs.htm>[2006].
VILLAVICENCIO, C.; G. ESTRADA y C. DOWNTOWN. 1997. Primer registro
de Alopias pelagicus en el Golfo de California, México. México. In:
Revista de Biología Tropical 44 (3)/ 45 (1), pg. 688 – 689. En línea: http:
//www.<rbt.ots.ac.cr/revistas/44-3y451/villavic.htm>[2005].

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
103
COMUNICACIONES PERSONALES:
BURGESS, G. 2006. Florida Museum of Natural History, University of Florida.
Gainsville, Florida, (USA). Certificado de la especie Alopias pelagicus
para aguas ecuatorianas, extendido como carta de respaldo para la tesis
en mención (07 de marzo del 2006). Guayaquil, Ecuador.
ESTUPIÑAN, C. 2006. Com. pers. Identificación de la especie
Alopias pelagicus para aguas ecuatorianas durante una charla del
Dr. Felipe Galván en Manta en el 2002. Manta , Ecuador.
GALVÁN, F. 2003. Instituto Politécnico Nacional (IPN), Centro Interdisciplinario
de Ciencias Marinas (CICIMAR). La Paz, Baja California Sur, México.
Com. pers. Reporte de la especie Alopias pelagicus para aguas
ecuatorianas durante una conferencia de biología de tiburones realizada
en el Instituto Nacional de Pesca (27 de agosto del 2003). Guayaquil,
Ecuador.
GALVÁN, F. 2004. Instituto Politécnico Nacional (IPN), Centro Interdisciplinario
de Ciencias Marinas (CICIMAR). La Paz, Baja California Sur, México.
Com. pers. Explicación de los métodos para la determinación de los
niveles tróficos de los organismos marinos (11 de noviembre del 2004).
Guayaquil, Ecuador.
GALVÁN, F. 2005. Instituto Politécnico Nacional (IPN), Centro Interdisciplinario
de Ciencias Marinas (CICIMAR). La Paz, Baja California Sur, México.
Com. pers. Certificado de la especie Alopias pelagicus para aguas
ecuatorianas, extendido como carta de respaldo para la tesis en
mención (11 de abril del 2005). Guayaquil, Ecuador.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
104
14. GLOSARIO
Amplitud del nicho trófico.- Rango de selección de la dieta de un depredador
con respecto a un número potencial de tipos de alimentos. De ahí que los
organismos pueden ser especialistas o generalistas.
Depredador.- Organismo que mata a otro para alimentarse de él (Cabrera,
2004).
Especialista.- Organismo que se alimenta de un número bajo de recursos y
presenta alta preferencia por algunos componentes en particular (Cabrera,
2004).
Espectro trófico.- Totalidad de componentes alimenticios de la dieta de un
organismo (Cabrera, 2004). Son las presas que ingiere un organismo (Galván
Com. pers., 2004).
Generalista.- Organismo que se alimenta de una gran variedad de recursos sin
ninguna selección o preferencia (Cabrera, 2004).
Hábito alimenticio.- Estudio detallado que provee una descripción del alimento
que es ingerido recientemente por los organismos con el fin de comprender la
interacción ecológica entre el depredador y la presa, por lo que es preciso
conocer la cantidad de alimento ingerido y la frecuencia alimenticia del
depredador (Harvey & Kitchel, 2000; Carrier, Musick & Hetithaus, 2004).
Nicho.- Papel o función de un organismo en su medio ambiente (Cabrera,
2004).
Nicho trófico.- Relación que tiene un organismo con todos los recursos
alimenticios que se encuentran a su disposición (Cabrera, 2004).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
105
Nivel trófico: Posición en la que se encuentra un determinado organismo en la
cadena trófica, dicha posición está clasificado mediante la manera en que
obtiene su energía (Cabrera, 2004).
Presa.- Organismo que sirve como alimento para otros.
Superposición de dieta.- Sinónimo de traslapamiento trófico.
Traslapamiento trófico.- Determinación de las relaciones interespecíficas o
intraespecíficas que pueden repercutir en competencia para aquellos individuos
que comparten el mismo tiempo y espacio en un ecosistema. Para aquellos
que no comparten el mismo tiempo y espacio determina si los depredadores se
alimentan de presas similares o distintas (Krebs, 1999). Se puede aplicar
también como la similaridad existente entre los componentes de la dieta de dos
organismos pertenecientes a una categoría de la misma especie.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
106
15. ANEXOS Anexo 1. Certificado de la partipación de tesistas Bolaño, Calle y Erazo en el proyecto Biología
de los Elasmobranquios (CICIMAR) e Investigaciones Biológicas de la Fauna
Incidental (CIAT).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
107
Anexo 2. Certificado de la especie Alopias pelagicus para aguas ecuatorianas, extendido
como carta de respaldo para la tesis en mención, por del Dr. Felipe Galván del IPN –
CICIMAR de La Paz, Baja California Sur, México.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
108
Anexo 2. Certificado de la especie Alopias pelagicus para aguas ecuatorianas, extendido como
carta de respaldo para la tesis en mención, por del Dr. Felipe Galván del IPN –
CICIMAR de La Paz, Baja California del Sur, México (Continuación…)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
109
Anexo 3. Registro de la presencia del Tiburón Zorro Alopias pelagicus en Aguas
Ecuatorianas. Villón y Aguilar, 2005 (con permiso de Aguilar).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
110
Anexo 3. Registro de la presencia del Tiburón Zorro Alopias pelagicus en Aguas
Ecuatorianas. Villón y Aguilar, 2005 (con permiso de Aguilar) (Continuación…)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
111
Anexo 3. Registro de la presencia del Tiburón Zorro Alopias pelagicus en Aguas
Ecuatorianas. Villón y Aguilar, 2005 (con permiso de Aguilar) (Continuación…)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
112
Anexo 3. Registro de la presencia del Tiburón Zorro Alopias pelagicus en Aguas
Ecuatorianas. Villón y Aguilar, 2005 (con permiso de Aguilar) (Continuación…)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
113
Anexo 4. Certificado de la especie Alopias pelagicus para aguas ecuatorianas, extendido
como carta de respaldo para la tesis en mención, por del Dr. George H. Burgess del
Museo de Historia Natural de Florida de la Universidad de Florida, Gainsville, Florida,
EE.UU.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2006
114
Anexo 5. Tabla de campo para los muestreos durante mayo – diciembre del 2004.
Simbología:
No. = Número de ejemplar; LT = Longitud total; LP = Longitud Precaudal; % LL = Porcentaje de llenado; LG = Longitud de gonopterigio; Rot = Rotación;
Cal = Grado de calcificación; MC = Marcas de copulación; AC = Abertura cloacal; NE = Número de embriones y LE = Longitud de embriones
No Especie Sexo LT LP % LL LG Rot Cal Rifiodón Semen MC AC NE LE Observación

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
115
Anexo 6. Datos principales de campo para Alopias pelagicus muestreados durante mayo –
diciembre del 2004.
Meses
No. Individuo
Sexo LT (cm) LP (cm) Estado de Madurez Sexual
MAYO 1 Hembra 267 154 Madura
2 Hembra 250 123 Inmadura 3 Hembra 256 135 Inmadura 4 Hembra 270 145 Madura 5 Hembra 290 150 Madura 6 Hembra 270 151 Madura 7 Hembra 273 144 Madura 8 Hembra 155 78 Inmadura 9 Macho 286 163 Maduro 10 Hembra 152 76 Inmadura 11 Hembra 153 78 Inmadura 12 Hembra 257 130 Inmadura 13 Hembra 251 130 Inmadura 14 Macho 229 118 Inmaduro 15 Macho 225 120 Inmaduro 16 Hembra 227 143 Inmadura 17 Hembra 320 180 Madura 18 Hembra 365 156 Madura 19 Hembra 270 145 Madura 20 Hembra 269 184 Madura 21 Hembra 272 183 Madura 22 Hembra 269 140 Madura 23 Hembra 266 130 Madura 24 Hembra 260 137 Madura 25 Hembra 250 134 Inmadura 26 Macho 298 158 Maduro 27 Macho 291 147 Maduro 28 Macho 178 149 Inmaduro 29 Macho 271 143 Maduro 30 Macho 270 145 Maduro 31 Hembra 155 81 Inmadura 32 Macho 251 221 Maduro 33 Macho 266 140 Maduro 34 Macho 267 138 Maduro 35 Macho 265 146 Maduro 36 Hembra 290 120 Madura
AGOSTO 37 Hembra 175 121 Inmadura

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
116
Anexo 6. Datos principales de campo para organismos muestreados durante mayo – diciembre
del 2004 (Continuación…)
Meses
No. Individuo
Sexo LT (cm) LP (cm) Estado de Madurez Sexual
AGOSTO 38 Macho 178.5 94 Inmaduro
39 Macho 192 104 Inmaduro 40 Hembra 173 92 Inmadura
41 Macho 190 100 Inmaduro
SEPTIEMBRE 42 Macho 176 93 Inmaduro 43 Macho 177 94 Inmaduro
OCTUBRE 44 Macho 195 105 Inmaduro
45 Hembra 210 108 Inmadura 46 Macho 207 107 Inmaduro
47 Macho 187 99 Inmaduro
48 Hembra 222 108 Inmadura 49 Hembra 245 139 Inmadura
50 Hembra 210 107 Inmadura
51 Hembra 206 107 Inmadura 52 Hembra 188 99 Inmadura
53 Macho 204 107 Inmaduro
54 Macho 206 105 Inmaduro 55 Macho 257 150 Maduro
56 Macho 202 100 Inmaduro
57 Hembra 289 150 Madura 58 Hembra 262 101 Madura
59 Hembra 226 108 Inmadura
60 Hembra 214 114 Inmadura 61 Hembra 190 90 Inmadura
62 Hembra 172 93 Inmadura
63 Hembra 275 144 Madura 64 Macho 305 161 Maduro
65 Hembra 280 147 Madura
NOVIEMBRE 66 Macho 283 153 Maduro 67 Hembra 308 160 Madura
68 Hembra 320 168 Madura
69 Hembra 322 172 Madura 70 Hembra 309 166 Madura
71 Macho 250 130 Maduro
DICIEMBRE 72 Hembra 213 150 Inmadura 73 Hembra 310 173 Madura
74 Hembra 161 135 Inmadura

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
117
Anexo 6. Datos principales de campo para organismos muestreados durante mayo – diciembre
del 2004 (Continuación…)
Meses
No. Individuo
Sexo LT (cm) LP (cm) Estado de Madurez Sexual
DICIEMBRE 75 Hembra 259 143 Madura
76 Hembra 265 140 Madura 77 Macho 251 140 Maduro
78 Hembra 270 140 Madura
79 Hembra 265 140 Madura 80 Macho 264 142 Maduro
81 Hembra 268 142 Madura
82 Hembra 270 145 Madura 83 Hembra 268 143 Madura
84 Macho 268 145 Maduro
85 Hembra 252 141 Inmadura 86 Hembra 267 138 Madura
87 Hembra 296 156 Madura
88 Macho 310 160 Maduro 89 Hembra 285 150 Madura
90 Macho 258 143 Maduro
91 Hembra 285 150 Madura 92 Hembra 293 154 Madura
93 Macho 268 142 Maduro
94 Hembra 280 149 Madura 95 Hembra 291 148 Madura
96 Macho 273 148 Maduro
97 Hembra 291 140 Madura 98 Hembra 285 148 Madura
99 Hembra 231 124 Inmadura
100 Hembra 210 114 Inmadura 101 Hembra 273 146 Madura
102 Macho 206 111 Inmaduro
103 Hembra 282 142 Madura 104 Hembra 264 142 Madura
105 Hembra 276 143 Madura
106 Hembra 233 125 Inmadura 107 Hembra 236 136 Inmadura
108 Hembra 273 148 Madura
109 Macho 181 98 Inmaduro 110 Hembra 270 157 Madura

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
118
Anexo 7. Diferencias morfológicas entre el tiburón zorro pelágico Alopias pelagicus y el
tiburón zorro común Alopias vulpinus.
El tiburón zorro pelágico A. pelagicus presenta 12 características que difieren enormemente
de su especie similar el tiburón zorro común A. vulpinus. Las descripciones están basadas en
Compagno (1984 y 2001); Compagno et al., (1995) Compagno, Dando and Fowler (2005).
A. pelagicus presenta:
a) Caracteres distintivos:
Ø Un surco horizontal poco evidente a cada lado de la nuca, el cual empieza al nivel de la
boca hasta las aletas pectorales.
Ø El hocico es largo y cónico; el perfil de la frente casi recto y el espacio ínter- orbital
fuertemente convexo.
Ø Surcos labiales rudimentarios o ausentes.
Ø Dientes pequeños de bordes cortantes, dispuestos en más de 29 hileras en cada
mandíbula, con una sola cúspide angosta, casi vertical u oblicua vista distalmente y a
menudo con una cúspide accesoria distal.
Ø Dos aletas dorsales, la primera más corta y con el ápice redondeado.
Ø Aletas pectorales son angostas, largas y casi rectas, no falciformes y de ápices
redondeados o anchos.
Ø Color en la parte dorsal que varía entre azul o gris, el vientre es blanco, con reflejos
plateados; sin embargo el color blanco del vientre no se extiende por encima de las
bases de las aletas pectorales (Fig. 7.1b y c).
b) Talla:
Ø Máxima: Por lo menos 3.3 m de longitud total (hembras adultas) la talla de madurez
para hembras oscila entre 2,8 – 2,9 m de longitud total y para machos entre 2,6 – 2,7 m
de longitud total (Fig. 7.1b y c).
c) Hábitat y Biología:
Ø Es una especie circumtropical, se encuentra desde la superficie hasta 152 m de
profundidad.
Ø Su biología es poco conocida.
Ø Es un organismo ovovivíparo, por lo menos con dos embriones, parece ser un caníbal
uterino.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
119
d) Distribución:
Ø El zorro pelágico sólo se encuentra en los Océanos Pacífico e Índico (Trejo, 2005).
Desde el Golfo de California hasta las I. Galápagos (Godknecht, 2004).
A. vulpinus presenta:
a) Caracteres distintivos:
Ø Sin surcos en la nuca.
Ø El hocico es corto y cónico; el perfil de la frente ampliamente convexa.
Ø Surcos labiales cortos.
Ø Dientes pequeños de bordes cortantes, dispuestos en más de 29 hileras en cada
mandíbula, con una sola cúspide ancha, de borde posterior recto o curvo distalmente y
por lo general sin cúspides accesorias.
Ø Dos aletas dorsales, la primera más larga y con el ápice puntiagudo.
Ø Aletas pectorales muy largas y falciformes, de ápices angostos, estrechamente
redondeados a puntiagudos.
Ø Color dorso y la superficie ventral del hocico marrón, gris, azul grisáceo o negruzco,
flancos más claros y abruptamente claros centralmente: un área blanca se extiende
desde el abdomen hasta por encima de las bases de las aletas pectorales; aletas
pectorales, pélvicas y dorsales negruzcas; ápices de las pectorales y pélvicas y extremos
de la caudal a veces con manchitas blancas (Fig. 7.1a y 6.2).
b) Talla:
Ø Máxima: Entre 5 y 6.09 m de longitud total; común entre 4.3 y 4.9 m; aparentemente de
talla mayor que A. superciliosus y A. pelagicus (Fig. 7.2).
c) Hábitat y Biología:
Ø Es una especie tropical, se encuentra desde la superficie hasta 366 m de profundidad.
Se encuentra en aguas subtropicales y templadas. A veces salta fuera del agua (Fig. 7.3)
Ø Su biología bastante conocida.
Ø Es un organismo ovovivíparo, de 2 a 4 (generalmente 2) embriones por camada,
parece ser un caníbal uterino.
d) Distribución:
Ø Es una especie cosmopolita.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
120
(a)
(b)
(c) Fig. 7.1. Morfología externa de (a) Alopias vulpinus (Fuente: National Marine Fisheries Service In: Field
Key to Sharks of North Carolina and Adjacent Waters), (b) Alopias pelagicus hembra de
294 cm (LT) y (c) de 305 cm (LT) en Santa Rosa de Salinas (Marzo, 2005).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
121
Fig. 7.2. Características de Alopias vulpinus de 4.1 m. (LT) capturado en Dinamarca (In: Trejo, 2005).
(a) (b) Fig. 7.3. Salto fuera del agua de Alopias vulpinus (a) en Bar Harbor, Maine (Leonard, 2002) y (b) (a y b,
In: Trejo, 2005).
e) Dentición:
v A. pelagicus presenta en la mandíbula superior expansiones laterales moderadamente
grandes en sus bases y la unión de la región superior de ésta describe la forma de un
arco. La mandíbula inferior posee pequeñas expansiones laterales en las bases y la
unión de la región inferior es a manera de una W. (Fig. 7.4a).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
122
v Por su parte, A. vulpinus en la mandíbula superior no tiene expansiones laterales en
las bases y la unión de la región superior de ésta describe la forma de una M. La
mandíbula inferior posee grandes expansiones laterales en las bases y la unión de la
región inferior es a manera de una U. (Fig. 7.4b).
(a) (b)
Fig. 7.4. (a) Mandíbula de A. pelagicus de Santa Rosa de Salinas (Noviembre del 2005) y (b) mandíbula
de A. vulpinus de México (Aguilar, enero del 2006).
v A. pelagicus presenta los dientes superiores e inferiores de la región central con una
sola cúspide angosta y oblicua con una cúspide accesoria (Figs. 7.5a y 7.6a).
v A. vulpinus presenta los dientes superiores e inferiores de la región central con una
sola cúspide ancha y recta sin cúspide accesoria (Figs. 7.5b y 7.6b).
(a) (b)
Fig. 7.5. Dientes superiores de la región central la mandíbula de (a) A. pelagicus y (b) A. vulpinus.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
123
(a) (b)
Fig. 7. 6. Dientes inferiores de la región central la mandíbula de (a) A. pelagicus y (b) A. vulpinus.
v A. pelagicus presenta los dientes laterales, superiores e inferiores, con una sola
cúspide angosta y oblicua con una cúspide accesoria (Figs. 7.7a y c).
v A. vulpinus presenta los dientes laterales, superiores e inferiores, con una sola
cúspide ancha y oblicua sin cúspide accesoria (Figs. 7.7b y d).
(a) (b)
(c) (d)
Fig. 7.7. Dientes superiores e inferiores de la región derecha (4º, 5º y 6º) de la mandíbula de (a)
A. pelagicus y (b) A. vulpinus.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
124
(a) (b)
(c) (d)
Fig. 7.8. Tesista con mandíbula de (a-c) Mandíbula de A. pelagicus de Santa Rosa de Salinas
(Tesista, noviembre del 2005) y (b - d) mandíbula de A. vulpinus de México (Aguilar,
enero del 2006).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
125
(a)
(b) Fig. 7.9. Esquema de las principales características morfológicas de (a) A. pelagicus y (b)
A. vulpinus, donde se observa las diferencias entre la región ventral de la cabeza, el
organismo y los dientes.

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
126
Anexo 8. Tabla de registro de laboratorio
Muestra No. _____ Sexo:___________ Fecha:_________
Peso Total (g) Especies encontradas Peso(g) – Peces
Longitud (cm) -Peces
Peso (g) – Restos de Peces (ojos y
otolitos)
Longitud (cm) Restos de Peces
Número de Ojos (peces)
Número de Otolitos (peces)
Peso (g) - Calamar
Longitud (cm) Calamar
Peso (g) - Resto de Calamar (ojos y
picos)
Longitud (cm) Resto de Calamar
Número de Picos (Calamar)
Número de Ojos (Calamar)
Testículo (ancho – largo) Calamar
Diámetro del Ovario
Observaciones

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
127
Anexo 9. Vista panorámica de Santa Rosa de Salinas (Marzo 3 del 2005)
Anexo 10. Crías de Alopias pelagicus (a) machos de 22.5 cm (arriba) y 23 cm (LT) (abajo)
sobre el dorso de una hembra adulta de 290 cm. (LT) y (b) gonopterigios de 3 cm
(LG) (Marzo 3 del 2005).
(a) (b)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
128
Anexo 11. Proceso de evisceración de Alopias pelagicus (a) corte de cabeza; (b) corte de
región ventral; (c) aparato digestivo; (d) aletas y (e) aleta pectoral de 40 cm. LA
(Marzo 3 del 2005).
(a) (b)
(c) (d)
(e)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
129
Anexo 12. Tiburón zorro de anteojos Alopias superciliosus (a) ejemplares desembarcados;
(b) vista frontal donde se observa el surco encefálico y los ojos grandes en un
organismo de 300 cm LT; (c) vista lateral y la disposición de sus aletas; (d) vista
lateral; (e) gonopterigio de 25 cm. (LG) y (f) apertura del rifiodón (Marzo 3 del 2005).
(a) (b)
(c) (d)
(e) (f)

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
130
Anexo 13. Tiburón puntas negras Carcharhinus limbatus de 255 cm LT.
Anexo 14. Tiburón tinto Isurus oxyrhinchus de 250 cm LT (con permiso de Erazo, 2005).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
131
Anexo 15. Boca de Tiburón tinto Isurus oxyrhinchus de 250 cm LT (con permiso de Erazo,
2005).
Anexo 16. Tiburón azul Prionace glauca de 270 cm LT (con permiso de Erazo, 2005).

Espectro trófico de Alopias pelagicus en Santa Rosa Calle, M. 2005
132
Anexo 17. Tiburón tollo Mustelus sp. macho de 60 cm LT.
Anexo 18. Detalle de la cabeza de Dorado Coryphaena hippurus macho de 140 cm LT.
Top Related
Copyright © 2022 FDOKUMEN

