







Bahasa
Halaman
Hukum


Plant Biotechnology Journal
(2005)
3
, pp. 571–582 doi: 10.1111/j.1467-7652.2005.00147.x
© 2005 Blackwell Publishing Ltd
571
Blackwell Publishing, Ltd.Oxford, UKPBIPlant Biotechnology Journal1467-7644© 2005 Blackwell Publishing Ltd? 20052?571Original Article
Root-specific repressor-operator gene complexTehryung Kim et al.
Engineering a root-specific, repressor-operator gene complex
Tehryung Kim
1,
†, Rebecca S. Balish
2,
†, Andrew C. P. Heaton
1
, Elizabeth C. McKinney
1
, Om Parkash Dhankher
3
and Richard B. Meagher
1,
*
1
Department of Genetics, University of Georgia, Athens, GA 30602
2
Department of Microbiology, Miami University, Oxford, OH 45056
3
Department of Plant, Soil, and Insect Science, University of Massachusetts Amherst, Amherst, MA 10062
Summary
Strong, tissue-specific and genetically regulated expression systems are essential tools in
plant biotechnology. An expression system tool called a ‘repressor-operator gene complex’
(ROC) has diverse applications in plant biotechnology fields including phytoremediation,
disease resistance, plant nutrition, food safety, and hybrid seed production. To test this
concept, we assembled a root-specific ROC using a strategy that could be used to construct
almost any gene expression pattern. When a modified
E. coli lac
repressor with a nuclear
localization signal was expressed from a rubisco small subunit expression vector,
S1pt::lacIn
,
LacIn protein was localized to the nuclei of leaf and stem cells, but not to root cells. A LacIn
repressible
Arabidopsis
actin expression vector
A2pot
was assembled containing upstream
bacterial
lacO
operator sequences, and it was tested for organ and tissue specificity using
β
-glucuronidase (
GUS
) and mercuric ion reductase (
merA
) gene reporters. Strong GUS
enzyme expression was restricted to root tissues of
A2pot::GUS/S1pt::lacIn
ROC plants,
while GUS activity was high in all vegetative tissues of plants lacking the repressor.
Repression of shoot GUS expression exceeded 99.9% with no evidence of root repression,
among a large percentage of doubly transformed plants. Similarly, MerA was strongly
expressed in the roots, but not the shoots of
A2pot::merA/S1pt::lacIn
plants, while MerA
levels remained high in both shoots and roots of plants lacking repressor. Plants with MerA
expression restricted to roots were approximately as tolerant to ionic mercury as plants
constitutively expressing MerA in roots and shoots. The superiority of this ROC over the
previously described root-specific tobacco RB7 promoter is demonstrated.
Received 11 November 2004
revised 10 May 2005;
accepted 18 May 2005.
*
Correspondence
(fax 706-542-1387;
e-mail [email protected])
†These two authors contributed equally in
a collaborative effort.
Keywords:
phytoremediation, plant
pathogens, nutrition, resistance, hybrid seed,
gene silencing.
Introduction
Strong, tissue-specific expression systems are essential plant-
biotechnology tools. We devised a strategy for building repressor-
operator gene-complexes (ROCs) that could be used to target
almost any expression pattern and tested it by constructing
a simple root-specific system with wide applications to plant
biotechnology. The general approach to creating a ROC is to
use the restricted expression of one promoter to limit the
expression pattern of a second, more widely expressed
promoter or to repress the expression of a promoter under
particular genetic circumstances. In this manuscript, we
utilized a shoot-specific promoter to limit the expression of a
vegetative constitutive promoter to root expression.
Our root-specific ROC combined the well-characterized
genetic regulatory elements from the
E. coli lac
operon and
two strongly expressed plant genes,
SRS1
and
ACT2
, as out-
lined in Figure 1. The strategy employs a modified
Lac
I repres-
sor protein for the permanent repression in shoots of target
genes expressed from the
ACT2
promoter. First, we assem-
bled a
Lac
I repressible vector,
A2pot
, in which the
ACT2
promoter region was modified to contain wild-type
E. coli lacO
sequences, which are the DNA binding sites for the repressor
.
The wild-type
Arabidopsis ACT2
actin promoter directs strong

572
Tehryung Kim
et al
.
© Blackwell Publishing Ltd,
Plant Biotechnology Journal
(2005),
3
, 571–582
constitutive gene expression in all vegetative shoot, root,
and inflorescence tissues and cell types in young and mature
plants (An
et al
., 1996; Kandasamy
et al
., 2002). Second, we
constructed the
S1pt::lacIn
gene in which a modified
E. coli
Lac
I repressor protein was expressed using
SRS1
regulatory
elements in the vector
S1pt
. Various configurations of the
Glycine max SRS1 ribulose bisphosphate carboxylase (RUBISCO)
small subunit
promoter direct strong light-induced expression
in shoots, but no expression has ever been detected from this
promoter in root tissues, even when assayed with sensitive
reporters (Shirley
et al
., 1987; Dhankher
et al
., 2002). Third,
β
-glucuronidase (
GUS
) and mercuric ion reductase (
merA
)
sequences were cloned into
A2pot
to test the repressible
expression directed by this ROC. Fourth, we demonstrate that
both reporter constructs were strongly expressed in shoots
and roots, except in the presence of
S1pt::lacIn
, which shut
down the
lacO
-modified promoter in shoots, leaving strong
activity in roots.
Results
Constructing a root-specific ROC
The
A2pt
expression vector contains promoter, 5
′
, and 3
′
UTR sequences, and polyadenylation signals from the
ACT2
gene (Kandasamy
et al
., 2002). The
A2pt
sequence
was modified into a
Lac
I repressible expression vector,
A2pot
,
by adding two 25-bp
lacO
operator sequences from the
E. coli lac
operon, as diagrammed in Figure 1. These sequences
were inserted 75 bp apart, one immediately after the
ACT2
TATA box sequence and the other immediately after the
multiple start sites of transcription, and replaced two 25-bp
stretches of native sequence. The two 25-bp regions of
ACT2
native promoter that were replaced are poorly conserved
with the closely related
Arabidopsis ACT8
constitutive actin
promoter sequence and, thus, were predicted to be non-
essential for the strong constitutive expression of
ACT2
(An
et al
., 1996).
The
S1pt
expression vector has promoter, 5
′
, and 3
′
UTR
sequences and polyadenylation signals from the light-induced
shoot-specific
RUBISCO small subunit
gene
SRS1
(Materials
and methods).
S1pt
was used to drive expression of a modi-
fied
E. coli lac
repressor,
LacIn
, in shoots, and hence limit
the expression of
A2pot
to roots, as diagrammed in Figure 1.
The
lacIn
DNA coding sequence was modified from the wild-
type
E. coli
sequence so that it encoded a protein with
the C-terminal eukaryotic nuclear localization signal (NLS),
SSVVHPKKKRKV. The
lacIn
sequence was cloned as an
in-frame translational fusion following the ATG initiation
codon in
S1pt
to make
S1pt::lacIn
(Figure 1).
Immunolocalization of LacIn protein
T
1
generation transgenic plants expressing
S1pt::lacIn
were
obtained by selecting for the linked hygromycin resistance
gene (
hptII
) marker. The tissue specificity of
LacIn
repressor
expression was assayed immuno-histochemically using a
Lac
I-specific antibody in T
2
generation transgenic
Arabidopsis
seedlings and plants
.
LacIn protein was localized to shoot
Figure 1 Construction of a root-specific repressor-operator gene complex based on shoot-repression of a constitutive promoter. (a) The S1pt::lacIn gene encoded a modified bacterial lac repressor that contained a nuclear localization signal (NLS). The lacIn sequence was transcribed from the strong, light-induced, shoot-specific regulatory elements from the SRS1 gene. The A2pot vector used a modified constitutive actin ACT2 promoter regulatory elements that contained two bacterial lacO operator sequences flanking the start site for transcription (ts). A2pot was used to express two target genes of interest, merA and GUS. When S1pt::lacIn was expressed in shoots, the lacIn protein (green) entered the nucleus and repressed A2pot::GUS or A2pot::merA transcription. In roots, where no repressor protein was expressed, and the A2pot::GUS and A2pot::merA constructs were still active. (b) When plants expressing LacIn (green) from the S1pt::lacIn gene in shoots were crossed with those constitutively expressing MerA (blue) from the A2pot::merA gene, the hybrids expressed MerA primarily in roots.

Root-specific repressor-operator gene complex
573
© Blackwell Publishing Ltd,
Plant Biotechnology Journal
(2005),
3
, 571–582
tissues only of
S1pt::lacIn
plants, shown in Figure 2. Compar-
ison of LacIn localization with DNA-specific DAPI staining
revealed that essentially all shoot nuclei contained repressor
protein. LacIn staining was concentrated in the nuclei with
little staining detected in the cytoplasm, suggesting the
composite NLS was functional in directing efficient nuclear
transport. LacIn was found in the nuclei of essentially all leaf
cells from seedlings and older plants. No LacIn protein was
detected in the nuclei or cytoplasm of root cells from
S1pt::lacIn
plants (Figure 2c), demonstrating the specificity of
the expression in shoot tissues. Furthermore, no immuno-
staining was observed in the shoots or roots of wild-type
plants (Figure 2b,d), demonstrating the specificity of the
immunolocalization assay.
Expression pattern of a GUS reporter with root-specific
ROC
The
β
-glucuronidase (GUS) reporter gene sequence was
cloned under control of A2pot sequences to test the tissue
specificity of the ROC system outlined in Figure 1. T1 genera-
tion transgenic plants were obtained by selecting for the
linked kanamycin resistance gene (nptII ). A2pot::GUS expres-
sion was examined in T2 generation transgenic seedlings and
plants. Histochemical staining for GUS activity showed that
A2pot::GUS was highly expressed in all vegetative tissues, as
shown at the top of Figure 3. A closer examination of these
vegetative tissues revealed strong expression in all cell types,
as reported previously for wild-type ACT2 promoter-GUS
fusions stained for 24 h (An et al., 1996). Longer staining of
the A2pot::GUS plants made the vegetative organs and
tissues opaque blue with significant blue pigment leaching
into the medium, further demonstrating the strength of GUS
enzyme expression. After 2 or 24 h of staining with substrate,
no significant blue GUS precipitate was observed in pollen,
ovules, or developing seeds (not shown), consistent with the
lack of expression of wild-type ACT2 promoter-GUS fusions
in reproductive tissues (An et al., 1996). Thus, it does
not appear that insertion of the lacO sequences in A2pot
significantly altered the strong, vegetative expression pattern
from that of the wild-type ACT2 promoter. Fifty of 50 T1
generation plants transformed with this construct showed
the same expression pattern, with a several-fold variation
in the levels of expression among the various lines.
Plants expressing both the A2pot::GUS and S1pt::lacIn
genes were obtained by cotransformation of the two con-
structs and coselection for the growth of seedlings expressing
both nptII and hptII markers, respectively. The majority of the
transgenic plant lines demonstrating resistance to both drugs
showed little or no GUS expression in shoot tissues, while
root expression was already quite strong in these plants after
2 h of histochemical staining. A typical doubly transformed
plant is shown in Figure 3c,d. Cross sections of histochemically
stained root confirmed that all tissue and cell types in roots
exhibited strong GUS activity, including root hairs, epidermis,
cortex, vascular cylinder (Figure 3g,h), and root tip (not shown),
consistent with the known root expression pattern of the
wild-type actin ACT2 promoter (An et al., 1996). Root stain-
ing was indistinguishable from that in A2pot::GUS plants
lacking the repressor. Among this majority of plants showing
Figure 2 Expression of the LacIn protein in nuclei. Subcellular localization of LacIn in shoot (a, b) and root (c, d) cells of transgenic plants expressing S1pt::lacIn. The LacIn protein was localized in dissociated and fixed cells using the commercial anti-LacI rabbit polyclonal antibody (#217449 Stratatene) and FITC-goat-anti-rabbit antisera (Sigma) (left panel, a–d). LacIn protein was detected in the nucleus of the leaf (a, left), but not root (c, left) cells of S1pt::lacIn plants and was not detected in wild-type plants (b, d, left). Cells were costained with DAPI to localize DNA and nuclei (right panel a-d). Young leaves and roots of T2 generation plants were examined in these experiments.

574 Tehryung Kim et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
root-specific expression, occasional weak staining was observed
in cotyledons and at the distal margins of the shoots of newly
emerging leaves (not shown). Nontransgenic control plants
showed no GUS staining of shoots or roots (Figure 3e,f).
In order to quantify both expression and repression levels
we examined GUS activity in the roots and shoots of a
population of independently transformed plant lines using the
fluorescent substrate MUG that can be quantified over a wide
dynamic range. We compared the data for 60 T1 generation
plant lines transformed with both A2pot::GUS + S1pt::lacIn
to data from 15 T1 plants transformed with A2pot::GUS alone.
The 15 A2pot::GUS plants lacking repressor had shoot fluo-
rescence levels that averaged 150% of that in roots when data
were normalized for fresh shoot and root weights (Figure 4).
Figure 3 The root-specific expression of β-glucuronidase from the A2pot::GUS sequence in transgenic Arabidopsis expressing the LacIn repressor in leaves. Seedlings transformed with the A2pot::GUS gene alone (a and b); seedlings cotransformed with both A2pot::GUS and S1pt::lacIn (c and d); and wild-type seedlings were examined for GUS enzyme expression. GUS expression in shoots (a) was knocked down by the expression of S1pt::lacIn in shoots (c), but not in roots (d). Cross section of root tissue confirmed that all types of cells in root tissue exhibited GUS activity (d).

Root-specific repressor-operator gene complex 575
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
The doubly transformed plants showed a wide range of
fluorescence phenotypes from strong to weak repression of
shoot activity by LacIn. Therefore, we binned these plants
into three groups: those showing strong, intermediate, or
weak shoot repression based on the ratios of shoot to root
fluorescence (Figure 4a). Among the 60 cotransformants, the
17 plants with strong shoot repression showed shoot expres-
sion levels that averaged much less than 0.2% of root values
(Figure 4b). In other words, the average root levels were more
than 500 times higher than shoot levels. Some of these plants
showed root GUS expression with 1000- to 5000-fold lower
shoot expression levels as shown in Figure 4b for the two
plants in this group with the highest and lowest levels of total
root GUS activity. The 18 intermediate plants showed shoot
activity that was 10% to 15% of the root expression levels. The
25 plants in the weakly repressed group showed shoot levels
of GUS expression that were equal to the root levels and,
thus, only slightly repressed relative to control plants lacking
repressor. While the observed fluorescence values for plants
from each group varied widely (Figure 4b), the absolute levels
of root GUS activity did not appear to correlate with the pres-
ence or absence of the repressor construct S1pt::lacIn. This
suggested LacIn repressor was never expressed in sufficient
quantities in roots to repress root GUS expression. Stably
transformed T2 plant lines maintained similar levels of shoot
repression and strong root expression as shown for a plant
with strong repression in Figure 3c,d.
Root-specific expression of a merA reporter
We assembled a second reporter construct, A2pot::merA,
designed to express a modified E. coli mercuric ion reductase
merA gene that could be repressed by LacIn (see Materials
and methods). Plants were independently transformed with
S1pt::lacIn and A2pot::merA and lines were identified that
contained single T-DNA insertions based on segregation
analysis of T2 generation plants. Ten independent plant lines
with high levels of LacIn or MerA protein expression in shoots
were identified by Western blot analysis for protein expres-
sion. Representative plants with single A2pot::merA trans-
gene insertions are shown in Figure 5 (see lanes labelled
S1pt::lacIn-5, A2pot::merA-1, A2pot::merA-5, A2pot::merA-
10). These single insertion lines and their genetic hybrids with
S1pt::lacIn-5 were examined for organ-specific MerA protein
expression. The shoots and roots of the three lines expressing
A2pot::merA alone showed high levels of MerA protein. In
contrast, MerA protein expression was significantly repressed
in shoots of A2pot::merA × S1pt::lacIn hybrid plants. The low
levels of MerA expression in shoots corresponded with LacIn
Figure 4 Quantification of relative β-glucuronidase shoot and root activities. Fluorometric assays were used to quantify β-glucuronidase (MUG) activity in shoots and roots of T1 generation plants cotransformed with both A2pot::GUS and S1pt::LacIn and control plants containing the A2pot::GUS construct alone. A. Shoot fluorescence is expressed as a percent of root fluorescence of four-week-old plants. The 60 doubly transformed plants were grouped into those showing strong (99–100%), intermediate (70–90%), or weak repression (0–65%). Average root levels were defined as 100%. Standard errors are shown for shoot data. B. Shoot and root MUG fluorescence values are shown for the two individual plants with the weakest and strongest root MUG activity from each group in panel A along with the average values for each group. These data demonstrate the wide range of fluorescent activity obtained in populations of transformed plant lines. A & B. All GUS data were normalized to fresh shoot or root weights.

576 Tehryung Kim et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
protein expression from the S1pt::lacIn-5 parent repressor
line.
These Western results were quantified as summarized in
Figure 5f. The levels of shoot-specific MerA expression were
repressed more than 20-fold in some lines, when crossed
to the S1pt::lacIn-5 repressor line. For example, in the
S1pt::lacIn-5 × A2pot::merA-1 hybrid the levels of MerA were
not detectable over background as compared to the strong
MerA expression observed in the A2pot::merA-1 parent line.
Other lines like S1pt::lacIn-5 × A2pot::merA-10 were repressed
in shoots sevenfold below the A2pot::merA-10 parent line.
The average repression of MerA in shoots, among the three
shoot samples shown in frame a, was approximately nine-
fold. The limitation in quantifying repression in these experi-
ments is that shoot MerA levels below 5% of that shown for
the strong A2pot::merA lines in Figure 5 cannot be detected
on Western blot. Thus, the dynamic range of these measure-
ments, approximately 20-fold, underestimated the levels of
MerA repression in shoots. These levels of repression were
reproducible for each line in repetitions of this experiment.
Hybrid plants were examined through the T3 generation and
still showed the same strong, root-specific expression pat-
tern, suggesting this root-specific ROC is genetically stable.
The hybrid plants expressing MerA protein predominantly
in roots (A2pot::merA × S1pt::lacIn) were plated on media
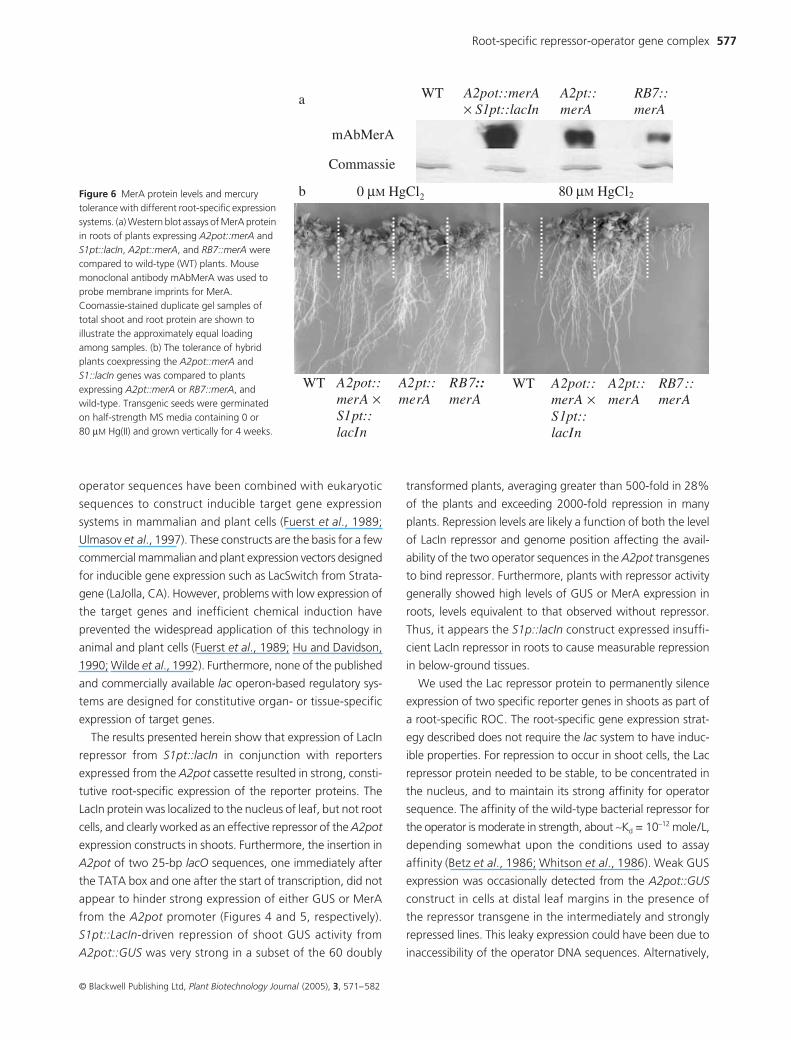
with 80 µM Hg(II) (Figure 6). These plants were as tolerant to
Hg(II) as were plants expressing MerA from the A2pot::merA
construct alone without LacIn repression. As expected from
previous studies, wild-type seeds and seedlings die on these
same Hg(II) concentrations. Thus, it appears that detoxifica-
tion of Hg(II) to Hg(0) in roots is sufficient to confer resistance
to above-ground portions of the plant.
ROC controlled MerA expression was compared with that
from the tobacco RB7 root-specific gene expression system
(Yamamoto et al., 1991). Twenty independent transgenic
Arabidopsis seedling expressing the RB7::merA fusion were
made and the most Hg(II) resistant plant line was selected
for further study. MerA expression levels in roots from ROC
system were approximately fivefold higher compared to those
with RB7::merA construct (Figure 6a). Root growth on the
Hg(II) containing medium was much higher for S1pt::lacIn
× A2pot::merA plants with than for the RB7::merA plants,
which barely grew on the mercury-containing medium
(Figure 6b).
Discussion
ROCs could be built using repressor proteins and operator
DNA binding sites from several known systems of gene
regulation. The lac operon is one of the best-characterized reg-
ulatory systems and was among the first discovered in E. coli
(Pardee et al., 1958; Jacob and Monod, 1959). The tetrameric
LacI repressor protein binds naturally to a lacO operator DNA
sequence in the E. coli lac promoter region, repressing tran-
scription of downstream genes for lactose utilization. Addi-
tion of a small molecule inducer releases the repressor and
activates transcription. The E. coli lacI repressor and lacO
Figure 5 Assays of root-specific MerA protein expression: Western blot assays of MerA protein in roots and shoots of plants expressing A2pot::merA and S1pt::lacIn were compared to plants expressing either gene alone and wild-type plants. The complementary expression of LacIn in shoots is shown for comparison. Mouse monoclonal antibodies were used to probe Western blots for MerA (a & b) and LacIn (e). Immunoreactions were enhanced and images quantified as described in Materials and methods. Coomassie-stained samples of total shoot and root protein are shown to illustrate the approximately equal loading among samples (c & d). All lanes of the polyacrylamide gels were loaded with 20 µg of total protein. The levels of MerA among transgenic lines lacking repressor, A2pot::merA-1, A2pot::merA-5, and A2pot::merA-10 varied by about 30%, being 58.6, 77.6, and 80.6 density units, respectively. f. Relative levels of MerA protein expression with and without the S1pt::LacIn gene. The levels of MerA protein in each line were normalized to the levels of MerA in the roots of the parent lines lacking LacIn. The data for the three sets of plant lines examined above (a & b) were combined to give standard errors. Image J was used to quantify the intensity of bands from Western blots (http://rsb.info.nih.gov/ig/).
Root-specific repressor-operator gene complex 577
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
operator sequences have been combined with eukaryotic
sequences to construct inducible target gene expression
systems in mammalian and plant cells (Fuerst et al., 1989;
Ulmasov et al., 1997). These constructs are the basis for a few
commercial mammalian and plant expression vectors designed
for inducible gene expression such as LacSwitch from Strata-
gene (LaJolla, CA). However, problems with low expression of
the target genes and inefficient chemical induction have
prevented the widespread application of this technology in
animal and plant cells (Fuerst et al., 1989; Hu and Davidson,
1990; Wilde et al., 1992). Furthermore, none of the published
and commercially available lac operon-based regulatory sys-
tems are designed for constitutive organ- or tissue-specific
expression of target genes.
The results presented herein show that expression of LacIn
repressor from S1pt::lacIn in conjunction with reporters
expressed from the A2pot cassette resulted in strong, consti-
tutive root-specific expression of the reporter proteins. The
LacIn protein was localized to the nucleus of leaf, but not root
cells, and clearly worked as an effective repressor of the A2pot
expression constructs in shoots. Furthermore, the insertion in
A2pot of two 25-bp lacO sequences, one immediately after
the TATA box and one after the start of transcription, did not
appear to hinder strong expression of either GUS or MerA
from the A2pot promoter (Figures 4 and 5, respectively).
S1pt::LacIn-driven repression of shoot GUS activity from
A2pot::GUS was very strong in a subset of the 60 doubly
transformed plants, averaging greater than 500-fold in 28%
of the plants and exceeding 2000-fold repression in many
plants. Repression levels are likely a function of both the level
of LacIn repressor and genome position affecting the avail-
ability of the two operator sequences in the A2pot transgenes
to bind repressor. Furthermore, plants with repressor activity
generally showed high levels of GUS or MerA expression in
roots, levels equivalent to that observed without repressor.
Thus, it appears the S1p::lacIn construct expressed insuffi-
cient LacIn repressor in roots to cause measurable repression
in below-ground tissues.
We used the Lac repressor protein to permanently silence
expression of two specific reporter genes in shoots as part of
a root-specific ROC. The root-specific gene expression strat-
egy described does not require the lac system to have induc-
ible properties. For repression to occur in shoot cells, the Lac
repressor protein needed to be stable, to be concentrated in
the nucleus, and to maintain its strong affinity for operator
sequence. The affinity of the wild-type bacterial repressor for
the operator is moderate in strength, about ∼Kd = 10−12 mole/L,
depending somewhat upon the conditions used to assay
affinity (Betz et al., 1986; Whitson et al., 1986). Weak GUS
expression was occasionally detected from the A2pot::GUS
construct in cells at distal leaf margins in the presence of
the repressor transgene in the intermediately and strongly
repressed lines. This leaky expression could have been due to
inaccessibility of the operator DNA sequences. Alternatively,
Figure 6 MerA protein levels and mercury tolerance with different root-specific expression systems. (a) Western blot assays of MerA protein in roots of plants expressing A2pot::merA and S1pt::lacIn, A2pt::merA, and RB7::merA were compared to wild-type (WT) plants. Mouse monoclonal antibody mAbMerA was used to probe membrane imprints for MerA. Coomassie-stained duplicate gel samples of total shoot and root protein are shown to illustrate the approximately equal loading among samples. (b) The tolerance of hybrid plants coexpressing the A2pot::merA and S1::lacIn genes was compared to plants expressing A2pt::merA or RB7::merA, and wild-type. Transgenic seeds were germinated on half-strength MS media containing 0 or 80 µM Hg(II) and grown vertically for 4 weeks.

578 Tehryung Kim et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
it may have been due to reduced transcription of S1pt::lacIn
gene or more rapid turnover of the repressor RNA or protein,
or some combination of these effects occurring in some small
number of cells. Looking toward improving this system fur-
ther, it is likely that tighter binding to lacO sequences of a fur-
ther modified LacIn protein, should lower the concentration
of repressor required for complete repression and, thus, lead
to more efficient repression in shoots. Mutant LacI repressor
proteins with 10 000-fold higher affinities for operator
sequences and mutant operator DNA sequences with 30-
fold higher affinities for repressor have been reported in
bacteria (Pfahl, 1979; Betz et al., 1986; Falcon and Matthews,
2000). Thus, there are many options for further improving ROC
repression stringency using elements from the lac operon.
The root-specific ROC developed has a wide potential
application, because strong, constitutive, root-specific pro-
moters are essential to many areas of plant biotechnology,
including enhanced plant nutrition, control of plant root dis-
eases, food safety, and phytoremediation of contaminated
soils. For example, the enhanced uptake of essential nutrients
such as iron, zinc, potassium, and/or nitrate into plant roots
should enhance plant yield and the nutritional quality of
foods (Guerinot and Eide, 1999; Guerinot, 2001). Root-specific
protection from pathogens would improve crop and food safety
by both restricting expression of antifungal and antinematode
agents to roots and preventing the synthesis of native toxins
by these and associated pathogens. Numerous fungal path-
ogens target roots, such as those causing sudden death syn-
drome in soybean, take-all disease in wheat, and wilt diseases
in vegetable crops (Iqbal et al., 2002; Anjaiah et al., 2003;
Gutteridge et al., 2003; Lievens et al., 2003). Nematodes nearly
all target roots and several diseases such as soybean cyst
nematode (Lewers et al., 2002) and nematode root-knot
(Giblin-Davis et al., 2003) cause significant annual loss of
crops to US agriculture. Targeting the synthesis of fungal-
specific and nematode-specific toxins with the root-specific
ROC described herein should improve the yield and quality of
many food, forage, and animal feed crops (Gao et al., 2000).
Another application with profound health and environmen-
tal implications is the phytoremediation of toxic pollutants in
the soil. Among the genes that need to be enhanced for root
expression are those encoding catabolic enzymes that degrade
organic pollutants, plasma membrane transporters that uptake
pollutants, transporters involved in the short-distance move-
ment of pollutants from root-epidermal cells to the root vas-
cular system, and enzymes involved in the transformation of
elemental pollutants like mercury and arsenic to less toxic
chemical species (Meagher, 2000). A key to many engineered
phytoremediation strategies is plant tolerance to toxins once
they enter plant cells (Salt et al., 1995). The root-specific
expression of MerA provided high levels of mercury tolerance
to transgenic plants. Based on data presented herein, we
have sufficient root expression of MerA in various root tissues
to protect these tissues from ionic mercury’s toxic effects by
converting reactive Hg(II) to relatively nontoxic, nonreactive
metallic mercury, Hg(0).
Considering the need for root-specific promoters, several
previously examined, native root-specific genes might have
been chosen as a source of promoter and terminator sequences,
but none of these could be depended upon to drive strong
target gene expression in all root tissues and cells. For exam-
ple, large numbers of P450 encoding genes may be root-
specific, but no single P450 of the more than 270 CYP genes
in Arabidopsis has been identified that is both strongly
expressed and expressed in all root tissues (Xu et al., 2001).
Other root-specific genes, including the Zea maize zmGRP3,
ZRP2, and ZRP4 genes (Held et al., 1993, 1997; Goddemeier
et al., 1998) and tomato LeRse-1 gene (Lauter, 1996) all
showed relatively weak levels of root-specific expression.
Localization of ZRP2 and ZRP4 mRNAs suggests their
expression is concentrated in cortical parenchyma cells of
young roots and the root endodermis, respectively. In
transgenic plants, a few plant promoters driving root-specific
expression of reporters have been described, such as Arabi-
dopsis SCARECROW (Sabatini et al., 2003) and tobacco
TobRD2 (also NtQPT1) (Conkling et al., 1990) and TobRB7
(Yamamoto et al., 1991). Again, reporter expression patterns
from these promoters are restricted to subsets of tissues: root
meristem, root cortex, and root vascular cylindar, respectively.
Finally, promoter constructs derived from the Arabidopsis
Pyk10 gene drive moderate levels of expression in transgenic
plants that is relatively root-specific reporter (Nitz et al., 2001).
We tested the TobRB7 system and found MerA root activity
undetectable in most plant lines (data not shown). While the
level of total MerA protein in the most active RB7::merA plant
line was reasonably high, these plants still were not appreci-
ably mercury resistant (Figure 6). Perhaps this is due to the
lack of RB7 promoter activity in root epidermal and vascular
tissues (Yamamoto et al., 1991) leading to the death of these
tissues from Hg(II) toxicity. We also tested merA reporter
gene expression in transgenic plants driven from the root
promoter, NPQpT1 (TobRD2) (Conkling et al., 1990) and did
not obtain significant mercury resistance in a number of
transgenic plants (not shown). The negative results from these
experiments with two tobacco promoters, including low levels
of MerA protein in roots, the lack of any significant mercury
resistance, and substantial leaky expression in shoots of nearly
all plants motivated our development of a root-specific ROC.

Root-specific repressor-operator gene complex 579
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
Engineered ROCs are needed to create a variety of strong
organ- or tissue-specific gene expression patterns and to
create genetic regulatory systems not found among natural
plant promoters. For example, it should be a simple extension
of this work to create a genetically regulated pollen-specific
ROC that would have wide application in the hybrid seed
industry. In the well-accepted BarStar/Barnase system (Beals
and Goldberg, 1997) male-sterile elite line plants are gener-
ated by the anther-specific expression of the toxic ribonucle-
ase, barnase, leading to male sterility. When this male-sterile
line is crossed to a second fully fertile elite plant line express-
ing barstar ribonuclease inhibitor the hybrid seeds are fully
fertile. The BarStar/Barnase system is very specific for these
two proteins. However, a much more widely applicable ROC
could be set up for hybrid seed production. The first male-
sterile elite parent A would be engineered by expressing a
toxic gene, such as that encoding a ribonuclese or RNA inter-
ference construct, from an anther-specific promoter contain-
ing lac operator sequences. A second elite parent, B, would
be engineered for the anther-specific expression of LacIn
repressor, but would remain fully fertile. Crosses between A
and B, with B as the male parent, would produce fully fertile
hybrid seeds and plants due to the repression of the toxic
gene in anthers. Variations on this mechanism could be used
to create fully sterile genetically modified plants that could be
released into the environment with safe containment of their
germplasm (Daniell, 2002).
The root-specific ROC described resulted in strong root
expression levels of GUS and MerA reporter proteins and
GUS reporter levels were repressed by three orders of mag-
nitude in the shoots of some plant lines. The wide variation
observed in both absolute expression levels and in the levels
of shoot repression suggests genome position effects play a
significant role in the performance of this ROC. In the future
we anticipate making improvements to our lac-based ROCs,
testing the root-specific system for application in useful field-
adapted plant species, and developing male-sterility systems
for hybrid seed production and complete-sterility for the
release of genetically modified plants. More immediately, we
will test the root-specific expression of transporters and
detoxifying enzymes that will assist with the development of
plants for the phytoremediation of mercury and arsenic.
Materials and methods
Plasmid construction
A 1072-bp lacI gene coding sequence (GenBank accession
#AY042185) was PCR amplified from plasmid pSK9118
(pMS421) (Grana et al., 1988) supplied by the Sidney Kush-
ner laboratory. The sense primer LacIs-NcoXho (TAGTAAG-
GAG GAACCACCTC GAGGCCATGG GTAAACCAGT
AACGTTATAC GAT) introduced sequences for XhoI and NcoI
sites at the AUG initiation codon on the 5′ end, and the anti-
sense primer LacIa-NLSBamHind (ÁTGTAAGCTT GGATCCTCAA
ACCTTTCTCT TCTTCTTAGG ATGAACAACA GAAGACTGCC
CGCTTTCCAG TCGGGAAA) introduced codons for the NLS
(SSVVHPKKKRKV-stop) and BamHI and HindIII sites at the 3′end of the lacI sequence. This artificial NLS was comprised of
sequences from the SV40 T-antigen (PKKKRKV) (Goldfarb
et al., 1986) and ARP7 (SVVHRK) (Kandasamy et al., 2003).
The amplified LacIn fragment was cleaved by XhoI and HindIII
and ligated into the compatible replacement region of pBlue-
script-SKII (Stratagene) to make the plasmid placIn. The lacIn
coding sequence was subcloned into the NcoI-HindIII replace-
ment region of the pS1pt plasmid to make S1pt::lacIn (Shirley
et al., 1987; Dhankher et al., 2002). The entire cassette
containing the lacIn encoding sequence and SRSI flanking
sequences was subcloned into the KpnI and SacI replace-
ment region of pCambia (Hajdukiewicz et al., 1994).
To construct A2pot, two copies of the 25-bp wild-type
bacterial lac operator sequence 5′GTGGAATTGT GAGCG-
GATAA CAATT were substituted for two sequences in the
ACT2 gene promoter: one immediately following the TATA
box and the other following the transcriptional start site as
shown in Figure 1. A two-fragment overlap extension PCR
mutagenesis strategy (Ho et al., 1989) was used to assemble
the modified ACT2 promoter sequence and replace the cor-
responding sequences of A2pt. The first round PCR paired
sense primer ACT2p-KpnS (GGTACCTGAT CTCAAATACA
TTGATA) with antisense primer Act2p-LacO1A (GCCGGA-
GATT CAAAACGGCT GATGAAAGTG AGGAGGACAA
CGAGACAATT CAATTGTTAT CCGCTCACAA TTCCACTTAT
ATAGGCGGGT TTATCTCTT), and sense primer Act2p-LacO2S
(AGCCGTTTTG AATCTCCGGC GACTTGACAG AGAAGAACAA
GGATGTGGAA TTGTGAGCGG ATAACAATTT AATCCAG-
GAGATTCATTCTC CGTTTTGAA) with antisense primer ACT2p-
1100 A (TAGCTATAAT CGAGCTAACT GAT). The resulting
814- and 310-bp fragments were assembled into a single
fragment in a second round of PCR using the same flanking
primers, ACT2p-KpnS and ACT2p-1100A. This fragment was
digested with KpnI and EcoRI and cloned into the corre-
sponding promoter replacement region of ACT2pt cassette
in bluescript, creating pA2pot.
Both the GUS and merA sequences were cloned into the
A2pot cassette to make the A2pot::GUS and A2pot::merA
genes. The 1851-bp GUS sequence was PCR amplified from
pBI221 (Tanaka et al., 1990) using the sense primer GUS-S

580 Tehryung Kim et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
(TAGAGTTCTA GAATAAAGGA GGAAAAACCG GTACCCCATG
GGATTACGTCCTG TAGAAACCCC AA) to add a XbaI site and
a NcoI site at the ATG codon and using the antisense primer
GUS-A (TTCGATCTCG AGGAGCTCGG ATCCTCATTG TTT-
GCCTCCC TGCTGCGGTT) to add XhoI, SacI and BamHI sites.
The merA77 sequence was cloned as a NcoI-BamHI fragment
into the corresponding replacement region of the A2pot vec-
tor to make A2pot::merA (Figure 1). The KpnI-SacI fragments
containing the A2pot::merA and A2pot::GUS sequences were
moved into the replacement region of pBIN19.
The TobRB7-5A promoter (Yamamoto et al., 1991) was cloned
as a XbaI-BamHI fragment into the compatible replacement
region upstream of the merA77 gene in a pBluescript-merA
construct. The TobRB7-5A promoter-merA gene fusion was
subcloned as a XbaI and KpnI fragment into the compatible
replacement region pBin19 vector with a nos terminator
(Bevan, 1984).
Plant and bacterial growth conditions
Plants were grown on agar media with MS salts or in soil with
16-h light and 8-h darkness. Agar media was supplemented
with various concentrations of mercury chloride (HgCl2) as
indicated.
The A2pot::GUS, A2pot::merA, and S1pt::lacIn constructs
were introduced alone and together into Arabidopsis thal-
iana (ecotype Columbia) by Agrobacterium mediated trans-
formation using the vacuum infiltration procedure (Ye et al.,
1999). T1 generation plants were selected for kanamycin
resistance (35 µg/mL) encoded on the pBIN19 (Bevan, 1984)
derived reporter clones and/or for hygromycin resistance
(50 µg/mL) encoded by the pCambia (http://www.cambia.org)
derived clone S1pt::lacIn. Selection was carried out in 1/2MS
media on agar plates for 7 days after plating seeds. Seedlings
were then moved to nonselective media for 2 weeks and
then transplanted to soil. T2 and T3 generation plants were
examined for the phenotypes described herein.
Histochemical and fluorometric β-glucuronidase assay
GUS activity was histochemically assayed using 4-week-old
seedlings. Seedlings were selected on the media with antibiotics
for 1 week and then transferred to the same media without
antibiotics. The incubation solution was modified from Stomp
(1992) and contained 10 mM EDTA, 0.1 M Na2HPO4, 0.1 M
NaH2PO4, 0.5 mM K ferricyanide, 0.5 mM K ferrocyanide,
and 1.0 mg/mL X-gluc. Tissue was added and incubated for
2 h at 37 °C with gentle shaking and thus modified from the
original staining protocol used for strong actin promoters
(An et al., 1996). Tissues were then treated with two
changes of 70% ethanol over 24 h to remove chlorophyll
and photographed under bright field lighting on a dissecting
microscope.
Root tissues were examined under a compound microscope
to evaluate the staining of individual cells. After staining
(as above) and before ethanol treatment, the tissues were
fixed with 2% glutaraldehyde in 0.2 M sodium phosphate
buffer pH 7.2, dehydrated by the sequence of ethyl glycerol
monoacetate–ethanol–propanol–butanol, infiltrated and
embedded in glycolmethacrylate (HistoResin, Co.), sectioned
at 8 micron thickness, then mounted on glass slides (Kim
et al., 1999).
GUS activity was quantified fluorometrically using 4-week-
old seedlings. GUS was extracted by grinding shoot and root
sections frozen in liquid nitrogen and suspending in an
extraction buffer that contained 50 mM NaHPO4 (pH 7.0),
10 mM β-mercaptoethanol, 0.1% sodium lauryl sarcosine,
10 mM Na2EDTA, and 0.1% Triton X-100. The extract was
reacted with 2 mM 4-methylumbelliferyl β-D-glucuronide
(MUG, Sigma, St Louis, MO) for 1 h at 37 °C, stopped with
0.2 M Na2CO3 stop buffer, and fluorescence was measured
using 360 nm excitation and 480 nm emission and a Synergy
HT Fluorometer (BioTec, Inc., Winooski, VT).
MerA and LacIn protein detection and localization
LacIn and MerA proteins were assayed on Western blots
using polyclonal rabbit antibody (Stratagene) and mono-
clonal antibody (Rugh et al., 1998), respectively. LacI and GUS
protein standards were obtained from (Stratagene) and
(G7896, Sigma), respectively. Total protein levels in plant
extracts were determined as described previously (Kandasamy
et al., 1999). LacIn protein was localized to shoot cell nuclei
using a previously described immunolocalization protocol in
which the fixed tissues were partially digested with sufficient
cell wall-degrading enzymes to release pieces of plant tissue,
but not individual cells (Kandasamy and Meagher, 1999;
Kandasamy et al., 1999).
Acknowledgements
We wish to thank Gay Gragson for editorial comments, Dilip
Shah for thoughts on root pathogens, and Sidney Kushner
for helpful insights into the E. coli lac operon. This work was
supported by USDA grant # 2001-35100-10652 to R.S.B.,
EPA Star Fellowship Program (U-915647-01-0) to A.C.P.H.,
and DOE EMSP grant #DE-FG07-96ER20257 and NIH grant
(GM 36397-18) to R.B.M.

Root-specific repressor-operator gene complex 581
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
References
An, Y.Q., McDowell, J.M., Huang, S., McKinney, E.C., Chambliss, S.and Meagher, R.B. (1996) Strong, constitutive expression of theArabidopsis ACT2/ACT8 actin subclass in vegetative tissues. PlantJ. 10, 107–121.
Anjaiah, V., Cornelis, P. and Koedam, N. (2003) Effect of genotypeand root colonization in biological control of fusarium wilts inpigeonpea and chickpea by Pseudomonas aeruginosa PNA1. Can.J. Microbiol. 49, 85–91.
Beals, T.P. and Goldberg, R.B. (1997) A novel cell ablation strategyblocks tobacco anther dehiscence. Plant Cell, 9, 1527–1545.
Betz, J.L., Sasmor, H.M., Buck, F., Insley, M.Y. and Caruthers, M.H.(1986) Base substitution mutants of the lac operator: in vivo andin vitro affinities for lac repressor. Gene, 50, 123–132.
Bevan, M.W. (1984) Binary Agrobacterium vectors for plant trans-formation. Nuc. Acids. Res. 12, 8711–8721.
Conkling, M.A., Cheng, C.-I., Yamamoto, Y.T. and Goodman, H.M.(1990) Isolation of transcriptionally regulated root-specific genesfrom tobacco. Plant Physiol. 93, 1203–1211.
Daniell, H. (2002) Molecular strategies for gene containment intransgenic crops. Nat. Biotechnol. 20, 581–586.
Dhankher, O.P., Li, Y., Rosen, B.P., Shi, J., Salt, D., Senecoff, J.F.,Sashti, N.A. and Meagher, R.B. (2002) Engineering tolerance andhyperaccumulation of arsenic in plants by combining arsenatereductase and gamma-glutamylcysteine synthetase expression.Nat. Biotechnol. 20, 1140–1145.
Falcon, C.M. and Matthews, K.S. (2000) Operator DNA sequencevariation enhances high affinity binding by hinge helix mutants oflactose repressor protein. Biochemistry, 39, 11074–11083.
Fuerst, T.R., Fernandez, M.P. and Moss, B. (1989) Transfer of theinducible lac repressor/operator system from Escherichia coli to avaccinia virus expression vector. Proc. Natl. Acad. Sci. USA, 86,2549–2553.
Gao, A.G., Hakimi, S.M., Mittanck, C.A., Wu, Y., Woerner, B.M.,Stark, D.M., Shah, D.M., Liang, J. and Rommens, C.M. (2000)Fungal pathogen protection in potato by expression of a plantdefensin peptide. Nat. Biotechnol. 18, 1307–1310.
Giblin-Davis, R.M., Williams, D.S., Bekal, S., Dickson, D.W., Brito, J.A.,Becker, J.O. and Preston, J.F. (2003) ‘Candidatus pasteuria usgae’sp. nov., an obligate endoparasite of the phytoparasitic nematodeBelonolaimus longicaudatus. Int. J. Syst. Evol. Microbiol. 53, 197–200.
Goddemeier, M.L., Wulff, D. and Feix, G. (1998) Root-specific expres-sion of a Zea mays gene encoding a novel glycine-rich protein,zmGRP3. Plant Mol. Biol. 36, 799–802.
Goldfarb, D.S., Gariepy, J., Schoolnik, G. and Kornberg, R.D. (1986)Synthetic peptides as nuclear localization signals. Nature, 322,641–644.
Grana, D., Gardella, T. and Susskind, M.M. (1988) The effects ofmutations in the ant promoter of phage P22 depend on context.Genetics, 120, 319–327.
Guerinot, M.L. (2001) Improving rice yields – ironing out the details.Nat. Biotechnol. 19, 417–418.
Guerinot, M.L. and Eide, D. (1999) Zeroing in on zinc uptake in yeastand plants. Curr. Opin. Plant Biol. 2, 244–249.
Gutteridge, R.J., Bateman, G.L. and Todd, A.D. (2003) Variationin the effects of take-all disease on grain yield and quality ofwinter cereals in field experiments. Pest Manag. Sci. 59, 215–224.
Hajdukiewicz, P., Svab, Z. and Maliga, P. (1994) The small, versatilepPZP family of Agrobacterium binary vectors for plant transforma-tion. Plant Mol Biol. 25, 989–994.
Held, B.M., John, I., Wang, H., Moragoda, L., Tirimanne, T.S., Wurtele, E.S.and Colbert, J.T. (1997) Zrp2: a novel maize gene whose mRNAaccumulates in the root cortex and mature stems. Plant Mol. Biol.35, 367–375.
Held, B.M., Wang, H., John, I., Wurtele, E.S. and Colbert, J.T. (1993)An mRNA putatively coding for an O-methyltransferase accumu-lates preferentially in maize roots and is located predominantly inthe region of the endodermis. Plant Physiol. 102, 1001–1008.
Ho, S.N., Hund, H.D., Horton, R.M., Pullen, J.K. and Pease, L.R. (1989)Site-directed mutagenesis by overlap extension using thepolymerase chain reaction. Gene, 77, 51–59.
Hu, M.C. and Davidson, N. (1990) A combination of derepression ofthe lac operator-repressor system with positive induction by glu-cocorticoid and metal ions provides a high-level-inducible geneexpression system based on the human metallothionein-IIA pro-moter. Mol. Cell Biol. 10, 6141–6151.
Iqbal, J., Afzal, J., Yaegashi, S., Ruben, E., Triwitayakorn, K., Njiti, N.,Ahsan, R., Wood, J. and Lightfoot, A. (2002) A pyramid of locifor partial resistance to Fusarium solani f. sp. glycines maintainsMyoinositol-1-phosphate synthase expression in soybean roots.Theor. Appl. Genet. 105, 1115–1123.
Jacob, F. and Monod, J. (1959) Genes of structure and genes of reg-ulation in the biosynthesis of proteins. C R. Hebd. Seances Acad.Sci. 249, 1282–1284.
Kandasamy, M.K. and Meagher, R.B. (1999) Actin–organelle inter-action: association with chloroplast in Arabidopsis leaf mesophyllcells. Cell Motil. Cytoskeleton, 44, 110–118.
Kandasamy, M.K., McKinney, E.C. and Meagher, R.B. (1999) The latepollen-specific actins in angiosperms. Plant J. 18, 681–691.
Kandasamy, M.K., McKinney, E.C. and Meagher, R.B. (2002)Functional non-equivalency of actin isovariants in Arabidopsis.Mol. Biol. Cell, 13, 251–261.
Kandasamy, M.K., McKinney, E.C. and Meagher, R.B. (2003)Cell cycle–dependent association of Arabidopsis actin-relatedproteins AtARP4 and AtARP7 with the nucleus. Plant J. 33, 939–948.
Kim, T., Choudhury, M.K.U. and Wetzstein, H.Y. (1999) A quantitativeand histological comparison of GUS expression with different pro-moter constructs used in microprojectile bombardment of peanutleaf tissue. In Vitro Cell. Dev. Biol. Plant, 35, 51–56.
Lauter, F.-R. (1996) Root-specific expression of the LeRse-1 gene intomato is induced by exposure of the shoot to light. Mol. Gen.Genet. 252, 751–754.
Lewers, K., Heinz, R., Beard, H., Marek, L. and Matthews, B. (2002)A physical map of a gene-dense region in soybean linkage groupA2 near the black seed coat and Rhg (4) loci. Theor. Appl. Genet.104, 254–260.
Lievens, B., Brouwer, M., Vanachter, A.C., Levesque, C.A.,Cammue, B.P. and Thomma, B.P. (2003) Design and developmentof a DNA array for rapid detection and identification of multipletomato vascular wilt pathogens. FEMS Microbiol. Lett. 223, 113–122.
Meagher, R.B. (2000) Phytoremediation of toxic elemental andorganic pollutants. Curr. Opin. Plant Biol. 3, 153–162.
Nitz, I., Berkefeld, H., Puzio, P.S. and Grundler, F.M. (2001) Pyk10,a seedling and root specific gene and promoter from Arabidopsisthaliana. Plant Sci. 161, 337–346.

582 Tehryung Kim et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2005), 3, 571–582
Pardee, A.B., Jacob, F. and Monod, J. (1958) The role of the induciblealleles and the constrtutive alleles in the synthesis of beta-galactosidase in zygotes of Escherichia coli. C R Hebd. SeancesAcad. Sci. 246, 3125–3128.
Pfahl, M. (1979) Tight-binding repressors of the lac operon: selec-tion system and in vitro analysis. J. Bacteriol. 137, 137–145.
Rugh, C.L., Senecoff, J.F., Meagher, R.B. and Merkle, S.A. (1998)Development of transgenic yellow poplar for mercury phytoreme-diation. Nat. Biotechnol. 16, 925–928.
Sabatini, S., Heidstra, R., Wildwater, M. and Scheres, B. (2003)SCARECROW is involved in positioning the stem cell niche in theArabidopsis root meristem. Genes Dev. 17, 354–358.
Salt, D.E., Blaylock, M., Kumar, N.P.B.A., Dushenkov, V., Ensley, B.D.,Chet, I. and Raskin, I. (1995) Phytoremediation: a novel strategyfor the removal of toxic metals from the environment using plants.Biotechnology, 13, 468–474.
Shirley, B.W., Berry-Lowe, S.L., Rogers, S.G., Flick, J.S., Horsch, R.,Fraley, R.T. and Meagher, R.B. (1987) 5′ proximal sequences of asoybean ribulose-1,5-bisphosphate carboxylase small subunit genedirect light and phytochrome controlled transcription. Nucl. AcidsRes. 15, 6501–6514.
Stomp, A.M. (1992) GUS Protocols: Using the GUS Gene as aReporter of Gene Expression (Gallagher, S.R., ed.), pp. 103–112.Palo Alto, CA: Academic Press.
Ye G.-N., Stone, D., Pang, S.-Z., Creely, W., Gonzalez, K. and Hinchee, M.
(1999) Arabidopsis ovule is the target for Agrobacterium in plantavacuum infiltration transformation. Plant J. 19, 249–257.
Tanaka, A., Mita, S., Ohta, S., Kyozuka, J., Shimamoto, K. andNakamura, K. (1990) Enhancement of foreign gene expression bya dicot intron in rice but not in tobacco is correlated with anincreased level of mRNA and an efficient splicing of the intron.Nucl. Acids Res. 18, 6767–6770.
Ulmasov, B., Capone, J. and Folk, W. (1997) Regulated expressionof plant tRNA genes by the prokaryotic tet and lac repressors.Plant Mol. Biol. 35, 417–424.
Whitson, P.A., Olson, J.S. and Matthews, K.S. (1986) Thermody-namic analysis of the lactose repressor-operator DNA interaction.Biochemistry, 25, 3852–3858.
Wilde, R.J., Shufflebottom, D., Cooke, S., Jasinska, I., Merryweather, A.,Beri, R., Brammar, W.J., Bevan, M. and Schuch, W. (1992) Controlof gene expression in tobacco cells using a bacterial operator-repressor system. EMBO J. 11, 1251–1259.
Xu, W., Bak, S., Decker, A., Paquette, S.M., Feyereisen, R. andGalbraith, D.W. (2001) Microarray-based analysis of gene expres-sion in very large gene families: the cytochrome P450 gene super-family of Arabidopsis thaliana. Gene, 272, 61–74.
Yamamoto, Y.T., Taylor, C.G., Acedo, G.N., Cheng, C.L. andConkling, M.A. (1991) Characterization of cis-acting sequencesregulating root-specific gene expression in tobacco. Plant Cell, 3,371–382.
Copyright © 2022 FDOKUMEN

