







Bahasa
Halaman
Hukum


[CANCER RESEARCH 54, 4398-4404, August 15. 1994]
Efficient Transfer of Selectable and Membrane Reporter Genes in HematopoieticProgenitor and Stem Cells Purified from Human Peripheral Blood1
M. Valtieri, R. Schirò, C. Chelucci, B. Masella, U. Testa, I. Casella, E. Montesoro, G. Mariani, H. J. Hassan,and C. Peschle2
Thomas Jefferson Cancer Institute, Thomas Jefferson University, Philadelphia, Pennsylvania 19107; Department of Hematology and Oncology, Istituto Superiore di Sanità , Rome,Italy; Pediatrie Clinic, University of Milan, S. Gerardo Hospital, Monza, Italy
ABSTRACT
We have utilized highly purified hematopoietic progenitor and stemcells (HPCs, HSCs) from normal peripheral blood to develop methodologyfor: (a) efficient transfer into HPCs of a non-hematopoietic membrane
reporter, i.e., the nerve growth factor receptor complementary DNA; andi in effective gene transduction of putative HSCs, i.e., cells initiating Dexter-type long-term culture (LTC-ICs).
Purified HPCs induced into cycling by growth factors (interleukin 3,interleukin 6, c-kit ligand) were transduced with the N2 retroviral vectorcontaining the neomycin resistance dico' \ gene. More than 80% of trans
duced HPCs were resistant to the toxic G418 level. Thereafter, the HPCswere effectively transduced with the LNSN retroviral vector containing anerve growth factor receptor complementary DNA; the nerve growthfactor receptor was detected on £18% of the transduced HPCs. Theseexperiments provide a new tool from which (a) to monitor expression ofa transduced membrane reporter on hematopoietic cells, particularly atthe level of HPCs/HSCs, and (b) to characterize the transduced cells bydouble- and triple-labeling membrane antigen analysis.
Purified HPCs/HSCs grown in Dexter-type LTC were transduced at 1
week by exposure to supernatant N2 retroviral particles in the absence ofexogenous hematopoietic growth factors. The procedure, devoid of toxiceffects, allowed an efficient neo' transduction into LTC-ICs. Thus, we
consistently detected neomycin-resistant mRNA in the clonal progeny ofHPCs produced in LTC at 5-8 weeks in both the nonadherent and
adherent fractions; this timing of expression coincides with that of HPCproduction by LTC-ICs, thereby indicating the effective transduction ofthe LTC-ICs. These experiments represent a first step toward develop
ment of preclinical models for gene transfer into human peripheral bloodHSCs by complex retroviral vectors.
INTRODUCTION
Human hematopoiesis is sustained by pluripotent HSCs,3 which can
extensively self-renew and feed into lineage(s)-committed HPCs (1).
HPCs, in turn, differentiate into morphologically recognizable precursors, which mature to terminal elements circulating in PB (1).
Gene transfer into human HSCs represents a potentially fundamental tool at both basic and clinical levels (2-5). Transfer of physiolog
ically significant genes into HSCs may allow analysis of the cellular/molecular basis of hematopoiesis, particularly in the early crucialproliferation and differentiation steps. Furthermore, gene therapy with
Received 3/11/94; accepted 6/13/94.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This study was supported in part by Thomas Jefferson University, Philadelphia, PA;
Fondazione Tettamanti, CNR (P.P. ACRO), Rome; Progetto Terapia dei Tumori, IstitutoSuperiore di Sanità , Rome.
2 To whom requests for reprints should be addressed, at Thomas Jefferson Cancer
Institute, Thomas Jefferson University, Bluemle Life Sciences Building, Room 528, 233S. 10th St., Philadelphia, PA 19107.
3 The abbreviations used are: HSC, hematopoietic stem cell; HPC, hematopoietic
progenitor cell; BM, bone marrow; HGF, human growth factor; LTC, long-term culture;LTC-IC, long-term culture-initiating cell; PB. peripheral blood; cDNA, complementaryDNA; NGFR, nerve growth factor receptor; mAb, monoclonal antibody; KL, c-kit ligand;CPU, colony-forming units; Ep, erythropoietin; IMDM, Iscove's modified Dulbecco's
medium; MoMuLV, Moloney murine leukemia virus; IL, interleukin; FCS, fetal calfserum; RT-PCR, reverse transcription-polymerase chain reaction; GM-CSF, granulocyte-macrophage colony-stimulating factor.
appropriately transduced HSCs may bypass the present requirementfor allogeneic HSC transplantation in a variety of genetic disorders(3).
Gene transfer into murine repopulating HSCs by retroviral vectorshas been reported (4-7). Conversely, attempts to effectively transduce
human HPCs/HSCs by retroviral vectors have been only partiallysuccessful.
Retroviral gene transfer has been described for HPCs from BM,normal or "mobilized" PB, and cord blood (8-13). However, target
HPCs were usually cocultivated with vector-producing cell lines(10-13), thus hindering clinical application of these procedures.
LTC on an irradiated allogeneic stroma has been introduced recently to assay human primitive HPCs with repopulating capacity;these cells, termed LTC-ICs, produce clonogenetic precursors for atleast 5-8 weeks. It is generally conceded that LTC-ICs represent a
subset of primitive HPCs closely related to the pluripotent repopulating HSC (14); in the mouse, primitive HPCs from Dexter-type LTCinduce long-term in vivo repopulation of the myeloid-lymphoid system (7). BM LTCs cocultivated with exogenous HGFs and vector-
producing cell lines have shown integration and expression of thetransduced gene in clonogenic precursors until 6 weeks (15).
Analysis and characterization of transduced hematopoietic cellswill be facilitated greatly by transfer of a non-hematopoietic mem
brane reporter gene. Recently, a NGFR cDNA has been inserted in PBlymphocytes (16).
In the present study we report efficient retroviral gene transfer ofboth the neor and NGFR genes into HPCs purified from normal PB.Furthermore, we have developed an efficient procedure for neor gene
transfer into LTC-ICs, purified from normal PB and coincubated with
retroviral particles in the absence of exogenous HGFs.
MATERIALS AND METHODS
Recombinant Human HGFs
IL-3 (1.7 X IO7 units/mg), IL-6 (5.1 X Iff units/mg), and GM-CSF(2.5 x IO7 units/mg) were supplied by the Genetics Institute (Cambridge,
MA), and Ep and KL were supplied by Amgen (Thousand Oaks, CA) andImmunex (Seattle, WA), respectively.
Cells
Adult PB was obtained from twenty 40-year-old male donors after informedconsent. Blood (450 ml ±10%) was collected in a preservative-free anticoagulant (citrate-phosphate-dextrose-adenine). A buffy coat was obtained by
centrifugatiun (Beckman J6 M/E; 1400 rpm/20 min at room temperature).Bone marrow was obtained from healthy young male or female donors after
informed consent.
HPC/HSC Purification
PB HPCs were purified according to a modification of the method reportedin Ref. 17. In Step I, PB-buffy coat samples were separated over a Ficoll-
Hypaque density gradient (density, 1.077 g/ml) (Pharmacia Fine Chemicals,Piscataway, NJ); PB mononuclear cells, resuspended in IMDM containing20% heat-inactivated FCS (GIBCO, Grand Island, NY), were treated with
three cycles of plastic adherence. In Step II, cells were separated by
4398
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
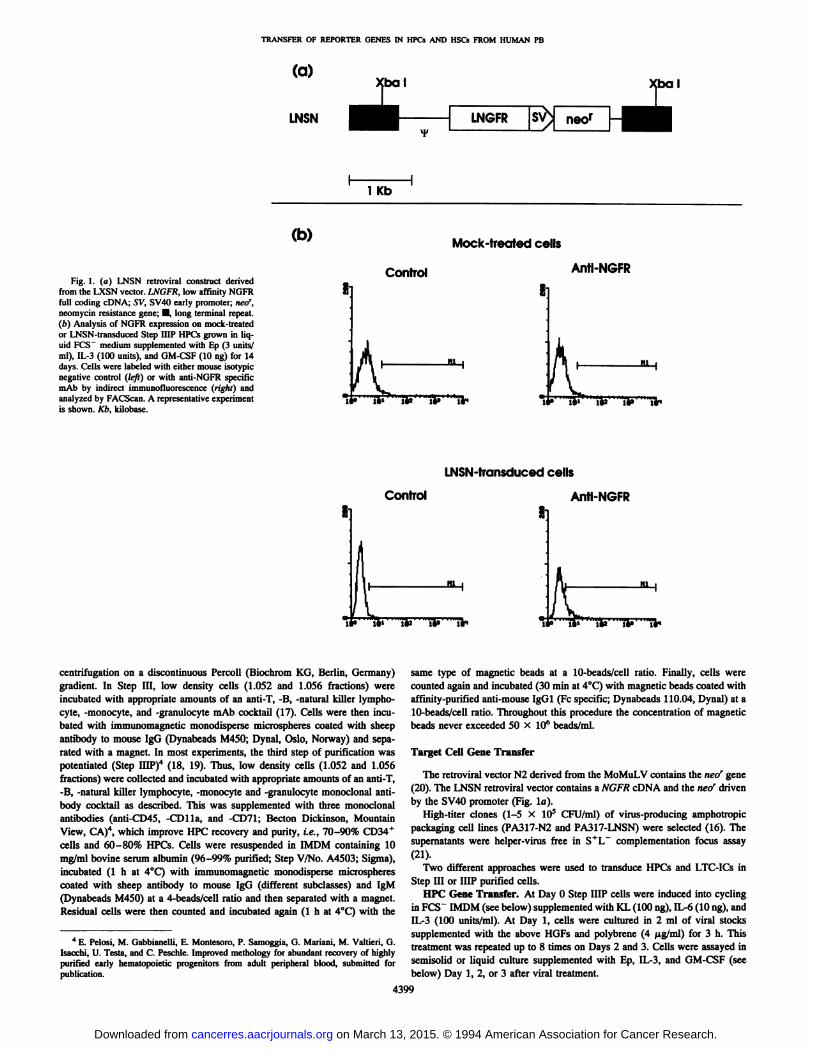
TRANSFER OF REPORTER GENES IN HPCs AND HSCs FROM HUMAN PB
(a)
LNSN
(bal
LNGFRsv;neor
Xbal
l Kb
Fig. 1. (a) LNSN retroviral construct derivedfrom the LXSN vector. LNGFR, low affinity NGFRfull coding cDNA; SV, SV40 early promoter; neo1,
neomycin resistance gene; •long terminal repeat.(b) Analysis of NGFR expression on mock-treatedor LNSN-transduced Step HIP HPCs grown in liquid FCS~ medium supplemented with Ep (3 units/
ml), IL-3 (100 units), and GM-CSF (10 ng) for 14days. Cells were labeled with either mouse isotypicnegative control (left) or with anti-NGFR specific
mAb by indirect immunofluorescence (righi) andanalyzed by FACScan. A representative experimentis shown. Kb, kilobase.
(b) Mock-treated cells
Control Anti-NGFR
Õ4« Õ4J Õ4» T4*
11
LNSN-transduced cells
Control Anti-NGFR
li»
centrifugation on a discontinuous Percoli (Biochrom KG, Berlin, Germany)gradient. In Step III, low density cells (1.052 and 1.056 fractions) wereincubated with appropriate amounts of an anti-T, -B, -natural killer lymphocyte, -monocyte, and -granulocyte mAb cocktail (17). Cells were then incu
bated with immunomagnetic monodisperse microspheres coated with sheepantibody to mouse IgG (Dynabeads M450; Dynal, Oslo, Norway) and separated with a magnet. In most experiments, the third step of purification waspotentiated (Step HIP)4 (18, 19). Thus, low density cells (1.052 and 1.056
fractions) were collected and incubated with appropriate amounts of an anti-T,-B, -natural killer lymphocyte, -monocyte and -granulocyte monoclonal anti
body cocktail as described. This was supplemented with three monoclonalantibodies (anti-CD45, -CDlla, and -CD71; Becton Dickinson, MountainView, CA)4, which improve HPC recovery and purity, i.e., 70-90% CD34*
cells and 60-80% HPCs. Cells were resuspended in IMDM containing 10mg/ml bovine serum albumin (96-99% purified; Step V/No. A4503; Sigma),incubated (l h at 4°C)with immunomagnetic monodisperse microspheres
coated with sheep antibody to mouse IgG (different subclasses) and IgM(Dynabeads M450) at a 4-beads/celI ratio and then separated with a magnet.Residual cells were then counted and incubated again (l h at 4°C)with the
4 E. Pelosi, M. Gabbianelli, E. Montesoro, P. Samoggia, G. Marìani,M. Valtieri, G.
Isacchi, U. Testa, and C. Peschle. Improved methology for abundant recovery of highlypurified early hematopoietic progenitors from adult peripheral blood, submitted forpublication.
same type of magnetic beads at a 10-beads/cell ratio. Finally, cells werecounted again and incubated (30 min at 4°C)with magnetic beads coated with
affinity-purified anti-mouse IgGl (Fc specific; Dynabeads 110.04, Dynal) at a10-beads/cell ratio. Throughout this procedure the concentration of magneticbeads never exceeded 50 X IO6 beads/ml.
Target Cell Gene Transfer
The retroviral vector N2 derived from the MoMuLV contains the neo' gene(20). The LNSN retroviral vector contains a NGFR cDNA and the neor driven
by the SV40 promoter (Fig. la).High-liter clones (1-5 X Vf CFU/ml) of virus-producing amphotropic
packaging cell lines (PA317-N2 and PA317-LNSN) were selected (16). Thesupernatants were helper-virus free in S+L~ complementation focus assay
(21).Two different approaches were used to transduce HPCs and LTC-ICs in
Step III or HIP purified cells.HPC Gene Transfer. At Day 0 Step HIP cells were induced into cycling
in PCS" IMDM (see below) supplemented with KL (100 ng), IL-6 (10 ng), and
IL-3 (100 units/ml). At Day 1, cells were cultured in 2 ml of viral stocks
supplemented with the above HGFs and polybrene (4 fig/ml) for 3 h. Thistreatment was repeated up to 8 times on Days 2 and 3. Cells were assayed insemisolid or liquid culture supplemented with Ep, IL-3, and GM-CSF (see
below) Day 1, 2, or 3 after viral treatment.
4399
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

TRANSFER OF REPORTER GENES IN HPCs AND HSCs FROM HUMAN PB
LTC-IC Gene Transfer. At Day 0, Steps III or HIP cells were seeded ontoDexter-type LTC (see below). On Day 7, one-half of the medium was replacedto induce LTC-IC cycling (22). On Day 8, LTC cells were incubated with viral
stocks (3 h incubation 3 times/day) by replacing directly in the culture flask,LTC medium with producer packaging line medium (10% PCS in Dulbecco'smodified Eagle's medium). Polybrene, protamine sulfate, and HGFs were not
added. Uninfected mock LTC were processed in parallel.
Clonogenic Assay
Step HIP HPCs transduced with the N2 vector were seeded at 1 X IO2 and1 x IO4 cells/ml/dish, respectively, in 0.9% methylcellulose, 40% PCS inIMDM (GIBCO) supplemented with a-thioglycerol (10~4 M) (Sigma) in the
presence of Ep (3 units/ml), KL (100 ng), IL-3 (100 units), and GM-CSF (10ng) to stimulate colony formation by erythroid burst-forming units, CFU-granulocyte-macrophage and multilineage CPU (granulocyte, erythrocyte,macrophage, megakaryocytic, CFU-granulocyte-erythrocyte-macrophage-monocyte) at 37°Cin a 5% CO2, 5% O2, and 90% N2 humidified atmosphere.
G418 (Sigma) was added at 1.6 mg/ml.
Liquid Phase Culture (23)
Purified Step HIP HPCs transduced with the LNSN vector or mock treatedwere incubated for 14 days in FCS~ liquid culture in the presence of Ep (3
units/ml), IL-3 (100 units), and GM-CSF (10 ng). PCS was replaced by (24)bovine serum albumin (10 mg/liier; Fraction V, Sigma), pure human trans-
ferrin (1 mg/ml), human low density lipoproteins (40 fig/ml), insulin (10/xg/ml), sodium pyruvate (10~4 mg/liter), L-glutamine (2 X 10~3 mg/liter),rare inorganic elements (25) supplemented with iron sulfate (4 x 10"" mg/
liter), and nucleosides (10 fig/ml of each). G418 selection was not performed.This methodology allows in 14-17 days, a a2 log expansion of the initial
HPC population to generate a progeny composed of 77% erythroblasts and23% granulocytic cells. At day 14, cells were collected, washed twice, andincubated for 30 min at 4°Cwith a human anti-NGFR mAb (clone 8211;
Boehringer Mannheim, Mannheim, Germany) specific for the ligand bindingepitope of NGFR or an irrelevant IgGl isotypic control. Cells were washedtwice and treated with a goat anti-mouse F(ab')2 fluoresceinated antibody
(Becton Dickinson, Oxnard, CA) as described, washed twice, and analyzed byFACScan (Becton Dickinson).
Dexter-type LTC
The LTC conditions have been described previously (26). Briefly, the BMaspirate Ficoll-Hypaque cut was inoculated (1.5-2 X IO6 cells/ml) in 25-cm2
tissue culture flasks (Falcon; Becton Dickinson, Plymouth, United Kingdom)in 10 ml total volume comprising 10% preselected PCS (GIBCO), 10%preselected horse serum (GIBCO), and 5 X 10~7 M hydrocortisone hemisuc-
cinate (Sigma) in IMDM. Flasks were incubated in a 5% CO, atmosphere at33°C.After formation of a confluent stromal layer in 2-4 weeks, flasks were
exposed to 15 Gy to ablate endogenous BM hematopoiesis and then used asstromal layers. A total of 10 experiments were performed with purified PBHPCs (6 Step III and 4 Step HIP) seeded at different cell concentrations.One-half of the medium was replaced weekly. Cultures were terminated byenzymatic treatment with a trypsin-EDTA solution (GIBCO) at 5 or 8 weeks.
HPCs in the nonadherent and adherent fractions were assayed in clonogeneticculture as above in the absence of G418.
Molecular Analysis
Total RNA was extracted from single colonies by the guanidine isothiocya-nate/CsCl method (27). Control RT-PCR of ß,-microglobulin mRNA in singlecolonies indicated a consistently reproducible RNA extraction (28). First-
strand cDNA was synthesized by MoMuLV reverse transcriptase (GIBCOBRL, Gaithersburg, MD) using the neo'-antisense oligomer 5'-ATTCGCCG-GCAAGCTCTC'l lCA-3' (20) as primer. Amplification was achieved by 30
PCR cycles in a total volume of 50 ¡il.Denaturation was performed at 95°Cfor1 min, annealing at 55°Cfor 90 s, and extension at 72°Cfor 90 s. The primersneo'-sense 5'-GCTCGCGCCAGC CGAACTGT-3' and neo'-antisense encom
pass a 215-base pair fragment. Thirty fil of each reaction were electrophoresed
in a 1.8% agarose gel, transferred onto a nylon filter, and hybridized withan internal probe (5'-TCTCGTCGTGACCCATGGCGAT GCCTGCTT-3').
Table 1 Effect of N2 viral particle treatment on Step /IIP HPCs, as indicated bypercentage of G418-resistant colonies
Experiment12345Dayofinfection11,21,21,2,31,2,31,2,31,2,3No.of infectioncycles1223(1
+ 1+1)6(1+2+3)6(1+2+3)8(1+3+4)G418-resistant
clones14%323645676080
RNA from the PA317-N2 packaging and Hep3B cell lines were used as
positive and negative controls, respectively.
RESULTS
neor Gene Transfer into Step HIP HPCs. Optimal conditions forneor gene transfer in Step HIP HPCs were evaluated in the first series
of experiments. Actively cycling cells are transduced preferentially byMoMuLV-derived retroviral vectors (29, 30). Thus, HPCs weretreated with a combination of HGFs (KL, IL-6, and IL-3), whichinduced a rapid rise of their cycling as evaluated by [3H]deoxythy-midine-thymidine suicide assay (31, 32).5 An increase in the number
of treatments, especially on days 2 and 3, significantly augmented thenumber of G418-resistant clones (from 14 to 60-80% of total colo
nies scored at day 14) (Table 1).NGFR Gene Transfer into Step HIP HPCs. LNSN is a more
complex retroviral vector containing both the NGFR reporter and neor
gene (Fig. la). Step HIP HPCs were treated with the LNSN vectorunder optimal conditions, as established above. At least 18% or moreof transduced cells, grown in PCS" liquid culture in the presence of
Ep (3 units), IL-3 (100 units,) and GM-CSF (10 ng) for 14 days,
expressed the NGFR as shown by indirect immunofluorescence analysis (Fig. Ib).
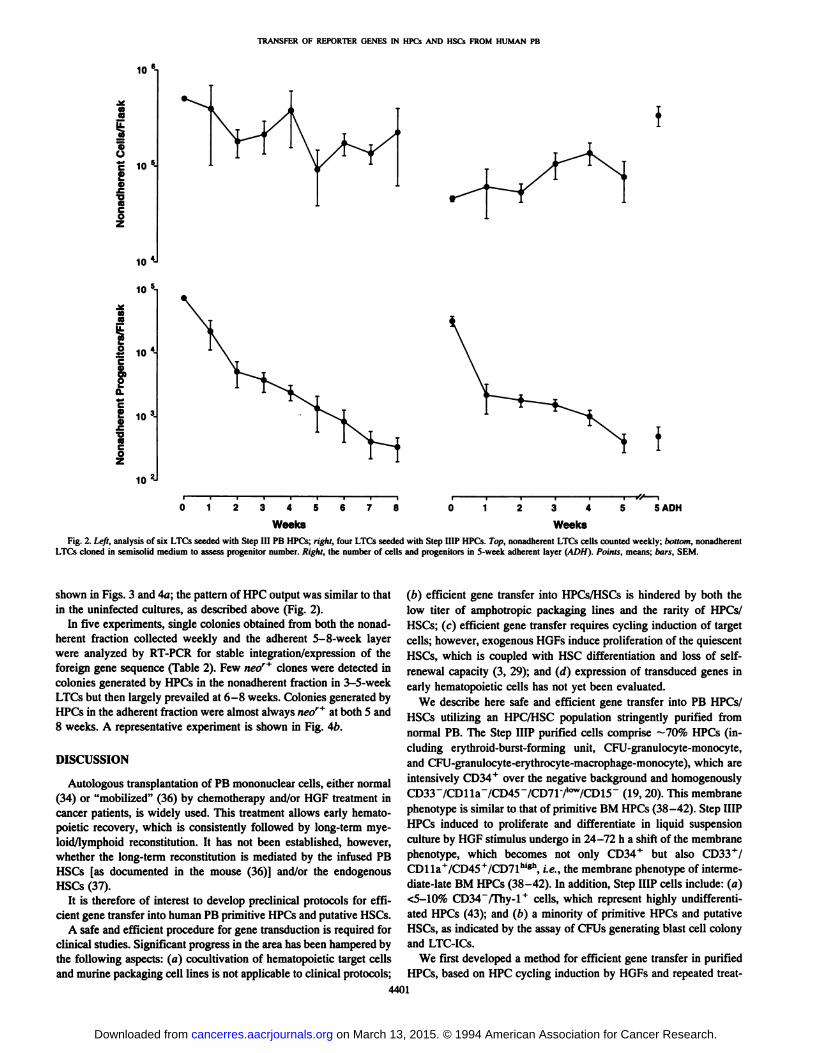
LTC-IC Assay. The production of clonogenic cells in LTC by StepHI HPCs/HSCs seeded onto a preestablished irradiated BM adherentlayer was evaluated for 8 weeks. Results from six individual experiments are shown in Fig. 2 (left). The purification index of the HPCwas 26.4 ±3.8% (SEM). Purified cells attached to the adherent layer.Observation by phase contrast under an inverted microscope showedsmall round cells closely associated with the stroma by 3 weeks ofLTC (not shown). Indirect evidence of attachment was provided bythe decrease in the total number of nonadherent cells during the first2 weeks of culture (Fig. 2, left). Clonogenic cells were detected in allcultures in both the nonadherent and adherent fractions up to 8 weeks(Fig. 2 and results not shown).
In four additional experiments with Step ¡UPHPCs/HSCs (purification index, 73.3 ±8.3%; CD34+ cells, 70-90%) clonogenic cell
number was assayed in both the adherent and nonadherent fractions(Fig. 2, right, bottom). The adherent/nonadherent progenitor ratioranged from 1.7 to 3.0, in line with recent LTC-limiting dilution
analysis (13).The consistent presence of a clonogenic progeny in all LTC through
5-8 weeks in both the adherent and nonadherent fractions indicatesthe presence of LTC-ICs in the initial inoculum (13, 33).
Gene Transfer into LTC-ICs. We attempted to insert the neo geneinto LTC-ICs present in primitive HPCs/HSCs purified from PB.Thus, we monitored the frequency and stability of neor gene insertion
in the clonal progeny of HPCs generated in 5-8-week LTCs.
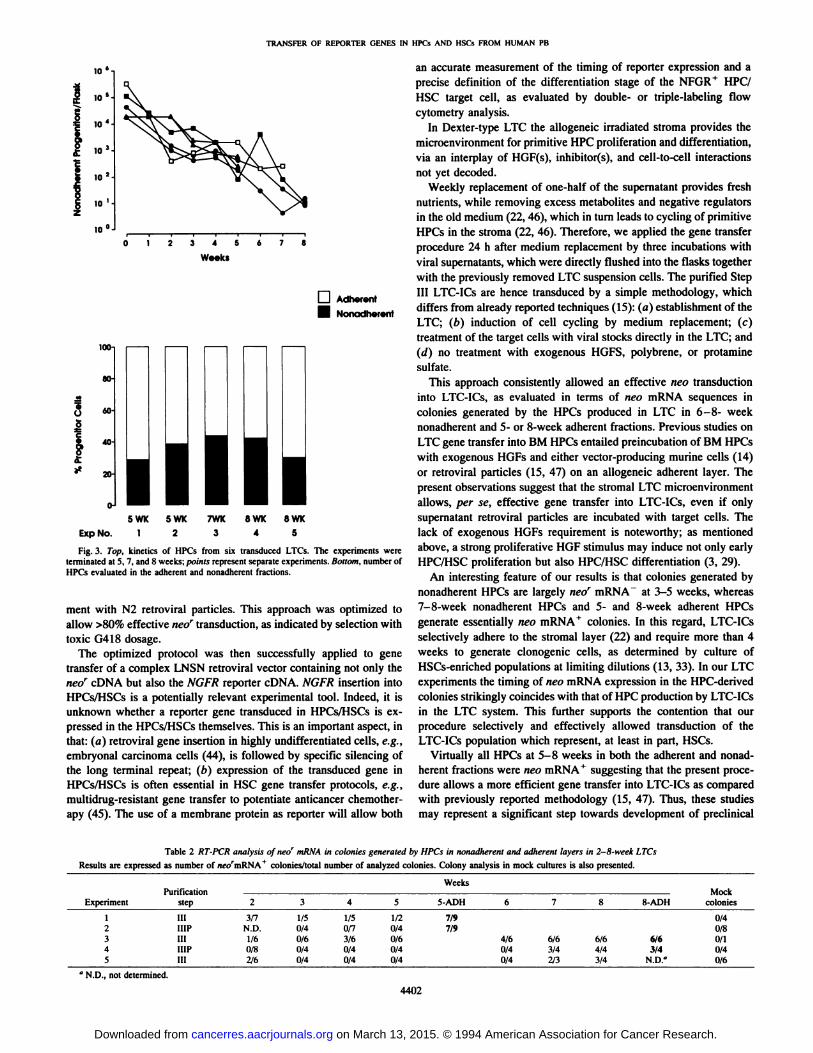
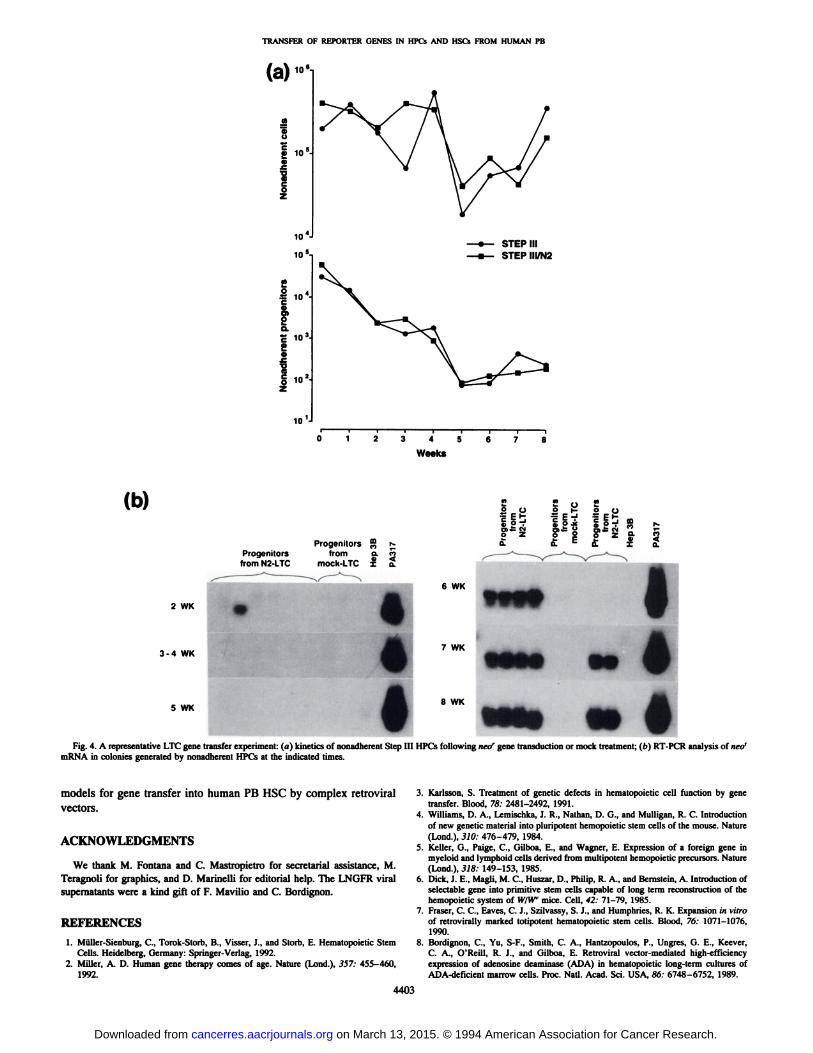
In seven independent experiments, LTCs were treated at 1 weekwith freshly filtered supernatant from the PA317-N2 cell line (Figs. 3,4fl). Clonogenic cell production of these LTCs through 5-8 weeks is
5 Unpublished data.
4400
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
TRANSFER OF REPORTER GENES IN HPCs AND HSCs FROM HUMAN PB
10
ii•aaO
105-
10
.i 10'H
g.!¡ «l
lI
345
Weeks3 4
Weeks
5ADH
Fig. 2. ¿f/r,analysis of six LTCs seeded with Step III PB HPCs; righi, four LTCs seeded with Step HIP HPCs. Top, nonadherent LTCs cells counted weekly; bottom, nonadherentLTCs cloned in semisolid medium to assess progenitor number. Right, the number of cells and progenitors in 5-week adherent layer (ADH). Points, means; bars, SEM.
shown in Figs. 3 and 4a; the pattern of HPC output was similar to thatin the uninfected cultures, as described above (Fig. 2).
In five experiments, single colonies obtained from both the nonadherent fraction collected weekly and the adherent 5-8-week layerwere analyzed by RT-PCR for stable integration/expression of theforeign gene sequence (Table 2). Few neor+ clones were detected in
colonies generated by HPCs in the nonadherent fraction in 3-5-weekLTCs but then largely prevailed at 6-8 weeks. Colonies generated byHPCs in the adherent fraction were almost always neor+ at both 5 and
8 weeks. A representative experiment is shown in Fig. 40.
DISCUSSION
Autologous transplantation of PB mononuclear cells, either normal(34) or "mobilized" (36) by chemotherapy and/or HGF treatment in
cancer patients, is widely used. This treatment allows early hemato-poietic recovery, which is consistently followed by long-term mye-loid/lymphoid reconstitution. It has not been established, however,whether the long-term reconstitution is mediated by the infused PBHSCs [as documented in the mouse (36)] and/or the endogenousHSCs (37).
It is therefore of interest to develop preclinical protocols for efficient gene transfer into human PB primitive HPCs and putative HSCs.
A safe and efficient procedure for gene transduction is required forclinical studies. Significant progress in the area has been hampered bythe following aspects: (a) cocultivation of hematopoietic target cellsand murine packaging cell lines is not applicable to clinical protocols;
(b) efficient gene transfer into HPCs/HSCs is hindered by both thelow titer of amphotropic packaging lines and the rarity of HPCs/HSCs; (c) efficient gene transfer requires cycling induction of targetcells; however, exogenous HGFs induce proliferation of the quiescentHSCs, which is coupled with HSC differentiation and loss of self-
renewal capacity (3, 29); and (d) expression of transduced genes inearly hematopoietic cells has not yet been evaluated.
We describe here safe and efficient gene transfer into PB HPCs/HSCs utilizing an HPC/HSC population stringently purified fromnormal PB. The Step HIP purified cells comprise -70% HPCs (including erythroid-burst-forming unit, CFU-granulocyte-monocyte,and CFU-granulocyte-erythrocyte-macrophage-monocyte), which areintensively CD34+ over the negative background and homogenouslyCD33-/CDlla-/CD45-/CD71Vlow/CD15- (19, 20). This membrane
phenotype is similar to that of primitive BM HPCs (38-42). Step HIPHPCs induced to proliferate and differentiate in liquid suspensionculture by HGF stimulus undergo in 24-72 h a shift of the membranephenotype, which becomes not only CD34"1"but also CD33+/CDlla+/CD45+/CD71high, i.e., the membrane phenotype of interme
diate-late BM HPCs (38-42). In addition, Step HIP cells include: (a)<5-10% CD34~/Thy-l+ cells, which represent highly undifferenti-
ated HPCs (43); and (b) a minority of primitive HPCs and putativeHSCs, as indicated by the assay of CPUs generating blast cell colonyand LTC-ICs.
We first developed a method for efficient gene transfer in purifiedHPCs, based on HPC cycling induction by HGFs and repeated treat-
4401
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
TRANSFER OF REPORTER GENES IN HPCs AND HSCs FROM HUMAN PB
10 S
I
l ,0
)0
10'
2345678
Weeks
Adherent
Nonodherent
80
«40- Illll
5WK 5WK 7WK 8 WK 8 WK
ExpNo. 12345
Fig. 3. Top. kinetics of HPCs from six transduced LTCs. The experiments wereterminated at 5, 7, and 8 weeks; points represent separate experiments. Bottom, number ofHPCs evaluated in the adherent and nonadherent fractions.
ment with N2 retroviral particles. This approach was optimized toallow >80% effective neor transduction, as indicated by selection with
toxic G418 dosage.The optimized protocol was then successfully applied to gene
transfer of a complex LNSN retroviral vector containing not only theneor cDNA but also the NGFR reporter cDNA. NCFR insertion into
HPCs/HSCs is a potentially relevant experimental tool. Indeed, it isunknown whether a reporter gene transduced in HPCs/HSCs is expressed in the HPCs/HSCs themselves. This is an important aspect, inthat: (a) retroviral gene insertion in highly undifferentiated cells, e.g.,embryonal carcinoma cells (44), is followed by specific silencing ofthe long terminal repeat; (b) expression of the transduced gene inHPCs/HSCs is often essential in HSC gene transfer protocols, e.g.,multidrug-resistant gene transfer to potentiate anticancer chemother
apy (45). The use of a membrane protein as reporter will allow both
an accurate measurement of the timing of reporter expression and aprecise definition of the differentiation stage of the NFGR ' HPC/
HSC target cell, as evaluated by double- or triple-labeling flow
cytometry analysis.In Dexter-type LTC the allogeneic irradiated stroma provides the
microenvironment for primitive HPC proliferation and differentiation,via an interplay of HGF(s), inhibitor(s), and cell-to-cell interactions
not yet decoded.Weekly replacement of one-half of the supernatant provides fresh
nutrients, while removing excess metabolites and negative regulatorsin the old medium (22, 46), which in turn leads to cycling of primitiveHPCs in the stroma (22, 46). Therefore, we applied the gene transferprocedure 24 h after medium replacement by three incubations withviral supernatants, which were directly flushed into the flasks togetherwith the previously removed LTC suspension cells. The purified StepIII LTC-ICs are hence transduced by a simple methodology, which
differs from already reported techniques (15): (a) establishment of theLTC; (h) induction of cell cycling by medium replacement; (c)treatment of the target cells with viral stocks directly in the LTC; and(d) no treatment with exogenous HGFS, polybrene, or protaminesulfate.
This approach consistently allowed an effective neo transductioninto LTC-ICs, as evaluated in terms of neo mRNA sequences incolonies generated by the HPCs produced in LTC in 6-8- weeknonadherent and 5- or 8-week adherent fractions. Previous studies on
LTC gene transfer into BM HPCs entailed preincubation of BM HPCswith exogenous HGFs and either vector-producing murine cells (14)
or retroviral particles (15, 47) on an allogeneic adherent layer. Thepresent observations suggest that the stromal LTC microenvironmentallows, per se, effective gene transfer into LTC-ICs, even if only
supernatant retroviral particles are incubated with target cells. Thelack of exogenous HGFs requirement is noteworthy; as mentionedabove, a strong proliferative HGF stimulus may induce not only earlyHPC/HSC proliferation but also HPC/HSC differentiation (3, 29).
An interesting feature of our results is that colonies generated bynonadherent HPCs are largely neor mRNA" at 3-5 weeks, whereas
7-8-week nonadherent HPCs and 5- and 8-week adherent HPCsgenerate essentially neo mRNA+ colonies. In this regard, LTC-ICs
selectively adhere to the stromal layer (22) and require more than 4weeks to generate clonogenic cells, as determined by culture ofHSCs-enriched populations at limiting dilutions (13, 33). In our LTCexperiments the timing of neo mRNA expression in the HPC-derivedcolonies strikingly coincides with that of HPC production by LTC-ICs
in the LTC system. This further supports the contention that ourprocedure selectively and effectively allowed transduction of theLTC-ICs population which represent, at least in part, HSCs.
Virtually all HPCs at 5-8 weeks in both the adherent and nonadherent fractions were neo mRNA+ suggesting that the present proce
dure allows a more efficient gene transfer into LTC-ICs as compared
with previously reported methodology (15, 47). Thus, these studiesmay represent a significant step towards development of preclinical
Table 2 RT-PCR analysis of neor mRNAin colonies generated by HPCs in nonadhereni and adherent layers in 2-8-week LTCsResults are expressed as number of n«/mRNA+ colonies/total number of analyzed colonies. Colony analysis in mock cultures is also presented.
Experiment1
234
5Purification
stepIIIHIPHI
HIPHI23/7
N.D.1/6(IS2/631/5
0/40/60/40/441/5
0/73/6040/451/2
0/40/60/40/4Weeks5-ADH7/9
7/964/6
0/40/476/63/42/386/6 4/43/48-ADH6/63/4N.D."Mock
colonies0/4
0/80/10/40/6
' N.D., not determined.
4402
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
TRANSFER OF REPORTER GENES IN HPCs AND HSCs FROM HUMAN PB
(a)«'-
10'J
10'
10 J-
S£•a
10'-i
STEP IIISTEP III/N2
23456
Weeks
(b)i i i:
2 WK
3-4 WK
5 WK
•Ei-
f El1^ a
o. z
6 WK
7 WK
8 WK
Fig. 4. A representative LTC gene transfer experiment: (a) kinetics of nonadherent Step III HPCs following neo' gene transduction or mock treatment; (b) RT-PCR analysis of neo'
mRNA in colonies generated by nonadherent HPCs at the indicated times.
models for gene transfer into human PB HSC by complex retroviralvectors.
ACKNOWLEDGMENTS
We thank M. Fontana and C. Mastropietro for secretarial assistance, M.Teragnoli for graphics, and D. Mannelli for editorial help. The LNGFR viralsupernatants were a kind gift of F. Mavilio and C. Bordignon.
REFERENCES
1. Müller-Sienburg, C., Torok-Storb, B., Visser, J., and Storb, E. Hematopoietic StemCells. Heidelberg, Germany: Springer-Verlag, 1992.
2. Miller, A. D. Human gene therapy comes of age. Nature (Lond.), 357: 455-460,
1992.
3. Karlsson, S. Treatment of genetic defects in hematopoietic cell function by genetransfer. Blood, 78: 2481-2492, 1991.
4. Williams, D. A., Lemischka, J. R., Nathan, D. G., and Mulligan, R. C. Introductionof new genetic material into pluripotent hemopoietic stem cells of the mouse. Nature(Lond.), 310: 476-479, 1984.
5. Keller, G., Paige, C., Gilboa, E., and Wagner, E. Expression of a foreign gene inmyeloid and lymphoid cells derived from multipotent hemopoietic precursors. Nature(Lond.), 318: 149-153, 1985.
6. Dick, J. E., Magli, M. C.. Huszar, D., Philip, R. A., and Bernslein. A. Introduction ofselectable gene into primitive stem cells capable of long term reconstruction of thehemopoietic system of W/W mice. Cell, 42: 71-79, 1985.
7. Fraser, C. C., Eaves, C. J., Szilvassy, S. J., and Humphries, R. K. Expansion in vitroof retrovirally marked totipotent hematopoietic stem cells. Blood, 76: 1071-1076,
1990.8. Bordignon, C., Yu, S-F., Smith, C. A., Hantzopoulos, P., Ungres, G. E., Keever,
C. A., O'Reill, R. J., and Gilboa, E. Retroviral vector-mediated high-efficiency
expression of adenosine deaminase (ADA) in hematopoietic long-term cultures ofADA-deficient marrow cells. Proc. Nati. Acad. Sci. USA, 86: 6748-6752, 1989.
4403
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

TRANSFER OF REPORTER GENES [N HPCs AND HSCs FROM HUMAN PB
9. Brcgni, M., Magni, M., Siena, S., Di Nicola, M., Bonadonna. G., and Gianni. A. M.Human peripheral hlixxl hcmopoietic progenitors are optimal targets of retroviral-mcdiated gene transfer. Blood. KO: 1418-1422, 1992. 28.
10. Dick, J. E., Kamel-Reid, S., Murdoch, B., and Doedens, M. Gene transfer into normalhuman hematopoietic cells using in vitro and in vivo assays. Blood, 78: 624—634,
1991.11. Fink, J. K., Correli. P. H., Perry, L. K.. Brady. R. O., and Karlsson, S. Correction of 29.
gluciKcrehrosida.se deficiency after retroviral mediated gene transfer into hematopoietic progenitor cells from patients with Gaucher disease. Proc. Nati. Acad. Sci. USA. 30.87: 2334-2338, 1990.
12. Cournoyer, D., Scarpa, M.. Milani. K., Moora, K. A., Markowitz, D., Bank, A..Belmont, J. W., and Caskey. C. T. Gene transfer of adenosine deaminase imo 31.primitive human hematopoietic progenitor cells. Hum. Gene Ther., 2: 203-211, 1991.
13. Sutherland. H. J.. Lansdorp, P. M.. Henckelman. D. H., Eaves, A. C., and Eaves, C. J.Functional characterization of individual human hemopoietic stem cells cultured at 32.limiting dilution on supportive marrow stromal layers. Proc. Nati. Acad. Sci. USA,«7:3584-3588, 1990.
14. Hughes, P. F. D.. Eaves, C. J.. Hogge, D. E.. and Humphries, R. K. High-efficiencygene transfer to human hemopoietic cells maintained in long-term marrow culture. 33.Blood, 74: 1915-1919, 1989.
15. Hughes, P. F. D., Thacker. J. D., Hogge, D. E., Sutherland, H. J., Thomas, T. E.,Lansdorp, P. M., Eaves, C. J., and Humphries. R. K. Retroviral gene transfer to 34.primitive normal and leukemic hematopoietic cells using clinically applicable procedures. J. Clin. Invest., 89: 1817-1824. 1992.
16. Mavilio. F.. Ferrari, G., Rossini. S., Bonini, C., Casorati, G., Traversar!, C., and 35.Bordignon, C. Peripheral blood lymphocytes as target of retroviral vector-mediatedgene transfer. Blood, 83: 1988-1997, 1994.
17. Gabbianelli, M., Sargiacomo, M., Pelosi, E., Testa, U., Isacchi, G., and Pésenle,C."Pure" human hemopoietic progenitors: permissive action of basic fibrohlast growth 36.
factor. Science (Washington DC). 249: 1561-1564, 1990.
18. Labbaye, C. Valtieri, M., Testa, U., Giampaolo, A.. Meccia, E., Sterpetti, P., Parolini,1., Pelosi E.. Bulgarini, D.. Cayre, Y. E., and Peschle, C. Retinole acid downmodu- 37.lates erythroid differentiation and GATA-1 expression in purified adult progenitorculture. Blood. 83: 651-656, 1994. 38.
19. Peschle, C., Testa. U., Valtieri, M., Gabbianelli, M., Pelosi, E., Montesoro, E., Sposi,N. M., Fossati, C., Camagna, A., and Care. A. Stringently purified human hematopoietic progenitors/stem cells: analysis of cellular/molecular mechanisms underlying 39.early hematopoiesis. Stem Cells. U: 356-370. 1993.
20. Armentano, D., Yu, S. F., Kantoff. P. W., von Rüden.T., Anderson. W. F., andCiilboa. E. Effect of internal viral sequences on the utility of retroviral vectors. J. 40.Virol.. 61: 1647-1650, 1987.
21. Miller, A. D.. and Buttimore. C. Redesign of retrovinis packaging cell lines to avoidrecombination leading to helper virus production. Mol. Cell. Biol.. 6: 2895-2903,
1986.22. Eaves, A. C.. and Eaves, C. J. Maintenance and proliferation control of primitive 41.
hcmopoietic progenitors in long-term cultures of human marrow cells. Blood Cells.14: 355-368. 1988.
23. Sposi. N. M.. Zon, L. I., Care, A.. Valtieri, M.. Testa, U.. Gabbianelli, M., Mariani, 42.G., Bollero. L., Mather. C.. Orkin. S. H., and Peschle, C. Cell cycle-dependentinitiation and lineage-dependent abrogation of GATA-1 expression in pure differentiating hematopoietic progenitors. Proc. Nati. Acad. Sci. USA, 89: 6353-6357. 43.1992.
24. Valtieri, M.. Gabbianelli. M., Pelosi, E., Bassano. E.. Petti, S., Russo, G.. Testa, U., 44.and Peschle, C. Erythropoietin alone induces erythroid burst formation by humanembryonic hut not adult BFU-E in unicellular serum-free culture. Blood, 74: 460- 45.470, 1989.
25. Eliason, J. Granulocyte-macrophagc colony-formation in serum-free culture: effects 46.of purified colony-stimulating factors and modulation by hydrocortisone. J. Cell.Physiol., ¡28:231-238, 1986.
26. Coutinho. L. H., Will, A.. Radford. J., Schirò, R.. Testa, N. D.. and Dexter, T. M. Theeffects of recombinant hG-CSF. hGM-CSF. hIL-3 on hemopoiesis in human long- 47.term bone marrow culture. Blood, 75: 2118-2129, 1990.
27. Chirgwin. J. M., Prebyla, A. E., MacDonald. R. J., and Rutter, W. T. Isolation of
biological active ribonucleic acid from sources enriched in rihonucleasc. Biochemistry, 18: 5294-5300, 1979.Hernandez, A., Osterholtz. J.. Price, C. M.. Wiedemann, L. M., Gordon. M. Y.,Goldman, J. M., and Morgan, G. J. Detection of the hybrid BCR/ABL mRNA insingle CFU-GM using the polymerase chain reaction. Exp. Hematol. (NY), 18:1142-1144, 1990.Mulligan, R. C. The basic science of gene therapy. Science (Washington DC), 260:926-932, 1993.Miller. D. G.. Adam, M. A., and Miller. A. D. Gene transfer hy retrovinis vectorsoccurs only in cells that are actively replicating at the time of infection. Mol. Cell.Biol., 10: 4239-4242, 1990.
Hamada. K., Akagi, T., Okano, A.. Taki, S.. Shimotohno, K.. and Takahara. Y. A newmethod of gene transfer into interleukin-3 and interleukin-6. J. Immunol. Methods.141: 177-186, 1991.
Valtieri, M.. Vcnturelli, D., Care, A., Fossati, C., Pelosi, E., Mattia, G., Gewirtz,A. M., Calabretta, B., and Peschle, C. Antisensc myh inhibition of purified erythroidprogenitors in development and differentiation is linked to cycling activity andexpression of DNA polymerase a. Blood. 77: 1181-1189. 1991
Udomsakdi. C. U., Lansdorp, P. M., Hogge. D. E., Reid, D. S., Eaves, A. C, andEaves, C. J Characterization of primitive hemopoietic cells in normal human peripheral blood. Blood, 80: 2513-2521, 1992.Kessinger, A.. Armitage, J. O., Landmark. J. D., Smith. D. M.. and Weisemhurger.D. D. Autologous peripheral hemopoietic stem cell transplantation restores hemopoietic function following marrow ablative therapy. Blood. 71: 723-727, 1988.
Siena, S.. Bregni. M.. Brando. B.. Gianni, M.. and Bonadonna. G. Circulation ofCD34f hcmopoietic stem cells in the peripheral blood of high dose cyclophospha-
mide treated patients: enhancement by intravenous rccombinant human granulocyte-macrophage colony-stimulating factor. Blood, 74: 1905-1914. 1989.
Molineux. G.. Pojda. Z.. Hampson. I. N.. Lord. B. !.. and Dexter T. M. Transplantation potential of peripheral blood stem cells induced by granulocyte colony-stimulating factor. Blood, 76: 2153-2161, 1990.Eaves, C. J. Peripheral blood stem cells reach new heights. Blood, 82: 1957-1959,
1993.Andrews, R. G., Singer. J. W., and Bernstein, I.D. Precursors of colony-forming cellsin humans can he distinguished from colony-forming cells by expression of the CD33and CD34 antigens and light scatter properties. J. Exp. Med.. 769: 1721-1731, 1989.Lansdorp, P. M.. Sutherland. H. J.. and Eaves. C. J. Selective expression of CD45isoforms on functional subpopulations of CD34 ' hemopoielic cells from human bone
marrow. J. Exp. Med.. 772: 363-373. 1990.Gunji. Y., Nakamura, M.. Hagiwara, T.. Hayakawa, K.. Ratsushita, H.. Osawa, H.,Nagayoshi, K.. Nakauchi, H., Yanagisawa. M., Miura, Y.. and Suda. T. Expressionand function of adhesion molecules on human hematopoietic stem cells: CD34 'LFA-r cells are more primitive then CD34*LFA-1+ cells. Blood, 80: 429-436,
1992.Lansdorp, P. M., and Dragowska, W. Long-term erythropoiesis from constant numbers of CD34 f cells in serum-free cultures initiated with highly purified progenitor
cells from human bone marrow. J. Exp. Med., 175: 1501-1512. 1992.Fritsch, G., Buchinger, P.. Printz, D., Fink. F. M., Mann. G.. Peters, C., Wagner. T..Adler. A., and Gadner. H. Rapid discrimination of early CD34 * myeloid progenitors
using CD45RA analysis. Blood, 81: 2301-2309, 1993.Craig, W., Kay, R.. Cutler, R. L., and Lansdorp. P. M. Expression of Thy-1 on humanhematopoietic progenitor cells. J. Exp. Med.. 777: 1331-1342, 1993.Petersen, R.. Kempler, G., and Barklis, E. A stem cell-specific silencer in theprimer-binding site of a retrovinis. Mol. Cell. Biol., 77: 1214-1221, 1991.
Guticrrezz. A. A.. Lemoine, N. R.. and Sikora, K. Gene therapy for cancer. Lancet (N.Am. Ed.). .î.?9:715-721, 1992.Eaves, C. J., Sutherland, H. J., Cashman, J. D.. Otsuka, T., Lansdorp. P. M..Humphries, R. K., Eaves, A. C., and Hogge, D. E. Regulation of primitive humanhematopoietic cells in long-term marrow cultures. Seminars in Hcmatology (NY), 28:126-131, 1991.Moore. K. A., Deisserolh. A. B., Reading. C. L.. Williams. D. E., and Belmont. J. W.Stromal support enhances cell-free retroviral vector transduction of human bonemarrow long-term culture-initiating cells. Blood, 79: 1393-1399. 1992.
4404
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1994;54:4398-4404. Cancer Res M. Valtieri, R. Schirò, C. Chelucci, et al. Human Peripheral Bloodin Hematopoietic Progenitor and Stem Cells Purified from Efficient Transfer of Selectable and Membrane Reporter Genes
Updated version
http://cancerres.aacrjournals.org/content/54/16/4398
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on March 13, 2015. © 1994 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
Top Related
Copyright © 2022 FDOKUMEN

