






Bahasa
Halaman
Hukum


Journal oflmmunologicalMethods, 134 (1990) 43-50 43 Elsevier
JIM05727
Efficient hybridization of mouse-human cell lines by means of hypo-osmolar electrofusion
U. Z i m m e r m a n n 1, p. Gessner 1, R. Schnet t ler ~, S. Perkins 2 and S .K.H. F o u n g 2
I Institute of Biotechnology, University of Wfirzburg~ ROntgenring 11, 8700 Wiirzburg, F.R.G., and 2 Department of Pathology, Stanford University, School of Medicine, Stanford, CA 94305, U.S.A.
(Received 8 May 1990, accepted 3 July 1990)
The fusion of a mouse-human heteromyeloma with a mouse hybridoma is used as a model to define parameters to generate human hybridomas. Electrofusion of these cells in 300 mosM and 75 mosM solutions showed that strong hypo-osmolar conditions resulted in a dramatic increase in the efficiency of hybridoma formation. In contrast to iso-osmolar electrofusion, a high hybrid yield could be obtained by injection of only a single field pulse. The field strength was adjusted in proportion to the increased size of the cells in hypo-osmolar solutions. Hypo-osmolar electrofusion allowed the generation of approximately 0.45% hybrids at a suspension density of 1.75.105 mouse-human cel ls /ml corresponding to an input number of 3.5 • 10 4 mouse-human cells. A further increase in the efficiency of hybridoma formation to about 0.6% was achieved by cell alignment in an alternating field of modulated field strength. Experiments in which the total cell number per fusion chamber was decreased at constant optimum suspension density showed that a further increase in the efficiency of hybridoma formation in hypo-osmolar solution was not possible because of the increasing influence of the heterogeneity of the cell lines with decreasing cell number. The results allow the conclusion that hypo-osmolar electrofusion is a potential tool to enhance successful immortalisation of human B lymphocytes.
Key words: Hypo-osmolar electrofusion; Mouse-human cell line; Hybridoma formation
Introduction
Somatic cell hybridization is the most widely used method in the generation of monoclonal anti- bodies to pre-defined antigens (Neil and Urnovitz, 1988). Whereas this technique has been proven
Correspondence to: S.K.H. Foung, Stanford University Blood Bank, 800 Welch Road, Palo Alto, CA 94304, U.S.A.
Abbreviations: AOT, alignment off time; BrdU, bromo-de- oxyuridine; BSA, bovine serum albumin; CGM, complete growth medium; FCS, fetal calf serum; HAT, hypoxanthine- aminopterin-thymidine; HMAbs, human monoclonal antibod- ies; 75 L3, hypo-osmolar fusion medium; 300 L3, iso-osmolar fusion medium.
successful for the production of monoclonal anti- bodies of murine origin, the production of human monoclonal antibodies (HMAbs) has been more difficult to achieve for a variety of reasons (re- viewed in James and Bell, 1987). General prob- lems are the lack of appropriate fusion partners to immortalize human B cells, poor fusion efficiency of human cells and rapid loss of immunoglobulin secretion. In spite of these limitations some suc- cesses have been documented in the production of HMAbs.
One approach to solve the problem of poor efficiency of human cells is the application of electrofusion (Bischoff et al., 1982; Ohnischi et al., 1987; Foung et al., 1989; Foung and Perkins,
0022-1759/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

44
1989). A further enhancement of fusion efficiency is expected by electrofusion in low ionic, hypo- osmolar solutions (75 mosM) which has been suc- cessful applied in the production of murine mono- clonal antibodies (Schmitt and Zimmermann, 1989; Schmitt et al., 1989a; Zimmermann et al., 1990). The yield of hybrids considerably exceeded the yield obtained by electrofusion in low ionic, iso-osmolar solutions or by polyethylene glycol- mediated fusion (Karsten et al., 1985; Vienken and Zimmermann, 1985; Gravekamp et al., 1987; Ohnishi et al., 1987; Glassy, 1988; Zimmermann et al., 1988; Foung et al., 1989; Schmitt et al., 1989b).
To achieve ultimately a high rate of human hybridoma formation with a small number of in- put human B lymphocytes, we explored the poten- tial of hypo-osmolar electrofusion. As a first step, this modified electrofusion technique was applied to the fusion of a mouse-human by mouse model cell system. This involved the fusion of a hypo- xanthine-aminopterin-thymidine (HAT)-sensitive mouse-human heteromyeloma suitable for the pro- duction of human hybridomas, H73Cl l , with the HAT-sensitive mouse hybridoma cell line, G8. Be- cause of the similar cell size of the mouse-human heteromyeloma and the murine hybridoma G8 strain the well known effect of cell radius on the fusion voltage requirements is mitigated. Hypo- osmolar electrofusions performed with this system should yield information about the optimum elec- tric field parameters necessary for generating hu- man hybridomas with mouse-human fusion partners (Zimmermann, 1986; Schmitt and Zim- mermann, 1989).
Materials and methods
Cells The murine hybridoma cell line G8 originated
from fusion of splenic B cells (C57BL/6 mice) to the non-secreting myeloma cell line SP2/0-Ag14 (Shulman et al., 1978). The resulting hybridomas were cultured in complete growth medium (CGM) supplemented with HAT for the selection of thymidine-kinase deficient variants. One of these variants was then grown for 1 week followed by subcloning in growth medium containing 20
/~g/ml of bromo-deoxyuridine (BrdU). HAT-sen- sitive clones were selected and passaged in BrdU- supplemented growth medium. This fusion partner was generously provided by Dr. Howard Urnovitz (Calypte Biomedical Company, Berkeley, CA, U.S.A.).
The mouse-human heteromyeloma cell line H73Cl l was produced by fusion between Epstein- Barr virus activated human B cells and the mouse myeloma, NS-1. The heteromyelomas obtained were subsequently placed in 6-thioguanine and ouabain. This step of selection resulted in the formation of the HAT sensitive and ouabain re- sistant cell line H73Cl l . This fusion partner was generously provided by Dr. James Larrick (Gene- labs, Redwood City, CA, U.S.A.).
Cell culture G8 and H73Cl l cells were cultured in RPMI
1640 medium supplemented with 10% fetal calf serum (FCS, Boehringer), 2 g/1 NaHCO 3, 2 mM sodium pyruvate (Biochrom, Berlin), 2 mM L- glutamine, non-essential amino acids (Biochrom, Berlin), 100 U / m l penicillin-streptomycin and 50 ktM 2-mercaptoethanol (complete growth medium, CGM) at 37°C in an atmosphere enriched with 5% CO 2. Every 2-3 days, the cell suspensions were diluted with growth medium in a ratio of 1 : 10 in the case of G8 cells, and 3 :7 in the case of H73Cl l cells. The diluted cell suspensions were transferred to new culture flasks in order to keep the cells in the log phase for the electrofusion. The HAT sensitivity of the cells was verified in experi- ments performed at regular intervals. The cell cultures in log phase were again split 3 : 7 in the case of H73Cl l and 1 : 10 in the case of G8 1 day before fusion. These cultures were grown to a density of about 3-5 - 105 cel ls /ml in CGM after this last propagation step. The cells were then harvested by 10 min centrifugation at 150 × g (for detailed description, see Zimmermann et al., 1990).
Fusion procedure The two cell types were counted using a hemo-
cytometer and mixed together in a ratio of 1 : 1 for fusion. The cell mixtures were washed twice with hypo-osmolar fusion medium (75 L3) containing 75 mosM sorbitot, 0.1 mM Ca2+-acetate, 0.5 mM Mg 2+ -acetate and 1 m g /m l bovine serum al-

bumin (BSA). For fusion in iso-osmolar fusion medium (300 L3) a solution containing 280 mosM sorbitol, 0.1 mM Ca2+-acetate, 0.5 mM Mg 2÷- acetate and 1 m g / m l BSA was used. After wash- ing in 75 L3 or in 300 L3 medium an aliquot of cells was resuspended with the appropriate volume of the fusion medium necessary to reach the cell density desired for the specific experiment. 200/zl of each adjusted cell suspension was pipetted into the fusion chamber. After connection of the fusion chamber to the electrofusion apparatus (Biojet CF manufactured by Biomed, Theres) an alternating field of 250 V cm-1 (peak-value) and of 1.5 MHz frequency was applied for 30 s for cell alignment unless otherwise stated. Fusion was induced sub- sequently by application of rectangular field pulses of 15 /~s duration. Field strength and number of pulses were tested in the range of 1.25 1.75 kV cm -1 respectively using one or three pulses. The time interval between consecutive pulses was ad- justed to 1 s and the alignment off time (AOT) of the alternating field during each pulse to 10 ms. The alignment was continued for 30 s (250 V c m - l , 1.5 MHz) after the final pulse. The field- treated cells and identically handled controls were maintained at the fusion temperature of 25°C without any further manipulation for 10 min.
The chambers were then gently opened and rinsed with 1 ml of C G M free of phenol red. Cell suspensions of 0.3 ml were pipetted into each of four wells of a 24 well Greiner cloning plate (Greiner, F.R.G.). Each of these wells was pre- filled with 1.0 ml of phenol red free CGM. The fused cells and corresponding controls were grown at 37°C in a 5% CO 2 enriched atmosphere. After 24 h, 1 ml HAT medium (Boehringer, Mannheim, F.R.G.) was added to each well (HAT-selection medium). The number of colonies which survived in the presence of HAT was scored after 5-7 days using an inverted microscope (Leitz Labovert, Wetzlar, F.R.G.).
Fusion chamber Helical chambers were used unless as stated
(Perkins et al., 1990; Zimmermann et al., 1990). The electrode assembly consisted of ten 200 # m diameter and 0.15 m length platinum wires (200 ~m space between each wire) wound in a parallel helix around a plexiglass tube and connected to
45
two protruding electrodes which were attached to the power supply.
200/~1 of the cell suspension was pipetted into a cylindrical receptacle of slightly larger inner diameter than the diameter of the electrode assem- bly. The tube was carefully inserted into the re- ceptacle and the chamber was closed by screwing the head of the electrode assembly to the top of the receptacle. Subsequently the chamber was con- nected to the power supply (for details see: Per- kins et al., 1990).
Results
Initial experiments using fusion media with dif- ferent osmolarities showed that the mouse-human cell line, H73Cl l , can be kept for more than 30 min in 75 L3 solution without any significant cellular deterioration and without changes in growth after transfer into CGM. Therefore, this strongly hypo-osmolar medium was selected for the following experiments because fusion of murine cells has shown (Schmitt and Zimmer- mann, 1989; see also D~iumler and Zimmermann, 1989) that fusion and hybrid yield increased with decreasing osmolarity.
Optical examination of the fusion process of H73Cl l and G8 cells between two parallel elec- trodes glued on a microslide (distance 200 ~m) revealed large differences in fusion efficiency in iso(300 mosM, 300 L3) and hypo-osmolar (75 mosM, 75 L3) media. When using standard condi- tions (three consecutive pulses of 2.25 kV cm -1 field strength and about 1 0 6 cel ls /chamber) , (Schmitt et al., 1989b) iso-osmolar electrofusion only resulted in (hetero- or homologous) fusion of a few cells (Fig. l a ) . Injection of a single pulse of the same field strength did not lead to any fusion events. At about 5 • 105 ce l l s /chamber the fusion frequency was also nearly zero, even when three pulses were applied. These results indicated that both cell lines resisted electrofusion. In contrast, under hypo-osmolar conditions, multiple fusions occurred between the aligned cells ( 1 .10 6 cells / chamber) in response to a single field pulse of 1.25 kV cm -1 strength (Fig. lb). Multiple fusion re- suited in the formation of many giant cells. Fig. lc shows giant cells generated by homologous fusion
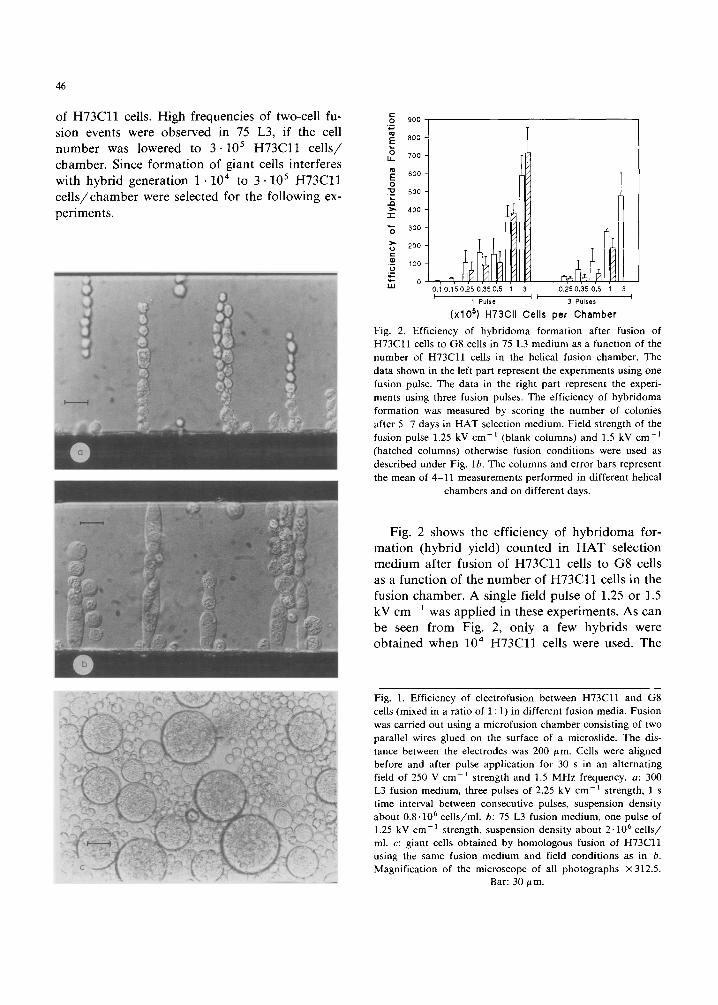
46
of H 7 3 C l l cells. High frequencies of two-cell fu- sion events were observed in 75 L3, if the cell number was lowered to 3 .105 H 7 3 C l l ce l l s / chamber. Since formation of giant cells interferes with hybrid generation 1 . 1 0 4 to 3" 10 5 H 7 3 C l l ce l l s /chamber were selected for the following ex- periments.
900 ',~
800
0 700 U-
600
O "~ 500
" ~ 400 - r
300
200
,-- 100 ,_u
0 UJ 0 .10 .150 .250 .350 .5 1 3 0 .250,350.5 1 3
1 Pulse 3 Pulses
(xlO s) H73CII Cells per Chamber Fig. 2. Efficiency of hybridoma formation after fusion of H73Cll cells to G8 cells in 75 L3 medium as a function of the number of H73Cll cells in the helical fusion chamber. The data shown in the left part represent the experiments using one fusion pulse. The data in the right part represent the experi- ments using three fusion pulses. The efficiency of hybridoma formation was measured by scoring the number of colonies after 5-7 days in HAT selection medium. Field strength of the fusion pulse 1.25 kV cm -1 (blank columns) and 1.5 kV cm -1 (hatched columns) otherwise fusion conditions were used as described under Fig. lb. The columns and error bars represent the mean of 4-11 measurements performed in different helical
chambers and on different days.
Fig. 2 shows the efficiency of hybridoma for- mation (hybrid yield) counted in H A T selection medium after fusion of H 7 3 C l l cells to G8 cells as a function of the number of H 7 3 C l l cells in the fusion chamber. A single field pulse of 1.25 or 1.5 kV cm -1 was applied in these experiments. As can be seen from Fig. 2, only a few hybrids were obtained when 10 4 H 7 3 C l l cells were used. The
Fig. 1. Efficiency of electrofusion between H73C11 and G8 cells (mixed in a ratio of 1 : 1) in different fusion media. Fusion was carried out using a microfusion chamber consisting of two parallel wires glued on the surface of a microslide. The dis- tance between the electrodes was 200 ,um. Cells were aligned before and after pulse application for 30 s in an alternating field of 250 V cm -1 strength and 1.5 MHz frequency, a: 300 L3 fusion medium, three pulses of 2.25 kV cm-1 strength, l s time interval between consecutive pulses, suspension density about 0.8.106 cells/ml, b: 75 L3 fusion medium, one pulse of 1.25 kV cm-1 strength, suspension density about 2-106 cells/ ml. c: giant cells obtained by homologous fusion of H73Cll using the same fusion medium and field conditions as in b. Magnification of the microscope of all photographs x312.5.
Bar: 30/~m.
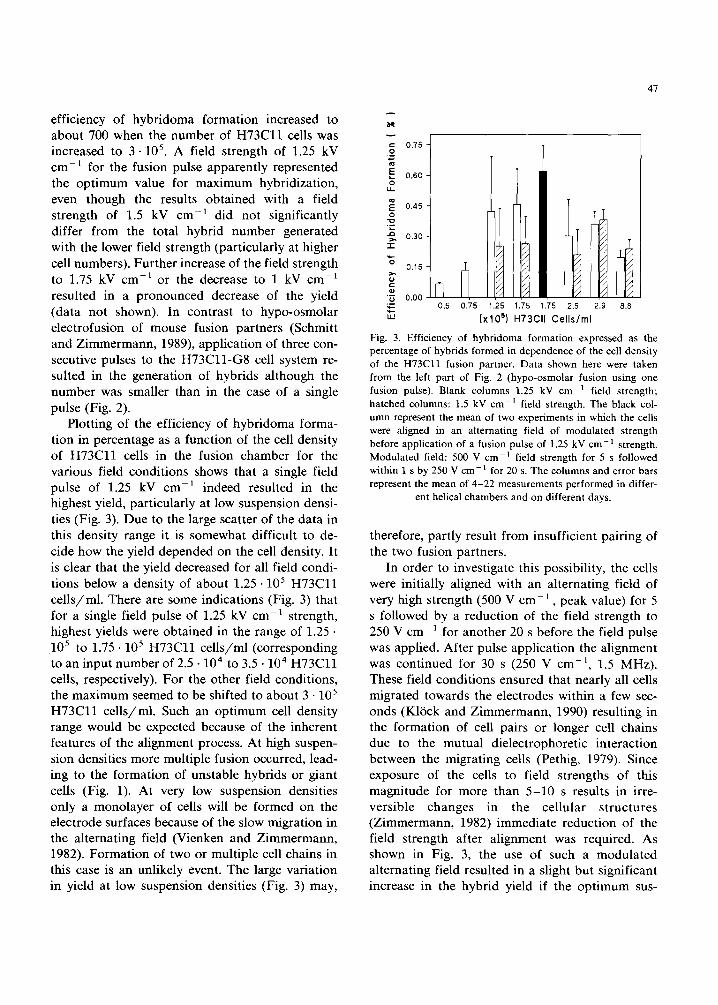
efficiency of hybridoma formation increased to about 700 when the number of H 7 3 C l l cells was increased to 3.105. A field strength of 1.25 kV cm-1 for the fusion pulse apparently represented the opt imum value for maximum hybridization, even though the results obtained with a field strength of 1.5 kV cm -1 did not significantly differ from the total hybrid number generated with the lower field strength (particularly at higher cell numbers). Further increase of the field strength to 1.75 kV cm -a or the decrease to 1 kV cm -a resulted in a pronounced decrease of the yield (data not shown). In contrast to hypo-osmolar electrofusion of mouse fusion partners (Schmitt and Zimmermann, 1989), application of three con- secutive pulses to the H 7 3 C l l - G 8 cell system re- sulted in the generation of hybrids although the number was smaller than in the case of a single pulse (Fig. 2).
Plotting of the efficiency of hybridoma forma- tion in percentage as a function of the cell density of H 7 3 C l l cells in the fusion chamber for the various field conditions shows that a single field pulse of 1.25 kV cm -1 indeed resulted in the highest yield, particularly at low suspension densi- ties (Fig. 3). Due to the large scatter of the data in this density range it is somewhat difficult to de- cide how the yield depended on the cell density. It is clear that the yield decreased for all field condi- tions below a density of about 1.25 • 105 H 7 3 C l l cel ls /ml . There are some indications (Fig. 3) that for a single field pulse of 1.25 kV cm-1 strength, highest yields were obtained in the range of 1.25 • 105 to 1.75 • 105 H 7 3 C l l cel ls /ml (corresponding to an input number of 2 .5 .10 4 to 3.5 • 10 4 H 7 3 C l l cells, respectively). For the other field conditions, the maximum seemed to be shifted to about 3 • 105 H 7 3 C l l cel ls /ml . Such an opt imum cell density range would be expected because of the inherent features of the alignment process. At high suspen- sion densities more multiple fusion occurred, lead- ing to the formation of unstable hybrids or giant cells (Fig. 1). At very low suspension densities only a monolayer of cells will be formed on the electrode surfaces because of the slow migration in the alternating field (Vienken and Zimmermann, 1982). Formation of two or multiple cell chains in this case is an unlikely event. The large variation in yield at low suspension densities (Fig. 3) may,
47
0.75
0.60 o [J_
0.45 o t 111
0.30 "1-
'~ ~ = ~" 0.15 [~
'~ 0.00 0.5 0.75 1.25 1.75 1.75 2.5 2.9 8.8
" ' [ x lO s) H73Cll Cel ls/ml
Fig. 3. Efficiency of hybridoma formation expressed as the percentage of hybrids formed in dependence of the cell density of the H73Cll fusion partner. Data shown here were taken from the left part of Fig. 2 (hypo-osmolar fusion using one fusion pulse). Blank columns 1.25 kV cm -1 field strength; hatched columns: 1.5 kV cm -1 field strength. The black col- umn represent the mean of two experiments in which the cells were aligned in an alternating field of modulated strength before application of a fusion pulse of 1.25 kV c m - l strength. Modulated field: 500 V cm 1 field strength for 5 s followed within 1 s by 250 V c m - l for 20 s. The columns and error bars represent the mean of 4-22 measurements performed in differ-
ent helical chambers and on different days.
therefore, partly result from insufficient pairing of the two fusion partners.
In order to investigate this possibility, the cells were initially aligned with an alternating field of very high strength (500 V c m - 1 , peak value) for 5 s followed by a reduction of the field strength to 250 V cm-1 for another 20 s before the field pulse was applied. After pulse application the alignment was continued for 30 s (250 V cm -a, 1.5 MHz). These field conditions ensured that nearly all cells migrated towards the electrodes within a few sec- onds (Kl~Sck and Zimmermann, 1990) resulting in the formation of cell pairs or longer cell chains due to the mutual dielectrophoretic interaction between the migrating cells (Pethig, 1979). Since exposure of the cells to field strengths of this magnitude for more than 5-10 s results in irre- versible changes in the cellular structures (Zimmermann, 1982) immediate reduction of the field strength after alignment was required. As shown in Fig. 3, the use of such a modulated alternating field resulted in a slight but significant increase in the hybrid yield if the opt imum sus-

48
pension and field conditions were used (a single pulse of 1.25 kV cm -1 strength with suspension density of 1.75 • 105 H73C l l cells/ml). The scatter in yield was, however, nearly as large under these conditions as in cell alignment with an alternating field of constant, but low field strength (Fig. 3). These results suggested that the (biological and biophysical) heterogeneity of the cell lines limited the efficiency of hybrid formation and that a certain number of hybrids cannot be exceeded under these conditions. The lower the total cell number, the greater will be the influence of the heterogeneous properties of the cell lines on hy- brid yield. Such a functional relationship can be established if the field-relevant volume of the heli- cal chamber for the cells is reduced by simulta- neous maintanance of the opt imum suspension density of 1.75 • 105 H 7 3 C l l cel ls /ml. To this end the receptacle was pre-filled with 100/~1 of a high density fusion medium (75 L3 plus 20% albumin). 100/~1 of the cell suspension containing 1.75 • 1 0 4
H73Cl l cells was then overlayered onto the surface of this cushion and the electrode assembly was carefully introduced into the receptacle. Such experiments showed that the percentage yield de- creased in average to 0.05% even though the sus- pension density was adjusted to the opt imum value. This dramatic reduction in yield can be explained in part if the cell populations were very heterogeneous.
Discussion
Because the number of antigen-specific human B cells is small in peripheral blood or lymphoid tissues, methods for high efficient fusion tech- niques are required. The results reported here for fusion of the mouse-human cell line H37C l l with the mouse hybridoma cell G8 demonstrate that this goal can be achieved by electrofusion in strongly hypo-osmolar solutions. This is a highly efficient alternative to electrofusion in iso-osmolar solutions. Our results are in contrast to the finding of Stenger et al., (1988). These authors reported that the yield of mouse hybridoma decreased when the osmolarity of the medium was lowered to 180 mosM. This is expected if the field strength of the fusion (breakdown) pulse is not matched to the
concomitant increase in cell volume with decreas- ing medium osmolarity. Field strengths in the range between 2.5-3.5 kV cm 1 (as used by those authors) are too high and will lead to irreversible destruction of most of the cells in the population. The average radius of the similar sized fusion partners used here increased from about 5.9 # m to about 8.8 btm when transferred to 75 mosM fusion medium (as measured with the hydrodynamically focussing particle analyzer, see Schmitt and Zimmermann (1989)). According to the integrated Laplace equation for spherical and freely sus- pended cells *, the theoretical field strength for breakdown of membrane sites located in field direction is calculated to be 0.76 kV cm-1 (assum- ing a value of 1 V for the breakdown voltage of the membranes at room temperature, see Zimmer- mann (1982, 1986)). The opt imum field strength of 1.25 kV cm -1 found experimentally is about 50% higher than this value. A somewhat higher field strength is consistent with the literature (Zimmermann, 1986; Schmitt and Zimmermann 1989) and results from the angular dependence of the voltage to induce breakdown (see footnote on this page). The cosine a term in the Laplace equation determines the magnitude of the per- meabilized area in and close to the contact zone of the aligned cells in response to a given field strength of the pulse. Experience has shown that electrofusion requires breakdown of some part of the membranes in order to establish a cytoplasmic continuum and to initiate membrane bridging be- tween the aligned cells.
The mechanism of facilitation of electrofusion by hypo-osmolarity is unknown. However, from work of Ahkong and Lucy (1986) and of D~iumler and Zimmermann (1989), it is conceivable that the volume increase with the concomitant increase in membrane tension under hypo-osmolar conditions causes membrane- and cell-skeleton proteins to become detached. This would increase the mobil- ity of the membrane components which is re-
* Accord ing to the in tegra ted Laplace equa t ion the genera ted m e m b r a n e po ten t i a l is given by V = 1.5. a . E - c o s a, where a is the radius of a spher ical cell, E the appl ied external field s t rength and a the angle be tween the no rma l vector of a given
m e m b r a n e si te and the d i rec t ion of the external field vector.

quired for intermingling of the membranes of the aligned cells. This explanation for facilitation of electrofusion under hypo-osmolar conditions is supported by the finding that the permeabili ty of cell membranes is increased slightly (and reversi- bly) at low osmolari ty (D~iumler and Zimmer- mann, 1989).
Laser scan microscopy (Spring, Trendlenburg, Z immermann , unpublished data) and electro- nmicroscopy (Scheer, KliSck, Z immermann, un- published data) of cells subjected to 75 mosM revealed that hypo-osmolar i ty resulted in swelfing of the irregularly shaped nuclei leading to a more uniform nuclear sphere. This may facilitate the subsequent fusion process of the nuclei in the fused cells which is the crucial step in hybridiza- tion (Bertsche et al. 1988). Changes in volume and shape of the nuclei may, therefore, be a further reason for the observed increase in hybrid number under condit ions of hypo-osmolari ty. Apar t f rom the need of hypo-osmolar conditions for efficient immortal izat ion of human B cells, the efficiency of hybr idoma format ion achieved in these studies demonstrates that the hehcal electrode assembly is a suitable chamber for product ion of hybrids with a relatively small number of input cells. Compared to other electrofusion chambers (for overview see Z immermann and Vienken, 1982) the helical chamber allows easy manipulat ions of the electro- fusion process under bo th sterile and well-defined conditions. The ability to work with a hquid spacer allows the use of the same type of helical chamber for large-scale fusion as well as for fusion of a very limited number of cells. In principle, electri- cal manipulat ion of very small numbers of cells is possible. However, our experience has shown that there are sufficient differences between individuals that one cannot expect to produce the highest number of hybrids in every fusion event. There is a great heterogeneity among such cells (Perkins et al., 1990) and additional variations may occur due to their growth condit ions and period in culture. As shown here, these heterogeneities could strongly effect the yield of hybrids. The reason is that it is not possible to establish osmotic and field condi- tions (see the Laplace equation) which are opti- mally adjusted to each cell in the suspension. However, in the light of the results reported here a total number of about 104-105 input human B
49
cells can be envisaged to be sufficient to facilitate generation of an t ibody secreting human hy- br idoma lines (see also Foung et al., 1990).
Acknowledgements
The authors are grateful to Michaela Wander for skillful technical assistance. We are thankful to Andreas Hein for his help to improve the micro- scale electrofusion technique and for taking the picture of Fig. lc.
This work was supported by grants of the Deutsche Forschungsgemeinschaf t (SFB 176) and of the Federal Ministry of Research and Technol- ogy, F.R.G. ( D F V L R 01QV 354) to U.Z. and by grants HL33811, AI22557, AI26031 and DA06596 from the Nat ional Institutes of Heal th to S.K.H.F.
References
Ahkong, Q.F. and Lucy, J.A. (1986) Osmotic forces in artifi- cially induced cell fusion. Biochim. Biophys. Acta 858, 206.
Bertsche, U., Mader, A. and Zimmermann, U. (1988) Nuclear membrane fusion in electrofused mammalian cells. Bio- chim. Biophys. Acta 939, 509.
Bischoff, R., Eisert, R.M., Schedel, I., Vienken, J. and Zimmer- mann, U. (1982) Human hybridoma cells produced by electrofusion. FEBS Lett. 147, 64.
Daumler, R. and Zimmermann, U. (1989) High yields of stable transformants by hypo-osmolar plasmid electro-injektion. J. Immunol. Methods 122, 203.
Foung, S.K.H. and Perkins, S. (1989) Electric field-induced cell fusion and human monoclonal antibodies. J. Immunol. Methods 116, 117.
Foung, S.K.H., Perkins, S., Bradshaw, P., Rowe, J., Robin, L.B., Reyes, G.R. and Lenette, E.T. (1989) Human mono- clonal antibodies to human cytomegalovirus. J. Infect. Dis. 159, 436.
Foung, S., Perkins, S., Kafadar, K., Gessner, P. and Zimmer- mann, U. (1990) Development of microfusion techniques to generate human hybridomas. J. Immunol. Methods 134, 35.
Glassy, M.C. (1988) Creating hybridomas by electrofusion. Nature 333, 579.
Gravekamp, C., Santoli, D., Vreugdenhil, R., Collard, J.G. and Bolhuis, R.L.H. (1987) Efforts to produce human cytotoxic T-cell hybridomas by electrofusion and PEG fusion. Hy- bridoma 6, 121.
James, K. and Bell, G.T. (1987) Human monoclonal antibody production, current status and future prospects. J. Im- munol. Methods 100, 5.
Karsten, U., Papsdorf, G., Roloff, G., Stolley, P., Abel, H., Walther, I. and Weiss, H. (1985) Monoclonal anti-cyto-

50
keratin antibody from a hybridoma clone generated by electrofusion. Eur. J. Cancer Clin. Oncol. 21,733.
K16ck, G. and Zimmermann, U. (1990) Facilitated electrofu- sion of vacuolated x evacuolated oat mesophyU protoplasts in hypo-osmolar media after alignment with an alternating field of modulated strength. Biochim. Biophys. Acta 1025, 87.
Neil, G. and Urnovitz, H.B. (1988) Recent improvements in the production of antibody-secreting hybridoma cells. TIBTECH 6, 209.
Ohnishi, K., Chiba, J., Goto, Y. and Tokumaga, T. (1987) Improvement in the basic technology of electrofusion for generation of antibody-producing hybridomas. J. Immunol. Methods 100, 181.
Perkins, S., Zimmermann, U., Gessner, P. and Foung, S.K.H. (1990) In: C. Borrebaeck and I. Hagen (Eds.), Electro- manipulation in Hybridoma Technology, Stockton Press, New York, pp. 47.
Pethig, R. (1979) Dielectric and electronic properties of bio- logical materials. Wiley, Chichester.
Schmitt, J.J. and Zimmermann, U. (1989) Enhanced hy- bridoma production by electrofusion in strongly hypo- osmolar solutions. Biochim. Biophys. Acta 983, 42.
Schmitt, J.J., Zimmermann, U. and Gessner, P. (1989a) Elec- trofusion of osmotically treated cells; high and reproduci- ble yields of hybridoma cells. Naturwissenschaften 76, 122.
Schmitt, J.J., Zimmermarm, U. and Neil, G. (1989b) Efficient generation of stable antibody forming hybridoma cells by electrofusion. Hybridoma 8, 107.
Shulman, M., Wilde, C.D. and K0hler, G. (1978) A better cell
line for making hybridomas secreting specific antibodies. Nature 276, 269.
Stenger, D.A., Kubiniec, R.T., Purucker, W.J., Liang, H. and Hui, S.W. (!988) Optimization of electrofusion parameters for efficient production of murine hybridomas. Hybridoma 7, 505.
Vienken, J. and Zimmermann, U. (1982) Electric field-induced fusion: Electro-hydraulic procedure for production of het- erokaryon cells in high yield. FEBS Lett. 137, 11.
Vienken, J. and Zimmermann, U. (1985) An improved electro- fusion technique for production of mouse hybridoma cells. FEBS Lett. 182, 278.
Zimmermann, U. (1982) Electric field-mediated fusion and related electrical phenomena (review). Biochim. Biophys. Acta 694, 227.
Zimmermann, U. (1986) Electrical breakdown, electropermea- bilization and electrofusion. Rev. Physiol. Biochem. Pharmacol. 105, 175.
Zimmermann, U. and Vienken, J. (1982) Electric field induced cell-to-cell fusion. J. Membrane Biol. 67, 165.
Zimmermann, U., Schmitt, J.J. and Kleinhans, P. (1988) In: R. Hubbard and V. Marks (Eds.), Clinical Applications of Monoclonal Antibodies, Plenum Press, New York and London, pp. 3.
Zimmermann, U., Gessner, P., Wander, M. and Foung, S.K.H. (1990) Electroinjection and electrofusion in hypo-osmolar solution. In: C. Borrebaeck and I. Hagen (Eds.), Electro- manipulation in hybridoma technology, Stockton Press, New York, pp. 1.
Top Related

Copyright © 2022 FDOKUMEN

