







Bahasa
Halaman
Hukum


Efficacy and mechanism of action of the proteasome inhibitor PS-341 in
T-cell lymphomas and HTLV-I associated adult T-cell leukemia/lymphoma
Rihab Nasr1, Marwan E El-Sabban2, Jose-Antonio Karam1, Ghassan Dbaibo3, Youmna Kfoury1,Bertrand Arnulf4, Yves Lepelletier4, Francoise Bex5, Hugues de The6, Olivier Hermine*,4
and Ali Bazarbachi*,1
1Department of Internal Medicine, Faculty of Medicine, American University of Beirut, Beirut, Lebanon; 2Department of HumanMorphology, Faculty of Medicine, American University of Beirut, Beirut, Lebanon; 3Department of Pediatrics, Faculty of Medicine,American University of Beirut, Beirut, Lebanon; 4CNRS UMR 8603 and Department of Hematology, Necker Hospital, Paris,France; 5Department of Molecular Biology, Universite Libre de Bruxelles, Brussels, Belgium; 6UPR 9051 CNRS, Laboratoire Associeau Comite de Paris de la Ligue contre le Cancer, Paris, France
HTLV-I associated adult T-cell leukemia (ATL) andHTLV-I-negative peripheral T-cell lymphomas are asso-ciated with poor prognosis. Using pharmacological con-centrations of the proteasome inhibitor PS-341, wedemonstrate inhibition of cell proliferation and inductionof apoptosis in fresh ATL cells, HTLV-I transformed andHTLV-I-negative malignant T cells, while normal restingor activated T lymphocytes were resistant. Combinationof PS-341 and doxorubicin or etoposide resulted in anadditive growth inhibition. In HTLV-I-negative malignantcells, PS-341 treatment significantly downregulated theantiapoptotic protein X-IAP and to a lesser extent c-IAP-1 and bcl-XL and resulted in caspase-dependent apoptosis.In HTLV-I transformed cells, the inhibition of theproteasomal degradation of Tax by PS-341 likelyexplains the relative protection of HTLV-I infected cellsagainst caspase-dependent apoptosis. PS-341 treatment ofthese cells stabilized IjBa, IjBb, IjBe, p21, p27 and p53proteins and selectively inhibited Rel-A DNA binding NF-jB complexes. In both HTLV-I-positive and -negativecells, PS-341 treatment induced ceramide accumulationthat correlated with apoptosis. We conclude that PS-341affects multiple pathways critical for the survival ofHTLV-I-positive and -negative malignant T cells support-ing a potential therapeutic role for PS-341 in both ATLand HTLV-I-negative T-cell lymphomas, whether aloneor in combination with chemotherapy.Oncogene (2005) 24, 419–430. doi:10.1038/sj.onc.1208212Published online 15 November 2004
Keywords: HTLV-I; leukemia; lymphoma; treatment;proteasome inhibitors; PS-341
Introduction
The retrovirus HTLV-I is the causative agent of adultT-cell leukemia/lymphoma (ATL), an aggressivemalignancy of CD4þ T lymphocytes (reviewed inBazarbachi and Hermine, 2001). Leukemia is precededby oligoclonal expansions of HTLV-I-infected activatedT cells (Wattel et al., 1995) as a result of the viraltransactivator protein Tax expression, which activatesvarious cellular genes including CREB/ATF, AP-1and NF-kB (Sun et al., 2000), functionally inactivatesp53 and interferes with several cell cycle regulatorsincluding cyclins and cdk-inhibitors (reviewed inYoshida, 2001).
ATL is resistant to chemotherapy and carries a verypoor prognosis (reviewed in Bazarbachi and Hermine,2001). High response rates were achieved with thecombination of zidovudine and interferon-a (IFN) (Gillet al., 1995; Hermine et al., 1995; Bazarbachi andHermine, 1996). However, most patients eventuallyrelapse (Hermine et al., 2002), which underlines theneed for new therapeutic approaches. Arsenic trioxide(As) synergizes with IFN to induce G1 arrest andapoptosis in ATL (Bazarbachi et al., 1999) throughshut-off of the NF-kB pathway and Tax degradation bythe proteasome (El-Sabban et al., 2000; Nasr et al.,2003).
PS-341 is a selective inhibitor of the 26S proteasome,a critical nuclear and cytoplasmic proteolytic systemthat regulates cell proliferation, differentiation andapoptosis (reviewed in Adams, 2003). This ubiquitin–proteasome pathway normally eliminates intracellulardamaged, mutant and misfolded proteins and a varietyof short-lived functional proteins such as cyclins andcyclin-dependent kinase inhibitors, NF-kB inhibitorsand p53 (Jentsch and Schlenker, 1995; Adams, 2003). Inunstimulated cells, NF-kB is inactive, complexed in thecytosol with inhibitory subunits known as IkB. WhenIkB kinase complexes phosphorylate IkB proteins, theyare ubiquitinated and degraded by the 26S proteasome(reviewed in Ghosh and Karin, 2002). Consequently,
Received 8 July 2004; revised 6 September 2004; accepted 16 September2004; published online 15 November 2004
*Correspondence: A Bazarbachi, Department of Internal Medicine,American University of Beirut, PO Box 113-6044, Beirut, Lebanon; E-mail: [email protected] or O Hermine, Department of Hematol-ogy, Hopital Necker, 149-161 rue de Sevres, 75743 Paris Cedex 15,France; E-mail: [email protected]
Oncogene (2005) 24, 419–430& 2005 Nature Publishing Group All rights reserved 0950-9232/05 $30.00
www.nature.com/onc

free NF-kB proteins translocate to the nucleus, bindspecific promoters and activate gene transcription.
PS-341 has demonstrated clinical effect in hematolo-gical malignancies (Gatto et al., 2003; Hideshima et al.,2003; Pham et al., 2003) and solid tumors (Adams et al.,1999; Ling et al., 2003). Although PS-341 antitumoractivity correlates with suppression of NF-kB activity(Ma et al., 2003), it also affects many intracellularregulatory molecules such as p53, p21 and p27 (An et al.,2000; Hideshima et al., 2003), blocks the antiapoptoticeffects of bcl-2 and sensitizes murine myeloid leukemiacells to TRAIL-mediated apoptosis (Sayers et al., 2003).PS-341 also blocks the cell cycle at G2–M phase(Adams et al., 1999; Ling et al., 2003). In a murinemodel of ATL, PS-341 synergized with humanizedanti-Tac monoclonal antibodies and at high doses(1 mM) decreased NF-kB DNA binding activity bypreventing degradation of IkB-a (Tan and Waldmann,2002).
In this report, we have studied the effect of clinicallyachievable concentrations of PS-341 and its combina-tion with doxorubicin or etoposide on cell growthand apoptosis of fresh ATL cells, HTLV-I transformedand HTLV-I-negative malignant T cells. Our datademonstrate selective effects on malignant cells andsupport a potential therapeutic role for PS-341,either alone or in combination with chemotherapy, inpatients with ATL and other HTLV-I-negative T-celllymphomas.
Results
PS-341 induces growth arrest of malignant T cells
We used four HTLV-I-transformed cell lines (HuT-102,MT2, C8166 and C91PL) and four HTLV-I-negativeT-cell lines (CEM, Molt-4, Jurkat and HuT-78).Achievable plasma concentrations of PS-341 rangingfrom 3 to 10 nM resulted in a dose- and time-dependentgrowth inhibition of all malignant T cells (Figure 1),while treatment at 1 nM had no significant effect. Ingeneral, HTLV-I-negative cells were more sensitive toPS-341 than HTLV-I transformed cells (Po0.01;Dunnett test). Complete growth inhibition wasobserved at 24 h post-treatment in three HTLV-I-negative cell lines CEM, Molt-4 and HuT-78 at PS-341concentrations of 3–5 nM, while Jurkat cells required48 h treatment at 10 nM for maximal growth inhibition.In HTLV-I infected cells, MT-2 cells were most sensitivewith complete growth inhibition by PS-341 treatment at5 nM for 24 h while HuT-102, C8166 and C-91PLcell lines required treatment at 10 nM concentrationfor 48–72 h to achieve complete growth inhibition.Significantly, PS-341 almost completely inhibited thegrowth of resting or PHA-stimulated freshly isolatedATL cells by 48 h at 5 and 10 nM. By contrast, normal Tcells from two healthy donors were completely resistantto PS-341 treatment up to 10 nM (Figure 1 and data notshown).
Figure 1 PS-341 inhibited growth of HTLV-I-positive and -negative human T cell lines and fresh ATL cells but not normal restingand activated T-lymphocytes. Effects of PS-341 treatment on the growth of HTLV-I-positive (HuT-102, MT-2, C8166 and C91-PL),HTLV-I-negative (CEM, Jurkat, Molt-4 and HuT-78) human T cell lines, resting and activated fresh ATL cells and normal resting orPHA-activated peripheral blood mononuclear cells (PBMC). Activated ATL cells or normal PBMC were supplemented with 2% PHA.PS-341 was added at the indicated concentrations for 24–72 h. Cell growth (% control) was assayed in quadruplicate wells with theCellTiter 96s nonradioactive cell proliferation kit
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
420
Oncogene

PS-341 treatment induces apoptosis in malignant T cells
Using TUNEL assay we investigated the involvement ofapoptosis in PS-341-induced growth inhibition ofHTLV-I-positive and -negative malignant T-cells. After48 h of culture, the percentage of TUNEL-positiveHuT-102 cells reached 25% with 10 nM PS-341 whileCEM cells were more sensitive to PS-341-inducedapoptosis reaching 92% TUNEL-positive cells with5 nM PS-341 (Figure 2a). Similarly, under fluorescencemicroscopy, PS-341 treatment resulted in a significantincrease in the number of TUNEL-positive apoptoticMT-2 and CEM cells (Figure 2b and data not shown).
PS-341-induced apoptosis was also associated with adose-dependent dissipation of mitochondrial membranepotential as shown by the decrease in Rhodamine-123fluorescence. This effect was evident in all tested ATL-derived and HTLV-I-negative cells (Po0.01; Dunnetttest) (Figure 2c). As expected, the dissipation ofmitochondrial membrane potential correlated withsensitivity to PS-341 with a more moderate decreaseobserved in HTLV-I-positive cells.
PS-341 treatment causes cell cycle arrest in malignantT cells
Cell cycle analyses of all tested cell lines showed thatPS-341 induced a significant dose-dependent increase inthe pre-G0/G1 region, regarded as apoptotic cells(Po0.01; Dunnett test). This increase was moresignificant in HTLV-I-negative cells in agreementwith their higher sensitivity to PS-341 as compared toHTLV-I-positive cells (data not shown).
PS-341 treatment also induced significant changes inthe cell cycle distribution. Figure 3 represents therelative distribution of nonapoptotic cells between G0/G1 phases (black boxes) and cycling cells, the sum of(SþG2/M) phases (white boxes). In HTLV-I-negativecells treated for 24 h, cycling CEM cells significantlyincreased from 40% in the control cells to 61 and 64%with 3 and 5 nM PS-341 respectively (Po0.01). Similarresults were observed with Molt-4 cells (Figure 3 anddata not shown), suggesting that PS-341 treatmentinduces a G2/M cell cycle arrest, as reported for othercell types (Adams et al., 1999; Ling et al., 2003).Interestingly, PS-341 treatment resulted in a completelydifferent pattern of changes in the cell cycle distributionof HTLV-I-positive cells. Indeed, after 24 h treatment,cycling HuT-102 and C91-PL cells diminished from 37and 47% respectively in the control cells to 29 and 36%,respectively, with 3 nM PS-341 and to 26 and 34%respectively with 5 nM PS-341, while treatment with10 nM PS-341 resulted in a secondary increase of cyclingcells to 33 and 47% respectively (Figure 3). The samedistribution was maintained in HuT-102 cells after 48 h.Similar results were also observed in C8166 cells whileMT-2 cells showed a decrease in cycling cells from 50 to47% with 3 nM PS-341 and a secondary increase to 54and 56% with 5 and 10 nM PS-341, respectively, again inagreement with their higher sensitivity to PS-341 ascompared to other HTLV-I-positive cells (data not
shown). These results suggest that PS-341 treatmentresults in a moderate G0/G1 arrest of HTLV-I-positivecells at concentrations insufficient to inhibit growthfollowed by a G2/M arrest at the growth inhibitoryconcentrations (10 nM for HuT-102, C-91PL and C8166;5 and 10 nM for MT-2).
PS-341-induced apoptosis is caspase-dependentin HTLV-I-negative cells
PS-341-induced loss of mitochondrial membrane poten-tial may result in the activation of the caspase cascade.Indeed, in the HTLV-I-negative cells CEM and Molt-4,PS-341-induced apoptosis was associated with caspaseactivation, as shown by complete PARP cleavage(113 kDa) into its death-associated fragments (89 and24 kDa). A partial cleavage was observed in HuT-102and MT-2 cells (Figure 4a). Furthermore, PS-341treatment resulted in complete cleavage of procaspase3, procaspase 8 and procaspase 9 in all tested HTLV-I-negative cells while only a modest cleavage could bedetected in some HTLV-I-positive cell lines, only at thehighest PS-341 concentration (Figure 4a and data notshown). Importantly, PS-341-induced growth inhibitionof the HTLV-I-negative cells Molt-4, CEM and Jurkatcells was partially but significantly reversed by thecaspase inhibitor zVAD (Po0.01) (Figure 4b). Incontrast, PS-341 induced growth inhibition of theHTLV-I-positive cells was not reversed by zVAD exceptfor MT-2 cells (Po0.05) (Figure 4b). Caspase inhibitorsare known to inhibit cell cycle progression in T and Blymphocytes (Alam et al., 1999; Olson et al., 2003).Hence, the protective effect of zVAD demonstrates theinvolvement of caspase activation and caspase-depen-dent apoptosis in PS-341-induced growth inhibition ofHTLV-I-negative cells but not ATL-derived cells.
PS-341 treatment downregulates the expression of theanti-apoptotic protein XIAP in HTLV-I-negative cells only
Tax has been reported to prevent apoptosis and caspaseactivation (Kawakami et al., 1999) through upregula-tion of the NF-kB-dependent XIAP (X-linked inhibitorof apoptosis), c-IAP-1 (inhibitor of apoptosis-1) andbcl-XL proteins. PS-341 treatment of HTLV-I-positivecells resulted in a delayed increase of bax at 48 h with10 nM PS-341 and a minimal decrease in bcl-XL andc-IAP-1 in some tested cell lines, whereas XIAP and bcl-2protein levels were unaffected (Figure 5a and data notshown). In HTLV-I-negative malignant T cells, PS-341treatment had no significant effect on bcl-2 protein level,increased bax in CEM cells but dramatically down-regulated the protein level of XIAP and to a lesser extentc-IAP-1, and bcl-XL in all tested HTLV-I-negative celllines likely explaining the caspase-dependent apoptosis.
PS-341 treatment stabilized IkB proteins and reversed theconstitutive NF-kB activation in HTLV-I-positive cells
Tax has been shown to target IkB-a, IkB-b and IkB-e forproteasomal degradation (Kanno et al., 1994; Sun et al.,
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
421
Oncogene
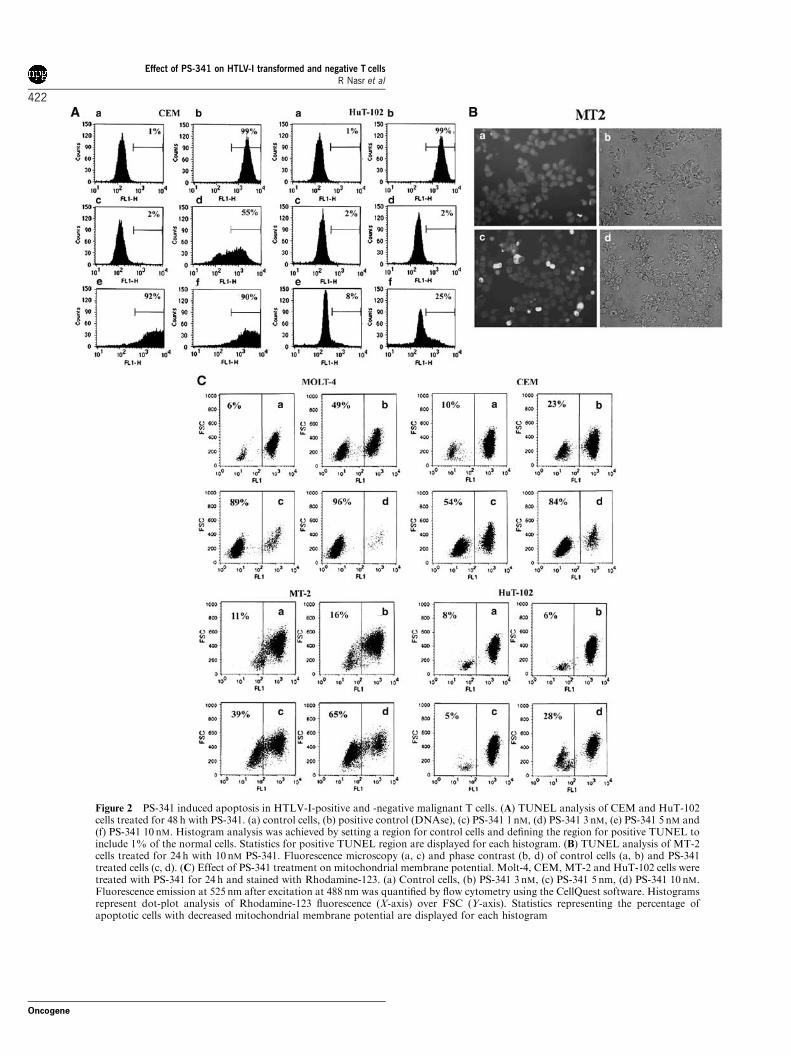
Figure 2 PS-341 induced apoptosis in HTLV-I-positive and -negative malignant T cells. (A) TUNEL analysis of CEM and HuT-102cells treated for 48 h with PS-341. (a) control cells, (b) positive control (DNAse), (c) PS-341 1 nM, (d) PS-341 3 nM, (e) PS-341 5 nM and(f) PS-341 10 nM. Histogram analysis was achieved by setting a region for control cells and defining the region for positive TUNEL toinclude 1% of the normal cells. Statistics for positive TUNEL region are displayed for each histogram. (B) TUNEL analysis of MT-2cells treated for 24 h with 10 nM PS-341. Fluorescence microscopy (a, c) and phase contrast (b, d) of control cells (a, b) and PS-341treated cells (c, d). (C) Effect of PS-341 treatment on mitochondrial membrane potential. Molt-4, CEM, MT-2 and HuT-102 cells weretreated with PS-341 for 24 h and stained with Rhodamine-123. (a) Control cells, (b) PS-341 3 nM, (c) PS-341 5 nm, (d) PS-341 10 nM.Fluorescence emission at 525 nm after excitation at 488 nm was quantified by flow cytometry using the CellQuest software. Histogramsrepresent dot-plot analysis of Rhodamine-123 fluorescence (X-axis) over FSC (Y-axis). Statistics representing the percentage ofapoptotic cells with decreased mitochondrial membrane potential are displayed for each histogram
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
422
Oncogene

1994; Nasr et al., 2003). Hence, proteasome inhibitionby PS-341 would be expected to diminish IkB proteo-lysis in Tax-positive cells. Indeed, in HuT-102, MT2,C8166 and C91-PL cells, Western blot analysis demon-strated that PS-341 treatment resulted in a significantincrease in IkB-a, IkB-b and IkB-e protein level in a dosedependent manner (Figure 6a and data not shown). Incontrast, the level of IkB-a, IkB-b and IkB-e was eitherunaffected or even decreased in the four tested HTLV-I-negative cells (Figure 6a and data not shown).
We then examined whether, following stabilization ofthe NF-kB inhibitors, PS-341 treatment could modulatethe NF-kB functional assembly by assessing DNAbinding NF-kB complexes. We first confirmed theconstitutive expression of NF-kB in the HTLV-I-positive cells HuT-102, MT2, C8166 and C91-PL cellsby EMSA. One predominant positive complex wasdetected by the 32P-labeled NF-kB consensus oligonu-cleotide (Figure 6b and data not shown). This complexdisappeared in the presence of excess of unlabeled NF-kB consensus oligonucleotide, but not with excessunlabeled mutant oligonucleotide (Figure 6b), confirm-ing its specificity. PS-341 exposure resulted in a dose-dependent inhibition of NF-kB complex formation inMT-2 cells where the NF-kB complex was slightlyreduced upon exposure to 3 nM PS-341 and drastically
reduced by exposure to 5–10 nM PS-341 (Figure 6b). InHuT-102 and C91-PL cells, PS-341 resulted in a drasticreduction of the NF-kB complexes at 10 nM only(Figure 6b and data not shown). As expected, inHTLV-I-negative cells (CEM, Jurkat and MolT-4), gelshift assays using NF-kB consensus oligonucleotideshowed a faint band complex reflecting the absence ofconstitutive NF-kB activation.
The specificity of these NF-kB complexes was furtherconfirmed using RelA and p50 specific antibodies toinduce a super-shift. As previously reported (El-Sabbanet al., 2000), in HTLV-I-positive cells, RelA and p50lead to super-shifting of a fraction of the NF-kBcomplexes (Figure 6c and data not shown). In all testedHTLV-I-positive cells (HuT-102, MT-2 and C91-PL),RelA containing complexes were drastically reducedupon exposure to 5–10 nM PS-341 (Figure 6c and datanot shown) while p50 containing complexes were slightlyreduced in HuT-102 cells and remained unaffected inC91-PL cells (data not shown). This dramatic andspecific decrease in RelA containing complexes isconsistent with enhanced expression of IkB-a, IkB-band IkB-e, which sequester RelA to the cytoplasm.Indeed, using anti-RelA antibodies, a mixed homoge-neous nuclear and cytoplasmic staining was observed inthe majority of untreated MT-2 cells, while cells treatedwith PS-341 showed exclusively cytoplasmic staining(Figure 6d).
PS-341 inhibited Tax degradation and upregulated p53,p21 and p27 proteins in ATL-derived cells
We have previously reported a significant stabilizationof Tax in HTLV-I infected cells treated with theproteasome inhibitor zLLL (Nasr et al., 2003). Similarresults were observed with PS-341. Treatment of HuT-102, MT-2 and C91PL cells with 5–10 nM PS-341resulted in a significant upregulation of Tax proteinlevel (Figure 5c and data not shown). These resultsconfirm our previous report of a continuous protea-some-mediated degradation of Tax in HTLV-I-positivecells. On the other hand, treatment of these cells with3–10 nM PS-341 resulted in a significant increase inp21, p27 and p53 protein level in a dose-dependentmanner, indicating an inhibition of their baselineproteasomal degradation (Figure 5b). In contrast, inthe HTLV-I-negative cells CEM, Molt-4 and Jurkat,PS-341 treatment slightly upregulated p53 while p21and p27 protein levels were unaffected. These resultssuggest a potential involvement of the p53 pathway inPS-341-induced growth inhibition of HTLV-I-positivecells.
PS-341 induced ceramide accumulation in malignantT cells
Ceramide, a coordinator of the cellular response tostress (Hannun, 1996), has several growth suppressiveeffects including cell cycle arrest (Dbaibo et al., 1995)and apoptosis (Obeid et al., 1993). In both HTLV-I-positive and -negative cells, PS-341 treatment resulted in
Figure 3 PS-341 induced G2/M cell cycle arrest. Effects of PS-341on the cell cycle distribution of HuT-102, MT-2, CEM and Molt-4cells. PS-341-treated cells were stained with propidium iodide (PI)(50mg/ml) and cell cycle analysis was performed using a FACScanflow cytometer. Histograms represent the relative distribution ofnonapoptotic cells between G0/G1 phases (black boxes) and thesum of (SþG2/M) phases (white boxes) after 24 h treatment unlessindicated otherwise. Results are representative of two independentexperiments
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
423
Oncogene

dramatic ceramide accumulation in a dose- and time-dependent manner that correlated with the induction ofcell death, potentially implicating the ceramide pathwayin PS-341-induced growth arrest and apoptosis. PS-341treatment of MT-2 cells increased intracellular ceramideto 4.5-fold at 48 h with 10 nM PS-341 (Po0.01)(Figure 5d), while HuT-102 cells, which are less sensitiveto PS-341, showed a lower increase in ceramide. HTLV-I-negative cells displayed a similar correlation betweensensitivity to PS-341 and ceramide accumulation reach-ing eight-fold at 24 h for CEM cells and seven-fold at48 h for Jurkat cells treated with 10 nM PS-341 (Po0.01)(Figure 5d).
PS-341 has an additive growth inhibition with doxorubicinand etoposide in ATL-derived and HTLV-I-negativemalignant T cells
NF-kB activation protects tumor cells from chemother-apy-induced apoptosis (Baeurle and Baltimore, 1996;Huang et al., 2000). Therefore, PS-341-induced reversalof NF-kB activation could restore the chemosensitivity ofATL-derived cells. We investigated the effect of PS-341 incombination with doxorubicin and etoposide. After 24htreatment, the growth of Molt-4 cells was reduced to 62%of control with 3 nM PS-341, 20% with 1mM etoposide,50% with 100 nM doxorubicin, 10% with the combina-tion PS-341 and etoposide and 20% with the combination
Figure 4 PS-341-induced apoptosis is caspase-dependent in HTLV-I-negative cells. (a) Effect of PS-341 on the cleavage of PARP(arrow) and procaspases in HTLV-I-positive and -negative cells. Cells were treated for 24 h with PS-341 at the indicated concentration.Total SDS protein lysates (30mg/lane) were prepared and immunoblotted against PARP and caspases 3, 8 and 9 antibodies. (b) Effectsof the general caspase inhibitor z-VAD (100mM) on the growth inhibition induced by PS-341. For each cell line, PS-341 was used at aconcentration and treatment duration that induced a significant but not complete growth inhibition: 3 nM for MOLT-4; 5 nM for HuT-102, MT-2, C-8166 and CEM; 10 nM for Jurkat; 24 h for HTLV-I-negative cells and 48 h for HTLV-I-positive cells. Cell growth wasassayed in quadruplicate wells with the CellTiter 96s nonradioactive cell proliferation kit. Results are expressed as percentage ofcontrol7s.d.
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
424
Oncogene

of PS-341 and doxorubicin (Po0.01) (Figure 7). CEMcells showed a similar additive effect of the combinationof PS-341 and doxorubicin (Po0.05) but not etoposide(Figure 7). In HTLV-I-positive cells, after 48h treatment,the growth of HuT-102 cells was reduced to 54% ofcontrol with 5nM PS-341, 46% with 1mM etoposide, 62%with 100nM doxorubicin, 34% with the combination PS-341 and etoposide, and 31% with the combination of PS-341 and doxorubicin (Po0.05) (Figure 7). These resultsindicate that the combination of PS-341 and doxorubicinresulted in an additive rather than synergistic effect inCEM, Molt-4 and HuT-102 cells, while the combination ofPS-341 and etoposide resulted in an additive effect mostlyin Molt-4 cells. The additive effect of these combina-
tions was lost when lower doses of PS-341 and/or chemo-therapy were used (data not shown). Interestingly, MT-2cells in which growth and NF-kB activity are more inhi-bited by PS-341 compared to other HTLV-I-positive cells,displayed a modest additive effect of PS-341 with etoposideand doxorubicin, suggesting that the constitutive activationof the NF-kB pathway in HTLV-I-positive cells plays alittle role in their resistance to chemotherapeutic agents.
Discussion
We show that clinically achievable concentrations of PS-341 induce growth inhibition, cell cycle arrest and
Figure 5 PS-341 affects multiple pathways critical for the survival of HTLV-I-positive and -negative malignant T cells. (a) Effect ofPS-341 treatment on the protein level of the apoptosis regulatory proteins bcl-2, bcl-xL and bax or the caspase inhibitors XIAP and c-IAP-1. Cells were treated with PS-341 at the indicated concentration. Total SDS protein lysates (30mg/lane) were prepared andimmunoblotted as described with specific antibodies. Protein levels of bax and c-IAP-1 in HuT-102 and MT2 cells are presented after48 h treatment, other results are those after 24 h treatment. (b) Effect of PS-341 treatment on the protein level of p21, p27 and p53. (c)Effect of PS-341 treatment on the protein level of Tax in HTLV-I-positive cells. (d) PS-341 treatment induced ceramide accumulation:cells were treated with PS-341 at the indicated concentration. Ceramide levels were determined after 24 and 48 h using the DGK assayas described in ‘Materials and methods’ and normalized to total cellular inorganic phosphate levels. Results are expressed as foldincrease in total ceramide levels7s.d.
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
425
Oncogene

apoptosis in all tested malignant T cells, while no effectwas observed on resting and activated normal T-lymphocytes. The mechanism for this differentialsensitivity of transformed cells to PS-341, which hasbeen reported in other cell types, remains to beelucidated.
In HTLV-I-negative malignant T cells, PS-341 treat-ment significantly downregulated the antiapoptoticprotein X-IAP and to a lesser extent c-IAP-1, andbcl-XL, induced ceramide accumulation and resulted ina significant loss of mitochondrial membrane potentialand caspase activation. Importantly, PS-341-inducedgrowth inhibition was partially but significantly reversedby caspase inhibitors, which inhibit cell cycle progres-sion in T and B lymphocytes (Alam et al., 1999; Olsonet al., 2003). Hence, the protective effect of zVAD in PS-341-treated HTLV-I-negative cells demonstrate theinvolvement of caspase activation and caspase-depen-dent apoptosis in PS-341-induced growth inhibition ofHTLV-I-negative cells.
In contrast, a different mechanism of action of PS-341was observed in HTLV-I infected cells. In agreementwith recent reports (Tan and Waldmann, 2002; Mitra-Kaushik et al., 2004; Satou et al., 2004), we show that inHTLV-I transformed cells, PS-341 treatment resulted ina dramatic stabilization of IkB-a, IkB-b and IkB-eproteins and reversal of NF-kB activation with a moreselective inhibition of Rel-A DNA binding NF-kBcomplexes. Previous studies clearly established theimportance of NF-kB activation and RelA containingcomplexes in both HTLV-I induced transformation andresistance to apoptosis (Kitajima et al., 1992; Yamaokaet al., 1996).
Although RelA appears to be a major target of PS-341, the effects of this proteasome inhibitor on HTLV-I-positive cells were not limited to the inhibition of theNF-kB pathway. PS-341 treatment resulted in adramatic stabilization of p21, p27 and p53 proteins ina dose-dependent manner. In HTLV-infected cells, p53is sometimes mutated but most frequently functionally
Figure 7 Additive inhibitory effect of PS-341 and chemotherapyon the growth of HTLV-I-positive and -negative cells. Cells weretreated with PS-341 (3 nM for MT-2, CEM and Molt-4; 5 nM forHuT-102) and/or etoposide (1mM) or doxorubicin (100 nM) for 24 h(CEM and Molt-4) or 48 h (HuT-102 and MT-2). For each cell line,PS-341 was used at a concentration and treatment duration thatdid not induce major growth inhibition with PS-341 alone. Cellgrowth was assayed in quadruplicate wells with the CellTiter 96s
nonradioactive cell proliferation kit. Results are expressed aspercentage of control7s.d.
Figure 6 PS-341 reversed NF-kB activation in HTLV-I-positivecells. (a) Effect of PS-341 treatment on the protein level of the NF-kB inhibitors IkB-a, b and e. Cells were treated with PS-341 at theindicated concentration for 24 h. Total SDS protein lysates (30mg/lane) were prepared and immunoblotted as described with specificantibodies. (b) Effects of PS-341 treatment on NF-kB DNAbinding activity assessed by EMSA using consensus oligonucleotideprobe. (c) Effects of PS-341 treatment on p65/RelA-containing NF-kB complexes assessed by adding anti RelA antibodies resulting inpartial ‘super-shift’ (arrow). (d) Effects of PS-341 treatment (10 nM)on the subcellular distribution of the NF-kB subunit RelA byimmuno-fluorescence
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
426
Oncogene

inactive (Cereseto et al., 1996; Mahieux et al., 2000) andTax-dependent activation of the NF-kB pathway wasreported to play a critical role in the inhibition of p53transactivation function (Pise-Masison et al., 2000).Hence, PS-341-induced p53 accumulation, together withthe reversal of NF-kB activation, should restore normalp53 function. The upregulation of p21 and the delayedupregulation of bax, two well characterized p53 targets,argue in favor of this hypothesis. Interestingly, PS-341effects on p21 and p27 proteins were limited to HTLV-I-positive cells, whereas no significant change was seen inHTLV-negative malignant T cells. Tax has beenreported to transactivate the p21 promoter and, conse-quently, p21 expression is upregulated at both thetranscript and protein levels in HTLV-I infected cells(Cereseto et al., 1996; De La Fuente et al., 2000).However, p21 effects on preventing apoptosis andstimulating growth of HTLV-I-positive cells (Kawataet al., 2003) are described in a cellular environment,where NF-kB is constitutively activated and p53transcriptional activity is inhibited. In the absence ofNF-kB activity and in the presence of active p53, PS-341-induced accumulation of p21 and p27 should resultin cell cycle arrest rather than growth promotion andprotection from apoptosis.
As for HTLV-I-negative cells, PS-341 treatment ofHTLV-I infected cells resulted in ceramide accumulationand loss of mitochondrial membrane potential, whichmay explain caspase activation. However, activation ofcaspase was only partial and importantly, PS-341-induced growth inhibition of HTLV-I-positive cellswas not reversed by zVAD, except partially for MT-2cells. That HTLV-I-negative cells are more sensitivethan their HTLV-I-positive counterpart to PS-341effects is in agreement with previous reports that Taxprotects HTLV-I-positive cells from caspase-dependentapoptosis (Kawakami et al., 1999) through upregulationof NF-kB-dependent apoptosis inhibitors such as bcl-XL
or caspase inhibitors such as c-IAP-1 and XIAP.However, despite complete reversal of NF-kB activa-tion, PS-341 treatment had no detectable effect on XIAPand bclXL proteins levels in ATL-derived cells, contraryto HTLV-I-negative cells. These results, together withthe inhibition of the proteasomal degradation of Tax byPS-341, suggest that Tax protects HTLV-I infected cellsagainst caspase-dependent apoptosis through an NF-kBindependent upregulation of XIAP. The mechanismunderlying downregulation of XIAP protein levels inPS-341-treated HTLV-I-negative cells remains to beelucidated.
Although Tax is undetectable in circulating ATL cells,our demonstration that Tax is continuously degraded bythe proteasome may account for the subsequentpresentation of Tax peptides on MHC class I molecules(Lomas et al., 2002) and for the high frequency ofcirculating Tax-specific cytotoxic T lymphocytes foundin most HTLV-I-infected individuals (reviewed inBangham, 2000). ATL cells may display a latent,transient and/or low level of Tax expression inpreferential sites involved by tumor cells such as skin,gastrointestinal tract and/or other lymphoid organs.
Since Tax is an essential activator of the viral geneexpression, which is absolutely indispensable to replica-tion, PS-341 treatment may be clinically useful not onlyin ATL but also in patients with tropical spasticparaparesis/HTLV-I associated myelopathy, in chronicHTLV-I carriers with high viral loads or during primo-infection.
Combination of PS-341 and the chemotherapeuticagents doxorubicin or etoposide resulted in an additiveinhibition of cell growth in most tested cells. Conven-tional chemotherapy has limited benefit in ATL patientsgiven that HTLV-1 cells carry an intrinsic resistance tochemotherapy due to frequent overexpression of themultidrug resistance protein (Lau et al., 1998) and thelung resistance protein (Ikeda et al., 1999). In addition,the constitutive activation of NF-kB, the inhibition ofp53 function and the downregulation of Fas-ligandexpression in HTLV-I-positive cells (Tamiya et al., 1998)likely protect these cells from chemotherapy-inducedapoptosis. Hence, the reversal of NF-kB activation byPS-341 and the potential restoration of p53 functionshould enhance the chemosensitivity of HTLV-I-posi-tive cells resulting in a synergistic inhibition of cellgrowth. In that sense, in murine models of ATL, PS-341synergized with humanized anti-Tac monoclonal anti-bodies (Tan and Waldmann, 2002) or with g-irradiation(Mitra-Kaushik et al., 2004) while single agents hadlittle effect. However, our results showed that combina-tion of PS-341 with doxorubicin and etoposide resultedin additive effect in most tested ATL-derived andHTLV-I-negative malignant T cells but no real synergywas noted. Some tested cell lines did not even showan additional growth inhibition with the combinedtherapy. This suggests that PS-341 and testedchemotherapeutic agents have distinct cellular targetsand that activation of NF-kB and inhibition of p53function play a marginal role in the chemo-resistance ofATL cells.
ATL remains of poor prognosis. Novel effective drugsare warranted to reduce the emergence of resistantclones. In addition, HTLV-I-negative peripheral T-celllymphomas are also associated with poor prognosis dueto a lower response rate to anthracyclin-based combina-tion chemotherapy and a higher relapse rate ascompared to B-cell lymphomas (Gisselbrecht et al.,1998). Our results demonstrate that PS-341 affectsmultiple cellular pathways critical for the survival ofboth HTLV-I-positive and -negative malignant T cellsresulting in growth arrest and apoptosis at clinicallyachievable concentrations. Hence, these findings sup-port a potential therapeutic role for PS-341 in both ATLand HTLV-I-negative T-cell lymphomas, either alone orin combination with chemotherapy.
Materials and methods
Cells, drugs and antibodies
The HTLV-I-infected CD4þ T cell lines HuT-102, MT-2,C91PL and C8166, and the HTLV-I-negative CD4þ T cell
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
427
Oncogene

lines CEM, Jurkat, HuT-78, and MOLT-4 were grown aspreviously described (El-Sabban et al., 2000). Peripheral bloodmononuclear cells (PBMC) were isolated, after informedconsent, from a patient with acute ATL and two healthyHTLV-I-negative donors, using Ficoll-Hypaque (Lympho-prep, Nyegaard, Norway). Before PS-341 addition, cells werecultured for 24 h at 1� 105 cells/ml in RPMI-1640 mediumcontaining 10% fetal calf serum (FCS) (Gibco, Invitrogen,Paisley, UK) and antibiotics. Activated PBMC were grown inHam’s F10 medium (Gibco) supplemented with 2% PHA(Gibco).
PS-341 was provided by Millenium Pharmaceuticals (Cam-bridge, MA, USA). The caspase inhibitor zVAD was obtainedfrom Bachem Bioscience (King of Prussia, PA, USA).Doxorubicine and etoposide were obtained from SigmaChemical (St Louis, MO, USA).
Antibodies: IkB-a (c-20), IkB-b (c-19), IkB-e (M-364), p21,p27, bcl-2, bcl-XL, bax, XIAP, c-IAP-1, PARP, caspases 3, 8and 9 were obtained from Santa Cruz Biotechnology Inc(Santa Cruz, CA, USA). Mouse monoclonal anti-GAPDHwas obtained from Biogenesis (Stinford Fload, UK). Mousemonoclonal anti-Tax (168-A51) was obtained from the NIHAIDS Research and Reagent Program. Rabbit polyclonalantiactin was obtained from Sigma. Rabbit polyclonal anti-p53 was purchased from Chemicon International (Temecula,CA, USA).
Growth and cytotoxicity assays
Cell growth was assessed by cell count using trypan blue dyeexclusion protocols and the CellTiter 96s cell proliferationassay kit (Promega Corp., Madison, WI, USA). Results areexpressed as cell growth relative to untreated controls and arederived from the mean of quadruplicate wells. Results arerepresentative of at least three independent experiments.
TUNEL assay
The terminal deoxytransferase (TdT)-mediated deoxyuridinetriphosphate (dUTP) nick-end labeling (TUNEL) assay(Boehringer, Mannheim, Germany) was used. Approximately10 000 cells per sample were acquired and analysed using flowcytometry (FACScan, Becton Dickinson, San Jose, CA, USA)or fluorescent microscopy. Results were analysed using theCellQuest software. Results are representative of two indepen-dent experiments.
Measurement of mitochondrial membrane potential
Quantification of mitochondrial membrane potential wasdetermined by rhodamine (R123) (Sigma) retention. PS-341-treated cells were loaded with 5 mM R123 for 30min at 371C,washed twice and assessed using flow cytometry. Results arerepresentative of two independent experiments.
Cell cycle analysis
PS-341 treated cells were washed twice with cold PBS, fixed inice cold 100% ethanol, and stored for 24 h at 41C.Subsequently, cells were rinsed with PBS, incubated for 1 hin Tris-HCl buffer (pH 7.4) containing 50 U RNase A and thenstained with propidium iodide (PI) (50 mg/ml) (Sigma). Cellcycle analysis was performed using a flow cytometer. Eachsample was collected as 10 000 ungated events and analysedusing Cell-Quest software. Results are representative of twoindependent experiments.
Immunoblot analysis
Total cellular proteins were electrophoresed, and transferredonto nitrocellulose membranes (Bio-Rad Laboratories). Theblots were blocked for 1 h at room temperature in 5%skimmed milk in TBS (50mM Tris-HCl and 150 mM NaCl)and probed overnight with primary antibody at 41C. Antibodybinding to specific proteins was visualized by enhancedchemiluminescence using the ECL system (Amersham, Bu-chinghamshire, UK).
Electrophoretic mobility shift assay (EMSA)
Nuclear extracts were prepared as described (El-Sabban et al.,2000). Protein concentration was determined using the Bioradassay (Bio-Rad Laboratories, Hercules, CA, USA). NF-kBconsensus oligonucleotide (Santa Cruz, CA, USA) was end-labeled with g-32P ATP using T4 polynucleotide kinase.Hybridization reaction was performed as described (El-Sabbanet al., 2000). The reaction mixture was electrophoresed on a5% nondenaturing polyacrylamide gel, dried at 801C undervacuum for 2 h on Whattman filter paper, and processed forautoradiography. Specific NF-kB bands were determined bycompetition experiments using a mutant oligonucleotide thathas lost its ability to bind to the transcription factor. Subunitspecificity was determined by using specific antibodies to theNF-kB components (anti-RelA, p50, c-Rel and p52) in theincubation step, known to super-shift NF-kB complexes.
Immunofluorescence
Cells were fixed in 70% ice-cold ethanol and processed forindirect immunofluorescence. Samples were blocked by in-cubation with normal goat serum (10% in PBS), reacted withanti-RelA antibodies for 2 h at room temperature, washed inPBS and incubated with FITC-conjugated secondary anti-bodies. Nuclear translocation of RelA was assessed byfluorescence microscopy (LSM 410, Zeiss, Germany).
Ceramide measurement
Ceramide was measured with a modified diacylglycerol kinaseassay using external ceramide standards as described (Blighand Dyer, 1959; Bielawska et al., 2001). Briefly, dried lipid wassolubilized in (7.5% octyl-b-D-glucoside, 25 mM dioleoylphosphatidylglycerol) solution. To the lipid micelles, reactionbuffer was added, and the reaction was started by adding[g-32P]ATP solution and allowed to proceed at 251C for30 min. Lipid extracts were resuspended in methanol/chloro-form (1 : 9, v/v) and spotted on silica gel thin layerchromatography plate. Plates were developed with chlorofor-m : acetone :methanol : acetic acid :H2O (50 : 20 : 15 : 10 : 5),air dried, and subjected to autoradiography. The radioactivespots corresponding to phosphatidic acid and ceramide-phosphate, the phosphorylated products of diacylglyceroland ceramide, respectively, were identified by comparison toknown standards, scraped and counted on a scintillationcounter.
Statistical analysis
Four statistical tests were performed: The Fisher test,the Dunnett test, the two-way analysis of variance(ANOVA) test, and the two-sample t-test as described (Zar,1999).
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
428
Oncogene

Acknowledgements
We thank Dr Kamal Badr for critical reading of thismanuscript. The expert assistance from the personnel of thecore laboratory facilities of the American University of Beirutis greatly appreciated. This paper is supported by the
American University of Beirut University Research Boardand Medical Practice Plan, the Lebanese National Council forScientific Research, the Diana Tamari Sabbagh Foundation,the CNRS, ARECA, HERN, the Eli Lilly InternationalFoundation and the Lady TATA memorial trust.
References
Adams J. (2003). Cancer Treat. Rev., 29, 3–9.Adams J, Palombella VJ, Sausville EA, Johnson J, Destree A,
Lazarus DD, Maas J, Pien CS, Prakash S and Elliott PJ.(1999). Cancer Res., 59, 2615–2622.
Alam A, Cohen LY, Aouad S and Sekaly RP. (1999). J. Exp.Med., 190, 1879–1890.
An WG, Hwang SG, Trepel JB and Blagosklonny MV. (2000).Leukemia, 14, 1276–1283.
Baeurle PA and Baltimore D. (1996). Cell, 87, 13–20.Bangham CR. (2000). Curr. Opin. Immunol., 12,
397–402.Bazarbachi A, El-Sabban ME, Nasr R, Quignon F,
Awaraji C, Kersual J, Dianoux L, Zermati Y, Haidar JH,Hermine O and de The H. (1999). Blood, 93,
278–283.Bazarbachi A and Hermine O. (1996). J. Acquir. Immune Def.
Syndrome Hum. Retrovirol., 13, S186–S190.Bazarbachi A and Hermine O. (2001). Virus Res., 78,
79–92.Bielawska A, Perry DK and Hannun YA. (2001). Anal.
Biochem., 298, 141–150.Bligh EG and Dyer WJ. (1959). Can. J. Biochem. Phys., 37,
911–917.Cereseto A, Diella F, Mulloy JC, Cara A, Michieli P,
Grassmann R, Franchini G and Klotman ME. (1996).Blood, 88, 1551–1560.
Dbaibo GS, Pushkareva MY, Jayadev S, Schwarz JK,Horowitz JM, Obeid LM and Hannun YA. (1995). Proc.Natl. Acad. Sci. USA, 92, 1347–1351.
De La Fuente C, Santiago F, Chong SY, Deng L, Mayhood T,Fu P, Stein D, Denny T, Coffman F, Azimi N,Mahieux R and Kashanchi F. (2000). J. Virol., 74,
7270–7283.El-Sabban ME, Nasr R, Dbaibo G, Hermine O, Abboushi N,
Quignon F, Ameisen JC, Bex F, de The H and BazarbachiA. (2000). Blood, 96, 2849–2855.
Gatto S, Scappini B, Pham L, Onida F, Milella M, Ball G,Ricci C, Divoky V, Verstovsek S, Kantarjian HM, KeatingMJ, Cortes-Franco JE and Beran M. (2003). Haematologica,88, 853–863.
Ghosh S and Karin M. (2002). Cell, 109, S81–S96.Gill PS, Harrington W, Kaplan MH, Ribeiro RC, Bennett JM,
Liebman HA, Bernstein-Singer M, Espina BM, Cabral L,Allen S and Levine A. (1995). N. Engl. J. Med., 332,
1744–1748.Gisselbrecht C, Gaulard P, Lepage E, Coiffier B, Briere J,
Haioun C, Cazals-Hatem D, Bosly A, Xerri L, Tilly H,Berger F and Diebold J. (1998). Blood, 92, 76–82.
Hannun YA. (1996). Science, 274, 1855–1859.Hermine O, Allard I, Levy V, Arnulf B, Gessain A and
Bazarbachi A. (2002). Hematol. J., 3, 276–282.Hermine O, Bouscary D, Gessain A, Turlure P, Leblond V,
Franck N, Buzyn-Veil A, Rio B, Macintyre E, Dreyfus Fand Bazarbachi A. (1995). N. Engl. J. Med.,332, 1749–1751.
Hideshima T, Mitsiades C, Akiyama M, Hayashi T, ChauhanD, Richardson P, Schlossman R, Podar K, Munshi NC,Mitsiades N and Anderson KC. (2003). Blood, 101,
1530–1534.Huang Y, Johnson KR, Norris JS and Fan W. (2000). Cancer
Res., 60, 4426–4432.Ikeda K, Oka M, Yamada Y, Soda H, Fukuda M, Kinoshita
A, Tsukamoto K, Noguchi Y, Isomoto H, Takeshima F,Murase K, Kamihira S, Tomonaga M and Kohno S. (1999).Int. J. Cancer, 82, 599–604.
Jentsch S and Schlenker S. (1995). Cell, 82, 881–884.Kanno T, Brown K, Franzoso G and Siebenlist U. (1994).
Mol. Cell. Biol., 14, 6443–6451.Kawakami A, Nakashima T, Sakai H, Urayama S,
Yamasaki S, Hida A, Tsuboi M, Nakamura H, Ida H,Migita K, Kawabe Y and Eguchi K. (1999). Blood, 94,
3847–3854.Kawata S, Ariumi Y and Shimotohno K. (2003). J. Virol., 77,
7291–7299.Kitajima I., Shinohara T, Bilakovics J, Brow DA, Xu X and
Nerenberg M. (1992). Science, 258, 1792–1795.Lau A, Nightingale S, Taylor GP, Gant TW and Cann AJ.
(1998). Blood, 91, 2467–2474.Ling YH, Liebes L, Jiang JD, Holland JF, Elliott PJ, Adams J,
Muggia FM and Perez-Soler R. (2003). Clin. Cancer Res., 9,1145–1154.
Lomas M, Hanon E, Tanaka Y, Bangham CR and Gould KG.(2002). J. Gen. Virol., 83, 641–650.
Ma MH, Yang HH, Parker K, Manyak S, Friedman JM,Altamirano C, Wu ZQ, Borad MJ, Frantzen M, Roussos E,Neeser J, Mikail A, Adams J, Sjak-Shie N, Vescio RAand Berenson JR. (2003). Clin. Cancer Res., 9,
1136–1144.Mahieux R, Pise-Masison CA, Lambert PF, Nicot C, De
Marchis L, Gessain A, Green P, Hall W and Brady JN.(2000). J. Virol., 74, 6866–6874.
Mitra-Kaushik S, Harding JC, Hess J and Ratner L. (2004).Blood, 104, 802–809.
Nasr R, Rosenwald A, El-Sabban ME, Arnulf B, Zalloua P,Lepelletier Y, Bex F, Hermine O, Staudt L, De The H andBazarbachi A. (2003). Blood, 101, 4576–4582.
Obeid LM, Linardic CM, Karolak LA and Hannun YA.(1993). Science, 259, 1769–1771.
Olson NE, Graves JD, Shu GL, Ryan EJ and Clark EA.(2003). J. Immunol., 170, 6065–6072.
Pham LV, Tamayo AT, Yoshimura LC, Lo P and Ford RJ.(2003). J. Immunol., 171, 88–95.
Pise-Masison CA, Mahieux R, Jiang H, Ashcroft M,Radonovich M, Duvall J, Guillerm C and Brady JN.(2000). Mol. Cell. Biol., 20, 3377–3386.
Satou Y, Nosaka K, Koya Y, Yasunaga JI, Toyokuni S andMatsuoka M. (2004). Leukemia, 18, 1357–1363.
Sayers TJ, Brooks AD, Koh CY, Ma W, Seki N, Raziuddin A,Blazar BR, Zhang X, Elliott PJ and Murphy WJ. (2003).Blood, 102, 3–10.
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
429
Oncogene

Sun SC, Elwood J, Beraud C and Greene WC. (1994). Mol.Cell. Biol., 14, 7377–7384.
Sun SC, Harhaj EW, Xiao G and Good L. (2000). Res. Hum.Retroviruses, 16, 1591–1596.
Tamiya S, Etoh K, Suzushima H, Takatsuki K and MatsuokaM. (1998). Blood, 91, 3935–3942.
Tan C and Waldmann TA. (2002). Cancer Res., 62, 1083–1086.
Wattel E, Vartanian JP, Pannetier C and Wain-Hobson S.(1995). J. Virol., 69, 2863–2868.
Yamaoka S, Inoue H, Sakurai M, Sugiyama T, Hazama M,Yamada T and Hatanaka M. (1996). EMBO J., 15, 873–887.
Yoshida M. (2001). Annu. Rev. Immunol., 19, 475–496.Zar JH. (1999). Biostatistical Analysis 4th edn. Prentice Hall:
Englewood Cliffs, NJ.
Effect of PS-341 on HTLV-I transformed and negative T cellsR Nasr et al
430
Oncogene
Top Related
Copyright © 2022 FDOKUMEN

