







Bahasa
Halaman
Hukum


91
Molecular and Cellular Biochemistry 259: 91–99, 2004.© 2004 Kluwer Academic Publishers. Printed in the Netherlands.
Effects of chronic quercetin treatment onantioxidant defence system and oxidative status ofdeoxycorticosterone acetate-salt-hypertensive rats
Milagros Galisteo,1 María Francisca García-Saura,1* Rosario Jiménez,1
Inmaculada Concepción Villar,1 Antonio Zarzuelo,1 Félix Vargas2 andJuan Duarte1
1Department of Pharmacology; 2Department of Physiology, School of Medicine, University of Granada, Granada, Spain
Received 30 April 2003; accepted 10 September 2003
Abstract
We investigated the potential of chronic administration of an oral daily dose (10 mg/kg) of the dietary flavonoid quercetin toprevent hypertension and oxidative stress induced by deoxycorticosterone acetate (DOCA)-salt in rats. We have compared itseffects to those produced by the well-known anti-hypertensive drug verapamil, administered orally (20 mg/kg/day). Quercetinand verapamil treatments reduced systolic blood pressure of DOCA-salt rats in approximately 67.6 and 63.3% respectively,producing no effect in control animals. Both drugs reduced significantly hepatic and renal hypertrophy induced by DOCA-saltadministration, while only quercetin prevented cardiac hypertrophy. Decreased endothelium-dependent relaxation to acetyl-choline of aortic rings from DOCA-salt-treated rats was improved by quercetin, but verapamil only enhanced it in the presenceof superoxide dismutase (SOD) plus catalase. Increased plasma and heart thiobarbituric acid reactive substances (TBARS)and total glutathione (GSH) levels in liver and heart, decreased liver glutathione peroxidase (GPX) and liver and kidney glu-tathione transferase (GST) activities were observed in DOCA-salt-treated rats compared to the control animals. The antihyper-tensive effect of quercetin was accompanied by normalisation of plasma TBARS values, improvement of the antioxidant defencessystem in heart and liver, restoring total GSH levels in both organs and altered liver GST and GPX activities, and improvingkidney GST activity. Verapamil treatment only restored GSH levels in heart, having no effect on other alterations induced byDOCA-salt chronic administration in the antioxidant defences analysed. In conclusion, quercetin shows both antihypertensiveand antioxidant properties in this model of mineralocorticoid hypertension, while verapamil exhibits only antihypertensiveeffects. (Mol Cell Biochem 259: 91–99, 2004)
Key words: flavonoids, verapamil, DOCA-salt hypertension, glutathione, glutathione enzymes, lipid peroxidation
Introduction
Hypertension is a risk factor for several cardiovascular dis-eases such as atherosclerosis or myocardium infarct. Both,production of reactive oxygen species (ROS) by vascularcells, as they contribute to decrease NO availability, promotevascular smooth muscle proliferation and induce neutrophilsinfiltration [1], and alterations of antioxidant enzymes have
been implicated in the pathogenesis of vascular diseases suchas hypertension. Most studies in hypertensive patients reportincreased levels of ROS such as superoxide anion, hydrogenperoxide and lipid peroxides in plasma, together with reducedlevels of natural antioxidant vitamin C and decreased activi-ties of antioxidant enzymes such as superoxide dismutase(SOD) or glutathione peroxidase (GPX) [2–4].
Because human hypertension is heterogeneous, several
Address for offprints: J. Duarte, Department of Farmacología, Fac. Farmacia, Universidad de Granada, 18071 Granada, Spain (E-mail: [email protected])*M. Galisteo and M.F. García-Saura are equal contributors to this work.

92
animal models have been developed to mimic its many fac-ets, and in most of them, i.e. spontaneously hypertensive rats(SHR), Dahl salt-sensitive rats and cyclosporine-inducedhypertension, oxidative stress has been demonstrated [5–7]. Heart, kidney and vessels are organs suffering impor-tant damages in hypertension and exposed to oxidative stress,which has been involved in myocardial and renal hypertro-phy [8] and endothelial dysfunction [1]. Because the liver isa key organ involved in the metabolism and elimination ofendogenous and exogenous substances, and because of themain role of the antioxidant enzymes in the protection of tis-sues against free radicals, it is interesting to know the hepaticmetabolic capacity in hypertension. The long-term adminis-tration of the mineralocorticoid deoxycorticosterone acetate(DOCA) induces sodium retention and, in the presence of ahigh salt intake, it produces a well-known volume-depend-ent type of hypertension in rats [9]. This experimental modelof hypertension is also associated with an increase in vascu-lar superoxide anion generation [10–12] and altered en-dothelium-dependent vascular relaxations [11]. A depressedantioxidant status, involving the glutathione (GSH) system,in the liver of DOCA-salt hypertensive rats has also beendescribed [13] but no data exist concerning the oxidativestatus of organs involved in the control of blood pressure suchas heart and kidney in this model. The GSH system is animportant and potent endogenous antioxidant, critical forscavenging peroxides and other lipid derived oxidants. Fur-thermore, oxidative stress-induced by GSH depletion causessevere hypertension in normal rats [14], suggesting that GSHis a necessary component of the natural antioxidant system.
ROS and blood pressure are intimately related. Thus, in-creased superoxide production has been proposed to be animportant factor contributing to endothelial dysfunction, tis-sue damage and hypertension [15]. Conversely, increasedblood pressure can lead to enhanced ROS production. Super-oxide and superoxide-producing enzymes are enhanced indifferent models of systemic hypertension, regardless of thehypertensive stimulus, and they can be restored by bloodpressure-lowering drugs [16].
The flavonoid quercetin is one of the most abundant poly-phenolic compounds found in the human diet. We have recentlyreported that quercetin reduced elevated blood pressure, en-dothelial dysfunction and cardiac and renal hypertrophy inthe spontaneously hypertensive rat (SHR) model [17, 18] andin hypertension induced by chronic inhibition of nitric ox-ide synthase by L-nitro-arginine-methyl ester [19]. In bothhypertension experimental models, featured by involvementof activation of the renin-angiotensin system in the establish-ment and maintenance of high blood pressure, the protectiveeffects exerted by quercetin were associated with an improve-ment of the oxidative status of the hypertensive animals,attributed to the antioxidant properties of the drug. Moreo-ver, there is no information about the effects of quercetin
on oxidative status in tissues in the mineralocorticoid modelof hypertension, which is associated with low plasma reninactivity [20]. The calcium channel blocker verapamil is awell-known antihypertensive drug, previously described forpreventing blood pressure in this hypertension experimentalmodel [21], but whose effects on GSH-dependent antioxidantsystem in hypertensive models are unknown.
Therefore, in the present study the main aim was to ana-lyse the effects of chronic administration of an oral daily doseof quercetin (10 mg/kg) on blood pressure, endothelial func-tion and oxidative status of DOCA-salt hypertensive rats, andto compare such effects with those of the calcium channelblocker verapamil (20 mg/kg).
Material and methods
Animals and experimental protocol
The experimental protocol followed the European Unionguidelines for animal care and protection. Twelve-week oldmale Wistar rats (220–250 g) were obtained from the Labo-ratory Animal Service of the University of Granada (Spain).All rats were maintained five per cage at a constant tempera-ture (24 ± 1ºC), with a 12-h dark/light cycle and on standardrat chow. An adaptation period of 2 weeks for vehicle admin-istration and blood pressure measurements was allowed be-fore the initiation of the experiment.
Animals were anaesthetised i.p. with 2.5 ml/kg equitensin(500 ml containing 43% w/v chloral hydrate in 81 ml etha-nol; 4.86 mg nembutal; 198 ml propylene glycol; 10.63 gMgSO
4; distilled water) and uninephrectomized. The next
day, they were treated with DOCA (12.5 mg/rat s.c.) or ve-hicle (soybean oil, 0.25 ml/rat). DOCA was administeredweekly at the same dose for 5 weeks. To study the effects ofquercetin on DOCA-salt-induced hypertension, rats were ran-domly assigned to an untreated hypertensive group (vehicle,1 ml of 1% methylcellulose) or a quercetin group (10 mg/kg,mixed in 1 ml of 1% methylcellulose). Another group of ani-mals received verapamil (20 mg/kg in 1 ml water) as a controldrug. Rats were treated orally by gavage for 5 weeks. Duringthe experimental period, DOCA-treated rats were allowed freeaccess to water containing 1% NaCl and chow. The quercetinor verapamil treatments were stopped 2 days before the endof the experiment in order to study their long-term effectswithout the involvement of acute administration effects.
Blood pressure measurements
Systolic blood pressure (SBP) was measured at the beginningof the experiment and just before killing, 18–20 h after admin-istration of quercetin or vehicle in conscious, pre-warmed,

93
restrained rats by tail-cuff plethysmography. At least 7 deter-minations were made in every session and the mean of thelowest three values within 5 mmHg was taken as the SBP level.
Samples collection and storage
At the end of the experimental period, animals were anaes-thetised i.p. with 2.5 ml/kg equitensin and blood was collectedin heparinized tubes from the abdominal aorta. Animals weresacrificed and organs were rapidly removed, rinsed in ice-coldphysiological saline solution, blotted, weighed, aliquoted andfrozen. The hepatic, cardiac and renal weight indices werecalculated by dividing the liver, heart and kidney weight bythe body weight. Hepatic weight ratio was expressed as %.Plasma was obtained by blood centrifugation at 2000 g for15 min, aliquoted and frozen. All biological samples werestored at –80ºC until analysis.
Ex vivo vascular relaxation to acetylcholine in thepresence or absence of SOD
Three-mm ring segments of the descending thoracic aortawere dissected and mounted in individual organ chambersfilled with Krebs buffer of the following composition (mM):NaCl 118, KCl 4.75, NaHCO
3 25, MgSO
4 1.2, CaCl
2 2,
KH2PO
4 1.2, glucose 11. The solution was continuously
gassed with a 95% O2 and 5% CO
2 mixture, and maintained
at 37ºC. Rings were stretched to 2 g of resting tension bymean of two L-shaped stainless-steel wires, which were in-serted into the lumen of the ring and attached to the chamberand to an isometric force-displacement transducer (Letigraph,model 2000, Letica S.A., Barcelona, Spain), as previously de-scribed [22]. Rings were equilibrated for 60–90 min, andduring this period tissues were re-stretched and washed every30 min with warm Krebs’ solution. Relaxant responses to ace-tylcholine (1 µM) were tested in rings pre-contracted by no-radrenaline (1 µM), and incubated in the presence or absenceof the antioxidant mixture of SOD (100 U/ml) plus catalase(100 U/ml), and were expressed as a percentage of pre-con-traction induced by noradrenaline.
Determination of total glutathione (GSH) content inorgans
Organ samples were deproteinized by homogenisation with 5%trichloroacetic acid. Homogenates were centrifuged at 4000 gfor 10 min and an aliquot of the supernatant was taken for themeasurement of GSH using the 5,5′-dithio-bis(2-nitrobenzoicacid) recycling assay described by Anderson [23]. Data wereexpressed as nmol GSH/g weight of tissue.
GSH-related enzymes activities in organs
Organ samples were homogenised with 5 vols of potassiumphosphate 0.1 M, ethylendiamine tetraacetic acid (EDTA)1 mM, pH 7.2, and 0.1% digitonine. They were centrifugedat 3000 g for 10 min and supernatants were used for enzymeactivity determinations. Protein concentration was deter-mined according to Bradford [24], using bovine serum albu-min as standard. Glutathione peroxidase (GPX) activity wasdetermined according to the method described by Flohé andGünzler [25], following the oxidation of NADPH at 340 nm.Liver homogenate samples were incubated at 37ºC for 10 minin the presence of 10 mM GSH, 1.5 mM NADPH, 40 mMNaN
3, glutathione reductase (0.24 U), and reaction started by
addition of 1.5 mM H2O
2 in sodium phosphate 0.1 M, pH 7.2.
For kidney and heart homogenates, incubation was monitoredin the same conditions but reaction started by addition of t-butyl hydroperoxide (12 mM) instead of H
2O
2. One unit of
GPX was defined as the amount of enzyme that catalyses theoxidation of 1 µmol of NADPH per min.
Glutathione reductase (GR) activity was assessed accord-ing to the method of Carlberg and Mannervik [26], follow-ing NADPH oxidation 340 nm, in a reaction which takesplace at 30ºC. Homogenate samples were incubated in thepresence of 20 mM GSSG and 2 mM NADPH, in potassiumphosphate buffer 0.2 M pH 7.2, containing 2 mM EDTA. Oneunit of GR was defined as the amount of enzyme that cataly-ses the oxidation of 1 µmol of NADPH per min.
Glutathione-S-transferase (GST) activity was determinedfollowing the method of Habig et al. [27]. This method isbased on the capacity of this enzyme to metabolise 1-chloro-2,4-dinitrobenzene in the presence of GSH at 25ºC. One unitof GST activity was defined as the amount of enzyme cata-lysing the formation of 1 µmol of product per min.
Measurement of thiobarbituric acid reactive substances(TBARS) in organs
Levels of TBARS in organs were determined as an indexof lipid peroxidation, following the method described byZingarelli et al. [28]. Organ samples were homogenised in1.15% KCl solution. An aliquot of the homogenate wasadded to a reaction mixture containing 200 µl of 8.1% so-dium dodecyl-sulphate, 1500 µl 20% acetic acid, pH 3.5,1500 µl 0.8% thiobarbituric acid and 700 µl distilled water.Samples were then boiled for 1 h at 95ºC and centrifuged at3000 g for 10 min. Supernatant absorbance was measured byspectrophotometry at 532 nm and compared with a stand-ard curve prepared from different concentrations of 1,1,3,3-tetramethoxypropane. Data were expressed as µmol/100 mgweight of tissue.

94
Measurement of thiobarbituric acid reactive substances(TBARS) in plasma
Plasma levels of TBARS were assessed following the methoddescribed by Esterbauer and Cheesseman [29]. An aliquot of300 µl of the sample was added to a reaction mixture con-taining 750 µl of an acetonitrile solution of 10.3 mM 1-me-thyl-phenylindole and 225 µl HCl 35%. Samples were warmedat 45ºC for 40 min, and centrifuged at 3000 g for 10 min. Theabsorbance of the supernatants was measured at 586 nm andcompared with a standard curve of 1,1,3,3-tetramethoxy-propane. Data were expressed as concentration of TBARS(µM).
Chemicals
All reagents were of analytical grade. DOCA was obtainedfrom ICN Biochemicals Inc. (Ohio, USA). The other drugs,including quercetin dihydrate and (±)-verapamil hydrochlo-ride, and chemicals were purchased from Sigma (Alcobendas,Madrid, Spain).
Statistics
Results are expressed as means ± S.E.M. of measurements.Statistically significant differences were analysed by anANOVA, followed by a post hoc Bonferroni test using theSigmastat software program. p < 0.05 was considered statis-tically significant.
Results
Blood pressure and body and organs weights
Table 1 shows SBP values before and after 5 weeks of treat-ment with quercetin or verapamil in control and DOCA-saltrats. Long-term quercetin or verapamil administration pre-vented the increase in SBP induced by DOCA, by approxi-mately 67.6 and 63.3% respectively. These drugs did notproduce any change in blood pressure values of the controlrats.
Values of body weight and hepatic, cardiac and renal in-dices are shown in Table 2. At the end of the experimentalperiod, animals of the DOCA-salt group showed significantlyincreased hepatic (~ 14%), renal (~ 123%) and cardiac (~ 42%)weight indices, as compared with the animals of the controlgroup. In the rats receiving DOCA-salt plus quercetin orverapamil, the hepatic and renal weight indices were signifi-cantly reduced as compared with the DOCA-salt group. How-ever, only quercetin reduced significantly cardiac hypertrophy.
In vitro vascular relaxation to acetylcholine in thepresence or absence of SOD
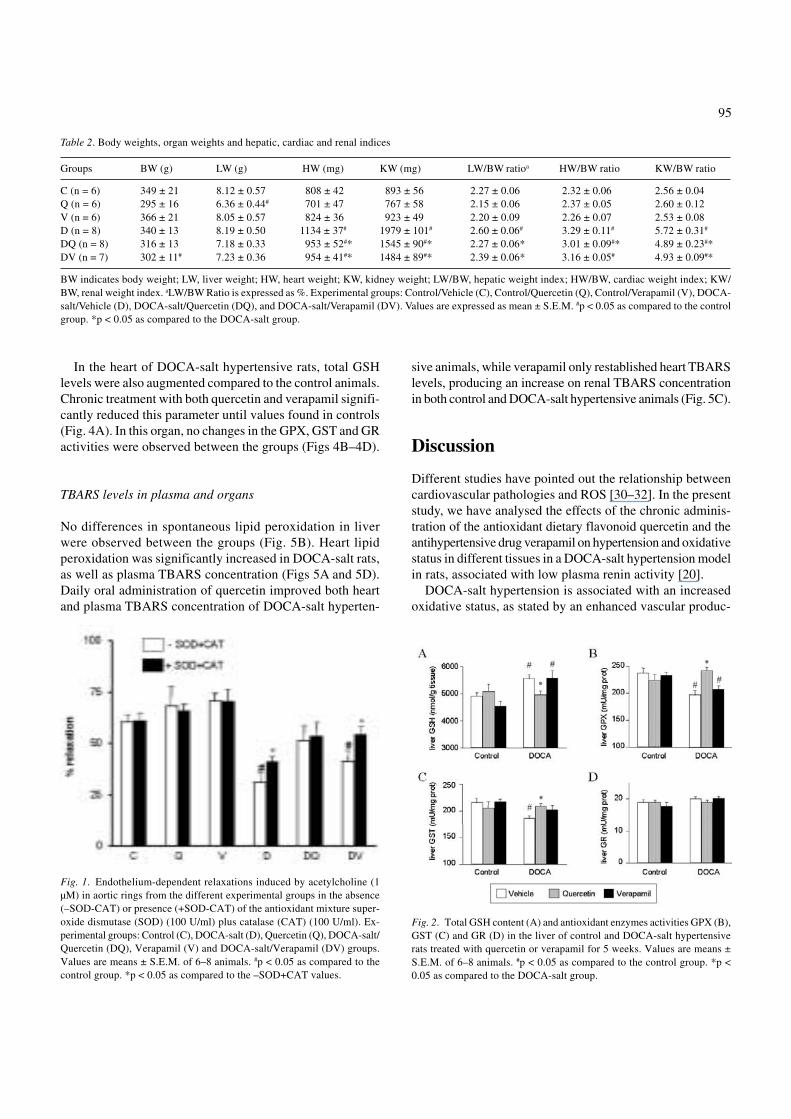
Endothelium-dependent relaxation to acetylcholine (1 µM)was reduced in aortic rings obtained from DOCA-salt hyper-tensive rats compared to those from the control group (p <0.05) (Fig. 1). Chronic treatment with quercetin, but not withverapamil, increased vasodilator response to acetylcholine inaortic rings from DOCA-salt rats. In the presence of the anti-oxidant mixture SOD (100 U/ml) plus catalase (100 U/ml),endothelium-dependent response to acetylcholine was sig-nificantly enhanced in aorta from DOCA-salt and DOCA-saltverapamil treated animals (p < 0.05) compared to rings in-cubated in the absence of this antioxidant mixture.
Total GSH content and GSH-related enzyme activities indifferent organs
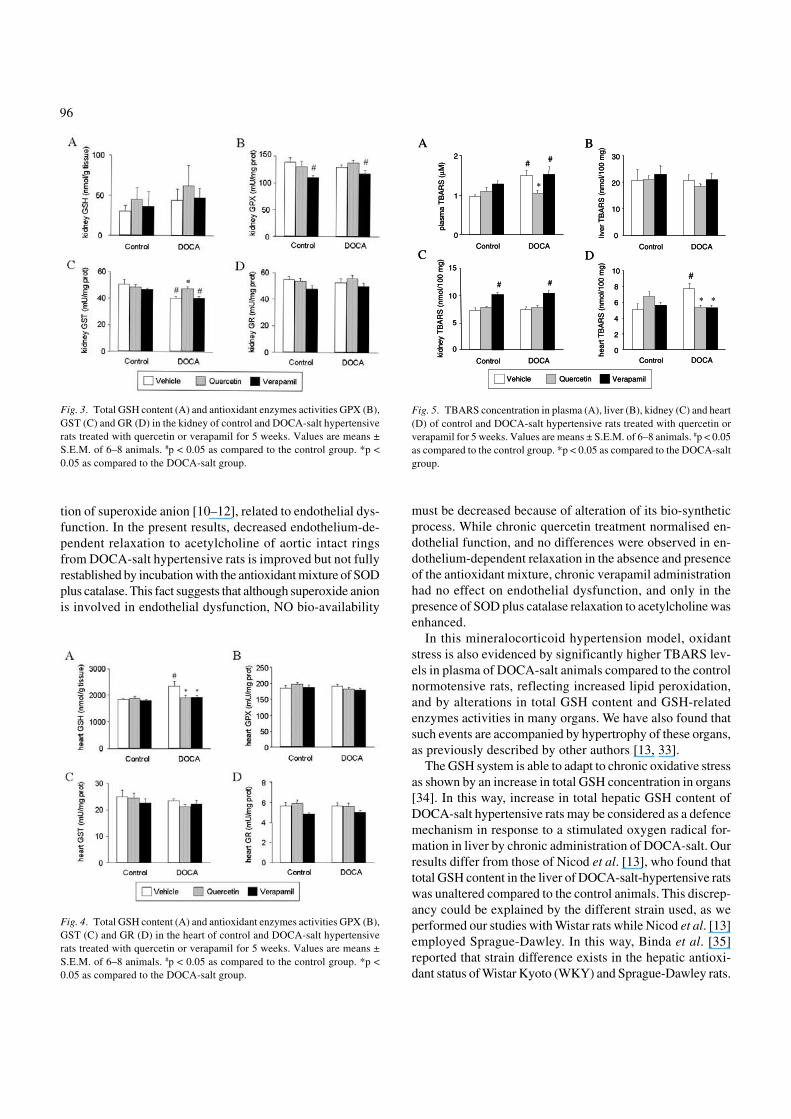
Liver total GSH content was significantly higher in DOCA-salt rats as compared to the control animals. Chronic treatmentwith quercetin, but not verapamil, resulted in a significantreduction of this parameter in the liver of DOCA-treated rats(Fig. 2A), reaching similar levels of GSH to those found inthe control group. Liver GPX and GST activities in DOCA-salt hypertensive rats were significantly reduced as comparedto the animals of the control group. Chronic treatment withquercetin augmented significantly both GPX and GST activi-ties in DOCA-salt liver, while verapamil only increased liverGST activity. No one of these drugs had any significant ef-fect on the control rats (Figs 2B and 2C). No differences werefound in the liver activity of GR between control normoten-sive and DOCA-salt hypertensive animals. Furthermore, GRactivity was not modified by the treatments with quercetinor verapamil (Fig. 2D).
No differences between the experimental groups were ob-served concerning kidney total GSH levels (Fig. 3A). Kid-ney GST activity was significantly diminished in DOCA-saltrats (Fig. 3C) and chronic treatment with quercetin improvedit, while verapamil treatment produced no changes.
Table 1. Effect of long-term quercetin or verapamil treatment on SBP (mmHg)
Group Baseline After 2 weeks After 5 weeks
C (n = 6) 139.3 ± 3.2 143.7 ± 2.4 145.3 ± 0.7Q (n = 6) 147.2 ± 2.5 143.2 ± 2.3 151.7 ± 2.3V (n = 6) 149.5 ± 3.9 141.8 ± 3.8 149.5 ± 2.1D (n = 8) 141.8 ± 1.6 167.0 ± 4.0# 188.9 ± 4.3#
DQ (n = 8) 147.5 ± 2.1 146.8 ± 2.1* 159.1 ± 4.4#*DV (n = 7) 143.3 ± 3.4 151.7 ± 1.9#* 161.0 ± 1.3#*
Experimental groups: Control/Vehicle (C), Control/Quercetin (Q), Control/Verapamil (V), DOCA-salt/Vehicle (D), DOCA-salt/Quercetin (DQ), andDOCA-salt/Verapamil (DV). Values are expressed as means ± S.E.M. #p <0.05 as compared to the control group. *p < 0.05 as compared to the DOCA-salt group.
95
In the heart of DOCA-salt hypertensive rats, total GSHlevels were also augmented compared to the control animals.Chronic treatment with both quercetin and verapamil signifi-cantly reduced this parameter until values found in controls(Fig. 4A). In this organ, no changes in the GPX, GST and GRactivities were observed between the groups (Figs 4B–4D).
TBARS levels in plasma and organs
No differences in spontaneous lipid peroxidation in liverwere observed between the groups (Fig. 5B). Heart lipidperoxidation was significantly increased in DOCA-salt rats,as well as plasma TBARS concentration (Figs 5A and 5D).Daily oral administration of quercetin improved both heartand plasma TBARS concentration of DOCA-salt hyperten-
sive animals, while verapamil only restablished heart TBARSlevels, producing an increase on renal TBARS concentrationin both control and DOCA-salt hypertensive animals (Fig. 5C).
Discussion
Different studies have pointed out the relationship betweencardiovascular pathologies and ROS [30–32]. In the presentstudy, we have analysed the effects of the chronic adminis-tration of the antioxidant dietary flavonoid quercetin and theantihypertensive drug verapamil on hypertension and oxidativestatus in different tissues in a DOCA-salt hypertension modelin rats, associated with low plasma renin activity [20].
DOCA-salt hypertension is associated with an increasedoxidative status, as stated by an enhanced vascular produc-
Table 2. Body weights, organ weights and hepatic, cardiac and renal indices
Groups BW (g) LW (g) HW (mg) KW (mg) LW/BW ratioa HW/BW ratio KW/BW ratio
C (n = 6) 349 ± 21 8.12 ± 0.57 808 ± 42 893 ± 56 2.27 ± 0.06 2.32 ± 0.06 2.56 ± 0.04Q (n = 6) 295 ± 16 6.36 ± 0.44# 701 ± 47 767 ± 58 2.15 ± 0.06 2.37 ± 0.05 2.60 ± 0.12V (n = 6) 366 ± 21 8.05 ± 0.57 824 ± 36 923 ± 49 2.20 ± 0.09 2.26 ± 0.07 2.53 ± 0.08D (n = 8) 340 ± 13 8.19 ± 0.50 1134 ± 37# 1979 ± 101# 2.60 ± 0.06# 3.29 ± 0.11# 5.72 ± 0.31#
DQ (n = 8) 316 ± 13 7.18 ± 0.33 953 ± 52#* 1545 ± 90#* 2.27 ± 0.06* 3.01 ± 0.09#* 4.89 ± 0.23#*DV (n = 7) 302 ± 11# 7.23 ± 0.36 954 ± 41#* 1484 ± 89#* 2.39 ± 0.06* 3.16 ± 0.05# 4.93 ± 0.09#*
BW indicates body weight; LW, liver weight; HW, heart weight; KW, kidney weight; LW/BW, hepatic weight index; HW/BW, cardiac weight index; KW/BW, renal weight index. aLW/BW Ratio is expressed as %. Experimental groups: Control/Vehicle (C), Control/Quercetin (Q), Control/Verapamil (V), DOCA-salt/Vehicle (D), DOCA-salt/Quercetin (DQ), and DOCA-salt/Verapamil (DV). Values are expressed as mean ± S.E.M. #p < 0.05 as compared to the controlgroup. *p < 0.05 as compared to the DOCA-salt group.
Fig. 1. Endothelium-dependent relaxations induced by acetylcholine (1µM) in aortic rings from the different experimental groups in the absence(–SOD-CAT) or presence (+SOD-CAT) of the antioxidant mixture super-oxide dismutase (SOD) (100 U/ml) plus catalase (CAT) (100 U/ml). Ex-perimental groups: Control (C), DOCA-salt (D), Quercetin (Q), DOCA-salt/Quercetin (DQ), Verapamil (V) and DOCA-salt/Verapamil (DV) groups.Values are means ± S.E.M. of 6–8 animals. #p < 0.05 as compared to thecontrol group. *p < 0.05 as compared to the –SOD+CAT values.
Fig. 2. Total GSH content (A) and antioxidant enzymes activities GPX (B),GST (C) and GR (D) in the liver of control and DOCA-salt hypertensiverats treated with quercetin or verapamil for 5 weeks. Values are means ±S.E.M. of 6–8 animals. #p < 0.05 as compared to the control group. *p <0.05 as compared to the DOCA-salt group.
96
tion of superoxide anion [10–12], related to endothelial dys-function. In the present results, decreased endothelium-de-pendent relaxation to acetylcholine of aortic intact ringsfrom DOCA-salt hypertensive rats is improved but not fullyrestablished by incubation with the antioxidant mixture of SODplus catalase. This fact suggests that although superoxide anionis involved in endothelial dysfunction, NO bio-availability
must be decreased because of alteration of its bio-syntheticprocess. While chronic quercetin treatment normalised en-dothelial function, and no differences were observed in en-dothelium-dependent relaxation in the absence and presenceof the antioxidant mixture, chronic verapamil administrationhad no effect on endothelial dysfunction, and only in thepresence of SOD plus catalase relaxation to acetylcholine wasenhanced.
In this mineralocorticoid hypertension model, oxidantstress is also evidenced by significantly higher TBARS lev-els in plasma of DOCA-salt animals compared to the controlnormotensive rats, reflecting increased lipid peroxidation,and by alterations in total GSH content and GSH-relatedenzymes activities in many organs. We have also found thatsuch events are accompanied by hypertrophy of these organs,as previously described by other authors [13, 33].
The GSH system is able to adapt to chronic oxidative stressas shown by an increase in total GSH concentration in organs[34]. In this way, increase in total hepatic GSH content ofDOCA-salt hypertensive rats may be considered as a defencemechanism in response to a stimulated oxygen radical for-mation in liver by chronic administration of DOCA-salt. Ourresults differ from those of Nicod et al. [13], who found thattotal GSH content in the liver of DOCA-salt-hypertensive ratswas unaltered compared to the control animals. This discrep-ancy could be explained by the different strain used, as weperformed our studies with Wistar rats while Nicod et al. [13]employed Sprague-Dawley. In this way, Binda et al. [35]reported that strain difference exists in the hepatic antioxi-dant status of Wistar Kyoto (WKY) and Sprague-Dawley rats.
Fig. 3. Total GSH content (A) and antioxidant enzymes activities GPX (B),GST (C) and GR (D) in the kidney of control and DOCA-salt hypertensiverats treated with quercetin or verapamil for 5 weeks. Values are means ±S.E.M. of 6–8 animals. #p < 0.05 as compared to the control group. *p <0.05 as compared to the DOCA-salt group.
Fig. 4. Total GSH content (A) and antioxidant enzymes activities GPX (B),GST (C) and GR (D) in the heart of control and DOCA-salt hypertensiverats treated with quercetin or verapamil for 5 weeks. Values are means ±S.E.M. of 6–8 animals. #p < 0.05 as compared to the control group. *p <0.05 as compared to the DOCA-salt group.
Fig. 5. TBARS concentration in plasma (A), liver (B), kidney (C) and heart(D) of control and DOCA-salt hypertensive rats treated with quercetin orverapamil for 5 weeks. Values are means ± S.E.M. of 6–8 animals. #p < 0.05as compared to the control group. *p < 0.05 as compared to the DOCA-saltgroup.
plas
ma
TB
AR
S (
µM)
Control DOCA
0
1
2# #
*
A
plas
ma
TB
AR
S (
µM)
Control DOCA
0
1
2# #
*
A
0
10
20
30
Control DOCA
liver
TB
AR
S (
nmol
/100
mg)
B
0
10
20
30
Control DOCA
liver
TB
AR
S (
nmol
/100
mg)
B
kidn
eyT
BA
RS
(nm
ol/1
00 m
g)
10
15
0
Control DOCA
5
# #
C
kidn
eyT
BA
RS
(nm
ol/1
00 m
g)
10
15
0
Control DOCA
5
# #
C
hear
tTB
AR
S (
nmol
/100
mg)
Control DOCA0
2
4
6
8
10#
* *
D
hear
tTB
AR
S (
nmol
/100
mg)
Control DOCA0
2
4
6
8
10#
* *
D
QuercetinQuercetin VerapamilVerapamilVehicleVehicle

97
Quercetin chronic treatment reduced liver total GSH contentof DOCA-salt rats, and this suggests that oxygen radical for-mation should be reduced in the liver of these animals. In thepresent study, we have also found diminished liver GPX andGST activities in DOCA-salt rats, as well as unaltered GRactivity, as previously described by Nicod et al. [13, 36].Chronic treatment with quercetin prevented the decrease inboth activities, while verapamil had no effect on them.
A consistent finding concerning GSH cycle alterations inhypertension is a decreased hepatic GPX activity, describedin other hypertension experimental models [18, 37, 38]. A re-duced GPX activity leads to increased cellular accumulationof ROS and lipid peroxidation. In fact, plasma accumulationof TBARS, a reliable marker of lipid peroxidation, was alsoincreased in DOCA-salt hypertensive rats, while chronictreatment with quercetin contributes to reduce this parameter,effect that should be attributed to its antioxidant properties.GST comprises a family of enzymes involved in the detoxi-fication processes of reactive metabolites and in limitingoxidative damage to tissues. This enzyme has been shown tobe critical in protecting cells against ROS due to redox cy-cling of exogenous and endogenous quinones [39].
Kidney damage in DOCA-salt hypertension is character-ised by hypertrophy and renal failure. We found that DOCA-salt induced severe renal hypertrophy, and quercetin andverapamil treatment of DOCA-salt animals prevented eleva-tion in renal weight index, indicating that both drugs protect,at least partially, from renal injury. There are no studies aboutthe renal oxidant status in mineralocorticoid hypertension. Inother models, i.e. stroke-prone-SHR, it has been described asignificant decrease of renal GPX compared to the normo-tensive WKY rats [16]. We found no difference in renal GPXactivity between untreated control and DOCA-salt animals,which correlated with similar kidney TBARS levels. How-ever, verapamil treatment significantly reduced renal GPXactivity in both control and DOCA-salt animals increasingthe kidney lipid peroxidation. Further investigations areneeded to clarify mechanisms involved in these deleteriouseffects induced by verapamil. Our results showed that chronicadministration of DOCA-salt significantly diminished renalGST activity in rats, suggesting a diminished rate in the renaldetoxification reactions, leaving kidneys more exposed to ox-idative injury. Quercetin, but not verapamil, treatment im-proved this renal enzymatic activity in hypertensive animals.
Increased oxidative stress in heart of DOCA-salt hyperten-sive rats was also evidenced in our study by significantlyhigher TBARS concentration and increased total GSH con-tent, compared to the control normotensive rats. In SHR, hearttotal GSH content has been found to be higher than in theirWKY counterparts, and this was correlated with an increasein the gamma-glutamylcysteine synthetase mRNA [40], en-zyme involved in the GSH synthesis. Both, quercetin orverapamil treatments prevented the increase of both TBARS
and total GSH content in heart from DOCA-salt animals.These results are in agreement to that found by Perna et al.[41] which showed that chronic verapamil inhibits the fattyacid oxidation of myocardium.
The flavonoid quercetin exerts antioxidant properties invitro, by inhibiting xanthine oxidase activity [42] and scav-enging radical species such as superoxide anion, hydroxylradical, and peroxynitrite [43]. When administered orally invivo, quercetin is bio-transformed and its metabolites retainits antioxidant properties [44]. This flavonol has been previ-ously described for improving hepatic antioxidant defencesin SHR and nitric oxide-deficient hypertensive rats [18, 19],and for reducing renal injury by preventing oxidative stressdependent on ischemia- reperfusion in kidneys [45].
Verapamil has been previously described for reducingblood pressure and ameliorating vascular lesions when ad-ministered chronically to rats with mineralocorticoid hyper-tension [21]. In the present study, its chronic administrationreduced significantly SBP in DOCA-salt treated rats. Any-way, despite antioxidant properties attributed to verapamil onin vitro studies [46], it failed to improve antioxidant de-fences in liver and even increased oxidative stress in kid-ney of DOCA-salt hypertensive animals. Furthermore, ourresults are in agreement with a recent clinical study, whichshows that calcium channel blockers have no antioxidantproperties in vivo in hypertensive patients [47].
In conclusion, our data demonstrate that chronic adminis-tration of quercetin reduces elevated blood pressure, andcontributes to enhance liver, kidney and heart antioxidantdefence system as well as to improve endothelial function inDOCA-salt hypertensive rats. Such effects were almost notobserved in the case of a chronic treatment with verapamil,which prevented the DOCA-salt-induced increase in bloodpressure in a similar extent to quercetin. These results sug-gest that increase in the oxidative status may be independentof the blood pressure control in this model of mineralocorti-coid hypertension. Anyway, antioxidant properties of quer-cetin contributed to endothelial dysfunction improvement,which is an essential step in the management of hypertension,effect lacking in the case of the calcium antagonist verapamil.
Acknowledgements
This work was supported by CICYT (SAF 98-0160, SAF-2001-2953) Grants. M. Galisteo was a recipient of a researchcontract from the University of Granada (Spain).
References
1. McIntyre M, Bohr DF, Dominiczak AF: Endothelial function in hyper-tension: The role of superoxide anion. Hypertension 34: 539–545, 1999

98
2. Lacy F, O’Connor DT, Schmid-Schonbein GW: Plasma hydrogen per-oxide production in hypertensives and normotensives subjects at ge-netic risk of hypertension. J Hypertens 16: 291–303, 1998
3. Russo C, Oliviero O, Girelli D, Faccini G, Zenari ML, Lombardi S,Corrucher R: Antioxidant status and lipid peroxidation in patients withessential hypertension. J Hypertens 16: 1267–1271, 1998
4. Redon J, Oliva MR, Tormos C, Giner V, Chaves J, Iradi A, Saez GT:Antioxidant activities and oxidative stress byproducts in human hyper-tension. Hypertension 41: 1096–1101, 2003
5. Lopez-Ongil S, Hernandez-Perera O, Navarro-Antolin J, Perez deLema G, Rodriguez-Puyol M, Lamas S, Rodriguez- Puyol D: Role ofreactive oxygen species in the signalling cascade of cyclosporine A-mediated up-regulation of eNOS in vascular endothelial cells. Br JPharmacol 1224: 447–454, 1998
6. Schnackenberg CG, Welch WJ, Wilcox CS: Normalisation of bloodpressure and renal vasculature resistance in SHR with a membrane-permeable superoxide dismutase mimetic: Role of nitric oxide. Hyper-tension 32: 59–64, 1998
7. Atarashi K, Ishiyama A, Takagi M, Minami M, Kimura K, Goto A,Omata M: Vitamin E ameliorates the renal injury of Dahl salt-sensi-tive rats. Am J Hypertens 10: 116S–119S, 1997
8. Sawyer DB, Siwik DA, Xiao L, Pimentel DR, Singh K, Colucci WS:Role of oxidative stress in myocardial hypertrophy and failure. J MolCell Cardiol 34: 379–388, 2002
9. Hollenberg NK, Coletti C, Passan D: Hypertension, volume and va-soconstriction: studies on the renal blood supply in SHR, WKY andDOCA-salt rat models. Hypertension Res 15: 3–11, 1992
10. Wu R, Millette E, Lingyun W, de Champlain J: Enhanced superoxideanion formation in vascular tissues from spontaneously hypertensiveand deoxycorticosterone acetate-salt hypertensive rats. J Hypertens 19:741–748, 2001
11. Somers MJ, Mavromatis K, Galis ZS, Harrison, DG: Vascular super-oxide production and vasomotor function in hypertension induced bydeoxycorticosterone acetate-salt. Circulation 101: 1722–1728, 2000
12. Beswick RA, Zhang H, Marable D, Catravas JD, Hill WD, Webb RC:Long-term antioxidant administration attenuates mineralocorticoidhypertension and renal inflammatory response. Hypertension 37: 781–786, 2001
13. Nicod L, Rodriguez S, Letang JM, Viollon-Abadie C, Jacqueson A,Berthelot A, Richert L: Antioxidant status, lipid peroxidation, mixedfunction oxidase and UDP-glucuronyl transferase activities in liversfrom control and DOCA-salt hypertensive male Sprague Dawley rats.Mol Cell Biochem 203: 33–39, 2000
14. Vaziri ND, Wanng XQ, Oveisi F, Rad B: Induction of oxidative stressby glutathione depletion causes severe hypertension in normal rats. Hy-pertension 36: 142–146, 2000
15. Hamilton CA, Brosnan MJ, McIntyre M, Graham D, Dominiczak AF:Superoxide excess in hypertension and aging: A common cause ofendothelial dysfunction. Hypertension 37: 529-534, 2001
16. Shou I, Wang LN, Suzuki S, Fukui M, Tomino Y: Effects of antihy-pertensive drugs on antioxidant enzyme activities and renal functionin stroke-prone spontaneously hypertensive rats. Am J Med Sci 314:377–384, 1997
17. Duarte J, Pérez-Palencia R, Vargas F, Ocete MA, Pérez-Vizcaino F,Zarzuelo A, Tamargo J: Antihypertensive effects of the flavonoid quer-cetin in spontaneously hypertensive rats. Brit J Pharmacol 133: 117–124, 2001
18. Duarte J, Galisteo M, Ocete MA, Pérez-Vizcaíno F, Zarzuelo A, TamargoJ: Effects of chronic quercetin treatment on hepatic oxidative status ofspontaneously hypertensive rats. Mol Cell Biochem 221: 155–160,2001
19. Duarte J, Jiménez R, O’Valle F, Galisteo M, Pérez-Palencia R, VargasF, Pérez-Vizcaíno F, Zarzuelo A, Tamargo J: Protective effects of the
flavonoid quercetin in chronic nitric oxide deficient rats. J Hypertens20: 1843–1854, 2002
20. Gavras H, Brunner HR, Larga JH, Vaughan ED Jr, Koss M, Cote LJGavras I: Malignant hypertension resulting from deoxycorticosteroneacetate and salt excess: Role of renin and sodium in vascular changes.Circ Res 36: 300–309, 1975
21. Aguas AP, Nickerson PA: Effect of verapamil on blood pressure andlesions in heart and kidney of rats made hypertensive by deoxycorti-costerone (DOC). Am J Pathol 110: 48–54, 1983
22. Duarte J, Pérez-Vizcaíno F, Zarzuelo A, Jiménez J, Tamargo J: Vasodi-lator effects of quercetin in isolated rat vascular smooth muscle. Eur JPharmacol 239: 1–7, 1993
23. Anderson ME: Determination of glutathione and glutathione disulfidein biological samples. Meth Enzymol 113: 548–555, 1985
24. Bradford MM: A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72: 248–254, 1976
25. Flohé L, Günzler WA: Assays of glutathione peroxidase. Meth Enzymol105: 114–121, 1984
26. Carlberg I, Mannervik B: Glutathione reductase. Meth Enzymol 113:484–490, 1985
27. Habig WH, Pabst MJ, Jakoby WB: Glutathione S-transferases. The firstenzymatic step in mercapturic acid formation. J Biol Chem 249: 7130–7139, 1974
28. Zingarelli B, Szabo C, Salzman AL: Blockade of polly(ADP-ribose)-synthetase inhibits neutrophil recruitment, oxidant generation andmucosal injury in murine colitis. Gastroenterology 116: 335–345, 1999
29. Esterbauer H, Cheeseman KH: Determination of aldehydic lipid per-oxidation product: Malonaldehyde and 4-hydroxynonenal. MethEnzymol 186: 407–421, 1990
30. Griendling KK, Alexander RW: Oxidative stress and cardiovasculardisease. Circulation 96: 3264–3265, 1997
31. Givertz MM, Colucci WS: New targets for heart-failure therapy:Endothelin, inflammatory cytokines, and oxidative stress. Lancet 52:SI34–SI38, 1998
32. Nakazono K, Watanabe N, Matsuno K, Sasaki J, Sato T, Inoue M: Doessuperoxide underlie the pathogenesis of hypertension? Proc Natl AcadSci USA 88: 10045–10048, 1991
33. Hoida G, Lindt S, Eichenberger E: The course of blood pressure andorgan changes in rats with DOCA-hypertension. Arznim Forsch 26:2003–2008, 1976
34. Toth KM, Beehler CJ, Parry SS, Halek MM, Berger EM, Repine JE:Erythrocyte antioxidants as scavengers and/or indicators of oxygenmetabolites in biological systems. In: G. Rotilio (ed). Superoxide andSuperoxide Dismutase in Chemistry, Biology and Medicine. Elsevier,Amsterdam, 1986, pp 391–394
35. Binda D, Nicod L, Viollon-Abadie C, Rodriguez S, Berthelot A,Coassolo P, Richert L: Strain difference (WKY, SPRD) in the hepaticantioxidant status in rat and effect of hypertension (SHR, DOCA). Exvivo and in vitro data. Mol Cell Biochem 218: 139–146, 2001
36. Nicod L, Rodriguez S, Jacqueson A, Viollon-Abadie C, Berthelot A,Richert L: Clofibric acid or diethylmaleate supplemented diet decreaseblood pressure in DOCA-salt treated male Sprague Dawley rats. Re-lation with liver antioxidant status. Mol Cell Biochem 213: 65–73, 2000
37. Yuan YV, Kitts DD, Godin DV: Variations in dietary fat and choles-terol intakes modify antioxidant status of SHR and WKY rats. J Nutr128: 1620–1630, 1998
38. Ma L, Johnson P: Antihypertensive drug therapy and antioxidant en-zyme mRNA levels in spontaneously hypertensive (SHR) rats. CompBiochem Physiol B Biochem Mol Biol 122: 119–126, 1999
39. Rushmore TH, Pickett CB: Glutathione-S-transferases, structure, regu-lation, and therapeutic implications. J Biol Chem 268: 11475–11478,1993

99
40. Carlos DM, Goto S, Urata Y, Iida T, Cho S, Niwa M, Tsuji Y, KondoT: Nicardipine normalizes elevated levels of antioxidant activity inresponse to xanthine oxidase-induced oxidative stress in hypertensiverat heart. Free Radic Res 29: 143–150, 1998
41. Perna AF, Smogorzewski M, Massry SG: Effects of verapamil on theabnormalities in fatty acid oxidation of myocardium. Kidney Int 36:453–457, 1989
42. Selloum L, Reichl S, Müller M, Sebihi L, Arnhold J: Effects of flavonolson the generation of superoxide anion radicals by xanthine oxidase andstimulated neutrophils. Arch Biochem Biophys 395: 49–56, 2001
43. Hanasaki Y, Ogawa S, Fukui S: The correlation between active oxy-gen scavenging and antioxidative effects of flavonoids. Free RadicalBiol Med 16: 845–850, 1994
44. Manach C, Morand C, Crespy V, Demigné C, Texier O, Régérat F,
Rémésy C: Quercetin is recovered in human plasma as conjugatedderivatives which retain antioxidant properties. FEBS Lett 426: 331–336, 1998
45. Inal M, Altinisik M, Bilgin MD: The effect of quercetin on renal is-chemia and reperfusion injury in the rat. Cell Biochem Funct 20: 291–296, 2002
46. Mason RP, Mak IT, Trumbore MW, Mason PE: Antioxidant propertiesof calcium antagonists related to membrane biophysical interactions.Am J Cardiol 84: 16L–22L, 1999
47. Baykal Y, Yilmaz MI, Celik T, Gok F, Rehber H, Akay C, Kocar IH:Effects of antihypertensive agents, alpha receptor blockers, betablockers, angiotensin-converting enzyme inhibitors, angiotensin re-ceptor blockers and calcium channel blockers, on oxidative stress. JHypertens 21: 1207–1211, 2003

100
Top Related
Copyright © 2022 FDOKUMEN

