







Bahasa
Halaman
Hukum


This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Deep-Sea Research II 54 (2007) 1864–1875
Diversity and distribution of Porifera in the bathyal and abyssalWeddell Sea and adjacent areas
Dorte Janussena,�, Ole Secher Tendalb
aForschungsinstitut und Naturmuseum Senckenberg, Senckenberganlage 25, D-60325 Frankfurt/Main, GermanybSNM, Zoological Museum, University of Copenhagen, Universitetsparken 15, DK-2100 Copenhagen O, Denmark
Accepted 6 July 2007
Available online 6 August 2007
Abstract
During the ANDEEP I–III expeditions, we obtained a rich and highly diverse sponge collection from the deep Weddell
Sea. All the three Poriferan classes, Calcarea, Demospongiae and Hexactinellida, were well represented. Among this
material, we have identified a total of 76 species from 47 genera and 30 families. Of these, 17 species (22%) are new to
science and 37 (49%) new for the Southern Ocean. Particularly remarkable is the considerable depth of the boundary
between bathyal and abyssal sponge faunas. Both Demospongiae and Hexactinellida show a strong shift in their
taxonomic composition from a typical shelf assemblage to a more cosmopolitan deep-sea fauna at around 2500m. Within
the Demospongiae, the families Polymastiidae and Cladorhizidae (carnivorous sponges) are particularly abundant and
very diverse. The Calcarea are recorded for the first time from the Antarctic deep sea. The type of sampling gear used,
especially the epibenthic sledge, was an important factor for the successful collection of deep-sea sponges during the
ANDEEP campaigns.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Sponges; Antarctic; Deep-sea; Biodiversity; ANDEEP
1. Introduction
Sponges are ubiquitous marine organisms and areoften very conspicuous members of macro- andmegafauna communities, especially on hard andsemihard substrates (Dayton et al., 1974; Van Soest,1994; Hooper and Van Soest, 2002; Klitgaard andTendal, 2004). When moving from the shelf downthe slope into the deep sea, there are certainconsistent trends in the composition, distribution
and biology of the sponge faunas: (1) Overall, thenumber of species decreases, although there arenarrow depth zones just beyond the shelf break andon the lower slope where species richness isenhanced (Vinogradova, 1962; Tabachnik, 1994;Maldonado and Young, 1996; Tendal, unpub-lished). (2) The relative proportions of the classeschanges; species of Hexactinellida become morenumerous and species of Calcarea are rare (Levi,1964). (3) Certain morphological features andbiological traits become more common, notablythe development of ‘‘roots’’, stalks and a life stylethat includes carnivory (Burton, 1928; Levi, 1964;Vacelet and Duport, 2004; Vacelet, 2006). (4) Thegeneral size, biomass and abundance of sponges
ARTICLE IN PRESS
www.elsevier.com/locate/dsr2
0967-0645/$ - see front matter r 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.dsr2.2007.07.012
�Corresponding author. Tel.: +49 69 7542 306;
fax: +4969 746238.
E-mail address: [email protected]
(D. Janussen).

Author's personal copy
decrease when going from the near-shore, eutrophicregions through oceanic eutrophic regions to theoligotrophic deep sea (Barthel and Tendal, 1993;Tabachnik, 1994; Sokolova, 2000; Ilan et al., 2003).
The Weddell, Scotia and Lazarev seas constitutethe Atlantic sector of the circumantarctic SouthernOcean, the sea extending from the Antarctic coastsnorthward to 601S (International HydrographicOrganization 2000). Because this limit approxi-mately coincides with the Antarctic Polar Front(the Antarctic Convergence), it has hydrographicand biological significance in the uppermost parts ofthe water column, which is dominated by Antarctic
Surface Water. At depths between 600m and theseafloor, however, the hydrographic conditions arequite different, being dominated by Antarctic
Bottom Water (Ostapoff, 1965; Pickard and Emery,1982). The Weddell and Lazarev seas are genuineparts of the Southern Ocean, while the Scotia Sea isdivided by the 601S line of latitude into a smallerSouthern Ocean part and a larger South Atlanticpart. The Weddell Sea covers approximately2.8� 106 km2 and the Scotia Sea approximately1� 106 km2, while the Lazarev Sea, which is ratherpoorly defined, has a surface area of about0.6� 106 km2. Over most of their area, the Weddelland Scotia seas are 4000–5000m deep, althoughsome minor ridges define a number of basins. Thecontinental shelf is generally narrow, the shelf edgeis located at 500–800m depth, and the slopes aremostly rather steep. The Lazarev Sea reaches thesame depths, but has more prominent ridges andgentler slopes.
Our present knowledge of the sponge fauna of thebathyal and abyssal parts of the Antarctic seasoriginates from a number of older Australian,British, French, German, Japanese, Scottish andRussian expeditions, some more recent Germancruises, and our investigation of the materialcollected during the ANDEEP I–III cruises in2002 and 2005. Here, we give an overview of thecomposition of the sponge fauna of the deepWeddell, Scotia and Lazarev seas and its geogra-phical and bathymetrical distribution patterns, andcomment on its possible biogeographical relation-ships and origin.
2. History of exploration of the Antarctic deep-sea
sponge fauna
The isostatically depressed Antarctic shelf extendsto greater depths than that of other continents. The
depth of the shelf break varies somewhat from placeto place, but is usually located between 500 and900m. The shelf fauna, including the sponges,follows the pattern of the bathymetry, probablybecause of the lack of hydrographical structure inthe water masses. The numbers of species decreasessubstantially below 500–700m, with a minor eur-ybathic element extending to considerably greaterdepths (Malyutina, 2004; McClintock et al., 2005;Arnaud and Hain, 1992; Ushakov, 1963).
Before the ANDEEP expeditions of 2002 and2005, little was known about the Porifera of thedeep Weddell Sea. Most previous expeditions tookvery few, if any, samples deeper than the shelfbreak. A survey is given in Table 1. We choose the800m depth contour as a convenient arbitrary limitbetween the shelf fauna and that of the slope. Thesamples were taken with a trawl or similar kind ofqualitative gear. In total, about 30 catches werereported to contain sponges. They are scattered overthe immense area from the Weddell Sea along EastAntarctica to the Ross Sea. Half of them are fromdepths below 2000m and the rest are equallydivided between the areas around the shelf breakand the slope.
The Scottish National Antarctic Expedition(1902–1904) was the first expedition to investigatethe great depths of the Weddell Sea. Along a ratherdirect track from Coats Land to the South OrkneyIslands they trawled 10 times at depths between2234 and 4795m (Wilton and Brown, 1908). Sixtrawls were successful, and four of these containedsponges. A total of 12 species (10 hexactinellids and2 demosponges) were recovered, all of them foundonly once (Topsent 1913, 1915a). The richest stationwas at 2580m (7 species), while the deepest stationat 4795m yielded only one species.
Almost 100 years later, one haul by the Agassiztrawl containing 18 species of sponges (6 hexacti-nellids and 12 demosponges) was taken at 2012mdepth in the inner part of the Weddell Sea duringthe 1989 ANTARKTIS VII/4 (EPOS, leg 3) cruiseon the German R.V. Polarstern. Only one of thespecies was also obtained during the ScottishNational Antarctic Expedition (Barthel and Tendal,1989, 1992).
3. Material and methods
Most of the ANDEEP sponges were collectedwith an Agassiz trawl (AGT) or an epibenthic sledge(EBS); a few were taken using a box corer and
ARTICLE IN PRESSD. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1865
Author's personal copy
multicorer (Janussen, 2003, 2006). The spongesfrom the trawls were sorted, and photographicallydocumented, immediately after their collection. TheEBS samples were preserved immediately in 96%ethanol, whereas the AGT samples were fixed in6% formalin and/or 96% ethanol. Subsamples ofsome specimens were frozen for later molecularinvestigations. In the laboratory, skeletal prepara-tions were made of all specimens, and some wereadditionally investigated by histological methodsand by SEM.
The collection is housed in the SenckenbergMuseum (Frankfurt am Main). After publicationof the ANDEEP sponges, this collection will beincorporated into the SESAM database and acces-sible on the Internet.
4. Collection gear and effects on the sampling success
The number of AGT and EBS deployments atANDEEP stations that yielded sponges are shown
in Table 2. The construction of the AGT had anobvious influence on the sampling success. DuringANDEEP I and II, the AGT used was 1.5m wide,and the net had a 10-mm mesh size. DuringANDEEP III, a 3-m-wide AGT was used. Atstations PS67/074-7, PS67/078-11 and PS67/081-9,the cod end mesh size was 10mm while at all otherstations an inner bag of 500-mm mesh size wasinserted to trap smaller sized animals. The trawlingcable length was 1.5 times the water depth, and thegear was hauled on the bottom at 1 knot for 10min.At station PS67/059-10, the AGT was hauled for20min (Linse et al., 2006). The effect on thecollection success becomes obvious when comparingthe results of ANDEEP II and III (Table 2),especially the stations in the Weddell Sea Basinthat were sampled during both expeditions. How-ever, the ANDEEP I operations in the Scotia Seaare difficult to compare with those of ANDEEP IIand III because of the very different geographicand oceanographic situation and the fact that no
ARTICLE IN PRESS
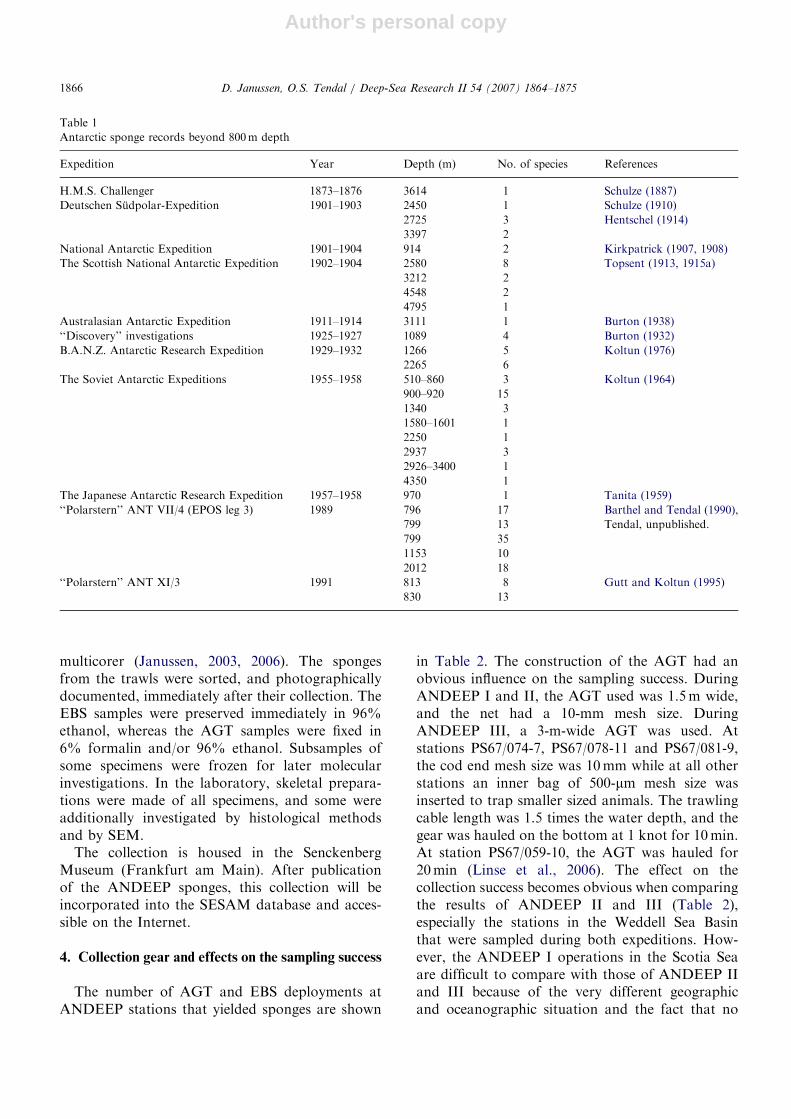
Table 1
Antarctic sponge records beyond 800m depth
Expedition Year Depth (m) No. of species References
H.M.S. Challenger 1873–1876 3614 1 Schulze (1887)
Deutschen Sudpolar-Expedition 1901–1903 2450 1 Schulze (1910)
2725 3 Hentschel (1914)
3397 2
National Antarctic Expedition 1901–1904 914 2 Kirkpatrick (1907, 1908)
The Scottish National Antarctic Expedition 1902–1904 2580 8 Topsent (1913, 1915a)
3212 2
4548 2
4795 1
Australasian Antarctic Expedition 1911–1914 3111 1 Burton (1938)
‘‘Discovery’’ investigations 1925–1927 1089 4 Burton (1932)
B.A.N.Z. Antarctic Research Expedition 1929–1932 1266 5 Koltun (1976)
2265 6
The Soviet Antarctic Expeditions 1955–1958 510–860 3 Koltun (1964)
900–920 15
1340 3
1580–1601 1
2250 1
2937 3
2926–3400 1
4350 1
The Japanese Antarctic Research Expedition 1957–1958 970 1 Tanita (1959)
‘‘Polarstern’’ ANT VII/4 (EPOS leg 3) 1989 796 17 Barthel and Tendal (1990),
Tendal, unpublished.799 13
799 35
1153 10
2012 18
‘‘Polarstern’’ ANT XI/3 1991 813 8 Gutt and Koltun (1995)
830 13
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–18751866

Author's personal copy
sponge specialist took part. The EBS has provenunsurpassed for the collection of very small speci-mens (e.g., Cladorhizidae, Fig. 1).
5. ANDEEP I, II and III results
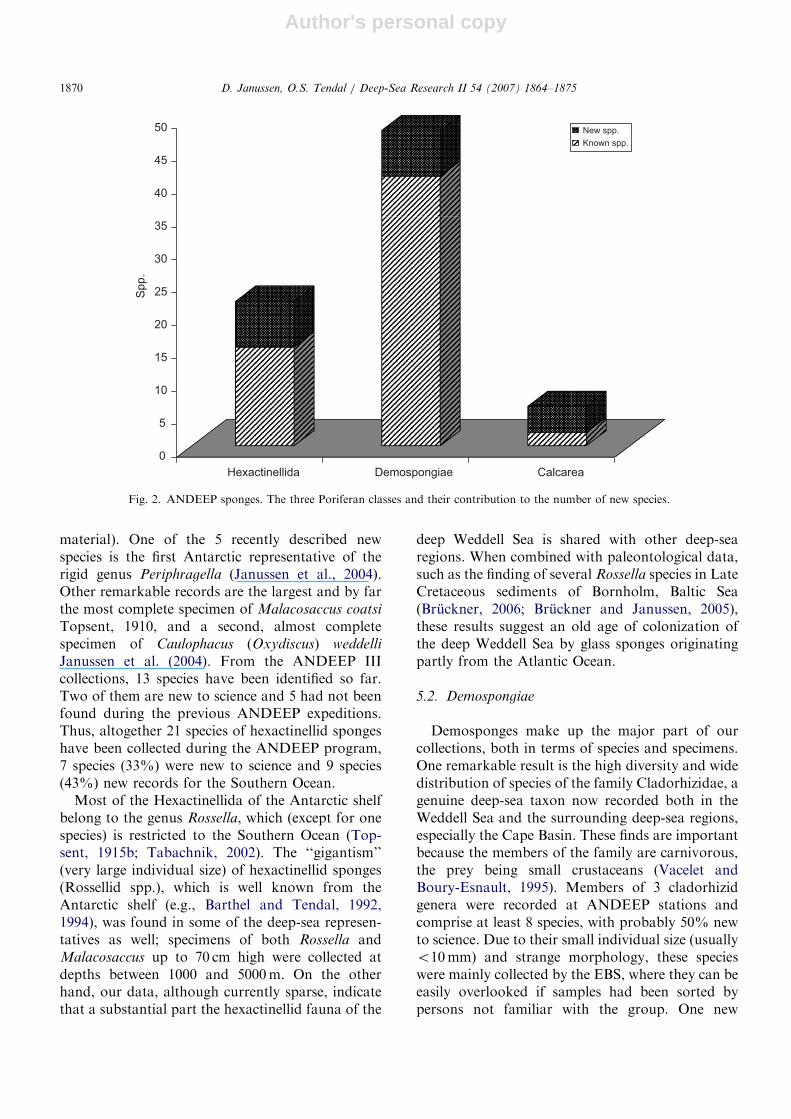
Little was known about the Poriferan fauna ofthe deep Weddell Sea prior to the ANDEEPexpeditions (Barthel and Tendal, 1989, 1992).Nevertheless, the high diversity of bathyal andabyssal sponges revealed by the new samplingprogram was surprising, both at the species andthe higher taxon level. A very preliminary list of theso-far-identified taxa collected during the ANDEEPprogram is given in Table 3. The first indicationscame in 2002 (ANDEEP I and II) when 49 spongespecies were found, including 30 species of Demos-pongiae (at least 2 new to science), 4 of Calcarea(3 new to science), and 16 of Hexactinellida (5 newto science) (Janussen et al., 2004). In 2005(ANDEEP III), these earlier results were extendedby the collection of a similarly rich Poriferan fauna,comprising 51 species. The fact that 32 (63%) ofthem were recorded for the first time during theANDEEP program demonstrates the large faunisticvariation between stations, as well as the potentialof these deep-sea expeditions for revealing novelbiodiversity.
The material from the ANDEEP expeditionscontributes very significantly to our knowledge ofPoriferan taxonomy in Antarctic waters and thedeep sea in general. In total, we have so faridentified 75 species from 50 genera and 24 familiesdistributed among the classes Calcarea, Demospon-giae and Hexactinellida (Table 3, Fig. 2). Ofthese, 17 species (22%) were new to science and 37(49%) new for the Southern Ocean. These figureswill certainly increase when the taxonomic work
on this large and complex sponge collection iscompleted.
There are great differences in this collectionbetween families in terms of abundance anddiversity. Some families, such as the Rossellidaeand Polymastiidae, include 5–10 species, whereasseveral families of the Calcarea are each representedby only 1 species. The same is true of the totalnumbers of individuals in different families. ThePolymastiidae and Suberitidae are very abundant,with 50–100 individuals found at one station. TheRossellidae, Mycalidae, Tedanidae and Myxillidae
ARTICLE IN PRESS
Table 2
Sponge catches by epibenthic sledge (EBS) and Agassiz trawl (AGT) during the ANDEEP expeditions
Expedition Collection gear Depths trawled during
expedition (m)
No. of hauls,
altogether
Hauls catching
sponges
In % of all
hauls
ANDEEP I (23 January
2002–3 February 02)
EBS 1799–5195 9 2 22
ANDEEP I AGT (1.5m) 2250–5100 9 0 0
ANDEEP II (28 February
2002–1 April 2002)
EBS 755–4980 13 4 31
ANDEEP II AGT (1.5m) 752–4995 12 10 83
ANDEEP III (21 January
2005–6 April 2005)
EBS 1048–4927 19 11 58
ANDEEP III AGT (3m) 1055–4931 19 16 84
Fig. 1. Small, mature specimen of Asbestopluma n. sp. with eggs
(visible in the central part of the sponge); from EBS station #16,
ANDEEP III (2005).
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1867
Author's personal copy
ARTICLE IN PRESS
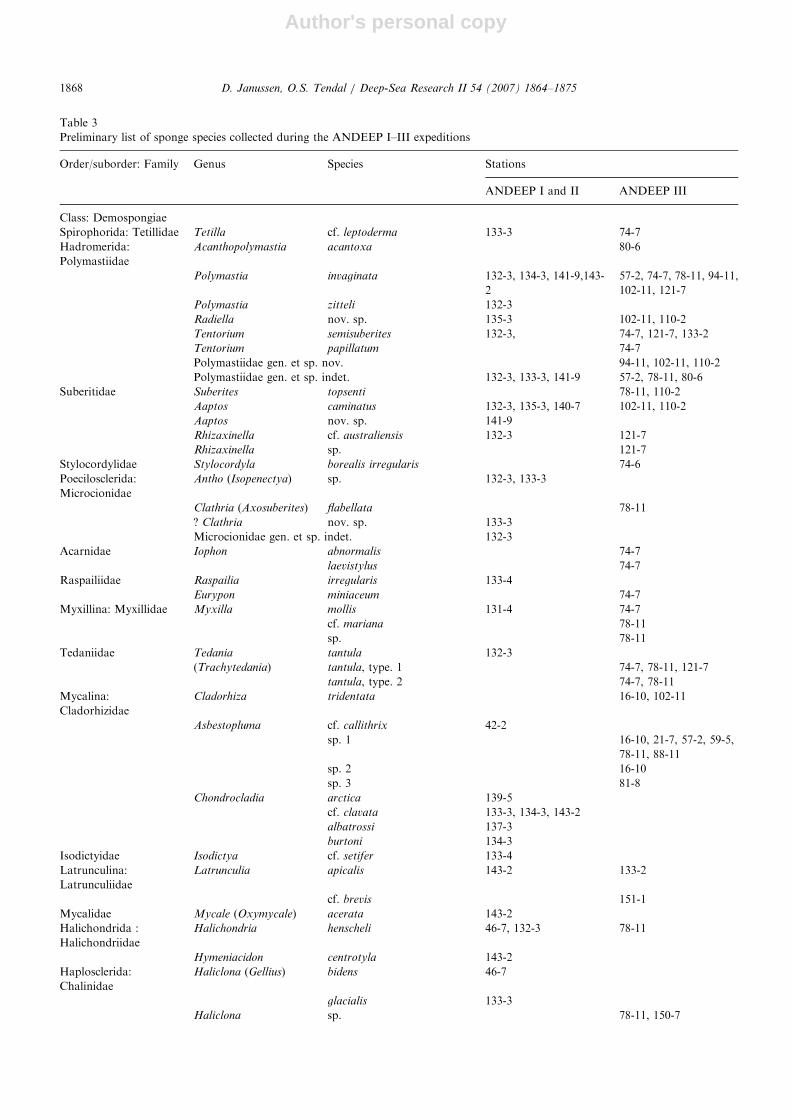
Table 3
Preliminary list of sponge species collected during the ANDEEP I–III expeditions
Order/suborder: Family Genus Species Stations
ANDEEP I and II ANDEEP III
Class: Demospongiae
Spirophorida: Tetillidae Tetilla cf. leptoderma 133-3 74-7
Hadromerida:
Polymastiidae
Acanthopolymastia acantoxa 80-6
Polymastia invaginata 132-3, 134-3, 141-9,143-
2
57-2, 74-7, 78-11, 94-11,
102-11, 121-7
Polymastia zitteli 132-3
Radiella nov. sp. 135-3 102-11, 110-2
Tentorium semisuberites 132-3, 74-7, 121-7, 133-2
Tentorium papillatum 74-7
Polymastiidae gen. et sp. nov. 94-11, 102-11, 110-2
Polymastiidae gen. et sp. indet. 132-3, 133-3, 141-9 57-2, 78-11, 80-6
Suberitidae Suberites topsenti 78-11, 110-2
Aaptos caminatus 132-3, 135-3, 140-7 102-11, 110-2
Aaptos nov. sp. 141-9
Rhizaxinella cf. australiensis 132-3 121-7
Rhizaxinella sp. 121-7
Stylocordylidae Stylocordyla borealis irregularis 74-6
Poecilosclerida:
Microcionidae
Antho (Isopenectya) sp. 132-3, 133-3
Clathria (Axosuberites) flabellata 78-11
? Clathria nov. sp. 133-3
Microcionidae gen. et sp. indet. 132-3
Acarnidae Iophon abnormalis 74-7
laevistylus 74-7
Raspailiidae Raspailia irregularis 133-4
Eurypon miniaceum 74-7
Myxillina: Myxillidae Myxilla mollis 131-4 74-7
cf. mariana 78-11
sp. 78-11
Tedaniidae Tedania
(Trachytedania)
tantula 132-3
tantula, type. 1 74-7, 78-11, 121-7
tantula, type. 2 74-7, 78-11
Mycalina:
Cladorhizidae
Cladorhiza tridentata 16-10, 102-11
Asbestopluma cf. callithrix 42-2
sp. 1 16-10, 21-7, 57-2, 59-5,
78-11, 88-11
sp. 2 16-10
sp. 3 81-8
Chondrocladia arctica 139-5
cf. clavata 133-3, 134-3, 143-2
albatrossi 137-3
burtoni 134-3
Isodictyidae Isodictya cf. setifer 133-4
Latrunculina:
Latrunculiidae
Latrunculia apicalis 143-2 133-2
cf. brevis 151-1
Mycalidae Mycale (Oxymycale) acerata 143-2
Halichondrida :
Halichondriidae
Halichondria henscheli 46-7, 132-3 78-11
Hymeniacidon centrotyla 143-2
Haplosclerida:
Chalinidae
Haliclona (Gellius) bidens 46-7
glacialis 133-3
Haliclona sp. 78-11, 150-7
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–18751868
Author's personal copy
are of intermediate abundance, being represented by10–20 individuals at some stations. Most deep-seasponge species, however, were rare, with 1–5individuals (if any) occurring at one station. TheCalcarea were particularly rare, all genera andthe four families being caught only once duringthe ANDEEP I–III expeditions.
5.1. Hexactinellida
The Hexactinellida plays a major role on theAntarctic shelf, where species occur in densepopulations, reach large body sizes and probably
high individual ages. The general taxonomy of theclass is very incomplete, and probably not even halfof the species have been recorded and described.Furthermore, many hexactinellid taxa are knownonly from one, sometimes fragmentary, specimen.Twenty hexactinellid species have been reportedfrom the deep Weddell Sea, 5 of which are alsoknown from other oceans.
The ANDEEP II Hexactinellida includes 14species belonging to 12 genera, of which 5 speciesand 1 subgenus are new to science (including 1 newspecies from another Weddell Sea expeditionstudied and described along with the ANDEEP
ARTICLE IN PRESS
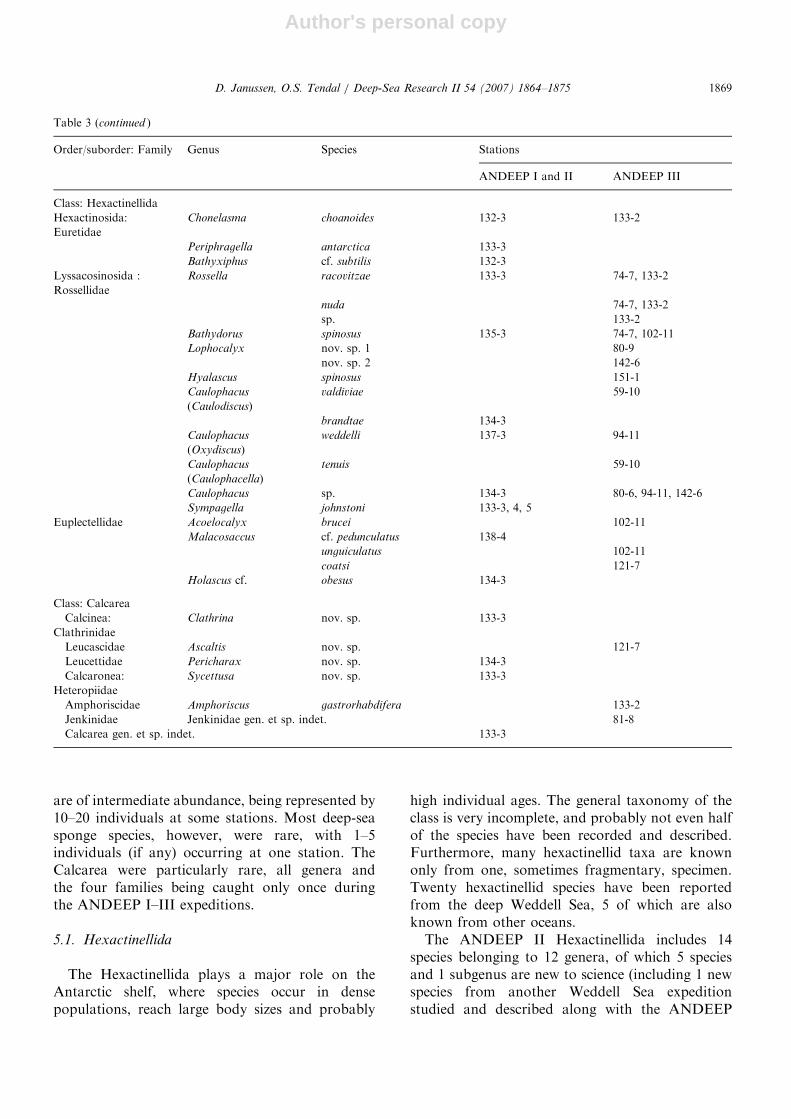
Table 3 (continued )
Order/suborder: Family Genus Species Stations
ANDEEP I and II ANDEEP III
Class: Hexactinellida
Hexactinosida:
Euretidae
Chonelasma choanoides 132-3 133-2
Periphragella antarctica 133-3
Bathyxiphus cf. subtilis 132-3
Lyssacosinosida :
Rossellidae
Rossella racovitzae 133-3 74-7, 133-2
nuda 74-7, 133-2
sp. 133-2
Bathydorus spinosus 135-3 74-7, 102-11
Lophocalyx nov. sp. 1 80-9
nov. sp. 2 142-6
Hyalascus spinosus 151-1
Caulophacus
(Caulodiscus)
valdiviae 59-10
brandtae 134-3
Caulophacus
(Oxydiscus)
weddelli 137-3 94-11
Caulophacus
(Caulophacella)
tenuis 59-10
Caulophacus sp. 134-3 80-6, 94-11, 142-6
Sympagella johnstoni 133-3, 4, 5
Euplectellidae Acoelocalyx brucei 102-11
Malacosaccus cf. pedunculatus 138-4
unguiculatus 102-11
coatsi 121-7
Holascus cf. obesus 134-3
Class: Calcarea
Calcinea:
Clathrinidae
Clathrina nov. sp. 133-3
Leucascidae Ascaltis nov. sp. 121-7
Leucettidae Pericharax nov. sp. 134-3
Calcaronea:
Heteropiidae
Sycettusa nov. sp. 133-3
Amphoriscidae Amphoriscus gastrorhabdifera 133-2
Jenkinidae Jenkinidae gen. et sp. indet. 81-8
Calcarea gen. et sp. indet. 133-3
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1869
Author's personal copy
material). One of the 5 recently described newspecies is the first Antarctic representative of therigid genus Periphragella (Janussen et al., 2004).Other remarkable records are the largest and by farthe most complete specimen of Malacosaccus coatsi
Topsent, 1910, and a second, almost completespecimen of Caulophacus (Oxydiscus) weddelli
Janussen et al. (2004). From the ANDEEP IIIcollections, 13 species have been identified so far.Two of them are new to science and 5 had not beenfound during the previous ANDEEP expeditions.Thus, altogether 21 species of hexactinellid spongeshave been collected during the ANDEEP program,7 species (33%) were new to science and 9 species(43%) new records for the Southern Ocean.
Most of the Hexactinellida of the Antarctic shelfbelong to the genus Rossella, which (except for onespecies) is restricted to the Southern Ocean (Top-sent, 1915b; Tabachnik, 2002). The ‘‘gigantism’’(very large individual size) of hexactinellid sponges(Rossellid spp.), which is well known from theAntarctic shelf (e.g., Barthel and Tendal, 1992,1994), was found in some of the deep-sea represen-tatives as well; specimens of both Rossella andMalacosaccus up to 70 cm high were collected atdepths between 1000 and 5000m. On the otherhand, our data, although currently sparse, indicatethat a substantial part the hexactinellid fauna of the
deep Weddell Sea is shared with other deep-searegions. When combined with paleontological data,such as the finding of several Rossella species in LateCretaceous sediments of Bornholm, Baltic Sea(Bruckner, 2006; Bruckner and Janussen, 2005),these results suggest an old age of colonization ofthe deep Weddell Sea by glass sponges originatingpartly from the Atlantic Ocean.
5.2. Demospongiae
Demosponges make up the major part of ourcollections, both in terms of species and specimens.One remarkable result is the high diversity and widedistribution of species of the family Cladorhizidae, agenuine deep-sea taxon now recorded both in theWeddell Sea and the surrounding deep-sea regions,especially the Cape Basin. These finds are importantbecause the members of the family are carnivorous,the prey being small crustaceans (Vacelet andBoury-Esnault, 1995). Members of 3 cladorhizidgenera were recorded at ANDEEP stations andcomprise at least 8 species, with probably 50% newto science. Due to their small individual size (usuallyo10mm) and strange morphology, these specieswere mainly collected by the EBS, where they can beeasily overlooked if samples had been sorted bypersons not familiar with the group. One new
ARTICLE IN PRESS
0
5
10
15
20
25
30
35
40
45
50S
pp.
CalcareaDemospongiaeHexactinellida
New spp.
Known spp.
Fig. 2. ANDEEP sponges. The three Poriferan classes and their contribution to the number of new species.
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–18751870

Author's personal copy
species of Asbestopluma was found to containmature eggs (Fig. 1). The ANDEEP material ofCladorhizidae substantially improves our knowl-edge of the distribution of this important deep-seasponge family in the Southern Ocean (Fig. 3).
The Polymastiidae is another common family ofdemosponges predominantly living in deep water.They are by far the most abundant sponges at somestations in the abyssal Weddell Sea. Samples fromANDEEP III stations #102 and #110 each providedmore than 100 specimens, both stations beinglocated at depths of about 4700m. Polymastia
invaginata Kirkpatrick, 1907, is the sponge speciesthat shows by far the most eurybathic distributionwithin the ANDEEP fauna. It was taken at manystations between 800 and 4900m and has beenreported earlier from the Antarctic as shallow as18m (Koltun, 1976). However, it is not common atthe abyssal stations. Here, it is replaced by a veryabundant species, which will probably be rede-scribed as a new species of Aaptos Gray, 1867, and asecond, less abundant new genus and species of thePolymastiidae (Plotkin et al., submitted). Amongother polymastiids, a new species of Radiella
represents the first record of this genus in theSouthern Ocean, while Tentorium is a eurybathicgenus common at the bathyal stations. It seems thatone species of the latter genus, Tentorium semisu-
berites (Schmidt, 1870), has a bipolar distribution.Geographically, many of the other demosponge
species collected are restricted to the SouthernOcean, where they mostly occur on the shelf. It isremarkable that during the ANDEEP expeditions,they were recorded at much greater depths. Theyseem to represent a eurybathic fauna elementextending down from the Antarctic shelf and slopeto depths of about 2500m. Thus, the slope (bathyal)demosponge fauna in the Weddell Sea is character-ized by a mixture of shallow-water and deep-seaspecies.
5.3. Calcarea
While the group is well known from the Antarcticshelf, it was a real surprise when Calcarea wererecorded for the first time at depths between 1120and 4400m during the ANDEEP II and IIIexpeditions (Janussen et al., 2003; Janussen, 2006).During ANDEEP II, 3 species new to science werefound, 2 at 1120m and 1 at 4065m depth. DuringANDEEP III, another 3 species, 1 of which is new,were collected at depths of about 1500, 2600 and
4380m. Thus, in total, 7 species have now beenfound. The 4 new species belong to the generaAscaltis, Clathrina, Pericharax and Sycettusa (Rappet al., in preparation). Amphoriscus gastrorhabdi-
fera, first described from shallow waters off Tristanda Cunha (Burton, 1932), is reported for the secondtime.
6. Implications and general considerations
(1) So far, 75 sponge species have been identifiedfrom the rich deep-sea samples from theANDEEP expeditions. Of these, 17 species(22%) are new to science and 37 (49%) new forthe Southern Ocean.
(2) The comparison of samples taken in the samelocal area shows the importance of sampling
ARTICLE IN PRESS
Fig. 3. Distribution of new and previous records of Cladorhizi-
dae in the Southern Ocean. Small squares represent one species,
the bigger square two species collected at the same station.
Records in the Weddell Sea and surrounding areas are from the
ANDEEP I–III expeditions; all other records are from literature
(Hentschel, 1914; Koltun, 1964; Ridley and Dendy, 1887;
Topsent, 1901, 1913). Two other ANDEEP III records outside
the Antarctic, one species in the Agulhas Basin and three spp. in
the Cape Basin, are not shown on this map.
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1871
Author's personal copy
with different kinds of gear in order to collectboth the small, often fragile but sometimesrather common species, and at the larger,robust but often less common species.
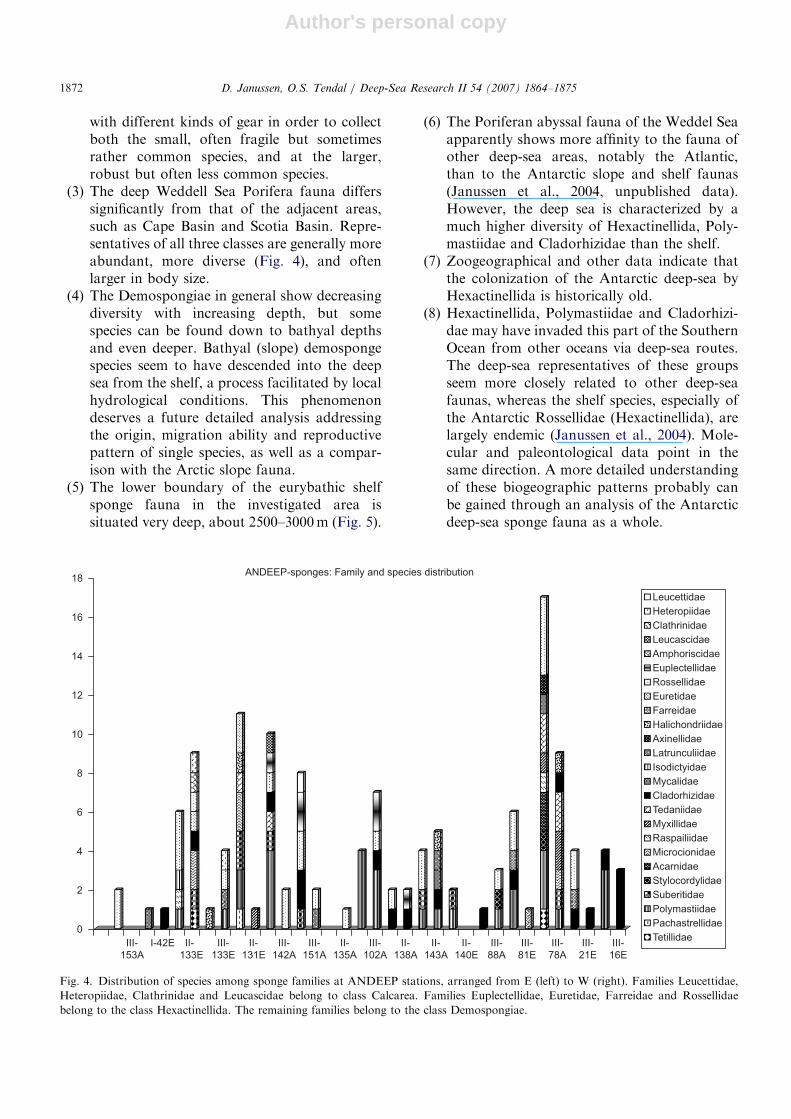
(3) The deep Weddell Sea Porifera fauna differssignificantly from that of the adjacent areas,such as Cape Basin and Scotia Basin. Repre-sentatives of all three classes are generally moreabundant, more diverse (Fig. 4), and oftenlarger in body size.
(4) The Demospongiae in general show decreasingdiversity with increasing depth, but somespecies can be found down to bathyal depthsand even deeper. Bathyal (slope) demospongespecies seem to have descended into the deepsea from the shelf, a process facilitated by localhydrological conditions. This phenomenondeserves a future detailed analysis addressingthe origin, migration ability and reproductivepattern of single species, as well as a compar-ison with the Arctic slope fauna.
(5) The lower boundary of the eurybathic shelfsponge fauna in the investigated area issituated very deep, about 2500–3000m (Fig. 5).
(6) The Poriferan abyssal fauna of the Weddel Seaapparently shows more affinity to the fauna ofother deep-sea areas, notably the Atlantic,than to the Antarctic slope and shelf faunas(Janussen et al., 2004, unpublished data).However, the deep sea is characterized by amuch higher diversity of Hexactinellida, Poly-mastiidae and Cladorhizidae than the shelf.
(7) Zoogeographical and other data indicate thatthe colonization of the Antarctic deep-sea byHexactinellida is historically old.
(8) Hexactinellida, Polymastiidae and Cladorhizi-dae may have invaded this part of the SouthernOcean from other oceans via deep-sea routes.The deep-sea representatives of these groupsseem more closely related to other deep-seafaunas, whereas the shelf species, especially ofthe Antarctic Rossellidae (Hexactinellida), arelargely endemic (Janussen et al., 2004). Mole-cular and paleontological data point in thesame direction. A more detailed understandingof these biogeographic patterns probably canbe gained through an analysis of the Antarcticdeep-sea sponge fauna as a whole.
ARTICLE IN PRESS
0
2
4
6
8
10
12
14
16
18
III-
153A
I-42E II-
133E
III-
133E
II-
131E
III-
142A
III-
151A
II-
135A
III-
102A
II-
138A
II-
143A
II-
140E
III-
88A
III-
81E
III-
78A
III-
21E
III-
16E
ANDEEP-sponges: Family and species distribution
Leucettidae
Heteropiidae
Clathrinidae
Leucascidae
Amphoriscidae
Euplectellidae
Rossellidae
Euretidae
Farreidae
Halichondriidae
Axinellidae
Latrunculiidae
Isodictyidae
Mycalidae
Cladorhizidae
Tedaniidae
Myxillidae
Raspailiidae
Microcionidae
Acarnidae
Stylocordylidae
Suberitidae
Polymastiidae
Pachastrellidae
Tetillidae
Fig. 4. Distribution of species among sponge families at ANDEEP stations, arranged from E (left) to W (right). Families Leucettidae,
Heteropiidae, Clathrinidae and Leucascidae belong to class Calcarea. Families Euplectellidae, Euretidae, Farreidae and Rossellidae
belong to the class Hexactinellida. The remaining families belong to the class Demospongiae.
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–18751872

Author's personal copy
(9) The ANDEEP samples provided the firstrecords of deep-sea Calcarea (mostly newspecies) from the Antarctic Ocean. Theyconstitute a rare, but stable, fauna componentin the deep Weddell Sea.
(10) The Southern Ocean deep-sea fauna, includingmegafauna such as sponges, is highly diverseand deserves further attention from marinebiologists.
Acknowledgments
We are indebted to many colleagues, especiallyto Hans Tore Rapp (Bergen), Alexander Plotkin(St. Petersburg), Henry M. Reiswig (Victoria) andKonstantin R. Tabachnick (Moscow), for the goodcooperation and valuable discussions. Thanks alsoto Andrew Gooday (Southampton) for correctionof the English text and to two anonymous reviewersfor critical comments. The drawing of an Antarcticmap (Figure XX) was made by Ms. Saskia Dimter(Senckenberg, Frankfurt/M.). Thanks are due tothe Deutsche Forschungsgemeinschaft (DFG) for
supporting the research of Antarctic sponges byDorte Janussen (JA 1063/1-1, JA 1063/11-2), to theDFG (436 RUS 17/92/06) and CeDAMar Taxono-mist Exchange Program for supporting the invita-tion of colleges from abroad to Senckenberg,Frankfurt/M., and to COBICE and SYNTHESYSfor financing research visits by Dorte Janussento the ZMUC. This is ANDEEP publicationnumber 92.
References
Arnaud, P.M., Hain, S., 1992. Quantitative distribution of the
shelf and slope molluscan fauna (Gastropoda, bivalvia)
of the Eastern Weddell Sea (Antarctica). Polar Biology 12,
103–109.
Barthel, D., Tendal, O.S., 1989. The sponge fauna of the deep
Weddell Sea: status and the need for further biological and
faunistic investigations. Deep-Sea Newsletter 16, 8–9.
Barthel, D., Tendal, O.S., 1990. Ecology and taxonomy of
sponges in the eastern Weddell Sea shelf and slope commu-
nities. Berichte zur Polarforschung 68, 120–130.
Barthel, D., Tendal, O.S., 1992. The Antarctic deep-sea
hexactinellid fauna. Deep-Sea Newsletter 19, 20–22.
ARTICLE IN PRESS
0
1
2
3
4
5
6
7
8
9
800-1000m 1000-2000m 2000-3000m 3000-4000m 4000-5000m
Tetillidae
Pachastrellidae
Polymastiidae
Suberitidae
Stylocordylidae
Acarnidae
Microcionidae
Raspailiidae
Myxillidae
Tedaniidae
Cladorhizidae
Mycalidae
Isodictyidae
Latrunculiidae
Axinellidae
Halichondriidae
Chalinidae
Farreidae
Euretidae
Rossellidae
Euplectellidae
Amphoriscidae
Leucascidae
Clathrinidae
Heteropiidae
Leucettidae
Fig. 5. Depth distribution of families of ANDEEP sponges. Families’ attribution to Poriferan classes as in Fig. 4.
D. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1873

Author's personal copy
Barthel, D., Tendal, O.S., 1993. The sponge association of the
abyssal Norwegian-Greenland Sea: species composition,
substrate relationships and distribution. Sarsia 78, 83–96.
Barthel, D., Tendal, O.S., 1994. Antarctic hexactinellida.
Synopses of the Antarctic Benthos, vol. 6, Koeltz Scientific
Books, pp. 1–154.
Bruckner, A., 2006. Taxonomy and paleoecology of lyssacinosan
Hexactinellida from the Upper Cretaceous (Coniacian) of
Bornholm, Denmark, in comparison with other Postpaleozoic
representatives. Abhandlungen der Senckenbergischen Nat-
urforschenden Gesellschaft 564, 1–103.
Bruckner, A., Janussen, D., 2005. The first entirely preserved
fossil sponge species of the genus Rossella (Hexactinellida)
from the Upper Cretaceous of Bornholm, Denmark. Journal
of Paleontology 79 (1), 21–28.
Burton, M., 1928. A comparative study of the characteristics of
shallow-water and deep-sea sponges, with notes on their
external form and reproduction. Journal of the Quekett
Microscopical Club 16, 49–50.
Burton, B., 1932. Sponges. Discovery Reports 6, 237–392.
Burton, M., 1938. Non-calcareous sponges. Scientific Reports of
the Australasian Antarctic Expedition 1911–1914. Ser. C. 9, 5,
1-22.
Dayton, P.K., Robilliard, G.A., Paine, R.T., Dayton, L.B., 1974.
Biological accommodation in the benthic community at
McMurdo Sound, Antarctica. Ecological Monographs 44,
105–128.
Gray, J.E., 1867. Notes on the arrangement of sponges, with the
descriptions of some new genera. Proceedings of the
Zoological Society of London 1867 (2), 492–558.
Gutt, J., Koltun, V.M., 1995. Sponges of the Lazarev and
Weddell Sea, Antarctica: explanations for their patchy
occurrence. Antarctic Science 7, 27–34.
Hentschel, E., 1914. Monaxone Kieselschwamme und Hornsch-
wamme. Deutsche Sudpolar-Expedition 15. Zoologie 7,
35–142.
Hooper, J.N.A., Van Soest, R.W.M., 2002. Preface. In: Hooper,
J.N.A., Van Soest, R.W.M. (Eds.), Systema Porifera. Kluwer
Academic, New York.
Ilan, M., Gugel, J., Galil, B.S., Janussen, D., 2003. Small bathyal
sponge species from east Mediterranean revealed by a non-
regular soft bottom sampling technique. Ophelia 57, 145–160.
Janussen, D., 2003. First report on the deep sea Porifera from the
Northern Weddell Sea and the slope of South Sandwich
Trench. Berichte zur Polar-und Meeresforschung 104–108,
470.
Janussen, D., 2006. Collection of Porifera (sponges) during
ANDEEP III. Berichte zur Polar-und Meeresforschung 533,
174–178.
Janussen, D., Rapp, H.T., Tendal, O.S., 2003. A myth vanished:
calcareous sponges are alive and well at abyssal depths. Deep-
Sea Newsletter 32, 17–19.
Janussen, D., Tabachnick, K.R., Tendal, O.S., 2004. Deep-sea
Hexactinellida (Porifera) of the Weddell Sea. Deep-Sea
Research II 51 (14–16), 1857–1882.
Kirkpatrick, R., 1907. Porifera 1. Hexactinellida. National
Antarctic Expedition 1901–1904. Natural History 3, 1–25.
Kirkpatrick, R., 1908. Porifera 2. Tetraxonida. National Ant-
arctic Expedition 1901–1904. Natural History 4, 1–56.
Klitgaard, A.B., Tendal, O.S., 2004. Distribution and species
composition of mass occurrences of large-sized sponges in the
northeast Atlantic. Progress in Oceanography 61, 57–98.
Koltun, V.M., 1964. Sponges of the Antarctic I. Tetraxonida and
Cornacuspongida. Biological reports of the Soviet Antarctic
Expedition (1955–1958) II, Academy of Sciences of the
USSR. Zoological Institute, pp. 1–133. (English translation
1966 by Israel program for scientific translations).
Koltun, V.M., 1976. Porifera Part I: Antarctic sponges. B.A.N.Z.
Antarctic Research Expedition 1929–1931. Reports ser. B.
IX(4), 147–198.
Levi, C., 1964. Spongiaires des zones bathyale, abyssale et hadale.
Galathea Report 7, 63–112.
Linse, K., Brandt, A., Bohn, J., Danis, B., De Broyer, C.,
Heterier, V., Hilbig, B., Janussen, D., Gonzales, P.J.L.,
Schwabe, E., Thomson, M.R.A., 2006. Megabenthos col-
lected by the Agassiz trawl. Berichte zur Polar-und Meer-
esforschung 533, 161–173.
Maldonado, M., Young, C.M., 1996. Bathymetric patterns of
sponge distribution on the Bahamian slope. Deep-Sea
Research 43, 897–915.
Malyutina, M., 2004. Russian deep-sea investigations of Antarc-
tic fauna. Deep-Sea Research II 51, 1551–1570.
McClintock, J.B., Amsler, C.D., Baker, B.J., van Soest, R.W.M.,
2005. Ecology of Antarctic marine sponges: an overview.
Integrated Comparative Biology 45, 359–368.
Ostapoff, F., 1965. Antarctic oceanography. In: Van Mieghen, J.,
van Oye, P. (Eds.), Biogeography and Ecology in Antarctica.
Dr. W. Junk Publishers, The Hague, pp. 97–126.
Pickard, G.L., Emery, W.J., 1982. Descriptive Physical Oceano-
graphy. Pergamon Press, pp. 1–249.
Plotkin, A.S., Janussen, D., Tendal, O.S. Polymastiidae and
Suberitidae (Porifera: Demospongiae: Hadromerida) of the
deep Weddell Sea, Antarctic. Zootaxa, submitted.
Ridley, S.O., Dendy, A., 1887. Report on the Monaxonida
collected by H.M.S. Challenger during the years 1873–76.
Report on the scientific results of the exploring voyage of H.
M. S. ‘Challenger’. Zoology 20 (59), 1–275 pp. 1–51.
Schmidt, O., 1870. Grundzuge einer Spongien-Fauna des
atlantischen Gebietes, Wilhelm Engelmann, Leipzig: iii–iv,
1–88, pls. I–IV.
Schulze, F.E., 1887. Report on the Hexactinellida. Report on the
scientific results of the voyage of H.M.S. Challenger. Zoology
21 (53), 514.
Schulze, F.E., 1910. Die Hexactinelliden. Deutsche Sudpolar-
Expedition 12. Zoologie 4, 1–62.
Sokolova, M.N., 2000. Feeding and Trophic Structure of the
Deep-Sea Macrobenthos. Smithsonian Institution Libraries,
Washington, DC, pp. 1–264.
Tabachnik, K.R., 1994. Distribution of recent Hexactinellida. In:
Van Soest, R.W.M., Van kempen, T.M.G., Braekman, J.-C.
(Eds.), Sponges in Time and Space. A.A. Balkema, Rotter-
dam, pp. 225–232.
Tabachnik, K.R., 2002. Family rosellidae Schulze, 1885. In:
Hooper, J.N.A., Van Soest, R.W.M. (Eds.), Systema Porifera.
Kluwer Academic, New York, pp. 1441–1505.
Tanita, S., 1959. Sponges. Special Publication from the Seto
Marine Biological Laboratory 1, 1–8.
Topsent, E., 1901. Spongiaires. Expedition Antarctique Belge.
Resultats du S. Y. Belgica en 1897–1897–1899. Rapports
Scientifiques Zoologie, 54pp.
Topsent, E., 1913 (1920). Spongiaires de l’Expedition Antarc-
tique Nationale Ecossaise. Report on the scientific Results of
the Voyage of S.Y. ‘‘Scotia’’ during the years 1902, 1903, and
1904, 7, pp. 3–72.
ARTICLE IN PRESSD. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–18751874

Author's personal copy
Topsent, E., 1915a. Spongiaires recueillis par la ‘‘Scotia’’ dans
l’Antarctique (1903–1904). Supplement. Transaction of the
Royal Society of Edinburgh 51, 35–43.
Topsent, E., 1915b. Une Rossella des Acores (Rossella nodas-
trella n. sp.). Bulletin de l’Institut Oceanographique 303, 1–6.
Ushakov, P.V., 1963. Quelques particularites de la bionomie
benthique de l’Antarctique de l’Est. Cahiers de Biologie
Marine 4, 81–89.
Vacelet, J., 2006. New carnivorous sponges (Porifera, Poeciloscler-
ida) collected from manned submersibles in the deep Pacific.
Zoological Journal of the Linnean Society 148, 553–584.
Vacelet, J., Boury-Esnault, N., 1995. Carnivorous sponges.
Nature 373, 333–335.
Vacelet, J., Duport, E., 2004. Prey capture and digestion in the
carnivorous sponge Asbestopluma hypogea (Porifera: Demos-
pongiae). Zoomorphology 123, 179–190.
Van Soest, R.W.M., 1994. Demosponge distribution patterns. In:
Van Soest, R.W.M., Van klempen, T.M.G., Braeksman, J.-C.
(Eds.), Sponges in Time and Space. A.A. Balkema, Rotter-
dam, pp. 225–232.
Vinogradova, N.G., 1962. Vertical zonation in the distribution of
deep-sea benthic fauna in the ocean. Deep-Sea Research 8,
245–250.
Wilton, D.W., Brown, R.N.R., 1908. Zoological log. Report on
the Scientific Results of the Voyage of S.Y. ‘‘Scotia’’ During
the Years 1902, 1903, and 1904, pp. 1–84.
ARTICLE IN PRESSD. Janussen, O.S. Tendal / Deep-Sea Research II 54 (2007) 1864–1875 1875
Top Related
Copyright © 2022 FDOKUMEN

