







Bahasa
Halaman
Hukum


Distinctive microRNA expression signatures in proton-irradiatedmice
Shahid Yar Khan • Muhammad Akram Tariq • James Patrick Perrott •
Christopher Drew Brumbaugh • Hyunsung John Kim •
Muhammad Imran Shabbir • Govindarajan T. Ramesh •
Nader Pourmand
Received: 16 April 2013 / Accepted: 14 June 2013 / Published online: 2 July 2013
� Springer Science+Business Media New York 2013
Abstract Proton particles comprise the most abundant
ionizing radiation (IR) in outer space. These high energy
particles are known to cause frequent double- and single-
stranded DNA lesions that can lead to cancer and tumor
formation. Understanding the mechanism of cellular
response to proton-derived IR is vital for determining
health risks to astronauts during space missions. Our
understanding of the consequences of these high energy
charged particles on microRNA (miRNA) regulation is still
in infancy. miRNAs are non-coding, single-stranded RNAs
of *22 nucleotides that constitute a novel class of gene
regulators. They regulate diverse biological processes, and
each miRNA can control hundreds of gene targets. To
investigate the effect of proton radiation on these master
regulators, we examined the miRNA expression in selected
mice organs that had been exposed to whole-body proton
irradiation (2 Gy), and compared this to control mice (0 Gy
exposure). RNA was isolated from three tissues (testis,
brain, and liver) from treated and control mice and sub-
jected to high-throughput small RNA sequencing. Bioin-
formatics analysis of small RNA sequencing data revealed
dysregulation of (p \ 0.05; 20 up- and 10 down-regulated)
14 mouse testis, 8 liver, and 8 brain miRNAs. The statis-
tically significant and unique miRNA expression pattern
found among three different proton-treated mouse tissues
indicates a tissue-specific response to proton radiation. In
addition to known miRNAs, sequencing revealed differ-
ential expression of 11 miRNAs in proton-irradiated mice
that have not been previously reported in association with
radiation exposure and cancer. The dysregulation of miR-
NAs on exposure to proton radiation suggest a possible
mechanism of proton particles involvement in the onset of
cell tumorgenesis. In summary, we have established that
specific miRNAs are vulnerable to proton radiation, that
such differential expression profile may depend upon the
tissue, and that there are more miRNAs affected by proton
radiation than have been previously observed.
Keywords miRNA � Differential expression �Proton radiation � Mice
Introduction
The space radiation environment consists mainly of trap-
ped particle radiation, solar particle radiation, galactic
cosmic radiation (GCR), and also ionizing radiation (IR).
IR is a well-known modality used in cancer treatment that
also induces oxidative genotoxic stress. It is known that IR
causes cellular damage by either directly damaging DNA,
or by the formation of free radicals [1]. Among IR, protons
S. Y. Khan � M. A. Tariq � J. P. Perrott �C. D. Brumbaugh � H. J. Kim � M. I. Shabbir �N. Pourmand (&)
Department of Biomolecular Engineering, University
of California Santa Cruz, Santa Cruz, CA 95064, USA
e-mail: [email protected]; [email protected]
M. A. Tariq
Center for Bionanotechnology and Environmental Research,
Texas Southern University, Houston, TX 77004, USA
Present Address:
M. I. Shabbir
Department of Bioinformatics and Biotechnology, Faculty
of Basic Applied Sciences, International Islamic University,
Sector H10, Islamabad, Pakistan
G. T. Ramesh
Molecular Toxicology Laboratory, Center for Biotechnology
and Biomedical Sciences, Norfolk State University,
700 Park Avenue, Norfolk, VA 23504, USA
123
Mol Cell Biochem (2013) 382:225–235
DOI 10.1007/s11010-013-1738-z

particles represent the largest fraction (85 %) of GCRs and
the understanding of the biological effects of high energy
protons is important in the neoplastic transformation that
can lead to cancer. A limited number of studies have been
performed to evaluate the impact of proton radiations on
the induction of neoplastic transformation in biological
systems. Recently, Sorokina and co-authors studied the role
of proton and c-radiation in the formation of double-
stranded breaks (DSB). They showed that protons and
X-rays at 2 Gy dose rate induced very similar number of c-
H2AX and 53BP1 foci in human umbilical vein cells [2]
and revealed that the biological effects of proton radiation
are similar to the effects of c-radiation and other low-linear
energy transfer (LET) radiation [2]. In another study, HeLa
6 skin fibroblast human hybrid cells were used for the
estimation of neoplastic transformation frequencies using
different proton doses (5–600 mGy) [3]. They showed
increase in transformation efficiency at doses of 100 and
300 mGy and significant neoplastic transformation fre-
quency was observed at 600 mGy proton radiation dose
[3]. Based on these recent findings, it is clear that a
molecular understanding of proton radiation role in cancer
initiation is vital for space exploration missions and safe
radiation therapies for cancer treatment.
IR can induce mRNA transcriptional changes and
aberrant expression of a variety of non-coding small RNA
in cells. Among these non-coding small RNAs, microRNA
(miRNA) plays a supremely important role in gene regu-
lation through the repression of target genes. Approxi-
mately, 60 % of human protein-coding genes are predicted
to contain miRNA binding sites within un-translated
regions (UTRs) [4]. It has been shown in some studies that
the levels of miRNAs change in response to IRs [5] and
also that miRNAs play an important role in cellular defense
[6, 7].
Several miRNAs have been previously reported as
important players in cancer biology as well as in IR-
induced changes in mouse and human cell lines. In litera-
ture, the up-regulation of miR-100 in mouse plasma after
whole-body c-irradiation (0.5, 2, and 10 Gy), in human
malignant glioma cells and in psoriasis, an autoimmune
disease has been reported [8–10]. In another study, up-
regulation of miR-100 has been shown to down-regulate the
expression of ataxia telangiectasia-mutated (ATM) in
human malignant glioma cells (M059J), which in turn
causes an increased sensitivity to IR [9]. It should also be
noted that the down-regulation of miR-100 was found in
human prostate cancer and normal fibroblast cell lines
exposed to IR/X-rays as compared to the up-regulation in
response to proton radiation [1, 11]. In human tumor
pathology, miR-100 was found to be up-regulated in acute
myeloblastic leukemia (AML) [12] and down-regulated in
some cancers like low-grade bladder cancer, oral cancer,
ovarian cancer, and hepatocellular carcinomas [13–16].
Another miRNA, miR-205, has been previously observed
to be up-regulated in cervical cancer tissues, head and neck
cancer cell lines, squamous cell carcinoma cell lines, and
non-small cell lung cancers [17–23] and down-regulated in
prostate cancer [24, 25]. Several studies have revealed
heterogeneity of miR-205 expression among different
breast cancer subtypes [26–30].
Two independent studies showed the over-expression of
let-7d in normal human fibroblast cell lines exposed to
X-ray (200 cGy/min) treatment, in human dermal micro-
vascular endothelial cells (HDMEC) irradiated with 2 Gy
photon radiation and in c-radiation-treated human thyroid
cells [1, 31, 32]. In another study, irradiation of normal
human lung epithelial line (CRL2741) along with three
lung cancer cell lines (A549, A427 (carcinoma), and
H4441 (adenocarcinoma)) revealed a down-regulation of
let-7 [33]. Three microarray-based cancer experiments
using human tissue also revealed let-7d up-regulation
in lung cancer and down-regulation in pancreatic cancer
[34–37]. In human cancer, miR-423-3p over-expression has
been found in head and neck squamous cell carcinoma
(HNSCC) patients [38], metastatic breast cancer patients
[39], in metastatic neuroblastoma (NB), and hepatocellular
carcinoma tissue (HCC) [40, 41].
Recently, two independent studies reported the down-
regulation of miR-451 in human non-small cell lung car-
cinoma (NSCLC) and gastric and colorectal cancer [42–44]
and significant over-expression of miR-451 among non-
relapsed and relapsed patients with HNSCCs [38].
MiR-146b has been previously found to be up-regulated
in adult papillary thyroid carcinomas (PTCs) with extra-
thyroidal invasion and BRAF mutation and in c-radiation-
treated human thyroid cells [45–49]. While down-regula-
tion of miR-146b in mouse plasma was reported following
whole-body c-radiation [8]. Furthermore, microarray and
qPCR analysis revealed a higher expression of miR-146b in
a murine model of acute and chronic asthma [50].
Recently, Wang and co-workers [42] reported a signif-
icant role of miR-138 on IR-induced DNA damage pathway
by targeting histone H2AX in a human osteosarcoma cell
line [51]. The miR-138 up-regulation and down-regulation
revealed a correlation with chromosomal instability and
cellular sensitivity to DNA-damaging agents [51]. The
down-regulation of miR-138 was also observed in ana-
plastic thyroid carcinoma (ATC) as compared to PTC cell
lines, in which the down-regulation of miR-138 has also
been associated with enhanced telomerase reverse trans-
criptase (TERT) expression [52]. Similarly, reduced miR-
138 level is also associated with enhanced cell growth and
invasion in HNSCC [53, 54].
Furthermore, up-regulation of miR-99b was also repor-
ted in mouse plasma after 2 Gy c-radiation [8]. In human
226 Mol Cell Biochem (2013) 382:225–235
123

tumor pathology, miR-99b was up-regulated in patients
with synovial sarcoma [55] and in older adults with
primary melanoma [56]. miRNA expression profiling
revealed a down-regulation of miR-409-5p in human brain
tumor [57], while in another recent study, Nikiforova
reported c-irradiated (1.0 or 10 Gy) human thyroid cells
revealing a high expression of miR-409-5p [32]. Up-regu-
lation of miR-409-5p was also observed in glioblastomas
(GBM) as compared to oligodendrogliomas (ODG) [57],
while a down-regulation of miR-409-5p was observed for
mouse model of Rett syndrome, an X-linked neurological
inherited disorder [58].
Despite the growing evidence of the differential expres-
sion of miRNAs in response to IR [5, 6, 59], little is known
about miRNA response to proton radiation in humans and
mice. Therefore, we sought to examine the effect of proton
irradiation on whole mice with respect to the possible
involvement of miRNA in the onset of cell tumorgenesis. To
this end, we analyzed the global miRNA expression profile in
the brain, liver, and radiation-sensitive testis tissue of proton-
irradiated mice. Our results show that proton radiation can
dysregulated miRNA levels in these three tissues and reveal a
potential pathway by which proton irradiation could induce
pathogenic mutations in miRNA that could lead to cancer.
While further studies are required to define a comprehensive
understanding of tissue-specific miRNA expression profiles
in response to proton radiation, these results show promise
for the possibility that miRNA expression profiles may one
day be useful as diagnostic markers.
Results
MicroRNA expression profiling
In this study, we employed a mouse model to analyze and
compare differential expression of miRNA in brain, liver,
and testis tissues induced by high energy, whole-body
radiation exposure. Eight mice were divided in two groups,
a non-irradiated ‘control’ (group 1) and an irradiated
‘exposed’ group (group 2). Four biological replicates were
used for the control four were used for exposed samples for
brain and testis tissues. These tissues were chosen because
they represent a highly protected tissue (brain) and a highly
sensitive tissue (testis). In the course of study, we recog-
nized that liver tissues would provide a possible interme-
diate exposure and began to collect those data as well. For
liver tissue, therefore, only two biological replicates are
presented for each of the control and exposed samples. The
Illumina HiSeq was used to generate small RNA reads
(2 9 50 bp) isolated from brain, liver, and testis of proton-
irradiated and non-irradiated mice for comprehensive
miRNA differential expression analysis.
Approximately, 87.47, 43.65, and 81.17 million small
RNA reads were obtained for brain, liver, and testis,
respectively (Fig. 1). Out of these total mapped small RNA
reads, 14.71 million (16.79 %) brain miRNA, 1.90 million
(4.59 %) liver miRNA, and 2.71 million (3.27 %) testis
miRNA were obtained after alignment against known
mouse miRNA with 0 mismatches (miRBase revision 17,
http://www.mirbase.org/). We observed a high proportion
(13.67 million; 8.99 %) of testis small RNA reads mapped
to Piwi-interacting RNA (piRNA). A surprisingly large
fraction of reads (28 million; 4.47 % in brain; 4.43 % in
liver and 4.32 % in testes) did not map to any known small
RNA moiety in brain, liver, and testis tissues (Fig. 1).
DESeq software was used for the differential expression
analysis of miRNA data obtained from high-throughput
sequencing. For analysis, strict parameters were used
including higher ‘‘fold change’’ and a p value lower than
0.05. Analysis revealed 30 differentially expressed miRNA
in three mouse tissues (Table 1; Fig. 2).
Mouse brain microRNA differential expression
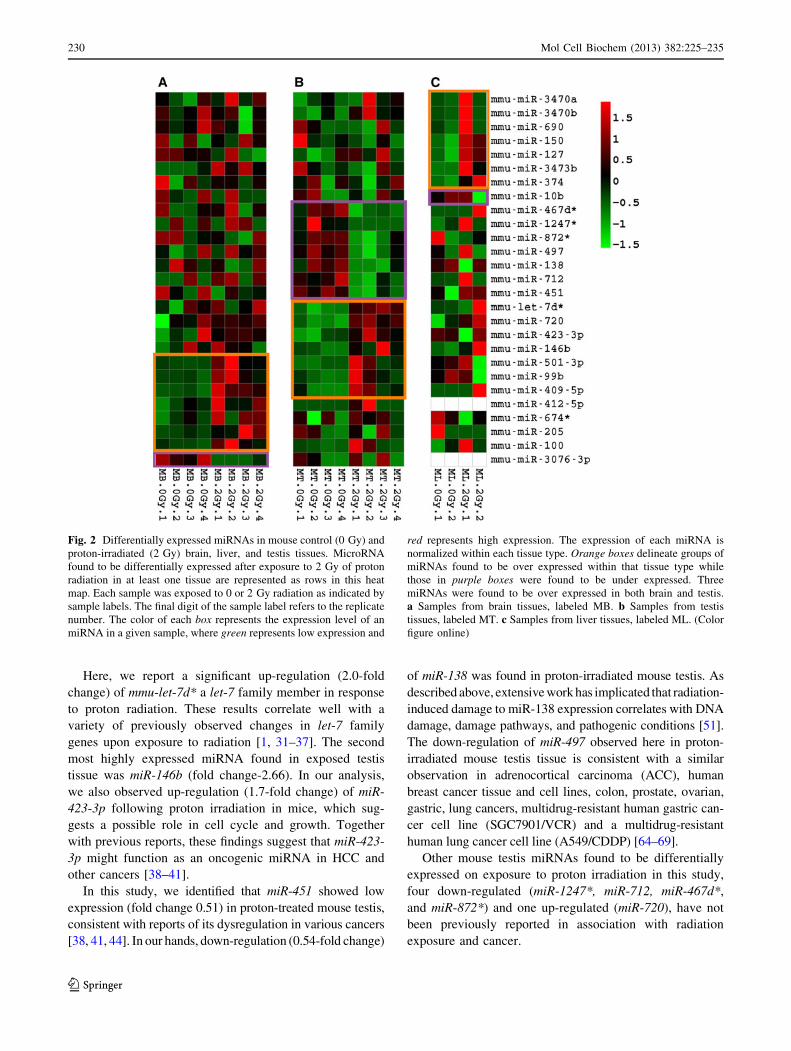
Cluster analysis revealed a significant dysregulation of eight
miRNAs in proton-irradiated mouse brain tissue compared to
non-irradiated brain tissue (Fig. 2a). The miRNA expression
data showed a good correlation among the four controls and
among the four treated brain biological replicates (0.95 for
0 Gy and 0.95 for 2 Gy). Among these differentially expres-
sed miRNA, one down-regulated (miR3076-3p) and seven up-
regulated (miR-409-5p, miR-205, miR-100, miR-501-3p, miR-
99b, miR-674* and miR-412-5p) miRNAs were observed
(Table 1; Fig. 2a).
Mouse testis microRNA differential expression
We also identified 14 testis miRNAs from the DESeq
analysis that we considered to be differentially expressed
by irradiation at a dose of 2 Gy (Fig. 2b). The miRNA
expression data showed a good correlation among the four
controls and also among the four treated biological repli-
cates (0.90 for 0 Gy and 0.89 for 2 Gy). Among these
differentially expressed miRNA, seven up-regulated (miR-
409-5p, miR-146b, miR-501-3p, miR-99b, let-7d*, miR-
423-3p and miR-720) and seven down-regulated (miR-
1247*, miR-712, miR-467d*, miR-451, miR-138, miR-872*
and miR-497) miRNAs were found (Table 1; Fig. 2b).
Mouse liver microRNA differential expression
Bioinformatics analysis also revealed differential expres-
sion of eight miRNAs among treated and control liver
tissue (Fig. 2c). The miRNA expression data showed a
Mol Cell Biochem (2013) 382:225–235 227
123

good correlation between the two controls and also the two
proton-treated biological replicates (0.88 for 0 Gy and 0.86
for 2 Gy). Among these differentially expressed miRNA,
two down-regulated (miR-3473b and miR-10b) and six
up-regulated (miR-3470b, miR-3470a, miR-690, miR-150,
miR-374 and miR-127) miRNAs were observed (Table 1;
Fig. 2c).
Mouse brain, testis, and liver microRNA editing
We performed alignments of mature miRNA reads with
one mismatch to find out miRNA editing patterns in mouse
control (0 Gy) and proton-treated (2 Gy) brain, testis, and
liver tissues. We used only high quality reads and align-
ment of at least 10 reads to remove false positive miRNA
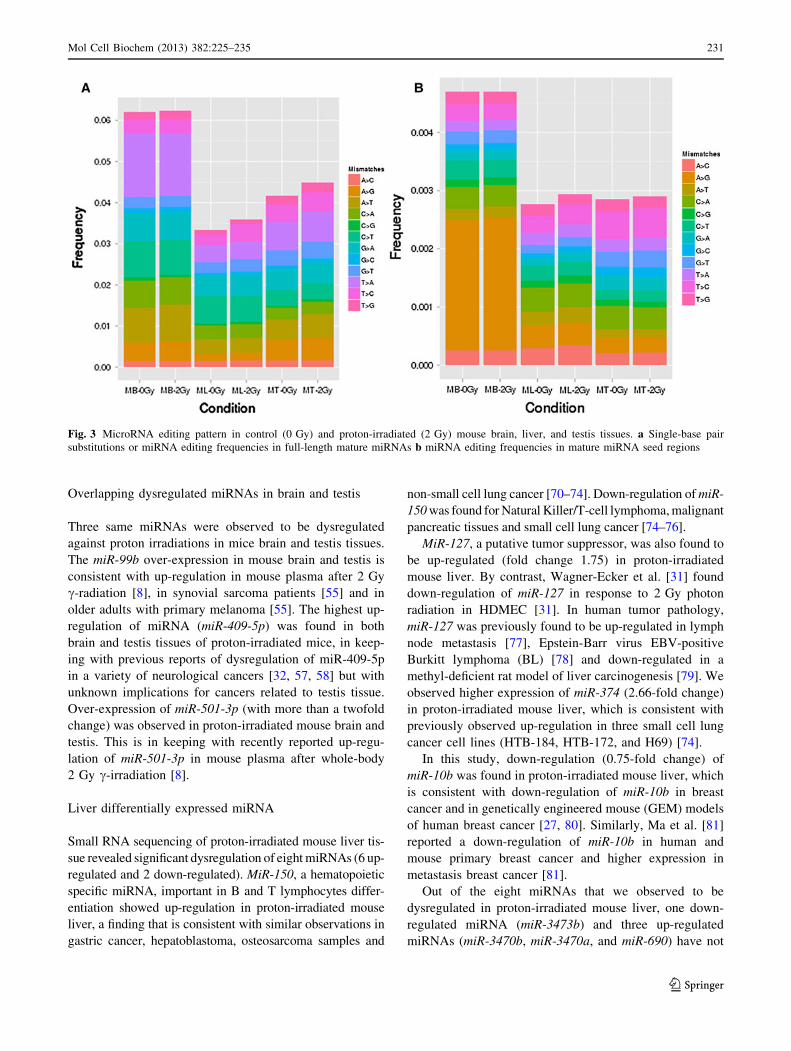
editing events. Approximately, 5.11 % of total reads hav-
ing a single mismatch in mature miRNA reads were
mapped (Fig. 3). We observed a high frequency of single
mismatch reads in brain (*5.61 %) and testis (*4.2 %)
tissues and slightly lower frequency of mismatch in liver
tissues (*2.5 %). We analyzed the frequencies of all
possible single-base substitutions, with particular focus on
the cytosine deamination (C?U (C?T) conversion) and
adenosine deamination (A?I (A?G) conversion).
We observed 24.7, 13.87, and 7.6 % single mismatch
frequencies for T?A, C?T, and A?G substitutions in
brain mature miRNA. We also identified a dominant ratio
(*45.5 %) for A?G single-base substitution in the seed
region of mature brain miRNA (Fig. 3a, b). Liver mature
miRNA reads revealed a higher frequency of C?T
(18.81 %) and A?G (5.22 %) in all substitutions. The
single-base pair mismatch analysis of the seed region
revealed a higher frequency for A?G (12.5 %) as com-
pared to C?T (8.0 %) (Fig. 3a, b). The single mismatch
alignment of testis miRNA reads revealed a higher fraction
of different substitutions. We observed over-representation
of T?A, T?C, A?G, and C?T substitutions, which
accounted for 24.70, 13.92, 12.02, and 8.78 % of single
Fig. 1 Pie charts illustrating the frequency of different non-coding
RNA species. a Pie chart representing brain non-coding RNA
frequencies. Small RNA sequencing revealed a high frequency
(19.86 and 16.79 %) for small nucleolar RNA (snoRNA) and
microRNA (miRNA), respectively, in brain tissue. b Pie chart
representing non-coding RNA frequencies for testes. Small RNA
sequencing revealed a high frequency (21.70 and 8.99 %) for
snoRNA and Piwi-interacting RNA (piRNA), respectively. c Pie
chart showing the liver non-coding RNA frequencies. Small RNA
sequencing revealed a high frequency (20.73 and 5.40 %) for
snoRNA and transfer RNA (tRNA), respectively. Small non-messen-
ger RNA (snmRNA); small cajal body specific RNA (scaRNA); small
nuclear RNA (snRNA); robosomal RNA (rRNA)
228 Mol Cell Biochem (2013) 382:225–235
123

mismatch reads, respectively (Fig. 3a, b). We did not
observe any significant differences in miRNA editing fre-
quencies among control (0 Gy) and proton-irradiated
(2 Gy) mouse brain, liver, and testis tissues (Fig. 3a, b).
Discussion
Mouse whole-body proton irradiation (2 Gy) revealed a
significant dysregulation of miRNA in the three tissues
examined (brain, liver, and testis). To approximate the
maximum dose that astronauts may receive from Solar
Particle Events during space missions [60, 61], we used a
2 Gy dose of proton radiation that is equivalent to this
exposure. The tissues used were selected on the basis of
their sensitivity to IR, with brain being the least sensitive,
liver as moderate, and testis as the most sensitive. This
study of selected tissues provides a glimpse of tissue
specificity and sensitivity, as well as several newly iden-
tified, miRNAs whose expression levels change in response
to proton radiation exposure.
Brain differentially expressed miRNA
Mouse brain tissue exposed to 2 Gy proton radiation
treatment revealed a significant up-regulation of seven
miRNAs and down-regulation of one miRNA. Interest-
ingly, we observed an entirely new miRNA expression
profile in proton-irradiated mice brain tissue, which dif-
fered from that observed by Koturbash and co-workers
[62]. This phenomenon suggests a unique miRNA signa-
ture, exclusive to brain tissue that is expressed only in
response to X-ray and proton radiation exposure. The dif-
ference among differentially expressed miRNA can be
attributed to both the radiation dose (1 Gy by Koturbash vs
2 Gy in our study) and the platform used for analysis
(microarray vs next generation sequencing).
Overall, we identified eight dysregulated miRNAs in
proton-irradiated mouse brain and out of these 8, 4 miR-
NAs (miR-99b, miR-100, miR-501-3p and miR-409-5p)
have previously been shown to be dysregulated after
exposure to IR in mice and various human cell lines (see
‘‘Introduction’’ section). The four novel miRNAs whose
expression changed in response to proton irradiation in
mice deserve further study to determine their roles in
IR-induced DNA damage, which could lead to a better
understanding of cancer pathogenesis and ultimately to
earlier diagnosis or new treatments.
Testis differentially expressed miRNA
Small RNA sequencing of testis tissue revealed significant
dysregulation of 14 miRNA (7 up-regulated and 7 down-
regulated). Perplexingly, we did not find any overlap
between these differentially expressed miRNAs and those
identified in a previous microarray study of differential
expression of miRNA in X-ray irradiated mouse testis [63].
The differences observed among differentially expressed
miRNAs may be attributable to the type (X-ray vs proton)
and dose of radiation (2.5 vs 2 Gy) as well as possible
skewing by the analysis platform used (microarray vs next
generation sequencing).
Table 1 MicroRNA species differentially expressed in mouse brain,
liver, and testis after exposure to proton radiation (2 Gy)
MicroRNA Fold change p value
Brain
mmu-miR-409-5pa 4.842 0.000
mmu-miR-205 3.072 0.021
mmu-miR-100 2.530 0.013
mmu-miR-501-3pa 2.380 0.010
mmu-miR-99ba 2.270 0.003
mmu-miR-674b 1.845 0.028
mmu-miR-412-5p 1.546 0.046
mmu-miR-3076-3p 0.064 0.009
Testis
mmu-miR-409-5pa 3.796 0.014
mmu-miR-146b 2.668 0.042
mmu-miR-501-3pa 2.360 0.017
mmu-miR-99ba 2.186 0.035
mmu-let-7db 2.016 0.001
mmu-miR-423-3p 1.780 0.004
mmu-miR-720 1.548 0.046
mmu-miR-1247b 0.000 0.039
mmu-miR-712 0.282 0.035
mmu-miR-467db 0.475 0.023
mmu-miR-451 0.514 0.002
mmu-miR-138 0.543 0.005
mmu-miR-872b 0.600 0.023
mmu-miR-497 0.606 0.017
Liver
mmu-miR-3470b 5.857 0.000
mmu-miR-3470a 5.115 0.001
mmu-miR-690 3.937 0.009
mmu-miR-150 3.014 0.011
mmu-miR-374 2.662 0.050
mmu-miR-127 1.756 0.030
mmu-miR-3473b Inf 0.003
mmu-miR-10b 0.755 0.031
a Indicates the overlapping microRNA expressing among brain and
testis tissuesb Indicates an miRNA expressed at low levels relative to the miRNA
in the opposite arm of a hairpin
Mol Cell Biochem (2013) 382:225–235 229
123
Here, we report a significant up-regulation (2.0-fold
change) of mmu-let-7d* a let-7 family member in response
to proton radiation. These results correlate well with a
variety of previously observed changes in let-7 family
genes upon exposure to radiation [1, 31–37]. The second
most highly expressed miRNA found in exposed testis
tissue was miR-146b (fold change-2.66). In our analysis,
we also observed up-regulation (1.7-fold change) of miR-
423-3p following proton irradiation in mice, which sug-
gests a possible role in cell cycle and growth. Together
with previous reports, these findings suggest that miR-423-
3p might function as an oncogenic miRNA in HCC and
other cancers [38–41].
In this study, we identified that miR-451 showed low
expression (fold change 0.51) in proton-treated mouse testis,
consistent with reports of its dysregulation in various cancers
[38, 41, 44]. In our hands, down-regulation (0.54-fold change)
of miR-138 was found in proton-irradiated mouse testis. As
described above, extensive work has implicated that radiation-
induced damage to miR-138 expression correlates with DNA
damage, damage pathways, and pathogenic conditions [51].
The down-regulation of miR-497 observed here in proton-
irradiated mouse testis tissue is consistent with a similar
observation in adrenocortical carcinoma (ACC), human
breast cancer tissue and cell lines, colon, prostate, ovarian,
gastric, lung cancers, multidrug-resistant human gastric can-
cer cell line (SGC7901/VCR) and a multidrug-resistant
human lung cancer cell line (A549/CDDP) [64–69].
Other mouse testis miRNAs found to be differentially
expressed on exposure to proton irradiation in this study,
four down-regulated (miR-1247*, miR-712, miR-467d*,
and miR-872*) and one up-regulated (miR-720), have not
been previously reported in association with radiation
exposure and cancer.
Fig. 2 Differentially expressed miRNAs in mouse control (0 Gy) and
proton-irradiated (2 Gy) brain, liver, and testis tissues. MicroRNA
found to be differentially expressed after exposure to 2 Gy of proton
radiation in at least one tissue are represented as rows in this heat
map. Each sample was exposed to 0 or 2 Gy radiation as indicated by
sample labels. The final digit of the sample label refers to the replicate
number. The color of each box represents the expression level of an
miRNA in a given sample, where green represents low expression and
red represents high expression. The expression of each miRNA is
normalized within each tissue type. Orange boxes delineate groups of
miRNAs found to be over expressed within that tissue type while
those in purple boxes were found to be under expressed. Three
miRNAs were found to be over expressed in both brain and testis.
a Samples from brain tissues, labeled MB. b Samples from testis
tissues, labeled MT. c Samples from liver tissues, labeled ML. (Color
figure online)
230 Mol Cell Biochem (2013) 382:225–235
123
Overlapping dysregulated miRNAs in brain and testis
Three same miRNAs were observed to be dysregulated
against proton irradiations in mice brain and testis tissues.
The miR-99b over-expression in mouse brain and testis is
consistent with up-regulation in mouse plasma after 2 Gy
c-radiation [8], in synovial sarcoma patients [55] and in
older adults with primary melanoma [55]. The highest up-
regulation of miRNA (miR-409-5p) was found in both
brain and testis tissues of proton-irradiated mice, in keep-
ing with previous reports of dysregulation of miR-409-5p
in a variety of neurological cancers [32, 57, 58] but with
unknown implications for cancers related to testis tissue.
Over-expression of miR-501-3p (with more than a twofold
change) was observed in proton-irradiated mouse brain and
testis. This is in keeping with recently reported up-regu-
lation of miR-501-3p in mouse plasma after whole-body
2 Gy c-irradiation [8].
Liver differentially expressed miRNA
Small RNA sequencing of proton-irradiated mouse liver tis-
sue revealed significant dysregulation of eight miRNAs (6 up-
regulated and 2 down-regulated). MiR-150, a hematopoietic
specific miRNA, important in B and T lymphocytes differ-
entiation showed up-regulation in proton-irradiated mouse
liver, a finding that is consistent with similar observations in
gastric cancer, hepatoblastoma, osteosarcoma samples and
non-small cell lung cancer [70–74]. Down-regulation of miR-
150 was found for Natural Killer/T-cell lymphoma, malignant
pancreatic tissues and small cell lung cancer [74–76].
MiR-127, a putative tumor suppressor, was also found to
be up-regulated (fold change 1.75) in proton-irradiated
mouse liver. By contrast, Wagner-Ecker et al. [31] found
down-regulation of miR-127 in response to 2 Gy photon
radiation in HDMEC [31]. In human tumor pathology,
miR-127 was previously found to be up-regulated in lymph
node metastasis [77], Epstein-Barr virus EBV-positive
Burkitt lymphoma (BL) [78] and down-regulated in a
methyl-deficient rat model of liver carcinogenesis [79]. We
observed higher expression of miR-374 (2.66-fold change)
in proton-irradiated mouse liver, which is consistent with
previously observed up-regulation in three small cell lung
cancer cell lines (HTB-184, HTB-172, and H69) [74].
In this study, down-regulation (0.75-fold change) of
miR-10b was found in proton-irradiated mouse liver, which
is consistent with down-regulation of miR-10b in breast
cancer and in genetically engineered mouse (GEM) models
of human breast cancer [27, 80]. Similarly, Ma et al. [81]
reported a down-regulation of miR-10b in human and
mouse primary breast cancer and higher expression in
metastasis breast cancer [81].
Out of the eight miRNAs that we observed to be
dysregulated in proton-irradiated mouse liver, one down-
regulated miRNA (miR-3473b) and three up-regulated
miRNAs (miR-3470b, miR-3470a, and miR-690) have not
Fig. 3 MicroRNA editing pattern in control (0 Gy) and proton-irradiated (2 Gy) mouse brain, liver, and testis tissues. a Single-base pair
substitutions or miRNA editing frequencies in full-length mature miRNAs b miRNA editing frequencies in mature miRNA seed regions
Mol Cell Biochem (2013) 382:225–235 231
123

been previously reported in association with radiation
exposure and cancer.
Editing of brain, liver, and testis miRNA
Post-transcriptional modification or editing of mammalian
primary-miRNAs (pri-miRNA) has a large impact on
processing and functionality of mature miRNAs and thus
affects expression and regulation of target genes [82, 83].
Efficient target mRNAs regulation requires a perfect base-
pairing of seed region (2–8 nucleotides) from the 50-end of
mature miRNA [84]. The editing events within the highly
conserved seed sequence can increase and decreases the
specificity as well as the numbers of target genes [82, 83,
85]. In this study, mature miRNA showed higher adenosine
deamination (A?I) frequency in mouse brain. The higher
adenosine deamination phenomenon can be attributed to
the brain-specific expression of the mammalian adenosine
deaminases (ADARs) responsible for A?I editing. We
observed a *6.0 % higher A?G substitution frequency in
the seed region than the full-length mature miRNA
(Fig. 3a, b). In contrast, the frequency of C?T substitu-
tions in the seed region was *3.0 % lower when compared
to mature miRNA (Fig. 3a, b). Similarly to brain miRNA,
we observed a *2.5 % increase in the A?G substitution
ratio and a *2.5 % decline in the C?T substitution ratio
for the seed region as compared to mature miRNA in liver
tissue (Fig. 3a, b). Previously, Joyce et al. [10] demon-
strated through deep sequencing of human psoriasis skin
that a higher frequency of adenosine deamination and a
lower frequency of cytosine deamination occurred in the
seed region as compared to mature miRNA [10]. In testis
miRNA reads, we observed a lower frequency of A?G
and C?T substitutions in mature miRNA sequence as well
as in the seed region. Meanwhile, we observed higher
frequencies (24.70 and 16.5 %) for T?A and T?C sub-
stitutions in full-length mature miRNA as well as the seed
region (Fig. 3a, b). The functional significance of miRNAs
A?I editing catalyzed by adenosine deaminases has been
proven in various research [83, 86]. Tissue specific A?I
editing within the seed region of mature hsa-miR-376
cluster transcript can lead to modifications in the target
recognition of mRNA targets in some tissues [83].
Conclusion
Deep sequencing of small RNA species revealed a unique
miRNA expression signature and post-transcriptional
modification for each tissue (brain, lever, and testis) from
whole-body proton-irradiated mice. To date, there have
been limited studies showing the response of these three
cell-types on miRNA expression following low doses of
proton radiation. All three tissues assessed in this study
showed differentially expressed profiles, with testis, the
most sensitive tissue, showing the greatest number of
responding miRNAs. Overall, this study has identified
several miRNAs that may be associated with tumorgenesis
in direct response from proton radiation. Further validation
and characterization studies on these altered miRNAs
could be helpful in elucidating the cellular and genetic
changes induced by proton irradiation, and in understand-
ing possible links between proton irradiation and cancer.
Materials and methods
Samples and tissues
Balb/C male mice were purchased from Harlan Laboratories
Inc. (Livermore, USA). The mice were irradiated with
protons after proper resting for 2 days at the Loma Linda
University Radiation Facility (Loma Linda, CA). Mice in
Group 1 served as a control (0 Gy), while mice in Group 2
were exposed (2 Gy) to a proton source at a dose rate of
1Gev/45 s. The controls and irradiated mice were eutha-
nized 4 h after exposure; brain, liver, and testis tissues were
then dissected out and immediately frozen in liquid nitrogen.
The animal care and treatment procedures were reviewed
and approved by the Institutional Animal Care and Use
Committees (IACUC) at the Loma Linda radiation facility.
RNA isolation and small RNA library preparation
for microRNAs
Total RNA was extracted from tissues (brain, liver, and
testis) using Trizol reagent (Invitrogen; Carlsbad, CA) per
manufacturer’s instructions. After passing quality controls
(quantification and quality check by Agilent 2100 Bioan-
alyzer), 1 lg of total RNA, without selection of small
RNAs, was used directly in Illumina TruSeq Small RNA
Sample Prep Kits (Catalog # RS-200-0012) to prepare
libraries for the three different tissues as well as controls.
Four biological replicates were used for brain and testis
tissue, while due to availability issue, only two biological
replicate were used for liver tissue. The quality of libraries
was determined on the Agilent 2100 Bioanalyzer (Agilent;
Palo Alto, CA) per manufacturer’s recommendation.
MicroRNAs sequencing
The bar-coded small RNA libraries for each irradiated and
non-irradiated mouse tissue (brain, liver, and testis)
including biological replicates were pooled together in
equal concentrations in one pool for sequencing analysis.
The pooled libraries were subjected to pair-end (2 9 50
232 Mol Cell Biochem (2013) 382:225–235
123

bp) sequencing using two different lanes on the HiSeq
2000 (Illumina, Inc) to analyze differential miRNAs
expression patterns.
Mapping and differential expression of microRNAs
sequencing data
The miRNA expression data was generated from two
Illumina HiSeq runs merged together to account for the
lack of counts identified from a single HiSeq run. For each
run, the paired end reads were first merged using SeqPrep
(https://github.com/jstjohn/SeqPrep) and then cleaned up
by removing any TruSeq adapters identified. After the
reads were merged, the data was filtered such that only the
size range for known mouse miRNAs from miRBase
revision 17 [87] remained. Bowtie [88] with miRBase v17
was run with no mismatches to identify mature miRNA
counts from the Illumina HiSeq data. Counts from both of
the runs were added together. Statistically significant
miRNAs for each tissue were identified using DESeq [89]
development version (R-devel build 09/01/2011) by
selecting for an unadjusted p value \0.05.
MicroRNA editing analysis
Possible RNA editing events were identified by looking for
substituted bases in Bowtie aligned reads. Mature miRNAs
from miRBase (revision 17, http://www.mirbase.org) were
used as a reference and a single substitution was allowed
during alignment. Observed substitution rates were gener-
ated separately for control (0 Gy) and proton-irradiated
(2 Gy) tissues by dividing the number reads aligning to
miRNAs which contained one substitution by the total
number of aligned reads (i.e. reads containing zero or one
substitutions).
Acknowledgments This study was supported in part by grants from
the National Aeronautics and Space Administration Cooperative
Agreements NCC9-165 and NNX08BA47A, National Institutes of
Health [P01-HG000205], the National Science Foundation [DBI
0830141].
References
1. Simone NL, Soule BP, Ly D, Saleh AD, Savage JE, Degraff W et al
(2009) Ionizing radiation-induced oxidative stress alters miRNA
expression. PLoS One 4:e6377. doi:10.1371/journal.pone.0006377
2. Sorokina S, Markova E, Gursky J, Dobrovodsky J, Belyaev I
(2013) Relative biological efficiency of protons at low and
therapeutic doses in induction of 53BP1/c-H2AX foci in lym-
phocytes from umbilical cord blood. Int J Radiat Biol. May 22.
[Epub ahead of print]
3. Elmore E, Lao XY, Kapadia R, Swete M, Redpath JL (2011)
Neoplastic transformation in vitro by mixed beams of high-energy
iron ions and protons. Radiat Res. 176(3):291–302 Epub 2011 Jul 6
4. Friedman RC, Farh KK, Burge CB, Bartel DP (2009) Most
mammalian mRNAs are conserved targets of microRNAs. Gen-
ome Res 19:92–105. doi:10.1101/gr.082701.108
5. Dickey JS, Zemp FJ, Martin OA, Kovalchuk O (2011) The role of
miRNA in the direct and indirect effects of ionizing radiation.
Radiat Environ Biophys 50:491–499. doi:10.1007/s00411-011-
0386-5
6. Ilnytskyy Y, Koturbash I, Kovalchuk O (2009) Radiation-induced
bystander effects in vivo are epigenetically regulated in a tissue-specific
manner. Environ Mol Mutagen 50:105–113. doi:10.1002/em.20440
7. Maes OC, An J, Sarojini H, Wu H, Wang E (2008) Changes in
microRNA expression patterns in human fibroblasts after low-LET
radiation. J Cell Biochem 105:824–834. doi:10.1002/jcb.21878
8. Cui W, Ma J, Wang Y, Biswal S (2011) Plasma miRNA as bio-
markers for assessment of total-body radiation exposure dosime-
try. PLoS One 6:e22988. doi:10.1371/journal.pone.0022988
9. Ng WL, Yan D, Zhang X, Mo YY, Wang Y (2010) Over-
expression of miR-100 is responsible for the low-expression of
ATM in the human glioma cell line: M059J. DNA Repair (Amst)
9:1170–1175. doi:10.1016/j.dnarep.2010.08.007
10. Joyce CE, Zhou X, Xia J, Ryan C, Thrash B, Menter A, Zhang W
et al (2011) Deep sequencing of small RNAs from human skin
reveals major alterations in the psoriasis miRNAome. Hum Mol
Genet 20:4025–4040. doi:10.1093/hmg/ddr331
11. Josson S, Sung SY, Lao K, Chung LW, Johnstone PA (2008)
Radiation modulation of microRNA in prostate cancer cell lines.
Prostate 68:1599–1606. doi:10.1002/pros.20827
12. Zhang H, Luo XQ, Zhang P, Huang LB, Zheng YS, Wu J, Zhou H
et al (2009) MicroRNA patterns associated with clinical prog-
nostic parameters and CNS relapse prediction in pediatric acute
leukemia. PLoS One 4:e7826. doi:10.1371/journal.pone.0007826
13. Catto JW, Miah S, Owen HC, Bryant H, Myers K, Dudziec E et al
(2009) Distinct microRNA alterations characterize high and low-
grade bladder cancer. Cancer Res 69:8472–8481. doi:10.1158/
0008-5472.CAN-09-0744
14. Henson BJ, Bhattacharjee S, O’Dee DM, Feingold E, Gollin SM
(2009) Decreased expression of miR-125b and miR-100 in oral
cancer cells contributes to malignancy. Genes Chromosomes
Cancer 48:569–582. doi:10.1002/gcc.20666
15. Yang H, Kong W, He L, Zhao JJ, O’Donnell JD, Wang J et al (2008)
MicroRNA expression profiling in human ovarian cancer: miR-214
induces cell survival and cisplatin resistance by targeting PTEN.
Cancer Res 68:425–433. doi:10.1158/0008-5472.CAN-07-2488
16. Thorgeirsson SS (2011) The almighty MYC: orchestrating the
micro-RNA universe to generate aggressive liver cancer. J Hep-
atol 55:486–487. doi:10.1016/j.jhep.2011.01.042
17. Jiang J, Lee EJ, Gusev Y, Schmittgen TD (2005) Real-time
expression profiling of microRNA precursors in human cancer cell
lines. Nucleic Acids Res 33:5394–5403. doi:10.1093/nar/gki863
18. Yu J, Ryan DG, Getsios S, Oliveira-Fernandes M, Fatima A,
Lavker RM (2008) MicroRNA-184 antagonizes microRNA-205
to maintain SHIP2 levels in epithelia. Proc Natl Acad Sci USA
105:19300–19305. doi:10.1073/pnas.0803992105
19. Fletcher AM, Heaford AC, Trask DK (2008) Detection of met-
astatic head and neck squamous cell carcinoma using the relative
expression of tissue-specific mir-205. Transl Oncol 1:202–208
20. Childs G, Fazzari M, Kung G, Kawachi N, Brandwein-Gensler
M, McLemore M et al (2009) Low-level expression of microR-
NAs let-7d and miR-205 are prognostic markers of head and neck
squamous cell carcinoma. Am J Pathol 174:736–745. doi:10.2353/
ajpath.2009.080731
21. Gottardo F, Liu CG, Ferracin M, Calin GA, Fassan M, Bassi P
et al (2007) Micro-RNA profiling in kidney and bladder cancers.
Urol Oncol 25:387–392. doi:10.1016/j.urolonc.2007.01.019
22. Wang X, Tang S, Le SY, Lu R, Rader JS, Meyers C et al (2008)
Aberrant expression of oncogenic and tumor-suppressive microRNAs
Mol Cell Biochem (2013) 382:225–235 233
123

in cervical cancer is required for cancer cell growth. PLoS One
3:e2557. doi:10.1371/journal.pone.0002557
23. Markou A, Tsaroucha EG, Kaklamanis L, Fotinou M, Georgou-
lias V, Lianidou ES (2008) Prognostic value of mature microR-
NA-21 and microRNA-205 overexpression in non-small cell lung
cancer by quantitative real-time RT-PCR. Clin Chem 54:
1696–1704. doi:10.1373/clinchem.2007.101741
24. Wszolek MF, Rieger-Christ KM, Kenney PA, Gould JJ, Silva
NB, Lavoie AK et al (2011) A MicroRNA expression profile
defining the invasive bladder tumor phenotype. Urol Oncol 29:
794–801. doi:10.1016/j.urolonc.2009.08.024
25. Schaefer A, Jung M, Mollenkopf HJ, Wagner I, Stephan C,
Jentzmik F et al (2010) Diagnostic and prognostic implications of
microRNA profiling in prostate carcinoma. Int J Cancer 126:
1166–1176. doi:10.1002/ijc.24827
26. Blenkiron C, Goldstein LD, Thorne NP, Spiteri I, Chin SF,
Dunning MJ et al (2007) MicroRNA expression profiling of
human breast cancer identifies new markers of tumor subtype.
Genome Biol 8:R214. doi:10.1186/gb-2007-8-10-r214
27. Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S et al
(2005) MicroRNA gene expression deregulation in human breast
cancer. Cancer Res 65:7065–7070. doi:10.1158/0008-5472.CAN-
05-1783
28. Mattie MD, Benz CC, Bowers J, Sensinger K, Wong L, Scott GK et al
(2005) Optimized high-throughput microRNA expression profiling
provides novel biomarker assessment of clinical prostate and breast
cancer biopsies. Mol Cancer 5:24. doi:10.1186/1476-4598-5-24
29. Gao W, Shen H, Liu L, Xu J, Xu J, Shu Y (2011) MiR-21
overexpression in human primary squamous cell lung carcinoma
is associated with poor patient prognosis. J Cancer Res Clin
Oncol 137:557–566. doi:10.1007/s00432-010-0918-4
30. Lee ST, Chu K, Im WS, Yoon HJ, Im JY, Park JE et al (2011)
Altered microRNA regulation in Huntington’s disease models.
Exp Neurol 227:172–179. doi:10.1016/j.expneurol.2010.10.012
31. Wagner-Ecker M, Schwager C, Wirkner U, Abdollahi A, Huber PE
(2010) MicroRNA expression after ionizing radiation in human
endothelial cells. Radiat Oncol 5:25. doi:10.1186/1748-717X-5-25
32. Nikiforova MN, Gandhi M, Kelly L, Nikiforov YE (2011)
MicroRNA dysregulation in human thyroid cells following
exposure to ionizing radiation. Thyroid 21:261–266. doi:10.1089/
thy.2010.0376
33. Weidhaas JB, Babar I, Nallur SM, Trang P, Roush S, Boehm M et al
(2007) MicroRNAs as potential agents to alter resistance to cytotoxic
anticancer therapy. Cancer Res 67:11111–11116. doi:10.1158/
0008-5472.CAN-07-2858
34. Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F
et al (2006) A microRNA expression signature of human solid
tumors defines cancer gene targets. Proc Natl Acad Sci USA
103:2257–2261. doi:10.1073/pnas.0510565103
35. Ozen M, Creighton CJ, Ozdemir M, Ittmann M (2008) Wide-
spread deregulation of microRNA expression in human prostate
cancer. Oncogene 27:1788–1793. doi:10.1038/sj.onc.1210809
36. Porkka KP, Pfeiffer MJ, Waltering KK, Vessella RL, Tammela TL,
Visakorpi T (2007) MicroRNA expression profiling in prostate
cancer. Cancer Res 67:6130–6135. doi:10.1158/0008-5472.CAN-
07-0533
37. Ali S, Almhanna K, Chen W, Philip PA, Sarkar FH (2010) Differ-
entially expressed miRNAs in the plasma may provide a molecular
signature for aggressive pancreatic cancer. Am J Transl Res 3:28–47
38. Hui AB, Lenarduzzi M, Krushel T, Waldron L, Pintilie M, Shi W
et al (2010) Comprehensive MicroRNA profiling for head and
neck squamous cell carcinomas. Clin Cancer Res 16:1129–1139.
doi:10.1158/1078-0432.CCR-09-2166
39. Farazi TA, Horlings HM, Ten Hoeve JJ, Mihailovic A, Halfwerk
H, Morozov P et al (2011) MicroRNA sequence and expression
analysis in breast tumors by deep sequencing. Cancer Res
71:4443–4453. doi:10.1158/0008-5472.CAN-11-0608
40. Guo J, Dong Q, Fang Z, Chen X, Lu H, Wang K et al (2010)
Identification of miRNAs that are associated with tumor metas-
tasis in neuroblastoma. Cancer Biol Ther 9:446–452
41. Lin J, Huang S, Wu S, Ding J, Zhao Y, Liang L et al (2011)
MicroRNA-423 promotes cell growth and regulates G(1)/S
transition by targeting p21Cip1/Waf1 in hepatocellular carci-
noma. Carcinogenesis 32:1641–1647. doi:10.1093/carcin/bgr199
42. Wang R, Wang ZX, Yang JS, Pan X, De W, Chen LB (2011)
MicroRNA-451 functions as a tumor suppressor in human non-
small cell lung cancer by targeting ras-related protein 14
(RAB14). Oncogene 30:2644–2658. doi:10.1038/onc.2010.642
43. Bandres E, Bitarte N, Arias F, Agorreta J, Fortes P, Agirre X et al
(2009) MicroRNA-451 regulates macrophage migration inhibitory
factor production and proliferation of gastrointestinal cancer cells. Clin
Cancer Res 15:2281–2290. doi:10.1158/1078-0432.CCR-08-1818
44. Ju X, Li D, Shi Q, Hou H, Sun N, Shen B (2009) Differential
microRNA expression in childhood B-cell precursor acute lym-
phoblastic leukemia. Pediatr Hematol Oncol 26:1–10. doi:10.1080/
08880010802378338
45. Chen YT, Kitabayashi N, Zhou XK, Fahey TJ III, Scognamiglio
T (2008) MicroRNA analysis as a potential diagnostic tool for
papillary thyroid carcinoma. Mod Pathol 21:1139–1146. doi:
10.1038/modpathol.2008.105
46. Pallante P, Visone R, Ferracin M, Ferraro A, Berlingieri MT,
Troncone G et al (2006) MicroRNA deregulation in human thy-
roid papillary carcinomas. Endocr Relat Cancer 13:497–508. doi:
10.1677/erc.1.01209
47. Nikiforova MN, Chiosea SI, Nikiforov YE (2009) MicroRNA
expression profiles in thyroid tumors. Endocr Pathol 20:85–91.
doi:10.1007/s12022-009-9069-z
48. Chou CK, Chen RF, Chou FF, Chang HW, Chen YJ, Lee YF et al
(2010) MiR-146b is highly expressed in adult papillary thyroid
carcinomas with high risk features including extrathyroidal
invasion and the BRAF(V600E) mutation. Thyroid 20:489–494.
doi:10.1089/thy.2009.0027
49. Mazeh H, Mizrahi I, Halle D, Ilyayev N, Stojadinovic A, Trink B
et al (2011) Development of a microRNA-based molecular assay
for the detection of papillary thyroid carcinoma in aspiration
biopsy samples. Thyroid 21:111–118. doi:10.1089/thy.2010.0356
50. Garbacki N, Di VE, Huynh-Thu VA, Geurts P, Irrthum A, Crahay
C et al (2011) MicroRNAs profiling in murine models of acute
and chronic asthma: a relationship with mRNAs targets. PLoS
One 6:e16509. doi:10.1371/journal.pone.0016509
51. Wang Y, Huang JW, Li M, Cavenee WK, Mitchell PS, Zhou X
et al (2011) MicroRNA-138 modulates DNA damage response by
repressing histone H2AX expression. Mol Cancer Res 9:
1100–1111. doi:10.1158/1541-7786.MCR-11-0007
52. Mitomo S, Maesawa C, Ogasawara S, Iwaya T, Shibazaki M,
Yashima-Abo A et al (2008) Downregulation of miR-138 is asso-
ciated with overexpression of human telomerase reverse trans-
criptase protein in human anaplastic thyroid carcinoma cell lines.
Cancer Sci 99:280–286. doi:10.1111/j.1349-7006.2007.00666.x
53. Liu X, Jiang L, Wang A, Yu J, Shi F, Zhou X (2009) MicroRNA-
138 suppresses invasion and promotes apoptosis in head and neck
squamous cell carcinoma cell lines. Cancer Lett 286:217–222.
doi:10.1016/j.canlet.2009.05.030
54. Jiang L, Liu X, Kolokythas A, Yu J, Wang A, Heidbreder CE
et al (2010) Downregulation of the Rho GTPase signaling path-
way is involved in the microRNA-138-mediated inhibition of cell
migration and invasion in tongue squamous cell carcinoma. Int J
Cancer 127:505–512. doi:10.1002/ijc.25320
55. Hisaoka M, Matsuyama A, Nagao Y, Luan L, Kuroda T, Akiy-
ama H et al (2011) Identification of altered MicroRNA expression
234 Mol Cell Biochem (2013) 382:225–235
123

patterns in synovial sarcoma. Genes Chromosomes Cancer
50:137–145. doi:10.1002/gcc.20837
56. Jukic DM, Rao UN, Kelly L, Skaf JS, Drogowski LM, Kirkwood
JM et al (2010) Microrna profiling analysis of differences
between the melanoma of young adults and older adults. J Transl
Med 8:27. doi:10.1186/1479-5876-8-27
57. Lages E, Guttin A, El AM, Ramus C, Ipas H, Dupre I, Rolland D
et al (2011) MicroRNA and target protein patterns reveal phys-
iopathological features of glioma subtypes. PLoS One 6:e20600.
doi:10.1371/journal.pone.0020600
58. Urdinguio RG, Fernandez AF, Lopez-Nieva P, Rossi S, Huertas
D, Kulis M et al (2010) Disrupted microRNA expression caused
by Mecp2 loss in a mouse model of Rett syndrome. Epigenetics
5:656–663. doi:10.4161/epi.5.7.13055
59. Baluchamy S, Zhang Y, Ravichandran P, Ramesh V, Sodipe A,
Hall JC et al (2010) Differential oxidative stress gene expression
profile in mouse brain after proton exposure. In Vitro Cell Dev
Biol Anim 46:718–725. doi:10.1007/s11626-010-9330-2
60. Shea MA, Smart DF (1994) Significant proton events of solar
cycle 22 and a comparison with events of previous solar cycles.
Adv Space Res 14:631–638
61. Smart DF, Shea MA (2002) A review of solar proton events
during the 22nd solar cycle. Adv Space Res 30:1033–1044
62. Koturbash I, Zemp F, Kolb B, Kovalchuk O (2011) Sex-specific
radiation-induced microRNAome responses in the hippocampus,
cerebellum and frontal cortex in a mouse model. Mutat Res
722:114–118. doi:10.1016/j.mrgentox.2010.05.007
63. Tamminga J, Kathiria P, Koturbash I, Kovalchuk O (2008) DNA
damage-induced upregulation of miR-709 in the germline
downregulates BORIS to counteract aberrant DNA hypomethy-
lation. Cell Cycle 7:3731–3736
64. Ozata DM, Caramuta S, Velazquez-Fernandez D, Akcakaya P,
Xie H, Hoog A et al (2011) The role of microRNA deregulation
in the pathogenesis of adrenocortical carcinoma. Endocr Relat
Cancer 18:643–655. doi:10.1530/ERC-11-0082
65. Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang Y et al (2011) Analysis
of MiR-195 and MiR-497 expression, regulation and role in breast
cancer. Clin Cancer Res 17:1722–1730. doi:10.1158/1078-0432.
CCR-10-1800
66. Lehmann U, Streichert T, Otto B, Albat C, Hasemeier B,
Christgen H et al (2010) Identification of differentially expressed
microRNAs in human male breast cancer. BMC Cancer 10:109.
doi:10.1186/1471-2407-10-109
67. Guo J, Miao Y, Xiao B, Huan R, Jiang Z, Meng D et al (2009)
Differential expression of microRNA species in human gastric
cancer versus non-tumorous tissues. J Gastroenterol Hepatol
24:652–657. doi:10.1111/j.1440-1746.2008.05666.x
68. Yan LX, Huang XF, Shao Q, Huang MY, Deng L, Wu QL et al
(2008) MicroRNA miR-21 overexpression in human breast can-
cer is associated with advanced clinical stage, lymph node
metastasis and patient poor prognosis. RNA 14:2348–2360. doi:
10.1261/rna.1034808
69. Zhu W, Zhu D, Lu S, Wang T, Wang J, Jiang B et al (2012) MiR-
497 modulates multidrug resistance of human cancer cell lines by
targeting BCL2. Med Oncol 29:384–391. doi:10.1007/s12032-
010-9797-4
70. Wu Q, Jin H, Yang Z, Luo G, Lu Y, Li K et al (2010) MiR-150
promotes gastric cancer proliferation by negatively regulating the
pro-apoptotic gene EGR2. Biochem Biophys Res Commun
392:340–345. doi:10.1016/j.bbrc.2009.12.182
71. Vasilatou D, Papageorgiou S, Pappa V, Papageorgiou E, Derve-
noulas J (2010) The role of microRNAs in normal and malignant
hematopoiesis. Eur J Haematol 84:1–16. doi:10.1111/j.1600-0609.
2009.01348.x
72. Magrelli A, Azzalin G, Salvatore M, Viganotti M, Tosto F,
Colombo T et al (2009) Altered microRNA Expression Patterns
in Hepatoblastoma Patients. Transl Oncol 2:157–163
73. Lulla RR, Costa FF, Bischof JM, Chou PM, de F Vanin M et al
(2011) Identification of Differentially Expressed MicroRNAs in
Osteosarcoma. Sarcoma 732690. doi:10.1155/2011/732690
74. Miko E, Czimmerer Z, Csanky E, Boros G, Buslig J, Dezso B et al
(2009) Differentially expressed microRNAs in small cell lung can-
cer. Exp Lung Res 35:646–664. doi:10.3109/01902140902822312
75. Watanabe A, Tagawa H, Yamashita J, Teshima K, Nara M,
Iwamoto K et al (2011) The role of microRNA-150 as a tumor
suppressor in malignant lymphoma. Leukemia 25:1324–1334.
doi:10.1038/leu.2011.81
76. Srivastava SK, Bhardwaj A, Singh S, Arora S, Wang B, Grizzle
WE et al (2011) MicroRNA-150 directly targets MUC4 and
suppresses growth and malignant behavior of pancreatic cancer
cells. Carcinogenesis 32:1832–1839. doi:10.1093/carcin/bgr223
77. Lee JW, Choi CH, Choi JJ, Park YA, Kim SJ, Hwang SY et al
(2008) Altered MicroRNA expression in cervical carcinomas. Clin
Cancer Res 14:2535–2542. doi:10.1158/1078-0432.CCR-07-1231
78. Leucci E, Onnis A, Cocco M, De FG, Imperatore F, Giuseppina A
et al (2010) B-cell differentiation in EBV-positive Burkitt lym-
phoma is impaired at posttranscriptional level by miRNA-altered
expression. Int J Cancer 126:1316–1326. doi:10.1002/ijc.24655
79. Tryndyak VP, Ross SA, Beland FA, Pogribny IP (2009) Down-
regulation of the microRNAs miR-34a, miR-127, and miR-200b
in rat liver during hepatocarcinogenesis induced by a methyl-
deficient diet. Mol Carcinog 48:479–487. doi:10.1002/mc.20484
80. Zhu M, Yi M, Kim CH, Deng C, Li Y, Medina D et al (2011)
Integrated miRNA and mRNA expression profiling of mouse
mammary tumor models identifies miRNA signatures associated
with mammary tumor lineage. Genome Biol 12:R77. doi:10.1186/
gb-2011-12-8-r77
81. Ma L, Teruya-Feldstein J, Weinberg RA (2007) Tumour invasion
and metastasis initiated by microRNA-10b in breast cancer.
Nature 449:682–688. doi:10.1038/nature06174
82. Heale BS, Keegan LP, McGurk L, Michlewski G, Brindle J,
Stanton CM et al (2009) Editing independent effects of ADARs
on the miRNA/siRNA pathways. EMBO J 28:3145–3156. doi:
10.1038/emboj.2009.244
83. Kawahara Y, Zinshteyn B, Sethupathy P, Iizasa H, Hatzigeorgiou
AG, Nishikura K (2007) Redirection of silencing targets by
adenosine-to-inosine editing of miRNAs. Science 315:1137–1140.
doi:10.1126/science.1138050
84. Bartel DP (2009) MicroRNAs: target recognition and regulatory
functions. Cell 136:215–233. doi:10.1016/j.cell.2009.01.002
85. Gommans WM (2012) A-to-I editing of microRNAs: regulating the
regulators? Semin Cell Dev Biol 23:251–257. doi:10.1016/j.
semcdb.2011.09.018
86. Yang W, Chendrimada TP, Wang Q, Higuchi M, Seeburg PH,
Shiekhattar R et al (2006) Modulation of microRNA processing
and expression through RNA editing by ADAR deaminases. Nat
Struct Mol Biol 13:13–21. doi:10.1038/nsmb1041
87. Kozomara A, Griffiths-Jones S (2011) miRBase: integrating
microRNA annotation and deep-sequencing data. Nucleic Acids
Res 39:D152–D157. doi:10.1093/nar/gkq1027
88. Langmead B, Trapnell C, Pop M, Salzberg SL (2009) Ultrafast
and memory-efficient alignment of short DNA sequences to the
human genome. Genome Biol 10:R25. doi:10.1186/gb-2009-
10-3-r25
89. Anders S, Huber W (2010) Differential expression analysis for
sequence count data. Genome Biol 11:R106. doi:10.1186/gb-2010-
11-10-r106
Mol Cell Biochem (2013) 382:225–235 235
123
Top Related
Copyright © 2022 FDOKUMEN

