







Bahasa
Halaman
Hukum


+ model ARTICLE IN PRESS
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
RECTED PROOFDiscovery of immune-related genes expressed in hemocytes
of the tarantula spider Acanthoscurria gomesiana
Daniel M. Lorenzini a, Pedro I. da Silva Jr b, Marcelo B. Soares c, Paulo Arruda d,1,
Joao Setubal e,2, Sirlei Daffre a,*
a Departamento de Parasitologia, Instituto de Ciencias Biomedicas, Universidade de Sao Paulo, Avenue Prof. Lineu Prestes,
1374, CEP 05508-900 Sao Paulo, SP, Brazilb Laboratorio de Artropodes, Instituto Butantan. CEP 05503-900 Sao Paulo, SP, Brazil
c Department of Pediatrics and Biochemistry, University of Iowa, Iowa City, IA 52242, USAd Instituto da Computacao, Centro de Biologia Molecular e Engenharia Genetica, Universidade Estadual de Campinas,
CEP 13083-970 Campinas, SP, Brazile Laboratorio de Bioinformatica, Instituto da Computacao, Universidade Estadual de Campinas, CEP 13083-970 Campinas, SP, Brazil
Received 5 July 2005; revised 28 August 2005; accepted 2 September 2005
Abstract
The present study reports the identification of immune related transcripts from hemocytes of the spider Acanthoscurria
gomesiana by high throughput sequencing of expressed sequence tags (ESTs). To generate ESTs from hemocytes, two cDNA
libraries were prepared: one by directional cloning (primary) and the other by the normalization of the first (normalized). A total of
7584 clones were sequenced and the identical ESTs were clustered, resulting in 3723 assembled sequences (AS). At least 20% of
these sequences are putative novel genes. The automatic functional annotation of AS based on Gene Ontology revealed several
abundant transcripts related to the following functional classes: hemocyanin, lectin, and structural constituents of ribosome and
cytoskeleton. From this annotation, 73 transcripts possibly involved in immune response were also identified, suggesting the
existence of several molecular processes not previously described for spiders, such as: pathogen recognition, coagulation,
complement activation, cell adhesion and intracellular signaling pathway for the activation of cellular defenses.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: Hemocytes; Innate immunity; Expressed sequence tags (ESTs); Hemocyanin; Coagulation; Lectins; Serine-proteases; Antimicrobial
peptides
UNCOR
0145-305X/$ - see front matter q 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.dci.2005.09.001
* Corresponding author. Tel.: C55 11 309 17272; fax: C55 11 309
17417.
E-mail address: [email protected] (S. Daffre).1 Alellyx Applied Genomics, CEP 13067-850, Campinas, SP,
Brazil.2 Virginia Bioinformatics Institute, Virginia Polytechnic Institute
and State University, Bioinformatics 1, Box 0477, Blacksburg, VA
24060, USA.
DCI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1
91
92
93
94
95
96
97
98
99
100
101
102
1. Introduction
Invertebrates developed an efficient immune system
to control infections. The hemolymph circulating cells,
named hemocytes, are essential components of this
system that play two important roles. One is the cellular
activity that causes the phagocytosis and/or encapsula-
tion of the pathogens [1]. The second, but not less
important, is the production of peptides and proteins
that participate in the immune response, as seen in the
following examples. The hemocytes of horseshoe crabs
Developmental and Comparative Immunology xx (xxxx) 1–12
www.elsevier.com/locate/devcompimm
–12
103
104

C
D
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–122
+ model ARTICLE IN PRESS
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
UNCORRE
store clotting factors, proteinase inhibitors and lectins
[2,3]. The insects and crustaceans’ hemocytes produce
phenoloxidase [4]. Antimicrobial peptides have been
isolated from hemocytes of animals that belong to
different phyla of invertebrates [5]. The constitutive
production and storage of these peptides and proteins in
the hemocytes keeps invertebrates ready to fight
invading pathogens.
To investigate the immune response of spiders,
initially, we worked in the characterization of anti-
microbial molecules. Two peptides (gomesin and
acanthoscurrin) and one acylpolyamine (mygalin)
with antimicrobial activity have been isolated from
the hemocytes of mygalomorph spider A. gomesiana.
The use of mygalomorph spiders as an experimental
model is very useful, because they are on of the oldest
species in the order Araneae. Gomesin is a cationic
peptide of 18 amino acids and two disulfide bridges,
produced from a precursor containing a signal peptide
and a anionic segment on the C-terminus, which is
constitutively synthesized in the hemocytes and stored
in their granules [6,7]. Acanthoscurrin is a glycine-rich
peptide, which is post-translationally processed by the
removal of the signal peptide and C-terminal amida-
tion. Acanthoscurrin is released into the cell free
hemolymph following immune challenge [8]. Mygalin
is a bis-acylpolyamine N1,N8-bis(2,5-dihydroxylben-
zoil)spermidine active against E. coli, and its activity is
inhibited by catalase [9].
The discovery of novel invertebrate genes related to
the immune response has been accelerated by high
throughput sequencing techniques combined with
searches for homologous sequences on public data-
bases. The sequencing of expressed sequence tags
(ESTs) is specially useful, since it allows simul-
taneously the novel gene discovery and gene expression
analysis. The sequencing of ESTs has been done with
hemocytes of shrimps [10,11], mollusks [12,13] and
insects [14], resulting in the identification of several
immune related genes. In the mosquito Anopheles
gambiae, the EST clones were used to prepare a cDNA
microarray, and the modulation of immune genes’
expression was analyzed under different immune
stimuli [15]. In this context, exploring novel genes on
invertebrates such as spiders, which are phylogeneti-
cally distant from the organisms mentioned above,
would produce meaningful information that can support
the search for immune features that are conserved
throughout the invertebrates.
This study reports the production of ESTs from the
hemocytes of the spider A. gomesiana obtained through
two cDNA libraries, one prepared by directional
CI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1–
TED PROOF
cloning (primary) and the other by the normalization
of the first (normalized). The random selection of
clones from the primary cDNA library resulted in the
identification of transcripts abundantly expressed in the
hemocytes, while the normalized library produced a
large number of unique sequences. In addition, several
of these sequences are related to invertebrate immune
response.
2. Material and methods
2.1. Spiders
A. gomesiana is a tarantula spider of the Therapho-
sidae family that is distributed in southeast Brazil.
Adults from this species are medium-sized (approxi-
mately 5 cm in length) and can live over 23 years. The
animals used in the experiments were adults of both
sexes and in the intermolt stage. These spiders were not
reared in the laboratory, but donated to the Arthropods
Laboratory of the Butantan Institute (Sao Paulo, Brazil)
by citizens of Sao Paulo and neighbor towns, where
they were kept.
2.2. Hemocyte isolation and RNA extraction
Hemolymph was collected from 15 spiders (approxi-
mately 0.5 ml/animal) as previously described [6].
Hemocytes were separated from cell free hemolymph
by centrifugation at 800!g for 10 min at 4 8C before
RNA extraction. Total RNA was isolated from
hemocytes using Trizol reagent (Gibco/BRL).
2.3. Construction of cDNA Libraries
2.3.1. Primary
One directionally cloned cDNA library was pre-
pared as described previously [16]. The mRNA (1 mg),
purified from total RNA (300 mg) with an oligo-(dT)
column, was annealed with 2-fold mass excess of a
NotI-(dT)18 primer and reverse transcribed with
Superscript Reverse Transcriptase (Life Sciences).
Following the second strand synthesis, the double-
stranded cDNAs longer than 350 bp were size selected
by gel filtration on a Bio-Gel A-50M column (Bio-
Rad), joined to a 500- to 1000-fold molar excess of
EcoRI adapter, digested with Not I, and size selected
over a second Bio-Gel column to remove the excess of
EcoRI adapter. The selected cDNAs were cloned into
the EcoRI and NotI sites of the pT7T3-Pac phagemid
vector and electroporated into E. coli DH10B host cells
(Invitrogen). The primary library plasmid DNA was
12

C
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–12 3
+ model ARTICLE IN PRESS
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
UNCORRE
purified using Qiagen-tip 100 (Qiagen) and stored at
K70 8C.
2.3.2. Normalized
The normalized library was prepared following the
method 4 described in [16]. The primary library
plasmid DNA was electroporated into E. coli
DH5aF 0, infected with the helper phage M13K07
(Pharmacia) and harvested to prepare the single-
stranded plasmids. An aliquot of the single-stranded
plasmids was amplified by PCR to produce the cDNA
inserts, which were hybridized in a 20-fold excess with
the single-stranded plasmids. Following hybridization
at a relatively low Cot (y5), the remaining single-
stranded circles (normalized library) were purified on a
hydroxyapatite (HAP) column, converted to double-
stranded circles by primer extension and electroporated
into E. coli DH10B host cells (Invitrogen). The
normalized library plasmid DNA was purified and
stored as described above.
2.4. Template preparation and DNA sequencing
E. coli DH10B host cells (Invitrogen) were
electroporated with plasmid DNA from the cDNA
libraries and spread on LB agar plates containing
60 mg/ml of ampicillin. Random selected colonies were
grown on Circle Grow medium (Qbiogene) containing
ampicillin at 100 mg/ml for 22 h at 37 8C. Plasmid DNA
from the transformed bacteria was prepared in 96-well
plates using a modified alkaline lysis method (http://
sucest.lbi.ic.unicamp.br/public/protocols.html). The 5 0
end of cDNA inserts was sequenced on an automatic
DNA sequencer ‘ABI 3100’ (Applied Biosystems)
using T3 primer and ABIe Big Dye terminator kit
(Applied Biosystems).
2.5. Sequence analysis
2.5.1. Sequence trimming and contaminant discarding
The chromatograms from sequenced clones were
automatically processed for base calling and low
quality trimming using Phred set to minimum quality
10 (non-default parameters: -trim_alt -trim_cutoff 0.09)
. Vector sequence trimming was done by Crossmatch
with the pT7T3-Pac sequence (non-default parameters:
-minmatch 10 -minscore 20) and contaminant
sequences were identified by BlastN set to e-value
cutoff 1!10K30 (non-default parameters: Ke 1!10K
30), using a database of possible contaminants
(ribosomal RNA, E. coli genome, mitocondrial and
plasmid sequences, all from Genbank). Sequences
DCI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1
containing more than 200 bp after trimming (quality
and vector) and not identified as contaminants were
considered as high quality sequences.
2.5.2. Sequence assembly
High quality sequences from both libraries were
assembled by sequence similarity using CAP3 set to
minimum of 40 overlap and 95% identity (non-default
parameters: -o 40 -p 95). The longest ORF size of each
assemble sequence was accessed by Flip (http://
megasun.bch.umontreal.ca/ogmp/aboutflip.html).
TED PROOF
2.5.3. Functional annotation
The assembled sequences (AS) were submitted to
similarity searches (BlastX—e-value cutoff 1!10K6,
InterProScan) against public databases (nr-NCBI,
SwissprotCTREmbl, and Interpro). The GeneOntol-
ogy (www.geneontology.org) terms associated with
Interpro, Swissprot or TREmbl sequences found in the
similarity searches were automatically annotated to the
corresponding AS. Two additional protein databases
with GeneOntology associations were prepared with
immune related sequences from horseshoe crabs [17]
and Drosophila melanogaster [18], and these databases
were used for automatic functional annotation as
described above. The AS containing at least one EST
from the primary library were manually annotated,
when possible, with one GeneOntology entry for
molecular function ontology. The manual annotation
was based on the results of automatic annotations and
similarity searches on public databases Increasing
cDNA coverage. The longest clones of AS related to
immune system were sequenced from 5 0 and 3 0 ends
(using T3 and T7 primers, respectively) and manually
assembled together with corresponding ESTs, using the
software Seqman (Lasergene package, DNAStar, USA)
3. Results
For the discovery of genes expressed in the spider
hemocytes, 7584 ESTs were sequenced from primary
and normalized cDNA libraries (Table 1). After
sequence trimming and contaminant rejection, almost
90% of these ESTs (High Quality ESTs) had enough
information for sequence analysis. The low frequency
of ESTs rejected for the presence of contaminants
indicated the high quality of the cDNA libraries. The
elevated efficiency of the DNA sequencing is also
attested by the low frequency of sequences discarded
due to quality and the high average length of the
sequences (Table 1). The high quality ESTs were
–12

C
Table 1
Statistics of expressed sequence tags (EST) from cDNA libraries of
Acanthoscurria gomesiana hemocytes
Normalized Primary Total
Sequenced ESTs 5760 1824 7584
High quality ESTs 5156 1634 6790
Discarded ESTs 604 190 794
Average length of
sequencesa
524.7 651.0
Discarded reason
Low qualityb 540 100 640
Sizec 6 4 10
No insert 13 1 14
Contaminants
E. coli 7 1 8
Mitocondrial 1 1 2
Ribossomal 14 1 15
Vector 23 82 105
a Sequences length after quality trimming.b Sequences shorter than 200 bp with Phred quality above 10.c Sequences shorter than 200 bp after vector and low quality
trimming.
Table 2
Automatic functional annotation statistics of assembled sequences
(AS)
Database ASs with match on
Database
ASs annotated with
GO
NRa 2192 –
Swisprot C
TREmbl 2171 1937
Interpro 873 378
Imune Drosophila 106 106
Imune Lımulus 91 91
Total 2411 1971
a Protein sequence database from NCBI.
D
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–124
+ model ARTICLE IN PRESS
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
UNCORRE
deposited in the GenBank (accession codes
DR442119–DR448908).
The sequence assembly of the high quality ESTs
revealed 3723 assembled sequences (AS), which
correspond to the estimated number of transcripts
identified. From this total, 814 AS contain at least one
EST from the primary library (Primary Assembled
Sequences—PAS). When the ESTs of each library were
assembled separately, the 20 most abundant AS from
the primary library corresponded to 34% of the ESTs
sequenced from this library, while in the normalized
library only 6% of the ESTs sequenced from this library
were found in the 20 most abundant AS. This
demonstrates that the abundance of transcripts found
in the primary library was significantly reduced by the
normalization process. Consequently, the discovery of
new transcripts was more efficient in the normalized
library, as verified by the higher number of unique
sequences obtained from this library (819 primary,
1308 normalized) when equivalent number of ESTs
from each library were used on separate identical
sequence assemblies.
In order to assign function to all putative transcripts
obtained, the assembled sequences were submitted to
similarity searches with several public sequence
databases (Table 2). The number of AS with matching
sequences on NR and SwisprotCTREmbl was very
similar, and much higher than on Interpro. However,
213 AS had matches only with Interpro search.
Using the Gene Ontology (GO) information avail-
able for entries of SwissprotCTREmbl and Interpro
CI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1–
TED PROOF
databases, the assembled sequences were automatically
associated with GO terms. Most of the AS with matches
on these databases were associated with GO terms
(Table 2). Several AS were associated with more than
one term, making it difficult to analyze the distribution
of AS on GO terms. On the other hand, this annotation
was very useful for the manual annotation of PAS and
identification of immune related transcripts.
Each PAS was manually annotated with only one
term of the molecular function ontology (Gene
Ontology). In this way, 459 of the 814 PAS were
annotated. The number of primary library ESTs in each
PAS was used to evaluate the abundance of the
corresponding transcripts in the spider hemocytes, and
the GO annotation grouped these ESTs by function
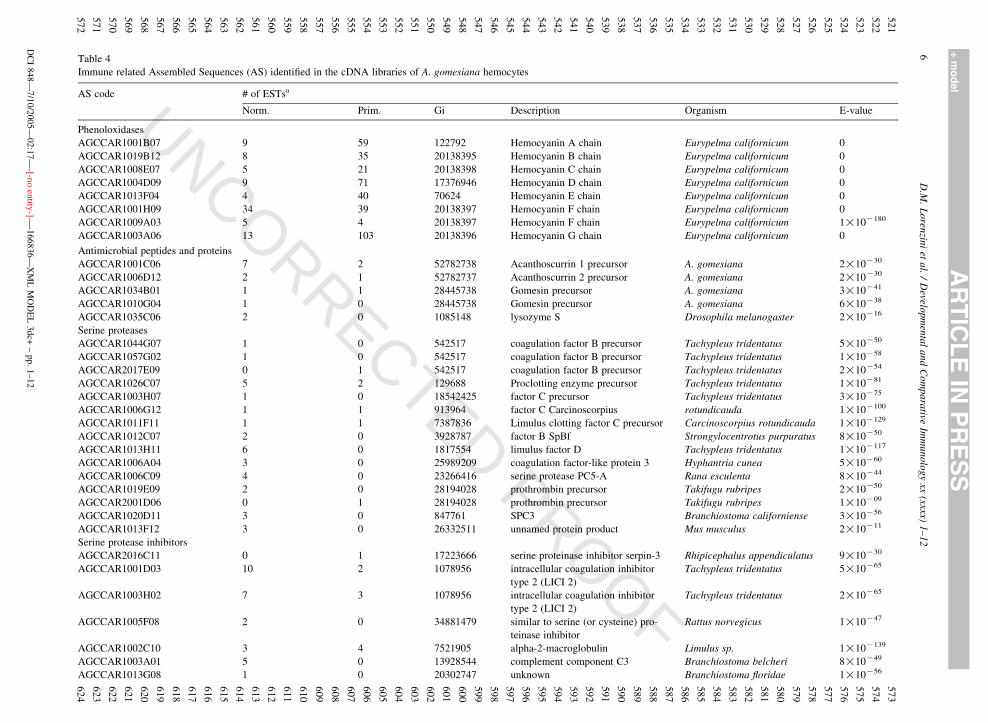
(Fig. 1). The functional class related to hemocyanins
has the highest number of ESTs. Components of
ribosome and cytoskeleton were also very abundant.
In the sugar binding (lectin) class, it was found one PAS
containing 27 ESTs from the primary library (AGC-
CAR1041B12, Table 4). There was also an elevated
number of ESTs related to transposable elements.
For the identification of immune related AS, a list of
functional classes was prepared from literature dedi-
cated to this issue (Table 3). The AS annotated with GO
terms that correspond to these functional classes were
individually analyzed. Special attention was given to
AS annotated through the immune related sequences
from horseshoe crabs and D. melanogaster. This
analysis led to the identification of 123 AS, which
was reduced to 73 after sequencing both ends and
manually assembling each AS. These AS represent
several functional classes involved in the innate
immune response (Table 4).
4. Discussion
The present study presents the identification of
transcripts related to immune system by
12

C
PROOF
356
92
5548 47
35 35 34 3430
26 24 23
504
10
100
1000
hemoc
yanin
struc
tural
cons
titue
nt of
cytos
kelet
on
struc
tural
cons
titue
nt of
ribo
some
recep
tor
DNA bind
ing
suga
r bind
ing
RNA bind
ing
trans
posa
se
cell
adhe
sion m
olecu
le
pepti
dase
GTP bind
ing
trans
lation
regu
lator
trans
cripti
on fa
ctor
unkn
own f
uncti
on
Prim
ary
Lib
rary
EST
s
Fig. 1. Gene expression profile of Acanthoscurria gomesiana hemocytes using Gene Ontology. Values indicate the number of sequenced clones
from the primary library grouped in categories of Molecular Function ontology. Only the fourteen most abundant categories are presented.
Table 3
GeneOntology (GO) terms related to the immune system
GO:0004252 Serine-type endopeptidase
GO:0003810 Protein-glutamine gamma-glutamyltransferase
GO:0003796 Lysozyme
GO:0006961 Antibacterial humoral response (sensu Inverteb-
rata)
GO:0008329 Pattern recognition receptor
GO:0003823 Antigen binding
GO:0005530 Lectin
GO:0017114 Wide-spectrum protease inhibitor
GO:0004867 Serine protease inhibitor
GO:0004888 Transmembrane receptor
GO:0003700 Transcription factor
GO:0006952 Defense response
GO:0004503 Monophenol monooxygenase
GO:0003793 Defense/immunity protein
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–12 5
+ model ARTICLE IN PRESS
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
UNCORRE
high-throughput sequencing of hemocyte ESTs of the
tarantula spider A. gomesiana. The sequencing of
clones from both a primary and a normalized cDNA
library yielded a total of 3723 transcripts that included
at least 20 abundantly expressed. The number of
transcripts is considered over estimated, because the
sequence assembly software (CAP3) often separates
identical ESTs into different AS [19]. This separation
may be due to errors or polymorphisms in the
sequences, or to regions of the transcripts with low
coverage of ESTs [19].
The number of transcripts without match in the NR
database was high (1531, 41%), suggesting the finding
of a great number of novel genes. However, the
percentage of novel genes is much higher on transcripts
with ORFs shorter than 400 bp than on transcripts of
longer ORFs (Fig. 2). Not all short ORFs without
matches in public databases should be considered as
novel genes, since it is difficult to find significant
matches (e-value !1!10K6) on database searches
with short protein sequences. As reported before,
transcripts with low protein coding capacity (short
ORFs) may correspond to ESTs of either very short
sequences or sequences with long 5 0 UTR (Untranslated
Region) [20]. Therefore, the estimated percentage of
DCI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1
TED novel genes should be closer to that found with ORFs
longer than 900 bp (20%).
The gene expression profile of spider hemocytes was
characterized by the abundance of primary library ESTs
(Fig. 1). Among the sequences with functional
attribution, the AS related to hemocyanin subunits
were remarkably abundant (356 ESTs), indicating the
involvement of hemocytes in the production of this
oxygen transport protein. In the tarantula spider
Eurypelma californicum, the hemocyanins are
–12
UNCORRECTED PROOF
Table 4
Immune related Assembled Sequences (AS) identified in the cDNA libraries of A. gomesiana hemocytes
AS code # of ESTsa
Norm. Prim. Gi Description Organism E-value
Phenoloxidases
AGCCAR1001B07 9 59 122792 Hemocyanin A chain Eurypelma californicum 0
AGCCAR1019B12 8 35 20138395 Hemocyanin B chain Eurypelma californicum 0
AGCCAR1008E07 5 21 20138398 Hemocyanin C chain Eurypelma californicum 0
AGCCAR1004D09 9 71 17376946 Hemocyanin D chain Eurypelma californicum 0
AGCCAR1013F04 4 40 70624 Hemocyanin E chain Eurypelma californicum 0
AGCCAR1001H09 34 39 20138397 Hemocyanin F chain Eurypelma californicum 0
AGCCAR1009A03 5 4 20138397 Hemocyanin F chain Eurypelma californicum 1!10K180
AGCCAR1003A06 13 103 20138396 Hemocyanin G chain Eurypelma californicum 0
Antimicrobial peptides and proteins
AGCCAR1001C06 7 2 52782738 Acanthoscurrin 1 precursor A. gomesiana 2!10K30
AGCCAR1006D12 2 1 52782737 Acanthoscurrin 2 precursor A. gomesiana 2!10K30
AGCCAR1034B01 1 1 28445738 Gomesin precursor A. gomesiana 3!10K41
AGCCAR1010G04 1 0 28445738 Gomesin precursor A. gomesiana 6!10K38
AGCCAR1035C06 2 0 1085148 lysozyme S Drosophila melanogaster 2!10K16
Serine proteases
AGCCAR1044G07 1 0 542517 coagulation factor B precursor Tachypleus tridentatus 5!10K50
AGCCAR1057G02 1 0 542517 coagulation factor B precursor Tachypleus tridentatus 1!10K58
AGCCAR2017E09 0 1 542517 coagulation factor B precursor Tachypleus tridentatus 2!10K54
AGCCAR1026C07 5 2 129688 Proclotting enzyme precursor Tachypleus tridentatus 1!10K81
AGCCAR1003H07 1 0 18542425 factor C precursor Tachypleus tridentatus 3!10K75
AGCCAR1006G12 1 1 913964 factor C Carcinoscorpius rotundicauda 1!10K100
AGCCAR1011F11 1 1 7387836 Limulus clotting factor C precursor Carcinoscorpius rotundicauda 1!10K129
AGCCAR1012C07 2 0 3928787 factor B SpBf Strongylocentrotus purpuratus 8!10K50
AGCCAR1013H11 6 0 1817554 limulus factor D Tachypleus tridentatus 1!10K117
AGCCAR1006A04 3 0 25989209 coagulation factor-like protein 3 Hyphantria cunea 5!10K60
AGCCAR1006C09 4 0 23266416 serine protease PC5-A Rana esculenta 8!10K44
AGCCAR1019E09 2 0 28194028 prothrombin precursor Takifugu rubripes 2!10K50
AGCCAR2001D06 0 1 28194028 prothrombin precursor Takifugu rubripes 1!10K09
AGCCAR1020D11 3 0 847761 SPC3 Branchiostoma californiense 3!10K56
AGCCAR1013F12 3 0 26332511 unnamed protein product Mus musculus 2!10K11
Serine protease inhibitors
AGCCAR2016C11 0 1 17223666 serine proteinase inhibitor serpin-3 Rhipicephalus appendiculatus 9!10K30
AGCCAR1001D03 10 2 1078956 intracellular coagulation inhibitor
type 2 (LICI 2)
Tachypleus tridentatus 5!10K65
AGCCAR1003H02 7 3 1078956 intracellular coagulation inhibitor
type 2 (LICI 2)
Tachypleus tridentatus 2!10K65
AGCCAR1005F08 2 0 34881479 similar to serine (or cysteine) pro-
teinase inhibitor
Rattus norvegicus 1!10K47
AGCCAR1002C10 3 4 7521905 alpha-2-macroglobulin Limulus sp. 1!10K139
AGCCAR1003A01 5 0 13928544 complement component C3 Branchiostoma belcheri 8!10K49
AGCCAR1013G08 1 0 20302747 unknown Branchiostoma floridae 1!10K56
DC
I848—
7/1
0/2
00
5—
02:1
7—
-[-no
entity
-]-—16683
6—
XM
LM
OD
EL
3dc+
–pp.
1–12
D.M
.L
oren
zini
eta
l./
Develo
pm
enta
la
nd
Co
mp
ara
tiveIm
mu
no
log
yxx
(xxxx)1
–1
26 +model
ARTICLEIN
PRESS
52
1
52
2
52
3
52
4
52
5
52
6
52
7
52
8
52
9
53
0
53
1
53
2
53
3
53
4
53
5
53
6
53
7
53
8
53
9
54
0
54
1
54
2
54
3
54
4
54
5
54
6
54
7
54
8
54
9
55
0
55
1
55
2
55
3
55
4
55
5
55
6
55
7
55
8
55
9
56
0
56
1
56
2
56
3
56
4
56
5
56
6
56
7
56
8
56
9
57
0
57
1
57
2
57
3
57
4
57
5
57
6
57
7
57
8
57
9
58
0
58
1
58
2
58
3
58
4
58
5
58
6
58
7
58
8
58
9
59
0
59
1
59
2
59
3
59
4
59
5
59
6
59
7
59
8
59
9
60
0
60
1
60
2
60
3
60
4
60
5
60
6
60
7
60
8
60
9
61
0
61
1
61
2
61
3
61
4
61
5
61
6
61
7
61
8
61
9
62
0
62
1
62
2
62
3
62
4

UNCORRECTED PROOF
AGCCAR1013A03 1 0 25149822 thrombospondin Caenorhabditis elegans 6!10K11
AGCCAR1056G07 3 0 25149822 thrombospondin Caenorhabditis elegans 4!10K11
AGCCAR1037F11 1 3 13346812 thrombospondin Haemonchus contortus 2!10K35
AGCCAR2006A03 0 1 22901764 Kunitz-like protease inhibitor pre-
cursor
Ancylostoma caninum 1!10K31
AGCCAR1013B05 1 0 2133556 cystatin precursor Tachypleus tridentatus 2!10K14
Lectins
AGCCAR1041B12 2 27 1346296 Hemocytin precursor Bombyx mori 1!10K104
AGCCAR1010G07 1 0 6630613 Hemolectin Drosophila melanogaster 1!10K21
AGCCAR1012D08 2 0 21666693 hemolectin-like protein Penaeus monodon 2!10K10
AGCCAR1053A05 1 0 1346296 Hemocytin precursor Bombyx mori 5!10K31
AGCCAR1036E09 3 0 1346296 Hemocytin precursor Bombyx mori 1!10K25
AGCCAR1045A09 1 0 17942826 Tachylectin 5a Tachypleus tridentatus 4!10K68
AGCCAR1046E07 1 0 5851893 Tachylectin 5a Tachypleus tridentatus 7!10K75
AGCCAR1005B06 1 0 17942826 Tachylectin 5a Tachypleus tridentatus 2!10K79
AGCCAR1006D08 2 0 5851893 Tachylectin 5a Tachypleus tridentatus 6!10K71
AGCCAR1009B07 1 0 17942826 Tachylectin 5a Tachypleus tridentatus 6!10K61
AGCCAR1017A07 4 1 5851897 Tachylectin 5b Tachypleus tridentatus 7!10K76
AGCCAR1017E06 1 0 17942826 Tachylectin 5a Tachypleus tridentatus 1!10K73
AGCCAR1018D08 1 0 17942826 Tachylectin 5a Tachypleus tridentatus 5!10K72
AGCCAR1052G07 1 0 5851893 Tachylectin 5a Tachypleus tridentatus 1!10K70
AGCCAR1018F02 6 1 5851893 Tachylectin 5a Tachypleus tridentatus 3!10K80
AGCCAR1008C06 6 0 31217088 ENSANGP00000012978 Anopheles gambiae 2!10K44
AGCCAR2018F05 0 1 27808640 peptidoglycan recognition protein Bos Taurus 6!10K36
AGCCAR1044F01 1 0 4878035 neurocan core protein precursor Gallus gallus 6!10K18
AGCCAR1014G02 3 0 4505245 mannose receptor C type 1 precursor Homo sapiens 1!10K28
AGCCAR1015G05 1 0 4505245 mannose receptor C type 1 precursor Homo sapiens 2!10K23
AGCCAR1027E01 2 0 4505245 mannose receptor C type 1 precursor Homo sapiens 1!10K25
AGCCAR1025B08 4 0 12738842 polydomain protein Mus musculus 4!10K49
AGCCAR2010G02 0 1 84651 C-reactive protein chain 3.3 Limulus sp. 2!10K15
AGCCAR1036D05 3 2 6981152 lectin, galactose binding Rattus norvegicus 3!10K18
AGCCAR1043F05 1 0 9857647 galectin LEC-4 Caenorhabditis elegans 5!10K08
AGCCAR1055H06 1 0 2833353 Galectin-4 (Lactose-binding lectin 4) Sus scrofa 4!10K16
Other immune related molecular functions
AGCCAR1010C07 1 0 22651842 Toll-related protein; AeTehao Aedes aegypti 5!10K17
AGCCAR1028H03 1 1 22651842 Toll-related protein; AeTehao Aedes aegypti 9!10K29
AGCCAR2016F04 0 1 22651842 Toll-related protein; AeTehao Aedes aegypti 1!10K40
AGCCAR1015B09 2 1 9965396 Toll/IL-1 receptor binding protein
MyD88
Xenopus laevis 7!10K34
AGCCAR1031C10 1 0 24650493 spatzle CG6134-PI Drosophila melanogaster 2!10K10
AGCCAR1009B05 3 1 15718457 Peroxinectin Penaeus monodon 1!10K89
AGCCAR1049A04 1 0 345423 protein-glutamine gamma-glutamyl-
transferase (EC 2.3.2.13)
Tachypleus tridentatus 2!10K58
The BLASTX hits on the protein sequence database from NCBI (NR) with the lowest E values (implying the most significant similarities) are indicated in the table.a Number of ESTs in each assembled sequence obtained from Normalized (Norm.) and Primary (Pri.) cDNA libraries.
DC
I848—
7/1
0/2
00
5—
02:1
7—
-[-no
entity
-]-—16683
6—
XM
LM
OD
EL
3dc+
–pp.
1–12
D.M
.L
oren
zini
eta
l./
Develo
pm
enta
la
nd
Co
mp
ara
tiveIm
mu
no
log
yxx
(xxxx)1
–1
27
+model
ARTICLEIN
PRESS
62
5
62
6
62
7
62
8
62
9
63
0
63
1
63
2
63
3
63
4
63
5
63
6
63
7
63
8
63
9
64
0
64
1
64
2
64
3
64
4
64
5
64
6
64
7
64
8
64
9
65
0
65
1
65
2
65
3
65
4
65
5
65
6
65
7
65
8
65
9
66
0
66
1
66
2
66
3
66
4
66
5
66
6
66
7
66
8
66
9
67
0
67
1
67
2
67
3
67
4
67
5
67
6
67
7
67
8
67
9
68
0
68
1
68
2
68
3
68
4
68
5
68
6
68
7
68
8
68
9
69
0
69
1
69
2
69
3
69
4
69
5
69
6
69
7
69
8
69
9
70
0
70
1
70
2
70
3
70
4
70
5
70
6
70
7
70
8
70
9
71
0
71
1
71
2
71
3
71
4
71
5
71
6
71
7
71
8
71
9
72
0
72
1
72
2
72
3
72
4
72
5
72
6
72
7
72
8

C
OF0
100
200
300
400
500
600
300 400 500 600 700 800 900 1000 1500 2000 Mais
ORF Length (bp)
Num
ber
of A
Ss
0
10
20
30
40
50
60
70
80
Nov
el G
enes
(%
)
Fig. 2. Distribution of Assembled Sequences (AS) according to longest ORF length and to percentage of novel genes. The AS considered as novel
genes had no matches on NR database with e-value cut-off 1!10K6.
D
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–128
+ model ARTICLE IN PRESS
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
764
765
766
767
768
769
770
771
772
773
774
775
776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
793
794
795
796
797
798
799
800
801
802
803
804
805
806
807
808
809
810
811
812
813
814
815
816
817
818
819
820
821
822
823
824
825
826
827
828
829
830
831
832
UNCORRE
synthesized in hemocytes attached to the inner heart
wall, and the hematopoiesis is induced subsequent to
bleeding the animal [21]. Hemocyanins in circulating
hemocytes have never been described, suggesting that
the ESTs identified in A. gomesiana may be originated
from hemocytes detached from heart wall during
bleeding. The frequency of hemocyanin ESTs also
demonstrates the efficiency of the cDNA library
normalization. In the primary library hemocyanin
represented 228 of every 1000 sequenced ESTs,
followed by 17 in the normalized library. Therefore,
the normalization reduced 13-fold the abundance of
hemocyanins ESTs, and a similar result is expected for
other abundant transcripts.
Other abundant transcripts were identified for
functional classes related to protein biosynthesis,
cytoscheleton organization, energy metabolism, regu-
lation of gene expression and DNA transposition
(Fig. 1). Interestingly, one single transcript related to
a lectin, AGCCAR1041B12, contained 27 ESTs from
the primary library and other four single transcripts,
each containing more than 10 ESTs, were assigned to
unknown function.
The functional annotation of spider transcripts aided
the identification of several sequences related to
immune response. These sequences represent various
functional classes and may be involved in diverse
processes of the immune response.
The transcripts with similarity to hemocyanin
subunits, besides the participation on oxygen transport,
may participate in the immune response as
CI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1–
TED PROphenoloxidase (Table 4). In insects and crustaceans,
phenoloxidases are enzymes responsible for melanin
formation on wounds or invading organisms. This
enzyme is synthesized as an inactive precursor,
prophenoloxidase, and activated by the removal of a
fragment by serine-proteases [4]. The phenoloxidases
and hemocyanins from arthropods are similar in both
amino acid sequences and physico-chemical properties
of the active site [22]. Differing from other arthropods,
the chelicerates do not have phenoloxidases, and some
reports have demonstrated phenoloxidase activity by
hemocyanins [23–25]. The hemocyanin of the tarantula
spider E. californicum acquires phenoloxidase activity
after limited proteolysis with trypsin or chymotrypsin
[23], while in horseshoe crabs this conversion is
observed with non-enzymatic interaction of hemocya-
nin with clotting factors or antimicrobial peptides
[24,25]. A. gomesiana hemocyanin presents phenolox-
idase activity when incubated with the detergent
sodium dodecylsulfate (SDS), but no activity is found
after incubation with trypsin or chymotrypsin (Daffre,
personal communication). The similarity between the
antimicrobial peptides gomesin from A. gomesiana and
tachyplesin from horseshoe crabs [6] suggests that
gomesin may induce the phenoloxidase activity in the
spider hemocyanins as observed for its analog in
horseshoe crabs [25].
The alignment of hemocyanin subunits’ sequences
from A. gomesiana and E. californicum suggests the
finding of an eighth hemocyanin sequence (AGC-
CAR1009A03, data not shown). The hemocyanin of E.
12

C
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–12 9
+ model ARTICLE IN PRESS
833
834
835
836
837
838
839
840
841
842
843
844
845
846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
865
866
867
868
869
870
871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
886
887
888
889
890
891
892
893
894
895
896
897
898
899
900
901
902
903
904
905
906
907
908
909
910
911
912
913
914
915
916
917
918
919
920
921
922
923
924
925
926
ORREcalifornicum contains only seven subunits, which have
been completely sequenced [21]. For each A. gomesi-
ana hemocianin AS it was found one matching E.
californicum subunit with sequence identity O96%,
except for AGCCAR1009A03 that showed less than
50% sequence identity to any of the E. californicum
subunits. We are investigating if product of this cDNA
is a component of the A. gomesiana hemocyanin
complex.
Among the sequences related to antimicrobial
proteins and peptides (Table 4) we found the sequences
of the previously isolated antimicrobial peptides,
gomesin, acanthoscurrin 1 and 2 [6–8]. A novel isoform
of gomesin (AGCCAR1010G04) was found, presenting
89% amino acid sequence identity to the gomesin
precursor, as well as transcript of the original gomesin
(AGCCAR1034B01). The alignment of these
sequences shows that the novel gomesin differs from
the original by five substitutions within the mature
peptide region and other four substitutions in the
precursor regions (Fig. 3). Two substitutions in the
mature gomesin region conserved the hydrophobic
nature (residues 30(Y to F) and 35(V to L), probably
maintaining the hydrophobic patch of the gomesin
structure [26]. In addition, two conservative substi-
tutions were observed in the mature gomesin region
(residues 31(K to R) and 32(Q to N)). The only non-
conservative substitution in the mature gomesin region
(residue 39(R to S)) is in the C-terminal of the mature
peptide. In a structure-activity relationship study of
gomesin, a similar substitution (R to A) did not affect
the antimicrobial activity [27]. Therefore, the substi-
tutions in the sequence of the novel gomesin indicate
that this peptide presents antimicrobial activities
similar to the original gomesin.
The transcript identified with similarity to lysozyme
may participate in bacterial killing through hydrolysis
of its cell wall. Lysozyme gene expression in
hemocytes was observed in shrimps [28] and ticks
[29]. In mammals, the lysozymes are stored in granules
of macrophages and neutrophils, and are involved in
killing of Gram-positive bacteria [30].
UNCFig. 3. Sequence alignment of gomesin precursor isoforms. The spider
Assembled Sequences (AGCC.) are compared with the previously
described gomesin precursor [7]. Positions containing substitutions
are marked in gray for conservative and in black for non-conservative.
DCI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1
927
928
929
930
931
932
933
934
935
936
TED PROOF
Several spider transcripts were found with similarity
to serine proteases or serine protease inhibitors
(Table 4). The proteases involved in invertebrate
immune response are members of the chymotrypsin
family that are produced as zymogens and activated by
limited proteolysis [31]. These proteases are organized
in cascades and controlled by specific inhibitors. Four
different spider transcripts (AS) have similarity to
proteases (Factor B and Proclotting Enzyme) that
contain two conserved domains: CLIP and Chymo-
trypsin. These domains are observed in proteases of
crustaceans and insects that activate phenoloxidase [4]
or trigger the Toll signaling pathway through the
cleavage of Spaetzle [32].
The clotting cascade from horseshoe crabs contains
four proteases (Factors C, G, B and Proclotting
Enzyme), three inhibitors (LICI 1, 2 and 3) and the
clottable coagulogen. This cascade is started with the
autoactivation of Factor C and G, induced by their
binding to components of bacterial and fungal surfaces,
respectively [2]. The finding of several transcripts with
similarity to components of the horseshoe crabs clotting
cascade (Factors C and B, Proclotting Enzyme, LICI 2)
suggests the presence of a homologous cascade in A.
gomesiana, that may participate in blood coagulation.
Interestingly, no spider sequence with similarity to the
clottable coagulogen was found.
Besides the clotting cascade, the spider transcripts
with similarity to Factor C could also act as hemocyte
receptors. In horseshoe crabs, the Factor C present in
the cell surface is activated by LPS, which triggers
a cell-signaling cascade that leads to the degranulation
of hemocytes [33]. The corresponding spider sequences
contain the SUSHI conserved domains, which are the
LPS binding regions [34].
The sequences of one serine protease and one
protease inhibitor from the spider show similarity to
components of the vertebrate complement system. The
serine protease (AGCCAR1012C07) is similar to
Factor B from the alternative pathway for complement
activation, and the protease inhibitor (AGC-
CAR1003A01) is similar to complement factor C3. In
vertebrates, these proteins form the C3 convertase of
the alternative pathway, which cleave C3 to C3a and
C3b. The C3b binds to the surface of pathogens to
promote phagocytosis, and released C3a participates on
chemotaxis and activation of leucocytes [35]. A
complement-like protein from A. gambiae hemocytes
also promotes phagocytosis of Gram-negative bacteria
[36].
Another spider transcript (AGCCAR1002C10) pre-
sented a conserved domain of alpha-2-macroglobulins.
–12

C
D
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–1210
+ model ARTICLE IN PRESS
937
938
939
940
941
942
943
944
945
946
947
948
949
950
951
952
953
954
955
956
957
958
959
960
961
962
963
964
965
966
967
968
969
970
971
972
973
974
975
976
977
978
979
980
981
982
983
984
985
986
987
988
989
990
991
992
993
994
995
996
997
998
999
1000
1001
1002
1003
1004
1005
1006
1007
1008
1009
1010
1011
1012
1013
1014
1015
1016
1017
1018
1019
1020
1021
1022
1023
1024
1025
1026
1027
1028
1029
1030
1031
1032
1033
1034
1035
1036
1037
1038
1039
1040
UNCORRE
These wide-spectrum protease inhibitors are present at
high concentration in the plasma of horseshoe crabs,
and assist the hemocytes in the clearance of circulating
proteases [37].
Several transcripts with similarity to lectins were
identified (Table 4). Lectins are proteins with sugar
binding activity that, in animals, participate in the
immune response as recognition molecules. Different
glycoconjugates present on the surface of pathogens are
recognized by specific lectins, and the absence of these
glycoconjucates on host cells serves as a marker to
distinguish between self and non-self [38]. The most
abundant lectin transcript has 27 ESTs from the primary
library (AGCCAR1041B12), and presents very signifi-
cant similarity (1!10K104) to hemocytin from Bombyx
mori hemocytes [39]. This protein presents sugar
binding and hemocyte aggregating activities, and its
synthesis is induced upon infection [39].
Ten spider transcripts present similarity to tachy-
lectin 5 from horseshoe crab (Table 4). In this
organism, this protein is found in the cell free
hemolymph at high concentration (O10 mg/ml) and
agglutinates bacteria. The tachylectin 5 presents a
fibrinogen conserved domain, also found in vertebrate
ficolins (involved in the lectin pathway of complement
system activation) and fibrinogen (involved in blood
clotting) [40]. In mollusks, fibrinogen-like proteins are
present as different isoforms in the mucous glands of
Limax flavus [41] and in the hemolymph of Biompha-
laria glabrata, where they are produced after infection
with a trematode [42]. The variety of transcripts
containing fibrinogen domain found in the spider was
also observed in the genomes of A. gambiae and D.
melanogaster [18].
Some transcripts are related to other lectins
(Table 4), such as: peptidoglycan recognition protein
(PGRP), C type lectins, pentraxins and galectins.
PGRPs show binding activity to bacterial cell wall,
and are involved in the phagocytosis of Gram-negative
bacteria in insects [43], activate the phenoloxidase
cascade and trigger the Toll [44] and Imd signaling
pathways [45]. The C-type lectin conserved domain
was found in a protein present in the prophenoloxidase
activating complex of Manduca sexta [46] and in two
vertebrate proteins: colectins (involved in lectin path-
way of complement activation) and selectins (involved
in leukocyte traffic to infected tissues) [38]. Pentraxins
are found in the serum of vertebrates, and in horseshoe
crabs it is one of the most abundant proteins of the free
cell hemolymph [38]. Some galectins are secreted from
mammal macrophages and are involved in activation
and recruiting of leukocytes [47].
CI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1–
TED PROOF
Five spider transcripts were found with similarity to
components of the Toll signaling pathway (Table 4),
including the Toll receptor, the MyD88 Toll adaptor
and Spatzle. This pathway activates the synthesis of
antimicrobial peptides in the fat body of D. melanoga-
ster and the production of inflammatory mediators
(citokines, chemokines) on vertebrate macrophages
[48,49]. In vertebrates, the Toll receptors bind to
microbial surface molecules directly, while in D.
melanogaster the activated Spatzle is the Toll target
[48]. After the binding, the Toll receptors interact with
MyD88 and start the intracellular signaling pathway
that regulates the transcription of immune related
genes. The spider sequence with similarity to Spatzle
suggests a Toll activation mechanism similar to the one
observed in D. melanogaster.
Finally, transcripts with similarity to peroxinectins
and transglutaminases were found (Table 4). Perox-
inectins are adhesion molecules stored in hemocyte
granules of crustaceans. Following the degranulation
induced by infection, the released peroxinectins are
activated by serine-proteases to stimulate cell adhesion,
phagocytosis and encapsulation. The sequence of this
protein contains a peroxidase domain followed by a
C-terminal domain, which is involved in adhesion [50].
Transglutaminases are enzymes that catalyze the
covalent binding of glutamine residues to lysine
residues or other primary amines. In crustaceans, this
protein is released from hemocytes and polymerizes the
clot protein, forming a stable clot around the wound
[51]. The horseshoe crab’s transglutaminase promotes
the attachment of the hemocytes over the clot through
the covalent binding of a hemocyte surface protein,
proxins, to coagulin [52].
The sequences from the spider A. gomesiana
produced for this work increased immensely the
diversity of araneae genes deposited in public
databases, specially the immune related genes. These
sequences will be an useful material for comparative
and evolutionary studies. In addition, this material will
support the characterization of the spider hemocyte
proteome by a mass spectrometry approach (in
progress).
Acknowledgements
This work was supported by grants from Fundacao
de Amparo a Pesquisa do Estado de Sao Paulo
(FAPESP) (Brazil), Conselho Nacional de Desenvolvi-
mento Cientıfico e Tecnologico (CNPq) (Brazil). We
are thankful to Dr Ana Teresa R. de Vasconcelos
(LNCC/MCT) for processing the Interpro searches,
12

D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–12 11
+ model ARTICLE IN PRESS
1041
1042
1043
1044
1045
1046
1047
1048
1049
1050
1051
1052
1053
1054
1055
1056
1057
1058
1059
1060
1061
1062
1063
1064
1065
1066
1067
1068
1069
1070
1071
1072
1073
1074
1075
1076
1077
1078
1079
1080
1081
1082
1083
1084
1085
1086
1087
1088
1089
1090
1091
1092
1093
1094
1095
1096
Almir Samuel Zanca (CBMEG/UNICAMP) for DNA
sequencing, Renato Vicentini dos Santos (CBMEG/U-
NICAMP) and Apua Cesar de Miranda Paquola
(ICB/USP) for valuable discussions.
C
1097
1098
1099
1100
1101
1102
1103
1104
1105
1106
1107
1108
1109
1110
1111
1112
1113
1114
1115
1116
1117
1118
1119
1120
1121
1122
1123
1124
1125
1126
1127
1128
1129
1130
1131
1132
1133
1134
1135
1136
1137
1138
1139
1140
1141
1142
1143
1144
UNCORRE
References
[1] Ratcliffe NA, Whitten MMA. Vector immunity. In:
Gillespie SH, Smith GL, Osbourn A, editors. Microbe-vector
interactions in vector-borne diseases. Cambridge: Cambridge
University Press; 2004. p. 240–71.
[2] Iwanaga S, Kawabata S, Muta T. New types of clotting factors
and defense molecules found in horseshoe crab hemolymph:
their structures and functions. J Biochem (Tokyo) 1998;123(1):
1–15.
[3] Iwanaga S, Lee BL. Recent advances in the innate immunity of
invertebrate animals. J Biochem Mol Biol 2005;38(2):128–50.
[4] Cerenius L, Soderhall K. The prophenoloxidase-activating
system in invertebrates. Immunol Rev 2004;198:116–26.
[5] Bachere E, Gueguen Y, Gonzalez M, de Lorgeril J, Garnier J,
Romestand B. Insights into the anti-microbial defense of marine
invertebrates: the penaeid shrimps and the oyster Crassostrea
gigas. Immunol Rev 2004;198(1):149–68.
[6] Silva Jr PI, Daffre S, Bulet P. Isolation and characterization of
gomesin, an 18-residue cysteine-rich defense peptide from the
spider Acanthoscurria gomesiana hemocytes with sequence
similarities to horseshoe crab antimicrobial peptides of the
tachyplesin family. J Biol Chem 2000;275(43):33464–70.
[7] Lorenzini DM, Fukuzawa AH, da Silva Jr PI, Machado-
Santelli G, Bijovsky AT, Daffre S. Molecular cloning,
expression analysis and cellular localization of gomesin, an
anti-microbial peptide from hemocytes of the spider Acanthos-
curria gomesiana. Insect Biochem Mol Biol 2003;33(10):
1011–6.
[8] Lorenzini DM, da Silva Jr PI, Fogaca AC, Bulet P, Daffre S.
Acanthoscurrin: a novel glycine-rich antimicrobial peptide
constitutively expressed in the hemocytes of the spider
Acanthoscurria gomesiana. Dev Comp Immunol 2003;27(9):
781–91.
[9] Pereira LS. Study of two antimicrobial compounds from the
spider Acanthoscurria gomesiana. In: Departamento de Para-
sitologia. Universidade de Sao Paulo: Sao Paulo; 2004. p. 125.
[10] Supungul P, Klinbunga S, Pichyangkura R, Jitrapakdee S,
Hirono I, Aoki T, et al. Identification of Immune-Related Genes
in Hemocytes of Black Tiger Shrimp (Penaeus monodon). Mar
Biotechnol (NY) 2002;4(5):487–94.
[11] Gross PS, Bartlett TC, Browdy CL, Chapman RW,
Warr GW. Immune gene discovery by expressed sequence
tag analysis of hemocytes and hepatopancreas in the Pacific
White Shrimp, Litopenaeus vannamei, and the Atlantic
White Shrimp, L. setiferus. Dev Comp Immunol 2001;
25(7):565–77.
[12] Jenny MJ, Ringwood AH, Lacy ER, Lewitus AJ, Kempton JW,
Gross PS, et al. Potential indicators of stress response identified
by expressed sequence tag analysis of hemocytes and embryos
from the American oyster, Crassostrea virginica. Mar
Biotechnol (NY) 2002;4(1):81–93.
[13] Raghavan N, Miller AN, Gardner M, FitzGerald PC,
Kerlavage AR, Johnston DA, et al. Comparative gene analysis
DCI 848—7/10/2005—02:17—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1
TED PROOF
of Biomphalaria glabrata hemocytes pre—and post-exposure to
miracidia of Schistosoma mansoni. Mol Biochem Parasitol
2003;126(2):181–91.
[14] Dimopoulos G, Casavant TL, Chang S, Scheetz T, Roberts C,
Donohue M, et al. Anopheles gambiae pilot gene discovery
project: identification of mosquito innate immunity genes from
expressed sequence tags generated from immune-competent cell
lines. Proc Natl Acad Sci USA 2000;97(12):6619–24.
[15] Dimopoulos G, Christophides GK, Meister S, Schultz J,
White KP, Barillas-Mury C, et al. Genome expression analysis
of Anopheles gambiae: responses to injury, bacterial challenge,
and malaria infection. Proc Natl Acad Sci USA 2002;99(13):
8814–9.
[16] Bonaldo MF, Lennon G, Soares MB. Normalization and
subtraction: two approaches to facilitate gene discovery.
Genome Res 1996;6(9):791–806.
[17] Iwanaga S. The molecular basis of innate immunity in the
horseshoe crab. Curr Opin Immunol 2002;14(1):87–95.
[18] Christophides GK, Zdobnov E, Barillas-Mury C, Birney E,
Blandin S, Blass C, et al. Immunity-related genes and gene
families in Anopheles gambiae. Science 2002;298(5591):
159–65.
[19] Liang F, Holt I, Pertea G, Karamycheva S, Salzberg SL,
Quackenbush J. An optimized protocol for analysis of EST
sequences. Nucleic Acids Res 2000;28(18):3657–65.
[20] Whitfield CW, Band MR, Bonaldo MF, Kumar CG, Liu L,
Pardinas JR, et al. Annotated expressed sequence tags and
cDNA microarrays for studies of brain and behavior in the
honey bee. Genome Res 2002;12(4):555–66.
[21] Voit R, Feldmaier-Fuchs G, Schweikardt T, Decker H,
Burmester T. Complete sequence of the 24-mer hemocyanin
of the tarantula Eurypelma californicum. Structure and
intramolecular evolution of the subunits. J Biol Chem 2000;
275(50):39339–44.
[22] Decker H, Jaenicke E. Recent findings on phenoloxidase activity
and antimicrobial activity of hemocyanins. Dev Comp Immunol
2004;28(7–8):673–87.
[23] Decker H, Rimke T. Tarantula hemocyanin shows phenolox-
idase activity. J Biol Chem 1998;273(40):25889–92.
[24] Nagai T, Kawabata S. A link between blood coagulation and
prophenol oxidase activation in arthropod host defense. J Biol
Chem 2000;275(38):29264–7.
[25] Nagai T, Osaki T, Kawabata S. Functional conversion of
hemocyanin to phenoloxidase by horseshoe crab antimicrobial
peptides. J Biol Chem 2001;276(29):27166–70.
[26] Mandard N, Bulet P, Caille A, Daffre S, Vovelle F. The
solution structure of gomesin, an antimicrobial cysteine-rich
peptide from the spider. Eur J Biochem 2002;269(4):
1190–8.
[27] Miranda A, Fazio MA, Miranda MTM, Daffre S, Lamas WT.
Alanine series of the antimicrobial peptide gomesin: a structure-
activity relationship study. J Pep Sci 2004;(Suppl 10):179.
[28] Rojtinnakorn J, Hirono I, Itami T, Takahashi Y, Aoki T. Gene
expression in haemocytes of kuruma prawn, Penaeus japonicus,
in response to infection with WSSV by EST approach. Fish
Shellfish Immunol 2002;13(1):69–83.
[29] Simser JA, Macaluso KR, Mulenga A, Azad AF. Immune-
responsive lysozymes from hemocytes of the American dog tick,
Dermacentor variabilis and an embryonic cell line of the Rocky
Mountain wood tick, D. andersoni. Insect Biochem Mol Biol
2004;34(12):1235–46.
–12

C
D
D.M. Lorenzini et al. / Developmental and Comparative Immunology xx (xxxx) 1–1212
+ model ARTICLE IN PRESS
1145
1146
1147
1148
1149
1150
1151
1152
1153
1154
1155
1156
1157
1158
1159
1160
1161
1162
1163
1164
1165
1166
1167
1168
1169
1170
1171
1172
1173
1174
1175
1176
1177
1178
1179
1180
1181
1182
1183
1184
1185
1186
1187
1188
1189
1190
1191
1192
1193
1194
1195
1196
1197
1198
1199
1200
1201
1202
1203
1204
1205
1206
1207
1208
1209
1210
1211
1212
1213
1214
1215
1216
1217
1218
1219
1220
1221
1222
1223
1224
1225
1226
1227
1228
1229
1230
1231
[30] Ganz T, Gabayan V, Liao HI, Liu L, Oren A, Graf T, et al.
Increased inflammation in lysozyme M-deficient mice in
response to Micrococcus luteus and its peptidoglycan. Blood
2003;101(6):2388–92.
[31] Jiang H, Kanost MR. The clip-domain family of serine
proteinases in arthropods. Insect Biochem Mol Biol 2000;
30(2):95–105.
[32] Ligoxygakis P, Pelte N, Hoffmann JA, Reichhart JM. Activation
of Drosophila Toll during fungal infection by a blood serine
protease. Science 2002;297(5578):114–6.
[33] Ariki S, Koori K, Osaki T, Motoyama K, Inamori K,
Kawabata S. A serine protease zymogen functions as a
pattern-recognition receptor for lipopolysaccharides. Proc Natl
Acad Sci USA 2004;101(4):953–8.
[34] Tan NS, Ng ML, Yau YH, Chong PK, Ho B, Ding JL. Definition
of endotoxin binding sites in horseshoe crab factor C
recombinant sushi proteins and neutralization of endotoxin by
sushi peptides. Faseb J 2000;14(12):1801–13.
[35] Fujita T, Matsushita M, Endo Y. The lectin-complement
pathway–its role in innate immunity and evolution. Immunol
Rev 2004;198:185–202.
[36] Levashina EA, Moita LF, Blandin S, Vriend G, Lagueux M,
Kafatos FC. Conserved role of a complement-like protein in
phagocytosis revealed by dsRNA knockout in cultured cells of
the mosquito, Anopheles gambiae. Cell 2001;104(5):709–18.
[37] Melchior R, Quigley JP, Armstrong PB. Alpha 2-macroglo-
bulin-mediated clearance of proteases from the plasma of the
American horseshoe crab, Limulus polyphemus. J Biol Chem
1995;270(22):13496–502.
[38] Kilpatrick DC. Animal lectins: a historical introduction and
overview. Biochim Biophys Acta 2002;1572(2–3):187–97.
[39] Yamakawa M, Tanaka H. Immune proteins and their gene
expression in the silkworm, Bombyx mori. Dev Comp Immunol
1999;23(4–5):281–9.
[40] Gokudan S, Muta T, Tsuda R, Koori K, Kawahara T, Seki N,
et al. Horseshoe crab acetyl group-recognizing lectins involved
in innate immunity are structurally related to fibrinogen. Proc
Natl Acad Sci USA 1999;96(18):10086–91.
[41] Kurachi S, Song Z, Takagaki M, Yang Q, Winter HC,
Kurachi K, et al. Sialic-acid-binding lectin from the slug
Limax flavus—cloning, expression of the polypeptide, and tissue
localization. Eur J Biochem 1998;254(2):217–22.
UNCORRE
CI 848—7/10/2005—02:18—-[-no entity-]-—166836—XML MODEL 3dc+ – pp. 1–
TED PROOF
[42] Adema CM, Hertel LA, Miller RD, Loker ES. A family of
fibrinogen-related proteins that precipitates parasite-derived
molecules is produced by an invertebrate after infection. Proc
Natl Acad Sci USA 1997;94(16):8691–6.
[43] Ramet M, Manfruelli P, Pearson A, Mathey-Prevot B,
Ezekowitz RA. Functional genomic analysis of phagocytosis
and identification of a Drosophila receptor for E. coli. Nature
2002;416(6881):644–8.
[44] Michel T, Reichhart JM, Hoffmann JA, Royet J. Drosophila Toll
is activated by Gram-positive bacteria through a circulating
peptidoglycan recognition protein. Nature 2001;414(6865):
756–9.
[45] Takehana A, Katsuyama T, Yano T, Oshima Y, Takada H,
Aigaki T, et al. Overexpression of a pattern-recognition
receptor, peptidoglycan-recognition protein-LE, activates
imd/relish-mediated antibacterial defense and the prophenolox-
idase cascade in Drosophila larvae. Proc Natl Acad Sci USA
2002;99(21):13705–10.
[46] Yu XQ, Jiang H, Wang Y, Kanost MR. Nonproteolytic serine
proteinase homologs are involved in prophenoloxidase acti-
vation in the tobacco hornworm, Manduca sexta. Insect
Biochem Mol Biol 2003;33(2):197–208.
[47] Linehan SA, Martinez- Pomares L, Gordon S. Macro-
phage lectins in host defence. Microbes Infect 2000;2(3):
279–88.
[48] Beutler B. Toll-like receptors: how they work and what they do.
Curr Opin Hematol 2002;9(1):2–10.
[49] Hoffmann JA, Reichhart JM. Drosophila innate immunity: an
evolutionary perspective. Nat Immunol 2002;3(2):121–6.
[50] Johansson MW. Cell adhesion molecules in invertebrate
immunity. Dev Comp Immunol 1999;23(4–5):303–15.
[51] Hall M, Wang R, van Antwerpen R, Sottrup-Jensen L,
Soderhall K. The crayfish plasma clotting protein: a vitello-
genin-related protein responsible for clot formation in
crustacean blood. Proc Natl Acad Sci USA 1999;96(5):
1965–70.
[52] Osaki T, Okino N, Tokunaga F, Iwanaga S, Kawabata S.
Proline-rich cell surface antigens of horseshoe crab
hemocytes are substrates for protein cross-linking with a
clotting protein coagulin. J Biol Chem 2002;277(42):
40084–90.
12
1232
1233
1234
1235
1236
1237
1238
1239
1240
1241
1242
1243
1244
1245
1246
1247
1248
Top Related
Copyright © 2022 FDOKUMEN

