







Bahasa
Halaman
Hukum


Developmental& ComparativeImmunology
ARTICLE IN PRESS
0145-305X/$ - se
doi:10.1016/j.dc
Abbreviations
BlyS, B cell stim
Differential dete
ontology; HSP,
electrospray ion
identification te
electrophoresis;
exchange; SDS,
X-100; VSGP/F�CorrespondiE-mail addre
Developmental and Comparative Immunology 30 (2006) 1070–1083
www.elsevier.com/locate/devcompimm
Differential detergent fractionation for non-electrophoreticbovine peripheral blood monocyte proteomics reveals proteins
involved in professional antigen presentation
S.-R. Lee, G.T. Pharr, A.M. Cooksey, F.M. McCarthy, B.L. Boyd, L.M. Pinchuk�
Department of Basic Sciences, College of Veterinary Medicine, Mississippi State University, P. O. Box 6100, Mississippi State,
MS, 39762-6100, USA
Received 26 August 2005; received in revised form 8 February 2006; accepted 9 February 2006
Available online 13 March 2006
Abstract
Professional antigen presenting cells (APC), dendritic cells (DC) and their myeloid progenitors, monocytes/macrophages
are critical controllers of innate and adaptive immunity. Here we show that differential detergent fractionation (DDF)
analysis of bovine monocytes reveals proteins related to antigen pattern recognition, uptake and presentation to
immunocompetent lymphocytes. We identify 53 bovine proteins involved in immune function of professional APC. In
particular, 13 adhesion molecules, three toll-like receptors (TLR1, 6 and 8), three antigen uptake-related proteins
(including mannose receptor [MR] precursor), and eight actin-like proteins involved in active endocytosis were identified.
In addition, MHC class I and II-related proteins, cytokines, active substances and growth factors have been identified. We
conclude that the DDF approach can provide interpretable and meaningful functional information concerning protein
expression profiles associated with monocyte activation, transformation into macrophages and/or immature DC, and
maturation of monocyte-derived DC in the presence of multiple bovine pathogens.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Monocytes; Progenitors; DC; DDF; TLR; Lectin; Actin; MHC; Cytokines; Growth factors
e front matter r 2006 Elsevier Ltd. All rights reserved.
i.2006.02.002
: 1D, one-dimensional; 2D, two- dimensional; APC, Antigen presenting cells; APRIL, A proliferation-inducing ligand;
ulator protein; BVDV, Bovine Viral Diarrhea Virus; CTL, Cytotoxic T cells; DB, Database; DC, Dendritic cells; DDF,
rgent fractionation; DOC, Deoxycholate; GM-CSF, Granulocyte macrophage colony stimulating factor; GO, Gene
Heat shock protein; IFN, Interferon; Ig, Immunoglobulin; IL, Interleukin; LC-ESI/MS2, Liquid chromatography
ization tandem mass spectrometry; LPS, Lipopolysaccharide; MR, Mannose receptor; MudPIT, Multidimensional protein
chnology; NCBI, National Center for Biotechnology Information; NK, Natural killer; PAGE, Polyacrylamide gel
PBMC, Peripheral blood mononuclear cells; RP, Reverse phase; SCO, Subcommissural organ; SCX, Strong cation
Sodium dodecyl sulfate; Th1, T helper cell type 1; TNF-a, Tumor necrosis factor a; TLR, Toll-like receptor; TX, Triton
, Vascular smooth muscle cell growth-promoting; vWF, Von Willebrand factor
ng author. Tel.: +1 662 325 1436; fax: +1 662 325 1031.
ss: [email protected] (L.M. Pinchuk).

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1071
1. Introduction
Cells of the monocyte/macrophage lineage areimportant elements of the immune defense systembecause these cells can phagocytose foreign materi-al, present antigens to immunocompetent cells andproduce cytokines, including tumor necrosis factora (TNF-a) and interleukins 1 and 6 (IL-1, IL-6).Cytokine expression by monocytes is efficientlytriggered by stimulation with bacterial productssuch as lipopolysaccharide (LPS) acting via theTLR [1–4]. Bovine monocytes, as professional APC,express relatively high levels of MHC class I andclass II molecules, costimulatory molecules inducedupon activation, receptors for endo- and phagocy-tosis, and adhesion molecules [5]. Recent datasuggest that bovine monocytes are capable ofdirectly inducing immunoglobulin (Ig) secretion inactivated bovine peripheral blood B cells [5,6] aswas previously demonstrated in humans [7,8].Before entering the tissues, under the influence ofcertain cytokines, monocytes are capable of differ-entiating into macrophages or DC [9]. DC, highlyspecialized and the most powerful APC, are pivotalin immune responses, and are considered to beterminally differentiated cells derived from hemato-poietic progenitors or monocytes [10,11]. Cells withphenotypic and functional characteristics of DChave been produced from fully differentiated bovinemonocytes using granulocyte macrophage colonystimulating factor (GM-CSF) and IL-4 [12,13].
Recent studies have provided important cluesabout the mechanisms of TLR-mediated control ofadaptive immunity orchestrated by DC populations[14]. Professional APC antigen uptake and presen-tation of antigenic peptides on the relevant MHCmolecules involve: phagocytosis, the upregulation ofcostimulatory and MHC molecules, a switch inchemokine receptor expression, and the secretion ofcytokines [14–16]. Furthermore, chemokines areregulated through the recognition of pathogens viareceptors expressed by monocyte-derived DC lead-ing to the generation of effector responses includingT helper cell type 1 (Th1) [15] and cytotoxic T cells(CTL) responses [16]. Activation of monocyte-derived DC through TLR modulated the expressionof a common set of genes in response to differentpathogens [14]. After pathogen recognition, DCdownregulate mRNA levels for mannose receptor(MR) [17] and upregulate mRNA for the pro-inflammatory cytokines TNF-a, IL-1b and chemo-kines IL-8 and MIP-1a [18]. In the later stages of
inflammation, the pathogen-induced differentiationand maturation of DC from monocytes becomesmore apparent with the expression of chemokinereceptors CCR7 and CXCR4, and cell-adhesionproteins integrin, LFA-1 and E-cadherin which arerequired for migration to secondary lymphoidtissues [19,20]. Finally, in the secondary lymphoidtissues, mature DC express genes associated with Tcell activation and differentiation [19,20]. ThemRNA levels for CD80, MHC, as well as compo-nents of the proteosome Imp7 and Tap1 areupregulated at the mature DC stage [20–22].
Given the disconcordance between mRNA andprotein levels [23], we have chosen to identifyproteins in subcellular fractions of bovine mono-cytes using a proteomics-based approach. Once theproteins have been identified and characterized, itwill then be possible to address the modulation inexpression of these proteins during the conversionof monocytes to macrophages or DC by cytokinesor infectious agents.
DDF of whole cells yields four electrophoreticallydistinct fractions enriched in cytosolic, membrane-organelle, nuclear membrane and cytoskeletal-ma-trix markers, respectively [24]. Furthermore, multi-dimensional protein identification technology(MudPIT) couples two-dimensional (2D) chroma-tography of peptides with tandem mass spectro-metry (MS2), allowing for the identification ofproteins from highly complex mixtures [25]. Thecombination of DDF-MudPIT was used to increasethe proteome coverage over that of standard DDFand MudPIT [26]. Here we provide the baselineinformation on expression of protein profiles relatedto immune function, in particular, professionalantigen presentation, in bovine peripheral bloodmonocytes.
2. Materials and methods
2.1. Animals
Two conventionally reared and healthy Jerseycattle from the Mississippi State University DairyFacility (Mississippi State, MS) were used. TheMississippi State University Institutional AnimalCare and Use Committee approved all animal use.
2.2. Cell preparation
Bovine peripheral blood mononuclear cells(PBMC) were separated as described elsewhere

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831072
[12,13,27]. Briefly, PBMC were isolated usingHistopaque gradients (1.077 g/ml, Amersham Bios-ciences) and resuspended in RPMI-1640 supple-mented with 10% FBS, 1% Glutamax-1 (Gibco LifeTechnologies), 5� 10�5M 2-mercaptoethanol and100 IU/ml gentamicin (Gibco Life Technologies).Monocytes were separated from PBMC as de-scribed elsewhere [5]. Briefly, sheep red blood cellswere used to separate E-rosette negative andpositive fractions of PBMC. To isolate monocytes,the E-rosette negative fractions of PBMC wereincubated for 2 h at 37 1C and after removing non-adherent populations (mostly B cells), adherent cellswere incubated with mAbs to CD14 (MM61A,VMRD) followed by the addition of magnetic beadsconjugated with mouse anti-IgG1 (Miltenyi Biotech,Auburn, CA) [12,27]. CD14+ monocytes werepositively selected by using magnetic cell separationtechnique according to the manufacturer’s instruc-tions (Miltenyi Biotech). CD14+ monocytes wereobtained from two animals and stored at –70 1Cseparately. The bleeding procedures for the sameanimals have been repeated until the desirablenumber of 2� 107 cells was achieved. Then all thesamples were thawed and mixed followed byimmediate protein separation procedure.
2.3. Protein extraction, trypsin digestion and 2D-LC
ESI MS2
Proteins were isolated using DDF as described[26]. Briefly, DDF sequentially extracts proteinsfrom cellular compartments using a series ofdetergents. Cytosolic proteins were isolated byrepeated washes in digitonin buffer. After thedigitonin washes, the isolation of membrane,nuclear and cytoskeletal proteins were performedwith triton X-100 (TX), deoxycholate (DOC),Tween 40, and SDS buffers, respectively. Toevaluate the quality of isolated proteins, 1% ofthe protein samples were compared using 10%SDS-PAGE. For each of the detergent fractions,equal amounts of protein were precipitated with25% tricholoroacetic acid to remove salts anddetergents. Protein pellets were solubilized andthen digested with 100 ng of trypsin (50:1 ratioof substrate to enzyme) overnight at 37 1C.Peptides were desalted using a peptide microtrap(Michrom BioResources, Inc.) and eluted by a 0.1%triflouroacetic acid, 95% acetonitrile solution.Desalted peptides were dried and resuspended in0.1% formic acid.
2D LC ESI MS2 was done as described elsewhere[26]. Briefly, LC analysis was accomplished bystrong cation exchange (SCX) followed by reversephase (RP) liquid chromatography (LC) coupleddirectly in line with electrospray (ESI) ion trap MS.Samples were loaded into a LC gradient ionexchange system including a Thermo SeparationsP4000 quaternary gradient pump (ThermoElectronCorporation) coupled with a 0.32� 100mm BioBa-sic SCX column. A flow rate of 3 mL/min was usedfor both SCX and RP columns. A salt gradient wasapplied in steps of 0, 5, 10, 15, 20, 25, 30, 35, 40, 45,50, 57, 64, 71, 79, 90, 110, 300, and 700mMammonium acetate in 5% acetonitrile, 0.1% formicacid and the resultant peptides were loaded directlyinto the sample loop of a 0.18� 100mm BioBasicC18 RP LC column of a Proteome X workstation(ThermoElectron). The RP gradient used 0.1%formic acid in acetonitrile and increased theacetonitrile concentration in a linear gradient from5% to 30% in 30min and then 30% to 65% in 9minfollowed by 95% for 5min and 5% for 15min.TheLC peak ion chromatograms of four bovinemonocyte DDF are presented as Supplementarydata online (see Appendix A).
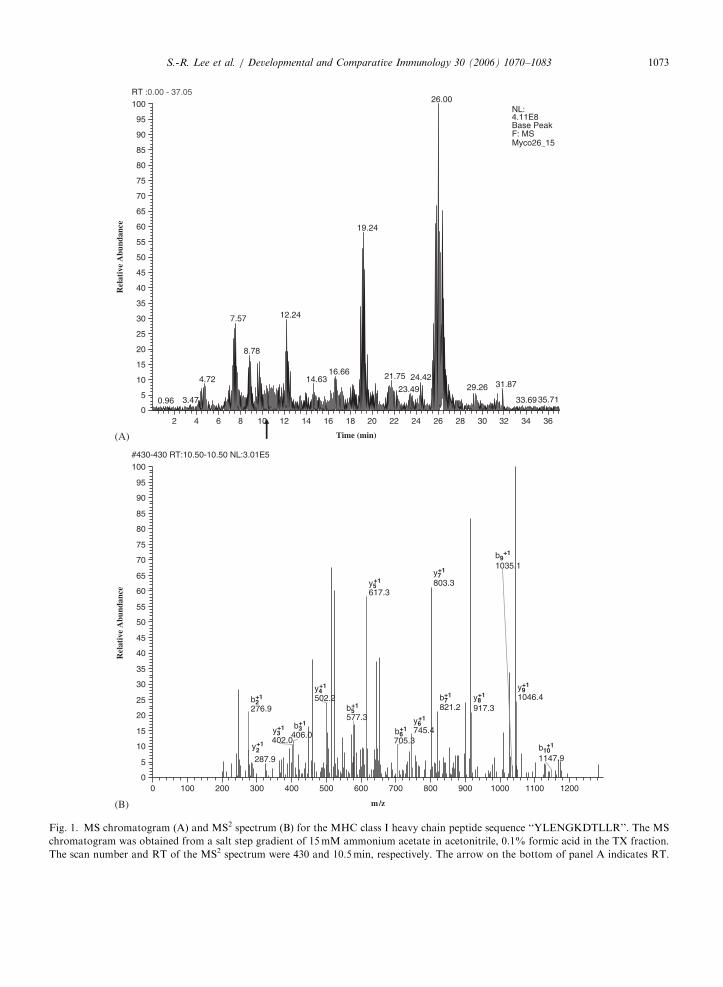
The spectrum collection time was 59min for everySCX step. The LCQ Deca ion trap mass spectro-meter was configured to optimize the duty cyclelength with the quality of data acquired byalternating between a single full MS scan followedby three tandem MS scans on the three most intenseprecursor masses from full scan. The collisionenergy was normalized to 35%. Dynamic massexclusion windows were set at 2min, and all of thespectra were measured with an overall mass/charge(m/z) ration range of 200–2000. Shown in Fig. 1 isone example of a MS chromatogram (Fig. 1A) andMS2 spectrum (Fig. 1B) for MHC class I heavychain peptide, GI# 1864812.
2.4. Protein identification and analysis
Proteins were identified and analyzed as pre-viously described [26]. The non-redundant proteindatabase (DB) downloaded from the NationalCenter for Biotechnology Information (NCBI;3/29/05) by TurboSEQUEST (Bioworks Browser3.1; ThermoElectron) was used to create a bovinesubset (bovine DB; search terms: bos taurus and bos
indicus). Trypsin digestion was applied in silico tobovine DB and mass changes due to cysteinecarbamidomethylation and methionine oxidation
ARTICLE IN PRESS
RT :0.00 - 37.05
2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 360
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100 26.00
19.24
12.247.57
8.78
16.66 21.75 24.424.72 14.6331.8729.2623.49
3.47 33.6935.710.96
NL:4.11E8Base PeakF: MSMyco26_15
Rel
ativ
e A
bund
ance
Time (min)
#430-430 RT:10.50-10.50 NL:3.01E5
0 100 200 300 400 500 600 700 800 900 1000 1100 12000
5
10
15
20
25
30
35
40
45
50
55
60
65
70
75
80
85
90
95
100
y5+1
617.3
y9+1
1046.4y4+1
502.2b2+1
276.9y8+1
917.3
y6+1
745.4b3+1
406.0
b7+1
821.2
y3+1
402.0b6+1
705.3
y7+1
803.3
b5+1
577.3
y2+1
287.9b10
+1
1147.9
b9+1
1035.1
m/z
Rel
ativ
e A
bund
ance
(A)
(B)
Fig. 1. MS chromatogram (A) and MS2 spectrum (B) for the MHC class I heavy chain peptide sequence ‘‘YLENGKDTLLR’’. The MS
chromatogram was obtained from a salt step gradient of 15mM ammonium acetate in acetonitrile, 0.1% formic acid in the TX fraction.
The scan number and RT of the MS2 spectrum were 430 and 10.5min, respectively. The arrow on the bottom of panel A indicates RT.
S.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1073

ARTICLE IN PRESS
DOC 9
SDS2
Digitonin1
1
4
30
0 0
0
0
Triton X-10033
Fig. 2. Distribution of immune-related proteins identified from
bovine monocyte DDF. The MS2 data from each of the DDF
samples were analyzed using the bovine DB subset and the
resulting proteins were classified with the respect to their
subcellular location.
Table 1
Proteins related to major cellular functions identified in bovine
monocytes by DDF-MudPIT
Biological functions Number of
protein
DDF
Proteins related to immune
functions
53 D/T/TD/S
Sodium ion transport 2 T
Protein transport 1 T
Thyroid hormone
generation; transport
1 T
ATP synthesis coupled
proton transport
14 T/TD/S
Lipid transport 5 T
Oxygen transport 7 T
Electron transport 27 T/TD/S
Protein folding 9 T/TD
Lactose biosynthesis 14 D/T/TD
Protein biosynthesis 12 T
Cytochrome-c oxidase
activity
6 T/TD/S
Nucleosome assembly 5 T/TD/S
Signal transduction 10 D/T/TD
Proteolysis and peptidolysis 2 T/TD
Gene regulation 7 T/TD
Blood coagulation 3 T/S
Protein amino acid
dephosphorylation
4 D/T/TD/S
Regulation of apoptosis 1 T
Muscle contraction 3 T/TD/S
Metabolism 7 T
Unannotated 91 D/T/TD/S
Total 284
Key: DDF, Differential detergent fraction; D, Digitonin; T,
Triton X-100; TD, Tween 40-deoxycholate; S, Sodium dodecyl
sulfate.
S.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831074
were included. The bovine DB was used to searchtandem MS using peptide (MS precursor ion) masstolerance of 1.5Da and a fragment ion (MS2) masstolerance of 1.0Da. Peptide matches were consid-ered genuine if they were X6 amino acids with a X
correlation values of 1.5, 2.0 and 2.5 (+1, +2, and+3 ions, respectively) and DCn (X0.1) values [28].Each protein was classified with respect to itsbiological process using gene ontology (GO) anno-tation [29]. GO: biological process was determinedfor each protein by searching the UniProt database(http://www.pir.uniprot.org/).
3. Results and discussion
3.1. Subcellular fractionation of bovine CD14+
peripheral blood monocytes
A widely used 2D separation technique inproteomics has been 2D gel electrophoresis. How-ever, this technique is limited by its inability todetect and identify low abundance proteins, pro-teins with extremes in isoelectric point and mole-cular weight and very hydrophobic proteins [30]. Toovercome these obstacles, several comprehensivetechniques have been developed [31], includingMudPIT. The combination of DDF and MudPITallows obtaining a comprehensive cellular proteomeas well as information about a protein’s subcellularlocalization [26]. We identified a total of 284proteins from monocytes using this non-electro-phoretic technique. In comparison, only 250 pro-teins were identified from bovine neutrophils usingone-dimensional (1D) electrophoresis coupled withMudPIT [32].
In our study proteins from four different sub-cellular fractions were obtained using differentdetergent buffers. Different proteins are isolatedfrom the different detergent fractions, and aftereight washes with DDF buffer 1, most of thecytosolic proteins had been extracted (Fig. 2).
In this study tandem mass spectra were analyzedusing a bovine DB and the identified proteins wereclassified according to their GO biological process(Table 1). We found a total of 284 proteins of which91 (32.0%) were not annotated and not related tothe immune function based on the availableliterature. Of the remaining proteins, 53 wererelated to immune function (27.5%), including 13unannotated bovine proteins that have been as-signed to this group based on the available literaturein humans, mice and cattle (Table 2). Proteins

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1075
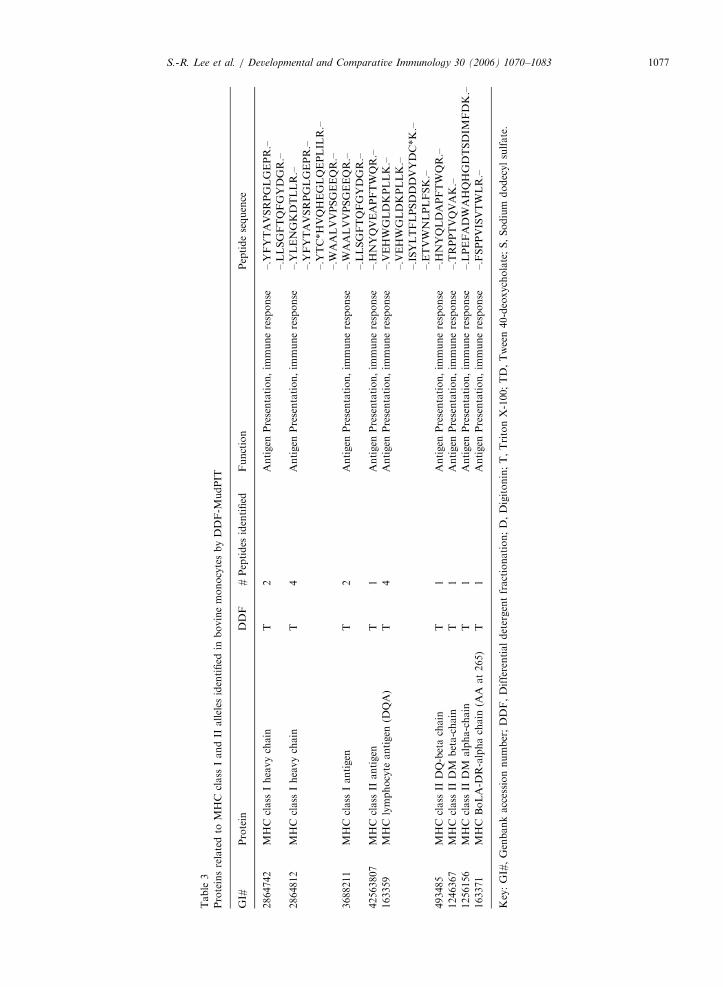
annotated to immune response were examined inmore detail to determine the specific biologicalprocess such as antigen uptake, processing andpresentation (Tables 2 and 3).
Information on the immunologically importantproteins is summarized in Tables 2 and 3. Weidentified eight categories of immune responseproteins: antigen uptake; innate antigen recognition;immune response; cell proliferation; adhesion,integrin-mediated signal pathway; actin cytoskele-ton organization and biogenesis; antigen presenta-tion, immune response; response to unfoldedprotein (Tables 2 and 3). Most of the proteinsinvolved in immune function were found in the TXfraction (62.3%) (Fig. 2).
Overall, 13 bovine proteins related to cell adhe-sion, three TLR proteins (bovine TLR1, 6, 8)involved in innate antigen pattern recognition, threeproteins related to antigen uptake including the MRprecursor, and eight actin-related proteins involvedin active endocytosis in monocytes/macrophages andimmature DC were identified (Table 2). In addition,a total of nine MHC class I- and II-related proteinswere identified in bovine monocytes (Table 3).Finally, cytokines, including active substances andgrowth factor proteins were also found in bovineCD14+ monocytes. Namely, interferon- g (IFN- g),transforming growth factor beta 1 precursor, vascu-lar endothelial growth factor C and heat shockprotein (HSP) have been identified by DDF-MudPIT(Table 2).
3.2. Proteins involved in antigen uptake, processing
and presentation identified by DDF
Different mechanisms of antigen uptake are usedby different APC types and determine their relativeefficiency in priming MHC restricted T cells [27,33].Capture of antigens by surface receptors, such asIgR, FcR or MR allows efficient delivery of antigento the processing compartment via receptor-mediated endocytosis [34]. Recently, we haveidentified MR as the important receptor involvedin endocytosis in bovine monocytes [27]. We havealso demonstrated that antigen uptake throughMR-mediated endocytosis, an important APCfunction, is affected in monocytes in the early stageof Bovine viral diarrhea virus (BVDV) infectionduring the first 24 h, with both BVDV biotypes [27].Endocytosis can occur by several mechanisms, inparticular, clathrin-, non-coated pit-, caveolar-,membrane ruffles-dependent and others [34]. In
our research, we identified three proteins related toantigen uptake: the galactose binding lectin, MyD-1, and MR precursor (Table 2). In addition, weidentified eight proteins related to cytoskeleton suchas actin, beta actin and gamma actin (Table 2).Previous data in humans suggest that lectins, inparticular galectins (galactose binding lectins) andtheir ligands, regulate immune cell homeostasiswhereas some members of this family, such asgalectin-3, behave as amplifiers of the inflammatorycascade [35]. MyD-1, found in bovine monocytes,macrophages, granulocytes, and a subset of afferentlymph veiled cells, plays a role in regulation of theinnate immunity in humans by pathogen productssuch as LPS, purified protein derivative andzymosan [36,37]. MR, a C-type lectin containingcarbohydrate recognition domains with broadspecificity for sugars, mediates phagocytosis ofmannose-coated particles, such as yeast and en-docytosis of mannosylated glycoproteins in macro-phages and immature DC [38–41]. MR can beviewed as a pattern recognition molecule that canprovide APC with some capacity of self non-selfdiscrimination at the level of antigen uptake on thebasis of glycosylation and perhaps hydrophobicity[27,38]. Antigens that fail to bind to cell surfacereceptors can still be taken up by micro- andmacropinocytosis [42]. While micropinocytosis oc-curs constitutively in all cells, macropinocytosis islimited to few cell types, such as macrophages andendothelial cells stimulated by growth factors[43–45]. In this study, we found two growth factorsthat could function in antigen uptake: transforminggrowth factor beta 1 precursor and vascularendothelial growth factor C (Table 2).
One of the most important functions in profes-sional APC is an innate form of antigen recognition[46]. TLR recognize specific molecular patterns thatare present in microbial components, and triggerDC maturation. Mature human DC induce costi-mulatory molecule expression and increase antigenpresenting capacity [46]. Therefore, TLR help todirect adaptive immune responses to antigensderived from microbial pathogens [46]. Stimulationof different TLR induces distinct patterns of geneexpression which both lead to the activation ofinnate immunity and instruct the development ofantigen specific acquired immunity in humans [47].In this study, three bovine TLR were identified:TLR 1, 6 and 8 (Table 2). In addition, cathelicidin[48] and fetuin [49], which are involved in innateantigen recognition, were also identified (Table 2).

ARTICLE IN PRESS
Table 2
Proteins related to professional antigen presentation identified in bovine monocytes by DDF-MudPIT
GI# Protein DDF # Peptides
identified
Function
28189272* Similar to galactose-binding lectin T 1 Antigen uptake
2842388* MyD-1 antigen T/TD 4 Antigen uptake
163336 Mannose-6-phosphate receptor
precursor
T 3 Antigen uptake
48995741* Toll-like receptor 1 T 1 Innate antigen recognition
44804291* Toll-like receptor 6 T 1 Innate antigen recognition
61875304* Similar to Toll-like receptor 8, partial T/TD/S 4 Innate antigen recognition
463 Cathelicidin T 2 Innate antigen recognition
344 Fetuin T 1 Innate antigen recognition
508 Clathrin light chain A T 1 Innate antigen recognition
514 Clathrin light chain B TD 1 Innate antigen recognition
28195120 Gamma interferon T 1 Immune response
61864535 Predicted: Similar to CD80 antigen T 1 Immune response
163748 Transforming growth factor-beta-1
precursor
S 1 Cell proliferation
4850314 Vascular endothelial growth factor C TD 1 Cell proliferation
520521 Beta-1 integrin subunit T 3 Adhesion, integrin-mediated signaling pathway
53680685 Integrin beta-2 T 3 Adhesion, integrin-mediated signaling pathway
162819 Antigen CD18 T 18 Adhesion, integrin-mediated signaling pathway
4996449 CD34 T 1 Adhesion, integrin-mediated signaling pathway
187 Bovine CD44 T 2 Adhesion, integrin-mediated signaling pathway
33086939* Brain microvascular membrane
cofactor protein CD46
T 2 Adhesion, integrin-mediated signaling pathway
37811992* CD46 splice variant T 1 Adhesion, integrin-mediated signaling pathway
1044811 Metalloproteinase D 1 Adhesion, integrin-mediated signaling pathway
11320818 VSGP/F-spondin TD 1 Adhesion, integrin-mediated signaling pathway
20145484 SCO-spondin TD 1 Adhesion, integrin-mediated signaling pathway
2230771 Von Willebrand factor TD 1 Adhesion, integrin-mediated signaling pathway
20336613 Plakoglobin TD 1 Adhesion, integrin-mediated signaling pathway
6707084* Low-affinity IgE receptor (CD23) TD 1 Adhesion, integrin-mediated signaling pathway
162606 Actin T/TD/S 11 Actin cytoskeleton organization and biogenesis
2239063 Actin-binding protein CP3 TD 1 Actin cytoskeleton organization and biogenesis
22655316* Beta-actin T/TD 14 Actin cytoskeleton organization and biogenesis
28189611* Similar to beta actin S 1 Actin cytoskeleton organization and biogenesis
409694 Alpha skeletal actin precursor T/S 9 Actin cytoskeleton organization and biogenesis
59858375* Actin, gamma 2 T/TD/S 10 Actin cytoskeleton organization and biogenesis
2852385 14-3-3 protein gamma TD 1 Actin cytoskeleton organization and biogenesis
1838956 Capping protein, beta 3 isoform T/TD/S 10 Actin cytoskeleton organization and biogenesis
116 Beta 2 microglobulin precursor T/TD 6 Antigen presentation, immune response
28189424* Similar to beta 2-microglobulin T 3 Antigen presentation, immune response
32306980* Beta-2-microglobulin T 3 Antigen presentation, immune response
163161 70 kDa heat shock protein T 1 Response to unfolded protein
414975 70 kDa heat shock protein-2 T 1 Response to unfolded protein
28189915 Similar to 70 kDa heat shock cognate
protein
T 1 Response to unfolded protein
32364477 Heat-shock 70-kilodalton protein 1A T 1 Response to unfolded protein
32364478 Heat-shock 70-kilodalton protein 1B T 1 Response to unfolded protein
34392343 90-kDa heat shock protein alpha T 1 Response to unfolded protein
Key: GI#, Genbank accession number; DDF, Differential detergent fractionation; D, Digitonin; T, Triton X-100; TD, Tween 40-
deoxycholate; S, Sodium dodecyl sulfate; *, Proteins not annotated with GO biological process.
S.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831076
ARTICLE IN PRESS
Table
3
Proteinsrelatedto
MHC
class
IandII
alleles
identified
inbovinemonocytesbyDDF-M
udPIT
GI#
Protein
DDF
#Peptides
identified
Function
Peptidesequence
2864742
MHC
class
Iheavychain
T2
Antigen
Presentation,im
muneresponse
–.Y
FYTAVSRPGLGEPR.–
–.LLSGFTQFGYDGR.–
2864812
MHC
class
Iheavychain
T4
Antigen
Presentation,im
muneresponse
–.Y
LENGKDTLLR.–
–.Y
FYTAVSRPGLGEPR.–
–.Y
TC*HVQHEGLQEPLIL
R.–
–.W
AALVVPSGEEQR.–
3688211
MHC
class
Iantigen
T2
Antigen
Presentation,im
muneresponse
–.W
AALVVPSGEEQR.–
–.LLSGFTQFGYDGR.–
42563807
MHC
class
IIantigen
T1
Antigen
Presentation,im
muneresponse
–.H
NYQVEAPFTWQR.–
163359
MHC
lymphocyte
antigen
(DQA)
T4
Antigen
Presentation,im
muneresponse
–.VEHWGLDKPLLK.–
–.VEHWGLDKPLLK.–
–.ISYLTFLPSDDDVYDC*K.–
–.ETVWNLPLFSK.–
493485
MHC
class
IIDQ-betachain
T1
Antigen
Presentation,im
muneresponse
–.H
NYQLDAPFTWQR.–
1246367
MHC
class
IIDM
beta-chain
T1
Antigen
Presentation,im
muneresponse
–.TRPPTVQVAK.–
1256156
MHC
class
IIDM
alpha-chain
T1
Antigen
Presentation,im
muneresponse
–.LPEFADWAHQHGDTSDIM
FDK.–
163371
MHC
BoLA-D
R-alphachain
(AA
at265)
T1
Antigen
Presentation,im
muneresponse
–.FSPPVISVTWLR.–
Key:GI#,Genbankaccessionnumber;DDF,Differentialdetergentfractionation;D,Digitonin;T,TritonX-100;TD,Tween40-deoxycholate;S,Sodium
dodecylsulfate.
S.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1077

ARTICLE IN PRESS
Gi# 2864812 ---------------LTETRAGSHSLRYFYTAVSRPGLGEPRFISVGYVD 35 Gi# 3688211 MGPRALFMLLLGALVLIETRAGSHFLRYFHTAVSRPGLREPLFITVGYVD 50
* ******* ****:******** ** **:*****
DTQFVRFDSDAPDPRMEPRAPWIEKEGPEYWDRETRISKENTLVYRGSLN 85 DTQFVRFDSDARDPRKEPRQPWMEKEGPEYWDRETQISKENALWYREALN 100 *********** *** *** **:************:*****:* ** :**
NLRGYYNQSEAGSHTLQLMYGCDVGPDGRLLRGYRQDAYDGRDYLALNED 135 NLRGYYNQSEAGSHTYQRMYGCDVGPDGRLLSGFTQFGYDGRDYIALNED 150 *************** * ************* *: * .******:*****
LRSWTAADTAAQITKRKWEAEGYAESLRNYLEGRCVEGLRRYLENGKDTL 185 LRSWTAADTAAQITKRKWEAAGEAERFRNYVEGRCVEGLRRYLENGKDAL 200 ******************** * ** :***:*****************:*
LRADPPKAHVTHHSISDREVTLRCWALGFYPEEISLTWQHNGEDQTQDME 235 LRADPPKAHVTHHPISEHEVTLRCWALGFYPEEISLTWQRNGEDQTQDME 250 *************.**::*********************:**********
LVETRPSGDGTFQKWAALVVPSGEEQRYTCHVQHEGLQEPLILRWEPPQT 285 LVETRPSGDGNFQKWAALVVPSGEEQRYTCRVQHEGLQEPLTLRWEPPQT 300 **********.*******************:********** ********
SFLIMGIIVGLVLLVVAVVAGAVIWRKKRSGEKGRIYTQAASSDSTQGSD 335 SFLTMGIIVGLVLLVVAVVAGAVIWMKKHSGEKRRTYTQAASNDSAQGSD 350 *** ********************* **:**** * ******.**:****
MSLTVPKV 343 VSLTVPKV 358 :*******
Fig. 3. The identification of different MHC class I protein alleles with peptide sequences detected in MHC class I a 1 domain. Two MHC
class I heavy chain alleles (GI# 2864812 and GI# 3688211) were aligned using ClustalW (http://www.ebi.ac.uk/clustalw/) to reveal the
allelic differences between the proteins. MHC class I peptide sequences identified in bovine monocytes by DDF-MudPIT are marked with
gray background (GI# 2864812—dark gray, GI# 3688211—light gray). *, identical residues; conserved substitutions; semi-conserved
substitutions; gaps were introduced to maximize homology.
S.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831078
Several reports that characterize different TLR areavailable. In particular, it is known that TLR 1recognizes triacyl lipopeptides of bacteria andmycobacteria, and soluble factors of Neisseria
meningitis [47]. Another report suggests that diacyllipopeptides, lipoteichoic acid and zymosan, whichare originally released from Mycoplasma, grampositive bacteria and fungi, respectively, are recog-nized by TLR 6 [47]. Finally, TLR 8 recognizesimidazoquinoline, synthetic compounds, and single-stranded RNA of viruses [47]. At present, 11 TLRhave been reported in mammals [47]. Complete andpartial sequences of 10 bovine TLR are available inNCBI database. TLR 2 and TLR 6 mRNAtranscripts have been detected in bovine macro-phages and monocyte-derived DC [4]. The bovinegene, encoding the key pathogen recognitionreceptor, TLR 9 was characterized and mapped tobos taurus chromosome A22 and used to establishreal-time PCR quantification assays to measure themRNA abundances of TLR 9 in bovine mammarygland [50]. In this study, we identified for the first
time TLR 1, 6 and 8 proteins in bovine monocytes(Table 2), thus expanding the knowledge aboutinnate immune responses in cattle.
MHC genes encode class I and II molecules.Recent data indicate that MHC class I moleculesinteract with a range of natural killer (NK) cellreceptors, in addition to CD8+T cells [51]. Wefound nine proteins, including MHC class I, MHCclass II DQ alpha and beta, DM and DR alpha,related to antigen presentation (Table 3). Each ofthe MHC genes exists in an extremely large numberof allelic variations and the exact number of allelesis not known [52]. For example, GI] 2864812 andGI] 3688211 are both MHC class I heavy chainproteins, but they arose from two different MHCclass I alleles (Fig. 3). Both proteins shared the samepeptide sequence WAALVVPSGEEQR (Fig. 3).However, both MHC class I proteins had uniquepeptide sequences to confirm that peptides identifiedby DDF-MudPIT experiments arose from twodifferent MHC class I alleles (Fig. 3). The mostessential features of MHC gene regulation is that

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1079
the expression of these molecules is controlled bythe level of their transcription, and is responsive to anumber of cytokines [52]. We found the cytokineIFN -g protein, which enhances the expression ofboth MHC class I and II molecules [52].
A previous study demonstrated that humanmonocytes and DC induce CD40 independent Igclass switching through the B cell stimulator protein(BLyS) and a proliferation-inducing ligand(APRIL) in humans [8]. Furthermore, monocytesand DC upregulate BLyS and APRIL uponexposure to IFN-a, IFN-g or CD40 ligand [8]. Inthis study, we were not able to identify themolecules involved in direct monocyte-B cell inter-actions in cattle. Although recent data suggest thatbovine monocytes and monocyte-derived DC di-rectly regulate Ig secretion in peripheral blood Bcells [5,6,13], the possible bovine counterparts forAPRIL and BLyS have yet to be identified.
CD80 but not CD86 proteins have been detectedin the TX fraction of bovine monocytes (Table 2).Both CD80 and CD86 proteins are crucial forprofessional APC to provide costimulatory signalsto naıve T cells [53]. At present there is no clearmodel for understanding which ligands promote orinhibit specific immune responses [54]. Surfaceexpression of bovine CD80 and CD86moleculeshave been characterized in cattle by flow cytometrywith marker specific antibodies in monocytes andDC [5,13,55]. Low abundant molecules such asCD86 are difficult to detect at the protein levelbecause the expression of the genes encodingcostimulatory molecules is tightly regulated: theyare often expressed at low levels and for the shortperiods of time [55]. Overall, DDF-MudPIT wasvery useful to detect the low abundant cytosolicproteins in our study, suggesting high sensitivity ofthis approach.
Some of the anti-inflammatory proteins known tobe important for signaling in monocytes have notbeen identified in our study. Cytokines such asTNF-a, IL-1, IL-6, and IFN-a and b are difficult todetect on the protein level because they are oftenexpressed at low amounts and their production isefficiently triggered with bacterial products [9]. Inour study, CD14+ monocytes were obtained fromhealthy animals that most likely had not beenchallenged with bacterial infection at the time ofblood drawing. Therefore, we suggest that thesesoluble signaling proteins were downregulatedbelow the detectable level under our experimentalconditions.
3.3. Proteins related to cell adhesion
DC and their myeloid progenitors, monocytesand macrophages, serve as a bridge between theinnate and adaptive immune systems and are criticalin host protection against infectious agents. Theseprofessional APC can contribute to local sites ofinflammation where, after being activated bypathogenic molecules, they uptake antigens and/ormigrate to secondary lymphoid organs [52]. Adhe-sion molecules play an important role in themigration of monocytes from blood to the tissuesand from tissues to the lymph nodes [56]. Thismigration is mediated by adhesive interactionsbetween leukocytes and vascular or lymphaticendothelium [56]. We have identified 12 proteinsrelated to adhesion: b integrins, CD18, 34, 44 and46, metalloproteinase, vascular smooth muscle cellgrowth-promoting (VSGP/F)-spondin, subcommis-sural organ (SCO)-spondin, von Willebrand factor(vWF), plakoglobin, and the low-affinity IgEreceptor (CD23) (Table 2).
Integrins were named for their ability to link theextracellular and intracellular skeletons [57]. In ourstudy two integrins, b1 and b2 have been identifiedin the detergent fractions (Table 2). In bovinesystem, b1 integrin was characterized in bovinetrophoblasts by mRNA PCR and immunohisto-chemistry with anti-b1 integrin specific mAbs [58].In addition, specific mAbs were used to detectbovine b2 integrin expressed in neutrophils [59].Recent work has revealed that the integrin-actincytoskeleton connection is highly dynamic and is asubject to many regulatory processes [60]. Forexample, loss of b1-integrin on keratinocytes leadsto impaired as well as non-directed migrationresulting in severely delayed re-epithelializtion [60].Importantly, the CD18 antigens and b2-integrinscomprise a family of three closely related cell surfaceglycoproteins with CD11 a- and CD18 b-chains [61].Four additional CD molecules related to celladhesion were identified in this study (Table 2).One of them, CD34, a surface expressed glyco-protein [62], was detected in membrane fraction(Table 2). Bovine CD34 was characterized in bonemarrow cells based on the CD34 mRNA level [63].In this study, CD34 protein was detected andcharacterized in bovine monocytes for the first time.In humans CD34 plays a role in adhesion to thestromal microenvironment and may be involved incell transformation [62]. Other members of theadhesion protein family identified in this study were

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831080
CD44, CD46, and CD23 (Table 2). Previously,CD44 cDNA was detected in bovine [64]. CD44 is asurface receptor that has been implicated in celladhesion to a variety of matrix components andmay be associated with cytoskeletal proteins andintracellular signal pathways [65]. CD46 is acomplement regulatory protein that also functionsas the cellular receptor in humans [66]. Recentstudies indicate that CD46 may posses signalingfunctions that modulate cellular response [66]. Inaddition, cross-linking of CD46 with antibody orwith the complement activation product C3binhibited IL-12, which is critical for the generationof cell mediated and humoral immunity, by provid-ing a regulatory link between the complementsystem and cellular immune responses [66]. Impor-tantly, CD46 may act as a cellular receptor for theBVDV strains, in particular, the cytopathic strainNADL in cattle [67]. Finally, CD23 (FceRII), thelow-affinity receptor for IgE is expressed on thesurface of B cells prior to the isotype switching aswell as on monocytes, eosinophils and platelets [67].In cattle, CD23 plays a role in IgE associatedimmune responses, including allergies [68]. How-ever, it may also be important in endocytosis andadhesion because of its relation to the lectin family,which is involved in endocytosis and antigenpresentation [69]. Interestingly, metalloproteinasewas the one adhesion protein found in digitoninfraction (Table 2). Metalloproteinase is critical inpathological processes that depend on dysregulatedcell-cell and cell-matrix interactions in monocytes[69]. Metalloproteinase was also assessed by-Northern analysis and Western blotting in bovinebrain [70]. Our study confirmed that bovine mono-cytes also expressed the metalloproteinase protein.Furthermore, two spondins in bovine system,involved in protein-protein interactions, SCO-spon-din [71] and VSGP/F-spondin [72], were identifiedin our study (Table 2). VSGP/F-spondin is a majorfactor for vascular SMC proliferation promotingfactor, and vWF is a multimeric adhesive glycopro-tein with cystein-rich domains [72,73]. Finally,we found plakoglobin (g-catenin), a close homologof b-catenin, in the membrane protein fraction(Table 2). Plakoglobin shares some of the samefunctions of b-catenin [74]. Plakophilin, the memberof plakoglobin family, was examined in varioustissues and cell cultures including bovine withspecific antibodies and cDNA probes [75].
In conclusion, the data presented in this manu-script are important because the identification of the
proteins present in professional APC is instrumentalto understanding the mechanisms involved in thefunction of monocytes/macrophages and monocyte-derived DC as controllers and stimulators of T andB cell specific immune responses. In particular, thetransformation of monocytes into immature DCand their further maturation to become the mostpowerful APC is a very critical bridge betweeninnate and adaptive immune responses. In futurestudies, comparative protein profiling could be usedto elucidate the mechanisms involved in pathogeninterference of DC maturation to avoid effectiverecognition and elimination by innate and adaptiveimmune responses which are controlled and regu-lated by the most powerful professional APC, DC.Finally, the unannotated proteins will be annotatedin the future with database and literature searches toidentify homologs that are publicly released throughAgBase [76].
Acknowledments
This work was supported by the MississippiAgricultural and Forestry Experiment Station(MAFES) under project ] MIRS-13788 and theCollege of Veterinary Medicine, Mississippi StateUniversity. This paper is MAFES ] J-10791. Theauthors wish to acknowledge Dr. T. Pechan andMSU LSBI facility for the protein analysis ofbovine monocyte DDF. The authors also wish tothank Drs. C. Panuska, M. Lawrence, and L.Hanson for the helpful critique and suggestion onthe manuscript.
Appendix A. Supplementary data
Supplementary data associated with this articlecan be found in the online version at doi:10.1016/j.dci.2006.02.002.
References
[1] Medzhitov R, Preston-Hurlburt P, Janeway Jr CA. A
human homologue of the Drosophila Toll protein signals
activation of adaptive immunity. Nature 1997;388(6640):
394–7.
[2] Poltorak A, He X, Smirnova I, Liu MY, Huffel CV, Du X,
et al. Defective LPS signaling in C3H/HeJ and C57BL/
10ScCr mice: mutations in Tlr4 gene. Science 1998;
282(5396):2085–8.
[3] Krutzik SR, Sieling PA, Modlin RL. The role of Toll-like
receptors in host defense against microbial infection. Curr
Opin Immunol 2001;13(1):104–8.

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1081
[4] Werling D, Hope JC, Howard CJ, Jungi TW. Differential
production of cytokines, reactive oxygen and nitrogen by
bovine macrophages and dendritic cells stimulated with Toll-
receptor agonists. Immunol 2004;111(1):41–52.
[5] Kruger EF, Boyd BL, Pinchuk LM. Bovine monocytes
induce immunoglobulin production in peripheral blood B
lymphocytes. Dev Comp Immunol 2003;27(10):889–97.
[6] Bajer AA, Garcia-Tapia D, Jordan KR, Haas KM, Werling
D, Howard C, et al. Peripheral blood-derived bovine
dendritic cells promote IgG1-restricted B cell responses in
vitro. J Leukoc Biol 2003;73(1):100–6.
[7] Dubois B, Vanbervliet B, Fayette J, Massacrier C, van
Kooten C, Briere F, et al. Dendritic cells enhance growth
and differentiation of CD40-activated B lymphocytes. J Exp
Med 1997;185(5):941–51.
[8] Litinskiy MB, Nardelli B, Hilbert DM, He B, Schaffer A,
Casali P, et al. DCs induce CD40-independent immunoglo-
bulin class switching through BLyS and APRIL. Nat
Immunol 2002;3(9):822–9.
[9] Banchereau J, Steinman RM. Dendritic cells and the control
of immunity. Nature 1998;392(6673):245–52.
[10] Caux C, Massacrier C, Vanbervilet B, Dubois B, Durand I,
Cella M, et al. Cd34+ hematopoietic progenitors from
human cord blood differentiate along two independent
dendritic cell pathways in response to granulocyte-macro-
phage colony-stimulating factor and tumor necrosis factor
alpha: II. Functional analysis. Blood 1997;90(4):1458–70.
[11] Dauer M, Pohl K, Obermaier B, Meskendahl T, Robe J,
Schnurr M, et al. Interferon-alpha disables dendritic cell
precursors: dendritic cells derived from interferon-alpha-
treated monocytes and defective in maturation and T-cell
stimulation. Immunol 2003;110(1):38–47.
[12] Werling D, Hope JC, Chaplin P, Collins RA, Taylor G,
Howard CJ. Involvement of Caveolae in the Uptake of
Respiratory Syncytial Virus Antigen by Dendritic Cells.
J Leukoc Biol 1999;66(1):50–8.
[13] Pinchuk LM, Boyd BL, Kruger EF, Roditi I, Furger A.
Bovine dendritic cells generated from monocytes and bone
marrow progenitors regulate immunoglobulin production in
peripheral blood B cells. Comp Immunol Microbiol Infect
Dis 2003;26(4):233–49.
[14] Kadowaki NSH, Antonenko S, Malefyt RW, Kastelein RA,
Bazan F, Liu YJ. Subsets of human dendritic cell precursors
express different Toll-like receptors and respond to different
microbial antigens. J Exp Med 2001;194(6):863–9.
[15] Schnare M, Barton GM, Holt AC, Takeda K, Akira S,
Medzhitov R. Toll-like receptors control activation of
adaptive immune responses. Nat Immunol 2001;2(10):
947–50.
[16] Palliser D, Huang Q, Hacohen N, Lamontagne SP, Gullien
E, Young RA, et al. A role for Toll-like receptor 4 in
dendritic cell activation and cytolitic CD8+ T cell differ-
entiation in response to a recombinant heat shock fusion
protein. J Immunol 2004;172(5):2885–93.
[17] Hacker C, Kirsch RD, Ju X- S, Hieronymus T, Gust TC,
Kuhl C, et al. Transcriptional profiling identifies Id2
function in dendritic cell development. Nat Immunol
2003;4(4):380–6.
[18] Tureci O, Bian H, Nestle FO, Raddrizzani L, Rosinski JA,
Tassis A, et al. Cascades of transcriptional induction during
dendritic cell maturation revealed by genome-wide expres-
sion analysis. FASEB J 2003;17(8):836–47.
[19] Granucci F, Vizzardelli C, Virzi E, Rescigno M, Ricciardi-
Castagnoli P. Transcriptional reprogramming of dendritic
cells by differential stimuli. Eur J Immunol 2001;31(9):
2539–46.
[20] Huang Q, Liu D, Majewski P, Schulte LC, Korn JM, Young
RA, et al. The plasticity of dendritic cell responses to
pathogens and their components. Science 2001;294(5543):
870–5.
[21] Granucci F, Vizzardelli C, Pavelka N, Feau S, Persico M,
Virzi E, et al. Inducible IL-2 production by dendritic cells
revealed by global gene expression analysis. Nat Immunol
2001;2(9):882–8.
[22] Le Naour F, Hohenkirk L, Grolleau A, Misek DE, Lescure
P, Geiger JD, et al. Profiling changes in gene expression
during differentiation and maturation of monocyte-derived
dendritic cells using both oligonucleotide microarrays and
proteomics. J Biol Chem 2001;276(21):17920–31.
[23] Ideker T, Thorsson V, Ranish JA, Christmas R, Buhler J,
Eng JK, et al. Integrated genomic and proteomic analysis of
a systemically perturbed metabolic network. Science
2001;292(5518):929–34.
[24] Ramsby ML, Makowski GS. Differential detergent fractio-
nation of eukaryotic cells. Analysis by two-dimensional gel
electrophoresis. Methods Mol Biol 1999;112:53–66.
[25] Washburn MP, Wolters D, Yates III JR. Large-scale
analysis of the yeast proteome by multidimentional protein
identification technology. Nat Biotechnol 2001;19(3):242–7.
[26] McCarthy FM, Burgess SC, van den Berg BH, Koter MD,
Pharr GT. Differential detergent fractionation for non-
electrophoretic eukaryote cell proteomics. J Proteome Res
2005;4(2):316–24.
[27] Boyd BL, Lee TM, Kruger EF, Pinchuk LM. Cytopathic
and non-cytopathic bovine viral diarrhea virus biotypes
affect fluid phase uptake and mannose receptor-mediated
endocytosis in bovine monocytes. Vet Immunol Immuno-
pathol 2004;102(1–2):53–65.
[28] Link AJ, Eng J, Schieltz DM, Carmack E, Mize GJ, Morris
DR, et al. Direct analysis of protein complexes using mass
spectrometry. Nat Biotechnol 1999;17(7):676–82.
[29] Camon E, Barrell D, Lee V, Dimmer E, Apweiler R. The
Gene Ontology Annotation (GOA) Database—an inte-
grated resource of GO annotations to the UniProt knowl-
edgebase. In Silico Biol 2004;4(1):5–6.
[30] Washburn MP. Utilisation of proteomics datasets generated
via multidimensional protein identification technology
(MudPIT). Brief Funct Genomic Proteomic 2004;3(3):280–6.
[31] Liu H, Lin D, Yates III JR. Multidimensional separations
for protein/peptide analysis in the post-genomic era.
Biotechniques 2002;32(4):898–911.
[32] Lippolis JD, Reinhardt TA. Proteomic survey of bovine
neutrophils. Vet Immunol Immunopathol 2005;103(1–2):
53–65.
[33] Lanzavecchia A. Receptor-mediated antigen uptake and its
effect on antigen presentation to class II-restricted T
lymphocytes. Annu Rev Immunol 1990;8:773–93.
[34] Gonzalez-Gaitan M, Stenmark H. Endocytosis and signal-
ing: a relationship under development. Cell 2003;115(5):
513–21.
[35] Rabinovich GA, Baum LG, Tinari N, Paganelli R, Natoli C,
Liu F- T, et al. Galectins and their ligands: amplifiers,
silencers or tuners of the inflammatory response? Trends
Immunol 2002;23(6):313–20.

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–10831082
[36] Brooke GP, Parsons KR, Howard CJ. Cloning of two
membranes of the SIRP alpha family of protein tyrosine
phosphatase binding proteins in cattle that are expressed on
monocytes and a subpopulation of dendritic cells and which
mediate binding to CD4T cells. Eur J Immunol
1998;28(1):1–11.
[37] Smith RE, Patel V, Seatter SD, Deehan MR, Brown MH,
Brooke GP, et al. A novel MyD-1 (SIRP-1a) signaling
pathway that inhibits LPS-induced TNFa production by
monocytes. Blood 2003;102(7):2532–40.
[38] Sallusto F, Lancavecchia A. Efficient Presentation of
Soluble Antigen by Cultured Human Dendritic Cells is
Maintained by Granulocyte/Macrophage Colony-Stimulat-
ing Factor Plus Interleukin 4 and Down Regulated by
Tumor Necrosis Factor. J Exp Med 1994;179(4):1109–18.
[39] Stahl PD. The mannose receptor and other macrophage
lectins. Curr Opin Immunol 1992;4(1):49–52.
[40] Ezekowitz RA, Sastry K, Bailly P, Warner A. Molecular
characterization of the human macrophage mannose recep-
tor: demonstration of multiple carbohydrate recognition-like
domains and phagocytosis of yeasts in Cos-1 cells. J Exp
Med 1990;172(6):1785–94.
[41] Reis e Sousa C, Stahl PD, Austyn JM. Phagocytosis of
antigens by Langerhans cells in vitro. J Exp Med
1993;178(2):509–19.
[42] Watts C, Marsh M. Endocytosis: what goes in and how?
J Cell Sci 1992;103(Pt 1):1–8.
[43] Racoosin EL, Swanson JA. Macrophage colony-stimulating
factor (rM-CSF) stimulates pinocytosis in bone marrow-
derived macrophages. J Exp Med 1989;170(5):1635–48.
[44] Racoosin EL, Swanson JA. M-CSF-induced macropinocy-
tosis increases solute endocytosis but not receptor-mediated
endocytosis in mouse macrophages. J Cell Sci 1992;102(Pt
4):867–80.
[45] West MA, Bretscher MS, Watts C. Distinct endocytic
pathways in EGF-stimulated human carcinoma A431 cells.
J Cell Biol 1989;109(6 Pt 1):2731–9.
[46] Janeway Jr CA, Medzhitov R. Innate immune recognition.
Annu Rev Immunol 2002;20:197–216.
[47] Akira S, Takeda K. Toll-like receptor signalling. Nat Rev
Immunol 2004;4(7):499–511.
[48] Scocchi M, Wang S, Zanetti M. Structural organization of
the bovine cathelicidin gene family and identification of a
novel member. FEBS Lett 1997;417(3):311–5.
[49] Dziegielewska KM, Brown WM, Casey SJ, Christie DL,
Foreman RC, Hill RM, et al. The complete cDNA and
amino acid sequence of bovine fetuin. Its homology with
alpha 2HS glycoprotein and relation to other members of
the cystatin superfamily. J Biol Chem 1990;265(8):4354–7.
[50] Goldammer T, Zerbe H, Molenaar A, Schuberth H- J,
Brunner RM, Kata SR, et al. Mastitis increases mammary
mRNA abundance of b-defensin 5, toll-like-receptor 2
(TLR2), and TLR4 but not TLR9 in cattle. Clin Diagn
Lab Immunol 2004;11(1):174–85.
[51] Trowsdale J. Genetic and functional relationships between
MHC and NK receptor genes. Immunity 2001;15(3):363–74.
[52] Pinchuk G. The major histocompatibility complex. In:
Schaum’s outlines immunology. New York, NY: McGraw-
Hill; 2002. p. 77–93.
[53] Thery C, Duban L, Segura E, Veron P, Lantz O, Amigorena
S. Indirect activation of naıve CD4+Tcells by dendritic cell-
derived exosomes. Nature Immunol 2002;3(12):1156–62.
[54] Sansom DM, Manzotti CN, Zheng Y. What’s the difference
between CD80 and CD86? Trends Immunol 2003;24(6):
314–9.
[55] Norimatsu M, Harris J, Chance V, Dougan G, Howard CJ,
Villarreal-Ramos B. Differential response of bovine mono-
cyte-derived macrophages and dendritic cells to infection
with Salmonella typhimurium in a low-dose model in vitro.
Immunol 2003;108:55–61.
[56] Imhof BA, Aurrand-Lions M. Adhesion mechanisms
regulating the migration of monocytes. Nat Rev Immunol
2004;4(6):432–44.
[57] Blystone SD. Integrating an integrin: a direct route to actin.
Biochim Biophys Acta 2004;1692(2-3):47–54.
[58] MacLaren LA, Wildeman AG. Fibronectin receptors in
preimplantation development: cloning, expression, and
localization of the alpha 5 and beta 1 integrin subunits in
bovine trophoblast. Biol Reprod 1995;53(1):153–65.
[59] Nagahata H, Higuchi H, Teraoka H, Takahashi K, Inanami
O, Kuwabara M. Decreased apoptosis of beta 2- integrin-
deficient bovine neutrophils. Immunol Cell Biol 2004;82(1):
32–7.
[60] Grose R, Hutter C, Bloch W, Thorey I, Watt FM, Fassler R,
et al. A crucial role of X1 integrins for keratinocyte migration
in vitro and during cutaneous wound repair. Development
2002;129(9):2303–15.
[61] Van Amersfoort ES, Van Berkel TJ, Kuiper J. Receptors,
mediators, and mechanisms involved in bacterial sepsis and
septic shock. Clin Microbiol Rev 2003;16(3):379–414.
[62] Healy L, May G, Gale K, Grosveld F, Greaves M, Enver T.
The stem cell antigen CD34 functions as a regulator of
hemopoietic cell adhesion. Proc Natl Acad Sci USA
1995;92(26):12240–4.
[63] Zhou JH, Hikono H, Ohta M, Sakurai M. Cloning of bovine
CD34 cDNA. J Vet Med Sci 2001;63(9):1051–3.
[64] Bosworth BT, St John T, Gallatin WM, Harp JA. Sequence
of the bovine CD44 cDNA: comparison with human and
mouse sequences. Mol Immunol 1991;28(10):1131–5.
[65] Vivers S, Dransfield I, Hart SP. Role of macrophage CD44
in the disposal of inflammatory cell corpses. Clin Sci
2002;103(5):441–9.
[66] Karp CL, Wysocka M, Wahl LM, Ahearn JM, Cuomo PJ,
Sherry B, et al. Mechanism of suppression of cell-mediated
immunity by measles virus. Science 1996;273(5272):228–31.
[67] Maurer K, Krey T, Moennig V, Thiel H- J, Rumenapf T.
CD46 is a cellular receptor for bovine viral diarrhea virus.
J Virol 2004;78(4):1792–9.
[68] Watson JL, Jackson KA, king DP, Stott JL. Molecular
cloning and sequencing of the low-affinity IgE receptor
(CD23) for horse and cattle. Vet Immunol Immunopathol
2000;73(3–4):323–9.
[69] Galt SW, Lindemann S, Medd D, Allen LL, Kraiss LW,
Harris ES, et al. Differential regulation of matrix metallo-
proteinase-9 by monocytes adherent to collagen and plate-
lets. Circ Res 2001;89(6):509–16.
[70] Howard L, Lu X, Mitchell S, Griffiths S, Glynn P.
Molecular cloning of MADM: a catalytically active mam-
malian disintegrin-metalloprotease expressed in various cell
types. Biochem J 1996;317(Pt 1):45–50.
[71] Gobron S, Creveaux I, Meiniel R, Didier R, Herbet A, Bamdad
M, et al. Subcommissural organ/reissner’s fiber complex:
characterization of SCO-Spondin, a glycoprotein with potent
activity on neurite outgrowth. Glia 2000;32(2):177–91.

ARTICLE IN PRESSS.-R. Lee et al. / Developmental and Comparative Immunology 30 (2006) 1070–1083 1083
[72] Miyamoto K, Morishita Y, Yamazaki M, Minamino N,
Kangawa K, Matsuo H, et al. Isolation and characterization
of vascular smooth muscle cell growth promoting factor
from bovine ovarian follicular fluid and its cDNA cloning
from bovine and human ovary. Arc Biochem Biophys
2001;390(1):93–100.
[73] Janel N, Ribba A- S, Cherel G, Kerbiriou-Nabias D, Meyer
D. Primary structure of the propeptide and factor VIII-
binding domain of bovine von willebrand factor. Biochim
Biophys Acta 1997;1339(1):4–8.
[74] Zhurinsky J, Shtutman M, Ben-Ze’ev A. Plakoglobin and b-catenin: protein interactions, regulation and biological roles.
J Cell Sci 2000;113(Pt18):3127–39.
[75] Heid HW, Schmidt A, Zimbelmann R, Schafer S, Winter-
Simanowski S, Stumpp S, et al. Cell type-specific desmoso-
mal plaque proteins of the plakoglobin family: plakophilin 1
(band 6 protein). Differentiation 1994;58(2):113–31;
Heid HW, Schmidt A, Zimbelmann R, Schafer S, Winter-
Simanowski S, Stumpp S, Keith M, Figge U, Schnolzer M,
Franke WW. Cell type-specific desmosomal plaque proteins
of the plakoglobin family: plakophilin 1 (band 6 protein).
Differentiation 2005;4(2):316–24.
[76] Bridges SM, McCarthy FM, Luthe DS, Wang B, Nanduri B,
Burgess SC. Targeted gene ontology annotation databases
for agriculture. Fifth annual gene ontoligy users meeting.
Bergen, Norway. September 14–15, 2005.
Top Related
Copyright © 2022 FDOKUMEN

