







Bahasa
Halaman
Hukum


Ž .Biochimica et Biophysica Acta 1345 1997 327–337
Contrasting adrenergic effects on lipoprotein lipase gene expression inthe brown adipose tissue of intact mice and in cultured brown adipocytes
from mice
Pertti Kuusela a, Anders Jacobsson a, Martin Klingenspor b, Stefan Rehnmark a,Gerhard Heldmaier b, Barbara Cannon a, Jan Nedergaard a,)
a The Wenner-Gren Institute, The Arrhenius Laboratories F3, Stockholm UniÕersity, S-106 91 Stockholm, Swedenb Fachbereich Biologie, Philipps-UniÕersitat, D-3550 Marburg, Germany¨
Received 4 December 1996; accepted 9 January 1997
Abstract
Ž .To examine the regulation of lipoprotein lipase LPL gene expression, LPL mRNA levels in the brown adipose tissue ofintact mice and in mouse brown adipocyte cultures were examined. In intact mice, exposure to cold resulted in a rapid,
Ž .transient, 5-fold increase in LPL mRNA level. Norepinephrine NE injection could fully mimic the effect of acute exposureto cold, and LPL mRNA and enzymatic activity were increased in parallel after NE injection. These results indicatedpositive adrenergic control of LPL gene expression in the brown adipose tissue of intact mice. In cultured mouse brownadipocytes, the level of spontaneously expressed LPL mRNA decreased in parallel with the progression of brown adipocytedifferentiation. NE treatment of undifferentiated cells led to a decrease in LPL mRNA levels. In brown adipocytes that hadreached a mature state, NE had a small negative or no effect on LPL mRNA levels, irrespective of whether the experimentwas performed in the presence or absence of insulin or of newborn-calf serum. It was concluded that LPL gene expressionin brown adipose tissue in intact mice is under adrenergic control but that this gene is not under positive adrenergic controlin cultured brown adipocytes from mice, although these cells are otherwise adrenergically sensitive. The presence ofadditional factors may be necessary to confer adrenergic sensitivity to the LPL gene in the cultured brown adipocytes;alternatively, cells other than the mature brown adipocytes may confer the positive adrenergic sensitivity to the brownadipose tissue depots in situ. q 1997 Elsevier Science B.V. All rights reserved.
Ž .Keywords: Brown adipose tissue; Lipoprotein lipase; Norepinephrine; Gene expression regulation; Transcriptional control; Mouse
1. Introduction
In white adipose tissue, the expression of the geneŽ .for lipoprotein lipase LPL is under positive control
) Corresponding author: Fax: q46 8 156756. E-mail:[email protected]
of insulin, while adrenergic stimulation, if anything,w xhas an inhibitory effect on expression 1–3 . In con-
trast to this, we and others have earlier observed inbrown adipose tissue of rats that expression of the
w xenzyme is under positive adrenergic control 4 . Inw xrecent studies on rat brown-fat cells in culture 5 , we
have demonstrated that this is due to a direct effect ofŽ .norepinephrine NE on the rat brown-fat cells them-
0005-2760r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0005-2760 97 00008-8

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337328
selves. This may be considered physiologically mean-ingful, in that the same physiological stimulus thatincreases the utilization of fatty acids for thermogen-
w xesis in the brown adipocyte 6–9 also increases thesupply to the cell of fatty acids for combustion. Thepositive adrenergic control of LPL gene expressionthus also constitutes one of the qualitatively distin-guishing features between brown adipocytes and white
w xadipocytes 10 and indicates the occurrence of aramification point during the development of thesetwo types of adipocytes.
These conclusions concerning the control of LPLgene expression in rat brown adipocytes would beexpected to have general validity in other species,such as, e.g., the mouse. Also in the mouse, LPL
Žactivity increases in the tissue in the cold i.e., during. w xphysiological activation of thermogenesis 11 . It
would therefore be expected that LPL gene expres-sion also in mice would be under positive adrenergic
w xcontrol. However, in earlier preliminary studies 12we were unable to observe a positive effect of NE onLPL gene expression in mouse brown-fat cells inculture. Similarly, in brown adipocyte-like cell linesof mouse origin, no consistent positive effect of NEor other adrenergic agents on LPL gene expression
w xhas been reported 13,14 . Thus, the possibility ex-isted that expression of the LPL gene in mice wouldnot be under adrenergic control.
We have therefore found it of importance to exam-ine whether the physiologically induced increase inLPL activity in mice is due to an increase in LPLmRNA level or whether translational or posttrans-lational mechanisms must be invoked – and whetherLPL gene expression at all can be induced via anadrenergic pathway in the intact mouse. Further, wehave examined whether it is possible to observepositive adrenergic control of LPL gene expression inbrown adipocyte cultures.
2. Materials and methods
2.1. Experiments in intact mice
ŽAll experiments were performed with mice age.f5 weeks; body weight f25 g of the NMRI
outbred strain, obtained from a local supplierŽ .Eklunds . For all experiments in intact mice, themice were preacclimated to 288C for f1 week.
Acclimation experiments. The mice were placedŽ .two per cage at 48C cold-exposed or remained at
288C for the times indicated. The mice had freeŽaccess to food R3, Ewos; 26 energy % as protein,
.13% as fat and 61% as carbohydrate and water witha 6Lr18D light cycle. The acclimation experimentwas initiated early in the light phase. Further detailson experimental conditions, mouse growth, RNApreparation and slot blot techniques used can be
w xfound in 15 . For the measurements of LPL mRNAw xlevels, the LPL cDNA probe described in 16 was
nick-translated with a kit from Bethesda Researchw xLaboratories and hybridised as described in Ref. 15
for UCP.Acute experiments. On the day of experiment,
during the early light phase, mice were either injectedi.p. with saline or with NE as indicated in the figurelegends, or were left untreated, at 288C, or were
Ž .moved to 48C. Two or four hours later as indicated ,the mice were killed, and the interscapular brownadipose tissue depot dissected out. The tissue wasfrozen on dry ice or in liquid nitrogen and was storedat y808C until analysis.
For measurement of LPL activity, the rate of re-14 w 14 xlease of C-oleate from glyceryl tri 1- C oleate was
w xfollowed, as described previously 17 . Acetone-ethertissue extracts were incubated in a gum arabic-stabi-lized triglyceride emulsion. Liberated oleate was sep-
w xarated in a liquid–liquid partition system 18 . In afinal volume of 80 ml the assay contained 140 mM
Ž . w 14 xTris-HCl pH 8.6 , 5.1 mM glyceryl tri 1- C oleateŽ .32 000 dpm , 12 mM CaCl , 2% bovine serum2
albumin, 10% human serum, 0.4% gum arabic, 5USP units heparinrml " 1 M NaCl. The LPL activ-ity was calculated as the salt-sensitive lipolytic activ-ity.
For measurement of LPL mRNA levels, RNA wasextracted and Northern blots for LPL mRNA were
w xrun as described elsewhere 5 .
2.2. Experiments with mouse cell cultures
Brown adipocyte cultures were initiated from theinterscapular brown adipose tissue depots of 3- to4-week-old mice and grown in culture in DMEMmedium in the presence of 10% newborn-calf serum,4 nM insulin and 10 mM Hepes and with 50 IU
( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337 329
penicillin, 50 mg streptomycin and 25 mg sodiumw xascorbate per ml, as described earlier 19,20 .
For the individual experiments, the cell cultureswere treated as described in the legends to figures. Ifnot otherwise indicated, RNA was prepared by guani-dine-HCl extraction, and slot blot assays for LPL
w xmRNA were performed as earlier described 19,15 .In each experiment, each data point was routinely
Ž .obtained as the mean of 2 –4 -culture wells, equallytreated.
3. Results
3.1. Regulation of LPL gene expression in the brownadipose tissue of intact mice
3.1.1. Effect of acclimation to cold on LPL geneexpression in brown adipose tissue
In order to examine whether LPL gene expressionin brown adipose tissue could be physiologicallyinduced in mice, we followed the LPL mRNA levelduring cold acclimation. After preacclimation to 288CŽi.e., a temperature close to thermoneutrality for this
.species , the mice were transferred to 48C and theLPL mRNA levels in the brown adipose tissue werefollowed.
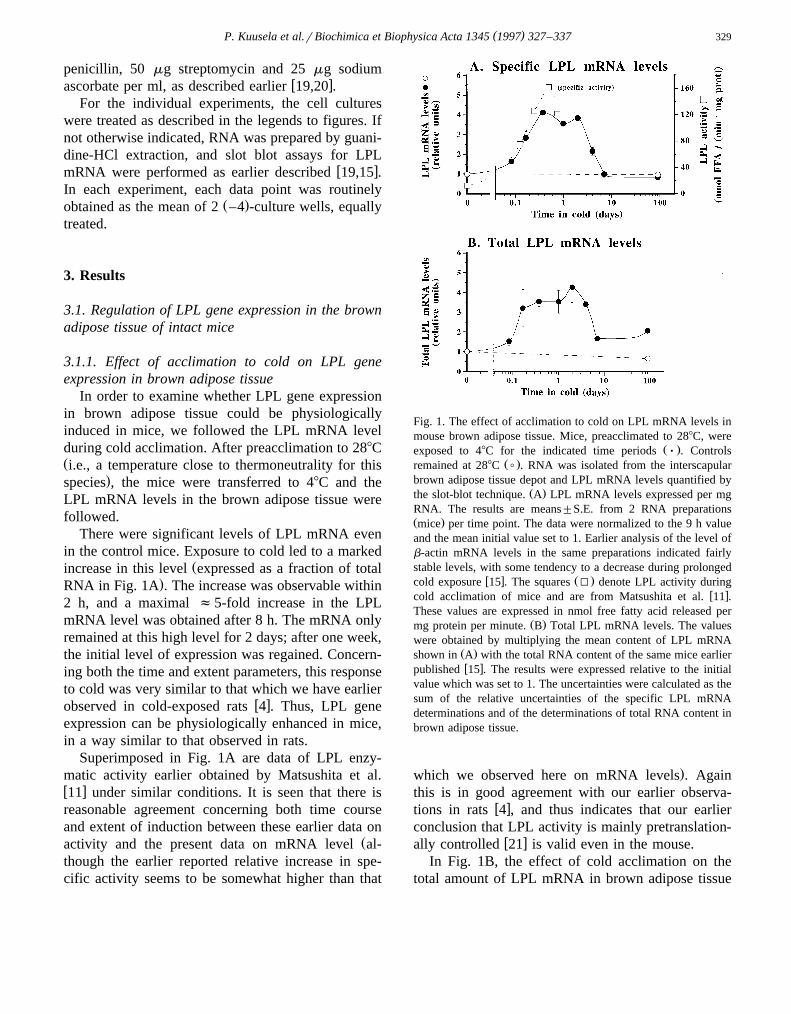
There were significant levels of LPL mRNA evenin the control mice. Exposure to cold led to a marked
Žincrease in this level expressed as a fraction of total.RNA in Fig. 1A . The increase was observable within
2 h, and a maximal f5-fold increase in the LPLmRNA level was obtained after 8 h. The mRNA onlyremained at this high level for 2 days; after one week,the initial level of expression was regained. Concern-ing both the time and extent parameters, this responseto cold was very similar to that which we have earlier
w xobserved in cold-exposed rats 4 . Thus, LPL geneexpression can be physiologically enhanced in mice,in a way similar to that observed in rats.
Superimposed in Fig. 1A are data of LPL enzy-matic activity earlier obtained by Matsushita et al.w x11 under similar conditions. It is seen that there isreasonable agreement concerning both time courseand extent of induction between these earlier data on
Žactivity and the present data on mRNA level al-though the earlier reported relative increase in spe-cific activity seems to be somewhat higher than that
Fig. 1. The effect of acclimation to cold on LPL mRNA levels inmouse brown adipose tissue. Mice, preacclimated to 288C, were
Ž .exposed to 48C for the indicated time periods Ø . ControlsŽ .remained at 288C ( . RNA was isolated from the interscapular
brown adipose tissue depot and LPL mRNA levels quantified byŽ .the slot-blot technique. A LPL mRNA levels expressed per mg
RNA. The results are means"S.E. from 2 RNA preparationsŽ .mice per time point. The data were normalized to the 9 h valueand the mean initial value set to 1. Earlier analysis of the level ofb-actin mRNA levels in the same preparations indicated fairlystable levels, with some tendency to a decrease during prolonged
w x Ž .cold exposure 15 . The squares I denote LPL activity duringw xcold acclimation of mice and are from Matsushita et al. 11 .
These values are expressed in nmol free fatty acid released perŽ .mg protein per minute. B Total LPL mRNA levels. The values
were obtained by multiplying the mean content of LPL mRNAŽ .shown in A with the total RNA content of the same mice earlierw xpublished 15 . The results were expressed relative to the initial
value which was set to 1. The uncertainties were calculated as thesum of the relative uncertainties of the specific LPL mRNAdeterminations and of the determinations of total RNA content inbrown adipose tissue.
.which we observed here on mRNA levels . Againthis is in good agreement with our earlier observa-
w xtions in rats 4 , and thus indicates that our earlierconclusion that LPL activity is mainly pretranslation-
w xally controlled 21 is valid even in the mouse.In Fig. 1B, the effect of cold acclimation on the
total amount of LPL mRNA in brown adipose tissue
( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337330
is shown. The successive decrease in LPL mRNAŽlevel when expressed as a fraction of total RNA
Ž ..Fig. 1A was partly compensated by the increase inw xtotal RNA levels in the tissue 15 . Thus, the total
LPL mRNA amount remained increased during pro-longed cold exposure.
3.1.2. Effect of NE injection on LPL gene expressionand actiÕity in brown adipose tissue
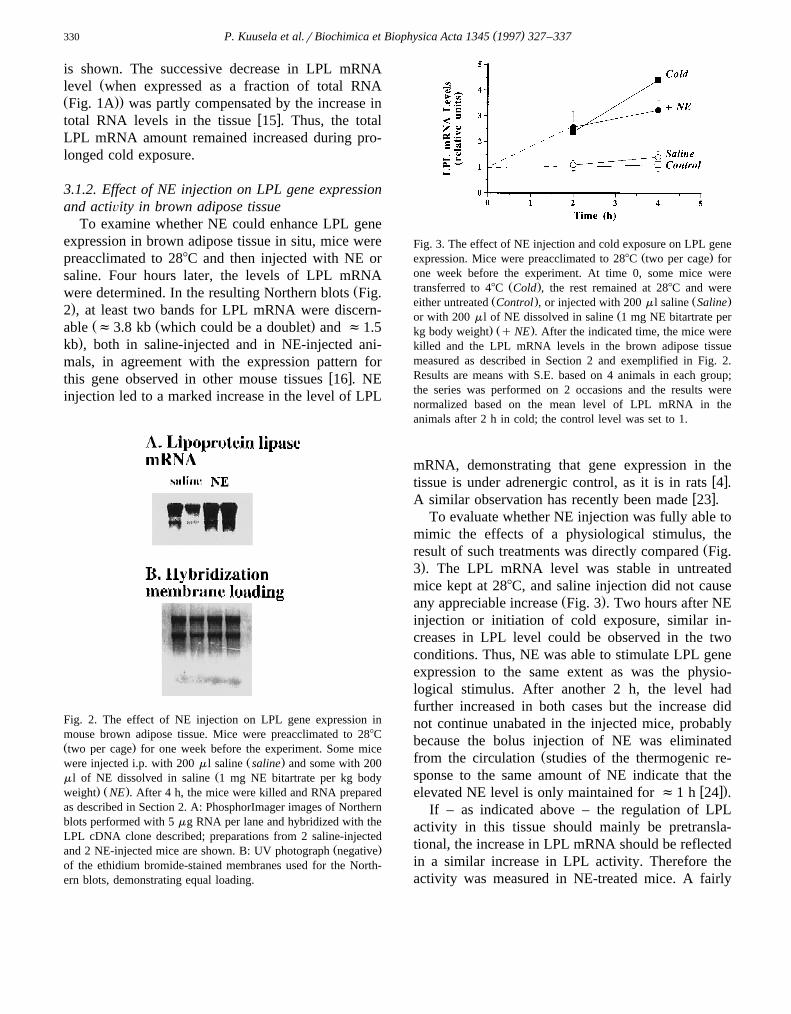
To examine whether NE could enhance LPL geneexpression in brown adipose tissue in situ, mice werepreacclimated to 288C and then injected with NE orsaline. Four hours later, the levels of LPL mRNA
Žwere determined. In the resulting Northern blots Fig..2 , at least two bands for LPL mRNA were discern-
Ž Ž .able f3.8 kb which could be a doublet and f1.5.kb , both in saline-injected and in NE-injected ani-
mals, in agreement with the expression pattern forw xthis gene observed in other mouse tissues 16 . NE
injection led to a marked increase in the level of LPL
Fig. 2. The effect of NE injection on LPL gene expression inmouse brown adipose tissue. Mice were preacclimated to 288CŽ .two per cage for one week before the experiment. Some mice
Ž .were injected i.p. with 200 ml saline saline and some with 200Žml of NE dissolved in saline 1 mg NE bitartrate per kg body
. Ž .weight NE . After 4 h, the mice were killed and RNA preparedas described in Section 2. A: PhosphorImager images of Northernblots performed with 5 mg RNA per lane and hybridized with theLPL cDNA clone described; preparations from 2 saline-injected
Ž .and 2 NE-injected mice are shown. B: UV photograph negativeof the ethidium bromide-stained membranes used for the North-ern blots, demonstrating equal loading.
Fig. 3. The effect of NE injection and cold exposure on LPL geneŽ .expression. Mice were preacclimated to 288C two per cage for
one week before the experiment. At time 0, some mice wereŽ .transferred to 48C Cold , the rest remained at 288C and were
Ž . Ž .either untreated Control , or injected with 200 ml saline SalineŽor with 200 ml of NE dissolved in saline 1 mg NE bitartrate per
. Ž .kg body weight q NE . After the indicated time, the mice werekilled and the LPL mRNA levels in the brown adipose tissuemeasured as described in Section 2 and exemplified in Fig. 2.Results are means with S.E. based on 4 animals in each group;the series was performed on 2 occasions and the results werenormalized based on the mean level of LPL mRNA in theanimals after 2 h in cold; the control level was set to 1.
mRNA, demonstrating that gene expression in thew xtissue is under adrenergic control, as it is in rats 4 .
w xA similar observation has recently been made 23 .To evaluate whether NE injection was fully able to
mimic the effects of a physiological stimulus, theŽresult of such treatments was directly compared Fig.
.3 . The LPL mRNA level was stable in untreatedmice kept at 288C, and saline injection did not cause
Ž .any appreciable increase Fig. 3 . Two hours after NEinjection or initiation of cold exposure, similar in-creases in LPL level could be observed in the twoconditions. Thus, NE was able to stimulate LPL geneexpression to the same extent as was the physio-logical stimulus. After another 2 h, the level hadfurther increased in both cases but the increase didnot continue unabated in the injected mice, probablybecause the bolus injection of NE was eliminated
Žfrom the circulation studies of the thermogenic re-sponse to the same amount of NE indicate that the
w x.elevated NE level is only maintained for f1 h 24 .If – as indicated above – the regulation of LPL
activity in this tissue should mainly be pretransla-tional, the increase in LPL mRNA should be reflectedin a similar increase in LPL activity. Therefore theactivity was measured in NE-treated mice. A fairly

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337 331
Table 1A comparison between the effect of NE injection on LPL enzy-matic activity and mRNA levels in brown adipose tissue of mice
Saline NE NErsalinebLPL mRNA 1.4"0.2 3.2"0.4 2.3"0.6
aLPL activity 14"3 26"4 1.9"0.7
Ž .Mice at 288C were injected as described in Section 2 and in theŽlegends to Fig. 2 and Fig. 3 60 ml injections were used for the
.activity measurements , and the LPL activity and mRNA levelsin the brown adipose tissue were determined. The values formRNA are the 4-h values from Fig. 3 and are thus based on 4mice. The values for activity are the means"S.E. of 6 mice andare expressed as nmol oleate released per min per mg protein. a
b Žand indicate significant effects of NE treatment P -0.05 and.P -0.01, respectively . The uncertainties given for the ratio
between NE-injected and saline-injected mice were calculated asthe sum of the relative uncertainties of each data set.
similar increase in mRNA level and enzymatic activ-Žity could be observed 4 h after NE injection Table
.1 . A tendency to a somewhat higher increase inŽ . Ž .mRNA level G2-fold than in activity F2-fold
may be related to the inherent delay between mRNAlevels and protein synthesis in a transient such as thisw x25,15 . The data are therefore consistent with theidea that regulation of LPL activity occurs predomi-nantly pretranslationally in this species.
3.2. Regulation of LPL gene expression in culturedbrown adipocytes
3.2.1. Effect of differentiation progression on LPLgene expression in brown adipocytes cultures
Based on the data above, it was clear that NE wasable to induce increased expression of the LPL genein brown adipose tissue. However, it was not evidentfrom this, which cell type in the tissue partook in theincrease.
To examine this, we used our culture system forw xmouse brown adipocytes 12,19,20,26–32 . In this,
undifferentiated precursors from the stromal-vascularfraction of the interscapular brown adipose tissue ofyoung mice are isolated and transferred to culturewhere the cells spontaneously advance in their degreeof differentiation and acquire characteristics that re-semble those observed in brown adipocytes differen-tiated in situ.
The effect of the progression in differentiationstate on spontaneous LPL gene expression in thesecells is shown on the Northern blot in Fig. 4A. Only
Ž .one band f3.6 kb hybridized with the LPL cDNAprobe; this is different from what was observed in the
Ž .preparations from intact mice Fig. 2 but in agree-ment with what has been observed in brown
w xadipocyte-like cell lines derived from mice 14,33 .The LPL mRNA level fell continuously throughout
Ž .the maturation period Fig. 4AB . Expression of theLPL gene is considered an early marker for differen-
Žtiation in developing white adipocytes or white-. w xadipocyte-like cell lines 34,35 ; the high expression
seen in the brown adipocytes on day 3 is in agree-ment with such a concept. Such a tendency to adecrease in basal LPL gene expression may also beobservable in spontaneously differentiating brown
w xadipocytes from Djungarian hamsters 36 , and LPLenzymatic activity also declines with time in cultures
w xof mouse brown adipocytes 37 .That the cells progressed in their differentiation
concomitantly with the decline in LPL mRNA levelsis demonstrated by the ability of the cells to respondto NE with an induction of expression of the brown-
Ž .fat-specific uncoupling protein thermogenin UCPŽ .Fig. 4C . Practically no spontaneous expression of
Ž .UCP was observed not shown . The addition of NEled to a vast increase in UCP mRNA level in the
Ž .mature cells at day 5 and later but had only a verysmall effect on the days preceding this developmental
Ž . Žswitch Fig. 4C all in agreement with earlier obser-w x.vations 19 .
However, despite our observations of NE-inducedexpression of the LPL gene in the brown adipose
Ž .tissue of intact mice Figs. 2 and 3 , and despite thedemonstrated ability of the brown adipocyte culturesto respond to NE addition by an increased level of
Ž .gene expression for the UCP gene Fig. 4C , theeffects of NE on LPL gene expression in the culturedbrown adipocytes were very small and in general
Ž .more inhibitory than stimulatory Fig. 4D .
3.2.2. Effects of NE in young cellsTo substantiate the apparently inhibitory effect of
ŽNE on LPL gene expression in young undifferenti-. Ž .ated brown adipocytes Fig. 4D , studies were made
with cells on day 4, i.e., before the developmentalswitch had turned the cells into mature brown
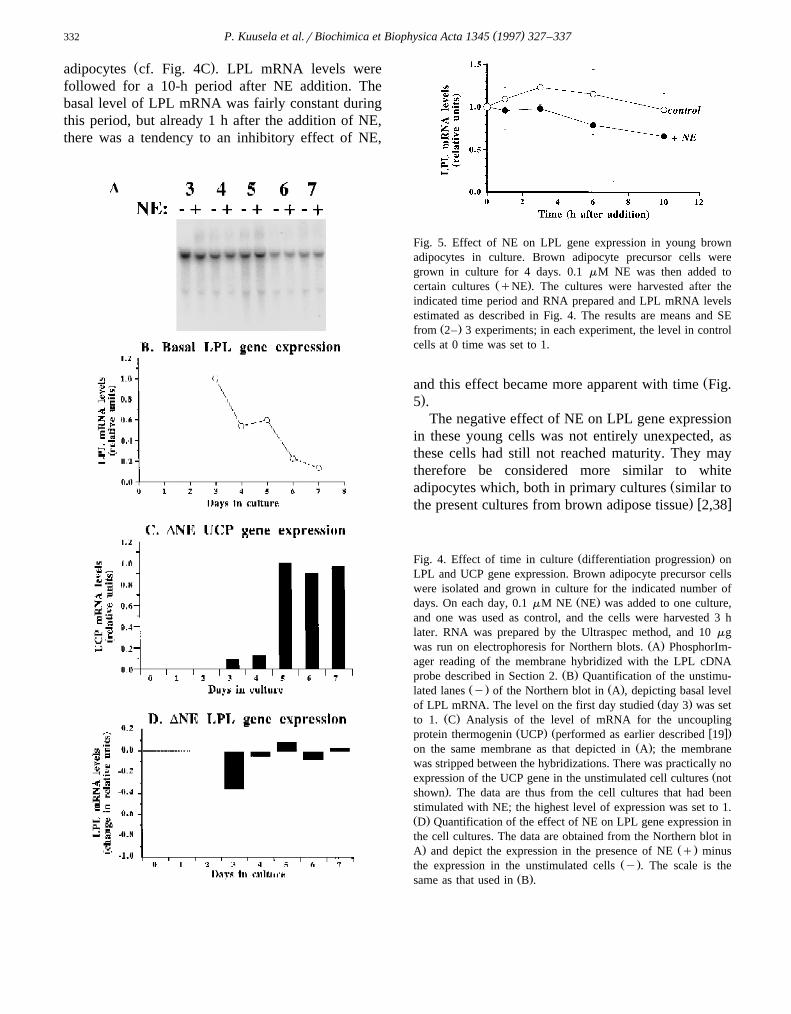
( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337332
Ž .adipocytes cf. Fig. 4C . LPL mRNA levels werefollowed for a 10-h period after NE addition. Thebasal level of LPL mRNA was fairly constant duringthis period, but already 1 h after the addition of NE,there was a tendency to an inhibitory effect of NE,
Fig. 5. Effect of NE on LPL gene expression in young brownadipocytes in culture. Brown adipocyte precursor cells weregrown in culture for 4 days. 0.1 mM NE was then added to
Ž .certain cultures qNE . The cultures were harvested after theindicated time period and RNA prepared and LPL mRNA levelsestimated as described in Fig. 4. The results are means and SE
Ž .from 2– 3 experiments; in each experiment, the level in controlcells at 0 time was set to 1.
Žand this effect became more apparent with time Fig..5 .
The negative effect of NE on LPL gene expressionin these young cells was not entirely unexpected, asthese cells had still not reached maturity. They maytherefore be considered more similar to white
Žadipocytes which, both in primary cultures similar to. w xthe present cultures from brown adipose tissue 2,38
Ž .Fig. 4. Effect of time in culture differentiation progression onLPL and UCP gene expression. Brown adipocyte precursor cellswere isolated and grown in culture for the indicated number of
Ž .days. On each day, 0.1 mM NE NE was added to one culture,and one was used as control, and the cells were harvested 3 hlater. RNA was prepared by the Ultraspec method, and 10 mg
Ž .was run on electrophoresis for Northern blots. A PhosphorIm-ager reading of the membrane hybridized with the LPL cDNA
Ž .probe described in Section 2. B Quantification of the unstimu-Ž . Ž .lated lanes y of the Northern blot in A , depicting basal level
Ž .of LPL mRNA. The level on the first day studied day 3 was setŽ .to 1. C Analysis of the level of mRNA for the uncoupling
Ž . Ž w x.protein thermogenin UCP performed as earlier described 19Ž .on the same membrane as that depicted in A ; the membrane
was stripped between the hybridizations. There was practically noŽexpression of the UCP gene in the unstimulated cell cultures not
.shown . The data are thus from the cell cultures that had beenstimulated with NE; the highest level of expression was set to 1.Ž .D Quantification of the effect of NE on LPL gene expression inthe cell cultures. The data are obtained from the Northern blot in. Ž .A and depict the expression in the presence of NE q minus
Ž .the expression in the unstimulated cells y . The scale is theŽ .same as that used in B .
( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337 333
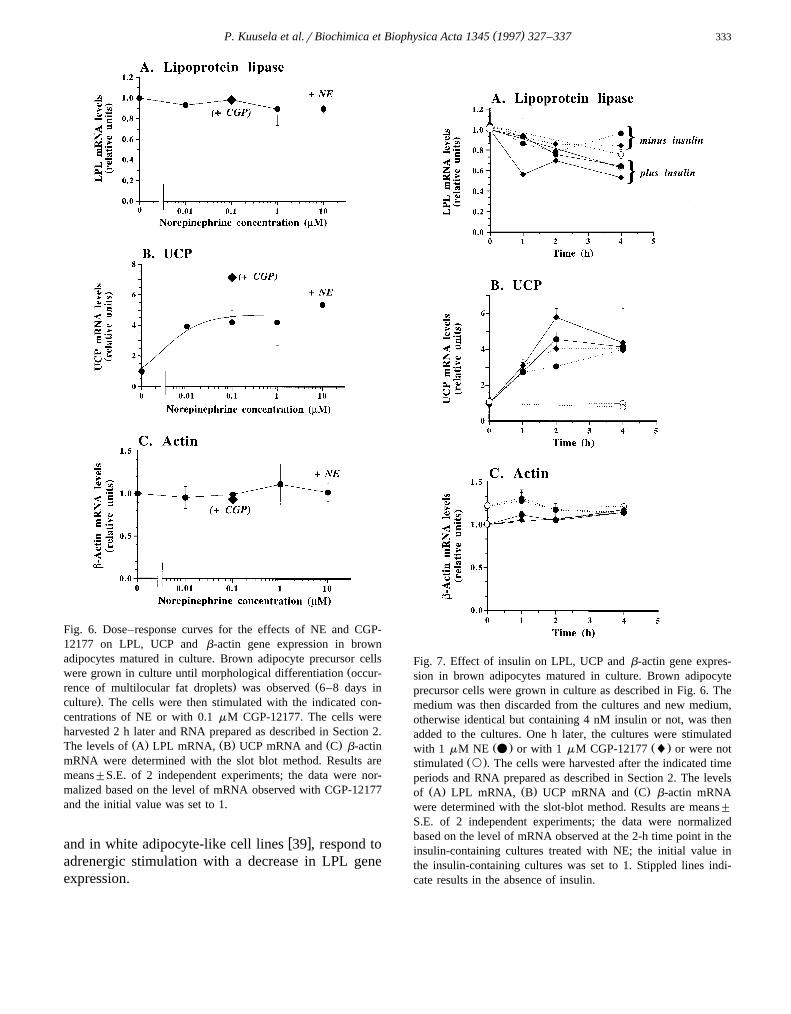
Fig. 6. Dose–response curves for the effects of NE and CGP-12177 on LPL, UCP and b-actin gene expression in brownadipocytes matured in culture. Brown adipocyte precursor cells
Žwere grown in culture until morphological differentiation occur-. Žrence of multilocular fat droplets was observed 6–8 days in
.culture . The cells were then stimulated with the indicated con-centrations of NE or with 0.1 mM CGP-12177. The cells wereharvested 2 h later and RNA prepared as described in Section 2.
Ž . Ž . Ž .The levels of A LPL mRNA, B UCP mRNA and C b-actinmRNA were determined with the slot blot method. Results aremeans"S.E. of 2 independent experiments; the data were nor-malized based on the level of mRNA observed with CGP-12177and the initial value was set to 1.
w xand in white adipocyte-like cell lines 39 , respond toadrenergic stimulation with a decrease in LPL geneexpression.
Fig. 7. Effect of insulin on LPL, UCP and b-actin gene expres-sion in brown adipocytes matured in culture. Brown adipocyteprecursor cells were grown in culture as described in Fig. 6. Themedium was then discarded from the cultures and new medium,otherwise identical but containing 4 nM insulin or not, was thenadded to the cultures. One h later, the cultures were stimulated
Ž . Ž .with 1 mM NE v or with 1 mM CGP-12177 l or were notŽ .stimulated ` . The cells were harvested after the indicated time
periods and RNA prepared as described in Section 2. The levelsŽ . Ž . Ž .of A LPL mRNA, B UCP mRNA and C b-actin mRNA
were determined with the slot-blot method. Results are means"
S.E. of 2 independent experiments; the data were normalizedbased on the level of mRNA observed at the 2-h time point in theinsulin-containing cultures treated with NE; the initial value inthe insulin-containing cultures was set to 1. Stippled lines indi-cate results in the absence of insulin.

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337334
3.2.3. Effects of NE in differentiated cellsIf NE should have a stimulatory effect on LPL
gene expression in the brown adipocytes, this maynot become apparent until the cells had reached a
Žmature state i.e., in cells cultured for more than 5.days, cf. Fig. 4C . However, in the initial studies
Ž .Fig. 4D , we did not see any clear positive effects ofNE on LPL gene expression, even in the maturebrown adipocytes. This could be due to less thanoptimal experimental conditions for this particulargene. We therefore varied the experimental condi-tions to investigate whether a positive effect could berevealed.
Dose–response. The dosage of NE tested abovecould be less than optimal; however, no dose of NE,from 10 nM to 10 mM, was able to enhance LPL
Ž .gene expression in these cultures Fig. 6A . Also thew xb -specific agonist CGP-12177 40 was without ef-3
fect. As expected, these agents were able to induceŽ .UCP gene expression Fig. 6B , and they were with-
Ž .out effect on b-actin expression Fig. 6C .Insulin. All the above experiments were conducted
in the presence of insulin. This could affect theresults: insulin could in itself stimulate LPL geneexpression to such an extent that NE could notfurther increase expression, or insulin could, throughits effects on e.g., the cAMP-phosphodiesterase,counteract any cAMP-dependent adrenergic effectsw x41 . However, acute omission of insulin only had asmall effect on LPL gene expression but, if anything,
Ž .led to a slightly higher level Fig. 7A . Further, evenin the absence of insulin, NE and CGP-12177 werestill unable to induce LPL gene expression. This wasnot due to a general insensitivity to adrenergic stim-uli: the response to UCP was only slightly lower in
Ž .the absence of insulin than in its presence Fig. 7B .Effect of acute serum omission. The above experi-
ments were all conducted in the presence of 10%newborn-calf serum. Inhibitory factors for LPL gene
Žexpression may be present in this serum cf. that suchfactors inhibit LPL activity in white adipocyte-like
w x.cell lines 42 . Albumin present in the serum mayalso influence LPL gene expression, e.g., by binding
Žfatty acids that have been shown to influence LPLw x.gene expression in certain cell lines 43 . However,
there was no measurable effect of acute serum omis-sion, neither on the basal LPL mRNA level, nor on
Ž .the level after NE addition Fig. 8 . In one of the
Fig. 8. Effect of acute serum omission on LPL gene expression inbrown adipocytes matured in culture. Brown adipocyte precursorcells were grown in culture as described in Fig. 6. The mediumwas then discarded from the cultures and new medium, otherwiseidentical but containing 10% newborn-calf serum or not, wasthen added to the cultures. One hour later, the cultures were
Ž . Ž .stimulated with 1 mM NE v, B or not (, I . ( and v
indicate results from cell cultures in the presence of serum; I
and B in the absence of serum. The cells were harvested afterthe indicated time periods and RNA prepared as described inSection 2. The levels of LPL mRNA were determined with theslot-blot method. Results are means"S.E. of 4 independentexperiments; in each experiment, the level of mRNA observed atthe 0-h time point in the serum-containing control cultures wasset to 1.
experimental series we checked whether acute serumomission would affect the ability of NE to induce
Ž .UCP expression. This was not the case not shown .Thus, in a variety of conditions in which NE was
able to induce UCP expression, NE failed to have apositive effect on LPL gene expression.
4. Discussion
In the present investigation, we have studied theregulation of LPL gene expression in the brownadipose tissue of mice and in mouse brown adipocytesdeveloping in culture. The in vivo and in vitro resultsmay be contrasted with each other and related towhat has been observed in other species.
4.1. LPL gene expression in intact mice
We found that in the brown adipose tissue of intactmice, LPL gene expression could be induced by aphysiological stimulation in the form of cold expo-sure. This increase in LPL mRNA levels could largelyexplain the increase in LPL enzymatic activity earlier

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337 335
observed in mouse brown adipose tissue during coldw xacclimation 11 . The early phase of physiological
induction of gene expression could be fully mimickedby NE injection, which also led to a commensurateincrease in enzymatic activity. The implication is thusthat in the mouse, the physiologically induced in-crease in LPL activity is mediated via activation ofthe sympathetic nervous system and a release of NEin the brown adipose tissue that in its turn leads to anincrease in LPL gene expression and through this toincreased synthesis of the LPL protein and a nearly
Žparallel increase in LPL activity i.e., activity is.primarily transcriptionally regulated .
In all these respects, the results obtained in theintact mouse are fully paralleled with those we have
w xearlier observed in the rat 4,21,44 . In other species,the regulation is not necessarily identical. Indeed,during the early phase of cold acclimation in theDjungarian hamster, LPL mRNA levels, protein lev-els and activity did not change in parallel, implying
w xmultiple sites of regulation 45 . However, even inthis species, in the resulting steady-state followingchronic cold acclimation, all three measures had in-
w xcreased in parallel 45 .
4.2. LPL gene expression in cultured brownadipocytes
In the cultured mouse brown adipocyte precursorcells, the basal expression of LPL was high and itdecreased with progressing differentiation. This is inaccordance with enhanced LPL gene expression be-ing an early marker of differentiation during adipocyte
w xdevelopment 34,35 .We found, however, that NE was unable to in-
crease expression in the cultured cells, irrespective ofthe degree of differentiation of the cells, the NEconcentration used, or the presence or absence ofinsulin or newborn-calf serum. The absence of apositive effect of NE on LPL gene expression earlierobserved in brown adipocyte-like cell lines from
w xmice 13,14 can therefore not be ascribed to a differ-ence in cellular behaviour between the immortalizedcell lines and brown adipocytes developed from pre-cursors directly obtained from the tissue, nor can it beascribed to a general adrenergic insensitivity of thegenes in the primary cultures of mouse brown
Žadipocytes, as these cells in other respects UCP gene.expression responded well to adrenergic stimulation.
There is a principal difference between the presentresults and those we have recently obtained in cul-
w xtured rat brown adipocytes 5 . In the rat cells, NEstimulation leads to a marked increase in the level ofLPL gene expression, mainly mediated via b -adren-3
oceptors and an increase in cAMP levels. Thus, thecultured rat brown adipocytes show the responseexpected from the in vivo experiments referred toabove. This is, however, clearly not the case for themouse brown adipocytes studied in the present inves-tigation.
4.3. Differences between LPL gene expression in ÕiÕoand in Õitro
Our observations here exclude the possibility thatthe absence of a positive response to adrenergicstimulation in cultured mouse brown adipocytes isdue to a species difference between rats and mice inthe regulation of LPL gene expression in the brownadipose tissue: in both species, in the tissue in theintact animal, the expression may be induced in vivoby a NE injection.
We have thus no simple explanation for the differ-ence between the regulation of LPL gene expressionin intact mice and in cell cultures. It may be notedthat this difference in regulation is parallelled by adifference in the transcript length pattern: in theintact animal, two LPL mRNA species are found, in
Ž .the cell cultures only one cf. Figs. 2 and 4 . It hasbeen discussed, at least concerning human LPL geneexpression, that such differences in transcript length
w xpattern may not be trivial 46 . The differences atleast imply that there are principal differences be-tween the control of LPL gene expression in the invivo and in the in vitro systems.
Two possibilities for the difference between the invivo and the in vitro data may be considered: that theabsence of adrenergic sensitivity in the cultures is atechnical artefact or that the result is principallycorrect: the mouse brown adipocytes are unable toincrease LPL gene expression due to adrenergic stim-ulation.
It is of course impossible to ensure that someŽdetails e.g., the absence of additional permissive
.factors in the culturing conditions may specifically

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337336
influence the control of LPL gene expression so thatthe NE signal is not transmitted to the gene. In otherrespects these cell cultures have, however, proven tobe good experimental models of the events occurringin brown adipocytes in the intact animal and in thisway they have led to new insights into the control ofbrown adipocyte development and differentiation.Still, the possibility remains that the culturing condi-tions can explain the absence of effect of NE on LPLgene expression.
An alternative suggestion would be that the con-clusions from the cell culture experiments are correct:that it is not the brown adipocyte itself that is respon-sible for the adrenergically induced increase in LPLgene expression observed in mouse brown adiposetissue. There are other cell types in the tissue, such asendothelial cells and other interstitial cells, and itcannot presently be excluded that it is these cells thatare the responsive ones. Indeed, in a tissue which inseveral respects is metabolically similar to brownadipose tissue, i.e., the heart, a discussion concerningthe identity of the cells expressing the gene for LPL
w xis ongoing 47,48 . It would therefore be of interest toexamine by in situ hybridization techniques the cellu-lar localization of the enhanced LPL gene expressionin the brown adipose tissue of adrenergically stimu-lated mice.
Acknowledgements
This study was supported by grants from theSwedish Natural Science Research Council and theDeutsche Forschungsgemeinschaft. The authors thankM.C. Schotz and T.G. Kirchgessner for access to theLPL cDNA clone, Sigrid Stohr for technical assis-¨tance concerning the LPL activity assays, and Mag-nus Fredriksson and Birgitta Leksell for preparationand analysis of certain RNA samples.
References
w x1 Ong, J.M., Kirchgessner, T.G., Schotz, M.C. and Kern, P.A.Ž .1988 J. Biol. Chem. 263, 12933–12938.
w x2 Raynolds, M.V., Awald, P.D., Gordon, D.F., Gutierrez-Hartmann, A., Rule, D.C., Wood, W.M. and Eckel, R.H.Ž .1990 Mol. Endocrinol. 4, 1416–1422.
w x Ž .3 Enerback, S. and Gimble, J.M. 1993 Biochim. Biophys.¨Acta 1169, 107–125.
w x4 Mitchell, J.R.D., Jacobsson, A., Kirchgessner, T.G., Schotz,Ž .M.C., Cannon, B. and Nedergaard, J. 1992 Am. J. Physiol.
263, E500–E506.w x5 Kuusela, P., Rehnmark, S., Jacobsson, A., Cannon, B. and
Ž .Nedergaard, J. 1997 Biochem. J. 321, 759–767.w x Ž .6 Nedergaard, J. and Lindberg, O. 1982 Int. Rev. Cytol. 74,
187–286.w x Ž . Ž .7 Trayhurn, P. and Nicholls, D.G. eds. 1986 Brown Adi-
pose Tissue, Edward Arnold, London.w x Ž .8 Himms-Hagen, J. 1990 in Thermoregulation: Physiology
Ž .and Biochemistry Schonbaum, E. and Lomax, P., eds. , pp.¨327–414, Pergamon Press, New York.
w x Ž . Ž9 Himms-Hagen, J. 1992 in Obesity Bjorntorp, P. and¨.Brodoff, B.N., eds. , pp. 15–34, J.B. Lippincott, Philadel-
phia.w x10 Nedergaard, J., Herron, D., Jacobsson, A., Rehnmark, S.
Ž .and Cannon, B. 1995 Int. J. Dev. Biol. 39, 827–837.w x Ž .11 Matsushita, H., Kobayashi, K. and Kusumi, H. 1980 Adv.
Physiol. Sci. 32, 523–526.w x12 Rehnmark, S., Kopecky, J., Jacobsson, A., Nechad, M.,´
Herron, D., Nelson, B.D., Obregon, M.J., Nedergaard, J.Ž .and Cannon, B. 1989 Exp. Cell Res. 182, 75–83.
w x13 Ross, S.R., Choy, L., Graves, R.A., Fox, N., Solevjeva, V.,Ž .Klaus, S., Ricquier, D. and Spiegelman, B.M. 1992 Proc.
Natl. Acad. Sci. USA 89, 7561–7565.w x14 Klaus, S., Champigny, O., Cassard-Doulcier, A.-M., Choy,
Ž .L., Spiegelman, B. and Ricquier, D. 1994 in Obesity inŽEurope 1993 Ditschuneit, H., Gries, F.A., Hauner, H.,
.Schusdziarra, V. and Wechsler, J.G., eds. , pp. 65–72, JohnLibbey, London.
w x15 Jacobsson, A., Muhleisen, M., Cannon, B. and Nedergaard,¨Ž .J. 1994 Am. J. Physiol. 267, R999–R1007.
w x16 Kirchgessner, T.G., Svenson, K.L., Lusis, A.J. and Schotz,Ž .M.C. 1987 J. Biol. Chem. 262, 8463–8466.
w x17 Klingenspor, M., Klaus, S., Wiesinger, H. and Heldmaier,Ž .G. 1989 Am. J. Physiol. 257, R1123–R1127.
w x Ž .18 Belfrage, P. and Vaughan, M. 1969 J. Lipid Res. 10,341–344.
w x19 Rehnmark, S., Nechad, M., Herron, D., Cannon, B. and´Ž .Nedergaard, J. 1990 J. Biol. Chem. 265, 16464–16471.
w x Ž .20 Bronnikov, G., Houstek, J. and Nedergaard, J. 1992 J.Biol. Chem. 267, 2006–2013.
w x Ž .21 Carneheim, C.M.H., Nedergaard, J. and Cannon, B. 1988Am. J. Physiol. 254, E155–E161.
w x Ž .23 Trayhurn, P., Duncan, J.S. and Rayner, D.V. 1995Biochem. J. 311, 729–733.
w x24 Sadurskis, A., Dicker, A., Cannon, B. and Nedergaard, J.Ž .1995 Am. J. Physiol. 269, E351–E360.
w x Ž .25 Hargrove, J.L. and Schmidt, F.H. 1989 FASEB J. 3,2360–2370.
w x26 Herron, D., Rehnmark, S., Nechad, M., Loncar, D., Cannon,´Ž .B. and Nedergaard, J. 1990 FEBS Lett. 268, 296–300.
w x27 Kopecky, J., Baudysova, M., Zanotti, F., Janıkova, D.,´ ´ ´Ž .Pavelka, S. and Houstek, J. 1990 J. Biol. Chem. 265,
22204–22209.w x28 Puigserver, P., Herron, D., Gianotti, M., Palou, A., Cannon,
Ž .B. and Nedergaard, J. 1992 Biochem. J. 284, 393–398.

( )P. Kuusela et al.rBiochimica et Biophysica Acta 1345 1997 327–337 337
w x29 Rehnmark, S., Antonson, P., Xanthopoulos, K.G. and Jacob-Ž .sson, A. 1993 FEBS Lett. 318, 235–241.
w x30 Pico, C., Herron, D., Palou, A., Jacobsson, A., Cannon, B.´Ž .and Nedergaard, J. 1994 Biochem. J. 302, 81–86.
w x31 Thonberg, H., Zhang, S.-J., Tvrdik, P., Jacobsson, A. andŽ .Nedergaard, J. 1994 J. Biol. Chem. 269, 33179–33186.
w x32 Uchida, Y., Irie, K., Tsukahara, F., Ohba, K.-i., Ogawa, A.,Ž .Fujii, E. and Muraki, T. 1995 Eur. J. Pharmacol. 291,
33–41.w x33 Klaus, S., Choy, L., Champigny, O., Cassard-Doulcier, A.-
Ž .M., Ross, S., Spiegelman, B. and Ricquier, D. 1994 J. CellSci. 107, 313–319.
w x Ž .34 Doglio, A., Dani, C., Grimaldi, P. and Ailhaud, G. 1986Biochem. J. 238, 123–129.
w x35 Ailhaud, G., Abumrad, N., Amri, E.-Z. and Grimaldi, P.-A.Ž . Ž1994 in Obesity in Europe 93 Ditschuneit, H., Gries,F.A., Hauner, H., Schusdziarra, V. and Wechsler, J.G.,
.eds. , pp. 51–57, John Libbey, London.w x Ž .36 Klaus, S., Cassard-Doulcier, A.-M. and Ricquier, D. 1991
J. Cell Biol. 115, 1783–1790.w x37 Forest, C., Doglio, A., Casteilla, L., Ricquier, D. and Ail-
Ž .haud, G. 1987 Exp. Cell Res. 168, 233–246.w x Ž .38 Ong, J.M., Saffari, B., Simsolo, R.B. and Kern, P.A. 1992
Mol. Endocrinol. 6, 61–69.
w x Ž .39 Antras, J., Lasnier, F. and Pairault, J. 1991 Mol. Cell.Endocrinol. 82, 183–190.
w x Ž .40 Mohell, N. and Dicker, A. 1989 Biochem. J. 261, 401–405.w x Ž .41 Nedergaard, J. and Lindberg, O. 1979 Eur. J. Biochem. 95,
139–145.w x42 Pradines-Figueres, A., Barcellini-Couget, S., Dani, C., Bau-`
Ž .doin, C. and Ailhaud, G. 1990 Biochem. Biophys. Res.Commun. 166, 1118–1125.
w x43 Amri, E.-Z., Teboul, L., Vannier, C., Grimaldi, P.-A. andŽ .Ailhaud, G. 1996 Biochem. J. 314, 541–546.
w x Ž .44 Carneheim, C., Nedergaard, J. and Cannon, B. 1984 Am.J. Physiol. 246, E327–E333.
w x45 Klingenspor, M., Ebbinghaus, C., Hulshorst, G., Stohr, S.,¨ ¨Ž .Spiegelhalter, F., Haas, K. and Heldmaier, G. 1996 J.
Lipid Res. 37, 1685–1695.w x46 Ranganathan, G., Ong, J.M., Yukht, A., Saghizadeh, M.,
Ž .Simsolo, R.B., Pauer, A. and Kern, P.A. 1995 J. Biol.Chem. 270, 7149–7155.
w x47 Stein, O., Stein, Y., Schwartz, S.P., Reshef, A., Chajek-Shaul, T., Ben-Naim, M., Friedman, G. and Leitersdorf, E.Ž .1991 Arteriosclerosis Thrombosis 11, 857–863.
w x48 O’Brien, K.D., Ferguson, M., Gordon, D., Deeb, S.S. andŽ .Chait, A. 1994 Arterioscler. Thromb. 14, 1445–1451.
Top Related
Copyright © 2022 FDOKUMEN

