







Bahasa
Halaman
Hukum


This article was downloaded by: [Ingenta Content Distribution (Publishing Technology)]On: 15 October 2014, At: 05:54Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Avian PathologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/cavp20
Comparison of the immune responses againstSalmonella enterica serovar Gallinarum infectionbetween naked neck chickens and a commercialchicken lineMaría Teresa Alvarez a b , Nestor Ledesma c , Guillermo Téllez c d , José Luis Molinari e &Patricia Tato ba Laboratorio de Parasitología , Facultad de Biología Universidad Michoacana de SanNicolás de Hidalgo , Michoacán, Moreliab Departamento de Microbiología y Parasitología, Facultad de Medicina , UniversidadNacional Autónoma de México , Universidad, 04510 3000 Avc Departamento de Producción Animal: Aves, Facultad de Medicina Veterinaria yZootecnia , Universidad Nacional Autónoma , Fayetteville, Méxicod Department of Poultry Science , University of Arkansas , Fayetteville, AR, 72701e Departamento de Genética Molecular , Instituto de Fisiología Celular UniversidadNacional Autónoma , MéxicoPublished online: 17 Jun 2010.
To cite this article: María Teresa Alvarez , Nestor Ledesma , Guillermo Téllez , José Luis Molinari & Patricia Tato (2003)Comparison of the immune responses against Salmonella enterica serovar Gallinarum infection between naked neckchickens and a commercial chicken line, Avian Pathology, 32:2, 193-203, DOI: 10.1080/0307945021000071605
To link to this article: http://dx.doi.org/10.1080/0307945021000071605
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Comparison of the immune responses againstSalmonella enterica serovar Gallinarum infectionbetween naked neck chickens and a commercialchicken line
Marıa Teresa Alvarez1,2, Nestor Ledesma
3, Guillermo Tellez
3,4, Jose Luis
Molinari5and Patricia Tato
2
1Laboratorio de Parasitologıa, Facultad de Biologıa, Universidad Michoacana de San Nicolas de Hidalgo,Morelia, Michoacan, Mexico, 2Departamento de Microbiologıa y Parasitologıa, Facultad de Medicina,Universidad Nacional Autonoma de Mexico, Av. Universidad 3000, 04510 Mexico, DF, 3Departamento deProduccion Animal: Aves, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autonoma deMexico, 4Department of Poultry Science, University of Arkansas, Fayetteville AR 72701, USA and5Departamento de Genetica Molecular, Instituto de Fisiologıa Celular, Universidad Nacional Autonoma deMexico, Mexico
The immune responses of indigenous naked neck (NaNa and Nana) and normally feathered (nana) chickensagainst a Salmonella Gallinarum (SG) infection were evaluated and compared with those of a commercial line
(B-380). Groups of 28-day-old chickens (NaNa , Nana , nana , and B-380) were immunized orally and
subcutaneously with 50 mg of SG antigens. Control non-immunized animals were inoculated with sterile saline
solution. All chickens were challenged with 1 LD50 of SG and mortality was recorded daily for 20 days.
Antibodies to SG were measured in sera before immunization, before the challenge, 10 days after the challenge,
and at sacrifice. Peripheral blood lymphocyte proliferation assays were performed using concanavalin A and
SG antigens. Results showed that non-immunized Nana chickens exhibited the best natural resistance to
Salmonella infection, since only 30% of them died. In contrast, all control B-380 chickens died by the 13th day.
Immunization with SG induced immunity in chickens of all genotypes. Indigenous naked neck and normally
feathered chickens showed a higher survival rate when compared with B-380 chickens. Immunized Nanachickens showed the highest antibody titres (PB/0.05) as well as the highest thymidine incorporation in
peripheral blood lymphocytes stimulated with con A or SG antigens (PB/0.05). The results show that Nanachickens are the most resistant to SG infection and the best responders to vaccination with SG antigens.
Introduction
The naked neck birds have received great attentionfor broiler production, because of their associationwith heat tolerance (Merat, 1986). It has beenreported that the reduction of feather coverageprovides relative heat tolerance and therefore, in
high ambient temperature, heterozygous nakedneck broilers are superior to their normally feath-ered counterparts (Cahaner et al ., 1993; Yalcin etal ., 1997; Deeb & Cahaner, 1999). The naked neckbroilers have exhibited higher food intake, growthrate, and meat yield than their normally feathered
*To whom correspondence should be addressed.
Tel.: �/52 55 5623-2388. Fax: �/52 55 5623-2382. E-mail: [email protected]
Received 1 July 2002. Provisionally accepted 22 August 2002. Accepted 15 November 2002
Avian Pathology (2003) 32, 193�/203
ISSN 0307-9457 (print)/ISSN 1465-3338 (online)/03/020193-11 # 2003 Houghton Trust LtdDOI: 10.1080/0307945021000071605
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

counterparts at normal and high temperatures(El-Attar & Merat, 1985; Cahaner et al ., 1993;Deeb & Cahaner, 1999). However, immune re-sponses against infectious agents have not beenstudied in these birds.Salmonella enterica serovar Gallinarum causes
the severe systemic disease known as fowl typhoid(FT) in domestic poultry. Specificity of the diseaseis expressed in vivo primarily at the level of thereticuloendothelial system (Barrow et al ., 1994).FT is host specific, causing a mortality rate of 50%or higher in the domestic fowl (Shivaprasad, 1997,2000). It is a septicaemic disease that basicallyaffects adult and growing chickens, althoughchicks are also affected (Wray et al ., 1996). Fowltyphoid is rare in countries with a modern poultryindustry; it has been eradicated in developedcountries and very recently in Mexico. In spite ofthe application of hygienic measures, serologicaltesting and slaughter of positive reactors (Barrow,1993), this disease has gained incidence in SouthAmerica and other countries throughout Africaand Asia (Lucio et al ., 1985; Silva, 1985; Mdegelaet al ., 2000).
The importance of genetic control in theresistance of infectious diseases is well recognized.It has been suggested that some strains of chickensare more genetically susceptible or resistantto infection with Salmonella than others (Benjaminet al ., 1991). It has been described that chicksare highly susceptible to Salmonella infectionduring the first 4 days post hatch, becomingincreasingly resistant to infection with age,such resistance requires cell-mediated immunity(Ziprin et al ., 1989). Differences in resistanceagainst Salmonella infections among differentcommercial lines (Kaiser et al ., 1998; Mdegelaet al ., 2000) and in inbred chicken lines havebeen reported (Bumstead & Barrow, 1993;Berchieri et al ., 2001). The host geneticbackground is known to have a considerable effecton the susceptibility to acute infection withSalmonella Gallinarum (SG) both in in-bred(Bumstead & Barrow, 1993) and commercial lines(Berchieri et al ., 2000). Recently, SAL 1 andNRAMP 1 genes have been related to Salmonel-losis resistance in fowls (Berchieri et al ., 2001;Mariani et al ., 2001; Girard-Santosuosso et al .,2002).
Immune responses against Salmonella infectionin naked neck chickens have not been explored.The first aim of this study was to study thedifferences in resistance to Salmonella infection(innate immunity) in homozygotic and heterozy-gotic naked neck (NaNa and Nana) chickens, andnormally feathered (nana and B-380) male chick-ens. The second aim was to evaluate and comparethe immune responsiveness against SG antigens inthe same genotypes.
Material and Methods
Animals and husbandry
Trials were conducted using one-day-old male homozygotic (NaNa )
and heterozygotic (Nana ) naked neck, and normally feathered indi-
genous (nana ) chickens, which were obtained from local flocks of
indigenous hens in Michoacan, Mexico. One-day-old male Babcock 380
(B-380) chickens were obtained from a commercial hatchery. The flocks
were Salmonella free as detected by serology and culturing methods
according to Davies et al . (1997). Twenty eight-day-old chickens in
groups of 10 animals each were used in all experiments. Groups I and
V, indigenous homozygotic naked neck (NaNa ) chickens; groups II and
VI, indigenous heterozygotic naked neck (Nana ) chickens; groups III
and VII, indigenous normally feathered chickens (nana ); and groups IV
and VIII, commercial line B-380 chickens. Groups I, II, III, and IV
were immunized with a SG antigen at day 28 of age, and a booster was
given 14 days later. Groups V, VI, VII and VIII received no
immunization and were used as control groups. The birds were
individually weighed and wing-banded before housing them in different
electrically heated commercial brooder batteries in isolation units
located within the Avian Medicine Department of the Faculty of
Veterinary Medicine, National Autonomous University of Mexico.
They were provided with a balanced unmedicated ration and water ad
libitum . Individual body weights of each chicken were registered on
days 28, 52, 62 and 72. Birds from immunized and non-immunized
groups were challenged with SG at day 52 (10 days after the booster).
Birds that survived were sacrificed 20 days after the challenge.
Bacteria and antigen preparation
A primary poultry isolate of SG (field strain OR-2) was selected for
resistance to nalidixic acid and maintained on nutrient agar. The
medium used to culture the resistant isolate in experimental studies
contained 20 mg/ml nalidixic acid to inhibit the growth of other
bacteria. The inoculum for challenge was prepared in sterile phosphate-
buffered saline. The viable cell concentration of the inoculum was
determined by counting the colony forming units (CFU) on brilliant
green agar plates. Salmonella stocks were maintained in casein-peptone
soy meal-peptone agar (Merck). Bacterial antigens were obtained
according to the method described by Tato et al . (1979). Briefly, SG
was grown in casein-peptone soy meal-peptone broth (Merck) and,
after 5 h of incubation at 378C, cells were harvested by centrifugation at
1100 g for 15 min at 48C in a Sorvall RC-2B centrifuge. The pellet was
suspended in cold acetone previously dehydrated with calcium chloride.
The suspension was stirred for 15 min and filtered using a glass filter.
After repeating this procedure twice, cells were cleaned with cold
chloroform and left at 48C until chloroform residues were evaporated.
Bacterial powder was suspended in sodium phosphate buffer (0.01 M,
pH 7.2) (PBS), containing magnesium chloride 0.012 M, sucrose 0.25
M, sodium deoxycholate 0.4% and potassium chloride 0.1 M.
Deoxyribonuclease (Sigma) was added to a final concentration of 20
mg/ml. After 1 h of incubation at 388C with constant stirring, the
material was dialysed for 3 days with frequent changes of PBS. The
dialysed mixture was centrifuged at 10,200 g for 15 min in a
refrigerated Sorvall centrifuge. The supernatant was carefully decanted,
filtered through a 0.22 mm Millipore membrane and lyophilized.
Protein concentration was determined according to the method of
Bradford (1976). This antigenic preparation contained proteins of the
whole bacteria including outer membrane proteins.
Immunizations and challenge of chickens
Chickens were immunized, by oral and subcutaneous routes, at day 28
of age with 50 mg protein/0.1 ml isotonic saline solution for each route.
A booster was given 14 days later in the same manner. Ten days after
the booster, all chickens were challenged orally with 108 CFU 1 (LD50)
of the virulent bacilli (strain OR-2) suspended in 0.2 ml PBS. Clinical
signs and mortality were recorded daily for 20 days.
Determination of specific IgY antibodies
Blood samples (1.0 ml) were obtained from the brachial vein of each
chick before the first immunization (28 days old), at the challenge (52
194 M.T. Alvarez et al.
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

days old), ten days after the challenge, and immediately before the
sacrifice (72 days old). Sera were separated by centrifugation (1150 g
for 10 min), aliquoted, and stored at �/48C. Specific IgY antibody titres
were determined by micro-agglutination test conducted in 96 rounded
(U-shaped) well plates (Costar, Cambridge, Mass) using a commercial
Salmonella antigen kit and according to the manufacturer’s instructions
(Salsbury Laboratories, Charles City, Iowa).
Proliferation assays
Lymphocyte proliferation assays were carried out according to
Arechavaleta et al . (1998), which allowed us to evaluate cell-mediated
immunity in all the groups at different times. Blood was obtained from
the brachial vein in an heparinized syringe, afterwards it was diluted
using an equal volume of Hank’s balanced saline solution (HBSS) and
homogenized. Blood was carefully deposited in a gradient with 70 and
80% of Percoll (Pharmacia) and centrifuged to 1150 g for 30 min at
room temperature as has been previously reported (Mills & Wilcox,
1993; Desmidt et al ., 1996). Cells were washed twice with HBSS by
centrifugation (1150 g for 10 min) and the pellet was suspended in
RPMI medium containing 10% fetal bovine serum and 10 UI/ml
penicillin, and 10 mg/ml streptomycin (complete RPMI medium). Cell
viability was determined using the Trypan blue staining and cell
suspensions were adjusted to 6�/106 cells/ml. Aliquots containing
5�/105 cells were plated in triplicate in 96-well plates (Costar, Cam-
bridge, Mass). Cells incubated in complete RPMI medium were
included as controls in each group. Twenty microlitres of (Con A) (5
mg/ml) were added to each well and incubated (378C, 5% CO2 for 2
days). In other experiments, SG antigens (2.5 mg of protein) were added
to each well and plates were incubated in the same conditions for 7
days. Sixteen hours before harvesting, 1 mCi [3H]-thymidine (45Ci/
mmol; Amersham Pharmacia Biotech Inc., Piscataway, New Jersey)
was added to each culture. Cells were harvested onto glass microfibre
filters (Whatman 934-AH) and washed with distilled water using a
multiple automated sample harvester (Nunc, Denmark). Tritiated
thymidine incorporation was measured by liquid scintillation counting
(Packerd Tri-card 300; Beckman Instruments Inc. Fullerton, CA). Data
were expressed as mean counts per minute9/standard error (SE).
Statistical analysis
Differences on antibody titres and lymphocyte proliferation assays were
determined by analysis of variance using the General Linear Models
procedure (Luginbuke & Schlotzhauer, 1987). Significant differences
were further separated by Duncan’s multiple range tests using
commercial SAS† statistical software. Chi-square analysis was used
to determine significant differences between groups in mortality rate
(Zar, 1984). Significant differences were set at PB/0.05.
Results
Clinical signs and mortality
Clinical signs were observed in all the birds,although they appeared earlier and more severelyin the commercial chickens B-380. Birds showedlethargy, ruffed feathers, drowsiness, anorexia,fever, dehydration, a watery to mucoid yellowdiarrhoea, loss of weight, and paleness of the crest.When animals were dying the paleness was fol-lowed by cyanosis, blindness, and seizures.
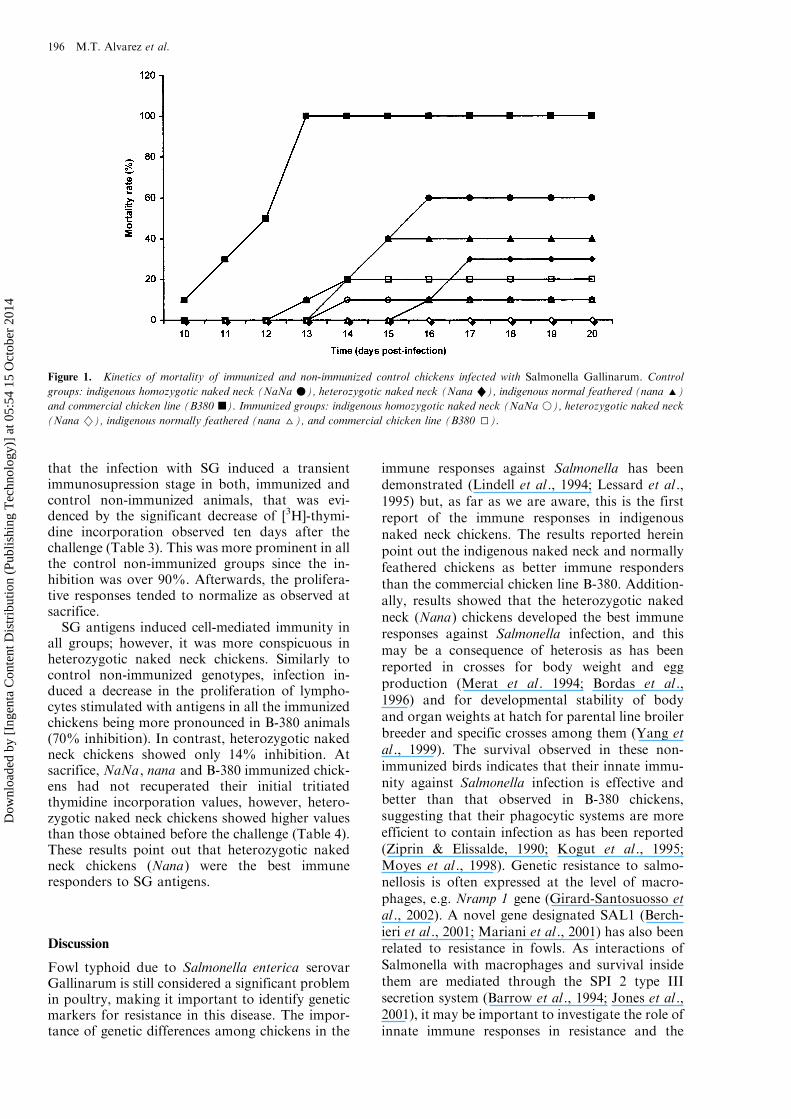
The results of mortality in immunized andcontrol non-immunized genotypes showed thatthe most resistant genotype was Nana , since thefirst death of these control chickens was observedat the 16th day and only 30% of them died at theend of the experiment. On the other hand, theBabcock (B-380) chickens showed the highestsusceptibility to Salmonella Gallinarun infection
since, 100% of control animals died (PB/0.05). Incontrast, 70% of Nana , 60% of nana and 40% ofNaNa control chickens survived. Kinetics of mor-tality showed that chickens began to die after the10th day, and B-380 control chickens were themost susceptible group to Salmonella infectionsince all the animals died by the 13th day (Figure1).
Immunization with Salmonella antigens inducedprotection against the challenge with SG in all thegenotypes. All the heterozygotic naked neck chick-ens (100% Nana), 90% of homozygotic (NaNa)and normally feathered (nana) chickens, and 80%of the commercial chicken line (B-380) survived.Only 2 chickens of the B-380 immunized groupdied on the 14th day, one of the NaNa group diedthat same day and one of the nana on the 16th day.The rest of the animals survived during the entireobservation period (Figure 1).
Body weight
All chickens gained weight with time but NaNaand Nana chickens were the heaviest at the end ofthe experiment as can be observed in normalanimal groups (Table 1). Before the challenge allanimals had similar body weights but afterwards,all non-immunized chickens lost weight. This losswas more conspicuous in the commercial chickenssince ten days after the challenge they had lost38.4% of their weight. At sacrifice NaNa hadrecovered their body weight but Nana and nanachickens gained weight. In contrast, the challengewith the virulent bacilli of the immunized animalsdid not affect the weight gain in indigenouschickens (naked neck or normally feathered), butagain B-380 chickens did not gain weight for thenext 10 days after the challenge, although theybegan to do it afterwards.
Antibody titres
The results of specific antibody titres in immunizedand control non-immunized genotypes are sum-marized in Table 2. Heterozygotic naked neckchickens produced the highest antibody titres inthe control non-immunized groups after the chal-lenge and at sacrifice, meanwhile B-380 chickensshowed the lowest titres. Immunization with SGantigens induced antibodies in all the genotypes,but higher titres were observed in naked neckchickens (Nana and NaNa). Immunized B-380chickens showed the lowest antibody titres afterthe second immunization, after the challenge, andat sacrifice (12299/190, 18909/404, and 35969/750,respectively).
Lymphocyte proliferation
Tritiated thymidine incorporation in peripheralblood lymphocytes stimulated with Con A showed
Immune responses in naked neck chickens 195
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014
that the infection with SG induced a transientimmunosupression stage in both, immunized andcontrol non-immunized animals, that was evi-denced by the significant decrease of [3H]-thymi-dine incorporation observed ten days after thechallenge (Table 3). This was more prominent in allthe control non-immunized groups since the in-hibition was over 90%. Afterwards, the prolifera-tive responses tended to normalize as observed atsacrifice.
SG antigens induced cell-mediated immunity inall groups; however, it was more conspicuous inheterozygotic naked neck chickens. Similarly tocontrol non-immunized genotypes, infection in-duced a decrease in the proliferation of lympho-cytes stimulated with antigens in all the immunizedchickens being more pronounced in B-380 animals(70% inhibition). In contrast, heterozygotic nakedneck chickens showed only 14% inhibition. Atsacrifice, NaNa , nana and B-380 immunized chick-ens had not recuperated their initial tritiatedthymidine incorporation values, however, hetero-zygotic naked neck chickens showed higher valuesthan those obtained before the challenge (Table 4).These results point out that heterozygotic nakedneck chickens (Nana) were the best immuneresponders to SG antigens.
Discussion
Fowl typhoid due to Salmonella enterica serovarGallinarum is still considered a significant problemin poultry, making it important to identify geneticmarkers for resistance in this disease. The impor-tance of genetic differences among chickens in the
immune responses against Salmonella has beendemonstrated (Lindell et al ., 1994; Lessard et al .,1995) but, as far as we are aware, this is the firstreport of the immune responses in indigenousnaked neck chickens. The results reported hereinpoint out the indigenous naked neck and normallyfeathered chickens as better immune respondersthan the commercial chicken line B-380. Addition-ally, results showed that the heterozygotic nakedneck (Nana) chickens developed the best immuneresponses against Salmonella infection, and thismay be a consequence of heterosis as has beenreported in crosses for body weight and eggproduction (Merat et al . 1994; Bordas et al .,1996) and for developmental stability of bodyand organ weights at hatch for parental line broilerbreeder and specific crosses among them (Yang etal ., 1999). The survival observed in these non-immunized birds indicates that their innate immu-nity against Salmonella infection is effective andbetter than that observed in B-380 chickens,suggesting that their phagocytic systems are moreefficient to contain infection as has been reported(Ziprin & Elissalde, 1990; Kogut et al ., 1995;Moyes et al ., 1998). Genetic resistance to salmo-nellosis is often expressed at the level of macro-phages, e.g. Nramp 1 gene (Girard-Santosuosso etal ., 2002). A novel gene designated SAL1 (Berch-ieri et al ., 2001; Mariani et al ., 2001) has also beenrelated to resistance in fowls. As interactions ofSalmonella with macrophages and survival insidethem are mediated through the SPI 2 type IIIsecretion system (Barrow et al ., 1994; Jones et al .,2001), it may be important to investigate the role ofinnate immune responses in resistance and the
Figure 1. Kinetics of mortality of immunized and non-immunized control chickens infected with Salmonella Gallinarum. Control
groups: indigenous homozygotic naked neck (NaNa m), heterozygotic naked neck (Nana "), indigenous normal feathered (nana ')
and commercial chicken line (B380 j). Immunized groups: indigenous homozygotic naked neck (NaNa k), heterozygotic naked neck(Nana 2), indigenous normally feathered (nana ^), and commercial chicken line (B380 I).
196 M.T. Alvarez et al.
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

Table 1. Effect of immunization with Salmonella Gallinarum antigens and infection with the virulent bacilli in the body weight of the different chicken genotypes
Normal chickens (Mean weight (g)9/SE) Non-immunized chickens (Mean weight (g)9/SE) Immunized chickens (Mean weight (g)9/SE)
Days post hatch
Group 28 52 62 72 28 52b 62c 72d 28a 52b 62c 72d
NaNa 5399/8B 8189/9B 15109/13B 19049/23B 5709/6B 8159/7B 6159/11B 8159/20B 5589/3B 8059/10B 11859/32B 16729/20B
Nana 5749/9A 8599/11A 15939/20A 20449/55A 6189/12A 8789/10A 7659/18A 12919/31A 5919/9A 8629/13A 13789/47A 18099/30A
nana 5059/5C 7619/10C 14599/18BC 17929/28C 5599/11B 7799/21B 6129/32B 8259/33B 5369/7BC 7609/21C 10909/37B 14779/39C
B-380 5099/10C 7809/13C 14369/22C 17719/17C 5509/17B 8039/8B 4959/13C ND 5239/6C 7909/9B 7849/12C 11929/41D
aBefore the immunization with Salmonella Gallinarum antigens.bImmediately before the challenge with Salmonella Gallinarum bacilli (108 CFU).cTen days after the challenge.dAt sacrifice.
Means in the same column with different superscripts differ significantly (PB/0.05).
ND, not determined.
N, number of animals.
Imm
un
eresp
on
sesin
nak
edn
eckch
icken
s197
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014
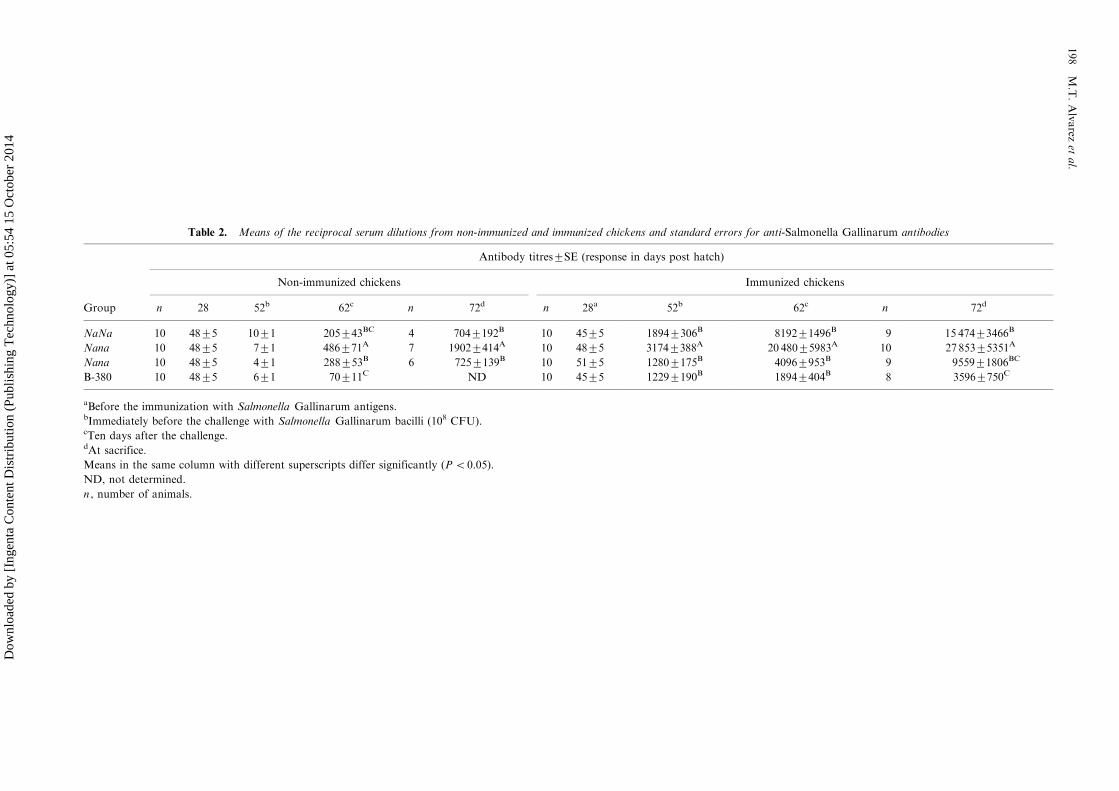
Table 2. Means of the reciprocal serum dilutions from non-immunized and immunized chickens and standard errors for anti-Salmonella Gallinarum antibodies
Antibody titres9/SE (response in days post hatch)
Non-immunized chickens Immunized chickens
Group n 28 52b 62c n 72d n 28a 52b 62c n 72d
NaNa 10 489/5 109/1 2059/43BC 4 7049/192B 10 459/5 18949/306B 81929/1496B 9 15 4749/3466B
Nana 10 489/5 79/1 4869/71A 7 19029/414A 10 489/5 31749/388A 20 4809/5983A 10 27 8539/5351A
Nana 10 489/5 49/1 2889/53B 6 7259/139B 10 519/5 12809/175B 40969/953B 9 95599/1806BC
B-380 10 489/5 69/1 709/11C ND 10 459/5 12299/190B 18949/404B 8 35969/750C
aBefore the immunization with Salmonella Gallinarum antigens.bImmediately before the challenge with Salmonella Gallinarum bacilli (108 CFU).cTen days after the challenge.dAt sacrifice.
Means in the same column with different superscripts differ significantly (PB/0.05).
ND, not determined.
n , number of animals.
198
M.T
.A
lvarez
etal.
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014
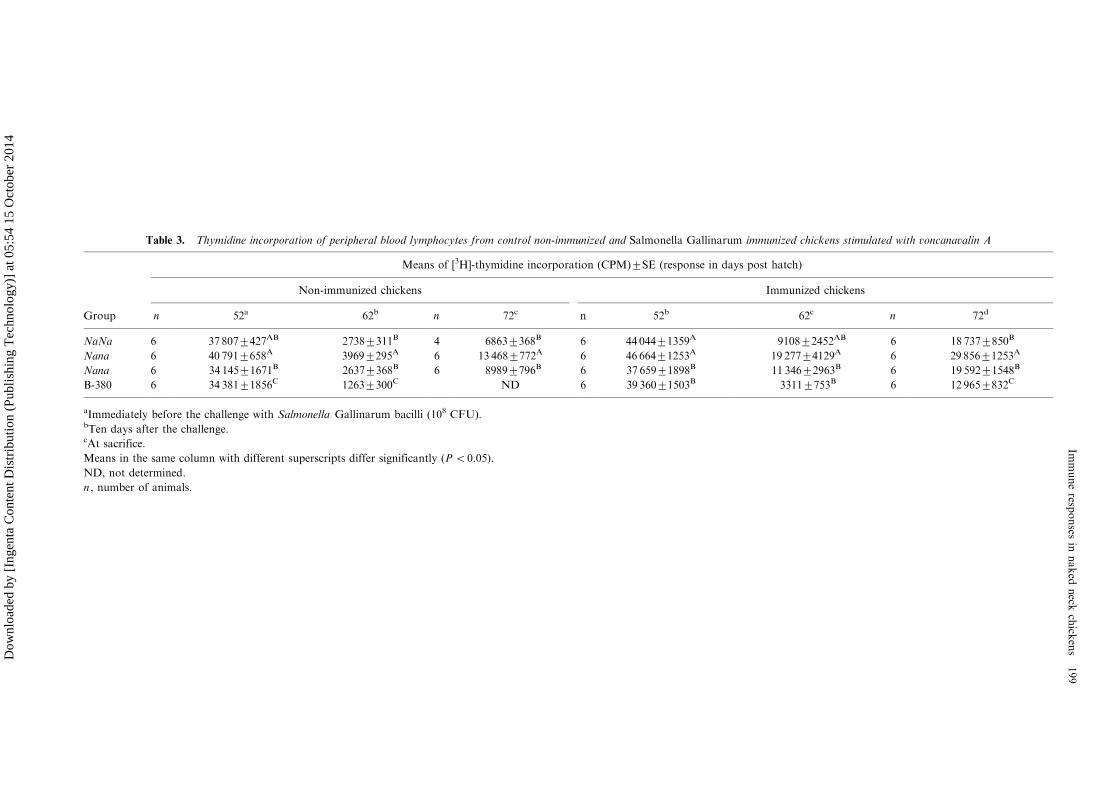
Table 3. Thymidine incorporation of peripheral blood lymphocytes from control non-immunized and Salmonella Gallinarum immunized chickens stimulated with voncanavalin A
Means of [3H]-thymidine incorporation (CPM)9/SE (response in days post hatch)
Non-immunized chickens Immunized chickens
Group n 52a 62b n 72c n 52b 62c n 72d
NaNa 6 37 8079/427AB 27389/311B 4 68639/368B 6 44 0449/1359A 91089/2452AB 6 18 7379/850B
Nana 6 40 7919/658A 39699/295A 6 13 4689/772A 6 46 6649/1253A 19 2779/4129A 6 29 8569/1253A
Nana 6 34 1459/1671B 26379/368B 6 89899/796B 6 37 6599/1898B 11 3469/2963B 6 19 5929/1548B
B-380 6 34 3819/1856C 12639/300C ND 6 39 3609/1503B 33119/753B 6 12 9659/832C
aImmediately before the challenge with Salmonella Gallinarum bacilli (108 CFU).bTen days after the challenge.cAt sacrifice.
Means in the same column with different superscripts differ significantly (PB/0.05).
ND, not determined.
n , number of animals.
Imm
un
eresp
on
sesin
nak
edn
eckch
icken
s199
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014
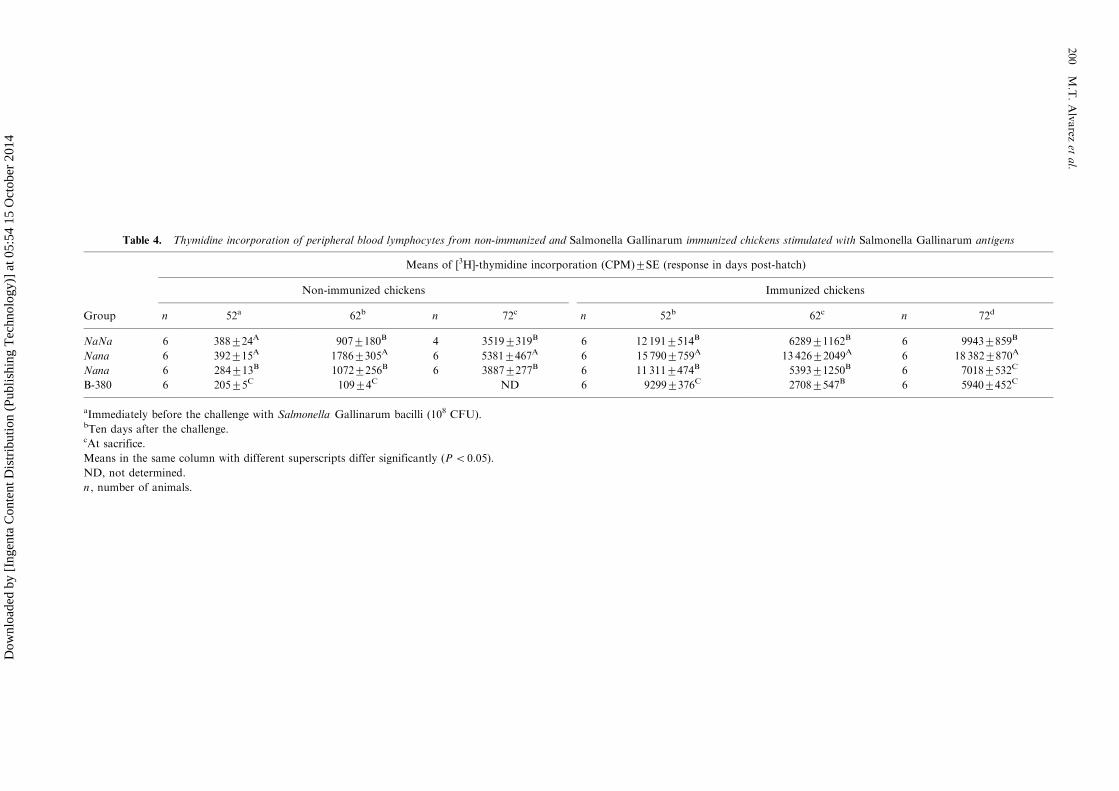
Table 4. Thymidine incorporation of peripheral blood lymphocytes from non-immunized and Salmonella Gallinarum immunized chickens stimulated with Salmonella Gallinarum antigens
Means of [3H]-thymidine incorporation (CPM)9/SE (response in days post-hatch)
Non-immunized chickens Immunized chickens
Group n 52a 62b n 72c n 52b 62c n 72d
NaNa 6 3889/24A 9079/180B 4 35199/319B 6 12 1919/514B 62899/1162B 6 99439/859B
Nana 6 3929/15A 17869/305A 6 53819/467A 6 15 7909/759A 13 4269/2049A 6 18 3829/870A
Nana 6 2849/13B 10729/256B 6 38879/277B 6 11 3119/474B 53939/1250B 6 70189/532C
B-380 6 2059/5C 1099/4C ND 6 92999/376C 27089/547B 6 59409/452C
aImmediately before the challenge with Salmonella Gallinarum bacilli (108 CFU).bTen days after the challenge.cAt sacrifice.
Means in the same column with different superscripts differ significantly (PB/0.05).
ND, not determined.
n , number of animals.
200
M.T
.A
lvarez
etal.
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

presence of these genetic regions in indigenousnaked neck chickens. It has been reported thatsusceptibility to cecal colonization by Salmonellain commercial chicks could be associated with thedelay in the establishment of mature native micro-bial flora that inhibits colonization by this patho-gen, due to the controlled conditions in which theyare raised (Nurmi & Rantala, 1973).
Chicks are highly susceptible to Salmonellainfection during the first four days post hatch,after that time they become increasingly resistantto infection (Ziprin et al ., 1989). Infection with SGbacilli induced loss of weight in all the non-immunized chickens, especially in the B-380 group.Death in some of the animals may be due to theloss of weight that was observed as a consequenceof infection. Mortality of commercial strain chick-ens infected with Salmonella varies according tothe doses of challenge, age, and sex (Prince &Garren, 1966; Bouzoubaa et al ., 1987). Our resultsshow that in 52-day-old male B-380 chicks chal-lenged with 1 LD50 virulent microorganism, themortality began during the second week afterinfection. There are good indications that cell-mediated immunity plays an important role incontrolling and clearing this facultative intracellu-lar bacterium (Tellez et al ., 1993; Kogut et al .,1994, 1995). The results show that cell-mediatedimmune responses were induced in chickens afterimmunization with SG antigens as well as highspecific antibody titres. Interestingly, all the sam-ples from 28-day-old chickens showed antibodiesagainst Salmonella , but these titres decreased withtime in control chickens, suggesting that theseantibodies could be passively transferred from themother (maternal antibodies). After immunization,all chickens showed specific antibodies against SGantigens, which increased with time.
We found that proliferation of peripheral bloodcells from immunized chickens stimulated withCon A decreased significantly after the challengein all the groups, this could be due to a decrease ofIL-2 production as reported by Kaiser et al . (2000)following S. enteritidis invasion, although this hasnot been shown in vivo . On the other hand, adecrease in the values of haematocrit and haemo-globin was observed 5 days after the challenge(data not shown), being more conspicuous in theB-380 chickens. This anaemia could be related tothe decrease in the proliferative response. How-ever, when cells were stimulated with SG antigens,Nana chickens did not show a significant decreaseof thymidine incorporation when compared withthe counts obtained after the challenge (15,7909/
759 vs. 13,4269/2,049). The immunosuppressionstage was transient in all the groups, since 20 daysafter the challenge the proliferative responsesbegan to recover. This effect was not observed inany control group. Further studies are necessary in
order to understand the sources and immunologi-cal meaning of this immunosuppression.
The results of the present study indicate thatindigenous heterozygotic Nana chickens had a highinnate immunity to SG infection. Besides, whenimmunized with SG antigens, they showed thehighest humoral and cell-mediated immune re-sponses. These results support the idea that thisgenotype should be considered in genetic selectionprogrammes for resistance to fowl typhoid causedby Salmonella Gallinarum.
Acknowledgements
Authors wish to thank Dr. David AlejandroMedina for manuscript revision. This researchwas supported by funds of the Universidad Na-cional Autonoma de Mexico.
References
Arechavaleta, F., Molinari, J.L. & Tato, P. (1998). A Taenia solium
cysticercus factor nonspecifically inhibits cytokine production. Para-
sitology Research , 84 , 117�/122.
Barrow, P.A. (1993). Salmonella control-past, present, and future.
Avian Pathology , 22 , 651�/669.
Barrow, P.A., Huggins, M.B. & Lovell, M.A. (1994). Host specificity of
Salmonella infection in chickens and mice is expressed in vivo
primarily at the level of the reticuloendothelial system. Infection and
Immunity , 62 , 4602�/4610.
Benjamin Jr., W.H., Briles, W.E., Waltman, W.D. & Briles, D.E.
(1991). Effects of genetic and prior Salmonella enteritidis infection on
the ability of chickens to be infected with S. enteritidis . In L.C.
Blankernship (Ed.), Colonization Control of Human Bacterial En-
teropathogens in Poultry (pp. 365�/369). New York: Academic Press.
Berchieri Jr., A., Oliviera, G.H., Pinheiro, L.A.S. & Barrow, P.A.
(2000). Experimental Salmonella Gallinarum infection in light laying
hens. Brazilian Journal of Microbiology , 31 , 50�/52.
Berchieri Jr., A., Murphy, C.K., Marston, K. & Barrow, P.A. (2001).
Observations on the persistence and vertical transmission of Salmo-
nella enterica serovars Pullorum and Gallinarum in chickens: effect
of bacterial and host genetic background. Avian Pathology , 30 , 221�/
231.
Bordas, A., Merat, P. & Minvielle, F. (1996). Heterosis in egg-laying
lines under divergent selection for residual feed consumption. Poultry
Science , 75 , 20�/24.
Bouzoubaa, K., Nagaraja, K.V., Newman, J.A. & Pomeroy, B.S.
(1987). Use of membrane proteins from Salmonella gallinarum for
prevention of fowl typhoid infection in chickens. Avian Diseases , 31 ,
699�/704.
Bradford, M.M. (1976). A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye-binding. Analytical Biochemistry , 72 , 248�/
254.
Bumstead, N. & Barrow, P. (1993). Resistance to Salmonella galli-
narum , S. pullorum and S. enteritidis in inbred lines of chickens.
Avian Diseases , 37 , 189�/193.
Cahaner, A., Deeb, N. & Gutman, M. (1993). Effects of plumage-
reducing naked neck (Na) gene on the performance of fast growing
broilers at normal and high ambient temperature. Poultry Science ,
72 , 767�/775.
Davies, R.H., Nicolas, R.A.J., McLaren, I.M., Corkish, J.D., Lanning,
D.G. & Wray, C. (1997). Bacteriological and serological investiga-
tion of persistent Salmonella enteritidis infection in an integrated
poultry organization. Veterinary Microbiology , 58 , 272�/293.
Immune responses in naked neck chickens 201
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

Deeb, N. & Cahaner, A. (1999). The effects of naked neck genotypes,
ambient temperature, and feeding status and their interactions on
body temperature and performance of broilers. Poultry Science , 78 ,
1341�/1346.
Desmidt, M., Van Neron, A., Haesebrouck, R. & Ysebaert, M.T.
(1996). Oxygenation activity of chicken blood phagocytes as
measured by luminol- and lucigenin-dependent chemiluminescence.
Veterinary Immunology and Immunopathology , 53 , 303�/311.
El-Attar, A. & Merat, P. (1985). Composition corporelle de poulets
‘‘Cou nu’’ normalement emplumes resultats dans un consement de
‘‘chair’’. Genetic Selection Science , 14 , 271�/279.
Girard-Santosuosso, O., Lantier, F., Lantier, I., Bumstead, N., Elsen,
J.M. & Beaumont, C. (2002). Heritability of susceptibility to
Salmonella enteritidis infection in fowls and test of the role of the
chromosome carrying the NRAMP 1 gene. Genetic Selection
Evolution , 34 , 211�/219.
Jones, M.A., Wigley, P., Page, K.L., Hulme, S.D. & Barrow, P.A.
(2001). Salmonella enterica serovar Gallinarum requires the Salmo-
nella pathogenicity island 2 Type III secretion system but not the
Salmonella pathogenicity island 1 Type III secretion system for
virulence in chickens. Infection and Immunity , 69 , 5471�/5476.
Kaiser, M.G., Wing, T. & Lamont, S.J. (1998). Effect of genetics,
vaccine dosage and post vaccinations sampling interval on early
antibody response to Salmonella enteritidis vaccine in broiler breeder
chicks. Poultry Science , 77 , 271�/275.
Kaiser, P., Rothwell, L., Galyov, E.E., Barrow, P.A., Burnside, J. &
Wigley, P. (2000). Differential cytokine expression in avian cells in
response to invasion by Salmonella typhimurium , Salmonella enter-
itidis and Salmonella gallinarum . Microbiology , 146 , 3217�/3226.
Kogut, M.H., McGruder, E.D., Hargis, B.M., Corrier, D.E. &
DeLoach, J.R. (1994). Characterization of the pattern of inflamma-
tory cell influx in chicks following the intraperitoneal administration
of live Salmonella enteritidis -immune lymphokines. Poultry Science ,
74 , 8�/17.
Kogut, M.H., McGruder, E.D., Hargis, B.M., Corrier, D.E. &
DeLoach, J.R. (1995). In vivo activation of heterophil function in
chickens following injection with Salmonella enteritidis -immune
lymphokines. Journal of Leukocyte Biology , 57 , 56�/62.
Lessard, M., Hutchings, D.L. & Spencer, J.L. (1995). Cell-mediated
immunity and humoral immune responses in chickens infected with
Salmonella typhimurium . Avian Diseases , 39 , 230�/238.
Lindell, K.A., Saeed, A.M. & McCabe, G.P. (1994). Evaluation of
resistance of four strains of commercial laying hens to experimental
infection with Salmonella enteritidis phage type eight. Poultry
Science , 73 , 757�/762.
Lucio, B., Padron, M. & Mosqueda, A. (1985). Fowl typhoid in
Mexico. In G.H. Snoeyenbos (Ed.), Proceedings of the International
Symposium on Salmonella (pp. 382�/383). Kennett Square, PA:
American Association of Avian Pathologists.
Luginbuke, R.C. & Schlotzhauer, S.D. (1987). SAS/STAT† Guide for
Personal Computers , 6th edn. Cary, NC: SAS Institute Inc.
Mariani, P., Barrow, P.A., Cheng, H.H., Groenen, M.M., Negrini, R.
& Bumstead, N. (2001). Localization to chicken chromosome 5 of a
novel locus determining Salmonellosis resistance. Immunogenetics ,
53 , 786�/791.
Mdegela, R.H., Yongolo, M.G.S., Minga, U.M. & Olsen, J.E. (2000).
Molecular epidemiology of Salmonella gallinarum in chickens in
Tanzania. Avian Pathology , 29 , 457�/463.
Merat, P. (1986). Potential usefulness of the Na (naked neck) gene in
Poultry production. World’s Poultry Science Journal , 42 , 124�/142.
Merat, P., Minvielle, F., Bordas, A. & Coquerelle, G. (1994). Heterosis
in normal versus dwarf laying hens. Poultry Science , 73 , 1�/6.
Mills, J.N. & Wilcox, G.E. (1993). Separation of phagocytes leukocytes
from the peripheral blood of chickens. Avian Pathology , 22 , 343�/
352.
Moyes, R.B., Kogut, M.H., Drolesky, R.E. & DeLoach, J.R. (1998).
Differential expression of adhesion molecules by chickens heterophils
activated in vivo with Salmonella enteritidis -immune lymphokines.
Veterinary Immunology and Immunopathology , 62 , 83�/95.
Nurmi, E. & Rantala, M. (1973). New aspects of Salmonella infection
in broiler production. Nature , 241 , 210�/211.
Prince, W.R. & Garren, H.W. (1966). An investigation of the resistance
of White Leghorn chicks to Salmonella gallinarum . Poultry Science ,
15 , 1149�/1153.
Shivaprasad, H.L. (1997). Pullorum disease and fowl typhoid. In B.W.
Calnek, H.J. Barnes, C.W. Beard, L.R. McDougald & Y.M. Saif
(Eds.), Diseases of Poultry , 10th ed. (pp. 82�/96). Ames: Iowa State
University Press.
Shivaprasad, H.L. (2000). Fowl typhoid and pullorum disease. Review
Science Technology , 19 , 405�/424.
Silva, E.N. (1985). The Salmonella gallinarum problem in Central and
South America. In G.H. Snoeyenbos (Ed.), Proceedings of the
International Symposium on Salmonella (pp. 150�/156). Kennett
Square, PA: American Association of Avian Pathologists.
Tato, P., Flisser., A., Gavilanes, M. & Molinari, J.L. (1979).
Immunogenic complexes obtained from Salmonella typhymurium
and Salmonella typhi Ty2 by the bacterial acetone powder method.
Annals of Microbiology Institute Pasteur , 130A , 47�/60.
Tellez, G.I., Kogut, M.H. & Hargis, B.M. (1993). Immunoprophylaxis
of Salmonella enteritidis infection by lymphokines in Leghorn
chickens. Avian Diseases , 37 , 1062�/1070.
Wray, C., Davies, R.H. & Corkish, J.D. (1996). Enterobacteriaceae. In
F.T.W. Jordan & M. Pattison (Eds.), Poultry Diseases , 4th edn (pp.
9�/43). London: Saunders Company Ltd.
Yalcin, S., Testik, A., .Ozkan, S., Settar, P., Celen, F. & Cahaner, A.
(1997). Performance of naked neck and normal broilers in hot,
warm, and temperate climates. Poultry Science , 76 , 930�/937.
Yang, A., Emmerson, D.A., Dunnington, E.A. & Siegel, P.B. (1999).
Heterosis and developmental stability of body and organ weights at
hatch for parental line broiler breeders and specific crosses among
them. Poultry Science , 78 , 942�/948.
Zar, J. (1984). Biostatistical Analysis , 2nd edn. Englewood Cliffs, NJ:
Prentice-Hall Inc.
Ziprin, R.L., Corrier, D.E. & Elissalde, M.H. (1989). Maturation of
resistance to salmonellosis in newly hatched chicks: inhibition by
cyclosporine. Poultry Science , 68 , 1637�/1642.
Ziprin, R.L. & Elissalde, M.H. (1990). Effect of T-2 toxin on resistance
to systemic Salmonella typhimurium infection on newly hatched
chickens. American Journal of Veterinary Reseach , 51 , 1869�/1872.
RESUME
Comparaison des reponses immunitaires vis-a-vis de l’infection a Salmo-
nella enterica , serovar Gallinarum chez les poulets a cou nu et chez des
poulets de lignee commerciale
Les reponses immunitaires de poulets indigenes a cou nu (NaNa et
Nana ) et de poulets emplumes normalement (nana) vis-a-vis de
l’infection a Salmonella Gallinarum (SG) ont ete evaluees et comparees
a celles de poulets de lignee commerciale (B-380). Des groupes de
poulets ages de 28 jours (NaNa, Nana, nana et B-380 ) ont ete
immunises oralement et par voie sous-cutanee avec 50 mg d’antigene
SG. Des poulets temoins non immunises ont ete inocules avec une
solution saline sterile. Tous les poulets ont ete eprouves avec 1 LD50 de
SG et la mortalite a ete enregistree tous les jours durant 20 jours. Les
anticorps anti SG ont ete mesures dans les serums, avant immunisation,
avant epreuve, 10 jours apres l’epreuve et lors de l’euthanasie. Des tests
de proliferation des lymphocytes du sang peripherique ont ete realises
en utilisant la Concavaline A et l’antigene SG. Les resultats montrent
que les poulets Nana non immunises ont presente la meilleure resistance
naturelle a l’infection salmonellique, puisque seulement 30% d’entre
eux sont morts. A l’oppose, tous les poulets temoins B-380 sont morts
avant le 13eme jour. L’immunisation avec SG a induit une immunite
chez les poulets de tous les genotypes. Les poulets indigenes a cou nu et
les poulets emplumes normalement ont montre un taux de survie
superieur compares aux poulets B-380. Les poulets Nana immunises
ont presente les titres en anticorps les plus eleves (PB/0,05) ainsi que
l’incorporation la plus importante de thymidine dans les lymphocytes
du sang peripherique stimules avec de la Con A ou de l’antigene SG
(PB/0,05). Les resultats montrent que les poulets Nana sont les plus
resistants a l’infection SG et ceux qui repondent le mieux a la
vaccination avec un antigene S. Gallinarum.
202 M.T. Alvarez et al.
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014

ZUSAMMENFASSUNG
Vergleich von Immunantworten auf eine Salmonella entrica Serovar
Gallinarum- Infektion bei Nackthalshuhnern und einer kommerziellen
Huhnerlinie
Die Immunantworten von einheimischen Nackthals (NaNa und Nana )-
und normal befiederten (nana ) Huhnern auf eine Salmonella gallinarum
(SG)-Infektion wurde ermittelt und verglichen mit denen einer kom-
merziellen Huhnerlinie (B-380). Gruppen von 28 Tage alten Kuken
(NaNa, Nana, nana und B-380) wurden oral und subcutan immunisiert
mit 50 g SG-Antigen. Nicht immunisierte Kontrolltiere wurden mit
steriler Kochsalzlosung inokuliert. Alle Huhner wurden mit 1 LD50 SG
testinfiziert und die Mortalitat wurde taglich uber einen Zeitraum von
20 Tagen protokolliert. Antikorper gegen SG wurden in den Seren vor
der Immunisierung, vor der Belastungsinfektion, 10 Tage nach der
Belastungsinfektion und am Versuchsende ermittelt. Lymphozytenpro-
liferationstests mit peripheren Blutlymphozyten wurden unter Verwen-
dung von Concanavalin A (ConA) und SG-Antigen durchgefuhrt. Die
Ergebnisse erbrachten, dass die nicht immunisierten Nana -Huhner die
beste naturliche Resistenz gegen die Salmonelleninfektion hatten, da
nur 30% von ihnen starben. Im Gegensatz dazu verendeten alle
Kontroll-B-380-Huhner bis zum 13. Tag nach der Infektion. Die
Vakzination mit SG induzierte eine Immunitat bei allen Genotypen.
Die einheimischen Nackthals- und die normal befiederten Huhner
hatten eine hohere Uberlebensrate als die B-380-Huhner. Die immuni-
sierten Nana -Huhner zeigten sowohl die hochsten Antikorpertiter
(pB/0,05) als auch die hochste Thymidin-Inkorporation in peripheren
Blutlymphozyten nach Stimulation mit ConA oder SG-Antigen
(pB/0,05). Die Ergebnisse zeigen, dass Nana -Huhner die großte
Resistenz gegenuber einer SG-Infektion und das beste Immunreak-
tionsvermogen auf eine Vakzination gegen Salmonella gallinarum
besitzen.
RESUMEN
Comparacion de la respuesta inmune frente a la infeccion con Salmonella
enterica serovar Gallinarum entre pollos de cuello desnudo y una lınea
comercial de pollos
La respuesta inmune de pollos indıgenas de cuello desnudo (NaNa y
Nana ) y pollos de plumaje normal (nana ) frente a la infeccion con
Salmonella Gallinarum (SG) fue evaluada y comparada con la de una
lınea comercial (B-380). Grupos de pollos de 28 dıas de edad (NaNa ,
Nana , nana y B-380) se inmunizaron oral y subcutaneamente con 50 mg
de antıgeno de SG. Animales control no inmunizados fueron inocu-
lados con una solucion salina esteril. Todos los pollos fueron desafiados
con 1 LD50 de SG y se registro la mortalidad diariamente durante 20
dıas. Se cuantificaron los anticuerpos frente a SG en el suero antes de la
inmunizacion, antes del desafıo, 10 dıas tras el desafıo y al sacrificio. Se
realizaron pruebas de proliferacion de linfocitos de sangre periferica
mediante el uso de Concanavalina A y antıgenos de SG. Los resultados
mostraron que los pollos Nana no inmunizados exhibıan la mejor
resistencia natural frente a la infeccion con Salmonella , ya que
unicamente el 30% de ellos murieron. En contraste, todos los pollos
controles B-380 murieron hacia el dıa 13. La inmunizacion con SG
indujo inmunidad en pollos de todos los genotipos. Los pollos
indıgenas de cuello desnudo y los de plumaje normal mostraron una
supervivencia mucho mayor que los pollos B-380. Los pollos Nana
inmunizados mostraron los niveles mas altos de anticuerpos (PB/0.05)
ası como los niveles de incorporacion de timidina mas altos en los
linfocitos de sangre periferica estimulados con Con A o antıgenos de
SG (PB/0.059. Los resultados demuestran que los pollos Nana son los
mas resistentes a la infeccion con SG y los que responden mejor a la
vacunacion con antıgenos de S. Gallinarum.
Immune responses in naked neck chickens 203
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
05:
54 1
5 O
ctob
er 2
014
Top Related
Copyright © 2022 FDOKUMEN

