







Bahasa
Halaman
Hukum


Molecular Cell, Vol. 15, 647–657, August 27, 2004, Copyright 2004 by Cell Press
An Atomic Resolution Model for Assembly,Architecture, and Function of the Dr Adhesins
coli is a notable exception, as its members are associ-ated with both diarrheal and urinary tract infections.These proteins are reported to form both fimbrial and
Kirstine L. Anderson,1,2,9 Jason Billington,3,9
David Pettigrew,3 Ernesto Cota,1,2
Peter Simpson,1,2 Pietro Roversi,3 Ho An Chen,1,2
Petri Urvil,4 Laurence du Merle,5 Paul N. Barlow,6 afimbrial structures at the bacterial cell surface andtarget a common host cell receptor, the decay-accel-M. Edward Medof,7 Richard A.G. Smith,8
Bogdan Nowicki,4 Chantal Le Bouguenec,5 erating factor (DAF or CD55). Using the newly solvedthree-dimensional structure of AfaE, we have con-Susan M. Lea,3,* and Stephen Matthews1,2,*
1Department of Biological Sciences structed a robust atomic resolution model that revealsthe structural basis for assembly by donor strand com-Wolfson Laboratories
Imperial College London plementation and for the architecture of capped sur-face fibers.South Kensington
London SW7 2AZUnited Kingdom Introduction2 Centre for Structural BiologyImperial College London The ability of pathogenic Escherichia coli to adhere toSouth Kensington and invade mucosal epithelial layers in humans andLondon SW7 2AZ other mammals is an essential prerequisite for disease.United Kingdom Bacterial proteins—called adhesins and invasins—are3 Laboratory of Molecular Biophysics involved in targeting the host. They are located on ex-Department of Biochemistry tended cell surface appendages known as fimbriae, asUniversity of Oxford individual membrane-tethered molecules, or as part ofSouth Parks Road an amorphous outer membrane-associated structureOxford OX1 3QU termed an afimbrial sheath (Soto and Hultgren, 1999).United Kingdom The Dr family of adhesins has been the subject of recent4 Department of Obstetrics and Gynecology and interest, as it was believed to contain representativesDepartment of Microbiology and Immunology having both fimbrial and afimbrial architectures (Bilge etThe University of Texas Medical Branch al., 1989; Labigne-Roussel and Falkow, 1988; Labigne-Galveston, Texas 77555 Roussel et al., 1984; Nowicki et al., 1989). The Dr adhes-5 Unite de Pathogenie Bacterienne des Muqueuses ins mediate a diffuse adherence pattern of bacteria toInstitut Pasteur epithelial cells by virtue of binding to decay-accelerating28 rue du Docteur Roux factor (DAF or CD55), a 70 kDa glycophosphatidylinosi-75724 Paris CEDEX 15 tol (GPI)-anchored glycoprotein that regulates comple-France ment activation (Medof et al., 1984, 1985). Members of6 Edinburgh Protein Interaction Centre this family are commonly isolated from uropathogenicUniversity of Edinburgh E. coli (UPEC), which causes urinary tract infections, andEdinburgh EH9 3JJ diffusely adherent E. coli (DAEC), a cause of intestinalScotland infections (Le Bouguenec et al., 2001).7 Institute of Pathology The afimbrial classification for some members of theCase Western Reserve University School of Medicine Dr family was based on the fact that surface fimbriaeCleveland, Ohio 44106 could not be detected by electron microscopy (EM) (Gar-8 Adprotech Ltd. cia et al., 1996; Labigne-Roussel and Falkow, 1988; La-Chesterford Research Park bigne-Roussel et al., 1984; Le Bouguenec et al., 1993).Little Chesterford Other Dr family members such as the Dr hemagglutininSaffron Walden and F1845 fimbriae (Bilge et al., 1989; Swanson et al.,Essex CB10 1XL 1991), however, have been subsequently shown to ex-United Kingdom hibit a fimbrial morphology. A diffusely adherent fimbrial
structure was also recently discovered in enteropatho-genic E. coli (EPEC) (Keller et al., 2002). Although theDr subunits themselves possess no sequence homologySummarywith orthodox fimbrial domains, the presence of fimbrialusher and chaperone paralogs suggests a commonPathogenic bacteria possess adhesion protein com-
plexes that play essential roles in targeting host cells mechanism of assembly for Dr and prototypical fimbrialadhesins (Barnhart et al., 2000; Sauer et al., 1999, 2000b).and in propagating infection. Although each family of
adhesion proteins is generally associated with a spe- Furthermore, recent structural analysis of the Caf1 anti-gen from Yersinia pestis suggests that bacterial append-cific human disease, the Dr family from Escherichiaages assembled via the chaperone/usher pathway arefibrillar (Zavialov et al., 2003).*Correspondence: [email protected] (S.M.L.); s.j.matthews@
Key members of the Dr family are the plasmid-borneimperial.ac.uk (S.M.)9 These authors contributed equally to this work. afa-3 operon, which encodes proteins that assemble
Molecular Cell648
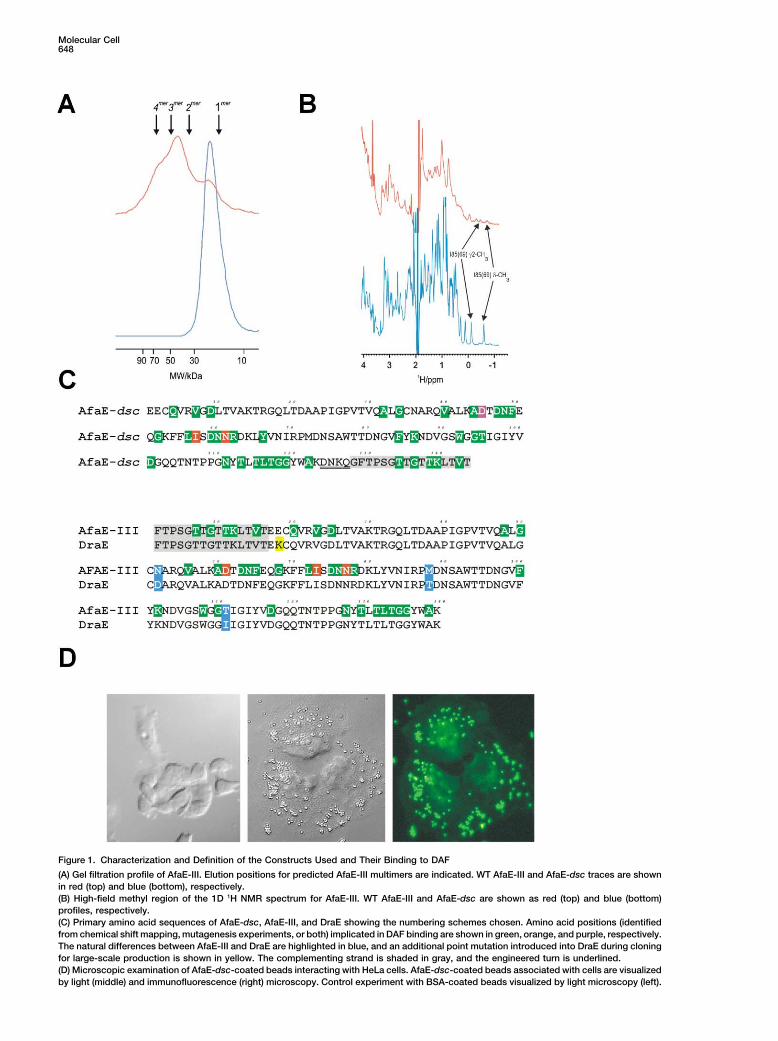
Figure 1. Characterization and Definition of the Constructs Used and Their Binding to DAF
(A) Gel filtration profile of AfaE-III. Elution positions for predicted AfaE-III multimers are indicated. WT AfaE-III and AfaE-dsc traces are shownin red (top) and blue (bottom), respectively.(B) High-field methyl region of the 1D 1H NMR spectrum for AfaE-III. WT AfaE-III and AfaE-dsc are shown as red (top) and blue (bottom)profiles, respectively.(C) Primary amino acid sequences of AfaE-dsc, AfaE-III, and DraE showing the numbering schemes chosen. Amino acid positions (identifiedfrom chemical shift mapping, mutagenesis experiments, or both) implicated in DAF binding are shown in green, orange, and purple, respectively.The natural differences between AfaE-III and DraE are highlighted in blue, and an additional point mutation introduced into DraE during cloningfor large-scale production is shown in yellow. The complementing strand is shaded in gray, and the engineered turn is underlined.(D) Microscopic examination of AfaE-dsc-coated beads interacting with HeLa cells. AfaE-dsc-coated beads associated with cells are visualizedby light (middle) and immunofluorescence (right) microscopy. Control experiment with BSA-coated beads visualized by light microscopy (left).

Assembly and Function of the Dr Adhesins649
al., 1984) and prevent subsequent amplification of theTable 1. Structural Statistics for AfaE-dsccomplement cascade (Medof et al., 1984, 1985). The
Construct and PDB Code for Deposited AfaE-dsc (1RXL)extracellular portion of DAF includes four N-terminalCoordinatescomplement control protein modules (CCPs), alterna-
Number of Experimental Restraints 4158 tively known as short consensus repeats or sushidomains (Caras et al., 1987; Medof et al., 1987). CCPTotal NOE-derived 4099
Ambiguous 1542 modules comprise short � strands in an elongated bar-Unambiguous 2557 rel-like arrangement with N and C termini at either endIntraresidue 864 of the long axis. The binding of classical pathway (CP)Sequential 587
convertases is localized within CCP-2 and CCP-3, whileMedium-range (|i � j| � 4) 194regulation of the alternative pathway (AP) extendsLong-range (|i � j| � 4) 912through to CCP-4 (Brodbeck et al., 1996). Further muta-TALOS (φ/�) 58genesis studies have identified CCP-3 as the CCP do-
RMSD from Experimental Restraintsmain central to binding of the Dr family adhesins (Hasan
Distance (A) 0.020 � 0.0025 et al., 2002; Le Bouguenec et al., 2001; Pham et al.,Dihedral angle (deg.) 0.52 � 0.11 1995; Selvarangan et al., 2000).
The high sequence similarity of the adhesin subunitsRMSD from Idealized Covalent Geometry
among the Dr family raises several fundamental ques-Bonds (A) 0.0045 � 0.00013tions: (1) How do they assemble into surface organelles?Angles (deg.) 0.59 � 0.02(2) What is the architecture of resultant afimbrial and
Energies (kcal mol � 1) fimbrial structures? (3) What is the basis for receptorENOE 84.8 � 22.5 binding? Despite much recent attention, the fine struc-Ebond 43.8 � 2.5 tural details of Dr adhesin architecture and function haveEangle 205 � 12.4 yet to be characterized. To address many of these is-Evdw �1259 � 20.3
sues, we have embarked on an atomic resolution studyCoordinate RMSD (A) of the Dr family. Our combined NMR and biophysical
studies of an engineered monomer of AfaE-III reveal theBackbone atoms in secondary structure 0.49 � 0.05structural basis for self-assembly into a fine flexible fiberHeavy atoms in secondary structure 0.91 � 0.08
Backbone atoms (all residues 1–143) 0.67 � 0.09 and its interaction with DAF.Heavy atoms (all residues 1–143) 1.13 � 0.09
Results and DiscussionRamachandran Plota
Residues in most favored regions (%) 75Determination of the Atomic Resolution StructureResidues in additionally allowed regions (%) 19for the AfaE-III SubunitResidues in generously allowed regions (%) 3.3Large, multimeric assemblies are difficult to study usingResidues in disallowed regions (%) 2.7high-resolution structural techniques. The gel filtrationa Structural quality was evaluated using PROCHECK_NMR (Laskow-profile of AfaE-III in combination with NMR spectra dem-ski et al., 1996).onstrate (Figures 1A and 1B) that the wild-type (WT)adhesin is a heterogeneous oligomer and therefore doesnot constitute a suitable candidate for structural an-
the afimbrial sheath, and the closely related dra operon, alysis. High-resolution structures of archetypal fim-which encodes the fimbrial Dr hemagglutinin (Le Bou- brial components have been successfully determinedguenec et al., 1993; Nowicki et al., 1989). The afa-3 (Choudhury et al., 1999; Sauer et al., 1999, 2002; Zavialovoperon encodes two subunit proteins, AfaE-III and AfaD- et al., 2003), and these studies provide useful clues forIII, that are directly linked to virulence and are coloca- experimental strategies to prevent the self-assembly oflized at the outer membrane (Garcia et al., 1996; Gounon the Dr adhesins.et al., 2000). AfaE-III mediates the primary adhesion One clue is provided by the similarity between chaper-event, while AfaD-III is necessary for efficient internaliza- ones encoded by afa-3 and dra operons and the peri-tion of the bacteria. The closely related dra operon en- plasmic chaperones that assist the assembly of manycodes the fimbrial subunits DraE and DraD, collectively fibrous bacterial organelles. These function by pro-known as the Dr hemagglutinin. AfaE-III and the equiva- tecting pilus subunits while they fold within the bacteriallent protein from the dra operon, DraE, differ in sequence periplasm and target them to the outer membrane usherby only three residues, and both confer cell adherence protein for export (Barnhart et al., 2000; Sauer et al.,by binding specifically to DAF (Garcia et al., 1996; No- 1999, 2000b). The subunits lack an antiparallel � strand,wicki et al., 2001). Unlike AfaE, however, DraE binds which is provided by the chaperone as a parallel strand,a second receptor, the 7s domain of type IV collagen a process termed donor strand complementation (DSC)(Westerlund et al., 1989). Another intriguing distinction (Choudhury et al., 1999; Sauer et al., 1999, 2000a; Zavia-between AfaE-III and DraE is that the latter carries a lov et al., 2003). Proteins that are destined to join thebinding site for chloramphenicol that inhibits adhesion lengthening fiber possess a free N-terminal strand thatto both DAF and type IV collagen (Carnoy and Moseley, allows attachment to another subunit by taking over1997; Nowicki et al., 1988; Swanson et al., 1991; Van the role previously performed by the chaperone in anLoy et al., 2002). antiparallel arrangement, a process termed donor strand
The primary function of DAF is to dissociate C3 con- exchange (DSE) (Choudhury et al., 1999; Sauer et al.,1999, 2002; Zavialov et al., 2003).vertases that assemble on self-cell surfaces (Medof et

Molecular Cell650
Figure 2. Three-Dimensional Structure ofAfaE-dsc
(A) Stereoview showing C� traces represent-ing the ensemble of NMR-derived structures.(B) Ribbon representation of a representativestructure for AfaE-dsc. All � strands areshown in blue, with the exception of the self-complementing strand, which is blue/white.(C) Sequence comparison of the engineeredC-terminal strand of AfaE-dsc with the N-ter-minal residues of members of the Dr familyof adhesins. The conserved G, T, and L arehighlighted.(D) The binding cleft of the self-complement-ing strand Gd. The body of the protein is repre-sented using a surface map, with the excep-tion of strand Gd in AfaE-dsc, which is shownas a ball and stick representation.(E) Plot of 2D 1H-15N steady-state hetero-nuclear NOE value against sequence numberfor AfaE-dsc.
The sequence similarity among these periplasmic in polymerization. Microscopic examination of AfaE-dsc-coated beads revealed a host cell adherence patternchaperones suggests that DSC/DSE is employed in as-
sembly of the Dr family adhesins. Based upon this hy- similar to WT AfaE-III (Figure 1D), and surface plasmonresonance (SPR) experiments confirmed that AfaE-dscpothesis, we engineered a self-complemented and
therefore monomeric soluble form of the adhesin. We binds DAF with comparable affinity to WT AfaE-III. Fur-thermore, SPR-based competition experiments clearlyconstructed this “donor strand-complemented” AfaE-III
(AfaE-dsc) by removing the first 16 N-terminal amino demonstrate that AfaE-dsc effectively competes for thesame DAF binding site as AfaE-III (Figure 5). We canacids from mature AfaE-III (i.e., the donor strand) and
inserting them at the C terminus after a four-residue therefore conclude that AfaE-dsc displays all the func-tional properties of AfaE-III.turn, thereby enabling the donor strand to fold back and
stabilize the structure (Figure 1C). This work was based Using a combination of manual and ARIA NMR assign-ment methods for analysis of AfaE-dsc (Linge et al.,on the approach used in studies of FimH whereby the
sequence was extended with the N-terminal peptide of 2003), a total of 4099 nuclear Overhauser effects (NOEs)were assigned in AfaE-dsc 15N/13C-edited NOESY spec-FimG, producing a mono-dispersed species, as tested
by light scattering (Barnhart et al., 2000). The resulting tra. The structure determination was also supplementedwith 58 φ and � dihedral angle restraints, amounting toAfaE-dsc is highly soluble and exhibits a gel filtration
profile consistent with a monomeric 17 kDa protein (Fig- an average of 30 restraints per residue. For the finaliteration, the 15 lowest energy structures were chosenure 1A). NMR analysis shows a dramatic improvement
in spectra compared to the WT form of the subunit, from a total of 50 calculated based on agreement withexperimental data and structural quality (Table 1). Allwith the substantial decrease in line widths being the
outstanding feature (Figure 1B). Moreover, the 15N relax- areas of secondary structure are very well defined (Fig-ure 2A); the average pairwise root-mean-square devia-ation data (not shown) for AfaE-dsc, which provide an
estimate for the overall correlation time of 10 ns, confirm tion (RMSD) for the water-refined final structures is0.49 A for the backbone atoms and 0.91 A for the heavyits monomeric status at mM concentrations and support
our hypothesis that a DSC/DSE mechanism is involved atoms of residues within secondary structure.

Assembly and Function of the Dr Adhesins651
Figure 3. Topology Diagrams of the Atomic Structure and Proposed Topology for All Dr Family Adhesin Polymers
(A) AfaE-dsc. All � strands are shown in sky blue, with the exception of the self-complementing strand, which is sky blue/white. 310 helicesare shown in dark blue, and loops are shown as black lines.(B) Topology diagram for the proposed DSC/DSE-induced AfaE-III oligomer.
The final structure of the AfaE-dsc exhibits an immu- Earlier characterization of the Dr family adhesive organ-elles has demonstrated that the E protein not only func-noglobulin-like topology (Figures 2B and 3A), in which
the two � sheets pack against each other in a similar tions as the adhesin, which is required for interactionwith the host cell, but also as the pilin, which is responsi-fashion to archetypal pilin domains (Choudhury et al.,
1999; Sauer et al., 1999, 2002; Zavialov et al., 2003). ble for construction of the organelle. Our structure ofAfaE-dsc implies that the organelle is constructed via aDespite sequence identities of only �10%, AfaE-dsc
superimposes with an RMSD of 3.2 A over 105 equiva- DSC/DSE-driven polymerization of E subunits (Figure3B). Some fimbrial structures possess an adhesin atlent backbone C� atoms of PapE from the P pilus and
the capsular F1 antigen from Yersinia pestis, Caf1 (Sauer their tip that caps the growing fiber, implying that asimilar mechanism may exist for the so-called afimbrialet al., 2002; Zavialov et al., 2003). The structure of AfaE-
dsc reveals that the remodeled C terminus performs the adhesive organelle. To test this hypothesis, we investi-gated whether the D protein could be presented at therole of the complementing strand, thereby mimicking
the intermolecular interactions observed between pilin tip of the structure, where it can efficiently perform itsrole as an invasin.domains and consequently confirming the DSC/DSE
mechanism for the assembly of an AfaE polymer (Figures Protein sequence analysis reveals conservation be-tween a region within AfaD-III and the canonical � strand2 and 3). This provides the first high-resolution structural
evidence for the engineered self-complementation F from AfaE-III, which pairs with the donated Gd strand(Figure 4A). Therefore, it is reasonable to propose that(Barnhart et al., 2000). In AfaE-dsc, the key side chains
of Thr11(138) and Leu13(140) (numbering is as for the this donating Gd strand, i.e., the N terminus of matureAfaE-III, would also complement an incomplete AfaD-native [dsc] constructs; Figure 1B) from the comple-
menting strand are conserved throughout the Dr family III domain. To test this hypothesis, we performed anNMR titration experiment with AfaD-III and a peptideof adhesins and form the necessary interactions within
the core of the AfaE fold (Figures 2C and 2D). Further- corresponding to the N-terminal 18 amino acids of AfaE-III (i.e., AfaEN1–18). On addition of the AfaEN1–18 peptide, amore, low 1H-15N heteronuclear NOE values for the engi-
neered turn (residues 123–128 in AfaE-dsc) indicate that number of resonances shift to new chemical shift posi-tions indicative of a highly structured species with athis region is highly flexible and does not induce nonna-
tive structure (Figure 2E). specific complex formed between AfaD-III and AfaEN1–18.Moreover, gel filtration profiles and the reduction in linewidths for AfaD-III/AfaEN1–18 resonances clearly demon-Assembly and Capping of the Dr Family Adhesins
Type 1 and P pili form heteropolymeric structures com- strate the loss of AfaD-III aggregation (Figure 4B). Al-though AfaD does not possess an obvious N-terminalposed of a rigid stalk and a thin flexible extension (fibril-
lium) that is connected to the adhesive tip. The adhesin extension that might perform the role of a donatingstrand, we performed an identical titration with the firstdomain is connected to a truncated pilin subunit, which
accepts a donor strand and caps the tip of the fiber. 18 amino acids of AfaD (AfaDN1–18). Both NMR and gel
Molecular Cell652
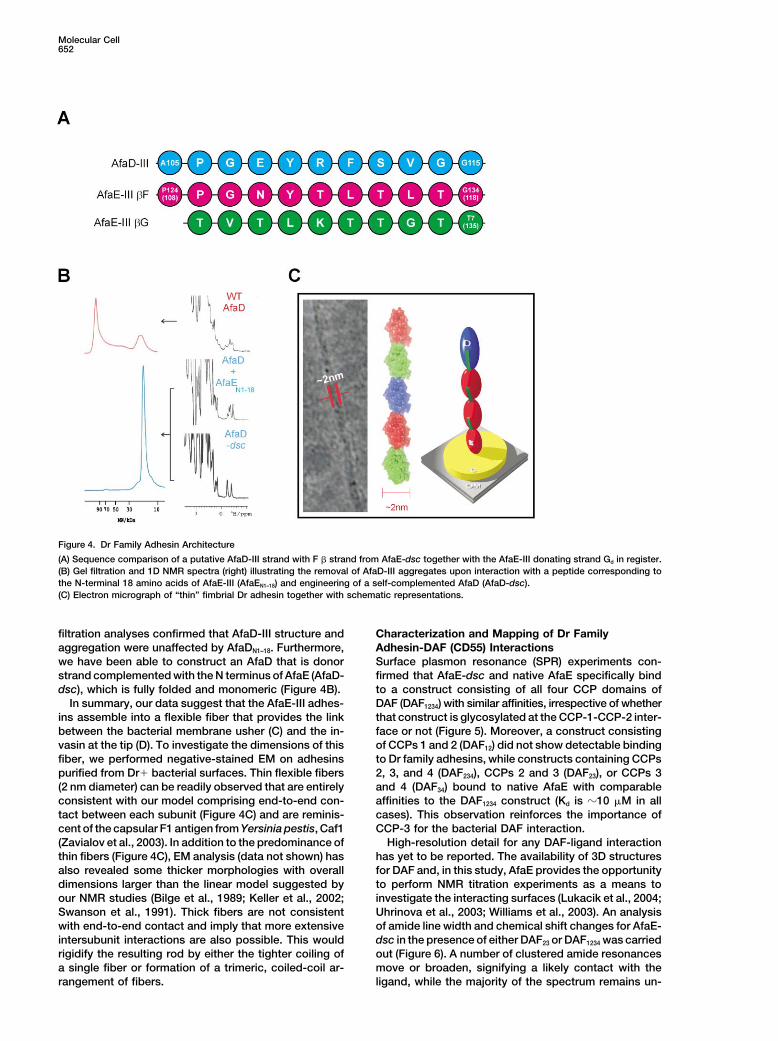
Figure 4. Dr Family Adhesin Architecture
(A) Sequence comparison of a putative AfaD-III strand with F � strand from AfaE-dsc together with the AfaE-III donating strand Gd in register.(B) Gel filtration and 1D NMR spectra (right) illustrating the removal of AfaD-III aggregates upon interaction with a peptide corresponding tothe N-terminal 18 amino acids of AfaE-III (AfaEN1–18) and engineering of a self-complemented AfaD (AfaD-dsc).(C) Electron micrograph of “thin” fimbrial Dr adhesin together with schematic representations.
filtration analyses confirmed that AfaD-III structure and Characterization and Mapping of Dr FamilyAdhesin-DAF (CD55) Interactionsaggregation were unaffected by AfaDN1–18. Furthermore,
we have been able to construct an AfaD that is donor Surface plasmon resonance (SPR) experiments con-firmed that AfaE-dsc and native AfaE specifically bindstrand complemented with the N terminus of AfaE (AfaD-
dsc), which is fully folded and monomeric (Figure 4B). to a construct consisting of all four CCP domains ofDAF (DAF1234) with similar affinities, irrespective of whetherIn summary, our data suggest that the AfaE-III adhes-
ins assemble into a flexible fiber that provides the link that construct is glycosylated at the CCP-1-CCP-2 inter-face or not (Figure 5). Moreover, a construct consistingbetween the bacterial membrane usher (C) and the in-
vasin at the tip (D). To investigate the dimensions of this of CCPs 1 and 2 (DAF12) did not show detectable bindingto Dr family adhesins, while constructs containing CCPsfiber, we performed negative-stained EM on adhesins
purified from Dr bacterial surfaces. Thin flexible fibers 2, 3, and 4 (DAF234), CCPs 2 and 3 (DAF23), or CCPs 3and 4 (DAF34) bound to native AfaE with comparable(2 nm diameter) can be readily observed that are entirely
consistent with our model comprising end-to-end con- affinities to the DAF1234 construct (Kd is �10 M in allcases). This observation reinforces the importance oftact between each subunit (Figure 4C) and are reminis-
cent of the capsular F1 antigen from Yersinia pestis, Caf1 CCP-3 for the bacterial DAF interaction.High-resolution detail for any DAF-ligand interaction(Zavialov et al., 2003). In addition to the predominance of
thin fibers (Figure 4C), EM analysis (data not shown) has has yet to be reported. The availability of 3D structuresfor DAF and, in this study, AfaE provides the opportunityalso revealed some thicker morphologies with overall
dimensions larger than the linear model suggested by to perform NMR titration experiments as a means toinvestigate the interacting surfaces (Lukacik et al., 2004;our NMR studies (Bilge et al., 1989; Keller et al., 2002;
Swanson et al., 1991). Thick fibers are not consistent Uhrinova et al., 2003; Williams et al., 2003). An analysisof amide line width and chemical shift changes for AfaE-with end-to-end contact and imply that more extensive
intersubunit interactions are also possible. This would dsc in the presence of either DAF23 or DAF1234 was carriedout (Figure 6). A number of clustered amide resonancesrigidify the resulting rod by either the tighter coiling of
a single fiber or formation of a trimeric, coiled-coil ar- move or broaden, signifying a likely contact with theligand, while the majority of the spectrum remains un-rangement of fibers.
Assembly and Function of the Dr Adhesins653
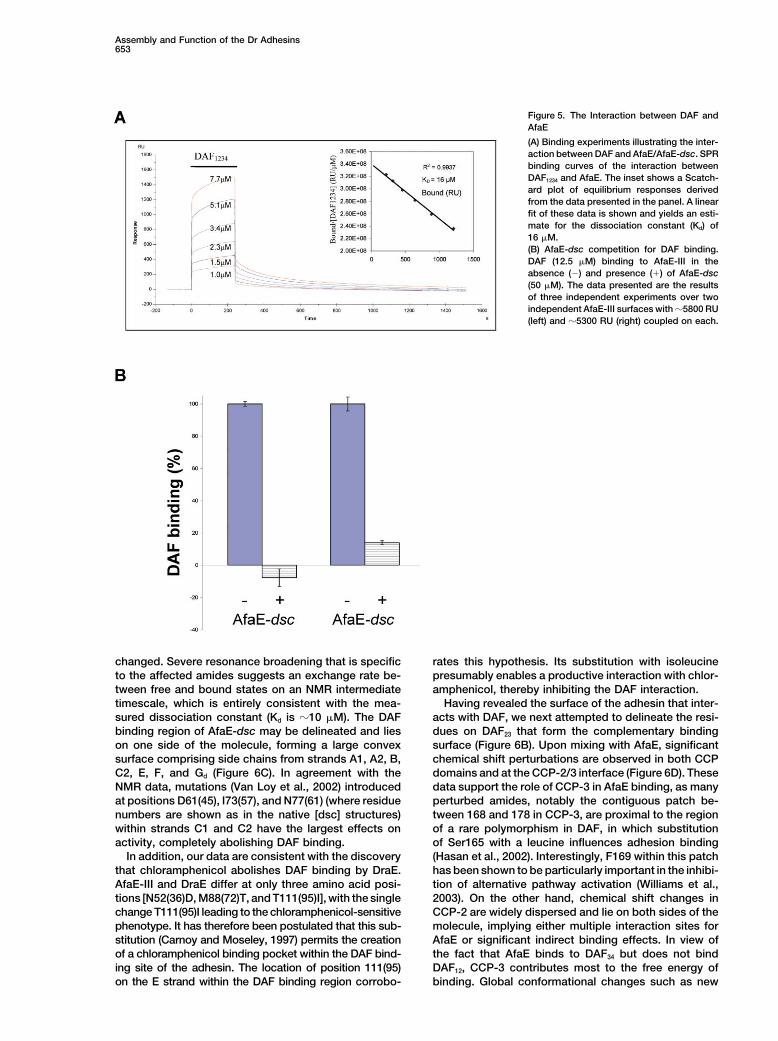
Figure 5. The Interaction between DAF andAfaE
(A) Binding experiments illustrating the inter-action between DAF and AfaE/AfaE-dsc. SPRbinding curves of the interaction betweenDAF1234 and AfaE. The inset shows a Scatch-ard plot of equilibrium responses derivedfrom the data presented in the panel. A linearfit of these data is shown and yields an esti-mate for the dissociation constant (Kd) of16 M.(B) AfaE-dsc competition for DAF binding.DAF (12.5 M) binding to AfaE-III in theabsence (�) and presence () of AfaE-dsc(50 M). The data presented are the resultsof three independent experiments over twoindependent AfaE-III surfaces with �5800 RU(left) and �5300 RU (right) coupled on each.
changed. Severe resonance broadening that is specific rates this hypothesis. Its substitution with isoleucinepresumably enables a productive interaction with chlor-to the affected amides suggests an exchange rate be-
tween free and bound states on an NMR intermediate amphenicol, thereby inhibiting the DAF interaction.Having revealed the surface of the adhesin that inter-timescale, which is entirely consistent with the mea-
sured dissociation constant (Kd is �10 M). The DAF acts with DAF, we next attempted to delineate the resi-dues on DAF23 that form the complementary bindingbinding region of AfaE-dsc may be delineated and lies
on one side of the molecule, forming a large convex surface (Figure 6B). Upon mixing with AfaE, significantchemical shift perturbations are observed in both CCPsurface comprising side chains from strands A1, A2, B,
C2, E, F, and Gd (Figure 6C). In agreement with the domains and at the CCP-2/3 interface (Figure 6D). Thesedata support the role of CCP-3 in AfaE binding, as manyNMR data, mutations (Van Loy et al., 2002) introduced
at positions D61(45), I73(57), and N77(61) (where residue perturbed amides, notably the contiguous patch be-tween 168 and 178 in CCP-3, are proximal to the regionnumbers are shown as in the native [dsc] structures)
within strands C1 and C2 have the largest effects on of a rare polymorphism in DAF, in which substitutionof Ser165 with a leucine influences adhesion bindingactivity, completely abolishing DAF binding.
In addition, our data are consistent with the discovery (Hasan et al., 2002). Interestingly, F169 within this patchhas been shown to be particularly important in the inhibi-that chloramphenicol abolishes DAF binding by DraE.
AfaE-III and DraE differ at only three amino acid posi- tion of alternative pathway activation (Williams et al.,2003). On the other hand, chemical shift changes intions [N52(36)D, M88(72)T, and T111(95)I], with the single
change T111(95)I leading to the chloramphenicol-sensitive CCP-2 are widely dispersed and lie on both sides of themolecule, implying either multiple interaction sites forphenotype. It has therefore been postulated that this sub-
stitution (Carnoy and Moseley, 1997) permits the creation AfaE or significant indirect binding effects. In view ofthe fact that AfaE binds to DAF34 but does not bindof a chloramphenicol binding pocket within the DAF bind-
ing site of the adhesin. The location of position 111(95) DAF12, CCP-3 contributes most to the free energy ofbinding. Global conformational changes such as newon the E strand within the DAF binding region corrobo-
Molecular Cell654
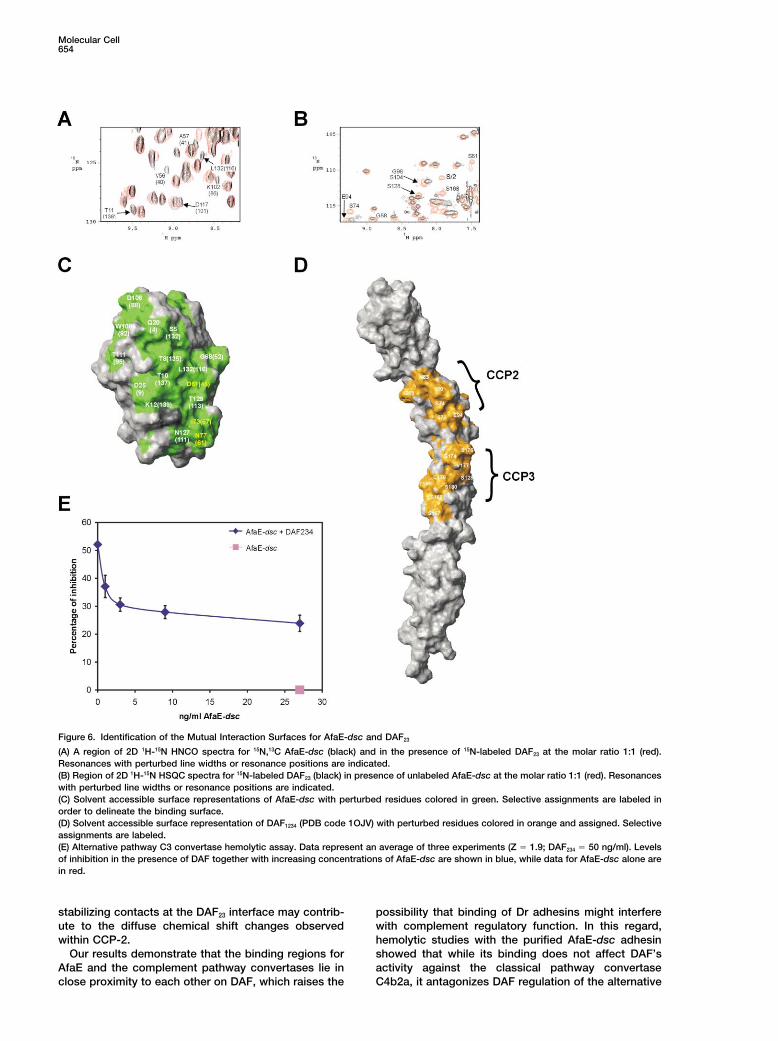
Figure 6. Identification of the Mutual Interaction Surfaces for AfaE-dsc and DAF23
(A) A region of 2D 1H-15N HNCO spectra for 15N,13C AfaE-dsc (black) and in the presence of 15N-labeled DAF23 at the molar ratio 1:1 (red).Resonances with perturbed line widths or resonance positions are indicated.(B) Region of 2D 1H-15N HSQC spectra for 15N-labeled DAF23 (black) in presence of unlabeled AfaE-dsc at the molar ratio 1:1 (red). Resonanceswith perturbed line widths or resonance positions are indicated.(C) Solvent accessible surface representations of AfaE-dsc with perturbed residues colored in green. Selective assignments are labeled inorder to delineate the binding surface.(D) Solvent accessible surface representation of DAF1234 (PDB code 1OJV) with perturbed residues colored in orange and assigned. Selectiveassignments are labeled.(E) Alternative pathway C3 convertase hemolytic assay. Data represent an average of three experiments (Z � 1.9; DAF234 � 50 ng/ml). Levelsof inhibition in the presence of DAF together with increasing concentrations of AfaE-dsc are shown in blue, while data for AfaE-dsc alone arein red.
stabilizing contacts at the DAF23 interface may contrib- possibility that binding of Dr adhesins might interferewith complement regulatory function. In this regard,ute to the diffuse chemical shift changes observed
within CCP-2. hemolytic studies with the purified AfaE-dsc adhesinshowed that while its binding does not affect DAF’sOur results demonstrate that the binding regions for
AfaE and the complement pathway convertases lie in activity against the classical pathway convertaseC4b2a, it antagonizes DAF regulation of the alternativeclose proximity to each other on DAF, which raises the

Assembly and Function of the Dr Adhesins655
a percentage. These data clearly show that AfaE-dsc effectivelypathway convertase C3bBb (Figure 6E). Thus, in addi-competes for DAF binding with both AfaE-III and DraE.tion to utilizing DAF as a convenient cellular attachment
point, it is possible that Dr adhesins block or exploitNMR Spectroscopy and Structure Calculationsome of the cellular functions of DAF—for example,Backbone and side chain assignments were completed using stan-
making host cells more susceptible to complement- dard double- and triple-resonance assignment methodology (Sattlermediated lysis may facilitate release and enhanced et al., 1999). H� and H� assignments were obtained using HBHA(CBC
ACO)NH (Sattler et al., 1999). The side chain assignments were com-spreading of bacteria.pleted using HCCH-total correlation (TOCSY) spectroscopy and(H)CC(CO)NH TOCSY (Sattler et al., 1999). 3D 1H-15N/13C NOESY-Concluding RemarksHSQC (mixing time 100 ms at 500 MHz and 800 MHz) experimentsThe solution structure of AfaE-dsc, in which a self-com-provided the distance restraints used in the final structure calcula-
plementing � strand was engineered into the C terminus, tion. Heteronuclear 1H-15N NOE data with minimal water saturationis strongly indicative of an analogous mode of assembly were acquired using the sequence described by Farrow et al. (1994).
A total of 321 long-range NOEs, providing unambiguous 3D infor-to that of fimbrial appendages. The E subunits assemblemation, were manually assigned from the NOESY data. The ARIAend to end to form thin filaments that are capped withprotocol (Linge et al., 2003) was used for completion of the NOEthe invasin (D) at the tip. The two-dimensional arrayassignment and structure calculation. A total of 4099 NOE-derivedimplied by the “afimbrial sheath” terminology for AfaEdistances were assigned from 13C- and 15N-edited spectra, which
presumably results from a collapse of the fine fibrillar comprised 2557 unambiguous and 1542 ambiguous restraints. Dihe-structures onto the bacterial surface. The structure of dral angle restraints derived from TALOS were also implemented.
The frequency window tolerance for assigning NOEs was �0.04AfaE, combined with binding studies and previouslyppm and �0.06 ppm for direct and indirect proton dimensionssolved structures of DAF, has allowed the mutual inter-and �0.7 ppm for nitrogen and carbon dimensions. The ARIA param-action surfaces of the E subunit/DAF complex to be de-eters p, Tv, and Nv were set to default values. The 15 lowest energyfined.structures had no NOE violations greater than 0.5 A and no dihedralangle violations greater than 5�. The structural statistics are pre-
Experimental Proceduressented in Table 1.
Cloning, Expression, and Purification of AfaE-dscChemical Shift Mapping for the AfaE and DAF Interactionand Native AfaE-IIIFor NMR mapping experiments, 15N,13C-labeled AfaE-dsc was pre-The construct AfaE-dsc was expressed cytoplasmically using thepared in 50 mM sodium acetate buffer at pH 5.3 at approximatelypRSETA plasmid (Promega) in the BL21(DE3) E. coli strain (Nova-30 M in 0.5 ml as mentioned above. 15N DAF in the same buffergen). 15N,13C double-labeled samples of AfaE-dsc were produced inwas introduced in several steps up to a molar ratio of 1:1, and 2Dminimal media, containing 0.07% 15NH4Cl and 0.2% 13C-glucose,15N-1H HNCO spectra were recorded at each stage under identicalsupplemented with 50 g/ml ampicillin. AfaE-dsc was purified inexperimental conditions. The titration was repeated with 15N-labeleddenaturing conditions (50 mM sodium phosphate buffer [pH 8.0],DAF and unlabeled AfaE-dsc, using the 2D 15N-1H HSQC experiment0.3 M NaCl, and 8 M guanidine hydrochloride) using the binding ofto detect the DAF amides. A further chemical shift mapping experi-the N-terminal hexahistidine tag to nickel bound agarose beads.ment was carried out as above using DAF1234 instead of DAF23.Purified protein was refolded by dialysis into 50 mM sodium acetate
buffer (pH 5.0) and concentrated to approximately 1 mM for NMR.Cell Adherence AssayNative AfaE-III was expressed as N-terminal hexahistidine fusionsThe coating of beads with AfaE-dsc and immunofluorescence ad-in E. coli strain M15 (Qiagen). The adhesins were purified from theherence assay were performed as described previously (Plancon etsupernatant using nickel affinity chromatography, followed by sizeal., 2003).exclusion chromatography in an S75 Sepharose column (Phar-
macia Biotech).Preparation and Analysis of Purified Dr FimbraeTo purify the natural, polymeric forms of the Dr adhesin, E. coli DH5Expression and Purification of DAF Constructs� (pCC90) (Carnoy and Moseley, 1997) was spread on dried surfacesDAF constructs were overexpressed either in Pichia pastorisof 10 LB agar plates (diameter 150 mm) with 100 g ampicillin per(DAF1234, DAF234, DAF12, DAF34 [Powell et al., 1997] and 15N-DAF23
ml and grown o/n at 37 C�. The bacteria were scraped from the[Uhrinova et al., 2003]) or refolded from E. coli (DAF1234 [White et al.,plates into 15 ml of phosphate buffered saline (PBS), washed with2004]). For the chemical shift perturbation experiments, 15N-DAF23
PBS, resuspended in 15 ml of PBS, and heated at 65 C� for 20 min.samples were prepared in 50 mM sodium phosphate buffer (pH 7.0)After 15 min of centrifugation at 18,000 g, the supernatant wasand concentrated to �0.5 mM.precipitated by adding an equal volume of 40% (w/v) ammoniumsulfate, gently mixed o/n at 4 C�, and pelleted at 18,000 g for 5 min.Surface Plasmon ResonanceThe pellet was resuspended and reprecipitated in 40% ammoniumNative AfaE-III was covalently coupled to the carboxylated dextransulfate and finally resuspended into 5 ml of 0.5% of sodium deoxy-matrix on the surface on an activated CM5 sensor chip using thecholate in 4 M urea, gently mixed o/n at 4 C�, and purified in Sepha-primary amine coupling kit (BIAcore AB). After activation accordingrose 4B column (Amersham Biosciences Corp., Piscataway, NJ)to the standard protocol, 0.15 mg/ml of AfaE-III in 10 mM sodiumwith the same buffer. The Dr adhesin preparation showed a hemag-acetate (pH 4.5) was injected. Differing levels were immobilizedglutination that was reversible by adding chloramphenicol at 10 M.(5,000–15,000 RU) by varying the volume of protein injected. AllFor electron microscopy, five microliter drops of the dispersed puri-interaction sensorgrams were collected at 20�C by flowing 80 l offied adhesin were absorbed to carbon-coated 200 mesh Ni grids.the various DAF constructs at a flow rate of 20 l/min.The grids were negatively stained with 2% phosphotungstic acidFor the competition experiments, AfaE-dsc remained in solution(pH 6.8) for 10 s, wick dried with filter paper, and observed in aand competed for DAF binding to AfaE-III bound on the chip surface.Philips 201 transmission electron microscope at 60 kV.The data presented are the results of three independent experiments
over two AfaE-III surfaces (5800 RU and 5300 RU coupled on eachsurface). Each experiment was performed as matched triplets where Alternative and Classical Pathway Activation Assays
Sheep erythrocytes (ESh; 1 109 ml�1) were sensitized with rabbitthree sequential injections were made of (1) 20 l of AfaE-dsc at aconcentration of 50 M, (2) 20 l of a mixture of DAF at a concentra- anti-sheep hemolysin (A) at 37�C for 30 min, followed by 4�C for 30
min. The resulting EShA (adjusted to 1 109 ml�1 in DGVB2) weretion of 12.5 M and AfaE-dsc at 50 M, and (3) 20 l of DAF at aconcentration of 12.5 M. Binding of DAF alone is shown as 100%, incubated at 30�C for 15 min with an equal volume of DGVB2 con-
taining 100 site-forming units (SFU) of C1. After washing, the re-and the binding obtained in the presence of AfaE-dsc is given as

Molecular Cell656
sulting EShAC1 (1 108 ml�1 in DGVB2) were incubated at 30�C for Garcia, M.I., Gounon, P., Courcoux, P., Labigne, A., and Le Bou-guenec, C. (1996). The afimbrial adhesive sheath encoded by the20 min with an equal volume of DGVB2 containing 100 SFU of C4
to yield EShAC14. For studies of classical pathway activation, 100 afa-3 gene cluster of pathogenic Escherichia coli is composed oftwo adhesins. Mol. Microbiol. 19, 683–693.l of EShAC14 (1 108 ml�1 in DGVB2) were incubated at 30�C for
5 min with 100 l of C2. Hemolytic sites then were developed by Gounon, P., Jouve, M., and Le Bouguenec, C. (2000). Immunocyto-addition of 1.3 ml of GVB-E containing 150 l guinea pig serum chemistry of the AfaE adhesin and AfaD invasin produced by patho-(C-EDTA) as a source of C3-9 and incubation at 37�C for 60 min. genic Escherichia coli strains during interaction of the bacteria with
For studies of alternative pathway activation, EShAC142 prepared HeLa cells by high-resolution scanning electron microscopy. Mi-as above (200 l at 5 107 ml�1 in DGVB2) were incubated at 30�C crobes Infect. 2, 359–365.for 20 min with 100 l of DGVB2 containing 2 SFU of C3. After Hasan, R.J., Pawelczyk, E., Urvil, P.T., Venkatarajan, M.S., Goluszko,washing, the resulting EShAC1423 (1 108 ml�1 in GVB-E) were P., Kur, J., Selvarangan, R., Nowicki, S., Braun, W.A., and Nowicki,incubated for 2–4 hr at 30�C until decay of C2 and C1 was complete B.J. (2002). Structure-function analysis of decay-accelerating factor:(as assessed by the absence of lysis upon addition of C2 and Identification of residues important for binding of the EscherichiaC-EDTA). The resulting E�shAC43 (1 108 ml�1 in DGVB2) then coli Dr adhesin and complement regulation. Infect. Immun. 70, 4485–were incubated at 30�C for 30 min with an equal volume of DGVB2
4493.containing 1.5 SFU of factor B and factor D. Hemolytic activity was
Keller, R., Ordonez, J.G., de Oliveira, R.R., Trabulsi, L.R., Baldwin,developed by addition of 1.3 ml of C-EDTA and incubation at 37�CT.J., and Knutton, S. (2002). Afa, a diffuse adherence fibrillar adhesinfor 1 hr. In all hemolytic assays, cells were pelleted and hemoglobinassociated with enteropathogenic Escherichia coli. Infect. Immun.color was read at 412 nm.70, 2681–2689.
Labigne-Roussel, A., and Falkow, S. (1988). Distribution and degreeCoordinates and Figure Preparationof heterogeneity of the afimbrial-adhesin-encoding operon (afa)Coordinates for the ensemble of NMR structures have been depos-among uropathogenic Escherichia coli isolates. Infect. Immun. 56,ited at the Protein Databank under the accession code 1RXL. Tables640–648.of NMR assignments and restraints are available as supplementary
material and have been deposited in the BioMagResBank in Madi- Labigne-Roussel, A., Lark, D., Schoolnik, G., and Falkow, S. (1984).son, WI (accession code 5947). Figures were prepared using pro- Cloning and expression of an afimbrial adhesin (AFA-I) responsiblegrams AESOP (Martin Noble, unpublished program), Jplot (Jeremy for P blood group-independent, mannose-resistant hemagglutina-Craven, unpublished program), and LIGPLOT (Wallace et al., 1995). tion from a pyelonephritic Escherichia coli strain. Infect. Immun.
46, 251–259.Acknowledgments Laskowski, R.A., Rullmann, J.A.C., MacArthur, M.W., Kaptein, R.,
and Thornton, J.M. (1996). AQUA and PROCHECK-NMR: ProgramsThe authors are indebted for the financial support of the Wellcome for checking the quality of protein structures solved by NMR. J.Trust (Research Leave Award to S.M.), BBSRC (43/B16601 to S.M.L.), Biomol. NMR 8, 477–486.Arthritis Research Campaign (L0534 to S.M.L.), MRC (Studentship Le Bouguenec, C., Garcia, M.I., Ouin, V., Desperrier, J.M., Gounon,to D.P.), EPSRC (Studentship to K.L.A.), and the NIH (AI23598 to P., and Labigne, A. (1993). Characterization of plasmid-borne afa-3M.E.M.). The authors would also like to thank Geoff Kelly and Tom gene clusters encoding afimbrial adhesins expressed by EscherichiaFrenkiel of the 800 MHz NMR service at NIMR. coli strains associated with intestinal or urinary-tract infections. In-
fect. Immun. 61, 5106–5114.Received: March 18, 2004
Le Bouguenec, C., Lalioui, L., Du Merle, L., Jouve, M., Courcoux,Revised: June 4, 2004P., Bouzari, S., Selvarangan, R., Nowicki, B.J., Germani, Y., Andre-Accepted: June 9, 2004mont, A., et al. (2001). Characterization of AfaE adhesins producedPublished: August 26, 2004by extraintestinal and intestinal human Escherichia coli isolates:PCR assays for detection of afa adhesins that do or do not recognizeReferencesDr blood group antigens. J. Clin. Microbiol. 39, 1738–1745.
Linge, J.P., Habeck, M., Rieping, W., and Nilges, M. (2003). ARIA:Barnhart, M.M., Pinkner, J.S., Soto, G.E., Sauer, F.G., Langermann,automated NOE assignment and NMR structure calculation. Bioin-S., Waksman, G., Frieden, C., and Hultgren, S.J. (2000). PapD-likeformatics 19, 315–316.chaperones provide the missing information for folding of pilin pro-
teins. Proc. Natl. Acad. Sci. USA 97, 7709–7714. Lukacik, P., Roversi, P., White, J., Esser, D., Smith, G.P., Billington,J., Williams, P., Rudd, P.M., Wormald, M.R., Crispin, M.D.M., et al.Bilge, S.S., Clausen, C.R., Lau, W., and Moseley, S.L. (1989). Molecu-(2004). Complement regulation at the molecular level: the structurelar characterization of a fimbrial adhesin, F1845, mediating diffuseof decay-accelerating factor. Proc. Natl. Acad. Sci. USA, in press.adherence of diarrhea-associated Escherichia coli to HEp-2 cells.
J. Bacteriol. 171, 4281–4289. Medof, M.E., Kinoshita, T., and Nussenzweig, V. (1984). Inhibitionof complement activation on the surface of cells after incorporationBrodbeck, W.G., Liu, D., Sperry, J., Mold, C., and Medof, M.E. (1996).of decay-accelerating factor (DAF) into their membranes. J. Exp.Localization of classical and alternative pathway regulatory activityMed. 160, 1558–1578.within the decay-accelerating factor. J. Immunol. 156, 2528–2533.Medof, M.E., Kinoshita, T., Silber, R., and Nussenzweig, V. (1985).Caras, I.W., Davitz, M.A., Rhee, L., Weddell, G., Martin, D.W., andAmelioration of lytic abnormalities of paroxysmal-nocturnal hemo-Nussenzweig, V. (1987). Cloning of decay-accelerating factor sug-globinuria with decay-accelerating factor. Proc. Natl. Acad. Sci. USAgests novel use of splicing to generate two proteins. Nature 325,82, 2980–2984.545–549.Medof, M.E., Lublin, D.M., Holers, V.M., Ayers, D.J., Getty, R.R.,Carnoy, C., and Moseley, S.L. (1997). Mutational analysis of receptorLeykam, J.F., Atkinson, J.P., and Tykocinski, M.L. (1987). Cloningbinding mediated by the Dr family of Escherichia coli adhesins. Mol.and characterization of cDNAs encoding the complete sequence ofMicrobiol. 23, 365–379.decay-accelerating factor of human complement. Proc. Natl. Acad.Choudhury, D., Thompson, A., Stojanoff, V., Langermann, S., Pink-Sci. USA 84, 2007–2011.ner, J., Hultgren, S.J., and Knight, S.D. (1999). X-ray structure ofNowicki, B., Moulds, J., Hull, R., and Hull, S. (1988). A hemagglutininthe FimC-FimH chaperone-adhesin complex from uropathogenicof uropathogenic Escherichia coli recognizes the Dr blood groupEscherichia coli. Science 285, 1061–1066.antigen. Infect. Immun. 56, 1057–1060.
Farrow, N.A., Muhandiram, R., Singer, A.U., Pascal, S.M., Kay, C.M.,Nowicki, B., Svanborgeden, C., Hull, R., and Hull, S. (1989). Molecu-Gish, G., Shoelson, S.E., Pawson, T., Formankay, J.D., and Kay,lar analysis and epidemiology of the Dr hemagglutinin of uropatho-L.E. (1994). Backbone dynamics of a free and phosphopeptide-genic Escherichia coli. Infect. Immun. 57, 446–451.complexed Src homology 2 domain studied by 15N NMR relaxation.
Biochemistry 33, 5984–6003. Nowicki, B., Selvarangan, R., and Nowicki, S. (2001). Family of Esch-

Assembly and Function of the Dr Adhesins657
erichia coli Dr adhesins: Decay-accelerating factor receptor recogni- the capsular F1 antigen from Yersinia pestis: Preserved folding en-ergy drives fiber formation. Cell 113, 587–596.tion and invasiveness. J. Infect. Dis. 183, S24–S27.
Pham, T., Kaul, A., Hart, A., Goluszko, P., Moulds, J., Nowicki, S.,Lublin, D.M., and Nowicki, B.J. (1995). dra-related X adhesins ofgestational pyelonephritis-associated Escherichia coli recognizeSCR-3 and SCR-4 domains of recombinant decay-accelerating fac-tor. Infect. Immun. 63, 1663–1668.
Plancon, L., du Merle, L., Le Friec, S., Gounon, P., Jouve, M., Guig-not, J., Servin, A., and Le Bouguenec, C. (2003). Recognition of thecellular �1-chain integrin by the bacterial AfaD invasin is implicatedin the internalization of afa-expressing pathogenic Escherichia colistrains. Cell. Microbiol. 5, 681–693.
Powell, R.M., Ward, T., Evans, D.J., and Almond, J.W. (1997). Interac-tion between echovirus 7 and its receptor, decay-accelerating factor(CD55): evidence for a secondary cellular factor in A-particle forma-tion. J. Virol. 71, 9306–9312.
Sattler, M., Schleucher, J., and Griesinger, C. (1999). Heteronuclearmultidimensional NMR experiments for the structure determinationof proteins in solution employing pulsed field gradients. Progressin NMR Spectroscopy 34, 93–158.
Sauer, F.G., Futterer, K., Pinkner, J.S., Dodson, K.W., Hultgren, S.J.,and Waksman, G. (1999). Structural basis of chaperone functionand pilus biogenesis. Science 285, 1058–1061.
Sauer, F.G., Barnhart, M., Choudhury, D., Knights, S.D., Waksman,G., and Hultgren, S.J. (2000a). Chaperone-assisted pilus assemblyand bacterial attachment. Curr. Opin. Struct. Biol. 10, 548–556.
Sauer, F.G., Mulvey, M.A., Schilling, J.D., Martinez, J.J., and Hult-gren, S.J. (2000b). Bacterial pili: molecular mechanisms of patho-genesis. Curr. Opin. Microbiol. 3, 65–72.
Sauer, F.G., Pinkner, J.S., Waksman, G., and Hultgren, S.J. (2002).Chaperone priming of pilus subunits facilitates a topological transi-tion that drives fiber formation. Cell 111, 543–551.
Selvarangan, R., Goluszko, P., Popov, V., Singhal, J., Pham, T.,Lublin, D.M., Nowicki, S., and Nowicki, B. (2000). Role of decay-accelerating factor domains and anchorage in internalization of Dr-fimbriated Escherichia coli. Infect. Immun. 68, 1391–1399.
Soto, G.E., and Hultgren, S.J. (1999). Bacterial adhesins: commonthemes and variations in architecture and assembly. J. Bacteriol.181, 1059–1071.
Swanson, T.N., Bilge, S.S., Nowicki, B., and Moseley, S.L. (1991).Molecular structure of the Dr adhesin nucleotide sequence andmapping of receptor-binding domain by use of fusion constructs.Infect. Immun. 59, 261–268.
Uhrinova, S., Lin, F., Ball, G., Bromek, K., Uhrin, D., Medof, M.E.,and Barlow, P.N. (2003). Solution structure of a functionally activefragment of decay-accelerating factor. Proc. Natl. Acad. Sci. USA100, 4718–4723.
Van Loy, C.P., Sokurenko, E.V., Samudrala, R., and Moseley, S.L.(2002). Identification of amino acids in the Dr adhesin required forbinding to decay-accelerating factor. Mol. Microbiol. 45, 439–452.
Wallace, A.C., Laskowski, R.A., and Thornton, J.M. (1995). LIGPLOT:A program to generate schematic diagrams of protein-ligand inter-actions. Protein Eng. 8, 127–134.
Westerlund, B., Kuusela, P., Risteli, J., Risteli, L., Vartio, T., Rauvala,H., Virkola, R., and Korhonen, T.K. (1989). The O75X adhesin ofuropathogenic Escherichia coli is a type IV collagen-binding protein.Mol. Microbiol. 3, 329–337.
White, J., Lukacik, P., Esser, D., Steward, M., Giddings, N., Bright,J.R., Fritchley, S.J., Morgan, B.P., Lea, S.M., Smith, G.P., and Smith,R.A.G. (2004). Biological activity, membrane-targeting modificationand crystallization of soluble human decay accelerating factor ex-pressed in E. coli. Protein Sci. 13, in press.
Williams, P., Chaudhry, Y., Goodfellow, I.G., Billington, J., Powell,R., Spiller, O.B., Evans, D.J., and Lea, S. (2003). Mapping CD55function. The structure of two pathogen-binding domains at 1.7 A.J. Biol. Chem. 278, 10691–10696.
Zavialov, A.V., Berglund, J., Pudney, A.F., Fooks, L.J., Ibrahim, T.M.,MacIntyre, S., and Knight, S.D. (2003). Structure and biogenesis of
Top Related
Copyright © 2022 FDOKUMEN

