







Bahasa
Halaman
Hukum


PRIMARY RESEARCH PAPER
Ammonium threshold values for groundwater qualityin the EU may not protect groundwater fauna: evidencefrom an alluvial aquifer in Italy
T. Di Lorenzo • M. Cifoni • P. Lombardo •
B. Fiasca • D. M. P. Galassi
Received: 10 June 2014 / Revised: 19 August 2014 / Accepted: 21 August 2014 / Published online: 29 August 2014
� Springer International Publishing Switzerland 2014
Abstract The European Union (EU) threshold val-
ues for NHþ4 in groundwater range from a minimum of
0.084 mg l-1 to the maximum allowed of 5 mg l-1.
The aim of our study was to determine whether these
values are adequate to protect groundwater copepods
in alluvial aquifers underlying intensive agriculture.
To this end, we analyzed abiotic (including NHþ4concentration) and biological patterns (copepod
assemblages) in an alluvial aquifer in an area of
intensive agriculture. Groundwater was collected from
pre-existing farmer-owned bores. Abiotic and biolog-
ical patterns were not related to seasonal variation in
agricultural practices, and pollutant concentrations
were typically below the legal maximum threshold
values. However, both abiotic and biological variables
differed significantly between two groups of bores,
with high and low NHþ4 conditions based on separate
ecotoxicological assays that set the NHþ4 toxicity
threshold for stygobiotic copepods at 2.59 below the
current EU lowest legal threshold. In particular,
phosphorus concentration was higher, and oxygen
and copepod abundance and richness were lower in
high NHþ4 bores. Our results suggest that the present
threshold value range for NHþ4 adopted by EU
Member States may not protect stygobiotic copepod
assemblages in alluvial aquifers underlying densely
cultivated areas.
Keywords Ammonium � Aquifer � Stygobiotic �Toxicity � Groundwater � Threshold
Introduction
The use of N-fertilizers in excess of crop needs is a
widespread concern in developed and developing
countries as, generally, no more than 50% of the
applied nitrogen (N) is taken up by crops (Mosier
et al., 2004). The unused N can be lost from the soil–
plant system by volatilization as NH3, by denitrifica-
tion as reduced form NHþ4 , and by leaching as the
oxidized forms NO�3 and NO�2 (Erisman et al., 2007).
The excess of N-fertilizers (both reduced and oxidized
forms), not removed by crops, leaches into
Handling editor: Diego Fontaneto
Electronic supplementary material The online versionof this article (doi:10.1007/s10750-014-2018-y) containssupplementary material, which is available to authorized users.
T. Di Lorenzo (&) � M. Cifoni
Istituto per lo Studio degli Ecosistemi - CNR, Via
Madonna del Piano 10, Sesto Fiorentino, 50019 Florence,
Italy
e-mail: [email protected]
M. Cifoni � P. Lombardo � B. Fiasca � D. M. P. Galassi
Department of Life, Health and Environmental Sciences,
University of L’Aquila, Via Vetoio, Coppito,
67100 L’Aquila, Italy
P. Lombardo
Limno Consulting, via Bedollo 303, 00124 Rome, Italy
123
Hydrobiologia (2015) 743:139–150
DOI 10.1007/s10750-014-2018-y

groundwater or reaches surface waters through runoff,
polluting water bodies. Therefore, intensive agricul-
ture is often associated with high concentrations of
nitrogen (N) compounds in groundwater (Di Lorenzo
et al., 2012; Melo et al., 2012; Morari et al., 2012;
Dahan et al., 2014), due to the use of fertilizers in the
forms of ammonium nitrate, urea, calcium nitrate,
ammonium bicarbonate, and mixtures of nitrogen,
potassium, and phosphorus (Erisman et al., 2007).
Like surface water ecosystems (e.g., Carpenter et al.,
1998; Tilman, 1999; Blann et al., 2009), groundwater
ecosystems are sensitive to changes in water quality
induced by agricultural practices (Wood et al., 2008;
Hartland et al., 2011; Korbel et al., 2013; Di Lorenzo
& Galassi, 2013; Di Lorenzo et al., 2013). Pollution
from pesticides and fertilizers and groundwater with-
drawal for irrigation purposes remain the major causes
of poor groundwater quality in several European
alluvial aquifers (Candela et al., 2008; European
Union, 2012; van Grinsven et al., 2012; Di Lorenzo &
Galassi, 2013; Caschetto et al., 2014; Poch-Massegu
et al., 2014).
Groundwater biota (i.e., stygobiotic organisms,
defined as species that are strictly confined to
groundwater and hence have evolved specific mor-
phological and physiological adaptations to cope with
the unique subterranean habitat: Gibert et al., 1994) is
essential to maintain groundwater ecosystem health
(Stein et al., 2010 and references therein). Both
groundwater microbes (such as bacteria, fungi, and
protozoa) and stygofauna (macro- and meiofaunal
invertebrates) provide important ecosystem services
which support aquifer self-purification (Kota et al.,
1999; Marshall & Hall, 2004; Boulton et al., 2008).
Stygobiotic copepods (by far the most abundant and
species-rich invertebrate group in groundwater: Ga-
lassi, 2001; Galassi et al., 2009) are highly sensitive to
NHþ4 (Di Lorenzo et al., 2014) as observed for several
other freshwater invertebrates (Jensen, 1995, 2003;
Weihrauch et al., 2004; Camargo et al., 2005;
Camargo & Alonso, 2006). However, the utility of
groundwater copepods as indicators of NHþ4 contam-
ination in groundwater is yet to be established. In the
laboratory, Di Lorenzo et al. (2014) found that the
NHþ4 chronic lethal concentration (CL) for the stygo-
biotic species Diacyclops belgicus Kiefer, 1936
(Copepoda Cyclopidae) is 0.032 mg l-1. This con-
centration is 2.59 lower than the more stringent
threshold value (TV) indicated by EU Member States
(MS) (European Commission, 2010). However, it is
unknown whether exposure to such NHþ4 concentra-
tion may affect other stygobiotic species of copepods
in their natural habitats.
The aim of our study was to determine whether
stygobiotic copepod assemblages may be affected, in
terms of species richness and/or abundance, by
exposure in natural groundwater habitats to NHþ4concentration equal to or higher than the laboratory-
determined CL concentration (0.032 mg l-1) assessed
for the stygobiotic D. belgicus by Di Lorenzo et al.
(2014). To this end, we analyzed biological data from
an alluvial aquifer in an area of intensive agriculture,
in Italy. We selected the alluvial aquifer of the River
Adige, because of its homogeneous geological and
hydromorphological features and absence of contam-
ination according to the TVs indicated in the Annexes
I and II of the Groundwater Daughter Directive 2006/
118/EC (European Commission, 2006). We tested the
hypotheses that: (1) stygobiotic copepod assemblages
are affected by environmental NHþ4 concentrations
C0.032 mg l-1; and (2) the current TVs for NHþ4 ,
indicated by EU Member States for groundwater
bodies (European Commission, 2010), are inadequate
to protect groundwater copepods in alluvial aquifers
underlying intensive agriculture.
Materials and methods
Study region
The study was carried out in the northernmost sector
(45 km2) of the alluvial plain of the River Adige in the
Trento Province, Italy (Fig. 1). The floodplain is
intensely used for agriculture, while industrial and
urban land use is less extensive. Floodplain soils are
similar across the study area, with sequences of clay,
sand, and gravel sediments (Distretto Idrografico delle
Alpi Orientali, 2010). The alluvials are 50–60 m thick
and overlie an impermeable bedrock. The distribution
of Quaternary sediments filling the plain varies
through the area with fine material (silt, clay) covered
by larger-grained sediments (gravel and sand), pro-
ducing a complex, locally multi-layered structure with
a hydraulic conductivity of 10-3–10-5 m s-1 (Dist-
retto Idrografico delle Alpi Orientali, 2010). However,
140 Hydrobiologia (2015) 743:139–150
123

the aquifer layers are hydrogeologically intercon-
nected, leading to the physico-chemical homogeniza-
tion of groundwater (Distretto Idrografico delle Alpi
Orientali, 2010). The aquifer is fed by rainfalls and
lateral discharge from the surrounding calcareous
groundwater bodies (Autorita di Bacino dell’Adige,
2008). Mean annual discharge of the aquifer is about
9 m3 s-1 (Autorita di Bacino dell’Adige, 2008). The
climate is continental, with cold winters and maxi-
mum rainfall typically in summer. The average annual
rainfall in the floodplain is about 400–500 mm year-1
(Distretto Idrografico delle Alpi Orientali, 2010).
Agriculture (predominantly orchards, especially the
renowned local apple trees) is the main activity
([85 % of the study area), with much less-extensive
industrial and urban land use, mainly within or
adjacent to agricultural areas. The aquifer is exploited
for orchard irrigation (mainly by sprinkle and drip
methods) from March through the end of September,
and to a lesser extent over the whole year for industrial
and urban uses (Distretto Idrografico delle Alpi
Orientali, 2010). Groundwater abstraction is sustain-
able (Autorita di Bacino dell’Adige, 2008). Orchard
cultivation requires nitrogen, phosphorus, and potas-
sium fertilizers, which are applied in spring and
summer, as well as herbicides and pesticides. Both
organic and synthetic fertilizers are used at a ratio of
about 10:1 (ISTAT, 2006).
Sampling regime
Groundwater biota and water samples were collected
in 33 bores, in June and again in October 2006. Study
bores were randomly selected from a pool of privately
owned bores drilled in aquifer layers of mixed gravel
and sands, and already in use for irrigation purposes.
Farmers did not use the bores during the week before
the planned sampling to allow the piezometric levels
to restore. We selected bores of various depth
(maximum depth: about 60 m), 16–20 cm in diameter,
lined with PVC casing, with a 2-m slotted screen at the
base. The range in bore depth allowed us to sample the
entire aquifer depth-wise. The distance of the study
bores from the River Adige was random, thus prox-
imity to the river was not included in our analyses as a
factor.
Sampling
Bores were equipped with immersion pumps that were
used to extract bore water, which was filtered through
a 60-lm mesh net to collect groundwater fauna. After
preliminary tests, representative samples of bore and
aquifer taxonomic richness and composition were
collected by pumping 1 m3 of groundwater (2–4 times
the bore volumes) from each bore. After fixation in the
field with 70% ethanol, specimens were sorted under a
stereomicroscope and identified to class/order level in
the laboratory, except for copepods that were identi-
fied to species. Each species, according to the degree
of adaptation to the groundwater environment (Galas-
si, 2001), was listed as stygobite (SB: species strictly
associated to groundwater habitats, where they com-
plete the life cycle) or as non-stygobite (nSB),
Fig. 1 Map of sampled bores in areas of different land use in
the River Adige alluvial aquifer, Italy (white agricultural, gray
wild vegetation, striped areas: urban areas)
Hydrobiologia (2015) 743:139–150 141
123

including stygophiles (capable of living in both
surface and groundwater habitats) and stygoxenes
(epigean species which accidentally or occasionally
enter groundwater).
After the biological sampling, specific electrical
conductivity (SEC), pH, and temperature of ground-
water were measured by a multiparametric probe
(ECM MultiTM; Dr. Lange GmbH, Dusseldorf,
Germany) in a vessel directly after pumping for three
times. During measurements, a continuous groundwa-
ter flow was ensured across the probe. Water samples
to be tested for other chemical parameters in the
laboratory were set aside after fauna had been
removed. Samples were transported to the laboratory
in a cooler at 4�C within a few hours after collection.
Groundwater quality analyses were performed by the
local environmental protection agency (APPAT).
Groundwater samples were analyzed for 30 pesti-
cides/herbicides, metals (Cu2þ and Zn2þ), calcium,
total hardness, chloride, sulfate, nitrate, nitrite, NHþ4 ,
total N, and SRP (soluble reactive phosphorus, an
operationally defined P fraction which is, to a large
extent, equivalent to ortho-phosphate).
Data analysis
We grouped bores according to the mean level of NHþ4in bore groundwater. The H (high NHþ4 concentration)
group included 16 bores with a mean NHþ4C0.032 mg l-1; the L (low NHþ4 concentration) group
included 17 bores with a mean NHþ4 \0.032 mg l-1,
where 0.032 mg l-1 is the chronic lethal concentration
of NHþ4 determined for the stygobiotic D. belgicus (Di
Lorenzo et al., 2014). Groundwater quality parame-
ters, richness, and abundance of copepods, were
analyzed using one-way analysis of variance
(ANOVA) between the two groups of bores.
As subsequent samples from the same sampling
unit (bore) were considered temporal pseudorepli-
cates according to Hurlbert (1984), in ANOVAs we
considered the mean value (data of June and
October) of each abiotic parameter and the sum of
the number of copepod individuals in the two
sampling surveys, for each sampling unit. To test if
the seasonality of the agricultural activities (irriga-
tion and fertilizer/pesticide applications) produced an
effect on groundwater quality, a one-sample t test
was performed for each groundwater quality param-
eter on the difference between June (i.e., before the
expected peak in irrigation and during fertilizer
application) and October sampling (after both the
peak in irrigation and fertilizer application), for each
bore. For the one-sample t test, we assumed a null
hypothesis that there was no difference between the
mean of each quality parameter and zero. ANOVAs
and one-sample t tests were applied only to variables
with normally distributed data (verified with the
Shapiro test). Variables with homogeneous variances
(verified with the Bartlett test) were tested with
untransformed data; data for variables with hetero-
geneous variances were log-transformed before
ANOVAs and/or one-sample t tests. Univariate
analyses were performed using R software v. 2.15.0
(R Development Core Team, 2008).
Patterns in water quality were also examined with
a multivariate approach using principal components
analysis (PCA). Patterns of copepod assemblages
were examined using non-metric multidimensional
scaling (nMDS) incorporating the Bray–Curtis sim-
ilarity coefficient. The BEST (Bio-Env ? Stepwise)
permutational procedure (Clarke et al., 2008) was
applied to examine whether the multivariate biotic
and environmental variables were correlated under
the null hypothesis of complete independence of
biotic and environmental patterns. Permutational
analyses of variance (PERMANOVA; Anderson
et al., 2008) were used to test for differences in
water quality conditions and copepod assemblages
with SEASON (levels: J = June and O = October)
and bore NHþ4 concentration (levels: H = high and
L = low) as orthogonal and fixed factors (number of
permutations = 9,999). SIMPER analysis (Clarke,
1993) was used to identify the species that contributed
mostly to the differences between groups. In nMDS,
BEST, and PERMANOVA, copepod abundance was
square-root transformed and a dummy variable of 1
was added to all samples to facilitate the inclusion of
otherwise empty (zero abundance) samples. Levene
tests were performed in order to check for homoge-
neity of variances. Water quality variables were
normalized before PCA, BEST, and PERMANOVA.
Multivariate analyses were performed using E Primer
version 6 (Clarke & Gorley, 2006). All statistical
analyses were conducted with a significance level (a)
of 0.05.
142 Hydrobiologia (2015) 743:139–150
123
Results
Abiotic variables
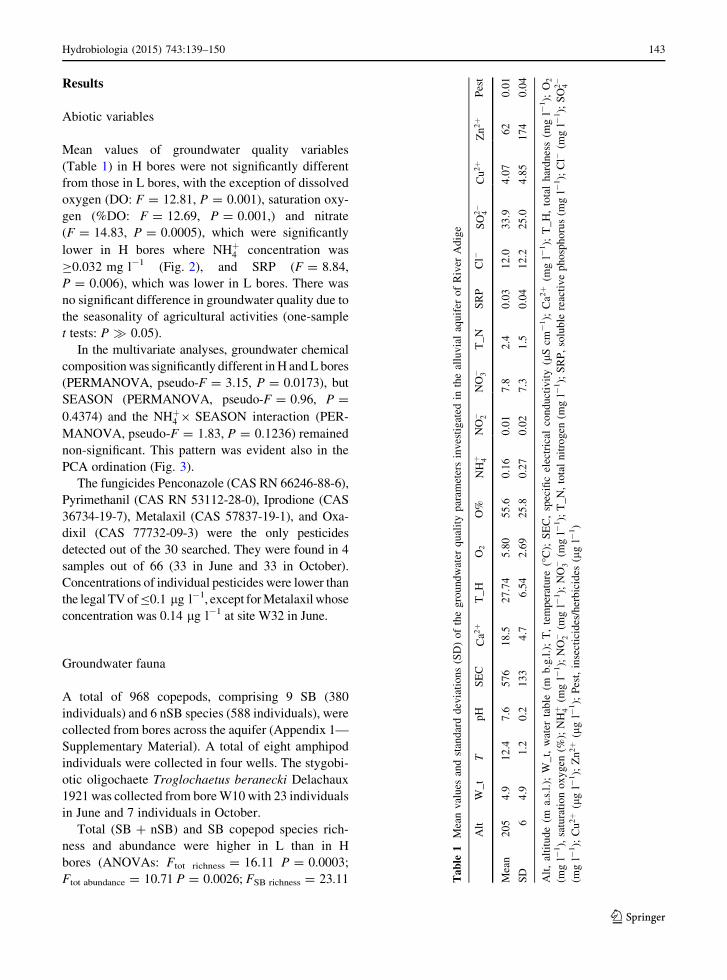
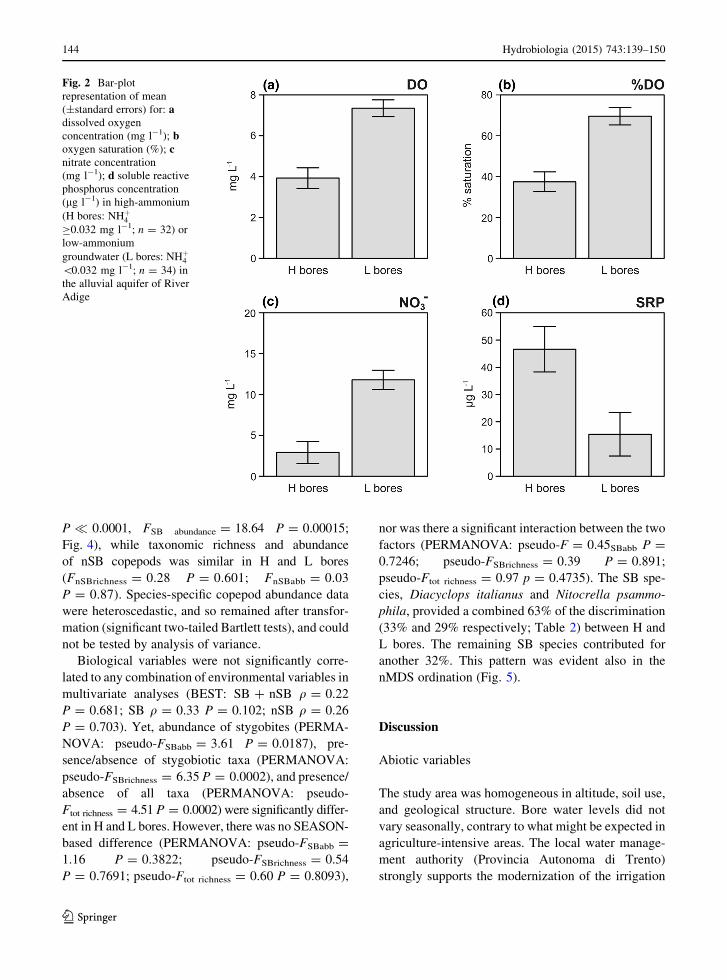
Mean values of groundwater quality variables
(Table 1) in H bores were not significantly different
from those in L bores, with the exception of dissolved
oxygen (DO: F = 12.81, P = 0.001), saturation oxy-
gen (%DO: F = 12.69, P = 0.001,) and nitrate
(F = 14.83, P = 0.0005), which were significantly
lower in H bores where NHþ4 concentration was
C0.032 mg l-1 (Fig. 2), and SRP (F = 8.84,
P = 0.006), which was lower in L bores. There was
no significant difference in groundwater quality due to
the seasonality of agricultural activities (one-sample
t tests: P � 0.05).
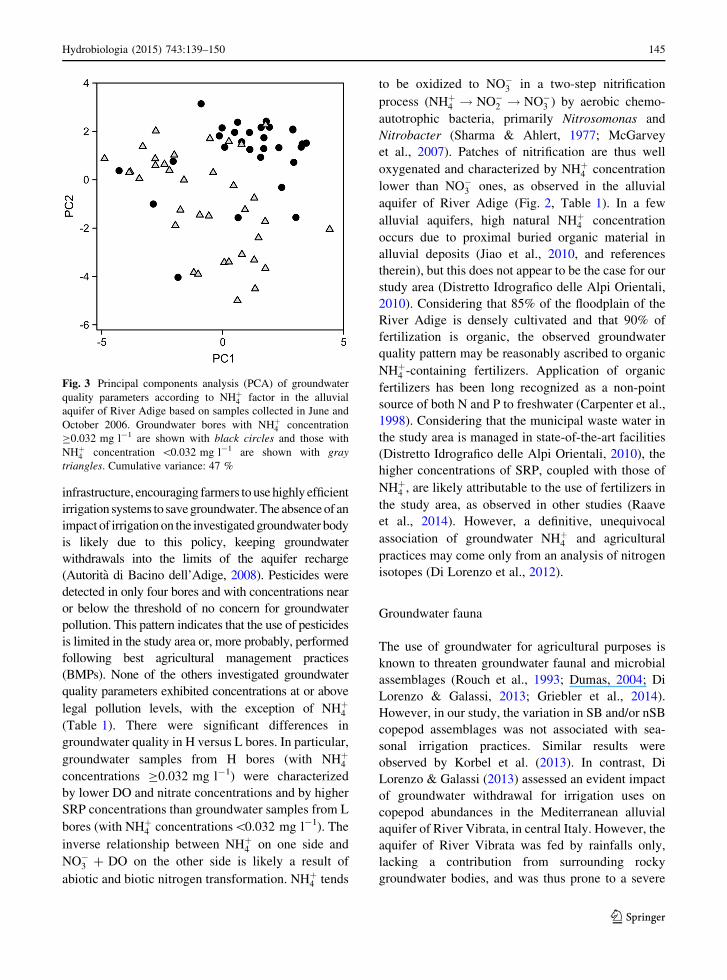
In the multivariate analyses, groundwater chemical
composition was significantly different in H and L bores
(PERMANOVA, pseudo-F = 3.15, P = 0.0173), but
SEASON (PERMANOVA, pseudo-F = 0.96, P =
0.4374) and the NHþ4 9 SEASON interaction (PER-
MANOVA, pseudo-F = 1.83, P = 0.1236) remained
non-significant. This pattern was evident also in the
PCA ordination (Fig. 3).
The fungicides Penconazole (CAS RN 66246-88-6),
Pyrimethanil (CAS RN 53112-28-0), Iprodione (CAS
36734-19-7), Metalaxil (CAS 57837-19-1), and Oxa-
dixil (CAS 77732-09-3) were the only pesticides
detected out of the 30 searched. They were found in 4
samples out of 66 (33 in June and 33 in October).
Concentrations of individual pesticides were lower than
the legal TV of B0.1 lg l-1, except for Metalaxil whose
concentration was 0.14 lg l-1 at site W32 in June.
Groundwater fauna
A total of 968 copepods, comprising 9 SB (380
individuals) and 6 nSB species (588 individuals), were
collected from bores across the aquifer (Appendix 1—
Supplementary Material). A total of eight amphipod
individuals were collected in four wells. The stygobi-
otic oligochaete Troglochaetus beranecki Delachaux
1921 was collected from bore W10 with 23 individuals
in June and 7 individuals in October.
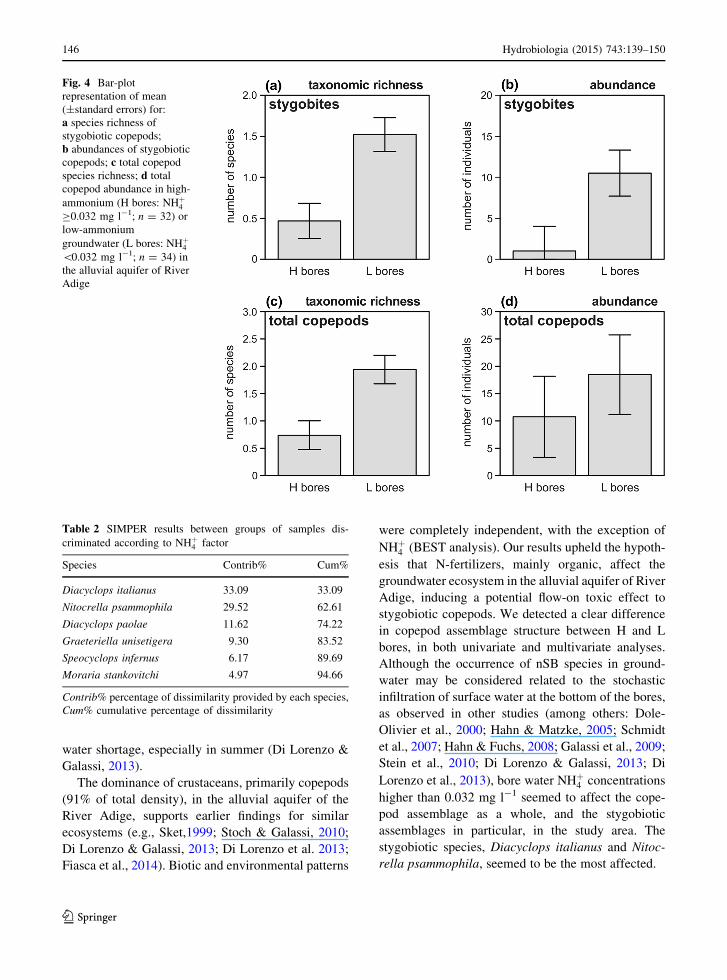
Total (SB ? nSB) and SB copepod species rich-
ness and abundance were higher in L than in H
bores (ANOVAs: Ftot richness = 16.11 P = 0.0003;
Ftot abundance = 10.71 P = 0.0026; FSB richness = 23.11 Ta
ble
1M
ean
val
ues
and
stan
dar
dd
evia
tio
ns
(SD
)o
fth
eg
rou
nd
wat
erq
ual
ity
par
amet
ers
inv
esti
gat
edin
the
allu
via
laq
uif
ero
fR
iver
Ad
ige
Alt
W_
tT
pH
SE
CC
a2þ
T_
HO
2O
%N
Hþ 4
NO� 2
NO� 3
T_
NS
RP
Cl�
SO
2�
4C
u2þ
Zn
2þ
Pes
t
Mea
n2
05
4.9
12
.47
.65
76
18
.52
7.7
45
.80
55
.60
.16
0.0
17
.82
.40
.03
12
.03
3.9
4.0
76
20
.01
SD
64
.91
.20
.21
33
4.7
6.5
42
.69
25
.80
.27
0.0
27
.31
.50
.04
12
.22
5.0
4.8
51
74
0.0
4
Alt
,al
titu
de
(ma.
s.l.
);W
_t,
wat
erta
ble
(mb
.g.l
.);
T,
tem
per
atu
re(�
C);
SE
C,
spec
ific
elec
tric
alco
nd
uct
ivit
y(l
Scm
-1);
Ca2þ
(mg
l-1);
T_
H,
tota
lh
ard
nes
s(m
gl-
1);
O2
(mg
l-1),
satu
rati
on
ox
yg
en(%
);N
Hþ 4
(mg
l-1);
NO� 2
(mg
l-1);
NO� 3
(mg
l-1);
T_
N,
tota
ln
itro
gen
(mg
l-1);
SR
P,
solu
ble
reac
tiv
ep
ho
sph
oru
s(m
gl-
1);
Cl�
(mg
l-1);
SO
2�
4
(mg
l-1);
Cu
2þ
(lg
l-1);
Zn
2þ
(lg
l-1);
Pes
t,in
sect
icid
es/h
erb
icid
es(l
gl-
1)
Hydrobiologia (2015) 743:139–150 143
123
P � 0.0001, FSB abundance = 18.64 P = 0.00015;
Fig. 4), while taxonomic richness and abundance
of nSB copepods was similar in H and L bores
(FnSBrichness = 0.28 P = 0.601; FnSBabb = 0.03
P = 0.87). Species-specific copepod abundance data
were heteroscedastic, and so remained after transfor-
mation (significant two-tailed Bartlett tests), and could
not be tested by analysis of variance.
Biological variables were not significantly corre-
lated to any combination of environmental variables in
multivariate analyses (BEST: SB ? nSB q = 0.22
P = 0.681; SB q = 0.33 P = 0.102; nSB q = 0.26
P = 0.703). Yet, abundance of stygobites (PERMA-
NOVA: pseudo-FSBabb = 3.61 P = 0.0187), pre-
sence/absence of stygobiotic taxa (PERMANOVA:
pseudo-FSBrichness = 6.35 P = 0.0002), and presence/
absence of all taxa (PERMANOVA: pseudo-
Ftot richness = 4.51 P = 0.0002) were significantly differ-
ent in H and L bores. However, there was no SEASON-
based difference (PERMANOVA: pseudo-FSBabb =
1.16 P = 0.3822; pseudo-FSBrichness = 0.54
P = 0.7691; pseudo-Ftot richness = 0.60 P = 0.8093),
nor was there a significant interaction between the two
factors (PERMANOVA: pseudo-F = 0.45SBabb P =
0.7246; pseudo-FSBrichness = 0.39 P = 0.891;
pseudo-Ftot richness = 0.97 p = 0.4735). The SB spe-
cies, Diacyclops italianus and Nitocrella psammo-
phila, provided a combined 63% of the discrimination
(33% and 29% respectively; Table 2) between H and
L bores. The remaining SB species contributed for
another 32%. This pattern was evident also in the
nMDS ordination (Fig. 5).
Discussion
Abiotic variables
The study area was homogeneous in altitude, soil use,
and geological structure. Bore water levels did not
vary seasonally, contrary to what might be expected in
agriculture-intensive areas. The local water manage-
ment authority (Provincia Autonoma di Trento)
strongly supports the modernization of the irrigation
Fig. 2 Bar-plot
representation of mean
(±standard errors) for: adissolved oxygen
concentration (mg l-1); boxygen saturation (%); cnitrate concentration
(mg l-1); d soluble reactive
phosphorus concentration
(lg l-1) in high-ammonium
(H bores: NHþ4C0.032 mg l-1; n = 32) or
low-ammonium
groundwater (L bores: NHþ4\0.032 mg l-1; n = 34) in
the alluvial aquifer of River
Adige
144 Hydrobiologia (2015) 743:139–150
123
infrastructure, encouraging farmers to use highly efficient
irrigation systems to save groundwater. The absence of an
impact of irrigation on the investigated groundwater body
is likely due to this policy, keeping groundwater
withdrawals into the limits of the aquifer recharge
(Autorita di Bacino dell’Adige, 2008). Pesticides were
detected in only four bores and with concentrations near
or below the threshold of no concern for groundwater
pollution. This pattern indicates that the use of pesticides
is limited in the study area or, more probably, performed
following best agricultural management practices
(BMPs). None of the others investigated groundwater
quality parameters exhibited concentrations at or above
legal pollution levels, with the exception of NHþ4(Table 1). There were significant differences in
groundwater quality in H versus L bores. In particular,
groundwater samples from H bores (with NHþ4concentrations C0.032 mg l-1) were characterized
by lower DO and nitrate concentrations and by higher
SRP concentrations than groundwater samples from L
bores (with NHþ4 concentrations\0.032 mg l-1). The
inverse relationship between NHþ4 on one side and
NO�3 ? DO on the other side is likely a result of
abiotic and biotic nitrogen transformation. NHþ4 tends
to be oxidized to NO�3 in a two-step nitrification
process (NHþ4 ! NO�2 ! NO�3 ) by aerobic chemo-
autotrophic bacteria, primarily Nitrosomonas and
Nitrobacter (Sharma & Ahlert, 1977; McGarvey
et al., 2007). Patches of nitrification are thus well
oxygenated and characterized by NHþ4 concentration
lower than NO�3 ones, as observed in the alluvial
aquifer of River Adige (Fig. 2, Table 1). In a few
alluvial aquifers, high natural NHþ4 concentration
occurs due to proximal buried organic material in
alluvial deposits (Jiao et al., 2010, and references
therein), but this does not appear to be the case for our
study area (Distretto Idrografico delle Alpi Orientali,
2010). Considering that 85% of the floodplain of the
River Adige is densely cultivated and that 90% of
fertilization is organic, the observed groundwater
quality pattern may be reasonably ascribed to organic
NHþ4 -containing fertilizers. Application of organic
fertilizers has been long recognized as a non-point
source of both N and P to freshwater (Carpenter et al.,
1998). Considering that the municipal waste water in
the study area is managed in state-of-the-art facilities
(Distretto Idrografico delle Alpi Orientali, 2010), the
higher concentrations of SRP, coupled with those of
NHþ4 , are likely attributable to the use of fertilizers in
the study area, as observed in other studies (Raave
et al., 2014). However, a definitive, unequivocal
association of groundwater NHþ4 and agricultural
practices may come only from an analysis of nitrogen
isotopes (Di Lorenzo et al., 2012).
Groundwater fauna
The use of groundwater for agricultural purposes is
known to threaten groundwater faunal and microbial
assemblages (Rouch et al., 1993; Dumas, 2004; Di
Lorenzo & Galassi, 2013; Griebler et al., 2014).
However, in our study, the variation in SB and/or nSB
copepod assemblages was not associated with sea-
sonal irrigation practices. Similar results were
observed by Korbel et al. (2013). In contrast, Di
Lorenzo & Galassi (2013) assessed an evident impact
of groundwater withdrawal for irrigation uses on
copepod abundances in the Mediterranean alluvial
aquifer of River Vibrata, in central Italy. However, the
aquifer of River Vibrata was fed by rainfalls only,
lacking a contribution from surrounding rocky
groundwater bodies, and was thus prone to a severe
Fig. 3 Principal components analysis (PCA) of groundwater
quality parameters according to NHþ4 factor in the alluvial
aquifer of River Adige based on samples collected in June and
October 2006. Groundwater bores with NHþ4 concentration
C0.032 mg l-1 are shown with black circles and those with
NHþ4 concentration \0.032 mg l-1 are shown with gray
triangles. Cumulative variance: 47 %
Hydrobiologia (2015) 743:139–150 145
123
water shortage, especially in summer (Di Lorenzo &
Galassi, 2013).
The dominance of crustaceans, primarily copepods
(91% of total density), in the alluvial aquifer of the
River Adige, supports earlier findings for similar
ecosystems (e.g., Sket,1999; Stoch & Galassi, 2010;
Di Lorenzo & Galassi, 2013; Di Lorenzo et al. 2013;
Fiasca et al., 2014). Biotic and environmental patterns
were completely independent, with the exception of
NHþ4 (BEST analysis). Our results upheld the hypoth-
esis that N-fertilizers, mainly organic, affect the
groundwater ecosystem in the alluvial aquifer of River
Adige, inducing a potential flow-on toxic effect to
stygobiotic copepods. We detected a clear difference
in copepod assemblage structure between H and L
bores, in both univariate and multivariate analyses.
Although the occurrence of nSB species in ground-
water may be considered related to the stochastic
infiltration of surface water at the bottom of the bores,
as observed in other studies (among others: Dole-
Olivier et al., 2000; Hahn & Matzke, 2005; Schmidt
et al., 2007; Hahn & Fuchs, 2008; Galassi et al., 2009;
Stein et al., 2010; Di Lorenzo & Galassi, 2013; Di
Lorenzo et al., 2013), bore water NHþ4 concentrations
higher than 0.032 mg l-1 seemed to affect the cope-
pod assemblage as a whole, and the stygobiotic
assemblages in particular, in the study area. The
stygobiotic species, Diacyclops italianus and Nitoc-
rella psammophila, seemed to be the most affected.
Fig. 4 Bar-plot
representation of mean
(±standard errors) for:
a species richness of
stygobiotic copepods;
b abundances of stygobiotic
copepods; c total copepod
species richness; d total
copepod abundance in high-
ammonium (H bores: NHþ4C0.032 mg l-1; n = 32) or
low-ammonium
groundwater (L bores: NHþ4\0.032 mg l-1; n = 34) in
the alluvial aquifer of River
Adige
Table 2 SIMPER results between groups of samples dis-
criminated according to NHþ4 factor
Species Contrib% Cum%
Diacyclops italianus 33.09 33.09
Nitocrella psammophila 29.52 62.61
Diacyclops paolae 11.62 74.22
Graeteriella unisetigera 9.30 83.52
Speocyclops infernus 6.17 89.69
Moraria stankovitchi 4.97 94.66
Contrib% percentage of dissimilarity provided by each species,
Cum% cumulative percentage of dissimilarity
146 Hydrobiologia (2015) 743:139–150
123

The toxic effect of NHþ4 and NH3 on crustaceans is
primarily, but not exclusively, related to the alteration
of respiratory metabolism (Aarset & Aunaas, 1990).
NHþ4 binds to crustacean hemocyanins (Romano &
Zeng, 2013). Under exposure to NHþ4 , the hemolymph
pH of aquatic crustaceans changes, altering the
detoxification mechanisms (Colt & Armstrong,
1981; Chen et al., 1994; Cheng et al., 2013), and thus
increasing organism vulnerability to other toxicants.
Ammonium compounds also alter osmoregulation,
affecting ATP-K?/Na? activities in the cell membrane
(Colt & Armstrong, 1981; Chen et al., 1994; Cheng
et al., 2013). Moreover, Di Marzio et al. (2013) argued
that NHþ4 acts as an endocrine disruptor, affecting
growth in epigean and hyporheic copepods. The
negative impact on survival or growth by NHþ4 occurs
via the disruption of various physiological mecha-
nisms, including osmoregulation, immunology, acid/
base balance, gas exchanges, as well as increased
oxidative stress, and pathogenic susceptibility, and
may induce histologic damage (Romano & Zeng,
2013).
There is not a unique European threshold value
(TV) for NHþ4 in groundwater. NHþ4 is one of the
pollutants indicated in the Annex II of the Directive
2006/118/EC for which TVs are to be provided by
each Member State. Twenty-one Member States have
so far provided highly different TVs for NHþ4 , ranging
from a minimum value of 0.084 mg l-1 to the
maximum allowed TV of 5 mg l-1 (European Com-
mission, 2010). The TV for NHþ4 in Italy is 0.5 mg l-1
(Repubblica Italiana, 2009). According to the results
of our study, such TVs do not seem to adequately
protect groundwater copepod assemblages in the
alluvial aquifer of River Adige.
Conclusions
Although agricultural N loading in excess of crop
needs is a long-recognized issue for Europe’s receiv-
ing water bodies, current EU threshold values for NHþ4do not address the potential adverse impact on
groundwater biota. Our results suggest that the com-
position, richness, and abundance of groundwater
copepods can be reliable indicators of groundwater
quality and ecosystem health related to NHþ4 contam-
ination. In particular, our study showed that levels of
NHþ4 lower than the present threshold values adopted
by EU Member States may not be protective of
stygobiotic copepod assemblages in some alluvial
aquifers underlying densely cultivated areas. Despite
copepod commonness and dominance in groundwater
habitats, to our knowledge, no information currently
Fig. 5 nMDS (non-metric
Multidimensional Scaling)
ordination of stygobiotic
copepod assemblages
according to NHþ4 factor in
the alluvial aquifer of River
Adige, based on June and
October abundances per
sample. Groundwater bores
with NHþ4 concentration
C0.032 mg l-1 are shown
with black circles and those
with NHþ4 concentration
\0.032 mg l-1 are shown
with gray triangles. Vectors
defining correlations
between each species and
the ordination are also
shown. Species
abbreviations as in
Appendix 1—
Supplementary Material
Hydrobiologia (2015) 743:139–150 147
123

exists on NHþ4 effects on groundwater copepod
reproduction, egg viability, hatching rates, or lipid
metabolism. We believe that this is an area worth
investigating, in order to estimate the true extent of the
NHþ4 -induced biological damage. The time needed for
the recovery of copepod assemblage in the contami-
nated sectors of the alluvial aquifer of River Adige,
after a hypothetical reestablishment of good ground-
water quality, also is difficult to estimate. The
recovery depends on several factors, such as ground-
water flow and population sizes in the unaffected areas
from which copepods can re-colonize. However, we
think that such a process may take some time, due to
low reproductive rates, long life spans (up to
3.5 years), and long post-embryonic development
time before sexual maturity, of stygobiotic copepods.
Moreover, we focused on the effect only of NHþ4because copepods assemblage structure proved not to
be related to other chemical patterns. However, while
such a situation applies to our study aquifer, it may not
apply to other scenarios, when potential interactions
with NHþ4 and other pollutants, such as pesticides,
should be explored.
Acknowledgements The project was funded by a grant from
the Basin Authority of the River Adige (Trento, Italy). We thank
Fabio Lazzeri for the design and concept of Fig. 1, and Andrea
Marchetti and Enrico Gattone for help in field sampling.
Constructive criticism by Dr. Grant C. Hose (Macquarie
University, NSW, Australia) and an anonymous reviewer also
improved the paper.
References
Aarset, V. & T. Aunaas, 1990. Effects of osmotic stress on
oxygen consumption and ammonia excretion of the Arctic
sympagic amphipod Gammarus wilkitzkii. Marine Ecology
Progress Series 58: 217–224.
Anderson, M. J., R. N. Gorley & K. R. Clarke, 2008. PER-
MANOVA? for PRIMER: Guide to Software and Statis-
tical Methods. PRIMER–E Ltd, Plymouth.
Autorita di Bacino dell’Adige, 2008. Quaderno sul bilancio
idrico superficiale di primo livello—Bacino idrografico del
fiume Adige. A cura della Segreteria Tecnico Operativa.
Trento, Maggio 2008. http://www.bacino-adige.it/sito/
index.php?option=com_content&view=article&id=104&
Itemid=324. Accessed 22 July 2014.
Blann, K. L., J. L. Anderson, G. R. Sands & B. Vondracek, 2009.
Effects of agricultural drainage on aquatic ecosystems: a
review. Critical Reviews in Environmental Science and
Technology 39: 909–1001.
Boulton, A. J., G. Fenwick, P. J. Hancock & M. S. Harvey, 2008.
Biodiversity, functional roles and ecosystem services of
groundwater invertebrates. Invertebrate Systematics 22:
103–116.
Camargo, J. A. & A. Alonso, 2006. Ecological and toxicological
effects of inorganic nitrogen pollution in aquatic ecosys-
tems: a global assessment. Environment International 32:
831–849.
Camargo, J. A., A. Alonso & M. de la Puente, 2005. Eutrophi-
cation downstream from small reservoirs in mountain riv-
ers of central Spain. Water Research 39: 3376–3384.
Candela, L., K. J. Wallis & R. M. Mateos, 2008. Non-point
pollution of groundwater from agricultural activities in
Mediterranean Spain: the Balearic Islands case study.
Environmental Geology 54: 587–595.
Carpenter, S. R., N. F. Caraco, D. L. Corell, R. W. Howarth, A.
N. Sharpley & V. H. Smith, 1998. Nonpoint pollution of
surface waters with phosphorus and nitrogen. Ecological
Applications 8: 559–568.
Caschetto, M., M. Barbieri, D. M. P. Galassi, L. Mastrorillo, S.
Rusi, F. Stoch, A. Di Cioccio & M. Petitta, 2014. Human
alteration of groundwater-surface water interactions (Sag-
ittario River, Central Italy): implication for flow regime,
contaminant fate and invertebrate response. Environmental
Earth Sciences 71: 1791–1807.
Chen, J.-C., C.-T. Chen & S.-Y. Cheng, 1994. Nitrogen excre-
tion and changes of hemocyanin, protein and free amino
acid levels in the hemolymph of Penaeus monodon
exposed to different concentrations of ambient ammonia-N
at different salinity levels. Marine Ecology Progress Series
110: 85–94.
Cheng, S.-Y., L.-W. Shieh & J.-C. Chen, 2013. Changes in
hemolymph oxyhemocyanin, acid-base balance, and elec-
trolytes in Marsupenaeus japonicus under combined
ammonia and nitrite stress. Aquatic Toxicology 130–131:
132–138.
Clarke, K. R., 1993. Non-parametric multivariate analyses of
changes in community structure. Australian Journal of
Ecology 18: 117–143.
Clarke, K. R. & R. N. Gorley, 2006. PRIMER v6: User Manual/
Tutorial. PRIMER-E, Plymouth.
Clarke, K. R., J. P. Somerfield & N. G. Raymond, 2008.
Testing of null hypotheses in exploratory community
analyses: similarity profiles and biota-environment link-
age. Journal of Experimental Marine Biology and Ecol-
ogy 366: 56–69.
Colt, J. E. & D. A. Armstrong, 1981. Nitrogen toxicity to
crustaceans, fish and molluscs. In: Allen, L. J & E.
C. Kinney (eds), Proceedings of the Bio-Engineering
Symposium for Fish Culture. Fish Culture Section,
American Fisheries Society, Northeast Society of Conser-
vation Engineers. Bethesda, Maryland: 34–47.
Dahan, O., A. Babad, N. Lazarovitch, E. E. Russak & D. Ku-
rtzman, 2014. Nitrate leaching from intensive organic
farms to groundwater. Hydrology and Earth System Sci-
ences 18: 333–341.
Di Lorenzo, T. & D. M. P. Galassi, 2013. Agricultural impact in
Mediterranean alluvial aquifers: do groundwater commu-
nities respond? Fundamental and Applied Limnology 182:
271–282.
Di Lorenzo, T., M. Brilli, D. Del Tosto, D. M. P. Galassi & M.
Petitta, 2012. Nitrate source and fate at the catchment scale
of the Vibrata River and aquifer (central Italy): an analysis
148 Hydrobiologia (2015) 743:139–150
123

by integrating component approaches and nitrogen iso-
topes. Environmental Earth Sciences 67: 2383–2398.
Di Lorenzo, T., F. Stoch & D. M. P. Galassi, 2013. Incorporating
the hyporheic zone within the river discontinuum: longi-
tudinal patterns of subsurface copepod assemblages in an
Alpine stream. Limnologica 43: 288–296.
Di Lorenzo, T., W. D. Di Marzio, M. E. Saenz, M. Baratti, A.
A. Dedonno, A. Iannucci, S. Cannicci, G. Messana & D.
M. P. Galassi, 2014. Sensitivity of hypogean and epigean
freshwater copepods to agricultural pollutants. Environ-
mental Science and Pollution Research 21: 4643–4655.
Di Marzio, W. D., D. Castaldo, T. Di Lorenzo, A. Di Cioccio, M.
E. Saenz & D. M. P. Galassi, 2013. Developmental endpoints
of chronic exposure to suspected endocrine-disrupting
chemicals on benthic and hyporheic freshwater copepods.
Ecotoxicology and Environmental Safety 96: 86–92.
Distretto Idrografico delle Alpi Orientali, 2010. Piano di
Gestione delle Acque. http://www.alpiorientali.it/new/
index.php/direttiva-2000-60/piano-di-gestione-acque-2009-
2015/documenti-preliminari-h2o. Accessed 22 July 2014.
Dole-Olivier, M.-J., D. M. P. Galassi, P. Marmonier & M.
Creuze des Chatelliers, 2000. The biology and ecology of
lotic microcrustaceans. Freshwater Biology 44: 63–91.
Dumas, P., 2004. Irrigation as a disturbance for interstitial
crustacean communities in a French Pyrenean alluvial
aquifer. International Journal of Limnology 40: 139–147.
Erisman, J. W., A. Bleeker, J. Galloway & M. S. Sutton, 2007.
Reduced nitrogen in ecology and the environment. Envi-
ronmental Pollution 150: 140–149.
European Commission, 2006. Directive 2006/118/EC of the
European Parliament and of the Council of 12 December
2006 on the protection of groundwater against pollution
and deterioration. Official Journal of the European Union L
372/19.
European Commission, 2010. Commission Staff Working
Document accompanying the Report from the Commission
in accordance with Article 3.7 of the Groundwater Direc-
tive 2006/118/EC on the establishment of groundwater
threshold values Brussels, SEC(2010) 166 final.
European Union, 2012. Statistical and Economic Information.
Report 2012. Directorate-General for Agriculture and
Rural Development. http://ec.europa.eu/agriculture/statistics/
agricultural/2012/pdf/full-report_en.pdf.
Fiasca, B., F. Stoch, M.-J. Olivier, C. Maazouzi, M. Petitta, A.
Di Cioccio & D. M. P. Galassi, 2014. The dark side of
springs: What drives small-scale spatial patterns of sub-
surface meiofaunal assemblages? Journal of Limnology
73: 55–64.
Galassi, D. M. P., 2001. Groundwater copepods (Crustacea:
Copepoda): diversity patterns over ecological and evolu-
tionary scales. Hydrobiologia 453(454): 227–253.
Galassi, D. M. P., R. Huys & J. W. Reid, 2009. Diversity,
ecology and evolution of groundwater copepods. Fresh-
water Biology 54: 691–708.
Gibert, J., D. L. Danielopol & J. A. Stanford (eds), 1994.
Groundwater Ecology. Academic Press, San Diego.
Griebler, C., F. Malard & T. Lefebure, 2014. Current develop-
ments in groundwater ecology—from biodiversity to eco-
system function and services. Current Opinion in
Biotechnology 27: 159–167.
Hahn, H. J. & D. Matzke, 2005. A comparison of stygofauna
communities inside and outside groundwater bores. Lim-
nologica 35: 31–44.
Hahn, H. J. & A. Fuchs, 2008. Distribution patterns of
groundwater communities across aquifer types in south-
western Germany. Freshwater Biology 54: 848–860.
Hartland, A., G. D. Fenwick & S. J. Bury, 2011. Tracing sew-
age-derived organic matter into a shallow groundwater
food web using stable isotope and fluorescence signatures.
Marine and Freshwater Research 62: 119–129.
Hurlbert, S. H., 1984. Pseudoreplication and the design of
ecological field experiments. Ecological Monographs 54:
187–192.
ISTAT, 2006. Agricoltura. Fertilizzanti distribuiti, per tipo di
produzione, stato, utilizzo consentito in agricoltura in
quintali, allo stadio solido. Dettaglio per Provincia. Anno
2006. http://www3.istat.it/dati/catalogo/20061109_00/PDF/
cap13.pdf. Accessed 22 July 2014.
Jensen, F. B., 1995. Uptake and effects of nitrite and nitrate in
animals. In Walsh, P. J. & P. Wright (eds), Nitrogen
Metabolism and Excretion. CRC Press, Boca Raton:
289–303.
Jensen, F. B., 2003. Nitrite disrupts multiple physiological
functions in aquatic animals. Comparative Biochemistry
and Physiology, Part A 135: 9–24.
Jiao, J. J., Y. Wang, J. A. Cherry, X. Wang, B. Zhi, H. Du & D.
Wen, 2010. Abnormally high ammonium of natural origin
in a coastal aquifer-aquitard system in the Pearl River
Delta, China. Environmental Science and Technology 44:
7470–7475.
Korbel, K. L., P. J. Hancock, P. Serov, R. P. Lim & G. C. Hose,
2013. Groundwater ecosystems vary with land use across a
mixed agricultural landscape. Journal of Environmental
Quality 42: 380–390.
Kota, S., R. C. Borden & M. A. Barlaz, 1999. Influence of
protozoan grazing on contaminant biodegradation. Feder-
ation of European Microbiological Societies Microbiology
Ecology 29: 179–189.
Marshall, M. C. & R. O. Hall, 2004. Hyporheic invertebrates
affect nitrogen cycling and respiration in stream sediment
microcosms. Journal of the North American Benthological
Society 23: 416–428.
McGarvey, J. A., W. G. Miller, R. Zhang, Y. Ma & F. Mit-
loehner, 2007. Bacterial population dynamics in diary
waste during aerobic and anaerobic treatment and sub-
sequent storage. Applied and Environmental Microbiology
73: 193–202.
Melo, A., E. Pinto, A. Aguiar, C. Mansilha, O. Pinho & I.
Ferreira, 2012. Impact of intensive horticulture practices
on groundwater content of nitrates, sodium, potassium, and
pesticides. Environmental Monitoring and Assessment
184: 4539–4551.
Morari, F., E. Lugato, R. Polese, A. Berti & L. Giardini, 2012.
Nitrate concentrations in groundwater under contrasting
agricultural management practices in the low plains of
Italy. Agriculture, Ecosystems & Environment 147: 47–56.
Mosier, A. R., J. K. Syers & J. R. Freney, 2004. Agriculture and
the nitrogen cycle: assessing the impacts of fertilizer use on
food production and the environment. Island Press,
Washington, DC.
Hydrobiologia (2015) 743:139–150 149
123

Poch-Massegu, R., J. Jimenez-Martinez, K. J. Wallis, F. Ra-
mirez de Cartagena & L. Candela, 2014. Irrigation return
flow and nitrate leaching under different crops and irriga-
tion methods in Western Mediterranean weather condi-
tions. Agricultural Water Management 134: 1–13.
R Development Core Team, 2008. A language and environment
for statistical computing. R Foundation for Statistical
Computing, Vienna, Austria. http://www.R-project.org.
Accessed 22 July 2014.
Raave, H., I. Keres, K. Kauer, M. Noges, J. Rebane, M. Tampere
& E. Loit, 2014. The impact of activated carbon on NO3--
N, NH4?-N, P, K leaching in relation to fertilizer use.
European Journal of Soil Science 65: 120–127.
Repubblica Italiana, 2009. Decreto Legislativo 16 Marzo 2009,
n. 30: Attuazione della direttiva 2006/118/CE, relativa alla
protezione delle acque sotterranee dall’inquinamento e dal
deterioramento. Gazzetta Ufficiale 79.
Romano, N. & C. Zeng, 2013. Toxic effects of ammonia, nitrite,
and nitrate to decapod crustaceans: a review on factors
influencing their toxicity, physiological consequences, and
coping mechanisms. Reviews in Fisheries Science 21:
1–21.
Rouch, R., A. Pitzalis & A. Descouens, 1993. Effets d’un
pompage a gros debit sur le peuplement des crustaces d’un
aquifere karstique. Annales de Limnologie 29: 15–29.
Schmidt, S. I., H. J. Hahn, T. J. Hatton & W. F. Humphreys,
2007. Do faunal assemblages reflect the exchange intensity
in groundwater zones? Hydrobiologia 583: 1–19.
Sharma, B. & R. C. Ahlert, 1977. Nitrification and nitrogen
removal. Water Research 11: 897–925.
Sket, B., 1999. The nature of biodiversity in hypogean waters
and how it is endangered. Biodiversity and Conservation 8:
1319–1338.
Stein, H., C. Kellermann, S. I. Schmidt, H. Brielmann, C. Steube,
S. E. Berkhoff, A. Fuchs, H. J. Hahn, B. Thulin & C.
Griebler, 2010. The potential use of fauna and bacteria as
ecological indicators for the assessment of groundwater
quality. Journal of Environmental Monitoring 12: 242–254.
Stoch, F. & D. M. P. Galassi, 2010. Stygobiotic crustacean
species richness: a question of numbers, a matter of scale.
Hydrobiologia 653: 217–234.
Tilman, D., 1999. Global environmental impacts of agricultural
expansion: the need for sustainable and efficient practices.
PNAS 96: 5995–6000.
van Grinsven, H. J. M., H. F. M. ten Berge, T. Dalgaard, B.
Fraters, P. Durand, A. Hart, G. Hofman, B. H. Jacobsen, S.
T. J. Lalor, J. P. Lesschen, B. Osterburg, K. G. Richards,
A.-K. Techen, F. Vertes, J. Webb & W. J. Willems, 2012.
Management, regulation and environmental impacts of
nitrogen fertilization in northwestern Europe under the
Nitrates Directive; a benchmark study. Biogeoscience 9:
5143–5160.
Weihrauch, D., J. C. McNamara, D. W. Towle & H. Onken,
2004. Ion-motive ATPases and active transbranchial NaCl
uptake in the red freshwater crab Dilocarcinus pagei
(Decapoda Trichodactylidae). Journal of Experimental
Zoology 207: 4623–4631.
Wood, P. J., J. Gunn & S. D. Rundle, 2008. Response of benthic
cave invertebrates to organic pollution events. Aquatic
Conservation 18: 909–922.
150 Hydrobiologia (2015) 743:139–150
123
Top Related
Copyright © 2022 FDOKUMEN

