
![[Human and wild mammal parasitosis in French Guiana]](https://static.fdokumen.com/doc/165x107/633660bb02a8c1a4ec022a28/human-and-wild-mammal-parasitosis-in-french-guiana.jpg)






Bahasa
Halaman
Hukum


This article was downloaded by: [Red de Bibliotecas del CSIC]On: 05 September 2014, At: 11:04Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Historical Biology: An International Journal ofPaleobiologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/ghbi20
A Late Pleistocene (MIS3) ungulate mammalassemblage (Los Rincones, Zaragoza, Spain) in theEurosiberian–Mediterranean boundaryVíctor Sauquéa, Ricardo García-Gonzálezb & Gloria Cuenca-Bescósa
a Grupo Aragosaurus-IUCA, Paleontología, Facultad de Ciencias, Universidad de Zaragoza,C/Pedro Cerbuna 12, E-50009 Zaragoza, Spainb Instituto Pirenaico de Ecología (IPE-CSIC), Avda. Nuestra Señora de la Victoria s/n, 22700Jaca, SpainPublished online: 03 Sep 2014.
To cite this article: Víctor Sauqué, Ricardo García-González & Gloria Cuenca-Bescós (2014): A Late Pleistocene (MIS3)ungulate mammal assemblage (Los Rincones, Zaragoza, Spain) in the Eurosiberian–Mediterranean boundary, HistoricalBiology: An International Journal of Paleobiology, DOI: 10.1080/08912963.2014.945926
To link to this article: http://dx.doi.org/10.1080/08912963.2014.945926
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

A Late Pleistocene (MIS3) ungulate mammal assemblage (Los Rincones, Zaragoza, Spain) in theEurosiberian–Mediterranean boundary
Vıctor Sauquea*, Ricardo Garcıa-Gonzalezb1 and Gloria Cuenca-Bescosa2
aGrupo Aragosaurus-IUCA, Paleontologıa, Facultad de Ciencias, Universidad de Zaragoza, C/Pedro Cerbuna 12, E-50009 Zaragoza,Spain; bInstituto Pirenaico de Ecologıa (IPE-CSIC), Avda. Nuestra Senora de la Victoria s/n, 22700 Jaca, Spain
(Received 29 March 2014; accepted 15 July 2014)
The Late Pleistocene archaeo-palaeontological sites in the Iberian Peninsula are located mainly on the coasts. Here, wepresent for the first time a palaeoenvironmental proxy for Upper Pleistocene locality (Marine Isotope Stage 3 MIS3) that isin the interior peninsular, in the Moncayo massif (Zaragoza). This is actually the boundary between Mediterranean andEurosiberian climatic regions. This study is based in the site ungulates: Capra pyrenaica, which is larger in size than thecurrent and fossil Capra from the Mediterranean area of the Iberian Peninsula. The horses have a small size, which is similarto that of the horses from Fontainhas and Casares. Significantly lesser in the number of specimens are the roe deer, thesouthern chamois and the auroch. The study and comparison of the faunal assemblage of the locality of Los Rincones withother sites of the Iberian MIS3 show a cluster of sites in the southern Peninsular, though separated, due to the abundance ofCapra pyrenaica. The association of Los Rincones is similar to the southern peninsular sites such as Nerja, Gorham Cave,Cova Beneito and Zafarraya. The ungulate assemblage of Los Rincones represents a landscape with temperate climate,presence of steppe and patches of forest, similar to the current landscape surrounding the cavity today.
Keywords: ungulates; Iberian Peninsula; Late Pleistocene; Capra pyrenaica; Equus ferus
1. Introduction
The Iberian Peninsula is an exceptional area for studying
the faunal associations of the Late Pleistocene, because on
account of its geographical configuration it displays two
distinct climatic ranges: the Atlantic and the Mediterra-
nean (Schmidt et al. 2012). Furthermore, the Iberian
Peninsula contains the southwestern limit to the cold
faunas that make up what is also known as the
‘Mammuthus–Coelodonta faunal complex’ (Kahlke
1999, Kahlke 2014). These faunas generally occur on a
sporadic basis, being most frequent during Marine Isotope
Stage 3 (MIS3) (Alvarez-Lao and Garcıa 2010, 2011).
Although there are many Late Pleistocene (MIS3) sites in
the Iberian Peninsula, most are concentrated in coastal
areas or in the Pyrenean region, and there are few
references to inland sites with faunas from the period in
question (d’Errico and Sanchez Goni 2003; Stewart 2007;
Schmidt et al. 2012).
The objective of this article is to undertake a
taxonomic and biometric study of the ungulates found in
the cave of Los Rincones (Zaragoza, Spain) and to carry
out a faunal analysis of the association present at the site,
comparing it with the bibliographical data. Through a
comparative study of the fauna, the aim is to define the
landscape of the inland area of the Iberian Peninsula
during the Late Pleistocene (MIS3), a landscape that was
inhabited by large predators such as the leopard and Homo
neanderthalensis.
2. Description of the study area
The cave of Los Rincones is located in the Sierra del
Moncayo, in the central part of the Iberian Range in the
north of the Iberian Peninsula (Figure 1(a)). On account of
the isolated relief it presents between the boundary of the
Ebro Valley and the Castilian Meseta as well as its high
altitude (2315m), its special orientation and its geological
diversity, the Moncayo massif is endowed with special
characteristics that are unique within its geographical
setting. It represents a confluence of three biogeographical
subregions (Uribe-Echebarrıa 2002). The northern slope of
the Moncayo is subject to the wet, cold influence of fronts
from the Atlantic, harbouring plant taxa of a northern
character, such as Quercus robur (Gomez Garcıa et al.
2003). By contrast, the surrounding land shows a marked
Mediterranean character, with vegetation dominated by
evergreen Quercus such as Quercus ilex and Quercus
coccifera. A few kilometres to the east lies the Ebro Valley,
with its notable aridity, where species characteristic of
semi-arid steppes such as Lygeum spartum can be found
(Hernandez and Valle 1989; Uribe-Echebarrıa 2002).
The climatological data from Agramonte de Moncayo
(Zaragoza), situated at an altitude of 1090m, show a mean
annual precipitation of 927mm and a mean annual
temperature of 9.48C (Hernandez and Valle 1989). The
pronounced altitude gradient and the NW–SE climatic
dissymmetry result in a great diversity of plant
environments, with three vegetation levels at present:
q 2014 Taylor & Francis
*Corresponding author. Email: [email protected]
Historical Biology, 2014
http://dx.doi.org/10.1080/08912963.2014.945926
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

meso-, supra- and oro-Mediterranean, with the existence
of a fourth level under discussion, the cryoro-Mediterra-
nean, which would be the equivalent of an Alpine level.
The only comparable reference as regards the predominant
vegetation during the period to which the fossil remains
from Los Rincones correspond (MIS3) would be Gabasa
Cave. A pollen study carried out on hyena coprolites
dating to between 50 and 40 ka reflects a mosaic glacial
landscape that includes Pinus and Juniperus woodlands
and steppes with Chenopodiaceae, Poaceae, Artemisia
and Asteraceae, suggesting an arid and cold/cool climate.
The presence of mesophilous and thermophilous tree and
shrub taxa (Quercus ilex, Quercus coccifera) also suggests
the existence of temperate Mediterranean refugia during
the period in question (Gonzalez-Samperiz et al. 2003,
2005).
The relief around the cave of Los Rincones is abrupt,
with the site surrounded by steep cliffs (Figure 1(b)),
which would foster the presence of rupicolous species
such as Rupicapra pyrenaica and Capra pyrenaica.
However, the ravine of Los Rincones opens into a broad
valley that could have hosted open-environment species
such as Equus and Bos primigenius and woodland species
such as Capreolus capreolus and Cervus elaphus.
Figure 1. (a) Geographical location of Los Rincones cave. (b) Panoramic view of the Los Rincones ravine. (c) Panoramic view of themain entrance of the cave. (d) Plan view of the cave. (e) Elevation view of the cave.
2 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

2.1. The site of Los Rincones: origin of theaccumulation
The cave of Los Rincones is situated at the head of the
ravine of Los Rincones (with a SW orientation), in the
municipality of Purujosa (Zaragoza, Spain). It opens at an
altitude of 1010m (see diagram of geographical location)
(Figure 1(c)). The cave is divided into a number of
chambers. The remains under study in this article appeared
at the surface of the ‘Ursus gallery’ and the ‘Leopard
gallery’, which are connected and are considered a single
site (Figure 1(d),(e)). The sample presents a similar pattern
of fragmentation, with similar tooth and cut marks, and
breaking pattern due to collapse of celling blocks. Also the
faunal composition, the skeletal survival profiles and
the degree of preservation of the remains, as well as the
distribution of fragments of the same anatomical element
in distinct galleries of the site, indicate that the process of
accumulation was similar: bones, with other clastic
sediments, were carried in from the surface (allogenic
transport) to the Ursus gallery, until the cone blocked the
mouth of the cave (Sauque et al. 2014). The faunal
association under study in this article date at least to the
Late Pleistocene, probably MIS3. We sent samples to a
radiocarbon laboratory (Beta Analytics, Miami, FL, USA)
but unfortunately we could not obtain radiometric ages by
the 14C method due to lack of collagen in the bones. Also
during the early stages of cleaning the gallery, a piece of
Mousterian industry was also discovered in the cave of Los
Rincones.The upper part of the sedimentary cone that
closes off the original entrance to the cave has been dated
on the basis of a microfaunal association consisting of
Microtus, Iberomys and Pliomys lenki. Such an association
is characteristic of the Late Pleistocene. The species
Pliomys lenki disappears between 50 ka and 40 ka from the
centre of the Iberian Peninsula, where it is only found at
Mousterian localities (Cuenca-Bescos et al. 2010; Sauque
and Cuenca-Bescos 2013).
Late Pleistocene faunal accumulations are generally
produced by the activity of humans or carnivores (Cruz-
Uribe 1991; Stiner 1991; Diedrich 2011; Enloe 2012).
Noteworthy among the latter are the accumulations
generated by hyenas, which are much more frequent
than those generated by leopards (Yravedra 2006;
Diedrich 2013; Sauque et al. 2014). Both hyenas and
humans tend to break bones in order to extract the marrow,
so the presence of whole bones is very rare at such sites.
The site of Los Rincones is one of the few sites where the
accumulation is attributed to the activity of a leopard as the
main accumulator (Sauque and Cuenca-Bescos 2013;
Sauque et al. 2014). Nonetheless, the site also testifies to
the sporadic presence of Neanderthals, who left their mark
in the form of cut marks in some bones, as well as remains
of lithic industry (Sauque et al. 2014). As a result, the
accumulation is composed mainly of small-sized herbi-
vores that fall within the range of leopard consumption.
The accumulation consists primarily of remains of Capra
pyrenaica (Table 1; Figure 2), thus providing a great
opportunity to carry out a biometric study of this
taxonomically controversial taxon (Garrido 2008;
Garcıa-Gonzalez 2011).
3. Materials and methods
For taxonomic identification, the following authors have
been followed: Bibikova (1958), Pales and Lambert
(1971), Eisenmann (1986), Gee (1993), Lister (1996) and
Fernandez (2001). Use has also been made of the reference
collections of the UZ (Universidad de Zaragoza) and the
Table 1. Composition of the Los Rincones ungulateassemblage.
Taxa NISP %NISP MNI %MNI
Cervus elaphus 13 2.11 2 6.25Capreolus capreolus 26 4.23 2 6.25Rupicapra pyrenaica 29 4.72 3 9.38Capra pyrenaica 528 85.85 20 62.5Bos primigenius 4 0.65 1 3.13Equus ferus 12 1.95 3 9.38Equus hydruntinus 2 0.33 1 3.13Total 615 100 32 100
Figure 2. Pie chart of ungulates from Los Rincones.
Historical Biology 3
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014
IPE (Instituto Pirenaico de Ecologıa). In order to assess the
skeletal representation of the assemblage from Los
Rincones, we have used the number of remains, the
number of identified specimens (NISP) and the minimum
number of individuals (MNI), which have been calculated
in accordance with Brain (1981) and Lyman (1994a).
In order to determine the age of death, dental
replacement and the degree of dental eruption have been
used (Morris 1978; Hillson 1992), as well as the degree of
fusion of the epiphyses in long bones (Morris 1972).
Furthermore, for each species the criteria proposed by
other specialists have been followed: e.g. those from
Perez-Ripoll (1988) and Vigal and Marchordom (1985) for
Capra pyrenaica; those from Tome and Vigne (2003) for
Capreolus capreolus; those from Perez-Barberıa (1994)
for Rupicapra pyrenaica and those from Aitken (1975),
Azorit et al. (2002), D’Errico and Vanhaeren (2002) and
Mariezkurrena (1983) for Cervus elaphus. The measure-
ments were taken with a digital calliper (Digimatic Caliper
CD-800CX) in accordance with von den Driesch (1976) and
Bibikova (1958). For the remains from Equus, the
methodology proposed by Eisenmann (1986) has been
followed.
All measurements are given in millimetres. The
morphometric data have been compared with data
collections from western Europe published by a variety
of authors (see citations in Tables 2–7), as well as with our
own data gathered directly from the collections of the
Museo Nacional de Ciencias Naturales in Madrid
(MNCN), the Museu de Prehistoria de Valencia (MPV),
the Instituto Pirenaico Ecologıa (IPE), the Zoologische
Staatssammlung Munchen (ZSM), the Museo Paleontolo-
gico de Zaragoza (MPZ), the Museo Arqueologico
Provincial de Huesca (MAPH), the Instituto Alaves de la
Naturaleza (IAN), Institut fur Palaoanatomie, Domestika-
tionsforschung und Geschichte der Tiermedizin (Ludwig
Maximilians university Munchen) (LMU) and the
Naturhistorisches Museum Basel (NMB).
To provide a statistical comparison of the site of Los
Rincones with the faunal associations of other sites, we
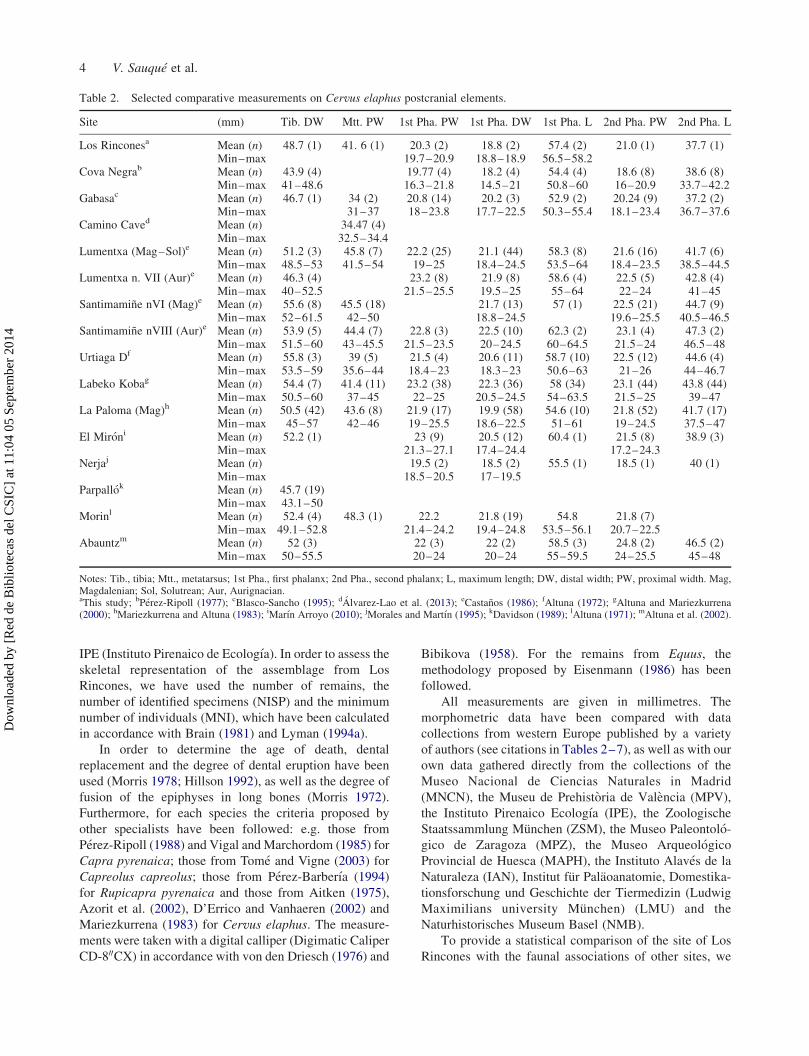
Table 2. Selected comparative measurements on Cervus elaphus postcranial elements.
Site (mm) Tib. DW Mtt. PW 1st Pha. PW 1st Pha. DW 1st Pha. L 2nd Pha. PW 2nd Pha. L
Los Rinconesa Mean (n) 48.7 (1) 41. 6 (1) 20.3 (2) 18.8 (2) 57.4 (2) 21.0 (1) 37.7 (1)Min–max 19.7–20.9 18.8–18.9 56.5–58.2
Cova Negrab Mean (n) 43.9 (4) 19.77 (4) 18.2 (4) 54.4 (4) 18.6 (8) 38.6 (8)Min–max 41–48.6 16.3–21.8 14.5–21 50.8–60 16–20.9 33.7–42.2
Gabasac Mean (n) 46.7 (1) 34 (2) 20.8 (14) 20.2 (3) 52.9 (2) 20.24 (9) 37.2 (2)Min–max 31–37 18–23.8 17.7–22.5 50.3–55.4 18.1–23.4 36.7–37.6
Camino Caved Mean (n) 34.47 (4)Min–max 32.5–34.4
Lumentxa (Mag–Sol)e Mean (n) 51.2 (3) 45.8 (7) 22.2 (25) 21.1 (44) 58.3 (8) 21.6 (16) 41.7 (6)Min–max 48.5–53 41.5–54 19–25 18.4–24.5 53.5–64 18.4–23.5 38.5–44.5
Lumentxa n. VII (Aur)e Mean (n) 46.3 (4) 23.2 (8) 21.9 (8) 58.6 (4) 22.5 (5) 42.8 (4)Min–max 40–52.5 21.5–25.5 19.5–25 55–64 22–24 41–45
Santimamine nVI (Mag)e Mean (n) 55.6 (8) 45.5 (18) 21.7 (13) 57 (1) 22.5 (21) 44.7 (9)Min–max 52–61.5 42–50 18.8–24.5 19.6–25.5 40.5–46.5
Santimamine nVIII (Aur)e Mean (n) 53.9 (5) 44.4 (7) 22.8 (3) 22.5 (10) 62.3 (2) 23.1 (4) 47.3 (2)Min–max 51.5–60 43–45.5 21.5–23.5 20–24.5 60–64.5 21.5–24 46.5–48
Urtiaga Df Mean (n) 55.8 (3) 39 (5) 21.5 (4) 20.6 (11) 58.7 (10) 22.5 (12) 44.6 (4)Min–max 53.5–59 35.6–44 18.4–23 18.3–23 50.6–63 21–26 44–46.7
Labeko Kobag Mean (n) 54.4 (7) 41.4 (11) 23.2 (38) 22.3 (36) 58 (34) 23.1 (44) 43.8 (44)Min–max 50.5–60 37–45 22–25 20.5–24.5 54–63.5 21.5–25 39–47
La Paloma (Mag)h Mean (n) 50.5 (42) 43.6 (8) 21.9 (17) 19.9 (58) 54.6 (10) 21.8 (52) 41.7 (17)Min–max 45–57 42–46 19–25.5 18.6–22.5 51–61 19–24.5 37.5–47
El Mironi Mean (n) 52.2 (1) 23 (9) 20.5 (12) 60.4 (1) 21.5 (8) 38.9 (3)Min–max 21.3–27.1 17.4–24.4 17.2–24.3
Nerjaj Mean (n) 19.5 (2) 18.5 (2) 55.5 (1) 18.5 (1) 40 (1)Min–max 18.5–20.5 17–19.5
Parpallok Mean (n) 45.7 (19)Min–max 43.1–50
Morinl Mean (n) 52.4 (4) 48.3 (1) 22.2 21.8 (19) 54.8 21.8 (7)Min–max 49.1–52.8 21.4–24.2 19.4–24.8 53.5–56.1 20.7–22.5
Abauntzm Mean (n) 52 (3) 22 (3) 22 (2) 58.5 (3) 24.8 (2) 46.5 (2)Min–max 50–55.5 20–24 20–24 55–59.5 24–25.5 45–48
Notes: Tib., tibia; Mtt., metatarsus; 1st Pha., first phalanx; 2nd Pha., second phalanx; L, maximum length; DW, distal width; PW, proximal width. Mag,Magdalenian; Sol, Solutrean; Aur, Aurignacian.aThis study; bPerez-Ripoll (1977); cBlasco-Sancho (1995); dAlvarez-Lao et al. (2013); eCastanos (1986); fAltuna (1972); gAltuna and Mariezkurrena(2000); hMariezkurrena and Altuna (1983); iMarın Arroyo (2010); jMorales and Martın (1995); kDavidson (1989); lAltuna (1971); mAltuna et al. (2002).
4 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

have undertaken correspondence analysis (CA) using
PAST v.2.14 (Hammer et al. 2001). CA is a way of
projecting a multivariate database onto two or three
dimensions in order to visualise the cluster patterns. It is
thus a good method for discovering geographical group-
ings and patterns and for bringing to light environmental
gradients (Greenacre 1984; Jongman et al. 1995; Legendre
and Legendre 1998).
The algorithms used by CA present two problems. The
first is the tendency to compress the end of the ordinate
axis, squeezing together the samples and taxa, which is not
useful. The second is the arch effect, which occurs when
the environmental gradient presents ‘losses’ in the first and
second ordination axes instead of being linked only with
the first axis as it should be. To avoid these problems, we
have used a detrended correspondence analysis (DCA) for
our comparison of faunal associations. First, this rescales
itself, removing the compression at the ends of the axes;
afterwards, it carries out the detrending, stretching and
bending the arch until it becomes straight. This type of
analysis is of great value for the ordination of ecological
databases (Hill and Gauch 1980).
4. Taxonomy
The ungulates from Los Rincones show a good state of
preservation due to the non-anthropic origin of the
accumulation (Sauque et al. 2014). Thanks to the low
levels of modification, it has been possible to identify
taxonomically a total of 615 ungulate remains (Table 1).
These remains correspond to the following taxa: Cervus
elaphus (red deer), Capreolus capreolus (roe deer), Capra
pyrenaica (Iberian wild goat), R. pyrenaica (Pyrenean
chamois), Equus hydruntinus (European ass), Equus ferus
(horse) and Bos primigenius (aurochs) (Table 1; Figure 2).
The main ungulate is Capra pyrenaica, constituting
86% of the total ungulate remains. The carnivores, which
are not included in this study, are Ursus arctos (brown
Table 3. Selected comparative measurements on Capreolus capreolus cranial and postcranial elements.
Site (mm) m3. L Hum. DW Ra.PW Mtcp. DW Tib. DW
Los Rinconesa Mean (n) 15.1 (1) 27.1 (1) 29.5 (1) 22.2 (1) 27.4 (1)Min–max
Gabasab Mean (n) 27.3 22.2 (1) 25.5 (3)Min–max 27–27.6 24.9–26
Jou Puertac Mean (n) 17.2 (2) 29.8 (5) 21.6 (1) 27.7 (4)Min–max 16.3–18.1 28.2–31.4 21–22.5 26.5–28.7
Urtiagad Mean (n) 16.5 (24) 31.1 (12) 24.4 (13) 30.7 (2)Min–max 14.5–17.6 28.2–33.4 21.3–27.9 29.9–31.5
Santimaminee Mean (n) 16.1 (27) 29.8 (13) 29.2 (3) 23.9 (8)Min–max 14.2–17.6 28–32.5 27–32.5 23–25
La Rieraf Mean (n) 16.9 (8) 29.4 (5) 24 (2)Min–max 15.5–18 22.5–32 23.5–24.5
Lumentxag Mean (n) 27 (1) 25 (2)Min–max 24–26
Notes: m3 L., lower third molar; Hum. humerus; Ra., radius; Mtcp., metacarpus; Tib., tibia; L, maximum length; DW, distal width; PW, proximal width.aThis study; bBlasco-Sancho (1995); cAlvarez-Lao (2014); dAltuna (1972); eCastanos 1984; fAltuna (1986); gCastanos (1986).
Table 4. Values of the index of lateral articular facet and distal trochlea astragalus of Bos and Bison according to different authors.
Astragalus
Lateral articular facet index Distal trochlea index
Reference Genus n Mean Variability N Mean Variability
Bibikova (1958) Bos .58 93–98.7Bison 50–54 73.3–89.2
Stampfli (1963) Bos 11 54.3 46.4–62.2 16 90 82–122.7Bison 12 45.9 41.6–56.6 12 81.7 76–100
Altuna (1972) Bos 3 52 46.5–55.8 8 89.7 81.2–95.5Buitrago-Villaplana (1992) Bos 8 53 43.3–69.4 8 95 83–108.5Sala et al. (2010) Bos 1 61.81 1 93.3
Bison 1 40.95 2 88.43 82.95–93.91Perez-Ripoll (1977) Bos 1 61.1 97.1Los Rincones Bos 1 60 1 110
Historical Biology 5
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Table
5.
SelectedcomparativemeasurementsonCapra
pyrenaicacranialandpostcranialelem
ents.
Site
(mm)
LP2–P4
LM1–M3
m3.L
Hum.DW
Ra.PW
Ra.DW
Mtcp.PW
Mtcp.DW
Mtcp.L
Tib.DW
Ast.L
Ast.W
Mtt.L
Mtt.PW
Mtt.DW
LosRincones
aMean(n)
23.1
(1)
46.1
(1)
27.2
(11)
39.3
(10)
39.9
(6)
42.4
(4)
33.7
(12)
38.1
(8)
152.6
(6)
34.2
(8)
36(15)
23.9
(16)
150.4
(1)
26.5
(4)
32(4)
Min–max
25.2–29.3
34.8–45.9
38.5–41.5
40.2–45.3
28.5–37.8
31.6–40.7
145.6–158.3
29.8–37.9
34.2–40.6
21.9–28.2
24.1–30.2
29.6–35.6
JouPuertab
Mean(n)
24.6
(1)
53.6
(1)
25.5
(2)
34.2
(4)
37.6
(3)
38.3
(2)
Min–max
24.54–26.5
27–36.3
36.7–38.1
37.7–38.8
TitoBustillo
cMean(n)
22.85(2)
40.8
(5)
42.5
(1)
35.5
(1)
35.8
(2)
33.5
(1)
35.7
(7)
23.4
(7)
32.2
(3)
Min–max
21.7–24
34.5–45
34–37.5
32–40
21–27
31–33.5
LaRiera
dMean(n)
24.5
(2)
23.8
(23)
29(2)
35.7
(3)
31(6)
Min–max
23.5–25.5
19.5–28.5
28–30
30–39
29.5–34.5
UrtiagaDe
Mean(n)
23.7
(7)
55.9
(3)
27(23)
43.7
(13)
41.5
(1)
34.9(6)
145(1)
36.5
(1)
37(29)
24.2
(27)
Min–max
22.1–26
50.3–62
24–29.3
39.5–48.5
33.3–39
34.3–40.5
21.5–28
Chaves
(Mag)f
Mean(n)
24(1)
54(1)
23.6
(29)
36.7
(5)
42.3
(4)
31.25(2)
35(1)
34.3
(5)
32(9)
34.1
(5)
147.5
(1)
28.8
(3)
Min–max
21–29.5
31–41.5
36.5–45
27.5–35
28.5–41.5
27–36.5
31–35.5
27.5–30
Gabasag
Mean(n)
29(58)
43(11)
40(8)
37.4
(3)
31.3
(12)
35.2
(5)
142.9
(5)
35.3
(12)
37.6
(34)
24.5
(30)
148.1
(4)
28(10)
32.2
(5)
Min–max
25.8–32.2
38.2–48.6
37.8–42.8
37–37.9
24.7–37.4
27.4–41.4
129–156
31.8–39.1
32.2–41.7
21.5–27.5
143–151.1
25.7–29.9
29.7–37
Santimam
ineh
Mean(n)
25.5
(4)
48.6
(4)
26.8
(8)
40.8
(2)
30.5
(1)
35.3(5)
35.3
(3)
37.8
(12)
23.5
(11)
179(1)
25.5
(1)
31(1)
Min–max
22.5–31.5
44.5–54
21.5–32.5
37–44.5
30.5–41
33.5–38.5
30–40.5
19.4–27.5
Bolinkobai
Mean(n)
27(1)
26(37)
38.8
(2)
29.7
(3)
39.5
(3)
36.6
(31)
23.9
(28)
Min–max
22–28.5
38.5–39
29.5–30
33–43.5
33.5–40.5
21–27.5
Lumentxah
Mean(n)
21.5
(1)
25.5
(1)
35.5
(3)
43(1)
40(1)
40.5
(2)
156.5
(1)
29.3
(2)
32.1
(8)
21.1
(9)
32.3
(2)
Min–max
32.5–41
35–46
29–29.5
30.5–33.5
18.5–23
30.5–36
Errallaj
Mean(n)
25.6
(4)
48.6
(4)
25.9
(87)
39.6
(7)
39.4
(6)
41(1)
34.7
(3)
37.5
(4)
149.7
(3)
34.5
(8)
38.8
(6)
26.2
(5)
152.3
(2)
29.4
(4)
35.2
(3)
Min–max
23.5–29
44.5–54
23–29
37–42.5
32–48
33.4–35.5
33–41
141–154
29–39
37.5–40.5
24.5–28
150–154.5
27–31.5
32.5–39
LosCasares
kMean(n)
51.1
(2)
28.2(4)
42(1)
Min–max
50.2–52
25.5–31.4
CovaNegra
(MIS
5)l
Mean(n)
22.1
(1)
51.1
(1)
37.2
(1)
43.4
(3)
41(2)
31.6
(8)
34.3
(7)
32.7
(4)
37.3
(6)
23.8
(5)
Min–max
41.1–48
37.5–44
29–37
31.6–42.5
31.3–35
32.2–41
20.3–26.4
Parpallo
(UPa)
m,n
Mean(n)
26.8
(144)
34.9
(259)
36(37)
38.4
(71)
29.5
(49)
33.9
(113)
137.6
(29)
31.7
(63)
149.4
(29)
22.5
(23)
30.1
(19)
Min–max
23.5–30.4
28.9–39.6
36.5–41
31.9–45.9
25.4–32.1
26–37.1
123.8–143.4
27.9–35.2
133–160
21–27.5
27–34.5
Recolduc(Sol-Gra)n
Mean(n)
27.3
(9)
30.6
(9)
128.6
(6)
143.4
(5)
24(5)
28.3
(5)
Min–max
26.3–28.9
27.2
234.7
123.4–143.1
132.2–157.5
21.6–26.6
26.7–31.6
Malladetes
(UPa)
nMean(n)
28.4
(6)
32.3
(7)
131.1
(1)
137.1
(2)
23.9
(2)
29.5
(8)
Min–max
25.7–30.1
31.6–33.8
133.4–140.9
23.1–24.7
27.9–31
Nerja
(Mag)o,n
Min–max
27.3
(1)
34.5
(3)
36(1)
23(1)
28.6
(1)
Mean(n)
33.1–37.2
Capra
pyrenaicaextantn
Mean(n)
28.1
(131)
33.9
(131)
125.4
(131)
32.5
(136)
20.7
(135)
133.6
(135)
23.5
(135)
27.4
(135)
Min–max
23.2–33.3
25.9–38
104–144.2
27.1–36.4
18–24.8
105.77–155.48
18.7–26.1
21.1–33
Notes:P2–P4,upper
premolarseries;M1–M1,upper
molarseries;m3L.,lower
thirdmolar;Hum.,humerus;Ra.,radius;Mtcp.,metacarpus;Tib.,tibia;Ast.,astragalus;Mtt.,metatarsus;L,maxim
um
length;W,maxim
um
width;DW,distalwidth;PW,proxim
alwidth.Mag,Magdalenian;Sol,Solutrean;Gra,Gravettian;Aur,Aurignacian;UPa,Upper
Palaeolihitc.
aThisstudy;bAlvarez-Lao
(2014);
cAltuna(1976);
dAltuna(1986);
eAltuna(1972);
f Castanos(1993);
gBlasco-Sancho(1995);
hCastanos(1984);
i Castanos(1986);
j AltunaandMariezkurrena(1985);
kAltuna
(1973);
l Perez
Ripoll(1977);
mDavidson(1989);
nSauque(unpublished
data);oMoralesandMartın(1995).
6 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Table
6.
SelectedcomparativemeasurementsonRupicapra
pyrenaicacranialandpostcranialelem
ents.
Site
(mm)
m3.L
Hum.DW
Ra.PW
Tib.DW
Ast.L
Ast.W
1stPha.L
1stPha.PW
2ndPha.L
2ndPha.PW
LosRincones
aMean(n)
17.1
(3)
28(2)
28.8
(1)
27.1
(1)
30.4
(4)
19.9
(4)
41.8
(1)
13.3
(1)
27.5
(4)
13.7
(4)
Min–max
16.9–17.4
26.4–29.6
29.6–31.2
19.1–20.8
25.8–28.5
12.8–14.2
Abauntzb
Mean(n)
18.2
(6)
32.6
(4)
28.5
(2)
31.3
(5)
20.4
(5)
46.3
(5)
14.2
(5)
30.4
(6)
13.8
(6)
Min–max
17.1–8.7
31.5–33.5
28–29
30.5–32
19–22
41.5–50
13.8–14.8
29–33
12.5–15.5
Santimam
inenIII–
V(M
ag-Sol)c
Mean(n)
18.5
(8)
31.3
(2)
33.5
(2)
25(1)
30.4
(4)
19.5
(3)
42.3
(6)
12.8
(7)
26(2)
11.1
(2)
Min–max
17–19.7
30.5–32
32.5–34.5
28–31.5
18.2–21
37.5–47.5
11.6–14.5
22–30
9.4–12.7
Gabasad
Mean(n)
19.2
(9)
27.8
(3)
28.7
(1)
31.2
20.5
(3)
46.6
(7)
13.6
(9)
28(4)
12.7
(4)
Min–max
17.8–21.5
27.6–28.1
29–32.3
20–20.9
44.9–49.1
12.8–14.3
23.8–32.3
10.4–13.9
CovaNegra
eMean(n)
29.5
(1)
29.1
(2)
31.8
(1)
21.1
(1)
44.2
(1)
12.2
(1)
Min–max
29–29.2
Chaves
fMean(n)
19(4)
31(2)
29(1)
31(1)
20.15(2)
13.13(3)
28(1)
13.3
(1)
Min–max
18.4–19.6
30.5–31.5
30.5–31.5
19.8–20.5
12.3–14
Bolinkoban.III–
VI(M
ag-G
ra)g
Mean(n)
18.8
(2)
29.2
(3)
19.8
(1)
40.8
(5)
12.7
(7)
28.5
(5)
12.7
(5)
Min–max
18.3–19.3
28–30.5
31–47
11.9–13.2
26–31.5
12–13.5
LumentxanIV
.V.VI(M
ag-Sol)g
Mean(n)
17.4
(1)
29.7
(3)
26.8
(2)
22.5
(1)
10.7
(1)
Min–max
29–30.5
25.5–28
CuevaMillanh
Mean(n)
17.8
(7)
30.4
(1)
20.5
(1)
12.1
(1)
27.3
(3)
12.1(4)
Min–max
17–19.2
25.7–29
11.6–13
Ekaini
Mean(n)
17.6
(8)
31(1)
31.5
(3)
31.5
(5)
20.3
(3)
46.8
(10)
13.7
(10)
30.5
(3)
13.2
(3)
Min–max
17.3–18.3
30.5–33
30–34
19.7–21.2
42.5–49.5
13–14
30–31
13–13.5
JouPuertaj
Mean(n)
19.3
(1)
32.2
(2)
27.22(3)
Min–max
31–33.4
26.4–28.9
Naranco
jMean(n)
17.46(4)
30.42(7)
26.3
(10)
Min–max
16.3–18.9
29.5–33.4
25.36–27.92
Lezetxikik
Mean(n)
18(19)
33.3
(2)
27.9
(6)
Min–max
16.4–19
32.1–34.4
26.3–29.5
Urtiagak
Mean(n)
18.9
(16)
29.9
(8)
Min–max
15.3–20.6
27.6–32.8
Amaldal
Mean(n)
17.8
(3)
28.75(2)
Min–max
17.3–18.5
28.5–29
LaPalomam
Mean(n)
16.7
(2)
31.5
(3)
28.5
(1)
Min–max
14.7–18.7
31–32
Valdegoban
Mean(n)
30.7
(8)
31.8
(19)
26.3
(27)
Min–max
28.8–33
24.6–38.9
23.2–28.5
Notes:m3.L,lower
thirdmolar;Hum.humerus;Ra.,radius;Tib.,tibia;Ast.,astragalus;1stPha.,firstphalanx;2ndPha.,secondphalanx;L,maxim
um
length;W,maxim
um
width;DW,distalwidth;PW,
proxim
alwidth.Mag,Magdalenian;Sol,Solutrean;Gra,Gravettian.
aThisstudy;bAltunaetal.(2002);
cCastanos(1984);
dBlasco-Sancho(1995);
ePerez-Ripoll(1977);
f Castanos(1993);
gCastanos(1986);
hPerez
LegidoandCerdeno(1992);
i AltunaandMariezkurrena(1984);
j Alvarez-Lao
(2014);
kAltuna(1972);
l Altuna(1990);
mCastanos(1980);
nArceredillo
etal.(2011).
Historical Biology 7
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Table
7.
SelectedcomparativemeasurementsonEquuspostcranialelem
ents.
Site
Taxa
(mm)
1stPha.L
1stPha.PW
1stPha.DW
2ndPha.L
2ndPha.PW
2ndPha.DW
3rd
Pha.LM
3rd
Pha.W
LosRincones
aEquusferus
Mean(n)
78.8
(4)
49.3
(3)
44.1
(3)
45.8
(5)
49.9
(4)
44.9
(5)
58.8
(1)
70.7
(1)
Min–max
75.2–84.1
47.4–50.3
43.7–44.7
41.9–48.1
48.8–51.6
41.4–48.7
Cuetodela
Minab
Equusferus
Mean(n)
84.3
(2)
58(1)
47(2)
50.2
(5)
54.4
(4)
48.9
(4)
48.25(4)
83.5
(1)
Min–max
80–88.5
45–49
46–51
50.5–58
47–53.5
44–54
Santimam
inec
Equuscaballus
Mean(n)
87.8
(3)
55.1
(4)
46.3
(3)
50.6
(5)
55.3
(5)
50(4)
Min–max
83.9–93
54.5–59.4
44–48.6
50–51.2
53.6–58.6
48.6–51.9
Lumentxac
Equuscaballus
Mean(n)
88(2)
59.8
(2)
50.3
(2)
50.3
(2)
56.3
(2)
53(2)
68.5
(2)
84.5
(2)
Min–max
84.5–91.5
59–60.5
50–50.5
48.5–52
54.5–58
52–54
68–69
83.5–84.5
Atxuric
Equuscaballus
Mean(n)
82(1)
54.5
(1)
47(1)
Min–max
Bolinkobac
Equuscaballus
Mean(n)
42(1)
49.1
(4)
56.3
(4)
48.8
(4)
66(1)
78.5
(1)
Min–max
47.5–50
54–58.5
43.5–51
San
Lorenzo
IIc
Equuscaballus
Mean(n)
42.5
(1)
47(1)
42.5
(1)
Min–max
AbricRomanıd
Equuscaballus
Mean(n)
85.8
(1)
52.6
(1)
43.3
(1)
Min–max
LosCasares
eEquuscaballuscasarensis
Mean(n)
77.3
(2)
46(1)
37(1)
Min–max
76.5–78
E.caballusgallicuse
Equuscaballusgallicus
Mean(n)
84.7
(25)
60.1
(25)
50(25)
Min–max
79–89
57.6–66
46.5–53
E.cfgallicuse
Equuscf
gallicus
Mean(n)
81.6
(47)
57.1
(47)
47.4
(47)
Min–max
74.5–91
53.5–65.5
42–53.5
UrtiagaDf
Equuscaballus
Mean(n)
82(1)
54.5
(1)
45(1)
50.65(2)
57(2)
51.75(2)
50.5
(2)
77(2)
Min–max
49–52.3
55–59
49.5–54
50–51
76–78
Aitzbartef
Equuscaballus
Mean(n)
55(1)
75.7
(1)
Min–max
Gruta
deFontainhas
gEquuscaballusantunensis
Mean(n)
79.6
(4)
52.8
(3)
41.7
(3)
46.2
(1)
48(1)
43(1)
50.5
(2)
70.75(2)
Min–max
78.5–82
52–54
41–42
47–57
70–71.5
Algar
deJoao
Ram
osg
Equuscaballusantunensis
Mean(n)
82(2)
52.75(2)
42.5
(1)
49.5
(1)
54(1)
45(1)
53.5
(2)
70.5
(2)
Min–max
82
52.5–53
42.5
53–54
69–72
PdreiradeSalem
asg
Equuscaballusantunensis
Mean(n)
87(3)
50.85(2)
42(2)
46.5
(1)
50(1)
47(1)
Min–max
84.5–89
50–51.7
41–43
Parpallo
hEquuscaballus
Mean(n)
83(12)
50.4
(11)
40.8
(20)
47.1
(29)
49(28)
44.5
(30)
Min–max
75–89
47–51
37.4–44.1
44–50.5
43–51.5
40.5–49.2
CovaNegra
iEquuscaballus
Mean(n)
51.6
(2)
55.1
(2)
53(2)
50(1)
74.8
(1)
Min–max
49.2–54
51.3–58.9
52.1–53.8
8 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

bear), Canis lupus (grey wolf), Panthera pardus spelaea
(Ice Age leopard) and Lynx sp.
Abundant microfaunal taxa are also present, including
the insectivoreCrocidura sp., the rodentsMicrotus arvalis,
Microtus agrestis, Chionomys nivalis, Arvicola terrestris,
Terricola sp., Pliomys lenki and Apodemus sp., and the
lagomorph Lagomorpha indet., as well as Aves, squamates,
the tortoise Testudo hermanni and fish. This small-
vertebrate association is typical of the Late Pleistocene of
the Iberian Peninsula (Cuenca-Bescos et al. 2010; Sauque
and Cuenca-Bescos 2013; Sauque et al. 2014).
4.1. Order Artiodactyla Owen, 1848
4.1.1. Family Cervidae Goldfuss, 1820
Cervids are scarce at Los Rincones, representing just 6.3%
of the ungulates. Although they only form a small
proportion of the total, their presence is interesting as it
indicates a woody environment (Table 1; Figure 2).
4.1.1.1. Cervus elaphus Linnaeus, 1758. Thirteen
remains were recovered, representing 2.1% of the
ungulates. The dental remains are scarce; only two incisors
were recovered. The most abundant remains are the
phalanges (six), two first phalanges, two second and two
third phalanges; also there are one metatarsus and one tibia
(Figure 3).
The metatarsus (Ri10/O-13/172) presents a clear
separation between grand cuneiform facet and escafocu-
boides facet such as in Cervus elaphus while in Dama
dama they appear to meet (Lister 1996). Also, Ri10/O-
13/172 shows a single large foramen typical of Cervus
elaphus while Dama dama presents a network of pores
(Lister 1996). First phalanx (Ri10/P13/1, Ri10/O-13/81) of
Los Rincones shows a posterior articular facet with a mid-
groove which is typical of Cervus elaphus (Lister 1996).
The dimensions of the postcranial material that was
found correspond to a medium-sized deer, one larger than
the deer from Cueva Camino and Cova Negra (MIS5), but
somewhat smaller than those from the north of the Iberian
Peninsula (the sites of Lumentxa, Santimamine, Urtiaga D,
Labeko Koba, La Paloma, El Miron, Morın and Abauntz)
that correspond to the last cold stage (MIS3 and MIS2).
The deer from Los Rincones is similar in size to deer from
Gabasa, Parpallo and Nerja (MIS3 and MIS2), which are
found in a Mediterranean climatic environment (Table 2).
The larger size of these deer from the cold stages was
already pointed out by Mariezkurrena (1983). This greater
size could be related to Bergmann’s rule (cold climates
select for the survival of large-sized animals), or it could
be due to differences in food quality, the high quality of the
vegetation during the cold periods allowing ruminants
such as deer to attain large sizes (Guthrie 1990).Gabasaj
Equuscaballus
Mean(n)
82.4
(4)
51(3)
41.5
(6)
48.3
(2)
49.1
(2)
46(2)
68.4
(1)
78.8
(1)
Min–max
80–88
46.4–54.6
39–43.9
47.5–49
46.6–51.5
43–49
Abauntzk
Equusferuscf.gallicus
Mean(n)
82.5
(3)
56.5
(1)
48.5
(2)
49(1)
54.5
(1)
52(1)
64(2)
77(1)
Min–max
78–85
48–49
63–65
Hornosdela
Penal
Equuscaballus
Mean(n)
80(1)
55(1)
42(1)
47(1)
57(1)
53(1)
Min–max
TitoBustillo
mEquusferusgallicus
Mean(n)
83(2)
55(2)
47.5
(2)
49.5
(1)
57(1)
49(1)
Min–max
53–57
42–53
Equusferusprzew
alskiiextantn
Equusferusprzew
alskii
Mean(n)
78.1
(27)
49.6
(27)
43.8
(27)
44.4
(85)
49(85)
43.3
(85)
64.6
(47)
70.7
(47)
Min–max
74–84
46–55
40.5–47.1
40–50
45–54
39.5–52
56–78
63.5–78
Notes:1stPha.,firstphalanx;2ndPha.,secondphalanx;3rd
Pha.,thirdphalanx;L,maxim
um
length;W,maxim
um
width;DW,distalwidth;PW,proxim
alwidth.
aThisstudy;bCastanos(1982);
cCastanos(1986);
dSanchez
(1990);
eAltuna(1973);
f Altuna(1972);
gCardoso
andEisenmann(1989);
hDavidson(1989);
i Perez-Ripoll(1977);
j Blasco-Sancho(1995);
kAltuna
etal.(2002);
l Yravedra
(2010);
mAltuna(1976);
nEisenmannonlinedata.
Historical Biology 9
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

4.1.1.2. Capreolus capreolus Linnaeus, 1758. Twenty-
six remains were recovered, representing 4.2% of the
ungulates. Cranial and postcranial remains were found
(Figure 3). The minimum number of individuals is two,
one adult and one subadult of roughly 19–20 months.
Among the remains recovered, an antler (Ri10/
O13/230) is particularly notable for its taxonomic value.
This is composed of a central branch that presents a tip that
grows forwards and upwards in its upper part. The rear tip
projecting backwards situated a third of the way along the
central branch is not observed, since the individual is
juvenile. It presents great development of the rosettes, with
pearled patterning. These characteristics are typical of
Capreolus capreolus (Sempere et al. 1996; Mateos-
Quesada 2011) (Figure 3).
The remains that could be measured have been
compared with the measurements from the bibliography
(Table 3). The remains recovered from Los Rincones are
similar in size to the roe deer from Gabasa (MIS3) but
smaller than those from the north of the Iberian Peninsula
(the sites of Jou Puerta, Urtiaga, La Riera, Santimamine
and Lumentxa) that correspond to the last cold stage
(MIS3 and MIS2) (Table 3).
The presence of Capreolus capreolus is interesting
since the species does not occur frequently in the sites of the
Quaternary of western Europe (Altuna 1972; Lister 1986;
Figure 3. (a) Right tibia Cervus elaphus (Ri10/O-13/14). (b) Right metatarsus Cervus elaphus (Ri10/O-13/172). (c) Left radiusCapreolus capreolus (Ri10/O-13/235). (d) Metacarpus Capreolus capreolus (Ri10/O-14/87). (e) Left antler Capreolus capreolus (Ri10/O-13/4; Ri10/O-14/45). (f) Right antler Capreolus capreolus (Ri10/O-13/230). (g) Right tibia Capreolus capreolus (Ri10/O-13/75). (h)Left mandible m2–m3 Capreolus capreolus (Ri10/O-13/73). (i) Right humerus Rupicapra pyrenaica (Ri10/N10/34). (j) Left tibiaRupicapra pyrenaica (Ri10/N10/188). (k) Left lower third molar Rupicapra pyrenaica (Ri10/N10/72). (l) Right lower third molarRupicapra pyrenaica (Ri10/N10/71).
10 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Alvarez-Lao 2014). In the Iberian Peninsula, it has mainly
been recovered in northern regions, though its presence
never represents more than 20% of the NISP of the
ungulates of the sites in question (see Appendix 1 %NISP).
4.1.2. Family Bovidae Gray, 1821
Bovids are predominant in the ungulate association from
Los Rincones, constituting 91.2% of the NISP of ungulates.
The most abundant taxon is the Iberian wild goat (Capra
pyrenaica), followed by much lower percentages of the
Pyrenean chamois (Rupicapra pyrenaica) and the aurochs
(Bos primigenius) (Figure 2; Table 1).
4.1.2.1. Bos primigenius Bojanus, 1827. The remains of
Bos primigenius are scarce, representing just 0.7% of the
NISP of ungulates.Moreover, they seem to belong to a single
individual, since theywere collectedwithin a very small area,
are similar in size and articulate with one another. The
remains recovered belong to the postcranial skeleton: a talus
bone, a metatarsus, a scaphocuboid and a third phalanx
(Figure 4).
The distinction between Bos and Bison is complex, and
the absence of the cranial or dental remains that are best
for distinguishing them obliged us to use a variety of
morphological and metric criteria. In spite of the
complexity, the distinction is relevant because the taxa
Figure 4. (a), (b) Right metatarsus Bos primigenius (Ri10/O-13/147). (c) Right astragalus Bos primigenius (Ri10/N10/14). (d) Firstphalanx Equus ferus (Ri10/N10/31). (e) First phalanx Equus ferus (Ri10/N11/1) (f) First phalanx Equus ferus (Ri10/O-13/71). (g) Thirdphalanx Equus ferus (Ri10/N10/64). (h) Second phalanx Equus ferus (Ri10/O-14/53). (i) Second phalanx Equus ferus (Ri10/N10/98). (j)Second phalanx Equus ferus (Ri10/N10/150). (k) Third phalanx Equus hydruntinus (Ri10/M10/10). (l) Third phalanx Equus hydruntinus(Ri10/N10/57).
Historical Biology 11
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

indicate different environments: Bison priscus is associ-
ated with steppe regions (Brugal 1985), whereas Bos
primigenius is found both in open and woodland areas
(Ekstrom 1993).
Only the proximal part of the metatarsus is preserved,
where the proximal articular facets corresponding to the great
cuneiform and the scaphocuboid can be seen to be slightly
separated, which is a trait characteristic of Bos, whereas in
Bison they are joined (Brugal 1985; Gee 1993). Moreover,
these facets are asymmetrical,which differentiates them from
Bison (Alvarez-Lao and Garcıa-Garcıa 2006). Another
criterion that supports an ascription of the fragment to Bos
is the lack of the medial tubercle (Gee 1993).
With the talus bone, we have followed morphological
and metric criteria. In the case of Bos, the morphology of
the plantar groove separating the articular surfaces of the
calcaneus and the scaphocuboid displays an angle of 908both in the plantar and medial area of the bone and on the
lateral margin, whereas in Bison it follows a gently arching
course that ends in an open angle (Altuna 1972; Buitrago-
Villaplana 1992; Gee 1993; Sala et al. 2010). Specimen
Ri10/N10/14 clearly presents straight angles; furthermore,
it does not display an articular facet for the scaphocuboid,
which is a character typical of Bison (Bibikova 1958;
Altuna 1972; Buitrago-Villaplana 1992; Gee 1993; Sala
et al. 2010). The metric criteria applied to the talus are the
index of the lateral articular facet and that of the distal
trochlea (Bibikova 1958). Table 4 presents the values for
these indices, as given by various authors. It can be seen
that there is an overlap between the two species in these
values (Sala et al. 2010), though values above 56.6 for the
articular facet index have only been published for Bos
(Stampfli 1963). For the distal trochlea index, the overlap
is substantial, but values below 81.2 correspond to Bison
(Altuna 1972), whereas values above 100 correspond to
Bos (Stampfli 1963). The value of the lateral articular facet
index for specimen Ri10/N10/14 is 60, and the distal
trochlea index has a value of 110. Both these values place
it clearly within the range of variability of Bos and exclude
it from that of Bison. In short, metric and morphological
characteristics enable us to assign the large-sized bovid
remains from Los Rincones to the species Bos primigen-
ius, which inhabited Europe until historical times
(Degerbøl and Fredskild 1970).
4.1.2.2. Capra pyrenaica (Schinz, 1838). The Iberian
wild goat is thepredominant taxonat the site ofLosRincones;
528 remains were recovered, which represent 85.9% of the
totalNISPofungulates (Table 1). Theage ofdeath reveals the
presence of 20 individuals at the site. Grouped by age, these
are one neonate individual, three juvenile individuals, seven
subadults, five adults and four old individuals.
The sample includes almost all the anatomical
elements, especially bones of the extremities, but also
cranial remains, mainly isolated teeth, axial elements,
scapulae and pelvis (Figures 5–7).
The taxonomy of the genus Capra is based principally
on the morphological characteristics of the horns. In the
absence of these elements, M3 has been one of the most-
used elements, in particular the presence or absence of the
metastylar wing and the width of the interstylar surface
(Cregut-Bonnoure 1992). On the basis of these characters,
Cregut-Bonnoure (1992, 2006) proposed that Capra
pyrenaica is derived from a common ancestor with the
Caucasian goat (Capra caucasica praepyrenaica), which
would have arrived at the Massif Central in France in the
course of the Eemian. Once there, it would have evolved
into Capra pyrenaica, moving towards the south of France
and reaching the Pyrenees in the Magdalenian. On this
theory, from the Pyrenees it would have subsequently
colonised the Iberian Peninsula. Currently, in the light of
the presence of Capra in Iberia for more than 100 kyr
(Altuna 1992), the high variability of the distinguishing
morphological characters proposed by Cregut-Bonnoure
(Magniez 2009) and the results of molecular studies
showing the kinship of Capra ibex and Capra pyrenaica
(Pidancier et al. 2006; Urena et al. 2011), the hypothesis of
the Caucasian origin of Capra pyrenaica has rather lost
plausibility (Garcıa-Gonzalez 2011). Although the exact
taxonomic classification of Iberian goats is far from being
resolved, however, it does seem very likely that Capra
pyrenaica was already present and differentiated in the
Late Pleistocene. Accordingly, it would make sense to
ascribe the remains from Los Rincones to this taxon,
irrespective of possible hybridisations with Capra ibex that
might have taken place in the northeast of the Iberian
Peninsula and south of France during this period (Garcıa-
Gonzalez 2012).
The comparative data for Capra pyrenaica presented
in Table 5 suggest the existence of two groups (clades?) in
the Late Pleistocene of the Iberian Peninsula, separable by
their dimensions: one northern group, characterised by
their large size, and another southern group that is smaller
in size (Table 5; Figure 8). The Capra pyrenaica remains
from Los Rincones would fall within the range of variation
of the first group. This size gradient would have been
reproduced in the Holocene (Castanos 2004) and at
present, for this and other characteristics have been
recognised in the present-day subspecies (Cabrera 1911).
Do the differences in size among Iberian goats in the Late
Pleistocene correspond to phylogenetic differences? The
current subspecific differentiation of Capra pyrenaica is a
matter of controversy (Acevedo and Cassinello 2009), and
the systematics should probably be revised in the near
future, even at the specific level (Garcıa-Gonzalez 2011).
The north–south size gradient of the Iberian goats does not
seem to be a temporary case of Bergmann’s rule, but seems
rather to be due to the possible abundance of trophic
resources in two contrasting environments: the Eurosiber-
12 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

ian (temperate and fertile) and the Mediterranean (arid and
poor). The increase in size that took place in Capra
pyrenaica in the interglacial period (MIS5) at Cova Negra
(Perez-Ripoll 1977; Table 5; Figure 8) or during the
Holocene (Garcıa-Gonzalez 2012) would support the
hypothesis of the abundance of resources.
4.1.2.3. Rupicapra pyrenaica Linnaeus, 1858. As
regards the Pyrenean chamois, 29 remains were recovered,
representing 4.7% of the NISP of the ungulates at the site
(Table 1; Figure 2). Both appendicular and cranial
elements were found. Among the cranial material, three
m3, one m2, one M2 and one M3 were recovered (Figure 3).
The m3 presents an external wall with a metastyle and
parastyle and an oval talonid with a practically polygonal
section. The distal edge is straight and not convex (Prat
1966).
The talus bones, with four elements recovered, and the
phalanges, with six elements recovered, are the most
numerous elements of Rupicapra pyrenaica at the site, and
their measurements have been compared with a sample of
Rupicapra pyrenaica from the Pleistocene of the Iberian
Peninsula (Table 6). The chamois from Los Rincones is
similar in size to that from Cueva Millan and smaller than
those from the sites in the north of the Iberian Peninsula
(Santimamine, Abauntz, Bolinkoba, Lumentxa, Ekain,
Naranco, Aitzbitatre, Lezetxiki, Urtiaga, Amalda,
La Paloma and Valdegoba) that correspond to the last
cold stage (MIS3 and MIS2). Furthermore, it is smaller
than those from the Ebro Valley sites of Gabasa and
Chaves (MIS3 and MIS2, respectively) (Table 6).
The genus Rupicapra (chamois) is a small-sized
mountain goat-antelope that inhabits Europe and western
Asia. The genus comprises two different species:
Rupicapra rupicapra, present in the Alps, the Balkans
and the Caucasus; and Rupicapra pyrenaica, present in the
Iberian Peninsula and Italy (Lovari and Scala 1980;
Nascetti et al. 1985; Perez et al. 2002).
The time of divergence between Rupicapra rupicapra
and Rupicapra pyrenaica has been estimated on the basis
of their genetic distance, but different methods yield
different ages. Molecular clocks show a separation that
took place at ca. 1.7Ma (Rodrıguez et al. 2010), whereas
phylogenetic analysis based on Y chromosomes has given
a more recent age of 655 ka (Perez et al. 2011). These
genetic studies imply that the chamois recovered from the
Late Pleistocene of the Iberian Peninsula belong to the
species Rupicapra pyrenaica (Alvarez-Lao 2014).
4.2. Order Perissodactyla Owen, 1848
4.2.1. Family Equidae Gray, 1821
The systematics of horses is under revision. To underline
the difference between domestic and wild species, the
Figure 5. Remains of Capra pyrenaica. (a) Left maxilla P2–M3 (Ri10/N10/41). (b) Right mandible of a juvenile p2-p4 (Ri10/O-13/92).(c) Left mandible p3–m3 (Ri10/N10/117). (d) Left M3 (Ri10/O-13/126). (e) Right M3 (Ri10/N10/83). (f) Right M3 (Ri10/GL9/9). (g)Right M3 (Ri10/O-13/201). (h) Right M3 (Ri10/O-13/203). (i) Right mandible p3–m3 (Ri10/O-13/27). (j) Atlas (Ri10/O-13/118). (k)Atlas (Ri10/N11/2). (l) Atlas (Ri10/O-13/82).
Historical Biology 13
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

International Commission on Zoological Nomenclature in
2003 proposed that the name of a wild species – in the
case of the horse Equus ferus Boddaert, 1785 – should
take precedence over the name of the domesticated
species. However, the name of the domesticated species,
Equus caballus Linnaeus, 1758, continues to be valid
(Gentry et al. 2004; Conti et al. 2010). Accordingly, we
here use the designation Equus ferus to refer to the horses
of the Pleistocene even though they may previously have
been classified as Equus caballus.
It is difficult to distinguish between the different
species of equids on the basis of morphological characters
alone (Eisenmann 1986; Dive and Eisenmann 1991;
Orlando et al. 2009). In the present case, this is particularly
difficult due to the lack of preserved cranial remains,
which are those that provide most systematic information.
Figure 6. Remains of Capra pyrenaica. (a) Right humerus (Ri10/N11/41). (b) Right humerus (Ri10/O-14/12). (c) Right humerus (Ri10/J10/1). (d) Left humerus (Ri10/O-13/179). (e) Left radius (Ri10/O-13/165). (f) Left radius (Ri10/N10/208).(g) Right metacarpus (Ri10/N10/195). (h) Right metacarpus (Ri10/O-13/199). (i) Left metacarpus (Ri10/N19/151). (j) Left metacarpus (Ri10/M9/17). (k) Leftmetacarpus (Ri10/GL7/19; Ri10/GL7/2; Ri10/GL7/5). (l) Right metacarpus (Ri10/O-13/184).
14 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

For this reason, the remains have been classified by means
of a biometric study.
4.2.1.1. Equus ferus Linnaeus, 1758. A fragment of
metapod and phalanges were recovered from the site: four
first phalanges, three of which are posterior, six second
phalanges and a third phalanx (Figure 4). The phalanges
were distinguished as posterior or anterior in accordance
with the criteria proposed by Prat (1957), Eisenmann and
De Giuli (1974) and Dive and Eisenmann (1991) (Figures 9
and 10).
As a whole, the horse from Los Rincones is small in
size compared with the other horses of the Late
Pleistocene of the Iberian Peninsula, and especially
compared with the horses of the Upper Palaeolithic of
the Basque Country from sites such as Atxuri, Santima-
mine and Lumentxa (Castanos 1984) (Table 7; Figure 11).
Its size is similar to that of the horse from Fontainhas,
which forms part of the subspecies Equus ferus antunesi,
characterised by its small dimensions (Cardoso and
Eisenmann 1989), and it is also close in size to the
subspecies Equus ferus casarensis described at Los
Casares (Altuna 1973). The measurements of the horse
from Los Rincones are very similar to the current
population of Equus ferus przewalskii (Table 7). The third
phalanx from Los Rincones presents a small maximum
width, which may be related to the type of land over which
Figure 7. Remains ofCapra pyrenaica. (a) Lumbar vertebras and sacrum (Ri10/O-13/90, Ri10/O-13/93). (b) Left scapula (Ri10/GL8/5).(c) Left scapula (Ri10/O-13/219). (d) Left tibia (Ri10/GL9/12). (e) Left tibia Ri10/M9/19). (f) Right tibia (Ri10/O-13/102). (g) Right tibia(Ri10/M10/3). (h) Right tibia (Ri10/P13/5). (i) Right femur (Ri10/O-13/218; Ri10/O-13/234; Ri10/O-14/15).
Historical Biology 15
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

the animals moved; large ungual phalanges are associated
with soft ground, whereas narrow ones indicate an adaptation
for movement over craggy reliefs (Eisenmann 1984).
Despite its small size, the horse from Los Rincones
does not fall within the range of variation for the
measurements of Equus hydruntinus; as can be seen in
Figure 9. It lies within the range of variation for Equus
ferus przewalskii as well as for the Equus ferus of the Late
Pleistocene.
4.2.1.2. Equus hydruntinus Regalia, 1907. Two remains
of Equus hydruntinuswere recovered, which correspond to
two third phalanges (Figure 4). This taxon is an indicator
for conditions of aridity and open spaces (Burke et al.
2003; Orlando et al. 2006; Feranec et al. 2010).
Equus hydruntinus was described by Regalia (1907) in
the Grotta Romanelli. As it showed characteristics similar
to the onager, he designated as Equus (Asinus) hydruntinus
(Conti et al. 2010). Subsequent studies of the systematic
Figure 8. Graph representing the maximum value, minimum and the mean of Capra pyrenaica from different Pleistocene sites andextant animals. Mtcp., metacarpus; Mtt., metatarsus; Tib., tibia; Hum., humerus; L, maximum length; DW, distal width.
16 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

position of this taxon classified it as an ass (Stehlin and
Graziosi 1935; Gromova 1949), a zebra (Davis 1980) and a
stenonian form (Forsten 1999). In recent years, however,
the study of material from Crimea has brought to light the
phylogenetic affinity between Equus hydruntinus and
Equus hemionus (Burke et al. 2003), confirmed by DNA
studies on remains from Crimea and Iran (Orlando et al.
2006). Equus hydruntinus was the last species of the
Pleistocene megafauna to go extinct in the Iberian
Peninsula, as there is evidence of its presence as late as
the 16th century (Quesada and von Lettow-Vorbeck 1992).
As there are no available biometric data for the third
phalanges of Equus hydruntinus and these display
similarly proportioned bones of the extremities to onagers
(Eisenmann and Baryshnikov 1995; Forsten 1999; Burke
et al. 2003; Orlando et al. 2006), we have used the
measurements from modern-day onagers taken from
Eisenmann’s collection (online data at http://www.
vera-eisenmann.com) for the purpose of biometric
comparison.
The bones of the extremities of Equus hydruntinus are
slender, and the third phalanges are pointed (Davis 2002).
Figure 9. Graph representing greatest length versus the proximal width of the first phalanx of fossil and extant Equus species. Speciesdata points and corresponding regression lines are plotted together.
Figure 10. Graph representing greatest length versus the greatest width of the third phalanx of fossil and extant Equus species. Speciesdata points and corresponding regression lines are plotted together.
Historical Biology 17
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

The third phalanges from Los Rincones with a size similar
to Equus hemionus (Figure 10) and a sharp-pointed end
belong to Equus hydruntinus.
5. Faunal composition
5.1. Analysis of the faunal composition in theEuropean context of MIS3–MIS2
The faunal composition, expressed as the percentage of
each herbivore species at the site of Los Rincones, was
compared with that from 89 European sites, with a total of
127 levels with different chronologies, but all of them
belonging to the isotope stages MIS3 and MIS2. Of these
sites, 56 are located in the Iberian Peninsula, and 33 in the
rest of Europe, both in western and central parts (France,
United Kingdom, Germany, the Czech Republic and
Switzerland; see Appendix 1). To compare the faunal
associations present at each site, the NISP was calculated
for the herbivores in each case. This parameter is
considered the most reliable (Lyman 1994b), as there is
no taphonomic bias towards any of the species in the
assemblages (Alvarez-Lao 2014). In order to minimise
the taphonomic biases that might be produced by the
accumulators of the remains, this study has taken account
Figure 11. Graph representing maximum value, minimum and the mean of Equus from different Upper Pleistocene sites and extantanimals. 1st Pha., first phalanx; 2nd Pha., second phalanx; L, maximum length; PW, proximal width.
18 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

both of sites where the accumulators are hunter-gatherers
(Chaves, Grotte de la Vache, Gabasa, Abric Romanı,
Santimamine and Nerja) and of those where they are
carnivores (Zafarraya, Los Rincones, Zarzamora and Las
Caldas). It has only used assemblages with an NISP
greater than 100, in accordance with the methodology
proposed by Alvarez-Lao (2014). Moreover, certain taxa
have been grouped together, as in the case of Capra
pyrenaica and Capra ibex (C.p/i) and Rupicapra pyrenaica
and Rupicapra rupicapra (R.r/p). We follow Deng et al.
(2011) in the taxonomic nomenclature of Stephanorhinus
hemitoechus.
The CA has allowed us to place the sites with a similar
taxonomic composition close to one another. At the same
time, the taxa with a similar distribution throughout the
sample should also be placed close to one another.
The DCA shows a clear separation of the sites of
central and Western Europe in relation to those of the
Iberian Peninsula. The sites of central and western Europe
(black dots) are situated on the right of the graph and are
characterised by the presence of the faunas of the
‘Mammuthus-Coelodonta faunal complex’, comprising
Mammuthus primigenius, Coelodonta antiquitatis, Mega-
loceros giganteus, Rangifer tarandus, Saiga tartarica,
Equus ferus and bovids (Bison priscus/Bos primigenius)
(Guthrie 1990; Kahlke 1999; West 2000). The mammoth
and the woolly rhinoceros have been recovered in the
Iberian Peninsula at 25 and 23 sites, respectively (Alvarez-
Lao and Garcıa 2011, 2012). However, none of the Iberian
sites presents this fauna at percentages that place it within
the area of sites with the typical faunal association of the
Mammuthus–Coelodonta complex. An exception among
the sites in the west is La Grotte de la Vache in the French
Pyrenees. Its position on the left of the graph is due to the
abundance of Capra pyrenaica, which represents 88% of
the NISP of the ungulates (Pailhaugue 1995) (Figure 12).
The sites of the Iberian Peninsula (grey dots) are
located on the left of the graph, with the exception of
Urtiagako Leizea. This site is situated on the right due to
its high numbers of reindeer, Rangifer tarandus (Altuna
1984), which bring it closer to the French sites of the
Massif Central, where the reindeer is very abundant
(Delpech 1983, 1990). The sites of the Iberian Peninsula
are characterised by the presence of taxa associated with
more temperate environments than the cold European
faunas. Such taxa are Capra pyrenaica, Cervus elaphus,
Figure 12. Ordination of palaeontological sites and species along the first two axes of the DCA on the basis of their ungulatecomposition. Grey, Iberian Peninsula sites; black, Western and Middle European sites; star, Los Rincones. Taxa abbreviations: R.t.,Rangifer tarandus; C.e., Cervus elaphus; M.g.,Megaloceros giganteus; C.c., Capreolus capreolus; C.i./p., Capra ibex/pyrenaica; R.r./p.,Rupicapra rupicapra/pyrenaica; S.t., Saiga tatarica; Bov., Bovinae indet.; S.s., Sus scrofa; E.f., Equus ferus; E.h., Equus hydruntinus; C.a., Coelodonta antiquitatis; S.h., Stephanorhinus hemitoechus; M.p., Mammuthus primigenius. Site abbreviations: Zaf, Zafarraya; Ne,Nerja; Vg, Vanguard Cave; Go, Gorham’s Cave; G.V, Grotte de la Vache; Ab R, Abric Romanı; Ge, Cova del Gegant; Mo, Cueva Morın;La, Labeko Koba; Lez, Lezika; Ar, Arbreda; Zar, Zarzamora; Du, L’abri Duruthy; Mor, Morancourt; Srb, Srbsko Chulum-Komin Cave; H.S, Hohle Stein; Pe, Perick Cave; Vo, Vogelherd (IV–V); Ly, Lynford; Gei, Geissenklosterle; Hor, L’Hortu; U.L, Urtiagako Leizea; Fac,Abri du Facteur; Com, Roc de Combe; Lau, Laugerie Haute Ouest.
Historical Biology 19
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Capreolus capreolus, Rupicapra pyrenaica, Sus scrofa and
Stephanorhinus hemitoechus (steppe rhinoceros)
(Figure 12).
The DCA shows an oblique general gradient of
increasing latitude and decreasing temperature, running
from the upper right to the lower left corner. In the upper
right corner are the sites of Lynford (UK) and
Geissenklosterle, Hohle Stein, Vogelherd and Perick
Cave (Germany), sites characterised by the presence of
larger-sized taxa of the Mammuthus–Coelodonta com-
plex, such as Mammuthus primigenius, Coelodonta
antiquitatis and Megaloceros giganteus. In the lower
right corner of the graph, we find the sites located in the
Massif Central in France, such as Abri Pataud, Roc de
Combe, Laugerie Haute-Ouest and Abri Facteur among
others, which are characterised by a predominance of
Rangifer tarandus and Saiga tatarica. On the far right of
the horizontal ordination of the graph, we find both French
sites (Morancourt, Duruthy and Hortus) and Middle
European sites (Srbsko Chulum-Komin Cave from Czech
Republic), characterised by the presence of Equus ferus
and large bovids. Close to these sites are Iberian sites such
as l’Arbreda, Abric Romanı and Cova del Gegant
(Catalonia), and Cueva Morın, Labeko Koba and Lezika
(Basque-Cantabrian region). Both the northern part of
Catalonia and the Basque Country functioned as faunal
corridors in the Pleistocene (Alvarez-Lao and Garcıa
2011), so it is hardly surprising that these areas are the
most similar to European ones. Despite their similarity to
European sites, these sites exhibit a greater proportion of
Equus hydruntinus, associated with arid environments
(Burke et al. 2003; Feranec et al. 2010). The sites of the
Iberian Peninsula hardly show any variation with respect
to axis 2 (horizontal alignment), whereas their distribution
in relation to axis 1 marks a transition from the
Mediterranean conditions of the sites in the south (Nerja,
Zafarraya, Gorham’s Cave and Vanguard Cave) on the left
of the graph, with a high proportion of Capra pyrenaica,
towards more Atlantic, mid-European conditions, exem-
plified by the sites of the Basque Country and Catalonia on
Figure 13. Ordination of palaeontological sites and species of the Iberian Peninsula along the first two axes of theDCAon the basis of theirungulate composition. White: Iberian Peninsula sites; Star: Los Rincones. Taxa abbreviations: C.e., Cervus elaphus; C.c., Capreoluscapreolus; C.i./p., Capra ibex/pyrenaica; R.i/p., Rupicapra rupicapra/pyrenaica; Bov., Bovinae; S.s., Sus scrofa; E.f., Equus ferus; E.h.,Equus hydruntinus; S.h., Stephanorhinus hemitoechus. Site abbreviations: Zar, Zarzamora; Bu, Cueva del Buho; Mo, Cueva Morın; Ge,Cova del Gegant; La, Labeko Koba; Fi, Figueira Brava (2–3); Ar, Arbreda; Es, Escoural; Ab R, Abric Romanı; Pe, Cueva del Pendo; Mi,Cueva Millan; Le, Lezetxiki (IIIa); HP, Hornos de la Pena; Ax, Axlor; Am, Amalda; Va, Valdegoba; Ai, Aizbiarte IV; Ab, Abauntz; Er,Cueva Ermita; JP, JouPuerta; Arr, Arrillor; PD, Pego doDiablo; GC,Grota Nova da Columbeira; Co, Covalejos, At, Atxagakoa; Li, Linares;Sa, Santimamine; Lu, Lumentxa n VII; Ca, Los Casares; Cal, Las Caldas; Mie, Pena Miel; Ga, Gabasa 1; Be, Cova Beneito; Vg, VanguardCave; Ne, Nerja; Zaf, Zafarraya.
20 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

the far right of the graph (with the exception of La Grotte
de la Vache). In spite of being located in an area that is
climatically at the boundary between Atlantic and
Mediterranean environments, the site of Los Rincones is
closer to the sites of the south in faunal terms (Figure 12).
5.2. The site of Los Rincones in the Iberian context of
MIS3
The herbivore association present at the site of Los
Rincones was compared with those at 37 sites in the Iberian
Peninsulawith a chronology close toMIS3 (Figure 13). The
CA shows that the sites with taxa associated with open
environments such as Equus hydruntinus, Equus ferus,
Stephanorhinus hemitoechus and the large bovids (Brugal
1985; Burke et al. 2003; Feranec et al. 2010; Sala et al.
2011) are situated in the left part of the graph. These sites
are Cueva de la Zarzamora, CuevaMorın, Cova del Gegant,
Labeko Koba and Figueira Brava. On the right, by contrast,
are the sites of the south of the Iberian Peninsula such as
Zafarraya, Nerja, Vanguard Cave and Cova Beneito, which
are characterised by a great abundance of Capra pyrenaica
associated with rocky areas. Furthermore, it should be
borne in mind that this graph shows a comparison of sites
from MIS3, at which time human hunter-gatherers had not
yet specialised in hunting Capra. This phenomenon
appeared in the Upper Palaeolithic (MIS2) (Freeman
1973; Straus 1987; Gamble 1995). As such, the high
percentages of Capra in the Mediterranean sites cannot be
attributed to the presence of specialised hunters, but rather
to the abundance of these taxa in the area of the
accumulation. In the central part of the graph, we find
mainly Basque–Cantabrian sites under Atlantic influence.
Axis 2 separates sites with woodland taxa such as Cervus
elaphus, Capreolus capreolus and Sus scrofa (Covalejos,
Linares, Lumentxa and Aitbiarte) in the lower part from
sites in the upper central part (Amalda, Valdegoba and
Hornos de la Pena) characterised by the presence of taxa
associated with rocky environments such as the chamois,
but with a more Atlantic climatic affinity than Capra
pyrenaica (Feranec et al. 2010).
Despite currently being located in a geographical
situation close to the boundary between the Mediterranean
and Atlantic climatic regions, in MIS3 the site of Los
Rincones was close to sites such as Zafarraya, Nerja, Cova
Beneito and Vanguard Cave situated in the Mediterranean
climatic region in the south of the Iberian Peninsula and
characterised by a strong presence of Capra pyrenaica, a
species with a high drought-tolerance (Feranec et al.
2010).
The faunal association from los Rincones is similar
to southern Iberian sites (Zafarraya, Nerja and
Vanguard). This fact may reflect similar environmental
conditions, but it can also be due to a bias produced by
the main accumulator agent. In Los Rincones, most of
the ungulates recovered were transported there by Ice
Age leopards, which used the cave as a refuge for
protecting their kills (Sauque et al. 2014). This predator,
as his relative the Panthera uncia (snow leopard), shows
a predilection by preys of small–medium size, especially
goats (see references in Diedrich 2013). This phenom-
enon, added to the placement of the cave in a rocky
wall, may produce an overrepresentation of Capra
pyrenaica. Anyway, although leopards focus mainly on
prey ranging from 20 to 80 kg in weight (Mills and
Harvey 2001), they are not selective hunters and have
the broadest diet of the larger predators with 92 prey
species recorded in sub-Saharan Africa (Mills and
Harvey 2001; Hayward et al. 2006), which may indicate
that a high percentage of Capra pyrenaica is related to
its great abundance in the landscape which surrounds the
cave. However, the low percentage of equids, rhinoceros
and bovids (Bos primigenius and Bison) found in Los
Rincones could not be related to a low abundance of
these taxa in the environment, but to the impossibility of
leopards to drag these preys to their dens because they
weigh more than the range the leopard could hunt.
Therefore, the abundance of these might not reflect the
‘real’ situation surrounding the cave and subsequently
the faunal association would be more similar to the
Iberian sites placed in the inland such as Zarzamora,
Cueva del Buho and Los Casares.
To sum up, the analysis of faunal associations is a great
tool to analyse the environmental patterns, though we have
to take these results carefully, especially when the
accumulator may generate bias in the association.
6. Palaeoenvironmental context
The herbivores at Los Rincones represent various types of
landscape. The horses (Equus ferus and Equus hydrunti-
nus) indicate open environments (Nowak 1999; Blasco
et al. 2011), and Equus hydruntinus is also indicative of
conditions of semi-aridity (Burke et al. 2003; Feranec et al.
2010) (Figure 14). However, Cervus elaphus and
Capreolus capreolus suggest woodland habitats (Delpech
and Prat 1980; Carranza et al. 1991; Mateos-Quesada
2011; Carranza 2011), whereas Bos primigenius can
inhabit both woody environments and plains (Ekstrom
1993). However, the best-represented herbivores both in
terms of NISP and MNI are those associated with high or
medium mountain areas with abrupt reliefs, such as Capra
pyrenaica and Rupicapra pyrenaica (Alados and Escos
1996; Granados et al. 2001; Perez-Barberıa et al. 2010)
(Figure 15). In this context, it is interesting to bear in mind
that the site has also yielded a peripheral bone from the
shell of a Hermann’s tortoise (Testudo hermanni), which is
Historical Biology 21
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Figure 14. In the forefront, a herd of horses (Equus ferus). Behind, to the right, three Equus hydruntinus might be seen in a steppesurrounding Los Rincones cave. In the background, the Moncayo massive (2315m a.s.l) (Illustration Gianfranco Mensi).
Figure 15. Ice Age leopard (Panthera pardus spelaea) chasing a herd of Spanish wild goat (Capra pyrenaica) in the ravine of LosRincones. Behind, the mouth of the cave (1000m a.s.l) (Illustration Gianfranco Mensi).
22 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

a good indicator of Mediterranean conditions (Morales
Perez and Serra 2009).
The great abundance of Capra pyrenaica and the
presence of Rupicapra pyrenaica indicate that in the
vicinity of the site there were rocky mountains. This
landscape would be similar to what we currently find in the
area around the cave, the mouth of which opens at an
altitude of some 1010m in a ravine with rocky walls, ideal
for both species (Figure 1).
7. Conclusions
The ungulate association from Los Rincones provides us
with palaeoenvironmental information about the region
around the Moncayo during MIS3. In spite of its current
importance as a boundary between Mediterranean and
Eurosiberian climatic regions, until now there were no
data available for a reconstruction of its landscape during
the Late Pleistocene.
The great diversity of ungulates present at the site of Los
Rincones, with seven species – Cervus elaphus,Capreolus
capreolus, Rupicapra pyrenaica, Capra pyrenaica, Bos
primigenius, Equus ferus and Equus hydruntinus –
associated with different environments, indicates a high
taxonomic and environmental diversity corresponding both
to the situation of the massif as an ecotone and to the
particular location of the site, not very different from
present-day conditions. Moreover, both Equus hydruntinus
andBos primigenius are species that are now extinct but that
survived into historical times in Europe.
The ungulate association from Los Rincones is
dominated by Capra pyrenaica. Furthermore, this taxon
is similar in size to the goats from the sites in the north of the
Iberian Peninsula, which are larger than present-day goats
and the fossil goats of the south of the Iberian Peninsula.
The horses present at Los Rincones are smaller in size than
those recovered from the Basque–Cantabrian region.
The faunal associations of the Iberian Peninsula during
MIS3 and MIS2 are clearly distinct from those present at
the sites of western and central Europe. The statistical
ordination analysis shows a latitudinal/climatic gradient
from sites with a greater presence of faunas of the
Mammuthus–Coelodonta complex towards sites with a
predominance of species with a Mediterranean affinity
(Capra pyrenaica, Sus scrofa, Cervus elaphus and
Stephanorhinus hemitoechus). At a midpoint are the sites
characterised by the presence of Equus and large bovids
associated with steppe environments.
Our study of faunal associations in the context of the
Iberian Peninsula during MIS3 indicates a cluster of sites
in the south of the peninsula that are separate from the
other sites in the peninsula due to their high presence of
Capra pyrenaica. These sites are Nerja, Gorham’s Cave,
Cueva Beneito and Zafarraya, as well as the site of Los
Rincones, which is included within this group.
The ungulate association of Los Rincones indicates a
landscape with a temperate climate characterised by the
presence of steppes, woodland and rocky areas in
the vicinity of the cave. This landscape would be similar
to the present-day landscape of the ravine of Los Rincones,
where the cave of the same name is located.
Acknowledgements
We thank the Centro Espeleologico de Aragon (SpeleologicalCentre of Aragon), in particular Mario Gisbert, for the discoveryand topography of the cave. The Government of Aragon, theCEA (Centro de Espeleologıa de Aragon) and the University ofZaragoza have partially subsidised the palaeontological activitiesat Los Rincones. This study forms part of the projects ‘GruposConsolidados H54’ of the Government of Aragon and IUCA(Instituto Universitario de Ciencias Ambientales) and CGL2012-38434-C03-01. The first author (VS) also thanks Alfred Sanchis(MPV), Josefina Barreiro and Patricia Perez (MNCN), JulioRamon (MAPH), Mario Laurino (IAN) Jose Ignacio Canudo(MPZ), Henriette Obermaier (LMU) and Caroline Lang (ZSM)for allowing him to study the material for comparison, as well asLourdes Montes, Pillar Utrilla and Mª Fernanda Blasco forgranting access to unpublished data on Gabasa as well as theirvaluable comments. He also thanks Raquel Rabal and CristinaSola for their comments and discussion on taphonomy; VeraEisenmann for her discussions and clarifications on horses; JorgeColmenar for his help with the photographs, and FranciscoGutierrez and Ira Sasowsky for their valuable comments aboutkarst and geology. Rupert Glasgow revised the English grammar.A particular thanks to palaeoartist Gianfranco Mensi for hisprecise reconstruction. The authors also thank Joan MadurellMalapeira and anonymous reviewer for the useful suggestionsthat greatly improved the manuscript. We are especially gratefulfor the comments and improvements of the editor Gareth Dyke.Finally, many thanks go to all the members of the excavationteam of the 2009 and 2010 field campaigns at Los Rincones.
Notes
1. Email: [email protected]. Email: [email protected]
References
Acevedo P, Cassinello J. 2009. Biology, ecology and status of Iberianibex Capra pyrenaica: a critical review and research prospectus.Mamm Rev. 39:17–32.
Aitken RJ. 1975. Cementum layers and tooth wear as criteria for ageingRoe deer (Capreolus capreolus). J Zool. 175:15–28.
Alados CL, Escos J. 1996. Ecologıa y Comportamiento de La CabraMontes y Consideraciones para su Gestion. Madrid: CSIC.
Altuna J. 1971. Los mamıferos del yacimiento prehistorico de Morın(Santander). In: Gonzalez Echegaray J, Freeman LG, editors. CuevaMorın. Excavaciones 1966–1968. Vol. 6. Santander: Publicacionesdel Patronato de Cuevas Prehist; p. 367–398.
Altuna J. 1972. Fauna de mamıferos de los yacimientos prehistoricos deGuipuzcoa. Con catalogo de los mamıferos cuaternarios delCantabrico y del Pirineo occidental. Munibe. XXIV:1–464.
Altuna J. 1973. Fauna de mamıferos del yacimiento prehistorico de LosCasares (Guadalajara). En: La Cueva de Los Casares. 1. Barandiaran.Excavaciones Arqueologicas en Espana. 76:97–116.
Altuna J. 1976. Los mamıferos del yacimiento prehistorico de TitoBustillo (Asturias). In: Moure JA, Cano M, editors. Excavaciones en
Historical Biology 23
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

la cueva de Tito Bustillo (Asturias). Oviedo: Boletın del Instituto deEstudios Asturianos; p. 149–194.
Altuna J. 1984. Primer hallazgo de mamut (Mammuthus primigeniusBlumenbach) en el Paıs Vasco Meridional. Munibe. 36:27–32.
Altuna J. 1986. The mammalian faunas from the prehistoric site of LaRiera. In: Straus LG, Clarck G, editors. La Riera Cave, Stone AgeHunter Gatherer adaptations in Northern Spain. AnthropologicalPapers, Vol. 36. Tucson: University of Arizona; p. 237–274.
Altuna J. 1990. Caza y alimentacion procedente de Macromamıferosdurante el Paleolıtico de Amalda. In: Altuna J, Baldeon A,Mariezcurrena K, editors. La Cueva de Amalda (Zestoa, Paıs Vasco).Ocupaciones paleolıticas y postpaleolıticas. Sociedad de EstudiosVascos, Serie B 4. San Sebastian: Sociedad de Estudios Vascos;p. 149–192.
Altuna J. 1992. El medio ambiente durante el Pleistoceno Superior en laregion cantabrica con referencia especial a sus faunas de mamıferos.Munibe. 44:13–29.
Altuna J, Mariezkurrena K. 1984. Bases de subsistencia de origen animalen el yacimiento de Ekain. El yacimiento prehistorico de la Cueva deEkain (Deba, Guipuzcoa). Sociedad de Estudios Vascos Serie B.1:211–280.
Altuna J, Mariezkurrena K. 1985. Bases de subsistencia de los pobladoresde Erralla: Macromamıferos. In: Altuna J, Baldeon A, MariezcurrenaK, editors. Cazadores magdalenienses en Erralla (Cestona, PaısVasco). Munibe (Antropologia-Arkeologia). Vol. 37. p. 87–117.
Altuna J, Mariezkurrena K. 2000. Macromamıferos del yacimiento deLabeko Koba (Arrasate, Paıs Vasco). Munibe. Ciencias naturales.52:107–181.
Altuna J, Mariezkurrena K, Elorza M. 2002. Arqueozoologıa de losniveles paleolıticos de Abauntz (Arraiz, Navarra). Saldvie. Estudiosde Prehistoria y Arqueologıa, 2. Zaragoza: Departamento deCiencias de la Antiguedad, Universidad de Zaragoza; p. 1–26.
Alvarez-Lao DJ. 2014. The Jou Puerta cave (Asturias, NW Spain): A MIS3 large mammal assemblage with mixture of cold and temperateelements. Palaeogeogr Palaeoclimatol Palaeoecol. 393:1–19.
Alvarez-Lao DJ, Arsuaga JL, Baquedano E, Perez-Gonzalez A. 2013.Last Interglacial (MIS 5) ungulate assemblage from the CentralIberian Peninsula: The Camino Cave (Pinilla del Valle, Madrid,Spain). Palaeogeogr Palaeoclimatol Palaeoecol. 374:327–337.
Alvarez-Lao DJ, Garcıa N. 2010. Chronological distribution ofPleistocene cold-adapted large mammal faunas in the IberianPeninsula. Quatern Int. 212(2):120–128.
Alvarez-Lao DJ, Garcıa N. 2011. Geographical distribution ofPleistocene cold-adapted large mammal faunas in the IberianPeninsula. Quatern Int. 233(2):159–170.
Alvarez-Lao DJ, Garcıa N. 2012. Comparative revision of the Iberianwoolly mammoth (Mammuthus primigenius) record into a Europeancontext. Quatern Sci Rev. 32:64–74.
Alvarez-Lao D, Garcıa-Garcıa N. 2006. A new site from the SpanishMiddle Pleistocene with cold-resistant faunal elements: La Parte(Asturias, Spain). Quatern Int. 142–143:107–118.
Arceredillo D, Gomez-Olivencia A, Garcıa-Perez A. 2011. Threestatistical methods for sex determination in extant and fossilcaprines: assessment of the Rupicapra long bones. J Archaeol Sci.38:2450–2460.
Azorit C, Analla M, Carrasco R, Calvo JA, Munoz-Cobo J. 2002. Teetheruption pattern in red deer (Cervus elaphus hispanicus) in southernSpain. Anales de Biologıa. 24:107–114.
Bibikova VI. 1958. Some distinguishing features in the bones of thegenera Bison and Bos. Bull. Mosk. Obschtschestwa Isp Privoda NSOtdel Biol. 63(6):23–35.
Blasco R, Rosell J, van der Made J, Rodrıguez J, Campeny G, Arsuaga JL,Bermudez de Castro JM, Carbonell E. 2011. Hiding to eat: the role ofcarnivores in the early Middle Pleistocene from the TD8 level ofGran Dolina (Sierra de Atapuerca, Burgos, Spain). J Archaeol Sci.38(2011):3373–3386.
Blasco-Sancho F. 1995. Hombres, fieras y presas. Estudio arqueozoo-logico y tafonomico del yacimiento del Paleolıtico Medio de laCueva de Gabasa 1 (Huesca). Monografıas Arqueologicas 38 [Tesis].Zaragoza: Universidad de Zaragoza.
Brain CK. 1981. The hunters or the hunted? An introduction to Africancave taphonomy. Chicago, IL: University of Chicago Press.
Brugal JP. 1985. Le Bos primigenius BOJ. 1827 du Pleistocene moyendes grottes de Lunel-Viel (Herault). Bulletin du Musee d’Anthro-pologie Prehistorique de Monaco. 28:7–62.
Buitrago-Villaplana AM. 1992. Estudio de los Artiodactilos delyacimiento del Pleistoceno medio de Pinilla del Valle (Madrid)[Tesis doctoral]. Madrid: Universidad Complutense de Madrid.
Burke A, Eisenmann V, Ambler G. 2003. The systematic position ofEquus hydruntinus, an extinct species of Pleistocene equid. QuaternRes. 59:459–469.
Cabrera A. 1911. The subspecies of the Spanish ibex. Proc Zool SocLond. 66:963–977.
Cardoso JL, Eisenmann V. 1989. Equus caballus antunesi, nouvelle sous-espece quaternaire du Portugal. Vera Montpellier: Lab. de Pal. desVertebres.
Carranza J. 2011. Ciervo – Cervus elaphus. In: Salvador A, Cassinello J,editors. Enciclopedia Virtual de los Vertebrados Espanoles. Madrid:Museo Nacional de Ciencias Naturales. http://www.vertebradosibericos.org
Carranza J, Hidalgo de Trucios SJ, Medina R, Valencia J, Delgado J.1991. Space use by red deer in a Mediterranean ecosystem asdetermined by radio-tracking. Appl Anim Behav Sci. 30(3–4):363–371.
Castanos P. 1980. La Macrofauna de la Cueva de La Paloma. La Cueva deLa Paloma. Soto de las Regueras (Asturias), ExcavacionesArqueologicas en Espana. 116:65–100.
Castanos P. 1982. Estudio de los Macromamıferos del yacimiento deCueto de La Mina. Bol Inst Estud Asturianos. 105(106):43–86.
Castanos P. 1984. Estudio de los Macromamıferos de la cueva deSantimamine (Vizcaya). Kobie (Serie Paleoantropologıa y CienciasNaturales). 14:235–318.
Castanos P. 1986. Los Macromamıferos del Pleistoceno y Holoceno deVizcaya. Faunas asociadas a los yacimientos arqueologicos[Unpublished PhD]. Universidad del Paıs Vasco.
Castanos P. 1993. Estudio de los macromamıferos de los nivelespaleolıticos de Chaves. Bolskan: Revista de Arqueologıa delInstituto de Estudios Altoaragoneses. 10:9–30.
Castanos P. 2004. Estudio arqueozoologico de los macromamıferos delNeolıtico de la Cueva de Chaves (Huesca). Salduie: Estudios deprehistoria y arqueologıa. (4):125–172.
Conti N, Petronio C, Salari L. 2010. The Equids of the Late Pleistocene of“Tana delle Iene” (Ceglie Messapica, Brindisi, Southern Italy). BollSoc Paleont It. 49:227–236.
Cregut-Bonnoure E. 1992. Interet biostratigraphique de la morphologiedentaire de Capra (Mammalia, Bovidae). Ann Zool Fennici. 28(3–4):273–290.
Cregut-Bonnoure E. 2006. European Ovibovini, Ovini and Caprini(Caprinae, Mammalia) from the Plio-Pleistocene: new interpret-ations. Cour Forsch-Inst Senckenberg. 256:139–158.
Cruz-Uribe K. 1991. Distinguishing Hyena from Hominid BoneAccumulations. J Field Archaeol. 18(4):467–486.
Cuenca-Bescos G, Rofes J, Lopez-Garcıa JM, Blain H-A, De Marfa RJ,Galindo-Pellicena MA, Bennasar-Serra ML, Melero-Rubio M,Arsuaga JL, Bermudez De Castro JM, Carbonell E. 2010.Biochronology of Spanish Quaternary small vertebrate faunas.Quatern Int. 212(2):109–119.
Davidson I. 1989. La Economıa del final del Paleolıtico en la Espanaoriental. Serie de Trabajos Varios del SIP 85. Valencia: DescargarPublicaciones del Museo.
Davis SJ. 1980. Late Pleistocene and Holocene equid remains from Israel.Zool J Linnean Soc. 70:289–312.
Davis SJ. 2002. The mammals and birds from the Gruta do Caldeirao,Portugal. Revista Portuguesa de Arqueologia. 5(2):29–98.
Degerbøl M, Fredskild B. 1970. The urus (Bos primigenius Bojanus) andneolithic cattle (Bos taurus domesticus Linne) in Denmark. DetKongelige Danske Videnskabernes Selskab, Biologiske Skrifter.17(1):234 pp.
Delpech F. 1983. Les faunes du Paleolithique Superieur dans le Sud-Ouest de La France. Cahiers du Quaternaire 6, Editions du CentreNational de la Recherche Scientifique.
Delpech F. 1990. The range distributions of some ungulate mammalsduring the Dryas I (recent Wurm) in Aquitaine. Paleoecologicalimplications. Geobios. 23(2):221–230.
24 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Delpech F, Prat F. 1980. Les grands mammiferes du Sud-Ouest de laFrance. In Problemes de stratigraphie quaternaire en France et dansles pays limitrophes. Bulletin de l’Association franc�aise pour l’etudedu quaternaire. 1(Supplement):268–297.
Deng T, Wang X, Fortelius M, Li Q, Wang Y, Tseng ZJ, Xie G. 2011. Outof Tibet: Pliocene woolly rhino suggests high-plateau origin of IceAge megaherbivores. Science. 333(6047):1285–1288.
d’Errico F, Sanchez Goni MF. 2003. Neandertal extinction and themillennial scale climatic variability of OIS 3. Quatern Sci Rev.22(8):769–788.
d’Errico F, Vanhaeren M. 2002. Criteria for identifying red deer (Cervuselaphus) age and sex from upper canines: application to the study ofUpper Palaeolithic and Mesolithic ornaments. J Archaeol Sci.29:211–232.
Diedrich CG. 2011. Periodical use of the Balve Cave (NW Germany) as aLate Pleistocene Crocuta crocuta spelaea (Goldfuss 1823) den:hyena occupations and bone accumulations vs. human MiddlePalaeolithic activity. Quatern Int. 233(2):171–184.
Diedrich C. 2013. Late Pleistocene leopards across Europe – northern-most European German population, highest elevated records in theSwiss Alps, complete skeletons in the Bosnia Herzegowina Dinaridsand comparison to the Ice Age cave art. Quatern Sci Rev.76:167–193.
Dive J, Eisenmann V. 1991. Identification and discrimination of the firstphalanges from Pleistocene and modern Equus, wild and domestic.In: Meadow RH, Uerpmann HP, editors. Equids in the ancient world.Vol. II. Beihefte zum Tubin.
Eisenmann V. 1984. Adaptations chez des Equus sauvages actuels etfossiles. C.E.R.E.O.P.A. (Institut National Agronomique), Paris,1Oeme journee d’etude. 27–37.
Eisenmann V. 1986. Comparative osteology of modern and fossil horses,half-asses, and asses. In: Meadow RH, Uerpmann H-P, editors.Equids in the ancient world. Vol. I. Wiesbaden: Dr. Ludwig Reichert;p. 67–116.
Eisenmann V, Baryshnikov GF. 1995. Equus cf taubachensis et E.hydruntinus de la Grotte de Prolom 2 (Crimee, Ukraine) 4, 16 1994.Bulletin du Museum national d’Histoire naturelle. 2–4:329–347.
Eisenmann V, De Giuli C. 1974. Caracteres distinctifs des premieresphalanges anterieures et posterieures chez certains Equides actuels etfossils. Bulletin de la Societe geologique de France 7e serie.XVI:352–361.
Ekstrom J. 1993. The late Quaternary history of the urus (Bos primigeniusBojanus 1827) in Sweden [Lundqua Thesis]. Sweden.
Enloe JG. 2012. Middle Palaeolithic cave taphonomy: discerning humansfrom hyenas at Arcy-sur-Cure, France. Int J Osteoarchaeol.22:591–602.
Feranec R, Garcıa N, Dıez JC, Arsuaga JL. 2010. Understanding theecology of mammalian carnivorans and herbivores from Valdegobacave (Burgos, northern Spain) through stable isotope analysis.Palaeogeogr Palaeoclimatol Palaeoecol. 297(2):263–272.
Fernandez H. 2001. Osteologie comparee des petites ruminantseurasiatiques sauvages et domestiques (genres Rupicapra, Ovis,Capra et Capreolus): diagnose differentielle du squelette appendi-culaire. Universite de Geneva, Facultat de Ciencies.
Forsten A. 1999. A review of Equus stenonis Cocchi (Perissodactyla,Equidae) and related forms. Quatern Sci Rev. 18(12):1373–1408.
Freeman L. 1973. The significance of mammalian faunas frompalaeolithic occupations in Cantabrian Spain. Am Antiq. 38:3–44.
Gamble C. 1995. Large mammals, climate and resource richness in UpperPleistocene Europe. Acta Zool Cracovien. 38(1):155–175.
Garcıa-Gonzalez R. 2011. Elementos para una filogeografıa de la cabramontes iberica (Capra pyrenaica Schinz, 1838). Pirineos.166:87–122.
Garcıa-Gonzalez R. 2012. New Holocene Capra pyrenaica (Mammalia,Artiodactyla, Bovidae) skulls from the southern Pyrenees. CRPalevol. 11:241–249.
Garrido G. 2008. Generalidades sobre los artiodactilos del Villafran-quiense superior en relacion con el registro fosil de Fonelas P-1.Cuadernos del Museo Geominero. 10:279–335.
Gee H. 1993. The distinction between postcranial bones of Bosprimigenius Bojanus, 1827 from the British Pleistocene and thetaxonomic status of Bos and Bison. J Quat Sci. 8(1):79–92.
Gentry A, Clutton-Brock J, Groves CP. 2004. The naming of wild animalspecies and their domestic derivates. J Archaeoll Sci. 31:645–651.
Gomez Garcıa D, Martınez A, Montserrat P, Uribe-Echebarria PM. 2003.EL roble (Quercus robur L.) y otras plantas boreales en crisis en elMacizo del Moncayo (Soria-Zaragoza). Collectanea Botanica(Barcelona). 26:141–157.
Gonzalez-Samperiz P, Montes L, Utrilla P. 2003. Pollen in hyenacoprolites from Gabasa Cave (northern Spain). Rev PalaeobotPalynol. 126(1):7–15.
Gonzalez-Samperiz P, Valero-Garces BL, Carrion JS, Pena-Monne JL,Garcıa-Ruiz JM, Martı-Bono C. 2005. Glacial and Late glacialvegetation in northeastern Spain: new data and a review. Quatern Int.140:4–20.
Granados JE, Perez JM, Marquez FJ, Serrano E, Soriguer RC, Fandos P.2001. La cabra montes (Capra pyrenaica, Schinz, 1838). Galemys.13(1):3–37.
Greenacre MJ. 1984. Theory and applications of correspondence analysis.London: Academic Press.
Gromova VI. 1949. Histoire des chevaux (genre Equus) de l’AncienMonde. Trudy Paleontologicheskogo Instituta Akademja NaukSSSR. 7:1–373.
Guthrie RD. 1990. Frozen Fauna of the Mammoth Steppe: the Story ofBlue Babe.
Hammer Ø, Harper DAT, Ryan PD. 2001. PAST: PaleontologicalStatistics software package for education and data analysis.Palaeontol Electron. 4(1):9.
Hayward MW, Henschel P, O’Brien J, Hofmeyr M, Balme GA, KerleyGIH. 2006. Prey preferences of the leopard (Panthera pardus).J Zool. 270(2):1–16.
Hernandez ML, Valle J. 1989. Los pisos bioclimaticos de la vertiente N-NE del Moncayo (Aplicacion de modelos de analisis). Tarazona:Institucion Fernando el Catolico, C.S.I.C.–C.E.T.
Hill MO, Gauch HG, Jr. 1980. Detrended correspondence analysis:An improved ordination technique. Vegetatio. 42:47–58.
Hillson S. 1992. Mammal bones and teeth: an introductory guide tomethods of identification. London: Institute of Archaeology,University College London.
Jongman RHG, ter Braak CJF, van Tongeren OFR. 1995. Data Analysisin community and landscape ecology. Cambridge, UK: CambridgeUniversity Press.
Kahlke RD. 1999. The history of the origin, evolution and dispersal of theLate PleistoceneMammuthus-Coelodonta faunal complex in Eurasia(large mammals). Rapid City, Fenske Companies.
Kahlke RD. 2014. The origin of Eurasian Mammoth Faunas (Mammuthus-Coelodonta Faunal Complex). Quatern Sci Rev. 96:32–49.
Legendre P, Legendre L. 1998. Numerical ecology. 2nd edn. Amsterdam:Elsevier.
Lister AM. 1986. New results on deer from Swanscombe, and thestratigraphical significance of deer in the Middle and UpperPleistocene of Europe. J Archaeol Sci. 13:319–338.
Lister AM. 1996. The morphological distinction between bones and teethof fallow deer (Dama dama) and red deer (Cervus elaphus). Int J Ost.6:119–143.
Lovari S, Scala C. 1980. Revision of Rupicapra genus L. a statisticalre-evaluation of couturier’s data on the morphometry of sixchamois subspecies. Bol Zool. 47:113–124.
Lyman RL. 1994a. Vertebrate taphonomy. Cambridge: CambridgeUniversity Press 524 pp.
Lyman RL. 1994b. Relative abundances of skeletal specimens andtaphonomic analysis of vertebrate remains. Palaios. 9:288–298.
Magniez P. 2009. Nouvelles donnees sur le genre Capra Linne, 1758(Mammalia, Bovidae) du Pleistocene superieur de la grotte Tournal(Bize-Minervois, France): Implications biochronologiques et evolu-tives. Quaternaire. 20:509–525.
Mariezkurrena K. 1983. Contribucion al conocimiento del desarrollo dela denticion y el esqueleto postcraneal de Cervus elaphus. Munibe.35:149–202.
Mariezkurrena K, Altuna J. 1983. Biometrıa y dimorfismo sexual en elesqueleto de Cervus elaphus wurmiense, postwurmiense y actual delCantabrico. Munibe. 35:203–246.
Marın-Arroyo AB. 2010. Archaeozoology in the eastern Cantabrianregion during the pleistocene/holocene transition: El Miron cave.Vol. 2. Cantabria: Universidad de Cantabria.
Historical Biology 25
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014

Mateos-Quesada P. 2011. Corzo – Capreolus capreolus. En: Enciclo-pedia Virtual de los Vertebrados Espanoles.
Mills MGL, Harvey M. 2001. African predators. Cape Town: Struik.Morales A, Martın JM. 1995. Los mamıferos de la Cueva de Nerja:
analisis de las cuadrıculas NM-80A, NM-80B, y NT-82. In: Trabajosde la Cueva de Nerja. Vol. 5. Malaga: Cave of Nerja Foundation;p. 59–159.
Morales Perez JV, Serra AS. 2009. The Quaternary fossil record of thegenus Testudo in the Iberian Peninsula: archaeological implicationsand diachronic distribution in the western Mediterranean. J ArchaeolSci. 36(5):1152–1162.
Morris P. 1972. A review of mammalian age determination methods.Mamm Rev. 2:69–104.
Morris P. 1978. The use of teeth for estimating the age of wild mammals.In: Butler PM, Joysey KA, editors. Development, function andevolution of teeth. London: Academic Press; p. 483–494.
Nascetti G, Lovari S, Lanfranchi P, Berducou C, Mattiucci S, Rossi L,Bullini L. 1985. Revision of Rupicapra genus. III. Electrophoreticstudies demonstrating species of chamois populations of the Alpsfrom those of the Apennines and Pyrenees. In: Lovari S, editor. Thebiology and management of mountain ungulates. London: CroomHelm; p. 56–62.
Nowak RM. 1999. Mammals of the world, 6th edon. Baltimore, MD:Johns Hopkins University Press.
Orlando L, Mashkour M, Burke A, Douady CJ, Eisenmann V, Hanni C.2006. Geographic distribution of an extinct equid (Equushydruntinus: Mammalia, Equidae) revealed by morphological andgenetical analyses of fossils. Mol Ecol. 15:2083–2093.
Orlando L, Metcalf JL, Alberdi MT, Telles-Antunes M, Bonjean D, OtteM, Cooper A. 2009. Revising the recent evolutionary history ofequids using ancient DNA. Proc Natl Acad Sci USA.106(51):21754–21759.
Pailhaugue N. 1995. La faune de la salle Monique, Grotte de la Vache(Alliat, Ariege). Bulletin de la Societe prehistorique de l’Ariege.50:225–289.
Pales L, Lambert C. 1971. Atlas osteologique pour servir a laidentification des mammiferes du quaternaire. Bordeaux: CNRS.
Perez T, Albornoz J, Domınguez A. 2002. Phylogeography of chamois(Rupicapra spp.) inferred from microsatellites. Mol PhylogenetEvol. 25:524–534.
Perez T, Hammer SE, Albornoz J, Domınguez A. 2011. Y-chromosomephylogeny in the evolutionary net of chamois (genus Rupicapra).BMC Evol Biol. 11:272.
Perez-Barberıa FJ. 1994. Determination of age in Cantabrian chamois(Rupicapra pyrenaica parva) from jaw tooth-row eruption and wear.J Zool. 233:649–656.
Perez-Barberıa FJ, Garcıa-Gonzalez R, Palacios B. 2010. Rebeco:Rupicapra pyrenaica. In: Salvador A, Cassinello J, editors.Enciclopedia Virtual de los Vertebrados Espanoles. Madrid: MuseoNacional de Ciencias Naturales.
Perez Legido B, Cerdeno E. 1992. Los macromamıferos del Pleistocenosuperior de Cueva Millan (Burgos). Estudios Geologicos.48(3–4):187–192.
Perez-Ripoll M. 1977. Los mamıferos del yacimiento musteriense deCova Negra (Jativa, Valencia). Valencia: Diputacion de Valencia.
Perez-Ripoll M. 1988. Estudio de la secuencia del desgaste de los molaresde la Capra pyrenaica de yacimientos prehistoricos. Archivo dePrehistoria Levantina. XVIII:83–127.
Pidancier N, Jordan S, Luikart G, Taberlet P. 2006. Evolutionary historyof the genus Capra (Mammalia, Artiodactyla): Discordance betweenmitochondrial DNA and Y-chromosome phylogenies. Mol Phylo-genet Evol. 40:739–749.
Prat F. 1957. Sur la discrimination des phalanges anterieures etposterieures d’Equides. Proces verbaux de la Societe linneenne deBordeaux. XCVII:22–25.
Prat F. 1966. In: Lavocat R, Faunes er Flores Prehistoriques, editors.Boubee, 489 pp. 67 fig., 137 pI. Paris.
Quesada CN, von Lettow-Vorbeck CL. 1992. La zoologıa historica comocomplemento de la arqueozoologıa. El caso del zebro. Archaeofauna.1:61–71.
Regalia E. 1907. Sull’Equus (Asinus) hydruntinus Regalia della grotta diRomanelli (Castro, Lecce). Archivio per l’antropologia e l’etnologia.37:375–390.
Rodrıguez F, Perez T, Hammer SE, Albornoz J, Domınguez A. 2010.Integrating phylogeographic patterns of microsatellite and mtDNAdivergence to infer the evolutionary history of chamois (genusRupicapra). BMC Evol Biol. 10:222.
Sala NMT, Arsuaga JL, Laplana C, Zapata BMa, Gil Garcıa J, Garcıa N,Aranburu A, Algaba M. 2011. Un paisaje de la Meseta durante elPleistoceno Superior. Aspectos paleontologicos de la Cueva de laZarzamora (Segovia, Espana). Boletın Real Sociedad EspanolaHistorıa Natural Seccion Geologıa. 105(1–4):67–85.
Sala N, Pantoja A, Arsuaga JL, Algaba M. 2010. Presencia de bisonte(Bison priscus Bojanus 1827) y uro (Bos primigenius Bojanus 1827)en la Cueva del Buho y de la Zarzamora (Segovia, Espana). Munibe.61:43–55.
Sanchez B. 1990. Analisis de la fauna demamıferos del Pleistoceno superiordel Abric Romanı (Capellades, Barcelona). Actas de Paleontologıa.Univ. de Salamanca. Acta Salmanticensia. 68:331–347.
Sauque V, Cuenca-Bescos G. 2013. The Iberian Peninsula, the lastEuropean refugium of Panthera pardus Linnaeus 1758 during theUpper Pleistocene. Quaternaire. 24(1):35–48.
Sauque V, Rabal-Garces R, Sola-Almagro C, Cuenca-Bescos G. 2014.Bone accumulation by leopards in the Late Pleistocene in theMoncayo massif (Zaragoza, NE Spain). PLoS One. 9(3):e92144.
Schmidt I, Bradtmoller M, Kehl M, Pastoors A, Tafelmaier Y, WeningerB, Weniger GC. 2012. Rapid climate change and variability ofsettlement patterns in Iberia during the Late Pleistocene. Quatern Int.274:179–204.
Sempere AJ, Sokolov VE, Donilkin AA. 1996. Capreolus capreolus.Mamm Spec. 538:1–9.
Stampfli HR. 1963. Wisent, Bison bonasus (Linne) 1758, Ur, Bosprimigenius Bojanus, 1827, und Hausrind, Bos taurus (Linne), 1758.Acta Bernensia, Beitrage zur prahistorischen, klassischen undjungeren Archaologie. 2:117–196.
Stehlin HG, Graziosi P. 1935. Ricerche sugli Asinidi fossili d’Europa.Memoires de la Societe Paleontologique Suisse. 56:1–73.
Stewart JR. 2007. Neanderthal extinction as part of the faunal change inEurope during Oxygen Isotope Stage 3. Acta Zoologica Cracov-iensia-Series A: Vertebrata. 50(1–2):1–2.
Stiner MC. 1991. Food procurement and transport by human and non-human predators. J Archaeol Sci. 18:455–482.
Straus LG. 1987. Upper Paleolithic ibex hunting in southwest Europe.J Archaeol Sci. 14(2):163–178.
Tome C, Vigne J. 2003. Roe deer (Capreolus capreolus) age at deathestimates: New methods and modern reference data for tooth eruptionand wear, and for epiphyseal fusion. Achaeofauna. 12:157–173.
Urena I, Arsuaga JL, Galindo-Pellicena MA, Gotherstrom A, ValdioseraC. 2011. Filogenia y evolucion local de la cabra montes (Caprapyrenaica) en el yacimiento Cuaternario de Chaves (Huesca,Espana). Bol R Soc Esp Hist Nat Sec Geol. 105:5–14.
Uribe-Echebarrıa PM. 2002. Vegetacion e itinerarios botanicos en elParque Natural del Moncayo. Zaragoza: Consejo de Proteccion de laNaturaleza de Aragon 143 pp.
Vigal CR, Machordom A. 1985. Tooth eruption and replacement in theSpanish wild goat. Acta Theriol. 30:305–320.
von den Driesch A. 1976. A guide to the measurement of animal bonesfrom archaeological sites. Harvard: Peabody Museum; p. 1–136.
West RG. 2000. Plant life of the Quaternary cold stages: Evidence from theBritish Isles. Vol. 1. Cambridge: Cambridge University Press 320 pp.
Yravedra J. 2006. Acumulaciones biologicas en yacimientos arqueolo-gicos: Amalda VII y Esquilleu III-IV. Trabajos de Prehistoria. 62(2):55–78.
Yravedra J. 2010. Zooarqueologıa y tafonomıa del yacimiento de Hornosde la Pena (San Felices de Buelna, Cantabria). Complutum. 21(1):69–86.
26 V. Sauque et al.
Dow
nloa
ded
by [
Red
de
Bib
liote
cas
del C
SIC
] at
11:
04 0
5 Se
ptem
ber
2014
Top Related
Copyright © 2022 FDOKUMEN

