
Volatile Dimethyl Polonium Produced by Aerobic Marine Microorganisms
Upload
independentCategory
view
0download
0
Global Distribution of Halogenated Dimethyl Bipyrroles inMarine Mammal Blubber
S. Tittlemier,1* A. Borrell,2 J. Duffe,3 P. J. Duignan,4 P. Fair,5 A. Hall,6 P. Hoekstra,7 K. M. Kovacs,8 M. M. Krahn,9
M. Lebeuf,10 C. Lydersen,8 D. Muir,11 T. O’Hara,12 M. Olsson,13 J. Pranschke,3 P. Ross,14 U. Siebert,15 G. Stern,16
S. Tanabe,17 R. Norstrom1,3
1 Centre for Analytical and Environmental Chemistry, Carleton University, Ottawa, Ontario, Canada2 Parc Cientific de Barcelona, Department of Animal Biology, University of Barcelona, Barcelona, Spain3 National Wildlife Research Centre, Environment Canada, Hull, Quebec, Canada4 New Zealand Wildlife Health Centre, IVABS, Massey University, Palmerston North, New Zealand5 National Oceanic and Atmospheric Administration, Center for Coastal Environmental Health and Biomolecular Research, Charleston, South
Carolina, USA6 Sea Mammal Research Unit, University of St. Andrews, Fife, Scotland7 University of Guelph, Guelph, Ontario, Canada8 Norwegian Polar Institute, Tromso, Norway9 National Oceanic and Atmospheric Administration/National Marine Fisheries Service, Environmental Conservation Division,
Seattle, Washington, USA10 Maurice Lamontagne Institute, Department of Fisheries and Oceans, Mont Joli, Quebec, Canada11 National Water Research Institute, Environment Canada, Burlington, Ontario, Canada12 Department of Wildlife Management, North Slope Borough, Barrow, Alaska, USA13 Swedish Museum of Natural History, Stockholm, Sweden14 Institute of Ocean Sciences, Department of Fisheries and Oceans, Sydney, British Columbia, Canada15 Forschungs- und Technologiezentrum Westku¨ste, University of Kiel, Bu¨sum, Germany16 Freshwater Institute, Department of Fisheries and Oceans, Winnipeg, Manitoba, Canada17 Center for Marine Environmental Studies, Ehime University, Matsuyama, Japan
Received: 13 October 2001/Accepted: 12 February 2002
Abstract. Four halogenated dimethyl bipyrroles (HDBPs), hy-pothesized to be naturally produced, were quantitated in marinemammal blubber from a number of species obtained fromvarious locations worldwide. HDBPs were found in samplesfrom all locations studied. Concentrations of total HDBPs(�HDBPs) ranged from 0.4 ng/g lipid weight in ringed seals(Phoca hispida) from the White Sea to 2,540 ng/g lipid weightin Dall’s porpoise (Phocoenoides dalli) from the northwesternNorth Pacific Ocean. At their highest levels,�HDBPs made up11% of the total quantitated organohalogen body burden ofadult male Dall’s porpoises. In two beluga (Delphinapterusleucas) data subsets, it was found that males contained signif-icantly higher concentrations of�HDBPs than females. Nosignificant effects of age or sex on�HDBPs were observed inharbor seal (Phoca vitulina) and bowhead whale (Balaenamysticetus) data subsets. The geographical distribution of con-centrations did not resemble that of the ubiquitous anthropo-genic organohalogen, polychlorinated biphenyl congener CB-
153. Higher concentrations of HDBPs and different patterns ofcongeners were observed in samples from Pacific as opposed tonon–Pacific Ocean influenced environments. Concentrations ofHDBPs in beluga from the Arctic and St. Lawrence River weresimilar. Their high abundance in north Pacific Ocean biota andwidespread occurrence suggest that HDBPs undergo extensivetransport from a source located primarily in the Pacific Ocean.Evidence from HDBP congener patterns indicates that bothocean currents and atmospheric transport likely play a role inthe movement of HDBPs. These results imply that HDBPs andanthropogenic organohalogens have different sources and sup-port the natural production hypothesis.
Halogenated dimethyl bipyrroles (HDBPs) are a family ofmixed halogenated nitrogen– containing heterocycles pro-posed to have a natural origin. The molecular structures ofthe four environmentally relevant HDBPs have recentlybeen determined (Tittlemieret al. 2002a) and are shown inFigure 1. The fundamental molecular structure of the com-pounds is a 1,1�-dimethyl-2,2�-bipyrrole moiety in whichthe rings are fully substituted with bromine and chlorine.This structure resembles that of a known marine bacterium
*Present address: Food Research Division, Health Canada, Ottawa,Ontario, Canada
Correspondence to: R. Norstrom;email: [email protected]
Arch. Environ. Contam. Toxicol. 43, 244–255 (2002)DOI: 10.1007/s00244-002-1175-6
A R C H I V E S O F
EnvironmentalContaminationa n d Toxicology© 2002 Springer-Verlag New York Inc.

product, hexabromo-2,2�-bipyrrole (Andersen et al. 1974)and suggests that HDBPs may also be naturally produced.There are numerous examples of naturally produced halo-genated pyrroles containing bromine, chlorine, or a combi-nation of these two halogens (Gribble 1996). The largenumber of naturally produced halogenated pyrroles is in partdue to the electronic configuration that makes pyrroles sus-ceptible to rapid electrophilic substitution (Linda andMarino 1968). In addition, other evidence, such as theirapparent absence in high-trophic-level biota from the highlyindustrialized Great Lakes region (Tittlemier et al. 1999),supports the hypothesis that HDBPs have a biogenic ratherthan anthropogenic origin.
The distribution of HDBPs in the environment is not wellknown because only two studies concerning their occurrencehave been completed (Tittlemier et al. 1999, 2002b). The initialstudy described the levels of the most abundant congener,1,1�-dimethyl-3,3�,4,4�-tetrabromo-5,5�-dichloro-2,2�-bipyr-role (DBP-Br4Cl2), in seabird eggs obtained from the GreatLakes, Pacific, and Atlantic coastal regions of Canada. DBP-Br4Cl2 was found at the highest concentrations in samples fromthe Pacific coast, followed by those from the Atlantic coast, andwas not observed in Great Lakes samples (Tittlemier et al.1999). All four HDBP congeners shown in Figure 1 were foundin a marine food web from the Canadian high Arctic consistingof three species of zooplankton (Calanus hyperboreus, Mysisoculata, and Sagitta sp.), Arctic cod (Boreogadus saida), fourspecies of seabirds (dovekie [Alle alle], black guillemot [Cep-phus grylle], black-legged kittiwake [Rissa tridactyla], andglaucous gull [Larus hyperboreus]), and ringed seal (Phocahispida) (Tittlemier et al. 2002b). The results of these studiesindicated that HDBPs are persistent, bioaccumulative, and
widely distributed throughout the marine environment of NorthAmerica.
Marine mammals can be good indicators of the distributionof persistent organohalogen compounds in marine ecosystemsbecause they tend to accumulate relatively high levels of hy-drophobic organohalogens. Many pinnipeds and odontocetesoccupy the upper levels of marine food webs and thus accu-mulate relatively high levels of organohalogens (Muir et al.1988; Nakata et al. 1997; Tanabe et al. 1997). It has also beenshown that pinnipeds and small cetaceans generally have lowercapabilities than birds and terrestrial mammals to metabolizeorganohalogens, such as polychlorinated biphenyls (PCBs) un-substituted by chlorine in adjacent meta-para position, which isconsistent with low or absent inducible CYP2B enzyme activ-ity in marine mammals (Tanabe et al. 1988; Boon et al. 1989).In general, organohalogens are less likely to undergo metabo-lism and subsequent excretion in marine mammals than interrestrial mammals and birds.
In addition, marine mammal blubber is a good samplematrix for the examination of the widespread distribution ofpersistent organohalogens, such as HDBPs, because marinemammals inhabit virtually every ocean and sea, as well assome land-locked lakes, such as Lake Baikal. In this work,the distribution of the four identified HDBP congeners in 20species of marine mammals from 27 different locations wasstudied. Levels of the four HDBP congeners were alsonormalized to concentrations of the persistent PCB congener2,2�,4,�,5,5�-hexachlorobiphenyl (CB-153), one of the mostubiquitous anthropogenic organohalogen compounds. Com-parison of this ratio facilitated comparison among speciesand geographical locations and provided additional evidenceabout the nature of the source.
Fig. 1. Structures of halogenateddimethyl bipyrroles (HDBPs)
HDBPs in Marine Mammal Blubber 245

Materials and Methods
Organohalogen Analyses
Blubber samples from a variety of cetacean and pinniped species wereobtained from the locations shown in Figure 2. Detailed informationconcerning sampling locations, sample size, age class, and sex issummarized in Table 1.
Because the samples were supplied by various research groups, themethod of organohalogen extraction varied among the samples. Thespecific method utilized for each sample is indicated in Table 1. Themajority of samples underwent column extraction (method A). In thismethod, accurately weighed amounts of blubber (about 2 g) were cutinto small pieces and mixed with a 10-fold amount of activatedNa2SO4. The mixtures were then spiked with an internal standardsolution of 13C-labeled organochlorines (13C6-pentachlorobenzene,13C6-hexachlorobenzene, 13C12-CB118, 13C12-CB153, 13C12-CB180,13C12-CB194). The mixtures were wet-packed with dichloromethane/hexane (1:1) into a glass column (2 cm ID), and after 45 min extractedwith 250 ml dichloromethane/hexane (1:1).
Method B involved accelerated solvent extraction of accuratelyweighed blubber samples (about 5 g) using dichloromethane/hexane(1:1) and toluene. The extracts from the two different solvents werecombined, reduced to 10 ml, and spiked with the internal standard usedin method A.
Method C also used accelerated solvent extraction. Accurateamounts of blubber (about 2 g) were extracted using dichloromethane.The extracts were initially reduced to approximately 20 ml and driedover activated Na2SO4. The Na2SO4 was removed by filtration andwashed with hexane. The hexane washings were combined with theextracts, spiked with the internal standard, and reduced to 10 ml.
In method D accurately weighed blubber samples (about 5 g) weremixed with activated Na2SO4 and Soxhlet-extracted with hexane. Theextracts were reduced to 10 ml and spiked with the internal standard.
Method E also utilized Soxhlet extraction. As with the other meth-ods, an accurately weighed amount of blubber (about 4 g) was mixedwith activated Na2SO4. The mixture was Soxhlet extracted with chlo-roform/hexane (1:1), reduced to 10 ml, and spiked with the internalstandard.
The final extraction method, method F, followed the techniqueoutlined in Muir et al. (1988) with the following modifications: blub-ber samples were combined with dichloromethane and extracted usinga Polytron mixer. After extraction, lipid determination by gravimetricmethods, and removal of lipids, organohalogens were separated intotwo fractions on a silica gel (100% activated) column. Fraction 1 waseluted with hexane and fraction 2 with dichloromethane/hexane (1:1).Fraction 2, containing the HDBPs, was reduced in volume and spikedwith volume corrector (13C12-CB 138) prior to analysis.
After extraction, all samples—aside from the bowhead whales,which were worked up according to method F—were subjected to thesame procedure. The sample extracts were first filtered using pre-cleaned PTFE syringe filters. Next, the lipid content of the sampleswas determined gravimetrically. Lipids were then removed by gelpermeation chromatography (GPC) (Letcher et al. 1995). The GPCeluates were reduced to approximately 1 ml and applied to a Florisilcolumn (8 g, 1.2% deactivated). Organohalogen compounds wereeluted using 110 ml dichloromethane/hexane (1:1). The Florisil col-umn eluate containing the organohalogens was reduced to 500 �l andspiked with volume corrector (13C12-CB 138) prior to analysis. A totalof 25 method blanks containing no sample were used. One blank wascarried through all steps of the sample preparation and analysis witheach set of samples.
The samples and blanks were analyzed for PCB congener 153(CB-153) using gas chromatography electron impact ionization in thesingle ion monitoring mode using the method outlined by Sandau et al.
(2000). An external standard was used to quantitate CB-153. In addi-tion, other organochlorines were quantitated in selected samples con-taining relatively high levels of HDBPs to examine the relative con-centrations of HDBPs and commonly analyzed organochlorinescompounds. Dall’s porpoise samples were analyzed for total PCBs(taken as the sum of congeners corresponding to those present in anAroclor mixture external standard), plus the following organochlorinecompounds: 1,2,4,5-tetrachlorobenzene, 1,2,3,4-tetrachlorobenzene,pentachlorobenzene, hexachlorobenzene, �-HCH, �-HCH, �-HCH,octachlorostyrene, heptachlor epoxide, oxychlordane, trans/cis-chlor-dane, trans/cis-nonachlor, p,p�-DDD/DDE/DDT, dieldrin, photomirex,mirex, and tris-(4-chlorophenyl)-methanol. An external standard wasused to quantitate the 21 organochlorine compounds in the selected ionmonitoring electron impact ionization mode.
HDBPs were analyzed using the method described in Tittlemier etal. (2002b) using gas chromatography electron-capture negative-ion-ization mass spectrometry in the single ion monitoring mode. Quan-titation was performed using synthesized 1,1�-dimethyl-3,3�,4-tri-bromo-4�,5,5�-trichloro-2,2�-bipyrrole (DBP-Br3Cl3), 1,1�-dimethyl-3,3�,4,4�-tetrabromo-5,5�-dichloro-2,2�-bipyrrole (DBP-Br4Cl2), 1,1�-dimethyl-3,3�,4,4�,5-pentabromo-5�-chloro-2,2�-bipyrrole (DBP-Br5Cl),and 1,1�-dimethyl-3,3�,4,4�,5,5�-hexabromo-2,2�-bipyrrole (DBP-Br6) asexternal standards (Tittlemier et al. 2002a).
Statistical Analyses
Statistical analyses were performed using Statistica (v 5.5) or SAS (v.8). SAS GLM procedure was used for all analyses of variance (ANO-VAs) and the model was considered significant if p � 0.05. Concen-tration data were lipid normalized because the analytes all have rela-tively high Kows (i.e., Kow � 6) (Tittlemier et al. 2002c) and wouldtherefore be associated mainly with lipid phases. Log transformationof both raw concentration data and ratio data was required to satisfyassumptions of normality and homoscedasticity. Specific geographic–species combinations that had n 1 had to be discarded from furtheranalysis due to nonreplication; this included grey and killer whalesfrom British Columbia, Canada; hump-backed dolphin from India;Steller sea lion from Japan and California; and Risso’s dolphin fromJapan.
In previous studies of organohalogens in marine mammals (Muir etal. 1988), age and sex have been shown to affect concentrations foundin blubber. Data subsets containing samples from only adult maleswere assembled as much as possible to limit variability in the datafrom these possible confounding factors. To avoid discarding samples,the hypothesis that HDBPs would vary with sex and age of the animalusing analysis of covariance (ANCOVA) or two-way ANOVA wastested using available data subsets. If no significant differences weredetected, the data were pooled for analysis of geographical patterns.However, when significant differences were encountered, they wereaccounted for before performing the geographical model.
To minimize confounding effects of feeding differences, metabolicpotential, or other ecological factors, analyses examining geographicaltrends of HDBPs were performed by grouping animals into fourgroups: beluga, dolphins and porpoises, sea lions, and seals. In addi-tion, where appropriate, animals were classified as originating fromeither a Pacific– or non–Pacific Ocean–influenced environment to testfor further geographical differences in HDBP concentrations.
Results
Table 2 lists the geometric means and ranges of �HDBP andCB-153 concentrations on a lipid weight basis and �HDBP/CB-153 ratios. Because recoveries of the 13C12-PCB internal
246 S. Tittlemier et al.

standards were 80 20%, �HDBP and CB-153 concentrationswere recovery corrected.
Effects of Age and Sex
St. Lawrence River beluga, Igloolik beluga, Wadden Sea har-bor seal, Alaskan harbor seal, and bowhead whale samplesubsets were used to test for the effects of age and sex on�HDBP and �HDBP/CB-153 concentrations. Length was usedto estimate age class for the bowhead whale samples (Georgeet al. 1999). These were the only subsets of the same speciesfrom a single location containing enough samples for validstatistical analyses.
There were no significant effects of age or sex observed onthe �HDBP and �HDBP/CB-153 values in the Wadden Seaharbor seal, Alaskan harbor seal, and bowhead whale samplesubsets. Sex had a significant effect on �HDBP concentrationsin the St. Lawrence River and Igloolik beluga sample subsets.Males had significantly higher �HDBP concentrations than didfemales (p � 0.02, p � 0.008) (Figure 3); there were nosignificant effects on �HDBP/CB-153 ratios.
Due to the lack of observed age and sex effects, the �HDBPand �HDBP/CB-153 data from male, female, adult, and im-mature seals and bowhead whales were combined and treated
as one group. Data for dolphins, porpoises, and sea lions weresimilarly combined because the majority of samples were male.Beluga �HDBP data were corrected for effects due to sex usinga general linear model.
Beluga
After correcting for sex, it was found that the Svalbard belugashad lower absolute �HDBP concentrations than the other fourbeluga groups studied (p � 0.006). In addition, there weresignificant (p � 0.001) differences in �HDBP/CB-153 ratios.Ratios were lower in samples from the St. Lawrence River andSvalbard regions than those from Igloolik and ChesterfieldInlet. Figure 4 shows the percentage contribution of eachHBDP congener to �HDBP for beluga samples.
Dolphins and Porpoises
Fraser’s and Spinner dolphins from the Mindanao Sea, Philip-pines, were treated as one group because these species havesimilar diets (Pauly et al. 1998). Absolute HDBP concentra-tions were significantly (p � 0.005) higher in the Dall’s por-poise samples from the northwestern northern Pacific Ocean
Fig. 2. Marine mammal sampling locations. Numbers refer to locations listed in Table 1
HDBPs in Marine Mammal Blubber 247
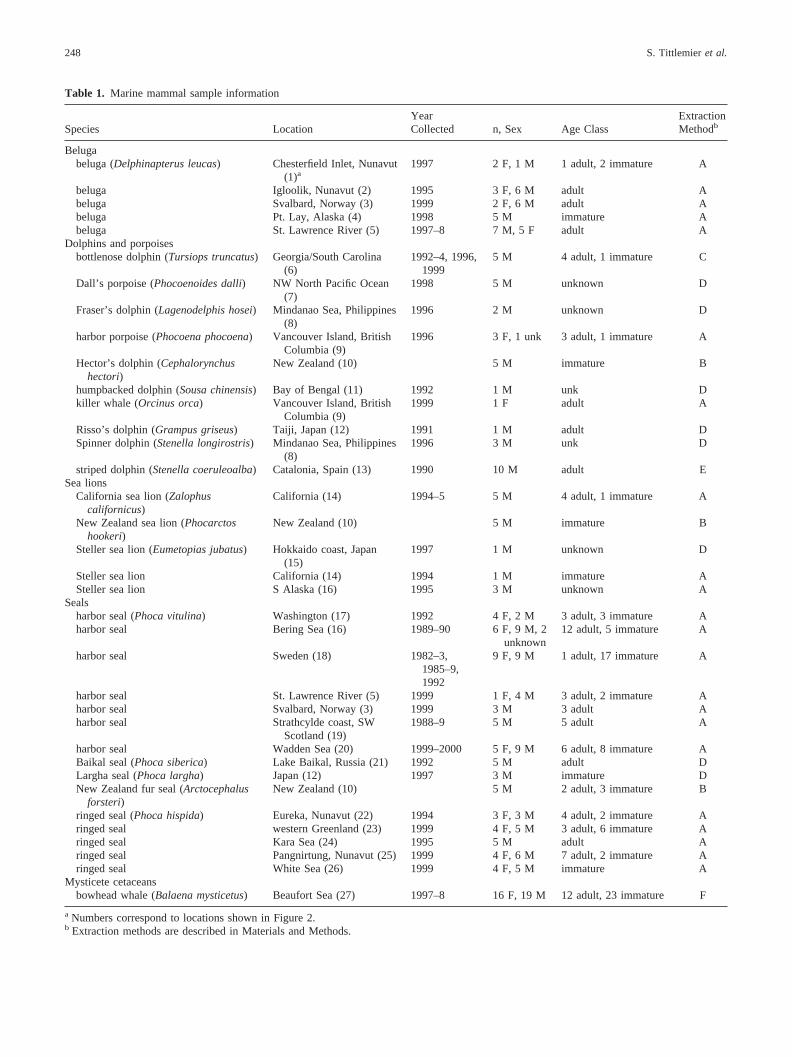
Table 1. Marine mammal sample information
Species LocationYearCollected n, Sex Age Class
ExtractionMethodb
Belugabeluga (Delphinapterus leucas) Chesterfield Inlet, Nunavut
(1)a1997 2 F, 1 M 1 adult, 2 immature A
beluga Igloolik, Nunavut (2) 1995 3 F, 6 M adult Abeluga Svalbard, Norway (3) 1999 2 F, 6 M adult Abeluga Pt. Lay, Alaska (4) 1998 5 M immature Abeluga St. Lawrence River (5) 1997–8 7 M, 5 F adult A
Dolphins and porpoisesbottlenose dolphin (Tursiops truncatus) Georgia/South Carolina
(6)1992–4, 1996,
19995 M 4 adult, 1 immature C
Dall’s porpoise (Phocoenoides dalli) NW North Pacific Ocean(7)
1998 5 M unknown D
Fraser’s dolphin (Lagenodelphis hosei) Mindanao Sea, Philippines(8)
1996 2 M unknown D
harbor porpoise (Phocoena phocoena) Vancouver Island, BritishColumbia (9)
1996 3 F, 1 unk 3 adult, 1 immature A
Hector’s dolphin (Cephalorynchushectori)
New Zealand (10) 5 M immature B
humpbacked dolphin (Sousa chinensis) Bay of Bengal (11) 1992 1 M unk Dkiller whale (Orcinus orca) Vancouver Island, British
Columbia (9)1999 1 F adult A
Risso’s dolphin (Grampus griseus) Taiji, Japan (12) 1991 1 M adult DSpinner dolphin (Stenella longirostris) Mindanao Sea, Philippines
(8)1996 3 M unk D
striped dolphin (Stenella coeruleoalba) Catalonia, Spain (13) 1990 10 M adult ESea lions
California sea lion (Zalophuscalifornicus)
California (14) 1994–5 5 M 4 adult, 1 immature A
New Zealand sea lion (Phocarctoshookeri)
New Zealand (10) 5 M immature B
Steller sea lion (Eumetopias jubatus) Hokkaido coast, Japan(15)
1997 1 M unknown D
Steller sea lion California (14) 1994 1 M immature ASteller sea lion S Alaska (16) 1995 3 M unknown A
Sealsharbor seal (Phoca vitulina) Washington (17) 1992 4 F, 2 M 3 adult, 3 immature Aharbor seal Bering Sea (16) 1989–90 6 F, 9 M, 2
unknown12 adult, 5 immature A
harbor seal Sweden (18) 1982–3,1985–9,1992
9 F, 9 M 1 adult, 17 immature A
harbor seal St. Lawrence River (5) 1999 1 F, 4 M 3 adult, 2 immature Aharbor seal Svalbard, Norway (3) 1999 3 M 3 adult Aharbor seal Strathcylde coast, SW
Scotland (19)1988–9 5 M 5 adult A
harbor seal Wadden Sea (20) 1999–2000 5 F, 9 M 6 adult, 8 immature ABaikal seal (Phoca siberica) Lake Baikal, Russia (21) 1992 5 M adult DLargha seal (Phoca largha) Japan (12) 1997 3 M immature DNew Zealand fur seal (Arctocephalus
forsteri)New Zealand (10) 5 M 2 adult, 3 immature B
ringed seal (Phoca hispida) Eureka, Nunavut (22) 1994 3 F, 3 M 4 adult, 2 immature Aringed seal western Greenland (23) 1999 4 F, 5 M 3 adult, 6 immature Aringed seal Kara Sea (24) 1995 5 M adult Aringed seal Pangnirtung, Nunavut (25) 1999 4 F, 6 M 7 adult, 2 immature Aringed seal White Sea (26) 1999 4 F, 5 M immature A
Mysticete cetaceansbowhead whale (Balaena mysticetus) Beaufort Sea (27) 1997–8 16 F, 19 M 12 adult, 23 immature F
a Numbers correspond to locations shown in Figure 2.b Extraction methods are described in Materials and Methods.
248 S. Tittlemier et al.
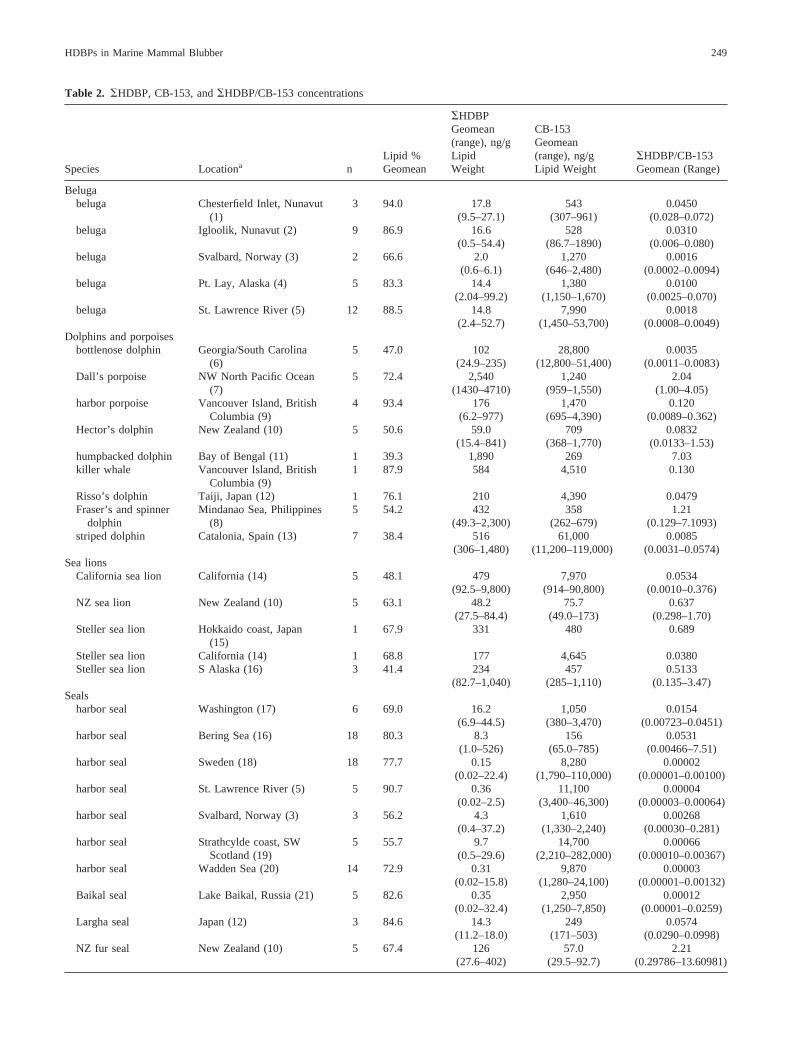
Table 2. �HDBP, CB-153, and �HDBP/CB-153 concentrations
Species Locationa nLipid %Geomean
�HDBPGeomean(range), ng/gLipidWeight
CB-153Geomean(range), ng/gLipid Weight
�HDBP/CB-153Geomean (Range)
Belugabeluga Chesterfield Inlet, Nunavut
(1)3 94.0 17.8
(9.5–27.1)543
(307–961)0.0450
(0.028–0.072)beluga Igloolik, Nunavut (2) 9 86.9 16.6
(0.5–54.4)528
(86.7–1890)0.0310
(0.006–0.080)beluga Svalbard, Norway (3) 2 66.6 2.0
(0.6–6.1)1,270
(646–2,480)0.0016
(0.0002–0.0094)beluga Pt. Lay, Alaska (4) 5 83.3 14.4
(2.04–99.2)1,380
(1,150–1,670)0.0100
(0.0025–0.070)beluga St. Lawrence River (5) 12 88.5 14.8
(2.4–52.7)7,990
(1,450–53,700)0.0018
(0.0008–0.0049)Dolphins and porpoises
bottlenose dolphin Georgia/South Carolina(6)
5 47.0 102(24.9–235)
28,800(12,800–51,400)
0.0035(0.0011–0.0083)
Dall’s porpoise NW North Pacific Ocean(7)
5 72.4 2,540(1430–4710)
1,240(959–1,550)
2.04(1.00–4.05)
harbor porpoise Vancouver Island, BritishColumbia (9)
4 93.4 176(6.2–977)
1,470(695–4,390)
0.120(0.0089–0.362)
Hector’s dolphin New Zealand (10) 5 50.6 59.0(15.4–841)
709(368–1,770)
0.0832(0.0133–1.53)
humpbacked dolphin Bay of Bengal (11) 1 39.3 1,890 269 7.03killer whale Vancouver Island, British
Columbia (9)1 87.9 584 4,510 0.130
Risso’s dolphin Taiji, Japan (12) 1 76.1 210 4,390 0.0479Fraser’s and spinner
dolphinMindanao Sea, Philippines
(8)5 54.2 432
(49.3–2,300)358
(262–679)1.21
(0.129–7.1093)striped dolphin Catalonia, Spain (13) 7 38.4 516
(306–1,480)61,000
(11,200–119,000)0.0085
(0.0031–0.0574)Sea lions
California sea lion California (14) 5 48.1 479(92.5–9,800)
7,970(914–90,800)
0.0534(0.0010–0.376)
NZ sea lion New Zealand (10) 5 63.1 48.2(27.5–84.4)
75.7(49.0–173)
0.637(0.298–1.70)
Steller sea lion Hokkaido coast, Japan(15)
1 67.9 331 480 0.689
Steller sea lion California (14) 1 68.8 177 4,645 0.0380Steller sea lion S Alaska (16) 3 41.4 234
(82.7–1,040)457
(285–1,110)0.5133
(0.135–3.47)Seals
harbor seal Washington (17) 6 69.0 16.2(6.9–44.5)
1,050(380–3,470)
0.0154(0.00723–0.0451)
harbor seal Bering Sea (16) 18 80.3 8.3 156 0.0531(1.0–526) (65.0–785) (0.00466–7.51)
harbor seal Sweden (18) 18 77.7 0.15(0.02–22.4)
8,280(1,790–110,000)
0.00002(0.00001–0.00100)
harbor seal St. Lawrence River (5) 5 90.7 0.36(0.02–2.5)
11,100(3,400–46,300)
0.00004(0.00003–0.00064)
harbor seal Svalbard, Norway (3) 3 56.2 4.3(0.4–37.2)
1,610(1,330–2,240)
0.00268(0.00030–0.281)
harbor seal Strathcylde coast, SWScotland (19)
5 55.7 9.7(0.5–29.6)
14,700(2,210–282,000)
0.00066(0.00010–0.00367)
harbor seal Wadden Sea (20) 14 72.9 0.31(0.02–15.8)
9,870(1,280–24,100)
0.00003(0.00001–0.00132)
Baikal seal Lake Baikal, Russia (21) 5 82.6 0.35(0.02–32.4)
2,950(1,250–7,850)
0.00012(0.00001–0.0259)
Largha seal Japan (12) 3 84.6 14.3(11.2–18.0)
249(171–503)
0.0574(0.0290–0.0998)
NZ fur seal New Zealand (10) 5 67.4 126(27.6–402)
57.0(29.5–92.7)
2.21(0.29786–13.60981)
HDBPs in Marine Mammal Blubber 249
than samples from the other locations. �HDBP/CB-153 valueswere significantly (p � 0.003) greater in the Dall’s porpoisefrom the North Pacific Ocean and dolphins from the Philip-pines than the other dolphin and porpoise samples. Figure 5illustrates the relative amount of HDBPs in the most contam-inated Dall’s porpoise compared to other organochlorine com-pounds.
Samples were then classified as having either a Pacific ornon-Pacific origin. There was no significant difference in�HDBP concentrations, but �HDBP/CB-153 ratios were lowerin the non-Pacific group (p � 0.01).
Sea Lions
There were no significant differences in �HDBP concentra-tions among the sea lions from the various sampling locations.In addition, �HDBP/CB-153 ratios were not significantly dif-
ferent among locations, although the ratio appeared lower inthe California samples due to higher CB-153 concentrationsthan the other areas.
Seals
Absolute �HDBP concentrations and �HDBP/CB-153 ratioswere not significantly different among seal samples from thevarious locations, although samples from New Zealand, Wash-
Fig. 3. �HDBP concentrations in male and female beluga from Ig-loolik and the St. Lawrence River
Fig. 4. HDBP congener profiles (expressed as a percentage of�HDBP) for beluga. Numbers in parentheses refer to locations shownon Figure 2
Table 2. Continued
Species Locationa n Lipid %Geomean
�HDBPGeomean(range), ng/gLipidWeight
CB-153Geomean(range), ng/gLipid Weight
�HDBP/CB-153Geomean (Range)
ringed seal Eureka, Nunavut (22) 6 89.3 1.2(0.1–46.4)
237(73.3–1,770)
0.00522(0.00137–0.0412)
ringed seal western Greenland (23) 9 89.3 0.8(0.3–1.5)
107(48.1–286)
0.00717(0.00232–0.0232)
ringed seal Kara Sea (24) 5 83.4 1.6(0.6–11.5)
235(140–376)
0.00693(0.00156–0.0355)
ringed seal Pangnirtung, Nunavut (25) 9 93.1 1.4(0.4–17.2)
109(44.2–402)
0.0125(0.00345–0.103)
ringed seal White Sea (26) 9 89.0 0.4(0.1–6.0)
367(158–1,070)
0.00115(0.00020–0.00560)
Mysticete cetaceansbowhead whale
Beaufort Sea (27)35 70.5
0.6(0.1–1.8)
29.7(9–197)
0.0200(0.00119–0.0794)
a Numbers correspond to locations shown in Figure 2.
250 S. Tittlemier et al.

ington, Japan, Alaska, and Scotland appeared higher than theremaining locations.
In addition, harbor, New Zealand fur, Baikal, and Larghaseals were separated into groups from Pacific– and non–PacificOcean–influenced areas. The samples originating from non-Pacific areas had significantly lower �HDBP and �HDBP/CB-153 levels (p � 0.00002, p � 0.000001) than samples obtainedfrom Pacific environments. Figure 6 shows the percentagecontribution of each HDBP congener to �HDBP for the samplegroups used in the Pacific versus non-Pacific comparison.
Discussion
Significance of HDBPs as Organohalogen Contaminants
HDBPs are just one of many classes of organohalogen com-pounds that have been found in marine mammals. They are,however, the first classes of persistent bioaccumulative orga-nohalogens hypothesized to have a natural origin. The presenceof another halogenated compound, also proposed to be natu-rally produced, at relatively high levels in marine mammalsamples has recently been described (Vetter et al. 2000). Thiscompound, C9H3N2Cl7, is thought to contain a methyl-bipyr-role moiety and may be related to HDBPs. The majority ofother organohalogen contaminants commonly analyzed are an-thropogenic or biotransformation products of anthropogeniccompounds.
A wide range of HDBP concentrations were observed acrossall of the sampling locations and species studied. Concentra-tions ranged from sub ng/g in Baikal seals and harbor sealsfrom the St. Lawrence River, Wadden Sea, and Sweden, to�g/g levels in Dall’s porpoise from the northwestern Pacific
Ocean. Similar to organochlorine compounds (O’Hara et al.1999), HDBPs were at lower concentrations in baleen whales(bowhead whales) compared to odontocetes (beluga) from anearby location. In addition to trophic level, metabolism mayalso affect interspecies differences in HDBP concentrations.Evidence from a food web study suggests that ringed seals mayhave the ability to metabolize HDBPs (Tittlemier et al. 2002b).Possible differential metabolic capabilities of pinnipeds andcetaceans toward HDBPs may account for the differences inHDBP concentrations observed in these groups. In all cases,though, geometric mean total HDBPs (�HDBPs) were at levelslower than total PCBs or total DDT compounds, the mostabundant organochlorine compounds, reported for the samespecies at similar locations (Tanabe et al. 1983, 1988; Kuehl etal. 1991; Blomkvist et al. 1992; Nakata et al. 1997, 1998;Prudente et al. 1997; Tilbury et al. 1999; Westgate and Tolley1999; Muir et al. 2000).
However, there were some instances in which HDBPs made asignificant contribution to an organism’s organohalogen bodyburden. The highest �HDBP concentrations were found in maleDall’s porpoises from the northwestern Pacific Ocean. The piechart in Figure 5 depicts the percent contribution of each contam-inant class to the amount of total organohalogens quantitated inthe Dall’s porpoise samples. HDBPs made up approximately 11%of the total mass of quantitated organohalogens and were thefourth largest contributor after PCBs (39%), DDT compounds(27%), and chlordanes (15%). For the remaining marine mammalsamples, HDBPs were less than 3% of the total quantitated orga-nohalogens and did not constitute a large portion of the organo-halogen body burdens. It is likely that the actual percent contri-bution of HDBPs to the total organohalogen body burden is lessthan calculated here, because other organohalogens (principallytoxaphene) were not analyzed.
Fig. 5. Relative contribution (byweight) of HDBPs to the total quan-titated organohalogen body burdenfor adult male Dall’s porpoises fromthe northwestern North PacificOcean. �PCBs includes 92 conge-ners; �DDT includes p,p�-DDT, p,p�-DDE, and p,p�-DDD; �CHL includestrans/cis-chlordane, trans/cis-nona-chlor, and oxychlordane; �HCHsincludes �/�/�-HCH, �CIBZs in-cludes 1,2,4,5-tetrachlorobenzene,1,2,3,4-tetrachlorobenzene, pentachlo-robenzene, and hexachlorobenzene;�other includes dieldrin, mirex, pho-tomirex, tris-(4-chlorophenyl)-metha-nol, and octachlorostyrene
HDBPs in Marine Mammal Blubber 251
Variation of �HDBP Concentrations with Age and Sex
The majority of sample groups were of inadequate size andcomposition to confidently test for relationships betweenHDBP concentrations, age, and sex. Belugas of the St. Law-rence River and Igloolik were the only cases for which signif-icant relationships between �HDBP concentrations and sexwere observed. The mean concentration in males was approx-imately five times higher than those in females. There was asimilar difference in CB-153 concentrations between males andfemales in the same samples. This difference has been ob-served on other occasions with many persistent organochlorinecompounds, such as DDT and PCBs (Stern et al. 1994) and isattributed to lactational and placental transfer of contaminantsin females to offspring.
Geographical Distribution
The general variation of �HDBP concentration within cetaceanand pinniped groups appears to be governed strongly by onefactor—location. There is a clear distinction between samplesobtained from Pacific and non-Pacific locations in the NorthernHemisphere and tropics. There are likely confounding factorspresent, such as species- and area-specific differences in feed-ing ecology, biotransformation capabilities, sample condition,and interindividual variability, which contribute to variations in
HDBP concentrations. Nevertheless, samples originating froma Pacific environment contained significantly higher concen-trations of HDBPs than the same or similar species from otherlocations. The highest �HDBP concentrations among all spe-cies and sites were found in Dall’s porpoise samples obtainedfrom northwestern Pacific Ocean environments. This geo-graphical difference has been noted previously for HDBPs inseabird eggs (Tittlemier et al. 1999).
A differentiation in �HDBP levels in Pacific and non-Pacificsamples is observed in the seal and dolphin/porpoise subsets,the only two groups of species with an adequate range ofsampling locations to allow for such a comparison. The dis-tinction between concentrations in Pacific and non-Pacific sealblubber is clear but is less apparent with the dolphins andporpoises. Striped dolphin blubber from the Mediterranean Seacontained relatively high concentrations of HDBPs comparedto the other European samples. When these concentrations arenormalized to CB-153 and compared to the other �HDBP/CB-153 ratios, it is clear that the Mediterranean samples weremuch more contaminated with organohalogens overall. Thesesamples were collected during the 1990 Mediterraneanepizootic and were previously shown to contain high levels ofPCBs when compared to healthy striped dolphins sampledbefore and after the epizootic (Aguilar and Borrell 1994). Thehigh HDBP levels may therefore be due to mobilization ofblubber reserves and subsequent inability to metabolize orexcrete compounds at a comparable rate, rather than a higherlevel of HDBPs in the Mediterranean environment. This se-quence of events has been proposed to explain differences inconcentrations of PCBs and DDT compounds in grey seals ofvarying nutritional condition from Sweden and Scotland(Blomkvist et al. 1992).
One of the most abundant and widespread anthropogenicorganohalogen compounds in the environment is CB-153, ahexachlorinated PCB congener. CB-153 is not readily biotrans-formed and bioaccumulates in biota. The water solubility,vapor pressure, and octanol/water partition coefficient (Kow) ofCB-153 are similar to those estimated for HDBPs (Sw 1 �10�5 to 3 � 10�5 g/L; P°L,25 6 � 10�6 to 2 � 10�4; Kow 6.4 to 6.7), indicating that CB-153 is a good marker of therelative distribution of semivolatile anthropogenic and naturalorganohalogen compounds (Tittlemier 2001). However, it mustbe noted that HDBPs are probably not as persistent in pinni-peds as CB-153 (Tittlemier 2001).
The geographical distribution of �HDBP concentrationsdoes not resemble the distribution patterns of some anthropo-genic organohalogens, such as CB-153, and PCBs in general. Atendency for levels of PCBs in Pacific to be higher than innon-Pacific organisms has not been observed. Studies monitor-ing these organohalogens in seals and marine fish have indi-cated that the highest levels of contaminants are in samplesfrom the Atlantic Ocean, North, and Baltic Seas, followed bythe Northwest Pacific Ocean, Arctic Ocean, and finally theAntarctic Ocean, as discussed in Muir et al. (1988).
In contrast to PCBs, HDBPs appeared to be distributeduniformly in beluga sampled from Arctic regions and the St.Lawrence River. The relatively uniform distribution of HDBPsis different from what was observed for CB-153 (Table 2) andtotal PCBs in beluga from western Greenland and the St.Lawrence River. The levels of CB-153 in St. Lawrence belugaranged from 5 to 15 times greater than those from the other
Fig. 6. HDBP congener profiles (expressed as a percentage of�HDBP) for seal samples excluding ringed seals. Numbers in paren-theses refer to locations shown in Figure 2
252 S. Tittlemier et al.

sites. St. Lawrence beluga also contained approximately 15times greater amounts of total PCBs than those from westernGreenland (Stern et al. 1994; Muir et al. 1996). In contrast,�HDBP concentrations were essentially identical in the Ches-terfield Inlet, Igloolik, Pt. Lay, and St. Lawrence River beluga;the Svalbard group contained lower levels.
There was also no consistent difference between concentra-tions of HDBPs in biota from the Northern Hemisphere andNew Zealand, the sole sampling site in the Southern Hemi-sphere. Hector’s dolphin and New Zealand sea lion samplescontained less HDBPs than ecologically similar species in theNorthern Hemisphere, but New Zealand fur seals containedsignificantly higher levels than did northern seal species. Thefew available studies concerning anthropogenic organohalo-gens in Southern Hemisphere marine mammals conclude thatlevels are lower than those found in the Northern Hemisphere(Bacon et al. 1992; Kemper et al. 1994; Connell et al. 1999).However, a recent study by Vetter et al. (2000) found thatlevels of Q1, an organohalogen hypothesized to be of biogenicorigin, were higher in seal blubber samples from the Antarcticcompared to those sampled from the Arctic.
CB-153 concentrations and ratios of �HDBP/CB-153 con-firm that HDBPs have a different distribution than PCBs. Thedata in Table 2 illustrate that samples with high levels ofHDBPs do not necessarily contain correspondingly high levelsof CB-153. The geographical distribution of CB-153 generallyfollows the distribution of industrialization and other humanactivity. The samples that contain the highest levels of CB-153within each group are from sites near intense human activity,such as the St. Lawrence River, the Mediterranean Sea, and theSoutheastern U.S. coast. Yet these same samples do not havethe highest �HDBP concentrations.
Similar physical properties of CB-153 and HDBPs suggestthat these compounds may have comparable capacities to movethroughout the environment. However, the potential of thesecompounds to accumulate in marine mammals is likely differ-ent in pinnipeds due to their ability to metabolize HDBPs(Tittlemier et al. 2002b). The relatively high �HDBP concen-trations observed in cetacean blubber samples combined withthe proposed low metabolic capabilities of cetaceans towardnonplanar organohalogens may suggest that cetaceans do notmetabolize HDBPs or do not metabolize them to the sameextent as pinnipeds (Tanabe et al. 1988; Boon et al. 1992,1994). If the potential to move and accumulate in a food webis similar between CB-153 and HDBPs, as is likely the casewith cetaceans, the difference in their distribution patterns maybe attributed to different sources and exposures. Sources ofPCBs are terrestrial and distributed in industrialized urbanareas. However, HDBPs appear to be formed primarily in thenorth Pacific Ocean. This provides more evidence to supportthe hypothesis that HDBPs are naturally produced.
Transport of HDBPs
It is very likely that the source of HDBPs exists predominantlyin the northern Pacific Ocean because the highest �HDBPlevels have been observed in biota from this area. If the HDBPsare produced mainly in this area, transport mechanisms must beat work, or else there would not be as wide a distribution of
HDBPs as was observed in marine mammals. Two possiblemechanisms regulating HDBP distribution are atmospheric andoceanic current transport.
Atmospheric transport of organohalogens of marine origin isgoverned by Henry’s law constant, which describes the air–water partitioning behavior of a substance (Wania and Mackay1993). Compounds with high Henry’s law constants will tendto volatilize from aqueous solution to air, whereas those withlow constants will tend to remain in solution. The Henry’s lawconstants of HDBPs have been estimated from the ratio ofvapor pressure to aqueous solubility and are on the order of0.005 Pa m3/mol (Tittlemier et al. 2002c). These values placeHDBPs in the same range as octa- to decachlorinated PCBcongeners and thus are not likely to undergo extensive atmo-spheric transport (Mackay et al. 1999). In a previous study,HDBPs were not detected in herring gull egg samples from theGreat Lakes, supporting the hypothesis that they are not atmo-spherically transported to a great extent (Tittlemier et al. 1999).
Evidence for extensive atmospheric transport does not existin the HDBP congener patterns observed in the samples. In-creasing proportions of more volatile congeners with an in-crease in latitude has been described as being indicative ofatmospheric transport of low volatility compounds. For exam-ple, tri-, tetra-, and pentachlorinated PCB congeners increasedin proportion to higher chlorinated congeners in burbot (Lotalota) obtained from remote Canadian lakes and rivers at in-creasing latitudes (Muir et al. 1990). When HDBP congenerpatterns in harbor, Largha, Baikal, and New Zealand fur sealsfrom locations at various latitudes are compared, no suchincrease in the more volatile congeners is observed (Figure 6).In fact, the relative amounts of the least volatile congener,DBP-Br6, are higher in samples from Alaska and Svalbard, thetwo most northern latitudes. The same results are found whenHDBP congener patterns in beluga are considered (Figure 4).Aside from the samples obtained from Pt. Lay, the locationclosest to the Pacific Ocean, the predominant congener is theleast volatile DBP-Br6.
It does appear, however, that limited atmospheric transportof HDBPs occurs, because HDBPs were found at low concen-trations in Baikal seals obtained from the freshwater environ-ment of Lake Baikal, Russia. The pattern of HDBP congenerswas similar to Pacific Ocean marine samples, and unlike thatfound in the North Atlantic Ocean and adjacent seas. Previ-ously, HDBPs were not detected in seabird eggs from thefreshwater environment of the Great Lakes, thereby indicatingthat they were present at much higher levels in marine envi-ronments (Tittlemier et al. 1999). The difference between thepresence of HDBPs in the freshwater environment of LakeBaikal and its apparent absence in the Great Lakes may beexplained by average surface wind flows. January and Julyaverage winds flow in an easterly direction over a considerablylarge distance across the North American land mass beforereaching the Great Lakes. On the other hand, July averagewinds flowing toward Lake Baikal originate over the north-eastern Pacific Ocean and cross a shorter section of land mass(Hasse and Dobson 1986). The low-vapor-pressure HDBPswould be more easily transported a shorter distance over landto Lake Baikal than the Great Lakes before removal from theair masses by deposition.
Overall, the evidence does not clearly indicate atmospherictransport as the major mode of HDBP global distribution.
HDBPs in Marine Mammal Blubber 253

Indirect evidence for ocean current transport can be found inthe congener patterns of HDBPs from Pacific– and AtlanticOcean–influenced environments, assuming that the primarysource is in the north Pacific Ocean. Figures 4 and 6 illustratethe differences between HDBP congener patterns in beluga andharbor seal samples from these two environments. Samplesoriginating in areas influenced by the Pacific Ocean haveDBP-Br4Cl2 as the predominant congener. This was similarlyobserved for seabirds (Tittlemier et al. 1999). Samples from theNorth Atlantic, or areas where there are inputs of North Atlan-tic waters such as the eastern Canadian Arctic, have eitherDBP-Br3Cl3 or DBP-Br6 as the most abundant congeners.These differences in HDBP congener patterns are not as clearin the dolphin and porpoise samples, presumably due to thenumber of diverse species sampled. HDBP congener patternshave been shown to be related to an organism’s feeding habits,e.g., whether the food chain is benthic- or pelagic-based (Tittle-mier et al. 2002b). There is also the possibility that differencesin metabolism of HDBPs exist among the species.
One would not expect the congener patterns to be so distinctbetween and conserved within Pacific and Atlantic regions ifatmospheric transport was the only mechanism of movement.Because atmospheric transport is governed in part by a com-pound’s vapor pressure, it would be expected that the morevolatile congeners would predominate the composition of totalHDBPs in all northern locations irrespective of their source’sHDBP composition. This would tend to minimize differencesin congener patterns between Pacific- and Atlantic-originatingsamples. As illustrated in Figures 4 and 6, this clearly does notoccur. It cannot be ruled out that the congener pattern differ-ences are produced by different organisms distributed in vari-ous locations.
In reality, HDBPs are likely transported by a combination ofatmospheric and ocean current transport. Evidence indicativeof the primary mode of movement has likely been obscuredover time. If HDBPs are naturally produced, they would havebeen present in the environment for numerous centuries andhave had the time to be transported great distances by bothmechanisms even if the rate of one mode of transport is muchless than the other.
Conclusions
HBDPs are globally distributed in the marine environment.They likely undergo extensive transport by a combination ofatmospheric and ocean current mechanisms. However, the pre-dominant mode of transport cannot be determined from theavailable evidence. HDBPs appear to be primarily produced inthe northern Pacific Ocean, as indicated by relatively highconcentrations in biota from this region. In some instances inthis area, HDBPs contribute significantly to an organism’s totalorganohalogen body burden. The variation of HDBP concen-tration with location is very different from that of anthropo-genic organochlorines, such as CB-153. This difference indistribution suggests a biogenic rather than anthropogenicsource of HDBPs. The widespread distribution of HDBPs,most likely occurring as a slow process as indicated by theirlow vapor pressure and water solubility, suggests that HDBPshave a long lifetime in the marine biosphere.
Acknowledgments. The Natural Sciences and Engineering ResearchCouncil of Canada, the Canadian Chlorine Coordinating Committee (C4),and the Canadian Chemical Producers Association provided financialsupport for this work. All groups involved in sample collection, includingnative subsistence hunters from Alaska, are gratefully acknowledged fortheir contributions. Laboratory work and analyses were performed at theNational Wildlife Research Centre, Hull, Quebec.
References
Aguilar A, Borrell A (1994) Abnormally high polychlorinated biphe-nyl levels in striped dolphins (stenella coeruleoalba) affected bythe 1990–1992 Mediterranean epizootic. Sci Tot Env 154:237–247
Andersen RJ, Wolfe MS, Faulkner DJ (1974) Autotoxic antibioticproduction by a marine Chromobacterium. Mar Biol 27:281–285
Bacon CE, Jarman WM, Costa DP (1992) Organochlorine and poly-chlorinated biphenyl levels in pinniped milk from the Arctic, theAntarctic, California and Australia. Chemosphere 24:779–791
Blomkvist G, Roos A, Jensen S, Bignert A, Olsson M (1992) Con-centrations of sDDT and PCB in seals from Swedish and Scottishwaters. Ambio 21:539–545
Boon JP, Eijgenraam F, Everaarts JM (1989) A structure-activityrelationship (SAR) approach towards metabolism of PCBs inmarine animals from different trophic levels. Mar Env Res 27:159–176
Boon JP, Van Arnhem E, Jansen S, Kannan N, Petrick G, Schulz D,Duinker JC, Reijnders PJH, Goksoyr A (1992) The toxicokineticsof PCBs in marine mammals with special reference to possibleinteractions of individual congeners with the cytochrome P450-dependent monooxygenase system: an overview. In Walker CH,Livingstone DR (eds) Persistent pollutants in marine ecosystems.Pergamon Press, Oxford, 119–160
Boon J, Oosingh I, van der Meer J, Hillebrand MTO (1994) A modelfor the bioaccumulation of chlorobiphenyl congeners in marinemammals. Eur J Pharm 270:237–251
Connell DW, Miller GJ, Mortimer MR, Shaw GR, Anderson SM(1999) Persistent lipophilic contaminants and other chemical res-idues in the Southern Hemisphere. Crit Rev Env Sci Tech 29:47–82
George JC, Bada J, Zeh J, Scott L, Brown SE, O’Hara T, Suydam R(1999) Age and growth estimates of bowhead whales (Balaenamysticetus) via aspartic acid racemization. Can J Zool 77:571–580
Gribble GW (1996) Naturally occurring organohalogen com-pounds—a comprehensive survey. Prog Chem Org Nat Prod 68:1–467
Hasse L, Dobson F (1986) Introductory physics of the atmosphere andocean. Reidel, Dordrecht, Netherlands
Kemper C, Gibbs P, Obendorf D, Marvanek S, Lenghaus C (1994) Areview of heavy metal and organochlorine levels in marine mam-mals in Australia. Sci Tot Env 154:129–139
Kuehl DW, Haebler R, Potter C (1991) Chemical residues in dolphinsfrom the U.S. Atlantic Coast including Atlantic bottlenose ob-tained during the 1987/88 mass mortality. Chemosphere 22:1071–1084
Letcher RJ, Norstrom RJ, BergmanÅ(1995) An integrated analyticalmethod for determination of polychlorinated aryl methyl sulfonemetabolites and polychlorinated hydrocarbon contaminants in bi-ological matrices. Anal Chem 67:4155–4163
Linda P, Marino G (1968) Electrophilic substitution in five-memberedheterocyclic systems, Part VI: kinetics of the bromination of the2-methoxycarbonyl derivatives of furan, thiophen, and pyrrole inacetic acid solution. J Chem Soc B392
Mackay D, Shiu W-Y, Ma K-C (1999) Physical-chemical properties andenvironmental fate handbook. CRCnetBASE, Boca Raton, FL
254 S. Tittlemier et al.

Muir DCG, Ford CA, Grift NP, Metner DA, Lockhart WL (1990)Geographic variation of chlorinated hydrocarbons in burbot (Lotalota) from remote lakes and rivers in Canada. Arch EnvironContam Toxicol 19:530–542
Muir DCG, Ford CA, Rosenberg B, Norstrom RJ, Simon M, Beland P(1996) Persistent organochlorines in beluga whales (Delphi-napterus leucas) form the St. Lawrence River estuary—I. Con-centrations and patterns of specific PCBs, chlorinated pesticidesand polychlorinated dibenzo-p-dioxins and dibenzofurans. Envi-ron Pollut 93:219–234
Muir DCG, Norstrom RJ, Simon M (1988) Organochlorine contami-nants in Arctic marine food chains: accumulation of specificpolychlorinated biphenyls and chlordane-related compounds. En-viron Sci Technol 22:1071–1079
Muir DCG, Riget F, Cleeman M, Skaare J, Kleivane L, Nakata H,Dietz R, Severinsen T, Tanabe S (2000) Circumpolar trends ofPCBs and organochlorine pesticides in the Arctic marine environ-ment inferred from levels in ringed seals. Environ Sci Technol34:2431–2438
Nakata H, Tanabe S, Tatsukawa R, Amano M, Miyazaki N, Petrov E(1997) Bioaccumulation profiles of polychlorinated biphenyls in-cluding coplanar congeners and possible toxicological implica-tions in Baikal seal (Phoca sibirica). Environ Pollut 95:57–65
Nakata H, Tanabe S, Tatsukawa R, Koyama Y, Miyazaki N, BelikovS, Boltunov A (1998) Persistent organochlorine contaminants inringed seals (Phoca hispida) from the Kara Sea, Russian Arctic.Environ Toxicol Chem 17:1745–1755
O’Hara TM, Krahn MM, Boyd D, Becker PR, Philo LM (1999)Organochlorine contaminant levels in Eskimo harvested bowheadwhales of arctic Alaska. J Wildl Dis 35:741–752
Pauly D, Trites AW, Capuli E, Christensen V (1998) Diet compositionand trophic levels of marine mammals. ICES J Mar Sci 55:467–481
Prudente M, Tanabe S, Watanabe M, Subramnian A, Miyazaki N,Suarez P, Tatsukawa R (1997) Organochlorine contamination insome odontoceti species from the North Pacific and Indian Ocean.Mar Env Res 44:415–427
Sandau CD, Ayotte P, Dewailly E, Duffe J, Norstrom RJ (2000)Analysis of hydroxylated metabolites of PCBs (OH-PCBs) andother chlorinated phenolic compounds in whole blood from Ca-nadian Inuit. Environ Health Perspect 108:611–616
Stern GA, Muir DCG, Segstro MD, Dietz R, Heide-Jorgensen MP(1994) PCBs and other organochlorine contaminants in whitewhales (Delphinapterus leucas) from west Greenland: variations
with age and sex, Meddelelser om Gronland. Bioscience, 39:245–259
Tanabe S, Madhusree B, Ozturk AA, Tatsukawa R, Miyazaki N,Ozdamar E, Aral O, Samsun O, Ozturk B (1997) Persistentorganochlorine residues in harbour porpoise (Phocoena pho-coena) from the Black Sea. Mar Pollut Bull 34:338 –346
Tanabe S, Mori T, Tatsukawa R (1983) Global pollution of marinemammals by PCBs, DDTs, and HCHs (BHCs), Chemosphere12:1269–1275
Tanabe S, Watanabe S, Kan H, Tatsukawa R (1988) Capacity andmode of PCB metabolism in small cetaceans. Mar Mam Sci4:103–124
Tilbury KL, Adams NG, Krone CA, Meador JP, Early G, Varanasi U(1999) Organochlorines in stranded pilot whales (Globicephalamelaena) from the coast of Massachusetts. Arch Environ ContamToxicol 37:125–134
Tittlemier SA, Blank DH, Gribble GW, Norstrom RJ (2002a)Structure elucidation of four possible biogenic organohalogensusing isotope exchange mass spectrometry. Chemosphere, 46:511–517
Tittlemier SA, Fisk AT, Hobson KA, Norstrom RJ (2002b) Examina-tion of the bioaccumulation of halogenated dimethyl bipyrroles inan Arctic marine food web using stable nitrogen isotope analysis.Environ Pollut 116:85–93
Tittlemier SA, Pranschke JL, Norstrom RJ (2002c) Vapour pressures,aqueous solubilities, and octanol/water partition coefficients ofHDBPs. in press
Tittlemier SA, Simon M, Jarman WM, Elliott JE, Norstrom RJ (1999)Identification of a novel C10H6N2Br4Cl2 heterocyclic compoundin seabird eggs. A bioaccumulating marine natural product? En-viron Sci Technol 33:26–33
Vetter W, Alder L, Kallenborn R, Schlabach M (2000) Determinationof Q1, an unknown organochlorine contaminant, in human milk,Antarctic air, and further environmental samples. Environ Pollut,110:401–409
Wania F, Mackay D (1993) Global fractionation and cold condensa-tion of low volatility organochlorine compounds in polar regions.Ambio 22:10–18
Westgate AJ, Tolley KA (1999) Geographical differences in or-ganochlorine contaminants in harbour porpoises Phocoenaphocoena from the western North Atlantic. Mar Ecol Prog Ser177:255–268
HDBPs in Marine Mammal Blubber 255
Copyright © 2022 FDOKUMEN




![Ethyl 2-(6-amino-5-cyano-3,4-dimethyl-2H,4H-pyrano[2,3-c]pyrazol-4-yl)acetate](https://static.fdokumen.com/doc/165x107/630bead9dffd3305850820dd/ethyl-2-6-amino-5-cyano-34-dimethyl-2h4h-pyrano23-cpyrazol-4-ylacetate.jpg)











![Synthesis and spectral properties of 7-( p -bromophenyl)-10,10-dimethyl-8-alkylthio-7,9,10,11-tetrahydro-benz[ c ]acridines and deprotection-aromatization of 7-[( o -; and p -substituted)phenyl]-10,10-dimethyl-7,8,9,10,11,12-hexahydrobenz[](https://static.fdokumen.com/doc/165x107/63138aacfc260b71020f2412/synthesis-and-spectral-properties-of-7-p-bromophenyl-1010-dimethyl-8-alkylthio-791011-tetrahydro-benz.jpg)



