







Bahasa
Halaman
Hukum


b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9
Avai lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ie r . com/ loca te / i ssn /15375110
Research Paper
A dynamic data-based model describing nephropathiaepidemica in Belgium
Sara Amirpour Haredasht a, Jose Miguel Barrios b, Piet Maes c, Willem W. Verstraeten b,f,g,Jan Clement c, Genevieve Ducoffre d, Katrien Lagrou e, Marc Van Ranst c, Pol Coppin b,Daniel Berckmans a, Jean-Marie Aerts a,*aMeasure, Model & Manage Bioresponses (M3-BIORES), Department of Biosystems, Katholieke Universiteit Leuven,
Kasteelpark Arenberg 30, B-3001 Leuven, BelgiumbM3-BIORES, Department of Biosystems, Katholieke Universiteit Leuven, Willem de Croylaan 34, B-3001 Leuven, Belgiumc Laboratory of Clinical Virology, Rega Institute, Katholieke Universiteit Leuven, Minderbroedersstraat 10, B-3000 Leuven, BelgiumdScientific Institute of Public Health, Epidemiology, Juliette Wytsmanstraat 14, B-1050 Brussels, BelgiumeDepartment of Experimental Laboratory Medicine, Katholieke Universiteit Leuven, Herestraat 49, B-3000 Leuven, BelgiumfRoyal Netherlands Meteorological Institute, Climate Observations, PO Box 201, NL-3730 AE, De Bilt, The NetherlandsgEindhoven University of Technology, Applied Physics, PO Box 513, 5600 MB, Eindhoven, The Netherlands
a r t i c l e i n f o
Article history:
Received 11 August 2010
Received in revised form
8 February 2011
Accepted 11 February 2011
Published online 31 March 2011
* Corresponding author. Tel.: þ32 16 321434;E-mail address: [email protected]
1537-5110/$ e see front matter ª 2011 IAgrEdoi:10.1016/j.biosystemseng.2011.02.004
Nephropathia epidemica (NE) is a human infection caused by Puumala virus (PUUV), which
is naturally carried and shed by bank voles (Myodes glareolus). Population dynamics and
infectious diseases in general, such as NE, have often been modelled with mechanistic SIR
(Susceptible, Infective and Remove with immunity) models. Precipitation and temperature
have been found to be indicators of NE, however most SIR models do not take them into
account. The objective of this paper was to develop a dynamic model of incidences of NE in
Belgium by taking into account climatological data. A multipleeinput, single-output (MISO)
transfer function was used to model the incidence of NE. In a first step, the NE cases were
modelled based on data from 1996 until 2003 with an R2T of 0.68. In the next step the MISO
model was validated for incidences of NE in Belgium from 2003 to 2008 (R2T of 0.54). The
output of the MISO models was the number of NE cases in Belgium over the time period
1996 until 2008 and the inputs were average measured monthly precipitation (mm), and
temperature (�C) as well as estimated carrying capacity (vole ha�1).
The monthly values of carrying capacity (K ) were estimated for the whole period by
using an existing mechanistic SIR model. K is related to the variation in seed production in
Northern Europe, which has an effect on the population of bank voles.
In the future, such modelling approaches may be used to predict and monitor forth-
coming NE cases based on climate and vegetation data as a tool for prevention of NE cases.
ª 2011 IAgrE. Published by Elsevier Ltd. All rights reserved.
fax: þ32 16 321480.leuven.be (J.-M. Aerts).. Published by Elsevier Ltd. All rights reserved.

Nomenclature
Bank vole dynamics
S susceptible vole density, vole ha�1
In newly infected vole density, vole ha�1
Ic chronically infected vole density, vole ha�1
P overall density of the vole population, i.e.
S þ In þ Ic, vole ha�1
b(t) birth rate at time t, y�1
m natural mortality rate, y�1
k(t) induced density-dependent effect and seasonal
variation on mortality rates, ha y�1 vole�1
K(t) carrying capacity, vole ha�1
bn transmission rate using density-dependent
transmission during the acute excretion phase,
vole�1 y�1
bc transmission rate using frequency-dependent
transmission during the chronic excretion phase,
y�1
3 indirect contamination rate (ground to rodents),
ha�1 y�1
s inverse of duration of high excretion period, y�1
Soil contamination dynamics
G contaminated proportion of the soil litter surface
4n ground contamination rate by newly infected
voles (rodents to ground), ha vole y�1
4c ground contamination rate by chronically infected
voles, ha vole y�1
d ground decontamination rate, ha�1 y�1
Human contamination dynamics
WSv susceptible number of workers in the village
WSf susceptible number of workers in the forest
HSv other inhabitants in the village
HSf other inhabitants in the forest
If infected individuals (workers and other
inhabitants combined) in the forest
Iv infected individuals in the village
R recovered individuals
rw worker frequency of displacements in the forest,
person�1 y�1
rh other inhabitant frequency of trip in the forest,
person�1 y�1
r inverse of standard time spent in the forest on
each visit, person�1 y�1
3w contamination rate for professionals, ha�1 y�1
3h contamination rate for other inhabitants, ha�1 y�1
u recovery rate from NE, person�1 y�1
Dynamic data-based model
H(t) number of NE cases reported in Belgium per
month, cases month�1
t discrete-time instants with a measurement
interval of 1 month, month
T(t) inputs of the model: average monthly
temperature, �CP(t) inputs of the model: average monthly
precipitation, mm
K(t) inputs of the model: average monthly carrying
capacity, vole ha�1
nti the number of the time delays between each input
i and their first effects on the output
A(z�1) is the denominator polynomial and equals to
1þ a1z�1 þ a2z�2 þ.þ anaz�na
Bi(z�1) are the numerator polynomials and equals to
b0i þ b1iz�1 þ b2iz
�2 þ.þ bnbiz�nbi, cases �C�1,
cases mm�1, cases ha vole �1 (depending on the
input i)
aj the model parameters to be estimated
bi the model parameters to be estimated, cases �C�1,
cases mm�1, cases ha vole�1(depending on the
input i)
z�1 the backward shift operator, defined as
z�1y ðkÞ ¼ y ðk� 1Þna the orders of the respective polynomials
nbi the orders of the respective polynomials
3(t) additive noise, cases
s_ 2
variance of the model residuals, cases2
sy2 variance of themeasured output around its mean,
cases2
b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 978
1. Introduction 1999; Clement, Maes, & Van Ranst, 2007) or remaining unno-
Hantavirusesare rodentor insectivore-bornevirusesandsome
of themare recognizedascausesofhumanhaemorrhagic fever
withrenal syndrome (HFRS). InwesternandcentralEuropeand
in western Russia one of the most important hantavirus is
Puumala virus (PUUV), which is transmitted to humans by
infected red bank voles (Myodes glareolus). PUUV causes
a general mild form of haemorrhagic fever with renal
syndromecallednephropathiaepidemica (NE) (Clement,Maes,
& Van Ranst, 2006).
In general, only 13% of all PUUV infections are sero-diag-
nosed, the others being interpreted as ‘a bad flu’ (Brummer-
Korvenkontio, Vapalahti, Henttonen, Koskela, & Vaheri,
ticed. HFRS, including NE, is now the most underestimated
cause of infectious acute renal failure worldwide, so the offi-
cially registered NE is only the tip of the iceberg.
Mechanistic models play an important role in analysing
the spread and control of infectious diseases (Anderson &
May, 1979; May & Anderson, 1979). Many attempts have
been made to build mathematical models describing the
dynamics of the bank voles’ population and spread and
survival of PUUV (Allen, Langlais, & Phillips, 2003; Sauvage,
Langlais, & Pontier, 2007; Sauvage, Langlais, Yoccoz, &
Pontier, 2003; Wolf, Sauvage, Pontier, & Langlais, 2006).
These models are typically based on components such as an
epidemiological compartment structure, the nature of the

b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 79
incidence, a demographical structure of the population, and
the interaction between the demographical structure and the
epidemiological incidence of the disease.
Mathematical models, such as the model of Sauvage et al.
(2007), developed to describe the population dynamics of
infectious disease are often dealing with the ecology of the
interaction between host and parasite. The dynamics of
animal population and disease behaviour involve four main
factors as described by these models: the host providing
a habitat for parasites, the degree to which parasites reduce
host mortality, the extent to which the host acquires immu-
nity, and the necessity of transmission from one host to the
next (Anderson & May, 1979). In reality, in wild animals, such
as bank voles, disease risk depends on many biological
complications. The population dynamics of host and infection
do not just depend on the abundance and nature of the
infections but are also influenced by the environmental
factors related to the abundance of hosts (bank voles) and
environmental factors related to the spread of the parasites
(PUUV).
Although these mechanistic models have important
scientific merits, they also have limitations. Often, these
models just show the demographic variability of the pop-
ulation without considering the environment and its impact
on the target population.
Taking into account the role of the climate and vegetation
conditions can assist in (i) a better understanding of the
transmission characteristics of the disease, (ii) making fore-
casts about the epidemics of the disease based on expected
trends in future climatological and environmental conditions
and (iii) analysing crucial data that influence the occurrence of
the disease. Therefore, models that consider the dynamics of
climate and of vegetation conditions may play an important
role in improving detection, control and planning of the inci-
dence of some infectious diseases.
The transmission dynamics of PUUV are very complex.
They involve the interaction between environment, tree
biology, the bank vole’s population cycle and human risk
behaviour (Bennet et al., 2006). It is not surprising that the
bank vole’s population dynamics, spread, and survival of
PUUV are complex. Consequently, mechanistic models
describing these complex interactions turn out to be compli-
cated although they are still an idealisation of reality which is
much more complex than the equations. These mechanistic
models can be compared with data-based mathematical
procedures, where the model is inferred, and the model
parameters are directly estimated from experimental data,
using more objective statistically-based methods (e.g. Costa,
Borgonovo, Leroy, Berckmans, & Guarino, 2009; Ferentinos &
Albright, 2003; Rastetter, 1987; Thanh, Vranken, Van Brecht,
& Berckmans, 2007; Ushada & Murase, 2006). The most
straightforward way to obtain a model based on experiments
is to describe the data statistically with some mathematical
relationships (Meinzer, 1982). Living organisms in general
respond dynamically to changes in their physical micro-
environment. Many attempts have already been made to
model these biological responses of living organisms to their
physical micro-environment (Aerts, Steppe, Boonen, Lemeur,
& Berckmans, 2007; Aerts, Wathes, & Berckmans, 2003; Cho
& Mostaghim, 2009).
Because of the dynamic nature of the bank vole’s pop-
ulation, a dynamic systems approach might be a valuable
alternative to mechanistic models to investigate the under-
lying mechanisms. The resulting dynamic data-based models
of this type are often simple in structure, inherently stochastic
in form and are characterised by the minimum number of
parameters required to justify the dynamic information
content of the available data (Young, 1984).
In order to predict the NE incidence in humans, it is there-
fore necessary to havemore knowledge about the dynamics of
hantavirus infections aswell as the bank vole’s population and
their interactions with their natural environment. Several
studies have been carried out to develop a tool for predicting
the epidemic years of NE incidence in Belgium, based on
environmental factors. Clement et al. (2009) showed, through
statistical analysis, that it was possible to predict the epidemic
years based on the precipitation and temperature of the
previous years. Tersago, Servais, Heyman, Ducoffre, and Leirs
(2009) showed a relation between annual NE incidence, based
on tree ecology, and the average air temperature and precipi-
tation of summer and autumn, by using a generalised linear
model (GLM). Both studies related the incidence of NE cases to
the mast year phenomenon that has a direct influence on the
number of bank voles in the forest.
We hypothesised that combining a data-based modelling
approach with a mechanistic SIR model would allow the
dynamics of the NE cases to be modelled with a compact
model structure that takes into account climatological data. In
this study, we aimed to build a multiple-input, single-output
(MISO) transfer function to model the incidence of NE cases in
Belgium from 1996 till 2003 as a function of measured average
monthly air temperature (�C),monthly precipitation (mm) and
carrying capacity (vole ha�1) estimated from the mechanistic
SIR model described by Sauvage et al. (2007). To validate the
data-basedmodel, we usedmeasurements of the NE cases, air
temperature, precipitation and carrying capacity from 2003
until 2008.
The outbreaks and spread of hantavirus have been ques-
tioned and studied for many years. An important added value
of modelling NE cases is that it can be used as a tool in future
to study themechanism bywhich the virus spreads, to predict
the future course of an occurrence and to evaluate strategies
to control the epidemics.
2. Materials and methods
2.1. Data
2.1.1. Nephropathia epidemica infectionsThe Scientific Institute of Public Health (IPH, Brussels) in
Belgium provided NE data. In Belgium, the weekly numbers of
NE cases per postal code (a spatial entity smaller than the
municipality) were available from 1994 until 2008.
2.1.2. Climate dataThe Royal Meteorological Institute of Belgium (RMI, Ukkel),
which is located in thecentre of Belgium,provideddailydataon
air temperature (�C) and precipitation (mm) from 1996 to 2008.

b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 980
To be capable of capturing the dynamics of the NE cases,
we calculated monthly average precipitation (mm) and
average temperatures (�C) based on the daily reported climate
data for Ukkel.
2.1.3. Tree seed production categoriesThe Tree Seed Centre of the Ministry of the Walloon Region
supplied categories of seed production of beech andnative oak
species (Quercus robur, Quercus petraea). Tree seed production
for each tree species is divided into four categories: “very good
year” (the species is fruiting throughout the Walloon territory
and practically all trees are bearing seed in high quantities),
“goodyear” (the species is fruiting throughout the territory, but
the trees are bearing much less seed and some trees do not
fruit), “moderate year” (there is a reduced number of trees
bearing seeds and sometimes only located in a portion of the
territory) and “low year” (years without fructification in
significant quantities).
2.2. Epidemiological model
The population SIR model presented here is based on the
equations proposed by Sauvage et al. (2007). Their model
consists of two sub-models. The first sub-model (bank vole’s
population sub-model) describes the bank vole’s demography
and infection and the second sub-model (human population
sub-model) describes the access of humans to the forest and
the dynamics of the subsequent human infections. In the
model, the bank voles contaminate the environment which
then spreads the virus into the human population.
The first sub-model describes the bank vole’s demography
and illustrates the interaction between the trees’ biology by
the K(t) factor (Eq. (5)),the bank vole’s population cycle and
environmental contamination (G). The humans sub-model
illustrates the interaction between the environment (G) and
human risk behaviour. The transmission of the PUUV to
humans occurs by inhalation of air-suspended particles of
urine, faeces, or saliva from infected bank voles (Lee et al.,
1981; Nozum, Rossi, Stephenson, & Leduc, 1988).
2.2.1. Bank vole’s population sub-modelThe epidemic model with one viral infection is implemented
using themodel of Sauvage et al. (2007). Themodel consists of
three differential equations with states S, In and Ic defined as
“susceptible vole density” (vole ha�1), “newly infected vole
density” (vole ha�1) and “chronically infected vole density”
(vole ha�1) respectively. The model equations are:
dSdt
¼bðSþInþIcÞ�ðmþkðSþInþIcÞÞS��bnInþ
bcIcP
�S�eGS (1)
Indt
¼�bnIn þ
bcIcP
�Sþ eGS� sIn � ðmþ kðSþ In þ IcÞÞIn (2)
Icdt
¼ sIn � ðmþ kðSþ In þ IcÞÞIc (3)
The model parameters are described in the Nomenclature
section as defined by Sauvage et al. (2007).
Parameters b and k are functions of time. Parameter b rep-
resented birth rate and k integrated the seasonal variations
linked to the reproductive season or the possiblemulti-annual
variations of the environmental carrying capacity, which is
responsible for multi-annual fluctuations of the bank vole
population density.
The function used to express the birth rate b(t) by Sauvage
et al. (2007) is:
b ¼ j20sinð2pðt� 0:15ÞÞj þ 20sinð2p� 0:15Þ (4)
Another point which needs further explanation is the
parameter k(t), the density-dependent effect and seasonal
variation onmortality rate. It corresponds to themean growth
rate of the bank vole population divided by the forest carrying
capacity:
k ðtÞ ¼ ð10�mÞ=K ðtÞ (5)
where K(t) is the environment carrying capacity.
Throughout the text, b (bn and bc) refers to a direct trans-
mission rate and e (e, ew and eh) to an indirect one. bn was the
direct transmission rate from newly infected rodents. bc was
the transmission rate from chronically infected voles, and is
directly affected by the number of different neighbours met
per vole per year. The direct transmission contact rates, bn and
bc, are unknown in bank vole populations. They were cali-
brated using the estimate from Begon et al. (1998) for the bank
voleecowpox system.
We took a conservative indirect contact rate assuming that
on average each point of the area is visited once every 73 days,
i.e. 5 (¼365/73) visits per unit of area per year (Sauvage et al.,
2007).
Site contamination/decontamination dynamics allowed
the spread of infection from voles to humans. The modelling
process was similar to that used by Berthier, Langlais, Auger,
and Pontier (2000).
dGdt
¼ ð4nIn þ 4cIcÞð1� GÞ � dG (6)
where 4n, the proportion of territory contaminated by one
newly infectious individual per year, and 4c, the equivalent
rate for chronically infected voles, were estimated from
Rozenfeld, Le Boulenge, and Rasmont (1987). The value of 4n is
higher than that of 4c because newly infected voles release
virus in higher quantities and through more routes than do
chronically infected voles do. Rozenfeld et al. (1987) showed
a strategic choice of scentmarks deposited bymale bank voles
on their territory borders, their feeding point’s area and even
their rivals’ burrows. These marked areas are more probably
explored by other bank voles. The ground decontamination
rate (d ) in the Sauvage model is taken as 30 ha-1 y-1 as it is
basedon the reciprocal of the virus survival time in the ground,
which is about 12 days (Kallio, 2003), calculated as 365/12.
The initial condition of S(0), In(0) and Ic(0) are non-negative.
The total population size is P ¼ S þ In þ Ic, which satisfied the
differential equations.
2.2.2. Human population sub-modelThe dynamics of NE infection in humans were written using
a SIR (Susceptible, Infective andRemovewith immunity)model
with indirect transmission and for a two-host sub-population,
forest workersW and others H (Sauvage et al., 2007).

b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 81
dWSv
dt¼ �rwWSv þ rWSf (7)
dHSv
dt¼ �rhHSv þ rHSf (8)
dWSf
dt¼ rwWSv � ew GWSf � rWSf (9)
dHSf
dt¼ rwHSv � ehGHSf � rHSf (10)
dIfdt
¼ �ewWSf þ ehHSf
�G� rIf (11)
dIvdt
¼ rIf � gIv (12)
dR
dt¼ gIv (13)
In the human population sub-model, G describes the
threshold value of the index of environment contamination
required for humans to become infected. The model param-
eters are described in the Nomenclature section.
We consider a threshold for human contamination as it is
described for tularaemia in both animals and humans (Patrick,
2001). Such a threshold results in zero values for ew and eh
when G� 0.08. This threshold is based on the assumption that
humans breathe 1.5 m further above the forest litter than the
voles. Moreover, the high host specificity of hantaviruses
(Monroe et al., 1999) should reduce the effective infectivity of
k ðtÞ ¼ ð4ndIn=dtþ 4cdIc=dt� 4nðBnIn þ BcIc=PÞS� 4neGS� ð4c � 4cÞsIn �mÞ�ðdG=d ðtÞ þ dGÞ=1� G
=P ðtÞ 15)
the virus when humans are exposed to infectious aerosols, as
is the case, for example, for tularaemia (Patrick, 2001). Ifrepresents the number of people acquiring the virus in the
forest. Iv and If represent the actual numbers of diseased
people in the forest and town respectively at the considered
moment. As NE is contracted only once in a lifetime, the
infected people (If and Iv) were removed from the system and
accumulate in the R class (Eq. (13)) as they recover. The R class
equals the total number of NE cases since the beginning of the
simulation.
2.3. Calculation of carrying capacity
Carrying capacity of a biological species K(t) in an environ-
ment is the population size of the species that the environ-
ment can sustain given the food, habitat, water and other
necessities available in the environment (Sayre, 2008).
The environmental carrying capacity K(t) of the bank voles
follows the variation in seed production in the northern part of
Europe. K(t) is one of the driving factors responsible for multi-
annual fluctuations of the bank vole’s population density
(Sauvage et al., 2003). It has beendemonstrated that increasing
amounts of beech and oak fruits, in so-called mast years,
increases bank vole numbers, and that this is associated with
outbreaks of NE in northwest Europe (Clement et al., 2009).
Recently, many attempts have been made to predict mast
years by analysing meteorological data and to link the
occurrence of the mast year to NE cases (Bennet et al., 2006;
Clement et al., 2009; Linard et al., 2007; Tersago et al., 2009).
However, none of these approaches quantified the environ-
mental carrying capacity K(t).
In our research, we estimated the carrying capacity by
using the mechanistic model described in Sauvage et al.
(2007). First, we calculated the ground contamination rate
from the human’s sub-model based on the number of cases in
Belgium from 1996 until 2007. As explained for the human
population sub-model, the numbers of infection cases in
humans is calculated in the mechanistic model by summing
Eqs. (11) and (12). In order to quantify the carrying capacity
based on the human cases, Eqs. (11) and (12) were replaced in
the mechanistic model by the known NE cases in Belgium on
a monthly basis from 1996 until 2007. In order to calculate the
equations, the human cases in Belgium were defined as
a Fourier transform function. By using themechanisticmodel,
the ground contamination rate, G(t) was defined based on
human cases by Eqs. (11) and (12) as:
G ðtÞ ¼ d�If þ Iv
�ð1þ gÞdt
��eWWSf þ ehHSf
�(14)
k(t) is calculated from the bank vole’s population sub-
model. The k(t) value was derived from Eqs. (2), (3) and (6). The
final formula was:
The equations were solved by using the ODE45 function in
Matlab� (R2008a, MathWorks Inc., US). The ODE45 function is
used to solve ordinary differential equations. The human
cases were described using the Fourier transform function in
Matlab�.
The estimated carrying capacity (Eq. (5)) was used in a next
step as one of the inputs in the data-based modelling phase.
2.4. Dynamic data-based modelling
The objective of the next step was to quantify the dynamics of
the incidence of NE in Belgium and to relate it with environ-
mental data. First, a multiple-input, single-output (MISO)
transfer function (TF) was used to model the incidence of the
disease from 1996 until 2008 as function of the climatology
data of temperature and precipitation only. The model
structure used could be described as follows (Young, 1984):
H ðtÞ ¼ BT ðz�1ÞA ðz�1Þ Tðt� ntTÞ þ BP ðz�1Þ
A ðz�1Þ P ðt� ntPÞ þ e ðtÞ (16)
where H(t) is the number of NE cases reported in Belgium
per month; t represents discrete-time instants with

b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 982
a measurement interval of 1 month; T(t) and P(t) represent the
two inputs of the model, namely average monthly air
temperature (�C) and average monthly precipitation (mm)
respectively; nti is the number of the time delays between each
input i and their first effects on the output; A(z�1) is the
denominator polynomial and equals
1þ a1z�1 þ a2z�2 þ.þ anaz�na; Bi(z�1) are the numerator
polynomials linked with the inputs i and are equal to
b0i þ b1iz�1 þ b2iz�2 þ.þ bnbiz�nbi; aj, bi are the model param-
eters to be estimated; z�1 is the backward shift operator,
defined as z�1y ðkÞ ¼ y ðk� 1Þ; na, nbi are the orders of the
respective polynomials; and e(t) is additive noise, a serially
uncorrelated sequence of random variables with variance s2
that accounts for measurement noise, modelling errors and
effects of unmeasured inputs to the process (assumed to be
a zero mean).
Second, the MISO model was extended with carrying
capacity as a third input variable. The carrying capacity was
estimated from a mechanistic model as described above. The
model structure can be described as follows (Young, 1984):
HðtÞ¼BT ðz�1ÞAðz�1ÞTðt�ntTÞ þBP ðz�1Þ
Aðz�1ÞPðt�ntPÞ þBk ðz�1ÞAðz�1ÞKðt�ntkÞþeðtÞ
(17)
where K(t) is the average monthly carrying capacity
(vole ha�1). The available time series of the three inputs and
the output of the MISO model are shown in Fig. 1.
1996 1997 1998 1999 2000 2001 20
50
100 O
Num
ber o
f NE
case
s
0
20
0
5
1995 1996 1997 1998 1999 2000 20
50
100
Ave
rage
tem
pera
ture
°C
Pre
cipi
taio
n m
mC
arry
ing
capa
city
rode
nt-1
Fig. 1 e Time series of availab
The model parameters were estimated using a refined
instrumental variable approach with the Captain toolbox in
Matlab� (Taylor, Pedregal, Young, & Tych, 2007; Young, 1984).
For each data set, themodel parameters of Eqs. (16) and (17)
were estimated. The resulting models were evaluated by the
coefficient of determination (R2T; Young & Lees, 1993).
The ability to estimate the parameters of a TF model
represents only one side of the model identification problem.
Equally important is the problem of objective model order
identification. This involves the identification of the best
choice of orders of the polynomials A(z�1) and Bi(z�1). The
process of model order identification can be assisted by
the use of well-chosen statistical measures that indicate the
presence of over-parameterisation. A good identification
procedure used to select the most appropriate model order
[na, nbi] is based on the minimisation of the Young Identifi-
cation Criterion (YIC; Young & Lees, 1993).
The YIC is a heuristic statistical criterion that consists of
two terms. The first term provides a normalised measure
of how well the model fits the data. The smaller the variance
of the model residuals in relation to the variance of the
measured output, the more negative this term becomes. The
second term is a normalised measure of how well the model
parameter estimates are defined. This term tends to become
less negative when the model is over-parameterised (more
complex) and the parameter estimates are poorly defined.
Consequently, the model that minimises YIC provides a good
002 2003 2004 2005 2006 2007 2008
utput
Year Inputs
001 2002 2003 2004 2005 2006 2007
Year
le input and output data.

b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 83
compromise between goodness of fit and parametric effi-
ciency (which is equivalent to complexity).
After YIC, the standard errors on the parameter estimates
were calculated as the root of the diagonal elements of the
covariance matrix. Based on the standard errors, the 95%
confidence interval (CI95%) for each parameter estimated (q_)
could be calculated as:
CI95% ¼ q_ � t0:025;N�npSE ðq_ Þ (18)
where t0.05,N-np is the value given by the two-tailed student t
distribution with N the number of data used to estimate the
parameters and np the number of parameters. In this study,
the value for t0.05,N�np was approximately 2. This means that
the parameter estimation was considered to be reliable when
the observed value of the parameter estimate was at least
twice the value of its standard error (meaning that the
parameter value was significantly different from 0).
Thirdly, the model stability was calculated for the selected
TF models as part of the model evaluation. Stability was
determined by quantifying the poles (roots of the A(z�1)
polynomial) of the models. The model is considered to be
stable when all the poles lie within the unity circle in the
complex plane or z-plane (Young & Wang, 1987).
In order to identify the models for the whole period,
different combinations for na, nbt, nbp, nbk, ntT, ntP and ntKwere calculated.More specifically for theMISOmodelwith two
inputs, na ranged from 1 to 4, nbT and nbP ranged from 1 to 4,
and ntT and ntP ranged from 0 to 4. Therefore, to identify the
first MISO model of two inputs and one output, in total 1600
(4� 4� 4� 5� 5) possible TFmodelswere calculated. The total
number of possible TFmodels for the secondMISOmodelwith
three inputs and one outputwas 6400 (4� 4� 4� 4� 5� 5� 1).
The delay ntk was predetermined to be 12 (only one value). It
was assumed that there is 12-month time delay between the
carrying capacity and the incidence of the NE cases, as sug-
gested by Sauvage et al. (2007). All these models were ranked
based on YIC (from low to high values). Only the first 20 best
models as indicated by YIC were selected for further evalua-
tion. Based on the resulting 20 models, the TF order identifi-
cation wasmade on the basis of the goodness of fit, expressed
as the coefficient of determination R2T, the stability of the
resulting model and the confidence interval of the estimated
model parameters. This approach was used to identify one
final model (i.e. model structure with specific model parame-
ters) for the whole period from 1996 until 2003. The identified
model was validated by applying the model to inputeoutput
data from2003 to 2008.Validationcanbedefinedas theprocess
that determines the accuracy with which a model represents
a real system (Young & Lees, 1993).
3. Results
The first aim of this study was to quantify the dynamics of the
incidence of NE in Belgium and to relate it to climatological
data (average monthly temperature and precipitation only).
When applying the modelling approach to the period from
1996 until 2008, the YIC criterion selected models that were
predominantly third order (na ¼ 3, Eq. (16)). Comparing the
resulting models for the period 1996e2003, there was one
model structure that (i) was selected by YIC as being one of the
20 best models, and (ii) was stable (all poles within the unit-
circle in the z-plane). This model structure was described by
na ¼ 3, nbT ¼ 1, nbP ¼ 1, ntT ¼ 0 and ntP ¼ 0. The resulting TF
model structure is represented in Eq. (19).
H ðtÞ ¼ bTz�1
1þ a1z�1 þ a2z�2 þ a3z�3T ðtÞ
þ bPz�1
1þ a1z�1 þ a2z�2 þ a3z�3P ðtÞ (19)
However, looking at themodelling results in Fig. 2, it is clear
that the model was not able to describe the periodic peaks in
NE cases (1996, 1999, 2001, 2003, 2005).
In an attempt to improve the data-based model, because
the climatological data alone were not able to explain the
dynamics of NE cases and since carrying capacity is ameasure
for, among other things, food availability for the bank voles,
we introduced the variable ‘carrying capacity’ as a third input.
Elasticity and sensitivity analyses were done for the SIR
(Susceptible, Infective and Remove with immunity) mecha-
nistic model and revealed that slight differences in the forest
carrying capacity values used in the model create very large
gradient in the predicted epidemic pattern in humans
(Sauvage et al., 2007). Therefore, it could be expected that
introduction of this variable could help explain the different
dynamics in the years considered. As described before, the
value of carrying capacity was estimated from the mecha-
nistic SIR model of Sauvage et al. (2007). The incidence of NE
cases per year versus the average estimated carrying capacity
(vole ha�1) is shown in Fig. 3. As demonstrated in previous
studies conducted by Clement et al. (2009), Sauvage et al.
(2007) and Tersago et al. (2009), high NE incidence were asso-
ciated with abundance of food in the previous autumn.
As can be seen in Fig. 3, there is an unexpected increase in
the number of NE cases in 2005. This pattern of increased NE
cases continued in the following years, as 163 cases were
confirmed in 2006 and again 298 in 2007. Until 2005, each
epidemic year (>100 NE cases annually) was followed by at
least one non-epidemic year with <60 annual NE cases
(Clement et al., 2009, 2010; Tersago et al., 2009).
Fig. 3 indicates further the category of seed production of
annual beech and native oak fruiting in southern Belgium.
Analysis of Tersago et al. (2009) confirmed the data in Fig. 3,
which showed that each peak year for number of NE cases is
preceded by a year with high seed production of at least native
oak, or beech or both. The extreme NE outbreak in 2005 was
probably a result of a heat wave occurring 2 years earlier in
summer 2003 (Tersago et al., 2009). Higher temperature from
summer to autumn in 2006 and 2007 increased the green
biomass of herbaceous plants that constituted a high
percentage of the bank vole’s diet. High temperature during
the reproductive season increases bank vole’s reproduction
and decreases the use of resources to maintain the high
metabolic rate of bank voles (Tersago et al., 2009). As a result,
from 2005 until 2008, the higher carrying capacity did not
coincide with seed production of beech and oak trees.
As described earlier we estimated in total 6400 models for
the case of three inputs and selected the 20 best models based

1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 20080
10
20
30
40
50
60
70
Year
Num
ber o
f NE
cas
es
Fig. 2 e The resulting model (——) of the data-based MISO model with 2 inputs (average monthly temperature and
precipitation) versus measured (–�–) number of NE cases in Belgium from January 1996 till September 2007.
b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 984
on YIC. The models that were selected were predominantly
third order (na ¼ 3). Comparing the resulting models for the
period 1996e2003, there was one model structure that (i) was
selected by YIC as being one of the 20 best models, (ii)
described the data accurately (R2T ¼ 0.69), and (iii) was stable
(all poles within the unit-circle in the z-plane) This model
structure was described by na ¼ 3, nbT ¼ 2, nb2 ¼ 2, nbK ¼ 2,
ntT ¼ 0, ntP ¼ 3 and ntK3 ¼ 12. The resulting TFmodel structure
is represented in Eq. (20).
H ðtÞ ¼ b0Tz�1 þ b1Tz�1
1þ a1z�1 þ a2z�2 þ a3z�3T ðtÞ
þ b0Pz�1 þ b1Pz�1
1þ a1z�1 þ a2z�2 þ a3z�3Pðt� 3Þ
þ b0Kz�1 þ b1Kz�1
1þ a1z�1 þ a2z�2 þ a3z�3Kðt� 12Þ (20)
The specific values for the model parameters (a1, a2, a3, b0T,
b1T, b0P, b1P, b0K, b1K) are presented in Table 1. As can be seen in
the table, the standard deviations on the parameter estimates
were small compared to the absolute parameter values,
providing confidence in the parameter estimates. The
modelling results for the period 1996e2003 are shown in Fig. 4.
The second part of the data set (January 2003 until
September 2007) was used for validation to determine
whether the model parameters and structure described in
Table 1 were adequate for the rest of the data set. The model
validation results are presented in Fig. 5. The model described
the data with a R2T of 0.54.
To evaluate the performance of the model further, for
both data sets the hypothesis that the slope of the
regression line of modelled versus measured data equalled
one and the intercept equalled zero was tested. A linear
regression analysis was performed. The result of this anal-
ysis is shown in Fig. 6 for the first data set and Fig. 7 for the
second (validation) data set. It is clear that the first data set
(1996e2003) was better modelled (slope of 0.74 and intercept
of 4.27), compared to the second part of the data set with
a slope of 0.68 and an intercept of 6.67. However, for both
data sets the null hypothesis was rejected as the values of
one for the slope and zero for the intercept were outside the
95% confidence interval (the CI95% of the slopes equalled to
0.74 � 0.12 and 0.68 � 0.18 for first and second data set
respectively and the CI95% of the intercepts equalled to
4.27 � 1.32, 6.67 � 3.86 for the first and second data set
respectively).
When looking at the modelling results in general, it could
be observed that the model described the data quite well, but
when looking in more detail it is clear that the model showed
particularly good results for the epidemic months in which
the data were more dynamic. To evaluate the performance of
the model in these more dynamic periods, we applied the
same analysis for each dataset, as reported above, for the
months with more than 10 NE cases. The results of this
analysis are shown in Figs. 8 and 9.
Using the 95% confidence interval, the null hypothesis was
now accepted for the two data sets. The 95% confidence
interval for the slope of the first data set was 0.86 � 0.24 and
for the second data set it was 0.75 � 0.26. The 95% confidence
interval for the intercept of the first data set was 2.26 � 4.64
and for the second data set it was 4.43 � 6.83.
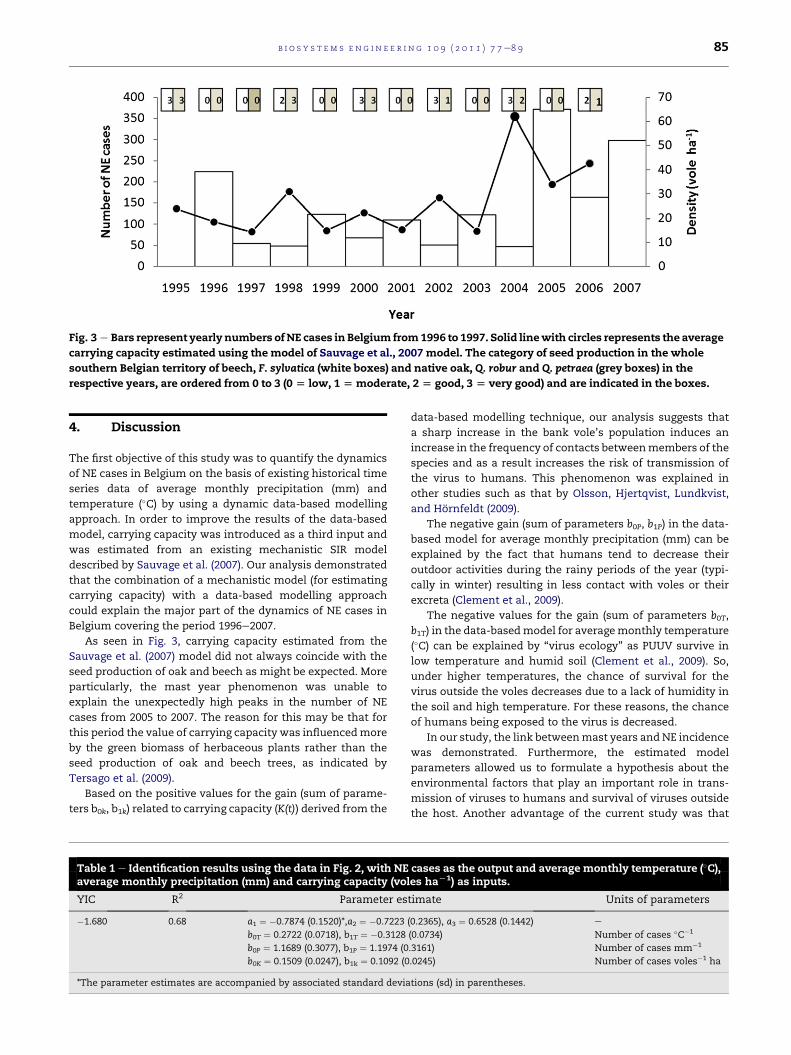
Fig. 3 e Bars represent yearly numbers ofNE cases in Belgium from1996 to 1997. Solid linewith circles represents the average
carrying capacity estimated using the model of Sauvage et al., 2007 model. The category of seed production in the whole
southern Belgian territory of beech, F. sylvatica (white boxes) and native oak, Q. robur and Q. petraea (grey boxes) in the
respective years, are ordered from 0 to 3 (0 [ low, 1[ moderate, 2[ good, 3 [ very good) and are indicated in the boxes.
b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 85
4. Discussion
The first objective of this study was to quantify the dynamics
of NE cases in Belgium on the basis of existing historical time
series data of average monthly precipitation (mm) and
temperature (�C) by using a dynamic data-based modelling
approach. In order to improve the results of the data-based
model, carrying capacity was introduced as a third input and
was estimated from an existing mechanistic SIR model
described by Sauvage et al. (2007). Our analysis demonstrated
that the combination of a mechanistic model (for estimating
carrying capacity) with a data-based modelling approach
could explain the major part of the dynamics of NE cases in
Belgium covering the period 1996e2007.
As seen in Fig. 3, carrying capacity estimated from the
Sauvage et al. (2007) model did not always coincide with the
seed production of oak and beech as might be expected. More
particularly, the mast year phenomenon was unable to
explain the unexpectedly high peaks in the number of NE
cases from 2005 to 2007. The reason for this may be that for
this period the value of carrying capacity was influencedmore
by the green biomass of herbaceous plants rather than the
seed production of oak and beech trees, as indicated by
Tersago et al. (2009).
Based on the positive values for the gain (sum of parame-
ters b0k, b1k) related to carrying capacity (K(t)) derived from the
Table 1 e Identification results using the data in Fig. 2, with NEaverage monthly precipitation (mm) and carrying capacity (vol
YIC R2 Parameter es
�1.680 0.68 a1 ¼ �0.7874 (0.1520)*,a2 ¼ �0.7223
b0T ¼ 0.2722 (0.0718), b1T ¼ �0.3128
b0P ¼ 1.1689 (0.3077), b1P ¼ 1.1974 (0
b0K ¼ 0.1509 (0.0247), b1k ¼ 0.1092 (0
*The parameter estimates are accompanied by associated standard devia
data-based modelling technique, our analysis suggests that
a sharp increase in the bank vole’s population induces an
increase in the frequency of contacts betweenmembers of the
species and as a result increases the risk of transmission of
the virus to humans. This phenomenon was explained in
other studies such as that by Olsson, Hjertqvist, Lundkvist,
and Hornfeldt (2009).
The negative gain (sum of parameters b0P, b1P) in the data-
based model for average monthly precipitation (mm) can be
explained by the fact that humans tend to decrease their
outdoor activities during the rainy periods of the year (typi-
cally in winter) resulting in less contact with voles or their
excreta (Clement et al., 2009).
The negative values for the gain (sum of parameters b0T,
b1T) in the data-basedmodel for averagemonthly temperature
(�C) can be explained by “virus ecology” as PUUV survive in
low temperature and humid soil (Clement et al., 2009). So,
under higher temperatures, the chance of survival for the
virus outside the voles decreases due to a lack of humidity in
the soil and high temperature. For these reasons, the chance
of humans being exposed to the virus is decreased.
In our study, the link betweenmast years and NE incidence
was demonstrated. Furthermore, the estimated model
parameters allowed us to formulate a hypothesis about the
environmental factors that play an important role in trans-
mission of viruses to humans and survival of viruses outside
the host. Another advantage of the current study was that
cases as the output and average monthly temperature (�C),es haL1) as inputs.
timate Units of parameters
(0.2365), a3 ¼ 0.6528 (0.1442) e
(0.0734) Number of cases �C�1
.3161) Number of cases mm�1
.0245) Number of cases voles�1 ha
tions (sd) in parentheses.

1996 1997 1998 1999 2000 2001 2002 20030
5
10
15
20
25
30
35
Year
Num
ber o
f NE
cas
es
Fig. 4 e The resulting model simulation (——) of the data-based MISO model with 3 inputs (average monthly temperature,
precipitation and estimated carrying capacity) versus measured (–�–) number of NE cases in Belgium from January 1996 till
January 2003. The model described the data with the R2T of 0.68.
2003 2004 2005 2006 2007 20080
10
20
30
40
50
60
70
Year
Num
ber o
f NE
cas
es
Fig. 5 e The simulation result (——) versus measured (–�–) number of NE cases in Belgium from January 2003 till January
2008.
b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 986

Fig. 8 e The MISO model result for epidemic months (>10
NE cases) versus number of NE cases in Belgium from
January 1996 till January 2003 (circles). The solid line
represents the regression line of the modelled versus
measured data.
Fig. 6 e The MISO model result versus number of NE cases
in Belgium from January 1996 till January 2003 (circles).
The solid line represents the regression line of the
modelled versus measured data.
b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 87
employing a data-based modelling approach allowed us to
quantify and predict the dynamics of NE cases on a monthly
basis. To the authors’ knowledge, this is the first time this has
been reported.
A study in Sweden performed a time series analysis for NE
cases in relation to bank vole’s population dynamics and the
North Atlantic Oscillation (NAO) index. Within this research,
no relationshipwas foundbetween theNAOand thenumberof
NE cases over two time series, totalling 37 years (Palo, 2009).
The analysis included a long-term data set on host abundance
(25 years). The incidence of Puumala virus infections in the
host was not measured. In order to model the dynamics of NE
cases, a multivariate stepwise model was applied with vole
index, winter mortality and NAO index at different time lags.
The analysis showed that only vole index contributed signifi-
cantly to the model of human NE. The authors acknowledged
that the NAO may be too coarse a measurement and that
temperature and precipitation individually may be better
predictors of NE incidence. In our study, we used average
measuredoutside temperatureandprecipitationaspredictors.
Fig. 7 e The simulation result versus number of NE cases in
Belgium from January 2003 till January 2008 (circles). The
solid line represents the regression line of the modelled
versus measured data.
Furthermore,weused theestimatedcarrying capacitywhich is
an indicator for abundance of bank voles in the forest. Our
model could describe the dynamics of NE in a more accurate
way despite the fact that Palo (2009) had a longer time series as
well asmeasurements of the temporal bank voles fluctuations.
In the study of Palo (2009), the NE cases were modelled with
a coefficient of determination (R2) between measured and
modelled data of 0.39. In our study, the R2T valuewas 0.68when
applying the model to the training data set. In contrast to the
work of Palo (2009) we also validated our model on a separate
dataset, resulting inaR2T valueof 0.54.AlthoughtheR2
T values in
our study were not very high, they were considerably higher
than values found by other researchers for similar modelling
approaches.
In another study, NE epidemics have been explained by
rodent host abundance (Olsson et al., 2009). The explanation is
based on the strong positive correlation and reported
temporal patterns linking bank voles’ abundance and NE
cases in Sweden. In the study a linear regression analysis was
Fig. 9 e The model simulation for epidemic months (>10
NE cases) versus number of NE cases in Belgium from
January 2003 till January 2008 (circles). The solid line
represents the regression line of the modelled versus
measured data.

b i o s y s t em s e n g i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 988
performed in the NE cases in Sweden (per year) from 1989
until 2007 based on the autumn bank voles trapping indices.
The NE cases (per year) were modelled with a coefficient of
determination (R2) between measured and modelled data of
0.63 but no validation was performed. In our study we
managed tomodel the NE cases in Belgium on amonthly basis
with a R2T of 0.68 without using any data about the cyclic phase
of the bank vole fluctuations, bank vole abundance, or PUUV
dynamics in the bank vole populations. Knowledge about the
dynamics of the bank vole population and NE cases was
introduced into our approach via the mechanistic model of
Sauvage et al. (2007). This can be considered as useful when
there is a lack of the trapping data.
Although the modelling results look promising when
comparedwith the available literature, several limitations can
be identified which are related to the applied modelling
methodology. The first limitation we faced when identifying
the data-based model was that the NE data in Belgium cover
a period of only 11 years. Although the number of sampleswas
theoretically sufficient to estimate the model parameters of
the model structure used, as indicated by the acceptable
standard errors on the parameter estimates (Table 1), a data-
set covering a longer period would be expected to improve the
modelling results. Another limitation is the quantification of
the carrying capacity. As shown in the work of Sauvage et al.
(2007), carrying capacity is one of the key factors in the
transmission of the disease. However, today there is no clear
way of quantifying this critical factor. Furthermore, it should
be noticed that the climatological values used as inputs in the
data-based model were measured at the weather station in
Ukkel, which is located close to Brussels, near to the centre of
Belgium. Since we used these climatological data tomodel the
NE cases for the whole country, it might be expected that
taking into account spatial characteristics of the climatolog-
ical data could potentially improve the modelling results.
In contrast to the mechanistic modelling approach, data-
based modelling techniques identify the dynamic character-
istics of processes based onmeasured data and as such are not
based on a priori knowledge. As a result, the choice of the
different input variables for explaining the output of the
considered system is crucial. Howeverwe know fromprevious
work (e.g. Sauvage et al., 2007; Tersago et al., 2009) that
climatological data and carrying capacity are important vari-
ables explaining the NE cases.
Based on the earlier critical reflection, several suggestions
can be made for improving this dynamic modelling concept.
Firstly, carrying capacity estimated from the mechanistic
model may be estimated from the vegetation index values
derived from satellite images. Using this method, carrying
capacity could be estimated more directly, provided that the
relationship between the information in the satellite images
(such as broad leaved forest phenology, e.g. Barrios et al., 2010)
and the carrying capacity can be established. Another way to
estimate the carrying capacity is the use of regression models
to investigate the effect of meteorological data and vegetation
indexes on carrying capacity. Secondly, the knowledge that is
described in mechanistic models could be used to explain the
modelparametersof thedynamicdata-basedmodels inamore
physically/physiologically meaningful way. Thirdly, in future
work the data-basedmodelling approachmay be improved by
integration of estimated bank vole population dynamics
measured in thefield.This couldgive thepossibility toquantify
the carrying capacity based on field measurements instead of
epidemiological models. Finally, it is expected that linking the
data-based modelling approach with existing mechanistic
models, such as the model of Sauvage et al. (2007), will create
added value since such data-based mechanistic models can
take advantage of the fact that they combine mechanistic
process knowledge with measured information (e.g. vegeta-
tion indexes, climatologicaldata, etc.)whichmakes themmore
understandable from a biological/ecological point of view, but
at the same time allows real-timemeasured information to be
taken into account to predict future NE outbreaks.
In this study we have focused on the temporal patterns in
the evolution of the emergence of the NE incidence. The
observed spatial features of NE incidence could be the subject
of future research.
5. Conclusion
The objective of this study was to quantify the dynamics of NE
cases using a dynamic data-based modelling approach.
Combining amechanisticmodel as described by Sauvage et al.
(2007) and a data-based modelling approach proved to be
valuable for describing the dynamics of NE cases in Belgium
on a monthly basis from 1996 until 2003 (R2T of 0.68).
The identified data-basedmodel was validated by applying
it to the inputeoutput data from 2003 to 2007.The validation
data were fitted with R2T of 0.54 and it was seen that the model
performedwell in simulating themonthly dynamics of the NE
cases in Belgium.
The results of the current study help to define significant
environmental factors on the spread of the disease. Deter-
mining a dynamic data-based model for NE which includes
factors such as vegetation coverage and abundance of food for
bank voles’ may provide an expert tool to predict (and aid
prevention of) the incidence of NE cases by making use of
remote sensing tools for measuring broad leaved forest
phaenology andmonitoring the vegetation dynamics together
with climatological data.
Acknowledgement
We are grateful to Professor James Taylor for reviewing the
modelling results of this study. This research has been sup-
ported by the Katholieke Universiteit Leuven (project IDO/07/
005). Piet Maes is supported by a postdoctoral grant from the
‘Fonds voorWetenschappelijk Onderzoek (FWO)-Vlaanderen’.
r e f e r e n c e s
Aerts, J.-M., Steppe, K., Boonen, C., Lemeur, R., & Berckmans, D.(2007). Simulation of sap flow in a beech tree by means ofa dynamic data-based model as influenced by a solar eclipse.Biosystems Engineering, 98(4), 446e454.

b i o s y s t em s e ng i n e e r i n g 1 0 9 ( 2 0 1 1 ) 7 7e8 9 89
Aerts, J.-M., Wathes, C. M., & Berckmans, D. (2003). Dynamic data-based modelling of heat production and growth of broilerchickens: development of an integrated management system.Biosystems Engineering, 84(3), 257e266.
Allen, L. J., Langlais, M., & Phillips, C. J. (2003). The dynamics oftwo viral infections in a single host population withapplications to hantavirus. Mathematical Biosciences, 186(2),191e217.
Anderson, R. M., & May, R. M. (1979). Population biology ofinfectious diseases: part I. Nature, 280, 361e367.
Barrios, J. M., Verstraeten, W.W., Maes, P., Clement, J., Aerts, J.-M.,Amirpour Haredasht, S., et al. (2010). Satellite derived forestphenology and its relation with nephropathia epidemica inBelgium. International Journal of Environmental Research and PublicHealth, 7(6), 2486e2500.
Begon, M., Feore, S., Bown, K., Chantrey, J., Jones, T., & Bennett, M.(1998). Population and transmission dynamics of cowpox inbank voles: testing fundamental assumptions. Ecology Letters,1(2), 82e86.
Bennet, E., Clement, J., Sansom, P., Hall, I., Leach, S., & Medlock, J.M. (2006). Environmental and ecological potential for enzooticcycles of Puumala hantavirus in Great Britain. Epidemiology andInfection, 4(1), 1e8.
Berthier, K., Langlais, M., Auger, P., & Pontier, D. (2000). Dynamicsof a feline virus with two transmission modes withinexponentially growing host populations. The Royal Society, 267(1457), 2049e2056.
Brummer-Korvenkontio, M., Vapalahti, O., Henttonen, H.,Koskela, P., & Vaheri, A. (1999). Epidemiological study ofnephropathia epidemica in Finland 1989e96. ScandinavianJournal of Infectious Diseases, 31, 427e435.
Cho, J., & Mostaghim, S. (2009). Dynamic agricultural non-pointsource assessment tool (DANSAT): model application.Biosystems Engineering, 102(4), 500e515.
Clement, J., Maes, P., & Van Ranst, M. (2006). Hantaviruses in theold and new world. Perspectives in Medical Virology, 16, 161e177,(Emerging viruses in human populations).
Clement, J., Maes, P., & Van Ranst, M. (2007). Acute kidney injuryin emerging, non-tropical infections. Acta Clinica Belgica, 62(6),387e395.
Clement, J., Maes, P., van Ypersele de Strihou, C., van derGroen, G., Barrios, J. M., Verstraeten, W. W., et al. (2010).Beechnuts and outbreaks of nephropathia epidemica (NE): ofmast, mice and men. Nephrology, Dialysis, Transplantation, 25(6), 1740e1746.
Clement, J., Vercauteren, J., Verstraeten, W. W., Ducoffre, G.,Barrios, J.M., Vandamme,A.M., et al. (2009). Relating increasinghantavirus incidences to the changing climate: the mastconnection. International Journal of Health Geographics, 8(1).
Costa, A., Borgonovo, F., Leroy, T., Berckmans, D., & Guarino, M.(2009). Dust concentration variation in relation to animalactivity in a pig barn. Biosystems Engineering, 1(104), 118e124.
Ferentinos, K. P., & Albright, L. D. (2003). Fault detection anddiagnosis in deep-trough hydroponics using intelligentcomputational tools. Biosystems Engineering, 84(1), 13e30.
Kallio, E. (2003). Stability of Puumala-virus outside the host. In:2nd European meeting on viral zoonoses. St Raphae.
Lee, H. W., French, G. R., Lee, P. W., Baek, L. J., Tsuchiya, K., &Foulke, R. S. (1981). Observations on natural and laboratoryinfection of rodents with the etiologic agent of Koreanhemorrhagic fever. The American Society of Tropical Medicine andHygiene, 30(2), 477e482.
Linard, C., Lamarque, P., Heyman, P., Ducoffre, G., Luyasu, V.,Tersago, K., et al. (2007). Determinants of the geographicdistribution of Puumala virus and Lyme borreliosis infectionsin Belgium. International Journal of Health Geographics, 6(15).
May, R. M., & Anderson, R. M. (1979). Population biology ofinfectious diseases: part II. Nature, 280.
Meinzer, F. C. (1982). Models of steady state and dynamic gasexchange responses to vapour pressure and light in Douglasfir (Pseudotsuga menziesii) saplings. Oecologia, 55(3), 403e408.
Monroe, M., Morzunov, S. P., Johnson, A. M., Bowen, M. D.,Artsob, H., Yates, T., et al. (1999). Genetic diversity anddistribution of Peromyscus-borne hantaviruses in NorthAmerica. Emerging Infectious Diseases, 5, 75e86.
Nozum, E. O., Rossi, C. A., Stephenson, E. H., & Leduc, J. W. (1988).Aerosol transmission of hantaan and related viruses tolaboratory rats. The American Journal of Tropical Medicine andHygiene, 38(3), 636e640.
Olsson, G. E., Hjertqvist, M., Lundkvist, A., & Hornfeldt, B. (2009).Predicting high risk for human hantavirus infections, Sweden.Emerging Infectious Diseases, 15(1), 104e106.
Palo, R. T. (2009). Time series analysis performed on nephropathiaepidemica in humans of northern Sweden in relation to bankvole population dynamic and the NAO index. Zoonoses andPublic Health, 56(3), 150e156.
Patrick, W. (2001). Biological warfare scenarios. In S. I. Layne, T. J.Beugelsdijk, & C. K. Patel (Eds.), Firepower in the lab automationin the fight against infectious diseases and bioterrorism(pp. 215e224). Washington: Joseph Henry Press.
Rastetter, E. B. (1987). Analysis of community interactions usinglinear transfer function models. Ecological Modelling, 36(1e2),101e117.
Rozenfeld, F. M., Le Boulenge, E., & Rasmont, R. (1987). Urinemarking by male bank voles (Clethrionomys glareolus Schreber,1780; Microtidae, Rodentia) in relation to their social rank.Canadian Journal of Zoology, 65, 2594e2601.
Sauvage, F., Langlais, M., & Pontier, D. (2007). Predicting theemergence of human hantavirus disease using a combinationof viral dynamics and rodent demographic patterns.Epidemiology and Infection, 135, 46e56.
Sauvage, F., Langlais, M., Yoccoz, N. G., & Pontier, D. (2003,January). Modelling hantavirus in fluctuating populations ofbank voles: the role of indirect transmission on viruspersistence. The Journal of Animal Ecology, 1e13.
Sayre, N. F. (2008). The genesis, history, and limits of carrying.Annals of the Association of American Geographer, 98(1), 120e134.
Taylor, C. J., Pedregal, D. J., Young, P. C., & Tych, W. (2007).Environmental time series analysis and forecasting with thecaptain toolbox. Environmental Modelling & Software, 22(6),797e814.
Tersago, K., Servais, A., Heyman, P., Ducoffre, G., & Leirs, H.(2009). Hantavirus disease (nephropathia epidemica) inBelgium: effects of tree seed production and climate.Epidemiology Infection, 137, 250e256.
Thanh, V. T., Vranken, E., Van Brecht, A., & Berckmans, D. (2007).Data-based mechanistic modelling for controlling in threedimensions the temperature distribution in a room filled withobstacles. Biosystems Engineering, 98(1), 54e65.
Ushada, M., & Murase, H. (2006). Identification of a moss growthsystem using an artificial neural network model. BiosystemsEngineering, 94(2), 179e189.
Wolf, C., Sauvage, F., Pontier, D., & Langlais, M. (2006). A multi-patch epidemic model with periodic demography, direct andindirect transmission and variable maturation rate.Mathematical Population Studies, 13, 153e177.
Young, P. C. (1984). Recursive estimation and time series analysis.Berlin: Springer-Verlag.
Young, P. C., & Lees, M. (1993). The active mixing volume: a newconcept in modelling environmental systems. In V. Barnet, R.Turkman, & K. Feridun (Eds.), Statistics for the environment (pp.2e43). Chichester: John Wiley.
Young, P. C., & Wang, C. (1987). ldentification and estimation ofmultivariable dynamic systems. In J. O’Reilly (Ed.),Multivariable control for industrial applications (pp. 244e270).London: Peter.
Top Related
Copyright © 2022 FDOKUMEN

