







Bahasa
Halaman
Hukum


제 5 장 광합성과 호흡
1. 광합성과 호흡을 통해 태양에너지를 포획하고 방출한다. 1-1. 광합성 1-2. 호흡작용(respiration)
2. 대사는 일련의 경로로 많은 종류의 화학반응이 관여 한다. 2-1. 화학반응은 자유에너지 감소반응과 증가반응이 있다. 2-2. 산화ㆍ환원 반응은 세포대사에서 아주 중요하다. 2-3. 대사에서 화학반응은 경로를 가진다.
3. 광합성은 지구상 생명을 유지하기 위해 태양에너지를 포획한다. 3-1. 광합성은 다세포 생물이 살 수 있도록 원시 지구를 변화시켰다. 3-2. 광합성은 생물체에게 식량과 연료를 공급한다. 3-3. 태양에너지의 포획은 빛과 색소의 상호작용이다. 3-4. 광합성은 조류와 식물의 엽록체에서 일어난다. 3-5. 광합성은 빛 에너지를 당에 저장된 화학에너지로 전환한다. 3-6. 광호흡은 광합성의 비효율적 경로이다. 3-7. C4 식물과 CAM 식물은 광호흡 기작을 감소시킨다.
4. 호흡과 발효는 세포대사를 위해 에너지를 방출한다. 4-1. 호흡은 세포질과 미토콘드리아에서 일어난다. 4-2. 해당작용은 포도당을 두 분자의 피루브산으로 나눈다. 4-3. 피루브산은 CO2와 조효소 A가 결합한 아세틸로 분리된다. 4-4. 크렙스 회로에서 아세틸기는 두 분자의 이산화탄소로 파괴된다. 4-5. 전자전달 연쇄는 미토콘드리아 내막을 경계로 양성자 기울기가 일어난다. 4-6. 화학삼투와 산화적 인산화는 미토콘드리아에서 ATP를 생성한다.

제 5 장 광합성과 호흡
밀은 세계에서 가장 널리 재배되는 농작물인 동시에 세계 인류가 가장 많이 소비하는 식량이다. 밀은 벼나 옥수수처럼 낟알(grain)이라고 하는 단단한 건과를 생산한다.
재배 밀의 원산지는 근동지역이며 기원은 9,000년 이전으로 거슬러 올라간다. 야생 밀 × 고우트 그래스(goat grass) = Emmer 밀(Triticum turgidum)이 탄생. 약 8,000년 전 에머 밀 × 고우트 그래스 = 참밀 (Triticum aestivum)을 탄생시켰다. 참밀은 단백질 함량, 특히 글루텐(gluten) 함량이 높다. 참밀의 밀가루는 글루텐 때문에 반죽이 잘 늘어난다. 이 반죽에 효모나 베이킹 소다를 가하면 발효된 빵이 만들어 진다.
밀과 같은 식물이 그렇게 많은 인간의 식량을 공급할 수 있게 된 것은 두 가지 중요한 대사과정으로 인하여 이러한 기능을 수행할 수 있다.
광합성(photosynthesis) 호흡작용(respiration)
본 장에서는 먼저 두 가지 과정과 이들 대사과정에서 일어나는 화학반응에 대하여 개괄적으로 살펴보고자 한다.

1. 광합성과 호흡을 통해 태양에너지를 포획하고 방출한다.
1-1. 광합성 식물은 태양 에너지를 이용하여 이산화탄소와 물로부터 탄수화물과 같은 고에너지 유기화합물을 생산하기 위해 광합성을 이용한다. 이 과정은 다음의 식으로 요약할 수 있다. 6CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O 이 식에서 이산화탄소와 물은 반응물 혹은 기질이고, 산소, 물, 단당류는 생성물이다. 이 반응은 태양 에너지에 의해 추진되며 그 에너지는 단당류의 공유결합에 포획된다.
1-2. 호흡작용(respiration)
호흡은 대부분의 생물들이 단당류의 고유결합에 든 에너지를 방출하는 과정이다. 호흡과정 역시 다음 식으로 요약할 수 있다. C6H12O6 + 6O2 → 6CO2 + 6H2O 이러한 호흡과정에서 방출된 에너지의 상당량은 ATP분자에게 전달된다. 포획된 태양 에너지는 ATP의 형태로 많은 생화학 반응을 수행하는데 사용된다. 그림 5-1은 태양 에너지가 광합성과 호흡을 경유하여 세포대사를 추진하기 위한 ATP 생성에 이르기 까지 에너지 흐름의 과정을 요약하고 있다.
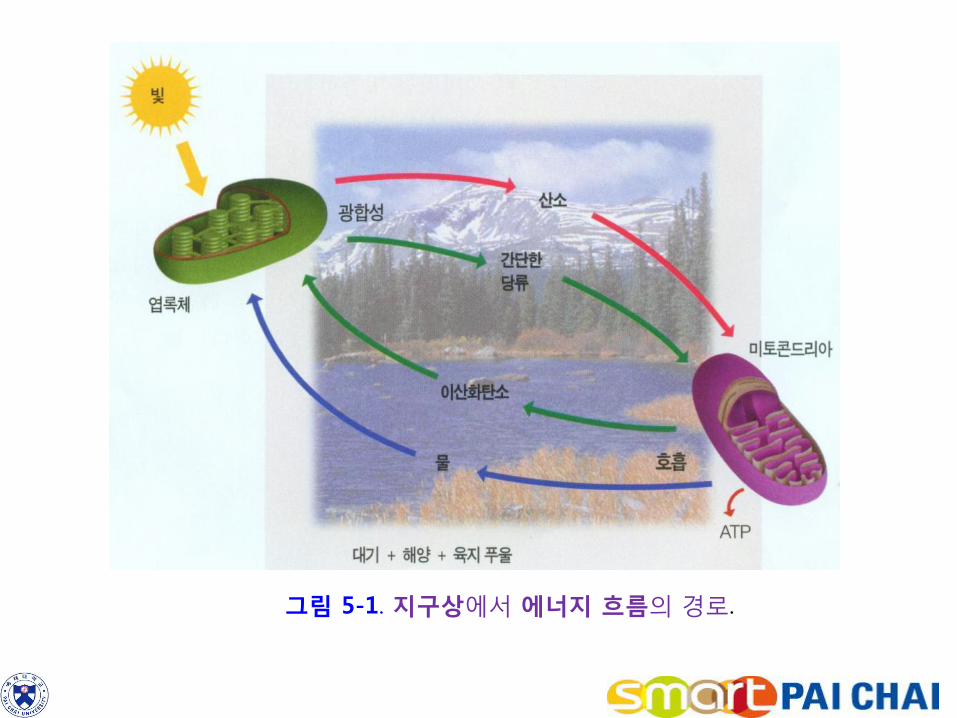
그림 5-1. 지구상에서 에너지 흐름의 경로.

2. 대사는 일련의 경로로 많은 종류의 화학반응이 관여 한다.
2-1. 화학반응은 자유에너지 감소반응과 증가반응이 있다.
2-2. 산화ㆍ환원 반응은 세포대사에서 아주 중요하다.
2-3. 대사에서 화학반응은 경로를 가진다.

2. 대사는 일련의 경로로 많은 종류의 화학반응이 관여 한다.
세포 대사(metabolism)를 구성하는 화학반응 즉 광합성과 호흡은 효소(enzyme)라
고 하는 생체촉매의 지시를 받는다.
대사의 화학반응들은 반응의 진행을 위해 에너지 투입을 필요로 하는 효소 촉매 반
응과 화학반응 진행과정에서 에너지 방출이 일어나는 반응이 있다.
세포대사의 많은 화학반응들에서는 반응 진행 동안 원자와 원자 간의 전자 이동이
일어난다.
이러한 반응을 산화-환원 반응이라고 하며, 광합성과 호흡에서 중요한 역할을
수행한다.

2-1. 화학반응은 자유에너지 감소반응과 증가반응이 있다. 1) 자유 에너지의 감소와 증가반응
화학반응이 일어나는 동안 에너지가 방출되는지 혹은 소모되는지에 따라 자유에너지 감소반응과 자유에너지 증가반응으로 구분한다. (1) 자유에너지 감소반응
기질의 퍼텐셜 에너지가 생성물보다 크다면 반응과정에서 에너지가 방출되며, 이것은 자유 에너지 감소반응이다.
에너지가 방출되는 과정은 계의 외부로부터 에너지를 투입하지 않아도 자발적으로 반응이 일어날 수 있다.
호흡은 자유에너지 방출과정이다.
(2) 자유에너지 증가반응
기질의 퍼텐셜 에너지가 생성물보다 적다면 반응을 진행시키기 위해 외부에서 에너지를 투입해야 하므로 이것은 자유 에너지 증가반응이다.
광합성의 많은 반응들과 기타 생합성 반응들은 자유 에너지 흡수과정이다.
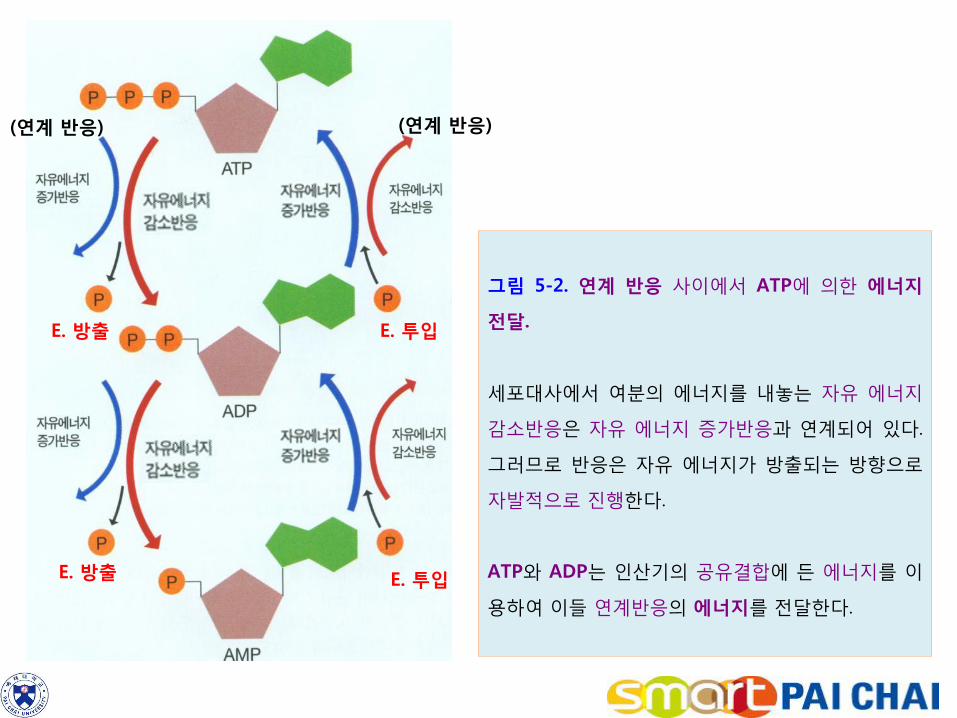
그림 5-2. 연계 반응 사이에서 ATP에 의한 에너지
전달.
세포대사에서 여분의 에너지를 내놓는 자유 에너지
감소반응은 자유 에너지 증가반응과 연계되어 있다.
그러므로 반응은 자유 에너지가 방출되는 방향으로
자발적으로 진행한다.
ATP와 ADP는 인산기의 공유결합에 든 에너지를 이
용하여 이들 연계반응의 에너지를 전달한다.
E. 투입 E. 방출
E. 방출 E. 투입
(연계 반응) (연계 반응)

2) ATP는 세포의 주된 에너지 운반체이다. (1) ATP의 에너지 운반
세포는 흔히 자유에너지 감소반응과 자유에너지 증가반응을 연계시킴으로서 자유 에너지 증가반응을 일으킨다.
대부분의 연계반응에서는 ATP가 두 반응 사이의 에너지 운반을 담당한다(그림 5-2). (2) ATP → ADP → AMP 로 인산결합이 절단되면 에너지를 방출한다.
ATP(adenosine triphosphate)는 세포의 주된 에너지 운반체이다.
ATP는 3개의 인산기(-PO4)를 갖는데 이 중에서 마지막 두 개의 인산기를 연결하는 공유결합들은 상당한 퍼텐셜 에너지를 갖고 있다. (3) 말단의 인산기(-PO4)는 단순히 제거되는 것이 아니라 에너지와 함께 다른 분자로 전달
인산기를 받은 분자는 에너지가 높아져 다른 반응에 참여할 수 있다.

2-2. 산화ㆍ환원 반응은 세포대사에서 아주 중요하다.
많은 화학반응에서 전자(e-)는 한 원자에서 다른 원자로 옮겨갈 수 있다. 이러한 화학반응을 산화-환원반응(oxidation-reduction reaction)이라고 한다. 1) 원자나 분자가 전자를 잃을 때 그 원자 혹은 분자는 산화되었다고 한다. 반대로 전자를 얻으면 환원이다(그림 5-3a).
산화ㆍ환원반응은 짝을 이루어 일어난다. 2) 세포의 대사반응에서 수소 이온은 전달된다. 이 때 수소 이온(H = e- + H+)은 자신의 퍼텐셜 에너지와 함께 전달되므로 전자가 함께 전달된다고 할 수 있다.
예를 들면 호흡에서 수소는 포도당으로부터 산소로 전달된다. 산소분자는 전자를 받아서 물로 환원되고, 반면에 포도당의 탄소원자들은 이산화탄소로 산화된다(그림 5-3b).
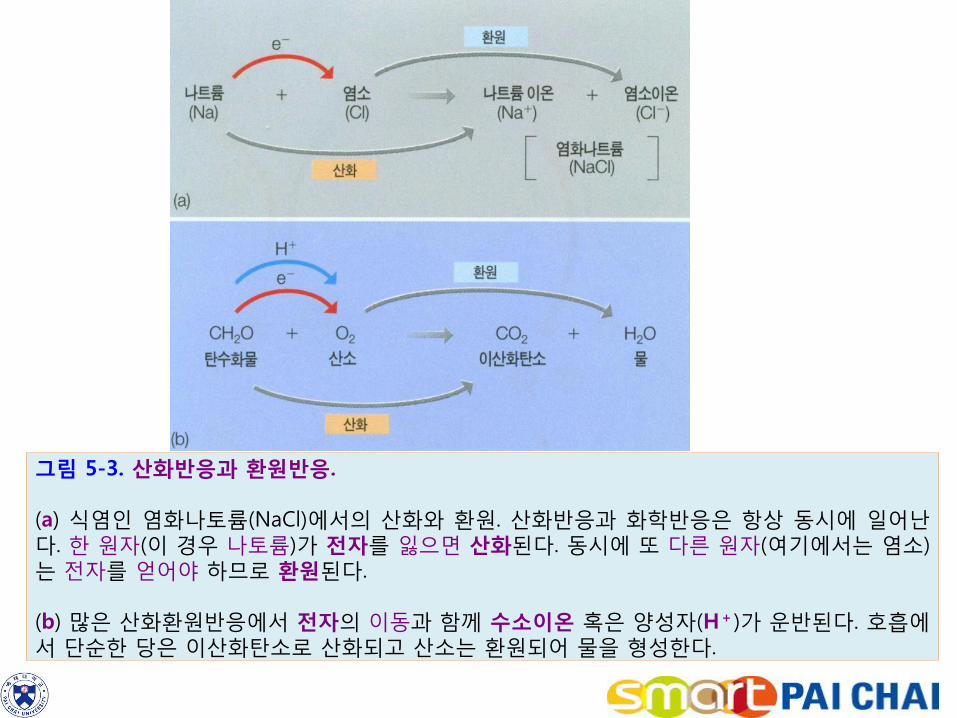
그림 5-3. 산화반응과 환원반응. (a) 식염인 염화나토륨(NaCl)에서의 산화와 환원. 산화반응과 화학반응은 항상 동시에 일어난다. 한 원자(이 경우 나토륨)가 전자를 잃으면 산화된다. 동시에 또 다른 원자(여기에서는 염소)는 전자를 얻어야 하므로 환원된다. (b) 많은 산화환원반응에서 전자의 이동과 함께 수소이온 혹은 양성자(H+)가 운반된다. 호흡에서 단순한 당은 이산화탄소로 산화되고 산소는 환원되어 물을 형성한다.

1. 산소와 수소에 의한 산화와 환원
산화 : 어떤 물질이 산소와 화합하거나,
수소화합물이 수소를 잃는 것
환원 : 어떤 물질이 수소와 결합하거나
산소화합물이 산소를 잃는 것
예)
2Mg + O2 → 2MgO
CH2O→CO2, 2H2O →2H2 + O2
O2→H2O, N2 + 3H3 → 2NH3
CuO + H2 → Cu +H2O
2. 전자 이동에 의한 산화와 환원
산화 : 원자, 분자, 이온 등이 전자를 잃는 것
환원 : 원자, 분자, 이온 등이 전자를 얻는 것
예)
Mg → Mg++ + 2e-
Cu+++ 2e- → Cu

3) 호흡이 일어나는 동안 포도당을 떠난 전자들은 직접 산소로 이동하지 않으며, 또한 광합성이 일어나는 동안 물을 떠난 전자들이 직접 이산화탄소로 이동하지 않는다. 이러한 전자와 양성자는 일부 에너지와 함께 전자 수용체 분자에게 전달된다. 전자수용체에는 NAD+(nicotinamide adenine dinucleotide)와 NADP+
(nicotinamide adenine dinucleotide phosphate)가 있다. NADP+는 광합성에서 주된 역할을 하며, NAD+는 호흡에서 중요한 역할을 한다.
4) 전자 수용체 분자들은 전자전달 연쇄라고 하는 계열에 무리지어 있다(그림 5-4). 전자전달 연쇄는 한 쪽 말단에서 전자를 받아 일련의 수용체 분자들을 거쳐 다른 쪽 말단으로 전달한다. 각 수용체 분자는 전자를 받으면 환원상태가 되고, 받은 전자를 다음 수용체 분자에게 넘기게 되면 자신은 산화한다. 따라서 전자전달 연쇄는 일련의 산화ㆍ환원 반응이라고 할 수 있다.
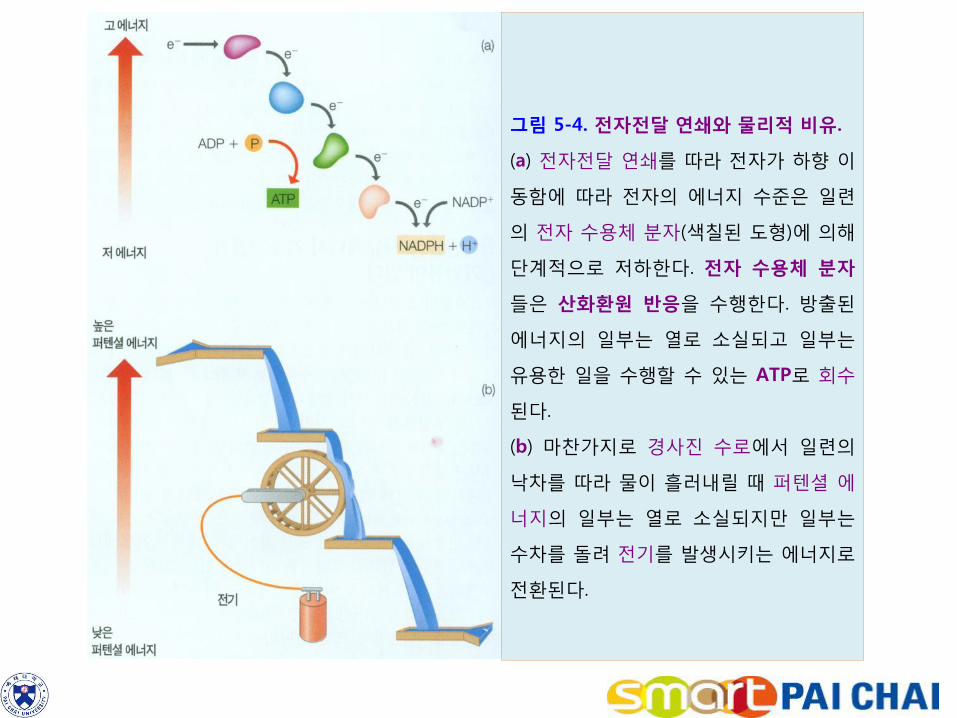
그림 5-4. 전자전달 연쇄와 물리적 비유.
(a) 전자전달 연쇄를 따라 전자가 하향 이
동함에 따라 전자의 에너지 수준은 일련
의 전자 수용체 분자(색칠된 도형)에 의해
단계적으로 저하한다. 전자 수용체 분자
들은 산화환원 반응을 수행한다. 방출된
에너지의 일부는 열로 소실되고 일부는
유용한 일을 수행할 수 있는 ATP로 회수
된다.
(b) 마찬가지로 경사진 수로에서 일련의
낙차를 따라 물이 흘러내릴 때 퍼텐셜 에
너지의 일부는 열로 소실되지만 일부는
수차를 돌려 전기를 발생시키는 에너지로
전환된다.

2-3. 대사에서 화학반응은 경로를 가진다. 1) 한 효소 반응의 생성물이 다음 효소반응의 기질이 된다(그림 5-5, 그림 5-6a).
2) 이러한 일련의 반응들은 대사경로를 구성한다. 대사 경로에는 3가지 기본 유형이 있다.
단순한 직선경로.
일부 경로에서는 분기점이 생겨나서 한 반응의 생성물이 두 경로 중 어느 한 쪽을
따라 진행되는 분기형 경로(그림 5-6b).
일부 경로에서는 경로의 출발물질이 마지막 단계에서 항상 재생되는 회로형
경로도 있다(그림 5-6c). 광합성과 호흡 과정에는 이러한 회로가 포함되어 있다.
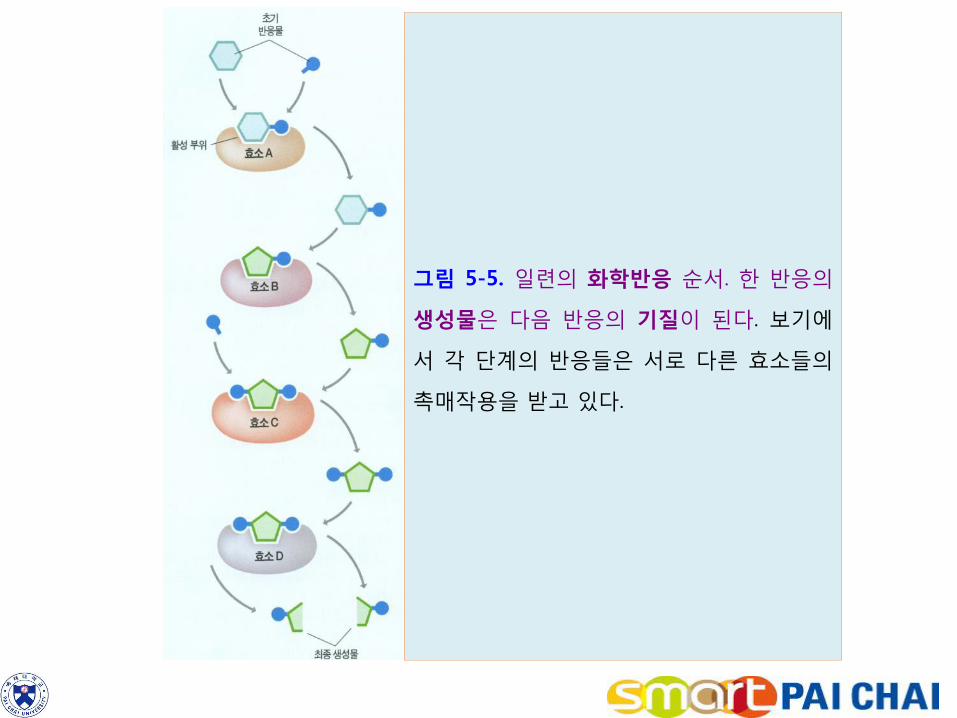
그림 5-5. 일련의 화학반응 순서. 한 반응의
생성물은 다음 반응의 기질이 된다. 보기에
서 각 단계의 반응들은 서로 다른 효소들의
촉매작용을 받고 있다.
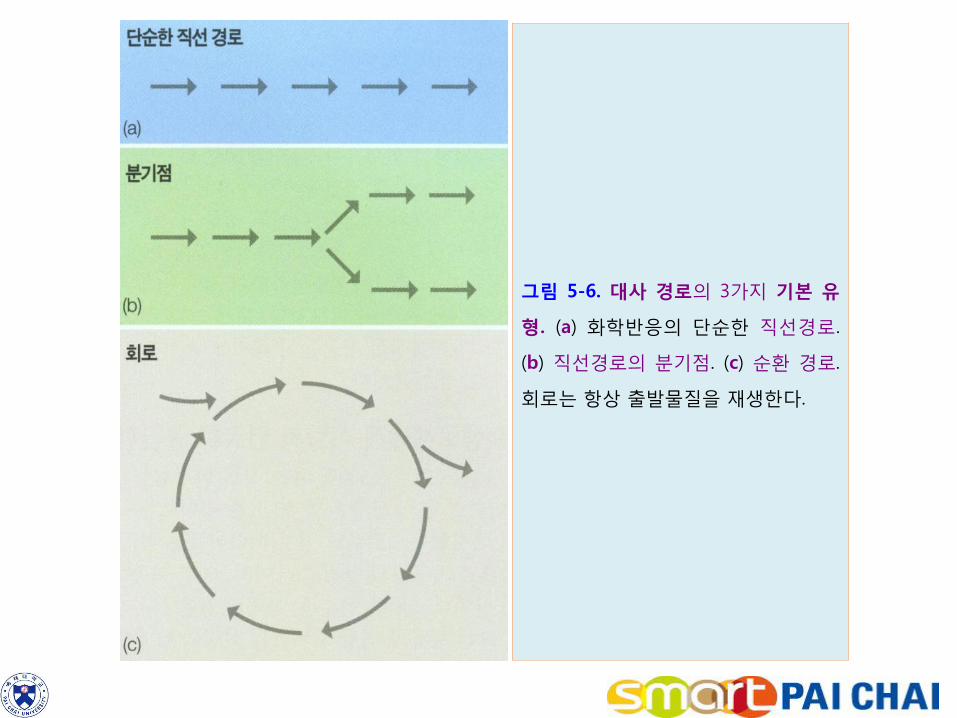
그림 5-6. 대사 경로의 3가지 기본 유
형. (a) 화학반응의 단순한 직선경로.
(b) 직선경로의 분기점. (c) 순환 경로.
회로는 항상 출발물질을 재생한다.

3. 광합성은 지구상 생명을 유지하기 위해 태양에너지를 포획한다.
3-1. 광합성은 다세포 생물이 살 수 있도록 원시 지구를 변화시켰다.
3-2. 광합성은 생물체에게 식량과 연료를 공급한다.
3-3. 태양에너지의 포획은 빛과 색소의 상호작용이다.
3-4. 광합성은 조류와 식물의 엽록체에서 일어난다.
3-5. 광합성은 빛 에너지를 당에 저장된 화학에너지로 전환한다.
3-6. 광호흡은 광합성의 비효율적 경로이다.
3-7. C4 식물과 CAM 식물은 광호흡 기작을 감소시킨다.
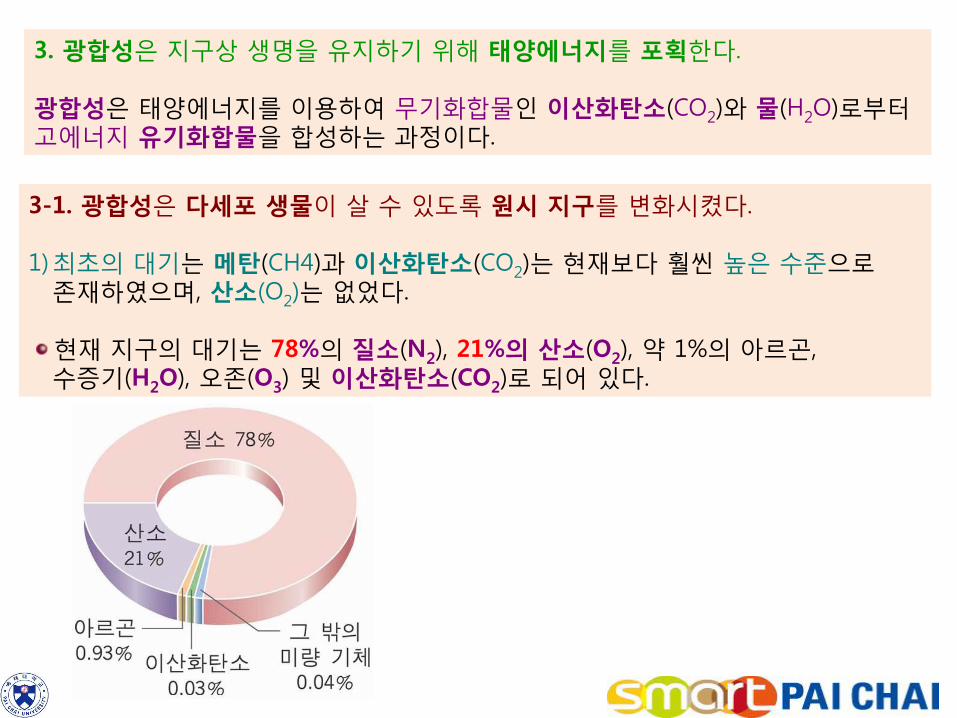
3. 광합성은 지구상 생명을 유지하기 위해 태양에너지를 포획한다. 광합성은 태양에너지를 이용하여 무기화합물인 이산화탄소(CO2)와 물(H2O)로부터 고에너지 유기화합물을 합성하는 과정이다.
3-1. 광합성은 다세포 생물이 살 수 있도록 원시 지구를 변화시켰다. 1)최초의 대기는 메탄(CH4)과 이산화탄소(CO2)는 현재보다 훨씬 높은 수준으로 존재하였으며, 산소(O2)는 없었다.
현재 지구의 대기는 78%의 질소(N2), 21%의 산소(O2), 약 1%의 아르곤, 수증기(H2O), 오존(O3) 및 이산화탄소(CO2)로 되어 있다.

2) 초기의 해양 광합성 미생물에서 육지의 복잡한 다세포생물의 탄생 약 35억 년 전 원시지구의 해양에서 무리를 이루었던 광합성의 미생물들은 풍부한 CO2를 섭취한 후 태양에너지를 사용하여 CO2를 간단한 탄수화물로 환원시켰다. 태양에너지는 탄수화물의 화학결합에 저장되었으며 반응의 부산물로 산소(O2)가 발생하였다. 장구한 세월 동안 대기의 CO2 농도는 점차 저하되고 O2의 농도는 상승하였다. 대기 중의 일부 산소는 오존(O3)을 형성하였다. O2와 O3농도가 상승하면서 초기의 광합성 미생물은 표층으로 이동하게 되었으며, 마침내 6억 년 전 O2와 O3은 해양의 복잡한 다세포 생물이 폭발적으로 번성할 수 있을 만큼 높은 농도로 축적되었다. 복잡한 다세포생물은 그 후 약 1억 5천만 년이 지나(즉 4억 5천만 년 전) 육지를 침입하기에 이르렀다.

3-2. 광합성은 생물체에게 식량과 연료를 공급한다.
광합성 생물들은 매년 2,500억 톤 이상의 탄수화물을 생산한다. 이러한 연 생산량은 약 50만종의 광합성 생물의 대사활동으로 얻어지며, 지구상의 나머지 300만종의 생물을 먹여 살린다. 광합성은 식량공급 이외에도 목화나 린넨(linen, 亞麻의 실로 짠 얇은 직물을 )같은 직물은 식물의 섬유에서 얻어지며, 건축, 가구생산 및 종이 산업 등은 목재를 이용한다. 광합성 산물은 또한 향신료, 향수, 의약품, 화장품, 왁스, 기름, 고무 등을 생산하는데도 쓰인다. 광합성은 인간이 사용하는 다량의 연료를 공급한다. 石炭, 石油, 천연가스와 같은 화석연료는 모두 과거의 광합성 생물에 포획된 태양에너지라고 할 수 있다. 현대인의 생활은 과거 광합성의 산물 및 현재 광합성의 산물에 의해 영위된다.
석유를 뜻하는 영어단어인 'Petroleum'도 그리스어 '페트로스(petros, 바위)', 라틴어 'Petra(바위,돌)', 그리고 라틴어 '올레움
'(oleum,기름)에서 유래되었습니다. 말그대로 바위와 기름을 합쳐서 만들어진 단어입니다(石油) , 無機成因說( inorganic
theory)??, 有機成因說( organic theory)??
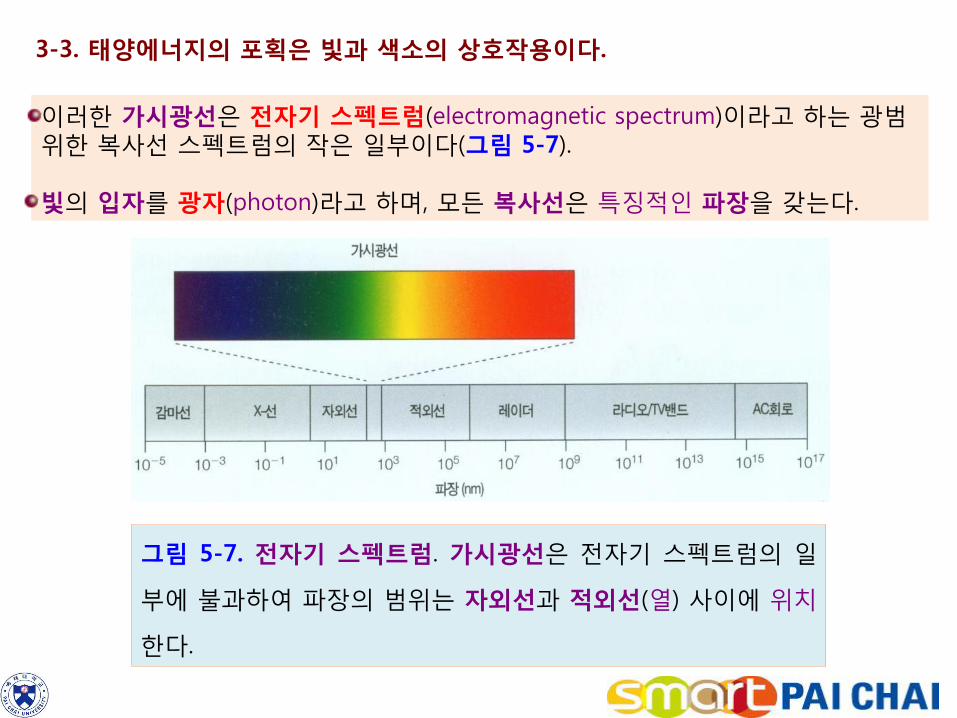
3-3. 태양에너지의 포획은 빛과 색소의 상호작용이다.
이러한 가시광선은 전자기 스펙트럼(electromagnetic spectrum)이라고 하는 광범위한 복사선 스펙트럼의 작은 일부이다(그림 5-7). 빛의 입자를 광자(photon)라고 하며, 모든 복사선은 특징적인 파장을 갖는다.
그림 5-7. 전자기 스펙트럼. 가시광선은 전자기 스펙트럼의 일
부에 불과하여 파장의 범위는 자외선과 적외선(열) 사이에 위치
한다.

1) 가시광선
가시광선은 400nm(보라색)부터 700nm(적색)의 범위에 있다. 파장이 길수록 광자 혹은 에너지 파장이 운반하는 에너지 함량은 낮다. 가시광선의 범위에서 가장 파장이 짧은 보라색광이 높은 에너지를 가지며, 가장 긴 파장의 적색광은 낮은 에너지 함량을 갖는다. 보라색보다 짧은 파장의 자외선이나 x선은 복잡한 유기분자를 손상시킬 만큼의 높은 에너지를 갖고 있다. 적외선의 파장을 가진 빛은 물에 의해 흡수되어 유기물 분자의 열이나 운동을 증가시킨다. 가시광선은 유기물 분자의 전자를 들뜨게 할 수 있는 적정한 량의 에너지를 가지고 있기 때문에 광합성에서는 가시광선을 사용한다. 전자가 최저 에너지 상태에 있을 때 이것을 바닥상태(ground state)라고 한다(그림 5-8a). 전자가 가시광선으로부터 에너지를 흡수하면 원자핵 주변의 더 높은 궤도로 올라가는데 (그림 5-8b), 이 상태를 여기상태(exited state)라고 한다. 여기상태에 있는 전자의 에너지가 진행할 수 있는 경로에는 3 가지가 있다 . a. 전자의 에너지는 열이나 빛으로 방출되고 낮은 에너지상태로 복귀한다(그림 5-8c). b. 전자가 보다 낮은 에너지 준위로 내려오는 것은 같으나, 에너지가 다른 분자에 전달되어 그 분자를 들뜨게 하는 경로(그림 5-8d). c. 여기된 전자 자체가 이동되어 전자수용체 분자에게 전달되는 경로(그림 5-8e) 등 이다.
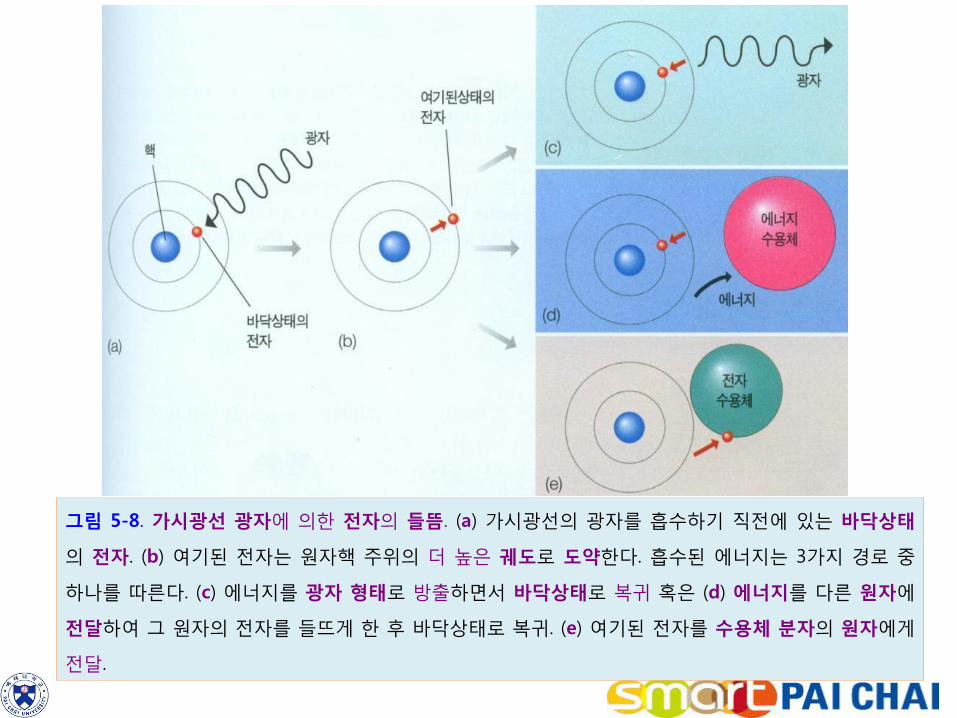
그림 5-8. 가시광선 광자에 의한 전자의 들뜸. (a) 가시광선의 광자를 흡수하기 직전에 있는 바닥상태
의 전자. (b) 여기된 전자는 원자핵 주위의 더 높은 궤도로 도약한다. 흡수된 에너지는 3가지 경로 중
하나를 따른다. (c) 에너지를 광자 형태로 방출하면서 바닥상태로 복귀 혹은 (d) 에너지를 다른 원자에
전달하여 그 원자의 전자를 들뜨게 한 후 바닥상태로 복귀. (e) 여기된 전자를 수용체 분자의 원자에게
전달.

2) 색소
색소란 빛을 흡수하는 물질을 말한다. 엽록소 a(chlorophyll a)는 식물 엽록체에 가장 풍부한 광 흡수 색소이다. 엽록소 a는 녹색 광을 반사하므로 식물이 녹색으로 보이는 것은 이 때문이다. 색소의 빛 흡수 패턴을 흡수 스펙트럼(absorption spectrum)이라고 한다. 엽록소 a의 흡수 스펙트럼을 보면 가시광선에서 주로 적색광과 청색 및 보라색 영역을 흡수함을 알 수 있다(그림 5-10). 엽록소 a가 이러한 광선을 흡수하면 전자가 들뜨게 되며, 여기된 전자는 엽록소 분자를 떠나 전자 수용체에 전달된다. 엽록소 b와 카로티노이드는 일종의 보조 색소들이다. 이러한 보조 색소들의 흡수 스펙트럼은 엽록소 a와 다르며, 보조 색소들은 광합성에 필요한 가시광선의 범위를 확장시킨다(그림 5-10 참조). 이들이 흡수한 에너지는 엽록소 a에 전달된다.
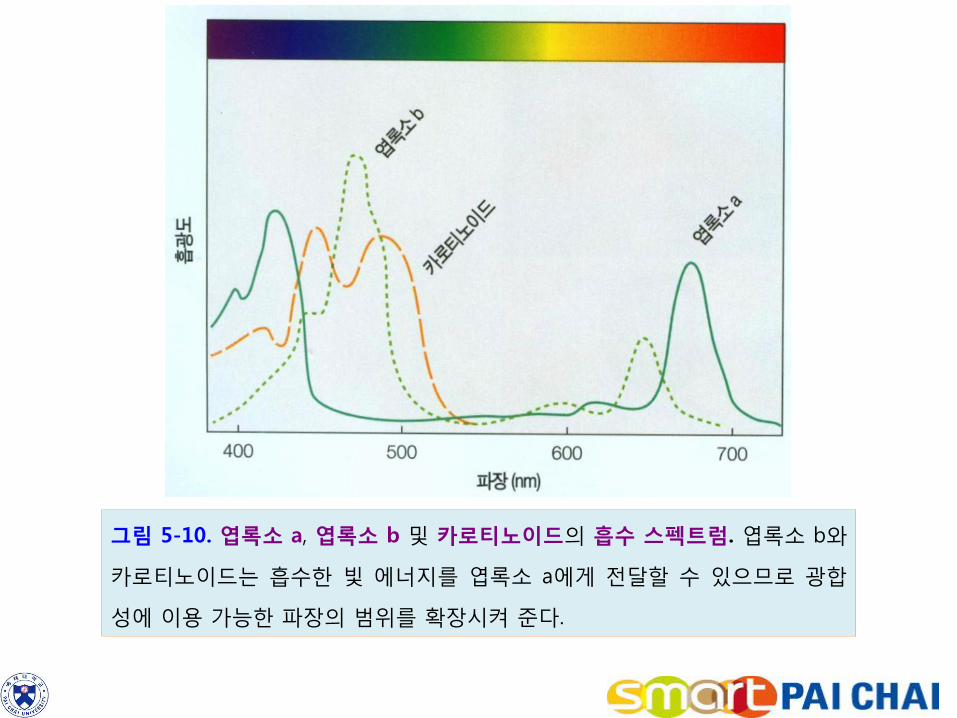
그림 5-10. 엽록소 a, 엽록소 b 및 카로티노이드의 흡수 스펙트럼. 엽록소 b와
카로티노이드는 흡수한 빛 에너지를 엽록소 a에게 전달할 수 있으므로 광합
성에 이용 가능한 파장의 범위를 확장시켜 준다.

1) 틸라코이드
틸라코이드는 얇고 평평한 판 모양의 주머니들이 망상구조를 이루어 교차되어 있으며, 틸라코이드에서 태양광선의 에너지가 ATP 및 NADPH의 화학에너지 형태로 전환된다. 2) 스트로마 엽록체의 내부를 채우고 있는 점액성의 액체로 기질이라고 한다. 스트로마에서는 화학에너지 형태로 전환된 에너지가 CO2를 당으로 환원시키는데 사용된다. 3) 엽록체 식물의 엽록체는 주로 엽육세포 내에 위치한다(그림 5-11 참조). 광합성이 진행되기 위해서는 잎의 엽육세포로 물 , 이산화탄소 (CO2), 태양광이 모두 공급되어야 한다.
3-4. 광합성은 조류와 식물의 엽록체에서 일어난다. 식물과 조류에서 광합성은 엽록체에서 일어난다(그림 5-11). 엽록체 내부는 스트로마라는 기질 속에 얇고 평평한 판 모양의 틸라코이드가 들어 차 있다.
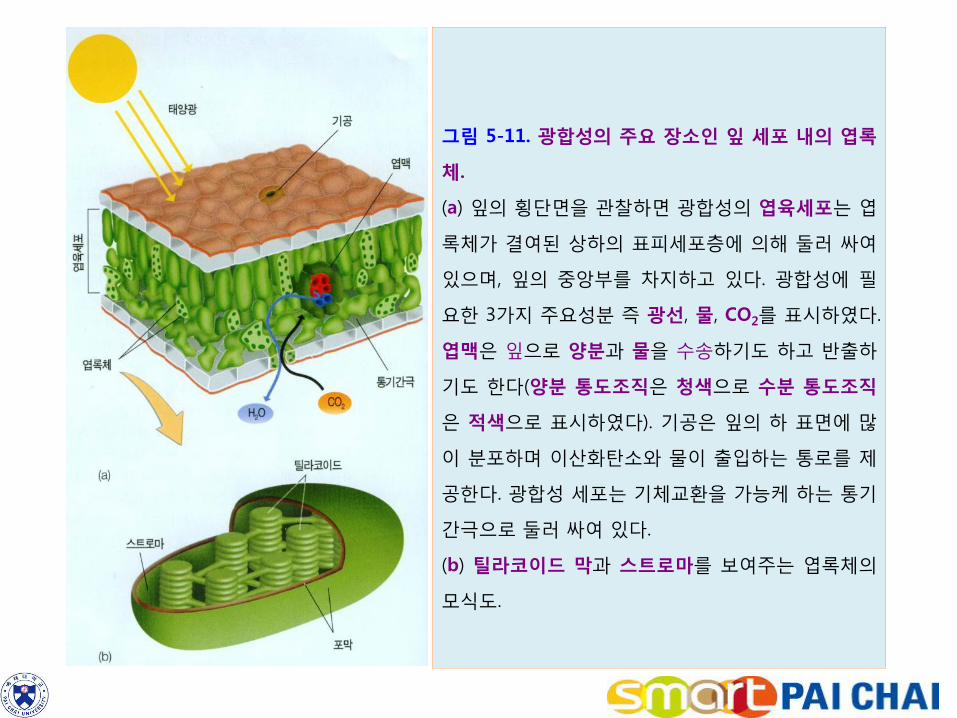
그림 5-11. 광합성의 주요 장소인 잎 세포 내의 엽록
체.
(a) 잎의 횡단면을 관찰하면 광합성의 엽육세포는 엽
록체가 결여된 상하의 표피세포층에 의해 둘러 싸여
있으며, 잎의 중앙부를 차지하고 있다. 광합성에 필
요한 3가지 주요성분 즉 광선, 물, CO2를 표시하였다.
엽맥은 잎으로 양분과 물을 수송하기도 하고 반출하
기도 한다(양분 통도조직은 청색으로 수분 통도조직
은 적색으로 표시하였다). 기공은 잎의 하 표면에 많
이 분포하며 이산화탄소와 물이 출입하는 통로를 제
공한다. 광합성 세포는 기체교환을 가능케 하는 통기
간극으로 둘러 싸여 있다.
(b) 틸라코이드 막과 스트로마를 보여주는 엽록체의
모식도.
3-5. 광합성은 빛 에너지를 당에 저장된 화학에너지로 전환한다.
광합성에서 이산화탄소(CO2)와 물(H2O)은 기질 혹은 반응물이고, 산소(O2)와 당
(C6H12O6)은 반응의 생성물이다. 종합 반응은 아래 식과 같다.
6CO2 + 12H2O → C6H12O6 + 6H2O
이 방정식만 가지고는 광합성이 어떻게 진행되는가에 대해서는 알 수 없다.
광합성은 실제로 2가지 반응에 의해서 즉 명반응(light reaction)과
탄소고정반응(carbonfixation reaction)으로 이루어져 있다.

1) 명반응
(1)명반응은 광계Ⅰ(photosynthesisⅠ) 및 광계Ⅱ(photosynthesisⅡ)라고 하는 2가지 색소계에 의하여 수행된다.
두 개의 광계는 모두 틸라코이드 막에 들어 있으며, 각기 2가지 성분을 포함한다. 즉 보조색소인 안테나 복합체와 반응중심이 그것인데 안테나 복합체는 빛 에너지를 포획하여 반응중심으로 전달한다. 각 반응중심에는 2개의 특별한 엽록소 a 분자가 들어 있는데, 이들은 전자를 전자수용체에게 전달할 수 있는 유일한 분자들이다. 전자전달 연쇄는 2개의 광계를 연결하며, 전자전달 연쇄의 말단에서 최종적으로 NADP+를 NADPH로 환원시킨다. 광계Ⅱ와 광계Ⅰ에서 전자전달 연쇄들의 물리적 위치는 엽록체의 틸라코이드이다(그림 5-14). (2) 물의 광분해는 틸라코이드 내부에서 광계Ⅱ에 가까운 내부공간 일명 틸라코이드 루멘에서 일어난다. 광분해는 루멘 내부에 양성자(H+)의 축적을 야기한다. 물이 분해되면서 방출된 전자들은 P680의 엽록소 a를 떠난 전자들을 대체한다. H2O → 2H+
+ 2e- + 1/2O2

(3) 광계Ⅱ(photosynthesisⅡ)
광계Ⅱ에서 반응이 시작되는 것은 안테나 색소가 빛의 광자를 흡수 한 후 에너지를 분자와 분자를 통해 이동시켜 마침내 반응 중심의 P680이라고 하는 특별한 엽록소 a 분자에게 넘겨주면서 부터이다(그림 5-13). P680분자가 여기되면 에너지가 높아진 전자는 P680을 떠나 일차 전자수용체와 결합함으로서 최초로 전자전달 연쇄에 진입한다(P680의 P는 pigment를 의미한다). 반응 중심에서 여기된 상태로 P680을 떠난 전자들은 일차 전자수용체에 전달되어 첫 번째 전자전달 연쇄에 진입한 후 플라스토퀴논(plastoquinone, PQ)이라고 하는 일련의 분자들을 거친다. 이들 전자는 이 후 시토크롬 복합체를 지나 플라스토시아닌(plastocyanin)이라고 하는 구리 함유 단백질에 도달한다. 시토크롬 복합체에는 몇 가지 서로 다른 시토크롬 분자들이 있으며, 전자운반 기능을 하는 철 원자를 갖고 있다. 전자는 플라스토시아닌을 떠나 광계Ⅰ로 들어간다.
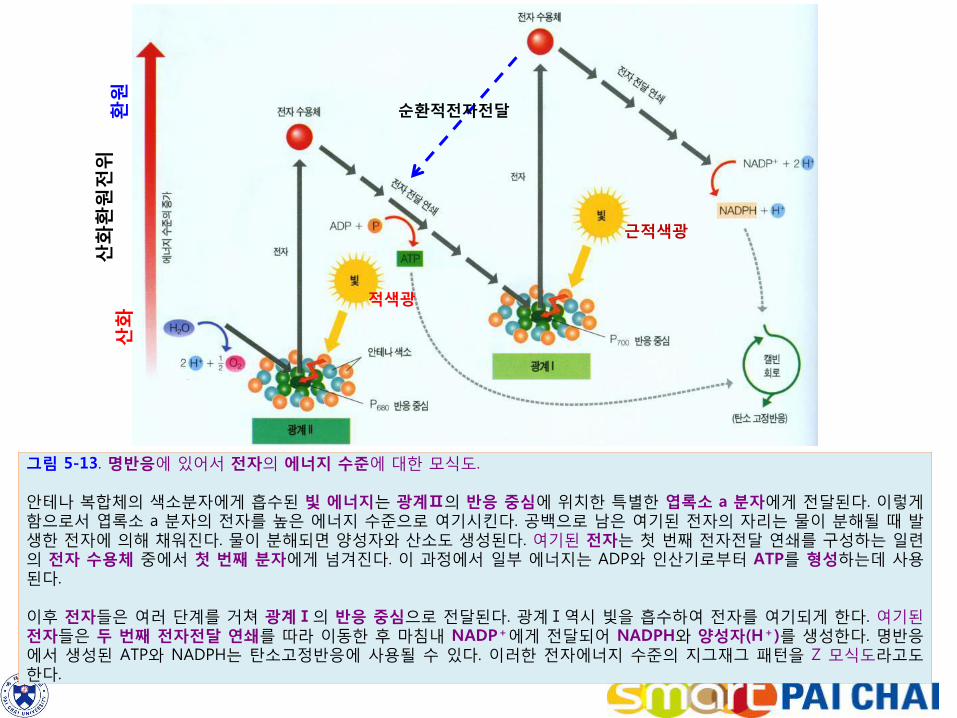
그림 5-13. 명반응에 있어서 전자의 에너지 수준에 대한 모식도. 안테나 복합체의 색소분자에게 흡수된 빛 에너지는 광계Ⅱ의 반응 중심에 위치한 특별한 엽록소 a 분자에게 전달된다. 이렇게 함으로서 엽록소 a 분자의 전자를 높은 에너지 수준으로 여기시킨다. 공백으로 남은 여기된 전자의 자리는 물이 분해될 때 발생한 전자에 의해 채워진다. 물이 분해되면 양성자와 산소도 생성된다. 여기된 전자는 첫 번째 전자전달 연쇄를 구성하는 일련의 전자 수용체 중에서 첫 번째 분자에게 넘겨진다. 이 과정에서 일부 에너지는 ADP와 인산기로부터 ATP를 형성하는데 사용된다. 이후 전자들은 여러 단계를 거쳐 광계Ⅰ의 반응 중심으로 전달된다. 광계Ⅰ역시 빛을 흡수하여 전자를 여기되게 한다. 여기된 전자들은 두 번째 전자전달 연쇄를 따라 이동한 후 마침내 NADP+에게 전달되어 NADPH와 양성자(H+)를 생성한다. 명반응에서 생성된 ATP와 NADPH는 탄소고정반응에 사용될 수 있다. 이러한 전자에너지 수준의 지그재그 패턴을 Z 모식도라고도 한다.
적색광
근적색광
산화
환원
전위
산
화
환원
순환적전자전달
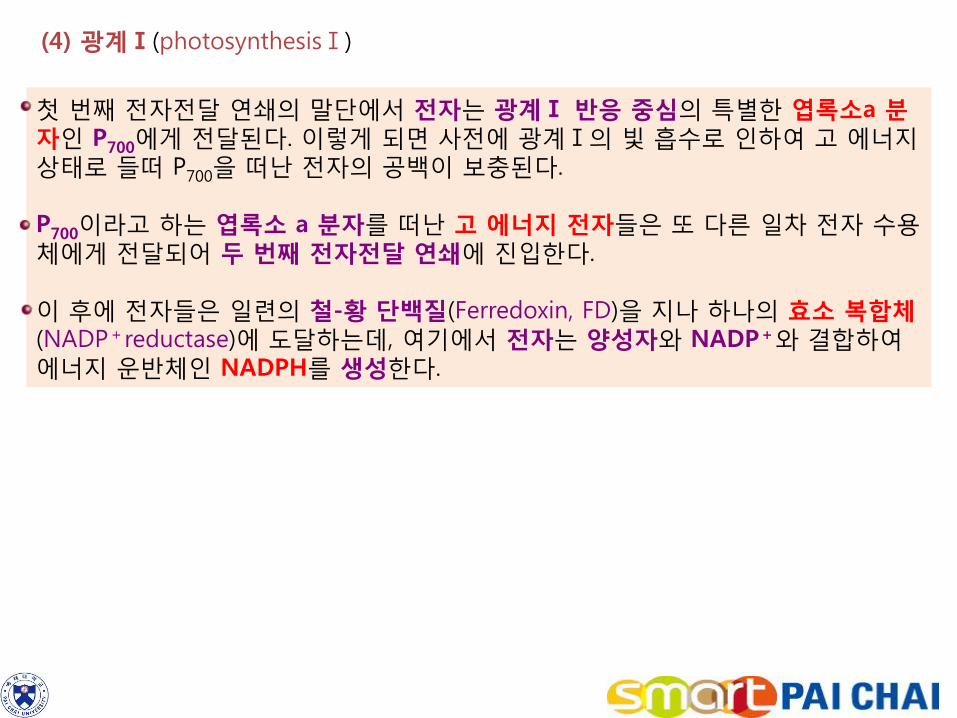
(4) 광계Ⅰ(photosynthesisⅠ)
첫 번째 전자전달 연쇄의 말단에서 전자는 광계Ⅰ 반응 중심의 특별한 엽록소a 분자인 P700에게 전달된다. 이렇게 되면 사전에 광계Ⅰ의 빛 흡수로 인하여 고 에너지 상태로 들떠 P700을 떠난 전자의 공백이 보충된다. P700이라고 하는 엽록소 a 분자를 떠난 고 에너지 전자들은 또 다른 일차 전자 수용체에게 전달되어 두 번째 전자전달 연쇄에 진입한다. 이 후에 전자들은 일련의 철-황 단백질(Ferredoxin, FD)을 지나 하나의 효소 복합체(NADP+reductase)에 도달하는데, 여기에서 전자는 양성자와 NADP+와 결합하여 에너지 운반체인 NADPH를 생성한다.
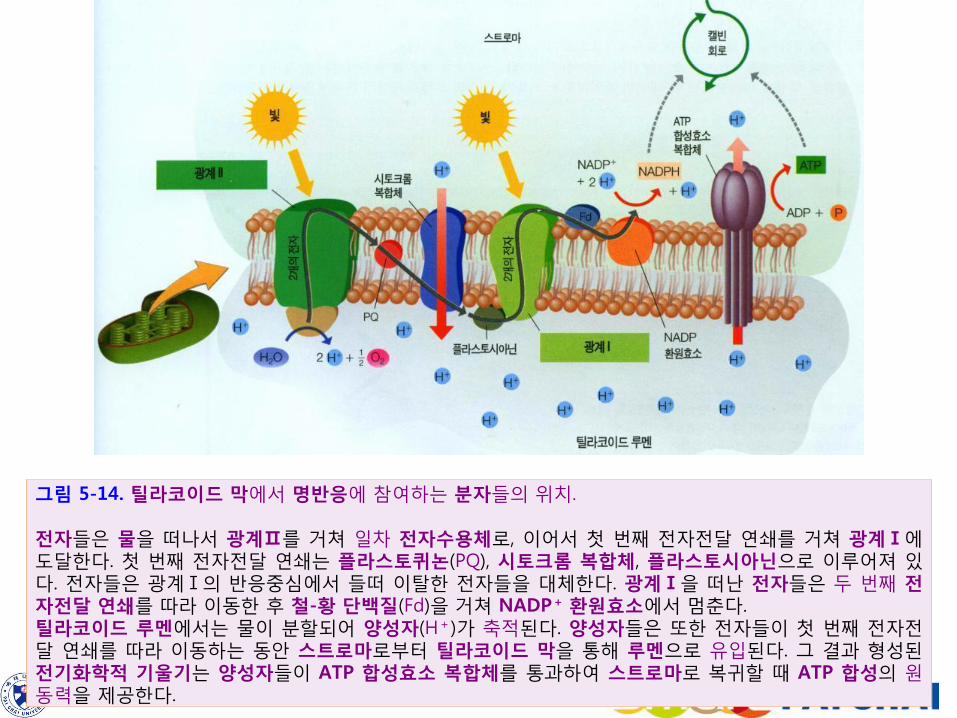
그림 5-14. 틸라코이드 막에서 명반응에 참여하는 분자들의 위치. 전자들은 물을 떠나서 광계Ⅱ를 거쳐 일차 전자수용체로, 이어서 첫 번째 전자전달 연쇄를 거쳐 광계Ⅰ에 도달한다. 첫 번째 전자전달 연쇄는 플라스토퀴논(PQ), 시토크롬 복합체, 플라스토시아닌으로 이루어져 있다. 전자들은 광계Ⅰ의 반응중심에서 들떠 이탈한 전자들을 대체한다. 광계Ⅰ을 떠난 전자들은 두 번째 전자전달 연쇄를 따라 이동한 후 철-황 단백질(Fd)을 거쳐 NADP+ 환원효소에서 멈춘다. 틸라코이드 루멘에서는 물이 분할되어 양성자(H+)가 축적된다. 양성자들은 또한 전자들이 첫 번째 전자전달 연쇄를 따라 이동하는 동안 스트로마로부터 틸라코이드 막을 통해 루멘으로 유입된다. 그 결과 형성된 전기화학적 기울기는 양성자들이 ATP 합성효소 복합체를 통과하여 스트로마로 복귀할 때 ATP 합성의 원동력을 제공한다.

(5) 요약하면 총 3분자의 ATP와 2분자의 NADPH를 생성한다.
물 분해로 방출된 전자는 광계Ⅱ의 반응중심을 거쳐 첫 번째 전자전달 연쇄를 따라 하향 이동한 후 광계Ⅰ의 반응 중심으로 들어가고, 이 후 두 번째 전자전달 연쇄를 따라 이동하여 마침내 NADP+와 H+와 결합함으로써 NADPH를 생성한다. 반응중심에서 8개의 광자가 흡수될 때마다 비순환적 전자전달을 통해 4개의 전자가 이동하고 총 3분자의 ATP와 2분자의 NADPH를 생성한다(2개의 전자 당 1개의 NADPH).

2) 화학침투와 광인산화는 엽록소에서 ATP를 만든다. (1) 여기된 전자가 전자전달 연쇄를 통과하여 내려옴에 따라 전자 에너지의 일부는
스트로마의 양성자를 틸라코이드 안의 공간(루멘)으로 유입시키는데 쓰인다.
따라서 루멘 내의 양성자의 축적이 일어나므로 내부는 더욱 산성화된다. (2) 틸라코이드막 내의 양성자 농도는 전기화학적 기울기를 형성한다. 이 기울기의 “전기적” 부분은 양성자의 (+)전하에 의해 발생하며, “화학적” 부분은 루멘을 더욱 산성화시키는 수소 이온(H+) 농도에 기인한다. 그 결과로 형성된 틸라코이드 막 내외의 전기화학적 기울기는 마치 건전지처럼 작용하는 퍼텐셜 에너지에 해당한다. (3) 이 퍼텐셜 에너지는 ATP 생산을 추진하는데 쓰일 수 있다. 양성자가 화학삼투라고 하는 기작에 의해 ATP의 합성효소 복합체의 내부에 있는 채널을 통과하여 스트로마로 되돌아간다. ATP의 합성효소 복합체의 채널을 따라 흘러간 양성자는 ADP와 인산으로부터 ATP를 합성한다. 현재 연구된 바에 따르면 ATP의 합성효소 복합체를 따라 이동한 양성자 4개당 하나의 ATP가 형성되는 것으로 알려져 있다. 이 와 같 은 유 형 의 인 산 화 는 그 에 너 지 가 태 양 에 서 유 래 하 기 때 문 에 광인산화(photophosphorylation)이라고 한다.

3) 탄소고정반응은 이산화탄소를 간단한 당으로 환원한다. 명반응에서 생성된 ATP와 NADPH의 화학에너지는 이산화탄소를 당으로 환원시키는 탄소고정반응에 사용된다. 이러한 일련의 반응을 캘빈회로(Calvin cycle)라고 한다. (1) RuBP는 CO2와 결합하여 2분자의 PGA를 형성한다.
a. 캘빈회로의 시작은 5탄소 단당류인 ribulose 1,5-bisphosphate(RuBP)라고 하
는 화합물이다. 이산화탄소는 루비스코(rubisco)라고 하는 효소의 작용에 의해 RuBP와 결합한다.
3 RuBP(C5H8O5ㆍⓟㆍⓟ) + 3 CO2 + 3H2O → 6 PGA(C3H5O4)ㆍⓟ b. 캘빈회로에서 광합성 최초의 안정산물이 3탄소 화합물인 3-포스포글리세르산
(3-phosphoglycerate, PGA)이므로 이 대사경로를 C3 경로(C3 pathway)라고 한다. 가장 일반적인 탄소고정경로로서 식물 종류 중 약 85%가 이 경로만을 이용한다.
c. 감자, 콩을 비롯하여 밀, 호밀, 귀리 등의 곡류들은 C3 경로를 이용하므로 C3 식
물(C3 plants)이라고 한다.
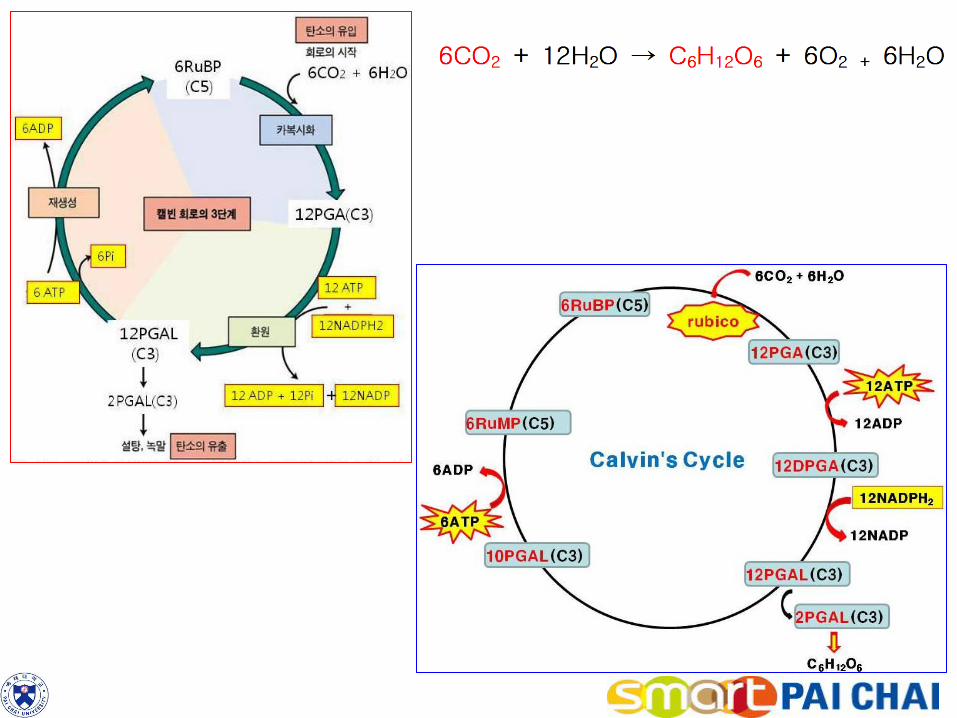
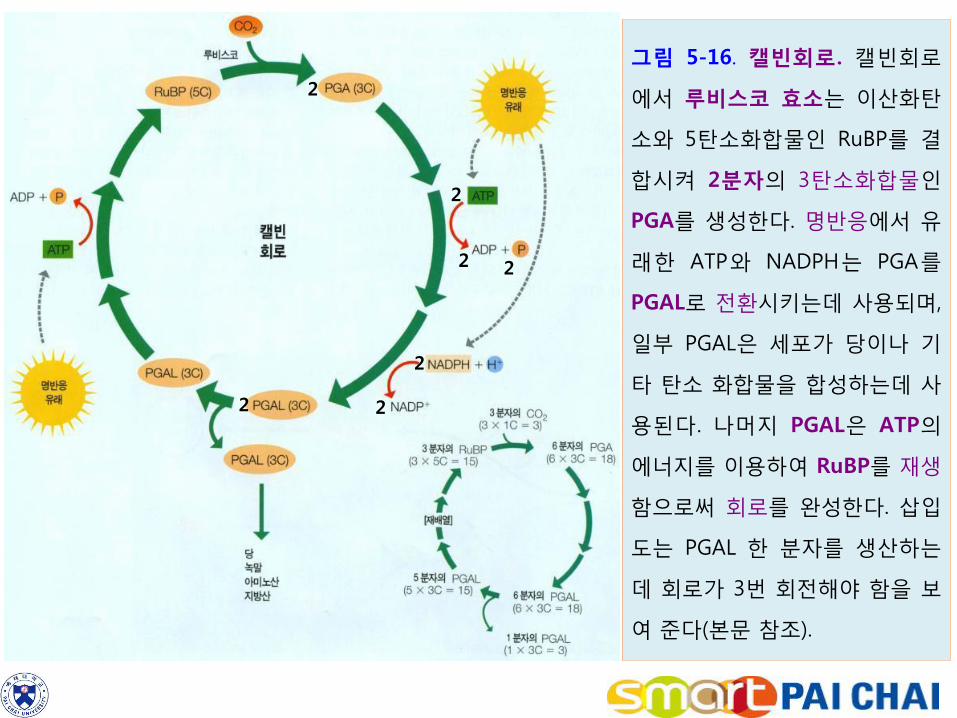
그림 5-16. 캘빈회로. 캘빈회로
에서 루비스코 효소는 이산화탄
소와 5탄소화합물인 RuBP를 결
합시켜 2분자의 3탄소화합물인
PGA를 생성한다. 명반응에서 유
래한 ATP와 NADPH는 PGA를
PGAL로 전환시키는데 사용되며,
일부 PGAL은 세포가 당이나 기
타 탄소 화합물을 합성하는데 사
용된다. 나머지 PGAL은 ATP의
에너지를 이용하여 RuBP를 재생
함으로써 회로를 완성한다. 삽입
도는 PGAL 한 분자를 생산하는
데 회로가 3번 회전해야 함을 보
여 준다(본문 참조).
2
2
2 2
2
2 2

(2) 일련의 화학반응을 거쳐 PGA는 3탄소 분자인 포스포글리세르알데히드(phosphoglycer-aldehyde, PGAL)로 환원된다.
6 PGA + 6 ATP → 6 DPGA + 6 ADP 6 DPGA + 6 NADPH → 6 PGAL + NADP+
PGAL이 진정한 광합성의 산물이다. 대부분의 PGAL은 재배열을 거쳐 RuBP를 재생시키는데 사용되지만 일부 PGAL은 회로를 벗어나 당 혹은 녹말을 만드는데 사용된다. (3) 캘빈회로가 3회전 할 때마다 PGAL 한 분자가 얻어지며, 6회전 할 때마다 두
분자의 PGAL이 얻어져 포도당이나 과당과 같은 6탄소 단당류를 만들 수 있다(그림 5-16의 삽입도 참조).
(4) 요약하면 엽록체 스트로마의 탄소고정반응은 명반응에서 생산된 ATP와
NADPH를 사용하여 이산화탄소를 3탄소분자인 PGAL로 환원시킨다. PGAL 분자들은 상호 결합하여 화학결합의 형태로 수송을 위한 설탕이 되거나 에너지 저장을 위한 녹말이 된다.

3-6. 광호흡은 광합성의 비효율적 경로이다. 1) C3 경로를 이용하는 식물의 광합성에는 식물의 기공을 통하여 대기로부터 이산
화탄소를 얻지만, 기공이 열려 있는 동안에는 기공을 통해 물을 잃기도 한다.
2) C3식물들은 덮고 건조한 날씨 때문에 스트레스를 받게 되면 수분손실을 막기 위해 기공을 닫게 되고, 그 결과로 잎 내부의 이산화탄소의 농도가 저하한다. 이럴 경우 루비스코는 이산화탄소 대신에 산소와 RuBP의 결합을 일으킨다.
3) 그 결과 이산화탄소를 결합시킬 때 얻어지는 두 분자의 PGA 대신에 한 분자의
PGA 및 또 다른 유기분자를 만들게 된다. 이 유기분자는 산소를 소모하고 이산화탄소를 방출하는 구출경로를 따라 변화해 간다(그림 5-17).
4) 이 과정은 햇빛 아래에서 산소 소모와 이산화탄소 방출을 수반하면서 진행되므로 광호흡(photorespiration)이라고 한다.
어떻게 해서 이러한 낭비적인 대사체계가 발달할 수 있었을까? 루비스코는 초기 지구의 대기조건에서 이산화탄소의 양이 훨씬 많고 산소가 거의 없을 때 진화되었을 것이다.
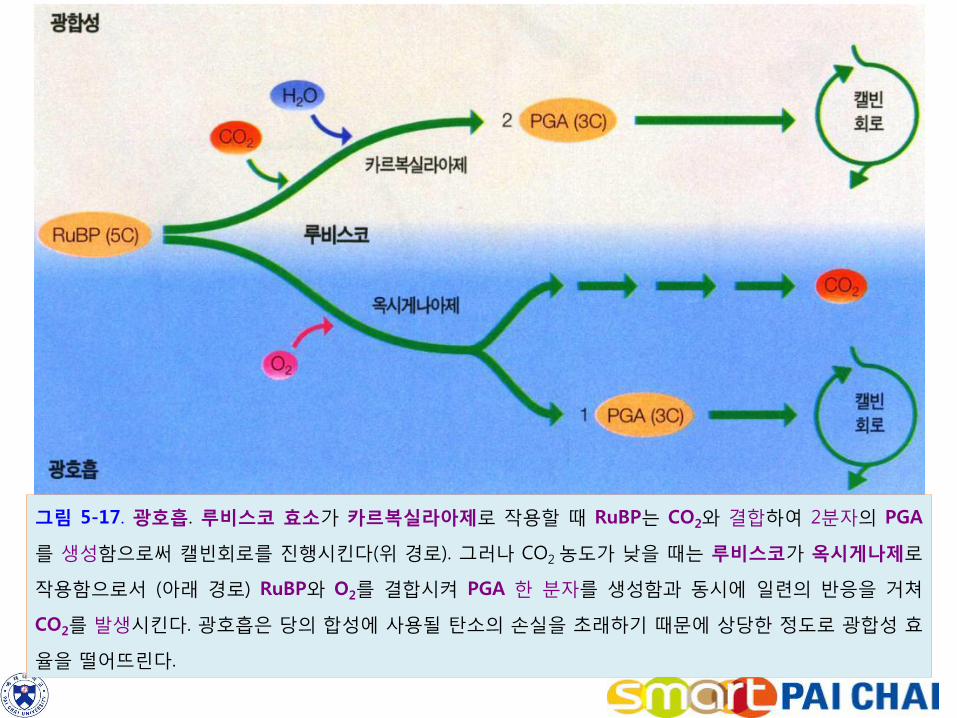
그림 5-17. 광호흡. 루비스코 효소가 카르복실라아제로 작용할 때 RuBP는 CO2와 결합하여 2분자의 PGA
를 생성함으로써 캘빈회로를 진행시킨다(위 경로). 그러나 CO2 농도가 낮을 때는 루비스코가 옥시게나제로
작용함으로서 (아래 경로) RuBP와 O2를 결합시켜 PGA 한 분자를 생성함과 동시에 일련의 반응을 거쳐
CO2를 발생시킨다. 광호흡은 당의 합성에 사용될 탄소의 손실을 초래하기 때문에 상당한 정도로 광합성 효
율을 떨어뜨린다.

3-7. C4 식물과 CAM 식물은 광호흡 기작을 감소시킨다.
C3 식물은 이산화탄소를 당으로 고정하기 위하여 캘빈회로만을 이용한다. 그러나
약 15%의 식물은 광합성 세포 내의 루비스코로 하여금 낮은 이산화탄소 환경에
노출되지 않도록 하는 대체기작을 발달시켰다.
1)C4 식물은 CO2를 이미 농축시켜 광호흡을 감소시킨다.
바랭이 (crabgrass), 옥수수 , 사탕수수와 같은 작물의 광합성 첫 안정산물은
옥살로아세트산(oxaloacetate)이다. 옥살로아세트산은 탄소를 4개 가지므로 이
경로를 C4 경로라 하며, 이 경로를 이용하는 식물을 C4 식물(C4 plant)이라고 한다.

(1)C4 식물의 엽육세포에서 옥살로아세트산을 형성한다.
C4 식물 잎의 엽육세포에서 이산화탄소는 PEP 카복실라아제(PEP carboxylase)의 작용으로 3탄소화합물인 포스포엔올피루브산(phosphoenolpyruvate, PEP)과 결합하여 옥살로아세트산을 형성한다(그림 5-18).
포스포엔올피루브산(C3H3O3ㆍⓟ) + CO2 + H2O → 옥살로아세트산(C4H4O5) + H3PO4
(2) 이 후 옥살로아세트산은 매우 신속하게 말산(malate, C4H5O4)으로 환원된다. 옥살로아세트산 + NADPH → 말산 + NADP+
(3) 말산은 엽맥을 둘러싸고 있는 특수한 세포인 유관속초세포로 수송되며, 여기에서 말산은 이산화탄소와 피루브산(pyruvate, C3H4O3)으로 전환된다. 말산 + NADP+
→ 피루브산 + CO2 + NADPH
이산화탄소는 캘빈회로에 들어가서 C3식물과 마찬가지로 루비스코에 의해 고정되어 PGA가 된다. 즉 이산화탄소의 고정이 2회 일어나는데 한 번은 엽육세포에서, 그리고 나머지 한 번은 유관속초세포에서 일어난다

(4) 이어서 피루브산은 엽육세포로 되돌아와 ATP를 사용하면서 PEP로 전환된다. C4 식물은 C4 경로와 캘빈회로를 공간적으로 분리시키고 있다. 즉 C4 경로는 엽육세포에서, 캘빈회로는 유관속초세포에서 일어난다. (5) C4 식물은 C4 경로를 이용함으로써 광호흡을 크게 감소시킨다. 유관속초세포 내에는 이산화탄소의 농도가 대기에 비해 10배나 높게 농축되어 있다. 이와 같이 높은 농도의 이산화탄소가 루비스코에 공급되면 기본적으로 광호흡이 없어지게 된다. 그 결과 높은 광도와 높은 온도 조건에서 옥수수, 사탕수수와 같은 C4 식물은 밀이나 벼와 같은 C3 식물에 비해 높은 순광합성율을 나타낼 수 있다.
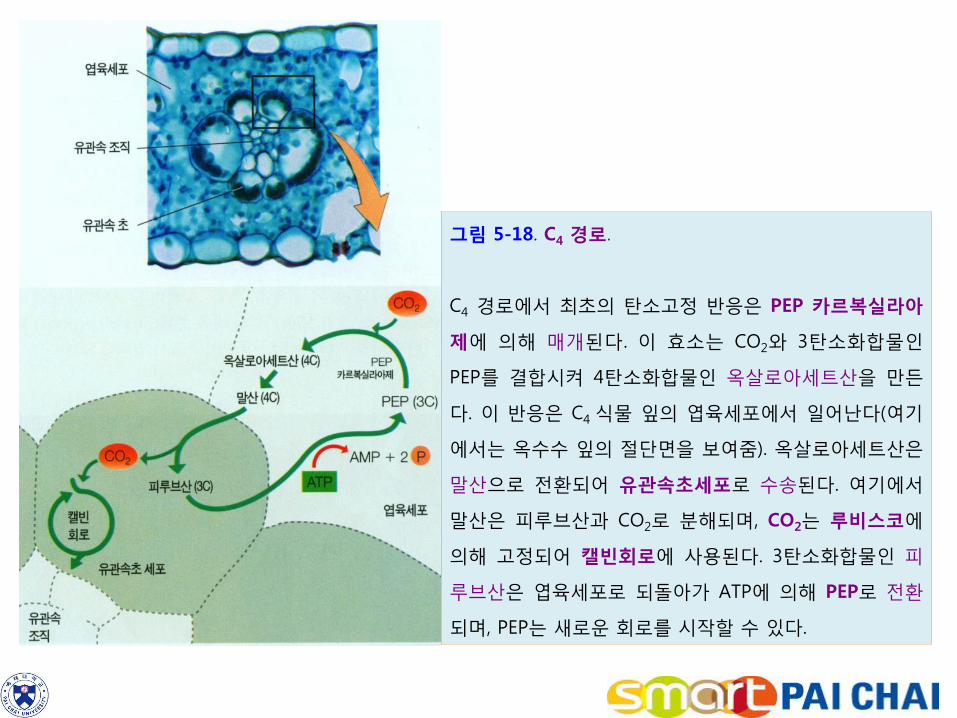
그림 5-18. C4 경로.
C4 경로에서 최초의 탄소고정 반응은 PEP 카르복실라아
제에 의해 매개된다. 이 효소는 CO2와 3탄소화합물인
PEP를 결합시켜 4탄소화합물인 옥살로아세트산을 만든
다. 이 반응은 C4 식물 잎의 엽육세포에서 일어난다(여기
에서는 옥수수 잎의 절단면을 보여줌). 옥살로아세트산은
말산으로 전환되어 유관속초세포로 수송된다. 여기에서
말산은 피루브산과 CO2로 분해되며, CO2는 루비스코에
의해 고정되어 캘빈회로에 사용된다. 3탄소화합물인 피
루브산은 엽육세포로 되돌아가 ATP에 의해 PEP로 전환
되며, PEP는 새로운 회로를 시작할 수 있다.

2) CAM 식물은 밤에 CO2를 고정하기에 광호흡을 감소시킨다.
사막에서 선인장이나 용설란과 같은 식물은 흔히 고온과 장기간의 수분 스트레스에
노출되어 있다. 수분손실을 줄이기 위하여 사막식물은 주간에 기공을 닫아야 한다.
많은 사막식물 특히 다육질의 줄기나 잎을 가진 사막식물은 광호흡을 줄이기 위하
여 C4 식물과는 다른 기작을 진화시켰다.
이러한 기작은 돌나물과(Crassulaceae) 식물인 꿩의비름에서 처음 발견되었으므로
이러한 기작을 크래슐산 대사(Crassulacean acid metabolism, CAM)라고 한다.
CAM 식물은 C4 식물과 마찬가지로 C4 경로와 캘빈회로를 모두 사용한다. 그러나
C4 식물은 이들 두 경로를 공간적으로 분리시켜 놓은 데 대하여 CAM 식물은 이들
을 시간적으로 분리시켜 놓았다(그림 5-19). 즉 CAM 식물은 기공을 개방하는 야간
에는 C4 경로를 운영하고, 기공을 폐쇄하는 주간에는 캘빈회로를 운영한다.

1) 온도가 낮은 야간에 기공을 열어 이산화탄소를 받아들인 후 엽육세포의 C4 경
로를 사용하여 이산화탄소를 말산으로 고정하고 이것을 액포에 저장한다.
포스포엔올피루브산(C3H3O3ㆍⓟ) + CO2 + H2O → 옥살로아세트산(C4H4O5) + H3PO4
옥살로아세트산 + NADPH → 말산 + NADP+
(2) 해가 뜨면 기공은 닫히면서 말산은 이산화탄소와 피루브산으로 분해되고,
말산 + NADP+ → 피루브산 + CO2 + NADPH
말산에서 분해된 이산화탄소는 엽록체로 들어 간 후 캘빈회로에 의해 고정된다.
3 RuBP(C5H8O5ㆍⓟㆍⓟ) + 3 CO2 + 3H2O → 6 PGA(C3H5O4)ㆍⓟ
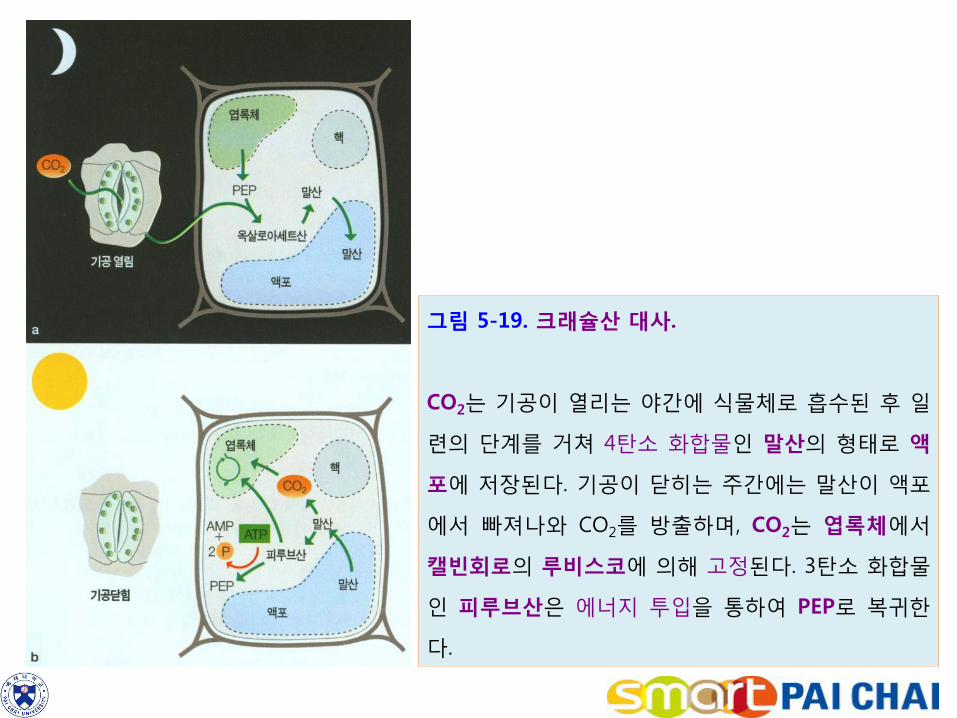
그림 5-19. 크래슐산 대사.
CO2는 기공이 열리는 야간에 식물체로 흡수된 후 일
련의 단계를 거쳐 4탄소 화합물인 말산의 형태로 액
포에 저장된다. 기공이 닫히는 주간에는 말산이 액포
에서 빠져나와 CO2를 방출하며, CO2는 엽록체에서
캘빈회로의 루비스코에 의해 고정된다. 3탄소 화합물
인 피루브산은 에너지 투입을 통하여 PEP로 복귀한
다.

(3) CAM 식물에는 선인장, 용설란, 산세비에리아, 꿩의비름(Crassula argentea) 및
돌나물속(Crassula) 식물들, 바랭이 등이 포함된다(그림 5-20).
(4) C3 식물은 C4 식물에 비해 보다 낮은 온도와 광도 그리고 보다 높은 습도에 잘
적응되어 있는 반면에
C4 식물은 C3 식물에 비해 덥고 건조한 기후에 더욱 잘 적응되어 있다.
CAM 식물 또한 덥고 건조한 기후에 더욱 잘 적응되어 있다.
(5) C4 식물과 CAM 식물의 탄소 농축 경로 가 아무 대가 없이 얻어지는 것은
아니다. 즉 탄소 농축 경로가 작동하기 위해서는 ATP 형태의 에너지가 여분으로
필요하다(그림 5-18 참조).

그림 5-20. CAM 식물의 예. (a) 산세베리아. (b) 꿩의비름(Crassula rupestris).

4. 호흡과 발효는 세포대사를 위해 에너지를 방출한다.
4-1. 호흡은 세포질과 미토콘드리아에서 일어난다.
4-2. 해당작용은 포도당을 두 분자의 피루브산으로 나눈다.
4-3. 피루브산은 CO2와 조효소 A가 결합한 아세틸로 분리된다.
4-4. 크렙스 회로에서 아세틸기는 두 분자의 이산화탄소로 파괴된다.
4-5. 전자전달 연쇄는 미토콘드리아 내막을 경계로 양성자 기울기가 일어난다.
4-6. 화학삼투와 산화적 인산화는 미토콘드리아에서 ATP를 생성한다.

4. 호흡과 발효는 세포대사를 위해 에너지를 방출한다. 식물은 종자를 만들 때 발아 시 빠른 유식물 생장을 개시하도록 전분 혹은 지질로 종자를 충전시킨다. 이러한 유기화합물을 분해하여 에너지를 방출하기 위하여 전분과 설탕은 먼저 포도당으로 분해되는데 이를 위하여 포도당은 호흡(respiration)이라고 하는 화학적 경로로 들어간다. 호흡이라고 하면 유산소 조건에서 포도당이 분해되어 이산화탄소와 물이 되면서 방출된 에너지가 ATP에 포획되는 대사과정이다. 호흡과정은 다음의 방정식으로 요약할 수 있다. C6H12O6 + 6O2 → 6CO2 + 6H2O 포도당 한 분자가 이산화탄소와 물로 산화될 때 36-38 분자의 ATP가 생성된다. 이들 ATP 분자는 포도당에 함유된 에너지의 약 38%에 해당한다. 남은 에너지는 열로 흩어져 버린다. 호흡은 일련의 효소반응을 통하여 포도당을 처리함으로서 포도당의 화학결합에 든 에너지를 포획한다. 이들 반응은 다음의 5가지 과정으로 구분되어 일어난다(그림 5-21). 1. 해당작용(glycolysis) 2. 아세틸 조효소 A(acetyl-CoA)의 형성 3. 크렙스 회로(Krebs cycle) 4. 전자전달 연쇄 5. 산화적 인산화반응.

4-1. 호흡은 세포질과 미토콘드리아에서 일어난다.
호흡에서 해당작용은 세포질의 액상 부분에서 일어난다. 나머지 반응들은 모두 미
토콘드리아에서 일어난다(제4장).
미토콘드리아는 외막 및 심하게 접혀진 구조를 가진 크리스테(cristae)로 되어 있다.
내막의 안쪽에는 액체상의 기질(matrix)이 있다.
아세틸 CoA의 형성 및 크렙스 회로는 미토콘드리아의 기질에서 일어난다.
전자전달 연쇄와 산화적 인산화반응의 효소들은 크리스테의 막에 들어 있다.
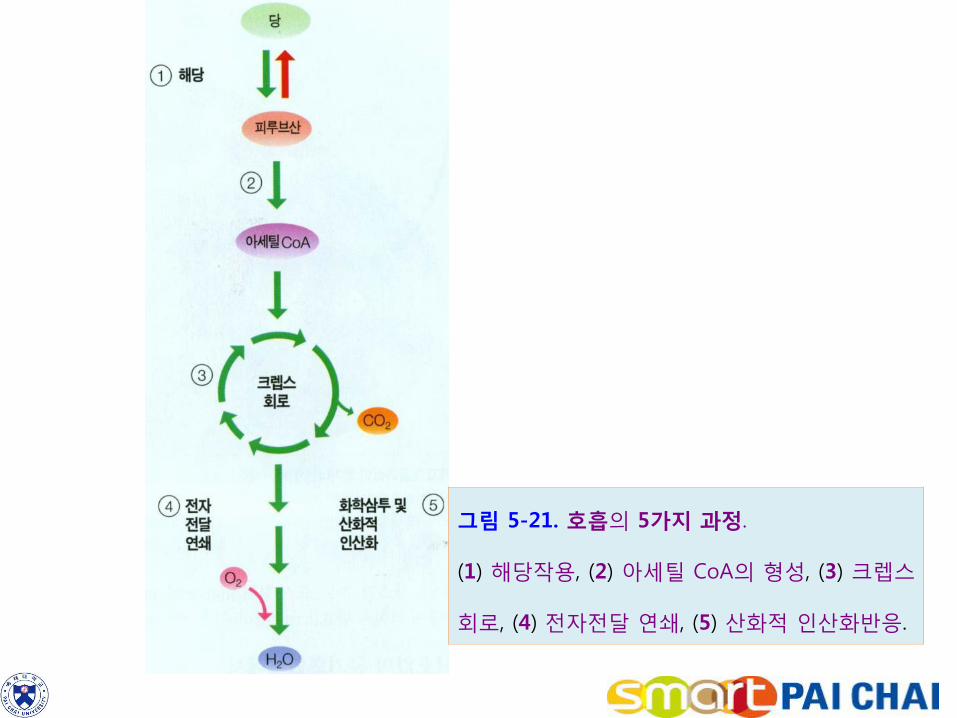
그림 5-21. 호흡의 5가지 과정.
(1) 해당작용, (2) 아세틸 CoA의 형성, (3) 크렙스
회로, (4) 전자전달 연쇄, (5) 산화적 인산화반응.
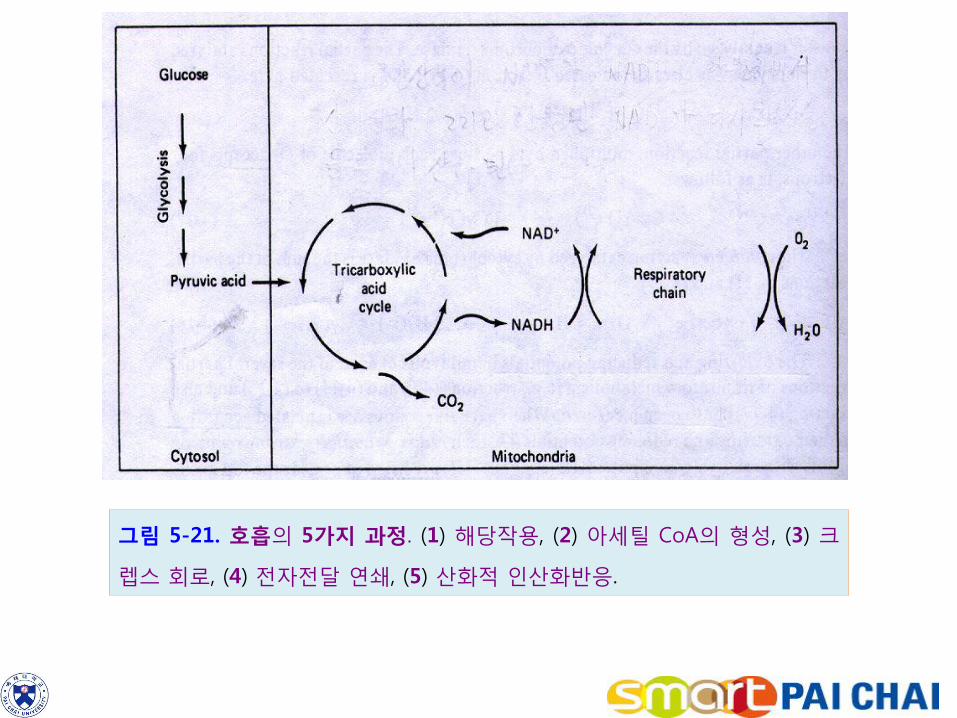
그림 5-21. 호흡의 5가지 과정. (1) 해당작용, (2) 아세틸 CoA의 형성, (3) 크
렙스 회로, (4) 전자전달 연쇄, (5) 산화적 인산화반응.
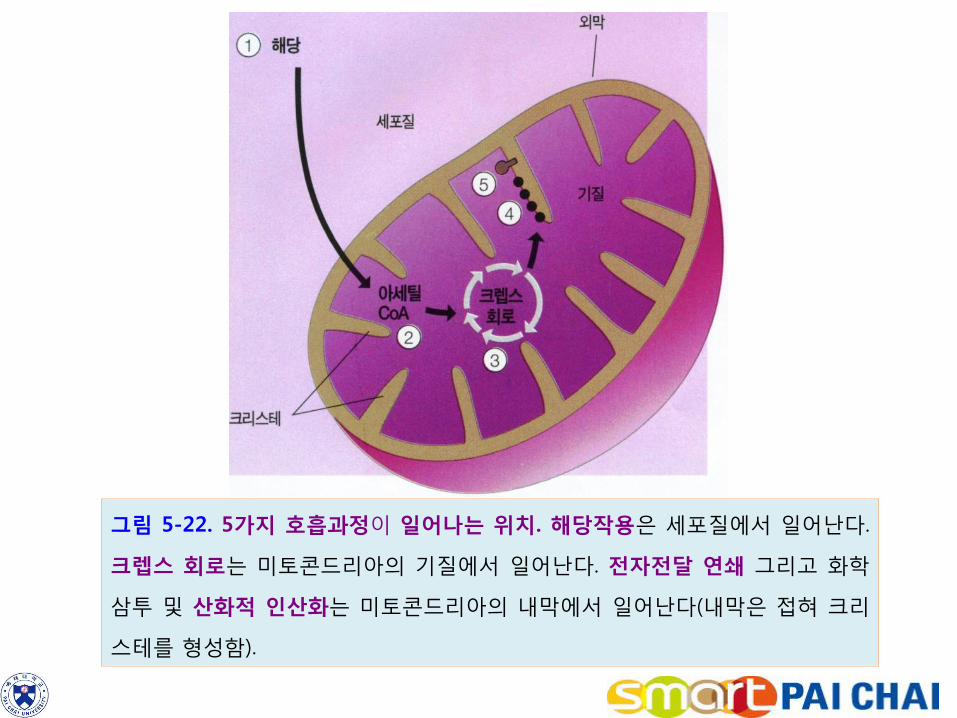
그림 5-22. 5가지 호흡과정이 일어나는 위치. 해당작용은 세포질에서 일어난다.
크렙스 회로는 미토콘드리아의 기질에서 일어난다. 전자전달 연쇄 그리고 화학
삼투 및 산화적 인산화는 미토콘드리아의 내막에서 일어난다(내막은 접혀 크리
스테를 형성함).

4-2. 해당작용은 포도당을 두 분자의 피루브산으로 나눈다.
모든 살아 있는 세포에서는 세포질에서 해당작용이 일어난다. 해당작용에서 6탄소 화합물인 포도당 한 분자는 3탄소 화합물인 피루브산 두 분자로 나누어진다. 해당작용은 10단계의 반응으로 구성되어 있으며, 각 반응은 각각 특정 효소에 의한 촉매작용을 받는다(그림 5-23). 1) 첫 단계는 포도당을 3탄소 화합물로 분할시키기 위하여 ATP의 도움을 받아 최초의
화합물이 형성되며, 이어서 포도당 분자를 재배열하는 준비 과정이다. glucose + ATP → glucoseㆍⓟ + ADP glucoseㆍⓟ → fructoseㆍⓟ 2) 나머지 단계는 이러한 3탄소 화합물인 피루브산(C3H3O3)으로 전환시키는 과정들이다. fructoseㆍⓟ → → → → → → → pyruvic acid 3) 세포에서 포도당 한 분자가 해당작용을 경유하면 2 ATP의 에너지가 순수하게 얻어진다.
ATP 2분자는 포도당에 함유된 전체 에너지의 2% 정도에 불과하다. 4) 해당작용에서는 또한 포도당으로부터 제거된 전자와 양성자로 NAD+를 환원시켜
NADH를 생성함으로서 일부 에너지를 포획하기도 한다. 해당작용에서 만들어진 두 분자의 피루브산에는 포도당의 대부분이 잔존한다.
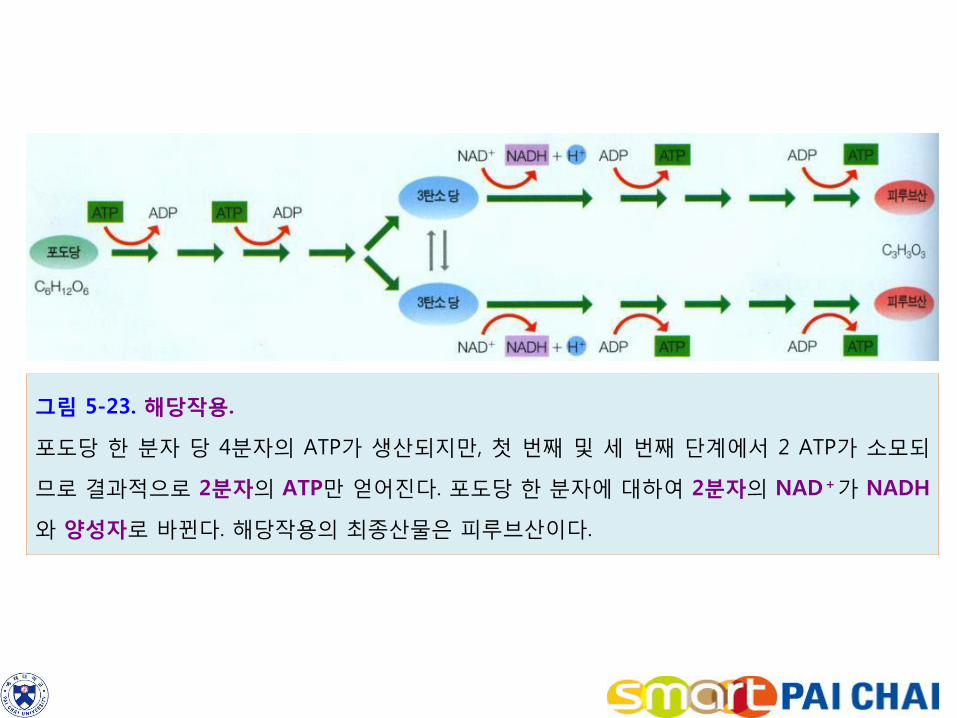
그림 5-23. 해당작용.
포도당 한 분자 당 4분자의 ATP가 생산되지만, 첫 번째 및 세 번째 단계에서 2 ATP가 소모되
므로 결과적으로 2분자의 ATP만 얻어진다. 포도당 한 분자에 대하여 2분자의 NAD+가 NADH
와 양성자로 바뀐다. 해당작용의 최종산물은 피루브산이다.

4-3. 피루브산은 CO2와 조효소 A가 결합한 아세틸로 분리된다..
피루브산 분자는 세포질을 떠나 미토콘드리아의 기질로 이동한다.
여기에서 피루브산은 한 분자의 이산화탄소를 내놓고 나머지 2탄소 화합물인 아세틸기(-
CH3CO)는 조효소 A(coenzyme A, Co A)와 결합하여 아세틸 Co A를 형성한다(그림 5-25). 하
나의 아세틸기가 형성될 때마다 전자수용체인 NAD+ 한 분자가 NADH로 환원되면서 에너
지가 포획된다.
Pyruvic acid + CoA + NAD+ → acetyl CoA + CO2 + NADH + H+
해당작용에서 포도당 한 분자는 피루브산 두 분자를 만들므로 두 개의 아세틸기가 크렙스
회로에 들어간다.

4-4. 크렙스 회로에서 아세틸기는 두 분자의 이산화탄소로 파괴된다.
식물 미토콘드리아에서 일어나는 크렙스 회로는 이산화탄소를 생성한다.
1) 첫 단계에서 아세틸 Co A는 아세틸기를 옥살로아세트산에게 전달하여
6탄소화합물인 시트르산(citric acid)을 만든다(그림 5-25 참조). 이와 같이
시트르산이 만들어지기 때문에 크렙스 회로를 시트르산 회로(citric acid
cycle)라고도 한다.
2) 아세틸기의 에너지 중 일부는 한 분자의 ATP로 포획되지만 대부분의 에너지는
NAD+와 FAD(flavin adenine dinucleotide)에게 전달된다 . 이들 전자수용체는
아세틸기로부터 전자와 양성자를 받아 NADH와 FADH2로 환원된다.
3) 하나의 아세틸기가 크렙스 회로를 따라 이동하는 동안 3분자의 NADH와
1분자의 FADH2가 만들어진다.

4) 회로가 끝날 때마다 옥살로아세트산이 재형성된다. 크렙스 회로에서 옥살로아세트산의 전구체는 말산이다.
5) 요약하면 하나의 포도당이 분해될 때마다 두 개의 아세틸기가 크렙스 회로에 들어가므로 각각의 포도당 분자가 크렙스 회로를 지나갈 때마다 2분자의 ATP, 6분자의 NADH와 2분자의 FADH2가 생산된다. 그리고 회로의 끝에서는 아세틸 Co A와 결합하는 옥살로아세트산이 재형성된다(해당작용 : 2ATP + 2NADP, 2Co A 형성 : 2NADP).
6) 지금까지의 호흡과정을 통해서 한 분자의 포도당이 완전히 CO2로 분해하는 동안 해당작용에서 2 ATP, 크렙스 회로에서 2 ATP가 만들어 지므로 총 4분자의 ATP가 생성된다.
나머지 한 분자의 포도당에 들어 있던 에너지의 대부분은 6분자의 NADH와 2분자의 FADH2에 포함되어 있다.
이들 분자에 들어있는 다량의 에너지는 직접 사용할 수는 없고 ATP로 전환해야 식물의 세포가 사용할 수가 있다.
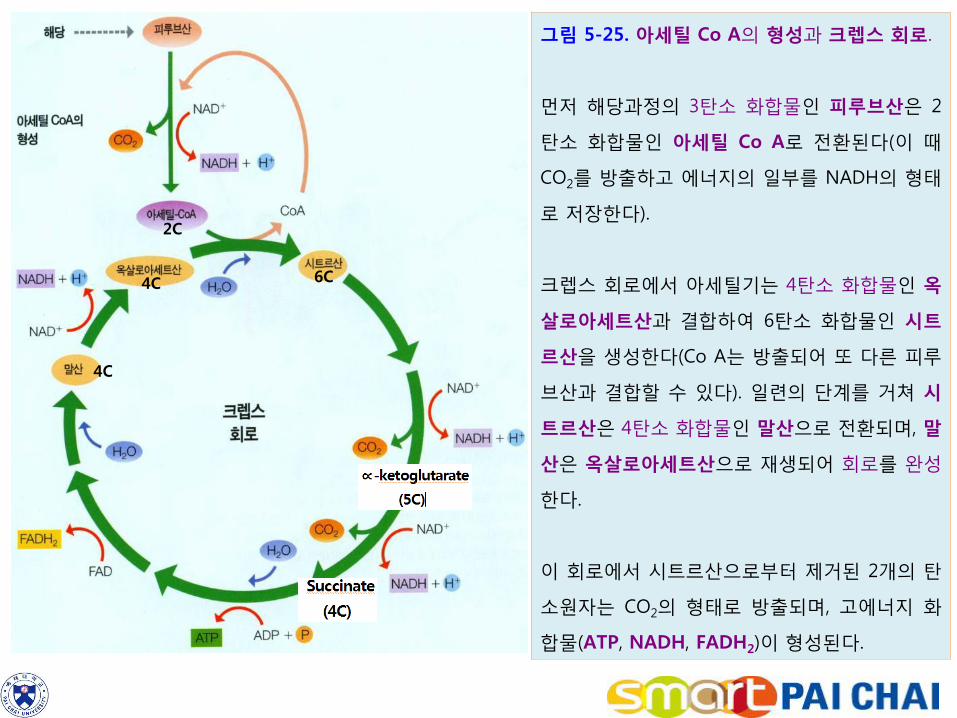
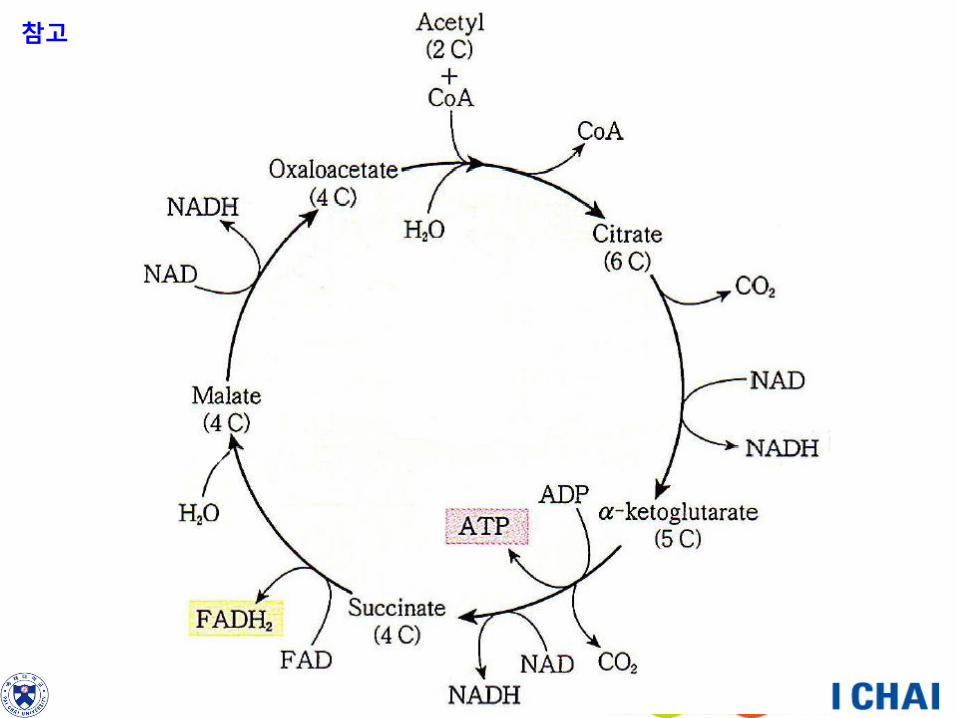
그림 5-25. 아세틸 Co A의 형성과 크렙스 회로.
먼저 해당과정의 3탄소 화합물인 피루브산은 2
탄소 화합물인 아세틸 Co A로 전환된다(이 때
CO2를 방출하고 에너지의 일부를 NADH의 형태
로 저장한다).
크렙스 회로에서 아세틸기는 4탄소 화합물인 옥
살로아세트산과 결합하여 6탄소 화합물인 시트
르산을 생성한다(Co A는 방출되어 또 다른 피루
브산과 결합할 수 있다). 일련의 단계를 거쳐 시
트르산은 4탄소 화합물인 말산으로 전환되며, 말
산은 옥살로아세트산으로 재생되어 회로를 완성
한다.
이 회로에서 시트르산으로부터 제거된 2개의 탄
소원자는 CO2의 형태로 방출되며, 고에너지 화
합물(ATP, NADH, FADH2)이 형성된다.
4C
4C
2C
6C

4-5. 전자전달 연쇄는 미토콘드리아 내막을 경계로 양성자 기울기가 일어난다.
에너지를 ATP로 전환하는 것이 호흡과정의 마지막 두 단계인 전자전달연쇄와 산화적 인산화이다. 이들 마지막 과정에서 포도당으로부터 포획된 에너지가 대량으로 회수된다. 전자전달 연쇄는 일련의 전자 운반체 분자로 이루어져 있으며, 이들은 미토콘드리아 내막에 위치한다(그림 5-26). 전자전달 연쇄에는 복합체Ⅰ부터 복합체Ⅳ까지4개의 단백질 복합체가 있다. 1) 복 합 체 Ⅰ 은 크렙스회로에서 생성된 NADH 를 NAD + 로 산 화 시키면서
유비퀴논(ubiquinone)을 환원시킨다.
2) 복합체Ⅱ는 크렙스 회로에서 생성된 FADH2를 FAD로 산화시키면서 유비퀴논을 환원시킨다. 환원된 유비퀴논은 전자를 복합체Ⅲ에게 운반한다.
3) 복합체Ⅲ은 유비퀴논으로부터 받은 전자를 시토크롬 c로 운반한다 . 시토크롬 c는 복합체Ⅲ로부터 복합체Ⅳ로 전자를 한 번에 하나씩 운반한다.
4) 복합체Ⅳ는 최종단계에서 산소가 환원되어 물이 되는 부위이다. 시토크롬 c를 통하여 전자가 4개씩 전달될 때마다 산소 한 분자가 환원되어 두 분자의 물이 된다.
요약하면 운반체 분자들은 NADH와 FADH2로부터 고에너지 전자를 취하여 일련의 산화 환원반응으로 이루어진 전자전달 연쇄를 따라 이들 전자를 넘겨준다
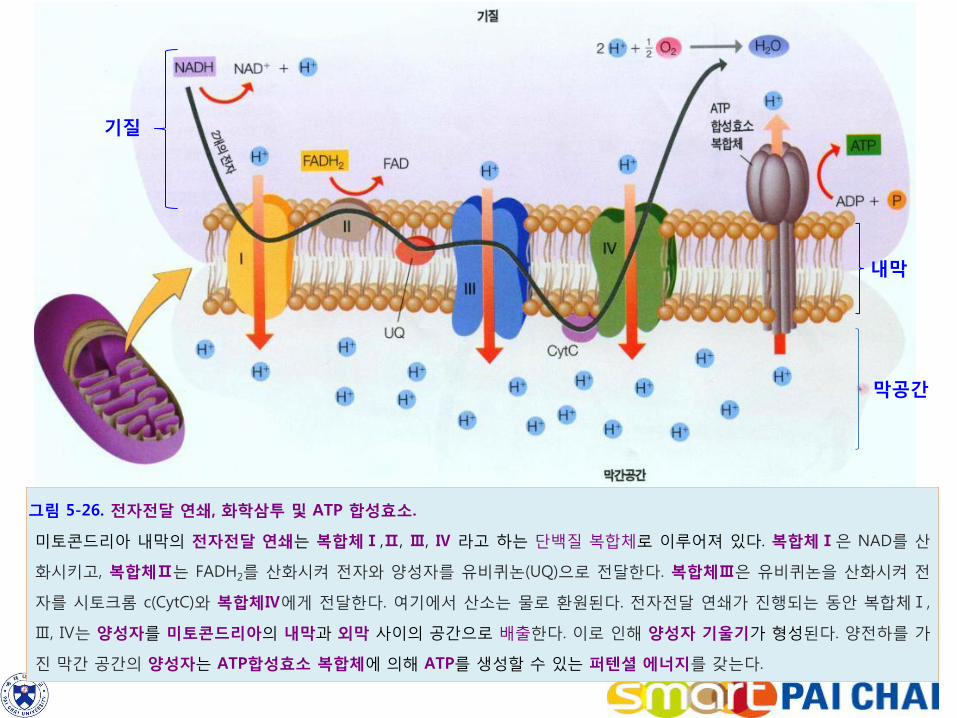
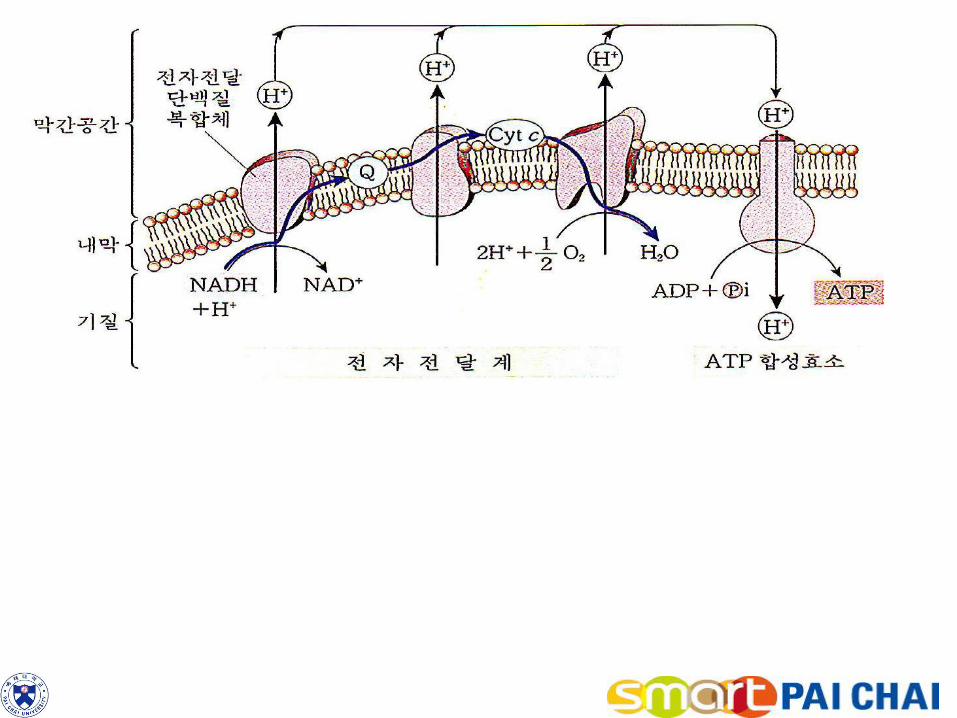
그림 5-26. 전자전달 연쇄, 화학삼투 및 ATP 합성효소.
미토콘드리아 내막의 전자전달 연쇄는 복합체Ⅰ,Ⅱ, Ⅲ, Ⅳ 라고 하는 단백질 복합체로 이루어져 있다. 복합체Ⅰ은 NAD를 산
화시키고, 복합체Ⅱ는 FADH2를 산화시켜 전자와 양성자를 유비퀴논(UQ)으로 전달한다. 복합체Ⅲ은 유비퀴논을 산화시켜 전
자를 시토크롬 c(CytC)와 복합체Ⅳ에게 전달한다. 여기에서 산소는 물로 환원된다. 전자전달 연쇄가 진행되는 동안 복합체Ⅰ,
Ⅲ, Ⅳ는 양성자를 미토콘드리아의 내막과 외막 사이의 공간으로 배출한다. 이로 인해 양성자 기울기가 형성된다. 양전하를 가
진 막간 공간의 양성자는 ATP합성효소 복합체에 의해 ATP를 생성할 수 있는 퍼텐셜 에너지를 갖는다.
내막
막공간
기질

1)미토콘드리아에서는 양성자 기울기의 퍼텐셜 에너지를 사용하기 위하여 ATP 합성효소 복합체를 활용한다.
이 복합체는 막간 공간의 양성자가 기질로 되돌아 올 수 있는 통로를 제공한다. 이러한 양성자의 흐름을 화학삼투(chemiosmosis)라고 불린다. 2) 양성자가 복귀함에 따라 이들의 에너지는 ADP를 ATP로 전환시켜 형성된 화학결합에 포획된다(그림 5-26 참조).
이 때 산소는 전자전달 연쇄의 최종 전자 수용체이고, 인산기가 ADP에 결합되어 ATP를 형성하는 인산화에서 에너지가 포획되므로 이 과정을 산화적 인산화라고 한다. 3) 전자전달 및 산화적 인산화에서 생산되는 ATP는 상당하다. 이들 최종 과정에서
포도당 한 분자로부터 얻어지는 ATP는 최대 34분자이다. 전자전달 및 산화적 인산화 이전의 과정에서 고작 4 ATP를 얻는데 불과했다.
따라서 포도당 한 분자를 완전한 호기성 호흡으로 분해할 때 얻을 수 있는 ATP의 최대량은 38분자이다.
4-6. 화학삼투와 산화적 인산화는 미토콘드리아에서 ATP를 생성한다.

4) 결론적으로 엽록체의 틸라코이드 막에서 광합성의 명반응은 태양 에너지를 ATP
와 NADPH 분자의 형태로 포획한다.
엽록체 스트로마의 탄소고정반응에서는 이 ATP와 NADPH를 이용하여 CO2를 3탄
당인 PGAL에 고정시킨다. 이 후 PGAL 분자들은 상호 결합되어 포도당, 설탕, 녹말
과 같은 당을 생산한다.
5) 호흡은 포도당과 같은 간단한 당을 취하여 이산화탄소로 완전 분해한다.
당의 분해에 따라 방출된 에너지는 ATP의 형태로 포획되어 세포 대사를 일으킨다.
광합성과 호흡은 모두 막을 가로 지르는 전기화학적 기울기를 이용하여 ADP와 인
산으로부터 ATP를 생산한다.
참고
참고
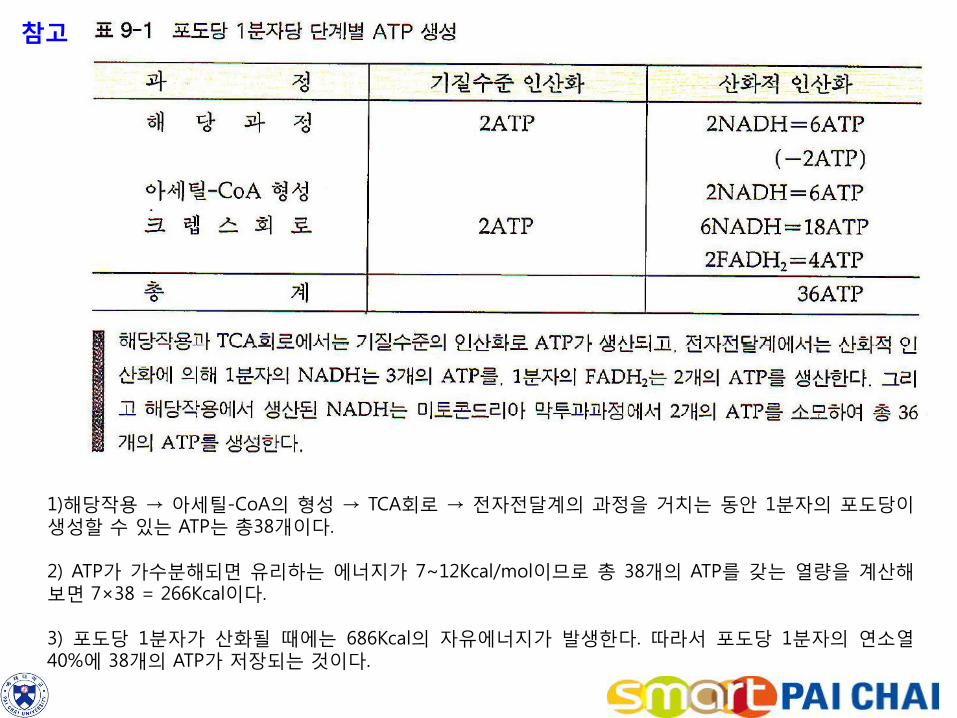
참고
1)해당작용 → 아세틸-CoA의 형성 → TCA회로 → 전자전달계의 과정을 거치는 동안 1분자의 포도당이 생성할 수 있는 ATP는 총38개이다. 2) ATP가 가수분해되면 유리하는 에너지가 7~12Kcal/mol이므로 총 38개의 ATP를 갖는 열량을 계산해 보면 7×38 = 266Kcal이다. 3) 포도당 1분자가 산화될 때에는 686Kcal의 자유에너지가 발생한다. 따라서 포도당 1분자의 연소열 40%에 38개의 ATP가 저장되는 것이다.
Copyright © 2022 FDOKUMEN

