
Mouse JunD negatively regulates fibroblast growth and antagonizes transformation by ras
Upload
independentCategory
view
1download
0
©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2129
Cell Cycle 10:13, 2129-2139; July 1, 2011; © 2011 Landes Bioscience
RepoRt RepoRt
*Correspondence to: Giovanni Morrone or Heather M. Bond; Email: [email protected] or [email protected]: 03/31/11; Revised: 04/26/11; Accepted: 04/26/11DOI: 10.4161/cc.10.13.16045
Introduction
The development of B lymphocytes from hematopoietic stem cells (HSCs) is an extensively investigated and well-characterized process that leads to the progressive lineage restriction of multi-potent stem and progenitor cells and to the acquisition of spe-cialized features. This is the result of the concerted activation/repression of specific repertoires of genes (reviewed in refs. 1–4) orchestrated by a network of transcription factors and epigen-etic effectors. Early B-cell factor 1 (EBF1) is acknowledged as a central player in this process. Alone or in co-ordination with E2A and/or Pax5, this factor activates the transcription of a number of B-cell-specific genes including CD79a (mb-1), CD79b (B29), Igll1 (lambda5), VpreB1 (VpreB) and CD19 (reviewed in ref. 5). The knockout of the Ebf1 gene results in abrogation of B-lymphopoiesis and accumulation of early lymphoid progeni-tors that lack signs of B-lineage priming but are endowed with
Zinc finger protein 521 (eHZF/ZNF521) is a multi-functional transcription co-factor containing 30 zinc fingers and an N-terminal motif that binds to the nucleosome remodelling and histone deacetylase (NuRD) complex. ZNF521 is believed to be a relevant player in the regulation of the homeostasis of the hematopoietic stem/progenitor cell compartment, however the underlying molecular mechanisms are still largely unknown. Here, we show that this protein plays an important role in the control of B-cell development by inhibiting the activity of early B-cell factor-1 (eBF1), a master factor in B-lineage specification. In particular, our data demonstrate that: (1) ZNF521 binds to eBF1 via its carboxyl-terminal portion and this interaction is required for eBF1 inhibition; (2) NuRD complex recruitment by ZNF521 is not essential for the inhibition of transactivation of eBF1-dependent promoters; (3) ZNF521 represses eBF1 target genes in a human B-lymphoid molecular context; and (4) RNAi-mediated silencing of ZNF521/Zfp521 in primary human and murine hematopoietic progenitors strongly enhances the generation of B-lymphocytes in vitro. taken together, our data indicate that ZNF521 can antagonize B-cell development and lend support to the notion that it may contribute to conserve the multipotency of primitive lympho-myeloid progenitors by preventing or delaying their eBF1-driven commitment toward the B-cell lineage.
Zinc finger protein 521 antagonizes early B-cell factor 1 and modulates
the B-lymphoid differentiation of primary hematopoietic progenitors
tiziana Mega,1 Michela Lupia,1 Nicola Amodio,1 Sarah J. Horton,3 Maria Mesuraca,1 Daniela pelaggi,1 Valter Agosti,1 Michele Grieco,4 emanuela Chiarella,1 Raffaella Spina,1 Malcolm A.S. Moore,2 Jan Jacob Schuringa,3 Heather M. Bond1,*
and Giovanni Morrone1,*
1Laboratory of Molecular Hematopoiesis and Stem Cell Biology; Department of experimental and Clinical Medicine “G. Salvatore”; University of Catanzaro Magna Græcia; University Campus “Salvatore Venuta;” Germaneto, Catanzaro, Italy; 2Moore Laboratory; Cell Biology program; Memorial Sloan-Kettering Cancer Center; New York, NY USA;
3Department of Hematology; Faculty of Medical Sciences; University Medical Center Groningen (UMCG); University of Groningen; Groningen, the Netherlands; 4Department of environmental Sciences; Second University of Naples; Caserta, Italy
Keywords: ZNF521, EBF1, transcription, B-lymphocytes, hematopoietic stem cells, differentiation
T-lymphoid, and considerable myeloid potential.6-8 Enforced expression of EBF1 in HSCs, even in the absence of Pax5, represses myeloid and T-lymphoid gene expression and restricts their differentiation to the B-cell lineage.8 Therefore, modulation of the expression and/or activity of EBF1 is likely to represent an important mechanism to preserve the multipotency of HSCs and early progenitors and to maintain the integrity of the immature hematopoietic cell compartment as well as to ensure the correct balance between T- and B-lymphopoiesis.
Known inhibitors of EBF1 include the Notch1 receptor,9 the Polycomb group protein Bmi1,10 and two related transcription co-factors, both containing 30 finger motifs, OAZ/ZNF423,11 and EHZF/ZNF521.12 The transcript encoding this latter factor was identified and cloned in our laboratory because of its selec-tive expression in human CD34+ cells compared with mature leukocytes.12 ZNF521 is highly abundant in the more primitive hematopoietic stem/progenitor cell fraction, and its levels rapidly

©2011 Landes Bioscience.Do not distribute.
2130 Cell Cycle Volume 10 Issue 13
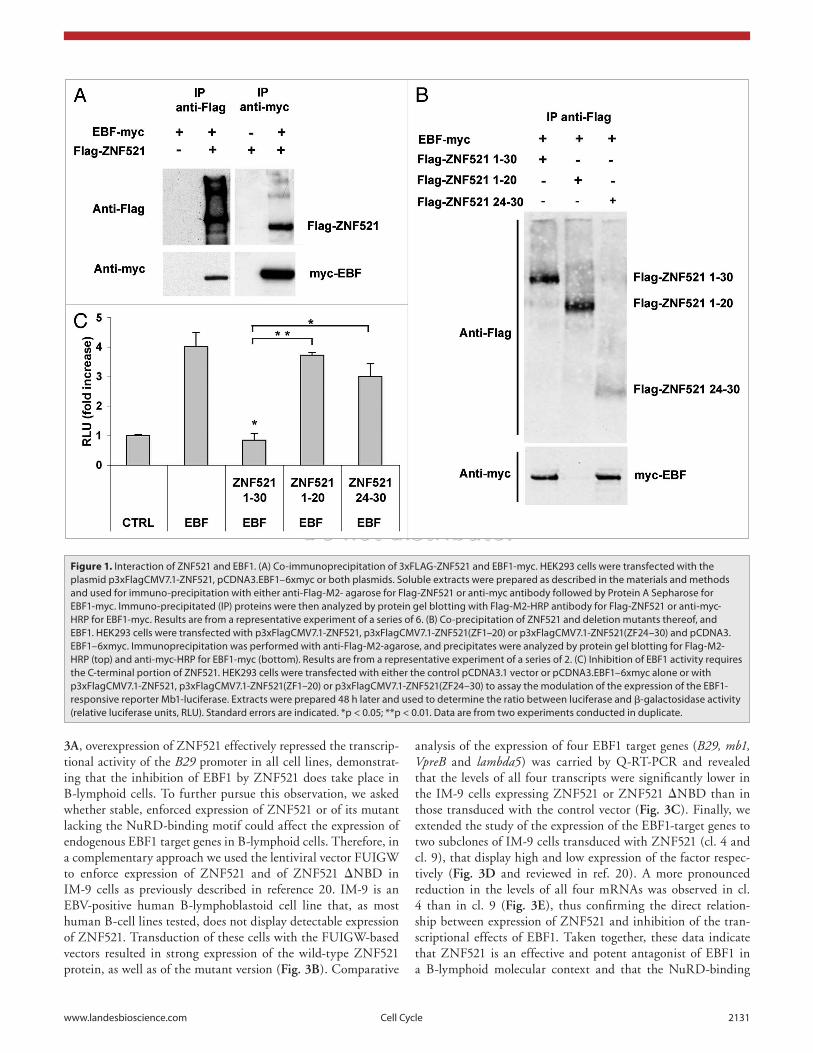
by antibodies to either tag sequence. Since the carboxyl-terminal zinc fingers of ZNF521 are highly homologous to those of OAZ/Zfp423, that are essential for binding to EBF1,15 we repeated the assay using 2 deletion mutants of ZNF521, comprising zinc-finger (ZF) motifs 1–20 and 24–30 respectively. Figure 1B shows that ZNF521(ZF 1–20) did not bind to EBF1, whereas the binding capacity of ZNF521(ZF 24–30) was comparable to that of the full-length protein. Thus, the carboxyl-terminal por-tion of ZNF521 is necessary for heterodimerization with EBF1. The inability of ZNF521(ZF 1–20) to interact with EBF1 was mirrored by its lack of inhibitory activity on the EBF-dependent promoter, mb1 (Fig. 1C). As could be expected, ZNF521(ZF 24–30), that contains only 7 of the 30 ZFs of the native pro-tein, had virtually no effect on EBF1-dependent transcription. Additional deletion mutants of ZNF521 spanning from the N-terminus to up to the 25th zinc finger motif were also tested and found unable to bind to and inhibit EBF1, but their expres-sion was poor, most likely owing to instability of the truncated protein (not shown).
The NuRD-binding domain of ZNF521 is not essential for EBF1 inhibition. As highlighted in the Introduction, ZNF521 contains an N-terminal, 12-amino acid motif shared by a num-ber of transcriptional co-repressors12,16 that is believed to function via the recruitment of histone deacetylases (HDACs) and other components of the NuRD complex to the promoter of target genes.17,18 To investigate the role of this motif, a deletion mutant was constructed (ZNF521 ΔNBD) that lacks 13-amino acids at the N-terminus and is indeed unable to bind members of the NuRD complex (reviewed in ref. 13 and Fig. 2A) but fully retains its ability to interact with EBF1 (Fig. 2A). Co-transfection exper-iments in HEK293 cells (Fig. 2B) showed that overexpression of ZNF521 ΔNBD effectively inhibited the EBF-induced trans-activation of the B29, mb1 and lambda5 promoters, although to a slightly lower extent than full-length ZNF521 indicating that the N-terminal domain may contribute to, but is not essential for, the inhibition of EBF1-mediated transactivation of all target promoters tested.
Repression of EBF1 target genes by ZNF521 and ZNF521 ΔNBD in a human B-lymphoid molecular context. Although a strong inhibitory effect of ZNF521 on EBF1 has been docu-mented in HEK293 and HeLa cells,12,19 it is entirely plausible that the functional interaction between the two factors may result in different, cell-type-specific, effects in diverse cell populations. In fact, in a study of B-cell malignancies in AKXD-27 mice, Hentges et al.19 observed an increase in the expression of Ebf1 target genes in leukemias overexpressing Zfp521 as a result of viral integration at the Evi3 locus. Based on this finding, they postulated a context-dependent scenario whereby the Zfp521-Ebf1 interaction in leukemic B lymphocytes, unlike in HEK293 cells or other cell types, may result in cooperative induction of the transcription of B-cell genes.
We therefore sought to verify if ZNF521 could inhibit the activity of endogenous EBF1 in cells of B-lymphoid origin. To this end, three human B-lymphoblastoid cell lines (IM-9, Roha9-MC3 and DeFew) were co-transfected with the plasmids 3x FLAG or 3x FLAG-ZNF521 and B29-Luc. As shown in Figure
decline during their erythroid, myeloid or lymphoid differentia-tion (reviewed in ref. 13), strongly suggesting a role for this factor in the maintenance of the immature hematopoietic cell compart-ment. ZNF521 is the human ortholog to murine Zfp521 (Evi-3), encoded by a gene adjacent to a common ecotropic viral inte-gration site on chromosome 18, associated with the occurrence of B-cell malignancies in mice of the AKXD strain.14 ZNF521 shares high sequence homology with ZNF423, another 30-ZF protein originally identified for its ability to interact with EBF1 in neural precursors of the olfactory epithelium and to inhibit its activity.11,15 Like ZNF423, ZNF521 strongly represses EBF1-dependent promoters in human embryonic kidney-derived HEK293 cells.12 Since ZNF521 is abundantly expressed in early hematopoietic progenitors, unlike ZNF423 (reviewed in ref. 12; Bond HM, unpublished) it is conceivable that it may have an important role in the control of fate choice of multipotent hema-topoietic progenitors by limiting their EBF1-driven commitment to the B-cell lineage.
Some functional properties of ZNF521 are likely to be medi-ated by a 12-amino acid motif, located at the N-terminal end of the protein and highly conserved in a number of transcrip-tion factors with repressor activity including friend of GATA (FOG)-1 and 2, and SALL proteins.12 This motif interacts with the nucleosome remodelling and histone deacetylation (NuRD) complex13,16 and is essential for the repression of GATA-1 tar-get genes by its molecular interactor, FOG-1, that requires the recruitment of the NuRD complex to the promoters of these genes.17,18 Thus, it could be hypothesized that the interaction of ZNF521 with NuRD proteins may be similarly necessary for its inhibitory effect on EBF1-dependent promoters.
We therefore deemed it of interest to further characterize the interaction between ZNF521 and EBF, to investigate the role of the NuRD-binding motif, and to elucidate the biological effects of ZNF521 in a B-lymphoid cellular context. The data illus-trated below provide the first evidence of a physical interaction between ZNF521 and EBF1, map the domain responsible for this interaction to the carboxyl-terminal region of ZNF521 and indicate that the NuRD recruitment by ZNF521 is not essen-tial for the inhibition of transactivation of EBF1 target promot-ers. Moreover, we show that the ectopic expression of ZNF521 in human B-lymphoblastoid cells inhibits EBF1-induced tran-scription and downregulates the expression of the endogenous B-lymphoid genes. Most importantly, we demonstrate that silencing of ZNF521 strongly enhances the generation of B lym-phocytes from human and murine pluripotent progenitors.
Results
Molecular interaction between ZNF521 and EBF1. Despite the functional evidence that ZNF521 inhibits the transactivation of EBF1-dependent promoters in HEK293 cells,12 a direct molec-ular interaction between these two factors has not been docu-mented thus far. To address this issue, co-immunoprecipitation assays were performed on extracts of HEK293 cells transfected with 3x FLAG-tagged ZNF521 and myc-tagged EBF1. As illus-trated in Figure 1A, both proteins were efficiently pulled down
©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2131
analysis of the expression of four EBF1 target genes (B29, mb1, VpreB and lambda5) was carried by Q-RT-PCR and revealed that the levels of all four transcripts were significantly lower in the IM-9 cells expressing ZNF521 or ZNF521 ΔNBD than in those transduced with the control vector (Fig. 3C). Finally, we extended the study of the expression of the EBF1-target genes to two subclones of IM-9 cells transduced with ZNF521 (cl. 4 and cl. 9), that display high and low expression of the factor respec-tively (Fig. 3D and reviewed in ref. 20). A more pronounced reduction in the levels of all four mRNAs was observed in cl. 4 than in cl. 9 (Fig. 3E), thus confirming the direct relation-ship between expression of ZNF521 and inhibition of the tran-scriptional effects of EBF1. Taken together, these data indicate that ZNF521 is an effective and potent antagonist of EBF1 in a B-lymphoid molecular context and that the NuRD-binding
3A, overexpression of ZNF521 effectively repressed the transcrip-tional activity of the B29 promoter in all cell lines, demonstrat-ing that the inhibition of EBF1 by ZNF521 does take place in B-lymphoid cells. To further pursue this observation, we asked whether stable, enforced expression of ZNF521 or of its mutant lacking the NuRD-binding motif could affect the expression of endogenous EBF1 target genes in B-lymphoid cells. Therefore, in a complementary approach we used the lentiviral vector FUIGW to enforce expression of ZNF521 and of ZNF521 ΔNBD in IM-9 cells as previously described in reference 20. IM-9 is an EBV-positive human B-lymphoblastoid cell line that, as most human B-cell lines tested, does not display detectable expression of ZNF521. Transduction of these cells with the FUIGW-based vectors resulted in strong expression of the wild-type ZNF521 protein, as well as of the mutant version (Fig. 3B). Comparative
Figure 1. Interaction of ZNF521 and eBF1. (A) Co-immunoprecipitation of 3xFLAG-ZNF521 and eBF1-myc. HeK293 cells were transfected with the plasmid p3xFlagCMV7.1-ZNF521, pCDNA3.eBF1–6xmyc or both plasmids. Soluble extracts were prepared as described in the materials and methods and used for immuno-precipitation with either anti-Flag-M2- agarose for Flag-ZNF521 or anti-myc antibody followed by protein A Sepharose for eBF1-myc. Immuno-precipitated (Ip) proteins were then analyzed by protein gel blotting with Flag-M2-HRp antibody for Flag-ZNF521 or anti-myc-HRp for eBF1-myc. Results are from a representative experiment of a series of 6. (B) Co-precipitation of ZNF521 and deletion mutants thereof, and eBF1. HeK293 cells were transfected with p3xFlagCMV7.1-ZNF521, p3xFlagCMV7.1-ZNF521(ZF1–20) or p3xFlagCMV7.1-ZNF521(ZF24–30) and pCDNA3.eBF1–6xmyc. Immunoprecipitation was performed with anti-Flag-M2-agarose, and precipitates were analyzed by protein gel blotting for Flag-M2-HRp (top) and anti-myc-HRp for eBF1-myc (bottom). Results are from a representative experiment of a series of 2. (C) Inhibition of eBF1 activity requires the C-terminal portion of ZNF521. HeK293 cells were transfected with either the control pCDNA3.1 vector or pCDNA3.eBF1–6xmyc alone or with p3xFlagCMV7.1-ZNF521, p3xFlagCMV7.1-ZNF521(ZF1–20) or p3xFlagCMV7.1-ZNF521(ZF24–30) to assay the modulation of the expression of the eBF1-responsive reporter Mb1-luciferase. extracts were prepared 48 h later and used to determine the ratio between luciferase and β-galactosidase activity (relative luciferase units, RLU). Standard errors are indicated. *p < 0.05; **p < 0.01. Data are from two experiments conducted in duplicate.

©2011 Landes Bioscience.Do not distribute.
2132 Cell Cycle Volume 10 Issue 13
motif is dispensable for this prop-erty of ZNF521.
Enhanced B-cell development following silencing of ZNF521/Zfp521 in murine and human hematopoietic progenitors. To assess the physiological relevance of zinc finger protein 521 in B-lymphopoiesis, it is important to establish whether the presence of this factor does indeed affect the B-lymphoid commitment of primary, multipotent hematopoi-etic progenitors. To address this issue we exploited the availability of stromal co-culture systems that permit efficient B-cell development in vitro.21,22
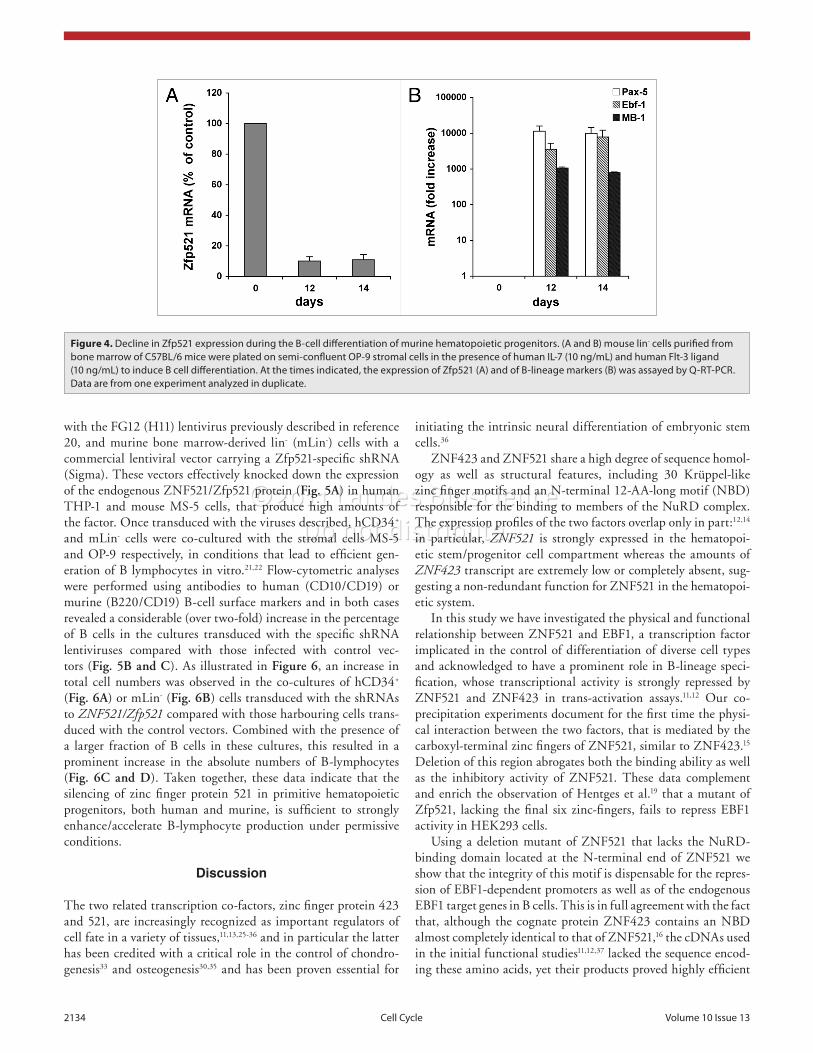
In initial experiments, we mea-sured the amounts of Zfp521 mRNA in murine Lin-negative cells co-cultured with OP-9 cells in the presence of interleukin 7 and Flt3 ligand. As shown in Figure 4A and B, a marked decline of the Zfp521 mRNA levels was evident in hematopoietic pro-genitors cultured in these condi-tions, that was paralleled by their B-lymphoid maturation as shown by the dramatic accumulation of mRNAs encoding B-cell-specific proteins (Ebf1, Pax5 and mb1). This is in full agreement with the observation of van Zelm et al. in human CD34+/lin- cells,23 where the expression of ZNF521 drops significantly during the transition to pro-B cells and is virtually absent in pre-B and immature B lympho-cytes. These data are also consistent
Figure 2. the NuRD-binding domain (NBD) of ZNF521 is not essential for the inhibition of eBF1-depen-dent promoters. (A) Deletion of the ZNF521 NBD abolishes binding to HDACs but not to eBF1. HeK293 cells were transfected with the pCDNA3.eBF1–6xmyc and with void p3xFlagCMV7.1, p3xFlagCMV7.1-ZNF521 or p3xFlagCMV7.1-ZNF521(ΔNBD). Immunoprecipitation was performed with anti-Flag-M2-aga-rose. precipitated proteins were detected by protein gel blotting with Flag-M2-HRp antibody for Flag-ZNF521, anti-myc-HRp for eBF1-myc, rabbit anti-HDAC-1 or goat anti-HDAC2 antibody followed by a HRp anti-rabbit/goat secondary antiserum. Results are from a representative experiment of a series of four. (B) Inhibition of eBF-1 activity by Flag-ZNF521 and Flag-ZNF521(ΔNBD). HeK293 cells were transfected with the reporter plasmids B29-Luc, MB1-Luc or l5-Luc and RSV-β-gal as an internal control, with the expression plasmid pCDNA3.eBF1–6xmyc and p3xFlagCMV7.1-ZNF521 or p3xFlagCMV7.1-ZNF521(ΔNBD) as indicated, and 48 h later extracts were assayed to determine the ratio between luciferase and β-galactosidase activity (relative luciferase units, RLU). Standard errors are indicated. *p < 0.05; **p < 0.01; ***p < 0.001. Data are from two experiments with triplicate and duplicate transfections respectively.
Figure 3 (See opposite page). ZNF521 represses eBF target genes in human B-lymphoid cells. (A) ZNF521 inhibits the activity of endogenous eBF1 in human B cell lines. the B cell lines, IM-9, Roha9-MC3 and DeFew were transfected by electroporation with the B29-luciferase reporter plasmid and with void p3xFlagCMV7.1 (control) or p3xFlagCMV7.1-ZNF521 plasmids. the renilla-luciferase plasmid pRL-tK was also transfected as an internal control. extracts were assayed in duplicate to determine the firefly/renilla luciferase activity ratio. the Relative Luciferase Units (RLU) are expressed as a percentage of the values measured in cells transduced with the control vector. Standard errors are indicated. **p < 0.01. Data are from two experi-ments conducted in duplicate. (B) expression of Flag-ZNF521 and Flag-ZNF521 ΔNBD in transduced IM-9 cells. the expression was detected in nuclear extracts by protein gel blotting with anti-flag M2-HRp. (C) enforced expression of ZNF521 or ZNF521 ΔNBD in IM-9 B lymphoblastoid cells inhibits the expression of eBF target genes. IM-9 cells were transduced with control lentiviral vector FUIGW or with FUIGW-ZNF521 or FUIGW-ZNF521 ΔNBD with an efficency of ranging between 76 and 99% as assessed by flow-cytometric measurement of eGFp+ cells (not shown). cDNA was prepared for Q-pCR analysis of mRNA expression of B29, mb1, VpreB and λ5 relative to the amplification of a housekeeping transcript (GAPDH). Data are represented as percentage of the expression of control FUIGW-transduced cells, and are from three distinct transductions assayed in duplicate. For ddct values, *p < 0.05; **p < 0.01. (D) expression of Flag-ZNF521 in IM-9 clones. the expression was detected in nuclear extracts by protein gel blotting with anti-flag M2-HRp. (e) Dose-dependent inhibition of eBF target genes in IM9 cells by ZNF521 Clones of ZNF521-transduced IM9 that expressed low (Clone 9) or high (Clone 4) amounts of ZNF521 (see D) were selected from methylcellulose cultures. the clones were analyzed by Q-Rt-pCR for the expression of B29, mb1, VpreB and λ5 transcripts relative to GAPDH. Data are represented as percentage of the expression of control FUIGW-transduced cells, and are from two experiments assayed in triplicate. For ddct values, *p < 0.05; **p < 0.01.

©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2133
We subsequently asked whether the RNAi-mediated silencing of the ZNF521/Zfp521 gene in primary hematopoietic progenitors could enhance the generation of B cells in culture. To this end we transduced human umbilical cord blood CD34+ (hCD34+) cells
with those of Chambers et al.24 who found a 42.8-fold higher expression of Zfp521 in hematopoietic stem cells compared with mature leukocytes including B cells (franklin.imgen.bcm.tmc.edu/loligag/lf/lt-hsc.php).
Figure 3. For figure legend, see page 2132.
©2011 Landes Bioscience.Do not distribute.
2134 Cell Cycle Volume 10 Issue 13
initiating the intrinsic neural differentiation of embryonic stem cells.36
ZNF423 and ZNF521 share a high degree of sequence homol-ogy as well as structural features, including 30 Krüppel-like zinc finger motifs and an N-terminal 12-AA-long motif (NBD) responsible for the binding to members of the NuRD complex. The expression profiles of the two factors overlap only in part:12,14 in particular, ZNF521 is strongly expressed in the hematopoi-etic stem/progenitor cell compartment whereas the amounts of ZNF423 transcript are extremely low or completely absent, sug-gesting a non-redundant function for ZNF521 in the hematopoi-etic system.
In this study we have investigated the physical and functional relationship between ZNF521 and EBF1, a transcription factor implicated in the control of differentiation of diverse cell types and acknowledged to have a prominent role in B-lineage speci-fication, whose transcriptional activity is strongly repressed by ZNF521 and ZNF423 in trans-activation assays.11,12 Our co-precipitation experiments document for the first time the physi-cal interaction between the two factors, that is mediated by the carboxyl-terminal zinc fingers of ZNF521, similar to ZNF423.15 Deletion of this region abrogates both the binding ability as well as the inhibitory activity of ZNF521. These data complement and enrich the observation of Hentges et al.19 that a mutant of Zfp521, lacking the final six zinc-fingers, fails to repress EBF1 activity in HEK293 cells.
Using a deletion mutant of ZNF521 that lacks the NuRD-binding domain located at the N-terminal end of ZNF521 we show that the integrity of this motif is dispensable for the repres-sion of EBF1-dependent promoters as well as of the endogenous EBF1 target genes in B cells. This is in full agreement with the fact that, although the cognate protein ZNF423 contains an NBD almost completely identical to that of ZNF521,16 the cDNAs used in the initial functional studies11,12,37 lacked the sequence encod-ing these amino acids, yet their products proved highly efficient
with the FG12 (H11) lentivirus previously described in reference 20, and murine bone marrow-derived lin- (mLin-) cells with a commercial lentiviral vector carrying a Zfp521-specific shRNA (Sigma). These vectors effectively knocked down the expression of the endogenous ZNF521/Zfp521 protein (Fig. 5A) in human THP-1 and mouse MS-5 cells, that produce high amounts of the factor. Once transduced with the viruses described, hCD34+ and mLin- cells were co-cultured with the stromal cells MS-5 and OP-9 respectively, in conditions that lead to efficient gen-eration of B lymphocytes in vitro.21,22 Flow-cytometric analyses were performed using antibodies to human (CD10/CD19) or murine (B220/CD19) B-cell surface markers and in both cases revealed a considerable (over two-fold) increase in the percentage of B cells in the cultures transduced with the specific shRNA lentiviruses compared with those infected with control vec-tors (Fig. 5B and C). As illustrated in Figure 6, an increase in total cell numbers was observed in the co-cultures of hCD34+ (Fig. 6A) or mLin- (Fig. 6B) cells transduced with the shRNAs to ZNF521/Zfp521 compared with those harbouring cells trans-duced with the control vectors. Combined with the presence of a larger fraction of B cells in these cultures, this resulted in a prominent increase in the absolute numbers of B-lymphocytes (Fig. 6C and D). Taken together, these data indicate that the silencing of zinc finger protein 521 in primitive hematopoietic progenitors, both human and murine, is sufficient to strongly enhance/accelerate B-lymphocyte production under permissive conditions.
Discussion
The two related transcription co-factors, zinc finger protein 423 and 521, are increasingly recognized as important regulators of cell fate in a variety of tissues,11,13,25-36 and in particular the latter has been credited with a critical role in the control of chondro-genesis33 and osteogenesis30,35 and has been proven essential for
Figure 4. Decline in Zfp521 expression during the B-cell differentiation of murine hematopoietic progenitors. (A and B) mouse lin- cells purified from bone marrow of C57BL/6 mice were plated on semi-confluent op-9 stromal cells in the presence of human IL-7 (10 ng/mL) and human Flt-3 ligand (10 ng/mL) to induce B cell differentiation. At the times indicated, the expression of Zfp521 (A) and of B-lineage markers (B) was assayed by Q-Rt-pCR. Data are from one experiment analyzed in duplicate.

©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2135
multiple interactions of ZNF521 with epigenetic effectors may mediate its corepressor functions in different molecular contexts. In future studies, it will be of interest to investigate whether either of these or other histone deacetylases are implicated in the repres-sion of EBF1-target genes by ZNF521. It will also be interesting to study whether ZNF521 may interfere with a novel epigenetic regulatory mechanism recently discovered in a genome-wide study of B-lymphoid differentiation by which, in addition to mediating transcriptional activation or repression of specific sets of target genes, EBF1 induces chromatin modifications in
in antagonizing the trans-activation of EBF1-dependent promot-ers. Thus, whereas this motif appears to be implicated in other activities of ZNF521, like the inhibition of erythroid differentia-tion of K-562 cells31 and the stimulation of clonogenic growth and tumorigenicity in medulloblastoma cells (R. Spina et al., in preparation), its contribution to EBF1 inhibition seems marginal or irrelevant. Interestingly, Correa et al.33 and Hesse et al.35 have recently demonstrated that the inhibition of Runx2 by Zfp521 in chondrocytes and osteoblasts depends on HDAC4 and HDAC3 respectively, but not on the NuRD complex, suggesting that
Figure 5. shRNA-mediated silencing of zinc finger protein 521 gene in mouse and human hematopoietic progenitors enhances B-cell differentiation. (A) Knock-down of ZNF521 and Zfp521 by RNAi. the human tHp1 and murine MS5 cell lines were transduced with the relevant shRNA to zinc finger protein 521, or with the control non target shRNA. protein gel blot analysis of the endogenous ZNF521/Zfp521 protein levels was performed on nuclear extracts from transduced cells. (B) Human CD34-positive cells were purified and transduced with the lentiviral control (CtRL) vector FG12 and with the ZNF521-specific shRNA vector FG12-H11, previously described (La Rocca et al. 2009). Cells were co-cultured with MS-5 cells to induce B-cell differentia-tion. After 14 and 21 d cells were analyzed by FACS for GFp, CD19-pe and CD10-pe-Cy5. the expression of the relevant antigens in GFp-positive cells, that represented 43–47% of the total population, is compared. Results are from a representative experiment of a series of two. (C) Mouse lin- cells puri-fied from bone marrow of C57BL/6 mice were transduced with the shRNA vectors, pLKo-puro-no-target (CtRL) and pLKo-puro-p15 (Zfp521-specific) and selected with puromycin for 24 h. Cells were then co-cultured with op-9 cells in the presence of human IL-7 (10 ng/mL) and human Flt-3 L (10 ng/mL) to induce B-cell differentiation. From day 5 to day 15 cells were assayed by FACS with CD45R/B220-FItC and CD19-pe. Results are from a represen-tative experiment of a series of six.

©2011 Landes Bioscience.Do not distribute.
2136 Cell Cycle Volume 10 Issue 13
an impact on B-lymphopoiesis. The data illustrated in this paper show that this is indeed the case: in transient overexpression experiments, ZNF521 downregu-lated the expression of an EBF1-reporter construct in a variety of B-lymphoid cell lines, and in addi-tion stable enforced expression of the protein in IM-9 cells repressed endogenous EBF1-target genes in a dose-dependent fashion. The study of primary human and murine hematopoietic progenitors further substantiated these data. In these cells, RNAi-mediated silencing of the endogenous ZNF521/Zfp521 gene resulted in a pronounced increase in the devel-opment of B cells compared with that of control cultures.
Zfp521 has been implicated in the development of B lymphoid neoplasias, including leukemias and lymphomas,14,20,43 in experi-mental animals. It is conceivable that aberrant expression of this gene in B-cell progenitors, in co-operation with concomitant oncogenic lesions or in mice with a genetic background predisposed to lymphoid malignancies, may contribute to the development and/or maintenance of immature leukemic cells by impeding dif-ferentiation. However, analysis
of publicly available leukemia gene expression data sets failed to detect significant levels of ZNF521 mRNA in human B-ALLs (ref. 13; M. Mesuraca et al. in preparation). This suggests that, if at all present, inappropriate expression of ZNF521 may be restricted to a minor subpopulation of malignant cells, probably those with more immature features. The sole case of a chromo-somal aberration involving ZNF521 detected in human B-ALL is that reported by Mullighan et al.44 who found a translocation that generated a PAX5/ZNF521 fusion transcript containing most of the coding sequences of the two factors. However, it is problematic to assess the contribution of ZNF521 to the patho-genesis of this leukemia since the translocation was associated with a PAX5 null allele. This would in turn result in severely reduced (or absent) PAX5 activity, that may per se impair the normal B-cell maturation.
Taken together, the results illustrated in this paper indicate that ZNF521 participates in the control of B-lymphopoiesis act-ing as a potent inhibitor of EBF1 in hematopoietic progenitors, and strongly support the view that this factor may contribute to maintain primitive lympho-myeloid progenitors in a multipotent
the regulatory regions of other genes, thereby priming them for expression at later stages of B-cell differentiation.38
ZNF521 is a transcriptional co-regulator with a complex struc-ture and a variety of molecular partners, and it is likely that its biological activities may differ in different cell types. For instance, Correa et al.33 have recently demonstrated that Zfp521 specifically stimulates cell proliferation in response to PTHrP, by inducing cyclin D1 expression, in chondrocytes but not in fibroblasts or osteoblasts. ZNF521 and EBF1 are co-expressed in a variety of cells, including differentiating hematopoietic progenitors, stria-tal neurons,39,40 cerebellar granule layer cells,13 medulloblastoma cells (Bond HM, unpublished), mesenchymal stromal cells and osteoblast precursors,30,33,41,42 and probably others. The biological significance of this co-expression is currently largely unknown and it is entirely plausible that the interaction of the two factors may result in either co-operation or antagonism, depending on the specific cell type where it occurs. It was thus of relevance to establish whether the ZNF521-mediated inhibition of EBF1, pre-viously investigated by co-transfection experiments in HEK293 cells, does take place in cells of B-lymphoid origin and does have
Figure 6. shRNA-mediated silencing of zinc finger protein 521 gene in mouse and human hematopoietic progenitors results in the generation of increased numbers of B cells. (A) the cells present in the superna-tant of the co-cultures of human CD34-positive cells illustrated in Figure 5B were counted at the indi-cated time points. the absolute numbers of total GFp-positive cells and of CD19/CD10-positive cells were calculated based on the percentage of total cells positive for the relevant markers as determined by FACS analysis. (B) the cells present in the supernatant of co-cultures of mouse lin- cells similar to those illustrated in Figure 5C were counted at the indicated time points and analyzed by flow cytometry with CD45R/B220-FItC and CD19-pe antibodies. the absolute numbers B220/CD19-positive cells were calculated based on the fraction of total cells expressing the relevant antigens.

©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2137
detect Flag-ZNF521 and mutants thereof in Protein gel blot-ting. The monoclonal for myc-tag and anti-myc-HRP were from Invitrogen. Rabbit anti-HDAC1 (Sigma-Aldrich) and goat anti-HDAC2 were used at a 1:5,000 dilution and detected with anti rabbit/goat secondary HRP-conjugated antibodies (from Santa Cruz).
A rabbit antibody to an internal ZNF521 peptide (VEAAPPIPKSRGRKR) was produced (Primm, Milan, Italy) and affinity-purified. In non-reducing conditions this antibody recognizes the dimeric form of ZNF521 at 300 kDa in Protein gel blotting.
Co-immunoprecipitation and protein gel blotting. HEK293 cells (2 x 107 cells) were transfected, by calcium phosphate, with combinations of plasmids containing the cDNA for Flag-ZNF521, Flag-ZNF521(ZF1–20), Flag-ZNF521(ZF24–30), FLAG-ZNF521 ΔNBD, EBF-myc or control plasmid 3xFlag 7.1-CMV. After 48 h the cells were collected, washed with PBS and lysed by sonication with IP buffer (15 mM Tris/HCl pH 7.5. 120 mM NaCl, 25 mM KCl, 1 mM EDTA, 0.1 mM DTT, 1% TritonX100) containing mammalian protease inhibitor cocktail (Sigma-Aldrich). Debris was removed by centrifugation (10,000 g, 20 min). Pre-clearing was performed with Protein A Sepharose. Soluble extracts were incubated with anti-flag M2-agarose (Sigma-Aldrich) or anti-myc antibody (Invitrogen) followed by Protein A Sepharose. Immuno-precipitates were washed five times with IP buffer at 1,000 g for 3 min. The soluble
state by antagonizing their irreversible commitment to terminal B-lymphoid differentiation orchestrated by EBF1.
The influence of ZNF521 on fate choice of hematopoietic progenitors is not limited to the B-lymphoid commitment. Matsubara et al.31 have recently reported that ZNF521 binds to GATA1 through the region encompassing ZFs 21–26 and represses a GATA1-responsive promoter, and that its silencing in the erythromyeloid cell lines, K-562 and HEL results in sponta-neous erythroid differentiation of these cells. Further studies will be required to draw a comprehensive picture of the full spectrum of molecular interactions and of biological functions of ZNF521. However, the data thus far available begin to delineate a role for this factor as one of the key regulators of the homeostasis of the immature cell compartment of the hematopoietic system.
Methods
Cell lines and culture conditions. HEK293T and HeLa cells were cultured in Dulbecco’s modified Eagle medium and the human B-cell lines, IM-9, ROHA-9 MC3, DeFew in RPMI 1640, supplemented with 10% fetal bovine serum, 50 U of peni-cillin and 50 μg of streptomycin/ml, at 37°C in 5% CO
2. All
tissue culture reagents were from Invitrogen.Expression vectors. The ZNF521 expression plasmids
p3xFlagCMV7.1-ZNF521 has been previously described in refer-ence 12. Three additional deletion mutants, ZNF521(ZF 1–20) ZNF521(ZF 24–30) and ZNF521 ΔNBD (lacking the NuRD-binding motif) were obtained by PCR, sequenced and subcloned into the HindIII and BamHI sites of the p3xFlag7.1-CMV vec-tor. The lentiviral vectors FUIGW-ZNF521 and FG12-H11 were previously described in reference 20. For mouse Zfp521 silenc-ing, the Mission shRNA vector p15 (target sequence: CGG CCC AUA UAU AUA UUU GUA, from Sigma-Aldrich) was found effective and used in the experiments described. As a control vector, the non-target Mission lentiviral vector was used. The plasmids B29-luciferase, mb-1-luciferase, lambda5-luciferase and pCDNA3.EBF-6xmyc were kindly provided by Dr. Mikael Sigvardsson (Linköping University, Sweden).
Quantitative RT-PCR. cDNA was synthesized from 1 μg total cellular RNA using SuperScript III reverse transcriptase and 2.5 μM random hexamers (Invitrogen). Q-PCR reactions were performed with the GoTaq qPCR SYBR green mastermix (Promega) in triplicate according to the manufacturer’s instruc-tions and analyzed using iQ5 multicolor detection system (Bio-Rad). One cycle of 3 min at 95°C was followed by 45 cycles of 10 sec at 95°C, 10 sec at 60°C and 20 sec at 72°C and then a melting curve. Relative gene expression was determined using the comparative threshold cycles Ct method, normalizing for endogenous GAPDH and the expression ratio was calculated as 2-ddCt. The specificity of amplification was confirmed by the dis-sociation curves of the amplicons. All data are mean values and standard deviations from at least three independent experiments using different RNA preparations. Primers used for Q-PCR are listed in Table 1.
Antibodies. The HRP-conjugated anti-flag monoclonal anti-body (M2, Sigma-Aldrich) was used at a 1:10,000 dilution to
Table 1. primers used for Q-Rt-pCR
Human genes
GApDH-FWD CAC CAt Ctt CCA GGA GCG AG
GApDH-ReV tCA CGC CAC AGt ttC CCG GA
ZNF521-FWD CCA CAt CCA AAC CAt CCA CCG
ZNF521-ReV CAG GtG GCA CtG GAG ttt GGC
B29-FWD GCA GAG CCC ACG ttt CAt
B29-ReV CGG AtG CCt tGG AtG Gt
VpReB-FWD AtG tCC tGG GCt CCt GtC C
VpReB-ReV tCA AGG GAC ACG tGt CCt G
MB-1-FWD CCA GCA tCA ttG AtG GtG AGC C
MB-1-ReV CAG CGt ACC Att GGG GtC Ct
LAMBDA 5-FWD CGt CCt CtG AGG AGC tCC A
LAMBDA 5-ReV AtC tCC ACG CCC tGG GtG A
Mouse genes
Gapdh-FWD tGA CGt GCC GCC tGG AGA AA
Gapdh-ReV AGt GtA GCC CAA GAt GCC Ctt CAG
Zfp521-FWD GCG ACA GCG CAG tCC AA
Zfp521-ReV GAt CtC GGt ttC GCt tGC tt
pax5-FWD CtA CAG GCt CCG tGA CGC AG
pax5-ReV GtC tCG GCC tGt GAC AAt AGG
Mb1-FWD tGG GAG ACG AtG CCA GGG GG
Mb1-ReV tGG CAC tGG GGG CCA tGt GA
ebf1-FWD ACC tGC ACC AGC ACC AAC GG
ebf1-ReV CCt GCt CtG GCA GAC CCC Ct

©2011 Landes Bioscience.Do not distribute.
2138 Cell Cycle Volume 10 Issue 13
Mouse Hematopoietic Progenitor Enrichment kit (StemCell Technologies) according to the manufacturer’s instructions. This purification, assessed by flow-cytometry, typically yielded >80% Lin-, c-kit+ cells. Cells were cultured in Opti-MEM1 (Gibco) containing 10% FBS (Biochrom), IL-3 (6 ng/mL), IL-6 (10 ng/mL) and SCF (50 ng/mL) (all from Peptrotech) over-night prior to, and during the course of, the lentiviral infection. This consisted of two consecutive rounds (8–12 h) of incubation with the virus-containing 293T supernatants in the presence of 6 μg/mL of polybrene (Sigma). In the first round, the cells were centrifuged at 2,000 rpm for 45 min at 32°C. The cells were then washed, subjected to 24-h puromycin selection (1 μg/ml) and plated in triplicate over sub-confluent OP-9 monolayers in 6-well plates (3 x 104 Lin- cells/well) in Opti-MEM1 supplemented with 10% FBS (Biochrom), hIL-7 (10 ng/mL) and hFlt-3 ligand (10 ng/mL) (Peprotech) to induce B cell differentiation. The cultures were demi-depopulated every 5 d for the first 10 d and then every other day, and the cells collected were assayed by flow cytometry for B-cell-specific antigens (CD45R/B220-PE or -FITC conjugated and CD19-PE, BD Biosciences) using a FACScan II (Becton-Dickinson). The data were analyzed with Flow Jo software (TreeStar, Inc.).
Statistics. The Student’s t-test assuming unequal variances between two samples was used to determine the significance of differences between the values of different samples where indi-cated. Groups were judged to differ significantly at p values lower than 0.05.
Acknowledgments
This work was supported in part by funds from: Italian Ministry for University and Research (Interlink to G.M. and PRIN to H.M.B.); Italian Association for Cancer Research (AIRC) to G.M.; Leukemia and Lymphoma Society Specialized Center of Research Program and Gar Reichman Cancer Research Institute Fund to M.A.S.M.; Leukemia Research Foundation to S.J.H. and J.J.S.; Dutch Cancer Foundation (KWF) and Netherlands Organization for Scientific Research (NWO-VIDI) to J.J.S. We are grateful to Dr. Mikael Sigvardsson (Linköping University, Sweden) for the constructs B29-luc, mb-1-luc, lambda5-luc and pCDNA3.EBF-6xmyc. TM is supported by a post-doctoral fel-lowship (assegno di ricerca) co-funded by the Medical School of the University of Catanzaro; M.L., R.S. and E.C. are fellows of the Ph.D., Programme in Molecular Oncology. We are indebted to Stefania Scicchitano, a graduate student in the laboratory of molecular hematopoiesis, for collaboration in some of the experi-ments described in the paper.
extracts and bound proteins were electrophoresed on NuPAGE Novex Bis-Tris 4–12% gels under reducing conditions, elec-troblotted onto nitrocellulose membranes, stained with Ponceau S and blocked with 5% blocking reagent (BioRad). Antigens were detected using directly conjugated antibodies, anti-Flag M2-HRP or anti-myc-HRP and Chemi-luminescence Luminol Reagent (Santa Cruz).
Cell transfections and luciferase assays. Cells were trans-fected by calcium phosphate (HEK293, HeLa) or electropora-tion (IM-9, ROHA-9 MC3, DeFew). After 48–72 h, extracts were prepared by repeated freezing and thawing in 250 mM Tris/HCl pH 7.5 and assayed for β-galactosidase activity with o-nitrophenyl-β-D-galactoside as a substrate or the Dual-Luciferase Reporter Assay System (Promega) to normalize for transfection and extraction efficiencies. Luciferase assays were performed using the Luciferase Reporter assay system (Promega). Measurements were performed using a TD-20/20 luminometer (Turner Designs).
Lentiviral transductions. The production of the lentivi-ral vectors described in this paper was performed as previously described in reference 20. The transduction of IM-9 cells and the isolation of the subclones 4 and 9 were also detailed in the same paper. The transduction of murine and human progenitors was performed as detailed below.
Isolation, transduction and MS-5 co-culture of human CD34+ cells. Neonatal cord blood was obtained after informed consent from the obstetrics departments of the University Medical Centre Groningen. After Ficoll separation of mononu-clear cells, CD34+ cells were isolated using MiniMACS (Miltenyi Biotech). CD34+ cells were pre-stimulated for 48 h in serum-free HPG medium (Lonza) containing 100 ng/mL stem cell factor, 100 ng/mL Flt3 ligand (Amgen) and 100 ng/mL thrombopoi-etin (Kirin). CD34+ cells were transduced in two consecutive rounds of 8–12 h on retronectin-coated plates (Takara). After transduction 50,000 cells were plated in T25 flasks containing pre-plated MS-5 cells and grown in α-MEM supplemented with 12.5% heat-inactivated FCS, penicillin and streptomycin, 57.2 μM β-mercaptoethanol and 50 μg/mL vitamin C. Half of the cultures were harvested weekly for analysis.
Isolation, transduction and OP-9 co-culture of murine lin- cells. All the mouse studies were conducted in compliance with the Italian and European regulations for animal studies, and in particular with the protocols approved by the Institutional Review Board of the institutions where the procedures were per-formed. Bone marrow hematopoietic progenitors from C57BL/6 mice were enriched by lineage depletion with the Easy Sep
References1. Welner RS, Pelayo R, Kincade PW. Evolving views on
the genealogy of B cells. Nat Rev Immunol 2008; 8:95-106; DOI: 10.1038/nri2234.
2. Mandel EM, Grosschedl R. Transcription control of early B cell differentiation. Curr Opin Immunol 2010; 22:161-7; DOI: 10.1016/j.coi.2010.01.010.
3. Bryder D, Sigvardsson M. Shaping up a lineage-lessons from B lymphopoesis. Curr Opin Immunol 2010; 22:148-53; DOI: 10.1016/j.coi.2010.02.001.
4. Ramírez J, Lukin K, Hagman J. From hematopoietic progenitors to B cells: mechanisms of lineage restriction and commitment. Curr Opin Immunol 2010; 22:177-84; DOI: 10.1016/j.coi.2010.02.003.
5. Lukin K, Fields S, Hartley J, Hagman J. Early B cell factor: Regulator of B lineage specification and com-mitment. Semin Immunol 2008; 20:221-7; DOI: 10.1016/j.smim.2008.07.004.
6. Lin H, Grosschedl R. Failure of B-cell differentiation in mice lacking the transcription factor EBF. Nature 1995; 376:263-7; DOI: 10.1038/376263a0.
7. Zandi S, Mansson R, Tsapogas P, Zetterblad J, Bryder D, Sigvardsson M. EBF1 is essential for B-lineage prim-ing and establishment of a transcription factor network in common lymphoid progenitors. J Immunol 2008; 181:3364-72.
8. Pongubala JM, Northrup DL, Lancki DW, Medina KL, Treiber T, Bartolino E, et al. Transcription factor EBF restricts alternative lineage options and promotes B cell fate commitment independently of Pax5. Nat Immunol 2008; 9:203-15; DOI: 10.1038/ni1555.

©2011 Landes Bioscience.Do not distribute.
www.landesbioscience.com Cell Cycle 2139
34. Masserdotti G, Badaloni A, Green YS, Croci L, Barili V, Bergamini G, et al. ZFP423 coordinates Notch and bone morphogenetic protein signaling, selectively upregulating Hes5 gene expression. J Biol Chem 2010; 285:30814-24; DOI: 10.1074/jbc.M110.142869.
35. Hesse E, Saito H, Kiviranta R, Correa D, Yamana K, Neff L, et al. Zfp521 controls bone mass by HDAC3-dependent attenuation of Runx2 activity. J Cell Biol 2010; 191:1271-83; DOI: 10.1083/jcb.201009107.
36. Kamiya D, Banno S, Sasai N, Ohgushi M, Inomata H, Watanabe K, et al. Intrinsic transition of embry-onic stem-cell differentiation into neural progenitors. Nature 2011; 470:503-9; DOI: 10.1038/nature09726.
37. Hata A, Seoane J, Lagna G, Montalvo E, Hemmati-Brivanlou A, Massague J. OAZ uses distinct DNA- and protein-binding zinc fingers in separate BMP-Smad and Olf signaling pathways. Cell 2000; 100:229-40; DOI: 10.1016/S0092-8674(00)81561-5.
38. Treiber T, Mandel EM, Pott S, Györy I, Firner S, Liu ET, et al. Early B cell factor 1 regulates B cell gene networks by activation, repression and transcription-independent poising of chromatin. Immunity 2010; 32:714-25; DOI: 10.1016/j.immuni.2010.04.013.
39. Lobo MK, Karsten SL, Gray M, Geschwind DH, Yang XW. FACS-array profiling of striatal projection neuron subtypes in juvenile and adult mouse brains. Nat Neurosci 2006; 9:443-52; DOI: 10.1038/nn1654.
40. Lobo MK, Yeh C, Yang XW. Pivotal role of early B-cell factor 1 in development of striatonigral medium spiny neurons in the matrix compartment. J Neurosci Res 2008; 86:2134-46; DOI: 10.1002/jnr.21666.
41. Lagergren A, Månsson R, Zetterblad J, Smith E, Basta B, Bryder D, et al. The Cxcl12, periostin and Ccl9 genes are direct targets for early B-cell factor in OP-9 stroma cells. J Biol Chem 2007; 282:14454-62; DOI: 10.1074/jbc.M610263200.
42. Kieslinger M, Folberth S, Dobreva G, Dorn T, Croci L, Erben R, et al. EBF2 regulates osteoblast-dependent differentiation of osteoclasts. Dev Cell 2005; 9:757-67; DOI: 10.1016/j.devcel.2005.10.009.
43. Yamasaki N, Miyazaki K, Nagamachi A, Koller R, Oda H, Miyazaki M, et al. Identification of Zfp521/ZNF521 as a cooperative gene for E2A-HLF to develop acute B-lineage leukemia. Oncogene 2010; 29:1963-75; DOI: 10.1038/onc.2009.475.
44. Mullighan CG, Goorha S, Radtke I, Miller CB, Coustan-Smith E, Dalton JD, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukemia. Nature 2007; 446:758-64; DOI: 10.1038/nature05690.
21. Rawlings DJ, Quan SG, Kato RM, Witte ON. Long-term culture system for selective growth of human B-cell progenitors. Proc Natl Acad Sci USA 1995; 92:1570-4; DOI: 10.1073/pnas.92.5.1570.
22. Vieira P, Cumano A. Differentiation of B lymphocytes from hematopoietic stem cells. Methods Mol Biol 2004; 271:67-76.
23. van Zelm MC, van der Burg M, de Ridder D, Barendregt BH, de Haas EF, Reinders MJ, et al. Ig gene rearrangement steps are initiated in early human precursor B cell subsets and correlate with spe-cific transcription factor expression. J Immunol 2005; 175:5912-22.
24. Chambers SM, Boles NC, Lin KY, Tierney MP, Bowman TV, Bradfute SB, et al. Hematopoietic fin-gerprints: an expression database of stem cells and their progeny. Cell Stem Cell 2007; 1:578-91; DOI: 10.1016/j.stem.2007.10.003.
25. Warming S, Rachel RA, Jenkins NA, Copeland NG. Zfp423 is required for normal cerebellar development. Mol Cell Biol 2006; 26:6913-22; DOI: 10.1128/MCB.02255-05.
26. Alcaraz WA, Gold DA, Raponi E, Gent PM, Concepcion D, Hamilton BA. Zfp423 controls pro-liferation and differentiation of neural precursors in cerebellar vermis formation. Proc Natl Acad Sci USA 2006; 103:19424-9; DOI: 10.1073/pnas.0609184103.
27. Cheng LE, Reed RR. Zfp423/OAZ participates in a developmental switch during olfactory neurogen-esis. Neuron 2007; 54:547-57; DOI: 10.1016/j.neu-ron.2007.04.029.
28. Cheng LE, Zhang J, Reed RR. The transcription factor Zfp423/OAZ is required for cerebellar development and CNS midline patterning. Dev Biol 2007; 307:43-52; DOI: 10.1016/j.ydbio.2007.04.005.
29. Huang S, Laoukili J, Epping MT, Koster J, Holzel M, Westerman BA, et al. ZNF423 is critically required for retinoic acid-induced differentiation and is a marker of neuroblastoma outcome. Cancer Cell 2009; 15:328-40; DOI: 10.1016/j.ccr.2009.02.023.
30. Wu M, Hesse E, Morvan F, Zhang JP, Correa D, Rowe GC, et al. Zfp521 antagonizes Runx2, delays osteoblast differentiation in vitro, and promotes bone formation in vivo. Bone 2009; 44:528-36; DOI: 10.1016/j.bone.2008.11.011.
31. Matsubara E, Sakai I, Yamanouchi J, Fujiwara H, Yakushijin Y, Hato T, et al. The role of zinc finger protein 521/early hematopoietic zinc finger protein in erythroid cell differentiation. J Biol Chem 2009; 284:3480-7; DOI: 10.1074/jbc.M805874200.
32. Gupta RK, Arany Z, Seale P, Mepani RJ, Ye L, Conroe HM, et al. Transcriptional control of preadipocyte determination by Zfp423. Nature 2010; 464:619-23; DOI: 10.1038/nature08816.
33. Correa D, Hesse E, Seriwatanachai D, Kiviranta R, Saito H, Yamana K, et al. Zfp521 is a target gene and key effector of parathyroid hormone-related peptide signaling in growth plate chondrocytes. Dev Cell 2010; 19:533-46; DOI: 10.1016/j.devcel.2010.09.008.
9. Smith EM, Akerblad P, Kadesch T, Axelson H, Sigvardsson M. Inhibition of EBF function by active Notch signaling reveals a novel regulatory pathway in early B-cell development. Blood 2005; 106:1995-2001; DOI: 10.1182/blood-2004-12-4744.
10. Oguro H, Yuan J, Ichikawa H, Ikawa T, Yamasaki S, Kawamoto H, et al. Poised lineage specification in multipotential hematopoietic stem and progenitor cells by the polycomb protein Bmi1. Cell Stem Cell 2010; 6:279-86; DOI: 10.1016/j.stem.2010.01.005.
11. Tsai RY, Reed RR. Cloning and functional charac-terization of Roaz, a zinc finger protein that interacts with O/E-1 to regulate gene expression: implications for olfactory neuronal development. J Neurosci 1997; 17:4159-69.
12. Bond HM, Mesuraca M, Carbone E, Bonelli P, Agosti V, Amodio N, et al. Early hematopoietic zinc finger protein (EHZF), the human homolog to mouse Evi3, is highly expressed in primitive human hematopoietic cells. Blood 2004; 103:2062-70; DOI: 10.1182/blood-2003-07-2388.
13. Bond HM, Mesuraca M, Amodio N, Mega T, Agosti V, Fanello D, et al. Early hematopoietic zinc finger protein-zinc finger protein 521: a candidate regulator of diverse immature cells. Int J Biochem Cell Biol 2008; 40:848-54; DOI: 10.1016/j.biocel.2007.04.006.
14. Warming S, Liu P, Suzuki T, Akagi K, Lindtner S, Pavlakis GN, et al. Evi3, a common retroviral inte-gration site in murine B-cell lymphoma, encodes an EBFAZ-related Kruppel-like zinc finger protein. Blood 2003; 101:1934-40; DOI: 10.1182/blood-2002-08-2652.
15. Tsai RY, Reed RR. Identification of DNA recognition sequences and protein interaction domains of the multiple-Zn-finger protein Roaz. Mol Cell Biol 1998; 18:6447-56.
16. Lin AC, Roche AE, Wilk J, Svensson EC. The N termini of Friend of GATA (FOG) proteins define a novel transcriptional repression motif and a superfam-ily of transcriptional repressors. J Biol Chem 2004; 279:55017-23; DOI: 10.1074/jbc.M411240200.
17. Gregory GD, Miccio A, Bersenev A, Wang Y, Hong W, Zhang Z, et al. FOG1 requires NuRD to promote hematopoiesis and maintain lineage fidelity within the megakaryocytic-erythroid compartment. Blood 2010; 115:2156-66; DOI: 10.1182/blood-2009-10-251280.
18. Hong W, Nakazawa M, Chen YY, Kori R, Vakoc CR, Rakowski C, et al. FOG-1 recruits the NuRD repres-sor complex to mediate transcriptional repression by GATA-1. EMBO J 2005; 24:2367-78; DOI: 10.1038/sj.emboj.7600703.
19. Hentges KE, Weiser KC, Schountz T, Woodward LS, Morse HC, Justice MJ. Evi3, a zinc-finger protein related to EBFAZ, regulates EBF activity in B-cell leu-kemia. Oncogene 2005; 24:1220-30; DOI: 10.1038/sj.onc.1208243.
20. La Rocca R, Fulciniti M, Lakshmikanth T, Mesuraca M, Ali TH, Mazzei V, et al. Early hematopoietic zinc finger protein prevents tumor cell recognition by natu-ral killer cells. J Immunol 2009; 182:4529-37; DOI: 10.4049/jimmunol.0802109.
Copyright © 2022 FDOKUMEN




















