
Worms and Human Disease 2nd Edition
310
Worms and Human Disease 2nd Edition
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Worms and Human Disease 2nd Edition

Worms and Human Disease
2nd Edition
00Worms & H.D. - Prelims. 20/11/01 1:41 PM Page i

Dedicated to the memory of Annie
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page ii

Worms and Human Disease2nd Edition
Ralph Muller, DSc, PhD, FIBiol
Department of Infectious and Tropical DiseasesLondon School of Hygiene and Tropical Medicine
University of London, UK
and
Former DirectorInternational Institute of Parasitology
St Albans, Hertfordshire, UK
With contributions and the chapter on Immunology of Helminths from
Derek Wakelin, DSc, PhD, FRCPath
School of Life and Environmental SciencesUniversity of Nottingham, UK
CABI Publishing
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page iii

CABI Publishing is a division of CAB International
CABI Publishing CABI PublishingCAB International 10 E 40th StreetWallingford Suite 3203Oxon OX10 8DE New York, NY 10016UK USATel: +44 (0)1491 832111 Tel: +1 212 481 7018Fax: +44 (0)1491 833508 Fax: +1 212 686 7993Email: [email protected] Email: [email protected] site: www.cabi-publishing.org
© CAB International 2002. All rights reserved. No part of thispublication may be reproduced in any form or by any means, electronically, mechanically, by photocopying, recording or otherwise, without prior permission of the copyright owners.
A catalogue record for this book is available from the British Library, London, UK.
Library of Congress Cataloging-in-Publication DataMuller, Ralph, 1933-
Worms and human disease / Ralph Muller ; with contributions and the chapter onimmunology from Derek Wakelin.- - 2nd ed.
p. ; cm.Rev. ed. of: Worms and disease / Ralph Muller. c1975.Includes bibliographical references and index.ISBN 0-85199-516-0 (pbk.)
1. Medical helminthology. 2. Helminthiasis. I. Wakelin, Derek. II. Muller, Ralph,1933- Worms and disease. III. Title.
[DNLM: 1. Helminths--pathogenicity. 2. Helminthiasis. QX 200 M958w 2001]RC119.7 .M84 2001
2001025591
ISBN 0 85199 516 0
Typeset in Melior by Columns Design Ltd, Reading.Printed and bound in the UK by Biddles Ltd, Guildford and King’s Lynn.
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page iv

Contents
Acknowledgements ix
Introduction 1
1. The Trematodes 3Morphology 5Life Cycle Stages 6Classification 6Family Schistosomatidae 9
Schistosomes 9Family Paragonimidae 32
Paragonimus westermani 32Family Achillurbainiidae 37Family Opisthorchidae 38
Clonorchis sinensis 38Opisthorchis viverrini 43Opisthorchis felineus 44
Family Dicrocoeliidae 45Dicrocoelium dendriticum 45
Family Fasciolidae 46Fasciola hepatica 46Fasciolopsis buski 49
Family Heterophyidae 51Heterophyes heterophyes 51Metagonimus yokogawai 53
Family Paramphistomidae 55Gastrodiscoides hominis 55
Family Echinostomidae 56Echinostoma ilocanum 56Other Echinostomids 56
Other Occasional and Rare Human-parasitic Trematodes 58Family Diplostomidae 58Family Lecithodendriidae 58Family Plagiorchiidae 58Family Troglotrematidae 58Other families 59
v
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page v

2. The Cestodes 63Classification 64Order Pseudophyllidea 65
Diphyllobothrium latum 65Sparganosis 70
Order Cyclophyllidea 71Family Taeniidae 71
Taenia saginata 71Taenia solium 76Cysticercosis 80Taenia multiceps 83Echinococcus granulosus 85Echinococcus multilocularis 94Echinococcus oligarthrus 97Echinococcus vogeli 97
Family Hymenolepididae 98Hymenolepis nana 98Hymenolepis diminuta 101
Family Dipylididae 102Dipylidium caninum 102
Very occasional human tapeworms 102
3. The Acanthocephala 106Moniliformis moniliformis 106Macranthorhynchus hirudinaceus 107
4. The Nematomorpha 108
5. The Nematodes 109Classification 109A Key to Nematodes Parasitic in Humans 113Intestinal Nematodes 115Geohelminths 115Order Rhabditida 115
Family Strongyloididae 115Strongyloides stercoralis 115
Order Strongylida 126Family Ancylostomatidae 126
Necator americanus 126Ancylostoma duodenale 134
Other Strongyles 137Family Trichostrongylidae 138
Trichostrongylus spp. 138Family Chabertiidae 139
Ternidens deminutus 139Oesophagostomum bifurcum 140
Family Syngamidae 142Mammomonogamus laryngeus 142
Family Angiostrongylidae 143Parastrongylus cantonensis 143Parastrongylus costaricensis 145
vi Contents
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page vi

Order Ascaridida 147Family Ascarididae 147
Ascaris lumbricoides 147Lagochilascaris minor 153Baylisascaris procyonis 153
Family Anisakidae 154Anisakis and other anisakids 154
Larva Migrans 156Visceral larva migrans 156
Toxocara and Toxascaris 156Cutaneous marva migrans or ‘creeping eruption’ 159
Order Oxyurida 160Family Oxyuridae 160
Enterobius vermicularis 160Order Enoplida 164
Family Trichuridae 164Trichuris trichiura 164
Tissue Nematodes 173Order Enoplida 173
Family Trichuridae 173Aonchotheca philippinensis 173Calodium hepaticum 175Eucoleus aerophilus 175
Family Trichinellidae 176Trichinella spiralis 176
Family Dioctophymidae 184Dioctophyma renale 184
Superfamily Mermithoidea 184Order Spirurida 184
Superfamily Gnathostomoidea 184Gnathostoma spinigerum 184
Other Spirurids 187Superfamily Filarioidea: the Filariae 188
Wuchereria bancrofti 190Brugia malayi 202Loa loa 206Onchocerca volvulus 211Mansonella perstans 221Mansonella streptocerca 223Mansonella ozzardi 225Accidental filarial infections 226
Superfamily Dracunculoidea 228Dracunculus medinensis 228
6. Other (Non-helminth) Groups 240Pentastomes 240
Armillifer (= Porocephalus) armillata 240Linguatula serrata 240
Leeches 241Myiasis 241
Contents vii
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page vii

7. Immunology of Helminths 243
8. Epidemiological Aspects of Helminth Infections 252
9. Helminthological Techniques 255
Appendix 1 Summary of Some Landmarks in Medical Helminthology 271Appendix 2 Glossary of Helminthological Terms 273Appendix 3 Location of Helminths in the Human Body 279
General References and Further Reading 282
Index 287
viii Contents
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page viii

It is a pleasure to thank SmithKlineBeecham (now GlaxoSmithKline) for agrant that has made possible the plates ofcolour pictures, which have greatlyincreased the usefulness of the whole book,and Dr John Horton of SmithKlineBeecham for his advice and also for theprovision of colour photographs (Plates 24and 25). Thanks also to Dr R. Neafie forproviding photographs from the ArmedForces Institute of Pathology collection(Figs 54, 62, 66, 67, 68, 75, 77 and 96 andPlate 18), to Dr L. Savioli for photographsand a booklet from the World HealthOrganization (WHO) and to Professor T.Polderman for a photograph ofOesophagostum (Plate 17). Other pho-tographs were kindly provided by Dr J.Anderson (Fig. 100 and Plates 29 and 30),Dr Kemal Arab (Plate 23), Dr N. Ashton(Fig. 72), Prof. E.A. Bianco (Plate 27), Dr A.Bryceson (Fig. 46), Dr P. Choyce (Fig. 107),Dr D. Denham (Plate 21), Prof. D. Dennis(Plate 28), Dr F. Etges (Fig. 14), Dr R.Feachem (Plate 5), Dr D. Flavell (Plate 6),Dr H. Fuglsang (Fig. 106), Dr L.M. Gibbons(Figs 34, 57, 61 and 85), Prof. D.B.Holliman (Figs 64 and 71 and Plates 11, 12,13, and 16), Prof. S. Lucas (Plate 19), Dr D.
McLaren (Figs 5 and 80), Prof. C.Macpherson (Plate 10), Prof. Y. Matsukado(Fig. 20), Prof. M. Murray (Plate 7), DrNagano (Plate 21), Prof. G.S. Nelson (Fig.44), Dr M.J. Taylor (Fig. 95), Dr A.C.Templeton (Fig. 40), Dr S. Townson (Plate31), Dr S. Vajrasthira (Fig. 18), Dr E. Watty(Plate 17), Prof. G. Webbe (Plate 3) and theWHO (Plates 26 and 32). Thanks to EdwardArnold and Professor D. Crompton for per-mission to reproduce Fig. 65, to CABInternational, and Dr P. Jordan and Dr R. Sturrock for permission to reproduceFig. 17 and to Dr J. Baker and Harcourt forpermission to publish Fig. 132.
My daughter Harriet kindly helped withmaps and drawings and I would also liketo acknowledge the help and informationprovided by many colleagues.
Professor Derek Wakelin has, in addi-tion to writing the sections on immunologyfor various groups of parasites as well asthe general chapter on the subject, provideduseful criticism and advice on statementson immunology and immunodiagnosisincluded in the treatments of many of theindividual parasites (possible errors inthese parts are, of course, not his responsi-bility).
Acknowledgements
ix
00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page ix

00Worms & H.D. - Prelims. 14/11/01 4:24 PM Page x

While this book is the second edition ofWorms and Disease: a Manual of MedicalHelminthology (1975), because of the longtime that has elapsed since the publicationof the earlier book, it has been so exten-sively revised and brought up to date thatvirtually every chapter has had to bealmost completely rewritten. In the inter-vening years the importance to humans ofsome new helminths has emerged, such asOesophagostomum bifurcum and Para-strongylus costaricensis, but principallythe changes have been necessitated by thegreat strides that have been made in knowl-edge of the diagnosis, treatment, immunol-ogy and molecular biology of parasites. Thechapter on the immunology of helminths(now written by Derek Wakelin) has beengreatly amplified, with the addition ofmore detailed paragraphs in the appropri-ate sections, together with the latest infor-mation on the prospects for specificvaccines. There has also been excitingprogress in the field of global control ofvarious helminths, such as the schisto-somes, soil-transmitted nematodes, filariae(both those causing lymphatic filariasis andthose causing onchocerciasis) and theguinea worm. Most of these campaignshave been possible because of recentadvances in chemotherapy and, in somecases, of diagnosis; many have been linkedwith efforts to improve sanitation and general health.
The book is intended principally as apractical guide in human helminthologyfor physicians and medical technologists
concerned with tropical and exotic dis-eases and for students taking postgraduatedegrees and diplomas in aspects of tropicaland infectious diseases. It should alsoprove useful as an accessory text and refer-ence source for undergraduate medical,zoological and tropical health engineeringstudents, and for medical technologists,microbiologists and physicians in temper-ate climates. With increase in air travel,most hospitals and medical practitioners indeveloped countries are meeting cases ofparasitic infections that may have beenvery rare occurrences in the past, and it isbecoming increasingly necessary to ask ofalmost all patients ‘Unde venis?’. Also, ifglobal warming increases, it is likely thatthe endemicity of some helminth infec-tions will extend to higher latitudes.
The format of this book is fairly conven-tional, with parasites considered in orderof their zoological relationships rather thantheir location in the body. The latterapproach may be useful for diagnosis but isnot practical for other aspects of the sub-ject, as some parasites can occupy a widerange of sites in the body, so that therewould be a great deal of repetition, andalso because the relationships betweenmany similar parasites that occupy differ-ent organs would be obscured. However,the various possible locations in the bodyof all the important helminths are shownin Fig. 132 and alternative diagnoses arediscussed in the appropriate individualsections. An attempt has also been made tohave the best of both worlds, e.g. all the
Introduction
1
01Worms Intro. & Chap 01 14/11/01 4:24 PM Page 1

intestinal nematodes are considered in thesame section so that the new global mea-sures being advocated for all the geo-helminths can be considered together, eventhough they are not all closely related.
Most of the figures for infection rateshave been obtained from the CD-ROMPARASITE database produced by CABInternational or from MEDLINE. Mapshave concentrated principally on helminthinfections that have a focal distribution.
The term helminth (Greek ��µ��) meansworm, although it is usually restricted tothe parasitic worms. The term does notrefer to any one zoological taxon but thosemembers parasitic in humans belongalmost entirely to two main groups; thephylum Platyhelminthes, which includesthe trematodes (flukes) and the cestodes(tapeworms), and the phylum Nematoda,comprising the nematodes (roundworms).This book provides a comprehensiveaccount of all important helminths foundin humans, with a mention of all others
reported, however occasionally (a total of267 species), and includes a brief consider-ation of other metazoan parasites some-times found in humans, such as thepentastomids, dipteran fly larvae andleeches, which may be confused with thetrue helminths.
While the title is Worms and HumanDisease, it must not be assumed thathelminth infection invariably results indisease; most of the helminths that are pre-dominantly human parasites are patho-genic only when worm burdens are highand, as there is no multiplication withinthe body, light infections become clinicallyimportant only following reinfection. Themajority of helminth infections are lightand cause little morbidity (although insome cases more than was previouslythought), but many are so widespread thatthe low percentage of patients who suffersevere clinical disease represents a prob-lem of great medical and economicimportance.
2 Introduction
01Worms Intro. & Chap 01 14/11/01 4:24 PM Page 2

Adult trematodes, or flukes, may be foundin the intestinal tract, bile-ducts, lungs orblood of humans. Some details concerningthe medically most important species areshown in Table 1. All the trematodes men-tioned in the table are normal human para-sites, except some species of Paragonimusand Fasciola and some heterophyids andechinostomes, which are accidental para-sites with humans not being involved intheir transmission cycles. However, almostall trematodes are very catholic in theirchoice of definitive hosts (a notable excep-tion is Schistosoma haematobium) andhave a wide range of animal reservoirs; 144species that have been found in humansare mentioned in the text, most of whichare natural animal parasites. Not shown inthe table are various aberrant forms, suchas the cercarial larvae of animal and birdschistosomes, which can penetrate the skinof humans but are not able to mature.
Pre-eminent in medical and economicimportance are the schistosomes, or bloodflukes, which are the cause of one of themajor human diseases, schistosomiasis.This is a source of suffering in many warmcountries and is a major cause of morbid-ity. No other trematode is the cause of suchwidespread morbidity, but liver flukes(Clonorchis and the closely relatedOpisthorchis) and lung flukes (Paragonimus)are important parasites in areas of Asia andtheir presence may result in severe diseaseand possibly death.
It needs to be stressed that the presenceof trematode parasites in the body is by no
means synonymous with the presence ofdisease. In contrast to viruses, bacteria orprotozoans, trematodes do not multiplywithin the human body and the few organ-isms present in the great majority ofinfected persons are tolerated with theminimum of inconvenience and are oftennot diagnosed. It is the small percentage ofpatients with large worm burdens (so-called ‘wormy people’) or in whom the par-asites or their eggs are in ectopic sites inthe body who give cause for alarm.
The digenetic trematodes are membersof the phylum Platyhelminthes, which alsoincludes the cestodes (tapeworms), mono-geneans (ectoparasites of fishes andamphibians) and free-living turbellarians(planarians, etc.). Platyhelminthes, or flat-worms, are acoelomate bilateria (bilaterallysymmetrical and lacking a coelom). Theexcretory system is based on the flame cell,or protonephridium, and often the patternof flame cells can be of importance in clas-sification. Trematodes are characteristicallyflat and leaflike, or occasionally globular,hermaphroditic organisms (except for theschistosomes, which have a male foldedabout its long axis and a cylindrical female(Figs 3 and 4)). All have complicated lifecycles with alternating sexual and asexualdevelopment in different hosts. Asexualmultiplication takes place in a snail, andfor parasites of medical importance this isalways a gastropod snail. It is believed thatthe trematodes were originally parasites ofmolluscs and they are still always very specific in their choice of snail host;
1The Trematodes
3

4C
hapter 1Table 1. Trematodes of medical importance.
Situation of Eggs recovered Snail intermediate Other intermediate GeographicalHabitat Species adult from host or transport hosts distribution
Blood Schistosoma Mesenteric veins Faeces Biomphalaria spp. None (active penetration Africa, South Americamansoni by cercariae)
S. japonicum Mesenteric veins Faeces Oncomelania spp. None (active penetration China, South-East Asiaby cercariae)
S. mekongi Mesenteric veins Faeces Neotricula None (active penetration Cambodia, Laosby cercariae)
S. intercalatum Mesenteric veins Faeces Bulinus spp. None (active penetration Central Africaby cercariae)
S. haematobium Vesicular veins Urine Bulinus spp. None (active penetration Africa, Middle Eastby cercariae)
Lungs Paragonimus Cysts in lungs Sputum and faeces Semisulcospirura Edible crustaceans South-East Asia, westermani Thiara containing China, Japan
Oncomelania metacercariaeParagonimus spp. Cysts in lungs Sputum and faeces Various Edible crustaceans South-East Asia,
containing West Africa, South andmetacercariae Central America
Liver Clonorchis sinensis Bile and pancreatic Faeces Bulimus Freshwater food fish South-East Asiaducts Parafossarulus containing metacercariae
Opisthorchis felineus Bile and pancreatic Faeces Bithynia Freshwater food fish Siberia, East Europeducts containing metacercariae
O. viverrini Bile and pancreatic Faeces Bithynia Freshwater food fish Thailand, Laosducts containing metacercariae
Fasciola hepatica Bile ducts Faeces Lymnaea Metacercariae encysted Cosmopolitan (mainly on plants temperate areas)
Intestine Fasciolopsis buski Small intestine Faeces Segmentina Metacercariae on South-East Asia, Indiawater plants
Heterophyes Small intestine Faeces Pirenella Freshwater food fish South-East Asia, heterophyes Cerithidea containing metacercariae Middle East, Egypt,
southern EuropeMetagonimus Small intestine Faeces Semisulcospira Freshwater food fish South-East Asia, Russia
yokogawai containing metacercariae (Siberia), southern EuropeOther heterophyids Small intestine Faeces Various Freshwater food fish Worldwide in warm
containing metacercariae countriesEchinostomes Small intestine Faeces Various Freshwater fish or snails Mostly South-East
containing metacercariae Asia, IndiaGastrodiscoides Caecum and colon Faeces Helicorbis Metacercariae on South-East Asia
hominis water plants

identification and study of the biology ofthe particular snails involved in transmis-sion form an important aspect of the epi-demiology of trematode diseases, known asmedical malacology.
Detailed consideration of snail inter-mediate hosts is outside the scope of thisbook. For the practical field worker it is nec-essary to consult a specialized monograph(e.g. Malek, 1963; Brown, 1980) or sendspecimens to an expert, as in most habitatsthere are species of snails present which donot transmit human helminth infections butclosely resemble those that do. Some of theimportant snail intermediate hosts involvedin the dissemination of trematodes of med-ical importance are shown in Fig. 1.
Morphology
The outer surface or tegument is a non-cellular syncytial extension of thesunken tegumental cells, and may havespines embedded in it. During the lastfew years a great deal of interest hasbeen shown in the physiology and finestructure of the tegument of both trema-todes and cestodes, because of its impor-tance in nutrition and in antigenicstimulation (it is considered in moredetail on p. 63 in the section on cestodes).
Two suckers are present in all trema-todes found in humans, an anterior oralsucker into which the alimentary canal
The Trematodes 5
Fig. 1. Snails that act as first intermediate hosts of the trematodes of medical importance. (a) Biomphalaria glabrata from Brazil, host of Schistosoma mansoni. (b) Bulinus (Physopsis) globosusfrom Nigeria and (c) Bulinus (Bulinus) truncatus from Iran, hosts of S. haematobium. (d) Oncomelaniahupensis nosophora from Japan, host of S. japonicum. (e) Thiara granifera from China and (f) Semisulcospira libertina from China, hosts of Paragonimus westermani and Metagonimus yokogawai.(g) Polypylis hemisphaerula from China, host of Fasciolopsis buski. (h) Parafossarulus manchouricusfrom China, host of Clonorchis sinensis. (i) Codiella (= Bithynia) leachi from Germany, host ofOpisthorchis felineus. (j) Pirenella conica (from Egypt) host of Heterophyes heterophyes. (k) Lymnaeatrunculatula from England, host of Fasciola hepatica.

opens and a more posterior ventralsucker, or acetabulum, by which theworm attaches itself to its host. InHeterophyes there is also an accessorygenital sucker.
The features of importance in therecognition and classification of a trema-tode are shown in Fig. 2 and the morphol-ogy of the flukes of medical importance in Fig. 3.
Life Cycle Stages
Adults are hermaphrodite, except for theschistosomes, which have separate sexes –the egg reaches water (in the schistosomes,opisthorchiids and heterophyids the eggcontains a larva, termed a miracidium,when passed out; in other trematodes thelarva develops inside the egg over a fewweeks) – the ciliated miracidium larvahatches from the egg and penetrates a spe-cific freshwater snail (except inopisthorchiids, where the egg containing alarva is ingested by the snail) – inside thesnail the miracidium develops into anirregular sac-like sporocyst – germ cellsinside this primary sporocyst form the nextlarval stage (these are termed rediae inmost trematodes, where they have a rudi-mentary gut, but secondary sporocysts inschistosomes, where they are similar to theprimary sporocysts), which burst out andinvade new tissues of the snail (principallythe digestive gland) – germ cells insidethese in turn develop into the next larvalstages, the tailed cercariae, which escapefrom the snail into the water (in someforms there are two redial generations).Thus one miracidium can give rise to manythousands of cercariae, the process takingseveral weeks or even months. The cer-cariae actively penetrate through the skin,as in the schistosomes, or form cysts(metacercariae) in a second intermediatehost or on vegetation and are passivelyingested in all other trematodes.
Classification
The classification given below is based prin-cipally on that of La Rue (1957), in whichthe life history and the larval stages are con-sidered as well as the morphology of theadult; more conservative classifications werebased entirely on the adult. The divisions atthe family level are generally accepted bymost authorities, but those taxa above thislevel are still controversial (Gibson and Bray,1994). Recent studies utilizing computer-based cladistic analysis and molecular biol-ogy might alter the familiar groupings in the
6 Chapter 1
Fig. 2. Diagram of Clonorchis sinensis to showthe features of taxonomic importance in thedigenetic trematodes.

future (Brooks et al., 1985; Rohde et al.,1993), but changes are not generallyaccepted. Only a very few of the numerousfamilies comprising the subclass Digenea areincluded (Yamaguti, 1971) – those whichhave members of medical importance.
The modes of infection of trematodesof medical importance shown in Table 2reflect quite well the taxonomic divisions(the odd one out being Echinostoma, inwhich it might be expected that the cercariae would encyst on vegetation).
The Trematodes 7
Fig. 3. Diagrams of the shape and principal organ systems of the hermaphrodite trematodes of medicalimportance (schistosomes are shown in Fig. 4). Comparative sizes shown in silhouette.

KEY: Members of the group occur in M = mammals, B = birds, R = reptiles, A = amphibians, F = fish.
PHYLUM PLATYHELMINTHES
CLASS TREMATODA
SUBCLASS DIGENEAEndoparasites with an indirect life cycleutilizing a snail as an intermediate host.Uterus has numerous coils containingmany eggs. Single excretory pore at post-erior end of the body.
SUPERORDER ANEPITHELOICYSTIDAThe cercaria has a thin-walled, non-epithelialbladder.
Order StrigeidaCercaria fork-tailed.
SUPERFAMILY SCHISTOSOMATOIDEA
Family Schistosomatidae (MB)
DioeciousSchistosoma
SUPERFAMILY DIPLOSTOMOIDEA
Family Diplostomidae (MB)Body usually divided into two regions.Metacercariae in fishes or amphibians.Alaria, Neodiplostomum
SUPERFAMILY GYMNOPHALLIOIDEA
Family Gymnophallidae (MB)Large oral sucker, small ventral sucker.Tegument with spines.Gymnophalloides
Order EchinostomomidaEggs operculate. Cercariae encyst onherbage or in other molluscs.
SUPERFAMILY ECHINOSTOMATOIDEA
Testes in tandem behind ovary. Family Echinostomatidae (MBRF)Head collar with row of spines.Echinostoma
8 Chapter 1
Table 2. Mode of infection of trematodes of medical importance.
Cercaria liberated from snail
Active penetration Encystment to giveof skin of definitive host metacercaria, which is
ingested by definitive host
Schistosoma
Encystment on vegetation 2nd intermediate host requiredˇ
FasciolopsisFasciola
Gastrodiscoides?
Fish Crustacean Snail Insect
Clonorchis Paragonimus Echinostoma DicrocoeliumOpisthorchis PlagiorchisHeterophyesMetagonimus

Family Fasciolidae (M)Suckers close to each other. Large, spiny,lanceolate flukes, usually found in herbivores.Fasciolopsis, Fasciola
SUPERFAMILY PARAMPHISTOMATOIDEA (MB)Sucker at anterior and posterior extremitiesof body but no oral sucker.Family Paramphistomatidae (or Zygo-cotylidae)Body thick and fleshy. Testes in tandem infront of ovary. Metacercariae on vegetationor on snail.Gastrodiscoides, Watsonius
SUPERORDER EPITHELIOCYSTIDACercaria has an additional thick-walledepithelial bladder.
Order PlagiorchiidaEggs operculate.
SUPERFAMILY DICROCOELIOIDEA
Cercariae encyst in arthropods and have anoral stylet.Family Dicrocoeliidae (MBRA)Found in intestine, liver, gall-bladder andpancreas. The oral sucker is subterminal.Testes adjacent or in tandem, anterior toovary. Vitellaria posterior to ventralsucker.Dicrocoelium
SUPERFAMILY OPISTHORCHIOIDEA (MBR)Cercariae encyst in or on fish.Family Opisthorchiidae (MB)Suckers weak. Semi-transparent flukesfound in bile-ducts and gall-bladder. Testesin tandem behind ovary.Clonorchis, Opisthorchis, Metorchis
Family Heterophyidae (MB)This is a large family. All members arepotential parasites of humans. They areminute flukes with a spinose tegument.Testes adjacent behind ovary. Heterophyes, Metagonimus
SUPERFAMILY PLAGIORCHIOIDEA
Family Lecinthodendriidae (MBRA)Small spiny flukes with gonads in fore-body. Oral sucker large, ventral suckersmall. Metacercariae in aquatic insects.Phaneropsolus, Prosthodendrium
Family Paragonimidae (MB)Many are parasites of the lungs. Vitellariacompact and dense. Cuticle spinous. Testesadjacent behind ovary. Cercariae encyst incrustaceans.Paragonimus
Family Plagiorchiidae (MBRAF)Cuticle spinous. Suckers well apart. Testesin tandem behind ovary. Metacercariae ininsects.Plagiorchis
Family Troglotrematidae (M)Cercariae encyst in fish or crustaceans.Genital pore posterior to ventral sucker.Spinous body.Nanophyetus
Family Schistosomatidae
Schistosomes
At least seven species are parasites ofhumans: Schistosoma haematobium(Bilharz, 1852); Weinland, 1858; S. mansoniSambon, 1907; S. japonicum Katsurada,1904; S. intercalatum Fischer, 1934; S.malayensis Greer, Ow-Yang and Yong,1988; S. mekongi Voge, Bruckner andBruce, 1978; S. sinensium Pao, 1959.
SYNONYMS (for S. haematobium)Distoma haematobia Bilharz, 1852;Bilharzia haematobium Diesing, 1859.
LOCAL NAMES
Au chung (Chinese), Tsagiya (Hausa, S.haematobium), Katayamabayo, Suisho-choman or Harapari (Japanese), Laremo(Luo, S. haematobium haematuria), Kadidhig (Somali, S. haematobium haema-turia), Kichocho (Swahili), Pa-yard bai-mai lohit (Thai), Atosi eleje (Yoruba, S.haematobium).
The Trematodes 9

DISEASE AND POPULAR NAMESSchistosomiasis or schistosomosis*, uri-nary schistosomiasis (S. haematobium),intestinal schistosomiasis (S. mansoni, S.japonicum, S. intercalatum and S.mekongi), schistosomiasis haematobium,intercalatum, japonicum, mansoni ormekongi; Katayama disease (early phase ofS. japonicum); bilharziasis; bilharzia.
GEOGRAPHICAL DISTRIBUTION
It has been estimated that about 220 million people are infected in the worldin 74 countries, with 600 million at risk;of those infected 20 million have severedisease, 120 million have mild symptomsand 80 million are symptomless (WHO,1993).
S. haematobium. Africa: most of the coun-tries of North Africa; widespread in Centraland West Africa; in eastern Africa presentfrom Somalia to the Cape and on theislands offshore, including Madagascar andMauritius. Middle East: present in mostcountries. There might also be small foci inIndia around Bombay and in Madras State.A total of about 90 million people areinfected worldwide.
S. mansoni. Africa: North Africa (Morocco,Tunisia, Egypt, southern Sudan); EastAfrica (from Ethiopia down to South Africaand Madagascar); most countries of Centraland West Africa; Middle East (Lebanon,Oman, Saudi Arabia, Somalia, Yemen);Americas: in South America and some ofthe Caribbean islands.
S. intercalatum. There are limited foci inCentral Africa including Cameroon, Congo,Congo Democratic Republic (Zaire),Equatorial Guinea, Gabon, São Tomé andPrincipe, and possibly in Central AfricanRepublic, Chad, Mali and Nigeria.
S. japonicum. China, Indonesia,Philippines, Thailand (an S. japonicum-like parasite which is probably distinct).
S. malayensis. Malaysia.
S. mekongi. Cambodia, Laos.
S. sinensium. China, Thailand.
MORPHOLOGY
Unlike all the other trematodes of medicalimportance, the sexes are separate (dioe-cious). In all species the male worm ischaracteristically boat-shaped, with a cen-tral canal (gynaecophoric canal) in whichthe female lives. The cuticle of the male issmooth in S. japonicum (the adults of S.malayensis and S. mekongi are identical)but has tuberculations in the other threeimportant species. There are two smallsuckers and a varying number of testes inthe different species. The female is longerthan the male but much thinner and circu-lar in cross-section. The two suckers of thefemale are very small and weak. The char-acteristics of the various species are givenin Table 3 and Fig. 4.
S. haematobium. The male measures10–20 mm � 0.9 mm and the cuticle hasfine tuberculations. There are 4–5 testes.The female has a long uterus, with theovary in the posterior third of the body.There are 10–100 eggs in the uterus at onetime.
S. mansoni (Fig. 5). The adults are smallerthan those of the other species. The malemeasures 6–13 mm � 0.75–1.0 mm and thecuticle has coarse tuberculations. There are4–13 (usually 6–9) testes. The ovary of thefemale is situated anteriorly. There is usu-ally only 1 egg in the uterus at one time.
10 Chapter 1
*This is the standardized nomenclature for parasitic diseases advocated by the World Association forthe Advancement of Veterinary Parasitology (WAAVP) (Kassai et al., 1988. Veterinary Parasitology 29,299–326) and the World Federation of Parasitologists (WFP), but has not been widely adopted inmedical helminthology, particularly by the World Health Organization (WHO). For instance, of the titlesof references in this book, 112 have -iasis endings and 16 -osis, most of which are mentioning cysticercosis or echinococcosis – used in both systems.

S. japonicum (S. malayensis and S.mekongi). The male measures 12–20 mm �0.5–0.55 mm and has no cuticular tubercu-lations. There are 6–7 testes. The femalehas the ovary at about the middle of thebody. There are 5–200 eggs in the uterus atone time.
S. intercalatum. The male measures11–14 mm � 0.3–0.4 mm. There are 2–7testes. The female has 5–60 eggs in theuterus at one time.
Of the complete 270 Mb genome ofSchistosoma, 18–24% has so far beendeposited in a database (Williams andJohnston, 1999).
LIFE CYCLE
The eggs are passed in urine in S. haemato-bium and in the faeces in all the otherspecies and contain a fully formed miracid-
ium. Eggs of each species can be recognizedby differences in size and morphology(Table 3 and Fig. 6). On immersion in freshwater, particularly under conditions ofwarmth and light, they hatch almost imme-diately. The miracidium larvae (Fig. 7)swim actively by means of the cilia withwhich they are covered and attempt to pen-etrate any freshwater snail they come intocontact with. The miracidia die in 16–32 hif they do not succeed in reaching a suitablesnail intermediate host. Like all trematodesthey are extremely host-specific in regard tothe snails in which they will develop, oftenfar more so than in the definitive host. Thespecies of snail parasitized depends on thegeographical region, but S. haematobiumand S. intercalatum develop in snails of thegenus Bulinus, S. mansoni in Biomphalariaand S. japonicum in Oncomelania.Oncomelania differs from the other two
The Trematodes 11
Map 1. Distribution of Schistosoma haematobium, S. japonicum and S. mekongi.

genera of snails in that it is amphibious,rather than aquatic, and dioecious (withseparate sexes) and has an operculum onthe bottom surface of the foot (to preventdrying up). Once in a susceptible snail, themiracidium loses its outer ciliated epider-mal layer and develops into a mothersporocyst. This becomes filled with germballs, which burst out after about 8 days,and most of these migrate to the digestivegland, where each develops into a thin-walled daughter sporocyst. A furtherprocess of asexual multiplication takesplace and each daughter sporocyst becomesfilled with the final larval stages, the cercariae. Thus one miracidium can giverise to thousands of cercariae, all of thesame sex. At about 26°C, the cercariae of S.intercalatum begin to emerge 3 weeks afterinfection, those of S. mansoni after 4–5weeks, those of S. haematobium after 5–6
weeks and those of S. japonicum after 7weeks. The principal stimulus for emer-gence is light and different species emergeat various times in the day. The cercariaemeasure 300–400 µm � 50–70 µm andhave forked tails (furcocercus type) (Fig. 8).They swim around in the water, usually tailfirst, and often hang from the surface film.They are infective for only a day or so inwater. Not all the cercariae mature at thesame time and usually a proportion continue to emerge throughout the life ofthe snail, which may be many months.When humans enter the water the cercariaepenetrate the skin, often between the hairfollicles, by means of the anterior spinesand the cytolytic secretions of the cephalicglands. The tail is shed in the penetrationprocess, which takes 3–5 min, and theimmature schistosomes (known as schisto-somula) enter peripheral lymphatics or
12 Chapter 1
Map 2. Distribution of Schistosoma mansoni and S. intercalatum.

The Trem
atodes13
Table 3. Differential features of schistosomes of humans.
Schistosoma Schistosoma Schistosoma Schistosoma Schistosomajaponicum mansoni haematobium intercalatum mekongi
Situation in human Mesenteric veins Mesenteric veins Vesical veins Mesenteric veins Mesenteric veins
MaleLength (mm) 10–20 6–12 10–14 11–14 15Width (mm) 0.5 1.1 0.9 0.3–0.4 0.4No. of testes 6–7 4–13 (usually 6–9) 4–5 2–7 (usually 4–5) 6–9Tuberculations None Coarse Fine Fine NoneCaecal junction Posterior third of body Anterior third of body Middle Middle Posterior third of body
FemaleLength (mm) 20–30 10–20 16–20 10–14 12Width (mm) 0.3 0.16 0.25 0.15–0.18 0.23Uterus Anterior half of body Anterior half of body Anterior two-thirds of body Anterior two-thirds of body Anterior half of bodyNumber of eggs in
uterus 50–200 1–2 10–50 5–60 10+Position of ovary Middle Anterior third of body Posterior third of body Posterior half of body Posterior half of bodyMature egg shape Round with small knob Lateral spined Terminal spined Terminal spined Round, small knob
and mean size 85 µm � 60 µm 140 µm � 61 µm 150 µm � 62 µm 176 µm � 61 µm 57 µm � 66 µmMode of voiding eggs Faeces Faeces Urine Faeces FaecesEgg production per 3500 100–300 20–300 150–400 ?
female per day (inexperimental animals)
Reaction of egg to Positive Positive Negative Positive Positive?Ziehl–Neelsen stain
Intermediate Oncomelania Biomphalaria Bulinus Bulinus Triculahost snail
Distribution overlap None Over most of Africa Throughout range None

venous vessels and are carried to the lungs4–7 days after penetration (Figs 9 and 10).The schistosomula move from the lungs tothe portal vessels and there grow into adultschistosomes, which mate and remain in
pairs. The schistosomula are usuallyassumed to travel to the liver via the bloodsystem (against the blood flow) but, at leastfor those of S. japonicum, some penetratedirectly through the diaphragm. The adult
14 Chapter 1
Fig. 4. Diagram of the structures of the three major schistosome species. Only the male of S. mansoni is shown. The males of the other species differ principally in the number of testes, while that of S. japonicum has a smooth tegument.
Fig. 5. Integument and double outer membrane of S. mansoni. Electron micrograph. Original magnification � 83,500.

The Trem
atodes15
Fig. 6. Eggs of (a) S. haematobium, (b) S. mansoni and (c) S. japonicum. Actual size of (a) is 125 µm.

worm pairs migrate to the mesenteric veins(S. intercalatum, S. japonicum, S. mansoniand S. mekongi) or the veins of the vesicalplexus surrounding the bladder, migratingthrough the anastomoses between the portal and systemic veins in the pelvis (S.haematobium).
Eggs of S. japonicum and S. mansoni firstappear in the stools 25–28 days after cercar-ial penetration, those of S. intercalatumafter 50–60 days and those of S. haemato-bium in the urine after 54–84 days. Adultworms of each species can live for as long as25 years in people who have moved from anon-endemic area, but it is probable thatlongevity is usually considerably less inendemic areas (3–10 years) and in childrenis about 2–5 years. Large numbers of wormsmay be present, up to 400 having beenfound at autopsy, but in most cases there arefewer than ten worm pairs present. Eachfemale of S. japonicum produces about 3500eggs per 24 h, of S. mansoni 100–300, of S.haematobium 20–30 and of S. intercalatum150–400. The number of eggs produced byS. haematobium decreases with age (Agnewet al., 1996).
It has been reported that S. japonicumcan undergo vertical transmission (cf.Alaria; p. 59), but this appears to be coin-cidental (Shoop, 1994).
16 Chapter 1
Fig. 7. Ciliated miracidium of S. mansoni. Note anterior penetration glands. Dark field. Actual size 80 �m.
Fig. 8. Diagram of the cercaria of S. japonicum.

CLINICAL MANIFESTATIONS AND PATHOGENESIS
The clinical manifestations and pathogene-sis of S. haematobium infection differ fromthose caused by the other species and willtherefore be considered separately. Briefly,this is because with S. haematobium theeggs accumulate progressively in the blad-der and ureters and the reaction to themleads to cystitis, hydronephrosis, uretericobstruction and occasionally cancer of the
bladder, while with S. japonicum, S. man-soni, S. mekongi and, to a lesser extent, S.intercalatum, the granulomas surroundingthe eggs cause colitis, and eggs reachingthe liver cause presinusoidal block to thehepatic portal flow, leading to portal hyper-tension. However, with all species, thepathology depends on the target organs,intensity and duration of infection, hosthuman leucocyte antigen (HLA) type and
The Trematodes 17
HUMANADULTS maturein 6–12 weeks.In veins of bowel:S.m., S.j., S.i.In veins of bladder:S.h.
liver
lungs
lymph and veinsto heart
SCHISTOSOMULAin skin
Male and femaleCERCARIAEpenetrate skin
SNAILCERCARIAE emerge
SECONDARYSPOROCYSTS
in digestivegland
PRIMARYSPOROCYSTS
MIRACIDIApenetrate snail
Asexualreproductivestages insnail 4–7weeks
CERCARIAE (0.4 mm)infective for 8hours in water.Penetrate skin
MIRACIDIA (0.1 mm)hatch immediatelyin water and canlive for 24 h.Penetrate suitablesnail
EGGSIn urine: S. haematobiumIn faeces: all others
Some EGGS retainedin tissues
S.m.
S.j.
S.h.
and
S.i.
Biomphalaria spp.
Oncomelania spp.
Bulinus spp.
Fig. 9. The life cycle of schistosomes. Figure from Book of the Dead papyrus. S.m., S. mansoni; S.j., S. japonicum; S.h., S. haematobium; S.i., S. intercalatum.

race, host immunological responses andconcomitant infections such as hepatitis.
S. haematobium
Invasive stage. A cercarial dermatitis(‘swimmer’s itch’) may appear 24 h afterfirst infection but seldom lasts more than48 h. It is rarely met with in peopleindigenous to an endemic region and ismore common after penetration by cer-cariae of non-human species of schisto-somes.
Acute phase. There are usually no symp-toms until 5–10 weeks after infection,when there may be mild allergic manifesta-tions in visitors but these are rare inindigenous populations.
Chronic phase. Maximum egg productionbegins 10–12 weeks after infection. Inschistosomiasis it is the egg that is theimportant pathogenic agent. The majorityof eggs pass through the bladder but anunknown proportion are trapped in thebladder wall and ureters and eventuallydie and calcify. The earliest bladder lesionis the pseudotubercle, but in long-standinginfections nests of calcified ova (‘sandypatches’) (Fig. 11) are surrounded byfibrous tissue in the submucosa and makethe bladder wall visible on X-ray or byultrasonography (Hatz, 2000).
Haematuria (found in about 50% ofcases), dysuria and increased frequency of
micturition are typical clinical signs andmay persist intermittently for years. Cystitisis caused by hyperplasia of the epithelium(which occasionally becomes 2–3 cm thick),sometimes with papilloma formation, and amarkedly reduced bladder capacity oftenresults. This, together with the ureteritis,can lead to hydroureter, hydronephrosis(Fig. 12) and uraemia. Hydronephrosis isnot necessarily only a late sequel of infec-tion, as it has been observed in over 8% ofpreadolescent children in both East andWest Africa. It may be reversible after anti-schistosomal treatment (Subramanian et al.,1999) and is probably caused by oedema,congestion, inflammation and possibly pro-liferative lesions (although granulomas ofthe kidneys are very rare).
Fibrosis of the ureteral wall usually leadsto dilatation, while strictures occur in lessthan 1% of patients. Cancer of the bladder isparticularly common in Egypt andMozambique and is clearly predisposed toby urinary schistosomiasis (IARC WorkingGroup, 1994) but some other precipitating
18 Chapter 1
Fig. 10. Diagram of a 12-day-old schistosomulumof S. mansoni recovered from the lungs.
Fig. 11. Inner surface of the bladder showingnests of calcified ova (‘sandy patches’) (arrowed)of S. haematobium.

factors, such as the presence of nitrosaminesin the urine, are probably involved (Mostafaet al., 1995). Squamous carcinoma is morecommon than transitional carcinoma (Fig.13). It has been estimated that schistosomia-sis is responsible for about 16% of cases ofbladder cancer in Egypt. In the Nile Deltaregion, men do most of the agriculturalwork and thus become infected, resulting ina 12:1 male-to-female bladder cancer ratio.Pulmonary arteritis progressing to irrevers-ible and lethal cor pulmonale because ofcapillary damage by eggs sometimes occurswhen eggs are swept back into the lungs.The presence of adult worms in the lungsfollowing drug treatment is also a possiblecause of pulmonary damage. In women,eggs may cause lesions in the ovaries,Fallopian tubes and uterus or in the lower
parts of the genital tract, including thecervix, vagina and vulva, and about 6–27%of such cases result in sterility.
S. japonicum, S. mekongi, S. mansoni and S. intercalatum
Invasive phase. As for S. haematobium.
Acute phase. Allergic manifestations, suchas pyrexia, headache, oedema, cough,dysenteric symptoms, pruritus andurticaria, occur 3–8 weeks after infectionwith S. japonicum. This is known as theKatayama syndrome and may be accompa-nied by tenderness in the liver region, mildabdominal pain, lymphadenopathy andsplenomegaly, with an accompanying
The Trematodes 19
Fig. 12. Intravenous pyelogram of a woman infected with S. haematobium showing bilateral hydronephrosiswith deformity of both ureters.

eosinophilia. This is possibly caused by aform of ‘serum sickness’ (acute immunecomplex disease) resulting from an excessof antigen when eggs are first produced.The acute phase of S. mansoni and S. inter-calatum infection is rarely recognized inan indigenous population.
Chronic phase. In the majority of individu-als, infection is light and symptoms areentirely absent, but in heavy infectionsabout 50% of the eggs are trapped in themucosa and submucosa of the colon, result-ing in the formation of pseudotubercles,which coalesce and form larger granuloma-tous reactions and pseudopapillomas. Eggsof S. japonicum, in particular, are inclinedto die and eventually to calcify in the colonand many thousands of calcified eggs maybe found in the thickened mucosa (Plate 1).Intestinal damage, however, is usuallyaccompanied by few symptoms except avague feeling of ill health, with perhapsheadache, abdominal pain and diarrhoea.Papillomas and inflammatory polyps oftendevelop, and in severe cases, can lead toobstruction of the lumen of the colon. Theulceration of the colon caused by the eggsof S. mansoni can result in a blood loss ofup to 12.5 ml day�1 by patients in Egypt,where papillomas are particularly common.Carcinomas of the large intestine are also
associated with chronic lesions of S. japon-icum in a small proportion of cases (IARCWorking Group, 1994). The increasing fibro-sis of the colon wall means that eggs arerepeatedly carried to the liver in the portalveins and become lodged in the portaltracts or, less frequently, in the lobules orsinusoids. The eggs become the centre of apseudotubercle, in which they are engulfedby multinucleate giant cells and sur-rounded by inflammatory cells (eosinophils,macrophages and polymorphonuclearleucocytes). In the early stages there is amicroabscess surrounding each egg, resem-bling miliary tuberculosis (Fig. 14).However, the cell types differ from a typicaltubercle. As the lesion heals, some degreeof fibrosis is usually left. Finally themiracidium in the egg dies and themononuclear cells form Langhans-type giantcells. Experiments in mice have demon-strated that CD4+ T lymphocytes are neces-sary for the formation of granulomas. Whilethe formation of granulomas is usuallytaken to be a host response to sequester theeggs, it is possible that mobile granulomascan aid in ‘transporting’ the eggs across tis-sues to reach the faeces or urine (Doenhoffet al., 1986; Damian, 1987). Pigment, chemi-cally but not structurally indistinguishablefrom malarial pigment, becomes depositedin the Kupffer cells, portal tracts and
20 Chapter 1
Fig. 13. Section of bladder. Eggs of S. haematobium (many are calcified or ‘black’ eggs) can be seenwith epithelial squamous cell metaplasia (arrowed).

lobules. Liver enlargement is common andsplenomegaly often follows the portal hyper-tension (Fig. 15). Hepatosplenic schisto-somiasis mansoni is more common inBrazil and Egypt than in Africa south of the
Sahara and signs of portal hypertension arealways present in such cases. Anaemia maybe found when there is splenomegaly and ismore severe after repeated haematemesis.The reaction to the eggs in the liver may
The Trematodes 21
Fig. 14. Egg granuloma of S. mansoni in liver with surrounding epitheloid cells and some leucocytic infiltration (see Plate 1 for more advanced stage).
Fig. 15. Two boys with advanced schistosomiasis mansoni. Note collateral venous circulation in nearestpatient.

eventually cause the periportal fibroticreaction termed ‘Symmer’s clay pipestem’fibrosis (Fig. 16). Liver function tests, how-ever, are not altered in schistosome fibrosis,as they are in true cirrhosis, and, althoughthere may be severe pathological lesions,there is no liver failure. Severe disease,with hepatosplenomegaly, occurs in about10% of cases of schistosomiasis mansoni,but takes 5–15 years to develop, and inchildren infection can have effects on nutri-tion (Stephenson, 1993).
As the portal pressure increases, a collateral venous circulation becomesestablished and severe or even fatalepisodes of bleeding can result from theoesophageal varices. The portal systemicshunt results in the eggs bypassing theliver and being deposited in the lungs. Anobstructive and destructive arteritis mayfollow, which can lead to systematic arter-ial hypertension and eventually to hyper-trophy of the right ventricle.
The adult worms in the blood-vessels dolittle damage when living but their deathcan lead to focal necrosis of the liver cellsand to granulomas. The more severepathology thought to be produced by infec-tion with S. japonicum is usuallyexplained by the greater egg production,the spherical shape of the egg lacking alarge spine resulting in more eggs being
distributed throughout the body, and thegreater longevity of the adult worms.
Brain involvement is most common in S.japonicum infections and two types of brainlesions have been reported. In the first typethere is diffuse involvement with scatteredlesions, probably caused by eggs being car-ried to the brain in the bloodstream, andthis type usually results in no symptoms. Inthe second type a localized granulomatousmass is present, containing large numbers ofeggs deposited by ectopic adult worms inthe blood-vessels of the brain. These granu-lomas can cause a wide range of symptoms,depending on the anatomical location of thelesion in the brain.
A transverse myelitis can result from thepresence of eggs in the spinal cord and ismost commonly found in infections with S.mansoni and S. haematobium.
In order for the eggs to make their waythrough the tissues into the gut, themiracidia release proteolytic enzymes andother material, which diffuses throughpores in the eggshell. This material (solu-ble egg antigen (SEA)) is highly immuno-genic, and the immune response madeagainst it leads to the formation of the largegranulomas that are responsible for most ofthe pathology in this phase. Evidence forthis and analysis of the mechanisms involvedhave come largely from experiments in
22 Chapter 1
Fig. 16. ‘Symmer’s clay pipestem’ fibrosis caused by eggs of S. japonicum surrounding portal veins in liver.

mice. Granulomas can be induced in thelungs of mice by intravenous injection ofeggs, and this process is prevented byimmunosuppressive treatments that inter-fere with cellular responses. The degree ofresponse to eggs and the pathology thatresults, following infection in mice, arestrongly influenced by genetic factors, andthis also reflects different degrees ofimmune responsiveness. It has recentlybeen shown in humans that at least someof the variation in pathology seen ininfected populations is due to the activityof particular major genes, which are associ-ated with T-cell function (e.g. a codomi-nant major gene, SM1, on chromosome 5).The immunology of granuloma formationis complex and involves the activity ofboth major subsets of CD4+ T-helper (Th)cells. SEA is a powerful inducer of Th2responses, but Th1 cells also play a role.The cellular response is initiated and con-trolled by the cytokines released and inmice the phenomenon of immunomodula-tion occurs – i.e. early granulomas tend tobe much larger than those formed later ininfection.
Antigen–antibody complexes have beenshown by fluorescent antibody studies tobe the cause of the Splendore–Hoeppliphenomenon that occurs in sensitizedhosts. Glomerulonephritis has beenreported as an immune-complex disease inschistosomiasis mansoni.
IMMUNOLOGY OF SCHISTOSOMES
Although it has been established for manyyears that laboratory animals, including pri-mates, develop immunologically mediatedresistance to experimental infections, defi-nite proof that acquired immunity to schisto-some infections occurs in humans has beenvery difficult to demonstrate. It is clear thatintensity of infection and prevalence with S.haematobium and S. mansoni is greatest in10–14-year-olds and declines in older age-groups, but this could be explained by adecrease in water contact. However, longitu-dinal studies and detailed monitoring ofreinfection after elimination of an existinginfection by chemotherapy have demon-strated that age-dependent resistance does
occur against these species (Wilkins et al.,1984, 1987; Butterworth, 1998; Kabatereineet al., 1999), and studies in Brazil link resis-tance to particular genetic characteristics(Abel and Dessein, 1997). Infections with S.japonicum do not show this picture,although there appears to be an age-relatedreduction in pathology (Ross et al., 2000).The slow acquisition of immunity against S.haematobium and S. mansoni correlateswith a change in the balance betweenantiparasite immunoglobulin E (IgE) andIgG4, the former promoting protectiveresponses against incoming larvae, perhapsby antibody-dependent cellular cytotoxic(ADCC) mechanisms directed against anti-gens expressed on the surface tegument. Asshown by work carried out in the 1960s,adult schistosomes are largely unaffected byimmunity and continue to survive in hostsimmune to larval stages (concomitant immu-nity). One way in which this is achieved isby the adult schistosomes masking the for-eign nature of their surface antigens byincorporating host-derived molecules intothe tegument and by rapid replacement ofthe tegument (antigenic disguise).Schistosomes, like many other helminths,exert profound effects on the host’s immuneresponse. Not only do these parasites tend topolarize the T-cell response selectivelytowards Th2 activity, but they may alsomodulate other components in ways thatresult in a more general suppression ofimmune and inflammatory responses.
Immunomodulation can influence theability of the host to respond successfullyto other infections, and there is recent evi-dence that schistosomes and soil-transmit-ted nematodes may increase susceptibilityto both AIDS and tuberculosis (Bundy etal., 2000).
DIAGNOSIS
Parasitological. The presence of eggs in thefaeces or urine is still the most widely usedmethod of diagnosis. This usually poses noproblems in heavy infections but diagnosismay not be so easy in light infections, asoccur in the majority of cases, particularlyin tourists, who have probably been infected
The Trematodes 23

only once. In general, it is only the presenceof eggs containing live miracidia thatindicates an active infection requiring treat-ment.
The eggs of S. haematobium, which occurin the urine, can be detected by sedimenta-tion of a 10 ml sample (best collected aroundmidday) in a urinalysis flask, the depositbeing examined under the microscope foreggs. Alternatively, a 10 ml sample can bepassed through a polycarbonate or poly-amide membrane, washed, removed andstained in 1% trypan blue (or 2.5% ninhy-drin, 5% iodine, eosin or methylene blue)for a few minutes. After washing again (anddrying at 37°C if wished), the filter is exam-ined in a few drops of saline on a micro-scopic slide. It is probably not wise to reusefilters. In about 5% of patients, eggs may alsobe found in the faeces.
Cytoscopy may be performed as a finaldiagnostic measure when no eggs can befound in the urine, but the danger ofsecondary infection makes this a possiblyhazardous procedure.
The eggs of the other species occur inthe faeces and concentration techniques,such as the formol–ether or Kato–Katzthick-smear methods (p. 258), are necessary.A modification of the thick-smear methodcan be used for quantitative determinationsand, in some studies, has given a goodindication of the number of adult worms,as can filtration staining techniques. In thelatter, eggs from faeces are stained withninhydrin on filter-paper and counted.
Rectal biopsy is often effective (even inabout 50% of cases of S. haematobium), anunstained squash being examined underthe microscope.
Clinical. In the field, haematuria can providea quick and inexpensive indicator of urinaryschistosomiasis; microhaematuria (measuredby means of a reagent strip) or proteinuriacorrelates well with the intensity of infec-tion. Even self-reporting by children canprovide a useful epidemiological record ofcommunity prevalence, although this islikely to give an overestimate, depending onthe species of worm and level of infection(Ansell and Guyatt, 1999); using models for
each species more accurate estimates arepossible (Guyatt et al., 1999).
Portable ultrasound can be used fordiagnosis of the degree of pathology, partic-ularly in the liver or bladder, and canscreen populations at the community level;it can also be used for determining theeffect of chemotherapy (Hatz, 2000).
Immunological. There are numerousimmunological tests for diagnosis but thoseinvolving various ELISA and immunoblot-ting techniques are the most convenient. Adipstick ELISA for urine samples, using anSEA can effectively diagnose schistosomia-sis and correlates well with quantitativeegg counts. Circulating cathodic antigen(CCA) is the dominant antigen in the urineof S. mansoni patients and can be detectedby a monoclonal antibody (mAb) of 41/42kDa. There are also circulating solubleadult-derived antigens (Sm31/32 andSj31/32) in serum, which can be detectedby dot ELISA, and an IgG ELISA can beused to detect antibodies against them.
TREATMENT
Chemotherapy. Praziquantel (usually in asingle oral dose of 40 mg kg�1) is the drug ofchoice for all species of schistosome and hasvirtually superseded almost all the agentspreviously used. It has few side-effects(except sometimes in patients with veryheavy worm loads), although it is probablybetter not to give it during early pregnancy.Praziquantel is a heterocyclic pyrazine-isoquinoline unrelated to otheranthelminthics – chemical formula 2-(cyclo-hexylcarbonyl)-1,2,3,6,7,11-b-hexahydro-4H-pyrazino[2,1a]isoquinolin-4-one. It is quicklymetabolized, crosses the blood– brain barrierand appears to act by causing spastic paraly-sis and vacuolization of the tegument of theworm, possibly from interference with inor-ganic ion transport and an increase in mem-brane permeability; it may also destroylysosomes. Two target antigens are alsoaffected (a surface tubercle glycoprotein of200 kDa and an esterase of 27 kDa) andmaybe epitopes become exposed so that thedrug evokes an effective immune response(Brindley, 1994; Dupre et al., 1999).
24 Chapter 1

Metrifonate is cheap and safe to useagainst S. haematobium infection. Its maindisadvantage is that repeated treatment isnecessary (usually 7.5–10 mg kg�1 givenorally in each of three doses at fortnightlyintervals). As an organophosphate com-pound, it inactivates the enzyme, destroy-ing acetylcholinesterase, but its mode ofaction against the worms is obscure.Atropine sulphate can be used if symptomsof cholinergic activity occur.
Oxamniquine is a tetrahydroquinolinederivative that has activity only against S.mansoni (given at a single oral dose of 15mg kg�1 ) and can be used for advancedcases of the disease. It should be avoidedin early pregnancy and in cases of epilepsy,and can cause drowsiness. Resistance tothe drug has also been reported, and it is inlimited supply. The mode of action isunknown.
The antimalarial compound artemetheror artesunate (oil and water soluble formsof an artemisinin derivative) is being testedas a prophylactic against S. japonicum inChina and in one trial was shown to reduceegg production of S. mansoni and S.haematobium by 15% after 12 weeks whengiven in the usual dose for malaria. It alsoacts principally against developing worms,unlike the two compounds at present inuse; cyclosporin has similar modes ofaction but would be too expensive for gen-eral use. Caution needs to be exercised inrelation to malaria resistance.
Surgery. In advanced cases with liver fibro-sis, portocaval shunt and occasionallysplenectomy have been used to relieve theportal hypertension, but the results havebeen disappointing.
EPIDEMIOLOGY
Schistosomiasis, particularly when due toS. haematobium, is very much a focal dis-ease and many prevalence figures are basedon limited surveys, which are not alwaystypical of the country as a whole. However,estimates in millions of overall prevalencein each endemic country have been made
recently (Chitsulo et al., 2000).
S. japonicum. China 1.06 (0.09% of totalpopulation), Philippines 0.43 (0.6%),Indonesia (Sulawesi) 0.0002 (0.0001%),Thailand very rare (‘S. japonicum-like’).
S. mansoni. Antigua fewer than 0.0001*,Brazil 7.01 (4%), with a mortality rate of0.44 per 100,000, Burundi 0.84 (13%),Dominican Republic 0.23 (3%), Eritrea 0.26(7%), Guadeloupe 0.033 (8%)*, Martinique0.005 (1.3%)*, Puerto Rico 0.015 (0.4%)*,St Lucia 0.0019 (1.2%)*, Rwanda 0.38(6%), Surinam 0.0037 (0.9%) and rising inchildren, Venezuela 0.0338 (0.2%).
S. haematobium. Algeria 2.1 (7.5%), India(Bombay area) very rare, Iran 0.042(0.07%), Iraq 0.042 (0.2%), Jordan 0.0001(0.002%), Lebanon very rare*, Libya 0.27(5%), Mauritius 0.016 (1.5%)*, Syria 0.003(0.02%)*, Turkey 0.0006 (0.001%)*.
S. mansoni and S. haematobium. Angola 4.8(44%), Benin 1.95 (35%), Botswana 0.15(10%), Burkina Faso 6.24 (60%), Cameroon3.02 (23%), Central African Republic 0.33(10%), Chad 2.78 (43%), Congo 0.89 (34%),Congo Republic (Zaire) 13.84 (28%), Côted’Ivoire 5.6 (40%), Egypt 10.06 (17%),Equatorial Guinea 0.008 (2%), Ethiopia 4.0(7%), Gabon 0.5 (45%), Gambia 0.33 (30%),Ghana 12.4 (73%), Guinea 1.7 (26%),Guinea Bissau 0.33 (30%), Kenya 6.14(23%), Liberia 0.648 (24%), Madagascar7.54 (55%), Malawi 4.2 (43%), Mali 5.88(60%), Mauritania 0.63 (27%), Mozambique11.3 (70%), Namibia 0.009 (0.6%), Niger2.4 (27%), Nigeria 25.83 (23%), Senegal 1.3(15%), Sierra Leone 2.5 (60%), South Africa4.5 (11%), Swaziland 0.23 (26%), Togo 1.03(25%), Uganda 6.14 (32%), Tanzania 15.24(51%), Yemen 2.23, Zambia 2.39 (27%),Zimbabwe 4.4 (40%).
S. intercalatum. Cameroon locally in forestareas, Congo (16% in children in one area),Congo Republic (Zaire) sporadic,Equatorial Guinea (13% in Bata city),
The Trematodes 25
*It is probable that in these countries transmission has now ceased and these are residual cases.

Gabon (29% in one area), São Tomé andPrincipe 0.005 (4%). Confirmation neededfor presence in Central African Republic,Chad, Mali and Nigeria.
S. mekongi. Cambodia 0.07 (0.7%), Laos0.12 (2%) and recent cases in refugeecamps in Thailand.
S. malayensis. Malaysia very rare (Shekar,1991).
In most areas endemic for S. haematobiumthere is a very high prevalence rate, withalmost everyone in the community lightlyinfected before the age of 10 years. Studiesin Brazil, St Lucia, Tanzania and Ugandademonstrated a relationship between eggoutput and prevalence of S. mansoni inchildren (Fig. 17). Above 50–60% preva-lence, a small extra rise is accompanied bya considerable increase in egg output andthus in the severity of disease. Post-mortemstudies and blood-filtration studies afterchemotherapy for counting adults haveshown a good relationship between the lasttwo parameters so that intensity of infec-tion can be expressed in terms of eggs pergram of faeces or per 10 ml of urine. Todetermine the intensity of infection of dif-ferent age-groups, the frequency of occur-rence of the different levels of egg outputcan be conveniently plotted and provides agood indicator of the proportion of the com-
munity that has levels of egg output likelyto cause morbidity (Jordan et al., 1993).This demonstrates the importance of quan-titative determinations of egg output inschistosomiasis (p. 260). The ecological fac-tors associated with the transmission ofschistosomiasis vary markedly with thespecies of schistosome, owing to the differ-ing habitats of the snails involved. Theamphibious snail hosts of S. japonicum livemainly in rice paddies and muddy habitatsbeside rivers; the aquatic snail hosts of S.mansoni live principally in gently flowingrunning water (less than 0.3 m s�1 ) such asirrigation channels, streams, lakes andponds (Plate 2); those transmitting S.haematobium live almost entirely in stillwater, such as ponds, pools, lakes andmarshy areas. Snail populations usuallyshow marked seasonal fluctuations, themost important factors influencing numbersbeing rainfall and temperature. The effect of rainfall varies, depending on the yearlyrainfall figures. In drier regions of Africarainfall stimulates the production of youngbulinid and planorbid snails, while inequatorial regions with a higher rainfall,snails are washed away during the rainyseason and breeding takes place principallyin the months following. Temperature isanother factor limiting population densi-ties, the temperature range for optimalexpansion of snail numbers being 20–28°C.
26 Chapter 1
200 400 600 800
100
75
50
25
Geometric mean of egg output (e.p.g. faeces)
Pre
vale
nce
(%)
EGYPT BRAZIL
KENYA
ZAMBIAST LUCIA
KENYAETHIOPIA
BURUNDIBURUNDI
ETHIOPIA
ZAIRE
xx
xx
xx
x
x
x
x
x
Fig. 17. Relationship between overall prevalence and intensity of S. mansoni infection as determined byegg output (measured by Kato–Katz technique) in various endemic areas. Each determination is basedon studies by different workers. e.p.g., eggs per gram. (From Jordan et al., 1993.)

Seasonal variations in the cercarial trans-mission rates in snails are most marked withthe bulinid hosts of S. haematobium livingin temporary bodies of water. In hyper-endemic habitats, such as canals, there maybe large numbers of snails producing cer-cariae of S. mansoni throughout the year.
Diurnal fluctuations in the productionof cercariae occur; cercariae of S. japon-icum (which is adapted for transmissionamong the nocturnal animal reservoirhosts) are mostly produced in the evening,while those of S. mansoni and S. haemato-bium are produced in the middle of theday. This factor is not usually of practicalimportance except where S. mansoni trans-mission occurs in running water, in whichcase washing and bathing are likely to bemuch safer in the morning and evening,when cercariae are less abundant.
SNAIL HOSTS OF SCHISTOSOMES
S. japonicum. Oncomelania is a conical,amphibious, prosobranch snail measuring3–10 mm and having 4–8 whorls, a dextralopening and an operculum covering the foot.Species involved are O. hupensis from main-land China, O. nosophora from south-westChina (and Japan), O. formosana fromTaiwan, O. quadrasi from the Philippinesand O. lindoensis from Sulawesi.
S. mekongi. Neotricula aperta is an aquaticsnail found in the Mekong River and itprobably also transmits S. sinensium.Robertsiella spp. are found in Malaysia andtransmit S. malayensis.
S. mansoni. Biomphalaria is a flattened pul-monate, planorbid snail with 3.5–7 whorls,measuring 7–22 mm. Four main groups arefound in Africa: (i) the pfeifferi group (fourspecies), which include the most importanthosts in Africa and the Middle East; (ii) thesudanica group (three species), which occurin both East and West Africa; (iii) thechoanomphala group (three species), whichlive in the great lakes and act as hosts inLake Victoria; and (iv) the alexandria group(two species), which occur sporadically inNorth, East and South Africa. In the western
hemisphere B. glabrata is the most impor-tant snail host, with seven other specieslocally involved.
S. haematobium. Bulinus is a turreted pul-monate snail with a left-handed openingwhen looked at with the spire upwards(Fig. 1). The height varies from 4 to 23 mm.The subgenus B. (Physopsis) is differenti-ated from B. (Bulinus) by having a truncatecolumella and a pointed end to the foot inliving specimens. Two main groups ofbulinid snails are important as hosts: (i) theafricanus group (ten species) of the sub-genus Physopsis is the more importantgroup in eastern and southern Africa andin most parts south of the Sahara; and (ii)the truncatus, forskali and reticulatusgroups (27 species) of the Bulinus sub-genus of snails act as hosts in the Near East(Iran, Egypt and Sudan), in Madagascarand in parts of both East and West Africa.
S. intercalatum. B. (Physopsis) africanus actsas snail intermediate host in Congo Republicand B. (Bulinus) camerunensis in Cameroon.
It must be borne in mind when attemptingto identify a snail that there are usuallymany other types of snail present in thesame body of water as the ones transmit-ting schistosomiasis, and they may bedifficult to differentiate. Specialized mono-graphs need to be consulted (Brown, 1980)or expert advice sought. It is also importantto distinguish clearly the cercarial speciesemerging, because of the large number ofanimal and bird schistosomes that use thesame snail hosts and produce cercariaesimilar to the human schistosomes. Indetailed epidemiological studies, it may benecessary to expose laboratory animals tothe cercariae and to identifiy any adultschistosomes that develop.
PREVENTION AND CONTROL
Personal prevention is by avoidance ofwater sources containing cercariae, impos-sible for fishermen to carry out andextremely difficult for children to be con-vinced of the necessity thereof. The most
The Trematodes 27

common source of infection in tourists toAfrica appears to be swimming in LakeMalawi. Thorough and speedy towellingafter swimming or wading in marshes mayprevent cercariae from penetrating.
Before attempting any control measure,it is essential to define its objectives – i.e.whether it is intended to prevent apread ofthe disease, to reduce morbidity or trans-mission or to permanently interrupt trans-mission (elimination in an area oreradication in a country or worldwide).Until the aims of a project have beenclearly defined, it is difficult to work outtechniques and costings, or even to evalu-ate its eventual success. While eradicationmust be the ultimate objective, it is not yeta feasible proposition in many areas.
There are five principal methods of con-trol: mass chemotherapy, destruction ofsnails, environmental sanitation, preven-tion of water contact and the possible useof mass vaccination. Which method ormethods are to be employed will dependvery much on the conditions where theyare to be applied but it is usually essentialto provide alternatives or to use two ormore methods simultaneously (e.g.chemotherapy, snail control and healtheducation). It is also important to deter-mine at the beginning how the success ofthe campaign is to be measured, and vari-ous indices can be used. The most commonassessment is that of prevalence, but inci-dence (measurement of the rate at whichpeople become infected) is more sensitiveand is particularly useful in masschemotherapy campaigns. Other measuresof assessment that can be used are theeffect on egg output (i.e. on intensity ofinfection) or on severity of clinical disease(morbidity), immunological methods andvarious indirect biological methods. Themost common biological method of assess-ing the success of measures against theintermediate host is a periodic snail countbut determinations of the infection rates insnails have been used.
Mass (universal) chemotherapy. The primaryobjective should be the reduction and pre-vention of morbidity, although a decrease in
the intensity of transmission is also impor-tant. Campaigns have been carried out inmany countries, often with great success,and at the outset are vertical, involving spe-cialized control units that have identifiedand mapped transmission sites, assessed thelevel of morbidity and educated healthworkers and the community about the con-trol programme (WHO, 1993; Savioli et al.,1997). Later, community-based control pro-grammes can be instigated, utilizing trainedcommunity health workers. If transmissionis intense (over 50% in children), then thewhole population can be treated; otherwise,7- to 14-year-old schoolchildren can betargeted, since they can be easily tested,treated and followed up and are likely to bethe most heavily infected section of the pop-ulation. A small number of heavily infectedindividuals produce a large percentage ofthe eggs reaching the environment (in oneclassic study with S. mansoni, 51% ofpatients had fewer than ten worm pairswhile only 7% had more than 80 – althoughone girl who died had 1600 worm pairs). Inall campaigns periodic re-treatment isessential in order to have a lasting effect.After campaigns with praziquantel in China,Indonesia and the Philippines, reduction inthe size of the liver and spleen was reported(WHO, 1993), while in patients with schisto-somiasis mansoni, hepatomegaly and peri-portal fibrosis have regressed after treatmentas measured by ultrasound (Boisier et al.,1998; Frenzel et al., 1999).
CONTROL OF SNAILS
Chemical control. The relationship betweenthe population dynamics of snail interme-diate hosts and disease transmission tohumans is poorly understood and completeeradication of snails from large areas is notfeasible. Mollusciciding is likely to be mosteffective in targeted focal sites, say wherechildren habitually bathe. Infection van-ished from Tunisia in 1983 after treatmentof oases.
Niclosamide (Bayluscide) can be used ata concentration of 8 p.p.m. hours (e.g. 0.33p.p.m. for 24 h) for aquatic snails (Plate 3)
28 Chapter 1

or 0.2 g m�2 on moist soil for amphibioushosts of S. japonicum.
Biological control. Carnivorous snails, suchas Marisa cornuarietis, or competitors,such as Melanoides tuberculata andTarebia granifera, have been added to habi-tats in Guadeloupe, Martinique, PuertoRico and St Lucia, with the result thatBiomphalaria has vanished from all oralmost all areas (Giboda et al., 1997;Pointier and Jourdane, 2000).
Ecological methods of control. Habitats canbe made unsuitable for snails by alternateflooding and drying of water channels orcovering and lining of canal systems, as inEgypt and Sudan, and filling in of marshyareas. In many areas such measures arelikely to be permanently successful, butthey require close cooperation betweenirrigation engineers and public healthworkers. In rice-growing areas of mainlandChina and in Leyte Island (Philippines)widespread cleaning of irrigation ditchesand filling in of ponds have resulted in agreat reduction in schistosomiasis japon-icum infection in recent years.
Prevention of water contact. The provisionof piped water supplies and alternativebathing and clothes-washing places can beeffective in preventing human contact withinfested water and, while expensive, doeshave wide-ranging health benefits.
For visitors and tourists, prevention canbe achieved by avoidance of contact withwater in ponds, canals, slow-flowingstreams and the shallow edge of lakes inendemic areas (although in Lake Victoria inEast Africa the snail Biomphalariachoanomphala transmits S. mansoni inwater 2 m deep). For tourists, swimming inLake Malawi or Lake Kariba is often fol-lowed by infection. Dibutyl phthalate orhexachlorophene spread over the skin willprotect for about 4 h.
Health education. Knowledge of the role ofindiscriminate defecation and urination inspreading the disease to members of a com-munity and the personal importance of
avoiding contaminated water needs to beeffectively communicated, particularly tochildren and adolescents. While it is prob-ably impossible in Africa to prevent chil-dren swimming in the water, they could betaught to urinate (and perhaps defecate)before entry.
Present position and future outlook for con-trol. Present control methods are capable ofsubstantially reducing transmission andparticularly morbidity in most areas pro-vided that they are properly designed andcarried out and if adequate funds are avail-able. However, new irrigation and hydro-electric schemes in many endemiccountries have increased the number ofsnail habitats. New human-made lakeshave followed the building of dams inmany countries: Côte d’Ivoire (LakesKossou and Taasbo), Egypt and Sudan(Lake Nasser); Ghana (Volta Lake); Nigeria(Kainji Lake); Zambia and Zimbabwe (LakeKariba); Senegal (Lake Guiers below theDiama dam and Manantali dam in Mali).The incidence of schistosomiasis in thepopulations bordering these lakes or theirrigation channels coming from them (asin Egypt and the Gezira scheme in Sudan)almost always rises. In Côte d’Ivoirearound Lake Kossou there used to be aninfection rate of 14% with S. haemato-bium, which has now risen to 53%, andaround Lake Taabo the rate has risen from0% to 73% (although the 2% infection ratewith S. mansoni has not changed so far);around Lake Kainji the infection rate withS. haematobium is now 14% in children(from nothing); around Lake Kariba there isnow a 70% infection rate in children withS. haematobium; infection has spreadgreatly in the Senegal River basin since thebuilding of the dams and there is now aprevalence of 91% with S. mansoni (from1%) and of 28% with S. haematobium(Southgate, 1997). One trend in Africacaused by changes in ecology following thebuilding of dams in Africa (e.g. in deltaareas of Cameroon, Ghana and Egypt) is thereplacement of S. haematobium by S. man-soni. The new opportunities for agricultureare also attracting possibly infected
The Trematodes 29

migrants and this is particularly apparentin the Gezira and in Ethiopia (although thebuilding of the Koka dam initially reducedtransmission).
Infection has been eliminated fromJapan, Tunisia and Montserrat recently andcampaigns against S. haematobium arebeing mounted in Mauritius (1.3% in adultsonly), Morocco (2.3% overall) and Tanzania(45% in Zanzibar), against S. mansoni inBrazil, Burundi, St Lucia and Venezuela(risen from 0.1% to 0.9% in under-10-year-olds in the last few years) and against bothspecies in Egypt, Ghana, Madagascar,Malawi, Mali, Nigeria, Saudi Arabia, Sudanand Zimbabwe; against S. japonicum inChina, Indonesia (1% overall infection rate)and the Philippines (3.6%); and against S.mekongi in Laos (72%) (WHO, 1993).
China has had control campaigns involv-ing mass treatment of people and cattle withanthelminthics (nowadays praziquantel),snail control and environmental engineeringfor the last 50 years but there were still esti-mated to be 1.06 million cases in humansand 0.25 million cases in cattle in 1997.However, S. japonicum has been eliminatedfrom 150 of the formerly 378 endemic coun-ties in 12 provinces south of the YangtzeRiver (Ross et al., 1997), while in 1955 therewere estimated to be over 11 million cases.Unfortunately, recent flooding has increasedthe extent of snail habitats. The ThreeGorges dam under construction on theYangtze River will, within the next 10 years,create the largest lake in the world in ahighly populous region where schistosomia-sis japonicum already occurs and wherethere will have to be about 2 million extramigrants from highly endemic areas, whichwill be flooded. The building of the damshould reduce the overall density of snailsalong the river banks but will provide manymore suitable, contaminated, marshlandhabitats and an increased contact with water.
Towards a vaccine. The need for a vaccineagainst schistosomiasis is underscored bythe fact that, although chemotherapy withpraziquantel is very effective, in theabsence of an effective immune responsereinfection tends to occur quite quickly.
Progress to date indicates that a vaccine isfeasible, since protection against sheepschistosomiasis (a problem in Sudan) usingirradiated cercariae has been successfullyachieved. However, any human vaccinewill have to consist of defined components,as cercariae might introduce viruses.
Various types of vaccine are being inves-tigated (Capron, 1998). These include thefollowing.
1. Peptide components of protective anti-gens. These can be used to generatedifferential responses, e.g. one elicitseosinophil-dependent ADCC, another stim-ulates delayed-type hypersensitivity (DTH)reactions with macrophage activation.Recently a multiple antigen peptide (MAP)vaccine has shown very promising results,inducing both T-cell and B-cell responsesin mice and in monkeys, reducing parasitefecundity and egg viability and decreasingliver pathology by over 70%. 2. Excretory/secretory (ES) antigens. ESmaterial obtained from postcercarial schisto-somula kept in vitro is effective in immu-nizing rats to subsequent infection. Thesecretions contained proteins in the range22–26 kDa (amplified by PCR) and inducean IgE-dependent ADCC.3. Membrane antigens. Molecules with amolecular weight of 23 kDa (e.g. Sm23 andSj23) occur in the tegument of all stages ofthe parasite but are often species-specific. 4. Paramyosin. This 97 kDa internal anti-gen (e.g. Sm97), obtained from schisto-somula or adults, induces responses thataffect the muscles and tegument of adults(Gobert, 1998). 5. DNA vaccines. Direct injection of thecoding sequence for known antigens isbeing investigated. Such a vaccine can beused alone or combined with chemother-apy, e.g. immunization with DNA for Sm28glutathione-S-transferase (GST) together withpraziquantel prevented the formation ofpathological lesions in mice (Dupre et al.,1999). 6. Irradiated live cercarial vaccines.Although these are not directly suitable forhuman use, they might provide usefulpointers to promising leads (Coulson, 1997).
30 Chapter 1

The most promising six antigens wererecently selected by the World HealthOrganization (WHO) for testing as vaccinesbut at best gave only a maximum of 60%protection (Katz, 1999).
A general problem facing the develop-ment of vaccines against any infectious dis-ease is the generation of an appropriateT-cell response (e.g. CD4+ or CD8) and, ifCD4+, then the appropriate Th subset, Th1or Th2. In addition, where the disease asso-ciated with infection is immune-mediated,as in schistosomiasis, vaccination must notinduce pathological responses. There iscontroversy about whether Th1 or Th2responses are most important and appro-priate for vaccine-induced resistance toschistosome infection, but it is probablethat a balanced response would be best(Wynn and Hoffmann, 2000).
The route of administration of any vac-cine is also very important, as is the choiceof a suitable adjuvant. Presentation of anti-gens via an engineered vector organism canovercome both problems. For example,Bacillus Calmette–Guérin (BCG) has beenengineered to produce an intracellularschistosome GST and this has induced astrong and long-lasting response in micewhen given by the intranasal route (amixed response of GST-specific immuno-globulin – IgG2a, IgG2b and IgA in serumand IgA in bronchial lavage fluids). A vac-cine against schistosomiasis haematobium(Sh28GST – Bilhvax) together with prazi-quantel treatment is undergoing field trialsin Senegal.
ZOONOTIC ASPECTS
Animal hosts are extremely important in thetransmission of S. japonicum and S.mekongi and in some countries can maintainthe parasite in the absence of humans. Inmainland China infection occurs in 40species of domestic and wild animals(Jordan et al., 1993) and is important as aserious cause of morbidity and mortality incattle and goats. In the Dongting Lake area ofHunan, 60% of cattle and buffaloes, 24% ofpigs, 9% of dogs and 7% of people werefound to be infected. Dogs and pigs play animportant part in maintaining transmission
on the island of Leyte in the Philippines,although many other species of animals areinfected, as they are on Sulawesi (Indonesia)and used to be in Japan. Molecular studiesindicate that S. japonicum is a species com-plex of many sibling species, and in Taiwanthe local form of S. japonicum occurs onlyin animals and is not infective to humans.The dog appears to be the only reservoir hostof S. mekongi in Laos. S. sinensium is a littleknown species with a laterally spined eggfound a few times in humans and rodents inChina and Thailand (Greer et al., 1989),while the ‘Tonle Sap schistosome’ has beennewly discovered in humans living aroundLake Tonle Sap in Cambodia (Pecoulas et al.,1995).
S. mansoni has been reported frequentlyfrom animals (of 38 species) but in manycases they do not appear to have any epi-demiological significance. However, baboonsand green monkeys are apparently main-taining infections among themselves inareas of East Africa and rodents may act asreservoir hosts in East Africa, Senegal andGuadeloupe and particularly in Brazil,where up to 15 species appear to be ofincreasing importance in maintaininginfection in urban and periurban areas ofthe north-east (Mott et al., 1995).
Animals appear to be of little or noimportance in the transmission of S.haematobium, although various primateshave been found infected, nor are they forS. intercalatum.
The schistosomes found in humans andanimals in Africa can be divided into twogroups of sibling species. Hybridizationcan occur experimentally within membersof each group but is not common in nature.In the S. haematobium/S. intercalatumgroup, S. mattheei Veglia and Le Roux,1929, is a parasite of sheep, cattle, wildanimals and occasionally humans in south-ern Africa. The eggs in the faeces or urinehave a terminal spine and measure120–180 µm in length (Fig. 124). Curiously,in all human cases, the eggs have occurredtogether with those of S. haematobium orS. mansoni; the females of S. mattheei areknown to be capable of producing eggsparthenogenetically and are perhaps carried
The Trematodes 31

by excess males of the other species. S.bovis (Sonsino, 1876) Blanchard, 1895, issimilar to S. haematobium and is found incattle in southern Europe, in Iraq and overmuch of Africa, but there have been veryfew genuine infections in humans(Mouchet et al., 1988). S. margrebowiei LeRoux, 1933, is a common parasite ofantelopes in Africa but the few humancases reported might be spurious. S. rod-haini Brumpt, 1931, is closely related to S.mansoni and is found in rodents and carni-vores in Central Africa; it has beenreported once from a human. S. curassoni,Brumpt 1931, is a parasite of domesticruminants in West Africa and has beendoubtfully reported from humans.
The cercariae of various mammal andbird schistosomes penetrate the skin ofhumans in many areas of the world but donot develop further. In a sensitized hostthey cause dermatitis at the site of entry,with irritation, pruritus, macules andpapules (‘swimmers’ itch’ or ‘schistosomedermatitis’). Examples are the cercariae ofthe bird parasites Austrobilharzia terri-galensis (Australia), Bilharziella polonica(Europe), Gigantobilharzia sturniae (Japan),G. huttoni (Florida, USA), Microbilharziavariglandis (Delaware, USA), Tricho-bilharzia brevis (Japan), T. maegraithi(Thailand), T. ocellata (worldwide) and T.stagnicolae (USA) and cercariae of mam-malian species of schistosomes, such asHeterobilharzia americana (Louisiana,USA), Orientobilharzia turkestanica (Chinaand Iran), Ornithobilharzia turkestanicum(Iran), Schistosoma bovis (Africa), S. spin-dale (Asia) and Schistosomatium douthitti(North America).
Schistosome dermatitis is more of a nui-sance than a serious medical problemwhen it occurs in bathers or water-sportsenthusiasts using lakes frequented bywaterfowl, but is of real economic impor-tance among rice planters in China,Malaysia and Vietnam.
Family Paragonimidae
Paragonimus westermani (Kerbert, 1878) Braun, 1899
SYNONYMS
Distoma westermani (D. westermanii)Kerbert, 1878; D. ringeri Cobbold, 1880.
DISEASE AND COMMON NAMES
Paragonimiasis or paragonimosis; pul-monary distomiasis, Oriental lung flukeinfection.
LOCAL NAMES
Pa-yard bai-mai nai pod (Thai).
GEOGRAPHICAL DISTRIBUTION
Paragonimus westermani is widely distrib-uted in the Chekiang Province of China,Korea, Laos, the Philippines, Taiwan andThailand and also occurs in smaller areas ofIndia, Indonesia, Japan, Malaysia andVietnam, with one or two cases in Russiaand Poland. Although there are at least 14other species that have been reported fromhumans, most of the estimated 21 millioncases in 39 countries worldwide are due to P.westermani, with 195 million people at risk.
LOCATION IN HOST
The adults are typically found in pul-monary cysts, usually in pairs.
MORPHOLOGY
A living Paragonimus is a thick fleshy flukemeasuring 8–16 � 4–8 mm and is reddish-brown in colour, with a tegument covered insimple or toothed spines. The ventral suckerlies just anterior to the middle of the bodyand is about the same size as the oral sucker.The testes are almost side by side in the pos-terior part of the body. The large excretorybladder is usually apparent in sections ofthe worm (Fig. 18 and Plate 4). The yellow-ish-brown egg is ovoid and thick-shelled. Itmeasures 80–110 µm � 50–60 µm, with arather flattened operculum, which gives theimpression of being too small, and is unem-bryonated when laid (Fig. 123).
32 Chapter 1

The Trem
atodes33
Map 3. Distribution of Paragonimus spp. and Opisthorchis felineus.

LIFE CYCLE
The eggs are usually swallowed and passout in the faeces but may also escape insputum. For further development theyhave to reach water. The miracidia hatchafter 3 weeks in water (at an optimum tem-perature of 27°C) and penetrate freshwatersnails. These are operculate genera, ofwhich the most important is Semisulcospira(S. libertina is an intermediate host speciesin China, Japan, Taiwan and Korea). Otherimportant snails are S. amurensis, Thiaragranifera and Oncomelania nosophora (seeFig. 1).
In the snail the miracidia develop intosporocysts followed by two generations ofrediae, and finally, in about 3 months, giverise to very short-tailed cercariae. The cer-cariae emerge from the snail and swim in
the water, where they can survive for only24–48 h.
If cercariae find freshwater crabs, princi-pally Eriocheir, the mitten crab in Asia andalso Potamon (Fig. 19) and Sesarma, orfreshwater crayfish (Astacus), they penetrateand encyst in the gills or muscles as meta-cercariae (measuring 250–500 µm).Freshwater crustaceans can probably alsobecome infected by ingesting unencystedcercariae in the water or even inside aninfected snail.
The crustacean intermediate hosts areeaten by humans as food. If they are eatenraw, the metacercariae hatch in the duode-num and young worms penetrate throughthe intestinal wall and pass across to theabdominal wall in about 6 h. The immatureflukes burrow through the diaphragm to the
34 Chapter 1
Fig. 19. Potamon, a crab second intermediate host of Paragonimus in Asia.
Fig. 18. Cut portion of lung (of a tiger) with cysts containing pairs of adult Paragonimus. Note largeexcretory bladder in each fluke.

The Trematodes 35
pleural cavity in 6–10 days and enter thelung capsule in 15–20 days. The first eggsare passed 60–70 days after infection.Although Paragonimus is hermaphrodite,two worms are necessary for fertilization totake place (Fig. 19 and Plate 4). This is truefor the usual diploid form of P. westermani,but there is also a triploid form, in whichparthenogenesis can occur; the adults of thelatter are larger and more pathogenic andthe eggs differ in size and shape (see Blairet al., 1999). The adult worms can live for20 years but usually die after about 6 years.
CLINICAL MANIFESTATIONSThere are no recognizable symptomsaccompanying the migratory phases.
The first signs are usually fever with adry cough, sometimes accompanied bybloodstained sputum containing eggs afterrupture of the cysts. Chest pain can besevere and, if the infection is heavy, thereis sometimes increasing dyspnoea andbronchitis. Many patients with haemo-ptysis are diagnosed as suffering frompulmonary tuberculosis.
Cerebral paragonimiasis may result inepileptic seizures, as well as headaches,visual disturbances and symptoms ofmeningitis.
PATHOGENESIS
Pulmonary. As flukes grow in the lungsthey cause an inflammatory reaction and
become surrounded by a cyst, possiblycaused by softening followed by fibrosis(Plate 4). However, as the cyst wall is usu-ally lined with columnar epithelium, it isprobable that, in most cases, it representsthe expanded wall of a bronchus. The cap-sule is collagenous and oedematous, withplasma cells, eosinophils, neutrophils,macrophages and fibroblasts. The cyst mea-sures about 1 cm in diameter after 90 daysand is filled with a thick brownish fluidcontaining eggs. Bronchopneumonia mayresult when a cyst bursts or retains anopening into a bronchus. Eggs are oftenpresent in the lung tissues and causepseudotubercles similar to those caused byschistosome eggs, with infiltration byeosinophils and lymphocytes, followed bygiant cells and fibroblasts.
Extrapulmonary. Adults are sometimesfound in many parts of the body, particu-larly in the organs of the abdominal cavityor subcutaneous tissues. In these sites theworms are rarely fertilized and presumablyhave developed from larvae that lost theirway. This occurs more frequently withspecies of Paragonimus which are lessadapted to humans than P. westermani.Most serious complications follow thepresence of flukes in the brain (Fig. 20).This may be common in areas of highendemicity (e.g. over 5000 cases perannum in South Korea). Adults found in
Fig. 20. Radiograph of head of a girl with soap-bubble type of calcification surrounding cysts containingadult Paragonimus in occipital lobes.

36 Chapter 1
the brain usually have eggs, unlike thoserecovered from other extrapulmonarysites; it is believed that they migrate fromruptured lung cysts along the soft tissuessurrounding the veins into the cranium.The flukes cause necrosis of the surround-ing brain substance and are eventuallyencapsulated, with granulomas surround-ing released eggs lying along the fibrouswall of the cyst. When a fluke dies, thecavity of the lesion becomes filled withpurulent material. Lesions may occur inthe cerebral cortex, cerebellum, basal gan-glia and medulla oblongata. Cerebralparagonimiasis can easily be confusedwith brain disease caused by otherhelminths, such as Schistosoma japon-icum, Gnathostoma spinigerum andParastrongylus cantonensis or larvae ofEchinococcus granulosus and Taeniasolium, as well as with viral or bacterialinfections and with brain tumours.
Levels of non-specific IgE are elevatedin infection and IgG4 levels are also veryhigh, possibly to obstruct IgE-mediatedimmunopathogenicity. Immune evasionmechanisms are probably employed by theparasites, with periodic sloughing of theouter calyx layer on the tegument.
DIAGNOSIS
Clinical. A cough with brown sputum butotherwise good health is often indicative ofa light infection in a patient from anendemic area. Characteristic nodular or ringshadows apparent on X-ray or computer-assisted tomography (CT) scan can be recog-nized by an expert, but the disease is oftenmistaken for tuberculosis.
Parasitological. A certain method of diagno-sis is to find the characteristic eggs in spu-tum or faeces, which need to bedifferentiated from those of Achillurbainia(see below). Various concentration tech-niques may be necessary for detection ofeggs in faeces (see p. 258) as, in chronicinfections, very few eggs may be passed;centrifugation with 2% NaOH is a methodthat can be used.
Immunological. The intradermal (ID) test hasbeen widely used for many years and is use-ful in differentiating paragonimiasis fromtuberculosis, but it is very non-specific andthe test lasts for many years after cure or thedeath of parasites. There have been numer-ous trials of ELISA tests in the last 20 years,some using crude antigens, but recentlymore specific ES antigens (probably cysteineproteases) and tests for parasite antigenusing mAbs have increased sensitivity andspecificity (Ikeda, 1998; Blair et al., 1999).Dot ELISAs using dried antigens dotted onnitrocellulose plates are also being used.Immunological techniques can differentiatebetween species of Paragonimus, particu-larly between P. heterotremus and P. wester-mani (Dekumyoy et al., 1998).
TREATMENT
The drug praziquantel has revolutionizedtreatment in the last few years when givenat 25 mg kg�1, three times a day after mealsfor 3 days. Side-effects are usually minorand short-lived. Triclabendazole at 5 mgkg�1 daily for 3 days has also proved to beeffective in cases in Africa and SouthAmerica, one of which was resistant topraziquantel.
Surgery, which used to be necessary forcerebral cases, is now rarely used since theadvent of effective drug therapy, except forsome cases of cutaneous infection whereflukes can be easily removed.
EPIDEMIOLOGY
Paragonimiasis can be contracted fromboth raw and pickled crabs and crayfish. Inthe Philippines a dish called kinagang ismade from crab juice and in the Chekiangprovince of China ‘drunken crabs’(Potamon sp.) are eaten alive after steepingin rice wine. In Japan crabs (usuallyEriocheir sp.) are usually eaten well-cooked but the metacercariae contaminateknives, hands and chopping-boards duringpreparation in the kitchen and are thenaccidentally transferred to salad vegetables.Crushed crayfish juice is used as a treat-ment of measles in Korea and in this wayyoung children become infected. In Japan,wild boar, hens and ducks appear to act as

paratenic hosts. Paratenic hosts (includingrodents) are likely to be important in trans-mission to large carnivores.
Metacercarial cysts accumulate duringthe life of the crab and in large specimensthere may be as many as 1000 cysts g�1 oftissue.
PREVENTION AND CONTROL
Metacercarial cysts in the crustaceans arekilled by heating in water at 55°C for 5 minand even light cooking renders them safe.However, cysts can live for up to 48 hwhen frozen at �20°C, although they die inminutes when dried.
Where zoonotic infections are rare, sani-tary improvements to prevent eggs reachingsnail habitats in water can be used. InKorea there has been a reduction in preva-lence caused by increasing industrializationand pollution of habitats.
National campaigns need to involvepublic awareness and community participa-tion, food control, sanitary waste disposaland adequate monitoring and surveillance(WHO, 1995).
ZOONOTIC ASPECTS
Paragonimus westermani is a natural parasiteof carnivores; in fact, the specific name hon-ours a director of Amsterdam zoo, where thefirst specimens were found in the lungs of atiger. It is found especially in the members ofthe cat family, such as the tiger, leopard anddomestic cat, in most parts of its range but hasalso been reported from dogs, pigs and mon-keys. In China, Japan and Korea, animal hostsare probably not of importance in transmis-sion to humans, while in Indonesia, Malaysiaand Sri Lanka infection is confined entirely toanimals and human cases do not occur.
Over 30 probably valid species ofParagonimus have been described (Blair etal., 1999) differing (often slightly) in mor-phology and life cycle. Many cause zoonoticinfections in humans in various parts of theworld (WHO, 1995), as outlined below.
Africa. P. africanus Voelker and Vogel,1965, and P. uterobilateralis Voelker andVogel, 1965, are parasites of the mongoose,civet cat and humans in Cameroon and
eastern Nigeria and occasionally also in 12other countries of West and Central Africa(WHO, 1995).
Americas. P. caliensis Little, 1968, is a nat-ural parasite of opossums in Central andSouth America. P. kellicotti Ward, 1908, isfound in many carnivores, pigs and opos-sums in North America, with a few cases inhumans in Canada and two in the USA. P.mexicanus Miyazaki and Ishii, 1968, isfound in many carnivores and opossums andoccasionally humans in North, Central andSouth America; most cases are in Ecuadorand cerebral infections are not infrequent.
Asia. P. bangkokensis Miyazaki andVajrasthira, 1967, is found in cats and othercarnivores in China, Russia and Thailand.P. heterotremus Chen and Hsia, 1964, isfound in carnivores and rodents in China,Laos, Thailand and Vietnam. P. miyazakiKamo et al., 1961, is a parasite of carni-vores in Japan, and adults do not mature inhumans. P. ohirai Miyazaki, 1939, is a para-site of rats and mice in Japan. P. philip-pinensis Ito et al., 1978, is primarily aparasite of dogs in the Philippines. P. pul-monalis (Baelz, 1883) Miyazaki, 1978, isfound in cats and dogs in Japan, Korea andTaiwan. P. sadoensis Miyazaki et al., 1968,has been found only in humans in Japan. P.(= Pagumogonimus) skrjabini Chen, 1960(= P. hueitungensis Chung et al., 1975) is aparasite of cats, dogs and palm civets inChina: in human infections worms do notmature and the young migratory worms areoften subcutaneous or cerebral.
Family Achillurbainiidae
Achillurbainia nouveli Dolfuss, 1939, wasfirst found in a palpebro-orbital abscess froma Malayan leopard in a zoo in France. Anadult was subsequently found in a 10-year-old Chinese girl. The adults are common inthe lungs of rats (Rattus muelleri) in WestMalaysia and freshwater crabs probably actas second intermediate hosts. A. reconditaTravassos, 1942, developmental stages havebeen found in humans in Brazil, Honduras
The Trematodes 37

and Louisiana, USA; adults are usually inopossums.
Poikilorchis (or probably Achillurbainia)congolensis Fain and Vandepitte, 1957, is asimilar parasite and was first found in aretroauricular cyst in Congo Republic.Similar eggs have been found in cysts intwo boys in Sarawak and in Cameroon andNigeria. The eggs are similar to those ofParagonimus but are smaller (63 µm � 40µm).
Family Opisthorchidae
Clonorchis sinensis (Cobbold, 1875) Looss, 1907
SYNONYMS
Distoma sinense Cobbold, 1875;Opisthorchis sinensis Blanchard, 1895.
Clonorchis was erected as a genus onthe basis of the shape of the testes, but it isdoubtful that this is sufficient to warrantseparation from the very closely relatedgenus Opisthorchis. However, it still per-sists in the literature and, unless there is aformal decision by the InternationalCommisssion on Zoological Nomenclature,it is reluctantly retained here.
DISEASE AND POPULAR NAMES
Clonorchiasis or clonorchiosis, opisthor-chiasis sinense; Chinese liver fluke disease.
LOCAL NAMES
Oogyubyo, Hareyamai or Suishobyo(Japanese).
GEOGRAPHICAL DISTRIBUTION
About 7 million cases in nine countries inthe world (with 290 million people at risk);in China (4.7 million cases), Hong Kong(0.3 million), India (rare in Assam), NorthKorea (0.95 million), Russia (Siberia),Taiwan and Vietnam (1 million).
LOCATION IN HOST
Adults inhabit the bile-ducts just under theliver capsule, but in massive infectionsmay be found in the gall-bladder, pancre-atic ducts and all bile-ducts as well.
MORPHOLOGY
Adult flukes measure 11–20 mm � 3–4.5mm and are elongated and extremely flat,tapering anteriorly and rounded post-eriorly; when living they are almost trans-parent. There can be a wide variation in thesize of adults, depending upon the intensityof infection and on the diameter of the bile-ducts in which they are living. The tegu-ment has no spines. The ventral sucker issmaller than the oral and is situated about aquarter of the length from the anterior end.The two deeply lobed testes lie one behindthe other at the posterior end. They providean important identification feature, particu-larly in stained specimens (Fig. 2).
LIFE CYCLE (Fig. 21)In most infections there are usually onlya few adults present but there can be upto 6000 and a daily egg production of1000 eggs ml�1 bile or 600 g�1 faeces maybe recorded.
The small yellowish-brown eggs (Fig.123) measure 23–35 µm � 10–20 µm andhave an operculum at one end and a smallknob at the other. The eggs contain a fullyformed miracidium when laid and pass fromthe bile-ducts into the intestine and out withthe faeces. They have to reach water, wherethey do not hatch but are ingested by variousspecies of snails (Alocinma longicornis insouthern China, Bulimus fuchsiana in north-ern China and Parafossarulus manchouricusin most endemic areas are commonlyinfected species). The miracidia hatch in therectum of the snail and develop into sporo-cysts in the hepatopancreas, where they giverise to numerous rediae.
Cercariae are produced 21–30 days afterinfection. These have an unforked keeledtail, and emerge into water, where they livefor only 1–2 days, unless they can pene-trate beneath the scales of freshwater fishand form a metacercarial cyst in the sub-cutaneous connective tissues, usually thosenear the caudal fin.
Over 100 species of fish have beenimplicated as secondary intermediatehosts of C. sinensis and up to 3000 cysts
38 Chapter 1

have been found in a single fish.Metacercarial cysts measure 140 µm �120 µm and are infective in about 23 days.When fish are eaten raw or undercooked,the metacercarial cysts hatch out in thehuman duodenum and the young flukesmigrate up the bile-ducts, maturing in 3–4weeks. The adult flukes can live for up to26 years but on average survive for 10years (Chen et al., 1994).
CLINICAL MANIFESTATIONS
The majority of infections (about 70%) aresymptomless and the presence of worms isonly diagnosed at autopsy, when up to 100flukes may be found in the bile-ducts.Moderate infections, due to the presence of100–1000 flukes, result in diarrhoea, abdomi-nal discomfort and some splenomegaly. Whena heavy infection, which may consist of manythousands of worms (the maximum recordedbeing 21,000), is contracted at one time, there
The Trematodes 39
Map 4. Distribution of Clonorchis sinensis and Opisthorchis viverrini.

are additional symptoms, such as a suddenonset of fever, acute pain in the upper rightquadrant, liver enlargement and tenderness,oedema, an increased erythrocyte sedimenta-tion rate and up to 40% eosinophilia; symp-toms of cholecystitis and hepatitis follow afew weeks later, and there may also be boutsof recurrent gall-bladder colic. There is usu-ally loss of weight, with slight jaundice, andlater there may be tachycardia, with palpita-tions and vertigo. However, in an endemicarea, parasites are often acquired progressivelyover a period of months or years, and the
onset of symptoms is more insidious.Portal hypertension may occur with
splenomegaly (this is found in 20–40% ofautopsies) and ascites is almost always pre-sent in fatal cases, although not usuallydiagnosed during life.
Prognosis depends largely on the numberof parasites present and, as the great majorityof cases have only light infections, no illhealth ensues. However, the prevalence can beso high in endemic regions that even the lowproportion of heavily infected individuals canprovide an important source of morbidity.
40 Chapter 1
oral sucker
uterus
testis
0.1
mm Cats and dogs
act asreservoir hosts
excretory bladder
ME
TAC
ER
CA
RIA
Ein
gest
edin
raw fishE
xcys
t in
duod
enum
ADULTS live in bile ducts,mature in 3–4 weeks
10 mmEmbryonated EGG(30�m � 16�m) infaeces reaches waterbut does not hatch.Can survive for over3 months in water
operculum
EGGS in faeces
EGGS ingeste
d
by snail
glanddigestive gland
gills
CERCARIAE leave snailand penetrate fresh water
fish
stages in snail
0.05
mm
excretorybladder penetration
glands
eye spot
0.3
mm
CERCARIAhangs in water then
attaches and penetratesinto any fish it comes
into contact with.Can live 1–2 days
in water
0.1
mm
0.1
mm
pharynxdeveloping
cercaria
REDIA indigestive gland
by 17th day
LATESPOROCYST
developingredia
EARLY SPOROCYSTin gut or rectum
of snail
MIRACIDIUMhatches in gut ofsnail (unlike mosttrematodes wheremiracidium hatchesin water and activelypenetrates snail)
0.01
mm
rectum
METACERCARIAFound in subcutaneoustissues and muscles of fish,particularly near caudal fin.
Fig. 21. Life cycle of Clonorchis sinensis. Figure of courtesan and cat by Kaigetsudo.

Death is only likely to occur where thereis an intercurrent bacterial infection (apartfrom the possibility of cholangiocarcinoma);salmonellae in particular flourish in bile-ducts attacked by Clonorchis. A similar rela-tionship has been reported with urinaryschistosomiasis, where it has been shownthat Salmonella typhi may live on the insideand outside of the body of the schistosomes.
PATHOGENESIS
At operation the presence of severe infec-tion is obvious from gross examination ofthe liver because the thickened bile-ductsappear as pale patches on the surface. If theliver is cut, the slimy, brownish flukesoften emerge from the ducts.
The adult worms presumably feed mainlyon the biliary epithelium but, unlikeFasciola hepatica, have no spines on thetegument to cause mechanical damage. Inthe early stages there is an inflammatoryreaction in the biliary epithelium and inheavy infections there is cellular infiltration,consisting mainly of lymphocytes and leuco-cytes, particularly in the outer part of thefibrous wall. This is followed by hyperplasiaand metaplasia of the epithelium, withmuch mucus, and there may be the forma-tion of new biliary ductules (Fig. 22).Because immature Clonorchis migrate up thebile-ducts, there is not the pathology associ-ated with early Fasciola infections, whichburrow through the liver parenchyma (see p.47). In the chronic stage there is an encapsu-lating fibrosis of the ducts and destruction ofthe adjacent liver parenchyma. A fibroticreaction surrounds eggs that reach the liverparenchyma, but there is not the granulomaformation around intact eggs which occursin schistosomiasis (Fig. 14).
Stones are common in the bile-ductsand gall-bladder in cases of C. sinensisinfection and were found in 70% of autop-sies in a study in Hong Kong. The remainsof adult worms are sometimes found insidethe stones. Recurrent pyogenic cholangitisis one of the most important complicationsof clonorchiasis, as well as of ascariasis (p.151). The cholangitis often follows a longrelapsing course, eventually leading towidespread fibrosis. Cirrhosis of the liver is
found in about 10% of autopsies, but thereis no evidence that it is directly related tothe presence of the parasite and it may becoincidental or accentuated by an accom-panying bacterial infection. Clonorchisoften enters the pancreatic ducts andcauses pancreatitis, with adenomatoushyperplasia; occasionally there may beacute inflammation.
Carcinoma of the bile-ducts is found inassociation with clonorchiasis in Cantonand Hong Kong and is far more commonthan in non-endemic regions (IARCWorking Group, 1994); it has also beenreported from experimental infections indogs and cats.
IMMUNOLOGY OF LIVER FLUKES
Immunological aspects of infection withFasciola hepatica (see p. 46) have beenextensively studied in domestic ruminantsand in laboratory rodents. Infections areassociated with blood eosinophilia, strongantibody responses and immune-mediatedinflammation in the liver and bile-ducts.Evidence sluggests that infection elicitsstrongly polarized Th2 responses but alsothat flukes are capable of down-regulatinghost immunity in various ways. Infectionswith Clonorchis sinensis similarly induceeosinophils and profound inflammatoryresponses in the liver (in some cases result-ing in cholangiocarcinoma) as well as para-site-specific IgA, IgE and IgG; it can beassumed therefore that a similar polariza-tion of the immune response occurs.Evidence for immunity to reinfection inhumans is not strong, there being no declinein prevalence or intensity of infection in chronically exposed populations.Experimental studies using Opisthorchisviverrini in hamsters have clarified someaspects of the immune response and sug-gested the operation of a degree of immuneprotection. This has also been proposed inhuman infections with this species. In anendemic area individuals who were egg-negative had higher levels of IgA, IgG andIgM than those who were egg-positive andthey showed recognition of specific anti-bodies not detected by antigens of infectedindividuals. Studies in hamsters have also
The Trematodes 41

shown that parasite antigen accumulates inthe tissues around the worms and is associ-atated with marked inflammatory responses.
DIAGNOSIS
Parasitological. The most important methodof diagnosis is examination of the faecesfor eggs (p. 254). When eggs are scanty,concentration methods (Kato–Katz or sedi-mentation techniques) can be used or aduodenal aspirate taken. In symptomlessinfections large numbers of eggs are usu-ally produced. Any eggs found need to bedistinguished from those of Heterophyes,Metagonimus, Metorchis or rarelyDicrocoelium, which are similar.
Immunodiagnosis. Intradermal and comple-ment fixation tests have been in use formany years but remain positive for manyyears after cure and they also cross-reactwith Paragonimus and Schistosoma.
An ELISA using a purified recombinantantigen of adult worms (mol. weight 28kDa) looks promising (Yong et al., 1998), asdoes application of a dot-immunogold–silver stain (Liu et al., 1995).
TREATMENT
Praziquantel at three times 25 mg kg�1
daily for 1–3 days or at a single dose of 40mg kg�1 (given in Korea to reduce side-effects) is stated to be effective for adultsor children over the age of 4. However,Clonorchis is not nearly as sensitive to theparalytic action of the drug as are theschistosomes.
EPIDEMIOLOGY
Infection is widespread in some endemicareas: 40% in adults in the Red Riverdelta region of North Korea, 24% in somevillages of the Amur River area of Siberia,23% in Hong Kong, 15% in Guangdong(China).
Infection is usually contracted from eat-ing raw or undercooked fish and fish dishes(usually cyprinids, such as Carassius caras-sius, Cyprio carpus and Pseudorasboraparva, but about 110 species have beenincriminated (see WHO, 1995)). However,the metacercarial cysts are very sticky andcan stick to the hands and to knives andboards, which are subsequently used forchopping salad vegetables, so that the para-site can sometimes be transmitted evenwhen fishes are cooked before eating.
42 Chapter 1
Fig. 22. Section of liver with two adult Clonorchis in bile-duct. There is marked epithelial proliferation,oedema of the mucosa and some fibrosis (see also Plate 6). The ventral sucker, uterus filled with manyeggs and testes (in fluke on left) are apparent.

In South Fukien province in China, fishis not commonly eaten raw, but decoctionsof raw crushed fish are used as a cure forasthma and freshwater shrimps (which canact as a secondary intermediate host in thisarea) are eaten as a cure for nosebleeds.Human cases in this province are mostly inchildren; the parasite being maintained indogs. In South Korea, infection formerlyoccurred in men, who traditionally eat rawfish to increase their thirst at rice winedrinking parties.
In rural areas of China, human faeces(‘night-soil’) was commonly used to enrichponds containing food fish and, as theseponds also often contain suitable snails,maximum opportunities for continuationof the life cycle were provided (Plate 5).Nowadays, however, untreated faeces arenot commonly used in mainland China.
There is a recent tendency in many peo-ple in Europe and North America to eatmore raw and lightly cooked fish and thereis a danger of cases of food-bornehelminths increasing in tourists and fromimported foodstuffs.
CONTROL AND PREVENTION
The most obvious method of prevention isthorough cooking of fish, but this is notalways feasible as raw fish is a traditionalitem of diet in endemic regions. Deep-freezing of fish at �30°C for 32 h is recom-mended for killing cysts.
Control is possible by sanitary disposalof faeces to prevent eggs reaching ponds orslow-flowing streams and canals. Evenstorage of faeces for a few days or additionof ammonium salts as fertilizer beforeadding to ponds is an effective measure.The use of molluscicides is difficultbecause of toxicity to fish.
National campaigns should follow thesame guidelines as for Paragonimus and otherfood-borne trematodes (see WHO, 1995).
ZOONOTIC ASPECTS
Dogs are the most important reservoir hostsbut infections occur in most fish-eatingcarnivores. Pigs and rats can also benaturally infected.
In the South Fukien province of China,
about 30% of dogs are infected and theseare almost entirely responsible for perpetu-ating infection.
Opisthorchis viverrini (Poirer, 1886) Stiles and Hassall, 1896
SYNONYM
Distoma viverrini Poirer, 1886. The nameshould more correctly have been O.viverrinae.
DISEASE AND LOCAL NAME
Opisthorchiasis or opisthorchiosis viverrini;Pa-yard bai-mai nai tub (Thai).
GEOGRAPHICAL DISTRIBUTION
Thailand, Cambodia and Laos, with about9 million cases (and 50 million at risk), ofwhich 7.3 million are in Thailand (see Map4, p. 39).
LOCATION IN HOST
Adults inhabit the bile-ducts.
MORPHOLOGY
The adults are very similar to those of C.sinensis but the testes are lobed rather thandendritic, and there are differences in theflame-cell pattern.
LIFE CYCLE
Very similar to that of C. sinensis. Snailsinvolved in Thailand are subspecies ofBithynia siamensis, with 15 species of fishbelonging to seven genera acting as secondintermediate hosts (WHO, 1995), such asPuntius orphoides (‘pla pok’ in Thai) andHampala dispar (‘pla suud’), which areeaten either raw (‘koi pla’) or semi-fermented (‘pla ra’).
CLINICAL MANIFESTATIONS AND PATHOGENESIS
The majority of infected individuals (about80%) show no symptoms or signs attribut-able to the parasite apart from eosinophiliaand there are likely to be less than 1000 eggsg�1 faeces. Although there may be localdamage to the bile-duct epithelium, there is
The Trematodes 43

no noticeable damage to liver function. Withheavier infections (10,000–30,000 eggs g�1
faeces), diarrhoea, epigastric and upperright-quadrant pain, anorexia, lassitude,jaundice and mild fever may slowlydevelop. In long-standing infections theremay be oedema of the legs, accompanied byascites, mild periportal fibrosis, hepato-megaly, inflammation and biliary epithe-lium hyperplasia.
In heavy infections the gall-bladderbecomes enlarged and non-functional, withcholecystitis, cholangitis, liver abscess andgallstones. Cholangiocarcinoma is stronglycorrelated with O. viverrini infection (Plate6) and few patients live longer than 6months after diagnosis (Elkins et al., 1990;IARC Working Group, 1994).
The immunology of liver-fluke infec-tions is discussed on p. 41.
DIAGNOSIS AND TREATMENT
Diagnosis is by finding eggs in the faeces.They are similar to those of C. sinensis butare slightly narrower (30 µm � 12 µm) andmore regularly ovoid, without such a clearshoulder at the operculum (Fig. 123). Theeggs of an intestinal fluke, Phaneropsolusbonnei (see p. 59), which has been reportedfrom humans in north-east Thailand, aresimilar in size but are smoothly ovoid andcontain an undifferentiated ovum whenpassed in the faeces.
Immunodiagnosis is not widely usedbut a mAb ELISA using mAbs that candetect an 89 kDa metabolic antigen from O.viverrini in faeces is stated to be both sensi-tive and specific (Sirisinha et al., 1995).
Praziquantel can be given as forClonorchis and is equally effective.
EPIDEMIOLOGY
There are prevalence rates of over 70% inethnically similar rural communities ofnorth-eastern Thailand (35% overall) andLaos, and it has been estimated that infectioncosts $120 million annually in lost wagesand medical care. Infection in humans occursprincipally at the end of the rainy season andthe beginning of the dry season (Septemberto February), as fish contain more metacer-cariae and are more easily caught before
ponds dry in March. Infection is very uncom-mon in children under the age of 5.
The metacercarial cysts are slightly lessresistant to pickling than those of C. sinen-sis, but thorough cooking or deep-freezingof fish is the only sure method of personalprevention.
ZOONOTIC ASPECTS
O. viverrini is a natural parasite of the civetcat but is common in domestic cats anddogs, which play a role in maintainingtransmission, although human-to-humantransmission is common.
Opisthorchis felineus(Rivolta, 1884) Blanchard, 1895
SYNONYMS
Distoma felineum Rivolta, 1884; D. sibir-icum Winogradoff, 1892 (first report fromhumans).
DISEASE AND POPULAR NAME
Opisthorchiasis or opisthorchiosis felineus;cat liver-fluke disease.
GEOGRAPHICAL DISTRIBUTION
About 1.5 million cases (with 14 million atrisk): Russia (Siberia with 1.2 millioncases), Belarus, Kazakhstan, Ukraine (0.3million) (see Map 3, p. 33).
LOCATION IN HOST
Adults are found in the distal bile-ductsand sometimes proximal ducts also.
MORPHOLOGY
Adults are identical to those of O. viverrini.Differentiation is based on differences inthe flame-cell patterns of the cercariae andin the life cycle.
LIFE CYCLE
Very similar to the previous two species.The snail intermediate hosts are species ofCodiella (= Bithynia), particularly C.leachi. Twenty-two species of fish belong-ing to 17 genera transmit infection (seeWHO, 1995).
44 Chapter 1

CLINICAL MANIFESTATIONS
Similar to the other two species, exceptthat symptoms are more likely to occur inthe acute phase (2–4 weeks after infection),such as high fever, abdominal pain andhepatitis-like symptoms, accompanied byeosinophilia (possibly because more infec-tions are in adult immigrants).
The connection with cholangiocarcinomais not as clear as with O. viverrini or C.sinensis (IARC Working Group, 1994),although the ratio of hepatic cell carcinomato bile-duct carcinoma in uninfected indi-viduals from an endemic area of Siberia wasfound to be 8:1, while in infected people theratio was reversed. In a fatal case in Russia, atotal of 7400 worms were recovered.
DIAGNOSIS AND TREATMENT
Eggs, which are identical to those of O.viverrini, can be recognized in the faeces.They are likely to be scanty in clinicallyimportant infections and concentrationtechniques may be necessary. Immuno-diagnostic tests are being investigated.
Treatment as for C. sinensis.
EPIDEMIOLOGY
In the central area of western Siberia thereare prevalence rates of 40–95% and thehighest infection rates in Russia in generalare found in river basin areas. In endemicareas of Ukraine rates vary from 5% to40%. There may be very heavy infectionsof metacercariae in food fish.
A popular raw fish dish, ‘stroganina’, isresponsible for many infections.
ZOONOTIC ASPECTS
Cats provide an important reservoir hostand may have high infection rates. Infectionin a range of animals, such as foxes, dogs,cats and pigs, is found over a wide area ofeastern Europe. O. guayaquilensis is a para-site of cats and dogs with a human focus inan area of Ecuador and O. noverca is a com-mon parasite of pigs and dogs with twohuman cases in India.
Metorchis albidus (Braun, 1893) Looss,1899, and M. conjunctus (Cobbold, 1864)Looss, 1899, are two liver flukes found inNorth American fish-eating carnivores. The
latter species was responsible for infectionin 19 people in Canada who ate sashimi ina restaurant. They suffered from upperabdominal pain, low-grade fever andeosinophilia, lasting for up to 4 weeks;praziquantel was very effective in treat-ment (MacLean et al., 1996).
Pseudamphistomum truncatum (Rudolphi,1819), Luhe, 1909 is a liver parasite of wildcarnivores, cats and dogs and has occasion-ally been found in humans in Russia(Siberia) from eating raw fish. P. aethiopicumPierantoni, 1942, was reported once from ahuman in Ethiopia.
Family Dicrocoeliidae
Dicrocoelium dendriticum (Rudolphi, 1818) Looss, 1899
SYNONYMS
Fasciola lanceolata, F. dendritica, D. lance-olatum.
DISEASE AND POPULAR NAME
Dicrocoeliasis or dicrocoeliosis; lancet-fluke infection.
GEOGRAPHICAL DISTRIBUTION
Dicrocoelium is a cosmopolitan parasite ofthe bile-ducts of sheep and other herbi-vores. True human infections are rare, butspurious infections from ingestion of eggsin liver are not uncommon. Real or spuri-ous infection was found in 0.2% of about27,000 patients in a hospital in Riyadh,Saudi Arabia, and in 0.2% of a rural popu-lation in Turkey, and occasional humancases are reported worldwide.
LOCATION IN HOST
Adults live in the bile-ducts.
MORPHOLOGY
Adult worms are flat and lancet-shaped,measuring 5–15 mm � 1.5–2.5 mm. Thetestes are in tandem just behind the ventralsucker. The vitellaria are restricted and theuterus takes up most of the posterior partof the body.
The Trematodes 45

LIFE CYCLE
This is of great interest zoologically, asDicrocoelium is one of the few trematodesthat utilize terrestrial snails as intermediatehosts (Zebrina or Helicella in Germany).The snail ingests the eggs containingmiracidia and eventually the cercariaeleave the snail in slime balls, each contain-ing 200–400 cercariae. These are theningested by ants, and the ants infected withmetacercariae are eaten by sheep (or rarelyhumans) on vegetation. Dicrocoelium waspresent in the livers of 37 autopsy cases inUzbekistan and infected ants were found onlocal mulberry trees (Azizova et al., 1988).
CLINICAL MANIFESTATIONS AND PATHOGENESIS
In the few proven cases detected in life,symptoms have been of mild hepatitis.
Pathology in animals is similar to that ofFasciola but less severe.
DIAGNOSIS AND TREATMENT
Characteristic eggs, measuring 38–45 µm �22–30 µm, may be found in the faeces (Fig.124). It is essential that patients are kept ona liver-free diet for a few days in order torule out spurious infections. Treatment ofone case with triclabendazole was notsuccessful.
Dicrocoelium hospes Looss, 1907, is foundin domesticated and wild herbivores inAfrica and human cases have beenreported from Ethiopia, Ghana, Kenya,Mali and Sierra Leone.
Eurytrema pancreaticum (Janson, 1889)Looss, 1907, is a parasite of the pancreaticducts of domestic herbivores in Asia andSouth America. Grasshoppers act as secondintermediate hosts. Three cases have beenreported from humans (Ishii et al., 1983),with worms in the dilated pancreatic ducts.
Family Fasciolidae
Fasciola hepatica Linnaeus, 1758
SYNONYM
Distomum hepaticum L., 1758.
DISEASE AND POPULAR NAMES
Fascioliasis or fasciolosis, distomiasis;liver-fluke infection, liver-rot (in sheep).
GEOGRAPHICAL DISTRIBUTION
About 2.4 million human cases in 61 coun-tries worldwide (with 180 million at risk).Fasciola is primarily an animal parasite ofgreat veterinary importance and is found insheep and cattle kept on damp and muddypastures in all the sheep- and cattle-raisingareas of the world. Human infections havebeen reported most frequently fromBolivia, Cuba, Ecuador, Egypt, France, Iran,Peru, Portugal and North Africa and aremuch more common than previouslythought.
LOCATION IN HOST
Adult worms inhabit the bile-ducts.
MORPHOLOGY
The hermaphroditic adult flukes measure20–30 mm � 13 mm wide (specimens areusually smaller in heavy infections) andare flat and leaflike, with a spiny tegument(Plate 7).
The oral sucker measures 1.0 mm indiameter and the ventral sucker, which liesclose behind it, about 1.6 mm. The maleand female reproductive systems are bothvery dendritic, as in Fasciolopsis buski, butin Fasciola the two gut caeca are alsohighly branched.
The dendritic testes lie one behind theother in the middle portion of the body.The small, highly branched, ovary is infront of the anterior testis. The dendriticvitellaria lie in the lateral fields and havevitelline ducts leading to the ootype. Theuterus, filled with eggs, is confined to theanterior third of the body.
LIFE CYCLE
In sheep the eggs are laid in the biliarytracts, reach the intestine and are passed outin the faeces. The undifferentiated ovumdevelops into a miracidium under moistconditions in 9–15 days at 22–25°C. Themiracidium that hatches out of the egg livesfor only about 8 h and can move in a film ofmoisture on damp pastures. For further
46 Chapter 1

development the free-living miracidiummust penetrate an amphibious snail, usuallya species of Lymnaea (mainly L. truncatulain Europe, L. tomentosa in north-westAfrica and L. viatrix in South America).
Once inside the snail, there is a sporo-cyst, followed by two redia stages; the cer-cariae, which emerge 30–40 days afterpenetration at 24°C, encyst as metacercariaeon vegetation (Fig. 23). This is usually wetgrass, but watercress, water mint (in Iran) orother salad vegetables are of most impor-tance in human infections (or perhapssometimes in drinking water). When cystsare ingested, the larvae excyst in the duode-num, migrate through the intestinal walland reach the bile-ducts by eating their waythrough the hepatic parenchyma from thesurface of the liver. The adult worms maturein 3–4 months and live for 5–12 years.
CLINICAL MANIFESTATIONS
There is an acute phase corresponding tothe period when the young flukes aremigrating through the liver tissues (4–8weeks). Symptoms of dyspepsia, nauseaand vomiting, abdominal pain localized tothe right hypochondrium or epigastriumand high fever (up to 40°C) ensue, oftenwith sudden onset. Allergic symptoms,such as pruritus and urticaria, can alsooccur. Anaemia is found in a proportion ofcases and the erythrocyte sedimentationrate is often raised. There is a leucocytosisin 50–100% of cases, with higheosinophilia (up to 60%).
The chronic phase ensues when theflukes reach the bile-ducts. There is oftenpainful liver enlargement and occasionallyan obstructive jaundice develops later;there may also be pains in the lower chest,with loss of weight. Symptoms are due toinflammation of the epithelial lining of theduct or to obstruction of its lumen and theyclosely resemble those of biliary disease,with obstructive jaundice and centrilobularcholestasis. Cholelithiasis can also occur asa complication. Many chronic cases areprobably asymptomatic and undiagnosed,so the proportion of such cases isunknown. Sometimes parasites in humansdevelop in ectopic sites in other organs or
in the wall of the peritoneal cavity, andcause granulomas.
The parasite is responsible for fewdeaths but in one case 40 flukes were foundto be blocking the bile-ducts at autopsy.
PATHOGENESIS
Most of the knowledge on pathology isbased on studies in animals. In sheep, thetracts of the migrating flukes show a coagu-lation necrosis of the liver cords, withnumerous neutrophils, lymphocytes anderythrocytes. The traumatic damage causedis roughly proportional to the number ofparasites present and, in sheep, death isusually caused by the presence of over 600worms. The mature flukes in the bile-ductscause a secondary adenomatous hyper-plasia and desquamation of the liningepithelium of the bile-ducts (whichbecomes over 2.0 mm thick) with exagger-ated Aschoff–Rokitansky crypts. Theplasma cells, lymphocytes and eosinophilsin the periductal tissues are replaced by afibrotic reaction leading to great thickeningof the walls and a much reduced lumen(Plate 7). Bile-duct hyperplasia and fibrosisof portal tracts has been seen in biopsyspecimens in human infections.
The immunology of liver-fluke infec-tions in general is discussed on p. 41.
The Trematodes 47
Fig. 23. Metacercariae of Fasciola hepatica onblades of grass (actual size 0.3 mm).

DIAGNOSIS
Clinical. Clinical signs are often variable buta triad of fever, liver enlargement and higheosinophilia is very suggestive, particularlyif combined with a history of having eatenhome-grown watercress a few weeksbefore.
CT and ultrasound can be of use in visu-alizing worms in the bile-ducts.
Parasitological. Eggs in the faeces are oftenscanty and duodenal or bile-duct aspiratescan be used. Eggs are large, measuring130–150 µm � 60–85 µm, and ovoid, withan inconspicuous operculum (Fig. 123).Faeces concentration techniques can beemployed, but in about 30% of humancases no eggs can be found in faeces orbile. Spurious infections from ingestion ofinfected liver can be ruled out by keepingthe patient on a liver-free diet for a week.
Immunological. An intradermal test hasbeen used for many years but is not veryspecific and only becomes positive after afew weeks. Passive haemagglutination isanother test that is well established. In thelast few years there have been many stud-ies utilizing ELISAs. For instance an IgG4ELISA, using either recombinant or nativeprotein (cathepsin L1 cysteine proteinase)as antigen, gives very sensitive results withfinger-prick blood (O’Neill et al., 1999). Anantigen-detection test using sandwichELISA detected infection by 1 week, withan optimum at 3 weeks. Westernimmunoblots are useful for confirmation ofinfection. Tests for detection of Fasciolacoproantigens also show great promise.These recent immunological tests can beused for detecting early infections and alsofor assessing the success of a cure.
TREATMENT
Praziquantel, which has been so effectiveagainst almost all other trematode infec-tions, has little effect in fascioliasis. Thestandard treatment for many years waswith bithionol (30–50 mg kg�1 given orallyon alternate days for 10–15 days or as asingle dose of 45 g for an adult and half
this for a child) and this is still used.Recently triclabendazole at 10 mg kg�1 for1 or 2 days has given cure rates of 80–90%and is now the drug of choice.
EPIDEMIOLOGY AND ZOONOTIC ASPECTS
There are occasional human cases reportedfrom many countries (61 in total) followingingestion of infected watercress or lettuceand epidemics may occur locally in partic-ularly wet years (e.g. in France a total of10,000 cases were recorded between 1956and 1982, principally in two wet and warmperiods). Infection is spreading in manyparts of the world and it is clear that thereare some human endemic regions wherehuman-to-human transmission takes place(Mas Coma et al., 1999). In the Andean alti-plano areas of Bolivia there are prevalencesof 15–100% and in children in central Peruof 10–34% and probably many cases alsoat high altitudes in Ecuador. There are alsomuch higher infection rates in the Niledelta, in the Tadzhik Republic (Samarkand)and in the Caspian region of Iran than pre-viously thought. In these areas of highintensity, infections are more common inchildren (Dalton, 1998).
Infection occurs in up to 50% of sheepand 70% of cattle in many parts of theworld and is a major veterinary problem inmany countries, but flukes may also befound in goats, donkeys, pigs, camels,horses, rabbits and deer.
The eggs of Fasciola do not usuallydevelop below 10°C (or over 30°C) and inmany temperate countries they rest overthe winter on pastures, the miracidiahatching out in the spring. However, eggscan develop at lower temperatures in thehigh Andes foci. Studies of the tempera-ture, of developmental factors in the inter-mediate and definitive hosts and of rainfallhave enabled accurate forecasts of thelikely build-up of infection in domesticstock in countries in Europe (Fig. 24).
PREVENTION AND CONTROL
In commercial watercress beds there mustbe precautions to prevent pollution byfaeces of domestic stock.
48 Chapter 1

A vaccine for infections in cattle andsheep is being investigated (Dalton, 1998).
Fasciola gigantica Cobbold, 1856, is acommon parasite of cattle, camels andother herbivores in Africa, in Asia and onsome Pacific islands. The eggs are largerthan those of F. hepatica (160–190 µm �70–90 µm) and the adults are longer(25–75 mm � 12 mm), with more anteriortestes, a larger ventral sucker and not sopronounced ‘shoulders’. Sporadic caseshave been reported from humans in tropi-cal Africa, China, Egypt, Hawaii, Iran,Thailand, Turkey and Uzbekistan. The lifecycle and clinical effects are similar tothose of F. hepatica except that the snailhosts, such as Lymnaea natalensis inAfrica, are often aquatic, not amphibious.
Immunodiagnostic tests, such as SDS-PAGE using an ES antigen of 27 kDa or asomatic antigen of 38 kDa, or IgM or IgGELISA, appear to be as effective as for F.hepatica and do not cross-react with schisto-somiasis.
Fasciola indica Varma, 1953, is found inbuffaloes and other herbivorous mammalsin Asia, with a very few human casesreported from India and Korea. In Russia
Lymnaea truncatula acts as first intermedi-ate host for all three species.
Fasciolopsis buski (Lankester, 1857) Odhner, 1902
SYNONYMS
Distoma buski Lankester, 1857, Distomacrassum Busk, 1859.
DISEASE, POPULAR AND LOCAL NAMES
Fasciolopsiasis or fasciolopsiosis; Busk’sfluke infection; Pa-yard bai-mai nai lumsi(Thai).
GEOGRAPHICAL DISTRIBUTION
The largest endemic foci of infection are inthe Yangtze basin provinces of China butinfection also occurs in Bangladesh,Cambodia, India (particularly Assam, butalso in Bihar, Madras and Orissa), Indonesia,Laos, Taiwan and central Thailand.
LOCATION IN HOST
Adult flukes are found in the small intes-tine, attached by the ventral sucker. Inheavy infections they may also be presentin the stomach and colon.
The Trematodes 49
Fig. 24. Effect of weather on the life cycle of F. hepatica in western Europe.

MORPHOLOGY
Fasciolopsis is the largest human trema-tode, measuring on average 30 (20–70) mm� 12 (8–20) mm (Fig. 3). It is a thick,ovoid worm without a cephalic cone and,when living, is pinkish in colour. Thetegument is covered with transverse rowsof small spines, which are most numerousaround the ventral sucker. The ventralsucker is a short distance behind the oralsucker and is about four times as large.The testes are highly branched and lie onebehind the other in the posterior part ofthe body. The organs of the female repro-ductive system are also branched, as istypical of members of the familyFasciolidae but, unlike the liver flukeFasciola, the two gut caeca areunbranched. The egg is large, 130–140 µm� 80–85 µm (Fig. 123); the yellowish-brown shell is clear and thick and has asmall operculum. The egg is unembry-onated when laid and is almost identicalto that of Fasciola hepatica; they cansometimes be differentiated with difficultywhen living by the distribution of the lipidgranules in the yolk cells, which are uni-formly arranged in Fasciolopsis and con-centrated around the nucleus in Fasciola,by differences in the number of yolk cellsand by the size of the germinal area. Dailyegg production is about 25,000 per fluke.
LIFE CYCLE
Eggs passed in the faeces need to reachwater for further development and theytake 3–7 weeks at 27–32°C to embryonatefully (they can survive for 3–4 months at4°C but do not develop). The miracidiumthat hatches out enters a freshwater plan-orbid snail, such as Gyraulis chinensis,Hippeutis cantori or Polypylis (=Segmentina) hemisphaerula, in which asporocyst and two redial stages are found.The xiphidocercous cercariae, whichemerge from the second-generation rediae,leave the snail and swim in the water. Theyencyst on the roots or fruits of variousfreshwater plants, such as the water caltrop(Trapa bicornis and T. natans) or waterchestnut (Eliocharis tuberosa), to give anencysted metacercarial stage.
When ingested on edible plants, themetacercariae excyst in the duodenum andthe young flukes take 3–4 months tomature in the ileum. The adult worms canproduce up to 28,000 eggs 24 h�1 but livefor only about 6–12 months in humans.
CLINICAL MANIFESTATIONS
In the majority of cases infection is light(fewer than 20 worms or 5000 eggs g�1
faeces) and there are no symptoms. With aheavy worm burden there is nausea anddiarrhoea, accompanied by intense gripingpains in the morning which are relieved byfood. Oedema, including a characteristicfacial swelling more prominent in the peri-orbital region, is often found. In childrenwith more than 100 worms the abdomen isusually distended and there may be stunt-ing of growth. The presence of more than400 worms usually proves fatal to youngchildren.
In about 40% of cases there is aneosinophilia of up to 30%.
PATHOGENESIS
The adult worms cause traumatic inflam-matory damage to the mucosa at the site ofattachment, often leading to ulceration andhaemorrhage, and make the mucosa hyper-aemic. They also provoke an excessivesecretion of mucus. Toxic by-products pro-duced by the adults are said to cause thefacial oedema that is sometimes found andcan result in a toxic diarrhoea, but this maybe due to a protein-losing enteropathy.When many worms are present, there isalso the possibility of obstruction of thebowel, with resultant stasis.
DIAGNOSIS
Recovery of eggs in faeces is the onlymethod used in practice and various con-centration methods can be employed if nec-essary. The eggs can be easily differentiatedfrom those of Gastrodiscoides and are largerthan those of echinostomes but cannot usu-ally be distinguished from those of Fasciola.
TREATMENT
Praziquantel (15–60 mg kg�1) and albend-azole have both been used successfully,
50 Chapter 1

although the latter had some failures withchildren.
EPIDEMIOLOGY
There are often high prevalence rates inendemic areas: 39% in Dacca, Bangladesh,70% in Jiangxi, China, 70% in Maharashtraand 29% in Bombay, India, 27% in SouthKalimantan, Indonesia (57% in children),3.8% in Vientiane, Laos, and 17% inSupanburi, Thailand (Yu and Mott, 1994).
The seed pods of water caltrop (alsoknown as water ling) are probably the mostimportant source of human infection,because this plant is grown in ponds fertil-ized with night-soil, while the other plantsinvolved are usually grown in runningstreams. Before the nuts of the water cal-trop or the bulbs of water chestnut areeaten, the outer covering is removed withthe teeth and this provides the principalmode of infection. This habit is particu-larly common in children, who are usuallymore heavily infected than adults. Inmarkets caltrop nuts are kept fresh bysprinkling with water and thus the cystsare prevented from drying.
In Vietnam the roots of the water bam-boo (Zizania aquatica, Salvinia natans andLemna polyrhiza) and water hyacinth(Eichornia crassipes) are also eaten, and inThailand lotus (Nymphoea lotus), a water-cress (Neptunia cleracea) and water morn-ing glory (Ipomea aquatica).
In Taiwan and Thailand water plants arecollected and fed to pigs, which helps tomaintain a reservoir of infection. InThailand infection is confined to regionsgrowing water caltrops although the nutsare grown in many other non-endemicareas also; this is probably because cystsdry up during transport or because the nutsare only eaten cooked in these otherregions.
PREVENTION AND CONTROL
Water vegetables need to be carefullypeeled and washed. Boiling for 1–2 min issufficient to kill the metacercarial cysts. Insome areas free metacercarial cysts havebeen found floating on the water surfaceand may be ingested in drinking water.
Night-soil, as once commonly used inChina, should be stored before being addedas fertilizer to submerged fields. Sanitarydisposal of faeces would probably be effec-tive, although there is still the possibilityof zoonotic infections. Molluscicides couldpossibly be used for snail control but theirefficacy has not been tested.
ZOONOTIC ASPECTS
Pigs act as important reservoir hosts. Theendemic region in central Thailand is aperennial flood basin where rice is grownand houses are built on stilts. Pigs are alsokept in pens on stilts and fed on waterplants; the faeces of the pigs passes into thewater below, in which snails live and foodplants are grown.
There are high prevalence rates in pigsin areas of Bangladesh, Burma (Myanmar),Cambodia and India but few human infec-tions, as the appropriate plants are noteaten raw.
Family Heterophyidae
Heterophyes heterophyes(von Siebold, 1852) Stiles and Hassall, 1900
SYNONYMS
Distoma heterophyes von Siebold, 1852;Heterophyes aegyptiaca Cobbold, 1866;Heterophyes nocens Onji and Nishio, 1915.
DISEASE AND POPULAR NAMES
Heterophyiasis, heterophyosis; von Siebold’sfluke infection, dwarf-fluke infection.
GEOGRAPHICAL DISTRIBUTION
Asia (southern China, Japan, Indonesia,Korea, the Philippines, Taiwan), NorthAfrica and Middle East (Egypt, Iran, Sudan,Tunisia, Turkey) and southern Europe(Greece and Romania).
LOCATION IN HOST
Adults are attached to the wall of thejejunum and upper ileum, often in thecrypts of Lieberkühn (Fig. 25).
The Trematodes 51
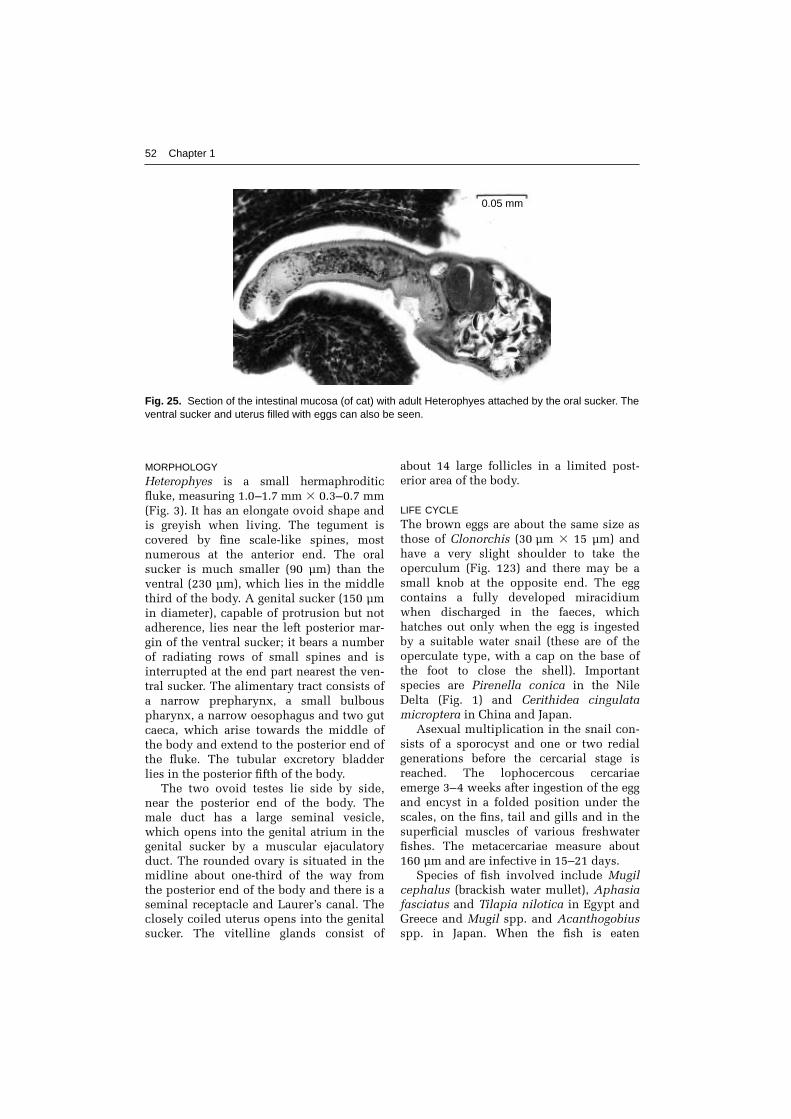
MORPHOLOGY
Heterophyes is a small hermaphroditicfluke, measuring 1.0–1.7 mm � 0.3–0.7 mm(Fig. 3). It has an elongate ovoid shape andis greyish when living. The tegument iscovered by fine scale-like spines, mostnumerous at the anterior end. The oralsucker is much smaller (90 µm) than theventral (230 µm), which lies in the middlethird of the body. A genital sucker (150 µmin diameter), capable of protrusion but notadherence, lies near the left posterior mar-gin of the ventral sucker; it bears a numberof radiating rows of small spines and isinterrupted at the end part nearest the ven-tral sucker. The alimentary tract consists ofa narrow prepharynx, a small bulbouspharynx, a narrow oesophagus and two gutcaeca, which arise towards the middle ofthe body and extend to the posterior end ofthe fluke. The tubular excretory bladderlies in the posterior fifth of the body.
The two ovoid testes lie side by side,near the posterior end of the body. Themale duct has a large seminal vesicle,which opens into the genital atrium in thegenital sucker by a muscular ejaculatoryduct. The rounded ovary is situated in themidline about one-third of the way fromthe posterior end of the body and there is aseminal receptacle and Laurer’s canal. Theclosely coiled uterus opens into the genitalsucker. The vitelline glands consist of
about 14 large follicles in a limited post-erior area of the body.
LIFE CYCLE
The brown eggs are about the same size asthose of Clonorchis (30 µm � 15 µm) andhave a very slight shoulder to take theoperculum (Fig. 123) and there may be asmall knob at the opposite end. The eggcontains a fully developed miracidiumwhen discharged in the faeces, whichhatches out only when the egg is ingestedby a suitable water snail (these are of theoperculate type, with a cap on the base ofthe foot to close the shell). Importantspecies are Pirenella conica in the NileDelta (Fig. 1) and Cerithidea cingulatamicroptera in China and Japan.
Asexual multiplication in the snail con-sists of a sporocyst and one or two redialgenerations before the cercarial stage isreached. The lophocercous cercariaeemerge 3–4 weeks after ingestion of the eggand encyst in a folded position under thescales, on the fins, tail and gills and in thesuperficial muscles of various freshwaterfishes. The metacercariae measure about160 µm and are infective in 15–21 days.
Species of fish involved include Mugilcephalus (brackish water mullet), Aphasiafasciatus and Tilapia nilotica in Egypt andGreece and Mugil spp. and Acanthogobiusspp. in Japan. When the fish is eaten
52 Chapter 1
0.05 mm
Fig. 25. Section of the intestinal mucosa (of cat) with adult Heterophyes attached by the oral sucker. Theventral sucker and uterus filled with eggs can also be seen.

uncooked the metacercariae escape anddevelop in the small intestine. The flukesmature in 15–20 days and the adults livefor about 2 months.
CLINICAL MANIFESTATIONS
In light infections there are usually nosymptoms. In heavy infections, nausea andintermittent chronic diarrhoea, sometimeswith the passage of blood and abdominaldiscomfort with colicky pains and tender-ness, have been reported beginning 9 daysafter ingestion of cysts. Symptoms resem-ble those of amoebiasis or heartburn andthere may be vomiting, simulating a pepticulcer. Slight eosinophilia is sometimesfound.
PATHOGENESIS
A mild inflammation of the mucosa at thesite of attachment and sometimes a superfi-cial necrosis occur if there are thousands ofworms present; up to 4000 have beenreported.
Eggs of many species of heterophyidshave been found at necropsy from varioussites in the body, either because the eggsthemselves have succeeded in filteringthrough the intestinal wall to the mesen-teric lymphatics or venules or because theadults have penetrated the gut wall andlaid eggs in the peritoneal cavity. Thisphenomenon appears to be a reaction tohumans as abnormal hosts, and emboliza-tion of the eggs may occur. Cardiac lesionscaused by Heterophyes or other membersof the same family include fibrosis sur-rounding eggs in the mitral valve andmyocardium, with consequent heart fail-ure. Eggs have been found in the brain on afew occasions, as have encapsulatedadults.
DIAGNOSIS
This is by recovery of eggs following sug-gestive clinical findings, either by directfaecal smears or by concentration methods.The eggs can be differentiated from thoseof opisthorchids by the less well-developedshoulders at the operculum giving a moreoval outline and by the smaller posterior
knob. The contained miracidium also hasbilaterally symmetrical penetration glands(Fig. 123).
TREATMENT
Niclosamide (2 g day�1 for 2 days for anadult) has been used for many years butmore recently praziquantel (a single doseof 20 mg kg�1) has become the drug ofchoice.
EPIDEMIOLOGY
Transmission is by eating fresh or salteduncooked fish. There may be heavy infec-tions in both rural and urban communitiesin the Nile Delta region of Egypt, who tra-ditionally eat pickled mullet (‘fessik’) atthe feast of Sham-el Nessim. An infectionrate of 10% has been reported from school-children in the area. However, most infec-tions are in 20–40-year-olds, who eatfreshly salted mullet; metacercariae canlive for up to 7 days in these.
PREVENTION AND CONTROL
Personal prevention is by thorough cookingof fish. Partial cooking (mostly of Tilapianilotica) carried out around Lakes Manzala,Borollos and Edco in the Nile Deltadestroys only about 70% of cysts.Preventing pollution of the water withfaeces, which in Egypt is mainly by fisher-men, is a possible long-term objective.
ZOONOTIC ASPECTS
Dogs, cats, foxes and other fish-eatingmammals act as reservoir hosts and canmaintain the infection in the absence ofhumans.
Metagonimus yokogawai Katsurada, 1912
SYNONYM
Heterophyes yokogawai Katsurada, 1912.
DISEASE AND POPULAR NAME
Metagonimiasis or metagonimosis;Yokogawa’s fluke infection.
The Trematodes 53

GEOGRAPHICAL DISTRIBUTION
This is probably the most common humanintestinal fluke in the Far East and isendemic in China, Indonesia, southernJapan, Korea, Manchuria, Taiwan andSiberia. It has also been reported fromIsrael, Serbia and Spain.
LOCATION IN HOST
Adults are found in the upper and middleparts of the jejunum, rarely in the duode-num, ileum and caecum. The flukes areembedded in the crypts of Lieberkühn inthe early stages or in the mucus found onthe surface.
MORPHOLOGY
This genus is closely related toHeterophyes, the adults being similar insize and structure. The principal differ-ences are that in Metagonimus the ventralsucker is situated to the right of the midlineand there is no accessory sucker (Fig. 3).
LIFE CYCLE
The eggs measure 27 µm � 16 µm and arevery similar to those of Heterophyes (Fig.123). The life cycle is also similar. InChina, Japan and Korea the snail speciesinvolved is Semisulcospira libertina. Thesecond intermediate fish hosts arePlecoglossus altivelis (a trout, the ‘ayu’ orsweetfish), Leuciscus hakuensis, Mugilcephalus, Odontobutis obscurus and Salmoperryi in Japan and Siberia and alsoPseudorasbora parva and Carrasius carra-sius (silver carp) in Korea. Each adult flukeproduces about 1500 eggs 24 h�1.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Similar to Heterophyes, with abdominalpain, diarrhoea and lethargy. A heavyinfection in Japan caused intractable diar-rhoea and weight loss and from the faecalegg count it was estimated that50,000–70,000 adult worms were present(63,587 have been recovered from onepatient who had only slight symptoms(Chai and Lee, 1990)). Headache, abdomi-nal and muscular pains, fever and urticariahave also been reported.
Pathological changes are characterizedby villous atrophy and hyperplasia of thecrypts, with inflammatory cell infiltration.Eggs may be carried by intestinal capillar-ies and lymphatics to the myocardium,brain, spinal cord and other tissues, wheregranulomatous changes or embolisms canbe provoked by their presence.
DIAGNOSIS AND TREATMENT
As for Heterophyes. The eggs can, with dif-ficulty, be differentiated from those ofHeterophyes, as the latter have a thinnershell and are light brown in colour.
EPIDEMIOLOGY
Since the eggs of this species are so similarto those of many other parasites in this fam-ily that can infect humans, it is difficult toknow the true prevalence. In ethnic minori-ties in Khabarovsk Territory of Siberia,there are high rates of 20–70% and, inendemic areas of coastal Korea, thereappear to be 10–20% of villagers infected.
In Japan the cercariae do not emergefrom the snail intermediate hosts below18–20°C and infection does not occur inareas where summer temperatures arebelow this figure.
ZOONOTIC ASPECTS
Dogs, cats, rats and pigs (and perhaps peli-cans and other fish-eating birds) act asreservoir hosts and infection in animals iscommon worldwide. Potentially all speciesof the family Heterophyidae could becomehuman intestinal parasites where raw fishis eaten and it is likely that many otherswill be recorded as occasional human para-sites (particularly in the Far East) whenexpelled adult worms are examined. Thefollowing species have been recorded fromhumans and, in some, eggs or occasionallyadults have been found in the heart, brainor spinal cord: Apophallus donicus (east-ern Europe, Canada and USA),Centrocestus armatus (Korea), C. caninus(Taiwan), C. cuspidatus (Egypt andTaiwan), C. formosanus (China and Japan),C. kurokawai (Japan), C. longus (Taiwan),Cryptocotyle lingua (Greenland),
54 Chapter 1

Diorchitrema amplicaecale (Taiwan), D.formosanum (Taiwan), D. pseudocirratum(Hawaii and Philippines), Haplorchismicrorchis (Japan), H. pleurolophocerca(Egypt), H. pumilio (Egypt, Iran and FarEast), H. taichui (Bangladesh, Egypt, Iranand Far East), H. vanissimus (Philippines)and H. yokogawai (Far East and Egypt),Heterophyes dispar (Korea and Thailand),H. equalis (Egypt), H. heterophyes nocens(Korea and Japan), H. katsuradai (Japan),Heterophyopsis continua (Far East),Metagonimus minutis (China and Taiwan),M. takahashi (Japan and Korea), Phagicolalonga (Brazil and Mexico), P. ornamenta, P.ornata and Pharyngostomum flapi (Egypt),Procerovum calderoni (China, Philippinesand Egypt), P. varium (Japan), Pygidiopsissumma (Japan and Korea), Stellantchasmusamplicaecalis (Egypt), S. falcatus (Korea,Laos, Thailand and Hawaii), Stictodora fus-cata and S. manilensis (Korea). See Coombsand Crompton (1991), Yu and Mott (1994),WHO (1995), Ashford and Crewe (1998)and Muller (2001), and for details of inter-mediate hosts, rarity and original refer-ences.
Family Paramphistomidae
Gastrodiscoides hominis(Lewis and McConnell, 1876) Leiper, 1913.
SYNONYMS AND DISEASE
Gastrodiscoidiasis or gastrodiscoidosis;Amphistomum hominis Lewis andMcConnell, 1876, Gastrodiscus hominisFischoeder, 1902.
GEOGRAPHICAL DISTRIBUTION
Infection is most common in India, particu-larly Assam, Bihar and Orissa, but alsooccurs in Burma (Myanmar), China,Kazakhstan, the Philippines, Thailand andVietnam.
LOCATION IN HOST
Adults attach to the wall of the caecum andascending colon by their suckers.
MORPHOLOGY
Gastrodiscoides is the only human-para-sitic member of the paramphistomes,which are important parasites of domesticherbivores. It is a dorsally convex flukemeasuring 5–8 mm � 3–5 mm and, whenliving, is reddish in colour. The body isdisc-shaped with an anterior conical pro-jection bearing the oral sucker (Fig. 3). Likeall other members of the family, the ventralsucker is close to the posterior end. Thetegument is smooth. The alimentary tractconsists of a pair of lateral pouches arisingfrom the oral sucker and a slightly tortuouspharyngeal tube, which bifurcates into twogut caeca that extend only to the middle ofthe discoid region. The large excretorybladder is in the midline behind the ven-tral sucker. The two lobate testes lie in tan-dem just behind the bifurcation of thecaeca. The common genital opening is on acone just anterior to the bifurcation. Theovoid ovary lies just posterior to the testesnear the midline. The vitelline glands arescattered and surround the caeca behindthe testes. The loosely coiled uterus openson the genital cone.
LIFE CYCLE
The eggs measure 146 (127–160) µm � 66(62–75) µm. They are rhomboidal in shape,have a small well-fitting operculum and aretransparently greenish in colour (Fig. 123).They contain about 24 vitelline cells and acentral unembryonated ovum. After theeggs are expelled in the faeces and reachwater, a miracidium develops in 9–14 daysat 24°C. The miracidium that emerges fromthe egg can develop experimentally in thesnail Helicorbis coenosus found in UttarPradesh and cercariae emerge 28–152 daysafter infection. The cercariae form meta-cercarial cysts on water plants; probablythe water caltrop is involved in humaninfection but the life cycle has not beenelucidated in nature.
ZOONOTIC ASPECTS
Pigs, monkeys (rhesus), apes (orang-utan),field rats and the Napu mouse deer arenatural hosts, but it is not known what the
The Trematodes 55

relationship is between human and animalhosts.
Watsonius watsoni (Conyngham, 1904)Stiles and Goldberg, 1910 (synonymAmphistomum watsoni, Conyngham,1904), has been reported once from north-ern Nigeria in a patient who died of severediarrhoea. Many worms were seen in thestools and after autopsy many more werefound attached to the wall of the duode-num and jejunum and a few in the colon.Presumably infection was contracted fromingestion of metacercarial cysts on vegeta-tion. Watsonius is a natural parasite ofmonkeys in West Africa and Singapore.
Fischoederius elongatus (Poirer, 1883)Stiles and Goldberger, 1910, is a parasite ofruminants in Asia, contracted from ingest-ing cysts on vegetation. Human infectionhas been reported from China.
Family Echinostomidae
Most of the members of this large andrather heterogeneous group are parasites ofthe gut of birds and lower vertebrates, but afew are found in mammals and at least 24species are occasional parasites of humans.Typically, members have a collar of spinesbehind the oral sucker.
Echinostoma ilocanum(Garrison, 1908) Odhner, 1911
SYNONYMS
Fascioletta ilocanum Garrison, 1908;Euparyphium ilocanum Tubangui andPasco, 1933.
DISEASE AND POPULAR NAME
Echinostomiasis or echinostomosis;Garrison’s fluke infection.
GEOGRAPHICAL DISTRIBUTION
Five human cases were found by Garrisonin prisoners in Manila jail in 1907 and
occasional cases have been reported sincefrom China (Yunnan), Indonesia, thePhilippines (11–44% in northern Luzon),Thailand (8.2% in one area) and recentlyIndia (Grover et al., 1998).
LOCATION IN HOST
Adults live in the jejunum; they attach byinsertion of the oral end into the mucosa.
LIFE CYCLE
This is a natural parasite of dogs and cats,and eggs (measuring 100 µm � 65–70 µm)in the faeces are undifferentiated. If theyreach water, the contained miracidiadevelop in about 8–14 days at 26°C. Theyemerge and penetrate into suitable fresh-water snails (particularly Hippeutis) andmultiply asexually. Cercariae that emergethen enter a wide range of other snails(such as Pila) and encyst as metacercariae.When these snails are eaten raw by dogs,cats or humans, they excyst and developinto adult worms. Other echinostomesutilize fish and amphibians (usually frogs)as second intermediate hosts, which arealso eaten raw. It appears that cercariae canalso be swallowed while swimming andcause infection.
Other Echinostomids
E. echinatum (= lindoense) (Zeder, 1803)formerly had a high infection rate(24–96%) around Lake Lindu in centralSulawesi but has been very uncommonsince the 1970s because of changes in eat-ing habits consequent on the introductionof tilapia into the lake. The first intermedi-ate host is a planorbid snail or a type ofmussel (Corbicula) and the metacercariaeencyst in edible pulmonate and bivalvesnails, which used to be eaten raw and arenow rare (partly because they are eaten bythe fish).
E. hortense Asada, 1926, has an infectionrate of up to 22.4% in southern Korea andis not uncommon in north-eastern China,where loach (the usual second intermediate
56 Chapter 1

host) are eaten raw as a cure for jaundice; ithas also been reported from Japan. The nat-ural definitive hosts are rats and dogs.
E. malayanum Leiper, 1911, has beenrecovered from humans in India, Indonesia(Irian Jaya and Sumatra), the Philippines(20% in areas of Luzon) and north-eastThailand (3–8%). It is probably a naturalparasite of dogs or rats and human infec-tion is through ingestion of edible molluscs(particularly Pila and Vivipara), which actas second intermediate hosts.
E. revolutum (Froelich, 1802) Looss, 1899,is a common parasite of the caeca of duck,goose and muskrat, and most human casescome from Taiwan (2.8–6.5% infection)and north-east Thailand, with reports fromChina and Indonesia. Edible molluscs,such as Corbicula, act as second intermedi-ate hosts.
Hypoderaeum conoideum (Bloch, 1872)Dietz, 1909, is a parasite of birds, but therecan be up to 55% infection rates in nort-east Thailand. Snails, such as Planorbisand Lymnaea spp., act as both first andsecond intermediate hosts.
There are many species of echinostomesthat can act as occasional or rare parasitesof humans, the numbers increasing as theuse of praziquantel allows voided adultworms to be examined (for further informa-tion, see Haseeb and Eveland, 2000):Artyfechinostomum malayanum (Thailand),A. mehrai (India), A. oraoni (India), A. (=Paryphostomum) sufrartyfex (four cases inIndia), Echinochasmus fujianensis (up to7.8% in southern Fujian, China), E. japoni-cus (China, Japan, Korea and Taiwan), E.jiufoensis (one fatal case in Canton, China),E. liliputanus (up to 23% in areas of Anhuiprovince, China), E. perfoliatus (Japan,Taiwan and Europe), Echinoparyphiumparaulum (Russia), E. recurvatum (Egypt,Indonesia and Taiwan), Echinostoma (=Echinochasmus) angustitestis (two cases inChina), E. cinetorchis (Japan, Korea andTaiwan), E. japonicum (Japan and Korea),E. macrorchis (Indonesia, Japan, Korea and
Taiwan), Echinostoma melis (= Euparyph-ium jassyense) (China, Romania andTaiwan), Episthmium caninum (Thailand)and Himasthala muehlensi (one case inColombia or USA).
MORPHOLOGY
These species of echinostomes are mostlysmall flukes measuring 2.5–8.0 mm and allhave a collar of spines behind the oralsucker (Fig. 3).
CLINICAL MANIFESTATIONS
In most infections there are no signs apartfrom the presence of eggs in the faeces. Inheavy infections (e.g. 14,000 worms wererecovered from one child at autopsy),symptoms are similar to those ofFasciolopsis buski, with vague abdominalmanifestations, such as flatulence, intesti-nal colic and diarrhoea (Graczyk and Fried,1998). Vomiting, diarrhoea, abdominalpain and swelling of the feet were reportedin one case in a child (Grover et al., 1998)and there may be anaemia and oedema.There have been fatalities in children fromdehydration. The adult worms attach to themucosa of the small intestine and may pro-duce inflammatory lesions and necrosis atthe site of attachment, with increased cel-lular infiltration in the intestinal mucosa.There may also be a generalized toxicprocess. In an experimental infection with113 metacercariae of Echinochasmusjaponicus, abdominal pain, diarrhoea andintestinal gurgling developed about 10days after infection (Lin et al., 1985).
DIAGNOSIS
Eggs found in the faeces are similar tothose of Fasciolopsis or Fasciola, althoughsomewhat smaller (most are about 100 µm� 70 µm); specific identification is almostimpossible without recovery of adults aftertreatment.
TREATMENT
Cure is simple with a single oral dose of10–20 mg kg�1 of praziquantel (or 5 mgkg�1 for Echinochasmus fujianensis).
The Trematodes 57

Other Occasional and Rare Human-parasitic Trematodes
Family Diplostomidae
Alaria americana Hall and Wigdor, 1918,and A. marcianae (La Rue, 1917) Walton,1950, are very similar parasites of wildcarnivores and dogs and cats in NorthAmerica, in which the anterior portion ofthe body is spoon-shaped, with an organthat secretes proteolytic enzymes. Theyhave an unusual life cycle. When eggs fromfaeces reach water, the miracidia hatch outand penetrate and multiply in suitablespecies of snails (including Heliosomaspp.). The emerging cercariae develop inamphibian tadpoles into a larval stageknown as a mesocercaria (midway betweena cercaria and a metacercaria). Frogs andsnakes ingest tadpoles and act as paratenichosts. When these are eaten by youngcarnivores, metacercariae develop in thelungs, are coughed up and become adultsin the small intestine in about 20 days.However, in pregnant animals the mesocer-cariae migrate to the mammary glands andinfect the suckling young through the milk,and adults develop in these (known as‘amphiparatenesis’). They can also infectsubsequent litters (Shoop, 1994). A fatalcase in Canada occurred in a man whoingested frogs’ legs while hiking. He wasthought to be suffering from gastric flu butdied 9 days later from extensive pulmonaryhaemorrhage and at autopsy was found tohave thousands of mesocercariae in manytissues. In other cases mesocercariae havebeen found in the eye (Smyth, 1995).Another species in carnivores, A. alata(Goeze, 1782), has occasionally infectedhumans with mesocercariae in the MiddleEast. Neodiplostomum (= Fibricola)seoulensis (Seo, Rim and Lee, 1964) Hongand Shoop, 1994, a parasite of rats, hasdeveloped to adulthood in the humansmall intestine in a few cases in SouthKorea and is contracted from eating rawamphibians or snakes (paratenic hosts).Diplostomum spathaceum (Rudolphi,1819) Olsson, 1876, is a cosmopolitan para-
site of birds, with metacercariae in the eyesof fish, and there have been a few cases ofswimmer’s itch in the former Czecho-slovakia, but more importantly cercariaehave entered the eye and caused cataractsin the lens.
Family Lecithodendriidae
Phaneropsolus bonnei Lie Kian Joe, 1951,and Prosthodendrium molenkampi Lie KianJoe, 1951, were both discovered in Indonesiaat autopsy and are also present in rural pop-ulations of north-east Thailand and Laos.Eggs of both species are similar to those of Opisthorchis and heterophyids, andinsects act as second intermediate hosts.Phaneropsolus spinicirrus, Paralecithoden-drium obtusum and P. glandulosum arethree other poorly characterized speciescommonly found in Thais (Yu and Mott,1994).
Family Plagiorchiidae
Plagiorchis harinasutai Radomyos, Bunnagand Harinasuta, 1989, P. javensisSandground, 1940, P. muris Tanabe, 1922,and P. philippinensis Sandground, 1940,have insects as second intermediate hostsand have been found in humans inThailand, Indonesia, Japan and thePhilippines, respectively.
Family Troglotrematidae
Nanophyetus salmincola Chapin, 1927, isan intestinal parasite of wild carnivores,birds and domestic dogs in the Pacificnorth of the USA and in Siberia.Metacercariae encyst in salmonid fishesand in the USA, in dogs that eat the fish.Adults cause a haemorrhagic enteritis, withhigh fever, anorexia, vomiting and lymph-adenopathy (‘salmon poisoning’), with ahigh mortality 10–14 days after infection,caused by a rickettsia (Neorickettsiahelminthoeca), which is transmitted by the
58 Chapter 1

trematode. Following ingestion of raw fish,some patients in both areas suffered fromdiarrhoea, weight loss, nausea, vomiting,fatigue, anorexia and a high eosinophilia ofup to 43%; in one case there was also ahigh fever but the possible role of the rick-ettsia was not investigated (Eastburn et al.,1987). Symptoms usually resolved after afew months. The parasites have been sepa-rated into two subspecies: N. s. salmincolain the USA and N. s. schikhobalowi inSiberia. Diagnosis is by finding eggs in fae-ces (measuring about 80 µm � 45 µm) andtreatment is with praziquantel. Preventionis by not eating raw or smoked salmon inendemic areas, although there has been onecase of infection through handling fish.There is a 4.2% infection rate in a focus onthe lower Amur River in Siberia (and up to60% in local ethnic minorities), and theranges of both subspecies appear to bespreading.
Other families
Species of trematodes belonging to variousother families that have occasionally beenfound in humans include: Carneophallus(= Spelotrema) brevicaeca (from shrimpsand has caused fatalities in thePhilippines), Cathaemasia cabreri(Philippines), Clinostomum complanatum(from fish in Japan and Middle East),Cotylurus japonicus (China), Gymnoph-alloides seoi (from oysters in a village inKorea), Isoparorchis hypselobagri (China,India), Microphallus minus (Japan),Paryphostomum bangkokensis (Thailand),P. sufrartyfex (India), Philophthalmuslacrymosus (in the eye in Serbia),Philophthalmus sp. (eye in Sri Lanka),Prohemistomum vivax (from fish in Egypt)and Psilorchis hominis (Japan). See Yu andMott (1994), WHO (1995), Ashford andCrewe (1998) and Muller (2000) for furtherdetails of intermediate and definitive hosts.
The Trematodes 59
References
Abel, L. and Dessein, A.J. (1997) The impact of host genetics on susceptibility to human infectiousdiseases. Current Opinion in Immunology 9, 509–516.
Agnew, A., Fulford, A.J.C., Mwanje, M.T., Gachuhi, K., Gutsam, V., Krijger, F.W., Sturrock, R.F.,Vennerwald, B.J., Ouma, J.H., Butterworth, A.E. and Deelder, A.M. (1996) Age dependent reduc-tion of schistosome fecundity in S. haematobium but not S. mansoni infections in humans.American Journal of Tropical Medicine and Hygiene 55, 338–343.
Ansell, J. and Guyatt, H. (1999) Self-reporting of blood in urine by schoolchildren may be the mostcost-effective diagnostic test for urinary schistosomiasis. Bulletin of Tropical Medicine andInternational Health 7, 5.
Ashford, R.W. and Crewe, W. (1998) The Parasites of Homo sapiens. Liverpool School of TropicalMedicine, Liverpool, UK.
Azizova, O.M., Sagieva, A.T., Israilova, S., Sadykov, K.M., Shirinova, N.Sh., Mukhidinov, Sh.M.,Ismatov, I., Adilova, N.B. and Saidaliev, T.S. (1988) Dicrocoelium lanceolatum infection in man(on autopsy data) [English summary]. Meditsinskaya Parazitologiya i Parazitarnye Bolezni 2,26–28.
Blair, D., Xu, Z.-B. and Agatsuma, T. (1999) Paragonimiasis and the genus Paragonimus. Advances inParasitology 42, 113–222.
Boisier, P., Ramarokoto, C.E., Ravaoalimalala, V.E., Rabrijaona, L., Serieye, J., Roux, J. and Esterre, P.(1998) Reversibility of Schistosoma mansoni-associated morbidity after yearly mass praziquan-tel therapy: ultrasonographic assessment. Transactions of the Royal Society of TropicalMedicine and Hygiene 92, 451–453.
Brindley, P.J. (1994) Relationships between chemotherapy and immunity in schistosomiasis.Advances in Parasitology 34, 134–161.
Brooks, D.R., O’Grady, R.T. and Glen, D.R. (1985) Phylogenetic analysis of the Digenea(Platyhelminthes: Cercomeria) with comments on their adaptive radiation. Canadian Journal ofZoology 63, 411–443.

Brown, D.S. (1980) Freshwater Snails of Africa and Their Medical Importance. Taylor and Francis,London, UK.
Bundy, D., Sher, A. and Michael, E. (2000) Helminth infections may increase susceptibility to TB andAIDS. Parasitology Today 16, 273–274.
Butterworth, A.E. (1998) Immunological aspects of human schistosomiasis. British Medical Journal54, 357–368.
Capron, A. (1998) Schistosomiasis: 40 years’ war on the worm. Parasitology Today 14, 379–384.Chai, J.Y. and Lee, S.H. (1990) Intestinal trematodes of humans in Korea: Metagonimus, heterophyids
and echinostomes. Korean Journal of Parasitology 28 (suppl.), 103–122.Chen, M.G., Lu, Y., Hua, X.J. and Mott, K.E. (1994) Progress in morbidity due to Clonorchis sinensis
infection: a review of recent literature. Tropical Diseases Bulletin 91, R7–R65.Chitsulo, L., Engels, D., Montresor, A. and Savioli, L. (2000) The global status of schistosomiasis and
its control. Acta Tropica 77, 41–51.Coombs, I. and Crompton, D.W.T. (1991) A Guide to Human Helminths. Taylor and Francis, London,
UK.Coulson, P.S. (1997) The radiation-attenuated vaccine against schistosomiasis in animal models: par-
adigm of a human vaccine? Advances in Parasitology 39, 272–336.Dalton, J.P. (ed.) (1998) Fasciolosis. CAB International, Wallingford, UK.Damian, R.T. (1987) The exploitation of host immune responses by parasites. Journal of Parasitology
73, 1–11. Dekumyoy, P., Waikagul, J. and Eom, K.S. (1998) Human lung fluke Paragonimus heterotremus: dif-
ferential diagnosis between Paragonimus heterotremus and Paragonimus westermani infectionsby EITB. Tropical Medicine and International Health 3, 52–56.
Doenhoff, M.J., Hassounah, O., Murare, H., Bain, J. and Lucas, S. (1986) The schistosome egg granu-loma: immunopathology in the cause of host protection or parasite survival? Transactions of theRoyal Society of Tropical Medicine and Hygiene 80, 503–514.
Dupre, L., Herve, M., Schacht, A.M., Capron, A. and Riveau, G. (1999) Control of schistosomiasispathology by combination of Sj28GST DNA immunization and praziquantel treatment. Journalof Infectious Diseases 180, 454–467.
Eastburn, R.I., Fritsche, T.R. and Terhune, C.A., Jr (1987) Human intestinal infection withNanophyetus salmincola from salmonid fishes. American Journal of Tropical Medicine andHygiene 36, 586–591.
Elkins, D.B., Haswell-Elkins, M.R., Mairang, E., Mairang, P., Sithithaworn, P., Kaewkes, S.,Bhudhisawasdi, V. and Uttaravichien, T. (1990) A high frequency of hepatobiliary disease andsuspected cholangiocarcinoma associated with heavy Opisthorchis viverrini infection in a smallcommunity in north-east Thailand. Transactions of the Royal Society of Tropical Medicine andHygiene 84, 715–719.
Frenzel, K., Grigull, L., Odongo-Aginya, E., Ndugwa, C.M., Loroni-Lakwo, T., Schweigmann, U.,Verster, U., Spannbrucker, N. and Doehring, E. (1999) Evidence for long-term effect of a singledose of praziquantel on Schistosoma mansoni-induced hepatosplenic lesions in northernUganda. American Journal of Tropical Medicine and Hygiene 60, 927–931.
Giboda, M., Malek, E.A. and Correa, R. (1997) Human schistosomiasis in Puerto Rico: reduced preva-lence rate and absence of Biomphalaria glabrata. American Journal of Tropical Medicine andHygiene 57, 564–568.
Gibson, D.I. and Bray, R.A. (1994) The evolutionary expansion and host–parasite relationships in theDigenea. International Journal for Parasitology 24, 1213–1226.
Gobert, G.N. (1998) The role of microscopy in the investigation of paramyosin as a vaccine candidateagainst Schistosoma japonicum. Parasitology Today 14, 115–118.
Graczyk, T.K. and Fried, B. (1998) Echinostomiasis: a common but forgotten food-borne disease.American Journal of Tropical Medicine and Hygiene 58, 501–504.
Greer, G.J., Kitikoon, V. and Lohachit, C. (1989) Morphology and life cycle of Schistosoma sinensiumPao, 1959, from northwest Thailand. Journal of Parasitology 75, 98–101.
Grover, M., Dutta, R., Kumar, R., Aneja, S. and Mehta, G. (1998) Echinostoma ilocanum infection.Indian Pediatrics 35, 549–552.
Guyatt, H., Brooker, S. and Donnelly, C.A. (1999) Can prevalence of infection in school-aged childrenbe used as an index for assessing community prevalence? Parasitology 118, 257–268.
Haseeb, M.A. and Eveland, L.K. (2000) Human echinostomiasis: mechanisms of pathogenesis and
60 Chapter 1

host resistance. In: Fried, B. and Graczyk, T.K. (eds) Echinostomes as Experimental Models forBiological Research. Kluwer, Dordrecht, The Netherlands, pp. 83–98.
Hatz, C.F.R. (2000) The use of ultrasound in schistosomiasis. Advances in Parasitology 48, 226–285.IARC Working Group (1994) Schistosomes, Liver Flukes and Helicobacter pylori. IARC Monographs
on the Evaluation of the Carcinogenic Risk of Chemicals to Humans, Vol. 6, World HealthOrganization, Geneva, Switzerland, 270 pp.
Ikeda, T. (1998) Cystatin capture enzyme-linked immunosorbent essay for immunodiagnosis ofhuman paragonimiasis and fascioliasis. American Journal of Tropical Medicine and Hygiene 59,286–290.
Ishii, Y., Koga, M., Fujino, T., Higo, H., Ishibashi, J., Oka, K. and Saito, S. (1983) Human infectionwith the pancreas liver fluke Eurytrema pancreaticum. American Journal of Tropical Medicineand Hygiene 32, 1019–1022.
Jordan, P., Webbe, G. and Sturrock, R. (eds) (1993) Human Schistosomiasis. CAB International,Wallingford, UK.
Kabatereine, N.B., Ouma, J.H., Kemijumbi, J., Butterworth, A.E., Dunne, D.W. and Fulford, A.J.C.(1999) Adult resistance to schistosomiasis mansoni: age-dependence of reinfection remains con-stant in communities with diverse exposure patterns. Parasitology 118, 101–105.
Katz, N. (1999) Schistosomiasis vaccines: the need for more research before clinical trials.Parasitology Today 15, 165–166.
La Rue, G.R. (1957) The classification of digenetic trematodes. Experimental Parasitology 6, 306–344.Lin, J.X., Chen, Y.Z., Liang, C.Z., Yang, L.B. and Zhuang, H.J. (1985) Epidemiological investigation
and experimental infection of Echinochasmus japonicus [in Chinese. English summary].Chinese Journal of Parasitology and Parasitic Diseases 3, 89–91.
Liu, Y.S., Du W.P., Wu, Y.M., Chen, Y.G., Zhang, K.Y., Shi, J.M., Hu, X.Z., Li, G.Y., You, C.F. and Wu,Z.X. (1995) Application of dot-immunogold–silver staining in the diagnosis of clonorchiasis.Journal of Tropical Medicine and Hygiene 98, 151–154.
MacLean, J.D., Arthur, J.R., Ward, B.J., Gyorkos, T.W., Curtis, M.A. and Kokoskin, E. (1996) Common-source outbreak of acute infection due to the North American liver fluke Metorchis conjunctus.Lancet, British Edition 347, 154–158.
Malek, E.A. (1963) Medical Malacology. Burgess, Minneapolis, USA.Mas Coma, M.S., Esteban, J.G. and Bargues, M.D. (1999) Epidemiology of human fascioliasis: a
review and proposed new classification. Bulletin of the World Health Organization 77, 340–346.Mostafa, M.H., Badawi, A.F. and O’Connor, P.J. (1995) Bladder cancer associated with schistosomia-
sis. Parasitology Today 11, 87–89.Mott, K.E., Nuttall, I., Desjeux, P. and Cattond, P. (1995) Geographical approaches to control of some
parasitic zoonoses. Bulletin of the World Health Organization 73, 247–257. Mouchet, F., Develoux, M. and Mamadou, B.M. (1988) Schistosoma bovis in human stools in
Republic of Niger. Transactions of the Royal Society of Tropical Medicine and Hygiene 82, 257. Muller, R. (2001) Dogs and trematode zoonoses. In: Macpherson, C.N.L., Meslin, F.-X. and Wandeler,
A.I. (eds) Dogs, Zoonoses and Public Health. CAB International, Wallingford, UK, pp. 149–176. O’Neill, S.M., Parkinson, M., Dowd, A.J., Strauss, W., Angles, R. and Dalton, J.P. (1999) Short report:
immunodiagnosis of human fascioliasis using recombinant Fasciola hepatica cathepsin L1 cysteine proteinase. American Journal of Hygiene and Tropical Medicine 60, 749–751 (also1998, 58, 417–423).
Pecoulas, P.E., Farhati, K. and Picot, H. (1995) Les schistosomes humaines d’Asie à l’exception deSchistosoma japonicum. Médecine et Maladies Infectieuses 25, 99–106.
Pointier, J.P. and Jourdane, J. (2000) Biological control of the snail hosts of schistosomiasis in areas oflow transmission: the example of the Caribbean area. Acta Tropica 77, 53–60.
Rohde, K., Hefford, C., Ellis, J.T., Baverstock, P.R., Johnson, A.M., Watson, N.A. and Dittman, S.(1993) Contributions to the phylogeny of the Platyhelminthes based on partial sequencing of 18sribosomal DNA. International Journal for Parasitology 23, 705–724.
Ross, A.G.P., Li-TueSheng, Sleigh, A.C., McManus, D.P. and Li, Y.S. (1997) Schistosomiasis control inthe People’s Republic of China. Parasitology Today 13, 152–155.
Ross, A.G.P., Sleigh, A.C., Li, Y.S., Williams, G.M., Aligui, G.D.L. and McManus, D.P. (2000) Is thereimmunity to Schistosoma japonicum? Parasitology Today 16, 159–164.
The Trematodes 61

Savioli L., Renganathan, E., Montresor, A., Davis, A. and Behbehani, K. (1997) Control of schisto-somiasis – a global picture. Parasitology Today 13, 444–448.
Shekar, K.C. (1991) S. malayensis: the biologic, clinical, and pathological features in man and experi-mental animals. In: Sun, T. (ed.) Progress in Clinical Parasitology, Vol. 2. Field and Wood,Philadelphia, USA, pp. 145–178.
Shoop, W.L. (1994) Vertical migration in the Trematoda. Journal of the Helminthological Society ofWashington 61, 153–161.
Sirisinha, S., Chawengkirttikul, R., Haswell-Elkins, M.R., Elkins, D.B., Kaewkes, S. and Sithithaworn,P. (1995) Evaluation of a monoclonal antibody-based enzyme linked immunosorbent assay forthe detection of Opisthorchis viverrini infection in an endemic area. American Journal ofTropical Medicine and Hygiene 52, 521–524.
Smyth, J.D. (1995) Rare, new and emerging helminth zoonoses. Advances in Parasitology 36, 1–47.Southgate, V.R. (1997) Schistosomiasis in the Senegal River basin: before and after the construction
of the dams at Diama, Senegal and Manantali, Mali and future prospects. Journal ofHelminthology 71, 125–132.
Stephenson, L. (1993) The impact of schistosomiasis on human nutrition. Parasitology 107 (suppl.),S107-S123.
Subramanian, A.K., Mungai, P., Ouma, J.H., Magak, P., King, C.H., Mahmoud, A.A.F. and King, C.L.(1999) Long-term suppression of adult bladder morbidity and severe hydronephrosis followingselective population chemotherapy for Schistosoma haematobium. American Journal ofTropical Medicine and Hygiene 61, 476–481.
WHO (1993) The Control of Schistosomiasis. Technical Report Series No. 830, World HealthOrganization, Geneva, Switzerland.
WHO (1995) Control of Foodborne Trematode Infections. World Health Organization, Geneva,Switzerland.
Wilkins, H.A., Goll, P.H., Marshall, T.F. de C. and Moore, P.J. (1984) Dynamics of Schistosomahaematobium infection in a Gambian community. III. Acquisition and loss of infection.Transactions of the Royal Society of Tropical Medicine and Hygiene 28, 227–232.
Wilkins, H.A., Blumenthal, U.J., Hagan, P., Hayes, R.J. and Tullock, S. (1987) Resistance to reinfec-tion after treatment of urinary schistosomiasis. Transactions of the Royal Society of TropicalMedicine and Hygiene 81, 29–35.
Williams, S.A. and Johnston, D.A. (with others) (1999) Helminth genome analysis: the current statusof the filarial and schistosome genome projects. Parasitology 118 (suppl.), 519–538.
Wynn, T.A. and Hoffmann, K.E. (2000) Defining a schistosomiasis vaccination strategy – is it reallyTh1 versus Th2? Parasitology Today 16, 497–501.
Yamaguti, S. (1971) Synopsis of Digenetic Trematodes of Vertebrates. Keigaku, Tokyo, Japan.Yong, T.S., Yang, H.J., Park, S.J., Kim, Y.K., Lee, D.H. and Lee, S.M. (1998) Immunodiagnosis of
clonorchiasis using a recombinant antigen. Korean Journal of Parasitology 36, 183–190.Yu, S.-H. and Mott, K.E. (1994) Epidemiology and Morbidity of Food-borne Intestinal Trematode
Infections. WHO document: WHO/SCHISTO/94.108. World Health Organization, Geneva,Switzerland.
Further Reading
Bergquist, N.R. (1992) Immunodiagnostic Approaches in Schistosomiasis. John Wiley & Sons,Chichester, UK.
Jordan, P., Webbe, G. and Sturrock, R. (eds) (1993) Human Schistosomiasis. CAB International,Wallingford, UK.
Rollinson, D. and Simpson, A.J.G. (eds) (1986) The Biology of Schistosomes: from Genes to Latrines.Academic Press, London, UK.
Wellcome Trust (1998) Schistosomiasis. Topics in International Health CD-ROM, CAB International,Wallingford, UK.
62 Chapter 1

All adult cestodes (or tapeworms) inhabitthe intestinal tract or associated tracts ofvertebrates.
They are flat, ribbon-shaped worms con-sisting of a chain of a few or many separateproglottides, the whole chain being knownas a strobila. At the anterior end is thescolex or holdfast organ, which usuallyattaches to the intestinal mucosa by meansof suckers or sucking grooves and often hashooks. Posterior to the scolex is the narrowundifferentiated neck region, from whichgrowth takes place to give rise to the devel-oping proglottides.
In the order Cyclophyllidea, to whichmost of the adult tapeworms of humansbelong, the proglottides become succes-sively mature and then gravid, when theyare filled with eggs. At the gravid stagethey become detached and pass out withthe faeces. All tapeworms of humans arehermaphroditic, the male organs develop-ing first. Thus male organs alone are foundin the mature proglottides nearer to thescolex; further back the organs of bothsexes are present but the female organsonly are functional. With large tapewormsthere is often a single specimen presentand cross-fertilization occurs betweenproglottides. The genital organs are welldeveloped in each proglottid. The malesystem consists of the testes (three in num-ber in Hymenolepis to over 500 in Taenia),the vasa efferentia, which unite to form avas deferens, and a protrusible, muscularcirrus or penis. The cirrus opens into acommon genital pore on the margin of the
proglottid just anterior to the vagina. Thefemale organs lie ventrally in the proglot-tid. The ovary is usually bilobed and liesposteriorly. The ova pass into the oviduct,which joins the spermatic duct from theterminal part of the vagina or seminalreceptacle. The ova then pass into theoötype, where fertilization occurs. Thevitelline ducts from the vitellaria also openinto the oötype and provide shell materialand yolk cells. The uterus extends anteri-orly from the oötype and ends blindly. Theuterus fills with eggs and, when gravid, hasa shape characteristic of its family (e.g.with lateral branches in taeniids and alobate transverse sac in hymenolepids).
Members of the order Pseudophyllideashow structural and life-history differencesfrom the Cyclophyllidea, which arementioned in the section dealing with theonly member that is an important parasiteof humans, the broad fish tapeworm,Diphyllobothrium.
As cestodes have no gut, all nutritiveand waste material must pass through theouter covering or tegument; this functionsalmost as an inside-out gut wall and con-tains specific systems of molecular and iontransport, e.g. of amino acids, hexose sug-ars, nucleotides, lipids, vitamins, purinesand pyrimidines. The tegument consists ofa syncytial distal cytoplasm with a superfi-cial anucleate zone and an inner nucleatedcytosomal zone. The external surface iscovered with a row of michrotriches,similar to intestinal microvilli, whichincreases the surface area in a similar way
2The Cestodes
63

(increasing the functional area by 1.7–11.8in some cestodes investigated, comparedwith a 26 times increase for mousemicrovilli). On the tip of each microtrichethere is a glycocalyx, composed ofmucopolysaccharide and glycoproteinacidic groups, which absorbs and incorpo-rates hexose sugars and amino acids, con-centrates inorganic and organic ions, bindshost amylase and carries out ‘contact diges-tion’ (Smyth and McManus, 1989).
In the great majority of infections theadult tapeworms that normally occur inhumans do not cause any serious pathol-ogy, although their presence can beextremely unpleasant. Accidental adulttapeworms contracted zoonotically areencountered rarely, principally in children,and are usually short-lived. However,infection with larval tapeworms is poten-tially very serious and in some countriescysticercosis (caused by the metacestodesof Taenia solium) and echinococcosis areimportant public health problems (56species of adult and larval cestodes areincluded as having occurred in humans).
Classification
KEY: Members are parasites of M = mammals,B = birds, R = reptiles, A = amphibians, F =fish.
PHYLUM PLATYHELMINTHES
CLASS CESTODANo digestive system. Presence of calcare-ous corpuscles. Usually hermaphrodite.
SUBCLASS EUCESTODABody typically divided into short proglot-tides with anterior holdfast organ, oftenwith suckers and hooks. Egg with 6 hooks.There are many orders of no medicalimportance, which are not included (seeKhalil et al., 1994).
Order Pseudophyllidea (MBRAF)Length very variable but may be up to30 m. Holdfast typically with dorsal andventral sucking grooves. Genital aperturesusually on ventral surface. Testes and vitel-laria scattered and follicular. Eggs usually
operculate. Life cycle includes procercoidand plerocercoid larvae.Diphyllobothrium, Spirometra
Order Cyclophyllidea (MBRA)Length variable, from 3 mm to 10 m.Holdfast typically with 4 large suckers.Apical end often with row of hooks.Proglottides usually wider than long.Genital apertures marginal. Vitellaria sin-gle, compact, postovarian. Gravid uteruswithout aperture and sometimes brokeninto capsules. Eggs non-operculate. Larvacommonly a cysticercus (bladder-worm) ina vertebrate, or a cysticercoid in an arthro-pod.
Family Taeniidae (MB)Rostellum of scolex non-retractable and usu-ally with 2 circles of hammer-shaped hooks.Testes numerous. Gravid proglottides longerthan broad. Gravid uterus has median stemand lateral branches. Larva a cysticercus,coenurus or hydatid in a vertebrate.Taenia, Echinococcus
Family Hymenolepididae (MB)Rostellum retractable, hooks thornlike.Testes rarely more than 4. Gravid uterus atransverse tube. Larva a cysticercoid ininsects (except H. nana).Hymenolepis
Family Dipylididae (M)Rostellum retractable, hooks typicallythorn-shaped. Genitalia single or double.Testes usually numerous. Mature proglot-tides longer than wide. Gravid uterus atransverse lobed sac or replaced by eggsacs. Larval stage a cysticercoid in insects.Dipylidium
Family Anoplocephalidae (MBR)No rostellum. Testes numerous. Gravidproglottides much wider than long. Graviduterus often replaced by paruterine organs.Eggs often with characteristic membranedrawn out into a pair of cross-taperedprocesses (pyriform apparatus). Larva acysticercoid in oribatid mites.Bertiella, Inermicapsifer
Family Davaineidae (MB)Rostellum retractable, hooks T-shaped.Suckers often spinose. Gravid uterus
64 Chapter 2

replaced by egg pouches. Larva a cysticer-coid in insects.Raillietina
Family Mesocestoididae (MB)No rostellum. Mature proglottides widerthan long, genital apertures median.Posterior paruterine organ. First larvalstage probably in oribatid mites, secondtetrathyridium in vertebrates.Mesocestoides
Order Pseudophyllidea
Diphyllobothrium latum(Linnaeus, 1758) Lühe, 1910
SYNONYMS
Taenia lata Linnaeus, 1758; Dibothrio-cephalus latus Lühe, 1899; Bothrio-cephalus latus Bremser, 1918.
DISEASE, POPULAR AND LOCAL NAMES
Diphyllobothriasis or diphyllobothriosis;broad fish tapeworm infection; Lapamato(Finnish), Breda binnikemaskens (Swedish).
GEOGRAPHICAL DISTRIBUTION
It is estimated that there are about 20 mil-lion cases worldwide. In Europe it is foundprincipally in countries with many lakes,such as Finland, Estonia, Russia (Don and
Volga Rivers), Poland and Ukraine, withsporadic cases in Italy/Switzerland (LakeMaggiore) and Slovakia (Danube basin); inAsia, in Japan, north-eastern Russia (AmurRiver basin) and with a few cases in China,India (one case) and Korea; in theAmericas, in the Great Lakes regions ofCanada and the USA, Argentina and Chile(Valdiva River basin).
LOCATION IN HOST
The adult worm or worms inhabit thelumen of the small intestine with thescolex embedded in the mucosa.
MORPHOLOGY
Diphyllobothrium is the longest tapeworminfecting humans. It measures 3–10 m inlength and may have 4000 proglottides.Each proglottid measures 2–7 mm �10–12 mm, and contains both male andfemale reproductive organs (Fig. 26).Diphyllobothrium differs from all otherhuman tapeworms in having a uterine porethrough which eggs are discharged (this istypical of the order Pseudophyllidea andsimilar to the situation in trematodes). Thispore and the cirrus and vagina open on theventral surface, not at the lateral margin asin the Cyclophyllidea. The scolex measures1–2 mm and has no suckers but attaches tothe mucosa by two sucking grooves (Fig. 27).
The Cestodes 65
testes
uterus
cirrusgenital pore
vaginavas deferens
vitellariavitelline duct
ootypeovary
1 mm
Fig. 26. Mature proglottid of Diphyllobothrium latum. The vitellaria are shown principally on the right-hand side, the testes on the left. Note genital uterine pore, through which eggs emerge.

LIFE CYCLE (Fig. 28)Up to 1 million eggs can be produced dailyand these pass out in the faeces. The ovoideggs measure 55–80 µm � 40–60 µm andare similar in structure to those of trema-todes (Fig. 123). They are yellowish-brown,operculate and with a small knob at theend opposite to the operculum. The ovumis unsegmented when laid and is sur-rounded by yolk cells. If the egg reachesfresh water, the ovum develops in 12 days
to many weeks, depending on the tempera-ture, into a ciliated larva (termed a coracid-ium) measuring 40–50 µm and with sixsmall hooks (Fig. 29a). The coracidiumlarva escapes into the water through theopened operculum and swims around inan apparently aimless manner. Thecoracidium can survive for 1–2 days but forfurther development must be ingested by asuitable planktonic copepod belonging tothe genus Diaptomus or, more rarely inEurope, Cyclops, living in large bodies ofwater (about 40 species including D. vul-garis, D. gracilis, C. vicinus and C.strenuus). Inside the stomach of the crus-tacean, the outer epithelium is lost and theenclosed larva penetrates the gut wall withthe aid of the hooks. In the body cavity aprocercoid larva, measuring 0.5 mm inlength, develops in 14–21 days (Fig. 29b).
66 Chapter 2
Fig. 27. Scolex with two sucking grooves.
ADULTS insmall intestinemature in 3–5weeks
Fish ingestedby human
Copepod ingested byfish: PLEROCERCOIDin muscles infectivein 4 weeks Ingested by copepod:
PLEROCERCOIDdevelops in 2–3 weeks
CORACIDIUMlarva lives for1–2 days
EGGS developin 2 to manyweeks in water
UnsegmentedEGGS (55–80�mx 40–60�m)in faeces
Fig. 28. Cartoon illustrating the life cycle of Diphyllobothrium latum (after Tiedoksianto Information,1971, 12, 3–9).

If the copepod is ingested by a plankton-eating freshwater fish, the procercoid larvapenetrates the intestinal wall in about 4 hand develops into a plerocercoid larva inthe connective tissues and muscles. Theplerocercoid is an elongate wormlike larvawith an inverted scolex and takes about 4weeks to become infective (Fig. 29c). Thelarvae eventually become encysted butusually remain infective throughout the
life of the fish. The small copepods (about1 mm long) are often ingested by small fishand, when these are eaten by a larger fish,such as pike, perch, burbot or salmon, thereleased plerocercoids can penetrate theintestinal wall and remain viable. In thisway the larval parasites become concen-trated in the larger carnivorous food fishes,which act as paratenic hosts (e.g. the pike).Human infection occurs when fish are eaten
The Cestodes 67
Fig. 29. Larval stages of D. latum. (a) Free-living coracidium. (b) Procercoid from copepod. (c) Plerocercoid from fish.

raw or semi-cooked. The released larvaattaches to the intestinal wall by the evertedscolex and develops proglottides. Diphyllo-bothrium reaches maturity in 3–5 weeks andcan live for up to 25 years in humans.Usually only a single worm is present. Aftervoiding eggs, a chain of the oldest proglot-tides breaks off and is often passed out in thefaeces (pseudoapolysis). A small form of D.latum, measuring about 60 cm � 2.3 mm,has been reported from Japan.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Often none. In a proportion of cases thereis abdominal pain, loss of weight, anorexiaand vomiting. Sometimes the wormbecomes lodged in the jejunum, where itcauses acute pain and vomiting of seg-ments.
The most important pathological effectof diphyllobothriasis is anaemia, whichoccurs only in Finland and is of the perni-cious megablastic (macrocytic) type. Itused to occur in about 20% of cases but isnot so common since diet has improved.The cause appears to be a competitiveuptake of vitamin B12 by the worm, ratherthan any substance released by it interfer-ing with Castle’s intrinsic factor (vonBonsdorff, 1977). Diphyllobothrium has avitamin B12 content 50 times that of Taeniasaginata and absorbs 80–100% of radio-active B12 added to the diet of symptomlesscarriers. The North American strain ofDiphyllobothrium takes up only 6–7%,however, and anaemia is very rare.Anaemia in Finland possibly occurs: (i)because the strain of worm is found higher
up the gut, in the jejunum and nearer to thestomach; (ii) because of a reduction inintrinsic factor resulting from damage tothe gastric mucosa; (iii) principally becauseof a low dietary intake of B12 (supported bythe finding that pernicious anaemia hasdecreased greatly since the diet hasimproved); (iv) because of human geneticfactors. There can be up to 25%eosinophilia in early infections.
Occasional cases of multiple infectionhave been known to cause obstruction ofthe small intestine.
DIAGNOSIS
This is by discovery of the characteristiceggs in the faeces but they need to be dif-ferentiated from similar trematode eggs,such as those of Paragonimus (Fig. 123).
Sometimes a chain of proglottides ispassed in the faeces at intervals and thesecan be recognized by the characteristicshape of the uterus (Fig. 30), but there is noregular expulsion of gravid proglottides, aswith taeniid tapeworms.
TREATMENT
Praziquantel is the drug of choice at a sin-gle oral dose of 15–30 mg kg�1, but it isimportant to make sure that the scolex hasbeen expelled .
Niclosamide (2,2�-dichloro-5,4�-nitro-salicylanilide) at 1 g first thing in the morn-ing and then 1 g 1 h later (scolex isdestroyed) and bithionol (50–60 mg kg�1,repeated a day or so later if no scolex isrecovered) have been used for many years.
68 Chapter 2
0.5 mmuterus
Fig. 30. Chain of proglottides as sometimes passed in the faeces. The characteristic branched uteruscan be recognized by the tanned eggs.

EPIDEMIOLOGY
Raw or smoked freshwater fish is an impor-tant item of diet in many countries. Inreservoirs in the Krasnoyarsk region ofSiberia the tipping of untreated sewage hascaused 100% infection in pike living inlakes and 5% in rivers, with 35,000 casesreported annually in Russia overall.‘Caviar’ made from the roe of pike is a pop-ular dish in that area and is the cause of aseasonal rise in the infection rate in thespring when the fish have roe. In theArchangel region (as in Finland) infectionhas been much reduced by improved treat-ment of sewage.
Around the Baltic Sea, slices of raw fishknown as ‘strogonina’ are a popular item ofdiet and are important in transmission, asis ‘sashimi’ in Japan.
PREVENTION AND CONTROL
The plerocercoid larvae in fish are killedby freezing at �10°C for 15 min. They arealso killed by complete drying or by thor-ough pickling.
Proper disposal and treatment of faecesare also of great importance in reducingincidence.
ZOONOTIC ASPECTS
Infection with D. latum has been recordedfrom many mammals, including dogs, catsand bears. However, with the exception ofdogs, animals are probably not of impor-tance in transmission; tapeworms in catslast only for a matter of weeks.
Numerous other species of Diphyllo-bothrium and related genera are occasion-ally found in humans, mostly contractedfrom marine fish (Ruttenber et al., 1984).However, the taxonomic status of specieswithin this genus is rather confused and itis often very difficult to differentiate themon adult characters alone, although thestructure and size of eggs can be useful.Diphyllobothrium pacificum (Nybelin,1931) Margolis, 1956, is a natural parasiteof fur seals and sea lions with plero-cercoids in marine fish (e.g. mackerel) andhas been found in humans in Chile,Ecuador and Japan. Other species with afew cases in humans include: D. cameroni
Rausch, 1969, a natural parasite of monkseal, in Japan; D. cordatum Cobbold, 1879,found naturally in seals and walrus, inAlaska; D. dalliae Rausch, 1956, and D.dendriticum (Nitzsch, 1824), parasites ofgulls and freshwater fish, in Alaska; D. ele-gans (Krabbe, 1865), naturally in seals andsea lion, in Japan; D. giljacicum (= D. luxi)Rutkevich, 1937, in freshwater fish andadults only known from humans in Siberia;D. hians (Diesing, 1850), naturally in seals,in Japan and Siberia; D. klebanovskii (= D.luxi) Muratov and Posokhov, 1988, natu-rally in brown bears and salmon, inBelarus and Siberia; D. lanceolatum(Krabbe, 1865), naturally in seals, inAlaska; D. minus Cholodkovsky, 1916, nat-urally in seagulls, in Russia; D. nenziPetrov, 1938, in Russia; D. nihonkaiense (=D. luxi) Yamane et al., 1986, is apparently ahuman parasite in Japan and Siberia con-tracted from salmon; D. orcini Hatsushikaand Shirouzu,1990, naturally in killerwhale, in Japan; D. scoticum (Rennie andReid, 1912), naturally in seals and sea lion,in Japan; D. skrjabini Plotnikov, 1932, indogs and freshwater fish, in Russia; D. tan-gussicum Podjapolskaia and Gnedina,1932, is apparently a human parasite inRussia; D. ursi Rausch, 1954, naturally inbears, in Canada and Alaska; D. yona-goense Yamane et al., 1981, natural cycleunknown, in Japan and Korea.
Diplogonoporus grandis (= D. balan-opterae) (Blanchard, 1894) Lühe, 1899, is aparasite of whales and has been reportedover 50 times from humans in Japan (caus-ing abdominal discomfort) and Korea, witha single case in Spain. Each proglottid hasa double set of reproductive organs and theeggs measure 63–68 µm � 50 µm. Otherspecies are D. brauni Leon, 1907, inRomania and D. fukuokaensis Kamo andMiyazaki, 1970, in Japan. Ligula intesti-nalis Linnaeus (1758) Bloch, 1782, a com-mon and widespread parasite of fish-eatingbirds, has been found in humans in Polandand Romania; Schistocephalus solidus(Mueller, 1776) Creplin, 1829, anothercommon bird parasite, has been recordedfrom humans in Alaska; Pyramicocephalusanthocephalus (Fabricius, 1780) Monticelli,
The Cestodes 69

1890, is a pseudophyllid tapeworm ofseals, which has been recorded fromhumans in Alaska and Greenland.
Sparganosis
Sparganosis is the term used to describeinfection with the plerocercoid larvae (=spargana) of diphyllobothriid tapeworms ofthe genus Spirometra (in some textbooksthey are included in the genusDiphyllobothrium, but it is clearly a validgenus, although more correctly it should beLuheella).
The adult tapeworms are parasites ofcarnivores in tropical and subtropicalregions of Africa, the Americas, Australia,Europe and Asia. The mature proglottidescan be easily differentiated from those ofDiphyllobothrium by the structure of theuterine coils, which form compact loops inthe midline rather than a rosette shape(Fig. 30). The eggs can also be differenti-ated.
The life cycle resembles that ofDiphyllobothrium except that the procer-coids develop in cyclopoid species thatinhabit small ponds rather than lakes(Eucyclops serrulatus and Mesocyclopsleuckarti and probably the other species
transmitting Gnathostoma and Dracunculus)(Fig. 31); also the plerocercoids do notdevelop in fish but, in most parts of theworld, in amphibians, reptiles and smallmammals and, in East Africa, in large gameanimals. A wide range of vertebrates,including humans, in which adult tape-worms will not develop, can also act asparatenic hosts. Human infection mayoccur in one of three ways:
1. By ingestion of cyclops containing pro-cercoid larvae in drinking water fromponds or open wells; this is the most likelymode of infection in many parts of theworld.2. By application of the flesh of an infectedfrog to ulcers, sores and infected eyes; theprocercoid is activated by warmth andenters the flesh. This form of traditionalpoultice is used only in areas of South-EastAsia, particularly Thailand and Vietnam,and spargana in the eye can result in severeoedema and eventually nodule formation,usually of the upper eyelid; when removedthe contained larva has often degeneratedinto a caseous mass. 3. By ingestion of raw or undercooked meatof infected frogs, snakes, mammals or birds;the contained plerocercoids burrowingthrough the wall of the intestine (Plate 8).
70 Chapter 2
Fig. 31. Cyclops with procercoids of Spirometra sp. in the haemocoel. Calcareous bodies can be seenin the bodies of the parasites.

Spargana wander in the deep tissuesand usually migrate to the subcutaneoustissues, where they eventually encyst,resulting in a fibrous nodule about 2 cm indiameter. Surgical excision of the nodule isusually possible, and the enclosed larva,which has a scolex similar to that ofDiphyllobothrium but with a tail many cen-timetres in length, is sometimes mistakenfor a guinea worm in Africa. The most seri-ous effects are caused by larvae in thebrain, which can cause repeated seizures(in 84% of patients), hemiparesis (59%)and headache (56%) with convulsions, andcan be diagnosed by computer-assistedtomography (CT) or magnetic resonanceimaging (MRI) scans (Chang et al., 1992).The lesions need to be removed surgically;high dosages of praziquantel have notalways been successful. Very rarely, larvaemay enter the eye and cause endoph-thalmitis, leading to blindness (Sen et al.,1989) and larvae may also enter variousvisceral organs. Sparganosis can be diag-nosed by a chemiluminescence ELISA.
Many species of Spirometra have beendescribed from various areas of the world,but most are morphologically variable andthe species position is very confused.Parasites from Asia or Europe are usuallytermed S. mansoni (Cobbold, 1882), S.houghtoni (Kellogg, 1927) or S. erinacei (=S. erinaceieuropei) (Rudolphi, 1819), withplerocercoids in frogs and snakes andadults occurring naturally in carnivores,including cats and dogs (16% in Thailand),those from Africa S. theileri (Baer, 1925),with plerocercoids in wild herbivores andadults in large carnivores, and worms fromthe Americas S. mansonoides (Mueller,1935), with plerocercoids in frogs andsnakes and adults in wild carnivores andcats and dogs (the adults of this species canbe differentiated morphologically from allthe others and produce a growth hormone-like cysteine proteinase that causes weightgain in experimentally infected mice).
The presence of spargana is becomingmore commonly identified with an increas-ing concern about strange growths, andthere is an ELISA using crude antigens
of adult or larval worms which works well.
‘Sparganum proliferum’ is a rare form,of unknown relationship, in which theplerocercoids become disorganized andproliferate in the tissues by lateral budding(DNA analysis shows that they are of thegenus Spirometra but of unknown species).In human cases found at autopsy manythousands of larvae have been present inthe viscera as well as in the subcutaneoustissues. It is possible that virus infection ofa normal sparganum may be responsible.Three fairly recent similar cases, probablyof proliferating spargana, have beendescribed, one in an abdominal mass froman Amerindian in Paraguay (Beaver andRolon, 1981) and two others from the lungsof a man and the spinal cord of a woman inTaiwan (Lo et al., 1987).
Order Cyclophyllidea
The majority of tapeworms of medical andveterinary importance belong to this order.
Family Taeniidae
This group of taeniid tapeworms includesthe most important cestode parasites ofhumans. The adults of two species, Taeniasaginata and T. solium, are only found inhumans but are usually of minor medicalimportance. The larval stages are always inmammals and consist of a fluid-filled cyst,which may contain a single or many proto-scolices, each of which is capable of devel-oping into an adult tapeworm in adefinitive host (Fig. 32). Human infectionwith larval cestodes can be a very seriousmedical problem indeed.
Taenia saginata Goeze, 1782
SYNONYMS
Taeniarhynchus saginatus Weinland, 1848;Taenia africana Linstow, 1900; T. confusa
The Cestodes 71

Ward, 1896; T. hominis Linstow, 1902. Inthe literature from eastern Europe this tape-worm is commonly called Taeniarhynchussaginatus, because some authorities believethat the lack of rostellum and hooks is suffi-cient to warrant a genus separate fromTaenia (Bessenov et al., 1994). However,there is evidence that there are rudimentaryhooks in the cysticerci and it is likely thatthese have been secondarily lost.
DISEASE AND POPULAR NAME
Taeniasis or taeniosis saginata; beef tape-worm infection.
LOCAL NAMES
Dooda shareatia or dedan (Arabic), Thankaung (Burmese), San-chik (Cantonese),Sanadu mushi (Japanese), Miliny (Luo),Tegu (Swahili), Nada puchi (Tamil), Pa-yard tua tued (Thai), Sonsono (Twi), Aráninú (Yoruba).
GEOGRAPHICAL DISTRIBUTION
Infection is cosmopolitan and occurs inalmost all countries where beef is eaten rawor undercooked. In most countries it is morecommon than the pork tapeworm (T.solium), with about 60 million cases in theworld.
LOCATION IN HOST
The scolex of the adult tapeworm isembedded in the mucosa of the wall of theileum, with the rest of the tape extendingthroughout the lumen.
MORPHOLOGY
The scolex is pear-shaped, 1–2 mm indiameter, but with no rostellum or hooks.The adult worm is very long, sometimesexceeding 20 m. Usually, however, it isabout 5 m long with 1000–2000 proglot-tides. Each mature proglottid measuresabout 12 mm wide and 10 mm long andeach gravid proglottid contains about 0.8–1� 105 eggs.
Female organs. The ovary is in the post-erior part of the proglottid and has twolarge lobes. The follicular vitellaria (whichproduce the eggshell and nutrient yolkcells) lie in a band behind the ovaries. Theuterus extends forwards as a tube, slightlydilated anteriorly. In contrast to the uterusof Diphyllobothrium, it has no opening tothe exterior. Gravid proglottides contain acharacteristic taeniid type of uterus filledwith eggs (Figs 33 and 34).
Male organs. The numerous follicular testesare scattered throughout the anterodorsalpart of the proglottid. The vas deferensopens through the muscular cirrus at thecommon genital pore, which is situatedirregularly on alternate lateral margins.
LIFE CYCLE
The eggs measure 43 µm � 31 µm and areidentical to those of T. solium and manyother taeniids. They are deposited in humanfaeces (about 0.7–1 � 106 24 h�1), ofteninside the gravid proglottid, or the latter
72 Chapter 2
Fig. 32. The various types of taeniid larvae. They contain one or many protoscolices.

The Cestodes 73
45
graviduterus
T. solium T. saginata
Either species
uterus
testes
ovaryvagina
Mehlis’ gland
vitellarium
1 mm
genitalopening
cirrus
vasdeferens
excretoryduct
nerve
3
T. solium
0.5 mm
1 2
T. saginata
6
7
Fig. 33. Comparative morphological features of Taenia solium and Taenia saginata.1 and 2. Scolices (6 and 7 are the scolices of Hymenolepis nana and H. diminuta at the same scale). 3. Mature proglottid of either species. 4 and 5. Gravid proglottides showing the typical numbers ofbranches of the uterus in each species.

may pass out through the anus and for fur-ther development need to be ingested bycattle, when the contained oncosphere larvahatches in the duodenum. The oncospherespenetrate through the intestinal wall withthe aid of the six hooklets and enter thevenous capillaries (or the mesenteric lym-phatics) in about 30 min. The embryos arecarried by the circulation until they reachthe voluntary muscles, where they lose thehooks and develop into an infective cys-ticercus larva in 10–12 weeks, or sometimeslonger. The fully formed cysticercus isovoid and white in colour and measuresabout 8 mm � 5 mm (it is known to veteri-narians as a ‘cysticercus bovis’). The invagi-nated protoscolex has four suckers but nohooks. The outer wall of the fluid-filledbladder has microvilli, possibly serving atrophoblastic function.
Humans become infected from eatingraw or undercooked beef. The protoscolexof the cysticercus evaginates in the duode-num, attaches to the wall of the ileum andbegins to form proglottides.
CLINICAL MANIFESTATIONS
Often the first and only sign of infection isthe presence of active proglottides in thefaeces, perhaps preceded by the feeling ofthem ‘crawling’ out through the anus.
As the tapeworm matures, 6–8 weeksafter infection, there may be epigastric or
umbilical pain, nausea, weakness, loss ofweight, alteration of appetite, and head-ache. These symptoms and signs are possi-bly caused by toxic products produced bythe worms or are allergic reactions, and arecharacteristically alleviated by taking food.
Occasionally, generalized allergic mani-festations, such as urticaria with pruritusani or widespread pruritus, result. Thepresence of a lump in the throat is a notuncommon symptom, reported particularlyby middle-aged women in Poland. Perhapsunsurprisingly, symptoms often increase infrequency and severity once patients areaware they have a tapeworm. Occasionallya worm causes obstruction due to bolusimpaction in the intestine and it has beenknown for proglottides to be vomited andthen aspirated.
A moderate eosinophilia occurs in5–45% of patients.
PATHOGENESIS
Little pathology can be directly attributedto the physical action of the scolex on themucosa, although inflammation of themucosa of the ileum sometimes occurs.Over 100 cases of intestinal obstruction,perforation or appendicitis have beenreported. In a recent case in Reunionisland, intestinal perforation led to deathfrom septic shock.
74 Chapter 2
Fig. 34. Section of proglottides of T. saginata in the appendix. The uterus contains eggs.

DIAGNOSIS
Parasitological. The presence of gravidproglottides with the characteristic taeniidbranched uterus in the faeces provides theusual method of diagnosis. These arepassed out in short chains of 5–8 proglot-tides, are very active and sometimes can befelt migrating out of the anus. Whenpressed between glass slides, the number oflateral branches of the uterus can becounted in order to differentiate T. saginata(15–32, usually 20–23, main branches) fromT. solium (7–13, usually 9, main branches).However, in South Africa at least somedegree of overlap was reported, but it is notclear how much. The two species can beclearly differentiated if the scolex is passedin the faeces after chemotherapy, because ofthe lack of rostellum and hooks in that of T.saginata. There are also small differences inthe mature proglottides: those of T. saginatahave a vaginal sphincter muscle and testesconfluent posteriorly to the vitellarium,while those of T. solium have a third lobe tothe ovary. Unfortunately, special prepara-tion is often necessary in order to see thesefeatures.
Free eggs may be found in the faeces ifthe gravid proglottides have been auto-lysed, particularly using concentrationtechniques, and may also be found by tak-ing possibly repeated anal swabs withsticky tape (p. 163) or a sticky glass spat-ula, as used in Russia, and examiningunder the microscope, although species oftaeniids cannot be differentiated.
Serological. The detection of coproantigens infaeces by capture ELISA techniques hasproved sensitive and also specific(Machnicka et al., 1996). There are varioustechniques for diagnosis of cysticerci incattle. The best probably recognize circulat-ing excretory/secretory (ES) antigens ofcysticerci by monoclonal antibody-based dotELISA, SDS-PAGE or immunoblotting (whichrecognizes antigens of 65, 87 and 100 kDa).
TREATMENT
Praziquantel at a single oral dose of 5–10mg kg�1 is usually completely effective.
Albendazole at a single oral dose of 800 mghas about 80% effectiveness. Niclosamide(2 g for an adult given in a divided doseover 2 days) has been in use for many yearsbut has (usually mild) side-effects and isnot always effective. With newer com-pounds the scolex is destroyed and so fol-low-up is necessary to determine that thecomplete worm has been expelled (if thescolex should remain embedded, a newworm can develop within weeks).
EPIDEMIOLOGY
The prevalence of T. saginata infection hasremained high in areas of Europe; variousrecent regional estimates are 0.4–9.5% inBelgium, 1.3–1.7% in northern Italy, 4.0%in northern Germany, 20,000 cases annuallyin Russia, 0.9% in Slovakia and 2.5% inTurkey. It is rare in the UK (627 cases inEngland and Wales between 1990 and1998). In other parts of the world estimatesare 1.3% in Afghanistan, 0.5% in Brazil,0.4–1.0% in Egypt, 1.0% in Ethiopia, 0.3%in Honduras, 7.7% in Iran and 2.3% in east-ern and 11.5% in northern Nigeria.Estimates of cysts in cattle are 1.9% inBrazil, 8.3–28.8% in Botswana, 0.1% inChile, 1.3% in Bulgaria, 0.72% overall inEgypt, 3.5–6.0% in northern Germany,0.43–15.9% in Kenya, 3.2% in Poland,1.7% in Sudan, 29% in Swaziland, 4.5% inSwitzerland, 0.6–16% in Tanzania,6.8–59% in Togo, 0.5–88% in Texas, USA,1.6% in Vietnam and 3.2% in Zimbabwe.Raw minced beef or very rare steaks arepopular items of diet in many countries.
The eggs can survive on pastures for60–180 days, in sewage for 16 days at18°C. and in river water for many weeksand they can survive for up to 1 h at 50°C.(Muller, 1983). In Russia eggs withstandthe winter better than summer (at the opti-mal temperature of �4°C, they survive for62–64 days). Moisture is a very importantfactor governing their survival and they dierapidly when desiccated (Muller, 1983).
Infection of cattle occurs mainly whenthey are young calves, sometimes from thehands of stockmen, but more usually fromfaecal matter spread on pastures.Proglottides can also migrate out through the
The Cestodes 75

anus and may contaminate pastures when aninfected person walks over them. In devel-oped countries transmission is primarily dueto inadequate treatment of sewage. Eggs arevery resistant and can survive the activatedsludge process; thorough drying or sand fil-tration is necessary to remove them. In someparts of the world improperly treated efflu-ent finds its way back to the fields.
The role of birds, particularly seagulls, inthe dissemination of T. saginata eggs fromsewage beds to fields has not been clearlyestablished but they have been implicatedin Denmark and England. Flies have alsobeen implicated in Russia and in one survey4.8% of all flies, particularly sarcophagidflies, were found to be infected.
Cysts in calves survive for longer (for upto 2 years) than those in mature cattle(maybe as little as 3 weeks). Cysticerci incattle are economically very important andare estimated to cost $1000 million annu-ally in Africa. In addition to cattle, cys-ticerci occur in buffalo and have beenreported from reindeer in Siberia, fromwild game animals in Africa and from pigsin Asia and Africa (but see below).
Taenia saginata asiatica Fan, Lin, Chenand Chung, 1995, is a recently named sub-species, in which cysticerci are found in theliver and mesenteries of pigs and wild boars,as well as in cattle (and goats and monkeys),in Burma (Myanmar), China, Indonesia(western islands), Korea (Cheju Island) andTaiwan (Ito, 1992); a similar form has beenreported from areas of Africa. The cysticer-cus has a sunken rostellum with two rows ofrudimentary hooks and there are manybranches to the uterus in gravid proglottides(Fan et al., 1995). It has been estimated tocause an annual economic loss of $34 mil-lion in Indonesia, Korea and Taiwan (Fan,1997). Cysticercosis can also occur inhumans, as with T. solium. In Taiwan, cystsof T. s. saginata, as well as T. s. asiatica, candevelop in pigs as well as in cattle.
PREVENTION AND CONTROL
Personal protection is by thorough cookingof all beef and pork (above 56°C.) or byfreezing at �10°C. for 10 days. Meat
inspection is the most important public-health measure possible. The most heavilyinfected muscles for inspection may varyfrom region to region but usually includethe heart, masseters, shoulder muscle andtongue, and also the liver of cattle or pigsin Asia. The cysts can be easily recognizedin ‘measly beef’ (or pork) about 6 weeksafter infection.
Infection rates have been reduced some-what in recent years in eastern Europe andEast Africa by sanitation measures, andmass diagnosis and treatment campaignshave been tried in Russia.
ZOONOTIC ASPECTS
In the strict sense this is an obligatorycyclo-zoonotic infection, the life cycle nec-essarily involving both cattle (or pig) andhumans (sometimes termed a euzoonosisor an anthropozoonosis). However, there isno animal reservoir host of the adult tape-worms, which occur only in humans.
Taenia solium Linnaeus, 1758
SYNONYM
Taenia cucurbitina Pallas, 1766.
DISEASE AND POPULAR NAME
Taeniasis or taeniosis solium; pork tape-worm infection.
LOCAL NAMES
As for T. saginata.
GEOGRAPHICAL DISTRIBUTION
Taenia solium infection has a cosmopolitandistribution in countries where pork is eatenraw or undercooked. It is most common inAfrica, Central and South America (particu-larly Mexico and Chile) and China.Prevalences fell greatly in eastern Europeancountries but with less efficient control arebeginning to rise again.
LOCATION IN HOST
The scolex is embedded in the mucosa ofthe jejunum, with the rest of the tapeextending through the ileum.
76 Chapter 2

The C
estodes77
Map 5. Distribution of Taenia solium (and cysticercosis) on a country basis.
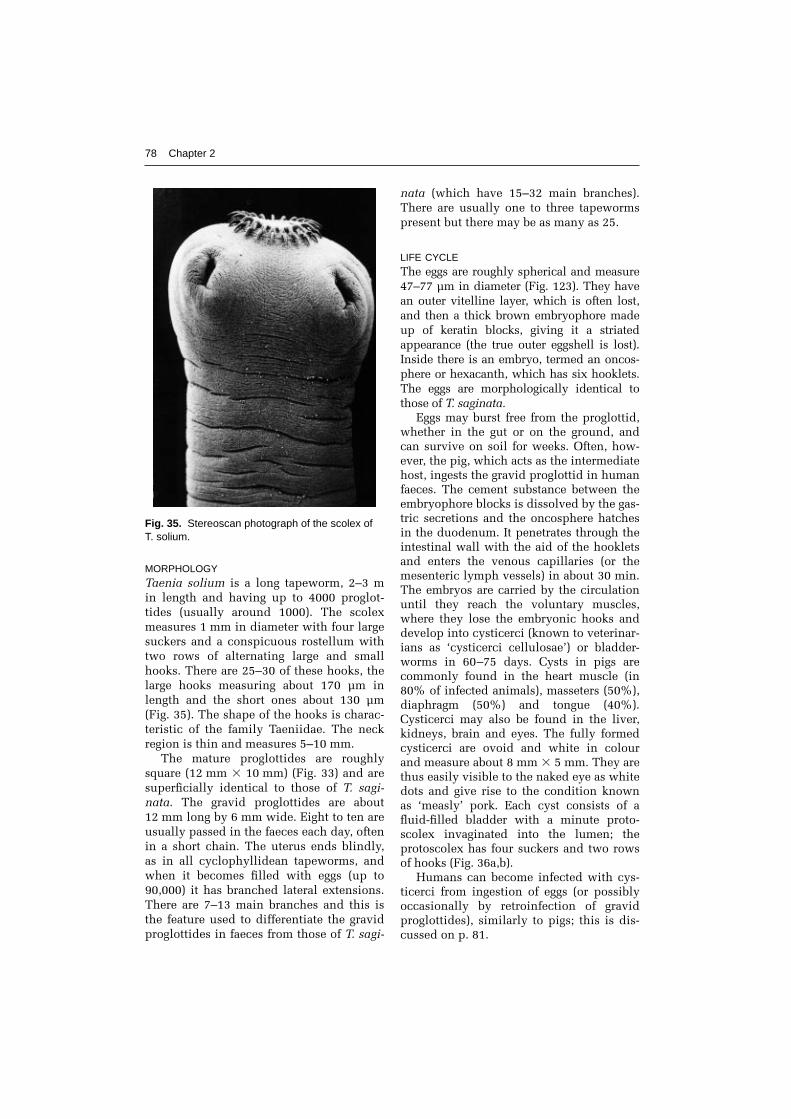
MORPHOLOGY
Taenia solium is a long tapeworm, 2–3 min length and having up to 4000 proglot-tides (usually around 1000). The scolexmeasures 1 mm in diameter with four largesuckers and a conspicuous rostellum withtwo rows of alternating large and smallhooks. There are 25–30 of these hooks, thelarge hooks measuring about 170 µm inlength and the short ones about 130 µm(Fig. 35). The shape of the hooks is charac-teristic of the family Taeniidae. The neckregion is thin and measures 5–10 mm.
The mature proglottides are roughlysquare (12 mm � 10 mm) (Fig. 33) and aresuperficially identical to those of T. sagi-nata. The gravid proglottides are about12 mm long by 6 mm wide. Eight to ten areusually passed in the faeces each day, oftenin a short chain. The uterus ends blindly,as in all cyclophyllidean tapeworms, andwhen it becomes filled with eggs (up to90,000) it has branched lateral extensions.There are 7–13 main branches and this isthe feature used to differentiate the gravidproglottides in faeces from those of T. sagi-
nata (which have 15–32 main branches).There are usually one to three tapewormspresent but there may be as many as 25.
LIFE CYCLE
The eggs are roughly spherical and measure47–77 µm in diameter (Fig. 123). They havean outer vitelline layer, which is often lost,and then a thick brown embryophore madeup of keratin blocks, giving it a striatedappearance (the true outer eggshell is lost).Inside there is an embryo, termed an oncos-phere or hexacanth, which has six hooklets.The eggs are morphologically identical tothose of T. saginata.
Eggs may burst free from the proglottid,whether in the gut or on the ground, andcan survive on soil for weeks. Often, how-ever, the pig, which acts as the intermediatehost, ingests the gravid proglottid in humanfaeces. The cement substance between theembryophore blocks is dissolved by the gas-tric secretions and the oncosphere hatchesin the duodenum. It penetrates through theintestinal wall with the aid of the hookletsand enters the venous capillaries (or themesenteric lymph vessels) in about 30 min.The embryos are carried by the circulationuntil they reach the voluntary muscles,where they lose the embryonic hooks anddevelop into cysticerci (known to veterinar-ians as ‘cysticerci cellulosae’) or bladder-worms in 60–75 days. Cysts in pigs arecommonly found in the heart muscle (in80% of infected animals), masseters (50%),diaphragm (50%) and tongue (40%).Cysticerci may also be found in the liver,kidneys, brain and eyes. The fully formedcysticerci are ovoid and white in colourand measure about 8 mm � 5 mm. They arethus easily visible to the naked eye as whitedots and give rise to the condition knownas ‘measly’ pork. Each cyst consists of afluid-filled bladder with a minute proto-scolex invaginated into the lumen; theprotoscolex has four suckers and two rowsof hooks (Fig. 36a,b).
Humans can become infected with cys-ticerci from ingestion of eggs (or possiblyoccasionally by retroinfection of gravidproglottides), similarly to pigs; this is dis-cussed on p. 81.
78 Chapter 2
Fig. 35. Stereoscan photograph of the scolex of T. solium.

Human infection with the adult tape-worms occurs through eating raw or inade-quately cooked pork. The protoscolexinvaginates in the duodenum (Fig. 36b, c),and attaches to the mucosa of the small intes-tine, and the worm matures in 5–12 weeks.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Usually the presence of an adult tapewormcauses no distinct clinical symptoms orsigns. Sometimes vague abdominal pain,with diarrhoea or constipation, is found.Excessive appetite is also occasionallyreported, with loss of weight and weak-ness. Symptoms may be psychological inorigin or perhaps due to toxic waste prod-ucts produced by the worm.
There is usually slight traumatic damageto the mucosa at the site of attachment ofthe scolex, although it has been known to
perforate the wall of the intestine, veryrarely causing death.
There is often an eosinophilia of10–13%.
Far more serious than the presence of anadult tapeworm of T. solium is humaninfection with cysticerci.
DIAGNOSIS
As for T. saginata. An immunoblot assayutilizing ES antigens is specific to adult T.solium infections (Williams et al., 1999),not reacting to T. saginata or to cases ofcysticercosis. There are also specific anti-body-based dot ELISAs for diagnosis ofcysticercus antigens.
TREATMENT
The theoretical risk with T. solium infec-tion that regurgitation of proglottides
The Cestodes 79
Fig. 36. The invaginated protoscolex of a cysticercus of T. solium. That of T. saginata is identical apartfrom the absence of hooks. (a) In section. (b) In solid view, arrow indicates beginning of evagination. (c)Whole cysticercus after invagination in stomach of new host.

caused by chemotherapy will lead tohatching of the eggs and subsequent cys-ticercosis does not appear to be importantin practice. None the less, it may be advis-able to give an antiemetic and follow treat-ment with a saline purge.
Praziquantel at 5–10 mg kg�1 in a singleoral dose is usually completely effectiveagainst adult worms.
Niclosamide at an oral dose of 2 g for anadult given in a divided dose over 2 days iseffective and causes only minor side-effects, although syncope has beenreported. It appears to disturb mitochon-drial phosphorylation.
EPIDEMIOLOGY
Infection with T. solium is nowhere near ascommon as with T. saginata, presumablybecause pork is not so often eaten under-cooked as beef. In Bali, however, ‘lawal’, adish of often uncooked spiced pork is popu-lar and taeniasis is common (although thisis probably due to T. saginata asiatica). Inmany areas the recorded infection rate inpigs (and sometimes also the cysticerci ratein humans) is very much higher than therecorded infection rate of adult tapewormsin humans would indicate and it is likelythat gravid proglottides are sometimes mis-taken for those of T. saginata.
Unhygienic disposal of human faeces isthe primary means of dissemination; theeggs surviving on pastures for many weeks.In a rural area of Mexico many households(38%) have pigs and all eat pork (oftenmeasly or ‘granos’ pork). There are poorpersonal and household standards ofhygiene and pigs routinely eat human fae-ces (Sarti et al., 1992, 1994).
It is not known how long cysticerci cansurvive in the pig, but, since degeneratedcysts are rare, it is probable that theyremain viable throughout the lifetime ofthe animal.
Estimated human infection rates in areasof various countries are: Brazil 13%, Bulgaria1.4% (0.7% in 1989), Cambodia 2.2%, Chile0.1% (0.7% in 1977), China (ShandungProvince) 0.3%, Guatemala 3.5%, Honduras0.3 (large-scale survey)–17%, India (UttarPradesh) 2.0%, Indonesia (Bali) 7.1%, (Irian
Jaya) 3–42.7%, (Sumatra) 9.5%, Mexico1.5%, Nigeria (plateau) 3.8%, (eastern) 8.6%,(northern) 11.5%, Peru 3.0% (8.6% inchicharroneros or pork sellers), Reunion1.4% and Zimbabwe 12%. Cysticerci rates inpigs are: Cameroon 24.6%, Colombia 26%,Egypt 0.1%, Honduras 4.0%–27%, India(Andhra Pradesh) 4.5–8.0%, (Karnataka)7.0%, (Uttar Pradesh) 9.3–15.5%, (Assam)27.5%, Indonesia (Bali) 13%, Mexico4–33%, Nigeria (northern) 3.3%, (eastern)20%, Tanzania 13.3% and Togo 17%.
PREVENTION AND CONTROL
Thorough cooking of pork and pork prod-ucts is essential. Cysticerci are killed by atemperature of 45–50°C, but may survive inlarge pieces of meat if the centre does notreach that temperature. Deep-freezing at�10°C for 4 days kills all cysts, but theycan survive for 70 days at 0°C. Pickling andsalting is not usually effective. At 18–20°C,cysts can survive the death of the host forup to 4 weeks.
Inspection of meat at the abattoir isimportant; the underside of the tongue canbe examined in life and the intercostal andcervical muscles after slaughter.
ZOONOTIC ASPECTS
Humans are the only definitive hosts forthe adult tapeworms. Both humans andpigs are necessary for the complete lifecycle. Cysticerci can develop in wild boarsand have been found in the lar gibbon(Hylobates lar), in which adult tapewormswill also develop experimentally. Cystshave also been reported from sheep, dogsand cats but these may not be those of T.solium.
Cysticercosis
In addition to infection with adult Taeniasolium, human infection with the cysticer-cus stage normally found in pigs can alsooccur and is the cause of an important andserious disease.
SYNONYM
Taenia solium cysticercosis, cysticercuscellulosae (in pigs).
80 Chapter 2

DISEASE AND LOCAL NAME
Cysticercosis; bladder-worm infection.
GEOGRAPHICAL DISTRIBUTION
All areas where adult T. solium infectionoccurs. Estimated cysticercosis infectionrates in some endemic areas are: Benin4.0%, Brazil (Sao Paulo) 0.27%, (MinasGerais) 18.3%, China (Shandung Province)0.7%, Ecuador 14.4%, Guatemala 3.5%,Indonesia (Irian Jaya) 17–43%, Madagascar(large-scale survey) 18%, Peru 6–10% inendemic areas, Reunion 8.2%, SouthAfrica (Transkei) 5.5–13.5% and Togo(north) 2.4%. Other endemic areas areColombia, India, Korea, Mexico, NewGuinea, West Africa and Zimbabwe.
MORPHOLOGY
The fully developed cysticercus measures10 mm � 5 mm with an opaque invagi-nated scolex (protoscolex) with four suck-ers and hooks and a fluid-filled bladder; itis identical to the cyst normally found inpigs. Development to the infective stageusually takes 9–10 weeks. A characteristicfeature, apparent in sections of most ces-tode tissues, is the presence of numerouscalcareous corpuscles.
TRANSMISSION
This is by the accidental ingestion of eggsof T. solium which have been passed inhuman faeces, presumably on vegetables orin water. In Indonesia, very young childrenmay have cysts, apparently from ingestingeggs on sweet potatoes, but rarely haveadult tapeworms below the age of 5 years(Simanjuntak et al., 1997). About 25% ofpatients with cysticercosis also harbour anadult tapeworm and this led to the beliefthat autoinfection may follow the regurgita-tion of proglottides into the stomach,caused by antiperistalsis; however, there isno proof of this. It has been shown experi-mentally that oncosphere (hexacanth) lar-vae in eggs must pass through the stomachbefore they can excyst and the usual courseof self-infection is likely to be from anus tofingers to mouth or from eggs on vegetablesor perhaps in water.
CLINICAL MANIFESTATIONS
Clinical disease is usually caused by thepresence of cysts in the brain (Plate 9). Theincubation period from ingestion of eggs tothe development of symptoms of cerebralcysticercosis is normally 5–15 years,although it can be much less.
Characteristic nervous manifestationsare epilepsiform attacks of the Jacksoniantype, but minor symptoms are also very fre-quent. Cysticerci scattered throughout thebrain may produce a variety of symptoms,motor, sensory or mental, with the result-ing classical features of any focal lesion,and various focal symptoms and signs may occur in the same patient. The mostfrequent of these are transient monoplegicparaesthesia, localized anaesthesia, visualand aural symptoms, aphasia and amnesia.Convulsions and seizures are common(1.5% of cysticercosis patients in Benin,9.5–14.2% in Brazil, 14.4% in Ecuador),but there may be unconsciousness withoutconvulsions, and headaches can be severe.There is some evidence that in LatinAmerica cases of neurocysticercosis havefewer subcutaneous cysts than similarcases in other parts of the world.
PATHOGENESISThis depends very much on the localiza-tion of the cysts in the body. They are nor-mally spread throughout the body roughlyin proportion to the weight of each organ,the most important serious effects beingcaused by cysts in the central nervous sys-tem.
DIAGNOSIS
Clinical and parasitological. Palpable subcu-taneous nodules can sometimes be felt anddiagnosis confirmed by biopsy (Fig. 37),although the hooks are not always seen intissue sections.
Radiological. X-rays will only detect oldcysts that have calcified (Fig. 38). CT andMRI scans are helpful, particularly for neu-rological cases, and portable ultrasoundhas been useful in the field (Macpherson,1992).
The Cestodes 81

Serological. There have been numerousrecent studies and an ELISA using recombinant antigens was found to be both
sensitive and specific (Hubert et al., 1999).A Western blot technique, using commer-cially available affinity purified glyco-protein antigen strips, is even moresensitive.
TREATMENT
Treatment with praziquantel at 50 mg kg�1
daily for 10–15 days has some effect whilethe cysts are still alive (treatment at doublethis dose for 21 days has been used in non-responding cases). Albendazole at 15 mgkg�1 daily for 30 days and flubendazole at40 mg kg�1 daily for 10 days have also beenused, with variable results..
Neurocysticercosis may need to betreated with anti-inflammatory agents andanticonvulsants.
EPIDEMIOLOGY
It has been estimated that in Latin America,where about 75 million people live inendemic areas there are 0.4 million sympto-matic cases, with 24,000–39,000 cases inPeru in highland and high jungle areas(Bern et al., 1999); in a survey of soldiers inMexico City, 14.7% were infected (theyoften ate pork from street vendors).
Taenia crassiceps (Zeder, 1800) Rudolphi,1810, is a tapeworm of dogs and wild car-
82 Chapter 2
Fig. 38. Radiograph of the thigh region of a youngadult with numerous calcified cysticerci (these canbe visualized while still living by CT or ultrasono-graphy (US) scans).
Fig. 37. Section of cysticercus in human muscle. Arrow points to the hooks.

nivores with cysticerci in various rodents,and human cases have been reported fromCanada and the USA. The cysticercus (‘cys-ticercus longicollis’) in the intermediatehost is unusual, as it multiplies by aprocess of asexual budding and producesmany hundreds of new cysticerci. In arecent patient who also had AIDS, therewas a painful spreading tumour in the sub-cutaneous tissues containing a tapioca-likematerial with numerous cysticercus-likesmall vesicles and with a marked granulo-matous reaction (Francois et al., 1998).
Small tapeworm cysts (up to 1 mm) wererecovered post-mortem from all the deeporgans of a man in the USA who died withHodgkin’s lymphoma (Connor et al., 1976).These might be multipying metacestodes ofan animal taeniid, such as T. crassiceps, oraberrant cysticercoids of Hymenolepisnana, which behave similarly in immuno-suppressed mice (Lucas et al., 1979).
Taenia multiceps Leske, 1780
SYNONYMS
Multiceps multiceps (Leske, 1789) Hall,1910; Coenurus cerebralis (Batsch, 1780)Rudolphi, 1808 (larvae); Multiceps gaigeriHall, 1916 (larvae in goats).
DISEASE
Coenurus infection; cerebral coenuriasis orcoenuriosis; multicepsosis (adults in carni-vores).
MORPHOLOGY AND LIFE CYCLE
The adult taeniid tapeworm is a natural par-asite of dogs and wild Canidae. The strobilameasures 40–60 cm and the scolex has a ros-tellum with two rows of hooks (large hooksmeasuring 150–170 µm, small ones 90–130µm) of typical taeniid shape. The gravidproglottid has 18–26 small branches to theuterus and is passed in the dog’s faeces. Theeggs are identical to those of Echinococcusin dogs or human Taenia and measure 31–36µm in diameter; when ingested by a suitableintermediate host, the oncosphere is liber-ated and bores through the wall of the intes-tine. It is then carried by the blood (or
lymph) and normally develops only in thebrain or spinal cord. Once lodged, theembryo grows into a bladder-worm stage,known as a coenurus. This differs from acysticercus in measuring about 3 mm in cir-cumference and in having as many as 100protoscolices invaginated from the wall intothe central cavity. Each protoscolex mea-sures 3 mm in length and has 26–36 hook-lets in two rows.
The normal intermediate hosts aresheep and goats, in which the coenurus inthe brain is known as a ‘gid worm’, produc-ing the ‘staggers’, but development mayalso occur in cattle, horses and other rumi-nants, as well as humans.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
In the majority of human cases from tem-perate countries, including Australia,France, Great Britain, Russia, South Africaand the USA (six cases), the coenurus hasbeen in the brain. However, in many casesfrom tropical Africa; in Ghana, Nigeria,Senegal and Uganda, a cyst often presentsas a subcutaneous or intramuscular nodule,diagnosed as a lipoma, ganglion or neurofi-broma (Fig. 39). A papillary oedema withpunctiform haemorrhages occurs about amonth before the onset of clinical signs.Symptoms include headache and vomiting.In many human cases (over 50%) cysts aresterile, without the presence of protoscol-ices, which makes diagnosis difficult.There is no clinical way of differentiatinginfection with this parasite from cases ofcysticercosis or echinococcosis (apart fromimaging techniques).
A fatal cerebral case in a Nigerianresulted in intermittent seizures, dizziness,headaches and blindness. At autopsynumerous cysts measuring 0.5–2.0 cm werefound in both cerebral hemispheres(Malomo et al., 1990).
Treatment has been with praziquantelbut concern has been expressed that itmight cause inflammation.
In Uganda, cases in the eye may be dueto Taenia brauni (Setti, 1897), with dogsand wild carnivores as normal definitivehosts and rodents as intermediate hosts(Fig. 40).
The Cestodes 83

Taenia serialis (Gervais, 1847) is a tape-worm of the dog and other carnivores withcoenuri in herbivorous mammals (rabbitsand hares) and primates in Africa (Senegal,South Africa and Uganda), the Americas(Brazil and the USA) and Europe (Franceand the UK). Six cases have been reported
from the USA, usually in nervous tissueand sometimes with extensive involvement(Ing et al., 1998). One case was treated withpraziquantel but this caused markedinflammation. Intramuscular cysts in a manin Nigeria were ascribed to Taenia glomer-atus Railliet and Henry, 1915.
84 Chapter 2
Fig. 40. Sagittal section through an eye with a coenurus in the vitreous chamber from Uganda (possiblyof Taenia brauni).
hooks
sucker0.5 mm
Fig. 39. Section of a portion of a coenurus cyst removed from the neck of a child.

Adults of Taenia longihamatusMorishita and Sawada, 1966, have beenrecovered from the intestine of humans inJapan and Okinawa, probably from ingest-ing cysts in rabbits.
Echinococcus granulosus (Batsch, 1786) Rudolphi, 1805
SYNONYMS
Taenia echinococcus Von Siebold, 1853and many others.
DISEASE, POPULAR AND LOCAL NAME
Cystic echinococcosis (CE); hydatid dis-ease. Espespes (Turkana).
GEOGRAPHICAL DISTRIBUTION
Echinococcus granulosus is found in mostsheep- and cattle-raising areas of the world(Table 4) and there are also wild carni-vore/wild herbivore cycles. Most humancases occur in the countries bordering theMediterranean (European, Asian and NorthAfrican), sub-Saharan Africa (East Africa,Nigeria and South Africa), South America(Argentina, Bolivia, Brazil, Chile, Peru andUruguay), eastern Europe and Eurasia(Bulgaria, Central Asian Republics, Poland,Romania and Russia) and in Asia(Bangladesh, north-western and centralChina, northern India and Nepal). It wasintroduced into the western USA (Arizona,California, New Mexico and Utah) about 60years ago and is important in Australia.
LOCATION IN HOST
The larval cysts (hydatid cysts) are foundin various sites in humans, principally theliver and lungs.
MORPHOLOGY
The adult tapeworm becomes attached to thesmall intestine of dogs and other carnivoresand comprises a scolex, neck and 3–5proglottides. It is always less than 1 cm inlength, usually 3–8.5 mm. The sphericalscolex has a rostellum with 28–40 hooks intwo rows (with large hooks measuring 30–40µm in length and small hooks 22–34 µm) andfour suckers.
The first one or two proglottides are
immature, the penultimate sexuallymature, with both male and female organs,and the last gravid, with a uterus filledwith eggs. The gravid proglottid measures2.0 mm � 0.5–1.0 mm, more than half thelength of the whole tapeworm, and con-tains about 5000 eggs. In the matureproglottid there are usually 45–60 follicu-lar testes scattered throughout. The cirrussac is well developed and the commongenital pore opens, irregularly alternating,on the lateral margin. The principal mor-phological features are shown in Fig. 41and the differences between the speciesthat occur in humans in Table 5.
The egg measures 30–37 µm in diameterand is indistinguishable from that ofTaenia species found in the dog. It can sur-vive for 6–12 months on soil and is theinfective stage for humans.
LIFE CYCLE
The gravid proglottides disintegrate in theintestine and the eggs are passed out in thefaeces of the dog. Eggs can live for 200 daysin the environment at 7°C, 50 days at 21°Cbut only a few hours at 40°C. They areingested on pasture by the intermediatehost, which is usually sheep in the straintransmissible to humans, but may be otherherbivores coming into contact with dogssuch as cattle, buffalo, goat, pig or camel.Ingested eggs hatch in the duodenum, theembryophore being digested by the actionof pancreatin and trypsin, and the con-tained oncosphere probably being activatedby the action of bile salts. The releasedoncosphere uses the hooklets to penetratethe mucosa and reach a blood-vessel. It iscarried by the bloodstream to all parts ofthe body but settles most frequently in theliver (in up to 76% of cases), due to capil-lary filtering. Arrived at its final destina-tion, the hooklets disappear and a smallcyst forms. The cyst (metacestode stage)measures about 1 mm in diameter after 1month and 10–55 mm after 5 months, bywhich time brood capsules and protoscole-ces are forming. The cyst wall consists of anexternal, anucleate, laminated cuticularmembrane (composed of a chitin-like sub-stance), about 1 mm thick, and an inner
The Cestodes 85

germinal layer, 10–25 µm thick, with manynuclei (Fig. 42). The germinal layer formssmall masses, which give rise to the broodcapsules. These enlarge and 5–20 smallbuds, about 0.1 mm in diameter, appear onthe inner surface of their walls. They
develop into invaginated protoscolices withfour suckers and a double row of hooks.Many small brood capsules and freed proto-scolices are released into the fluid of theoriginal cyst and, together with calcareousbodies, form the ‘hydatid sand’ (Fig. 43). In
86 Chapter 2
Table 4. Estimates of percentage prevalence of hydatid cysts in humans and domestic animals and ofadult Echinococcus granulosus in dogs in various countries.
Hydatid cysts (%)
Country Humans Sheep and cattle Adults in dogs
ArgentinaLa Pampa 0.03Chaco 0.019 2.3
Australia 170 cases in 10.23 years 48–93 (wild)
Azerbaijan 0.0014 20 15Bulgaria 0.003–0.006 32 7Chile 0.009 5–22 8China
Xinjiang 0.044 10–89 10–406.8–18.2 locally
Gansu 12 locally 2 (4 in yaks) 7–27Ningxa 56Quinghai 7.6 locally 22–80 11–47
GermanyBavaria 0.0011
India 60Iraq 10 20–56Iran 3 2–7 3–63Israel 0.01 8Italy
North 0.002Sardinia 0.017 50
Jordan 0.0029 9.4Kazakhstan 0.0044
0.015 locallyKenya 10–16 5–30 10–72Kuwait 0.0036Libya 0.0017–2 16 (48 in camels) 30Morocco 5–45 33Nepal
Kathmandu 1.8–5.7Peru 0.009 8–46Southern Sudan 0.5–3.5Switzerland 0.0004Tanzania
Maasailand 0.011 30Tunisia 0.057–3.5Turkey 3 (Izmir) 50 8–29UruguayUSA
Florida 0.0056 60 13.2 Mid-Wales 0.0023 13 9.2

addition to the sand grain-like proto-scolices, there may be many large, thin-walled, daughter cysts inside the outerhydatid cyst. The host tissues, especially inthe liver, form an adventitious fibrous wallaround the cyst. Fertile cysts may vary insize from 10 to 300 mm. The cyst cavity isfilled with a bacteriologically sterile fluid,which is clear or slightly yellowish, with aspecific gravity less than 1.012. The fluidcontains salts (with a higher concentrationof sulphur than in other parasitic cysts) andenzymes, plus a little protein and toxic sub-stances.
The cysts are ingested in offal by dogs andthe adult worms become mature in the intes-tine of the carnivore in 6–7 weeks. Theremay be many thousands of worms in wildcarnivores in tropical Africa and Australiabut usually only a few hundred in domesticdogs (Macpherson and Craig, 2000).
CLINICAL MANIFESTATIONS
These depend very much on how manyhydatid cysts are present and where theyare found. Cysts are usually single and inabout 50% of infected adults are in theright lobe of the liver towards its lower
The Cestodes 87
Matureproglottid
testes
eggs inuterus
genitalpore
(a)
(b)
0.5 mm
Gravidproglottid
ovary
vitellaria
Fig. 41. Diagrams of adult (a) E. multilocularis and (b) E. granulosus.

88 Chapter 2
brood capsule
germinallayer
laminatedcuticular layer
outer fibrouslayer
protoscolices
Fig. 42. Section of a portion of an early hydatid cyst of E. granulosus (the outer fibrous layer consists of host tissues).
Table 5. Differences between Echinococcus species.
E. granulosus E. multilocularis E. oligarthrus
Length of adult 3.0–6.0 mm 1.2–3.7 mm 2.2–2.9 (2.5) mmSize of gravid More than half Less than half More than half
segment body length body length body lengthPosition of genital pore Behind midline On or anterior Anterior to
in gravid segment to midline midlineAverage number of testes 56 (45–65) 22 (16–26) 29 (14–46)Number of testes anterior 15.8 (9–23) 2.3 (0–5) 8.9 (3–14)
to genital poreShape of ovary Bilobed or Two lobes Two lobes
kidney-shaped united by a fine tube united by a fine tubeShape of uterus Branching of central No side-branching No side-branching
column (12–15 of central column of central columnlateral branches)
Number of rostellar hooks 32 (28–40) 28 (20–36) 35 (26–40) (larger hooks than the others)
Number of segments 3–4 3–5 3Position in intestine Middle and lower Upper and middle Upper and middle
third ileum jejunum jejunumUsual definitive Dog, coyote, dingo, Red fox, Arctic fox, Puma, bobcat,
hosts wolf, hyena, jackal cat (dog) jaguarundiUsual intermediate Ruminants Microtine rodents Agouti
hosts and insectivoresTime for development 48–61 days 30–35 days ?
to maturity in final host (i.e. eggs in faeces)
Structure of cyst Usually a thick- Thin-walled multilocular, Polycysticwalled hydatid cyst with many small hydatid
interconnected cystsRate of development Slow. Adapted Rapidly developing. Probably
to long-lived Adapted to short-lived rapidlyintermediate hosts. intermediate hosts. developingProtoscolices Infective protoscolicesafter 1–2 years after 2–3 months

surface, while the lungs are involved in upto 40% of cases. In children cysts are some-times diagnosed more often in the lungsthan in the liver.
Symptoms and signs are those of slowlyincreasing pressure in the area of the cyst,resembling a slowly growing tumour. Theincubation period is usually at least 5 yearsand, except for cysts in the brain or orbit,symptoms rarely appear before adolescence
and sometimes not for 20 years after expo-sure (Fig. 44). In a proportion of cases (pos-sibly up to 60%; Eckert et al., 2000) thepresence of the hydatid never becomesapparent during life and the cysts eventuallycalcify and are only diagnosed at autopsy.
If a cyst ruptures, allergic reactions,such as pruritus and an urticarial rash,occur. If the patient has been previouslysensitized by slow leakage of antigenic
The Cestodes 89
50 �m
(a)
(b)
calcareousbodies
sucker
hooks
Fig. 43. Protoscolex of E. granulosus free inside a cyst (‘hydatid sand’). (a) Invaginated as in cyst.(b) Evaginated in gut of a new definitive host (dog).
Fig. 44. Group of Turkana people waiting for an operation to remove hydatid cysts.

material from the cyst, there may be acuteanaphylactic shock, with gastrointestinaldisturbances, dyspnoea and cyanosis.Death may result from pulmonary oedemacaused by the shock of a cyst rupturing inthe lung, the severity of the effect possiblybeing potentiated by anaesthesia. In a fewcases the anaphylactic shock reaction pro-vides the first intimation of disease.
Eosinophilia is present in less than aquarter of cases of hydatid.
PATHOGENESIS
The pathology caused by the presence of asingle unilocular cyst depends very muchon its site in the body.
In over 90% of liver cysts, the onco-sphere is trapped in the central veins of thehepatic lobules; only occasionally does itlodge in the portal vein. The resultant cystmay be deep or superficial and it causescompression of the liver cells, which canlead to biliary stasis and cholangitis. Thecholangitis arises because of secondaryinfection, sometimes following the ruptureof a cyst into the biliary tract. Because ofcompression, the cyst may take manyshapes and often gives the mistakenimpression of being multivesicular withmany finger-like processes.
Lung cysts are more spherical thanthose in the liver because of the spongynature of the pulmonary tissues (Fig. 45).The cysts are always intracapsular andtheir rupture may result in protoscolicesbeing coughed up in the sputum.Sometimes only hooks can be seen, whichcan be recognized microscopically. Thepresence of bile in the sputum is suggestiveof a hepatic cyst that has ruptured throughthe diaphragm. Haemoptysis (blood in thesputum) can result from rupture of the pul-monary capillaries.
About 3% of hydatid cysts occur in thebrain or spinal cord. The cysts are usuallysmall and cause symptoms earlier than inother sites. Most brain cysts are in thewhite matter and are surrounded by a thinadventitious layer.
Osseous cysts, which form 1–2% of thetotal, grow very slowly and have no wall asthey do not elicit any host reaction. As a
result, many small cysts form, whichdestroy the cancellous bone, long bonessometimes collapsing without warning.
Renal cysts sometimes occur and, if theyburst into the kidney pelvis, may becomesecondarily infected, but these and pancre-atic cysts are usually silent. Splenic cystsare often palpable and cause pain and dis-comfort in the left upper quadrant.
DIAGNOSIS
Clinical and parasitological. Symptoms of aslow-growing tumour if accompanied byeosinophilia in a patient from an endemicarea are suggestive of a hydatid.Occasionally, the hydatid ‘thrill’ can beelicited by tapping an abdominal cyst.Protoscolices or hooks may be found in thesputum following rupture of a pulmonarycyst. Needle aspiration is not to be advo-cated because of the danger of producingsecondary infection or anaphylactic shock,but sometimes protoscolices are foundunexpectedly on a subsequent stainedsmear.
90 Chapter 2
Fig. 45. Bronchogram showing blockage ofbronchus caused by a cyst of E. granulosus.

Radiological. The cavity of a cyst usuallyshows up as a characteristic shadow on X-ray, although cysts in the lungs cannotalways be differentiated from tumoursunless they become detached and float free.The water-lily sign (sign of Camelotte) iscaused by air filling the space between ecto-cyst and pericyst, and the characteristicallyshaped air cyst containing debris shows upon X-ray. In the last few years many new andmore accurate non-invasive techniques havebeen used. CT is widely used (Fig. 46) andboth CT and MRI can be used for diagnosingdeep-seated cysts and also for estimating thesize of cysts (this can be useful for evaluatingchemotherapy). Thin, ring-shaped or cres-centic calcifications surround the cysts in5–10-year-old infections. Completely calci-fied cysts are not likely to be pathogenic ifcausing no symptoms. Ultrasonography (US)is another cheaper technique that can bevery helpful for diagnosis of liver cysts and aportable apparatus can be employed in thefield, making it particularly useful for epi-demiological surveys in remote areas.
Immunodiagnosis. The indirect haemaggluti-nation (HA) test with cyst fluid as antigenhas been used for the last 25 years and givesa sensitivity of about 80%. More recently,ELISA and dot ELISA techniques, usinghydatid cyst fluid (fraction B, containing anantigen of 8 kDa) as antigen, give over 90%specificity and sensitivity and a commercial
ELISA kit is available (LMD Laboratories,Carlsbad, California). Immunoglobulin G4(IgG4) subclass antibodies appear to be mostimportant in serodiagnosis. E. granulosuscan be differentiated from E. multilocularisinfections by Western blotting using specificantigens and the test does not cross-reactwith any other parasitic infection or othercondition (Ito et al., 1999).
TREATMENT
Surgery. This is still the most commonform of treatment, particularly when thecysts are large and in a site such as thebrain or heart. Great care must be takenthat the cyst wall does not burst, asreleased living protoscolices can be carriedto new sites by the bloodstream and formnew cysts (Plate 10). The cyst cavity can besterilized with an injection of 0.5% silvernitrate or 2% formalin. For further safetyan exposed portion of the cyst can be deep-frozen before removing the sterilized con-tents. The laminated membrane of theparasite usually separates readily from thefibrous host tissue reaction. No attemptshould be made to remove the adventitiousfibrous layer in the liver, because of theserious danger of bleeding. If the cyst is toolarge for closure of the cavity to be possi-ble, marsupialization may be necessary.
Pulmonary cysts are preferably removedby enucleation of the intact cyst by making
The Cestodes 91
Fig. 46. A computer-assisted tomography (CT) scan of the liver visualizing a large cyst of E. granulosus.

an incision through the adventitia, aidedby increasing the intrapulmonary pressurewith an inflated cuff tube in the trachea.Cysts in the long bones are very difficult todeal with and may call for amputation.
Removal of a cyst is followed by a recur-rence in up to 25% of cases, possiblycaused by release of protoscolices duringoperation, but more probably by increasein size of another small silent cyst whenthe first is removed. Mortality is about 2%in primary operations but is much higher ifanother operation is required.
Percutaneous aspiration under US guid-ance, followed by irrigation with hyper-tonic (20%) saline, has been used for livercysts causing obstructive jaundice andendoscopy for evacuating biliary cysts andfor irrigating the main liver cyst. Similarguided percutaneous aspiration of livercysts in general, followed by injection of95% sterile ethanol, puncture, aspiration,injection and reaspiration (PAIR) and thena course of chemotherapy is proving to bean alternative to surgery.
Chemotherapy. This is with albendazole ormebendazole at high dosages over a longperiod; the former is slightly more effectiveas it is absorbed better. Albendazole is givenat 10 mg kg�1 daily and mebendazole at 50mg kg�1 daily, both for at least 3 months.Prognosis is uncertain but about one-thirdof patients appear to be cured and abouttwo-thirds improved. Side-effects are com-mon while the drugs are in use, includingpossible liver toxicity. The drugs will prob-ably be of use before and after surgery (toprevent the growth of new cysts).Praziquantel is also being evaluated but itseffectiveness has not been proved (Craig,1994); however, there is evidence that it hasa synergistic effect when used at 25 mg kg�1
week�1 for 7–14 days and then 40–50 mgkg�1 once a week for 8 weeks together withalbendazole.
EPIDEMIOLOGYAlthough echinococcosis is not a commondisease in most endemic areas, its seriousnature and the difficulty of cure make it animportant public health problem in manycountries.
Four species of Echinococcus are gen-erally recognized (E. granulosus, E. multi-locularis, E. oligarthrus and E. vogeli) butthe characteristics of E. granulosus differgreatly in various parts of the world and indifferent intermediate hosts (Thompsonand Lymberry, 1995; Thompson et al., 1995;Eckert and Thompson, 1996; Lymberry andThompson, 1996). These are usually desig-nated as strains, but with sequence datafrom mitochondrial and nuclear genes themolecular distances between some strainscan be greater than between the recognizedspecies (Bowles et al., 1995).
The principal strains are given below,with suggested specific names in squarebrackets (Rausch, 1995; Thompson et al.,1995):
1. Dog/sheep (also goat and other herbi-vores) strain, which is cosmopolitan anduniform (European biotype) [E. granulosus].2. Wolf/wild cervids (moose and reindeer)strain in northern Alaska, Canada, Eurasiaand Scandinavia (northern sylvatic bio-type) [E. granulosus].3. Dog/cattle or buffalo strain is found inAsia, Europe and South Africa and isgenetically very distinct from the others[E. ortleppi].4. Dog/horse strain is widely distributedbut does not appear to be infective tohumans [E. equinus].5. Dog/pig (and camel?) strain in Northand East Africa and the Middle East, east-ern Europe, Russia and Mexico is alsoprobably not infective to humans [E. suis?].6. Dingo/marsupial strain in Australia andwild carnivore/wild suid strain in Africaare probably the same as dog/sheep strain[E. granulosus].
PREVENTION AND CONTROL
Personal prevention is by taking hygienicmeasures to avoid eggs voided from dogsbeing ingested, particularly by children.Control measures should include the sani-tary disposal of offal following slaughter ofsheep, goats and cattle, both in small abat-toirs and at home (often for religous festi-vals), and campaigns to treat dogs (Meslinet al., 2000). Dogs can be successfully
92 Chapter 2

treated with praziquantel (at 5 mg kg�1) orthe older arecoline hydrobromide,although reinfection is common. Infectionsin dogs can be specifically diagnosed usingan ELISA for detection of coproantigens(Craig et al., 1996). In sheep-rearing areas,the treatment of farm dogs is particularlyimportant. In North African countries,there are often high infection rates in feraldogs which feed on discarded offal, andthese might have to be killed.
The disease has been eliminated fromIceland in the last 100 years by a combina-tion of treatment of dogs and public healthmeasures, particularly by preventing offalfrom reaching dogs. There used to be veryhigh infection rates and pamphlets pointingout the importance of control, first distrib-uted in 1863, provided welcome readingmatter in the long winter nights. Changes inanimal husbandry, involving slaughtering oflambs rather than older sheep, so that cystshave no time to develop, also aided eradica-tion (the last case was diagnosed in 1960).The disease was eradicated in New Zealand50 years ago by similar measures, includingcompulsory dosing strips, followed byinfection checks for dogs, and a successfulcampaign was carried out in Tasmania.Infection has returned (or was not com-pletely eliminated) in Cyprus, which is nowin the consolidation phase, and there arecontrol campaigns under way in regions ofArgentina and Chile, the Falkland Islands,northern Kenya, Extramadura (Spain) andwestern Australia. However, prevalences arerising in most countries of eastern Europe,as controls have been relaxed.
IMMUNOLOGY OF TAPEWORM INFECTIONS AND
POSSIBILITIES FOR VACCINES
There is little or or no evidence for immu-nity in humans against adult tapewormsand reinfection occurs readily. Infectedhumans do make immune responses, how-ever, and antibodies can be demonstratedin serum. Immunity can be demonstratedagainst intestinal tapeworms in rodents,but the underlying mechanisms are stillnot clear. Much more is known aboutimmunity and immune responses to larval
cestodes. There have been many experi-mental studies on taeniid metacestodes indomestic animals, such as sheep, as well asin rodents, and detailed studies have beenmade of hydatid disease and cysticercosisin humans. Humans infected with theseparasites make strong antibody responsesand these can be used in diagnosis. Theantigens of hydatid cysts and the cysticerciof T. solium have been analysed in detail;two (antigen 5 and antigen B) can be usedas diagnostic antigens for both conditions.The immunological relationships betweenhydatid cysts and the host are complex.There is evidence for antiparasiteresponses, and for specific and non-specificimmune suppression. Established cysts cansurvive in hosts that are resistant to incom-ing larvae (concomitant immunity) and evi-dence from animal studies suggests thatresistance operates against larvae at anearly stage, particularly invading oncos-pheres and developing cystic stages. Fullydeveloped cysts are surrounded by thickprotective layers and possess a variety ofdefensive mechanisms that reduce theeffectiveness of host responses (e.g. inacti-vation of complement).
Recombinant vaccines have been devel-oped that provide protection against thecysts of Taenia ovis in sheep, T. solium inpigs and E. granulosus in sheep. The anti-gen Eg95, a specific protein on the onco-sphere coat of E. granulosus has beenexpressed as a fusion protein with solubleglutathione-S-transferase (GST) from aschistosome. Purified and injected intosheep, this fusion protein gave 98% protec-tion against subsequent egg infection for atleast 12 months in field trials in Argentinaand Australia. The vaccine, which is stablefor over 1 year, protects against the estab-lishment of cysts in sheeps; human trials arealso envisaged. A similar vaccine againstthe larvae of T. ovis, which is of veterinaryimportance with a cycle involving dogs andsheep, is already commercially available,with 98% protection against natural infec-tions in pastures, and is also likely to havefuture relevance for human tapeworm infec-tions (Lightowers et al., 2000).
The Cestodes 93

Echinococcus multilocularis (Leuckart, 1863) Vogel, 1955
SYNONYMS
Echinococcus alveolaris Klemm, 1883; E.sibiricensis Rausch and Schiller, 1954;Alveococcus multilocularis Abuladse, 1959.
DISEASE
Alveolar echinococcosis (AE).
GEOGRAPHICAL DISTRIBUTION
Infection in animals is principally holarcticand occasional human cases occur inAustria (Tyrol), central Europe, southernGermany, Switzerland, Belarus, Russia(northern tundra areas, Caucasus andSiberia), Ukraine, Central AsiaticRepublics, China, Alaska (and nine otherstates in the USA), Canada, Bering Seaislands, Japan (Hokkaido and Rebun) andnorth-western Pacific.
MORPHOLOGY
The adult tapeworms are one-third to one-half the size of E. granulosus; other differ-ences are shown in Fig. 41 and listed inTable 5.
LIFE CYCLE
This is similar to that of E. granulosusexcept that adult tapeworms are found inabout seven species of wild carnivore, prin-cipally the Arctic (Alopex lagopus) and red
(Vulpes vulpes) fox, and occasionally thedog and cat, while the usual intermediatehosts are the field mouse in Europe and thefield or red-backed vole and lemming inAlaska and Siberia (and possibly 27 otherspecies of rodent). On Rebun Island, Japan,the cat and red fox act as hosts for E. multi-locularis but cannot be infected with E.granulosus.
When natural hosts, such as the vole(Microtus) ingest eggs on vegetation passedout by the definitive host, growth of the cystin the liver is rapid, with extensions from thegerminal membrane invading the adventitiaand forming numerous, minute, intercon-nected, secondary vesicles in about 9 days.By 150 days the alveolar vesicles are almostfilled with protoscolices and the weight ofthe liver may exceed that of the rest of thebody (Fig. 47). This rapid growth is an essen-tial adaptation to a short-lived intermediatehost. In the winter, when the hosts are hiber-nating, the cysts stop developing. It is proba-ble that when they continue growing in thespring they impair the vitality of the animal,so that it is more easily caught by a carnivore.Infected rodents rarely live for more than 5–6months. Ingested protoscolices take 26–28days to develop into mature adults in theintestine of the carnivores.
CLINICAL MANIFESTATIONS
Cysts resemble a slowly growing malignantgrowth in the liver, making this the most
94 Chapter 2
Fig. 47. Section of alveolar cyst of E. multilocularis from a liver of vole. The wall is thin and protoscolicesare embedded in a semi-solid matrix (cysts in humans are almost always sterile without protoscolices).

The C
estodes95
Map 6. Distribution of Echinococcus multilocularis.

dangerous helminth infection of humans.There are no symptoms due to pressureeffects, because of the absence of a thick-walled hydatid cyst surrounded by afibrous host reaction, although there is anouter laminated cyst layer, which appearsto protect the parasite from host immuneresponses.
The parasite destroys the liver paren-chyma, bile-ducts and blood-vessels, result-ing in symptoms of biliary obstruction andportal hypertension; necrosis of the cyst,with abscess formation may also occur.Hepatomegaly is often the first sign of infec-tion. There may be wasting in the later stagesand the condition is often mistaken for a car-cinoma. The cysts often metastase via thebloodstream to form new cysts, particularlyin the lungs and brain. Clinical symptomsmay not appear for 10–15 years after infec-tion but in untreated cases there is 94%fatality within 10 years. There is aneosinophilia of over 5% in 15–20% ofpatients.
PATHOGENESIS
In humans the multilocular or alveolar cystsform pseudo-malignant growths with aspongy mass of proliferating vesiclesembedded in a dense fibrous stroma, almostalways in the liver. They differ from thehydatid cyst by the thinness of the outerlayer, by exogenous budding and usually bythe absence of protoscolices. The germina-tive membranes are hyperplastic and foldedon themselves and the vacuole is filled witha gelatinous matrix rather than with fluid.The germinal layer is often absent in humaninfections, so that there are no protoscolicespresent in 85% of cases, and many cystsatrophy and disappear. Calcified cysts maybe found in Inuit at autopsy, undiagnosedduring life. The relatively slow growth andfew germinal cells indicate that humans arenot very suitable hosts.
In humans cysts cause necrosis of thesurrounding liver tissue with an oblitera-tive endarteritis. The outer zone consists ofan inflammatory response, with the pres-ence of giant cells and jaundice, and ascitesmight result from portal hypertension.Cysts sometimes metastase and secondary
cysts may be present in the lymph glands,lungs and brain – almost always fatal.
In patients susceptible to disease, thereis a specific lymphoproliferative hostresponse to cyst antigens containing pre-dominantly CD8+ cells and a T-helper(Th2) cell response develops slowly, with astill unknown functional potential of inter-leukin 5 (IL-5). In a proportion of cases thesymptomless cysts become calcified andcontain no living cells and there is amarked host response containing primarilyCD4+ T cells in the periparasitic granuloma(Gottstein and Felleisen, 1995).
DIAGNOSIS
As for E. granulosus. US can be helpful.Serological tests using antigen obtainedfrom protoscolices (Em2 or Em18) areextremely specific. Early diagnosis is veryimportant in determining the success ofsurgery.
TREATMENT
Surgical resection of the liver is very difficultand prognosis is rather poor; however, about90% of non-resectable patients die within 10years. Liver transplantation has beenattempted in patients with advanced infec-tions (mortality rate of 30%).
Very long-term chemotherapy withalbendazole (10–15 mg kg�1 day�1) ormebendazole (50 mg kg�1 day�1) inhibitsthe growth of cysts and prevents the occur-rence of metastases. Patients have a betterquality of life and greater longevity,although disease may recur, as both thesedrugs appear to be parasitostatic, not para-sitocidal. Albendazole is better absorbedbut can occasionally be hepatotoxic, whilethere should be breaks – say, every 28 days– with both drugs to prevent leucopenia(Tornieporth and Disko, 1994).
EPIDEMIOLOGY
Prevalence rates in various countries (per100,000 population) are: Alaska 7–98,China (Gansu) 2200, Central Europe0.02–1.4, Germany (Bavaria) 0.6, northernSiberia 170, northern Russia in general 10,Switzerland 0.18. There can be high infec-tion rates with adult tapeworms in the red
96 Chapter 2

fox: Austria 3.6%, Czech Republic 10.6%,Germany 18.4%, Japan (Hokkaido)52–67%, Poland 2.6%, Slovakia 10.7%and Switzerland 35%.
The life cycle of E. multilocularis,involving wild carnivores and rodents, usu-ally involves ecosystems separate fromhumans so that human infection is muchless likely than with E. granulosus.However, the distribution appears to bespreading in Europe, Japan and NorthAmerica (Eckert et al., 2000). In Arcticregions 50% of tundra voles (Microtusoeconomus) may be infected at some timeof the year and 100% of Arctic foxes mayhave up to 200,000 adult tapeworms. Manyhuman infections are reported from furtrappers, who ingest eggs when skinningwild canids. Eggs may also be ingested onsalad vegetables during Arctic summer, asthey can withstand very low temperatures(240 days at �18°C: a temperature as low as�70°C is necessary to be sure they arekilled). In Alaska and areas of Russia over50% of dogs are infected.
In woodland and agricultural areas ofcentral Europe, human infection is thoughtto be primarily from eating fruits, berriesand vegetables (wild strawberries, cran-berries, fallen apples and lettuce) pickedoff the ground and contaminated withfaeces from foxes or dogs.
PREVENTION AND CONTROL
Thorough washing of fruit and vegetableslikely to become contaminated. Chemo-therapy of dogs is possible, as for E. granu-losus.
Echinococcus oligarthrus (Diesing, 1863) Lühe, 1910
This species is in many features intermedi-ate between the other two (see Table 5).Adults occur in the jaguarundi (Felisyagouaroundi) in Panama, the puma (Pumaconcolor) in Panama and Brazil and thebobcat (Lynx rufus) in Mexico. The inter-mediate hosts are agoutis (Dasyproctaaguti). It is an infection of tropical sylvaticareas and identified human cases havebeen reported from Brazil (Minais Gerais),
India, Panama, Surinam and Venezuela;characteristic cysts have also been reportedfrom Argentina, Chile, Costa Rica, Mexico,Nicaragua and Uruguay (these might havebeen due to E. vogeli but are outside thehost range of the bush dog, its naturaldefinitive host).
Cysts in humans are usually in the liver(80% of cases), with most of the remainderin the lungs, although there have been twoorbital infections. Clinically, cysts in theliver have caused abdominal pain,hepatosplenomegaly, marked weight lossand fever, with the presence of hard, round,masses. Ascites and obstructive jaundicecan also occur. Portal hypertension is foundin about 25% of cases and often results indeath following biliary drainage or partialhepatectomy. The cysts are of the hydatidtype but form polylobed masses (polycysticechinococcosis (PE) spilling into the peri-toneal cavity (Plate 11); the rostellar hooksare larger than those of other species.
Treatment is with albendazole at 10 mgkg�1 daily for 2–8 months and has givenencouraging results in most cases. Side-effects stop when treatment is finished.
Echinococcus vogeli Rausch and Bernstein, 1972
Adults of this species are found in the bushdog (Spetheos venaticus), with cysts in thepaca (Cuniculus paca), in South andCentral America. Human cases have beenreported from Argentina, Brazil (Amazon),Ecuador and Surinam. Infection is veryrare in humans, partly because the bushdog avoids human habitations, and mayresult from dogs that have been fed on theviscera of pacas. The polycystic hydatidcysts (causing PE) are similar to those of E.oligarthrus but the outer membrane isthicker and the rostellar hooks on theprotoscolices are smaller and morphologi-cally distinct. Clinical effects are similaralso, with both exogenous budding of cystsinto the peritoneal cavity which grow inother organs and also endogenous buddingwith daughter cysts inside the main cyst.Treatment with albendazole at 10 mg kg�1
daily for up to 10 months has some effect.
The Cestodes 97

Family Hymenolepididae
Hymenolepis nana (Von Siebold, 1852) Blanchard, 1891
SYNONYMS
Taenia nana Von Siebold, 1852;Hymenolepis fraterna Stiles, 1906;Vampirolepis nana.
DISEASE AND POPULAR NAME
Hymenolepiasis or hymenolepiosis nana;dwarf tapeworm infection.
GEOGRAPHICAL DISTRIBUTION
Cosmopolitan, particularly in children.
LOCATION IN HOST
The adults inhabit the lumen of the upperthree-quarters of the ileum, with the scolexembedded in the mucosa. There may bemany thousands of tapeworms present in aheavy infection.
MORPHOLOGY
Hymenolepis nana is a very short tape-worm, measuring 15–40 mm �0.5–1.0 mm, and has approximately 200proglottides.
The scolex is spherical, about 0.3 indiameter (one-third the size of that of
Taenia solium) and has a short retractilerostellum with a single row of 20–30 smallhooks and four round suckers or acetabula.The hooks are spanner-shaped, which istypical of the family Hymenolepidiidae.The neck region is long and slender (Fig.48). Mature proglottides are broader thanlong (0.22 mm � 0.85 mm), with a singlecommon genital pore on one side of thestrobila or chain.
There are three round testes, which liein the posterior part of each proglottid. Thebilobed, coarsely granular, ovary lies post-eriorly between the testes with the compactvitelline gland behind it (Fig. 49). Thegravid uterus forms a sac filled with80–200 eggs.
LIFE CYCLE
This is the only known human tapewormthat does not require an intermediate host.
The gravid proglottides are usually bro-ken up in the colon so that free eggs arefound in the faeces. The eggs measure30–47 µm (Fig. 123). They are spherical orovoid and are enclosed in two transparentmembranes, the inner of which, theembryophore, is lemon-shaped and haspolar thickenings, from which arise 4–8slender filaments (these are not always evi-
98 Chapter 2
0.1 mm
H. nana H. diminuta
Fig. 48. Diagrams of the scolices of Hymenolepis nana and H. diminuta (see also Fig. 33).

dent when the eggs are preserved). Theenclosed oncosphere measures about 18µm in diameter and has the usual six hook-lets. The egg is immediately infective.
When ingested by another person, theoncosphere is liberated in the small intes-tine and penetrates a villus by means of thehooklets. It then loses the larval hookletsand becomes a tailless cysticercoid (some-times known as a cercocystis) in about 90h. This stage re-enters the intestinal lumenafter 4 days and develops into a tapewormwith mature proglottides in 10–12 days(Fig. 50). Eggs are passed in the faecesabout 30 days after infection.
Autoinfection is possible, eggs and freedoncospheres having been obtained byintestinal aspiration from an infected child.This accounts for reports of infections last-ing many years although, in childrenremoved from an endemic area, the adulttapeworms live for only a few months.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
In most cases no symptoms can be attrib-uted to a light infection but the presence ofmore than 2000 worms results in enteritis,
with abdominal pain, diarrhoea, irritabil-ity, loss of appetite, vomiting and dizzi-ness, presumably due to systemic toxaemiacaused by the waste products or to allergicresponses. Epileptoid fits have beenreported from children. Necrosis of themucosa has been found in rats but has notbeen demonstrated to occur in humans.
An eosinophilia of up to 15% occurs inabout 7% of infected individuals.
DIAGNOSIS
This is by the identification of eggs in thefaeces (p. 254 and Fig. 123).
TREATMENT
Praziquantel at a single dose of 15 mg kg�1
has about 80% effectiveness; the presenceof cysticercoids in the tissues may necessi-tate re-treatment.
Niclosamide can be given at 60–80 mgkg�1 for 5 days and perhaps repeated 10days later.
EPIDEMIOLOGY
H. nana is more common in children than inadults, particularly children from institutions
The Cestodes 99
vitellariaovary
cirrus
testis
0.1
mm
0.3
mm
0.25
mm
1 m
m
Fig. 49. Mature proglottides of H. nana (left) and H. diminuta (right). Note the different scales – theproglottides of H. diminuta are comparatively much larger than shown.

in tropical countries. Estimates in childrenin regions of various countries are:Argentina 10–15%; Bahrain 3%; Brazil(Amazon) 11%, (Minais Gerais) 7%, (Rio)1.4%, (Sao Paulo) 9–15%; Chile 0.4–4%;Côte d’Ivoire 1–7%; Egypt 6–20%; Ethiopia0.6–1%; Honduras 8%; India 1–12%; Japan2.3%; Mexico 3–5%; Nepal 7%; Pakistan7–18%; Peru 23%; Romania 5%; Sudan7–10%; Tunisia 2%; Turkey 6–14%;Uganda 0.5%; Venezuela 6–12%;Zimbabwe 21%. Large-scale estimates inthe general population are: Albania 1.5%;Australia (aboriginals) 55%; Bolivia 9%;Cuba (overall) 0.008%; DominicanRepublic 2%; Ecuador 3–4%; Egypt (Cairo)0.2%; Iraq 7%; Kenya 5%; Korea0.03–0.4%; Madagascar 9%; Mexico 5%;Namibia 1%; Nigeria (Plateau) 0.3–3%;Russia (Siberia) 0.1%; Saudi Arabia0.4–3%; Senegal 1.5%; Turkey 0.6–2%;Venezuela 2%.
The eggs are not resistant to heat or des-iccation and are viable for a maximum of
11 days. Infection is often from hand tomouth, although it may also be by food orwater.
PREVENTION AND CONTROL
Hygienic measures are not always immedi-ately effective and eradication has beenvery difficult in the past. As the eggs haveso little resistance to desiccation, measurescan be directed against direct hand-to-mouth transmission and to cleaning ofclothes and bed-linen. The much improvedchemotherapy available in the last fewyears makes control considerably easier.
ZOONOTIC ASPECTS
The role of animals in the spread of humaninfection is not clear. A morphologicallyidentical parasite, H. nana var. fraterna, iscommon in rats and mice and often has asintermediate hosts either fleas (Xenopsyllacheopis, Ctenocephalides canis) or beetles(Tenebrio molitor, Tribolium confusatum).However, the rodent variety differs physio-
100 Chapter 2
Fig. 50. Diagrammatic section of adult and cysticercoid of H. nana in a villus of the ileum. In heavy iinfections the mucosal lining may become abraded.
0.3 mm

logically and does not usually cause infec-tion in humans. The situation is furthercomplicated because the human tapewormcan also develop in insects and these aresubsequently infective when ingested, andpossibly rodents sometimes act as reservoirhosts of the human strain without the pres-ence of an intermediate host.
Hymenolepis diminuta (Rudolphi, 1819) Blanchard, 1891
SYNONYMS
Taenia diminuta Rudolphi, 1819; Taeniaflavopunctata Weinland, 1858; Taenia min-ima Grassi, 1886.
DISEASE AND POPULAR NAME
Hymenolepiasis or hymenolepiosis dimin-uta; the rat tapeworm.
GEOGRAPHICAL DISTRIBUTION
This is a common and cosmopolitan para-site of rats, mice and wild rodents in manyparts of the world. It is an occasional para-site of humans, most cases being reportedfrom children. In a few countries infectionin children is not uncommon (1.9% in theNew Guinea highlands; 1.3% in India(Uttar Pradesh); 0.4% in Morocco (Agadir);0.05–0.1% in Turkey).
LOCATION IN HOST
Adults are found in the upper three-quar-ters of the ileum, with the scolex usuallyembedded in the mucosa. However, in ratsit has been shown that the tapewormmoves backwards and forwards in theintestine and this may be generally true ofthis and other tapeworms in humans.
MORPHOLOGY
In spite of its name this species is consider-ably larger than H. nana. The entire tape-worm measures 300–600 mm � 4 mm andhas 800–1000 proglottides. The scolex(0.2–0.4 mm in diameter) is spherical andhas four small suckers and a retractablerostellum; but no hooks (Fig. 48).
The mature proglottides are broaderthan long and measure 0.75 mm � 0.25 mm
(Fig. 49). The reproductive organs are simi-lar in structure to those of H. nana,although the three testes are more widelyseparated.
The egg is slightly ovoid and has a thickyellow outer shell and a thin colourlessinner membrane (embryophore), with agranular intermediate layer. There are nopolar filaments. The egg measures 60–80µm in maximum diameter (Fig. 123).
LIFE CYCLE
For further development, the eggs passedin the faeces need to be ingested by aninsect intermediate host. They can survivein the environment for about 1 week.
The most important intermediate hostsare flour beetles (Tribolium confusatumand Tenebrio molitor) and rat fleas(Xenopsylla cheopis and Nosophyllus fas-ciatus). When ingested, often on contami-nated grain, the oncosphere larva insidethe egg develops into a tailed, solid, cysti-cercoid larva in the body cavity of theinsect. If the infected insect is ingested by arat (or a child), the liberated cysticercoidsattach to the intestinal mucosa and theworm matures in 18–20 days. There is notthe invasive tissue phase characteristic ofH. nana.
Infection in children appears to causeno clinical symptoms apart from diarrhoea.Some cases have been referred for neuro-psychiatric problems but these may becoincidental. Diagnosis is by finding thecharacteristic eggs in the faeces, and treat-ment is as for H. nana; repeated doses haveproved necessary in some cases. A singledose of 500 mg of niclosamide is an oldertreatment.
Although H. diminuta has little directmedical importance, it is of great scientificinterest as a laboratory model, and is beingused in many research studies on the phys-iology, biochemistry and immunology ofcestodes and for chemotherapeutic screen-ing tests.
Drepanidotaenia lanceolata (Bloch,1782) Railliet, 1892, is a hymenolepid par-asite of water-birds with cysticercoids incopepods and with a single case in a boy inGermany.
The Cestodes 101

Family Dipylididae
Dipylidium caninum (Linnaeus, 1758) Railliet, 1892
SYNONYMS, DISEASE AND POPULAR NAME
Taenia canina L., 1758 and many others;dipylidiasis or dipylidiosis; double-poreddog tapeworm.
GEOGRAPHICAL DISTRIBUTION
This is a very common and cosmopolitanparasite of dogs and cats and is foundsporadically in humans; a few hundredcases having been reported, almost all inchildren. Recent reported cases in childrenand infants have been from Austria,Bulgaria, Chile, China, India, Italy, Japan,Russia, Sri Lanka and the USA.
MORPHOLOGY
Dipylidium caninum is a short tapewormmeasuring 200–400 mm � 2.5–3 mm andhas 60–175 proglottides. The scolex mea-sures about 0.37 mm, has four prominentsuckers, a retractable rostellum and 30–150thorn-shaped hooks in 3–4 rows. Matureproglottides are longer than broad. Thegenital organs are in two sets, one set eachside of the midline. The two common geni-tal pores open at the centre of both lateralmargins.
The gravid proglottides are filled witheggs in egg capsules (Fig. 124), each con-taining up to 20 eggs, proglottides arepassed in the faeces. In shape, the proglot-tides are an elongate oval, measuring12 mm � 2.7 mm, and are very active whenpassed; frequently they are mistaken fortrematodes when seen in the faeces of dogsor children.
LIFE CYCLE
The egg capsules are liberated on theground or stick to the anal hairs of dogs.They cannot survive drying for more than1–2 days and live only 1–2 minutes inwater. The eggs are ingested by the larvalstages of ectoparasitic insects, such asCtenocephalides canis or C. felis (the dogand cat fleas) and develop in the bodycavity into tailed cysticercoids in about 18days.
Human infection is presumably fromingestion of fleas and is almost always inchildren. There is no definite pathogenesisassociated with this species and treatmentis with praziquantel (25 mg kg�1 in a singleoral dose). Diagnosis is by finding themobile, gravid, barrel-shaped, proglottides(with remains of double organs or pores) oregg capsules in the faeces (Fig. 124).
Very occasional human tapeworms
Bertiella studeri (Blanchard, 1891) Stilesand Hassall 1902 and B. mucronata
(Meyner, 1895) Stiles and Hassall, 1902 (family Anoplocephalidae)
These are parasites of monkeys with cysti-cercoids in oribatid mites. About 60 casesof B. studeri in humans, mostly in children,have been reported from Equatorial Guinea,India, Indonesia, Japan, Kenya, Malaysia,Mauritius, Nigeria, Philippines, Sri Lanka,Thailand and Yemen and of B. mucronatain Argentina, Brazil, Cuba and Paraguay(Denegri and Perez-Serrano, 1997). A chainof thick, very broad proglottides passed inthe faeces usually provides the first evi-dence of infection. A woman in EquatorialGuinea had suffered abdominal discomfortfor 6 years before proglottides were passedand she was cured by 40 mg kg�1 of prazi-quantel (Galan-Puchades et al., 1997).
Inermicapsifer madagascariensis (Davaine, 1870) Baer, 1956 (syn. I. cubensis)
(family Anoplocephalidae)
This is a small tapeworm (5 cm) of rodentsand hyraxes with cysticercoids presumablyin mites. Human cases have occurred inCuba, Comoros Islands, Congo, Kenya,Madagascar, South Africa, Venezuela,Zambia and Zimbabwe. It is not uncom-mon in children around Havana, Cuba, andis probably more common in East Africathan reported.
Mathevotaenia symmetrica (Baylis,1927) Akhumian, 1946, is widely distrib-uted in rodents in Thailand, with cysticer-
102 Chapter 2

coids in flour beetles and moths (and onecase in a human).
Mesocestoides lineatus (Goeze, 1782)Railliet, 1893 and M. variabilis Mueller, 1927
(family Mesocestoididae)
M. lineatus is a parasite of carnivores prob-ably with an arthropod as the first interme-diate host and amphibians, reptiles, smallmammals and birds as second (with a mod-ified cysticercoid known as a tetrathyrid-ium, of the plerocercoid type). Humancases have been reported from China (twocases), Japan (12), Korea (one) and SriLanka (one). The man infected in Korea ateraw chicken viscera and passed 32 wormsafter treatment with niclosamide. M. vari-abilis has a similar life cycle and humancases have been reported from Greenlandand the USA (6 cases).
Raillietina spp. (family Davaineidae)
Species found in humans include R. asiat-ica (Von Linstow, 1901) Stiles and Hassall(a single case in Russia, a doubtful case inIran); R. celebensis (Janicki, 1902)Fuhrmann, 1924, a parasite of rats andmice with cysticercoids in ants (a few casesin children from Australia, China, Japan,Moorea, Philippines, Tahiti and Taiwan);R. garrisoni Tubangui, 1931 (one case in achild in the Philippines); R. madagas-cariensis (Davaine, 1869) Joyeux and Baer,1919, a common parasite of rats in Asiawith cysticercoids probably in cockroaches(human cases from Comoros, Indonesia,Mauritius, Philippines and Thailand); R.siviraji Chandler and Pradatsundarasar,1957, a common parasite of rats inThailand (and with a few cases in chil-dren). Buginetta (= R.) alouattae Spaskii,1994 is found in South American monkeysand occasionally humans.
The Cestodes 103
References
Beaver, P.C. and Rolon, F.A. (1981) Proliferating larval cestode in a man in Paraguay: a case reportand review. American Journal of Tropical Medicine and Hygiene 30, 625–637.
Bern, C., Garcia, H.H., Evans, C., Gonzalez, A.E., Verastegul, M., Tsang, V.C.M. and Gilman, R.H.(1999) Magnitude of the disease burden from neurocysticercosis in a developing country.Clinical Infectious Diseases 29, 1203–1209.
Bessenov, A.S., Movsession, S.O. and Abuladze, K.I. (1994) On the classification and validity ofsuperspecies taxons of the cestodes of the suborder Taeniata Skrjabin et Schulz, 1937.Helminthologia 31, 67–71.
Bowles, J., Blair, D. and McManus, D.P. (1995) A molecular phylogeny of the genus Echinococcus.Parasitology 110, 317–328.
Chang, K.H., Chi, J.G., Cho, S.Y., Han, M.H., Han, D.H. and Han, M.C. (1992) Cerebral sparganosis:analysis of 34 cases with emphasis on CT features. Neuroradiology 34, 1–8.
Connor, D.H., Sparks, A.K., Strano, A.J., Neafie, R.C. and Juvelier, B. (1976) Disseminated parasitosisin an immunosuppressed patient – possibly a mutated sparganum. Archives of PathologicalLaboratory Medicine 100, 65–68.
Craig, P.S. (1994) Current research in echinococcosis. Parasitology Today 10, 209–211.Craig, P.S., Rogan, M.T. and Allan, J.C. (1996) Detection, screening and community epidemiology of
taeniid cestode zoonoses: cystic echinococcosis, alveolar echinococcosis and neurocysti-cercosis. Advances in Parasitology 38, 170–250.
Denegri, G.M. and Perez-Serrano, J. (1997) Bertiellosis in man: a review of cases. Revista do Institutode Medecina Tropical de Sao Paulo 39, 123–127.
Eckert, J. and Thompson, R.C.A. (1996) Intraspecific variation of Echinococcus granulosus andrelated species with emphasis on their infectivity to humans. Acta Tropica 64, 19–34.
Eckert, J., Conraths, F.J. and Taekmann, K. (2000) Echinococcosis: an emerging or re-emerging zoo-nosis? International Journal for Parasitology 30, 1283–1294.
Fan, P.C. (1997) Annual economic loss caused by Taenia saginata asiatica taeniasis in East Asia.Parasitology Today 13, 194–196.

Fan, P.C., Lin, C.Y., Chen, C.C. and Chung, W.C. (1995) Morphological description of Taenia saginataasiatica (Cyclophyllidea: Taeniidae) from man in Asia. Journal of Helminthology 69, 299–303.
Francois, A., Favennec, L., Cambon-Michot, C., Gueit, I., Biga, N., Tron, F., Brasseur, P. and Hemet, J.(1998) Taenia crassiceps invasive cysticercosis: a new human pathogen in acquired immuno-deficiency syndrome? American Journal of Surgical Pathology 22, 488–492.
Galan-Puchades, M.T., Frentes, M.V., Simarro, P.P. and Mas Coma, S. (1997) Human Bertiella studeriin Equatorial Guinea. Transactions of the Royal Society of Tropical Medicine and Hygiene 91,680.
Gottstein, B. and Felleisen, R. (1995) Protective immune mechanisms against the metacestode ofEchinococcus multilocularis. Parasitology Today 11, 320–326.
Hubert, K., Andriantsimahavandy, A., Michault, A., Frosch, M. and Muhlschlegel, F.A. (1999)Serological diagnosis of human cysticercosis by use of recombinant antigens from Taeniasolium cysticerci. Clinical and Diagnostic Laboratory Immunology 6, 479–482.
Ing, M.B., Schantz, P.M. and Turner, J.A. (1998) Human coenurosis in North America: case reportsand review. Clinical and Infectious Diseases 27, 519–523.
Ito, A. (1992) Cysticercosis in Asian-Pacific regions. Parasitology Today 8, 182–183.Ito, A., Ma-LiAng, Schantz, P.M., Gottstein, B., Liu-Yuetlan, Chai JunJie, Abdel-Hafez, S.K., Altintas,
N., Joshi, D.D., Lightowers, M.W., Pawlowski, Z.S., Ma-La, Liu, Y.H. and Chi, J.J. (1999)Differential serodiagnosis for cystic and alveolar echinococcosis using fractions of Echinococcusgranulosus cyst fluid (antigen B) and Echinococcus multilocularis protoscolex (Em18).American Journal of Hygiene and Tropical Medicine 60, 188–192.
Khalil, L.F., Jones, A. and Bray, R.A. (1994) Keys to the Cestode Parasites of Vertebrates. 768 pp. CABInternational, Wallingford, UK.
Lightowers, M.W., Flisser, A., Gauci, C.G., Heath, D.D., Jensen, O. and Rolfe, R. (2000) Vaccinationagainst cysticercosis and hydatid disease. Parasitology Today 16, 191–196.
Lo, Y.-K., Chao, D. and Yan, S.-H. (1987) Spinal cord proliferative sparganosis in Taiwan: a casereport. Neurosurgery 21, 235–238.
Lucas, S.B., Hassounah, O.A., Doenhoff, M. and Muller, R. (1979) Aberrant form of Hymenolepisnana; possible opportunistic infections in immunosuppressed patients. Lancet 22 Dec. (ii),1372–1373.
Lymberry, A.J. and Thompson, R.C.A. (1996) Species of Echinococcus: pattern and process.Parasitology Today 12, 486–491.
Machnicka, B., Dziemian, E. and Zwierz, C. (1996) Detection of Taenia saginata antigens in faeces byELISA. Applied Parasitology 37, 99–105 and 106–110.
Macpherson, C.N.L. (1992) Ultrasound in the diagnosis of parasitic disease. Tropical Doctor 22,14–20.
Macpherson, C.N.L. and Craig, P.S. (2000) Dogs and cestode zoonoses. In: Macpherson, C.N.L.,Meslin, F.X. and Wandeler, A.I. (eds) Dogs, Zoonoses and Public Health. CAB International,Wallingford, UK, pp. 177–212.
Malomo, A., Ogunniyi, J., Ogunniyi, A., Akang, A., Akang, E.E.U. and Shokunbi, M.J. (1990)Coenurosis of the central nervous system in a Nigerian. Tropical and Geographical Medicine 42,280–282.
Meslin, F.X., Miles, M.A., Vexenat, J.A. and Gemmell, M.A. (2000) Zoonoses control in dogs. In:Macpherson, C.N.L., Meslin, F.X. and Wandeler, A.I. (eds) Dogs, Zoonoses and Public Health.CAB International, Wallingford, UK, pp. 333–372.
Muller, R. (1983) Taenia, taeniasis and cysticercosis. In: Feachem, R.G., Bradley, D.J., Garelick, H.and Marr, D.D. (eds) Sanitation and Disease: Health Aspects of Excreta and WastewaterManagement. John Wiley & Sons, Chichester, UK, pp. 463–472.
Rausch, R.L. (1995) Life cycle patterns and geographic distribution of Echinococcus species. In:Thompson, R.C.A. and Lymberry, A.J. (eds) Echinococcus and Hydatid Disease. CABInternational, Wallingford, UK, pp. 89–119.
Ruttenber, D.J., Weniger, F., Sorvillo, R., Murray, R.A. and Ford, S.L. (1984) Diphyllobothriasis asso-ciated with salmon consumption in Pacific coast states. American Journal of Tropical Medicineand Hygiene 33, 455–459.
Sarti, E., Schantz, P.M., Plancarte, A., Wilson M., Gutierrez O., I., Lopez, A., Roberts, J. and Flisser, A.(1992) Prevalence and risk factors for Taenia solium taeniasis and cysticercosis in a village inMorelos, Mexico. American Journal of Tropical Medicine and Hygiene 46, 677–685.
104 Chapter 2

Sarti, E., Schantz, P.M., Plancarte, A., Wilson, M., Gutierrez, O., I., Aguilera, J., Roberts, J. and Flisser,A. (1994) Epidemiological investigation of Taenia solium taeniasis and cysticercosis in a ruralvillage of Michoacan State, Mexico. Transactions of the Royal Society of Tropical Medicine andHygiene 88, 49–52.
Sen, D.K., Muller, R., Gupta, V.P. and Chilana, J.S. (1989) Cestode larva (sparganum) in the anteriorchamber of the eye. Tropical and Geographical Medicine 41, 270–273.
Simanjuntak, G.M., Margono, S.S., Okamoto, M. and Ito, A. (1997) Taeniasis/cysticercosis inIndonesia as an emerging disease. Parasitology Today 13, 321–322.
Smyth, J.D. and McManus, D.P. (1989) The Physiology and Biochemistry of Cestodes. CambridgeUniversity Press, Cambridge, UK.
Thompson, R.C.A. and Lymberry, A.J. (eds) (1995) Echinococcus and Hydatid Disease. CABInternational, Wallingford, UK.
Thompson, R.C.A., Lymberry, A.J. and Constantine, C.C. (1995) Variation in Echinococcus: towards ataxonomic revision of the genus. Advances in Parasitology 35, 145–176.
Tornieporth, N.G. and Disko, R. (1994) Alveolar hydatid disease (Echinococcus multilocularis) –review and update. In: Sun, T. (ed.) Progress in Clinical Parasitology Vol. 4. Norton, New York,USA, pp. 55–76.
von Bonsdorff, B. (1977) Diphyllobothriasis in Man. Academic Press, London, UK.Williams, P.P., Allan, J.C., Verastegui, M., Acosta, M., Eason, A.G., Garcia, H.H., Gonzalez, A.E.,
Gilman, R.H. and Tsang, V.C.W. (1999) Development of a serological assay to detect Taeniasolium taeniasis. American Journal of Tropical Medicine and Hygiene 60, 199–204.
The Cestodes 105

Members of the phylum Acanthocephalaare all parasitic and show similarities instructure to both the nematodes and theplatyhelminthes (Crompton and Nickol,1985). Most species are under 1 cm inlength but some may be up to 70 cm. Theprincipal diagnostic character is the pres-ence of an eversible proboscis armed withrows of hooks (thorny-headed worms).Acanthocephalans are pseudocoelomateanimals (that is, they have a body cavitybut it is not homologous to a true coelom)with separate sexes and, like cestodes,have no digestive tract. The life cycle usu-ally involves an insect or crustacean asintermediate host. Many acanthocephalanshave a low specificity for the definitivehost but are not found in humans becausethe intermediate hosts are not likely to beingested (e.g. cockroaches).
Two genera, both belonging to the classArchiacanthocephala, are occasional para-sites of humans (Taraschewski, 2000).
Moniliformis moniliformis(Bremser, 1811), Travassos, 1915
SYNONYMS
Echinorhynchus moniliformis Bremser,1811; Moniliformis dubius Meyer, 1932.
DISTRIBUTION
Moniliformis is a very occasional humanparasite. Cases have been reported fromAustralia, Bangladesh, Belize, Iran, Israel,
Italy, Japan, Madagascar, Papua NewGuinea, Russia, the USA and Zimbabwe.Infection is common and cosmopolitan inrodents and also occurs in dogs and cats.
LOCATION IN THE HOST
Adults inhabit the small intestine, the pro-boscis being deeply embedded in themucosa.
MORPHOLOGY
The adult worms appear to be made up of aseries of bead-like annulations, but theseare only superficial. The anterior proboscisis cylindrical and has 12–15 rows ofcurved hooks, with 7–8 hooks per row.Females measure 10–25 cm in length,males 4–14 cm.
LIFE CYCLE
The ovoid eggs measure 70–120 µm �30–60 µm and are passed in the faeces. Theegg has three envelopes characteristic ofmembers of the phylum and contains aspiny acanthor larva when passed. Thelarva inside can survive for months in soilbut develops further if the egg is ingestedby various beetles or cockroaches. The lar-vae require several weeks before they reachthe infective stage. The encysted juvenilestages (cystacanths) are ingested by a mammalian host (usually the rat) anddevelop into adults in 5–6 weeks.
Human infection is probably usuallyfrom grain beetles and it is of interest that15% of coprolites from grain-eating prehis-toric humans in a cave in Utah (USA)
3The Acanthocephala
106
03Worms & H.D. - Chap 03 14/11/01 4:26 PM Page 106

The Acanthocephala 107
contained acanthocephalan eggs, whichwere probably of Moniliformis.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
The clinical manifestations have beenknown since Calandruccio in 1888 infectedhimself experimentally. Nineteen daysafter ingesting cystacanth larvae, he hadgastrointestinal disturbances with diar-rhoea, exhaustion and a ringing in the ears.Eggs appeared in his faeces about 2 weekslater. In a recent case in Florida, USA, aninfant had repeated episodes of diarrhoea,accompanied by coughing and fever; aftereach episode an adult worm was passed inthe faeces (Counselman et al., 1989).
TREATMENT
Mebendazole and pyrantel pamoate havebeen used in recent cases with variableresults.
Macranthorhynchus hirudinaceus(Pallas, 1781) Travassos, 1917
SYNONYMS AND POPULAR NAME
Taenia hirudinaceus Pallas, 1781;Gigantorhynchus gigas (Pallas, 1781); thegiant thorny-headed worm.
DISTRIBUTION
Occasional human cases have beenreported from Australia, Brazil, China,Czech Republic, Iraq, Russia and Thailand.
MORPHOLOGY AND LIFE CYCLE
These are large parasites, the female mea-suring 20–65 cm � 4–10 cm and the male5–10 cm � 3.5 cm. The most importantintermediate hosts are soil-inhabiting bee-tle larvae belonging to the familyScarabeidae. Development is similar to thatof Moniliformis. Recent cases in Chinahave had acute abdominal colic with mul-tiple perforations in the jejunum (Leng etal., 1983).
Macranthorhynchus ingens, normallyfound in the raccoon, has been reportedfrom a child in Texas, USA (Dingley andBeaver, 1985).
Other genera (belonging to the classPalaeacanthocephala) found once or twicein humans are: Acanthocephalus rauschiGolvan, 1969 from an Inuit in Alaska, witha fish probably acting as a paratenic host;Pseudoacanthocephalus bufonis (Shipley,1903) Southwell and Macfie, 1925, a toadparasite, with one case reported fromIndonesia; Corynosoma strumosum(Rudolphi, 1802) Luhe, 1904, one humancase being reported from Alaska (adults arenormally found in seals with fish acting asparatenic hosts).
Bolbosoma sp., a parasite of cetaceans, hasbeen recorded once from the jejunum of aJapanese, probably contracted from eatingraw marine fish.
References
Counselman, K., Fied, C., Lea, G., Nickol, B. and Neafie, R. (1989) Moniliformis moniliformis from achild in Florida. American Journal of Tropical Medicine and Hygiene 41, 88–90.
Crompton, D.W.T. and Nickol, B.B. (eds) (1985) Biology of the Acanthocephala. CambridgeUniversity Press, Cambridge, UK.
Dingley, D. and Beaver, P.C. (1985) Macranthorhynchus ingens from a child in Texas. AmericanJournal of Tropical Medicine and Hygiene 34, 918–920.
Leng, Y.J., Huang, W.D. and Liang, P.N. (1983) Human infection with Macranthorhynchus hirudi-naceus Travassos, 1916 [sic], in Guangdong Province with notes on its prevalence in China.Annals of Tropical Medicine and Parasitology 77, 107–109.
Taraschewski, H. (2000) Host parasite interactions in Acanthocephala – a morphological approach.Advances in Parasitology 42, 1–180.
03Worms & H.D. - Chap 03 14/11/01 4:26 PM Page 107

This phylum includes the gordiids orhair-worms, in which the adults are free-living in water and the larvae are para-sitic in insects. Often adults have beenfound in water in a toilet bowl from aninsect that fell into the water and werethought to have been passed in the faecesor urine. However, in isolated cases frommany parts of the world, worms do seemto have been passed in the faeces or perurethra or have been vomited out(Uchikawa et al., 1987) or removed fromthe eye or ear; in some cases, infectionwas probably acquired while swimming.
Numerous species have been implicated:Gordius aquaticus, G. chilensis, G. ges-neri, G. inesae, G. ogatai, G. perronciti,G. reddyi (a case in India with worms inthe orbit), G. robustus (abscess in orbit),G. setiger, G. skorikowi, Chordodescapensis, Neochordades columbianus(from the ear in Colombia),Parachordodes alpestris, P. pustulosus, P.raphaelis, P. tolusanus, P. violaceus, P.wolterstoffi, Paragordius cinctus, P. esa-vianus (in urethra), P. tanganyikae, P. tri-cuspidatus and P. varius (see Coombsand Crompton, 1991).
4The Nematomorpha
108
References
Coombs, I. and Crompton, D.W.T. (1991) A Guide to Human Helminths. Taylor and Francis, London,UK. pp. 177–190.
Uchikawa, R., Akune, K., Tinone, I., Akune, K., Inove, J., Kagei, N. and Sato, A. (1987) A human caseof hair worm (Gordius sp.) infection in Kagoshima, Japan. Japanese Journal of Parasitology 36,358–360.
04Worms & H.D. - Chap 04 14/11/01 4:26 PM Page 108

The nematodes are unsegmented pseudo-coelomate worms, usually cylindrical buttapering at the anterior and posterior ends.As a group they are remarkably uniform instructure, although differing in size andhabitat. Their uniformity is partly imposedby high internal hydrostatic pressure andhas resulted in fewer external protruber-ances than are found in most invertebrates.They are probably one of the most abun-dant and widespread of all animal groups,occurring in the sea, fresh water and soiland as parasites of vertebrates, inverte-brates and plants.
Species parasitic in humans (63 speciesin total here) vary in size from threadlikeobjects just visible to the naked eye(Strongyloides, Trichinella), to elongatestringlike worms attaining a length of50 cm (Dracunculus).
The outer layer of the body forms amultilayered proteinaceous cuticle. It con-tains collagen fibres arranged in a trellis-likepattern, thus allowing some contraction andextension of the body. The surface of the liv-ing worm is biologically inert (in contrast tothat of trematodes and cestodes) and themain antigenic stimulus to the host occursat various orifices with the production ofexcretory and secretory (ES) antigens.
During the life history the cuticle is shedperiodically, the process being termed amoult or ecdysis. There are four moultsduring development from ovum to adult,and in the majority of phasmidian speciesthe third-stage larva is responsible for infec-tion of a new host. The geographical distri-bution, location in the body and life cycles
of nematodes of most medical importanceare summarized in Tables 6 and 7.
All nematodes are dioecious and thefeatures typical of the male and femalephasmid nematode are shown in diagramsof the pinworm, Enterobius (Fig. 51).
Classification(mainly based on Anderson et al.,
1974–1983)
KEY: M = mammal, B = bird, R = reptile, A = amphibian, F = fish.
PHYLUM NEMATODAElongate, bilaterally symmetrical, cylindri-cal parasites. Unsegmented, with a gut andanus. Pseudocoelomate. Sexes separate.With four larval stages.
CLASS SECERNENTEA (‘Phasmidia’)These have the presence of small sensorystructures on the tail end (phasmids) and acharacteristic excretory system. Withnumerous caudal papillae.
Order Rhabditida (MBRA)Family StrongyloididaeFree-living and parasitic generations canalternate. The parasitic females areparthenogenetic. Small buccal capsulewithout teeth. Oesophagus of free-livingadults and of first-stage larva with posteriorbulb (rhabditoid).Strongyloides
Order Strongylida (MBRA, occasionally F)Male with cuticular copulatory bursa sup-ported by rays and with two similar
5The Nematodes
109

110 Chapter 5
Table 6. Outline of nematode life cycles.
1. Direct cycle: no intermediate host required
Vertebrate host
Egg contains Unembryonated egg Embryonated egg Rhabditiform larvarhabiditiform larva in faeces in faeces in faeceson perianal skin
Larva develops in Egg hatches on Free-living egg but does not ground generation with
hatch (L1 in Trichuris rhabditiformL2 in Ascaris) larvae in soil
Egg hatches in intestine when swallowed by humans Rhabditiform Larva develops to larvae develops to L3 in soil
L3 with sheath in soil
Larva (L1) develops Larva reaches L3 ingested L3 penetrates L3 penetrates directly to adult in intestine after skin skin
intestine entering bloodand via lungs
Enterobius Trichuris Ascaris Necator StrongyloidesAncylostoma
AncylostomaMammomonogamus*Oesophagostomum
Ternidens Trichostrongylus
2. Indirect cycle: intermediate host required
Vertebrate host
Rhabditiform L1 L1 larva in water Microfilaria L1 larva encysted larva in faeces (pre L1) larva in in muscles
blood or lymph
To L3 in land To L3 in To L2 in To L3 in bitingsnail or slug cyclopoid cyclopoid insect
L3 ingested L3 L2 ingested L3 enters skin L1 ingested ingested and to L3 in and to L3 in
fish, frog or snake intestine
L3 ingested
Parastrongylus* Gnathostoma* Trichinella*Dracunculus Brugia
LoaMansonellaOnchocercaWuchereria
*Humans are accidental or aberrant hosts.

The Nematodes 111
Table 7. Nematodes of medical importance.
Mode of infection and Geographical
Species infective Stage Site of adult distribution
Directly Enterobius Mouth (L1 in Large intestine Cosmopolitancontaminative vermicularis eggs)
Sometimes Skin (L3) Small intestine Africa, Asia, Strongyloides America, Pacificstercoralis
Soil-transmitted Strongyloides Skin (L3) Small intestine Africa, Asia,(geohelminths) stercoralis America, Pacific
Necator Skin (L3) Small intestine Africa, Asia,americanus America, PacificAncylostoma Mouth or skin Small intestine Asia, Africa, duodenale (L3) Middle East, PacificTrichostrongylus Mouth (L3) Small intestine Asia, Middle East, spp. AfricaAscaris Mouth (L2 in egg) Stomach and Cosmopolitanlumbricoides small intestineTrichuris trichiura Mouth (L1 in egg) Caecum Cosmopolitan
Aberrant Ancylostoma spp. Skin (L3) Larvae in skin Africa, America, infections (dermal larva Asia, Pacific
migrans)Toxocara, etc. Mouth (L2 in egg) Larvae in liver, Cosmopolitan(visceral larva eye, brainmigrans)
Insect-transmitted Onchocerca Simulium skin Subcutaneous Africa, Yemen, Central volvulus (L3) and South AmericaWuchereria Culex, Anopheles Lymphatics Asia, Africa, South bancrofti and Aedes skin (L3) America, PacificBrugia malayi Mansonia skin (L3) Lymphatics South-East AsiaLoa loa Chrysops skin (L3) Subcutaneous Africa (rain forest)Mansonella Culicoides skin (L3) Peritoneal cavity Africa, South AmericaperstansMansonella Culicoides skin (L3) Peritoneal cavity Africa (rain forest)streptocercaMansonella Culicoides, Peritoneal cavity South America, ozzardi Simulium skin (L3) West Indies
Crustacean- Dracunculus Cyclopoids mouth Subcutaneous Africa, Middle East, transmitted medinensis (L3) [India, Pakistan]
Aberrant Gnathostoma Cyclopoids mouth Subcutaneous South-East Asiainfection spinigerum (L2)
Snail-transmittedAberrant Parastrongylus Misc. snails and infection cantonensis crustaceans mouth Larvae in Pacific
(L3) meningesParastrongylus Misc. snails and Larvae in Americas, Japan, costaricensis crustaceans mouth mesenteric Africa (?)
(L3) arteries
Meat-transmittedAccidental Trichinella Mouth (L1) Small intestine Cosmopolitaninfection spiralis
Fish-transmittedAberrant Anisakis Mouth (L3) Intestine Europe, JapaninfectionAccidental Aonchotheca Mouth (L1) Small intestine Philippines, Thailandinfection philippinensis

spicules. Oesophagus club-shaped posteri-orly, but without definite bulb (rhabditoidin first-stage larva). Mouth without lips.Direct life cycle usually includes two free-living and feeding larval stages (except insuperfamily Metastrongyloidea).
SUPERFAMILY ANCYLOSTOMATOIDEA (M)Family AncylostomatidaeBuccal capsule well developed, usuallywith ventral teeth or cutting plates. Caudalbursa of male usually supported by rays.Ancylostoma, Necator
SUPERFAMILY STRONGYLOIDEA
Buccal capsule globular, strongly devel-oped and cuticularized.
Family Chabertiidae (M)Mouth surrounded by leaf crown.Oesophagostomum, Ternidens
Family Syngamidae (MB)Leaf crown rudimentary. Male and femaleattached.Mammomonogamus
SUPERFAMILY TRICHOSTRONGYLOIDEA (MBRA)Family Trichostrongylidae (MB)Filiform worms with the buccal capsuleabsent or feebly developed. Caudal bursawith reduced or absent dorsal lobe.Trichostrongylus
SUPERFAMILY METASTRONGYLOIDEA (M)Family Angiostrongylidae (M)Filiform with reduced buccal capsule.Caudal bursa of male usually asymmetricaland may be reduced. Parasites of repiratoryor circulatory system. Require an inter-mediate host.Parastrongylus
Order Ascaridida (MBRAF)Large stout intestinal worms. Mouth usuallywith three lips. Oesophagus without valvu-lar apparatus and not divided into two por-tions. Male and female similar in size. Malewith equal spicules. Life cycle either director indirect with intermediate host.
SUPERFAMILY ASCARIDOIDEA
Family AscarididaeDirect life cycle.Ascaris, Toxocara, Toxascaris
Family AnisakidaeIndirect life cycle.Anisakis, Pseudoterranova
Order Oxyurida (MBRAF)Small stout nematodes. Oesophagus withposterior bulb with valvular apparatus.Female with long pointed tail. Direct lifecycle, with two moults in egg. Female withlarge embryonated eggs, often flattened onone side. Parasites of colon or rectum.
SUPERFAMILY OXYUROIDEA
Family OxyuridaeEnterobius
Order Spirurida (MBRAF)Oesophagus in two parts, anterior muscularand posterior longer part glandular. Indirectlife cycle with larvae in arthropods.
SUPERFAMILIES GNATHOSTOMATOIDEA, GONGYLO-NEMATOIDEA, THELAZOIDEA, HABRONEMATOIDEA,PHYSALOPTEROIDEA (‘Spirurids’)Filiform. Mouth usually has two lips.Female longer than male, with vulva in theanterior region. Spicules of male veryunequal. Mainly non-intestinal.Gnathostoma, Gongylonema, Thelazia,Habronema, Physaloptera
112 Chapter 5
Fig. 51. Diagram of (a) a female (8–13 mm long) and (b) male (2.5 mm long) Enterobius vermicularis toillustrate typical nematode features (oesophagus = pharynx).

SUPERFAMILY FILARIOIDEA (MBRA) (‘Filariae’)Family OnchocercidaeFiliform. Mouth usually without lips.Female longer than male with vulva inanterior region and double uterus. Spiculesvery unequal in male. Non-intestinal para-sites. Usually ovoviviparous, with larvae inbloodsucking insects (or arachnids).Wuchereria, Brugia, Loa, Onchocerca,Mansonella, Dirofilaria
SUPERFAMILY DRACUNCULOIDEA (MBRF)Family DracunculidaeSimilar in structure to filariae but femalesvery much longer than males, with vulvaand anus atrophied in gravid female andsingle uterus. Ovoviviparous with larvae incopepods.Dracunculus
CLASS ADENOPHOREA (‘Aphasmidia’)Phasmids absent. Caudal papillae absentor few in male. Oesophagus usually form-ing a stichosome. First-stage larvae, oftenwith stylet and infective to final host.
Many members are parasites of plants andinsects.
Order Enoplida (MBRAF)SUPERFAMILY TRICHINELLOIDEA
Family TrichuridaeSexes separate. Anterior region of bodymuch thinner than posterior. Vulva near endof oesophagus. Male with single spicule.Trichuris, Aonchotheca, Calodium, Eucoleus
Family TrichinellidaeSexes separate. Anterior region of bodyslightly thinner than posterior. Male withsingle or no spicule. Female with oneovary and uterus, vulva near middle ofoesophagus. Ovoviviparous.Trichinella
SUPERFAMILY DIOCTOPHYMATOIDEA
Family DioctophymidaeMale tail modified to form ventral sucker.Well-developed oesophagus.Dioctophyma, Eustrongylides
The Nematodes 113
A Key to Nematodes Parasitic in Humans
1. Heterogenic. Parasitic form parthenogenetic with direct life cycle.Small buccal capsule without teeth RHABDITIDA
StrongyloidesNot heterogenic. Parasitic forms sexually differentiated 2
2. Pharynx consisting of a narrow tube running through the centre of a single row of cells ENOPLIDA
(TRICHUROIDEA)AonchothecaCalodiumEucoleusTrichurisTrichinella
Pharynx muscular and not consisting of intracellular tube 33. Males with copulatory bursa 4
Males without copulatory bursa 54. Copulatory bursa muscular and without rays ENOPLIDA
(DIOCTOPHYMATOIDEA)DioctophymaEustrongylides
Copulatory bursa cuticular and supported by rays STRONGYLIDAsee subkey below
5. Pharynx dilated posteriorly into a bulb, usually containing a valvular apparatus and often separated from rest of pharynx by constriction. Usually small forms OXYURIDA
EnterobiusPharynx not dilated posteriorly into a bulb 6

6. Relatively stout worms. Head with 3 large lips ASCARIDIDAAscarisToxocara
Relatively slender filiform worms. Head with 2 lateral lips or 4 or 6 small lips or absent. Arthropod intermediate hosts 7
7. Usually 2 lateral lips. Chitinous buccal capsule. Vulva in middle of body or posterior. Parasites of alimentary canal, respiratory system or orbital, nasal or oral canal SPIRURIDA
(except FILARIOIDEA and DRACUNCULOIDEA)
GnathostomaHabronemaPhysalopteraThelazia
Usually without lips. Buccal capsule absent. Vulva in pharyngeal region. Parasites of circulatory or lymphatic system, serous cavities, muscles or connective tissues 8
8. Females not more than three times longer than males. Vulva not atrophied SPIRURIDA
(FILARIOIDEA)Filariae
Females enormously longer than males. Vulva atrophied in gravid female SPIRURIDA
(DRACUNCULOIDEA)Dracunculus
Subkey to the order Strongylida
(a) Parasites of alimentary canal or rarely in renal tissue (b)Parasites of respiratory system (d)
(b) Buccal capsule feebly developed or absent. Threadlike worms TRICHOSTRONGYLOIDEA
TrichostrongylusBuccal capsule well developed (c)
(c) Buccal capsule with ventral teeth or cutting plates ANCYLOSTOMATOIDEA AncylostomaNecator
Buccal capsule without teeth, etc. but mouth typically with a leaf crown STRONGYLOIDEA
(Family Chabertiidae) OesophagostomumTernidens
(d) Well-developed chitinous buccal capsule. Male often attached to female STRONGYLOIDEA
(Family Syngamidae)Syngamus
Rudimentary buccal capsule or absent. Filiform METASTRONGYLOIDEAParastrongylus
114 Chapter 5

The section on the nematodes has beensplit into two parts, those that are intesti-nal and others that are tissue parasites.This has allowed the control of the com-mon intestinal nematodes to be consideredtogether. However, it has resulted in treat-ment of one of the main zoologicallyrelated group of nematodes being dividedby the other as below:
Intestinal nematodes | Tissue nematodes
Class Secernentea | Class Adenophorea | Class Secernentea
Intestinal Nematodes
Geohelminths
This is a group of intestinal nematodeswhich, while not closely related zoologi-cally, are all soil-transmitted and havegreat similarities in epidemiology and inmethods of control: it includesStrongyloides, the hookworms (Necatorand Ancylostoma), Ascaris and Trichuris.
Order RhabditidaFamily Strongyloididae
Strongyloides stercoralis (Bavay, 1876) Stiles and Hassall, 1902
SYNONYMSAnguillula stercoralis, Bavay, 1876 (larvaefrom faeces); A. intestinalis, Bavay, 1877(parasitic females from intestine);Pseudorhabditis stercoralis Perroncito, 1881(free-living adults).
DISEASE AND POPULAR NAMEStrongyloidiasis or strongyloidosis, anguil-losis; Cochin-China diarrhoea. Included asone of the geohelminths.
GEOGRAPHICAL DISTRIBUTION
Strongyloides infection is widely distrib-uted in tropical regions of Africa, Asia andCentral and South America (particularlyBrazil, Colombia and Guyana); it alsooccurs in eastern Europe and sporadicallyin the USA and southern Europe.Distribution is similar to that of hook-
worms but the prevalence is usually muchlower – an estimated 100 million casesworldwide (Grove, 1996).
LOCATION IN HOSTParthenogenetic females live embedded inthe mucosa of the small intestine – theduodenum and the first part of the jejunumin light infections, back to the terminalileum in heavy infections.
MORPHOLOGY
Parasitic females are small, transparent, fili-form nematodes, measuring 2.0–2.7 mm inlength by 0.03–0.075 mm in width. The cuti-cle is finely striated and the tail pointed. Theoesophagus occupies the anterior third of thebody and joins the midgut, which opens atthe ventral anus a short distance from theposterior end. The vulva opens ventrally atthe junction of the middle and posteriorthirds of the worm. The uteri, oviducts andovarian tubules are paired, one branchextending forwards from the vulva and theother backwards. About 10–20 eggs are pre-sent in the two uteri at one time. The eggs arethin-shelled, transparent and ovoid; theymeasure 50–60 µm � 30–35 µm when laid.The eggs are partially embryonated and usu-ally hatch in the tissues of the mucosa, sothat larvae rather than eggs are passed out inthe faeces (Figs 123 and 125).
The females produce eggs partheno-genetically and, although parasitic malessimilar to free-living ones described belowwere recorded twice, they have never beenseen since.
Strongyloides is also capable of undergo-ing a complete life cycle in the soil, livingas a free-living nematode with a distinctmorphology. The free-living female isshorter and stouter than the parasitic form(1.0 mm � 0.06 mm) and has a short rhab-ditiform oesophagus (muscular and with anenlarged posterior bulb, similar to that ofthe saprophytic soil nematode Rhabditis).
The free-living male measures about0.7 mm � 0.04 mm and has a pointed tail,curving ventrally, with two spicules and agubernaculum (this is a chitinous sheath inwhich the spicules can slide up and down;it is presumed to aid in the transference of
The Nematodes 115
116C
hapter 5
Map 7. Distribution of Strongyloides stercoralis (and S. fuellebornius in New Guinea).

sperm during copulation and is often oftaxonomic importance). The oesophagus isrhabditiform.
LIFE CYCLE (Fig. 52)
Parasitic (= direct) life cycle. Eggs are laid inthe mucosa and submucosa of the smallintestine. There they hatch as rhabditiformfirst-stage larvae, which escape into theintestinal lumen and pass out in the faeces,where they are very active. The larvae mea-
sure 250 µm � 20 µm and can be differenti-ated from those of hookworms by thesmaller buccal capsule (Fig. 53). The larvaefeed on bacteria in the soil and moult twiceto give the infective filariform stage, mea-suring 550 µm � 20 µm, in the surface lay-ers of the soil. The infective larvae differfrom those of hookworms by the lack of asheath, by the triradiate tip to the tail andin the greater length of the pharynx. Theyascend soil particles and damp vegetation
The Nematodes 117
Parthenogenic ADULTFEMALE in small intestine (2.0 mm)matures in 17 days
3RD-STAGE FILARIFORMLARVA (0.55 mm)lives about 12 daysin soil
1ST-STAGE RHABDITIFORMLARVA (0.25 mm)
FREE-LIVING MALE(0.8 mm)
spiculestestis
vulva
ovary
FREE-LIVINGFEMALE (1.0 mm)
Moults twice
Penetrates skin andreaches gut via blood,lungs, trachea,oesophagus
Or
indi
rect
DirectEither
Autoinfection can occurwhen larvae develop onperianal skin or rarelyin intestinal tract
3RD-STAGELARVA
1ST-STAGELARVA
in faeces andreaches soil
1ST-STAGERHABDITIFORMLARVA (0.25 mm)
Fig. 52. The possible life cycles of Strongyloides stercoralis. (Portrait of Dionysus from a Greek cylix.)

in the surface film, often standing on theirtails in tufts or aggregates. Under optimumconditions the larvae live in the soil forabout 2 weeks but, when they come intocontact with the skin, usually of the feet (ormore rarely the buccal mucosa), they pene-trate and reach the small cutaneous blood-vessels and then the lungs. Afterpenetration of the alveoli, the larvae moulttwice and the young adults pass up thebronchi and trachea and then down theoesophagus to reach the small intestine(although it has recently been suggestedthat they may use other routes). Once there,the female burrows into the mucosal tis-sues, becomes mature and lays its partheno-genetically produced eggs in the mucosaand crypts about 17 days after penetration.
Very unusually among human helminthsis the possibility of replication within thehuman host. Autoinfection is usuallybrought about because the rhabditiform lar-vae in faecal matter are deposited on theperianal skin and develop into filariform lar-vae, which penetrate through the skin. Thisaccounts for the persistence of strongyloidia-sis in patients up to 65 years after they haveleft an endemic region (most notably in for-mer Second World War prisoners in the FarEast, who are still suffering from recurrentlarva currens), although the adult worms live
for only a few months. In severely illpatients or in chronic carriers, rhabditiformlarvae passing down the gut may developinto filariform larvae without reaching theexterior at all, and these then penetrate theintestinal mucosa. Exceptionally, when thepatient’s resistance is low, all larval stagesmay be found in many of the visceral organsand rhabditiform and filariform larvae mayeven appear in the sputum.
Free-living (= indirect or heterogonic) lifecycle. A free-living cycle often occurs inwarm climates, provided that there is plentyof moisture and nutrients in the soil. Therhabditiform larvae passed out in faecesfeed and undergo four moults in the soil tobecome free-living adult males and femalesin 24–30 h. The eggs, which develop in theuterus of the free-living female after mating,occupy a large portion of the body and arelaid in a partly embryonated state. Theyhatch in a few hours after completing devel-opment and rhabditiform first-stage larvaeemerge. These first-stage larvae moult twiceand develop into non-feeding, filariform,third-stage larvae in a few days. The infec-tive larvae survive on soil and vegetation inwarm moist conditions and penetrate theskin when they come in contact. There is nosecond free-living cycle.
118 Chapter 5
Fig. 53. The anterior ends of the first-stage larva (L1) of Strongyloides (left) and hookworm (right) in soil.The former has a shorter buccal capsule.

The factors determining whether direct orindirect development occurs is not wellunderstood but it is believed that thehigher the immune status of the host themore likely that larvae passed in the faeceswill undergo an indirect cycle. In otherspecies at least there are genetically deter-mined strain differences and environmen-tal factors also help to determine whethermales and females or infective larvaedevelop in soil.
CLINICAL MANIFESTATIONS
There may be pruritic eruption at the siteof entry of the larvae (‘ground itch’) inheavy infections, followed by a cough aftera few days.
The majority of cases are of chronic,uncomplicated, strongyloidiasis and aresymptomless, with just a few worms presentand no alteration to the mucosa. There may,however, be abdominal discomfort, withintermittent gastric pains, accompanied bynausea, diarrhoea, weight loss and a gener-alized urticaria. Recurrent, linear, rapidlyprogressive, urticarial weals (‘larva currens’)can occur on the buttocks, groins or trunk.Lesions in the gluteal region indicate thatautoinfection is occurring and are likely tobe present in all clinically important cases.
In a proportion of heavily infectedcases, more serious clinical disease ensues,characterized by enteritis with malabsorp-tion syndrome and electrolyte imbalance(Plate 12). Such infections in children mayhave serious consequences, precipitatingkwashiorkor and emaciation.
Disseminated (complicated, hyperinfec-tive or overwhelming) strongyloidiasis isalways serious and often fatal and can havea variety of manifestations, depending onthe intensity of infection and the organsinvolved. Complications affect particularlythe bowel, lungs and central nervous system(CNS) and are often accompanied by sec-ondary bacterial infection. Parasitic ileus(pseudo-obstruction), pneumonitis withpulmonary infiltrates, meningitis, brainabscess and septicaemia are all possible.
Disseminated strongyloidiasis occurswhen the body’s cell-mediated immuneresponses are deficient, often in patients
with malignant lymphoma or acuteleukaemia, on immunosuppressive drugs,after renal transplantation, or suffering frommalnutrition. It does not seem to be stronglyassociated with cases of AIDS, althoughthere is a correlation with HTLV-1 infectionand it is possible that Strongyloides leads toimmunoglobulin E (IgE) deficiency (Phelpsand Neva, 1993). Hyperinfection may alsobe potentiated by a decrease in mobility ofthe intestine, which allows first-stage larvaeto mature into infective forms before beingpassed out in the faeces. This postulate issupported by the fact that, in cases of hyper-infection, ulcerations caused by infectivelarvae are always present in the colon aswell as the ileum. It has been postulatedthat hyperinfection is not an immune phe-nomenon but is caused by corticosteroidtherapy stimulating the production ofecdysteroids (moulting hormones) (Genta,1992). However, hyperinfection occurs inmany patients who have not received corti-costeroids.
There is evidence that the immunomod-ulatory effect of many helminths mayincrease susceptibility to tuberculosis (TB)and AIDS (Borkow and Bentwich, 2000;Bundy et al., 2000).
PATHOGENESISThis can be divided into three stages.
1. The penetration of the skin by filariformlarvae usually causes no symptoms in non-sensitized patients, provided infection islight. Larvae secrete a metalloprotease ofmolecular mass 40 kDa, which is immuno-genic; in heavier infections a rash oftenappears at the site of entry (‘ground itch’)and there may be severe pruritus. Whereautoinfection is occurring there may alsobe perianal and perineal ulceration. 2. The larvae that reach the lungs and passinto the alveoli can cause a pneumonitissimilar to Loeffler’s syndrome in ascariasis(p. 150). 3. The adult females live in the depths ofthe crypts of the ileum and lay their eggsthere. Larvae hatching from deposited eggsburrow through the lumen and causesuperficial catarrhal damage to the mucosa,often with excessive production of mucus.
The Nematodes 119

In severe cases of autoinfection, parasitesare present in the submucosa, whichbecomes oedematous. The mucosa becomesflattened and atrophic (Fig. 54), the patho-logical picture resembling that of tropicalsprue or of infection with Aonchothecaphilippinensis (p. 173). There may be severeulceration with a non-reversible fibrosis andoften a pronounced inflammatory responseto secondary bacterial invasion. Larvae inthe lymphatics cause a granulomatous lym-phangitis; this, coupled with the inflamma-tion, atrophic mucosa and fibrosis, causes amarked congestion and loss in elasticity ofthe gut. Paralytic ileus can result and therecan also be a severe haemorrhagic, ulcera-tive enterocolitis, with massive invasion ofthe whole bowel wall by larvae. In cases ofcomplicated disease, larvae can be found inalmost any organ, particularly the intestinallymph nodes, lungs, liver, heart, urinarytract and nervous system.
IMMUNOLOGICAL ASPECTS OF INTESTINAL
NEMATODES
Infections with intestinal nematodes generatecharacteristic patterns of immune responsesin their hosts. Typically, infected individualshave elevated levels of IgE, high peripheraleosinophilia and a variety of allergic symp-toms, some of which are generalized, othersbeing localized to the tissues invaded. Forexample, where infection occurs through theskin (e.g. hookworms, Strongyloides) theremay be dermatitis and pruritus, where para-site development involves the lungs (e.g.Ascaris, hookworms) there may be bronchitisor pneumonitis, while invasion of muscles(Trichinella) causes myositis. Collectivelythese symptoms imply that intestinal nema-todes generate responses dominated by the T-helper 2 (Th2) subset, and this is in agreementwith experimental studies in laboratoryrodents. Many species cause forms of intes-tinal pathology that again can be related toimmunological causes, e.g. the protein-losingenteropathy seen in patients infected withStrongyloides spp. or Aonchotheca philip-pinensis, and the dysentery syndrome seen inchildren heavily infected with Trichuristrichiura, although the extent to which theseare Th2-related is debatable. A major differ-ence between experimental studies and obser-vations on human infections is that, whereaslaboratory rodents mount strong protectiveresponses against the majority of species used(e.g. Nippostrongylus brasiliensis, Strongy-loides ratti and S. venezuelensis, Trichinellaspiralis, Trichuris muris), evidence for protec-tive responses in humans is very limited. Therapid reacquisition of infection with the com-mon geohelminths after chemotherapy sug-gests little immunity to reinfection, but,against this, in some species, is evidence of anage-related decline in infection intensity, ifnot in prevalence (Crompton, 1998; Fig. 55).
Experimental studies suggest that Th2cells mediate immunity against intestinalnematodes through changes in mucosalstructure and function, resulting in analtered parasite environment (Wakelin,1996). Changes in mucosal architecture(through villous atrophy) and epithelialfunction, development of mucosal masto-cytosis, increased secretion of mucus from
120 Chapter 5
Fig. 54. Section of colon of patient withhyperinfective strongyloidiasis. The section is atthe edge of an ulcer and shows inflammation.Adults of S. stercoralis are in the crypts andrhabditiform larvae in the lumen.

goblet cells and altered muscular activityhave all been implicated. Although infec-tion does elicit Th2-dependent antibodies(e.g. in mice IgA, IgE and IgG1), these arenot thought to play a major role. Little ornothing is known of comparable mucosalchanges in infected humans and, althoughantibody responses are made, these areoften positively correlated with the dura-tion and intensity of infection, suggestingno role in protection. Arguments have beenmade, however, for inverse correlations
between levels of IgA and level of infectionwith Trichuris and between the levels ofIgE and development of Necator.
The reasons for the apparent lack of pro-tective immunity against intestinal wormsare not fully understood, although manysuggestions have been made. In countrieswhere these infections are common, nutri-tional levels are often low and immunityimpaired as a consequence. The pathologi-cal consequences of the worm infection mayexacerbate nutritional problems, leading to
The Nematodes 121
Pre
vale
nce
(%)
(a)
(b)
Ascaris lumbricoides
100
80
60
40
20
00 20 40
Age (years)
(c)
Pre
vale
nce
(%)
Pre
vale
nce
(%) 100
Hookworm
80604020
00 1 2 3 4 5 6 7 8 9 10 11
Age (years)
Hookworm
3
2
1
00 1 2 3 4 5 6 7 8 9 10 11
Age (years)
e.p.
g. (
�10
–3)
e.p.
g. (
�10
–3)
Trichuris trichiura
3
2
1
00 20 40
Age (years)10 30
Trichuris trichiura
100
80
60
40
20
00 20 40
Age (years)10 30 50
(f)
(e)
Ascaris lumbricoides
20
16
12
8
4
010 20 60
(d)
40Age (years)
Mea
n w
orm
bur
den
Fig. 55. Left: Graphs showing the relationship between prevalence and age for (a) Ascaris, (b) Trichuris,(c) hookworms. Each graph is based on a different research study. Right: Graphs showing therelationships between intensity of infection and age for (d) Ascaris, (e) Trichuris, (f) hookworms. Curve(d) is based on recovery of worms after chemotherapy and (e) and (f) on egg counts. e.p.g., eggs pergram. Each graph is based on a different research study (all graphs from Crompton, 1998).

a vicious circle. Infections are sometimesassociated with reduced immune respon-siveness, and it is clear that worms them-selves can interfere with the protectiveresponses of the host in a variety of ways(Prichard, 1993). It is also possible thatgenetic factors influence the ability to con-trol infections, as has been amply demon-strated in experimental systems (Wakelin,1994). It is characteristic of infections withintestinal nematodes in populations thattheir distributions are highly aggregated, i.e.most individuals have low infections whilea few are heavily infected (Crompton, 1998).There has been much debate about thecause of such aggregation, with polarizedviews that it is due primarily to environ-mental exposure or that it is a consequenceof variation in immune responsiveness. Itwould be unlikely that genetic factors didnot influence the ability to respond protec-tively to intestinal worms, and indeed therehave been claims that possession of particu-lar major histocompatibility complex(MHC)-linked genes does correlate with anindividual’s likely level of infection.
DIAGNOSIS
Clinical. There is no well-defined clinical pic-ture, although watery diarrhoea with mucus,perhaps interrupted by constipation, abdomi-nal pain and with a high eosinophilia, maysuggest strongyloidiasis. The lesions in thegluteal region and larva currens are almostpathognomic if present. Imaging techniquescan show infiltration in the lungs (Fig. 56).
Parasitological. The presence of rhabditi-form first-stage larvae in a fresh faecal sam-ple provides a positive diagnosis (Figs 123and 125). In an old sample mixed with soil,hookworm larvae could have hatched fromeggs and must be differentiated (when liv-ing, Strongyloides larvae lash like a whip,while hookworm larvae move sinuously likea snake). Also in old faeces (30 h) free-livingadults of Strongyloides might be found.
To concentrate the larvae, Baermann fil-tration or formol–ether concentration (p. 263)can be used. In the Baermann technique 10 g (or more) of faeces is spread on a six-
layered gauze pad on a coarse sieve with awooden applicator and placed in a sedimen-tation flask that has been filled 25 mmbelow the rim with 0.9–1.2% saline or waterat 37°C. Alternatively, a funnel can be usedwith a length of plastic tubing clamped atthe bottom. If the faeces is watery, it can bespread on filter-paper, which is then placedface down on the gauze. After 2 h, 5 ml ofthe bottom sediment can be removed with aPasteur pipette and transferred to a 5 cmPetri dish. This is examined under a dissect-ing microscope for first-stage larvae. If nolarvae are found, the flask can be leftovernight at room temperature and re-examined; then a drop of iodine can beadded and the larvae examined on a micro-scope slide for generic identification.
There are various culture methods thatcan be used if no larvae can be found butdisease is clinically indicated.
1. The Harada–Mori, filter-paper, test-tubeculture and formol–ether concentrationmethods, which can also be used for hook-worms, are given on p. 263.
122 Chapter 5
Fig. 56. Radiograph of lungs in a patient withdisseminated strongyloidiasis, showing diffuseinfiltrations.

2. In the agar plate method, 2 g of faeces isplaced in the centre of an agar medium in a9 cm plastic Petri dish, covered and sealedround the edge with adhesive tape, andincubated at room temperature (about28°C) for 48 h. For greater safety, a double-walled Petri dish (or a smaller dish insidethe larger) can be used, the outer area con-taining 25% glycerine to prevent infectivelarvae from reaching the edge of the lid.The tracks of larvae can be seen with thenaked eye but the dish should also beexamined under a compound microscopewith a green filter at �40 magnification.For generic identification, a hole can bemade in the agar with hot forceps, 10% for-malin added, liquid removed from the bot-tom with a Pasteur pipette, sedimented andthe sediment examined on a microscopeslide at high power. This is probably themost sensitive method.3. In the watch-glass culture method sev-eral grams of faeces are spread out on awatch-glass, moistened with water, placedin a Petri dish containing a 2–3 mm layerof water, covered and re-moistened asrequired. Free-living adults may be seen onthe watch-glass after several days or infec-tive larvae after 3 days.4. A charcoal culture method is also some-times employed. Five to 10 g of faeces ismixed with activated charcoal in a 10 cmdiameter Petri dish, well dampened withdistilled water and left covered for 4 daysin an incubator at 26–28°C. The infectivelarvae can be recovered with a soft paint-brush from the condensation drops on thelid. For large quantities of faeces, a sealedjam jar can be used, 5 cm full, and largenumbers of larvae will be found in themeniscus around the edge of the mixturewhen left as before.5. The entero-test involves swallowing abrushed-nylon thread inside a gelatin cap-sule and then withdrawing it together withentrapped larvae, fixing in 10% formalinand examining under the microscope. Incases of hyperinfection, larvae may befound in sputum or even urine.
Immunological. These tests cannot usuallydifferentiate between current and past
infections. A skin test using extracts offilariform larvae has been in use for 70years. It measures IgE and is about 80%reliable, although, like other tests, it cross-reacts with filarial infection.
The ELISA is fast and efficient, usingstored antigen, and is in routine use insome laboratories (Mangali et al., 1991).The test gives sensitivity and specificity ofabout 95% when measuring IgG. Specificityof the indirect test can be improved byprior absorption with filarial and hook-worm antigens (Conway et al., 1993).
The indirect fluorescent antibody (IFA)test has been used with living larvae as anti-gen, preabsorbed against other nematodes,and recently a simple gelatine agglutinationtest has been proposed (Sato et al., 1991).
There is as yet no method for measuringcirculating antigens.
TREATMENTChemotherapy is principally by benzimida-zoles, which appear to act by binding totubulin and disrupting the assembly ofmicrotubules and by altering transmembraneproton discharge (Grove, 1996).
Thiabendazole. Given orally twice a day for3–7 days, 25 mg kg�1 body weight has acure rate of about 80%. This is still thedrug of choice – unfortunately, since side-effects (nausea, dizziness and anorexia) arecommon – chemical formula: (2-(4-thia-zolyl)-1-H-benzimidazole-2-yl)-carbamate.
Mebendazole. If given at 100–200 mg twicedaily for 3 days, it is probably effectiveagainst adult worms but not against migrat-ing larvae. Treatment for several weeks ismore effective, probably because it killsautoinfecting larvae when they developinto adults. Higher dosages may be moreeffective for cases of disseminated strongy-loidiasis – chemical formula: methyl(5-(benzyl)-1-H-benzimidazole-2-yl)-carbamate.
Albendazole. When given at 400 mg twicedaily for 3 days, it has given variableresults in trials, with 50–85% cure rates.Long-term treatment will probably prove tobe as effective as mebendazole, perhaps with
The Nematodes 123

fewer side-effects and greater action againstlarvae – chemical formula: methyl-(5-propylthio-1-H-benzimidazol-2-yl)-carbamate.
Ivermectin. In clinical trials of uncompli-cated cases, this macrocyclic lactone wasfairly effective (over 80% cure rate) againstadults and larvae at a single dose of 0.2 mgkg�1 body weight (Marti et al., 1996). Long-term treatment (0.2 mg kg�1 once a week for4 weeks) with ivermectin may prove to bethe best option for complete cure in patientswho fail to respond to initial treatment. Fordisseminated strongyloidiasis, treatmentover 3–4 weeks is necessary (Grove, 1996).
It is very difficult to treat cases of dissemi-nated strongyloidiasis with intestinalobstruction, where patients usually needsuction and drainage, intravenous fluids andthiabendazole given daily for a week, per-haps rectally, since there is no intravenousformulation. Since patients often die, par-enteral ivermectin or use of cyclosporin (3mg kg�1 daily) could be tried as a last resort.
EPIDEMIOLOGY
In many areas strongyloidiasis is coextensivewith hookworm infection since the environ-mental conditions favouring the free-livingstages and the mode of infection are similar,but this is not always true. Like hookworm,it is primarily a disease of poverty andinsanitary conditions, particularly wherethere are poor conditions for disposal ofhuman waste and inadequate water sup-plies. In a study in Bangladesh, occupants ofan urban slum community were more likelyto be infected if they used communallatrines rather than individual latrines, ifthey had an earth floor rather than a concretefloor and if they were aged between 7 and 10(Hall et al., 1994). Both are primarily ruralinfections, particularly common in coffee,cocoa and banana plantations. Infection isalso common in closed communities, suchas mental institutions. Infection appears tobe acquired progressively between the agesof about 6 and 20 and then prevalenceremains constant for the whole of adult life.The larvae of both parasitic and free-livingforms are unable to survive temperatures
below 8°C or above 40°C for more than a fewdays. They have little resistance to drying,excessive moisture or marked changes intemperature. Infective larvae live for about14 days in soil.
Where Strongyloides occurs togetherwith hookworms, the former is almostalways found to be rarer. This is rather sur-prising when it is considered that the para-sitic females are more prolific than femalehookworms and the presence of free-livinggenerations should increase the numbers oflarvae in the soil. There are three possibleexplanations: the first is that the first-stagelarvae in the faeces are not as resistant ashookworm eggs; the second that the filari-form larvae have shorter longevity; and thethird that reported prevalence rates aremuch too low because of the difficulties ofdiagnosis.
Infection rates reported recently fromsurveys in various countries are:Aborigines in Queensland, Australia (large-scale survey) 2%; north-eastern Brazil5.3% and 9.6%, Sao Paulo (infants) 2%;Dagestan (large-scale) 1–1.4%; Bihar, India10%; Jamaica 1%; Laos 19% (10% inunder-10s); Bauchi in Nigeria 0.4%;Okinawa (Ryuku Islands) 4–22%; USA(large-scale) 0.2%, Kentucky 0.9%,Tennessee 3%.
PREVENTION AND CONTROL
Personal prevention is aided by the wear-ing of shoes. Control is principally by theprovision of adequate sanitation facilitiesconsequent on a rise in living standards; itis sobering to realize that 2400 millionpeople (45% of the population of theworld) still do not have sanitation. In someareas water is recycled from sewage sys-tems and larvae can still survive on the soilor grass, and water thus needs further treat-ment or storage. Targeted chemotherapy ofthose groups most at risk may prove to beof use (Conway et al., 1995), but occasionalmass chemotherapy is unlikely to be effec-tive. However, the repeated mass chemo-therapy campaigns against all geohelminthsin general being carried out in many coun-tries may have some effect (see p. 134).
124 Chapter 5

ZOONOTIC ASPECTS
Transmission is almost entirely betweenhumans, although chimpanzees and dogshave been found naturally infected.
Another species, S. fuelleborni Linstow,1905, is a common parasite of monkeys inAfrica and Asia and occurs in humans inthe forest areas of Central Africa(Cameroon, Central African Republic,Congo, Congo Republic, Ethiopia, Malawi,Ruanda, Togo and Zimbabwe). In thesecountries it appears to be a natural parasiteof humans and in some areas is the pre-dominant species of Strongyloides, with aninfection rate of 25%.
S. fuelleborni can be differentiated fromS. stercoralis as eggs, not larvae, are foundin the faeces; these measure on average53 µm � 35 µm (range 48–61 µm � 30–40µm) (Fig. 124). The eggs are smaller thanhookworm eggs and in a later stage ofcleavage. The eggs hatch quickly in faecalsamples and in cases of doubt a specificdiagnosis can be made by culturing to thefree-living adult stage, since these differfrom those of S. stercoralis (Grove, 1996).Free-living generations of S. fuellebornitend to develop under semi-aerobic condi-tions.
An experimental infection resulted inan intense itching at the site of entry, fol-lowed by fever, lymphangitis, lymphadeni-tis and cough, with an eosinophilia of 48%in the early stages. Abdominal painoccurred after 20 days and eggs appearedin the faeces after 28 days.
What appeared to be a morphologicallyidentical parasite has been reported in thelast few years from Papua New Guinea(PNG) and Western New Guinea (IrianJaya), although there are no non-humanprimates on this island. In some areas ofPNG an almost invariably fatal disease(‘swollen belly syndrome’) was found inbabies 1–6 months old, with swollenabdomen, respiratory distress, enteric pro-tein loss and peripheral oedema. There wasno fever but over 100,000 eggs g�1 faeces,the eggs often being passed in strings(Ashford et al., 1992).
In other areas there are equally high egg
counts but no clinical disease. Clinical dis-ease might be potentiated by prenatalinfection from symptomless mothers, fol-lowed by a very high exposure to faecalcontamination in bags used to carry babies.Treatment with thiabendazole was veryeffective. The parasite has been designatedS. fuelleborni kellyi Viney, Ashford andBarnish, 1991, because of small differencesfrom the African parasite.
There are many species of Strongyloidesfrom a wide range of mammals in whichthe infective, third-stage, larvae can pene-trate into the skin of humans but developno further. They include: S. canis Brumpt,1922 in dogs, S. cebus Darling, 1911 inmonkeys (human case in Panama), S. felis(Chandler, 1925) in cats (Australia), S.myopotami Artigas and Pacheco, 1933 inrodents and carnivores (Brazil), S. papillo-sus (Wedl, 1956) Ransom, 1911 in sheep, S.planiceps Rogers 1943, in dogs, cats andfoxes (Japan), S. procyonis Little, 1965 incarnivores (USA, experimental), S. ransomiSchwarth and Alicata, 1930 in pig and rab-bit, S. simiae in primates (Guinea-Bissau,experimental), S. westeri Ihle, 1917 inhorse and pig. The free-living males of var-ious species can be differentiated by thestructure of the spicules (Grove, 1996).
Halicephalus (= Micronema) deletrix(Anderson and Bemrick, 1965) is a rhabditi-form saprophyte living in soil (familyCephalobidae) but on three occasions it hasinvaded the brain, liver and heart ofhumans in the USA through deep lacera-tions and caused death through meningo-encephalitis (Gardiner et al., 1981). It hasbeen found more often in horses (femaleworms only) in North America and Europe;albendazole with diethylcarbamazine waseffective in one cutaneous equine case.
Rhabditids (family Rhabditidae) aresmall, free-living, saprophytic nematodesthat are commonly found in soil and therehave been many sporadic reports of infec-tions in humans; adults are usually recov-ered from faeces and some infections maybe spurious. Species reported are:Cheilobus quadrilabiatus Cobb, 1924, infaeces of children in Slovakia, Diploscaptercoronata Cobb, 1913, in the stomach of
The Nematodes 125

patients with low gastric acidity in Texas,Pelodera strongyloides Schneider, 1866,found under the skin of an infant inAlabama, USA, and of a child in Poland(this is a widespread species often found inthe lachrymal fluid of voles), P. teresSchneider, 1866, found in faeces, Rhabditisaxei (Cobbold, 1884) in urine samples inChina and Zimbabwe, R. elongataSchneider, 1866, found twice in girls inSouth Korea, R. inermis (Schneider, 1860)Dougherty, 1955, in faeces of 17 school-children in Japan, R. niellyi Blanchard,1855, in skin of a patient in France, R. pel-lio (Schneider, 1866) Bütschli, 1873 in thevagina of a girl in Mexico, R. tauricaMireckij and Skrajabin, 1965, in the faecesof a child in Russia, R. terricola Dujardin,1845, in the faeces of children in Slovakiaand Rhabditis sp. in faeces of five childrenin South Korea.
Order Strongylida
The human hookworms comprise two par-asites, Necator americanus andAncylostoma duodenale. They both causesimilar disease, including anaemia, havesimilar life histories and require the samecontrol measures. Perhaps because of themovement of peoples, they also havewidely overlapping distributions. In addi-tion, there are animal hookworms that areoccasionally found in humans, and relatednematodes, which in some cases are locallyimportant, adults being found in the intes-tine or, in the metastrongyles includingParastrongylus, the respiratory passages.
Family Ancylostomatidae
Necator americanus (Stiles, 1902) Stiles, 1906
SYNONYMSUncinaria americana Stiles, 1902.
DISEASE AND POPULAR NAMEAncylostomiasis or ancylostomosis (neca-torosis), uncinariasis; New World hook-worm infection.
LOCAL NAMES
Chek kaung (Burmese), Au chung(Cantonese), Nsunsuma (Fanti), Aonobyo,Junishichochu, Kochu, Koekabure,Koemake, Sakanoshita or Tsuchikabure(Japanese), Njowni (Luo), Kokki pulu orKoku panuwa (Singalese), Gooryan(Somali), Njuka safura (Swahili), Puchi(Tamil), Pa-yard pak khow (Thai), Sondono(Twi), Aran inu (Yoruba).
GEOGRAPHICAL DISTRIBUTION
Necator was at one time confined almostentirely to the tropics and subtropics buthas spread widely and is now the domi-nant hookworm in Portugal, northernTurkey, the Caspian Sea area of Iran andareas of Japan. It is widespread in Africasouth of the Sahara, the Americas (Brazil,Colombia, Venezuela, West Indies, south-ern USA), where it was probably intro-duced with the slave trade (althoughAncylostoma has been reported from pre-Columbian mummies), the Far East (south-ern and eastern India, Sri Lanka, Malaysia,China, Indochina, Japan, Indonesia, north-ern Queensland) and the South Pacific. It isabsent from most arid zones and temperateclimates.
LOCATION IN HOST
Adult worms are present in the second andthird portions of the jejunum and rarely inthe duodenum with the buccal capsuleattached to the mucosa. In heavy infectionsthey may be found as far back as the cae-cum.
MORPHOLOGY
The adults are cylindrical, greyish-yellow,nematodes tapering anteriorly and slightlycurved, with the head bent back dorsally(forming a ‘hook’).
The cuticle is smooth with fine trans-verse striations and has lateral cervicalpapillae just behind the nerve ring, atabout the level of the middle of the oeso-phagus. The large buccal capsule opensslightly dorsally and has a pair of short tri-angular lancets at the bottom of the cavity(Figs 57 and 58). These aid in attachmentof the worm to the mucosa and also in cut-
126 Chapter 5

ting off portions for food. A pair of anti-coagulant-secreting glands open into thebuccal capsule, producing a histolytic pro-tease with a molecular mass of 37 kDa.
The muscular oesophagus is cylindricalwith a slight expansion posteriorly and isabout one-sixth of the total length of theworm. The intestine is straight and opensat the posterior end.
The male measures 7–9 mm � 0.3 mm.A single testis lies along the intestine andleads through the seminal vesicle and ejac-ulatory duct into the cloaca. Immediatelynoticeable is the broad, membranous, sym-metrical, caudal bursa, reinforced by rib-like rays. Within the bursa there are twobristle-like spicules about 2 mm long,which are fused at their distal ends to forma barb. The cloaca opens into the bursa.The shape of the bursa differs in Necatorand Ancylostoma and the two species canalso be differentiated by the dorsal ray,which in the former is split to its base (Fig.58). The female measures 9–11 mm �0.4 mm. The vulva usually opens just ante-rior to the middle of the body and from itpaired vaginae lead anteriorly and posteri-
orly to open into the oviducts, followed bylong, slender, coiled ovaries.
LIFE CYCLE (Fig. 59)The transparent egg is thin-shelled andovoid, with blunt, rounded ends; it mea-sures 60 (64–76) µm � 40 (36–42) µm. Theovum is unsegmented when discharged bythe worm but continues to develop and isusually at the 4- or 8-celled stage whenpassed in the faeces. It develops quicklyand may already be at the morula stage inold faecal samples. On reaching soil theembryo develops into a first-stage larva,which hatches out in 24–48 h. The condi-tions for maximum rate of development arewarmth (+25°C), shade, moisture and alight sandy loam. These larvae are termed
The Nematodes 127
Fig. 57. Stereoscan of the anterior end of thehookworm N. americanus.
Fig. 58. Anterior end and caudal bursa of male ofN. americanus (left) and A. duodenale (right).

rhabditiform because the structure of theoesophagus (with an anterior portion con-nected by a thin region to a posterior bulb)resembles that of the free-living nematodeRhabditis. They measure 280–300 µm � 17µm and resemble closely the first-stage lar-vae of Strongyloides, except that hook-worm larvae are slightly longer, lesspointed posteriorly and have a longer buc-cal cavity (Fig. 53). The larvae feed on bac-teria and organic debris in the soil andmoult twice; once on the 3rd day when 400µm long and again on the 5th day when500–700 µm long. The third-stage or filari-form larvae do not feed and retain the shedcuticle for a time. The mouth becomesclosed with a plug and the oesophagusoccupies one-third of the length of thebody. The larvae are now infective; those ofNecator and Ancylostoma can be differen-tiated by the apparent space between theoesophagus and intestine in Necator andby differences in the structure of theoesophageal spears (Fig. 60 and Table 8).
Hookworm larvae may also need to be dif-ferentiated from other nematode larvae insoil or cultured from faeces (Table 9).
The filariform larvae are most numerousin the upper 2 cm of soil, or they may besupported singly or in tufts in the surfacefilm of water on blades of grass. They wavetheir anterior ends in the air in a questing(nictating) manner. They thrive best inshady, moist open soils covered with vege-tation; drying, flooding or direct sunlightare rapidly fatal (Plate 2). Their average lifein the tropics is about 2–6 weeks but somelarvae may survive under favourable condi-tions for up to 15 weeks. The optimal tem-perature range is 28–35°C. They arecapable of ascending 60–90 cm of sandyloam but lateral movement is probably lim-ited to about 30 cm (Udonsi and Atata,1987).
On coming into contact with humanskin, the larvae become active and pene-trate, usually between the toes or fingers.The penetrating larvae respond to essential
128 Chapter 5
Via skin, venules,heart, lungs (alveoli,bronchi, trachea),oesophagus, ileum
Penetrate skin (ormay be ingested withAncylostoma)3RD-STAGE FILARIFORMLARVAE do notfeed and live insoil for about5 weeks
ADULTSin ileum
HUMAN
SegmentingEGGS (60 �m � 40 �m)
in faeces. Hatchin soil in24–48 h
SOIL
1ST-STAGE RHABDITIFORMLARVAE. Feed on
bacteria
Moult twice
3 and 5 days
Fig. 59. Life cycle of human hookworms.

fatty acids; they release proteolyticenzymes and secrete eicosanoids, whichare immunomodulative. They enter
venules and are carried by the venous cir-culation to the heart and then to the lungs.The larvae grow in the lungs, penetratethrough into the alveoli and ascend thebronchi and trachea. They are then swal-lowed and reach the small intestine withinabout 7 days after infection, where theyundergo a third moult; this is followedabout the 13th day by a fourth moult, bywhich time they are sexually differenti-ated. Fertilized females begin egg-laying4–7 weeks after infection and each pro-duces 2000–11,000 (usually 3000–6000)eggs per 24 h. Each female contains onlyabout 200 eggs at one time, so that the pro-tein turnover involved in egg production isenormous.
The adults can live for 1–14 years butusually for about 4. However, in endemicareas, continuous reinfection and lossoccur. This is demonstrated by the fact thatin many areas infection rates are muchhigher after the rainy season (e.g. in west-ern Tanzania there is a variation from 15 to85%).
CLINICAL MANIFESTATIONS
There is an intense itching and burning atthe site of entry. Pruritis may be followedby secondary infection, leading to the con-dition known as ‘ground itch’, withpapulovesicular dermatitis, subsiding inabout a fortnight (Fig. 61).
The larval migration through the lungscan cause a cough due to bronchitis orpneumonitis and patients may pass blood-stained sputum containing eosinophils, butpulmonary manifestations are less markedthan with Ascaris larvae and these earlysigns are not usually reported amongindigenous populations.
Enteritis, with Charcot–Leyden crystalsin the faeces (resulting from breakdown ofeosinophils) and a high eosinophilia char-acterize the early stages of intestinal infec-tion but there is not usually anyeosinophilia in the chronic phase.
A variety of gastrointestinal symptoms,such as abdominal pain, diarrhoea, nauseaand loss of appetite, have sometimes beenattributed to hookworm infection, but themost serious possible effect of an established
The Nematodes 129
Fig. 60. The third-stage infective larvae (L3s) ofNecator (left) and Ancylostoma (right). (a) and (b)Anterior ends; (c) and (d) junction of oesophagusand intestine; (e) and (f) tail ends.

infection is the development of ahypochromic microcytic anaemia, presentin about 36 million individuals worldwide.Hookworms are blood feeders and there isa daily loss of about 5 ml of blood in lightinfections to 100 ml in heavy infections(very roughly equivalent to 2 ml day�1 forevery 1000 eggs g�1 faeces). There may bemany symptoms accompanying anaemia,including lassitude, mental apathy, skinpallor, syncope and angina pectoris. Inchildren, stunting of growth is characteris-tic of severe chronic infections (Fig. 65)and there is evidence that infection alsolowers cognitive ability (Sakti et al., 1999).
PATHOGENESIS
In light and moderate infections anaemia isdue primarily to insufficient iron intakeexacerbated by the presence of the wormsbut in heavy infections it occurs evenwhen the dietary intake is adequate(Pawlowski et al., 1991). This applies toareas such as western Nigeria, where hook-worm anaemia only occurs when manyworms are present (i.e. where there are1000 worms with over 20,000 eggs g�1
faeces day�1). On the other hand, inMauritius, where the average iron intake isonly about 5–10 mg day�1, severe anaemiacan result from the presence of 100 worms.The time factor must also be considered, as
130 Chapter 5
Table 8. Differential features of Necator and Ancylostoma.
Necator Ancylostoma
AdultsSize � 5–9 mm � 0.3 mm � 8–11 mm � 0.45 mm
� 9–11 mm � 0.35 mm � 10–13 mm � 0.6 mmShape Head small, bent acutely in Bow-shaped curve; head
opposite direction to general curve in same line as body
Buccal capsule Almost spherical Elongated, pear-shapedMouth Guarded by 2 semilunar Guarded by two pairs of curved
cuticular cutting plates teeth
FemaleTail No spine Sharp spine (often lost)Vulva anterior to middle posterior to middle
MaleBursa Narrow, longer than wide Outspread, wider than longDorsal ray Deeply cleft. Tip of each cleft Shallow cleft. Tip of each cleft
divided into two divided into threeSpicules Tips usually united and recurved Tips not united
Eggs Indistinguishable
First- and second-stage larvae Indistinguishable
Third-stageinfective larvae
Size Body length 590 µm Body length 550 µmOverall length 660 µm Overall length 720 µm
Tail Short and pointed (63 µm) Long and blunt (85 µm)Sheath Marked striations, especially at Faint cuticular striations
posterior endHead Pointed BluntIntestine Apparent gap between intestine and No apparent gap between
oesophagus intestine and oesophagusOesophageal spears Prominent. Anterior and shaped like Not so prominent. Tips
thistle funnel do not diverge

anaemia will not become apparent untilthe iron reserves have been completelydepleted.
In many areas there is widespread infec-tion but hookworm anaemia is uncommon.
This is usually because of an adequate diet,as in Costa Rica or the southern USA but itmay also be because other resistance fac-tors limit the numbers of hookworms pre-sent, as in India.
The Nematodes 131
Table 9 . The differentiation of infective filariform larvae found in cultures from human faecal specimens.
Strongyloides Trichostrongylus Ancylostoma Nectator Ternidens Oesophagostomum
Length (�m) 500 750 660 590 680 (630–730) 720–950Sheath Absent Present Sheath 720 �m, Sheath 660 Present Present and
striations not �m, striatedclear striations
clear at tail end
Length of �� �� �� �� �� ��oesophagusas proportion of total body length
Intestine Straight Intestinal lumen Anterior end Anterior end Following Large triangularzigzagged narrower in as wide as the oesophagus cells and lumen
diameter than oesophagus is a pair of zigzaggedoesophageal bulb. Gap sphincter cells.bulb. No gap between the Intestinalbetween the oesophagus lumen oesophagus and the somewhatand the intestine intestine zigzagged
Tail Divided End of tail Blunt Sharply pointed Pointed Sharply pointedinto three knoblikeat the tip
Head Blunt Rounded RoundedMouth Mouth spears Mouth spears Buccal capsule +
not very clear clear and oesophagus makeand parallel divergent inverted Y shape
Fig. 61. ‘Ground itch’ at site of entry of larvae of Necator in a sensitized individual. With some non-human species of hookworms, the larvae continue to wander in the skin, causing creeping eruption (alsosee p. 160).

Anaemia occurs because the adultworms pass a steady stream of bloodthrough their intestines in order to obtainoxygen for respiration as well as food, witha consequent loss of erythrocytes and pro-tein-containing fluid. The amount of bloodpassed by an adult Necator (0.03–0.05 mlper worm day�1) is less than in an equiva-lent Ancylostoma infection (0.14–0.25 mlper worm day�1) because of its smallersize. Reabsorption of haemoglobin from theintestine occurs, but the amount (about11%) is less than often assumed in thepast; in many areas an adequate dietaccompanied by intravenous iron would bemore effective in suppressing the clinicalsigns than anthelminthic treatment.
There is no proof that hookwormsdepress haemopoiesis but they do appearto decrease erythrocyte life. In addition toanaemia, a characteristic hookwormoedema occurs in heavy infections, resem-bling the nephrotic syndrome. This iscaused by low plasma protein (particularlyalbumin) but usually only becomes appar-ent when there is a low iron intake.However, a heavy worm load of about 2000adults can cause a loss of 4 g of albumineven with an adequate dietary supply ofiron.
The extensive damage to the mucosa,including necrosis and haemorrhage, some-times recorded in cases of hookworm infec-tion is probably due to secondary causes,as is vitamin B12 and folic acid deficiency.Similarly, there does not appear to be adirect relationship between hookworminfection and malabsorption (contrastStrongyloides (p. 119)).
As with the other geohelminths(Haswell-Elkins et al., 1987a) and theschistosomes, a small minority of infectedindividuals have high worm burdens withserious disease and are responsible formost of the eggs reaching the environment(Anderson and Schad, 1985).
DIAGNOSIS
Clinical. Chronic anaemia and debility inpatients living in an endemic area are sug-gestive, but may also be caused by mal-
nutrition, malaria, amoebiasis and otherhelminthiases.
Parasitological. Recovery and recognition ofeggs in faecal samples (Fig. 123) are thestandard method of diagnosis. The eggs areidentical to those of Ancylostoma and verysimilar to those of Ternidens andOesophagostomum. Ova must be culturedto the infective larva stage if it is requiredto differentiate between the two hookwormspecies (Table 8 and Fig. 59). The Harada–Mori culture method is given on p. 262).
Serological. Not often employed.Complement fixation and indirect haemag-glutination tests work well with antigenobtained from adult worms. ELISA andradioimmunological assay (RIA, measuringIgE) both have good specificity when ESantigens are used.
Recently a gene for Ancylostomasecreted polypeptide (ASP), a protein pro-duced by infective larvae of A. caninum,has been cloned and the amino acidsequence determined. This is very similarto an insect venom that elicits a strongimmune response and is being researchedas a possible vaccine (Hotez and Prichard,1995).
TREATMENT
A single course of chemotherapeutic treat-ment aimed at permanent cure in outpa-tients living in an endemic area is unlikelyto be successful, but it is neverthelessimportant to reduce the number of wormspresent below the level causing clinicalsymptoms. This level will vary in differentparts of the world, depending mainly oniron intake and nutritional status. Anydrug given should also be supplemented byoral (or intravenous where supervision isnot possible) iron therapy. In light infec-tions a single annual treatment is suffi-cient, but in heavy infections 6- monthlytreatment is necessary. The properties ofthe various drugs are given in Albonico etal. (1998).
Albendazole. A single dose of 400 mg for allages above 2 (60–90% effective) is also
132 Chapter 5

active against most other intestinal nema-tode and cestode infections.
Mebendazole. A single dose of 500 mg forall ages above 2 or 100 mg twice daily for 3days (20–90% effective) is also effectiveagainst other intestinal nematodes.
Levamisole. A single dose of 3 mg kg�1
body weight is more effective againstAncylostoma than against Necator(20–60%).
Pyrantel pamoate. A daily dose of 10 mgkg�1 body weight (max. 1 g) for 3 days.More effective against Ancylostoma thanagainst Necator (20–90%) – chemical for-mula: (E)-1,4,5,6-tetrahydro-1-methyl-2-(2-(2-thienyl)vinyl]-pyrimidine.
Bitoscanate, bephenium and tetrachlor-ethylene can no longer be recommended,while ivermectin is not effective.
In a study of 3000 young children inZanzibar, it was estimated that 1260 casesof moderate and 276 cases of severeanaemia were prevented by treatment withmebendazole, with improved motor devel-opment. In children in East Africa learningpotential improves for months after treat-ment, with much higher school attendance(about 100 million children worldwide donot attend school because of soil-transmit-ted nematode infections).
EPIDEMIOLOGY
It has been estimated that there are from1050 to 1277 million cases worldwide(about two-thirds due to Necator) with90–130 million suffering morbidity and65,000 deaths annually, principally fromthe anaemia. Hookworm infection is essen-tially a rural disease of poverty and is par-ticularly common in cocoa, coffee andbanana plantations, owing to the denseshade and high rainfall (Plate 13). Heavyinfections are not found in areas withmuch less than 100 cm of rain per year andare rare in arid regions.
Recent estimates of infection rates invarious regions are: 46% in Cameroon, 6%in peninsular Malaysia, 14–50% in south-
ern Nigeria, 68% in Bobei County in China,2.5% in Burma (Myanmar), 69% inIndonesia and 54% in Surinam.
Soil type is of great importance to thelarvae, which migrate up against the waterflow and down again when the top layer ofsoil is drying. Clay soils are less well aer-ated and, where there is heavy rainfall, thelarvae are not able to survive so well as insandy soils. This is in contrast to the situa-tion with Ascaris and Trichuris eggs, whichsurvive better on clay soils, as they dry outless in drought and have better adhesiveproperties. Larvae are also better able tomigrate in well-filled soil that has a crumbstructure than in highly compacted soil.
PREVENTION AND CONTROL
Personal prevention is provided by thewearing of shoes.
The control of hookworms has a longhistory, beginning with the campaign of theRockefeller Sanitary Commission to elimi-nate the infection from the southern USAin 1909, followed by the internationalefforts of the Rockefeller Foundation in1913. The strategies used then of masschemotherapy and improvements in sanita-tion are still appropriate today. Such mea-sures were also very effective in Japan inthe 1950s–1980s (from 4.5% with hook-worms and 60% with Ascaris in 1950 to 0and 0.01% in 1987).
The newer single-dose, broad-spectrum,anthelminthics are a great advance formass campaigns against morbidity causedby all the geohelminths, which are underway in many countries, funded by manyinternational and national agencies and bynon-governmental organizations (NGOs).For instance, a campaign for annual treat-ment of all schoolchildren (6–14 years old)in Ghana with albendazole has begun, pri-marily to reduce disease rather than inter-rupt transmission, which is not feasible atpresent. The World Health Organization(WHO) target is that by 2010 regularchemotherapy will be provided for at least75% of schoolchildren in endemic coun-tries at risk of morbidity (a global schoolhealth initiative against both geohelminthsand schistosomes is being supported by
The Nematodes 133

many international agencies). However, thepossibility of resistance to drugs needs tobe carefully monitored, particularlybecause of the sale of substandard productsin some countries (Crompton, 2000).
There are three possible strategies forchemotherapy campaigns against all thegeohelminths.
1. Universal (= mass) campaigns, in whichcommunities are treated irrespective ofinfection status, age, sex or social charac-teristics.2. Targeted campaigns with group-levelapplication, irrespective of infection status.3. Selective campaigns with individualapplication, based on diagnosis of currentinfection or on level of infection.
The first is feasible if there are sufficientfunds; otherwise, the second is most cost-effective, with treatment aimed at high-riskgroups, such as school and preschool chil-dren and women of childbearing age, whilethe third is expensive, difficult to imple-ment and likely to be unacceptable to themajority of infected individuals.
Wider utilization of cheap and effectivesanitation technologies, such as the venti-lated improved pit (VIP) latrine, first intro-duced in Zimbabwe, or the pour-flushtoilet, which provides a water seal anddoes not require piped water, needs to beencouraged in addition to chemotherapy.Health education and the provision oflatrines are also needed to prevent indis-criminate defecation in the fields.
Integrated campaigns, together withimmunization or general school healthefforts, can often make the most effectiveuse of primary health-care workers.
There are large-scale control campaignsunder way in Mexico, Seychelles, SouthAfrica (Natal), Sri Lanka and Zanzibar(Albonico et al., 1998).
ZOONOTIC ASPECTSVirtually none, although Necator is foundoccasionally in chimpanzees and pigs.Young hamsters can be successfullyinfected in the laboratory, which is stimu-lating research into the physiology, pathol-ogy and particularly the immunology of
infection, although the fecundity of theparasites is suppressed.
Ancylostoma duodenale (Dubini, 1843) Creplin, 1845
SYNONYMSAgchylostoma duodenale Dubini, 1843;Ankylostomum duodenale; Uncinaria duo-denalis.
DISEASE AND POPULAR NAMES
Ancylostomiasis or ancylostomosis, unci-nariasis; Old World hookworm infection.
LOCAL NAMES
As for Necator.
GEOGRAPHICAL DISTRIBUTION
Ancylostoma extends into more temperateregions than Necator. It is present in theNorth African and EuropeanMediterranean littoral; Asia (northernIndia, Myanmar, Cambodia, Laos, Vietnam,Malaysia, Indonesia, China, Taiwan,Japan); the South Pacific; and the Americas(particularly Chile, Mexico, Venezuela).
LOCATION IN HOST
As for Necator.
MORPHOLOGY
Ancylostoma is a larger and more robustworm than Necator. The head is curveddorsally in a smooth curve so that speci-mens do not have the hook-like appearancecharacteristic of Necator. The large buccalcapsule has bilaterally symmetrical curvedcutting processes, consisting of two fusedteeth, and there is a pair of internal teeth atthe bottom of the capsule, with histolyticglands at the base. These aid the attach-ment of the worm to the mucosa and alsoassist in cutting off portions for food (Figs.62 and 63). The features differentiatingAncylostoma from Necator are shown inTable 8.
The male measures 8–11 mm �0.4–0.5 mm; the shape and structure of thecaudal bursa differs from that of Necator(Fig. 58).
134 Chapter 5
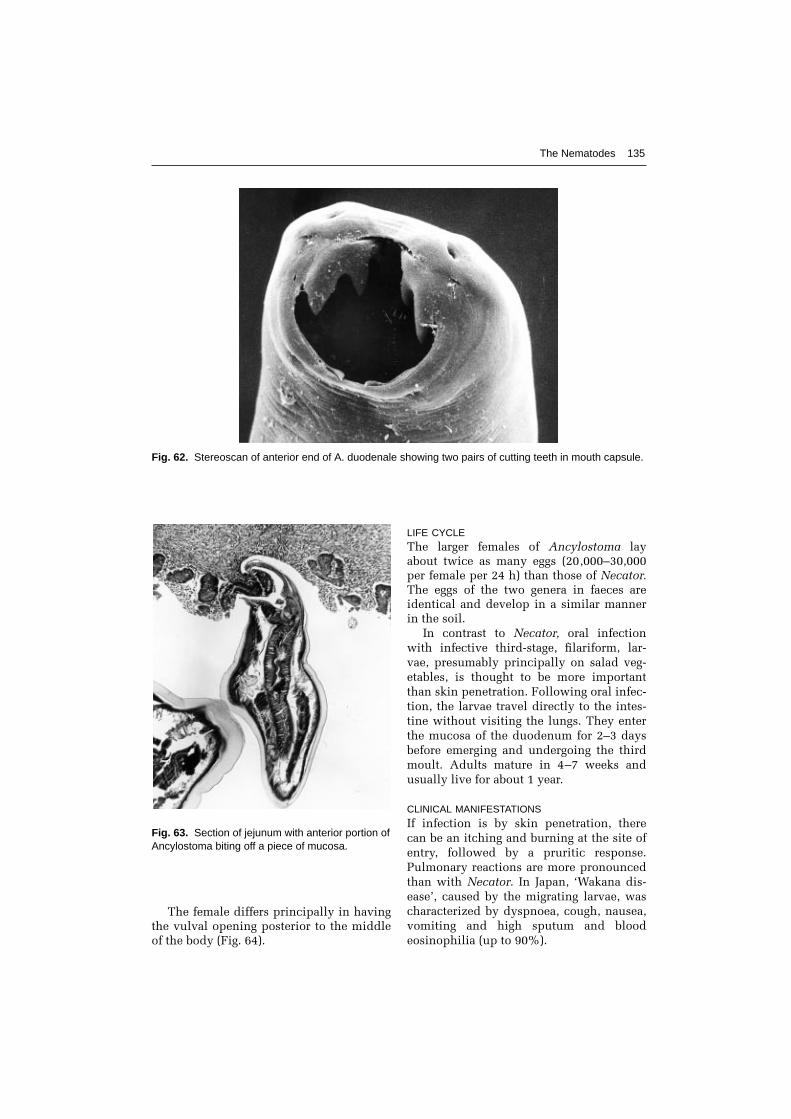
The female differs principally in havingthe vulval opening posterior to the middleof the body (Fig. 64).
LIFE CYCLEThe larger females of Ancylostoma layabout twice as many eggs (20,000–30,000per female per 24 h) than those of Necator.The eggs of the two genera in faeces areidentical and develop in a similar mannerin the soil.
In contrast to Necator, oral infectionwith infective third-stage, filariform, lar-vae, presumably principally on salad veg-etables, is thought to be more importantthan skin penetration. Following oral infec-tion, the larvae travel directly to the intes-tine without visiting the lungs. They enterthe mucosa of the duodenum for 2–3 daysbefore emerging and undergoing the thirdmoult. Adults mature in 4–7 weeks andusually live for about 1 year.
CLINICAL MANIFESTATIONS
If infection is by skin penetration, therecan be an itching and burning at the site ofentry, followed by a pruritic response.Pulmonary reactions are more pronouncedthan with Necator. In Japan, ‘Wakana dis-ease’, caused by the migrating larvae, wascharacterized by dyspnoea, cough, nausea,vomiting and high sputum and bloodeosinophilia (up to 90%).
The Nematodes 135
Fig. 62. Stereoscan of anterior end of A. duodenale showing two pairs of cutting teeth in mouth capsule.
Fig. 63. Section of jejunum with anterior portion ofAncylostoma biting off a piece of mucosa.

Weakness, anorexia and pica (dirt-eat-ing) are sometimes found in heavy chronicinfections.
PATHOGENESIS
This is similar to infection with Necatorbut, because an adult Ancylostoma islarger, the anaemia caused by an equivalentnumber of worms is more severe.
DIAGNOSIS AND TREATMENT
As for Necator.
EPIDEMIOLOGY
As for Necator. In many areas both types ofhookworm are present, possibly in thesame patient.
Infection rates recorded recently for var-ious areas where specific identification hasbeen made are: north-eastern Australia inAboriginals 31–77%, Benin 25–35%,
China (Fujien Province) 50%, (SichuanProvince equally with both species) 74%,Egypt 6–8%, Ethiopia 67%, Kenya(preschool children in Kilifi) 29%, south-ern Nepal 11–79%, Paraguay (school chil-dren) 59% with both species.
The ova can develop at a lower tempera-ture (a minimum of 14°C) than those ofNecator, so that infection is more commonin subtropical and temperate climates. Theoptimum temperature for hatching of eggsis 20–27°C and they fail to develop above40°C. Infection was formerly common intin miners in England and in workersbuilding the St Gotthard tunnel betweenSwitzerland and Italy.
PREVENTION AND CONTROL
As for Necator except that the thoroughwashing of salad vegetables is more impor-tant than the wearing of adequate footwear.
136 Chapter 5
testis
seminal vesicle
ejaculatory duct
cement gland
ovaryuterus
dorsal ray
gubernaculum
intestine
spicule
Fig. 64. Vulval region of female of A. duodenale and caudal bursa of male showing process of fertilization.

ZOONOTIC ASPECTS
None.
Ancylostoma ceylanicum Looss, 1911, is aparasite of dogs in Asia and South Americabut also occurs as a human parasite inIndia and the Philippines. Infection is bythe oral route, with a prepatent period ofabout 25 days. Infections are light andblood loss does not appear to have anyclinical significance; the larvae do notcause creeping eruption. Diagnosis is byculturing the larvae to the third stage.Chemotherapy is effective.
Recently, 200 cases of infection withthe dog hookworm, Ancylostomacaninum, have been diagnosed inQueensland, Australia, with another 200probable cases. All were Caucasians andpresented with severe abdominal pain,sometimes also with diarrhoea and weightloss, followed by blood eosinophilia(Croese et al., 1994; Prociv and Croese,
1996). In this area this is the leading causeof eosinophilic enteritis and patients mayhave aphthous ulcers of the terminalileum, caecum and colon. In 15 cases,adults were seen in situ and in nine thetypical buccal capsule with three teethcould be recognized in biopsy tissue sec-tions. The adults produced no eggs butwere diagnosed by biopsy or by an IgGELISA. Recently, a Western blot using ESantigens from adult worms to identify IgG4antibodies to a protein of 68 kDa (Ac68)was found to be more specific and sensi-tive than the ELISA. However, humanspecies are likely to cross-react and thespecific IgE ELISA for Necator may be nec-essary in order to differentiate.
All patients had dogs and walkedaround the garden in bare feet and it isassumed that infection was contractedfrom front lawns, which were watered withsprinklers and thus washed dog faeces intothe lawn. There were fewer cases in thewinter months. The patients were in anaffluent area with good hospital facilitiesfor the difficult diagnosis and it is likelythat infection with this species also occursin other parts of the world. One case hasbeen reported from New Orleans, USA, inan 11-year-old girl who had chronicabdominal pain, vomiting, anorexia andweight loss, which resolved after two dosesof mebendazole.
Various hookworms of animals veryoccasionally parasitize humans: A. japon-ica Fukuda and Katsurada 1925, possiblyin Japan; A. malayanum (Allesandrini,1905), a natural parasite of bears;Cyclodontostomum purvisi Adams, 1933,in Thailand and naturally in the rat;Necator argentinus Parodi, 1920 (doubtful);N. suillus Ackert and Payne, 1922, once ortwice in humans and widespread in pigs.Species causing cutaneous larva migrans(creeping eruption) are considered onp. 160.
Other Strongyles
As well as the hookworms, there are vari-ous other bursate intestinal nematodes that
The Nematodes 137
Fig. 65. Severe stunting of growth in children withvery heavy hookworm infections. The eldest childis aged 15 with a haemoglobin level of 1.8 g dl�1 .Height of windowsill equals 85 cm.

can sometimes be recovered from humans.Almost all are principally zoonotic infec-tions, but a few, such as Trichostrongylus,Ternidens and Oesophagostomum, do havehuman-to-human transmission in someareas.
Family Trichostrongylidae
Trichostrongylus spp. Looss, 1905
POPULAR NAME
Wireworms (when in domestic stock).
DISTRIBUTION
Species of Trichostrongylus occur locallyin humans in Iran (67% in Isfahan), Iraq(up to 10%), Israel, Egypt (11–60%),Ethiopia (23%), Armenia, Japan, Korea(80% in one area), Indonesia and the CapeVerde Islands. Sporadic cases may occur inmany areas where people and cattle are inclose proximity (e.g. Africa, Australia,India and South America).
LOCATION IN HOST
The adults attach to the wall of the duode-num and jejunum by the anterior end.
MORPHOLOGY
These are small thin strongyle worms (family Trichostrongylidae) measuring5–10 mm in length; most species are nor-mally parasitic in ruminants. They have nobuccal capsule and live with the headembedded in the mucosa of the small intes-tine. The males have a well-developed cop-ulatory bursa, and the structure of thespicules provides the most useful featureused to differentiate the 11 species (T. affi-nis, axei, brevis, calcaratus, capricola, col-ubriformis, lerouxi, orientalis, probolurus,skrjabini and vitrinus) recovered fromhumans; in all they are short, equal, stoutand spoon-shaped but have small specificdifferences.
Eggs found in the faeces are similar tothose of hookworms but are longer and nar-rower (75–100 µm � 30–50 µm) with more
pointed ends, and they usually contain asegmented ovum at the morula stage whenpassed in the faeces.
LIFE CYCLE
Eggs passed in the faeces at the morulastage hatch in the soil in about 24 h. Thelarvae reach the infective third stage inabout 3 days. These occasionally penetratethe skin, but in human infections they arealmost always ingested. When this hap-pens, the larvae develop into adults in 25days; in contrast to hookworms, they neverundergo a lung migration.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Infection is usually light and causes noclinical symptoms. In heavier infections (afew hundred worms) there may be abdomi-nal pain, diarrhoea and a slight anaemia,accompanied by a mild eosinophilia(under 10%).
The head of the adult worm producestraumatic damage to the mucosa, withinflammation at the site of attachment. Theworms may also suck blood but the amountis not sufficient to cause anaemia.
DIAGNOSIS
This is by recognition of eggs in the faeces(Fig. 124), which gives identification to thegeneric level.
The larvae can also be cultured, usingthe techniques for hookworms (p. 262).The infective third-stage larvae must bedifferentiated from those of hookwormsand Strongyloides (Table 9). The species oftrichostrongyle involved can only be deter-mined if adults are obtained afterchemotherapy, and veterinary textbooksshould be consulted.
TREATMENT
All the drugs used for hookworms workwell against trichostrongyles, which areusually easier to eliminate. Mebendazolehas been used successfully at 100 mg twicedaily for 3 days for adults.
EPIDEMIOLOGY, PREVENTION AND CONTROL
Larvae are usually ingested on contami-nated vegetables and these should be
138 Chapter 5

thoroughly washed or cooked. The eggsand infective larvae are both very resistantto desiccation, which makes control diffi-cult. In areas of Iran and Iraq cattle are keptin the backyards of houses (Plate 14) and aparticularly heavy build-up of larvaeoccurs after the rainy season.
It was reported in 1953 that there were48 million human cases worldwide; it isnot known if prevalence is now muchlower.
ZOONOTIC ASPECTS
In many areas infection is contracted fromlivestock, although with T. orientalis fromsheep and camels, which is the most com-mon species in humans in China, Japan,Iran, Iraq, Korea and Taiwan, there is prob-ably a human cycle involved.
Other trichostrongylids that are com-mon parasites of domestic stock in manyareas of the world, such as Haemonchuscontortus, Marshallagia marshalli,Mecistocirrus digitatus, Nematodirusabnormalis, Ostertagia circumcincta andO. ostertagi, may occasionally occur inhumans, particularly in Iran andAzerbaidjan. For further details of theseparasites, textbooks of veterinary medicineshould be consulted (see GeneralReferences, p. 282).
Family Chabertiidae
Ternidens deminutus Railliet and Henry, 1909
SYNONYMS
Triodontophorus deminutus Railliet andHenry, 1905.
DISEASE AND POPULAR NAME
Ternidens infection (ternidensiasis orternidensosis), false hookworm infection.
GEOGRAPHICAL DISTRIBUTION
Comoros Islands, Congo Republic,Mauritius, southern Tanzania andZimbabwe (up to 87% in one area).
LOCATION IN HOST
Adults occur in the colon and occasionallythe ileum, with the anterior end attached.
MORPHOLOGY
The adults superficially resemble hook-worms but are rather larger. The subglo-bose buccal capsule of Ternidens isdirected anteriorly and has forked teeth inits depths. The mouth is surrounded by adouble ‘leaf crown’ or corona radiata ofcuticular bristles.
Females measure 12–16 mm � 0.65–0.75 mm. The vulva opens just in front ofthe anus. Males measure 9.5 mm �0.55 mm and have a caudal bursa with therays arranged in a characteristic pattern.The spicules are long and equal, measuringabout 0.9 mm in length.
LIFE CYCLE
Eggs measure 81 (70–94) µm � 52 (47–55)µm. They pass out in the faeces, usually inthe 8-celled stage, and develop into rhab-ditiform larvae in about 30 h. The larvaefeed on bacteria and moult twice (once onthe 6th day and again in 8–10 days at29°C). The third-stage larva retains the castcuticle as a sheath, but it is not known howtransmission to humans occurs. Allattempted experimental infections eitherby mouth or by skin penetration havefailed. It is possible that an invertebrateintermediate host, such as the termite, isrequired (Goldsmid, 1991).
CLINICAL MANIFESTATIONS
The great majority of cases have a lightworm load and appear to be symptomless,although little is known of the effects of theparasites and they could be involved in theformation of intestinal nodules.
PATHOGENESIS
The parasites feed on blood and probablycause ulceration of the mucosa. While theloss of blood from the large bowel could bemore serious than that caused by hook-worms in the jejunum, as no reabsorptionoccurs, anaemia has not been reported.This might be because usually only a fewworms are present.
The Nematodes 139

In experimental studies in rhesus mon-keys, adults eventually become encapsu-lated in nodules, but it is not known if asimilar effect occurs in humans.
DIAGNOSIS AND TREATMENT
Diagnosis is by recognition of the eggs infaeces (Fig. 124); concentration methodscan be used if necessary. The eggs must bedifferentiated from those of hookworms,which can be done with difficulty by thelarger size and particularly by the largervolume occupied by the dividing ovum.For more positive identification the larvaecan be cultivated to the infective thirdstage (p. 262 and Table 9). These have char-acteristic paired sphincter cells at the junc-tion of the oesophagus and intestine.Ternidens is still frequently misdiagnosedas hookworm.
Chemotherapy is the same as for hook-worm; albendazole at 400 mg was success-ful in a trial in children (Bradley, 1990).
EPIDEMIOLOGY AND ZOONOTIC ASPECTS
Ternidens is a very common parasite ofmonkeys in tropical areas of Africa andAsia (China, Indonesia, Malaysia, southernIndia and Vietnam) and these probably actas reservoir hosts. It is not known whyhuman infection is restricted to areas ofAfrica, where it may be very common, butthis is presumably related to its (unknown)mode of transmission. One case has beendiagnosed from Surinam.
Oesophagostomum bifurcum(Creplin, 1849)
DISEASE AND POPULAR NAME
Oesophagostomiasis or oesophagostomosis;nodular worm (for species in veterinarypractice).
GEOGRAPHICAL DISTRIBUTION
Oesophagostomum bifurcum is astrongylid nematode (family Chabertiidae)and has been reported many times fromhumans in West Africa and also from EastAfrica and Asia. It is a common parasite ofmonkeys, while some other species may be
found in ruminants and pigs. O. aculeatum(Linstow, 1879) has been reported onceeach from a human in Brunei andIndonesia (Ross et al., 1989), Oesopha-gostomum sp. from the peritoneal cavity ofa patient in Malaysia and O. stephanosto-mum Railliet and Henry, 1909, very rarelyfrom Côte d’Ivoire, with a case from Braziland probably another from Surinam.
LOCATION IN HOST
Adults in nodules in the wall of the colonand caecum.
MORPHOLOGY
Species of Oesophagostomum resemblehookworms in shape and size but have abuccal capsule that opens forwards, with acuticular ‘leaf crown’ surrounding themouth opening, and a cylindrical ratherthan globose buccal capsule. The caudalbursa of the male is symmetrical and thelong spicules are equal. Each female hasbeen estimated to produce about 5000 eggs24 h�1.
LIFE CYCLE
The eggs closely resemble those of hook-worms, measuring 58–69 µm � 39–47 µm.Larvae hatch in 2 days at 30°C and developin the soil into third-stage infective larvaein 5–7 days. These are ingested, invade thewall of the colon and caecum and becomeencapsulated. A nodule forms, measuringabout 1.5 cm in diameter, with a fibrous tis-sue wall and having a living worm in thecentre of an unruptured abscess (Fig. 66).When the worms mature, they break out ofthe nodule in about 6–7 days and some-times become attached to the caecal wall.Shrunken third-stage larvae inside thesheath can survive for a considerableperiod in the environment under desic-cated conditions.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Some worms apparently mate in the lumenof the colon and cause no symptoms.However, others may stay in small nodulesin the intestinal wall or penetrate throughinto the abdominal cavity, where they cancause significant pathology. There can be
140 Chapter 5

dozens of nodules, each 2–3 cm in diame-ter. The wall of the colon becomes grosslythickened, although the mucosa and serosaremain intact. There is no definite cyst wallbut there is a layer of macrophages, epithe-lial cells and fibroblasts, and often manyplasma cells and eosinophils. Nodules con-tain thick pus or caseous material. In themajority of cases the infection is self-limiting and eventually resolves withouttreatment.
In northern Ghana and Togo and proba-bly southern Burkina Faso, infected indi-viduals often have an epigastric orperiumbilical swelling recognized by localpeople and known as a ‘tumeur deDapaong’ (Plate 15). These may be accom-panied by considerable pain, and abdomi-nal occlusion or the presence of theabscesses may require surgical interven-tion. Many cases are in children, mostabout 10 years old. The wall of the colon isoften oedematous and the whole of thebowel loop becomes hardened and sur-rounded by adhesions. There may also beinvolvement of the liver, and worms in theabdominal cavity can cause peritonitis
(Polderman et al., 1991, 1999; Poldermanand Blotkamp, 1995). Nodules sometimescalcify and then show up on X-ray. In acase of infection in Brunei, probablycaused by O. aculeatum, a single wormwas removed from a painless lump on theback measuring 3 cm and containing thickbrown pus. Infection may have been con-tracted by direct penetration (Ross et al.,1989).
DIAGNOSIS
Clinical diagnosis is often difficult andoesophagostomiasis has often been mis-taken for other conditions, such as carci-noma, appendicitis or amoebiasis;laparotomy will provide a positive diagno-sis and ultrasound can be useful.
In order to differentiate the eggs in fae-ces from those of hookworms, the larvaehave to be cultivated to the third stage(Table 9). An activated charcoal/faecesfilter-paper technique for 5–7 days workswell (p. 123). The larvae measure 720–950µm and have a cuticle with prominent stri-ations, 28–32 (or sometimes 20) large, char-acteristic triangular-shaped intestinal cells
The Nematodes 141
Fig. 66. Nodular lesion caused by O. bifurcum presenting as a para-umbilical mass in an 8-year-old girlin Nigeria. At laparotomy the mass was found adhering to muscles and fascia of the anterior abdominalwall of the ascending colon (magnification � 11). Sections of the nematodes can be seen.

and a buccal capsule that differs from thatof hookworm larvae (Blotkamp et al., 1993;Pit et al., 1999).
An ELISA based on detection of specificIgG4 in sera had a specificity of over 95%in one study and may prove useful indifferentiating Oesophagostomum fromhookworm in epidemiological studies(Polderman et al., 1993).
TREATMENTSurgery to resect the infected portion of thecolon is sometimes necessary.
Albendazole (400 mg or better 10 mgkg�1 body weight daily for 5 days) andpyrantel pamoate (two doses of 10 mgkg�1) are both effective but thiabendazoleand levamisole are not.
EPIDEMIOLOGY AND CONTROLIt is likely that infection with O. bifurcumin northern Ghana and Togo and maybeUganda now involves human-to-humantransmission and monkey reservoirs are nolonger necessary. In the focus identified inWest Africa it is estimated that there areabout a quarter of a million cases and insome villages in rural areas there are infec-tion rates of 30–60%, although infectionsare rarely found in young children(Polderman et al., 1991, 1999; Poldermanand Blotkamp, 1995). Transmission is lim-ited to the rainy season, but it is not clearexactly how humans become infected.Another species, O. apiostomum (Willach,1891) Railliet and Henry, 1905, a parasiteof monkeys in West Africa, has beenreported from the colon of a patient inTogo, but specific identification of humancases is difficult unless adults are expelled.
Infections are easier to treat than hook-worm infections and should be muchreduced by any hookworm control campaign.
Family Syngamidae
Mammomonogamus laryngeus(Railliet, 1899) Ryzhikov, 1948
SYNONYMS
Syngamus laryngeus Railliet, 1899.Syngamus kingi Leiper, 1913. The genus
name Syngamus is now used only for thespecies in birds, such as S. trachea causing‘gapeworm’ in poultry, although S. laryn-geus may still be found in some textbooks.
DISEASE NAME
Mammomonogamiasis or mammomonog-amosis.
GEOGRAPHICAL DISTRIBUTION
There have been over 90 reported casesfrom humans, the majority in SouthAmerica (25 from Brazil), with a few casesin North America, the West Indies andChina and single cases from Australia,Korea, the Philippines and Malaysia (orpossibly Thailand).
LOCATION OF ADULTS
Adults inhabit the air passages of the upperrespiratory tract, particularly the larynx orpharynx.
MORPHOLOGY AND LIFE CYCLE
The male worm (4 mm � 0.35 mm) ismuch smaller than the female (20 mm �0.5 mm) and the two are joined in perma-nent copulation, giving a characteristic Yshape (family Syngamidae). The buccalcapsule of both sexes is well developedand globular. It is used to attach the wormsto the air passages, where they suck blood.The copulatory bursa and supporting raysof the male have characteristic structuresthat are of specific importance.
The eggs measure 85–90 µm � 50 µmand have a sculptured appearance.Infective third-stage larvae develop fromeggs in 9–15 days at 26–30°C and are prob-ably ingested on salad vegetables. Larvaeare believed to reach the trachea via thebloodstream and lungs.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
There is an incubation period of 25–40days. Worms are found in the trachea, lar-ynx and sometimes the smaller bronchiand produce headache, nausea, irritationand a severe dry cough. There may also beblood in the sputum (haemoptysis). Aneosinophilia of 23% has been reported.
142 Chapter 5

DIAGNOSIS AND TREATMENT
Adults can usually be seen in the larynxand pharynx and eggs may be found in thesputum. Adults are sometimes expelledafter coughing and a recent Thai patientwho had worked in Malaysia coughed up96 worms.
Benzimidazoles, such as albendazole ormebendazole would probably be effective.Treatment with thiabendazole was effectivein one case.
ZOONOTIC ASPECTS
This is entirely a zoonotic infection.Natural hosts are cattle, water-buffaloes,goats and, in Thailand, wild felids. A sin-gle human case of infection with M. nasi-cola (Buckley, 1934), a similar speciesfound in a wide range of herbivores, hasbeen reported.
Family Angiostrongylidae
Parastrongylus cantonensis (Chen, 1935) Chabaud, 1972
SYNONYMS
Pulmonema cantonensis Chen, 1935; Angio-strongylus cantonensis Dougherty, 1946.
DISEASE
Parastrongyliasis or parastrongylosis,angiostrongyliasis. A cause of eosinophilicmeningitis.
GEOGRAPHICAL DISTRIBUTION
This is entirely a zoonosis, occasionalcases occurring in Indonesia, Laos,Malaysia, many Pacific islands, thePhilippines, Taiwan, Thailand andVietnam. In recent years it has also beenreported from humans in Australia, China,Côte d’Ivoire, Cuba, Dominican Republic,India, Japan, Mauritius, Puerto Rico,Reunion and the USA and the infectionappears to be spreading.
LOCATION IN HOST
P. cantonensis does not usually develop tomaturity in humans, the third-stage larvaemigrating to the brain, CNS or eyes.
MORPHOLOGY
The third-stage larva measures0.46–0.5 mm � 26 µm when ingested. Ithas small lateral alae and characteristicchitinous rods in the buccal cavity.
The adult worms are normally found inthe main branches of the pulmonary arteryof rats, but not in humans. They areslender strongyloid nematodes (superfam-ily Metastrongyloidea; many species inlivestock are known as lungworms), with asmall buccal capsule containing two lateraltriangular teeth. Females measure22–34 mm � 0.34–0.56 mm. The vulvaopens near the anus and the red, blood-filled gut and white uterus give a character-istic ‘barber’s pole’ effect. The malemeasures 20–25 mm � 0.32–0.42 mm andhas a well-developed caudal bursa, withsubequal spicules (1.02–1.25 mm long).
LIFE CYCLE
In rats, the eggs hatch in the lungs and first-stage larvae are passed out in the faeces;they can survive up to 2 weeks in water. Forfurther development the larvae need to beingested by or to penetrate into snails orslugs. They moult twice in the muscles andthe infective larvae remain coiled up in themantle. They are infective in 2–3 weeks at26°C. When an infected mollusc is eaten byanother rat, the larvae penetrate the intesti-nal wall and are carried by the circulatorysystem via liver, heart and lungs to the mainarterial circulation. They then reach theCNS and congregate in the spinal cord,medulla, cerebellum and diencephalon.After moulting, the fourth-stage larvaemigrate into the subarachnoid space and26–33 days after infection they reach thepulmonary arteries. Here they become sexu-ally mature in another 7 days.
In humans the third-stage larvaeingested in snails or slugs can migrate viathe liver, heart and lungs to reach the CNS.
CLINICAL MANIFESTATIONS
There is an incubation period of 12–36days before symptoms of severe headache,back and neck stiffness, extreme tiredness,photophobia, nausea, vomiting and vertigooccur. There is a generalized hyperaesthesia
The Nematodes 143

of the skin, with signs of paralysis of one ormore nerves and eye muscles and facialparaesthesia (burning, tingling, etc.). Thesymptoms usually last for about 2 weeksbut may be prolonged.
Mortality due to the disease is generallylow (although in Thailand a mortality rateof 1.2% has been reported) and most casesare self-limiting, with recovery occurringin about 4 weeks.
PATHOGENESIS
There are two distinct forms of the disease;more typical is the cerebral form, with ameningitic syndrome, but there is also amuch rarer ocular form, with acute inflam-mation of the eye but no signs of meningi-tis (Wariyapola et al., 1998).
Most of the knowledge of the pathology ofthe disease has been obtained from experi-mental infections in animals. However, inautopsy cases, worms have been found inthe brain, with inflammation spreadingalong the perivascular spaces into the brainsubstance. The inflammatory reaction in thebrain and meninges is localized, particularlyin the vicinity of the parasites under thearachnoid (Fig. 67). Eosinophils, Charcot–Leyden crystals and giant cells are apparenthistologically. Dead larvae are found inside
granulomatous lesions with necrotic debrisin the cerebellum, pons and medulla, deathin these cases probably being due to impair-ment of the bulbar functions.
Eosinophilia is common in this infec-tion and there is a raised cell count in thecerebrospinal fluid (in one fatal case therewere 880 cells mm�3, of which 92% wereeosinophils). In experimental animals aninflammatory reaction develops after a fewweeks, with leucocytic infiltration and apreponderance of eosinophils in themeninges and the parenchyma of the CNS.The severity of the disease is related to thenumber of third-stage larvae that reach theCNS, most of the pathology being causedby the dead worms.
DIAGNOSIS
Clinical diagnosis is usually possible, par-ticularly if there is evidence of raw snailshaving been eaten. Particularly importantis the eosinophilic pleocytosis of the cere-brospinal fluid (at least 25% eosinophils)with Charcot–Leyden crystals; foreign-body giant cells and sometimes larvalworms can also be seen in a lumbar punc-ture (about 10% of patients).
The meningitis must be differentiatedfrom that caused by a cerebral tumour or
144 Chapter 5
Fig. 67. Larvae of P. cantonensis migrating through the meninges of the central nervous system, excitingmeningeal infiltration, eosinophilic pleocytosis in the cerebrospinal fluid and sometimes subarachnoidalhaemorrhage (magnification � 50).

haemorrhage or from the presence of otherhelminths, such as T. solium cysticerci andGnathostoma, or possibly Paragonimus,Schistosoma eggs, Echinococcus, Trichinellaand Toxocara. Isolated instances ofeosinophilic meningitis have been reportedfrom parts of the world where P. cantonen-sis is not known to occur and may be due toother helminths, although this is clearly ahelminth that is being carried to new partsof the world inside the rat hosts (Cross,1987; Kliks and Palumbo, 1992).
Recently there has been a good deal ofwork on serodiagnostic tests. Two mono-clonal antibodies (AcJ1 and AcJ20) are ableto recognize the epitope on an antigen witha molecular mass of 204 kDa. A double-antibody sandwich ELISA showed goodspecificity with sera, but not with CNSfluid (Chye et al., 1997), and animmunoblot technique is promising.
Larvae have been recovered from theeye on three occasions (a recent case in SriLanka may have been of a new species).
TREATMENT
It is not certain that chemotherapy is advis-able, as live worms cause far less tissuereaction than dead ones and the infectionis usually self-limiting.
EPIDEMIOLOGY
Human infection usually occurs from eatingthe giant African land snail (Achatinafulica), which is popular among people ofChinese descent and has been introduced bythem to Hawaii and other Pacific islands.Another snail, Pila, is similarly eaten inThailand, as are freshwater prawns inTahiti. In addition, small slugs (e.g.Veronicella), snails and planarians may beingested inadvertently in salads. It is poss-ible that larvae escape from the invertebratesin faeces or slime and are ingested on veg-etables or fruit. At least six species of terres-trial snails, nine species of aquatic snailsand five species of slugs, freshwater prawns,frogs and toads have been found naturallyinfected, and many other invertebrates andprobably fish can serve as paratenic hosts.In some Asian countries some of these hostsare ingested for medicinal purposes.
The natural hosts are many species ofrodents, for human infections principallyRattus norvegicus and R. rattus (now com-monly infected in ports such asAlexandria, New Orleans and PortHarcourt, Nigeria, although human infec-tion has not yet been proved). The bandi-coot rat is another natural host.
Parastrongylus mackerrasae possiblycauses eosinophilic meningitis in Australiaand P. malaysiensis in South-East Asia(Prociv et al., 2000). Another species, P.vasorum, which occurs in dogs and foxesin Europe and South America, does nothave a neurotropic cycle.
Parastrongylus costaricensis(Morera and Céspedes, 1971) Ubelaker,
1986
SYNONYM AND DISEASE
Angiostrongylus costaricensis Morera andCéspedes, 1971; Morerastrongylus costa-ricensis Chabaud, 1972. Abdominal para-strongyliasis or parastrongylosis (orabdominal angiostrongyliasis).
GEOGRAPHICAL DISTRIBUTION
This parasite was discovered by Moreraand Céspedes in the mesenteric arteries ofpatients in Costa Rica in 1971. In the fol-lowing 30 years it has been found inhumans in Argentina, Brazil, Colombia,Costa Rica, Dominican Republic, Ecuador,El Salvador, Guadeloupe, Guatemala,Honduras, Japan, Martinique, Nicaragua,Panama, Peru, the USA and Venezuela,with a possible case in Congo Republic(Zaire).
LOCATION IN HOST
Adult worms that lay eggs but do not pro-duce larvae are found in the ileocaecalbranches of the anterior mesenteric arteryor occasionally in the liver or testicles. Fulldevelopment occurs in the natural host, thecotton rat, Sigmodon hispidus.
MORPHOLOGY
Adult males measure 17.4–22.2 mm �0.28–0.31 mm. The caudal bursa is well
The Nematodes 145

developed and symmetrical with six papil-lae and equal spicules. Females measure28.2–42.0 mm � 0.32–0.35 mm, with thevulval opening about 0.26 mm from the tipof the tail. Both sexes lack a buccal cap-sule.
LIFE CYCLE
Twelve species of rodents can act as finalhosts, as well as marmosets, dogs and thecoatimundi. First-stage larvae (L1s) in thefaeces of these hosts enter slugs as interme-diate hosts, where they develop into theinfective third stage (L3) and are oftenexcreted into the mucus left on vegetation.In Central America the veronicellid slug,Vaginulus plebeius, which is common up toan altitude of 2000 m, is commonlyinfected (50% overall in Costa Rica) some-times with large numbers of larvae (Morera,1994). When these slugs are ingested bycotton rats or other natural hosts, the L3spenetrate the intestinal wall within 4 h andmigrate to the lymphatics of the intestinalwall and mesentery. Following a furthertwo moults, the young adults (females mea-suring 28–44 mm and males 20 mm inlength) migrate to the mesenteric arteriesafter 10 days. They mature and release eggsby 18 days. The eggs are deposited in theintestinal wall, where they hatch, and theL1s are excreted in the faeces by 24 dayspostinfection. Humans become infected byingesting infected slugs, either intention-ally, in the case of young children, or acci-dentally, on salad vegetables. The L1s inmucus can also be ingested from food orfrom the hands of children after playingwith slugs.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Infection is found predominantly in children,but all ages can be susceptible. If worms arepresent in the tissues of the caecal region,there is usually pain in the right flank andiliac fossa. There may be fever, often accom-panied by anorexia and vomiting and consti-pation or diarrhoea in children. In manypatients a tumour-like mass can be palpatedin the lower right quadrant (although thisprocedure may be painful), and this may bemistaken for a malignant tumour. There is
usually some degree of leucocytosis, with20–50% eosinophilia. In children, symptomsmay mimic appendicitis, Crohn’s disease orMeckel’s diverticulum. Some cases havebright red blood in the faeces.
The adult worms and eggs in the arteriesprovoke damage to the endothelium, caus-ing thrombosis and necrosis of tissues, andthe eggs cause inflammatory reactions in thesmall vessels. The whole wall of the intes-tine can become thickened and hardened,with yellowish granulatomous areas ofinflammation (Fig. 68). In lighter infectionsonly the appendix is damaged but in heavyinfections the terminal ileum, caecum andascending colon may have to be excised.
If the liver is infected, there is likely to bepain in the upper right quadrant, with somedegree of hepatomegaly. Palpation is painfuland there are enzyme abnormalities and ahigh eosinophilia; the syndrome resemblesvisceral larva migrans (p. 157). The presenceof adult worms in the spermatic cords ofmales can result in acute pain from obstruc-tion by adult worms, leading to haemor-rhagic necrosis of the testicular parenchyma.
Even without treatment, recovery usuallytakes place in a few weeks to many months.
DIAGNOSIS
Symptoms in children of an appendicitis-like illness with a high eosinophilia aresuggestive. Clinical diagnosis can be greatlyaided by X-ray. Contrast medium shows fill-ing defects in the terminal ileum, caecumand ascending colon, with irritability andthickening of the wall. There is also a sud-den contraction of the caecum observed byfluoroscopy after a barium enema, typical ofchronic granulomatous inflammatory reac-tions and found also in tuberculosis andamoebic abscesses. Newer imaging tech-niques would probably be even more use-ful. Parasitological diagnosis is onlypossible following biopsy or resective surgerysince L1s are not produced in the faeces.
Serological diagnosis is thus veryimportant. There is a latex agglutinationtest and an ELISA, both of which are sensi-tive and specific, and immunoelectrophore-sis or Ouchterlony immunodiffusion canalso be employed.
146 Chapter 5

TREATMENT
Surgery is the most common form of treat-ment, but with more experience manycases have been found to resolve in timewithout it, and conservative palliativetreatment can be employed unless there isan acute flare-up.
Chemotherapy with thiabendazole, alben-dazole and diethylcarbamazine has beentried and may have some effect but there isevidence from experimental infections inrats that it causes the parasites to wandererratically and cause further lesions.
EPIDEMIOLOGY
This infection is widespread in rodents inthe Americas and the numbers of humancases is rising in many countries, probablybecause it is being increasingly recognizedas a public health problem rather than arare and curious infection. Most cases havebeen reported from Costa Rica, more than650 in 1993 in a population of 3 million(Morera, 1994). Almost all of these are inchildren, and boys are twice as likely to beinfected as girls; both are more likely to beinfected in the wet season. In Costa Rica,43% of cotton rats were found to be infected.
Metastrongylus elongatus Railliet andHenry, 1911 (Family Metastrongylidae), aparasite of pigs with earthworms as inter-mediate hosts, has been recovered oncefrom the human respiratory tract.
Order Ascaridida
Family Ascarididae
Ascaris lumbricoides Linnaeus, 1758
DISEASE AND POPULAR NAMES
Ascariasis or ascariosis; roundworm infec-tion. One of the geohelminth infections.
LOCAL NAMES
Dedan (Arabic), Cacing (Bahasa Malay),Than kaung (Burmese), San’chik (Canton-ese), Nsunsuma (Fanti), Agan afo (Ibo),Kaichu (Japanese), Njowni (Luo), Watapanuwa (Singalese), Gooryan (Somali),Lumbris (Spanish), Safura njoka (Swahili),Nakupoochi (Tamil), Pa-yard sai-doen(Thai), Aràn inú (Yoruba).
The Nematodes 147
Fig. 68. Cross-sections of five adult P. costaricensis in a mesenteric vessel. The inflammatory exudateand eosinophils surrounding them are undergoing necrosis (perhaps following the death of the worms), suggesting both immediate and delayed hypersensitivity responses.

GEOGRAPHICAL DISTRIBUTIONCosmopolitan. Ascaris infection is com-mon in both temperate and tropical coun-tries where there are both adequatemoisture and low standards of hygiene andsanitation. It is one of the most commonhuman helminths with an estimated 1400million infected persons in the world (23%of the world population), with about60,000–100,000 deaths annually.
LOCATION IN HOSTAdults are normally found in the lumen ofthe small intestine but do not attach to themucosa.
MORPHOLOGYThese are large, stout nematodes, taperingat both ends. The females (200–400 mm �3–6 mm) are slightly larger than the males(150–300 mm � 2–4 mm) but may showgreat variation in size, depending on ageand worm load. The mouth opens termi-nally and has three lips, each of which hasa pair of sensory papillae on its lateral mar-gin, and rows of small denticles or teeth.The principal structural protein of the cuti-cle is collagen, stabilized by disulphidebonds.
In the female the paired, tortuous, tubu-lar ovaries, oviducts, seminal receptaclesand uteri lead into a vagina, which opens
by a small vulva on the ventral surfaceabout a third of the body length from theanterior end. The average daily output ofeggs is about 200,000 per female and eachmay lay a total of 25 million during herlifetime. The posterior end of the male iscurved ventrally and there are two almostequal copulatory spicules (measuring2.0–3.5 mm) and numerous preanal andpostanal papillae. There is no gubernacu-lum.
The eggs are broadly ovoid and measure45–70 µm � 35–50 µm. The shell is thickand transparent and usually has a coarselymammilated outer albuminous coat,stained light brown with bile pigments(Fig. 69). The ova are unsegmented whenpassed in the faeces. Occasionally eggs arepassed without the outer coat and a pro-portion of eggs (about 15%) are infertile;these are longer and narrower (90 µm � 40µm), with a thin shell, more irregular outercovering and disorganized contents (Fig.69). They occur either because the rapidproduction of eggs allows some to passthrough unfertilized or because onlyfemale worms are present.
LIFE CYCLE (Fig. 70)The unsegmented, fertilized, ova, passed inthe faeces, take 10–15 days to develop tothe infective stage at 20–30°C in moist soil
148 Chapter 5
Fig. 69. Egg of Ascaris. Left: Fertile unembryonated egg as seen in faeces. Centre: Infertile egg fromfaeces. Right: Second-stage infective larva in egg after at least 10–15 days’ development in soil (originalmagnification � 350).

or water. The first-stage larva inside the eggmoults after about a week and it is the eggcontaining the second-stage larva that isinfective (Fig. 69). In moist loose soil withmoderate shade, infective eggs may survivefor up to 7 years (Plate 2).
When the infective eggs are ingested,the larvae hatch in the duodenum. Theeggshell is dissolved when there is a tem-perature of 37°C, high CO2 concentration,low oxidation-reduction potential and a pHof around 7.0. The second-stage larvaemeasure 0.2–0.3 mm � 0.014 mm and havea rhabditiform oesophagus extending backabout a quarter of the length of the body.They penetrate the mucosa and are carriedin the lymphatics or veins through the liverand heart to reach the lungs in 3 days.With A. suum in pigs, at least, the larvaepenetrate the colon and caecum, not theileum, and reach the liver in 6 h causinglesions (Murrell et al., 1997). The larvaemoult twice in the lungs (at 5–6 days and10 days) and then penetrate the alveolae,ascend the trachea and pass down the
oesophagus to reach the ileum. Theydevelop into adult males and females inabout 65 days and live for 1–2 years.
CLINICAL MANIFESTATIONS
Although the majority of infections areapparently symptomless (about 85%), thepresence of even a few worms can bepotentially dangerous and there are stillover 200 million cases showing morbidity.
The first clinical signs and symptoms area pneumonitis with cough, dyspnoea, sub-sternal pain, fever and sometimes a blood-stained sputum (which may contain larvae).These symptoms begin 5 to 6 days after infec-tion and usually last 10–12 days. This isknown as Loeffler’s syndrome and the X-rayfindings often show dense pulmonary infil-trations, resembling acute tuberculosis. Thereis moderate eosinophilia (which increases toa maximum of 40% 3 weeks after infection).
The severity of symptoms depends bothon the number of eggs ingested and on theprevious infection history; hypersensitiveindividuals may have severe symptoms,
The Nematodes 149
swallowed and hatch
Duodenum, blood orlymphatics, lungs(bronchi, trachea),oesophagus, ileum
2ND-STAGE LARVAin egg. Can survivein soil for up to 7 years
ADULTSin ileum
HUMAN
Unsegmented EGGS(60�m � 40�m) in
faeces reach soil
SOIL
1ST STAGE LARVAdevelops in egg
10–15 daysOne moult in egg
Fig. 70. Life cycle of Ascaris lumbricoides.

with perhaps urticaria and asthma from thepresence of only a few larvae. The relation-ship between pneumonitis and ascariasiswas well shown by a study in SaudiArabia, where there is a short transmissionseason in spring when infection can occur.In young adults (and probably children)Loeffler’s syndrome occurs at this time ofyear, with eggs appearing in the faeces 2–3months later. However, in Colombia, wherethere is constant transmission, this syn-drome does not occur.
The second phase of clinical disease isdue to the presence of the adult worms inthe intestinal lumen (Fig. 71). The usualsymptoms of heavy infections are digestivedisorders, nausea, vomiting and colic and,in children, restless sleep and tooth grind-ing.
Intestinal obstruction is a serious com-plication of Ascaris infection, which usu-ally occurs in children, often as young as 2(Plate 16). Mortality in such cases admittedto hospital is about 9%. Because of thedanger of causing obstruction, it is advis-able in an endemic area to examine thefaeces, followed by appropriate treatment ifnecessary, before carrying out an intestinaloperation or before giving chemotherapyfor other parasites, which may irritateAscaris. The symptoms of intestinalobstruction are recurrent abdominal pain,loss of appetite, nausea and vomiting.
Adult worms causing obstruction can oftenbe seen outlined as a shadow on X-raysafter a barium meal. In many areas this isthe commonest cause of intestinal obstruc-tion in children. Occasionally worms maycause peritonitis or may wander in theappendix and possibly give rise to appen-dicitis (but see p. 163).
Adult Ascaris appear to be particularlyprone to a ‘wanderlust’, perhaps an attemptto repeat the migrations of their youthwhen the larvae were only 0.3 mm long(Plate 17). They may migrate up the bile-duct or even up the pancreatic ducts. Inchildren this causes severe colicky pain inalmost all patients, with nausea and vomit-ing. After a few days fever develops,accompanied by a slight jaundice.Complications include ascending cholangi-tis (in about 15% of cases), obstructivejaundice (1%), acute pancreatitis (4%), andoccasionally multiple liver abscesses. Suchworms can be removed endoscopically. InChina ascariasis has been estimated toaccount for about 13% of all cases of bil-iary obstruction (Zhou Xianmin et al.,1999).
Sometimes adults may develop inectopic sites such as the abdominal cavityand infrequently worms pass out throughvarious orifices – mouth, nose, lachrymalduct, urethra, vagina – much to the con-sternation of the infected person.
150 Chapter 5
Fig. 71. Portion of ileum removed at autopsy containing many adult Ascaris.

Surgical complications are more com-mon in areas where patients have greaterworm loads, which in turn are foundwhere the intensity of transmission andprevalence are high.
The amount of host food materialremoved by Ascaris is not negligible andthis, together with concomitant anorexiaand malabsorption, probably potentiateskwashiorkor in areas where protein intakeis only just adequate. Parasitism is often sowidespread in many developing countriesthat the effect on the nutrition of alreadymalnourished children should not beunderestimated. It has been estimated thata child who has 26 worms may lose one-tenth of its total daily intake of protein.There is also some evidence that ascariasisin children can contribute to vitamin Aand C deficiency. The most importanteffect of disease in children aged 2–10 is itseffect on height and weight, developmentand probably cognitive ability (Cromptonet al., 1989; Thien Hlang, 1993; Crompton,2000).
PATHOGENESIS
The larvae cause lesions in the liver, spleenand lymph nodes. Sometimes necrotic fociare also found, encapsulated larvae causinga granulomatous reaction. In the lungs theycause petechiae and in severe reactionsthere is also ulceration of the bronchialmucosa and abundant mucus. Charcot–Leyden crystals may be present from thebreakdown of eosinophils.
High IgE production may function toprotect both the host and the parasite.Ascaris induces a Th2 cell-mediatedresponse, producing interleukin 4 (IL-4)and resulting in production of IgE. It ispossible that the parasite also produces anIL-4-type molecule that stimulates the hostto produce non-specific superfluous IgE,thus swamping the immune response(Prichard, 1993). Attempts to demonstratethat immune responses can explain theclear predisposition to infection have notbeen successful (see Crompton, 2000). Theimmunology of intestinal nematodes is dis-cussed on p. 120.
DIAGNOSIS
Clinical. The pneumonitis caused by themigrating larvae is often not diagnosed, butthe accompanying high eosinophilia is sug-gestive. The adult worms can sometimes beseen as clear spaces on X-rays after a bar-ium meal (the gut of the worms may showas a thin central line of contrast medium)or on a computer-assisted tomography (CT)scan. Ultrasound is particularly useful forworms in the biliary or pancreatic ducts,showing up as long, narrow, movingimages maybe with two inner parallellines. Endoscopy is also useful.
Parasitological. Eggs are usually plentiful inthe faeces, although infertile eggs may notbe recognized. Direct faecal smears, usingabout 2 mg of faeces, are usually adequate,while the Kato–Katz thick-smear technique(p. 257) works well. Concentration meth-ods using sedimentation or brine flotationmay be used but are not really necessary.Unlike most other nematodes, the numberof eggs in the faeces often gives a goodindication of the number of adults present(although a few parasites produce propor-tionally more eggs and very many parasitestoo few). However, since there are markedvariations in fecundity in different coun-tries, the WHO threshold of 50,000 eggs g�1
faeces indicating a heavy infection needingregular re-treatment may not be universallyapplicable (Hall and Holland, 2000).
Immunodiagnosis. All immunodiagnostictests are very non-specific and none areused in practice. A 25 kDa antigen from thebody fluids of larvae and adults of A. lum-bricoides and the pig ascarid A. suum(ABA-1) is highly allergenic against mastcells in vitro and could possibly provide thebasis for a test, at least at the communitylevel – preferably if it could utilize saliva. Itis possible to identify Ascaris metabolites inurine but this is not so far a practical test.
TREATMENT
Clinical. Intestinal obstruction in childrencan usually be treated conservatively withintravenous fluids, but acute infections
The Nematodes 151

may require surgical removal. Biliaryobstruction, which is more common inadults than in children, can sometimes betreated by using retrograde endoscopy.
Chemotherapy
● AlbendazoleThis is 100% effective at a single oralstandard dosage of 400 mg for all indi-viduals except pregnant women and theunder-5s and will have action against allthe geohelminths.
● Mebendazole This can similarly be used at a standardsingle oral dose of 400 mg for all geo-helminths although best against Ascaris(100%).
● Pyrantel pamoate (= embonate)This is given as a single oral dose at5–10 mg kg�1 body weight. It is noteffective against Strongyloides andTrichuris (0–20% cure rate) and onlypartially against hookworms (20–90%);possibly it may also encourage obstruc-tion by paralysed worms.
● LevamisoleWhen given at 150–250 mg (2.5–5 mgkg�1) it is completely effective (para-lysing Ascaris) but less so against otherintestinal nematodes.
EPIDEMIOLOGY
Ascariasis is prevalent in most tropicalcountries, with estimated infections of:China 523–539 million, Côte d’Ivoire 17%and Kenya 38%. Infection rates are oftenhigh in urban (particularly slum) areas:25% in Brazil (Rio de Janeiro), 47% inEgypt (Alexandria), 35% in India(Hyderabad), 64% in Malaysia (KualaLumpur), 55% in Mexico (Coatzacoalcos),68% in Nigeria (Lagos), 80% in thePhilippines (Manila) and 43% in SierraLeone (Freetown).
The adults are not usually very long-lived and there is often an annual cycle ofinfection. Evidence from people who moveto a non-endemic area shows that eggscease to be passed in under 18 months.Eggs may continue to be passed in the
faeces up to 7 days after the adults havebeen expelled. Males are expelled first, sothat in long-established infections withoutreinfection there is a preponderance offemales.
The use of human faeces as a fertilizer(‘night-soil’) is common in some parts ofthe world and can result in eggs getting onto salad vegetables. In China and otherareas raw faeces is now being stored. Over90% of ova are killed by storage for 15 daysat 29°C or 6 days at 33°C; but for completedestruction 40 days at 30°C is required(Muller, 1983).
Untreated sewage is sometimes dis-charged into rivers and lakes, which repre-sents a clear health hazard. Even morecommon is discharge into the sea and eggsare able to continue development in seawater, as is said to occur in coastal villagesof Samoa.
Properly designed modern sewage treat-ment plants will remove or destroy allAscaris eggs, the bacterial bed treatmentnot being so effective as the activatedsludge process or sand filtration. However,eggs need to be retained for 10–12 monthsin septic tanks and cesspits before they areall destroyed. Eggs will not embryonatebelow 18°C but can survive for manyweeks at much lower temperatures andcontinue development when the tempera-ture is raised. In experiments in Russiaeggs have survived in soil for 7 years, butin most natural habitats they will bedestroyed by sunlight, fungi or acarines in2–6 months and in temperate climates theysurvive for shorter periods in summer.
The soil type is important in transmis-sion; clay soils favour survival of eggs,since they retain water better. Completelack of aeration is not lethal, ova remainingviable for months without oxygen,although their development is interrupted(Mizgajska, 1993).
PREVENTION AND CONTROL
Where there is a marked rise in standardsof sanitation over a large area, Ascarisinfection falls quickly, usually within ayear. At the personal level, thorough wash-ing and preferably cooking of vegetables
152 Chapter 5

and supervision of children’s play areas areimportant. However, it is difficult to pre-vent infection and reinfection in youngchildren, particularly where pica (geopha-gia) is common (Geissler et al., 1998). Masschemotherapy campaigns for all geo-helminths are discussed on p. 133.
ZOONOTIC ASPECTS
The relationship of human A. lumbricoidesto the very common parasite of the pig stillremains unresolved. Slight morphologicaldifferences have been described, such asthe labial denticles being concave in theformer and straight-sided in the latter, butthese have been challenged by other workers(Gibson, 1983). Cross-infection canundoubtedly occur but in Guatemala, forinstance, there appears to be little geneflow between infections of humans and ofpigs, which is encouraging for control cam-paigns. In North America, however, a non-endemic region, molecular evidenceindicates that most human cases are of pigorigin (Anderson and Jaenike, 1997). Therealso appear to be genetic differencesbetween the two forms (Xingquan et al.,1999) and pig parasites are usuallyregarded as a separate species, A. suumGoeze, 1782. It is probable that humaninfections originally came when pigs werefirst domesticated in neolithic times, possi-bly in China (Peng Weidong et al., 1998;Crompton, 2000); infections of A. lumbri-coides in other primates appear to havebeen contracted from humans (cases of‘nosozootic’ infections).
Lagochilascaris minorLeiper, 1909
There have been occasional human casesrecorded from Bolivia, Brazil, Costa Rica,Ecuador, Mexico, Surinam, Trinidad andVenezuela.
In humans the first sign of infection is apustule, usually on the neck, which growsin size to give a large swelling. After theskin breaks, living adults, larvae and eggsare expelled at intervals in the pus. Eggsare continually developing into larvae andthen adults in the tissues at the base of
abscesses, so that repeated life cycles occur(de Aguilar Nasumento et al., 1993). In themastoid region, temporal bone can beinvaded, with numerous mastoid sinusesand perhaps involvement of the CNS. Inone woman worms emerged into the mouthand in a young girl a worm was recoveredfrom a tooth abscess. The zone of indura-tion grows deeper and sinus tracts develop,the openings oozing pus. Eventually achronic granulomatous inflammationensues, which can last for months. Somecases are fatal, including one with wormsin the lungs.
The natural definitive hosts areunknown but are probably carnivores oropossums. Cats have been found naturallyinfected in Argentina and Brazil and, incats experimentally infected with cystsfrom mice containing third-stage infectivelarvae, the larvae excysted in the stomachand penetrated the oesophagus, trachea,pharynx and cervical lymph nodes, devel-oping to adults in 9–20 days postinfection.However, the worms did not produce eggs,although they did in animals fed infectedmice. In mice infected orally with ova fromhumans, the first-stage larvae hatched inthe gut, penetrated the wall and migratedto the liver, lungs, skeletal muscles andsubcutaneous tissues, where they encystedas third-stage larvae. How humans becomeinfected is not known, although presum-ably it is either from ingestion of wild ani-mals containing cysts or from ingestion ofeggs in the environment.
Long-term treatment with levamisoleand thiabendazole has not been effective,but albendazole (200–400 mg each day for5 days) and perhaps mebendazole (200 mgeach day) have some (although perhaps atemporary) effect. Ivermectin (300 µg kg�1
daily for 4 weeks) has resulted in long-termcure in single cases in Ecuador and Brazil.
Baylisascaris procyonis (Stefanski and Zarnowski, 1951)
This is a common parasite of racoons in theUSA. In humans and small rodents, suchas squirrels, or porcupines, rabbits, birdsand dogs, it causes a form of visceral,
The Nematodes 153

neural or ocular larva migrans. There areabout a dozen species of Baylisascaris fromcarnivores and the full life cycle is knownonly for a species in badgers, whichrequires an intermediate host. It is likelythat racoons ingest small mammals andbirds with granulomas containing larvae inthe somatic and particularly CNS tissues(although eggs can also be infective); theworms then develop into adults in theintestinal lumen and eggs are evacuated inthe faeces, with second-stage larvae devel-oping inside them in soil. They are theninfective to the mammalian and bird inter-mediate hosts, where development to thethird stage takes about 2 weeks.
In a fatal case in a 10-year-old boy, thelarvae evoked a sessile polyploid mass inthe left ventricular myocardium, protrud-ing into the ventricular lumen (Boschettiand Kasznica, 1995). In other cases, withlarvae in the brain, a sometimes fataleosinophilic meningoencephalitis ensues(Plate 18). In an infant who recovered aftertreatment with a single dose of ivermectinand multiple doses of thiabendazole andprednisolone, there was permanent neuro-logical damage (Cunningham et al., 1994).Other cases in adults have resulted in lar-vae in one eye causing a diffuse neuro-retinitis. In most cases diagnosis is basedsolely on ELISA or immunoblot techniquesalthough L3 larvae have been found in usu-ally granulomatous biopsy specimens.Larvae measure 930–1300 µm � 50–65 µmand have well-developed lateral alae.Neurological cases present with extremelethargy, with 30–40% eosinophilia(Kazacos, 1983).
The adult worms resemble those ofAscaris in size but have cervical alae and anarea rugosa, while the eggs are pitted. Thelarvae inside eggs can survive for long peri-ods in soil, and infection is widespread inracoons throughout North America; similarspecies parasitize skunks, martens and bears.
Parascaris equorum (Goeze, 1782) Yorkeand Maplestone, 1926, is a widespread par-asite of horses and other equines that hascaused occasional cases of human viscerallarva migrans and pneumonitis from inges-
tion of eggs in the environment. A biologi-cal quirk, shared with A. suum, is thatnuclear DNA is greatly reduced in allsomatic cells but not in germ cells.
Family Anisakidae
Anisakis and other anisakids
DISEASE AND POPULAR NAMES
Anisakiasis or anisakidosis; herring-wormdisease, cod-worm disease (fromPseudoterranova).
GEOGRAPHICAL DISTRIBUTION
Anisakis simplex (Rudolphi, 1809), syn-onym A. marina, is a natural parasite ofmarine fish-eating mammals. Human casesoccur where fish are eaten raw and wasfirst recognized in The Netherlands in1960. Cases (almost 34,000 in total) havebeen reported from Japan (over 16,000),Korea (196 up to 1992), Canada, France,Germany, Italy, The Netherlands, Russia,Spain, the UK and the USA.
LOCATION IN HOST
Larvae are found in abscesses in the wall ofthe intestinal tract, usually the stomach inJapan but also the small intestine andcolon.
MORPHOLOGY AND LIFE CYCLE
L3s in humans measure about 29 mm �0.45 mm, with the typical ascarid three lipsand a dorsal boring tooth at the anteriorend. The tail is short and rounded, with asmall projection at the end. Occasionallylarvae in humans develop to the fourthstage but never to adults.
Thin-shelled eggs (50 µm � 40 µm) aredischarged in the faeces by the adults,which inhabit the stomach of marine mam-mals. Larvae develop inside the egg andmoult once to give the L2. The sheathedlarvae hatch and can live in sea water formany weeks. If ingested by planktoniceuphausid crustaceans (krill), the larvaepenetrate the gut wall and develop to theL3 stage. When krill is ingested by many(164) species of fish, such as herring, mack-
154 Chapter 5

erel, cod and salmon, or by squid, the lar-vae encyst in the muscles or viscera. Theseact as paratenic hosts, since the larvae donot develop further, but if they are eaten byany of many species of marine mammals(26 cetacean and 12 species of pinniped)the contained larvae develop into adults inthe stomach.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
If the parasite is in the stomach, the mostcommon symptoms are nausea, vomitingand epigastric pain, simulating a pepticulcer or a tumour, beginning 1–12 h afteringestion of infected fish (Plate 19).Sometimes there may be symptoms ofacute chest pain. The parasites can elicitsevere IgE-mediated hypersensitivity,which might require emergency treatment.There is sometimes urticaria, angioedemaand even anaphylaxis. If abscesses are pre-sent in the intestine or colon, there is likelyto be lower abdominal pain, but sometimesit can resemble acute appendicitis (Plate20). This develops after about 2 days andlasts for a further few days before ileus orperhaps ascites results. Gastric cancer ismuch more common in Japan than in anyother country and the presence of Anisakislarvae has been suggested as a cofactor(Petithory et al., 1990). Occasionallyworms may penetrate through to theabdominal cavity and in one patient a larvawas discovered in the liver.
In the early stages there is an abscessinfiltrated by neutrophils and later aneosinophilic granulomatous reaction. Theaffected portion of the intestinal tractbecomes oedematous (up to 1.5 cm thick),with ulcerations and petechial haemor-rhages in the mucosa, each containing alarval worm. Dead worms become sur-rounded by foreign-body giant cells.Submucosal lesions of the ileum or colonalso have larvae in them, surrounded by anarea of necrosis.
DIAGNOSIS AND TREATMENT
Clinical and parasitological. Endoscopy canbe very useful for viewing the stomachabscess, which is oedematous, and there
may be severe erosive gastritis, hyperaemiaand bleeding. There are usually only one tonine worms, but in one patient 56 wormswere removed (Kagei and Isogaki, 1992).Contrast X-rays can reveal filling defectsand oedematous, obstructed, areas in thestomach wall (Ishikura et al., 1992).
Immunodiagnosis. Various monoclonal anti-bodies have been discovered that recognizespecific epitopes from A. simplex larvae.One (An2) recognizes epitopes from intesti-nal cells and ES products and has beenused in a micro-ELISA. Another IgG1κantibody (UA3) of 139 kDa is more specific,but both only detect the presence of theworms about 4 weeks after infection. Animmunoblot technique with larval whole-body antigen to detect IgE is positive after afew days (Garcia et al., 1997). An antigenic�-galactosidase helminth-derived recombi-nant fusion protein (FP) obtained byrecombinant DNA techniques and used inan ELISA is stated to be completely spe-cific, in contrast to soluble antigens(Sugane et al., 1992).
The most effective method of treatmentfor gastric worms is by endoscopy andremoval of the larval worms using biopsyforceps; symptoms clear up within hours ofremoval.
EPIDEMIOLOGY AND PREVENTION
With the ability to easily remove larvalworms, it is becoming increasingly evidentthat other anisakids may be involved, aswell as Anisakis simplex. There is A. phy-setis Baylis, 1923 (previously known asAnisakis type II larvae), Pseudoterranovadecipiens (Krabbe, 1878) in Japan andNorth America, Contracaecum osculatum(Rudolphi, 1802) and Hysterothylacium(syn. C.) aduncum (Rudolphi, 1802). Larvaeof all are found in marine fish; C. oscula-tum caused an infection in Spain from eat-ing sardines and H. aduncum, with adultsin piscivorous fish, not mammals, wasreported from Chile. All cause similarlesions in the gastric and possibly theintestinal wall. A fourth-stage larva of P.decipiens that had been removed measured29.7 mm � 0.94 mm (Sohn and Seol, 1994)
The Nematodes 155

and an adult worm (reported once only)33 mm � 1.0 mm. This is a natural parasiteof seals and requires two crustacean inter-mediate hosts (a copepod and an amphipod).
The various larvae can be distinguished.Those of Anisakis have no lateral alae andin section have characteristic Y-shaped lat-eral cords. A. simplex larvae measure onaverage 28.4 mm � 0.45 mm and have ashort tail with a terminal spine, whilethose of A. physetis measure 27.8 mm �0.61 mm and have a long tail without a ter-minal spine. Larvae of Pseudoterranovadecipiens measure 32.6 mm � 0.8 mm andhave a short tail with a spine and theproventricular–caecal junction is notstraight but diagonal.
A high proportion of food fish may beinfected with anisakid larvae, up to 90% inherring and cod. Fish should be gutted assoon as caught, since larvae are thought tomove into the tissues after death, and deep-freezing (�20°C) will kill the larvae. Raw(green) herrings are popular in TheNetherlands but infection appears to havevanished, since all fish sold must now bedeep frozen first. Thorough cooking willalso kill the larvae, but raw fish are eatenin some countries, particularly Japan,where ‘sashimi’ is popular. Also, it hasbeen reported recently that cases of severeallergic symptoms, such as urticaria, dysp-noea and even anaphylaxis, have occurredin patients after eating thoroughly cookedfish and it is probable that the individualshad become sensitized by previous infec-tion with live larvae.
Larva Migrans
This is a term used to describe humaninfection with helminth larvae that do notgrow to maturity but wander in the body,sometimes with serious results. The condi-tion is caused by natural parasites of ani-mals, with humans acting as abnormalparatenic hosts, in which the parasites donot develop but are not destroyed, and mayalso result from invasion by larvae belong-ing to zoophilic strains of species normallyfound in humans.
Infection can be divided into two cate-gories:
1. Visceral larval migrans results wheningestion of eggs or larvae leads to the pres-ence of wandering larvae in the deeporgans. Potentially, this group constitutesthe most serious danger to humans.Species that may be involved include thelarvae of Toxocara canis, T. cati,Lagochilascaris major, Parascaris equorum,Gnathostoma spinigerum and otherspirurids, Parastrongylus cantonensis, P.costaricensis, Anatrichosoma cutaneum,Alaria spp. and Spirometra spp. This defi-nition does not strictly include Ascarissuum and Calodium hepaticum, whichreach maturity in the human body,Baylisascaris procyonis, which develops inhumans as in its usual intermediate hosts,or Anisakis simplex, which develops nor-mally in humans until it dies in the tissues,although these species are often included.2. Cutaneous larva migrans is caused byskin-penetrating larvae that continue towander in the superficial layers of the body.Larvae that may be responsible includehookworms, Strongyloides, Gnathostomahispidum, fly larvae and spargana ofSpirometra spp. Some animal schistosomes,including zoophilic strains of Schistosomajaponicum, Heterobilharzia andOrientobilharzia and bird parasites, such asAustralobilharzia, Bilharziella, Gigantobil-harzia, Microbilharzia, Schistosomatiumand Trichobilharzia, cause ‘swimmers’ itch’at the site of penetration but without wan-dering in the superficial layers.
Visceral larva migrans
Only the dog and cat ascarids will be con-sidered in this section, the other possiblecausative agents being considered in theirappropriate sections.
Toxocara and Toxascaris
DISEASE AND POPULAR NAMEToxocariasis or toxocarosis; ‘arrowheadworms’. Most important cause of viscerallarva migrans (VLM).
156 Chapter 5

Toxocara canis (Werner, 1782) Johnston,1916, is a very common ascarid nematodeof dogs in many parts of the world. T. cati(synonym T. mystax) (Schrank, 1788)Brumpt, 1927, is found in cats and foxesand Toxascaris leonina (von Linstow,1902) Leiper, 1907, in dogs, cats and manywild carnivores. Each of these parasites ismore common in young animals during thefirst few months of life.
MORPHOLOGY
The adults are smaller than those ofAscaris and have characteristic cervicalalae or wings (Table 10).
LIFE CYCLE
The egg is unsegmented when passed, butin soil the ovum inside embryonates togive the infective second-stage larva in 9days at 26–30°C and 11–18 days at 20°Cwith a relative humidity above 85%.
The natural hosts are usually infectedbefore birth by the mother ingesting eggscontaining second-stage larvae. Thehatched larvae penetrate the gut wall andthen undergo a ‘somatic’ migration, reach-ing the lungs, liver and kidneys and, inpregnant females, penetrating through theplacenta to infect the fetus. Young puppiesor kittens may also be infected with larvaethrough the milk or by ingesting eggs,
when a ‘tracheal’ type of migration similarto that of Ascaris results. Adult females inthe ileum produce eggs in the faeces after4–5 weeks.
Eggs hatch as second-stage larvae inmice (or in the case of Toxocara cati orToxascaris leonina in invertebrates such ascockroaches and earthworms also) andthese act as paratenic hosts, the larvaeinvading many tissues and becomingencapsulated after 12 days. Further devel-opment occurs if these are eaten by thedefinitive hosts.
In humans the eggs are ingested and thesecond-stage larvae hatch out in the intes-tine and undergo a similar ‘somatic’ migra-tion but do not develop further. Theymeasure 350–530 µm and are considerablythinner than the larvae of Ascaris and thismay explain why they are not filtered outin the liver and lungs.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Disease occurs mainly in young children(typically 1–4 years old) who have hadclose contact with infected household petsor have been exposed to eggs in the envi-ronment and is due mainly to T. canis. Inover 80% of cases there is a markedeosinophilia (over 50% with 20,000eosinophils mm�3) and hepatomegaly. Inover 50% of cases, pulmonary symptoms,
The Nematodes 157
Table 10 . Differentiation of the dog and cat ascarids.
Toxocara canis Toxocara cati Toxascaris leonina
Length: Length: Length:Females 6–18 cm Females 4–12 cm Females 4–10 cmMales 4–10 cm Males 3–6 cm Males 3–7 cm
Head bent ventrally Head bent ventrally Head straight
Cervical alae elliptical Cervical alae broad and end Cervical alae ellipticalabruptly posteriorly
Oesophagus with a Oesophagus with a posterior Oesophagus with noposterior muscular bulb muscular bulb posterior bulb
Tail of male with digitiform Tail of male with digitiform Tail of male conical with no appendage and caudal alae appendage and caudal alae caudal alae
Eggshell pitted Eggshell pitted Eggshell smooth(75–85 µm) (65–70 µm) (75–85 µm)
Specific to dog Specific to cat Found in dog or cat

such as coughing and wheezing with dysp-noea and fever (Loeffler’s syndrome), last-ing up to 3 weeks, are found and there maybe acute bronchitis with pneumonia. ChestX-rays often show bilateral infiltrates.These are followed by gastrointestinal dis-turbances, with larvae present in the liverand maybe the brain as well as the lungs,causing eosinophilic granulomas.Hepatomegaly and eosinophilia can last formany months. Larvae in the brain cancause convulsions or hemiplegia (Plate 21).There is the possibility that Toxocara lar-vae are responsible for carrying viral(including poliomyelitis and Japanese Bencephalitis), bacterial and protozoalpathogens into the brain and other organs.Very rarely there can be myelitis.
Another form of the disease, which usu-ally occurs in older children or adultswhen larvae invade the eye but do notcause the typical visceral symptoms, canbe termed ocular larva migrans (OLM). Thelarvae cause unilateral chronic granuloma-tous endophthalmitis (in 60% of cases) orperipheral retinitis (in 10% of cases).These granulomata cause distortion or evendetachment of the retina, and blindness iscommon (Gillespie, 1993; Gillespie et al.,1993). There can also be papillitis, leadingto glaucoma. One danger is that the lesionwill be mistaken for a retinoblastoma andthe eye hurriedly removed. However, theeye lesions in toxocariasis are raised andalways unilateral (Fig. 72).
A few human infections with adult T.
158 Chapter 5
Fig. 72. Top: Ophthalmoscope view of the retina with raised area near the optic disc (haemorrhage onleft). Bottom: Section of granulomatous lesion of retina, with a larva in the centre of fibrinoid necrosis.

canis and T. cati have been recorded, possi-bly from ingestion of larvae in an inverte-brate intermediate host.
Histopathology. In experimental animals,petechial haemorrhages occur in the liverwhen the larvae leave the sinusoids andburrow through the tissues, leavingnecrotic trails behind. These are inflamma-tory at first but then become granuloma-tous, containing eosinophils, lymphocytes,fibroblasts, epitheloid cells and giant cells.The live larvae become encapsulated andcan sometimes live for months or evenyears but usually soon die. Similar lesionsoccur in the lung and there can also bemyocardial or CNS involvement. In experi-mentally infected mice, larvae of T. canisentered the brain in all cases and the eye in5%, while those of T. cati migrated to thebrain in 8% of cases but never to the eye.
The migrating larvae evoke bothdelayed-type and immediate-type hyper-sensitivity reactions. The levels of both IgGand IgM are raised greatly but much is non-specific. There is a carbohydrate (trisaccha-ride) cuticular-associated epitope that isimmunogenic (Kayes, 1997).
DIAGNOSIS
Clinical. Hepatomegaly with eosinophilia issuggestive, as is elevated gammaglobulinand isohaemoagglutinin titres. Ultra-sonography and magnetic resonance imag-ing (MRI) show hepatic granulomas ascharacteristic multiple hypoechoic nodularlesions.
Parasitological. Liver biopsies may showgranulomatous lesions but it is rare to finda portion of a larva in tissue sections. Thesecond-stage larva measures 350–450 µm �18–21 µm and, if one is seen, can be differ-entiated from an Ascaris larva in sectionsby its smaller diameter.
Immunodiagnosis. ELISA, using antigensfrom second-stage larvae or their ES prod-ucts, has about 90% specificity and 80%sensitivity. Low-molecular mass (26.9 to38.9 kDa) polypeptides from L2s recog-
nized by infected sera show a very highspecificity in an SDS-PAGE or enzymeimmunotransfer blot assay.
TREATMENT
Albendazole (400 mg or 10–15 mg kg�1
day�1 for 10–21 days) or mebendazole (100mg daily for 10–21 days) have been suc-cessfully used in recent trials. Diethyl-carbamazine can no longer berecommended, although thiabendazole isstill used. The majority of cases resolve ina few months even without treatment,although corticosteroids (e.g. 30 mg ofprednisone day�1 for 21 days) can be givento reduce inflammation and relieve symp-toms in severe cases.
EPIDEMIOLOGY, PREVENTION AND CONTROL
Infection is very common in dogs and catsin most parts of the world, e.g. Chile (19%T. canis, 65% T. cati), Slovakia (70% T.cati). Many children also react positivelyin serological tests, e.g. 33% in Corunna,Spain, 35% in Malaysia. It is clear that thegreat majority of children exposed do notsuffer clinical disease (Gillespie, 1988).
Puppies and kittens should always bedewormed as a routine. There are oftenmany infective eggs around houses fromthe faeces of household pets and measuresshould be taken to prevent indiscriminatedefecation and also to prevent young chil-dren from playing in and ingesting contam-inated soil (pica). In many developedcountries children’s play sandpits are nowprotected with a fence.
Toxocara pteropodis Baylis, 1926, is a par-asite of fruit bats on the Pacific islandscomprising Vanuatu. It has been suggestedas the cause of a disease outbreak known asPalm Island mystery disease with hepatitis-like symptoms, but this may have been dueto poisoning (Prociv, 1989).
Cutaneous larva migrans or ‘creepingeruption’
Of the various possible causes of this con-dition, Spirometra is considered on page
The Nematodes 159

70, Strongyloides on page 115, Gnatho-stoma on page 184 and fly larvae on page241, and only the animal hookworms areconsidered here.
Ancylostoma braziliense Gomez de Faira,1910, is a hookworm of dogs and cats inmany tropical regions of the Americas. Thebuccal capsule has a smaller aperture thanthat of A. duodenale and has a small innertooth and a large outer one. When theinfective filariform larvae penetrate theskin of humans, they follow a tortuouspath under the skin, progressing at 2–5 cmday�1. An erythematous inflammatoryreaction follows the tunnel made by thelarvae between the germinal layer of theepidermis and the dermis. The tunnel fillswith serum and the surrounding oedema-tous tissues contain many polymorphs andeosinophils. The larval movement resultsin intense itching and pruritus, and oftensecondary bacterial infection is caused byscratching; the larvae die after a few weeks,even without treatment, and the symptomssubside (Jelinek et al., 1994). Other speciesof animal hookworms, such as A. caninum,A. tubaeforme, Bunostomum phlebotomumor Uncinaria stenocephala may also beresponsible. The condition is similar to the‘ground itch’ that sometimes follows pene-tration by the larvae of human species ofhookworms and by Strongyloides sterco-ralis, but lasts longer. Local treatment withan ethyl chloride spray or topical applica-tion of thiabendazole is effective.
A. caninum (Ercolani, 1859) is a commonand important parasite of dogs and cats inmost tropical and semi-tropical parts of theworld and the larvae can be a cause of folli-culitis; development to adults in humans isdiscussed on p. 138, together with A. cey-lanicum, which is morphologically verysimilar to A. braziliense and is found indogs in Asia and South America, but doesnot appear to cause creeping eruption.
A. tubaeforme (Zeder, 1800) is a cos-mopolitan parasite of the cat, Uncinariastenocephala (Railliet, 1884), a similarparasite to Necator, is found in the dog, cat
and fox in Europe and North America andBunostomum phlebotomum (Railliet,1900) is a widely distributed parasite ofcattle.
Order Oxyurida
Family Oxyuridae
Enterobius vermicularis (Linnaeus, 1758) Leach, 1853
SYNONYMS
Ascaris vermicularis L., 1758; Oxyuris ver-micularis Lamark, 1816.
DISEASE AND POPULAR NAMES
Enterobiasis or enterobiosis, oxyuriasis;pinworm, seatworm or threadworm infec-tion.
Local names
Deedan dabousia (Arabic), Cacing kerawit(Bahasa Malay), Toke kaung (Burmese),Kira panuwa (Singalese), Njoka safura(Swahili), Keeri pulu (Tamil), Pa-yard sen-deri or Kem-mood (Thai), Nnoko-boa (Twi).
GEOGRAPHICAL DISTRIBUTION
Taking into account the difficulty of diag-nosis, enterobiasis is undoubtedly one ofthe most common human helminth infec-tions worldwide, certainly in temperate cli-mates, with an estimate of 1000 millioncases worldwide (Cook, 1994). In general,infection is less common in tropical andless developed countries, presumablybecause of the fewer clothes worn.
LOCATION IN HOST
The adults are located in the lumen of thecaecum and appendix, sometimes with theanterior end attached to the mucosa. Inheavy infections they may also be presentin the ascending colon and ileum.
MORPHOLOGY
Pinworms are small cylindrical nematodes,pointed at either end. They have three lipsaround the mouth but no buccal capsule.
160 Chapter 5

The oesophagus is muscular and has a pos-terior bulb characteristic of the orderOxyurida. The cuticle has cervical alaecontinued as lateral projections, whichenable Enterobius to be easily recognizedin sections of appendix, etc. (Figs 51 and73). The female measures 8–13 mm �0.3–0.5 mm and has a long pointed tail(about one-third of the total length). Thevulva opens midventrally almost a third ofthe way down the body. The paired uteriare usually filled with thousands of eggsand the whole body becomes distended.The male is much smaller, measuring 2–5mm � 0.2 mm. The posterior end is curvedventrally and has a single copulatoryspicule 100–140 µm long. There is nogubernaculum. The bursa is reduced andthere are various caudal papillae. Themales die after copulation and often appearin the faeces.
Another species, E. gregorii Hugot,1983, is usually present together with E.vermicularis and has been described fromFrance, England, Japan, New Guinea andSouth Korea (Hugot and Tourte-Schaefer,1985; Chittenden and Ashford, 1987).Males of this species differ by having ashorter, simple, spicule (60–80 µm longrather than 100–140 µm) and other slightmorphological differences. The eggs andfemales cannot be differentiated.
LIFE CYCLE
The gravid female migrates out of the anusin the evening or at night and lays numer-ous sticky eggs on the perianal and per-ineal skin. The female normally dies afteroviposition. Often she bursts, releasing allthe eggs, which number about 11,000(4000–17,000).
The eggs measure 50–60 (average 54) µm� 20–30 µm and have a thin semi-transpar-ent shell composed of an outer albuminouslayer, two chitinous layers and an innerlipoidal membrane. They are ovoid and flat-tened on one side (Fig. 123). Eggs are usuallylaid partially embryonated and containinfective rhabditiform larvae in 4–7 h at35°C (they require oxygen for completedevelopment and do not develop belowabout 22°C). However, if the female hasburst, eggs may be only at the four-celledstage and then require about 48 h at 25°C tobecome infective. The outer albuminouslayer of the egg is very sticky and in sensi-tized individuals can cause an intense itch-ing, as does the adult female. This oftenleads to scratching and eggs then lodgeunder the fingernails. They are also carriedin dust and, when ingested, the larvae (mea-suring 145 µm � 10 µm) hatch in the duode-num. From comparison with other membersof the order, they are presumably at the thirdstage. The larvae lodge in the crypts of the
The Nematodes 161
Fig. 73. Section of appendix with adult female Enterobius vermicularis in lumen.

ileum and moult twice. The young adultsmove down to the colon, attach to themucosa and become mature in 15–43 days.
Retroinfection can occur if the larvaehatch on the perianal skin and migrateback up the anus. The adult females livefor 5–13 weeks and males for 7 weeks, butinfection in humans is usually maintainedfor very much longer by repeated reinfec-tion. Retroinfection is more common inadults than in children and results inrecurrent infections with a few wormsevery 40–50 days.
Biologically, this group of nematodes isinteresting since it appears to be the onlyendoparasitic haplodiploid taxon, i.e.females are diploid while males are hap-loid and derived from unfertilized eggs(Adamson, 1989).
CLINICAL MANIFESTATIONS
The majority of cases are symptomless,although children often suffer from loss ofappetite and irritability; it is usuallyregarded as primarily an infection of chil-dren.
The most common symptoms are due tothe pruritus ani, which can be very severeand lead to disturbed sleep. In females thevulva can also be involved from vaginalmigration of adult worms and in girls is thecommonest cause of pruritus vulvae; theremay also be acute urinary tract infection.Scratching may lead to secondary infectionof the anal region.
Mild catarrhal inflammation, with nauseaand diarrhoea, sometimes occurs and symp-toms of appendicitis have been reported.Low eosinophilia of 4–12% may be found,but eosinophilia is not so pronounced as inmany other intestinal nematodes, perhapsbecause there is no tissue phase.
If female worms reach the peritonealcavity from intestinal perforations or fol-lowing migration into the female reproduc-tive tract, worms can die and becomesurrounded by granulomas (Gilman et al.,1991).
PATHOGENESIS
Little internal pathology can be directlyattributed to the presence of Enterobius,
apart from a mild inflammation. Ulcerativeand haemorrhagic lesions of the small andlarge intestine have been reported but maybe coincidental.
The relationship of pinworm infectionto acute appendicitis has not been clearlydemonstrated, although Enterobius hasbeen found in many patients with symp-toms of appendicitis and where no othercause has been found. Sometimeseosinophilic granulomas are found (AlRabia et al., 1996), but recent large-scaleexaminations have not supported an asso-ciation between Enterobius, or any otherhelminth, and appendicitis. In one study inVenezuela, 3500 removed appendices wereexamined and Enterobius found in 11%,Trichuris in 33% and Ascaris in 25%, butthere were just as many in normal organsas in diseased ones (Dorfman et al., 1995).Indeed, in another study in Denmark, look-ing at 2267 appendices, 4.1% had E. ver-micularis but in 72% they were in normalorgans and only 7.4% were in inflamedorgans. Possibly E. vermicularis is thecause of appendicitis-like symptoms(pseudo-appendicitis) or, on the otherhand, worms may leave an appendix that isinflamed (Addiss and Juranek, 1991). Afew cases of perianal mass have beenreported with recurrent cellulitis.Occasionally worms lodge in ectopic sites,such as the Fallopian tubes or the ovaryand in peritoneal granulomatous nodules.In about 20% of infected young girls gravidfemale worms migrate into the vagina anduterus, occasionally evoking an endometri-tis (Sun et al., 1991). Eggs are sometimesfound in cervical Papanicolaou smears.
DIAGNOSIS
Adult worms are often observed on the sur-face of stools. Sometimes they can also beseen around the anus or in the vulva in theevening.
Ova are rarely found in the faeces andare better looked for in the perianal andperineal regions, using a swab technique.The National Institutes of Health cello-phane swab has been in use since 1937, butnowadays cellophane is replaced by adhe-sive tape (Sellotape or Scotch tape). The
162 Chapter 5

tape is usually stuck by one end to a micro-scope slide and turned back over a tonguedepressor or finger to make a swab(Graham swab). The sticky tape is rubbedover the anal region and stuck to the slide.A very convenient single-use flexible plas-tic slide is manufactured in Japan. InRussia an adhesive-covered glass ocularspatula is stated to be more sensitive andcheaper. Ideally the swab should be takenfirst thing in the morning and repeated for7 days before a negative diagnosis can bemade (and even then only at about the 90%level). If it is necessary to confirm whetherthe eggs are viable, the slide can be gentlyheated, when the enclosed larvae shouldbecome active. Where there is a lot ofextraneous material, a drop of toluene maybe placed on the slide first to increasetransparency.
TREATMENT
Enterobius is one of the easiest of helminthinfections to treat and a wide range ofanthelminthics are effective. The benzimi-dazole compounds inhibit microtubulefunction and glycogen depletion and arethe most effective. However, whicheverdrug is used, it is advisable to treat theentire family or group on several occasions;otherwise reinfection is almost inevitable.Where the cost is important, treatment isprobably not worthwhile for symptomlesscases unless used primarily for otherhelminths.
Albendazole. The standard treatment of asingle dose of 400 mg, or a divided dose of200 mg twice a day, usually gives 100%cure rates in children over 2 and adults(but should not be given to pregnant women).
Mebendazole. At a single dose of 100 mgwith the same proviso, this is equally effec-tive.
Pyrantel pamoate. Given in a single dose of10 mg kg�1 body weight or with a repeatdose 1 week later, this is also effective.
Ivermectin. At a single dosage of 50–200 µgkg�1 body weight or two doses of 100–200
µg kg�1, has given about 85% activity inclinical trials.
Levamisole (2.5 mg kg�1 body weight in asingle dose) and piperazine adipate (9 mgkg�1 daily for 7 days) have been used formany years and can still be employed.
EPIDEMIOLOGY
Enterobiasis is particularly a group infec-tion, being most common in large familiesand in institutions, such as boarding-schools, hospitals, mental homes andorphanages. Transmission may be termedcontaminative, as the eggs are immediatelyinfective, and normally takes placeindoors; this is in contrast to the soil-transmitted geohelminths, in which theeggs or larvae continue development in thesoil. Eggs may be found in large numbersinside houses, particularly in soiled bed-clothes, in night clothes and in dust in bed-rooms and lavatories (up to 50,000 m�2).Once embryonated (at above 22°C), theeggs can survive for 6–8 weeks under cool,moist, conditions but are killed in a fewdays by a dry atmosphere. Direct contami-nation from anus to fingers to mouth is alsoa common mode of infection.
Recent estimates of infection rates invarious countries using sticky tape are:preschool institutes in Azerbaijan 36%;Brisbane, Australia 43% (adults 23%);mental patients in Tasmania, Australia19%; Bulgaria 6.5%; Chile 16–22%; chil-dren’s homes in Santiago in Chile 87%;Denmark 12.5%; the Dominican Republic12.5%; Shoubra, Egypt 45%; children inhospital in England 55%; Germany 34%;Greece 22%; adults in Bengal, India 12%;Tamil Nadu, India 14%; Basrah, Iraq 18%;children in Lithuania 21%; Macao 23%;Penang, Malaysia 58%; Quetta, Pakistan4%; children in Peru 43%; Poland 30%;South Korea 20�77%; orphanages in SouthKorea 74–84%; Slovenia 40%; children inMoscow, Russia 30%; Tainen City, Taiwan38%; Turkey 40–97%; primary schools inCalifornia, USA 12–22%; Venezuela12.5%. Numerous other studies have beencarried out using faecal examination onlyand these have given much lower infectionrates.
The Nematodes 163

As with other intestinal nematode infec-tions, there appears to be a predispositionto infection, with a small percentage ofpatients having very heavy infections andbeing responsible for most of the eggsreaching the environment (Haswell-Elkinset al., 1987a, b).
Enterobius eggs have also been found in10,000-year-old coprolites from caves inTennessee and Utah, USA, and Chile, so itis a well-established human parasite.
PREVENTION AND CONTROL
Hygienic measures are of primary impor-tance and include frequent washing ofhands and changing and washing of sheetsand night garments every morning duringan outbreak. The area around the anusshould be washed every morning. Dustshould not be allowed to collect and thereshould be good ventilation. The eggs arevery light and easily become airborne.
ZOONOTIC ASPECTS
None. Almost exclusively a parasite ofhumans, although monkeys (chimpanzee,baboon, gibbon and marmoset) have beenfound infected in zoos. Other species arefound in apes and monkeys and relatedgenera occur in rabbits, rats, mice andhorses.
Syphacia obvelata (Rudolphi, 1802)Seurat, 1916, is a common and cosmopoli-tan parasite of rats and mice, both in thewild and in the laboratory. Three caseshave been reported from children in thePhilippines.
Order Enoplida
Family Trichuridae
Trichuris trichiura (Linnaeus, 1771) Stiles, 1901
SYNONYMS
Trichocephalos trichiura (Goeze, 1782)Trichocephalus trichiurus (Shrank, 1788);T. dispar (Rudolphi, 1802); T. hominis(Shrank, 1788); T. intestinalis; (Hooper,1799) Ascaris trichiura (Linnaeus, 1771).
The generic name Trichocephalus isprobably more correct as the originaldescription by Roederer and Wagler in1761 naming the organism Trichuris didnot obey the rules of binary nomenclatureand probably represented the speciesname. The fact that the name Trichuris(‘hair tail’) is not so apt as the later correc-tion Trichocephalus (‘hair head’) is irrele-vant to the argument, however. The nameTrichuris is preferred in this edition partlybecause it was chosen by the Committee onNomenclature of the American Society ofParasitologists in 1941 (although there hasbeen no ruling from the InternationalCommission on Zoological Nomenclature)but mainly because it is becoming morefirmly established each year, except per-haps in eastern Europe.
DISEASE AND POPULAR NAME
Trichuriasis or trichuriosis, trichocephalia-sis; whipworm infection. One of the geo-helminths.
LOCAL NAMES
Cacing kerawit (Bahasy Malay), Njowni(Luo), Keeri pulu (Tamil), Wakanabbyo(Japanese).
GEOGRAPHICAL DISTRIBUTION
Cosmopolitan but more common in warmhumid countries. There are about 800 mil-lion (Bundy and Cooper, 1989) to 1300million (Chan et al., 1994) cases in theworld, with one individual in about 500showing evidence of morbidity and 10,000deaths annually (8–13%; Bundy, 1994).
LOCATION IN HOST
The adults usually occur in the caecum butmay also be present in the appendix, rec-tum and upper colon with the thin anteriorend embedded in the mucosa (Figs 74–76).
MORPHOLOGY
The male measures 30–45 mm, with atightly coiled posterior end and a singlespicule. The female is slightly longer(30–50 mm) and both sexes have a narrowanterior portion and a much wider poste-rior end (hence the popular name of whip-
164 Chapter 5

worm, as they resemble the coaching whiponce used in Europe or a stock-whip (Fig.74). The mouth is simple, without lips, butwith a stylet for penetration of the mucosa.In the living worm the stylet is constantlyprobing and slashing, like a fencer wield-ing a rapier. The mouth is followed by asimple non-muscular oesophagus, sur-rounded by a single row of cells with anintracellular lumen and extending about
two-thirds from the anterior end. (Thewhole row is called a stichosome and eachcell is known as a stichocyte.) The oesoph-agus opens into the midgut at the end ofthe anterior thin part of the worm. Theanus is terminal. These features are typicalof the class Adenophorea, which have nophasmids.
In the female the vulva opens at thejunction of the oesophagus and intestine,the reproductive organs being confined tothe wider posterior portion. The uteruscontains about 60,000 eggs at one time.
LIFE CYCLE
The female lays about 2000–20,000 eggs 24 h�1. The characteristically shaped egg isbarrel-shaped with transparent terminalpolar prominences with a high chitin con-tent, and measures 57–58 µm � 26–28 µm,with an outer, brown bile-stained, shelland two inner shells (Fig. 123). The veryresistant egg contains an unsegmentedovum when laid, which takes from 2 weeksto many months to develop in moist soil,depending on the ambient temperature(4–6 months at 15°C, 3–4 weeks at 26°C, 17days at 30°C, 11 days at 35°C).Embryonated eggs are usually ingested oncontaminated food, such as salad vegeta-bles, or ingested in soil by infants. Thecontained second-stage larvae, measuring260 µm � 15 µm, hatch in the distal por-tion of the small intestine or colon. Theypenetrate the crypts of Lieberkühn of thecolon or caecum, where they live coiled upand after about a week move under theepithelium to the tip of the villus. The thinanterior end of each worm remains buriedin a tunnel in the mucosa, while the restemerges and hangs freely in the lumen.After the usual four moults, the femalesmature and lay eggs about 60–90 days afteringestion of the eggs.
The adult worms normally live for about1–2 years where there is repeated reinfec-tion, but can survive for much longer insome circumstances.
CLINICAL MANIFESTATIONS
In the great majority of infected individuals,only a few eggs can be found in a faecal
The Nematodes 165
Fig. 74. Adult male of Trichuris trichiura. Insertshows two females and a male (with coiled tail)attached to the mucosa of the colon by theiranterior ends.

sample and no overt symptoms can beattributed to the presence of the worms,except perhaps for diarrhoea; there is alsousually an eosinophilia of up to 25% inthe early stages. In heavy infections,Trichuris causes dysentery with blood inthe stools, abdominal or epigastric pain,headache, anorexia and weight loss, withprolapse of the rectum in children (inabout 30% of cases) (Plate 22). Iron-deficiency anaemia, pallor, malnutrition,stunting of growth, clubbing of fingers andpoor school performance are associatedwith heavy chronic infections in children(Forrester et al., 1990; Simeon et al., 1994;
Cooper et al., 1995; Ramdath et al., 1995;Gardner et al., 1996).
PATHOGENESIS
The anterior portions of the worms embed-ded in the mucosa cause petechial haemor-rhages and there is evidence that thepresence of T. trichiura predisposes toamoebic dysentery, presumably becausethe ulcers provide a suitable site for tissueinvasion by Entamoeba histolytica, and itmay also be involved in the spread of viraland bacterial pathogens (particularlyShigella). Cases have been reported inwhich the lumen of the appendix was
166 Chapter 5
Fig. 76. Section of colon with adult T. trichiura. Note thinner anterior end embedded in mucosa.
Fig. 75. Adults of T. trichiura in colon showing the slender anterior ends ‘threaded’ beneath theepithelium.

filled with worms and consequent irrita-tion and inflammation has led to appen-dicitis (but see p. 163) or granulomas. Themucosa of the large intestine becomeschronically inflamed, the extent dependingon the number of worms present. In heavyinfections there may be at least 1000 adultworms present (with about 30,000 eggs g�1
faeces), which can cause an insidiouschronic colitis, similar to Crohn’s disease,with inflammation extending to the rec-tum. A flexible fibre-optic colonoscopeshows that the wall of the caecum andascending colon is also hyperaemic andoedematous, with ulceration and bleeding.The bleeding might be partly due to bloodingestion by the parasites but is mainlycaused by enterorrhagia. Blood loss hasbeen estimated at about 0.005 ml per wormday�1. The rectal prolapse results fromstraining (tenesmus) consequent on irrita-tion and oedema of the lower colon simu-lating a full bowel (Plate 22).
Immune responses in humans have onlyrecently been demonstrated; ES antigens of16–17 kDa appear to be immunogenic inchildren (Needham and Lillywhite, 1994).Previously all knowledge on immunologyof infection was based on T. muris in themouse. In this host, parasites are rapidlyexpelled, with subsequent immunity tochallenge infection, unlike the situation inhuman infection. However, predispositionto infection has been demonstrated for allthe geohelminths, some individuals notbecoming infected in spite of being exposedas much as those who do (Crompton,1998). The immunology of all the intestinalnematodes is discussed on p. 122.
DIAGNOSIS
This is by finding the characteristic eggs inthe faeces. In very light infections concen-tration techniques can be used. More than1000 eggs in a 50 mg Kato–Katz faecalsmear (20,000 eggs g�1 faeces) (p. 257) rep-resents a heavy infection with overt symp-toms. It has been estimated that 2000 eggsg�1 faeces indicates the presence of 5–15adult worms but the relationship is notreliable. Each female worm produces about200–370 eggs g�1 stool 24 h�1.
In heavy infections sigmoidoscopy mayshow the white bodies of adult wormshanging from the inflamed mucosa – theso-called coconut-cake rectum.
TREATMENT
This is entirely by chemotherapy and,because of the sites of the adults, completeelimination used to be difficult. Recently,the newer benzimidazoles are proving to bevery effective.
Albendazole is given at a single oraldose of 200 mg but for symptomatic casesshould be given each day for 3 days. It isnot recommended for pregnant women orinfants under 1 year old (Savioli, 1999).Combined treatment with albendazole andivermectin, as recommended for filariasis(p. 199), appears to have a synergistic effectalso in trichuriasis.
Mebendazole at a single oral dose of400–500 mg is similarly effective.
Oxantel pamoate with pyrantel (forother intestinal nematodes), at 10–15 mgkg�1 body weight, has the advantage that itdoes not cause ectopic migration ofAscaris, but it does require weight mea-surements – chemical formula: (E)-3-(2-(1,4,5,6-tetrahydro-1-methyl-2-pyrimidinyl)-ethenyl)-phenol.
Thiabendazole is unpleasant to take andnot very effective and is no longer recom-mended.
EPIDEMIOLOGY
Trichuris infection is very common inmany countries where there is heavy rain-fall, dense shade and a constant tempera-ture between 22 and 28°C.
Recent estimates of infection rates are:25% in South and Central America, 31% inAfrica (with 76% in Nigeria and 70% inCameroon), 12% in the Middle East, 12%in southern India, 36% in Bangladesh,58% in South-East Asia except China and0.01% in Japan.
Fertile eggs are ingested with soil or onsalad vegetables. This is similar to the modeof infection in Ascaris and the two worms arecommonly present together. They are bothparticularly common where human faeces isused as a fertilizer (‘night-soil’). The larvae
The Nematodes 167

inside the eggs develop only under moistconditions and are killed by direct sunlight ora low relative humidity. Under conditions ofpoor sanitation both infections are peri-domestic and the areas around houses may beheavily contaminated with eggs (Plate 2) upto a depth of 20 cm; the habit of eating earth isan important reason why infants are ofteninfected by the age of 1. In the past, infectionwas also common in temperate countries,even though the eggs took a long time tobecome infective in the soil. Infection isoverdispersed with a small proportion of theinfected population having a high wormburden and producing most of the eggs reach-ing the environment (Bundy and Medley,1992).
Eggs of Trichuris were found in the rec-tum of the Alpine ‘ice man’, who lived about5300 years ago. He also had a bag containinga fungal purgative and anthelminthic(Capasso, 1998).
PREVENTION AND CONTROL
Personal protection is by the thoroughwashing of salad vegetables.
Until recently, control was almostentirely by sanitary disposal of faeces, butmass chemotherapy has recently becomemuch more practical, with the develop-ment of broad-action agents that can beused in a standard single oral dose againstmany intestinal helminths, and campaigns
are taking place against all the soil-transmitted nematodes (see p. 135).
Sanitation measures are likely to be veryslow to act. For instance, a campaign innorthern Italy in the 1950–1960s took 25years to reduce prevalence from 25 to 5% inschoolchildren. The basic reproductive rateof Trichuris is much higher than that ofhookworms or Ascaris, which makes it moreresistant to control. Children acquire approx-imately 90 new parasites per year and, in theabsence of constraints, each female wormwould produce four to six new egg-layingfemales (Bundy and Cooper, 1989).
ZOONOTIC ASPECTS
The same species occurs in monkeys andprimates, such as lemurs, catarrhine mon-keys and the orang-utan, but they are of noimportance as reservoir hosts. The pig par-asite, T. suis Schrank, 1788, is almost iden-tical to the human parasite, although theeggs are stated to be slightly larger (64 µm� 30 µm). Experimental infections inhumans are possible and it is likely thatnatural infections also occur where there isintensive pig farming. A few cases ofhuman infection with the dog and foxwhipworm, T. vulpis (Froelich, 1789), havebeen reported. Other species occur insheep and cattle (T. ovis) and mice (T.muris) and over 70 species have beenreported altogether; all from mammals.
168 Chapter 5
References
Adamson, M.L. (1989) Evolutionary biology of the Oxyurida (Nematoda): biofacies of a haplodiploidtaxon. Advances in Parasitology 28, 175–228.
Addiss, D.G. and Juranek, D.D. (1991) Lack of evidence for a causal association between parasiticinfections and acute appendicitis. Journal of Infectious Diseases 164, 1036–1037.
Albonico, M., Crompton, D.W.T. and Savioli, L. (1998) Control strategies for human intestinal nema-tode infections. Advances in Parasitology 42, 278–343.
Al Rabia, F., Halim, M.A., Ellis, M.E. and Abdulkareem, A. (1996) Enterobius vermicularis and acuteappendicitis. Saudi Medical Journal 17, 799–803.
Anderson, R.C., Chabaud, A.C. and Willmott, S. (eds) (1974–1983) CIH Keys to the NematodeParasites of Vertebrates, Vols. 1–10. CAB International, Wallingford, UK.
Anderson, R.M. and Schad, G.A. (1985) Predisposition to hookworm infection in humans. Science228, 1537–1540.
Anderson, T.J.C. and Jaenike, J. (1997) Host specificity, evolutionary relationships and macrogeo-graphical differentiation among Ascaris populations from humans and pigs. Parasitology 115,325–342.

Ashford, R.W., Barnish, G. and Viney, M.E. (1992) Strongyloides fuelleborni kellyi: infection and dis-ease in Papua New Guinea. Parasitology Today 8, 314–318.
Blotkamp, J., Krepel, H.P., Kumar, V., Baeta, S., Van’t Noordende, J.M. and Polderman, A.M. (1993)Observations on the morphology of adults and larval stages of Oesophagostomum sp. isolatedfrom man in northern Togo and Ghana. Journal of Helminthology 67, 49–61.
Borkow, G. and Beutwich, Z. (2000) Eradication of helminthic infections may be essential for suc-cessful vaccination against HIV and tuberculosis. Bulletin of the World Health Organization 78,1368–1369.
Boschetti, A. and Kasznica, J. (1995) Visceral larval migrans induced eosinophilic cardiac pseudo-tumour: a cause of sudden death in a child. Journal of Forensic Sciences 40, 1097–1099.
Bradley, M. (1990) Rate of expulsion of Necator americanus and the false hookworm Ternidensdeminutus Railliet and Henry, 1909 (Nematoda) from humans following albendazole treatment.Transactions of the Royal Society of Tropical Medicine and Hygiene 84, 720.
Bundy, D.A.P. (1994) Immunoepidemiology of intestinal helminthic infection. 1. The global burdenof intestinal nematode disease. Transactions of the Royal Society of Tropical Medicine andHygiene 88, 259–261.
Bundy, D.A.P. and Cooper, E.S. (1989) Trichuris and trichuriasis in humans. Advances inParasitology 28, 108–174.
Bundy, D.A.P. and Medley, G.F. (1992) Immunoepidemiology of human geohelminthiasis: ecologicaland immunological determinants of worm burden. Parasitology 104 (suppl.), S105–S119.
Bundy, D., Sher, A. and Michael, E. (2000) Helminth infections may increase susceptibility to TB andAIDS. Parasitology Today 16, 273–274.
Capasso, L. (1998) 5300 years ago the Ice Man used natural laxatives and antibiotics. Lancet, BritishEdition 352 (9143), 1864.
Chan, M.S., Medley, G.F., Jamison, D. and Bundy, D.A.P. (1994) The evaluation of potential globalmorbidity attributable to intestinal nematode infections. Parasitology 109, 373–387.
Chittenden, A.M. and Ashford, R.W. (1987) Enterobius gregorii Hugot, 1983: first report in the UK.Annals of Tropical Medicine and Parasitology 81, 195–198.
Chye, S.M., Yen, S.M. and Chen, E.R. (1997) Detection of circulating antigen by monoclonal antibod-ies for immunodiagnosis of angiostrongyliasis. American Journal of Tropical Medicine andHygiene 56, 408–412.
Conway, D.J., Atkins, N.S., Lillywhite, J.E., Bailey, J.W., Robinson, R.D., Lindo, J.F., Bundy, D.A.P.and Bianco, A.E. (1993) Immunodiagnosis of Strongyloides stercoralis infection: a method forincreasing the specificity of the indirect ELISA. Transactions of the Royal Society of TropicalMedicine and Hygiene 87, 173–176.
Conway, D.J., Lindo, J.F., Robinson, R.D. and Bundy, D.A.P. (1995) Towards effective control ofStrongyloides stercoralis. Parasitology Today 11, 420–427.
Cook, G.C. (1994) Enterobius vermicularis infection. Gut 35, 1159–1162.Cooper, E.S., Duff, E.M.W., Howell, S. and Bundy, D.A.P. (1995) ‘Catch up’ growth velocities after
treatment for the Trichuris dysentery syndrome. Transactions of the Royal Society of TropicalMedicine and Hygiene 89, 653.
Croese, J., Loukas, A., Opdebeeck, J., Fairley, S. and Prociv, P. (1994) Human enteric infection withcanine hookworms. Annals of Internal Medicine 120, 369–374 (also Gasteroenterology 106, 3–12).
Crompton, D.W.T. (1998) Gastrointestinal nematodes. In: Cox, F.E.G., Kreier, J.P. and Wakelin, D.(eds) Topley and Wilson’s Microbiology and Microbial Infections. Vol. 5. Parasitology, 9th edn.Arnold, London, UK, pp. 561–584. .
Crompton, D.W.T. (2000) Ascaris and ascariasis. Advances in Parasitology 48, 286–376.Crompton, D.W.T., Nesheim, M.C. and Pawlowski, Z.S. (1989) Ascariasis and its Prevention and
Control. Taylor and Francis, London, UK.Cross, J.H. (1987) Public health importance of Angiostrongylus cantonensis and its relatives.
Parasitology Today 3, 367–369.Cunningham, C.K., Kazacos, K.R., McMillan, J.A., Lucas, J.A., McAuley, J.B., Wozniak, E.J. and
Weiner, L.B. (1994) Diagnosis and management of Baylisascaris procyonis infection in an infant,with nonfatal meningoencephalitis. Clinical Infectious Diseases 18, 868–872.
de Aguilar Nasumento, J.E., Silva, G.M., Tadano, T., Valadores Filho, M., Akiyama, A.M.P. andCastelo, A. (1993) Infection of the soft tissue of the neck due to Lagochilascaris minor.Transactions of the Royal Society of Tropical Medicine and Hygiene 87, 198.
The Nematodes 169

Dorfman, S., Talbot, I.C., Torres, R., Cardozo, J. and Sanchez, M. (1995) Parasitic infestation in acuteappendicitis. Annals of Tropical Medicine and Parasitology 89, 99–101.
Forrester, J.E., Scott, M.E., Bundy, D.A.P. and Golden, M.H.N. (1990) Predisposition of individualsand families in Mexico to heavy infection with Ascaris lumbricoides and Trichuris trichiura.Transactions of the Royal Society of Tropical Medicine and Hygiene 84, 272–276.
Garcia, M., Moneo, I., Audicana, M.T., del Pozo, M.D., Munoz, D., Fernandez, E., Diez, J.,Etxenagusia, M.A., Ansotegni, I.J. and de Corres, L.F. (1997) The use of IgE immunoblotting as adiagnostic tool in Anisakis simplex allergy. Journal of Allergy and Clinical Immunology 99,497–501.
Gardiner, C.H., Koh, D.S. and Cardella, T.A. (1981) Micronema in man: third fatal infection.American Journal of Tropical Medicine and Hygiene 30, 586–589.
Gardner, J.M., Grantham-McGregor, S., Baddeley, A. and Meeks-Gardner, J. (1996) Trichuris trichiurainfection and cognitive function in Jamaican schoolchildren. Annals of Tropical Medicine andParasitology 90, 55–63.
Geissler, P.W., Mwaniki, D., Thiong’o, F. and Friis, H. (1998) Geophagy as a risk factor for geo-helminth infections: a longitudinal study of Kenya schoolchildren. Transactions of the RoyalSociety of Tropical Medicine and Hygiene 92, 7–11.
Genta, R.M. (1992) Dysregulation of strongyloidiasis: a new hypothesis. Clinical MicrobiologyReviews 5, 345–355.
Gibson, D.I. (1983) Ascaridoid nematodes – a current assessment. In: Stone, A.R. and Khalil, L.F.(eds) Concepts in Nematode Systematics. Academic Press, London, UK, pp. 321–338.
Gillespie, S.H. (1988) The epidemiology of Toxocara canis. Parasitology Today 4, 180–182.Gillespie, S.H. (1993) The clinical spectrum of human toxocariasis. In: Lewis, J.W. (ed.) Toxocara and
Toxocariasis: Clinical, Epidemiological and Molecular Perspectives. Institute of Biology, London,UK, pp. 55–61.
Gillespie, S.H., Dinning, W.J., Voller, A. and Crowcroft, N.S. (1993) The spectrum of ocular toxocaria-sis. Eye 7, 415–418.
Gilman, R.H., Marquis, G.S. and Miranda, E. (1991) Prevalence and symptoms of Enterobius vermicu-laris infections in a Peruvian shanty town. Transactions of the Royal Society of Tropical Medicineand Hygiene 85, 761–764.
Goldsmid, J.M. (1991) The African ‘hookworm’ problem: an overview. In: Macpherson, C.N.L. andCraig, P.S. (eds) Parasitic Helminths and Zoonoses in Africa. Unwin Hyman, London, UK, pp.101–137.
Grove, D.I. (1996) Human strongyloidiasis. Advances in Parasitology 38, 251–309.Hall, A. and Holland, C. (2000) Geographical variation in Ascaris lumbricoides fecundity and its
implication for helminth control. Parasitology Today 16, 540–544.Hall, A., Conway, D.J., Anwar, K.S. and Rahman, M.D. (1994) Strongyloides stercoralis in an urban
slum community in Bangladesh: factors independently associated with infection. Transactions ofthe Royal Society of Tropical Medicine and Hygiene 88, 527–530.
Haswell-Elkins, M.R., Elkins, D.B. and Anderson, R.M. (1987a) Evidence for predisposition inhumans to infection with Ascaris, hookworm, Enterobius and Trichuris in a South Indian fishingcommunity. Parasitology 95, 323–338.
Haswell-Elkins, M.R., Elkins, D.B., Manjula, K., Michael, E. and Anderson, R.M. (1987b) The distrib-ution and abundance of Enterobius vermicularis in a South Indian fishing community.Parasitology 95, 339–354.
Hotez, P.J. and Prichard, D.I. (1995) Hookworm infection. Scientific American 272, 42–48.Hugot, J.P. and Tourte-Schaefer, C. (1985) Etude morphologique des deux oxyures parasites de
l’homme: Enterobius vermicularis et E. gregorii. Annales de Parasitologie Humaine et Comparée60, 57–64.
Ishikura, H., Kikuchi, K., Nagasawa, K., Ooiwa, T., Takamiya, H., Sato, N. and Sugane, K. (1992)Anisakidae and anisakidosis. In: Progress in Clinical Parasitology, Vol. 3. Springer Verlag, NewYork, USA, pp. 43–102.
Jelinek, T., Maiwald, H., Nothdurft, H.D. and Loscher, T. (1994) Cutaneous larva migrans in trav-ellers: synopsis of histories, symptoms and treatment of 98 patients. Clinical Infectious Diseases19, 1062–1066.
Kagei, N. and Isogaki, H. (1992) A case of abdominal syndrome caused by the presence of a largenumber of Anisakis larvae. International Journal for Parasitology 22, 251–253.
170 Chapter 5

Kayes, S.G. (1997) Human toxocariasis and the visceral larval migrans syndrome: correlativeimmunopathology. In: Freedman, D.O. (ed.) Chemical Immunology, Vol. 66. S. Karger, Basle,pp. 99–124.
Kazacos, K.R. (1983) Raccoon Roundworms (Baylisascaris procyonis): a Cause of Animal andHuman Disease. Bulletin No. 422, Agriculture Experimental Station, Purdue University, WestLafayette, USA.
Kliks, M.M. and Palumbo, N.E. (1992) Eosinophilic meningitis beyond the Pacific basin: the globaldispersal of a peridomestic zoonosis caused by Angiostrongylus cantonensis, the nematode lung-worm of rats. Social Science and Medicine 34, 199–212.
Mangali, A., Chaicumpa, W., Nontasut, P., Chantavanij, P. and Viravan, C. (1991) Enzyme-linkedimmunosorbent assay for diagnosis of human strongyloidiasis. Southeast Asian Journal ofTropical Medicine and Public Health 22, 88–92.
Marti, H., Haji, H.J., Savioli, L., Chwaya, H.M., Mgeni, A.F., Ameir, J.S. and Hatz, C. (1996) A compar-ative trial of a single-dose ivermectin versus three days of albendazole for treatment ofStrongyloides stercoralis and other soil-transmitted helminth infections in children. AmericanJournal of Tropical Medicine and Hygiene 55, 477–481.
Mizgajska, H. (1993) The distribution and survival of eggs of Ascaris suum in six different naturalsoil profiles. Acta Parasitologia 38, 170–174.
Morera, P. (1994) Importance of abdominal angiostrongylosis in the Americas. In: Özcel, M.A. andAlkan, M.Z. (eds) Parasitology for the 21st Century. CAB International, Wallingford, UK, pp.253–260.
Muller, R. (1983) Ancylostoma, Necator and ancylostomiasis. In: Feachem, R.G., Bradley, D.J.,Garelick, H. and Mora, D.D. (eds) Sanitation and Disease: Health Aspects of Excreta andWastewater Management. John Wiley & Sons, Chichester, UK, pp. 359–373.
Murrell, K.D., Eriksen, L., Nansen, P., Slotred, H.C. and Rasmussen, T. (1997) Ascaris suum: a revi-sion of its early migratory path and implications for human ascariasis. Journal of Parasitology 83,255–260.
Needham, C.S. and Lillywhite, J.E. (1994) Immunoepidemiology of intestinal helminthic infections.2. Immunological correlates with patterns of Trichuris infection. Transactions of the Royal Societyof Tropical Medicine and Hygiene 88, 262–264.
Pawlowski, Z.S., Schad, G.A. and Stott, G.J. (1991) Hookworm Infection and Anaemia: Approachesto Prevention and Control. World Health Organization, Geneva, Switzerland.
Peng Weidong, Zhou Xianmin and Crompton, D.W.T. (1998) Ascariasis in China. Advances inParasitology 41, 109–148.
Petithory, J.C., Pangam, B., Buyet-Rousset, P. and Pangam, A. (1990) Anisakis simplex, a co-factor ofgastric cancer? Lancet 336, 1002.
Phelps, K.R. and Neva, F.A. (1993) Strongyloides hyperinfection in patients coinfected with HTLV-Iand Strongyloides stercoralis. American Journal of Medicine 94, 447–449.
Pit, D.S.S., de Graaf, W., Snoek, H., de Vlas, S.J., Baeta, S.M. and Polderman, A.M. (1999) Diagnosisof Oesophagostomum bifurcum and hookworm infection in humans: day-to-day and within-specimen variation of larval counts. Parasitology 118, 283–288.
Polderman, A.M. and Blotkamp, J. (1995) Oesophagostomum infections in humans. ParasitologyToday 11, 541–456.
Polderman, A.M., Krepel, H.P., Baeta, S., Blotkamp, J. and Gigase, P. (1991) Oesophagostomiasis, acommon infection of man in Northern Togo and Ghana. American Journal of Tropical Medicineand Hygiene 44, 336–344.
Polderman, A.M., Krepel, H.P., Verweij, J.J., Baeta, S. and Rotmans, J.P. (1993) Serological diagnosisof Oesophagostomum infections. Transactions of the Royal Society of Tropical Medicine andHygiene 87, 433–435.
Polderman, A.M., Anemana, S.D. and Asigri, V. (1999) Human oesophagostomiasis; a regional publichealth problem in Africa. Parasitology Today 15, 129–130.
Prichard, D.I. (1993) Immunity to helminths: is too much IgE parasite- rather than host-protective?Parasite Immunology 15, 5–9.
Prociv, P. (1989) Toxocara pteropodis and visceral larva migrans. Parasitology Today 5, 106–109. Prociv, P. and Croese, J. (1996) Human enteric infection with Ancylostoma caninum: hookworms
reappraised in the light of a ‘new’ zoonosis. Acta Tropica 62, 23–44.Prociv, P., Spratt, D.M. and Carlisle, M.S. (2000) Neuro-angiostrongyliosis: unresolved issues.
International Journal for Parasitology 30, 1295–1303.
The Nematodes 171

Ramdath, D.D., Simeon, D., Wong, M.S. and Grantham-McGregor, S.M. (1995) Iron status of school-children with varying intensities of Trichuris trichiura infection. Parasitology 110, 347–351.
Ross, R.A., Gibson, D.J. and Harris, E.A. (1989) Cutaneous oesophagostomiasis in man. Journal ofHelminthology 63, 261–265.
Sakti, H., Nokes, C., Hertanto,W.S., Hendratno, S., Hall, A., Bundy, D.A.P. and Satoto (1999) Evidenceof an association between hookworm infection and cognitive function in Indonesian school chil-dren. Tropical Medicine and International Health 4, 322–334.
Sato, Y., Toma, H., Kiyuna, S. and Shiloma, Y. (1991) Gelatin particle indirect agglutination test formass examination for strongyloidiasis. Transactions of the Royal Society for Tropical Medicineand Hygiene 85, 515–518.
Savioli, L. (1999) Treatment of Trichuris infection with albendazole. Lancet (British edition) 353(9148), 237.
Simeon, D., Callender, J., Wong, M., Grantham-McGregor, S.M. and Ramdath, D.D. (1994) School per-formance, nutritional status and trichuriasis in Jamaican school children. Acta Pediatrica 83,1188–1193.
Sohn, W.M. and Seol, S.Y. (1994) A human case of gastric anisakiasis by Pseudoterranova decipienslarva. Korean Journal of Parasitology 32, 53–56.
Sugane, K., Sin, S. and Matsuura, T. (1992) Molecular cloning of the cDNA encoding a 42 kDa anti-genic polypeptide of Anisakis simplex larvae. Journal of Helminthology 66, 25–32.
Sun, T., Schwartz, N.S., Sewell, C., Lieberman, P. and Gross, S. (1991) Enterobius egg granuloma ofthe vulva and peritoneum: review of the literature. American Journal of Tropical Medicine andHygiene 45, 249–253.
Thien Hlang (1993) Ascariasis and childhood malnutrition. Parasitology 107, S125–S136.Udonsi, J.K. and Atata, G. (1987) Necator americanus: temperature, pH, light, and larval develop-
ment, longevity, and desiccation tolerance. Experimental Parasitology 63, 136–142.Wakelin, D. (1994) Host populations: genetics and immunity. In: Scott, M.E. and Smith, G. (eds)
Parasitic and Infectious Diseases. Academic Press, London, UK, pp. 83–100.Wakelin, D. (1996) Immunity to Parasites: How Parasitic Infections are Controlled. Cambridge
University Press, Cambridge, UK.Wariyapola, D., Goonesinghe, N., Priyamanna, T.H.H., Fonseka, C., Ismail, M.M., Abeyewickreme, W.
and Dissanaike, A.S. (1998) Second case of ocular parastrongyliasis from Sri Lanka. Transactionsof the Royal Society of Tropical Medicine and Hygiene 92, 64–65.
Xingquan, Z., Chilton, N.B., Jacobs, D.E., Boes, J. and Gasser, R.B. (1999) Characterisation of Ascarisfrom human and pig hosts by nuclear ribosomal DNA sequences. International Journal forParasitology 29, 469–478.
Zhou Xianmin, Peng Weidong, Crompton, D.W.T. and Xing Jianquin (1999) Treatment of biliaryascariasis in China. Transactions of the Royal Society of Tropical Medicine and Hygiene 93,561–564.
Further Reading
Bundy, D.A.P., Chan, M.S., Medley, C.F., Jamison, D. and Savioli, L. (2001) Intestinal nematode infec-tions. In: Murray, C.J.L. and Lopez, A.D. (eds) The Global Epidemiology of Infectious Diseases.Harvard University Press, Cambridge, Massachusetts, USA (in press).
Farthing, M.J.G., Keusch, G.T. and Wakelin, D. (eds) (1995) Enteric Infections 2: Intestinal Helminths.Chapman and Hall, London, UK.
Gilles, H.M. and Ball, P.A. (eds) (1991) Hookworm Infections. Human Parasitic Diseases, Vol. 4.Elsevier, Amsterdam, The Netherlands.
Grove, D.I. (ed.) (1989) Strongyloidiasis: a Major Roundworm of Man. Taylor and Francis, London, UK. Holland, C. (1987) Neglected infections – trichuriasis and strongyloidiasis. In: Stephenson, L.S. (ed.)
Impact of Helminth Infections on Human Nutrition. Taylor and Francis, London, UK, pp.161–201.
Miller, T.A. (1979) Hookworm infection in man. Advances in Parasitology 17, 315–384.Sakanari, J.A. and McKerrow, J.H. (1989) Anisakiasis. Clinical Microbiology Reviews 2, 278–284.Schad, G.A. and Warren, K.S. (eds) (1990) Hookworm Disease: Current Status and New Directions.
Taylor and Francis, London, UK.
172 Chapter 5

Tissue Nematodes
All the nematodes considered in this sec-tion invade the tissues of the host, includ-ing those of the intestine, and have a lifecycle involving an intermediate host.Members of the first group, includingTrichinella, are only distantly related to allthe rest and have vertebrate intermediatehosts, in contrast to the others where theseare always arthropods and all of whichbelong to the order Spirurida.
Order Enoplida
Family Trichuridae
Aonchotheca philippinensis (Chitwood, Velasquez and Salazar, 1968)
Moravec, 1982
SYNONYM
Capillaria philippinensis Chitwood,Velasquez and Salazar 1968.
DISEASE AND COMMON NAME
Intestinal capillariasis or capillariosis; acause of non-vibrio cholera.
GEOGRAPHICAL DISTRIBUTION
This parasite was first described 30 yearsago from villages bordering the China Seaon the west coast of the island of Luzon inthe Philippines. It has been reported sincefrom the islands of Bohol, Mindanao andLeyte. Numerous cases have also beenfound in Thailand and others from Egypt,India, Indonesia, Iran, Japan and SouthKorea.
LOCATION IN HOST
The adults become embedded in themucosa of the small intestine, especiallythe jejunum.
MORPHOLOGY
Aonchotheca is related to Trichinella andTrichuris and there are many species of cap-illarids that are parasites of a wide range ofvertebrates. The adult worms are very small,females measuring 2.5–5.3 mm in length and
males 1.3–3.9 mm. In the female, the vulvalopening is just behind the oesophagus andthe male has a single very long spicule in asheath. Both sexes have a small spear at theanterior end.
LIFE CYCLE
The life cycle has been determined experi-mentally in monkeys and Mongolian jirds.Freshwater fishes indigenous to thePhilippines were fed eggs from patientsembryonated for 5–10 days in water.Larvae hatched in the intestines of the fishand reached the infective stage in about 3weeks but could live for months; larvaedeveloped into adults in 12–14 days whenfed to monkeys or jirds.
Following copulation and fertilization,the females produce larvae. The first-generation larvae are retained and developinto second-generation males and femalesin another 2 weeks. The second-generationfemales produce eggs passed out in the fae-ces about 26 days after fertilization.
Autoinfection occurs in jirds and almostcertainly in humans, as all stages of devel-opment, including viviparous andlarviparous females, have been found atautopsy. In one experiment, two larvaefrom fish produced 5353 worms in a jird.
Several species of freshwater and brack-ish-water fish have been experimentallyinfected; Hypeleotris bipartita, Ambassismiops and Eleotris melanosoma are ofteneaten raw in the Philippines and Cyprinuscarpio in Thailand.
The natural definitive hosts are probablyfish-eating birds, which can be infectedexperimentally and one bittern has beenfound naturally infected.
CLINICAL MANIFESTATIONS
In the initial epidemic in 1967, when over100 people died, all patients with eggs inthe faeces had intermittent diarrhoea, withborborygmi and diffuse recurrent abdomi-nal pain. The syndrome resembled tropicalsprue, with five to ten voluminous stoolsper day. Weight loss, anorexia and vomit-ing were common. Patients who recoveredfrom the initial attack usually suffered sev-eral relapses and, in a study conducted 3
The Nematodes 173

years after the initial epidemic, patientswho were still symptomatic were found tohave depression of the D-xylose test andimpairment of fat and protein digestionand absorption. A mortality rate of 35% inuntreated males and 19% in untreatedfemales was reported from one village inthe initial epidemic.
Nowadays, only the early symptoms ofborborygmi, diarrhoea and abdominal painare present before treatment is sought.
PATHOGENESIS
The stools contain large amounts of fat,protein and minerals, particularly potas-sium, which results in a marked protein-losing enteropathy. Malabsorption alsooccurs. Muscle wasting, emaciation, weak-ness and cardiopathy were reported from73% of early patients. Without treatment,death often ensues 2 weeks to 2 monthsafter the onset of symptoms, due to cardiacfailure or to intercurrent infection.
Necropsy studies on ten Philippinosshowed the presence of adults and larvaeembedded in the mucosa of the jejunum,together with numerous eggs (Fig. 77).
DIAGNOSIS
Clinical. The early symptoms are typicaland easily recognized.
Parasitological. Eggs, larvae and adults maybe found in the faeces. Eggs measure 36–45µm and have a flattened bipolar plug ateach end (Fig. 124). They could be mis-taken for the eggs of Trichuris but differ inhaving a pitted shell, a more ovoid shape, aslightly smaller size and no protuberanceof the plugs. Some eggs may contain larvae.The eggs of both parasites may be foundtogether in the faeces.
Serological. ELISA, double diffusion andindirect haemagglutination assay (IHA)have been tried but are not satisfactory.
TREATMENTMebendazole at 400 mg day�1 in divideddoses for 20–30 days is the current drug ofchoice. If treatment is curtailed there islikely to be a relapse, since larvae may not
be affected and there is multiplicationwithin the host.
Albendazole: In the few patients treatedthis drug appears to affect larvae as well asadults at a dose of 200 mg twice daily for14–21 days and may be more effective.
Thiabendazole can be given at 25 mgkg�1 body weight day�1 for at least amonth, but relapses are common.
EPIDEMIOLOGYRaw fish are a popular item of diet in thePhilippines and Thailand and were impli-cated in the cases in Japan and Iran.
Presumably infection could be pre-vented by thorough cooking of fish or evenby removing the intestines.
Capillarid eggs passed over a period bya patient in Irian Jaya differed in size fromthose of all the species considered here(Bangs et al., 1994).
174 Chapter 5
Fig. 77. Section of ileum with femaleAonchotheca (cut tangentially) invading the villiand containing many unembryonated thick-shelledeggs in the vagina. A section of the same wormabove shows a portion of the stichosome andoesophagus.

Calodium hepaticum (Bancroft, 1893) Moravec, 1982
SYNONYMS
Trichocephalus hepaticus Bancroft, 1893;Hepaticola hepatica (Bancroft, 1893) Hall,1916; Capillaria hepatica (Bancroft, 1893)Travassos, 1915.
DISEASE AND POPULAR NAMES
Hepatic capillariasis or capillariosis; thecapillary liver worm.
GEOGRAPHICAL DISTRIBUTION
About 25 human cases have been reportedfrom many countries, including Brazil,England, Germany, India, Italy, SouthKorea, Mexico, Nigeria, Slovakia, SouthAfrica, Turkey, continental USA andHawaii. C. hepaticum is a natural parasiteof the liver tissues of rats, mice, prairiedogs, muskrats, beavers, hares and mon-keys.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Female worms measure about 70 mm longby 0.12 mm wide, males 22 mm by0.05 mm. The anterior portion of the wormcontains the stichosome, oesophagus andbacillary bands, the posteror part the intes-tine and reproductive organs. Eggs measure48–66 µm � 28–38 µm (larger than thoseof Trichuris) and are thick-shelled, withsurface pits and bipolar caps that do notprotrude. The adults live in the liverparenchyma. The females lay eggs in thetissues and these accumulate and formlarge whitish spots under the liver surface.The eggs do not pass out in the faeces butreach the soil after the death of the host orafter the host is eaten by another and theeggs are passed in the faeces.
Clinical symptoms resemble those ofvisceral larva migrans, with an initialhepatitis. The liver is tender and enlarged,with persistent fever, anorexia, weight lossand a very high (over 90%) eosinophilia.There is focal destruction of parenchymaand fibrosis in chronic infections.
Diagnosis is by needle liver biopsy orusually at post-mortem. The presence of
eggs in the faeces is evidence of a spuriousinfection from eating an infected animalliver. A recent case was treated success-fully with albendazole at 10–20 mg kg�1
body weight day�1 for 20 days and thenthiabendazole at 25 mg kg�1 day�1 for 27days, although hepatomegaly was still pre-sent 2 years later (Choe et al., 1993). Mostinfections have been in children and arecent case in a 14-month-old girl wastreated. Eggs are presumably ingested invegetables or on soil after passing throughthe gut of a predator.
Eucoleus aerophilus (Creplin, 1839) Dujardin, 1845
SYNONYM AND DISEASE
Capillaria aerophila Creplin, 1839.Pulmonary capillariasis or capillariosis.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
This is a worldwide parasite of wild carni-vores and 11 human cases have beenreported from France, Iran, Morocco andRussia. Adults live buried under themucosa of the respiratory tract, the femalemeasuring 20 mm � 0.1 mm and the male18 mm � 0.07 mm. Eggs reach the intestinefrom the lungs and are passed out in thefaeces. Infection is common in dogs in theUSA but this might be a separate species(E. boehmi).
Clinical symptoms include a painful drycough, fever, dyspnoea and acute tracheo-bronchitis. There is some eosinophilia. Inone biopsy there were numerous smallgranulomatous lesions in the wall of asmall bronchus. Ivermectin at 200 µg kg�1
has had encouraging results and repeateddoses of thiabendazole have been tried.
Three cases of Anatrichosoma cutaneum(Swift, Boots and Miles, 1922) Chitwoodand Smith, 1956, a parasite of the rhesusmonkey, have been reported from humansin Japan, Malaysia (Marwi et al., 1990) andVietnam. Adults and eggs are found inlesions in the skin. Eggs removed from thefacial skin were similar to those ofTrichuris but larger (59 µm � 28 µm).
The Nematodes 175

Family Trichinellidae
Trichinella spiralis (Owen, 1835) Railliet, 1895
SYNONYM
Trichina spiralis Owen, 1835.
DISEASE AND POPULAR NAME
Trichiniasis or trichinellosis, trichinosis;trichina worm infection.
GEOGRAPHICAL DISTRIBUTION
Infection with Trichinella species is world-wide in carnivorous or scavenging animalsbut is particularly important in humans inAlaska and Canada in Inuit, Chile, easternEurope, Kenya and Russia. Other endemicareas include Central and South America(including the Caribbean), central andwestern Europe, China, Indonesia, Japan,New Zealand, the Pacific area (particularlyHawaii) and the Aleutians.
LOCATION IN HOST
The short-lived adults are partially embed-ded in the mucosa of the ileum. The larvaeare present in cysts in all the striated mus-cles.
MORPHOLOGY
The adults are minute, threadlike worms,the females measuring 2.8 (2.4–3.4) mm �0.06 mm and the males 1.5 (1.3–1.6) mm �0.03 mm. Both sexes are wider posteriorlythan anteriorly. The oesophagus differsfrom that of all other animal-parasiticnematodes (apart from Trichuris and capil-larids), as its posterior part is non-muscular but is surrounded by a column ofcuboidal cells, known as a stichosome orcell body (Zellenkörper), with an intracel-lular canal. The oesophagus connects witha thin-walled intestine, which ends in aterminal anus. The male has two copula-tory appendages at the posterior end but nospicules. The reproductive system of thefemale consists of a single ovary, oviduct,seminal receptacle, uterus, vagina andvulva. Trichinella is ovoviviparous, so thatin the mature female the uterus is filledwith larvae (Figs 78–80).
LIFE CYCLE
The muscle cysts containing larvae areingested in raw or semi-cooked meat. Thecyst wall is digested out in the stomachand the larvae emerge in the duodenum.The released first-stage larvae invade themucosa of the duodenum and jejunum andundergo four moults. The immature wormsemerge into the lumen of the intestine22–24 h after ingestion of cysts and, inmice, fertilized females have been observedat 30 h. In most experimental hosts,approximately twice as many females asmales are present and both sexes re-enter
176 Chapter 5
Fig. 78. Adult female and male of Trichinellaspiralis.

the mucosa. The female begins to producefirst-stage larvae (measuring 80–129 µm �5–6 µm) approximately 5 days after inges-tion of cysts. Each female produces200–2000 larvae during its lifetime. Thelarvae are deposited in the mucosa andreach the active skeletal muscles, travelling
by the lymphatics and blood-vessels. Afterreaching the muscles and penetrating thesarcolemma with the help of a stylet, thelarvae grow, measuring about 1 mm inlength 17–20 days after infection. Thesexes can be differentiated at this time. Thelarvae coil and by 17–21 days after infec-tion are capable of transmission to a newhost, as they can resist the action of diges-tive juices. The normal lifespan of thefemale worms in the intestine of humans isnot known but from autopsy studiesappears to be less than 2 months.
From the point of view of the parasite,humans are dead-end hosts. Transmissionnormally takes place from one flesh-eatinganimal (including carrion eaters) toanother, often of the same species, as withrats and mice.
CLINICAL MANIFESTATIONS
The first signs and symptoms can becaused by the development of the adultsand production of larvae in the intestine;in heavy infections they can be proteanand infection is often misdiagnosed. Withabout 1000 larvae g�1 muscle tissue, theremay be a sudden onset of illness, with nau-sea, vomiting, epigastric pain and severewatery diarrhoea, followed by pyrexia(38–40°C), myalgias, facial or generalized
The Nematodes 177
Fig. 79. Portion of the stichosome of T. spiralis.
Fig. 80. An electron micrograph of the cuticle and epicuticle of T. spiralis (original magnification � 82,000).

oedema (Plate 23) and urticarial manifesta-tions. There are also likely to be cardiovas-cular, renal and central nervous systemsymptoms. Eosinophilia is often the firstclinical sign and becomes apparent about10 days after infection, reaching a maxi-mum (usually 25–50%, but may be as highas 90% with 4000 eosinophils mm�3) dur-ing the 3rd or 4th week, and slowly dimin-ishing over a period of months. There isalso some degree of leucocytosis. Whenonly a few cysts are ingested, these may bethe only signs and symptoms, and themajority of cases of trichiniasis are diag-nosed only at routine autopsy.
Larvae in the muscles can cause manyeffects (Fig. 81). Transient cardiac manifes-tations (feeble pulse, cardiac dilatation andapical systolic murmur and palpitations),various nervous system manifestations(meningitis or meningoencephalitis, hemi-plegia, coma and neuritis with muscularparalysis) and pulmonary manifestations(effort dyspnoea, cough, hoarseness,bronchopneunomia, chest pain and pul-monary oedema) are all possible (Capo andDespommier, 1996). Ocular signs andsymptoms occur in over one-third of clini-cal cases and characteristically includeorbital oedema and waxy yellow chemosisof the bulbar conjunctiva, with photo-
phobia and blurred vision. Scleral or reti-nal haemorrhages with pain on pressure ormovement of the eyes and diplopia mayalso occur.
Flame-shaped splinter haemorrhagesunder the fingernails are sometimes seenand are almost diagnostic.
About 90% of infections are light, withfewer than ten larvae g�1 of muscle. One ofthe heaviest infections known was a fatalone in a young boy in East Africa, who hadover 5000 cysts g�1 of tongue muscle and6530 cysts g�1 in another boy (see T. nel-soni below). With massive infections likethis, there is a greater volume of cyst sub-stance than of muscle fibre. However, deathmay result with as few as 100 cysts g�1, theseverity of the reaction probably beingmore important than the number of larvaepresent. In very heavy infections, death ismore likely between the 4th and 8th weeks,from exhaustion, pneumonia or cardiacfailure.
Some infections in Inuit in northernCanada and Alaska from eating raw walrusor seal show atypical symptoms of pro-longed diarrhoea, with no fever or oedema,short myalgia, but a high eosinophilia (thisis caused by T. nativa – see below); thispossibly occurs in patients who have beenpreviously infected (infections are likely to
178 Chapter 5
Fig. 81. Section of skeletal muscle with cysts of Trichinella and interstitial inflammatory cells between themuscle cells. From a fatal case with severe myositis (original magnification � 65).

be high since they are almost entirely meateaters).
There is usually eventual completerecovery in most patients, but muscularpain (in 60% of sufferers), fatigue (in 52%)and cardiovascular pains may persist for1–6 years. The presence of two cysts pergram of muscle is likely to result in mildclinical symptoms.
A few cases are recorded where deathhas occurred about 10 days after infectionowing to the action of the stages in theintestine.
PATHOGENESIS
In the early stages the mucosa of the diges-tive tract may be hyperaemic, with inflam-mation consisting of eosinophils,neutrophils and lymphocytes and petechialhaemorrhages at the site of attachment.Antigen–antibody complexes form in thetissues and are probably involved in theoccasional severe gut reactions found inheavy infections (antibodies are producedparticularly to the stichocyte contents).
Most of the important pathology is dueto the larval stages in the muscles. Thesecause myositis, which makes the muscletougher than normal, and there may also becontractures. The extraocular muscles,masseter, muscles of the tongue and larynx,diaphragm, muscles of the neck, intercostalmuscles, deltoid muscles and sites ofattachment to tendons and joints are mostoften involved.
Larvae that invade other organs may beresponsible for more specific pathology, asshown below.
Heart. Larvae often invade the myocardiumbut cardiac muscle cannot form nurse cellsand cysts are never found. At autopsy offatal cases, the myocardium is usually softand flabby and there may be fatty degenera-tion.
Nervous system. Migratory larvae are oftenpresent in the cerebrospinal fluid andmeninges and the brain substance becomesoedematous and hyperaemic. There mayalso be many punctate haemorrhages.Microscopic nodules, with a clear area of
necrosis around each parasite are found inthe subcortical white matter.
Lungs. There may be oedema, disseminatedhaemorrhages and eosinophilic abscessessurrounding larvae, indicative of an aller-gic response.
In general many more migrating larvae aredestroyed in humans than in experimentalanimals and this is accompanied by morepronounced inflammatory changes.
Histopathology of muscle stages. Within 2days of larval invasion, the muscle fibreslose their cross-striations and become morebasophilic. The fibres are changed bio-chemically and become oedematous andswell. About 17 days postinfection, theremains of the muscle fibres become moredense and apparently help to form‘Nevinny’s basophilic halo’ surroundingeach larva. This halo area is actually fluidand contains proliferating, enlarged andaltered nuclei. The redifferentiationchanges in the muscle fibres near eachlarva, with an increase in nuclear numbersand in mitochondria and with enlargementof the Golgi apparatus, are followed by aninflammatory response, with invasion bylymphocytes, polymorphs and histiocytes.The larvae become progressively encapsu-lated and the oedema subsides by the 5thweek.
The outer homogeneous capsule thatsurrounds each larva is derived from thesarcolemma. It is usually translucent and isprobably composed of collagen. The size ofthe cysts formed varies with the host andwith the muscle involved. In humans, thecysts measure on average 0.4 mm �0.26 mm. There is usually only one larvaper cyst, but two may be present and infre-quently even more. The Trichinella larvaein the cyst may live for years but the cystwalls usually calcify a few months or yearsafter infection (Fig. 82).
The cysts in meat are very resistant andthe larvae can probably remain viable untildecomposition occurs (at least 10 days).They have large glycogen reserves and ananaerobic type of respiration.
The Nematodes 179

Trichinella down-regulates the hostimmune response by antigen-dependentmechanisms (molecular mimicry, antigenshedding and renewal, anatomical seclu-sion of the larvae in the cysts and stagespecificity) and by immunomodulation(Bruschi, 1999).
DIAGNOSIS
Clinical. Blood eosinophilia is usually theearliest sign and almost invariably hasappeared by the 10th day of infection; thelevel can be extremely high (90%). It usu-ally continues to rise for 2–5 weeks,remains stable for a further few weeks andslowly falls by the 9th week. The combina-tion of eosinophilia, orbital oedema, mus-cle pain, fever and gastrointestinaldisturbances following ingestion of pork isvery suggestive. None the less, trichiniasisis often misdiagnosed as typhoid fever orinfluenza.
Parasitological. Occasionally adults and lar-vae can be seen in the stools and larvae inthe blood during the 2nd to 4th weeks afterinfection. More certain is the recovery oflarvae by muscle biopsy. A sample of 1 g ofbiceps or gastrocnemius is sufficient forcompression of one half (Fig. 83) and sec-
tioning and staining of the other. The besttime for obtaining a positive biopsy speci-men is after the 4th week – rather late forchemotherapy aimed at killing the adultworms to be of much use. Digestion ofpooled muscle samples is of use in epi-demiological surveys of prevalence in ani-mals or in autopsy material. It is carriedout for 30 min in 1% pepsin in salinecontaining 1% HCl at 37°C and the freedlarvae are examined under the lower powerof the microscope.
Immunological. Because of the difficulties ofparasitological diagnosis, particularly inthe early stages, immunological methodsare more important for individual diag-nosis than in most other helminth infec-tions.
Antibodies can be detected by an ELISAas early as 12 days after infection(Ljungstrom, 1983) and there is a commer-cial kit available, or by Western blotting ofa characterized fraction of sonicated larvaein an immunoglobulin G (IgG) ELISA witha molecular mass of 109 kDa, reacted withspecific antibodies (Chan and Ko, 1990).Recently the use of recombinant antigens(with an excretory/secretory (ES) antigen of53 kDa) and of DNA probes has givenencouraging results in trials.
180 Chapter 5
Fig. 82. Calcified cysts seen in the voluntary muscles of a silver fox.

TREATMENT
Albendazole (at 14 mg kg�1 or 400–800 mgdaily for 3–8 days), albendazole sulphox-ide or mebendazole (200 mg daily for 5days) are effective in removing adultworms from the small intestine but unfor-tunately diagnosis is not usually madebefore the majority of larvae have alreadybeen produced and moved to the muscles(3–5 weeks after infection). Thiabendazole(at 25–45 mg kg�1 for 5–6 days) is equallyeffective but has more side-effects. Pyrantelpamoate at 5 mg day�1 for 5 days has alsobeen used successfully.
Massive doses of corticosteroids (e.g.40–60 mg daily of prednisolone, 200 unitsof adrenocorticotrophic hormone (ACTH)or 25 mg of cortisone, four times daily for4–5 weeks) have been given to relieve theeffects of the allergic response to the muscle phase but are most important forrelieving neurological manifestations.Anthelminthics should always be givenfirst, as corticosteroids could prolong thepresence of adult worms in the intestine.
EPIDEMIOLOGY
Some estimates of prevalences are: Belarus0.06%, Chile 0.26% (1989–1995), Bolivia
3.0% (locally by ELISA: this is a newendemic area), Lithuania 0.02% andPoland 0.03%. Infection is spreading inBelarus, Bulgaria, China (YunnanProvince), Lithuania, Romania, Russia,Ukraine (ten cases year�1 between 1984and 1996), former Yugoslavian countries(there was an outbreak in Bosnia in 1996)and northern Canada. There have beenrecent epidemics in France (there were 538cases in 1993 from a horse imported fromCanada) and Italy from eating raw horse-meat; the horses had probably been fed onanimal products. A recent outbreak inLebanon was caused by eating ‘kubeniye’, alocal uncooked pork dish, at New Year anda source in China is from eating under-cooked pork dumplings. Infection is com-mon in pigs in some countries: 11% inBolivia, up to 2% in Chile, up to 57% inChina, 4% in Costa Rica, up to 22% inCroatia, 4% in Egypt, up to 30% in Mexico,7% in Nigeria and up to 4% in Poland(Murrell and Bruschi, 1994).
While various strains of Trichinella havebeen recognized for many years based oninfectivity to domestic or wild animals(Fig. 84), recent studies based on biologi-cal, biochemical and genetic data of 300
The Nematodes 181
larva
remnant of degenerated muscle fibre
0.1
mm
Fig. 83. Cysts of Trichinella as seen in muscle squash.

geographical isolates indicate that there arefive separate species (Pozio et al., 1992;Kapel et al., 1998), although more conserv-ative authorities (e.g. Bessonov, 1998)believe that there are only the two gener-ally agreed (T. spiralis and T. pseudo-spiralis Garvaki, 1972), while the others aresubspecies of T. spiralis.
Trichinella britovi has a sylvatic cycle inwild carnivores in Europe (Estonia, France,Italy, Netherlands, Poland and Spain),often in national parks with no exposure tohumans. This species (or subspecies) issimilar to T. nativa but is less resistant tofreezing for 3 weeks, differs in distributionand genetic make-up and has less severeintestinal symptoms.
T. nativa is found in holarctic sylvaticanimals (polar bear, Arctic fox, mink, wolf,seal and walrus), but not in mice, rats orpigs. It has a high resistance to freezing. Ithas still not been proved how marine mam-mals become infected; seals may possibly eatinfected amphipod crustaceans that ingestlarval cysts in fragments of meat when feed-ing on dead walruses or polar bears.
T. nelsoni is found only in Africa inwild suids and has no resistance to freez-ing. It is also found in many large wild car-nivores in East Africa (hyena, jackal,leopard and lion), while human infectionis from eating uncooked bushpig or warthog.
T. pseudospiralis is cosmopolitan inwild birds and mammals and has no resis-tance to freezing. This form differs from allothers, as the larvae do not form cysts inthe muscles. It is not common in humansbut can cause muscle pain and fatigue withpolymyositis and a raised creatine kinaselevel (Andrews et al., 1994).
T. spiralis is cosmopolitan, mainly indomestic pigs and sometimes rodents, andis responsible for the great majority ofhuman cases, the symptoms and pathogen-esis of which are described above.
There have been recent outbreaks fromeating wild boar in Croatia (5.7% boarsinfected), Ethiopia, France, Italy, Poland(0.5%), Spain (0.2%), northern Canada andthe USA. In the USA, bear and cougar‘jerky’ are another source of infection inhunters.
PREVENTION AND CONTROL
Larvae in pork are killed by heating above55°C or (for T. spiralis) by deep-freezing at�15°C for 20 days. The number of cases inthe USA has dropped recently (with only38 cases each year for 1991–1996, withthree deaths), chiefly because most pork isdeep-frozen and pig swill is sterilized.However, it is estimated that inspectioncosts $1000 million annually in the USAand $570 million in the European Union(Murrell and Pozio, 2000).
Pork can be examined for infection bymeans of the trichinoscope at the abattoir;this is a projection microscope that projectsan image of cysts in compressed samples ofmuscle (best from the tongue). A pooleddigestion technique with muscle samplesfrom groups of pigs has replaced thetrichinoscope in those countries which stillundertake routine examinations, since it ismore suitable for low prevalences. AnELISA test is good for diagnosing infectionin live animals. It is not possible to recog-nize infected pigs from clinical symptoms,as there is often an absence of morbidity,even with large numbers of cysts.
Most developed countries now havelaws to prevent feeding pigs withuncooked swill (primarily to control vesic-ular erythema in pigs) and this has loweredthe incidence of infection. Rats in piggeriesshould be destroyed and meat from wildcarnivores should never be fed to pigs.
All pork and pork products should bethoroughly cooked, as should the meatfrom wild suids or carnivores.
ZOONOTIC ASPECTS
This is an entirely zoonotic infection,except possibly among the Turkana of EastAfrica, who put dead bodies out to be eatenby wild carnivores.
Rats and other rodents are frequentlyinfected with T. spiralis subspecies, but arenot usually important in transmission;dogs and cats may become infected fromingesting them but do not maintain trans-mission. However, wild carnivores areimportant in transmitting T. nativa and T.nelsoni. T. murrelli has recently beendescribed from wildlife in the USA (Pozio
182 Chapter 5

and La Rosa, 2000), although not so farfrom humans.
In some areas, more than one species (orsubspecies) is present: for instance inEurope, T. spiralis and T. britovi occur inFrance and Spain, T. britovi alone in Italyand Switzerland, T. spiralis alone inBelgium and Germany, T. spiralis, T. britoviand T. nativa in Sweden, T. nativa in
Norway and T. spiralis and T. nativa inFinland (Pozio, 1998).
Experimentally a wide range of mam-mals can be infected, but cysts do not formin Chinese hamsters.
Another non-cyst-forming species, T.papuae, has been recently described frompigs in Papua New Guinea, but not so farfrom humans (Pozio et al., 1999).
The Nematodes 183
BEAR, FOX,WILD CATT ,
MAR YNX,Y
RATATA DOMESTICPIG
HUMAN
HUMAN
HUMAN
WILD BOAR
BUSHPIGWARTHOG
WALRUS, SEAL
HYENA, JACKAL,LEOPARD, LION
POLAR BEAR,R,RARCTIC FOX,
WOLF, MINK, DOG
ARCTIC
AFRICA
TEMPERATE
Fig. 84. Life cycles of Trichinella species from various regions. Top: T. spiralis; centre: T. nelsoni; bottom:T. nativa.

Family Dioctophymidae
Dioctophyma renale (Goeze, 1782) Stiles, 1901
SYNONYMS, DISEASE AND POPULAR NAME
Dioctophyme renale; Eustrongylus gigas(Rudolphi, 1802) Diesing, 1951. Dioc-tophymosis. Giant kidney worm infection.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Dioctophyma is a natural parasite of thepelvis of the kidney or the body cavity ofcarnivores, including the dog, fox, mink,ottter, racoon and rat. About 20 humancases have been reported from Europe,North and South America, Iran, China andThailand.
These are very large nematodes, reddishin colour when alive. The female measures20–100 cm � 5–12 mm and the male14–20 cm � 4–6 mm. The male has asingle spicule set in the middle of a post-erior bursal cup. The unembryonated eggs(measuring 64–68 µm � 40–44 µm) have athick, deeply pitted shell (Fig. 124) and arepassed out in urine. Embryonated eggsmust reach water for further development,where they are ingested by aquaticoligochaetes. These are then ingested byfinal hosts; fish or amphibians may also actas paratenic hosts and this is probably thesource of human infections.
Adult worms in the kidney cause renalcolic and later dysfunction, the worms eat-ing the kidney tissue. Larvae have alsobeen found in subcutaneous nodules.
A related parasite, Eustrongylides ignotus(or E. tubifex), a parasite of fish-eatingbirds and with larvae in oligochaetes andfish has been reported from humans a fewtimes along the Atlantic coast of the USA.Symptoms of acute pain in the right lowerquadrant with vomiting, diarrhoea andfever indicating acute abdomen or appen-dicitis, required surgery. Red-colouredfourth-stage larvae measuring 4–6 cm havebeen removed from a perforated caecumand from the peritoneum. One patient inNew York had eaten raw oysters and goosebarnacles (Wittner et al., 1989) and others
raw fish. In one patient in New Jersey, rawminnows were eaten when being used asfish bait – 48% of the minnows in the areacontained larvae (Eberhard et al., 1989).
Superfamily Mermithoidea
The mermithids are free-living, aphasmidnematodes with larvae that are parasitic ininsects. There have been occasional reportsof recovery of larvae from humans(Agamomermis hominis once in faeces, A.restiformis once in the urethra, Mermisnigrescens once in the mouth, Mermis sp.once in urine), but these are probably spu-rious infections (Hasegawa et al., 1996),with one presumably genuine report of amale mermithid beneath the tunica in acase of hydrocele of the scrotum in aNigerian (Hunt et al., 1984); one can onlyspeculate how this occurred.
Order Spirurida
Superfamily Gnathostomoidea
Gnathostoma spinigerum Owen, 1836
DISEASE AND POPULAR NAMES
Gnathostomiasis or gnathostomosis; acause of visceral larva migrans and ofcreeping eruption (the latter usually due toanother species, G. hispidum).
GEOGRAPHICAL DISTRIBUTION
Most common in Japan (Kyushu, Shikokuand south Honshu) and Thailand(Daengsvang, 1980); occasional cases occurin Bangladesh, Burma, China, Ecuador,India, Indonesia, Malaysia, Mexico, NewGuinea and Vietnam.
LOCATION IN HOST
Adults live in small nodules in the stom-ach wall of carnivores but are rarely foundin humans. Third-stage larvae in humansmigrate through the tissues, usually subcu-taneously, and may be present in any partof the body.
184 Chapter 5

MORPHOLOGY
Adults measure 1.1–3.3 cm and have acharacteristic head bulb with eight rows ofhooks (Fig. 85). The mouth has two largelips. The anterior part of the body is cov-ered with cuticular spines. Third-stage lar-vae migrating in humans measure about4 mm and also have a head bulb with fewerrows of hooks (Fig. 86).
LIFE CYCLE
The eggs in the faeces of carnivores areyellowish-brown and contain an ovum in
the one- or two-celled stage. They measure69 (62–79) µm � 39 (36–42) µm and have afinely granulated surface and a characteris-tic polar thickening (Fig. 124). The eggsdevelop in water and the first-stage larvaehatch in 7 days at 27°C. The larvae areingested by species of cyclopoid copepods(e.g. Cyclops varicans, C. vicinus, C.strenuus, Eucylops agilis, E. serrulatus,Mesocyclops leuckarti, Thermocyclops sp.),which live in ponds, moult in the body cav-ity in 7–10 days (Plate 24), and develop fur-ther when the freshwater microcrustaceans
The Nematodes 185
Fig. 85. Stereoscan of head bulb of adult Gnathostoma hispidum.
Fig. 86. Third-stage larva of G. spinigerum removed from human subcutaneous tissues.

are ingested by a second intermediate host.These are usually freshwater fish, frogs(such as Rana limnocharis) or snakes. Thesecond-stage larvae pierce the gastric walland moult in about 1 month, and the third-stage larvae are eventually surrounded by acyst in the muscles, measuring around1 mm in diameter. The natural definitivehosts are the dog, cat and large carnivores,such as the tiger and leopard, which areinfected by eating fish or by eating aparatenic host (possibly a rodent) that haseaten fish. Suitable paratenic hosts areother fish, amphibians, snakes, rats, mice orchickens, new cysts being formed in these.Twenty-five species of animals from allvertebrate classes can act as paratenic hostsand multiple transfers are also possible.
Prenatal transmission of larvae frommother to offspring can occur in mice andmay account for human cases of gnatho-stomiasis reported from babies.
The adult worms take about 100 days tobecome mature in the stomach of carnivores.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Many patients suffer from epigastric pain,fever, nausea and diarrhoea 24–48 h afteringestion of the larvae, presumably caused bythe larvae penetrating through the intestinalwall. There is often a very high eosinophiliaof up to 90%, with leucocytosis.
Some of the pathology is apparentlycaused by toxic substances secreted byGnathostoma larvae; these include ahaemolytic substance, acetylcholine,hyaluronidase-like substances and a proteo-lytic enzyme.
The larvae migrate through the liver andcause disturbances of liver function. Theythen usually reach the subcutis, moving ata rate of about 1 cm h�1, where they causea painless, migrating oedema. The oedemais generally erythematous and remains forabout 2 weeks in one place (Plate 25). Afew cases of ocular and pulmonarygnathostomiasis have also been reportedand most importantly, an eosinophilicmyelencephalitis, in which the cerebro-spinal fluid is grossly bloody or xantho-chromic. Some fatal cases have presentedwith paraplegia.
Mature adult worms in intestinal nod-ules have also been reported in a few cases.
DIAGNOSIS
Clinical. A painless, migrating, intermittent,subcutaneous oedema is characteristic,particularly when there is an eosinophiliaof over 50%.
Immunological. A purified antigen fromadvanced third-stage larvae (L3s) of 24 kDacan be used in an indirect ELISA, which isstated to be 100% sensitive and specific(Nopparatana et al., 1991). A Westernimmunoblot technique with an antigen of16 kDa has also given good results.
TREATMENT
Surgical excision of worms is often possi-ble when they are in subcutaneous sites.
Albendazole at 400 mg twice daily for10–21 days has been used successfully in afew cases (Kraivichian et al., 1992) and isstated to stimulate larvae to move to thesurface so that they can be easily excised(Suntharasamai et al., 1992). Mebendazoleat 300 mg daily for 5 days was also effec-tive in one case.
EPIDEMIOLOGY AND PREVENTION
Infection in humans usually follows eatingraw fish dishes, such as ‘sashimi’ in Japan,‘somfak’ in Thailand or ‘ceviche’ (if madewith freshwater fish) in Latin America.Important species of food fish in Japan arecrucian carp, Ophiocephalus spp. or goby(Acanthogobius hasta); chicken can also bea source of infection to humans. Theencysted larvae in fish are killed by cook-ing for 5 min in water at 70°C or by pick-ling in vinegar for 6 h. Skin penetration ispossible experimentally in cats and mightoccur during preparation of fish dishes.
Prevention is by thorough cooking offish and chicken.
There are at least seven other species ofGnathostoma that could infect humans. G.doloresi Tubangui, 1925, and G. hispidumFedchenko, 1872, are parasites of pigs inAsia and there have been a few cases of
186 Chapter 5

human infection in China and Japan fol-lowing ingestion of raw freshwater fish orsnakes, usually causing creeping eruption,sometimes accompanied by stomach con-vulsions (Akahane et al., 1998). In one caseG. doloresi caused colonic ileus, due toeosinophilic nodular lesions. G. nippon-icum Yamaguti, 1941, has been recentlydescribed from humans in Japan, where ithas caused itching and creeping eruptionafter eating freshwater fish. (Ando, 1989;Ando et al., 1991). There has been a ‘rash’of cases of creeping eruption in Japanrecently after eating raw marine fish orsquid, with one patient having larvae inthe eye (‘type x’ spirurid larvae).
Larvae of the different species can bedifferentiated, although this is likely to beimpossible in tissue sections (Ando et al.,1991).
Other Spirurids
Thelazia callipaeda Railliet and Henry,1910, is a parasite of the conjunctival sacand lachrymal ducts of dogs in the FarEast. It has been recorded about 60 timesfrom humans in China, Indonesia, Japan,Korea, Thailand and Russia (Siberia). Theadults of both sexes are filariform and havea serrated cuticle. The female measures7–19 mm, the male 4–13 mm. The move-ment of the adults under the eyelids in theconjunctival sac causes intense irritationand the scratching of the cornea can lead toscar formation and opacity. Worms can beremoved after local anaesthesia.
The intermediate hosts are dipteranflies, including Musca domestica (Shi etal., 1988).
Thelazia californiensis Price, 1930, an eyeparasite of ruminants, particularly deer,has been reported five times from humansin the USA, causing keratoconjunctivitis,and is also transmitted by flies.
Gongylonema pulchrum Molin, 1857 (thegullet worm), is a natural parasite of theoesophagus of herbivores (sheep, cattle,goats, camels, equines, pigs and deer). Ithas been reported about 40 times from
humans in Austria, Bulgaria, China,Hungary, Italy, Morocco, New Zealand,Russia, Serbia, Sri Lanka and the USA.Adult females measure about 145 mm �0.5 mm, males 63 mm � 0.3 mm and theanterior end of both sexes is covered withcuticular thickenings. The worms live inserpentiginous tunnels in the squamousepithelium of the oesophagus and oral cav-ity, where they can cause painful abscessesof the buccal mucosa (Jelinek and Loscher,1994), sometime accompanied by pharyn-gitis, vomiting and irritability. Eggs pro-duced in these tunnels reach the mouthcavity and are passed out fully enbry-onated in the faeces. The eggs from naturalhosts containing first-stage larvae areingested by dung-beetles or, experimentallyat least, cockroaches, which act as interme-diate hosts. Adults take about 80 days todevelop when third-stage larvae areingested inside these insects. Diagnosis isby identifying the characteristic tunnelsand by extracting adult worms after givinganti-inflammatory drugs (Jelinek andLoscher, 1994): eggs have been seen in spu-tum but not in faeces.
Abbreviata (= Physaloptera) caucasica(von Linstow, 1902), is a natural parasite ofmonkeys in the tropics. The adult femalesmeasure 20–100 mm � 2.5 mm, males14–34 mm � 1.0 mm, and both live withtheir heads embedded in the mucosa of thestomach or intestinal wall. Cockroachesand grasshoppers can act as experimentalintermediate hosts. It has been reportedfrom humans in Brazil, Colombia, CongoRepublic (Zaire), India, Indonesia, Israel,Namibia, Panama, Zambia and Zimbabwe.In a recent case in an Indonesian woman,adult worms were recovered from the bil-iary duct, where they had caused biliarypain, jaundice and fever. Another species,Physaloptera transfuga Marits andGrinberg, 1970, a parasite of cats and dogs,has been recovered once from the lip of awoman in Moldova.
Cheilospirura sp. Diesing, 1861 (orAcuaria sp.), a parasite of the gizzard ofbirds with insect intermediate hosts, hasbeen recovered once from a nodule on theconjunctiva of a Philippine farmer.
The Nematodes 187

Spirocerca lupi (Rudolphi, 1809) is a com-mon parasite of dogs and other carnivoresin warm countries, forming tumours in thestomach wall, with dung-beetles as inter-mediate hosts and birds or mammals asparatenic hosts. It has been found onceembedded in the wall of the ileum of anewborn baby in Italy; it is assumed thatinfection was prenatal.
Rictularia spp. Froelich, 1802, are parasitesof rodents or bats, and a gravid female wormhas been recovered once from a section of anappendix of a man in New York.
Superfamily Filarioidea: the Filariae
Members of the superfamily Filarioidea ofthe order Spirurida are tissue-dwellingnematodes, responsible for some of the
most important helminth diseases ofhumans. Bancroftian filariasis (caused byinfection with Wuchereria bancrofti) andonchocerciasis (due to Onchocerca volvu-lus) in particular are the cause of muchmorbidity and the former shares withschistosomiasis the dubious distinctionamong helminth infections of increasing inincidence each year. All human filariae aretransmitted by the bite of a bloodsuckinginsect (Fig. 87). The adults are ovovivi-parous and the early first-stage larvae,known as microfilariae, are found in theblood or skin. Microfilariae measure from150 to 350 µm in length and some retainthe much expanded eggshell as a thinsheath; their specific identification can beof great importance in diagnosis (Figs 88and 89).
188 Chapter 5
Fig. 87. The life cycles of human filariae.
INSECT MAN
2nd-STAGE LARVAE
3rd-STAGELARVAE
1st-STAGELARVAE
Injected on to skinnext to bite
Develop in fat body:LoaDevelop in thoracicmuscles:all others
MICROFILARIAESkin tissue fluids:OnchocercaM. streptocercaBlood:all others
ADULTSLymphatics:WuchereriaBrugiaSubcutaneoustissues:LoaOnchocercaMansonellastreptocercaPeritoneal orpleural cavity:Mansonella ozzardiM. perstans
Fig. 88. A generalized microfilaria showing the features of use in identification (some features can beseen only with special staining techniques and the stylet and sheath are not always present).
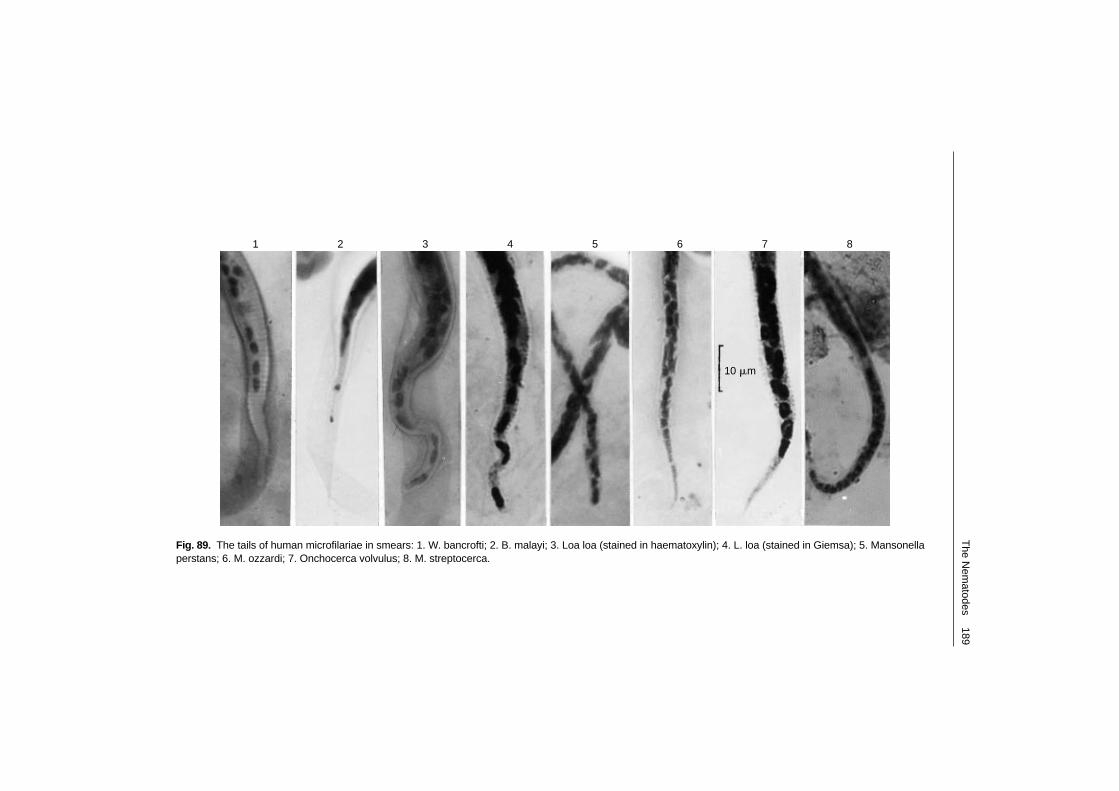
The N
ematodes
189
1 2 3 4 5 6 7 8
10 �m
Fig. 89. The tails of human microfilariae in smears: 1. W. bancrofti; 2. B. malayi; 3. Loa loa (stained in haematoxylin); 4. L. loa (stained in Giemsa); 5. Mansonellaperstans; 6. M. ozzardi; 7. Onchocerca volvulus; 8. M. streptocerca.

190 Chapter 5
Wuchereria bancrofti (Cobbold, 1877) Seurat, 1921
SYNONYMS
Filaria sanguinis hominis; Filaria bancroftiCobbold, 1877; Wuchereria pacifica.
DISEASE
Lymphatic filariasis or filariosis (LF);Bancroftian filariasis or filariosis; cause ofelephantiasis.
LOCAL NAMES
Jakute (Burmese), Etow = hydrocele andBarawa gyeprim = elephantiasis (Fanti),Wagaga or Wagana (filarial fever) (Fijian),Tundermi (Gwaina = hydrocele) (Hausa),Kusafurui (filarial fever) or Kusa in Kyushuand Ryuku Island (Henki, Senki orOogintama = hydrocele) (Japanese), Apipa =elephantiasis (Luo), Mumu (filarial fever)(Samoan), Watharoga = hydrocele(Singalese), Sheelo = hydrocele and Lugomarodi = elephantiasis (Somali), Matende(Mshipa = hydrocele) (Swahili), Fee fee andMariri (Tahitian), Yanaikal (Tamil), Gypeym(Etwo = hydrocele) (Twi).
GEOGRAPHICAL DISTRIBUTION
It has been estimated recently that 119 mil-lion people are infected with LF, of whom107 million are due to W. bancrofti.
Infection is present in most equatorialcountries of tropical Africa between 20°Nand 20°S and in Madagascar, Asia(Bangladesh, China, India, Indonesia,Japan, Korea, Myanmar, Pakistan,Philippines, Sri Lanka, Thailand), thePacific (New Guinea, Melanesia, Polynesia,including Fiji, Samoa, Tahiti), the WestIndies and northern South America.
LOCATION IN HOST
The adults live in the lymphatics, either ofthe lower limbs and groin region or of theupper limbs, and breasts in women.Sheathed microfilariae are found in thebloodstream.
MORPHOLOGY
Like all filariae of humans the adults ofWuchereria are long, threadlike worms
(hence the name ‘filaria’ from Latin). Thehead end is slightly swollen and has tworows of ten sessile papillae. The cuticle issmooth, with very fine transverse striations,and the posterior end of the female is finelytuberculated. The female measures80–100 mm � 0.25 mm, the male 40 mm �0.1 mm. The ovoviviparous female has twoposterior ovaries; from these two oviductslead into the paired uteri, which coil backand forth through the greater part of the bodybefore they fuse and open by a short vagina atthe vulva, 0.8 mm from the anterior end. Theproximal portions of the uteri are filled witheggs and embryos, while near the vulva themicrofilariae are crowded and remain in thesheath formed by the elongation of the eggmembrane. Cross-sections of females of allhuman filariae show this double-uterinestructure. The male has the posterior endcurved ventrally and on the ventral surface ofthe tail are 12–15 pairs of sessile papillae.There are two unequal spicules, measuring0.6 and 0.2 mm, and a gubernaculum (thenumber and arrangement of the papillae andthe structure and lengths of the spicules ofthe male worms are of great taxonomicimportance among the filariae).
The microfilariae are found in thebloodstream and can live for 3–6 months;they measure 210–320 µm by 7.5–10 µm(the mean lengths of microfilariae from var-ious geographical regions vary from 250 to290 µm). The anterior end is rounded andthe posterior end pointed. Each micro-filaria is surrounded by a long sheath,which stains pale pink in Giemsa stain. Inwet preparations the microfilaria movesvigorously in a serpentine manner; instained preparations it often assumesgraceful curves. The posterior end is free ofnuclei and thus differs from the microfilar-ial larvae of Brugia malayi (Fig. 90) andLoa loa.
The microfilariae have various struc-tures (nerve ring, excretory pore and cell,inner body or innenkörper, rectal cells andanus) whose size and position can be ofhelp in recognition. However, it must beremembered that these features are only ofuse in well-fixed and stained preparationsand some individual variation may occur.

The N
ematodes
191
Map 8. Distribution of Wuchereria bancrofti.

192 Chapter 5
Microfilariae described from easternMadagascar (Malagasy Republic) had somecharacters resembling W. bancrofti and oth-ers resembling those of Brugia malayi andwere designated as belonging to W. ban-crofti var. vauceli.
LIFE CYCLE
The sheathed microfilariae circulate in thebloodstream and, in most parts of the world,show a marked periodicity; they are foundin the peripheral circulation from 22.00 to02.00 h, while during the day they hide inthe capillaries of the lungs. In areas of thePacific east of longitude 160°, such as Fiji,Samoa and Tahiti, where the vectors are day-biting mosquitoes, the periodicity is partiallyreversed and the strain is said to be diur-nally subperiodic. Investigations on themechanism of microfilarial periodicity haveshown that during the day, in the periodicform, the microfilariae are held up in the
small blood-vessels of the lungs principallybecause of the difference in oxygen tensionbetween the arterial and venous capillariesin the lungs. Perhaps the peripheral circula-tion represents an unfavourable environ-ment and the microfilariae remain there forthe minimum time necessary to maintaintransmission.
When the microfilariae are ingested witha blood meal by a suitable species of mos-quito, they lose their sheath within 15–30min in the stomach of the insect. A propor-tion manage to penetrate the stomach wallbefore the formation of a peritrophic mem-brane and migrate to the thoracic musclesin 1–24 h. Two days later they have meta-morphosed into sausage-shaped larvae,measuring 150 µm � 10 µm, and by the endof a week (usually at 5 days) the larvae havemoulted, the alimentary canal is developedand the second-stage larvae then measure250 µm � 25 µm. During the 2nd week
Fig. 90. Microfilariae of (top) Wuchereria bancrofti and (bottom) Brugia malayi.

The Nematodes 193
(9–10 days) the larvae moult again, growmuch longer (1.2–1.8 mm) and migrate tothe head, where they enter the labium andemerge through the tips of the labella whilethe mosquito is feeding. The infective third-stage larvae can be differentiated from otherfilarial larvae, if the mosquito is dissectedin a drop of saline, by size and by the threesubterminal papillae (Fig. 91) (there may belarvae of various animal filariae in mosqui-toes, such as Dirofilaria and Setaria). Theparasite cannot be transmitted from oneperson to another until the larvae haveundergone this essential development tothe infective stage in the mosquito.
Optimal conditions for filarial develop-ment in mosquitoes are about 26°C (range16.5–31°C) and 90% relative humidity. Manymosquitoes are killed by heavy infectionsand in others larvae often fail to mature, sothat the infection rate in mosquitoes is usu-ally low (below 0.5%), though under someconditions it can be much higher.
Development in the mosquito takes 11 ormore days and the infective larvae escapethrough the labella when the insect bites anew individual and enter the skin throughthe puncture wound (this is likely to bemore successful in areas of high humidity,where the skin will be moist, although themosquito does deposit a drop of fluid on theskin while feeding). After entering the skinof a human, the larvae migrate to the lymphvessels and glands, where they moult twice
more and mature and the females producemicrofilariae within 1 year (206–285 days inexperimental infections in leaf monkeys).The adult female worms probably producemicrofilariae for about 5 years (Vanamail etal., 1996), although greater longevity hasbeen reported in the past (10–17 years).
CLINICAL MANIFESTATIONS
It has been estimated that currently about44 million people suffer from overt mani-festations of lymphatic filariasis. The other76 million cases appear to be symptomless,a condition that may last for years, and theonly overt evidence of infection is the pres-ence of microfilariae in the blood.However, recent studies using lym-phoscintigraphy indicate that even theseapparently symptomless cases are probablysuffering from subclinical lymphatic andrenal damage (Freedman et al., 1994;Noroes et al., 1996).
Acute disease. From the 3rd month afterentry of the larvae, there may be recurrentattacks of acute lymphangitis (inflamma-tion of the lymph ducts), with inflamed,tender lymph nodes, headache, nausea andsometimes urticaria (‘filarial fever’), eachattack lasting from 3 to 15 days. At thistime microfilariae vanish from the blood(amicrofilaraemia) and there may be manyattacks in a year. There is ofteneosinophilia accompanied by chyluria (the
10 �m
Wuchereria bancrofti Brugia malayi
Fig. 91. Tails of third-stage larvae of W. bancrofti and B. malayi from mouth-parts of mosquito.

194 Chapter 5
presence of whitish lymph containing fattyacids from the intestinal lymph vessels inthe urine). In many areas acute symptomsstart at the age of about 6, with a peak atage 25. In many areas infection probablyoccurs in very young children. In a studyin Haiti, 6% of 2-year-olds and 30% of 4-year-olds had filarial antigens but only onechild was microfilaraemic; some of thesechildren already had dilated lymphatics,shown by ultrasound (Lammle et al., 1998).The most serious clinical manifestationsare due to disturbances of the lymphaticsystem (Fig 92), which may result inlymphadenitis (inflammation of the lymph
nodes), lymphoedema, lymphoceles, lym-phuria, chyluria and, in males, genitallesions, including hydrocele and lymphscrotum, in which lymph drips from thespermatic cord. In Ghana, hydrocele butnot adenolymphangitis was associatedwith a high infection prevalence and inten-sity (Gyapong, 1998).
Chronic disease. True elephantiasis, inwhich the oedema is hard and non-pitting,takes about 10–15 years to develop.Secondary bacterial and fungal infectionoften follows, with the develop of verru-cous growths (Fig. 93).
Fig. 92. Radiograph showing blocked lymph ducts.
Fig. 93. Cases of elephantiasis in Guyana caused by W. bancrofti. It is not so evident in males becauseof the trousers worn. In the man third from the right there are verrucous lesions of the foot (‘mossy foot’).

The Nematodes 195
The changes in the lymphatics are oftenstated to be due to blockage caused by thedead worms but recent experimental stud-ies indicate that it is caused mainly by anallergic response to the still-living worms.
The nature of the lesions varies in dif-ferent parts of the world, reflecting particu-larly the different sites of obstruction. InIndia, chyluria is rarely reported, but it wascommon in Japan. In Africa, elephantiasisis less common than in the Pacific and isusually confined to the legs. Hydrocele(Figs. 94 and 95), with probable reductionin fertility, is particularly common inmales in East Africa (up to 50% of infectedindividuals) and India (with about 27 mil-lion cases) and has very serious social/sex-ual implications (Dreyer et al., 1997). InWest Africa, there are often high micro-filarial rates (40–50% of the population)but few clinical manifestations. In thosePacific islands where the subperiodicstrain of the parasite occurs, slowly devel-oping gross elephantiasis of the arms, legs,breasts, scrotum or vulva was commonuntil transmission was reduced in the lastfew years.
Tropical pulmonary eosinophilia iscaused by an abnormal host reaction to the
presence of a lymphatic filarial infectionand is sometimes known as occult filaria-sis. The symptoms include bouts of severe,paroxysmal, dry, wheezy coughing, resem-bling an asthma attack, with dyspnoea,
Fig. 94. A case of filarial hydrocele.
Fig. 95. Section of dilated lymphatic of the testis with adult female of W. bancrofti. The vessel wall isthickened and there is a granulomatous reaction to the worm (magnification � 55).

196 Chapter 5
chest pain and fever, resulting from denseeosinophilic infiltrations in the lungs last-ing for weeks or months. It occurs particu-larly in India or Singapore (in persons ofIndian extraction). Patients are amicrofila-raemic but have strong filarial serology,with a high eosinophilia (about 35%).
A similar reaction is known asMeyers–Kouwenaar’s syndrome, found inthe Pacific and East Indies. There is a benignlymphadenitis with enlargement of thelymph nodes, liver and spleen and with pul-monary symptoms but no microfilaraemia.Histologically the lymph nodes are hyper-plastic with dead microfilariae surroundedby eosinophils and then hyaline material toform nodular, granulomatous, abscesses(‘M–K bodies’). The condition can easily bemistaken for Hodgkin’s disease and thesymptoms of both filarial conditions arediagnostically rapidly alleviated by treat-ment with diethylcarbamazine (DEC).
PATHOGENESIS
The first phase of clinical disease is charac-terized by dilatation of the afferent lym-phatic vessels reacting to the presence ofmasses of adult worms. Initially thesinuses of the smaller lymph vesselsincrease in size (lymphangiectasia), but fol-lowing the repeated attacks of adenolym-phangitis the enlarged lymph nodesbecome progressively firm and fibrotic. Thenodes become hyperplastic and micro-scopical sections show inflammation of theendothelial lining, with many lympho-cytes, plasma cells and particularlyeosinophils, and there may be foci ofnecrosis, with sections of adults in thelumen. Lymphatics often become invadedby bacteria, but some clinical (Dissanayakeet al., 1995) and experimental (Denhamand Fletcher, 1987; Lawrence, 1996)studies indicate that this is not the primarycause of pathogenesis, which is caused byan allergic reaction to the adult worms,allied to cofactors such as a high wormburden, lymphangiectasia, location of adultworm nests (e.g. in spermatic cord) andchronology of adult worm deaths. How-ever, recurrent secondary bacterial infec-tions superimposed on the damage to the
lymphatics probably play an importantpart in the development of lymphoedemaand elephantiasis (Dreyer et al., 2000). Inchronic infections, lymphoid aggregatesand germinal centres develop and eventu-ally cause complete occlusion of thelumen, so that the lymph vessels are nolonger functional. Worms die and becomesurrounded by fibrotic tissue, which even-tually calcifies. It is not known what trig-gers the initial inflammatory response.Chyluria is caused by rupture of lymphvarices into any part of the urinary tract,with fatty lymph appearing in the urine. Incases of renal chyluria there may be pas-sage between the renal pelvis and a dilatedlymphatic to the cisterna chyli. Such casesshow up on intravenous or retrograde pyel-ograms but are not serious and do notrequire surgery. The rupture of varicoselymph vessels near the skin of the scrotumresults in lymph oozing on to the surface(lymph scrotum).
Adults of Wuchereria and Brugia con-tain a bacterium, Wohlbachia, in the lateralcords and it has recently been postulatedthat these bacteria are responsible forinflammatory responses, inducingcytokines (Fig. 96). The chronic release ofWohlbachia might lead to desensitizationof the innate immune response and thebacterial catalase may protect the filariaefrom the action of hydrogen peroxide-mediated damage (Taylor, 2000). The pres-ence of the bacterium appears to beessential for the long-term survival of thefilaria.
Immune responses. Many of the symptomsassociated with filarial infection (e.g.eosinophilia, urticaria, lymphangitis andlymphoedema) are immunological in originand confirm that infection induces strongimmune responses. Epidemiological stud-ies show that, in endemic areas, there areindividuals who appear to be parasitologi-cally negative, despite evidence of expo-sure to infection (the so-called endemicnormals). A key question is whetherimmune responses have the potential to behost-protective as well as pathogenic.Analysis of immune responses has been

carried out in many infected populationsand there has been a large amount of labo-ratory research with experimental models.However, it has to be said that no clear pic-ture has emerged.
Immunological relationships betweenfilarial nematodes and their hosts are com-plex and the cause of much contention.Particular difficulties are caused by thefact that in filarial-infected populationssome individuals apear to be immune, oth-ers suffer immune-infected pathology,while yet others (microfilaraemic individ-uals) show a depression of responses tofilarial antigens that is reversed afterchemotherapy. T cells appear to be impor-
tant in all of these phenomena and therehave been many attempts to explain eachin terms of differential T-helper 1 (Th1)and Th2 responses. However, there is nostraightforward relationship between sub-set activity and host response. It is truethat patients with high microfilarial loadstend to show reduced Th1 responses andhigh Th2 (e.g. reflected in high IgG4 levels,possibly acting as blocking antibody), butnot all type 2 cytokines are produced nor-mally. T cells from microfilaraemic indi-viduals show much reduced proliferationto worm antigens, but can still releaseinterleukin 4 (IL-4), though IL-5 responsesare reduced. The cytokine IL-10 may be
The Nematodes 197
Fig. 96. Section of W. bancrofti with the bacterium Wohlbachia in the lateral cords (the numerous bodieson right side of electron micrograph).

involved in down-regulating proliferativeresponses, although antigen overload fromthe large numbers of circulating micro-filariae may also be involved. Filarialworms are also thought to releaseimmunomodulatory molecules that depressspecific T-cell functions, such as prolifera-tion, gamma interferon (IFN-�) secretionand, surprisingly, IL-5 production.Children born to mothers who are microfi-laraemic may show a profound T-cellunresponsiveness, both Th1 and Th2responses being down-regulated, whichpresumably results from placental transferof antigen or immunomodulatory factors.
Immune-mediated protection againstinfection may act independently againstthe three major life-cycle stages (infectiveL3s, adults and microfilariae). Individualswho remain uninfected in endemic areastend to show Th1-biased responses, andmice lacking the capacity to make IL-4 (amajor Th2 cytokine) retain the ability tokill incoming infective larvae of B. malayi.However, neither Th1 nor Th2 responsesappear to eliminate adult B. malayi fromthe body cavity. It is clear that the differentstages also induce quite different patternsof host responses. Thus, in mice, adultworms and L3s elicit Th2 responses withhigh IL-4 and IgG4 production, as well asIgG1 and IgE antibodies (although IgEresponses are depressed), whereas expo-sure to microfilariae alone can induce Th1responses, with high initial IFN-� produc-tion and IgG2 antibody. How worms arekilled is still controversial, despite clearevidence from in vitro studies that anti-body and antibody-dependent cellularcytotoxic (ADCC) mechanisms can damageor eliminate parasites. Eosinophils partici-pate in ADCC mechanisms against infec-tive L3s, and IgM and IgG2a antibody canclear microfilariae. It is likely that nitricoxide mechanisms are important in attacksagainst larvae (Maizels et al., in Nutman,2000). People probably become resistantfollowing repeated reinfection and, as inschistosomiasis, may develop a state ofconcomitant immunity, with existing adultworms remaining alive but invading larvaebeing destroyed.
DIAGNOSIS
Parasitological. Routine diagnosis is byfinding microfilariae, usually in stainedthick blood films (see p. 262 for varioustechniques). Blood should be taken fromthe ear or finger at 22.00–02.00 h for thenocturnally periodic forms and at10.00–14.00 h for the subperiodic Pacificform. Microfilariae can sometimes beobtained during the day by taking bloodsmears about 1 h after giving two tablets(100 mg) of diethylcarbamazine (DEC)(provocation test). Concentration tech-niques can be employed (counting chamberor Nuclepore filter (p. 264)).
Clinical. Lymphangitis, lymphadenitis ormanifestations of lymph stasis in a patientin an endemic area, often witheosinophilia, are very suggestive. Recentstudies, particularly in Brazil, using ultra-sound have enabled direct visualization ofadult worms in the lymphatics (Amaral etal., 1994; Noroes et al., 1996). It has beenfound that the worms are in ceaselessmovement, undergoing what has beentermed a ‘filarial dance’.
Immunological. These tests are most suc-cessful in cases without microfilariae in theblood. A new simple immunochromato-graphic test (ICT) card kit is commerciallyavailable, using 100 µm of whole-bloodfinger-prick samples (or serum or plasma),which can be taken at any time byuntrained staff and put on a nitrocelluloselayer on a sample pad (Weil et al., 1997).The pink sample pad contains a driedpolyclonal antibody (AD12.1) specific to W.bancrofti attached to colloidal gold andmigrates up the pad to reach a specificmonoclonal antibody immobilized in a lineon the pad; this traps free antibody–antigencomplexes to form an intense pink line,which can be compared with a control line.The test takes 5–15 min and detects earlyinfections in children, which is veryimportant for assessing the success of anycontrol campaign, as well as for individualdiagnosis, and is negative for all filarialinfection apart from Bancroftian filariasis
198 Chapter 5

(this kit is commercially available fromwww.amrad.com.au or [email protected]). A commercially available dotELISA with monoclonal antibodies (usu-ally Og4C3), using whole dried blood sam-ples, can even give an indication of thenumber of parasites present but is not easyto use in the field (from JCU TropicalBiotechnology, Townsville, Queensland).Circulating parasite DNA detection by aPCR is both sensitive and specific, particu-larly when microfilariae are present, but ishighly labour-intensive and expensive.
TREATMENT
Chemotherapy. Recent ultrasonic studieshave enabled the action of chemotherapeu-tic agents to be directly observed and wormnests can be surgically removed and wormsexamined to ascertain drug-inducedchanges in somatic tissues or in the abilityto produce new larvae (embryogenesis)(Dreyer et al., 1995). Previously the effectson adult worms required studies overmany years.
DEC has been used for the last 50 years,usually at an oral dose of 72 g over 3 weeks,and was thought to kill microfilariae butnot adults. Recent studies indicate that itdoes kill about 50% of adult worms as well.Recently, much smaller doses (6 mg kg�1)have been found to be effective. However, itcannot be used in patients from Africa (seep. 201). In vitro DEC has no effect on micro-filariae and its action may be mediated byplatelets; excreted filarial antigens are alsonecessary to trigger the reaction.
Ivermectin at an oral dose of 400 µgkg�1 every fortnight for 6 months (totaldose of 4.8 mg kg�1) completely inhibitsproduction of microfilariae but has noeffect on adults when examined by ultra-sonography, even at the end of long-termtreatment. For mass chemotherapy, 200 µgkg�1 once yearly has been advocated. Itprobably acts by binding to glutamate-gated chloride channels.
Albendazole in a single oral dose of 400mg can be given with DEC or ivermectinand appears to have some action againstadults (possibly synergistic).
Long-term treatment with antibiotics,such as rifampicin or oxytetracycline, isshowing promise in research and clinicaltrials (Townson et al., 2000).
Surgical. Traditionally hydrocele has beentreated by adding sclerosing agents andlymphoedema by bandaging and elevationof limbs. Radical surgery has also been usedto remove adventitious tissue in elephantia-sis. Recently, regular washing and hygieniccare of inflamed lymph vessels and hydro-celes has been shown to resolve manifesta-tions and prevents further development.Following 5 years of chemotherapy andsimple medical treatment in New Guinea,87% of patients were cured of hydroceleand in 69% lymphoedema had resolved.
EPIDEMIOLOGY
Of the 107 million cases of Bancroftianfilariasis in the world, 49% are in South-East Asia (particularly India, with an esti-mated 48 million infections), 31.3.% inAfrica, 16% in the western Pacific, 3.4% inthe Americas and 0.3% (or now probablynil) in the eastern Mediterranean; alto-gether it occurs in over 80 countries, where750 million people are at risk. The geo-graphical distribution of the disease isdetermined largely by climate, being foundin areas with a hot, humid climate; thelocal distribution can be forecast by geo-graphical information systems (GIS) imag-ing (Brooker and Michael, 2000) and by thedistribution of the vectors, but other factorsmay be involved, such as local variationsin the density, susceptibility, longevity andhuman-biting habits of the vector and inthe density, sleeping habits and naturaland acquired resistance of the human pop-ulation.
Many species of mosquitoes have beenshown to act as vectors in various parts ofthe world (WHO, 1992; Kettle, 1995;Service, 2001), including at least 31 speciesof Anopheles (An. aquasalis, bellator, dar-lingi in the Americas; An. funestus, gam-biae, arabiensis, bwambae, melas, merus,nili in Africa; An. aconitus, anthropopha-gus, kewiyangensis, nigerrimus, sinensiscomplex, letifer, whartoni, flavirostrus,
The Nematodes 199

minimus, candidiensis, balabacensis, leu-cosphyrus, maculatus, philippinensis, sub-pictus, tesselatus, vagus in Asia; An.bancrofti, farauti, koliensis, punctulatus, inPapua New Guinea), five species of Culex(C. quinquefasciatus in the Americas,Africa and Asia; C. pipiens molestus in theMiddle East; C. bitaeniorhynchus, sitienscomplex, pipiens pallens in Asia; C. pipi-ens pallens in the western Pacific; C.annulirostris, bitaeniorynchus in Papua) 15species of Aedes (Ae. scapularis in theAmericas; Ae. niveus group, harinasutai,togoi, poicilius in Asia; Ae. togoi in thewestern Pacific; Ae. fijiensis, oceanicus,samoanus, vigilax, futunae, polynesiensis,pseudoscutellaris, tabu, tongae, upolensisin the South Pacific) and two species ofMansonia (M. titillans in the Americas; M.uniformis in Asia and Papua).
Vector Anopheles species breed inponds, slow-flowing streams, rice paddiesand fresh- or salty-water swamps, and LFtransmitted by them is principally a ruraldisease. Members of the An. gambiaegroup (An. gambiae, arabiensis, melas,merus) are very susceptible vectors andshow facilitation, whereby the penetrationof one larva through the stomach wall facil-itates the entry of others; this can result inhigh mortality of mosquitoes when microfi-larial rates are high (Dye, 1992). In ruralareas of East Africa, infection rates of1–1.5% are found in An. gambiae, An. ara-biensis and An. funestus, but 0–0.3% in C.quinquefasciatus (although this is the onlyvector in urban areas). In West Africa,transmission is entirely due to Anopheles;although C. quinquefasciatus is common inurban areas, it cannot be infected.
Culex quinquefasciatus is the mostimportant vector in urban areas of Asia andEast Africa, as it can breed in very smallbodies of water contaminated with organicmatter (especially leaking septic tanks, pitlatrines and drains). The adult femalesreadily enter houses, tend to rest indoorsand are night biters; they can sometimes flyfor up to 1 km. They are almost entirelyanthropophilic and in Singapore infectionrates of 20% occur. Culex species show thephenomenon of limitation, whereby pene-
tration of one larva inhibits the passage ofothers, so that only a few larvae are usuallyfound in each mosquito (it also has a pha-ryngeal armature, which destroys manymicrofilariae). However, there may be manymosquitoes biting each night (over 100 pernight is common) and in Calcutta it hasbeen estimated that one person may be bit-ten by 6000–7000 infected C. quinquefas-ciatus per year. This species is extending itsrange alarmingly in both Asia and Africa.
On Polynesian islands (east of longitude160°), such as Fiji, Samoa and Tahiti, adiurnally periodic form of LF occurs andthe vectors are species of Aedes, which canbreed in small bodies of water, such asopened coconut shells, leaf bracts, crabholes and manufactured containers, suchas pots, tin cans and car tyres. In Thailandthere is a nocturnally periodic form of LFtransmitted by Ae. niveus.
To determine transmission rates in mos-quitoes they can be dissected individually(Davies, 1995) or, where rates are very low,a pool method using 1000–2000 (possiblydried) mosquitoes can be employed andthe presence of larvae shown by DNAdetermination after PCR amplification.
PREVENTION AND CONTROL
Lymphatic filariasis was identified recentlyas one of six potentially eradicable diseasesby an International Task Force (1993), andthe World Health Organization (WHO) iscoordinating campaigns to eliminate it as apublic health problem in the next fewyears (Ottesen et al., 1997; WHO, 1999; seealso WHO web site: www.filariasis.org).The reason for optimism is partly becauseof new regimes of chemotherapy and fromthe success of campaigns already carriedout in various countries. It is possible thatthere is a window of opportunity such asin retrospect can be seen to have con-tributed to the success of the smallpoxcampaign: just after its eradication massvaccination would not have been possiblebecause of the spread of AIDS.
Control is aimed at both alleviating thesuffering caused by the disease (morbiditycontrol) and stopping the spread of infec-tion (transmission control).
200 Chapter 5

Morbidity control. Mass treatment cam-paigns with DEC have been tried withvarying degrees of success over the last 40years. China, where compliance is good, isnow in the final stages of a long-term cam-paign: in 1950 there were estimated to be30 million cases, while there were onlyabout 1.65 million in 1995. In China andother countries, such as Samoa, SolomonIslands and Tahiti and particularly in theRyuku Islands (Japan), South Korea andTaiwan, DEC has been added to cookingsalt (at a dose of about 3 mg kg�1 ) and hasgreatly reduced disease. In the past it wasthought that DEC only acted against micro-filariae (at a total dose of 72 g usuallygiven over 12 days), but recently studieshave indicated that a single annual dose iseffective and also has some action againstadults. If microfilarial rates fall below acritical breakpoint, then infection shoulddie out (this level probably differs in dif-ferent areas and with different popula-tions).
The real breakthrough in mass treatmentwas the recent finding that combined treat-ment schedules are much more effectivethan the use of a single drug (Ottesen et al.,1999; Plaisier et al., 2000). In the newWHO initiative two main regimes are rec-ommended:
1. All areas except sub-Saharan Africa:(a) DEC (6 mg kg�1 body weight) combinedwith albendazole (400 mg) once yearly for4–6 years is the preferred regime;(b) or DEC (6 mg kg�1) once a year for 4–6years alone (this has fewer side-effects thanthe traditional 12-day treatment;(c) or DEC in salt (0.2–0.4% w/w) for 6–12months.2. Sub-Saharan Africa: DEC cannot be given because there mayalso be infections with loiasis or oncho-cerciasis and this drug can then be danger-ous (see p. 210) or very unpleasant (seep. 218).(a) Single yearly dose of ivermectin (200µm kg�1) plus albendazole (400 mg) for4–6 years (optimal);(b) or ivermectin alone at same dose.
Campaigns using these schedules havestarted in American Samoa and westernSamoa and Niue (25 countries by August2001). It is intended that 50 million indi-viduals will have been treated at least onceby the end of 2001 and that the disease willbe eliminated as a problem in any countryby 2020. There is a concerted campaignstarting in all Pacific islands that hopes toeliminate infection there by 2010. This areaconsists of 22 island states with a totalpopulation of 7 million but extends overone-third of the world’s surface. The manu-facturers of both albendazole (GSK) andivermectin (Merck) have donated enoughof these agents to carry out the programme.It is very important in ivermectin treatmentthat coverage and pattern of attendance atmass clinics should be satisfactory (Plaisieret al., 2000).
To treat adenolymphangitis and hydro-cele, thorough and frequent washing andpossibly antibiotic and antifungal treat-ment can help to prevent bacterial infec-tion and this appears to preventsubsequent elephantiasis also.
Transmission control. There is no animalreservoir host and so all efforts can bedirected to breaking the human transmis-sion cycle.
1. Insecticide-impregnated (usually perm-ethrin) bed nets and curtains, as used formalaria control, which can be effective formonths even if the net has a few holes.Spraying walls is not as popular as it oncewas, as many mosquitoes, particularlyCulex species, are exophilic and do not restinside houses. Screening of doors and win-dows and provision of ceilings reduced thebiting rate from 200 to 5 per night in a trialin Tanzania.2. The use of long-lasting polystyrenebeads in enclosed breeding sites, such aslatrines, cesspools and ponds to preventthe air breathing of mosquito larvae andpupae. The ventilated improved pit (VIP)latrine with a screened air pipe, whichtraps emerging C. quinquefasciatus adultsas they move towards the light, can help in controlling mosquito numbers. New
The Nematodes 201

202 Chapter 5
biocides, such as Bacillus sphaericus, aself-reproducing, toxin-producing bac-terium specific against C. quinquefasciatus,will also kill larvae.3. Environmental engineering can bedesigned to prevent open drains andsewage systems and to remove unwantedwater and solid waste safely to preventmosquito breeding. Cesspools should alsobe covered. Measures against Culex areoften welcomed by the local population asit is an important nuisance biter (Plate 26).Where the same Anopheles vectors areinvolved, malaria control measures canhelp to control LF also and, indeed, in theSolomon Islands the latter has almost van-ished in this way, although malaria is stillcommon. The situation is similar inVanuatu, which at present has a prevalenceof 2.8%. In Polynesia, clearing up smallcontainers containing water or adding safeorganophosphorus insecticides to drinking-water pots can help to control Aedes vec-tors.
The use of larvivorous fish has longbeen advocated and might have a limitedeffect against Anopheles vectors.
Integrated control measures are likely tobe necessary for successful controlschemes, involving community participa-tion and possibly primary health-care ini-tiatives.4. Chemotherapy also plays a large part inreducing transmission and in New Guineathe annual transmission potential (ATP) ofthe vector Anopheles punctulatus wasreduced enormously after the first year’streatment.
ZOONOTIC ASPECTS
None, although microfilariae of Wuchereriatype have been reported once from a potto(Peridicticus potto). In experimental infec-tions, complete development has beenobtained in leaf monkeys (Presbytiscristata) and macaques (Macaca cyclopis)and slight development in male jirds.Another species, W. kalimantani, is a nat-ural parasite of the leaf monkey inKalimantan (Borneo) and is used in drugtrials but has not been recorded fromhumans. Microfilariae from humans in
Brazil differ in nuclear number and weregiven the name W. lewisi Schacher, 1969,but no differences have been found inadults recovered from the same area.
Brugia malayi (Brug, 1927) Buckley, 1960
SYNONYM
Wuchereria malayi Brug, 1927.
DISEASE
Lymphatic filariasis or filariosis (LF),Malayan filariasis or filariosis, cause of ele-phantiasis.
LOCAL NAMES
As for W. bancrofti. Baku (Japanese inHachijo Koshima Island).
GEOGRAPHICAL DISTRIBUTION
Infection is confined to Asia and occurs inCambodia, southern China, southern India,Indonesia, Korea, Laos, Malaysia,Philippines (Palawan Islands), Vietnamand until recently in Japan.
LOCATION IN HOST
The adults inhabit the lymphatic glandsand lymph vessels, usually of the lowerlimbs and groin. Sheathed microfilariaeoccur in the peripheral bloodstream.
MORPHOLOGY
The adults are very similar to those of W.bancrofti but there are small differences inthe papillae and spicules at the tail end ofthe male worm.
The sheathed microfilariae in the bloodcan be clearly differentiated from those ofW. bancrofti. They measure 170–260 µm �5–6 µm and in stained smears appear stiffand kinky, unlike the smooth curves of W.bancrofti microfilariae (Fig. 90). The sheathis much longer than the body and stainsbright pink in Romanowsky stains, such asGiemsa, at a pH of 6.8 (the sheaths ofmicrofilariae are sometimes lost in theprocess of making a smear, so more thanone should be examined). The round

The N
ematodes
203
Map 9. Distribution of Brugia malayi, B. timori, Loa loa and Mansonella ozzardi.

anterior end is free of nuclei for 12–16 µmand has a double stylet process. The tail ispointed and has two distinctive nuclei atthe posterior extremity. Of the complete100 Mb genome of B. malayi, 33% has nowbeen deposited in a database (Williams andJohnston, 1999).
LIFE CYCLE
The microfilariae are usually nocturnallyperiodic: a subperiodic form occurs inpeninsular Malaya, the Palawan Islands,Sabah and Thailand.
On being ingested by a mosquito vectorfrom the blood capillaries of an infectedperson, the microfilariae lose their sheathsin the stomach and penetrate the wall of thegut. The larvae develop similarly to those ofW. bancrofti and moult twice in the tho-racic muscles before migrating to thelabium in 6–12 days. Infective third-stagelarvae, measuring 1.3–1.7 mm in length,migrate out through the tips of the labellawhen the mosquito bites another individualand enter the skin through the puncturewound (Figs 91 and 97). The larvae reachthe lymph glands, moult twice more andreach maturity in about 3 months.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Brugia malayi causes similar disturbancesof the lymphatic system to those alreadydescribed under W. bancrofti. Elephantiasisis usually confined to the lower leg belowthe knee and is a common complication inMalaysia (Plate 27), while hydrocele isuncommon in males. There is also a higherproportion of apparently symptomless car-riers than with bancroftian filariasis (proba-bly because of a lower intensity ofinfection).
In animals experimentally infected withBrugia species, developing worms cancause dilatation of the lymph vessels andenlargement of the nodes within weeks ofinfection (Denham and Fletcher, 1987);ultrasound studies could help to determinewhether this occurs in humans.
DIAGNOSIS
As for W. bancrofti, although new commer-cial immunological tests for the former do
not cross-react. Specific tests for B. malayiare also being investigated, one of whichuses recombinant ES antigens (Kumari etal., 1994).
In blood films microfilariae of the noc-turnally periodic form often lose theirsheaths, which can be seen separately.
TREATMENT
As for bancroftian filariasis, except thatwith DEC only one-third of the normaldose should be given on the first 2 days,due to likely febrile reactions. These areprobably more severe in malayan filariasisbecause the drug is more effective againsttheir microfilariae and the count is usuallyhigher.
EPIDEMIOLOGY
In all endemic areas of malayan filariasisexcept for Sulawesi (Borneo), bancroftian
204 Chapter 5
Fig. 97. Infective larva of Brugia emerging fromlabella palp of mosquito.

filariasis also occurs. However, the infec-tion patterns of the two infections differ, asexplained below.
The nocturnally periodic form ofmalayan filariasis is transmitted by night-biting species of mosquito; by Mansoniauniformis in most areas, by Anopheles bar-birostris and Anopheles campestris insouthern China, India and Japan, and byAedes togoi, which breeds in swampyareas, in Cheju Island (Korea) and coastalareas of China. Species of Mansonia (M.anullifera, indiana, uniformis), whichtransmit the nocturnally periodic form ofinfection, lay their eggs in small batches onthe undersurface of the leaves of water-plants, such as the water hyacinth inAssam, mangrove in western Malaysia orwater lettuce (Pistia stratiotes) in southernIndia and Sri Lanka. The larvae and pupaeattach to the underwater stems of theseplants by the specially adapted respiratorysiphon through which they obtain air fromthe plant. This form of B. malayi is alsotransmitted by species of Anopheles (An.aconitus, anthropophagus, barbirostris,campestris, donaldi, kewiyangensis, niger-rimus, letifer, sinensis complex, whartoni)and by Aedes togoi. These species, unlikethe Culex vectors of W. bancrofti, cannotbreed in small temporary bodies of waterand so malayan filariasis is a more ruraldisease than bancroftian. The periodicform is highly adapted to humans andreservoir hosts are not important.
The subperiodic form, with microfilar-iae occurring in the peripheral circulationduring the day, is transmitted by species ofMansonia (M. annulata, annulifera, bon-nae, dives, indiana, uniformis), which biteby day as well as by night and are predomi-nantly zoophilic. This form is probably anatural parasite of monkeys and civet catsand human infections occur when rice-fields adjoin areas of, often cleared, forest.
PREVENTION AND CONTROL
Clearing of the water-weeds necessary forthe development of Mansonia larvae can bedifficult as they re-establish so quickly.However, in Sri Lanka, they have beenremoved from the ponds used for treating
copra and, with the help of weed-killersand insecticides against adults (used prin-cipally for malaria control), infection hasbeen virtually eliminated. Unfortunately,bancroftian filariasis is still spreading inSri Lanka.
The worldwide measures being startedagainst LF should be particularly effectiveagainst the nocturnally periodic form ofmalayan filariasis: a mass treatment cam-paign in north-west Malaysia reducedmicrofilarial rates from 26% to below 1%.
ZOONOTIC ASPECTS
Subperiodic B. malayi is one of the mostcatholic of filarial worms and natural infec-tions have been reported from four speciesof monkeys (particularly the leaf monkey,Presbytis obscurus), civet cats, domesticcats and pangolins. Experimentally it willalso develop to maturity in variousrodents.
Brugia pahangi (Buckley and Edeson,1956) is a natural parasite of cats and wildcarnivores in western Malaysia and hasdeveloped to maturity in humans in exper-imental infections. The different forms ofB. malayi may reflect the presence of aspecies complex with the sibling species B.pahangi. The microfilariae of subperiodicB. malayi occurring in wild animals can bedifferentiated from those of B. pahangi bythe fact that only the former will developin Mansonia bonneae.
Brugia timori Partano, Purnomo, Dennis,Atmosoedjono, Oemijati and Cross, 1977,causes timoran filariasis or filariosis inhumans on the islands of Timor, Flores andAlor (see Map 9, p. 203). Microfilariae arenocturnally periodic and in blood smearscan be differentiated from those of B.malayi as the sheath does not stain inGiemsa stain.
Clinical manifestations are similar tothose of B. malayi but the presence of adultworms in the lymphatics often causesabscesses to develop in the groin region(Plate 28). It is transmitted by mosquitoesof the Anopheles barbirostris group (proba-bly An. campestris).
The Nematodes 205

206 Chapter 5
Brugia beaveri Ash and Little, 1964, is anatural parasite of wild carnivores (bobcat,mink and racoon) in Louisiana, USA, andan adult male worm has been recoveredonce from an enlarged human lymph node(Schlesinger et al., 1977).
Brugia guyanensis Orihel, 1964: adultshave been reported once from a human cer-vical lymph node in Peru (Baird andNeafie, 1988). It is a natural parasite of thecoatimundi (Nasua nasua).
Loa loa (Cobbold, 1864) Castellani and Chalmers,
1913
SYNONYMS
Filaria oculi humani, Microfilaria diurna.
DISEASE AND POPULAR NAMES
Loiasis or loaosis; Loa filariasis or filariosis.Eye-worm infection. Cause of Calabarswellings.
LOCAL NAMES
Agan anya (Ibo), Aroro or Aján oju(Yoruba).
GEOGRAPHICAL DISTRIBUTION
Confined to forest areas of West andCentral Africa, particularly where equator-ial rain forest has been cleared and on thefringes of guinea savannah. In northernAngola, southern Cameroon, CentralAfrican Republic, Congo, DemocraticRepublic of Congo (Zaire), Gabon, Ghana,Guinea-Bissau, Ivory Coast, Nigeria (Nigerdelta region), Sierra Leone, southern Sudanand western Uganda, with a few infectionsreported from Ethiopia (see Map 9, p. 204).
LOCATION IN HOST
Adults are found in the subcutaneous con-nective tissues and the sheathed microfilar-iae circulate in the peripheral blood duringthe day.
MORPHOLOGY
The adults are thin transparent worms, thefemales measuring 70 mm � 0.5 mm, the
males 30–35 mm � 0.3–0.4 mm. The headis truncated and has a ring of six papillaejust behind the mouth. The cuticle hasnumerous, randomly arranged, smooth,round bosses; they are absent from thehead end of the female and from both endsof the male (these bosses must be differen-tiated from the regular annulations foundin Onchocerca (Fig. 98), as portions ofeither species could be found in biopsymaterial). The lateral cords are much moreconspicuous than in other human filariae.
In the female the vulva opens 2.5 mmfrom the anterior end. The posterior end isrounded and has a pair of terminal papil-lae. The females are ovoviviparous, thetwin uteri containing all stages of develop-ing eggs and larvae enclosed in egg mem-branes (the sheath). The male has a tailcurved ventrally, with two lateral, posteri-orly placed, cuticular expansions and fivepairs of asymmetrically placed, peduncu-lated papillae around the cloaca and threepairs of small, sessile papillae. There aretwo unequal spicules, measuring 150 and100 µm.
The microfilariae are sheathed and mea-sure 250 (230–300) µm � 6–8 µm. Theyhave a kinked appearance in stained prepa-rations (in contrast to the smooth curves ofW. bancrofti microfilariae). The tail is shortand relatively thick, with large nuclei con-tinuing to the tip. The sheath does notstain with Giemsa or Wright’s stain(although it does with haematoxylin) sothat the microfilariae are sometimes mis-takenly identified as being unsheathed(Figs 99 and 109).
LIFE CYCLE
Microfilariae found in the peripheral bloodshow a diurnal periodicity, being mostplentiful from 08.00 to 17.00 h, when up to40 ml�1 may be present.
The vectors are large (7 mm long), day-biting, tabanid flies, principally Chrysopsdimidiata and C. silacea ( also C. centurio-nis, distinctipennis, langi, longicornis,zahrai and possibly streptobalius).Chrysops, known as the mango, mangroveor softly-softly fly, usually bites (painfully)below the knee and is a pool feeder. The

The Nematodes 207
microfilariae are picked up in a bloodmeal, lose their sheaths inside the stomachof the fly and penetrate the gut wall afterabout 6 h. They develop in the cells of thefat body and moult twice. The third-stagelarvae, measuring 2 mm � 25 µm, migratefirst to the thorax and then to the head andare infective after 10–12 days. The larvaemove down the labium when the fly bites anew host and enter at the site of thewound. Because the vector takes such alarge blood meal, up to 100 larvae can befound in one fly. They migrate through thehuman subcutaneous connective tissuesand muscle fasciae, moulting twice anddeveloping into adults in 6–12 months.The adults live for 4–17 years.
CLINICAL MANIFESTATIONS
A weal appears at the site of the bite if theindividual has been previously infected.
The adult worms migrate through theconnective tissues in any part of the body,the resulting hypersensitivity reaction some-times causing painless Calabar swellings,
which disappear in a few hours or days toreappear elsewhere. These non-pittingoedematous swellings, which can be easilydifferentiated from onchocercal nodules bytheir fugitive nature, appear 6–12 monthsafter infection, most commonly in the handor ankle, and reappear at intervals foranother year or so. The Calabar swellingsare sometimes accompanied by intenseitching, with arthralgia, pruritus and fever,and, if near a joint, may cause difficulty inflexion. In chronic infections the worms godeeper and may migrate through all theviscera of the body; they do not then causeswellings and microfilariae may no longerbe found in the blood, but there have usu-ally been often undiagnosed symptoms offatigue, recurrent fever and perhapsarthritic pains for a long time. In general,indigenous inhabitants have higher micro-filaraemias but fewer swellings and symp-toms than visitors or expatriates. Adultworms sometimes cross the conjunctiva ata rate of about 1 cm min�1 and can then beextracted (Fig. 100); they usually pass
Loa loa
Onchocerca volvulus
0.1 mm
Fig. 98. Portions of Loa and Onchocerca as may be recovered in tissue biopsies.

208 Chapter 5
across the eye in minutes but can remainfor days, causing considerable discomfort.In most cases loiasis is a relatively benigndisease with a good prognosis.
A hypereosinophilia of 60–90% (20,000–50,000 eosinophils mm�3) is frequentlyfound and may contribute to the endocar-dial fibrosis sometimes reported; pul-monary infiltrations have also beenreported rarely. IgE levels are typicallyraised, although it is not clear what rolethis plays in the host response.
PATHOGENESIS
Biopsy sections of swellings sometimesshow adults in the centre of an oedematousreaction. In chronic cases the skin maybecome lichenified. Dying worms in thesubcutaneous tissues can cause a chronicabscess, followed by a granulomatous reac-tion and fibrosis (Fig. 101); worms havebeen excised from many parts of the body.
It is likely that in chronic sufferers whoare amicrofilaraemic, microfilariae are beingdestroyed by immune responses in these
Fig. 100. Adult of Loa moving under the conjunctiva.
20 �m
Fig. 99. Microfilaria of Loa loa (stained in haematoxylin – compare Fig. 109).

hypersensitized individuals, and the endo-myocardial fibrosis and nephrotic syndromesometimes reported from such cases resem-ble the situation in the collagen diseases.
Various visceral lesions have beenattributed to loiasis, but the most seriouscomplication is meningoencephalitis,which can be fatal and is not uncommon inareas of high endemicity. It may be accom-panied by retinal haemorrhages and isassociated with DEC in patients with veryhigh microfilaraemia. Microfilariae may befound in the cerebrospinal fluid of suchcases and encephalitis is presumablycaused by an allergic reaction to dead ordying microfilariae obstructing the capillar-ies of the brain (Pinder, 1988).
The relatively mild pathology usuallyassociated with this infection includes anumber of allergic manifestations. Thesemay include pruritus, oedema and arthri-tis, all indicative of immune-mediatedreactions to the parasite. Studies in man-drills have shown both Th1 and Th2responses during infection, although thereis some stage specificity (L3s preferentiallystimulated Th2, while microfilariae pro-voked both Th2 and Th1 cytokines). Theadverse reactions to DEC seen in loiasispatients are, as in onchocerciasis patients,associated with immunological hyper-responsiveness to microfilarial antigens(Wahl and Georges, 1995).
DIAGNOSIS
The most common method of diagnosis isby finding microfilariae in thick bloodsmears taken during the day. In chronicinfections and those in visitors, microfilar-iae may be very scanty and concentrationtechniques can be used (see p. 263). Themicrofilariae also live for many years andcan continue to circulate even after adultsare dead.
The presence of the fugitive (‘waspsting’) swellings with high eosinophilia ina patient who lives in or who has visitedan endemic region is characteristic of anearly infection. Biopsy sections ofswellings sometimes show portions ofadults in the centre of an oedematous reac-tion. Adult worms seen moving across theconjunctiva can be removed and identified.
Detection of antibodies is possible but isnot very specific. Recently, methods fordetection of circulating antigens have beenmore successful. An ELISA using parasite-specific IgG4 can recognize specific Loaantigens of low molecular mass (12–30kDa) and works in patients with or withoutmicrofilaraemia. The test can also differen-tiate from other filarial infections, particu-larly Mansonella perstans (Akue et al.,1994). Antigen detection by coelectro-syneresis (Co-ES) has also shown promise.A PCR amplification method to detect LoaDNA using a Southern blot technique is
The Nematodes 209
Fig. 101. Section of subcutaneous tissues with dying male Loa causing a host reaction.

stated to be very sensitive and specific(Toure et al., 1997).
TREATMENT
Chemotherapy. DEC kills both adults andmicrofilariae when given at 2–6 mg kg�1
(usually 300 mg for an adult) for 14–21days. However, side-reactions, such astransient oedema, generalized eruptions,arthralgia, nausea, diarrhoea and fever,often occur and, where there is a highmicrofilaraemia (over 1000 mm�3), moreserious effects are possible, such as meningo-encephalitis, coma and renal damage. It iswise to commence treatment at a very lowdosage in all patients with more than 20microfilariae ml�1, perhaps under corticos-teroid cover.
Ivermectin (200 µg kg�1 body weightevery 3 months for 2 years) decreasedmicrofilarial levels by 90% and reducedprevalence from 30% to 10% in a trial inCameroon, but these slowly returned toprevious levels after about 2 years. Side-effects of pruritus, headache, arthralgia andmaybe fever are common (Chippaux et al.,1992). Rare reports (1 per 10,000 personstreated in one trial) of encephalopathy,coma and renal damage have been noted.The use of a single dose of ivermectin (200µg kg�1 ) to reduce microfilaraemias beforeDEC treatment has also been advocated.
Albendazole (200 mg twice daily for 21days) causes microfilarial numbers to fallslowly and may have an adulticidal(macrofilaricidal) action, since immunolog-ical tests become negative after a fewmonths and eosinophilia falls.
Surgery. When a worm is seen movingacross the eye, a curved, bayonet-edge,surgical needle can be passed under theworm and it can then be removed with amounted needle. The conjunctiva andworm are first anaesthetized with a fewdrops of 10% novocaine.
EPIDEMIOLOGY
The disease is restricted to equatorial forestregions and prevalence is falling in someareas as forest is cleared. Recent estimates
of infection are 30% in Calabar and 10% inGongola State in Nigeria, 30% inCameroon, 19% in Bantu and 11% in pyg-mies in Congo, 10–70% in the Republic ofCongo, 12% in Equatorial Guinea, 30% inGabon; everywhere infection rates arehigher in adults than in children.
Chrysops breeds in densely shadedstreams covered with leaves, in which thesandy bottom is overlaid with mud anddecaying vegetation, on which the slowlydeveloping larvae feed. In West Africa,adult Chrysops are particularly common inthe rainy season (June–September). Theadult female flies live in the high canopy ofthe forest and possibly feed on monkeys.They are attracted to movement below or towood smoke rising, and humans get bittenmost often in cleared areas, such as rubber,cocoa, teak or oil-palm plantations on theforest fringes, where the flies can see abreak in the canopy. C. dimidiata and C.silacea are the principal vectors in WestAfrica and C. distinctipennis in CentralAfrica. Flies bite during the day and(except sometimes for C. silacea) do notusually enter native houses as they are toodark inside. Chrysops ingests 10–20 µl ofblood per meal; it probably feeds aboutonce a fortnight, since it requires one fullblood meal in each gestation period ofabout 12 days.
PREVENTION AND CONTROL
For individual protection, repellents andprotective clothing can be used. It has alsobeen reported that DEC kills infective larvaeand 5 mg kg�1 daily for 3 days every monthhas been suggested as a prophylactic.
In the past dieldrin sucessfully con-trolled larvae and pupae in forest streamsin Cameroon, but treatment (nowadaysprobably with temephos) is not practicalover large areas. A mass campaign withDEC in Nigeria was not successful, butprevalence was reduced from 30% to 10%in an area of Cameroon after mass treat-ment with ivermectin.
ZOONOTIC ASPECTS
A sympatric but ecologically separate cycleof Loa loa is also found in many species of
210 Chapter 5

The Nematodes 211
forest-dwelling primates, such as the drill(Papio = Mandrillus leucophaeus), themona monkey (Cercopithecus mona) andthe putty-nosed guenon (C. nictitans).However, the microfilariae in these hostsare larger and show a nocturnal periodic-ity, while the species of Chrysops acting asvectors (C. centurionis and C. langi) arealso night biters. The parasite is sometimesgiven subspecific status as L. loa papionis,with the human parasite called L. l. loa.The monkey and human strains have beenhybridized experimentally, but it isunlikely that humans become infected withthe monkey strain, because the flies bitealmost entirely in the upper canopy; thereis also some evidence that humans cannotbe infected with the monkey strain (or sub-species). It is possible, though, that mon-keys living near the ground sometimesbecome infected with the human form andcould then act as reservoir hosts.
A similar parasite, Loaina sp., of rabbits,transmitted by mosquitoes, has been recov-ered once from the anterior chamber of theeye in a man in Colombia (Orihel andEberhard, 1998).
Onchocerca volvulus (Leuckart, 1893) Railliet and Henry, 1910
SYNONYMS
Filaria volvulus Leuckart, 1893; Oncho-cerca caecutiens Brumpt, 1919.
DISEASE AND POPULAR NAMES
Onchocerciasis or onchocercosis; riverblindness, craw-craw (West Africa), blind-ing filaria.
LOCAL NAMES
Amar eljur (N. Sudan), Aràn oju (Yoruba),Firkaw and Ekraw (Twi), Kirci (Hausa),Sowda (Yemen), Ungoujwa ya usinyi(Sambaa).
GEOGRAPHICAL DISTRIBUTION
Africa (including Angola, Benin,Cameroon, Central African Republic, Chad,
Congo, Côte d’Ivoire, Equatorial Guinea,Ethiopia, Gabon, Ghana, Guinea, Liberia,Malawi, Mali, Nigeria, Republic of Congo,Senegal, Sierra Leone, Sudan, Tanzania,Togo and Uganda, with limited foci inBurkina Faso, Guinea-Bissau, Niger andYemen) and Central and South America(Brazil, Guatemala, southern Mexico,Venezuela and possibly still Colombia andEcuador). It is estimated that there areabout 17.5 million cases in Africa and140,000 in the Americas, of whom 0.8 mil-lion have visual impairment and 0.27 mil-lion are blind.
LOCATION IN HOST
The adult worms live in the subcutaneoustissues and in long-standing infections theyform tangled masses inside fibrous nodules(onchocercomas). Microfilarial larvae arefound in the skin (probably mostly in lym-phatic channels in the dermis) but not usu-ally in the blood.
MORPHOLOGY
Both ends of the filiform adults are taperedbut the tail terminates bluntly. The anteriorend has two circles of four papillae and alarge lateral pair. The cuticle has conspicu-ous annular thickenings (rugae), which areimportant in identification as usually onlyportions of worms can be obtained fromnodules (Fig. 98).
The adult female measures 300–500 mm� 0.25–0.4 mm and the male 20–40 mm �0.15–0.20 mm. The vulva of the femaleopens about 0.85 mm from the anterior endand the uterus is bicornate. The male has avariable number of anal papillae and twounequal spicules.
The microfilariae measure 280–330 µm� 6–9 µm. They are unsheathed and have atapered tail, and the head and tail are freeof nuclei (Fig. 102). The head is character-istically expanded and the cuticle is stri-ated. The microfilariae wander in the skinbut are not usually found in the blood-stream, except after DEC treatment; theycan survive for 6–24 months. Sometimesmicrofilariae are also present in the urineand sputum.

212 Chapter 5
20 �mOnchocerca
volvulus
Mansonellastreptocerca
Fig. 102. Microfilariae of Onchocerca volvulus and Mansonella streptocerca.
Map 10. Distribution of Onchocerca volvulus.

The Nematodes 213
LIFE CYCLE
Microfilariae in the skin are ingested by bit-ing flies of the genus Simulium (known asblack-flies or buffalo gnats. Figs 103 and104). The microfilariae are probablyattracted to the site of the bite by salivainjected into the puncture wound. The flieshave a short scarifying proboscis and arepool feeders on tissue fluids released bytheir bite. Some of the microfilariaeingested by the fly leave the midgut beforethe formation of the peritrophic membraneand penetrate the thoracic muscle cells,particularly the flight muscles; others aretrapped in the midgut and die. After twomoults in the muscles of the fly, the firstmoult giving rise to a sausage stage, the fewinfective larvae move to the head and pro-boscis, complete development taking 6–12days, depending on the ambient tempera-ture (no development occurs below 18°C).For studies of transmission rates, the infec-tive third-stage larvae must be differenti-ated from other animal filarial larvae found
in Simulium (including the cattle speciesO. ochengi, which occurs in West Africaand has morphologically identical larvae);they measure 560 (440–700) µm � 19 µmand have a single small knob at the tail.These larvae probably enter through thepuncture wound when an infected fly bitesanother individual. They migrate to thesubcutaneous tissues, moult twice andreach sexual maturity in about 10–15months. Microfilariae can first be found inthe skin by 18 months after infection andtake 4–6 weeks to reach the skin after leav-ing the vulva of the adult worm (about 1000actively leave the vulva of each female perday). The adults live on average for 9–11years and very rarely survive for more than13 years, while microfilariae can live in theskin for up to 2 years.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Most of the severe manifestations are dueto the presence of the microfilariae; theadults are of secondary importance (contrast
Fig. 103. Diagram of head of Simulium feeding on human skin with infective larvae of O. volvulusemerging from the labium (it is probable that they usually emerge from the posterior surface of the labiumor from the hypopharynx).

214 Chapter 5
infections with the lymphatic filariae). Inlong-standing infections the adults areoften found in characteristic non-tendernodules, known as onchocercomata, whichcan be felt and often seen, especially overbony prominences (Fig. 105). There may bea single nodule or many present. In Africanodules are usually found around the
pelvic region, knees and lateral chest andspine, while in the Americas (except forVenezuela) the nodules are more com-monly seen in the upper part of the body,including the head.
Death of adult worms deep in the tis-sues can lead to the formation of an abscessand nodules on the scalp may erode thebone.
As in many helminth infections, there isa blood eosinophilia, in this case of10–75% (usually with 700–1500 eosino-phils mm�3 blood).
The majority of lightly infected indige-nous individuals living in an endemic areashow few or no signs, although at sometime many will have pruritus, with skinlesions leading to the characteristic presby-dermia. In the early stages, or in lightlyinfected persons, the presence of the adultworms cannot be detected, but the micro-filariae usually cause pruritus with a per-sistent and intensely itchy rash of the skinin their vicinity (filarial itch), often con-fined to one anatomical quarter of the body.The rash consists of many raised papules,1–3 mm in diameter, and often becomessecondarily infected following scratching(‘craw-craw’ or ‘gale filarienne’). There isusually lymphadenopathy in the groin oraxilla of the affected side, sometimesaccompanied by aches and pains. The
Fig. 104. The black-fly Simulium damnosum s.l.feeding on human.
Fig. 105. Section of subcutaneous nodule with adult female and microfilariae (one arrowed) in the fibrous connective tissues.

The Nematodes 215
onchodermatitis can lead in months oryears to a secondary stage of thickeningdue to intradermal oedema (often giving a‘peau d’orange’ effect) and to pachydermia(lichenification or crocodile skin). The lossof elastic fibres in the skin can lead to‘hanging groin’, pendulous sacs containinginguinal or femoral glands (Fig. 106) orhernias, which are particularly found inmales in East Africa, and probably to theleonine facies encountered in Guatemala.Elephantiasis of the scrotum can also occurin males but, in contrast to that caused byBancroftian filariasis, there is not usuallyan associated hydrocele.
The last stage is characterized by atro-phy of the skin, with loss of elasticity, giv-ing a striking prematurely aged appearanceand a paper-thin skin. Mottled depigmenta-tion (‘leopard skin’), particularly of the
shins, is common and can resemble leprosy(Fig. 106). Skin changes often mimic thoseseen in vitamin A deficiency. Progressiveskin lesions can be roughly divided intostages: acute and then chronic papularonchodermatitis (Fig. 107); lichenifiedonchodermatitis; atrophy and depigmenta-tion (Murdoch et al., 1993).
In Central America, patients, particu-larly children, may have reddish-mauvelesions on the face, known as erisipela dela costa. The condition of ‘sowda’ is foundin Yemen and in East and West Africa, inwhich there are very few microfilariae inthe skin, but the skin of a localized area,usually one or both legs, is itchy, papularand pustular, hyperpigmented and thick-ened, with accompanying groin lymph-adenopathy, distinguishable clinically from‘hanging groin’. This hyperreactive oncho-dermatitis, with frequent acute exacerba-tions, is also encountered in visitors ornew residents to an endemic region.
The most important consequence of
Fig. 107. Section of skin with marked dyskeratitisand presence of microfilariae of O. volvulus.
Fig. 106. A case of ‘hanging groin’ with loss ofelasticity in the skin. There is also vitiligo of theshins (‘leopard skin’).

infection is blindness, which has led to thepopular name for the disease of river blind-ness. Blindness occurs more commonly insavannah areas of Africa and America thanin forest zones, where it is seldom over 1%.In some savannah areas of Africa andGuatemala, over 10% of the population maybe completely blind and another 20% haveimpaired vision on account of onchocercia-sis. In endemic savannah areas the sight of asmall boy leading a band of blind men totheir work in the fields is not uncommon. Afew years ago there were reckoned to beabout 70,000 adults with ‘economic blind-ness’ in the upper Volta basin.
The eye lesions can be separated intotwo types.
1. Anterior lesions are caused by micro-filariae reaching the cornea from the skin ofthe face through the conjunctiva. By theuse of the slit lamp, live microfilariae maysometimes be seen in the cornea, usuallyjust below Bowman’s membrane, withoutcausing any reaction. When microfilariaedie they first cause a punctate keratitiswith numerous opacities, with a diameterless than 0.5 mm, around each dead micro-filaria (Fig. 108). The fluffy opacities areoften symptomless, but there may be photo-
phobia and watering, with chronic con-junctivitis. This can be followed by a scle-rosing keratitis, usually starting in thelower half of the cornea, with an ‘apron’coming in from the sides and below (Plate29). Microfilariae may also be seen in theanterior chamber of the eye and their deathmay produce iridocyclitis and uveitis, withloss of the pigment ruff and a distortedpupil. Secondary glaucoma and cataractmay follow. 2. The aetiology of the posterior-segmentlesions is not so clear but they are associ-ated with heavy O. volvulus infection inboth savannah and forest areas. The‘Ridley–Hissette’ fundus is a well-definedpatch, usually bilateral, in which all retinalelements have vanished except for the reti-nal vessels and in which the choroidal ves-sels may show marked sclerosis (Plate 30).Optical atrophy is of the postneuritic type,with dense sheathing of the retinal vessels.Active optic neuritis appears to be com-moner than once thought (WHO, 1995) andlasts for several weeks to over 1 year, withscarring and pigment disturbance at thedisc margin. It has a reported prevalence of1–4% in hyperendemic communities inCameroon and 6–9% in the guinea savan-nah of northern Nigeria. Complete blind-
216 Chapter 5
Fig. 108. Snowflake opacities in onchocercal keratitis (slit-lamp view and section).

ness may result or tubular vision mayremain. Onchocerciasis can cause severereduction in peripheral visual fields, sothat functional blindness might be muchhigher than the standard ‘below 3/60 visualacuity in the better eye’ definition shows.A new field test, the Wu–Jones computer-ized visual function test, should make iteasier to obtain accurate and reproduciblemeasurements of paracentral visual fields,as well as acuity.
IMMUNOLOGY OF ONCHOCERCIASIS
There are some overall similarities betweenthe immunobiology of onchocerciasis andlymphatic filariasis, but some importantdifferences associated with the differencesin parasite location.
As with lymphatic filariasis, infectedindividuals show a spectrum of pathologi-cal responses and these may reflect geneticas well as environmental influences.Individuals with high microfilarial loadsmay show a degree of both specific andnon-specific immune unresponsivenessand low-grade skin inflammatory changes.A small percentage of patients show hyper-reactive responses, with severe localizedskin changes typical of sowda. When testedagainst worm antigens, patients withhyperreactive onchocerciasis showed evi-dence of a Th2 bias in their antibodyresponses – for example, making greaterIgE and IgG1 and lower IgG4 and IgMresponses than those with generalizedonchocerciasis. These differences inclasses of antibody associated with differ-ences in pathology may also be relevant toprotection – IgE and IgG1 participating inADCC-mediated killing, while IgG4 andIgM antibodies may block effector activity.Th1-biased responses have been implicatedin the development of immunity, althoughboth infected patients and endemic nor-mals appear to make mixed Th1/Th2responses. Production of both Th1 and Th2cytokines in putatively immune individu-als may allow the involvement of bothmacrophages and eosinophils in ADCC-mediated killing of infective L3 stages.
Chemotherapeutic treatment of oncho-cerciasis with drugs, such as DEC and iver-
mectin, carries the risk of adverse reac-tions. In part, these follow increased pro-duction of proinflammatory mediators,such as IL-6, tumour necrosis factor-α(TNF-α) and C-reactive protein, but thereare also changes in T cells and eosinophils.
Major histocompatibility complex(MHC)-linked influences on patterns ofinfection and pathology have been estab-lished and the interesting idea proposedthat possession of one human leucocyteantigen (HLA) locus (DQB1*0501), associ-ated with protection against severe malaria,may carry the cost of reduced resistance toonchocerciasis – an example of a trade-offbetween two endemic diseases in WestAfrica (Meyer and Kremsner, 1996).
DIAGNOSIS
Clinical. The acute pruritic onchocercalrash has to be distinguished from thosecaused by scabies, insect bites, pricklyheat, contact dermatitis, hypersensitivityreactions, post-traumatic or inflammatorydepigmentation, tuberculoid leprosy, dermato-mycoses, trepanonemoses and the similarparasite Mansonella streptocerca (seep. 223). The presence of the typical nod-ules measuring 0.5–10 cm in diameter,from which portions of adult worms maybe obtained on excision, allows a positivediagnosis to be made if no microfilariae canbe found in skin snips. Ultrasound can alsobe used to confirm whether a nodule isonchocercal. The presence of nodules inmore than 20% of young males over theage of 20 in a savannah area is estimated togive a sensitivity of 94% and specificity of50% (Whitworth and Gemade, 1999) inepidemiological surveys, while skin depig-mentation might give a better rapid, butrough, method in forest regions. In chronicinfections, skin and eye changes are usu-ally pathognomic.
The Mazzotti or DEC provocation test isstill sometimes used, in which 50 mg ofDEC is given orally and, hopefully short-lived, itching, rash and lymphadenitisoccur 1–24 h later. A much better recentmodification uses a DEC-impregnatedpatch and results in a small localized rash.
The Nematodes 217

Parasitological. Microfilariae may some-times be seen in the cornea and anteriorchamber of the eye and, in heavy infec-tions, in urine and blood.
The most widely used diagnostic proce-dure in onchocerciasis is the skin snip.After cleaning with spirit, a needle is usedto raise a small cone of skin, which is thencut off under the needle point with a sharprazor-blade or fine pair of scissors (theWalser or Holth corneosceral punch is usedto give a standard-sized sample of skin andenables quantitative microfilarial counts tobe made). It is important to take the snipfrom an area of maximum microfilarialdensity – from above the iliac crest inAfrica, from the trapezius in Guatemalaand from the lower calf in Yemen. Thebloodless skin snip, measuring about 3 mmin diameter and 0.1–1 mm deep (preferablytaken in the afternoon), is placed in a dropof physiological saline or water and, aftergentle teasing, is allowed to stand for 30min before being examined under themicroscope. Microfilariae show activethrashing movements and in West Africamust be separated from those of M. strepto-cerca, also found in the skin. Microfilariaeof the latter have a shivering mode of activ-ity and in stained smears have a thinnershape, small caudal space and non-over-lapping of the anterior nuclei (Fig. 102).Recently, skin scarification is being widelyused as an alternative to skin snips.
Immunological. Superficial skin scrapings,which can be dried on microscope slides,can also be used in a PCR-based assay toshow the presence of onchocercal DNA(Toe et al., 1998), but is still expensive.This test can also be used to specificallyidentify infections in black-flies and to dif-ferentiate between savannah and foreststrains of parasite (WHO, 1995).
Most immunological tests are more use-ful for epidemiological studies than forindividual diagnosis. ELISAs using a cock-tail of recombinant antigens (e.g. MBP/10,11 and 29) have been tested in many areaswith different levels of endemicity (WHO,1995; Bradley and Unnasch, 1996; Bradleyet al., 1998), and tests have been devised
for recognizing antigens in tears for ocularonchocerciasis, in dermal fluid for skinmicrofilariae and in urine for microfilariae(Ngu et al., 1998).
TREATMENT
Surgery. Nodulectomy under local anaes-thetic (2% lignocaine) has been widelyused in Central America and has beenclaimed to markedly reduce the number ofmicrofilariae in the skin and more impor-tantly around the eyes (Plate 31). Resultshave not been so striking in Africa, wherenodules are often impalpable or the largerones are merely fibrous reactions arounddead and dying worms.
Chemotherapy. Ivermectin (Mectizan – amacrocyclic lactone) is the only drug inwidespread use and has completelyreplaced DEC, which is no longer advo-cated. It is given as a single oral dose of150 µg kg�1 of body weight once or twice ayear. It should not be given in cases ofsevere illness. Side-effects are very rarelysevere but can include itching and rashes,fever and glandular pain (severe allergicreactions used to be common after treat-ment with DEC, which kills microfilariaemore rapidly). Ivermectin appears to havelittle effect against adult female wormswhen given at the standard dose but doesprevent the release of microfilariae fromthe uterus for over 6 months. Moxidectin, apromising long-persisting relation of iver-mectin, is now undergoing clinical trials,as is long-term treatment with antibiotics,probably directed against Wohlbachia (seep. 197. (Townson et al., 2000).
Intravenous suramin (Antrypol or Bayer205) is the only drug which has a clearmacrofilaricidal effect, but it is toxic andshould only be given rarely and underclose medical supervision in hospital(WHO, 1995).
EPIDEMIOLOGY
Persons involved in agriculture and otheroutdoor activities, such as fishing, are atthe highest risk, and in some parts of WestAfrica in the past onchocerciasis has ren-
218 Chapter 5

dered whole areas bordering rivers unin-habitable.
In Africa, the most important vectors areblack-flies belonging to the Simuliumdamnosum complex, small (4 mm long)nematocerous flies, which breed on vegeta-tion and on rocks in a great variety of riversystems, including the largest rivers, suchas the Niger, Volta and Nile, as well assmall streams, provided that they are welloxygenated. Members of the S. damnosumsubcomplex acting as vectors (S. damno-sum sensu stricto (s.s.), S. sirbanum, S.rasyani and possibly S. dieguerense) arefound in savannah areas, while those of theS. sanctipauli (S. sanctipauli s.s., S.soubrense, S. leonense and S. konkourense)and S. squamosum (S. squamosum s.s., S.yahense, S. mengense and S. kilibanum)subcomplexes are mostly found in forestareas. Most of these sibling species are mor-phologically identical and can only be dif-ferentiated by the banding patterns of thelarval chromosomes; many othercytospecies not known to act as vectorshave also been described. Members of theS. neavei complex (S. neavei s.s., S. woodiand S. ethiopense) are local vectors in east-ern Congo Republic (Zaire), Tanzania andEthiopia; all members breed in small high-land streams and have an obligatory associ-ation with freshwater crabs or othercrustaceans. S. albivirgulatum is a localvector in central Congo Republic.
In the Americas, members of the S.ochraceum complex are high-biting fliesthat are the principal and efficient vectorsin highland foci in Mexico and Guatemala.They breed in small rivers, streams andsmall trickles of water flowing throughhighly wooded volcanic slopes of coffee-growing areas between 1000 and 2000 mabove sea level. S. callidum and S. metal-licum sensu lato (s.l.) are secondary vec-tors; the latter is low-biting and mainlyzoophilic and is also found in Venezuela.Various other species (S. exiguum s.l., S.guianense, S, incrustatum, S. oyapockenses.l., S. quadrivittatum and probably S. lim-batum) act as vectors in South America(WHO, 1995).
Variations in the localization of nodules
in Africa and Central America are probablycaused by differences in the biting habits ofthe species of black-flies.
Although all species of Simulium areconfined to running water for breeding,adult flies have been found many kilo-metres from water and are probably carriedby winds. In savannah areas of West Africa,flies have a shorter flight range than thosein forest areas, so that infections tend to beheavier but more localized.
The epidemiology of onchocerciasis dif-fers over its range because different diseasepatterns are associated with differentstrains of parasite, with differences in theabundance, anthropophilic nature and vec-tor capacity of vectors and with differencesin the responses of the human host to theparasite. Because of strain differencesbetween parasites in Africa, microfilariae ofsavannah forms will not infect forest fliesand vice versa; also, anterior eye lesions aremore common in savannah than in forestareas in infections of comparable severity.
PREVENTION AND CONTROL
Control campaigns in the past have con-centrated on the vectors. The disease waseradicated from Kenya and from large areasof Uganda in the 1950s by using the insec-ticide (dichlorodiphenyltrichloroethane:DDT). In Kenya the Simulium speciesresponsible (S. neavei) has a short flightrange and lived on land crabs which inhab-itated water-filled holes, and these wereeasy to treat. In Uganda infection was con-fined to the Nile, which could be treated.
The largest control campaign is that ofthe Onchocerciasis Control Programme(OCP) (funded principally by the WorldBank, Food and Agriculture Organization(FAO), United Nations DevelopmentProgramme (UNDP) and WHO), whichcommenced in 1974, dedicated to control-ling the disease in savannah regions ofseven West African countries of the VoltaRiver basin, comprising a land area of 0.7million km2 (Benin, Burkina Faso, Côted’Ivoire, Ghana, Mali, Niger and Togo). Insome infected villages in the control area,more than 10% of the infected populationwas blind and as many as 30% had
The Nematodes 219

220 Chapter 5
irreversible eye lesions. At first, controlwas entirely by the addition of the larvi-cide temephos (Abate) to rivers andstreams, mostly from helicopters and fixed-wing aircraft (Plate 32). In 1980 insecticideresistance was encountered and the bacter-ial agent Bacillus thuringiensis H14replaced it for a while until black-fly popu-lations became susceptible again. By 1986vector control had interrupted transmis-sion almost completely in 90% of the area,with blindness reduced by 40%, while theincidence of infection in children wasreduced by 99%. However, in the remain-ing 10% western area, reinvasion by fliesborne on the monsoon winds occurred, sothe control area was extended to a furtherfour countries (Guinea, Guinea-Bissau,Senegal and Sierra Leone). Since 1987 themicrofilaricide ivermectin (Mectizan) hasbeen donated by the manufacturers andthis has augmented the use of insecticides.To date, over 20 million people havereceived at least one dose and some up tonine doses. The drug is given at 150 µmkg�1 once or twice a year, as it does not killadult worms. With the use of both regularMectizan and larviciding, treatment needsto be continued for about 12 years toreduce infection by 99% (this representsthe average lifespan of adult worms).
The campaign currently costs about $27million per annum but is due to end in2002, when responsibility will be handedover to the African Programme forOnchocerciasis (APOC). The maintenancephase of control will be almost entirely byrepeated community-based administrationof Mectizan (Remme, 1995) and a commu-nity-directed treatment programme isalready under way in 19 African countries,which will become part of primary healthcare. It has been realized recently that thesevere skin lesions, which also occur in theforest zones, can be very debilitating andtreatment will be much more widespreadthan just the OCP area.
The benefits of the OCP to date can beenumerated as below (Samba, 1994).
1. 10 million children prevented frombecoming infected.
2. 125,000–200,000 adults prevented frombecoming infected, with 1.5 million hav-ing lost their infection.3. 25 million ha that can now be used foragriculture – enough to support 17 millionpeople.4. Infection no longer being a problem inBurkina Faso, Niger, northern Benin, Côted’Ivoire, Ghana, Togo or south-easternMali.5. An estimated 20% return on the invest-ment in the programme.
In Central America insecticidal controlhas not been successful because of the dif-ficulties in treating all the small mountainstreams in which Simulium larvae breed.However, mass treatment campaigns withMectizan are proving very effective in bothCentral and South America.
ZOONOTIC ASPECTS
Probably none, although a natural infectionhas been found in a gorilla, and chim-panzees can be experimentally infected (aninfection reported from a spider monkeyappears to have been mistaken). Otherspecies of Onchocerca are found in a widerange of herbivores (O. gutturosa in thenuchal cartilage, O. lienalis in the gastro-splenic region and O. gibsoni in the sub-cutaneous tissues of cattle and O. cervicalisand O. reticulata in the ligaments of horsesoccur in many parts of the world and O.armillata parasitizes the aorta of cattle inAfrica and Asia). The horse species aretransmitted by species of Culicoides, butthe cattle species are transmitted bySimulium species and so could be con-fused with O. volvulus in the vector. Thisis most important for a subcutaneous cattlespecies in West Africa, O. ochengi, as theinfective larvae are almost identical tothose of the human species and are foundin the same species of black-fly (differentia-tion requires DNA probes and can becarried out on large pooled samples offlies).
A few amicrofilaraemic infections, pre-sumably zoonotic from cattle or horsespecies, have been reported from Canada,Japan, Russia, Switzerland and the USA.

The Nematodes 221
Mansonella perstans (Manson, 1891) Orihel and Eberhard, 1982
SYNONYMS
Filaria sanguinis hominis minor, F. s. h.perstans and F. perstans Manson, 1891;Acanthocheilonema perstans Railliet,Henry and Langeron, 1912; Dipetalonemaperstans Yorke and Maplestone, 1926;Tetrapetalonema perstans Chabaud andBain, 1976. This species and M. strepto-cerca (below) have had many rather confus-ing name changes. For many years theywere included in the very large and hetero-geneous genus Dipetalonema and weresplit off as Tetrapetalonema just beforeadults of Mansonella ozzardi were recov-ered from experimental animals, when they
were all found to belong the same genus(Muller, 1987). M. perstans and M. strepto-cerca are included in the subgenusEsslingeria while M. ozzardi is in the sub-genus Mansonella, i.e. the full names are M.(Esslingeria) perstans, M. (Esslingeria)streptocerca and M. (Mansonella) ozzardi.
DISEASE
Perstans filariasis or filariosis, mansonellia-sis or mansonellosis perstans.
GEOGRAPHICAL DISTRIBUTION
West and Central Africa, including Benin,Cameroon, Congo, Republic of Congo,Equatorial Guinea, Ethiopia, Gabon, Ghana,Guinea, Guinea-Bissau, Mali, Mozambique,Nigeria, Sierra Leone, Togo, western
Map 11. Distribution of Mansonella perstans.

222 Chapter 5
Uganda, Zimbabwe and limited areas ofSouth America (Colombia, Guyana andVenezuela).
LOCATION IN HOST
Adults inhabit the body cavities, mesen-teric, retroperitoneal and perirenal tisues;unsheathed microfilariae can be found inthe peripheral bloodstream at any time.
MORPHOLOGY
The adult females measure 70–80 mm �0.12 mm, males 35–45 mm � 0.06 mm;both sexes have a smooth cuticle and aventrally curved tail, which has a trilobedappearance (Baird et al., 1987). The tail ofthe male has four pairs of preanal and onepair of postanal papillae and there are two,very unequal, rodlike spicules. The micro-filariae in the blood are unsheathed andmeasure 200 (190–240) µm � 4.5 µm (theylook appreciably smaller and thinner thanthe sheathed microfilariae of Wuchereria,Brugia or Loa); they assume graceful curveswhen fixed and stained. The tail endsbluntly and contains nuclei to the tip (Figs89 and 109).
LIFE CYCLE
There is no marked periodicity but microfi-lariae are slightly more numerous in theperipheral circulation at night. The microfi-lariae are ingested in a blood meal by small(2 mm) female ceratopogonid midges of the
genus Culicoides, which are pool feeders.The larvae enter the thoracic muscles, moulttwice and reach the infective third stage by 9days, when they migrate to the head. Theyexit from the labium of the midge while it isfeeding. The parasites take a few months tobecome mature, moulting twice more, in thehuman body.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
There are often none, but infection hasbeen associated with allergic symptoms,such as itching, pruritus, joint pains,enlarged lymph glands and vague abdomi-nal symptoms, accompanied by higheosinophilia.
DIAGNOSIS
By identification of the microfilariae instained blood smears. More than ten micro-filariae per millilitre may be present, oftentogether with those of L. loa and W. ban-crofti. Differences in the morphology ofmicrofilariae in various African countrieshave resulted in the name M. semiclarumbeing given to a longer microfilaria (198 µm� 5.2 µm in thick films, 221 µm � 5.0 µmin thin) found in the blood of a man inCongo Republic (Dujardin et al., 1982).
TREATMENT
DEC has no effect. Albendazole andmebendazole given for 10 days have givenequivocal results. Ivermectin appears to
Fig. 109. Microfilariae of Loa loa and Mansonella perstans in same blood smear (stained in Giemsa sothat sheath of Loa has not stained).

The Nematodes 223
reduce microfilaraemia for a period butprobably has no long-term effect.
EPIDEMIOLOGY
Recent reports of infection rates are south-ern Cameroon 15–27%, southern Congo16% in Bantu and 81% in pygmies, CongoRepublic (Zaire) 42%, Equatorial Guinea52%, Gabon 25–80%, Abia and Imo States29%, Bauchi State 1.4%, Gongola State11%, Jos Plateau 13% (all in Nigeria).
In Africa the vector species areCulicoides grahami and C. milnei. Thesehave a short flight range of about 100 m.Eggs are laid in mud or wet soil in batchesor in water in banana stumps.
There is usually only one larva perinfected midge, but midges are numerousand a high percentage may be infected.This results in a high infection rate in thehuman population, but almost all infec-tions are light.
ZOONOTIC ASPECTS
Possibly identical microfilariae are foundin chimpanzees and gorillas in CentralAfrica and similar ones in New Worldmonkeys. Recently, new species have beenreported from African primates.
Meningonema peruzzii has been describedfrom the leptomeninges of Cercopithecusspp. in Zimbabwe. The microfilariae arealmost identical to those of M. perstans buthave an indistinct sheath that does notstain with Giemsa. Recently, a fourth-stagefemale larva of Meningonema was recov-ered from the cerebrospinal fluid of apatient in Cameroon (Boussinesq et al.,1995). It is possible that the nervous mani-festations reported from humans inRhodesia (now Zimbabwe), with the pres-ence of microfilariae in the spinal fluid,which led Patrick Manson to postulate thatM. perstans was the cause of sleeping sick-ness, was due to this species.
Microfilariae found in the blood of apatient in Thailand may be of Mansonelladigitatum (Sucharit, 1988).
Mansonella streptocerca(Macfie and Corson, 1922) Orihel and
Eberhard, 1982
SYNONYMS
Agamofilaria streptocerca Macfie andCorson, 1922; Acanthocheilonema streptocerca Faust, 1949; Dipetalonemastreptocerca Peel and Chardome, 1946;Tetrapetalonema streptocerca Chabaud andBain, 1976.
DISEASE
Streptocercal filariasis or filariosis; strepto-cerciasis or streptocercosis.
GEOGRAPHICAL DISTRIBUTION
Confined to the rainforest areas of Benin,Cameroon, Central African Republic, Congo,Congo Republic, Côte d’Ivoire, EquatorialGuinea, Gabon, Ghana, Nigeria, Togo andwestern Uganda.
Map 12. Distribution of Mansonella streptocerca.

224 Chapter 5
LOCATION IN HOST
The adults are tightly coiled in the subcu-taneous tissues of the dermis, microfilariaeoccur in the skin (Fig. 110).
MORPHOLOGY
Adult worms are very thin and femalesmeasure 27 mm � 0.08 mm. Males havebeen recovered only once from patients inthe Congo Republic in the dermal collagen(Meyers et al., 1977) and measured 17 mm� 0.05 mm. Microfilariae are unsheathedand measure 180–240 µm � 3–4 µm. Thetail is tapered and curved to form a hook; itends bluntly and has a quadrate nucleus towithin 1 µm of its bifid end. The cuticle isstriated.
LIFE CYCLE
Complete development has been observedexperimentally in the midge Culicoides gra-hami. Ingested microfilariae moved rapidlyto the thoracic muscles and moulted twice,and infective third-stage larvae, measuring574 µm, were seen in the mouth-parts on the8th day. Natural infection rates of up to 1%have been found in these insects.
CLINICAL MANIFESTATIONS AND PATHOGENESIS
Used to be regarded as non-pathogenic, butit is responsible for mild pruritus, withunpigmented spots (in about 60% of cases),papules and oedema, and thickening of theskin has been reported. Infection is oftenmistaken for onchocerciasis and sometimesleprosy.
DIAGNOSIS
By finding the microfilariae in skin snipstaken in the shoulder region. Living speci-mens in saline can be differentiated frommicrofilariae of Onchocerca by their shiv-ering mode of activity in contrast to theactive twisting of the latter. In stainedsmears they are thinner with a hooked andbluntly rounded tail, small caudal spaceand non-overlapping anterior nuclei (Figs.89 and 102).
TREATMENT
DEC (2–6 mg kg�1 body weight day�1 for 21days) kills microfilariae and probablyadults but can cause unpleasant itching.Ivermectin (150 µg kg�1) effectively removesmicrofilariae for long periods.
Fig. 110. Section of subcutaneous tissues with adult M. streptocerca above and microfilariae below(compare with Figs 105 and 107 showing microfilariae of O. volvulus).

The Nematodes 225
EPIDEMIOLOGY
Infection rates were 36% in children inwestern Uganda, 0.4% in Nigeria (BauchiState). M. streptocerca is transmitted byCulicoides grahami and possibly by C. mil-nei, so the epidemiology is similar to thatof M. perstans.
ZOONOTIC ASPECTS
Adults have been found in chimpanzeesand gorillas in Congo Republic, but it isunlikely that they act as reservoir hosts.
Sheathless microfilariae of Mansonella rod-haini, measuring 300 µm � 2.2 µm, havebeen recovered from skin snips taken fromboth sexes in Gabon. It was impossible todetermine pathogenicity since most alsohad M. streptocerca and M. perstans(Richard-Lenoble et al., 1988). It is a naturalparasite of chimpanzees in Congo Republic.
Mansonella ozzardi (Manson, 1897) Faust, 1929
SYNONYMS
Filaria demarquayi Manson, 1897; F.ozzardi Manson, 1897; F. tucumana Biglieriand Araoz, 1917.
DISEASE
Mansonelliasis or mansonellosis ozzardi,Ozzard’s filariasis or filariosis.
GEOGRAPHICAL DISTRIBUTION
Central America, including Mexico(Yucatán) and Panama; South America,including northern Argentina, Bolivia,Brazil, Colombia, Dominican Republic,
French Guiana, Guyana, Surinam andVenezuela; Caribbean, including DesiradeIsland, Dominica, Guadeloupe, Haiti,Martinique, St Lucia, St Vincent andTrinidad (see Map 9, p. 203).
LOCATION IN HOST
Adults inhabit the abdominal mesenteriesand subperitoneal tissues, microfilariae cir-culate in the blood.
MORPHOLOGY
Adult females from monkeys measure49 mm � 0.15 mm, with a smooth cuticleand two lappets at the caudal extemity,while males measure 26 mm � 0.07 mm.
The unsheathed microfilariae in theblood are similar in size to those of M. per-stans, measuring 175–240 µm � 4.5 µm.They differ in that the posterior end of thebody is free of nuclei for 3–4 µm, givingthe appearance of a sharp tail (Fig. 111).
LIFE CYCLE
The microfilariae in the blood show noperiodicity.
The intermediate host in St Vincent isthe minute (1 mm) female midge Culicoidesfurens, in Trinidad C. phlebotomus andexperimentally in Argentina C. paraensis.However, in Brazil the vector is a memberof the black-fly Simulium amazonicumcomplex. In this area microfilariae are alsofound in the skin as well as the blood.
The microfilariae penetrate the stomachwall of the insect and develop in the tho-racic muscles, moulting twice. Infective,third-stage larvae migrate to the head andproboscis by 8 days. Nothing is known of the
Fig. 111. Microfilaria of M. ozzardi.

226 Chapter 5
development in humans, but it has recentlybeen followed experimentally in patas mon-keys, in which patency took 163 days.
CLINICAL MANIFESTATIONS AND PATHOGENESISThe majority of cases appear to be symp-tomless, but allergic reactions have beenreported, particularly from Brazil. Theseinclude pruritus, joint pains, headache,hypopigmented blotchy papules andenlarged and painful inguinal lymph nodes,accompanied by a blood eosinophilia.
DIAGNOSISBy finding the characteristic microfilariaein stained blood smears.
TREATMENTDEC (2–6 mg kg�1 body weight for 21 days)kills microfilariae for a period. Ivermectinreduces microfilarial numbers greatly, butthey often return after a few years. Side-effects are common but minor.
EPIDEMIOLOGY AND ZOONOTIC ASPECTSInfection is most common in Amerindiansliving in rainforest areas. In Brazil it isfound particularly in outdoor workers ofboth sexes but is rare in children under 8.
Infection rates of over 50% in theRoraima area of Brazil, 81–94% over theborder in Venezuela and 22% in Trinidadhave recently been reported. Older figuresare 45% in Panama, 96% in Amerindiansin forests of southern Colombia, 85% inYucatán (Mexico) and 4–15% in Surinam.
Little is known of the epidemiology ofthis parasite in the Amazon basin.Although no reservoir host is known, mon-keys can be experimentally infected.
Accidental filarial infections
DIROFILARIAThose members of the genus Dirofilariathat have been found as parasites ofhumans are all natural parasites of dogs or,more rarely, primates in tropical and sub-tropical areas of the world. They can bedivided into two groups, depending ontheir structure and on their location in thebody:
1. Subgenus Dirofilaria: Dirofilaria (Diro-filaria) immitis, D. (D.) magalhaesi, D. (D.)louisianensis and D. (D.) spectans. Theseparasites have been recovered from thelungs, heart and blood-vessels and are rela-tively thin (diameter 0.1 mm) and imma-ture with a smooth cuticle.2. Subgenus Nochtiella: Dirofilaria (Noch-tiella) repens, D. (N.) tenuis (D. (N.) con-junctivae is a synonym of either of these),D. (N.) striata, D. (N.) ursi. Members of thisgroup from humans have also been recov-ered from the subcutaneous or conjunctivaltissues. They are usually stouter (0.2–0.5 mm in diameter) than those in the firstgroup and have longitudinal ridges in thecuticle with transverse striations.
Dirofilaria immitis (Leidy, 1856). This is animportant and common parasite of the rightventricle and pulmonary artery of the dog inmost warm parts of the world and is trans-mitted by both anopheline and culicinemosquitoes. Immature or unfertilized adultworms have been found about 230 times inlung abscesses or in the pulmonary artery ofhumans, but microfilariae are not present inthe peripheral blood. They usually cause apulmonary infarct, which is seen on X-rayand often mistakenly removed as a neo-plasm. In sections the parasite can be recog-nized by the smooth laminated cuticle, longmuscle cells, broad lateral cords and, inmature females, by the typical filarial twinuteri (Fig. 112). In the dog this is a largerfilaria than D. repens (250–300 mm � 1 mmas against 100–170 mm � 0.4–0.7 mm), butspecimens from humans are usually veryimmature. One case of a gravid female ofwhat was probably D. immitis was reportedfrom the lung of a man who had lymphoidleukaemia (Beaver et al., 1990).
Dirofilaria repens Railliet and Henry, 1911.This species is a natural parasite of thesubcutaneous tissues of dogs in Europe,Asia and possibly Africa and is transmittedby mosquitoes, particularly Aedes (proba-bly A. albopictus in Italy). It has beenreported about 400 times from 30 coun-tries, most (168) from Italy (Muro et al.,1999). There are also a few cases reportedeach year from Spain, France and Russia

The Nematodes 227
(Pampiglione et al., 1995). D. repens inhumans produces a painful migratory sub-cutaneous nodule, which at biopsy shows athreadlike parasite (0.3 mm in diameter) inthe centre of an inflammatory reaction withmany tissue eosinophils (Fig. 113). Manycases are under the conjunctiva or aroundthe eye, probably because these are mostlikely to be diagnosed. A technique usingPCR amplification of DNA can differentiatebetween D. immitis and D. repens infec-tions (Faria et al., 1996).
Dirofilaria tenuis Chandler, 1942. About 40human cases have been reported from theUSA, mostly from women in Florida. Thenatural host is the racoon and 14–45% werefound to be infected in Florida, with mos-quito intermediate hosts. It causes subcuta-neous lesions, often around or in the eye,similar to those of D. repens (Beaver, 1987).
Dirofilaria striata (Molin, 1858). Thisspecies is a natural parasite of various wildcats and has been found a few times in the
Fig. 113. Section of breast from a woman with adult of D. repens, which has evoked a granulomatousreaction. Note ridges on the cuticle of the worm.
Fig. 112. Section of lung with immature Dirofilaria immitis in a branch of the pulmonary artery. Apulmonary infarct may follow the death of the worm.

228 Chapter 5
subcutaneous tissues and eye of humans inthe USA. It is transmitted by mosquitoes.
Dirofilaria ursi Yamaguti, 1941. This is anatural parasite of the bear and subcuta-neous lesions have been found very rarelyin humans in Canada and the USA. It istransmitted by the black-fly Simulium.
Superfamily Dracunculoidea
Dracunculus medinensis (Linnaeus, 1758) Gallandant, 1773
SYNONYMS
Gordius medinensis Linnaeus, 1758;Filaria medinensis L.; Fuellebornius medi-nensis Leiper, 1926.
DISEASE AND COMMON NAMES
Dracunculiasis or dracunculosis, dracontia-sis; guinea or medina worm, le dragonneu,filaire de Medine.
LOCAL NAMES
Farentit (Arabic), Mfa (Fanti and Twi),Kurkunu (Hausa), Naru (Hindi), Reshteh orPiyook (Persian), Rishta (Uzbek), Orok alMai (Yemen), Sobiya (Yoruba).
GEOGRAPHICAL DISTRIBUTION
Dracunculus is now found only in thecountries of West and Central Africa just
south of the Sahara (Benin, Burkina Faso,Central African Republic, Cote d’Ivoire,Ghana, Mali, Mauritania, Niger, Nigeria,Sudan, Togo and Uganda). There are alsovery small foci in Cameroon and Ethiopiaand possibly still in Chad, Kenya andSenegal. Infection has been officially elimi-nated from the formerly endemic areas inIndia (1999) and Pakistan (1994) in the lastfew years and the disease has probablynow vanished from Saudi Arabia andYemen.
LOCATION IN HOST
Adult females emerge from the subcuta-neous tissues, usually of the foot or lowerlimbs but sometimes from any part of thebody.
MORPHOLOGY
The mature female measures 500–800 mm �1.0–2.0 mm. The mouth has a triangular ovalopening surrounded by a quadrangularcuticularized plate, with an internal circle offour double papillae. The vulva openshalfway down the body but is non-func-tional in the mature worm. The uterus hasan anterior and a posterior branch and isfilled with 1–3 million embryos; it fills theentire body cavity (pseudocoel), the gutbeing entirely flattened.
Males recovered from experimentalinfections in animals measure 15–40 mm� 0.4 mm (Fig. 114). The tail has 4 (3–6)
Fig. 114. Adult female and smaller male of D. medinensis.

The Nematodes 229
pairs of preanal and 4–6 pairs of postanalpapillae; the subequal spicules are490–750 µm long, with a gubernaculummeasuring about 117 µm. Males remain inthe connective tissues surrounding deepermuscles, usually in the thoracic region,and are either absorbed or eventually cal-cify; they have only doubtfully been recov-ered in human infections.
LIFE CYCLE
The mature female worm moves to the sur-face of the skin and forms a blister, whichbursts. The first-stage larvae are expelledfrom the ruptured uterus when the affectedpart of the body (usually the foot or leg)comes into contact with water. The larvaeon average measure 643 (490–737) µm �23 (18–24) µm and have a fully formed gut,although they do not feed. The tail is longand pointed and the cuticle is striated. Thelarvae in water are very active, their thrash-ing movements resembling those of a free-living nematode (Fig. 115). They live for4–7 days in pond water and for furtherdevelopment have to be ingested by vari-ous predatory species of small (1–2 mm)microcrustacean cyclopoids. Intermediatehosts belong to the genera Mesocyclops (M.
aequatorialis and M. kieferi), Metacyclops(M. margaretae) and Thermocyclops (T.crassus, T. incisus, T. inopinus and T.oblongatus). The larvae penetrate the gutwall of cyclops (Fig. 116) and, after twomoults in the haemocoel, reach the infec-tive third stage in 14 days at 24°C. Theinfective larva measures on average 450 µm� 14 µm and has a short bilobed tail. Wheninfected cyclops are ingested in drinkingwater, the released larvae burrow into thewall of the duodenum, migrate across theabdominal mesenteries and penetratethrough the abdomen and thorax in about15 days. There is no increase in size duringthis early migration but, after two moults,the sexually mature worms meet and matein the subcutaneous tissue, approximately100 days after infection. After fertilization,the males become encapsulated and even-tually die. They are usually absorbed,although occasionally calcified specimenshave been seen on X-ray. The females movedown the muscle planes, reaching theextremities 8–10 months after infection; by10 months the uterus contains fully formedlarvae. The mature females emerge fromthe tissues about 1 year after ingestion oflarvae.
Fig. 115. First-stage larvae of Dracunculus medinensis in water (phase contrast) after being expelled byadult female (actual size 0.66 mm).

CLINICAL MANIFESTATIONS
There are usually none while the femaleworms are moving freely through the con-nective tissues, but when about to emergeto the surface a few larvae are released intothe subdermis through a rupture at theanterior end of the worm (the uterus isnon-functional). The host reaction resultsin the formation of a burning, painful, blis-ter, which bursts in a few days to give ashallow ulcer, and there is then a markedinflammatory response against the cuticleof the entire worm, preventing its removal
(Fig. 117). In many patients (30–80%),urticaria, sometimes accompanied by fever,giddiness, gastrointestinal symptoms,dyspnoea and infraorbital oedema, appearsthe day before the blister forms but van-ishes in a few hours. The blister is the firstsign in about 60% of cases. It is minutewhen first noticed but may grow to a fewcentimetres in diameter before it bursts,usually in 1–3 days. The formation of theblister is accompanied by local itching andoften an intense pain, which can berelieved by immersion of the affected partin cold water. A portion of the worm isextruded through the ulcer formed by thebursting of the blister, the edges of whichconsist of pinkish granulation tissue. Insimple cases the anterior end of the wormis exposed by sloughing of the white cen-tral eschar of the ulcer and more of theworm protrudes, particularly after immer-sion in water; after many thousands of lar-vae are expelled the portion of wormoutside the body becomes flaccid.Complete expulsion of the worm occurs,on average, in 4 weeks and the lesion thenrapidly resolves.
Secondary bacterial infection of thetrack of an emerging worm occurs in about50% of cases and patients then becomeseriously incapacitated (Fig. 118). In astudy in an area of Nigeria, 58% of cases,almost all of whom belonged to the 14–40-
230 Chapter 5
Fig. 116. Cyclops with larvae of Dracunculus inthe haemocoel.
Fig. 117. Section of subcutaneous tissues with portion of an adult female. Removed just after blisterformation so that there is a strong tissue reaction.

The Nematodes 231
year-old working population or wereschoolchildren, were disabled for an aver-age of 12.7 weeks during the yam and riceharvest time and about 12,000 were perma-nently incapacitated (Smith et al., 1989).Permanent physical impairment occurs inabout 0.5% of infected individuals and inone study in Ghana 28% of patients hadcontinuing pain 12–18 months after infec-tion (Hours and Cairncross, 1994), while inanother study in Benin there was 0.3%mortality from tetanus and septicaemia(Chippaux and Massougbodi, 1991).Infection is a major cause of school absen-teeism and the parasite is unusual amongparasitic helminths as it does not appear toconfer any immune response, so that indi-viduals can be reinfected year after year.
Sometimes female worms fail to reachthe surface, usually when unfertilized, andbecome encysted and calcify and are thenonly apparent on X-ray. This occurs in a
proportion of apparently uninfected per-sons who live in an endemic area.Occasionally an encysted worm may causemore serious effects if in an unusual site;these include constrictive pericarditis,paraplegia, lymph node abscess and uro-genital complications.
PATHOGENESIS
The blister fluid consists of a bacteriologi-cally sterile fluid containing lymphocytes,neutrophils, eosinophils and larvae.
In a proportion of cases the femaleworm bursts in the tissues before emer-gence, releasing many thousands of larvae.This results in an intense tissue reaction,with myositis and formation of an abscesscontaining up to 0.5 l of pus, which canlead to chronic ulcerations, bubo or epi-didymo-orchitis in males. Adult femalessometimes enter joints and liberate larvaeinto the synovial fluid, causing oedema,congestion and plasma-cell infiltration ofthe synovial membrane. This can lead toarthritis or fibrous ankylosis of joints andcontractures of tendons, but in most casesthe condition resolves eventually.
Once larvae have been released fromthe emerging female worm, there is astrong adhesive reaction at the cuticlealong the whole length of the worm. In theearly stages, the tissue reaction consists ofinflammatory cells and is followed by a for-eign-body giant-cell reaction. This makesextraction of the worm difficult andincreases the chances of secondary bacter-ial infection along the track of the worm,with cellulitis.
In many endemic areas, the track of aguinea worm on the foot provides animportant mode of entry for tetanus spores.
DIAGNOSIS
Clinical and parasitological. Patients in anendemic area usually have no doubt of thediagnosis as soon as, or even before, thefirst signs appear. Local itching, urticariaand a burning pain at the site of a smallblister are usually the first signs of infec-tion, although sometimes a palpable andsometimes moving worm may have been
Fig. 118. Female of D. medinensis emerging fromankle of woman. There is secondary bacterialinfection up the track of the worm.

232 Chapter 5
recognized earlier. The blister bursts inabout 4 days and active larvae, obtained byplacing cold water on the resulting smallulcer, can be recognized under a low-pow-ered microscope.
Immunological. Not useful in practice.ELISA, dot ELISA and SDS-PAGE/Westernblotting worked well for patent infections.The most specific reaction appears to be fordetection of IgG4 antibodies and might beable to detect infections up to 6 monthsbefore patency (Saroj-Bapna andRenapurkar, 1996; Bloch and Simonsen,1998). The fluorescent antibody test usingdeep-frozen first-stage larvae was shown todemonstrate prepatent infections in mon-keys 30 years ago. No evidence has beenfound for the presence of circulating anti-gens (Bloch et al., 1998).
TREATMENT
Surgery. Guinea worms have been woundout on sticks since antiquity (e.g. in the RgVeda of about 1350 BC and possibly theyrepresent the fiery serpents afflicting theIsraelites in the wilderness, which Mosesrecommended should be set upon a pole).Provided that bacterial infection or othercomplications have not occurred, regularwinding out of the worm on a small stick,combined with sterile dressing and acri-flavine cream, usually results in completeexpulsion in 3–4 weeks with little pain.This should be commenced as soon afteremergence as possible.
Surgical removal of mature femaleworms before emergence is sometimes pos-sible using a small incision. At this stagethere is no host reaction to the cuticle ofthe worm, as occurs once emergence com-mences, but care must be taken that theworm is not wound around a ligament.
Chemotherapy. There is no evidence thatany chemotherapeutic agent has a directaction against guinea worms. However,many compounds, including thiabend-azole, niridazole, metronidazole, mebend-azole and albendazole have been reportedas hastening the expulsion of worms and
may act as anti-inflammatory agents.Ivermectin had no action in one field trialor against pre-emergent D. insignis inferrets.
EPIDEMIOLOGY
Dracunculiasis is typically a disease ofpoor rural communities and is the onlyinfectious agent that is transmitted entirelythrough drinking water. In all areas trans-mission is markedly seasonal and in mostparts of Africa (and formerly in India) themaximum incidence coincides with theplanting season, resulting in great eco-nomic hardship. In semi-desert (Sahel)areas of Africa (Burkina Faso, Chad, north-ern Cameroon, Mauritania, Niger, northernNigeria, Senegal and Sudan), drinkingwater is obtained from ponds during therainy season but from deep wells for therest of the year when the ponds are dry.However, in the humid (guinea) savannahregions of West Africa where rainfallexceeds 150 cm year�1 (Benin, Coted’Ivoire, Ghana, southern Nigeria andTogo), there is no transmission during therainy season, when ponds turn intostreams and cyclops densities are lowbecause of the large volume and turbidityof the water. Similarly, infection was high-est during the dry season in step wells inIndia.
The life history of the parasite is welladapted to provide the maximum chancesof transmission, as the female takes almostexactly a year to mature and release its lar-vae at the optimum period each year.
PREVENTION AND CONTROL
The United Nations World HealthAssembly in 1991 endorsed a campaign toeradicate dracunculiasis from the worldand originally set a date of the end of 1995.Although this date has had to be postponedby another 10 years, the prevalence and dis-tribution of the infection have diminishedmarkedly in the last few years and it hasalready been eliminated from Asia. The sit-uation in most African countries is not sofar advanced, but active campaigns arebeing carried out in all endemic countriesand some are free of the disease, while most

are at the stage of case containment,whereby all infected individuals can bequickly identified and stopped from enter-ing ponds.
There are various possible interventionsonce active surveillance measures havebeen initiated, based on knowledge of thelife cycle of the parasite. It has been esti-mated that passive surveillance identifiesabout 2% of cases.
The provision of safe drinking water,principally from tube-wells, is a priority inrural areas of Africa and Asia, and assis-tance is being provided by many aid agen-cies in order to prevent other water-bornediseases, such as typhoid, infantile diar-rhoea, etc., as well as guinea worm. It hasrecently been estimated that 1100 millionpeople (about 21% of the world’s popula-tion) do not have access to clean water.
There are various interventions possiblebut the use of chemotherapy has no part incontrol, except sometimes in aiding patientcompliance. Interventions which are beingmade include the following.
1. Filtering or boiling all drinking-water.Boiling water is not usually feasible,because wood fuel is a scarce commodity,but filters are playing an important role inall countries now mounting campaigns.The manufacturer of a monofilament nylonmaterial has provided enough to providefilter nets for all endemic villagers. Thesenets are long-lasting, have a regular poresize, are easily washed and dry quickly andare being widely used. 2. Persuading or preventing infected per-sons with an emerging worm from enteringponds used for drinking water. The identi-fication and bandaging of very early lesionscan also help to prevent subsequentimmersion. The cooperation of local teamsand of schoolchildren can be useful in con-vincing infected persons of the need for achange in behaviour.3. Treating water sources. The chemicaltreatment of ponds can prove a useful adju-vant to other measures, particularly whenthere is only a low level of transmissionremaining. The insecticide temephos haslow toxicity to mammals and fish and at 1
p.p.m. kills cyclops for 5–6 weeks but, inareas with a long transmission season, hasto be added a few times a year. The amountof temephos estimated to be needed fortotal eradication in Africa has beendonated by the manufacturer.
Health education interventions by localhealth workers are a very important com-ponent of any control or eradication cam-paign, and village development committeesare able to assist in other primary healthproblems.
Guinea worm was eliminated from theBokhara area of Uzbekistan in the early1930s and from the south of Iran, whereinfection was confined to large covered cis-terns (known as ‘birkehs’), in the 1970s. Itis interesting to note that infection has notreturned to Bokhara in spite of the fact thatstill only 33% of the population has pipedwater (Cairncross et al., 2002). It was elimi-nated from Pakistan in 1994, India in 1997,Kenya and Senegal in 1996 and Cameroon,Chad and Yemen by 1999. There are alsovery few cases left in Benin, Burkina Faso,Central African Republic, Côte d’Ivoire,Ethiopia, Mali, Mauritania, Niger, Togo andUganda. However, there are three countrieswhich still have appreciable numbers ofcases. These are Nigeria, which had 11,617cases in 1999 (but 650,000 cases in 1988),Ghana, with 5524 cases in 1999 (but180,000 cases in 1989), and Sudan, with56,226 cases reported in 1999. The appar-ent number of cases went up substantiallyin 2000 in Ghana, which demonstrates thedifficulties in identifying the many hun-dreds of small isolated endemic villages.However, there are now intensive cam-paigns in both Ghana and Nigeria and it islikely that elimination will follow in thenext few years. The future for eliminationin Sudan is more uncertain. Because ofpolitical problems it has been difficult toactively survey, let alone treat, manyinfected villages and there is also the prob-lem of migrants from Sudan reintroducinginfection into neighbouring countries.Thirty-seven other countries where the dis-ease used to be present in historical timeshave been certified as free by the WHO.
The Nematodes 233

ZOONOTIC ASPECTS
Female worms of the genus Dracunculushave been reported as emerging from awide range of mammals and reptiles frommany parts of the world, both endemic andnon-endemic for the human disease(Muller, 1971). Those found in reptilesclearly belong to other species but the situ-ation in regard to those in mammals is notclear. For instance guinea worm is commonin wild carnivores in North America andwas named D. insignis by Leidy in 1858but there is very little morphological evi-dence for separating this from the humanspecies.
There have been three documented casesof human zoonotic infections, from Japan in1986 (Kobayashi et al., 1986) and fromKorea and Indonesia in 1926. In both casesthe patients had eaten raw freshwater fishwhich have been proved experimentally tobe capable of acting as paratenic hosts.
In most highly endemic areas occasionalinfections, principally in dogs and occa-sionally other mammals, have beenreported, but there is no evidence that theyhave any part in maintaining transmission.What is presumably the human parasitecan still be found in dogs in the formerlyendemic areas of Tamil Nadu in India andUzbekistan, but no new human cases havebeen reported. Infection has also beenfound in dogs and cats in previously non-endemic Central Asian Republics;Azerbaijan, Kazakhstan and possiblyTurkmenia (Cairncross et al., 2002). Therehas also been a recent report of D. medi-nensis from a cat in China (Fu et al., 1999)and it is very likely that in many parts ofthe world there are widespread but under-reported animal cycles completely inde-pendent of human infection. However, it isunlikely that zoonotic infections have anyimportance in human transmission.
234 Chapter 5
References
Akahane, H., Sano, M. and Kobayashi, M. (1998) Three cases of human gnathostomiasis caused byGnathostoma hispidum, with particular reference to the identification of parasitic larvae.Southeast Asian Journal of Tropical Medicine and Public Health 29, 611–614.
Akue, J.P., Egwang, T.G. and Devaney, E. (1994) High levels of parasite-specific IgG4 in the absence ofmicrofilaraemia in Loa loa infection. Tropical Medicine and Parasitology 45, 246–248.
Amaral, G., Dreyer, G., Figueredos-Silva, J., Noroes, J., Cavalcanti, A., Samico, S.C., Santos, A. andCoutinho, A. (1994) Live adult worms detected by ultrasonography in human bancroftian filaria-sis. American Journal of Tropical Medicine and Hygiene 50, 753–757.
Ando, K. (1989) Two cases of infection with Gnathostoma nipponicum in humans [English sum-mary]. Saishin Igaku 44, 804–806.
Ando, K., Hatsushika, R., Akahane, H., Taylor, D., Mura, K. and Chinzei, Y. (1991) Gnathostoma nip-ponicum infection in the past human cases in Japan. Japanese Journal of Parasitology 40,184–186.
Andrews, J.R.H., Ainsworth, R. and Abernethy, D. (1994) Trichinella pseudospiralis in humans:description of a case and its treatment. Transactions of the Royal Society of Tropical Medicineand Hygiene 88, 200–203.
Baird, J.K. and Neafie, R.C. (1988) South American brugian filariasis: report of human infectionacquired in Peru. American Journal of Tropical Medicine and Hygiene 39, 185–188.
Baird, J.K., Neafie, R.C., Lanoie, L. and Connor, D.H. (1987) Adult Mansonella perstans in the abdom-inal cavity of nine Africans. American Journal of Tropical Medicine and Hygiene 37, 578–584.
Bangs, M.J., Purnomo and Andersen, E.M. (1994) A case of capillariasis in a highland community ofIrian Jaya, Indonesia. Annals of Tropical Medicine and Parasitology 88, 685–687.
Beaver, P.C. (1989) Intraocular filariasis: a brief review. American Journal of Tropical Medicine andHygiene 40, 40–45.
Beaver, P.C., Orihel, T.C. and Leonard, G. (1990) Pulmonary dirofilariasis: restudy of worms reportedgravid. American Journal of Tropical Medicine and Hygiene 43, 167–169.
Bessonov, A.S. (1998) Taxonomic position of nematodes of the genus Trichinella Railliet, 1895. [inRussian, English Summary]. Meditsinskaya Parazitologiya I Parazitarnye Bolezni 1, 3–6.

Bloch, P. and Simonsen, P.E. (1998) Studies on immunodiagnosis of dracunculiasis. I. Detection ofspecific serum antibodies. Acta Tropica 70, 73–86.
Bloch, P., Vennervald, B.J. and Simonsen, P.E. (1998) Studies on immunodiagnosis of dracunculiasis.II. Search for circulating antigens. Acta Tropica 70, 303–315.
Botero, D., Aguledo, L.M. and Uribe, F.J. (1984) Intraocular filaria: a Loaina species from man inColombia. American Journal of Tropical Medicine and Hygiene 33, 578–582.
Boussinesq, M., Bain, O., Chabaud, A.G., Gawdon-Wendel, N., Kamno, J. and Chippaux, J.P. (1995) Anew zoonosis of the cerebrospinal fluid of man, probably caused by Meningonema peruzzii, afilaria of the central nervous system of Cercopithecidae. Parasite 2, 173–176.
Bradley, J.E. and Unnasch, T.R. (1996) Molecular approaches to the diagnosis of onchocerciasis.Advances in Parasitology 37, 58–106.
Bradley, J.E., Atogho, B.M., Elson, L., Stewart, G.R. and Boussinesq, M. (1998) A cocktail of recombi-nant Onchocerca volvulus antigens for serological diagnosis with the potential to predictendemicity of onchocerciasis infection. American Journal of Tropical Medicine and Hygiene 59,877–882.
Brooker, S. and Michael, E. (2000) The potential of geographical information systems and remotesensing in the epidemiology and control of human helminth infections. Advances in Parasitology47, 246–288.
Bruschi, F. (1999) How can Trichinella escape host immune response? Helminthologia 3, 179–184.Cairncross, S., Muller, R. and Zagarian, N. (2002) Dracunculiasis (guinea worm disease) and its eradi-
cation. Clinical Microbiology Reviews (in press).Capo, V. and Despommier, D.D. (1996) Clinical aspects of infection with Trichinella spp. Clinical
Microbiology Reviews 9, 47–54.Chan, S.W. and Ko, R.C. (1990) Serodiagnosis of human trichinosis using a gel filtration antigen and
indirect IgG-ELISA. Transactions of the Royal Society of Tropical Medicine and Hygiene 84,721–722.
Chippaux, J.-P. and Massougbodi, A. (1991) Evaluation clinique et épidemiologique de la dracuncu-lose au Benin. Médecine Tropicale 51, 269–274.
Chippaux, J.-P., Ernould, J.C., Gardon, J., Gardon-Wendel, N., Chadre, F. and Barbieri, N. (1992)Ivermectin treatment of loiasis. Transactions of the Royal Society of Tropical Medicine andHygiene 86, 289.
Choe, G., Lee, H.S., Seo, J.K., Chai, J.Y., Lee. S.H., Eom, K.S. and Chi, J.G. (1993) Hepatic capillaria-sis: first report in the Republic of Korea. American Journal of Tropical Medicine and Hygiene 48,610–625.
Davies, J.B. (1995) A rapid staining and clearing technique for detecting filarial larvae in alcohol-preserved vectors. Transactions of the Royal Society of Tropical Medicine and Hygiene 89, 280.
Denham, D.A. and Fletcher, C. (1987) The cat infected with Brugia pahangi as a model of humanfilariasis. CIBA Foundation Symposium 127, 225–235.
Dissanayake, S., Watawana, L. and Piessens, W.F. (1995) Lymphatic pathology in Wuchereria ban-crofti microfilaraemic infections. Transactions of the Royal Society of Tropical Medicine andHygiene 89, 517–521.
Dreyer, G., Amaral, F., Noroes, J., Medeiros, Z. and Addiss, D. (1995) A new tool to assess the adulti-cidal efficacy in vivo of antifilarial drugs for bancroftian filariasis. Transactions of the RoyalSociety of Tropical Medicine and Hygiene 89, 225–226 (also 1994, 88, 558).
Dreyer, G., Noroes, J. and Addiss, D. (1997) The silent burden of sexual disability associated withlymphatic filariasis. Acta Tropica 63, 57–60.
Dreyer, G., Noroes, J., Figueredo-Silva, J. and Piessens, W.F. (2000) Pathogenesis of lymphatic diseasein bancroftian filariasis: a clinical perspective. Parasitology Today 16, 544–548.
Dujardin, J.P., Fain, A. and Maertens, K. (1982) Survey on the human filariasis in the region ofBwamanda in Northwest Zaire. Annales de la Société belge de Médecine tropicale 62, 315–342.
Dye, C. (1992) Does facilitation imply a threshold for the eradication of lymphatic filariasis?Parasitology Today 8, 109–110.
Eberhard, M.L., Hurwitz, H., Sun, A.M. and Coletta, D. (1989) Intestinal perforation by larvalEustrongylides (Nematoda: Dioctophymatoidae) in New Jersey. American Journal of TropicalMedicine and Hygiene 40, 648–650.
The Nematodes 235

Faria, G., Lanfrancotti, A., Della Torre, A., Cancrini, G. and Coluzzi, M. (1996) Polymerase chainreaction-identification of Dirofilaria repens and Dirofilaria immitis. Parasitology 113, 567–571.
Freedman, D.O., de Almeida-Filho, P.J., Besh, S., Maia-e-Silva, M.C., Braga, C. and Maciel, A. (1994)Lymphoscintigraphic analysis of lymphatic abnormalities in symptomatic and asymptomatichuman filariasis. Journal of Infectious Diseases 170, 927–933.
Fu, A.Q., Tao, J.P., Wang, Z.X., Jiang, B.L., Liu, Y.X. and Qui, H.H. (1999) Observations on the mor-phology of Dracunculus medinensis from a cat in China [in Chinese, English summary]. ChineseJournal of Zoonoses 15, 35–38.
Gyapong, J.O. (1998) The relationship between infection and disease in Wuchereria bancrofti infec-tion in Ghana. Transactions of the Royal Society of Tropical Medicine and Hygiene 92, 390–392.
Hasegawa, H., Korenaga, M., Kumazawa, H. and Imamawa, K. (1996) Mermis sp. found from humanurine in Kochi Prefecture, Japan (Nematoda: Mermithidae). Japanese Journal of Parasitology 45,43–46.
Hours, M. and Cairncross, S. (1994) Long-term disability due to guinea worm disease. Transactionsof the Royal Society of Tropical Medicine and Hygiene 88, 559–560.
Hunt, D.J., Muller, R., Hellyar, A.G. and Payne, S.R. (1984) Mermithid in man. Transactions of theRoyal Society of Tropical Medicine and Hygiene 78, 698–700.
International Task Force (1993) Recommendations for disease eradication. CDC Morbidity andMortality Weekly Report 42, 1–38.
Jelinek, T. and Loscher, T. (1994) Human infection with Gongylonema pulchrum: a case report.Tropical Medicine and Parasitology 45, 329–330.
Kapel, C.M.O., Webster, P., Lind, P., Pozio, E., Henriksen, S.A., Murrell, K.D. and Nansen, P. (1998)Trichinella spiralis, T. britovi and T. nativa: infectivity, larval distribution in muscle, and anti-body response after experimental infection of pigs. Parasitology Research 84, 264–271.
Kettle, D.S. (1995) Medical and Veterinary Entomology. CAB International, Wallingford, UK.Kobayashi, A., Katatura, A., Hamada, A. and Suzuki, T. (1986) Human case of dracunculiasis in
Japan. American Journal of Tropical Medicine and Hygiene 35, 159–161.Kraivichian, P., Kulkumthorn, M., Yingyourd, P., Akarabovorn, P. and Paireepai, C.C. (1992)
Albendazole for the treatment of human gnathostomiasis. Transactions of the Royal Society ofTropical Medicine and Hygiene 86, 418–421.
Kumari, S., Lillibridge, C.D., Bakeer, M., Lowrie, R.C., Jr, Jayaraman, K. and Philipp, M.T. (1994)Brugia malayi: the diagnostic potential of recombinant excretory/secretory antigens.Experimental Parasitology 79, 489–505.
Lammle, P.J., Reiss, M.D., Dimock, K.A., Streit, T.G., Roberts, J.M. and Eberhard, M.L. (1998)Longitudinal analysis of the development of filarial infection and antifilarial immunity in acohort of Haitian children. American Journal of Tropical Medicine and Hygiene 59, 217–221.
Lawrence, R.A. (1996) Lymphatic filariasis: what mice can tell us. Parasitology Today 12, 267–271.Ljungstrom, I. (1983) Immunodiagnosis in man. In: Campbell, W.C. (ed.) Trichinella and Trichinosis
Plenum Press, New York, USA, pp. 403–424.Marwi, M.A., Omar, B., Mohammod, C.G. and Jeffrey, J. (1990) Anatrichosoma sp. egg and Demodex
folliculorum in facial skin scrapings of Orang Aslis. Tropical Biomedicine 7, 193–194.Meyer, C.G. and Kremsner, P.G. (1996). Malaria and onchocerciasis: on HLA and related matters.
Parasitology Today 12, 179–186.Meyers, W.M., Neafie, R.C., Moris, R. and Bourland, J. (1977) Streptocerciasis: observation of adult
male Dipetalonema streptocerca in man. American Journal of Tropical Medicine and Hygiene 26,1153–1155.
Muller, R. (1971) Dracunculus and dracunculiasis. Advances in Parasitology 9, 73–151.Muller, R. (1987) A Dipetalonema by any other name. Parasitology Today 3, 358–359.Murdoch, M.E., Hay, R.J., McKenzie, C.D., Williams, J.F., Ghalib, H.W., Cousens, S., Abiose, A. and
Jones, B.R. (1993) A clinical classification and grading system of the cutaneous changes inonchocerciasis. British Journal of Dermatology 129, 260–269.
Muro, A., Genchi, C., Cordero, M. and Simón, F. (1999) Human dirofilariasis in the European Union.Parasitology Today 15, 386–389.
Murrell, K.D. and Bruschi, F. (1994) Trichinellosis. In: Sun, T. (ed.) Progress in Clinical Parasitology,Vol. 4. Norton, New York, USA, pp. 117–150.
Murrell, K.D. and Pozio, E. (2000) Trichinellosis: the zoonosis that won’t go quietly. InternationalJournal for Parasitology 30, 1339–1349.
236 Chapter 5

Ngu, J.L., Nbenfou, C., Capuli, E., McMoli, T.E., Perler, F., Mbwagbor, J., Tume, C., Nlatte, O.B.,Donfack, J. and Asonganyi, T. (1998) Novel, sensitive and low-cost diagnostic tests for ‘riverblindness’ – detection of specific antigens in tears, urine and dermal fluid. Tropical Medicine andInternational Health 3, 339–348.
Nopparatana, C., Setasuban, P., Chaicumpa, W. and Tapchaisri, P. (1991) Purification of Gnathostomaspinigerum specific antigen and immunodiagnosis of human gnathostomiasis. InternationalJournal for Parasitology 21, 677–687.
Noroes, J., Addiss, D., Santos, A., Medeiros, Z., Coutinho, A. and Dreyer, G. (1996) Ultrasonographicevidence of abnormal lymphatic vessels in young men with adult Wuchereria bancrofti infectionin the scrotal area. Journal of Urology 156, 409–412.
Nutman, T.B. (ed.) (2000) Lymphatic Filariasis. Imperial College Press, London, UK.Orihel, T.C. and Eberhard, M.L. (1998) Zoonotic filariasis. Clinical Microbiology Reviews 11,
366–381.Ottesen, E.A., Duke, B.O.L., Karam, M. and Behbehani, K. (1997) Strategies and tools for the con-
trol/elimination of lymphatic filariasis. Bulletin of the World Health Organization 75, 491–503.Ottesen, E.A., Ismail, M.M. and Horton, J. (1999) The role of albendazole in programmes to eliminate
lymphatic filariasis. Parasitology Today 15, 382–386.Pampiglione, S., Canestri-Trotti, G. and Rivasi, F. (1995) Human dirofilariasis due to Dirofilaria
(Nochtiella) repens: a review of world literature. Parassitologia 37, 149–193. Pinder, M. (1988) Loa loa – a neglected filaria. Parasitology Today 4, 279–284.Plaisier, A.P., Stolk, W.A., van Oortmarsen, G.J. and Hobbema, J.D.F. (2000) Effectiveness of annual
ivermectin treatment for Wuchereria bancrofti infection. Parasitology Today 16, 298–302.Pozio, E. (1998) Trichinellosis in the European Union: epidemiology, ecology and economic impact.
Parasitology Today 14, 35–38.Pozio, E. and La Rosa, G. (2000) Trichinella murrelli n.sp.: etiological agent of sylvatic trichinellosis
in temperature areas of North America. Journal of Parasitology 186, 134–139.Pozio, E., La Rosa, G., Murrell, K.D. and Lichtenfels, J.R. (1992) Taxonomic revision of the genus
Trichinella. Journal of Parasitology 78, 654–659.Pozio, E., Owen, I.L., la Rosa, G., Sacchi, L., Rossi, P. and Corona, S. (1999) Trichinella papuae n.sp.
(Nematoda), a new non-encapsulated species from domestic and sylvatic swine of Papua NewGuinea. International Journal for Parasitology 29, 1825–1839.
Remme, J.H.F. (1995) The African Programme for Onchocerciasis Control: preparing to launch.Parasitology Today 11, 403–406.
Richard-Lenoble, D., Kombila, M., Bain, O., Chandenier, J. and Mariotte, O. (1988) Filariasis inGabon: human infections with Microfilaria rodhaini. American Journal of Tropical Medicine andHygiene 39, 91–92.
Samba, E.M. (1994) The Onchocerciasis Control Programme in West Africa. World HealthOrganization, Geneva, Switzerland.
Saroj-Bapna and Renapurkar, D.M. (1996) Immunodiagnosis of early dracunculiasis. Journal ofCommunicable Diseases 28, 33–37.
Schlesinger, J.J., Dubois, J.G. and Beaver, P.C. (1977) Brugia-like filarial infection acquired in theUnited States. American Journal of Tropical Medicine and Hygiene 26, 204–207.
Service, M.W. (ed.) (2001) The Encyclopedia of Arthropod-transmitted Infections. CAB International,Wallingford, UK.
Shi, Y.E., Han, J.J., Yang, W.Y. and Wei, D.X. (1988) Thelazia callipaeda (Nematoda: Spirurida): trans-mission by flies from dogs to children in Hubei, China. Transactions of the Royal Society ofTropical Medicine and Hygiene 82, 627.
Smith, G.S., Blum, D., Huttly, S.R.D., Okeke, N., Kirkwood, B.R. and Feachem, R.G. (1989) Disabilityfrom dracunculiasis: effect on mobility. Annals of Tropical Medicine and Parasitology 83,151–158.
Sucharit, S. (1988) Mansonella digitatum in Thailand. Journal of the Medical Association ofThailand 71, 587–588.
Suntharasamai, P., Riganti, M., Chittamas, S. and Desakoru, V. (1992) Albendazole stimulates out-ward migration of Gnathostoma spinigerum to the dermis in man. Southeast Asian Journal ofTropical Medicine and Public Health 23, 716–722.
Taylor, M.J. (2000) Wohlbachia bacteria of filarial nematodes in the pathogenesis of disease and as atarget for control. Transactions of the Royal Society of Tropical Medicine and Hygiene 94,596–598.
The Nematodes 237

Toe, L., Boatin, B.A., Adjami, A., Back, C., Merriweather, A. and Unnasch, T.R. (1998) Detection ofOnchocerca volvulus infection by 0–150 polymerase chain reaction analysis of skin scratches.Journal of Infectious Diseases 178, 282–285.
Toure, F.S., Egwang, T.G., Wahl, G., Millet, R., Bain, O. and Georges, A.J. (1997) Species-specificsequence in the repeat 3 region of the gene encoding a putative Loa loa allergen: a diagnostic toolfor occult loiasis. American Journal of Tropical Medicine and Hygiene 56, 57–60.
Townson, S., Hutton, D., Siemenska, J., Hollick, L., Scanlon, T., Tagboto, S.K. and Taylor, M.J. (2000)Antibiotics and Wolbachia in filarial nematodes: antifilarial activity of rifampicin, oxytetracyclineand chloramphenicol against Onchocerca gutturosa, Onchocerca lienalis and Brugia pahangi.Annals of Tropical Medicine and Parasitology 94, 801–816.
Vanamail, P., Ramalah, K.D., Pani, S.P., Das, P.K., Grenfell, B.T. and Bundy, D.A.P. (1996) Estimationof the fecund life span of Wuchereria bancrofti in an endemic area. Transactions of the RoyalSociety of Tropical Medicine and Hygiene 90, 119–121.
Wahl, G. and Georges, A.J. (1995) Current knowledge on the epidemiology, diagnosis, immunologyand treatment of loiasis. Tropical Medicine and Parasitology 46, 287–291.
Weil, G.J., Lammie, P.J. and Weiss, N. (1997) The ICT filariasis test: a rapid-format antigen test fordiagnosis of bancroftian filariasis. Parasitology Today 13, 401–404.
Whitworth, J.A.G. and Gemade, E. (1999) Independent evaluation of onchocerciasis rapid assessmentmethods in Benue State, Nigeria. Tropical Medicine and International Health 4, 26–30.
WHO (1992) Lymphatic Filariasis and its Control. Technical Report Series 821, World HealthOrganization, Geneva, Switzerland.
WHO (1995) Onchocerciasis and its Control. Technical Report Series 852, World HealthOrganization, Geneva, Switzerland.
WHO (1999) Building Partnerships for Lymphatic Filariasis: Strategic Plan, September 1999. WorldHealth Organization, Geneva, Switzerland. 61 pp.
Williams, S.A. and Johnston, D.A. (with others) (1999) Helminth genome analysis: the current statusof the filarial and schistosome genome projects. Parasitology 118 (suppl.), 519–538.
Wittner, M., Turner, J.W., Jacquette, G., Ash, L.R., Salgo, M.P. and Tanowitz, H.B. (1989)Eustrongylidiasis – a parasitic infection acquired by eating sushi. New England Journal ofMedicine 320, 1124–1126.
Further Reading
Cairncross, S. (1992) Guinea Worm Eradication: a Selected Bibliography. Bureau of Hygiene andTropical Diseases, London, UK.
Chippaux, J.-P. (1994) Le Ver de Guinée en Afrique: Méthodes de Lutte pour l’Èradication. ORSTOM,Paris, France.
Cross, J.H. (1995) Capillaria philippinensis and Trichostrongylus orientalis. In: Farthing, M.J.G.,Keusch, G.T. and Wakelin, D. (eds) Enteric Infection 2: Intestinal Helminths. Chapman and Hall,London, UK, pp. 151–163.
Daengsvang, S. (1980) A Monograph on the Genus Gnathostoma and Gnathostomiasis in Thailand.Seamic, Tokyo, Japan.
Dissanaike, A.S. (1986) A review of Brugia sp. with special reference to Brugia malayi and tozoonotic infections. Tropical Biomedicine 3, 67–72.
Goddard, J. (1996) Physicians’ Guide to Arthropods of Medical Importance, 2nd edn. CRC Press,Boca Raton, Florida, USA.
Grenfell, B.T. and Michael, E. (1992) Infection and disease in lymphatic filariasis: an epidemiologicalapproach. Parasitology 104 (suppl.), S81-S90.
Hinz, E. (1996) Helminthiasen des Menschen in Thailand. Peter Lang, Frankfurt am Main, Germany,pp. 137–141.
Hopkins, D.R. and Ruiz-Tiben, E. (1991) Strategies for dracunculiasis eradication. Bulletin of theWorld Health Organization 69, 533–540.
Muller, R. (1979) Guinea worm disease; epidemiology, control and treatment. Bulletin of the WorldHealth Organization 57, 683–689.
238 Chapter 5

Peters, W. (1992) Colour Atlas of Arthropods in Clinical Medicine. Wolfe, London, UK.RSTM&H (2000) Elimination of lymphatic filariasis as a public health problem. Transactions of the
Royal Society of Tropical Medicine and Hygiene 94, 589–602.Sasa, M. (1976) Human Filariasis. University Park Press, Tokyo, Japan.
The Nematodes 239

Included in this section are a hetero-geneous collection of endoparasites thatare not strictly helminths, but which maybe confused with them, particularly inbiopsy tissue sections.
Pentastomes
The members of this small group of para-sites form a separate phylum(Pentastomida), usually placed somewherebetween the annelids and the arthropods.Larvae of seven species have been reportedfrom humans but only two are of anyimportance.
Armillifer (= Porocephalus) armillata
This is a parasite of the respiratory tract ofsnakes in Africa and Asia, while the larvaeare found encysted in the tissues of rodentsand other mammals. Human infection ispresumed to be by ingestion of eggs on veg-etation or in water. The egg contains anembryo (0.2 mm in length), which boresthrough the intestinal wall and encysts inthe mesenteries of the peritoneal cavity(particularly the liver). The coiled larvamoults twice and grows until it measures20–25 mm in length. These third-stage lar-vae have characteristic annulations, givinga beaded appearance (Fig. 119). Larvae mayproduce a considerable inflammatoryresponse but usually cause no symptomsand are only seen on X-rays taken for other
reasons when calcified, or probably areabsorbed and never diagnosed. Severepathology is rare and is associated withmigration of the primary larvae before theybecome encysted, or in a few cases consid-erable tissue damage has been caused bythe third-stage larvae leaving the fibrouscapsule and undergoing another migration;in both cases a form of visceral larvamigrans results.
Human infection is common in someareas of the world, such as Malaysia andWest Africa (Benin, Cameroon, Congo,Gabon, Nigeria and Senegal).
On X-ray Armillifer may be mistaken forguinea worm and in tissue sections theyare superficially similar to spargana,gnathostomes, fly larvae or the chigoe flea(Tunga penetrans). In tissue sections penta-stomes may be differentiated by the pres-ence of a body cavity and intestine (as haveall the others except spargana), chitinoustracheae (as have fly larvae and Tunga),striated muscle fibres and the presence ofreproductive organs.
Armillifer agkistrodontis, a parasite of theviper, has been recovered once in Chinaafter treatment with mebendazole at 400mg three times daily for 3 days in a patientwith abdominal pain who had drunk snakebile and blood (Zhang et al., 1997).
Linguatula serrata
Known as the ‘tongue worm’ this is a nat-ural parasite of the nasal passages of dogs
6Other (Non-helminth) Groups
240
06Worms & H.D. - Chap 06 14/11/01 4:29 PM Page 240

and other carnivores, with the larval stagesoccurring in herbivores. In Lebanon a typeof pharyngitis known as ‘halzoun’ occursin humans from ingestion of insufficientlycooked liver of sheep and goats containinglarvae; in Sudan the similar ‘Marrara syn-drome’ is found, with an allergicnasopharyngitis accompanied by itching inthe throat, deafness, tinnitus and facialpalsy (Yagi et al., 1996). Larvae can migratefrom the stomach to the tonsils and symp-toms often include sneezing and coughingalso. The condition usually recovers spon-taneously in about a week when the para-sites become dislodged. Ocular cases havebeen reported from Ecuador, Israel and theUSA (five cases), with conjunctivitis and alarval worm in the anterior chamber. Alarva has been found in the pericardial sacof a woman in Georgia, USA, two caseswith larvae causing tortuous tunnels andlymphatic nodules in the abdomen inCosta Rica and larvae in the bile-duct,causing jaundice and gallstones, in apatient in Turkey.
Leeches
Leeches belong to the phylum Annelidaand many species are adapted to suckingthe blood of vertebrates. Most are externalblood feeders and, although they may be amajor nuisance in areas of Asia, are notlikely to be mistaken for parasitichelminths. There are a few forms causinginternal infection, of which the best known
genus is Limnatis, reported from humansin southern Europe, North Africa, NorthAmerica and Asia. Limnatis nilotica is aparasite of cattle and lives in water-holesand springs. Adults may measure 15 cm inlength but are capable of great contractionand extension. The body is greenish-blackin colour with thin orange stripes, andthere are two suckers, one around themouth, the other at the posterior end.When a very young worm is ingested indrinking water, it attaches to the mucousmembranes of the mouth, pharynx, vocalcords or trachea, where it may remainattached for days or weeks. The leech feedson blood and secretes an anticoagulant,which can cause excessive bleeding andeven death. Usually, however, clinicalsymptoms result from the mechanicalaction of worms, which cause an oedema-tous reaction with paroxysmal coughingand dyspnoea. Various species of leecheshave attached to the conjunctiva (in Turkeyand the USA), causing periorbitalhaematomas, and invaded the nose (India),vaginal wall (India) and urethra (Malaysia).
Engorged leeches can be removed withforceps; otherwise an anaesthetic (0.5%tetracaine) or irritant substance may benecessary to induce them to detach.
Myiasis
Accidental myiasis may be caused by lar-vae of flies belonging to various families,such as the blowflies, flesh-flies and house-flies. The larvae or eggs are usuallyingested with contaminated food or waterand the larvae are found in the intestinaltract or passed out in the faeces.
The larvae of some flies are obligate par-asites in the tissues of various vertebrates,the adults being known as warble flies orbot-flies. The larvae can be mistaken forhelminths when causing larva migrans,although they can be recognized easilywhen removed. In tissue sections fly larvaecan be differentiated by the chitinoustracheae, lack of reproductive organs andnon-striated muscles.
Dermatobia hominis (the tropical warble
Other (Non-helminth) Groups 241
Fig. 119. Early encysted larva of Armilliferarmillata on intestinal wall (� 6).
06Worms & H.D. - Chap 06 14/11/01 4:29 PM Page 241

fly of South America) is the only genus thatcan complete its entire larval developmentin humans. The larvae are found in a widerange of mammalian hosts and in humansoccur in unprotected parts of the body. Theeggs are laid by the female fly on mosqui-toes, other flies or ticks, which transportthem to humans. The eggs hatch when themosquito feeds and the larvae penetrate theskin, with a channel remaining to the sur-face. The posterior ends of the larvae pokeout from an ulcer on the skin at intervalsbefore they drop out in 6–12 weeks afterinfection and pupate in the soil. Larvaeproduce a severe pruritus, and a suppurat-ing wound may result from scratching.
Cochliomyia hominovorax (the NewWorld screw-worm) is widely distributed incattle in the Americas. Eggs are depositedon the skin next to a wound or on mucousmembranes of the nose, mouth or vagina.The hatched maggots (up to 400) tunneldeeply to produce a communal boil-likelesion, oozing pus, which often becomessecondarily infected and leads to death inabout 8% of untreated cases. After 1 week offeeding the mature maggots exit and drop tothe ground to pupate. One case was success-fully treated with topical application of 1%ivermectin, which killed all larvae by 24 h(Victoria et al., 1999). This fly was acci-dently introduced into Libya in 1988, but
fortunately was eradicated before it spreadto the rest of Africa (Hall and Wall, 1995).Chrysomya bezziana (the Old World screw-worm) is a similar fly in Asia and Africa.
Gasterophilus sp. (the horse bot-fly)first-stage larvae occasionally wanderunder the skin of humans (particularly inRussia, with a recent case in an infant inthe USA). The dark transverse bands of thelarvae can sometimes be seen under theskin (particularly if mineral oil is rubbedinto the skin to make it transparent) insidea serpiginous tunnel. The larvae may moveseveral millimetres a day, producing a pru-ritus similar to creeping eruption.
Hypoderma bovis (the cattle bot-fly) lar-vae may bore down deeply into the tissues,while those of Oestrus ovis (the sheep bot-fly) are usually found in humans in theexternal membranes of the eye, and thoseof Rhinoestrus purpureus (the Russian gad-fly) in the nose.
Cordylobia anthropophaga (the Africantumbu fly) belongs to a different groupfrom the previous forms and has larvae thatproduce a boil-like swelling at the site ofpenetration in many animals. The eggshatch in moist soil and penetration is usu-ally through the feet. The larvae leave thebody in 8–9 days. A recent case in thebreast of a Nigerian woman appeared to bea tumour on a mammogram.
242 Chapter 6
References
Hall, M. and Wall, R. (1995) Myiasis of humans and domestic animals. Advances in Parasitology 35,257–334.
Victoria, J., Trujillo, R. and Barreto, M. (1999) Myiasis: a successful treatment with topical iver-mectin. International Journal of Dermatology 38, 142–144.
Yagi, H., El Bahari, S., Mohamed, H.A., Ahmed, E.R.S., Mustafa, M., Saad, M.B.A., Sulaiman, S.M.and El Hassan, A.M. (1996) The Marrara syndrome: a hypersensitivity reaction of the upper respi-ratory tract and buccopharyngeal mucosa to nymphs of Linguatula serrata. Acta Tropica 62,127–134.
Zhang, Q.Y., Wang, B.F., Huang, M. and Cheng, T.F. (1997) Viper’s blood and bile. Lancet British edi-tion 349, 250.
Further Reading
Hall, M. and Wall, R. (1995) Myiasis of humans and domestic animals. Advances in Parasitology 35,257–334.
06Worms & H.D. - Chap 06 14/11/01 4:29 PM Page 242

Introduction
Parasites, like all pathogens, elicit a widerange of immune responses in their hosts.The nature of these responses is deter-mined both by the host and by the parasite.Even within a single host species there isvariation in the capacity to expressimmune responses, which reflects geneticdifferences as well as differences in suchvariables as age, sex, nutrition, physiologi-cal status and the presence of concurrentinfections. Parasite-determined variation inresponse arises from differences in thelevel and frequency of infection, as well asfrom the genetic variation that existswithin widely distributed species. Much ofour current knowledge of antiparasiteresponses has come from experimentalstudies in laboratory rodents, and theextent to which this can be extended tohumans is sometimes controversial.However, there is no doubt that the essen-tial principles of immune responsivenessto parasites apply to all mammalian hosts.An important principle, which has particu-lar relevance to helminths, is that very fewof the responses elicited by infection arelikely to be functionally protective. Manyare irrelevant to protection and some, asseen very clearly in schistosomiasis, mayhave severe pathological consequences.One of the important goals for immuno-parasitologists is to identify thoseresponses that are beneficial and deviseways of promoting these while reducingimmunopathological responses.
The Immune Response
Immune responses to infection can bedivided into two categories, innate andadaptive. In general terms, the first coversresponses that are expressed immediatelyon exposure to infection, show little mem-ory or enhancement on re-exposure, havebroad molecular specificity (termed patternrecognition) and are associated with theactivities of non-lymphoid cells. In con-trast, adaptive responses depend upon theactivity of lymphocytes, are induced in anantigen-specific manner, show memory andconsequently are enhanced when the hostis re-exposed to the same infection. Thefocus here will be on adaptive responses,but it should be emphasized that the dis-tinction between the two categories is oftenblurred, that innate responses may beessential for the correct induction of adap-tive responses, and that both can con-tribute to the overall response be itprotective or pathological.
Adaptive Responses
Adaptive immunity is dependent upon therecognition of and the response to specificsequences (antigenic determinants or epi-topes) associated with molecules that areforeign to the body (i.e. are ‘non-self’).Recognition is absolutely dependent uponlymphocytes, a unique population of cells,characterized by possession of epitope-specific receptors on their cell membranes.
7Immunology of Helminths
243
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 243

244 Chapter 7
Lymphocytes are divided into two majorpopulations – T (thymus-dependent) cells,which carry T-cell receptors (TCR) and B(bursa- or bone-marrow-derived) cells, inwhich the B-cell receptor is immunoglobu-lin (Ig). As far as parasitic infections areconcerned most adaptive immuneresponses are initiated when T cells recog-nize epitopes, and this requires the suc-cessful completion of two distinctprocesses. The first is presentation of epi-topes to T cells by non-lymphoid antigen-presenting cells, such as macrophages,dendritic cells and B cells, and the secondis recognition of those epitopes by TCR.Recognition is a complex process thatinvolves multiple receptor–ligand inter-actions between antigen-presenting cellsand the T cell concerned, as well asepitope–receptor binding. These inter-actions involve a variety of molecules car-ried on each cell surface, as well as bindingof receptors on the T-cell surface to solublesignalling molecules (cytokines). Once acti-vated, T cells themselves release cytokinesand interact directly and indirectly withother cells.
T cells can be categorized into a numberof subsets by the surface molecules theycarry (e.g. CD4+ helper cells or CD8+ cyto-toxic cells) and by the cytokines theyrelease (e.g. helper cells can be dividedinto at least three subsets on this basis).Different T-cell subsets regulate differentcomponents of the immune system’s effec-tor and memory responses. Key effectormechanisms are mediated via antibodyproduction, cytotoxicity and inflammation;all three involve the coordinated activity oflymphocytes and non-lymphoid cells.
The interactions of these components inthe initiation of the immune response areshown in Fig. 120.
Antibody Production
Antibodies (circulating Ig) are produced byB cells that have been appropriately stimu-lated and have gone through a sequence ofdevelopmental steps to produce plasmacells. Although it is possible for B cells to
respond directly to certain types of antigen(particularly polysaccharides), in almost allcases involving parasites B cells require T-cell help, the T cells helping to determinethe class of antibody by the pattern ofcytokines they produce. The specificity ofthe antibodies produced is the same as thatof the surface receptor carried by the Bcells involved initially, but plasma cellsrelease large amounts of the molecule in asoluble form and modify the molecules toperform a variety of biological functions.
Ig molecules are formed from pairedheavy and light glycoprotein chains andcan be divided into two functional regions,the Fab region, which determines epitoperecognition, and the Fc region, whichdetermines other biological functions (Fig.121). Differences in the Fc regions definethe five major classes of antibodies (IgM,IgD, IgG, IgA, IgE). Binding of antibody to atarget epitope may itself have protectivevalue, e.g. if the target is carried by a cru-cial enzyme or sensory cell; more fre-quently, antibody binding serves toidentify the target immunologically andthus act as a focus for other effector mecha-nisms. An important mechanism involvesthe activation of complement, a complexseries of plasma proteins capable of trigger-ing non-lymphoid effector cells, of increas-ing phagocytic activity and of damagingcell membranes to bring about cell lysis.Bound antibody also acts to increase inter-actions between the target and non-lym-phoid cells that carry the appropriate Fcreceptor. Antibody binding and comple-ment activation can therefore both beinvolved in what is collectively termedantibody-dependent cellular cytotoxicity(ADCC), a mechanism thought to beinvolved in immune protection against anumber of helminths (Fig. 122).
Cytotoxicity
In addition to ADCC, cytotoxicity can bebrought about directly by T cells, particu-larly CD8+ cells and specialized naturalkiller (NK) cells These bind to targetsthrough the TCR or other receptors and
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 244

Immunology of Helminths 245
release a number of potent molecules thatdamage cell membranes or cause intra-cellular damage. T-cell cytotoxicity is wellestablished as a protective mechanism ininfections involving intracellular proto-zoans, but there is little evidence for itsimportance in helminths.
Inflammation
Parasites bring about inflammatory changesin a variety of ways. Their invasion andactivities in tissues cause mechanical andchemical damage, and their interactionswith non-lymphoid cells cause the releaseof potent mediators, which themselves can
damage tissues, but it is probable thatimmune-mediated inflammation plays amajor role in a majority of infectionsinvolving helminths. This form of inflam-mation can be mediated by antibodies, par-ticularly when complement is involved,and by T cells. Antibody of the IgE class isparticularly important in this respect, as itbinds to cells that have the potential torelease powerful biological mediators (e.g.amines) when the bound antibody becomescomplexed with its specific antigen. T-cell-mediated inflammation is the result ofcytokine release and the increased produc-tion, or increased activity, of a wide varietyof non-lymphoid cells. Again it is therelease of potent mediators that initiates
WORM
ESantigen
Somaticantigen
Effector function
Processingand bindingto MHC II
CD4
ANTIGEN-PRESENTING
CELL Costimulatorymolecules
T CELL
ActivationCytokinereleaseand clonalexpansion
MACROPHAGEACTIVATIOATIOA NANTIBODY
ANTIBODYINFLAMMATIOATIOA N
Type 2Type 2T
Type 1Type 1T
Cytokinerelease
EFFECTOR CELLS
MEMORY CELLS
Fig. 120. Initiation of the immune response to a parasitic worm. Worms release antigens in theirexcretions/secretions (ES antigens), from their cuticle, and from their bodies (somatic antigens) when theworm is killed. Antigen is taken up by antigen-presenting cells and presented by major histocompatibilitycomplex (MHC) molecules to T cells. Recognition of antigen by the T-cell receptor and ligation ofaccessory and costimulatory molecules activate the T cell. The activated cells release cytokines andundergo clonal expansion to produce clones of cells with identical T-cell receptors. These cells, actingthrough cytokines, become effector cells or memory cells. Effector cells, through release of type 1 or type2 cytokines, coordinate a variety of effector activities against the parasite. Key to symbols: MHCmolecule; � Antigen; T-cell receptor.
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 245

246 Chapter 7
the inflammatory damage. Some of themediators released during inflammation,however caused, can damage parasitesdirectly; in other cases the tissue changesassociated with inflammation may affectthe parasite indirectly, essentially by bring-ing about dramatic changes in its environ-ment.
Non-lymphoid Cells
Lymphocytes and non-lymphoid cells botharise from precursors in the bone marrow,but then follow divergent pathways ofdevelopment – the lymphoid and themyeloid. The latter pathway gives rise tothe two major categories of non-lymphoid
cells, the macrophage–monocyte line andthe granulocyte line. Monocytes circulatein the blood and mature as macrophages inthe tissues. Three groups of granulocytes(neutrophils, eosinophils and basophils)mature in the bone marrow and move intotissues from the blood, while the fourth(mast cells) matures in tissues. All non-lymphoid cells can participate in innateand adaptive responses through theirphagocytic activities and through theirability to release powerful mediators.Macrophages, neutrophils and eosinophilsoften infiltrate tissues around worms andmay form granulomas when parasite stagesbecome trapped in tissues. Basophils andmast cells – the amine-containing cells –are involved primarily through the capac-
X
X
X CH2 CH3
Hinge region
CLVL
VH CH1
Fab Fc
Light chain
Heavy chain
Antigen-bindingsite
Variable region
Constant region
Disulphide bond
Carbohydrate group
Fig. 121. Diagrammatic representation of an immunoglobulin molecule (based on IgG). The molecule ismade up of two light (L) chains and two heavy (H) chains, held by disulphide bonds. The bond betweenthe paired chains can be split by enzymes to break up the molecule into the Fab and Fc fragments. Eachchain forms a number of regions (domains): the variable (V) regions form the antigen-binding site of theFab fragments; the constant (C) regions of the Fc fragment confer the biological properties of themolecule, e.g. binding to receptors and activation of complement.
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 246

Immunology of Helminths 247
ity to release a variety of mediators.Whereas phagocytosis is an effectivedefence mechanism against protozoans,worms, even larvae, are too large to betaken in by single cells. A more character-istic response is for cells to bind closely tothe parasite surface, through antibody- orcomplement-mediated mechanisms, andthen to release mediators directly on to theworm surface. In that way the outer surfaceis damaged, often irreversibly, and theworm killed.
Parasites and their Antigens
Parasites, and helminths in particular, arecomplex organisms, often with complexlife cycles. The presence of an infectiontherefore exposes the host to a battery of
parasite-derived foreign molecules, whichcan be recognized by the immune system.These are commonly proteins, glycopro-teins or carbohydrates, but molecules suchas glycolipids can also be antigenic. In thecase of helminths, antigens may be presenton the parasite surface, released from theworm as it develops, reproduces andmetabolizes (collectively referred to asexcretory/secretory (ES) antigens) orreleased when the worm dies and isdestroyed. Antigens of intracellular proto-zoans may be expressed on the surface ofthe host cell, and this also occurs in thenematode Trichinella spiralis during itsintramuscular phase.
The antigens presented by helminthsfrequently change as the parasite goesthrough its life cycle, i.e. there is stagespecificity in antigen expression. This can
FcR- and/or C3R-mediatedadhesion
Adhesion notinvolving
FcR or C3R
EOSINOPHILS, MACROPHAGES, NEUTROPHILS(NK cells and Mast cells)
C3R Fc FcR C3R STEP 2
STEP 1Complement activatedby alternative pathway
IgG, IgA, IgEbound to antigen
Complement activatedby classical pathway
WORM
OOOOOOOOOOOOOO XXXXXXXXXXXXXX XXXXOOOXXXXOOO
OOOOOOOOOOOOOOOOOOXXXXXXXXXXXXXXXXXX X
Immune complexes boundto cell through FcR;
EOSINOPHILS,MACROPHAGES
Strong adherenceby activatedMACROPHAGES
Weak adherence(e.g. sugar–lectin)byMACROPHAGES
Fig. 122. Mechanisms by which cells mediate cytotoxic activity against parasitic worms. Adhesionbetween cells and parasites can involve cellular receptors for the Fc portion of immunoglobulinmolecules (FcR), or receptors for components of complement such as C3(C3R), when these moleculesare bound to the parasite’s surface. Adhesion can also occur through mechanisms not involving directbinding of immunoglobulin or complement to the parasite (e.g. via immune complexes, lectin–sugarbinding and intermolecular forces).
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 247

248 Chapter 7
result in the responses raised against onelife-cycle stage being ineffective against alater stage. Equally life-cycle stages maydiffer in their immunogenicity (their abilityto generate immune responses), althoughthis is also influenced by their location.Metabolically active stages that migratethrough tissues, or remain there for longperiods, are often potently immunogenic,Ascaris larvae and schistosome eggs beingexcellent examples. Resting larval stages,particularly those that become encysted orenter immunologically privileged sites,such as the central nervous system (CNS),may generate much less response – tape-worm larvae are good examples. Someparasites living in the intestine (e.g. hook-worms) can stimulate strong immuneresponses, their antigens being taken upacross the mucosa, but others, notably thetapeworms, appear much less immuno-genic. Many nematodes and larval trema-todes release antigens that act as allergens– i.e. stimulate the formation of IgE – andthus the host develops hypersensitivity tothe parasite, allergic symptoms being com-mon in many helminth infections.
Not all molecules released by helminthsact as immunogens, in the sense of stimu-lating positive immune responses. Manyworms are now known to releaseimmunomodulatory molecules, whichsuppress, divert or inactivate componentsof the immune response. Good exampleshave been described in infections withfilarial worms and with hookworms, butimmunomodulation is probably a wide-spread phenomenon. Helminths alsoexpress, or acquire, molecules that act asimmunological disguises (glycolipids inadult schistosomes, albumin in micro-filaria larvae, host proteins in cestode lar-vae), which enable them to reduce thedegree of ‘foreignness’ they present to thehost’s immune system, effectively beingseen as self rather than non-self. Somemolecules, although immunogenic, havebeen shown to elicit antibody responses(e.g. IgM antibodies to carbohydrate anti-gens of S. mansoni) that block the protec-tive action of other antibody classes and
thus decrease protective immunity (‘block-ing antibodies’).
Expression of Immune Responses
The detailed understanding of the immunesystem that we now have enables us tointerpret in mechanistic terms many of thecharacteristics of the immune responsesthat parasitic infections generate. It seems ageneral rule that, whereas many protozoa,particularly the intracellular species, elicitlymphocyte responses dominated by the T-helper 1 (Th1) subset, helminth infectionselicit Th2-dominated responses. Th1responses are associated with cellularimmunity (e.g. activation of macrophages)and production of specific classes of IgGantibodies. In contrast, Th2 cells coordinatea pattern of antibody responses, whichincludes IgE, and regulate the productionand activity of inflammatory cells, particu-larly eosinophils and mast cells. The reasonfor this Th subset bias is not fully under-stood, but it reflects the nature of particularantigens, the ways in which these are pre-sented to T cells and the cytokine environ-ment in which T-cell responses take place.Thus T-cell responses occurring in the pres-ence of the cytokine interleukin 12 (IL-12)(derived from previously activated NKcells) become polarized to the Th1 subset,while those occurring in the presence of IL-4 (derived from cells such as mast cells)become polarized to the Th2 subset.
The nature and degree of the immuneresponse elicited by infection are not onlydependent on the level and frequency ofinfection and the characteristics of the para-site concerned, they are also influenced bythe location of the parasite. Helminthsoccupy a very wide variety of habitats inthe host’s body, and the capacity of these torespond to infection and the nature of theresponse expressed can differ quitemarkedly. A useful distinction in immuno-logical terms is seen in the responses toparasites living within the intestine andthose elsewhere in the body, i.e. betweenparasites living in mucosal and systemic
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 248

locations. Although the mucosal and sys-temic immune systems are clearly intercon-nected, the populations of lymphocytes ineach and their patterns of migration are dis-tinct. The responses occurring within theintestinal wall itself are similar in manyrespects to those elsewhere in the body, buta major difference is seen in the predomi-nant class of Ig produced there and secretedinto the gut lumen (IgA), whereas the sys-temic immune system is dominated by IgGand IgM antibodies. The mucosal tissuesare readily infiltrated by inflammatory cells,characteristically including a discrete pop-ulation of mast cells. Interactions betweenthese cells and IgE can trigger hypersensi-tivity reactions, which lead to majorchanges in intestinal structure and func-tion. The mucosal immune system operatesnot only in the intestine, but also in thelungs and urogenital tissues. Systemicimmunity has different characteristics indifferent locations, and these influence theantiparasite responses generated by infec-tion. Important differences are seen, forexample, between responses in the CNS,usually regarded as an immunologicallyprivileged site (i.e. with reduced respon-siveness), and those in the skin, where aseparate population of antigen-presentingcells (Langerhans cells) operates and wherecellular inflammatory responses are readilygenerated. In contrast to protozoans, rela-tively few helminths live for any length oftime in the blood, adult schistosomes andmicrofilaria larvae being exceptions.Despite being bathed in antibodies andcomplement and potentially exposed to avariety of non-lymphoid cells, both survivefor prolonged periods, because they possesseffective evasion mechanisms which pre-vent them from being recognized.
Protective Immune Responses inInfected Hosts
Although it is easy to demonstrate thatparasitic infections elicit immuneresponses in their human hosts, it is moredifficult to demonstrate the relation of theseresponses to protective immunity. Immune
responses in parasitized humans can bemeasured in a variety of ways. Theseinclude tests that demonstrate the presenceof specific antiparasite antibody, lympho-cyte responses and cytokine production,skin sensitivity to antigen, increased pro-duction of ‘non-specific’ IgM and increasednumbers of eosinophils, as well ashistopathological demonstration of thepresence of immune cells in infected tis-sues. In addition, the capacity of antibodiesand cells taken from infected individuals tokill parasites or parasite-infected cells canbe tested in vitro. Demonstrable protectionagainst reinfection, in individuals or inpopulations (e.g. Leishmania major), isclear evidence of immunity, but is rela-tively rare in helminth infections. Evidenceof age-related declines in infection rates inendemic areas (e.g. in malaria and inhelminth infections) suggests the operationof host-protective immunity, as does protec-tion against the pathological effects ofinfection (e.g. malaria), but neither is neces-sarily conclusive. ‘Experiments of nature’ –i.e. study of infections in individuals withcompromised immune systems – can indi-cate the importance of immune competencein preventing the consequences of infection(e.g. giardiasis, cryptosporidiosis orstrongyloidiasis). However, these lines ofevidence can often only be deduced fromdetailed and extensive population studiesand sophisticated statistical analysis. Moredirect evidence that host-protective immu-nity can operate effectively against para-sites can be gained from studies inexperimental models, although it is oftennot possible to use in such models (particu-larly rodents) the species of parasite thatare of interest in humans.
There are experimental models for mostof the major human helminth infections, themajority of these being in rodent hosts.Under controlled conditions, it is possibleto show that, in many cases, infections elicitstrong immune responses and that thesemay confer substantial protection on thehost. Protection developing during an initialinfection can be quantified by determiningthe numbers of worms recovered at intervalsafter infection or by the levels and duration
Immunology of Helminths 249
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 249

of larval and egg output. In other cases pro-tection is not seen, and worms persist forlong periods in the presence of specificimmune responses. Good examples fromwork with mouse models include the long-term survival of schistosomes, larval ces-todes, liver flukes, microfilariae and certainintestinal nematodes. Immunity to reinfec-tion can be seen in models where there is noresistance to the primary infection, but thenature of this reinfection immunity needs tobe examined carefully. For example, theresistance of mice to reinfection withSchistosoma mansoni reflects the pathologi-cal changes that the primary infection hasbrought about in the liver, rather than spe-cific immune destruction of the incomingworms. Resistance to reinfection withTaenia taeniaeformis in mice carryingmature cysticerci reflects the operation ofimmune responses against the penetratingoncospheres and early cysticerci beforethese have developed resistance to comple-ment-mediated damage. Resistance of miceto reinfection with Heligmosomoides poly-gyrus (an intestinal nematode widely usedas a model of chronic infection) is expressedonly if the previous adult infection and itsassociated immunosuppressive effects havebeen removed by anthelminthic treatment.Once the existence of host-protective immu-nity has been established, potential mecha-nisms of immune-mediated resistance andthe response components involved can bestudied using in vitro approaches. Goodexamples include analysis of ADCC mecha-nisms against schistosomula and microfilar-iae. However, the demonstration of effectiveimmune killing under in vitro conditionsdoes not necessarily correlate with in vivopatterns of response. The capacity of hoststo become immune can also be studied inexperimental models by using attenuatedparasites or isolated parasite molecules asvaccine antigens. For example, mice can beeffectively vaccinated against S. mansoniusing irradiated cercariae, rats against thesame species with defined recombinantantigens (e.g. glutathione-S-transferase) andmice against intestinal nematodes withdefined antigens or synthetic peptides fromthese antigens.
Demonstration of protective immunityin human populations that are naturallyexposed to helminth infections is muchmore difficult than it is in the laboratory,because of the number of confounding fac-tors that have to be taken into account.Unlike experimental animals, individualhumans are exposed to varying intensitiesand frequencies of infection and differ fromone another in factors that are known toinfluence immunity (e.g. age, sex, infection,nutritional and physiological status andgenetics), as well as access to anthelminthictreatment. What was a general view thatthere is little immunologically mediatedresistance to helminths, despite abundantevidence of immune responses as such, hasbeen considerably modified as a result ofimmunoepidemiological and in vitro stud-ies. For example, it now seems quite clear,from detailed treatment/reinfection studies,that humans do develop protective immu-nity to infection with S. mansoni and S.haematobium. Correlative studies havelinked resistance to levels of parasite-spe-cific IgE and to eosinophil activity, data thatare supported by in vitro work. Althoughthe picture is much less clear with themajor gastrointestinal nematodes, there aredata suggesting that a level of protectiveimmunity is acquired, at least in hookwormand Trichuris infections. Knowledge of thecellular and molecular mechanisms under-lying immune responses has increased thepredictive value of studies that attempt tocorrelate infection status with particularimmune components. This is best seen withthe many studies that now use T-cell andcytokine data as analytical tools. The vari-ous clinical categories found in populationsexposed to lymphatic filariasis (from theputatively immune, parasite-negativeendemic normals to infected individualswith obstructive disease) have been corre-lated with particular patterns of Th-cellresponse and cytokine production,although the relationship is clearly com-plex. Epidemiological and genetic studieson populations exposed to S. mansoni havepinpointed a major gene that controls theexpression of protective immunity, andhave revealed positive correlations between
250 Chapter 7
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 250

Immunology of Helminths 251
resistance and the genetic control of Th2-related cytokines. For many helminth infec-tions, however, evidence for protectiveimmunity is slight, and there is little infor-mation about the underlying basis of theimmune responses that infection elicits.
Exploitation of Immune Responses
Diagnosis
Whether or not immune responses conferany protection against helminths theirpresence can be used as a means of diag-nosing infection. A majority of diagnostictests make use of antibody responses, andthese can be measured by many tech-niques. Perhaps the most widespread atpresent is the ELISA, which can be usedwith very small quantities of serum, can beautomated for large-scale surveys and isread using colorimetric assays. One draw-back of antibody-based tests is that thepresence of antiparasite antibody does notnecessarily imply the presence of a currentinfection, although measurement of partic-ular classes of antibody (e.g. IgG4) is a goodindicator. The ELISA can be adapted to testfor antigen rather than antibody, and ofcourse this is a positive indicator of para-sites in the body. Antigen detection hasbeen widely used in schistosome and filar-ial infections. An exciting new develop-ment is the development of non-invasivedetection of ES antigens by an antigen-cap-ture ELISA in faeces (coproantigens), urineand perhaps saliva (e.g. see p. 24 for schis-tosomiasis in urine and p. 77 for cestodesin faeces).
Conventionally, ELISA tests have usedmultiwell plates to provide the solid-phasesubstrate on which antigen or antibody canbe bound, and they require spectrophoto-metric readouts. Recent development of thedipstick technique, where reagents arebound to a nitrocellulose strip and coupledwith built-in controls, allows immediateidentification of a positive or negativeresponse and can be used in the absence ofsophisticated laboratory equipment. A lim-
iting factor in all diagnostic tests is theavailability of purified antigens to givehigh levels of both sensitivity and speci-ficity. The use of recombinant and syn-thetic techniques for antigen production isovercoming this limitation.
A number of older diagnostic tests usedcellular responses to injected antigen as anindicator of infection. These have largelybeen phased out, largely because of lowspecificity and the danger of adverse reac-tions.
The immunology of trematodes is dis-cussed on p. 23 and p. 41, of cestodes onp. 93, of intestinal nematodes on p. 121 andof filarial nematodes on pp. 196 and 217.
Vaccination
As with all infectious diseases, vaccinationagainst helminths is seen as one of the mosteffective ways in which infections could becontrolled. Until quite recently, success inthis field has been negligible comparedwith the progress made with viral and bac-terial vaccines. Although many of the latterwere initially developed quite empirically,this approach has not been successful withhelminths, where the problems of elicitingthe required protective responses are muchgreater. Experimental vaccines using atten-uated larval stages have proved effective inrodents against parasites such as schisto-somes, hookworms and some filarial nema-todes. Attenuated larvae have also beenused to reduce disease in schistosome-exposed cattle, but the ethical problemsinvolved with the use of live vaccines inhumans have prevented further develop-ment. Most progress has been achievedwith the development of recombinant vac-cines, which were first used successfullyagainst larval cestodes in sheep, showpromise against gastrointestinal nematodesin sheep and are being tested in trials foruse against S. mansoni in humans. The lat-ter vaccine is based on the use of recombi-nant glutathione-S-transferase, a moleculethat also has protective value against otherschistosome species.
07Worms & H.D. - Chap 07 14/11/01 4:34 PM Page 251

Epidemiology may be defined as the studyof the factors influencing the occurrenceand distribution of infection. In general, itis concerned with populations rather thanwith individual cases.
Epidemiological studies serve two mainfunctions: first, they provide the necessarydata for determining the medical, eco-nomic and social importance of an infec-tion, in order to evaluate and plan theeffort and funds that should be devoted toit; secondly, they can help to discover therelevant factors whose manipulation couldlead to prevention or control of disease.There have been many recent studies thathave provided important findings on therelative importance of helminth infectionsas causes of the morbidity discussed in thevarious sections and these are helping inthe evaluation of the benefits (includingeconomic) of control campaigns.
Unlike microorganisms, such as viruses,bacteria and protozoa, in which a feworganisms can multiply quickly and causeserious disease, helminths are large para-sites which, in most cases, do not multiplydirectly within the body of their hosts. Forthis reason the number of parasites presentat any one time can be of great importancein determining the likelihood of morbidity,and thus helminth infections tend to bechronic and insidious, with increase in theintensity of infection possible throughrepeated reinfection. For microorganismsthe infected host provides the most conve-nient unit of study in epidemiological sur-veys, while the individual helminth
provides the most convenient basic unit ofstudy, and measurements of worm burden(e.g. directly by counting worms in faecesafter chemotherapy, or indirectly, by eggcounts or recently by immunological esti-mates of circulating worm antigens) can bevery important.
Epidemiological studies can be dividedinto two main categories. Descriptive stud-ies can be used in order to determine thefrequency and intensity of infections,where and when they occur and what kindof people become infected; repeated stud-ies over a period of time (longitudinalcross-sectional studies) are often more use-ful than studies at one period in time (hori-zontal cross-sectional studies) and areessential for determining the rate of acquir-ing infection (incidence).
In general, epidemiology is a quantita-tive science and properly designed andstatistically valid techniques are of criticalimportance and there should be standard-ization of the methods employed. Changesin technique can often make it difficult tocompare older with more recent figures.For instance, newer serological techniquesare often considerably more sensitive thanolder parasitological methods (e.g. micro-filarial counts compared with theimmunochromatographic (ITC) test in lym-phatic filariasis).
Attempts to control or eradicate humanhelminth infections are usually aimed atbreaking the life cycle by manipulating oneor more of the various transmission factors.However, with a well-established endemic
8Epidemiological Aspects of
Helminth Infections
252
08Worms & H.D. - Chap 08 14/11/01 4:30 PM Page 252

Epidemiological Aspects of Helminth Infections 253
parasite, this might be very difficult toaccomplish. Helminths are almost invari-ably overdispersed or aggregated (a nega-tive binomial distribution) within theirhosts, so that many hosts harbour just afew parasites while a small proportion(about 15% in the case of ascariasis) haveheavy infections. Thus large sample sizesare necessary in order to obtain accuratefigures of intensity of infection.
The regulation of the abundance andprevalence of helminth infections is mostlyby density-dependent (or negative-feed-back) factors in the host population or indi-vidual host, so that herd immunity, degreeof input of susceptibles and fluctuations intransmission rates do not tend to regulatethe number of parasites present and infec-tions can be long-lived and stable. Whilethe likelihood of heavy infections is obvi-ously affected by differences in host behav-iour, there is also clear evidence, at leastfor the schistosomes, that it is geneticallydetermined. The age of the host is often animportant factor in the occurrence andseverity of infection and age prevalenceand age intensity curves can be calculated(see Fig. 55).
The basic reproductive rate or, moreaccurately, number (R0) determines theability of the parasite to perpetuate itself. Itcan be defined as the average number ofoffspring (or of female offspring in the thecase of dioecious parasites) producedthroughout the lifetime of a female para-site, which themselves achieve maturity inthe absence of density-dependent con-straints. R0 must be > 1 for the parasite tomaintain itself, while, if there were no con-straints, the numbers of parasites wouldincrease exponentially (provided R0 was >1). In practice, helminth populations tendto remain stable over time (Anderson,
1998), although R0 will vary within thehost population from one geographicallocality to another.
The search for a more quantitativeexpression of the factors involved in thetransmission of parasitic diseases has con-tinued since Ronald Ross applied a systemof differential equations (known as a math-ematical model) to the description ofmalaria transmission. A model can be con-structed based on the life cycle of the para-site and, provided that the parasitepopulation is at equilibrium, the net repro-ductive rate should equal unity. Forinstance, with the schistosomes, this ratecan be expressed as the product of variousfactors:
net reproductive = (net reproductive rate rate of parasite of parasite in snail
population) �
(probability of infecting a definitive host) � (net reproductive rate of parasite in definitive host) � (probability of infecting a snail)
Provided that figures are available in orderto calculate these various components, itshould be possible to predict the effect ofvariations in each on the overall transmis-sion picture. Preferably any model shouldinclude all the factors that have a signifi-cant influence on transmission, even if thiscomplicates the analysis.
The transmission threshold is the levelof parasite transmission, reproduction andmortality rates that maintain infection andbelow which the parasite will die out (R0 <1) and, by manipulation of various parame-ters in the model, the effect of each on thelikely ease of elimination of the parasitecan be forecast.
References and Further Reading
Anderson, R.M. (1998) Epidemiology of parasitic infections. In: Cox, F.E.G., Kreier, J.P. and Wakelin,D. (eds) Topley and Wilson’s Microbiology and Microbial Infections, 9th edn, Vol. 5. Arnold,London, UK, pp. 39–55.
Anderson, R.M. and May, R.M. (1985) Helminth infections of humans: mathematical models, popula-tion dynamics, and control. Advances in Parasitology 24, 1–101.
08Worms & H.D. - Chap 08 14/11/01 4:30 PM Page 253

254 Chapter 8
Bundy, D.A.P. and Medley, G.F. (1992a) Epidemiology and population dynamics of helminth infec-tion. In: Moqbel, R. (ed.) Allergy and Immunity to Helminths: Common Mechanisms or DivergentPathways. Taylor and Francis, London, UK, pp. 17–37.
Bundy, D.A.P. and Medley, G.F. (1992b) Immuno-epidemiology of human geohelminthiasis: ecologi-cal and immunological determinants of worm burden. Parasitology 104 (suppl.), S105-S110.
Giesecke, J. (1994) Modern Infectious Disease Epidemiology. Arnold, London, UK.Woolhouse, M.E.J. (1992) On the application of mathematical models of schistosome transmission
dynamics. II. Control. Acta Tropica 50, 189–204.
08Worms & H.D. - Chap 08 14/11/01 4:30 PM Page 254

The most important laboratory techniquesin medical helminthology are those con-cerned with the diagnosis of infection.Routine diagnostic techniques include therecovery of eggs or larvae in faeces or, forthe filarial worms, identification of micro-filariae in blood or skin snips.
Immunodiagnostic methods are advanc-ing at a rapid rate and are now the methodsof choice for the diagnosis of fascioliasis,cysticercosis, echinococcosis, aberrant tis-sue helminths and lymphatic filariasis,although not, as yet, for the diagnosis ofintestinal helminths. Immunodiagnostictests are discussed on page 250 and in thediagnosis sections of various helminthsand details of relevant texts (such as Wallsand Schantz, 1986; Garcia and Bruckner,1997; Garcia, 1999) are given in the generalreferences.
While not strictly a helminthologicaltechnique, it is often useful to staineosinophils and mast cells in tissues andthe methods of Ball and Hay orKermanizadeh and colleagues (see Cassellaand Hay, 1995) can be used.
Faecal Examination
The eggs of helminths commonly encoun-tered in the faeces of humans are shown inFig. 123 and those of rarer or spurious par-asites in Fig. 124. The eggs most commonlyfound in spurious infections (i.e. when noadults are present) are those of the sheepliver flukes, Dicrocoelium and Fasciola,
which are passed after eating infected liver,those of free-living or plant-parasitic nema-todes, such as Heterodera, which areingested in or on vegetables, and mite eggs.
The inexperienced operator needs toexercise caution in identifying helmintheggs. Provided eggs are present and areseen, it is usually a comparatively simplematter to recognize them for what they are.However, when eggs are absent, all too fre-quently other objects serve as acceptablesubstitutes. Particularly popular are miteeggs, starch grains, pollen grains and plantcells; but sometimes even air bubbles aremistaken for eggs. Starch grains and plantcells can be differentiated from eggs bytheir lack of internal structures and bytheir irregular outlines and great size range.Pollen grains are more difficult to distin-guish, as they appear to have an opercu-lum, but they are kidney-shaped and muchdenser than the Clonorchis type of egg,with which they are usually confused. Anexpert system has been devised for identifi-cation of helminth eggs (Esterre et al.,1987).
In order to minimize mistakes, the sizeof objects seen under the microscopeshould always be considered. As a roughguide, an egg measures about 1/15(Clonorchis) to 1/3 (Fasciolopsis) of thediameter of the microscope field at a mag-nification of �400 (�40 objective and �10objective).
Any larvae found in fresh faeces will bethose of Strongyloides (Fig. 125), but it ispossible for larvae of hookworms to be
9Helminthological Techniques
255
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 255

256 Chapter 9
Fig. 123. Helminth eggs and larva commonly found in human faeces. T1. Schistosoma haematobium (inurine); T2. S. mansoni; T3. S. japonicum; T4. Paragonimus westermani; T5. Fasciolopsis buski (egg ofFasciola hepatica is identical); T6. Gastrodiscoides hominis; T7. Clonorchis sinensis; T8. Opisthorchisfelineus (egg of O. viverrini is identical); T9. Metagonimus yokogawai; T10. Heterophyes heterophyes;C1. Diphyllobothrium latum; C2. Taenia saginata or T. solium ; C3. Hymenolepis nana; N1. Ascarislumbricoides; N2. A. lumbricoides (infertile); N3. Enterobius vermicularis; N4. Trichuris trichiura; N5.Hookworms (eggs of Necator and Ancylostoma are identical); N6. Larva of Strongyloides stercoralis.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 256

Helminthological Techniques 257
Fig. 124. Rarer eggs found in human faeces. T11. Schistosoma intercalatum; T12. S. mattheei; T13.Dicrocoelium dendriticum (often spurious from eating infected liver); C4. Dipylidium caninum egg mass;C5. Hymenolepis diminuta; N7. Strongyloides fuelleborni; N8. Ternidens deminuta (egg ofOesophagostomum bifurcum is similar); N9. Trichostrongylus spp.; N10. Physaloptera sp.; N11.Aonchotheca philippinensis; N12. Gnathostoma spinigerum (rarely reaches adult stage in humans); N13.Toxocara canis (not found in human faeces, although the similar egg of T. cati has been found a fewtimes); N14. Dioctophyma renale; N15. Ascaris lumbricoides (decorticated); N16. Hookworms(embryonated as sometimes found in very old faeces); N17. Meloidogyne sp. (root-knot nematodes,spurious infection from eating root vegetables).
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 257

258 Chapter 9
present in old faeces or even adults ofcoprophilic nematodes in old stools col-lected from the ground.
Freshly passed stool specimens shouldbe collected in waxed paper or plastic stoolcontainers (although the latter are oftenfound very useful for other purposes andnot returned), using a wooden applicator,and can be stored for a few days in therefrigerator or preserved in 10% formalin.Alternatively large numbers of samples canbe collected in sealed polyethylene tubing(12.5 cm � 7.6 cm � 0.1 mm thickness) andpreserved in merthiolate/iodine/formalin(MIF) stain and preservative (200 ml of1: 1000 merthiolate, 25 ml of 40% formalde-hyde solution, 5 ml of glycerol and 250 mlof distilled water; 0.15 ml of 5% Lugol’siodine can be added just before use for thedetection of protozoal cysts (great careshould be taken with formaldehyde as it isan irritant and can be carcinogenic). Thesimplest method for the detection of eggs isdirect microscopical examination of a smearof about 5 mg of faeces, diluted with freshlyfiltered tap water or saline, on a slide cov-ered with a coverslip.
The use of small disposable polyethyleneenvelopes 7 cm � 4 cm � 0.03 mm thick anda single microscope slide has been advocatedfor mass surveys to save expense of slidesand coverslips. With this method a larger100 mg sample can be smeared. However,these direct methods are not very sensitive.
Sodium acetate/acetic acid/formalin(SAF) fixative can also be used for preserv-ing eggs and protozoa in faeces (sodiumacetate 1.5 g, glacial acetic acid 2.0 ml,40% solution of formaldehyde 4.0 ml anddistilled water 92.0 ml).
Kato–Katz cellophane thick-smeartechnique
In this method, a faecal sample on scrappaper is pressed through a small screen and,with a spatula, a roughly 50 mg sample ofsieved faeces is placed on a microscopeslide (Fig. 126a, b, c and d). For a quantita-tive measurement, the faeces sample isplaced on a template with a central holeover the slide, excess faeces removed withthe spatula and the template carefullyremoved, leaving a cylinder of faeces (Fig.126e). The template should be 1 mm thick,with a hole of 9 mm, which will deliver 50mg of faeces, and it can be made of metal,plastic or cardboard; many should be made.A cellophane square (obtainable from ciga-rette packets, etc.) measuring 25 mm � 30or 35 mm which has been soaked in a mix-ture of 100 ml glycerine, 100 ml of distilledwater and 1 ml of 3% aqueous malachitegreen or 3% methylene blue is placed overit as a coverslip (Fig. 126f and g). The micro-scope slide can be turned over and pressedagainst a hard surface until newsprint can
Fig. 125. First-stage larva of Strongyloides stercoralis as found in human faeces (actual size 250 �m).
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 258

Helminthological Techniques 259
be read through the faecal sample (Fig.126h); alternatively, a rubber bung can beused. Carefully remove the slide by gentlysliding it sideways and leaving it for 20–30min at 37°C or 1 h at room temperature forthe faeces to clear progressively. Hookwormeggs clear first and will not be visible by 2 h,while Ascaris, Trichuris and schistosomeeggs will still be visible weeks or evenmonths later. The slide can be turned overon to filter-paper to prevent drying tem-porarily. In a modification to allow almostimmediate observation and long storage, amixture of 100 ml polyethylene glycol,100 ml formalin and 200 ml saturated saline(plus 2 ml colour, as above, if desired) canbe substituted.
The smear should be examined in a sys-tematic manner and the number of eggs ofeach species recorded. Multiply by 20 togive eggs per gram. (A hole of 6.5 mm in a0.5 mm thick template will give a samplesize of 20 mg, which should be multipliedby 50. This has the advantage that clearingtime is reduced to 15 min.) Naturally, ifquantitative measurements are notrequired, the template is unnecessary.
Concentration methods
Concentration methods are useful forprevalence surveys, but are not always use-
ful for measuring the intensity of an infec-tion because so many eggs may be presentin a sample that they cannot be easilycounted.
Formol–ether technique
The advantage of this method is that it isquick and can be used to search for proto-zoal cysts and Strongyloides larvae, as wellas helminth eggs. Using an applicator stick,1 or 2 g of faeces is emulsified in a cen-trifuge tube with 10 ml of 10% formalin.The suspension is then passed through ametal or plastic sieve (400 µm mesh) ortwo layers of gauze (for single use) into abeaker, evaporating dish or another cen-trifuge tube. Replace into another cen-trifuge tube (if necessary). Add more 10%formalin to bring up to 10 ml again, and3 ml of ether or ethyl acetate (or evenpetroleum, if the others are unavailable).Ether is very volatile and can explode inthe refrigerator so great care is needed. Thetube is then stoppered and shaken vigor-ously for at least 30 s. It is then centrifugedat 300–500 r.p.m. for 1–3 min until thereare four layers: a top layer of ether or ethylacetate, a second layer consisting of a fattydeposit, a third layer of formalin and asmall deposit containing the eggs at thebottom. The fatty deposit at the interface ofthe liquids is loosened with a glass rod and
Fig. 126. Illustrations of Kato–Katz technique: (a) Equipment, (b) Strips of hydrophilic cellophane and ajar with lid, (c, d, e, f, g. and h). Pressing faeces through sieve so that an amount above sieve can be putinto template, covered with cellophane and compressed so that a newspaper can be read through it.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 259

260 Chapter 9
the centrifuge tube quickly inverted at anangle of 45° to the ground so that the liq-uids run out (Fig. 127). Drops of water froma tap can be run down the glass tube whilethe debris is being cleaned off from thetube with the rod (the liquid from manysamples can be poured into a small beaker,centrifuged and the ether recovered andused again if in short supply). Some waterwill run up the rod by capillary action andthe deposit at the bottom of the tube isshaken and an aliquot removed with aPasteur pipette when the tube is uprightand placed on a microscope slide, coveredwith a large coverslip and examined usinga �10 objective. If there is no tap handy,the debris at the interface can be loosenedwith an applicator stick and the contents ofthe tube carefully poured away, allowingone or two drops to return to the bottom.For protozoal cysts, a drop of 1% Lugol’siodine, or Sergeaunt’s stain for amoebae,can be added and the sample examinedunder a higher power.
Zinc sulphate centrifugal flotation method
This method is not suitable for trematodeor Diphyllobothrium eggs, as they becomevery distorted.
1. A faecal suspension is made by emulsi-fying 5–10 g of faeces with about ten times
its volume of tap water, using a pestle andmortar.2. About 10 ml is strained through a gauzeor sieve (350 µm pore size) into a round-bottomed centrifuge tube.3. After centrifugation for 1 min the super-natant is discarded and 2–3 ml of wateradded. After breaking up the deposit bytapping the bottom of the tube, more wateris added to fill the tube.4. The previous step is repeated until thesupernatant is clear.5. The last supernatant is poured off and2–3 ml of 33% zinc sulphate (or saturatedsalt solution) added. The sediment is brokenup by tapping the tube and then this is filledwith more of the zinc sulphate solution.6. The tube is centrifuged rapidly for 1min and brought to a stop.7. A loopful of the material floating on thesurface is taken up and placed on a slidefor examination. The loop should be about6 mm in diameter and of wire that is stiffenough not to bend in routine handling.The loop is bent at right angles to the wireand must not be dipped below the surfaceof the liquid.
Quantitative techniques in faecalexaminations
For the schistosomes and most of theintestinal nematodes, figures have beengiven of the average egg production perfemale per 24 h. Thus, in theory at least, byestimating the number of eggs in the faeces,a measure of the number of adults presentcan be obtained. An estimate of the numberof worms present can be of importance indetermining the success of chemotherapyand in epidemiological studies. The mostcommonly used quantitative egg-countingtechniques are given below.
Kato–Katz and formol–ether techniques
Kato–Katz can detect as few as 100 eggs g�1
of faeces and is now regarded as the stan-dard. As is true for all such methods, itassumes that eggs are randomly dispersedin the faeces, which appears to be true in
Fig. 127. Running supernatant off down a glassrod, leaving sediment behind at bottom of centri-fuge tube (a conical tube is often used).
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 260

Helminthological Techniques 261
most cases; allowance could be made forthe state of the faeces, as shown below forthe Stoll method, but this is not usuallydone. The formol–ether method can beused as a semi-quantitative estimate andhas the advantage that it is quick, manysamples being treated simultaneously, andthat it also picks up protozoal cysts. Thefaecal sample size should be 0.2 g and allthe deposit counted (or a specified numberof drops of the total in the pipette). Fig. 128shows good correlation between the twotechniques.
The Stoll dilution egg-counting technique
The method was originally devised in 1926for the study of hookworm infection and isstill one of the best available today.
However, it is not suitable for less thanabout 200 eggs g�1 of faeces.
1. Add 0.1 N sodium hydroxide solutionup to the 56 ml mark in a Stoll dilutingflask. The diluting flask is made from aEhrlenmeyer flask with a neck of 20 mmdiameter glass tube fused on the top andgraduated at 56 and 60 ml (Fig. 129). Theyare closed with a rubber stopper.2. Faeces is carefully added with an appli-cator stick until the meniscus has been dis-placed to the 60 ml mark. For greateraccuracy 4 g of faeces can be weighed out.3. A few glass beads are added, a rubberstopper inserted and the flask well shaken.It is set aside for several hours preferablyovernight, and then shaken again to thor-oughly comminute the specimen.
5
4
3
2
1Light Mod. Heavy V. heavy
54321Occ.
Estimate (formol/ether)
0
Log
no. o
f egg
s g–
1 (K
ato)
Ascaris
Necator
Trichuris
Fig. 128. Comparison of eggs per gram (e.p.g.) of faeces as determined by Kato–Katz technique andestimate by formol–ether method on the same 40 faecal samples (occasional = 1–500 e.p.g.; light =501–2500; moderate = 2501–10,000; heavy = 10,001–20,000; very heavy = 20,000 +). The goodcorrelation shows that a quick approximate procedure is likely to be satisfactory in many surveys.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 261

262 Chapter 9
4. The flask is shaken well to resuspendthe specimen and 0.075 ml removed with awide-bore Pasteur pipette graduated at thislevel. This must be done with one squeezeof the bulb to prevent loss of eggs.5. The drop is expelled on to a microscopeslide and covered with a 22 mm � 40 mmcoverslip. All the eggs in the preparation arecounted, using a low-powered objective. Itis desirable to count several drops to obtainan average count from each specimen.6. The stool samples can be classified asformed (F), soft formed (SF), mushy (M) ordiarrhoeic (D) and the egg counts multi-plied by the following factors – SF � 1��, M � 2 and D � 3 – to compensate for theamount of fluid in these types of specimen.The results are multiplied by 200 forexpression as eggs per gram. Also 24 h fae-cal samples can be weighed and the totaloutput per 24 h obtained.
McMaster counting chamber
The chamber is commercially available andconsists of two glass slides kept 1.5 mmapart by glass bridges at the ends and inthe centre. A 1 cm2 square is ruled on theunderside of the top slide of the two cham-bers. The volume beneath each ruled area
is therefore 0.15 cm3. It is used principallyin veterinary studies and is not suitable forheavy eggs such as those of Fasciola.
1. 2 g of the well-mixed faecal sample isweighed out and placed in a glass jar con-taining a few glass beads. Alternatively, apestle and mortar can be used to mix thefaeces.2. 58 ml of a saturated salt solution (or a40% sugar solution or a 33% salt solution)is added and the faeces mixed to form asmooth emulsion.3. The suspension is passed through a400 µm metal or plastic sieve or gauze anda sample removed with a wide-borePasteur pipette.4. The suspension is run into the countingchamber, which is then set aside for 1–2min to allow the eggs to float to the surface.5. The eggs come to rest against the under-surface of the top slide and are in the samefocal plane as the two ruled areas. They arecounted using a low-power objective.
Macdonald pipette
This is an extremely rapid and simple fieldprocedure provided there is a glass-blowerwho can make lots of the special pipettes(Fig. 130).
Five grams of faeces are weighed out and45 ml of tap water measured out. The faecesis mixed with some of the water in a pestleand mortar, transferred to a closable jar andwashed out with more water. The total vol-ume is made up to 45 ml, the jar closed andshaken vigorously and 0.1 ml samples arewithdrawn using a Macdonald pipette andexamined on a microscope slide under lowpower. The mean figure is multiplied by100 to give eggs per gram. With freshlypassed faeces the pestle and mortar can bereplaced by shaking with 45 ml of waterand some glass beads.
Diagnosis of Larvae in Human Faecesby Culturing to the Infective Stage
It is necessary to culture larvae to the infec-tive third stage before a differential diagno-Fig. 129. Stoll flask.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 262

sis of hookworm species (and possiblyOesophagostomum, Ternidens andTrichostrongylus in some areas) can bemade. A simple method of accomplishingthis is the Harada–Mori test-tube filter-paper method.
1. Test tubes are filled with 7 ml of dis-tilled water and placed in a rack.2. About 0.5 g of the fresh faecal sample issmeared along the length of a strip of filter-paper (15 cm � 1.5 cm) creased longitudi-nally, leaving 5 cm clear at one end. Thestrip is placed in the test-tube with theunsmeared portion reaching to the bottomof the tube.3. The top of each is covered with a cello-phane strip, held in place by a rubber bandand the rack of tubes incubated at 28°C for10 days.4. The top and filter-paper are removed withforceps and both destroyed (use caution inhandling as the larvae are now infective).5. The larvae will be in the water at thebottom of the tube and can be seen withthe aid of a magnifying glass.
6. The tube can be placed in water at 50°Cfor about 15 min in order to kill the larvaewithout distorting them.7. When cool, the tubes are shaken and thecontents transferred to a centrifuge tube,which is spun gently for 2 min.8. The supernatant is pipetted off and thesediment examined on a microscope slide,using a coverslip.
Alternative methods for Strongyloides lar-vae are given on p. 121 and the differentialfeatures used in identification are shown inTable 9 (p. 131).
Methods in Filariasis
Staining of microfilariae
Wet preparations made from a drop ofblood under a coverslip can first be exam-ined for the presence of microfilariae underthe microscope and are easily seen movingunder the low power (�100 magnification).This method avoids the lengthy process ofstaining negative material, but many lightinfections will be missed.
Method A: Giemsa
There are various Romanowsky stains thatmay be used for staining blood parasites(Wright, Leishman and May-Grunwald), butGiemsa is one of the best for microfilariae.
The sheaths of Loa loa microfilariaenever stain by this method. The sheaths ofWuchereria stain faintly, while those ofBrugia malayi stain an intense pinkcolour.
1. A large drop of blood (20 mm3 ) from aSahli-type pipette is spread out to form athick smear (25 mm � 7 mm) on a grease-free microscope slide by means of a needle.2. The smear is dried overnight (be carefulof cockroaches and ants in the tropics),dehaemoglobinized in distilled water forabout 2 min and, when completely colour-less, left to dry.3. The smear is preserved with absolute ormethyl alcohol by flushing for 30 s andallowed to dry.
Helminthological Techniques 263
Fig. 130. Macdonald pipette.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 263

4. Ten drops of concentrated Giemsa stainare added to 10 ml of distilled water (pH7.2) and placed into a staining dish. Theblood film is left in the stain for 30 min.5. The slide is removed from the stain,flushed briefly and very gently under run-ning water and left to dry.6. When completely dry, the film can bemounted in a neutral mounting medium(DPX or euparol) with a cover glass.
Method B: haematoxylin
1. A thick drop made from peripheralblood is spread over an area of about 2 cm2
on a microscope slide and allowed to dry inair, preferably overnight. (For special pur-poses or if it is necessary to measure micro-filariae, a sequestrinated blood film can beleft in a humid chamber overnight at 10°Cand then left to dry. Alternatively, a thinsmear can be fixed while wet in three partsof methyl alcohol to one part of glacialacetic acid.) It is important when reportingmeasurements of microfilariae to state themethod of preparation and fixation. 2. The smear is dehaemoglobinized in tapwater and again dried. It is then fixed for afew minutes in methanol or absolute alco-hol (fixed smears can be stored formonths). 3. When dry, the slide is stained with hothaematoxylin (Mayer, Delafield, Ehrlich orHarris). The staining time is variable,depending on the batch of stain and on theage of the film, but 10–15 min is usuallysufficient.4. The slide is rinsed in tap water andexamined under the low power of a micro-scope. If understained, more hot haema-toxylin can be applied and, if overstained,the slide can be differentiated in acid alco-hol (1% hydrochloric acid in 70% alcohol).It is better to slightly overstain and thendifferentiate as this removes the blue back-ground from the film.5. The slide is blued in tap water (adddrop of ammonia if not alkaline) for 15 minand allowed to dry.6. The slide can be mounted in a neutralresin mountant.
Microfilarial counting techniques
1. For quantitative studies, 20 mm3 of bloodcan be taken in a haemoglobin pipette tomake a thick film, stained as above and theresult expressed in microfilariae per millil-itre of blood. However, this is time-consum-ing and a proportion of microfilariae are lostduring the staining procedure.2. One of the simplest and most sensitivemethods of detecting microfilariae (downto one microfilaria 100 mm3) is to mix ameasured quantity of blood with about tentimes its volume of distilled water in a sim-ple slide counting chamber and then toexamine immediately. The chamber can bemade by sticking thin strips of a glass slidewith an adhesive to form a rectangle onanother slide at about �30 magnification. Itis difficult to identify microfilariae by thismethod and, if necessary, stained prepara-tions may need to be made as well. 3. If very few microfilariae are expected, asin cases of amicrofilaraemic filariasis,1–10 ml of blood can be filtered through aNuclepore or Millipore membrane filter(pore size 5 µm) fitted to a hypodermicsyringe by a Swinney adaptor and the mem-brane examined either fresh or stained asabove. Using the Nuclepore membrane(General Electric Company, Pleasanton,California), sequestrinated or citrated bloodis passed through the filter on a syringe, fol-lowed by 0.85% saline solution. Finally, airis passed through to partially dry the mem-brane, which can be examined directlyunder a coverslip using �50–70 magnifica-tion, or fixed in methanol, stained in hothaematoxylin, blued, dried and a drop ofchloroform added in order to make themembrane transparent before viewing. Thismethod can detect microfilariae in a con-centration 1/50 that of a thick film. 4. Knott’s concentration technique hasbeen widely used for the last 60 years but isnot as sensitive as the previous techniques.In a quick modification, 1 ml of blood islysed with 10 ml of 1% formalin in a cen-trifuge tube, centrifuged at 500 g for 1 minand the supernatant poured off. The sedi-ment is then spread out on microscopeslides and dried and stained as above.
264 Chapter 9
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 264

5. Another modification of the Knott tech-nique involves spinning in QBC® tubesand examining the buffy layer under fluo-rescent light (Bawden et al., 1994).
Staining of filarial larvae in insects beforedissection
1. Insects are killed and identified andplaced in 80% alcohol, where they can bekept indefinitely.2. They are taken through descendingdilutions of alcohol to distilled water andare then stained for 3 days at room temper-ature in Mayer’s acid haemalum stain.3. Insects are removed and differentiatedin distilled water for 3 days. They are thenstored in glycerol until required for dissec-tion; 100–200 mosquitoes can be stored ina McCartney bottle.4. Insects are dissected under the stereo-scopic microscope (�35 magnification) byreflected light, using fine needles. Allstages of developing larvae are stained andcan be easily detected.5. Larvae should be mounted in glycerolfor detailed morphological study.
Technique for mass dissection of filarialvectors and mounting of filarial larvae
obtained
1. Insects are immobilized by cold or by apyrethrum spray and then sorted into sepa-rate species. All females of one species arecrushed by gently rolling with a test-tubeon a glass or Perspex plate in physiologicalsaline. This is repeated for each speciesseparately.2. Insects are transferred to sieves (boltingsilk with a pore size of 50 µm is suitable),glued across the bottom of the top half of acut plastic cup and suspended in aclamped glass funnel containing physiolog-ical saline. Care must be taken that thereare no air bubbles below the sieve. Eachsieve is left for 3 h at room temperature(the procedure can also be used for obtain-ing larvae for infection experiments, whena tissue culture medium should be used,with incubation at 37°C).
3. About 1 ml is run into a Syracusewatch-glass (these can be made by a glass-blower by pressing in the end of a heatedtest tube and cutting the end off) andexamined for larvae under a stereoscopicmicroscope. 4. Most of the saline is removed with aPasteur pipette under the microscope andreplaced by fixative. TAF (7 ml of 10% for-malin, 2 ml of triethanomaine and 2 ml ofdistilled water) is excellent but 70% alco-hol containing 10% glycerol can be used.Fixation is for 5 min.5. Fixative is removed with a fine pipetteand replaced by 96% alcohol containing5% glycerol and the watch-glass is left inan incubator for 30 min at 37°C (orovernight at room temperature) until onlypure glycerol is left. Larvae can be stainedwith new fast blue R or examined whenmounted under phase contrast.6. Larvae are transferred with a mountedcat’s whisker to a small drop of anhydrousglycerol on a clean 22 mm square cover-slip. Excess glycerol is removed with avery fine pipette, leaving only a smear.7. The coverslip is turned over on to a cav-ity slide and sealed around and under theedge with mounting medium. For cheap-ness, three short pieces of glass wool canbe used instead of using a cavity slide or asmall chamber can be made.
Caution needs to be exercised when thistechnique is used for Onchocerca volvulusin Simulium as larvae may be of the morpho-logically indistinguishable O. ochengi.
Preservation and Examination ofAdult Helminths
Before fixing, specimens should preferablybe washed in 0.85% saline.
Trematodes and cestodes
For examining the various organ systems inwhole mounts of these organisms, somedegree of flattening is usually necessarybut, as this causes distortion, materialshould also be fixed unflattened for
Helminthological Techniques 265
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 265

subsequent sectioning. Extension of trema-todes can be obtained by constant shakingand of cestodes by suspension or by wind-ing around a test-tube, while fixing in hot5–10% formalin.
Staining trematodes or tapeworm segments
METHOD1. The trematode or proglottides should beflattened between glass microscope slidesheld at the ends with rubber bands or, forsmaller specimens, with a dab of petro-leum jelly at each end of the slide. Theyare fixed in 80% alcohol or 10% formolsaline (a few drops of acetic acid added toeither of these fixatives improves the stain-ing) or Carnoy’s fixative.2. The flattened specimen is taken frombetween the slides when fixed andimmersed in the diluted aceto alum carminestain in a small glass dish. The staining timedepends in each case on the size of the spec-imen – from 20–30 min for a small trema-tode to several hours for larger specimens.3. Specimens are washed in 50% alcoholto remove excess stain and examinedunder the microscope.4. If understained the specimen can bereturned to the stain, and if overstained itcan be differentiated in acid alcohol (100 ml70% alcohol and 2 ml hydrochloric acid). 5. When stained, the specimen is washedwell in water to remove acid, dehydrated inseveral changes of alcohol, cleared in beech-wood creosote and mounted in Canada bal-sam or another mounting medium.
RESULT
The internal structures of the specimensare stained in precise shades of red againsta clear to pink background of the connec-tive tissues.
The above technique may be carried outusing alum haematoxylin instead of thecarmine. Ehrlich’s haematoxylin dilutedwith an equal volume of water has provedsatisfactory for this method.
ACETO ALUM CARMINE STAINBoil together:
Potassium aluminium sulphate 27 g
Carmine 17 gDistilled water 200 g
These should be gently heated in a largeflask, as the mixture is liable to froth whenboiling. The mixture is boiled until thecolour changes from the bright red of thecarmine to a deep red colour. When cool,20 ml of glacial acid is added. For use thestain is diluted to a rose-pink colour (1 in 5to 1 in 10).
Nematodes
If possible, nematodes should be looked atalive and measurements made; small speci-mens can often be immobilized by adding adrop of propylene phenoxetol or tetramisole.
Fixation
1. Hot 70% alcohol with constant shaking.Simple and usually satisfactory.2. Hot formol–acetic 4:1 (formalin 40% 10ml; glacial acetic acid 1 ml; distilled waterup to 100 ml) over a boiling water-bath.3. TAF (formalin 40% 7 ml; triethanolamine2 ml; distilled water to 100 ml). Hygroscopicso prevents drying of specimens.
Storage
Storing in 70% alcohol with 10% glycerolor pure glycerol will prevent drying up;10% formalin is not as satisfactory, asspecimens go mushy after a time.
Mounting
1. Temporary (these mounts are probablythe most satisfactory).(a) Glycerine. Worms are left overnight ina partly opened Petri dish or solid watchglass in 70% alcohol plus 10% glycerol inan oven at about 40°C, mounted in pureglycerol and returned to the bottle afterexamination. Specimens can be turnedover for examination of all sides and exam-ined under phase contrast.(b) Lactophenol. Some lactophenol is addedto the specimen in 70% alcohol and slowlyreplaced with pure lactophenol over a fewhours until specimen clears (hot lactophenol
266 Chapter 9
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 266

at 65°C may be used for rapid clearing).Worms are replaced in 70% alcohol afterexamination.2. Permanent.(a) Mounts are made in Canada balsam orDPX after dehydration slowly through alco-hols and xylene. Specimens can be stainedwith haematoxylin if desired; however, theresults often leave much to be desiredbecause of distortion.(b) Specimens are transferred to polyvinyllactophenol from lactophenol. However,specimens go on clearing until completelytransparent. Cotton blue can be dissolvedin the polyvinyl lactophenol for staining.(c) Glycerine. A tiny drop of glycerol on acoverslip with a cavity slide is used for lar-vae, and ringed slides for adults. Resultsare good but the preparations are ratherfragile.(d) Glycerine jelly. A tiny warmed drop ona coverslip is used for en face mounts ofthe anterior end on a cavity slide(Aquamount needs no heat).
Recognition of Helminths in Sections
Before a tentative diagnosis is made of pos-sible helminth material seen in a section, itis important that the size of the specimenis considered. It is useful first to look at theslide against the light before placing it onthe microscope stage; the cysticercus larvaof Taenia, for instance, can often be recog-nized in this way.
The descriptions given below are basedon sections stained in haematoxylin andeosin.
Trematodes
In sections of living trematodes an outertegument, often with spines, is always pre-sent. Depending on the region of the wormcut through, one or more of the followingorgans should also be seen embedded in aloose ground tissue (parenchyma): vitel-laria; gut caeca (usually double); ovaries;testes; suckers; uterus, probably filled witheggs. The diagram of a complete fluke (Fig.
2) should be consulted in order to under-stand which organs will be present incross-sections cut at different levels (thereis some variation in different species, par-ticularly in the relative positions of theovaries and testes).
If the organism in the tissues is deadand surrounded by a host reaction, thenidentification is more difficult. However, aparasite can usually be identified as aplatyhelminth or nematode even if noorgans are recognizable.
Cestodes
Larval cestodes in humans may be found inany tissue. The most important larvaeencountered in sections are the hollowcysts of taeniid tapeworms, which includethe cysticercus of Taenia solium, the coenu-rus of Multiceps spp. and the hydatid ofEchinococcus spp. In sections the proto-scolex (or protoscolices) will usually showclearly demarcated suckers and possiblyhooks (the latter are birefringent and showup well under a polarizing microscope).The characteristic structure of each type ofcyst can be seen from Fig. 32. Two solid lar-vae may also be found in tissues (ignoringthe cysticercoid of Hymenolepis nana,which occurs only in the villi of the ileum):the tetrathyridium of Meocestoides and thesparganum (plerocercoid) of Spirometra.These may show few diagnostic features,but the former have many large calcareouscorpuscles and perhaps suckers and the lat-ter the circular muscles which separate theparenchyma into inner and outer zones,typical also of adult tapeworms.
Nematodes
Adults encountered in sections can usuallybe easily recognized as belonging to thephylum. In cross-section they are seen tohave an outer acellular cuticle, enclosing athin hypodermis layer, which often con-sists of longitudinal cords that separate thesomatic muscles into longitudinal fieldsand contain the excretory canals (when
Helminthological Techniques 267
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 267

268 Chapter 9
present). The number and form of the mus-cle cells are an important feature in deter-mining to which order a specimen belongs.The gut is separated from the outer layersby a body cavity (pseudocoel).
The characteristic structures seen incross-sections of the various groups ofnematodes are shown in Fig. 131. The mostcommon adult nematodes recovered from
the tissues are filariae; females of non-human species are usually not fertilized buthave the double uterus typical of the group.A key for differentiating the larvae ofAscaris, Toxocara, Necator, Ancylostomaand Strongyloides in tissue sections can bemade based on their relative diameters, thestructure of the excretory cells and the pres-ence or absence of lateral alae.
Fig. 131. Cross-sections of females belonging to the various groups of nematodes. The dorsal andventral cords are not shown (Trichocephalus = Trichuris).
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 268

Helminthological Techniques 269
Identification Key for Sections ofNematodes
1. A. Hypodermal cords few to manyarranged asymmetrically. Lateral excre-tory canals absent. In the ovaries andtestes the germinal region extends theentire length of the gonad and gametesat various stages of development can befound at any level. Oesophagus non-muscular. TRICHURIDS
(Aonchotheca, Calodium, Eucoleus,Trichinella, Trichuris)
B. Hypodermal cords 2–4 arranged sym-metrically. Excretory canals in lateralcords except at extremities. In theovaries and testes the germinal regionconfined to proximal extremity of gonadand gametes at uniform stage of develop-ment at any one level. 2
2. A. Intestine composed of a few largemultinucleate cells rarely with morethan two cells in a cross-section.Anterior paired excretory gland cellspresent. STRONGYLIDS
(hookworms, trichostrongyles,Oesophagostomum, Ternidens)
B. Intestine composed of few to manysmall uninucleate cells. Excretoryglands single, paired or absent. 3
3. A. Somatic muscle cells few, large andflat. Lateral alae present or absent. 4
B. Somatic muscle cells many, U-shaped in cross-section. Lateral alaeusually absent. 5
4. A. Lateral alae present. Vagina long andmuscular. Oesophagus has posteriorbulb. Many eggs in uterus. Excretorygland cells absent. Enterobius
B. Lateral alae absent. Vagina very short.Ova large. Ovoviviparous, with eggsdeveloping into larvae. Excretory cellssingle or paired. Very small and often lar-vae (or eggs) in tissues. Strongyloides
5. A. Somatic musculature divided bylarge lateral cords (often very wide andflattened) into two crescent-shapedfields. 6
B. Somatic musculature divided intofour fields. 7
6. A. Females have double uterus oftencontaining microfilariae. Diameter lessthan 0.5 mm. FILARIIDS
(Brugia, Dirofilaria, Loa, Mansonella,Wuchereria)
B. Females with single uterus contain-ing larvae with long pointed tails.Diameter more than 1 mm. Dracunculus
7. A. Lateral cords very large, stalked andproject into body cavity; frequentlyunequal in size. SPIRURUDS
(Gnathostoma, Thelazia)
B. Lateral cords large and project intobody cavity but not stalked; usuallyunequal in size . ASCARIDS
(Anisakis, Ascaris, Toxascaris, Toxocara)
References and Further Reading
Ash, L.R. and Orihel, T.C. (1991) Parasites: a Guide to Laboratory Procedures and Identification.ASCP Press, Chicago, USA.
Bawden, M., Slaten, D. and Malone, J. (1994) QBC®: rapid filaria diagnoses from blood. Mansonellaozzardi and Wuchereria bancrofti. Transactions of the Royal Society of Tropical Medicine andHygiene 88, 66.
Cassella, J.P. and Hay, J. (1995) Tissue eosinophil and mast cell staining in helminth infections.Parasitology Today 11, 464–465.
Chitwood, M.B. and Lichtenfels, J.R. (1972) Identification of parasitic metazoa in tissue sections.Journal of Parasitology 32, 407–519.
Esterre, P., Vignes, R. and Lebbe, J. (1987) Identification assistée par ordinateur de principaux oeufsd’helminthes en coprologie humaine. Bulletin de la Société Française de Parasitologie 5, 241–244.
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 269

Gillespie, S.H. and Hawkey, P.M. (eds) (1995) Medical Parasitology: a Practical Approach. IRL Press,Oxford, UK.
Petithory, J.C. and Ardoin-Guidon, F. (1995) Parasitologie: vrais et faux parasites en coprologiemicroscopique. Cahier de Formation – Biologie Médicale no. 3. Bioforma, Paris, France.
Price, D.L. (1993) Procedure Manual for the Diagnosis of Intestinal Parasites. CRC Press, Boca Raton,USA.
WHO (1991) Basic Laboratory Methods in Medical Parasitology. World Health Organization, Geneva,Switzerland.
270 Chapter 9
09Worms & H.D. - Chap 09 14/11/01 4:30 PM Page 270

1500 BC Ebers papyrus possibly described various helminth diseases.400 BC–AD 1200 Taenia, Ascaris, Enterobius and Dracunculus described in Greek,
Roman, Arabic and Chinese writings.AD 1379 Jehan de Brie illustrated Fasciola hepatica from sheep.1674 Textbook by Velschius on Dracunculus.1684 Redi described many new helminths from domestic and wild animals.1700 Textbook by Andry on human helminths.1835 Paget discovered larval Trichinella spiralis in a man, which was described
by Owen.1845 Dujardin elucidated the relationship between adult and cycticercus of
Taenia saginata.1851 Küchenmeister, Van Beneden, etc. described the life cycle of Taenia solium.1851 Adult Schistosoma haematobium were found in the vesical veins by
Bilharz in Cairo.1852 Life cycle of Echinococcus granulosus determined by Von Siebold.1857–1898 Textbooks by Küchenmeister (1857), Davaine (1860), Cobbold (1864),
Leuckart (1867) and Manson (1898).1865 The life cycle of Enterobius was worked out by Leuckart.1870 Fedchenko observed larval development of Dracunculus in Cyclops.1876 Adult Wuchereria bancrofti found in lymph glands by Bancroft; surpris-
ingly in Australia.1878 Life cycle of Wuchereria bancrofti shown to involve mosquito by Manson.1882–1883 Leuckart and Thomas showed that the life cycle of Fasciola hepatica
involves a snail.1887 Hymenolepis nana shown not to need an intermediate host by Grassi
and Rovelli.1896 Looss demonstrated the active penetration of hookworm larvae through
the skin (by accidentally infecting himself).1897 Eosinophilia first associated with Trichinella spiralis infection by Brown.1900 W. bancrofti infective larvae shown to enter through the puncture
wound of mosquito by Low.1901–1913 Weinland’s studies on Ascaris biochemistry and physiology.1904 Adult Schistosoma japonicum found by Katsurada.1907 Schistosoma mansoni differentiated from S. haematobium by Sambon.1909 Rockefeller Sanitary Commision established to combat hookworm disease.1911 An intradermal test for the diagnosis of hydatid disease described by
Casoni.
Appendix 1Summary of Some Landmarks in
Medical Helminthology
271
10W. & H.D. - Appendix 1 14/11/01 4:30 PM Page 271

1912 Complement fixation test used for diagnosis of Trichinella.1911–1919 Role of snail and fish in the transmission of Clonorchis and of snail and
crab in that of Paragonimus determined in Japan.1913 Life cycle of S. japonicum involving snail discovered by Miyagawa.1914–1921 Life cycle of Loa loa involving Chrysops shown by Leiper, Kleine and Connal.1915 Leiper elucidated the life cycle of S. haematobium.1918 Connection between blindness and onchocerciasis recognized by
Pacheco-Luna and Robles in Guatemala. Antimony tartrate advocated for treatment of schistosomiasis byCristopherson.
1918 Life cycle of Diphyllobothrium latum determined by Rosen and Janicki.1920 Chandler suggested copper sulphate for the control of snail intermediate
hosts of trematodes.1921 Migration of Ascaris larvae through body discovered by Koino and Koino.
Life cycle of Fasciolopsis buski worked out by Nagagawa.1926 The vector of Onchocerca volvulus shown to be Simulium by Blacklock.1927 Brug found microfilariae of Brugia malayi in blood.1928 Stoll described ‘self-cure’ phenomenon in gastrointestinal nematodes of
domestic stock.1929 Faust’s Human Helminthology published.1930s–1950s Work of Culbertson, Taliaferro, Vogel and Minning on mechanisms of
immunity to helminth infections.1932 Löffler described pulmonary lesions and eosinophilia due to larval
migration of Ascaris.1934 Development of Mansonella in Culicoides shown by Buckley.1947 Diethylcarbamazine (DEC) first used in treatment of filariasis.1951 Piperazine used for treatment of Ascaris and Enterobius.
Role of Angiostrongylus cantonensis in causation of eosinophilic menin-gitis elucidated by Alicata and by Rosen and coworkers.
1952 Importance of Toxocara larvae in visceral larva migrans shown by Beaver.1954 Onchocerciasis eradicated from Kenya by McMahon.
Increasing use of new synthetic insecticides (chlorinated hydrocarbonsand organophosphates) in control of filariasis. Discovery of new synthetic molluscicides for the control of schistosomiasis.
1956 Work by Buckley and Edeson on tropical pulmonary eosinophilia andzoonotic aspects of Brugia infection (genus Brugia erected by Buckley in1959).
1964 Discovery of intestinal capillariasis in the Philippines.1971 Discovery of abdominal capillariasis in Costa Rica.
For a much fuller account the book by Grove (1990) (see p. 284) should be consulted.It is not always easy to assess the importance of recent work but some of the most
notable advances in medical helminthology have been in the fields of immunity, immuno-diagnosis and immunopathology, the search for human vaccines, the action of broad-action anthelminthics, such as praziquantel, the benzimidazoles and ivermectin, theimportance of strain differences in many human helminths, such as Echinococcus,Trichinella and Onchocerca, the epidemiology and medical importance of soil-transmit-ted nematodes, molecular biology, including the sequencing of the complete genome ofthe free-living nematode Caenorhabditis elegans, and the global control campaigns,whether under way or envisaged, for onchocerciasis, dracunculiasis, lymphatic filariasis,schistosomiasis and geohelminths.
272 Appendix 1
10W. & H.D. - Appendix 1 14/11/01 4:30 PM Page 272

aberrant parasite A parasite that is nevertransmitted between humans and whichdevelops abnormally in humans (e.g.Echinococcus multilocularis, Parastrongylus,Toxocara).accidental parasite One that is not usuallytransmitted between humans but whichdevelops in a similar manner to that in itsnormal host (e.g. Fasciola, Dipylidium,Taenia solium cysticerci, Trichinella).aggregation See overdispersal.alae (‘wings’) Extensions of the cuticle ofnematodes (e.g. in Enterobius); sometimespresent only at the head (cervical alae, e.g.Toxocara) or tail end (caudal alae, e.g. maleDirofilaria).amphid One of a pair of receptors situatedat the anterior end of nematodes.anthelminthic Any chemotherapeutic sub-stance directed against a helminthicinfection.antibody An immunoglobulin moleculethat recognizes a specific antigen.antigen A molecule that is foreign to thebody and is recognized. Because of thelarge size of helminths in relation to invad-ing microorganisms, most functional anti-gens of helminths are excretory orsecretory. The tegument of Platyhelminthesis also a site for attack by host defences,but the tough cuticle of nematodes islargely resistant.autoinfection Reinfection without expo-sure from the environment; self-infection
(e.g. Strongyloides, Aonchotheca andHymenolepis nana).carrier A host that harbours a particularpathogen without manifestations of disease.caudal bursa (= bursa copulatrix)Umbrella-like expansion of the caudal endof the male in strongylid nematodes, usedto clasp the female when mating.cercaria The free-living larva, usually pos-sessing a tail, which escapes from a sporocystor redia generation of a trematode within thesnail host, and constitutes the transfer stageto the next (usually definitive) host.cirrus Retractile muscular organ at the ter-minal end of the male reproductive systemof Platyhelminthes, which serves as a penisduring fertilization.coenurus Larval cystic stage of the tape-worm Multiceps, containing an inner ger-minal layer producing multiple scoliceswithin a single fluid-filled cavity (seecysticercus and hydatid cyst).commensal A symbiotic relationship inwhich one individual gains without havingbeneficial or detrimental effects on its partner.concomitant immunity A state of immu-nity that prevents establishment of a newinfection but is ineffective against anyexisting parasites that may be present (e.g.as in schistosomiasis).control Attack on an infection so that itwill no longer be clinically or economi-cally important (e.g. projected global cam-paigns against lymphatic filariasis or
Appendix 2Glossary of Helminthological Terms
These terms should be regarded as explanations rather than as strict definitions and donot necessarily apply to all other helminths not found in humans.
273
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 273

schistosomiasis plus intestinal helminths.See elimination or eradication).cuticle The outer chitinous body layer ofnematodes (see tegument).cysticercoid The solid second larval stageof tapeworms containing an invaginatedscolex. Usually tailed and found in inverte-brates (e.g. Dipylidium and Hymenolepis).cysticercus (= bladderworm) The hollowfluid-filled second larval stage of Taenia,containing a single invaginated scolex (seecoenurus, hydatid cyst and metacestode).digenetic Three or more generations (liter-ally two) required for completion of onelife cycle, as in digenetic trematodes.dioecious Having female and malereproductive organs in differentindividuals (e.g. schistosomes andnematodes).domestic cycle (= pastoral or synanthropiccycle) Life cycle of a zoonotic parasiteinvolving domestic stock or pets, with ahigh risk of infecting humans (e.g. usuallyin Echinococcus granulosus and Trichinellaspiralis).ectoparasite Living upon or in the superfi-cial tissues of a host.ectopic site Outside the normal location,as in the position of a parasite that lodgesin an atypical part of the body.egg count A quantitative measure of thenumber of eggs present in the faeces, usu-ally expressed as eggs per gram, in order toestimate the number of adults present.ejaculatory duct The muscular terminus ofthe male genitalia of nematodes, openinginto the cloaca.elimination Campaign resulting in thecomplete absence of an infection in a coun-try or region (e.g. Dracunculus in Asia). Canalso refer to absence of disease rather thanof the causative parasite (e.g. intention ofglobal elimination of lymphatic filariasiscampaign). (See control and eradication.)embryo The stage in development follow-ing cleavage of the zygote up to, but notincluding, the first larval stage.embryophore In tapeworms, the envelopeimmediately around the oncosphere andderived from it. In taeniids it forms a thickprotective layer composed of a keratin typeof protein.
endoparasite Living within another organ-ism, including the digestive and respira-tory tract of the latter.eradication Complete elimination of ahelminth infection throughout the world(e.g. hopefully Dracunculus soon. (See con-trol and elimination.)evasion of the immune responseMechanisms by which helminths avoid theimmune response of the host either byoccupying a privileged site or by coveringthemselves with host or host-like antigens(e.g. schistosomes).feral cycle (= sylvatic cycle) Life cycle of azoonotic parasite involving wild animalsand having a low risk of infecting humans(except hunters) (e.g. Echinococcus multi-locularis) (see domestic cycle).filariform larva A non-feeding nematodelarva characterized by its delicate elongatestructure and by its slim, capillary oeso-phagus without a posterior bulb (e.g. inhookworms, Strongyloides and filariae).flame cell See solenocyte.furcocercous cercaria Fork-tailed, as inthe cercaria of schistosomes. genital atrium In Platyhelminthes, thechamber collecting the terminal ends of thegenital tubules and opening to the exteriorby the genital pore.geohelminth A term applied to any nema-tode that is transmitted to humans afterdevelopment in the soil (e.g. Strongyloides,hookworms, Ascaris and Trichuris).gonotyl Genital sucker, retractile and asso-ciated with or incorporated into the ventralsucker, in some genera of trematodes (e.g.Heterophyes).gravid Filled with eggs, as in the terminalproglottides of a cyclophyllidean tape-worm or Enterobius.gubernaculum A small, sclerotinized,accessory structure in male nematodes,which serves to guide the spicules out ofthe cloaca.gymnocephalous cercaria Literally naked-headed; refers to cercariae without orna-mentation of body or tail, as in that ofFasciola hepatica.gynaecophoric canal In male schisto-somes, the ventral incurved fold of thebody extending from the ventral sucker to
274 Appendix 2
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 274

the caudal extremity, for carrying thefemale.hermaphroditic Containing both male andfemale organs in the same individual;monoecious (e.g. cestodes and trematodesexcept schistosomes).heterogonic Development in which bothmales and females are present in a colony(see hologonic).hexacanth embryo ‘Six-hooked’ embryo,the mature embryo containing six hookletswithin the egg of many tapeworms, includ-ing all species that parasitize humans.hologonic Development in which only onesex (usually the female) is present in acolony, as in parasitic Strongyloides.host An organism that harbours and nour-ishes a parasite.
definitive host One that harbours thesexually mature stage of the parasite (e.g.humans are the only definitive hosts forTaenia saginata).intermediate host An obligate host thatalternates with the definitive host andharbours the larval stage of the parasite(e.g. snails are intermediate hosts for allhuman trematodes).paratenic host A carrier or transporthost. One in which the parasite remainsviable but does not develop.reservoir host An animal host in whichinfection usually resides; also one whichharbours the infection when humans arenot infected (e.g. ruminants are reservoirhosts for many Trichostrongylus species).
hydatid cyst Larval cystic stage of thetapeworm Echinococcus granulosus, con-taining an inner germinal layer producingmany protoscolices, which when set freeinto the cystic cavity, develop into daugh-ter cysts and usually have a thick lami-nated wall surrounded by an outer fibrouscapsule of host origin (see coenurus andcysticercus).hydatid cyst, alveolar (= multilocular)Larval cystic stage of the tapewormEchinococcus multilocularis (AE), similarto the hydatid cyst (above) but in whichbudding is external (exogenous) as well asinternal (endogenous), so that many smallcysts are formed. There is no thick outercapsule.
hyperendemic High prevalence of infec-tion in a human community.hypodermis In helminths, the layer of tis-sue immediately below the tegument(trematodes and cestodes) or cuticle(nematodes).immunomodulation An alteration, eithernegative or positive, of the immuneresponse (see immunosuppresion).immunopathology Damage to tissuescaused directly or indirectly by immunemechanisms (the importance of this inhelminth infections is being increasinglyrecognized).immunosuppression A down-regulation ofthe immune response.incidence of infection The number of newcases of an infection reported in an area ina unit of time (taken to be a year if notgiven) (see prevalence of infection).infestation Existence of parasitic organ-isms on the outside of the body of a host orin the superficial tissues; ectoparasitism(e.g. lice and fleas).intensity of infection Number of parasitesharboured by a host. The mean intensity iscalculated for the whole host population(including those uninfected).larva The postembryonic stage of ahelminth, in which internal organs aredeveloping or are developed and are atleast partially functional.Laurer’s canal In trematodes, a tubuleleading from the dorsal surface to theregion of the ootype and seminal recepta-cle; it may be patent, vestigial or lacking. Itprobably functions as a copulatory canal.LD50 (= lethal dose 50) and LC50 (= lethalconcentration 50). The dose or concentra-tion of a chemotherapeutic, insecticidal ormolluscicidal agent estimated statisticallyto kill 50% of the helminths, insects orsnails being treated (also LD90 and LC90).Mehlis’ glands In Platyhelminthes, theglands surrounding the ootype. They pro-duce mucus to lubricate the uterus andprobably participate in eggshell formation.metacercaria The stage of trematodes fol-lowing the cercaria, with loss of tail, whichbecomes encysted and awaits passive trans-fer to the definitive host (present in alltrematodes of humans except schistosomes).
Glossary of Helminthological Terms 275
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 275

metacestode A collective term for any ofthe various larval stages of cestodes whichare found in the intermediate host (e.g.oncosphere, procercoid, plerocercoid,tetrathyridium, cysticercus, cysticercoid,coenurus, hydatid).microcercous cercaria Cercaria with ashort, stumpy tail (e.g. cercaria ofParagonimus westermani).microfilaria The first prelarval stage of afilaria, which either escapes from theeggshell (i.e. is unsheathed) or stretches theshell into an elongated sac (i.e. is sheathed)and is found in the blood or skin.miracidium The ciliated free-living first-stagelarva hatched from the eggs of trematodes.nosogeography Knowledge concerning thegeographical distribution of diseases. oncosphere The stage that hatches fromthe eggshell and later escapes from theembryophore of tapeworms; in humantapeworm infections it is six-hooked (i.e. ahexacanth embryo).ootype The chamber in the reproductivesystem of Platyhelminthes where the sev-eral components of the eggs are assembled.ovejector A muscular organ in somefemale nematodes that forces eggs from theuterus into the vagina.overdispersal Situation where most hostsharbour a few parasites while a few hostsharbour many parasites. Gives a negativebinomial distribution.oviparous Producing and laying eggs (seeovoviviparous).ovoviviparous Discharging living youngrather than eggs (e.g. Dracunculus, filariae,Strongyloides and Trichinella) (seeoviparous).papillae Small sensory knobs, either at theanterior end (dierids) or at the posteriorend in the male, where their number andposition may be of taxonomic importance.parasite An organism that lives at theexpense of another larger organism, itshost; which is physiologically dependenton its host, has a higher reproductivecapacity than it and may kill the host inheavy infections.
facultative One that may employ eithera free-living or a parasitic mode of life.e.g. Strongyloides.
obligate One that necessarily lives a par-asitic existence, e.g. all the other humanhelminths.
parasitocoenosis (= parasite mix) Theassemblage of parasites associated with ahost population. parenchyma In Platyhelminthes, theloose, usually undifferentiated tissue thatforms a matrix in which the viscera areembedded.parthenogenesis Production of progenyfrom the ovum without fertilization (e.g.parasitic female of Strongyloides).pastoral cycle See domestic cycle.patent infection Open or apparent, as indi-cated by unmistakable signs, like eggs inthe faeces or microfilariae in circulatingblood (see prepatent period).pathogen A parasite causing injury to ahost (see commensal and parasite).phasmid One of a pair of caudal organelles(sensory receptors?) found in most para-sitic nematodes (i.e. the Phasmidia),including all those in humans exceptAonchotheca, Dioctophyma, Trichinellaand Trichuris.pipestem fibrosis (Symmer’s) A fibroticreaction formed around a vein in schistoso-miasis, so called because of its resemblancein section to the stem of an old-fashionedclay pipe.plerocercoid larva A second larval stageof pseudophyllid tapeworms in which thescolex is embedded in a greatly enlargedtail (e.g. larva of Dyphyllobothrium latum)(see sparganum and procercoid larva).prepatent period The period that a para-site is present in the body before it givesrise to parasitological evidence of its pres-ence (e.g. eggs in the faeces or microfilariaein the blood).prevalence of infection The number of casesof an infection present in an area (actuallyper unit of population) at a fixed point intime. Prevalence = incidence � averageduration (see incidence of infection). proboscis In acanthocephalans and in thedog tapeworm Dipylidium caninum ananterior protrusible organ, typically stud-ded with hooklets.procercoid larva The first larval stage of pseudophyllidean tapeworms, which
276 Appendix 2
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 276

develops from the oncosphere; it consists ofa body proper and a caudal vestige of theoncosphere containing the hooklets called acercomer (see plerocercoid larva).proglottid One complete unit of a tape-worm, commonly called a segment. Incyclophyllideans the most anterior areimmature; further back they are mature,containing the male and female sex organs;and the posterior proglottides are gravid,being filled with eggs.pseudocoel Body cavity of nematodes, notlined completely with mesothelium andnot equivalent to a coelomic cavity.pseudotubercle A foreign-body reactionresembling a tubercle but not provoked bytubercle bacilli (e.g. by schistosome eggs).refractory host One that is not readilyinfectible by a particular parasite (see sus-ceptible host).retrofection A variety of autoinfection inwhich larvae hatch from eggs in the analregion, migrate up the large bowel anddevelop into adults (e.g. Enterobius).rhabditiform larva A feeding first-stagenematode larva (of strongylids andStrongyloides) in which the oesophagus isfunctional, is usually muscular and has anenlarged posterior bulb.rostellum The somewhat protuberant api-cal portion of the scolex of certain tape-worms, frequently bearing a circle ofhooklets, as in Taenia solium.schistosomulum Immature stage of schis-tosomes, from the time of entry into thedefinitive host until the worm reaches sex-ual maturity.scolex Attachment end of a tapeworm,commonly referred to as the anterior end orholdfast, which often bears suckers andhooks.seminal receptacle (= Receptaculum sem-inis) The storage reservoir for spermatozoain the female.seminal vesicle (= Vesicula seminalis) Thestorage reservoir for spermatozoa in themale.sensitivity (of diagnostic procedure) Theprobability that the procedure will diag-nose a real positive reaction.sign Objective evidence of disease (seesymptom).
solenocyte Literally canal cell. InPlatyhelminthes, the cell with a tuft of ciliaat the head of each capillary in the excre-tory system; commonly called a flame cell.sparganum The second plerocercoid larvalstage of the pseudophyllidean tapewormSpirometra, which occurs in mammals(particularly when in humans), character-ized by its elongated shape and lack of acystic cavity.specificity (of an immunodiagnostic test)The probability that a real negative host isrecognized by the test to be negative (i.e.number of real negatives divided by num-ber of real negatives plus number of falsepositives).spicules (copulatory) Two (or sometimes,in aphasmids, one) bristle-like, lanceolatesclerotinized structures in the outer genitalchamber of male nematodes, introducedinto the vulva or vagina of the female dur-ing copulation to guide the sperm.strobilization The production of proglot-tides from the neck region of tapeworms.superinfection New infection superimposedon an existing one of the same species.susceptible host A host that is readilyinfected by a particular parasite (see refrac-tory host).sylvatic cycle See feral cycle.symbiosis State of two organisms livingtogether for mutual advantage, the relation-ships being so intimate that the participat-ing species rely on one another for theirexistence. Mutualism is similar but the twoorganisms can live apart.symptom Any subjective evidence of dis-ease in a patient (see sign).tegument The outer layer of the body wallof helminths.tetrathyridium In the tapeworm genusMesocestoides, the second larval stage, inwhich the scolex with its four suckers isinvaginated into the anterior end of a plero-cercoid type of body.uterus The tubule in helminths containingthe fully formed eggs before they arevoided.vagina An outer chamber of the femalegenitalia in nematodes; also the tubuleleading from the genital atrium to theootype in cestodes (see vulva).
Glossary of Helminthological Terms 277
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 277

vas deferens The common male duct aris-ing from one or more vasa efferentia andleading into the seminal vesicle.vas efferens The male duct conveying sper-matozoa from the testis to the vas deferens.vector An intermediate host that carriesthe parasite to the definitive host.
biological vector A host essential to thedevelopment and transmission of a para-site, in which the parasite undergoesdevelopment (e.g. insects and filariae).mechanical vector A non-essential dis-seminator of parasites, in which the par-asite does not develop (e.g. fliessometimes carrying the eggs of Taenia orAscaris).
vermicide Therapeutic agent producing thedeath of a helminth (see anthelminthic).vermifuge Therapeutic agent producingevacuation of a helminth from the intestinewithout necessarily causing its death (e.g.
the action of piperazine against Ascarisand Enterobius).vitellaria (= vitelline glands) The glandsin Platyhelminthes which produce yolkmaterial and the shell of the egg.vulva The outermost, unpaired chamber ofthe female genitalia in nematodes.worm burden The number of worms pre-sent in the host (this is often estimatedfrom the egg count).xiphidiocercaria Cercaria with a stylet,median dorsal in position in the oralsucker; having associated penetrationglands with duct openings on either side ofthe stylet (as in Paragonimus).zoonosis Infection or disease naturallytransmitted between humans and othervertebrates.zooprophylaxis The prevention or amelio-ration of disease in humans by prior expo-sure to a heterologous infection of animals.
278 Appendix 2
11W. & H.D. - Appendix 2 14/11/01 4:30 PM Page 278

Appendix 3Location of Helminths in the Human Body
279
TRACHEAMammomonogamus* → irritation & cough
STOMACHGnathostoma* (larva) → abscessAnisakis* (larva) → abscess
SMALL INTESTINEFasciolopsis → enteritisHeterophyes → enteritisMetagonimus → enteritisDiphyllobothrium → anaemia*Taenia spp. → nauseaHymenolepsis spp. → diarrhoeaHookworm → anaemiaTrichostrongylus → diarrhoeaStrongyloides → dysentery (malabsorption)
Ascaris → obstructionTrichinella → dysenteryCapillaria philippinensis* → dysentery (malabsorption)Pseudoterranova* (larva) → abscess
COLON AND CAECUMSchistosoma spp. (eggs) → dysenteryGastrodiscoides* → diarrhoeaEnterobius → anal pruritusTrichuris → dysenteryTernidens → diarrhoea?Angiostrongylus → necrosis & costaricensis* granulomas (adults + eggs)
URINOGENITAL SYSTEMS. haematobium (eggs) → bladder fibrosis
Wuchereria funiculitis & hydroceleBrugia → in males, chyluria
*RARE
(a)
Fig. 132. Location in the human body of helminths and other metazoa with the possible pathology thatmay result. Organs through which a parasite passes without causing any pathology are ignored. Notincluded: Oesophagostomum bifurcum in the colon and caecum causing nodules. (Name changes:Angiostrongylus = Parastrongylus; Capillaria philippinensis = Aonchotheca philippinensis; Capillariahepatica = Calodium hepaticum.) (After Muller and Baker, 1990.)
12W. & H.D. - Appendix 3 14/11/01 4:30 PM Page 279

280 Appendix 3
SUBCUTANEOUS TISSUESSpirometra* (larva) → oedemaT. solium (larva) → nodulesWucheraria → elephantiasisBrugia → elephantiasisLoa → fugitive (Calabar) swellingOnchocerca → nodulesDirofilaria* (larva) → noduleDracunculus → cellulitisGnathostoma* (larva) → oedemaDermatobia (larva) → abscessCochliomyia (larva) → abscess
SKINBird schistosome (larva) → creeping eruptionOnchocerca (mf) → thickening then atrophy
Dracunculus → ulcerationMansonella streptocerca (mf) → oedema & papulesHookworm (larva) → ground itchAnimal hookworm (larva) → creeping eruptionStrongyloides (larva) → larva currensSarcoptes → scabiesTunga (larva) → boilDemodex → dermatitis*
MUSCLESTaenia solium (larva) → oedemaTrichinella (larva) → oedema
BONE MARROWEchinococcus granulosus → space-occupying lesion* (larva)
BLOODSchistosoma spp. → fever?Microfilariae of:WuchereriaBrugiaLoa Mansonella
→ fever?
LYMPHATICS
WuchereriaBrugia
lymphangitis→ lymphoedema ephantiasis
EYESTaenia solium (larva) → blurring of vision*T. multiceps (larva) → space-filling lesion
Loa → conjunctivitisOnchocerca volvulus (mf) → keratitis & retinopathy
& nerve atrophyToxocara canis (larva) → granuloma*Dirofilaria* (larva) → conjunctivitisOestrus (larva) → inflammation
*RARE
(b)
Fig. 132. Continued.
12W. & H.D. - Appendix 3 14/11/01 4:30 PM Page 280

Location of Helminths in the Human Body 281
BRAIN + CNSParagonimus (+eggs)Schistosoma spp. (+eggs)E. granulosus (larva)T. solium (larva)T. multiceps (larva)Gnathostoma* (larva) → myelencephalitisTrichinella (larva) → encephalitis*Angiostrongylus → meningitis cantonensis* (larva)Loa (mf) → encephalitis*Toxocara canis (larva) → granuloma*
LUNGSParagonimus → pneumoniaE. granulosus (larva) → space-occupying
lesionLarvae of:Ascaris, hookwormStrongyloides, Toxocara,Trichinella, Schistosomaspp., ParagonimusWuchereria (mf)Brugia (mf)Dirofilaria immitis* (larva) → coin lesion
HEARTLarvae of:T. soliumTrichinellaDirofilaria immitis*Loa (mf) → endomyocardial fibrosis*
LIVERClonorchisOpisthorchisFasciolaAscaris → cholangitisSchistosoma spp. (eggs) → portal hypertension
Larvae of:Fasciola ToxocaraCapillaria hepatica*AscarisE. granulosus (larva) → space-occupying
lesionE. multilocularis* (larva) → pseudomalignant
growthsArmillifer (larva) → inflammation*
KIDNEYSS. haematobium (eggs) → hydronephrosisE. granulosus (larva) → space-occupying lesion*Dioctophyma* → renal colic
SPLEENE. granulosus (larva) → space-occupying lesion*
PANCREASClonorchis → pancreatitisOpisthorchis → pancreatitisAscaris → pancreatitis
*RARE
→ abscess (& granulomas)
space-→ occupying
lesions
→ diffuse pneumonitis
→ tropical pulmonary eosinophilia
→ cardiomyopathy
→ biliary pathology
→ necrosis
(c)
Fig. 132. Continued.
12W. & H.D. - Appendix 3 14/11/01 4:30 PM Page 281

Abdi, Y.A. (1994) Handbook of Drugs for Tropical Parasitic Diseases, 2nd edn. Taylor and Francis,London, UK.
Ash, L.R. and Orihel, T.C. (1990) Atlas of Human Parasitology. American Society of ClinicalPathologists, Chicago, USA.
Ashford, R.W. and Crewe, W. (1998) The Parasites of Homo sapiens. Liverpool School of TropicalMedicine, Liverpool, UK.
Barnard, C.J. and Behnke, J.M. (eds) (1990) Parasitism and Host Behaviour. Taylor and Francis,London, UK.
Bennet, E.-M., Behm, C. and Bryant, C. (eds) (1989) Comparative Biochemistry of ParasiticHelminths. Chapman and Hall, London, UK.
Bogitsch, B.J. and Cheng, T.C. (1998) Human Parasitology, 2nd edn. Academic Press, San Diego, USA.Boothroyd, J.C. and Komuniecki, R. (1995) Molecular Approaches to Parasitology. Willey-Liss, New
York, USA.Brooker, S. and Michael, E. (2000) The potential of geographical information systems (GIS) and
remote sensing in the epidemiology and control of human helminth infections. Advances inParasitology 47, 246–288.
Bryant, C. and Behm, C. (1989) Biochemical Adaptation in Parasites. Chapman and Hall, London, UK.Campbell, W.C. and Rew, R.S. (1986) Chemotherapy of Parasitic Diseases. Plenum, New York, USA.Chowdhury, N. and Tada, I. (eds) (1994) Helminthology. Springer-Verlag, Berlin, Germany.Combes, C. (1995) Interactions durables: Écologie et évolution du parasitisme. Masson, Paris, France.Cook, G.C. (1990) Parasitic Disease in Clinical Practice. Springer-Verlag, Berlin, Germany.Cook, G.C. (ed.) (1996) Manson’s Tropical Diseases. Saunders, London, UK.Coombs, I. and Crompton, D.W.T. (1991) A Guide to Human Helminths. Taylor and Francis, London, UK.Cox, F.E.G., Kreier, J.P. and Wakelin, D. (eds) (1998) Topley and Wilson’s Microbiology and Microbial
Infections. Vol. 5 Parasitology, 9th edn. Arnold, London, UK.Dailey, M.D. (1996) Meyer, Olsen and Schmidt’s Essentials of Parasitology, 6th edn. Brown,
Dubuque, Iowa, USA.Despommier, D.D., Gwadz, R.W. and Hotez, P.J. (1994) Parasitic Diseases, 3rd edn. Springer-Verlag,
New York, USA.Esch, G.W. and Fernández, J.C. (1993) A Functional Biology of Parasitism: Ecological and
Evolutionary Implications. Chapman and Hall, London, UK.Euzeby, J. (1981) Diagnostic expérimental des helminthoses animales (animaux domestiques – ani-
maux de laboratoire – primates), 2 vols. Ministère de l’Agriculture, Paris, France.Farthing, M.J.G. and Keusch, G.T. (eds) (1995) Enteric Infections 2: Intestinal Helminths. Chapman
and Hall, London, UK.Freedman, D.O. (1997) Immunopathogenic Aspects of Disease Induced by Helminth Parasites.
Chemical Immunology, Vol. 66, S. Karger, Basle, 236 pp.Garcia, L.S. (1999) Practical Guide to Diagnostic Parasitology. American Society for Microbiology,
Washington, USA.
General References and Further Reading
282
13W. & H.D. - Further Reading 14/11/01 4:31 PM Page 282

Further Reading 283
Garcia, L.S. and Bruckner, D.A. (1997) Diagnostic Medical Parasitology. American Society forMicrobiology, Washington, USA.
Gentilini, M. (1993) Médecine Tropicale. Flammarion, Paris, France.Gillespie, S.H. and Hawkey, R.M. (1995) Medical Parasitology: a Practical Approach. Oxford
University Press, New York, USA.Goldsmith, R. and Heyeman, D. (1989) Tropical Medicine and Parasitology. Appleton and Lange,
Hemel Hempstead, UK.Gutterierrez, Y. (2000) Diagnostic Pathology of Parasitic Infections with Clinical Correlations, 2nd
edn. Oxford University Press, New York, USA.Hall, A. (1982) Intestinal helminths of man: the interpretation of egg counts. Parasitology 85,
605–613.Halton, D.W., Behnke, J.M. and Marshall, I. (eds) (2001) Practical Exercises in Parasitology.
Cambridge University Press, Cambridge, UK.Hugh-Jones, M.E., Hubbert, W.T. and Hagstad, H.V. (1995) Zoonoses: Recognition, Control and
Prevention. Iowa State University Press, Ames, USA.Hyde, J.E. (1990) Molecular Parasitology. Open University, Buckingham, UK.Hyde, J.E. (1993) Protocols in Molecular Parasitology. Humana Press, Totowa, New Jersey, USA.James, D.M. and Gilles, H.M. (1985) Human Antiparasitic Drugs: Pharmacology and Usage. John
Wiley & Sons, Chichester, UK.Kassai, T., Cordero del Campo, M., Euzeby, J., Gaafar, S., Hiepe, Th. and Himonas, C.A. (1988)
Standardized nomenclature of animal parasitic diseases (SNOAPAD). Veterinary Parasitology 29,299–326.
Leventhal, R. and Cheadle, R. (1996) Medical Parasitology: a Self-instructional Text. Davis,Philadelphia, USA.
Littlewood, T. and Bray, R. (eds) (2001) Interrelationships of the Platyhelminthes. Taylor and Francis,London, UK.
Macpherson, C.N.L. and Craig, P.S. (eds) (1991) Parasitic Helminths and Zoonoses in Africa. UnwinHyman, London, UK.
Maizels, R.M., Blaxter, M.L., Robertson, B.D. and Selkirk, M.E. (1991) Parasite Antigens, Parasite Genes.A Laboratory Manual for Molecular Parasitology. Cambridge University Press, Cambridge, UK.
Markell, E.K., John, D.T. and Krotoski, W.A. (1999) Markell and Voge’s Medical Parasitology, 8th edn.Saunders, Philadelphia, USA.
Marquardt, W.C., Demaree, R.S. and Grieve, R.B. (2000) Parasitology and Vector Biology. 2nd edn.Academic Press, Orlando, Florida, USA.
Marr, J.J. and Muller, M. (1995) Biochemistry and Molecular Biology of Parasites. Academic Press,London, UK.
Meyers, W. (2000) Pathology of Infectious Diseases. Vol. 1. Helminthiases. Armed Forces Institute ofPathology, Washington, USA.
Miyazaki, T. (1991) An Illustrated Book of Helminthic Zoonoses. International Medical Foundationof Japan, Tokyo, Japan.
Muller, R. (1997) Parasitology. In: Wyatt, H.V. (ed.) Information Sources in the Life Sciences, 4th edn.Bowker-Sauer, East Grinstead, UK, pp. 195–202.
Muller, R. and Baker, J.R. (1990) Medical Parasitology. Lippincott, Philadelphia, USA.Neva, F.A. and Brown, H.W. (1994) Basic Clinical Parasitology, 6th edn. Appleton and Lange,
Norwalk, Connecticut, USA.Nokes, C. and Bundy, D.A.P. (1994) Does helminth infection affect mental processing and educa-
tional achievement? Parasitology Today 10, 14–18.Orihel, T.C. and Ash, L.R. (1995) Parasites in Human Tissues. American Society of Clinical
Pathologists, Chicago, USA.Palmer, P.E.S. and Reeder, M.M. (2000) The Imaging of Tropical Diseases, 2 vols. Springer,
Heidelberg, Germany. Palmer, S.R., Lord Soulsby and Simpson, D.I.H. (eds) (2000) Zoonoses. Oxford University Press, New
York, USA.Peters, W. and Gilles, H.M. (1999) A Colour Atlas of Tropical Medicine and Parasitology, 5th edn.
Mosby, London, UK.Price, D.L. (1993) Procedure Manual of the Diagnosis of Intestinal Parasites. CRC Press, Boca Raton, USA. Rogan, M.T. (ed.) (1997) Analytical Parasitology. Springer-Verlag, Berlin, Germany.
13W. & H.D. - Further Reading 14/11/01 4:31 PM Page 283

284 Further Reading
Salfelder, K., Liscans, T.R. and de Sauerleig, E. (1992) Atlas of Parasitic Pathology. Kluwer,Dordrecht, The Netherlands.
Schmidt, G.D. and Roberts, L.S. (1989) Foundations of Parasitology. Mosby, Missouri, USA.Scott, M.E. and Smith, G. (eds) (1994) Parasitic and Infectious Diseases: Epidemiology and Ecology.
Academic Press, New York, USA.Smyth, J.D. (ed.) (1990) In vitro Cultivation of Parasitic Helminths. CRC Press, Boca Raton, Florida, USA.Smyth, J.D. (ed.) (1994) Introduction to Animal Parasitology. Cambridge University Press, Cambridge, UK.Soulsby, E.J.L. (ed.) (1987) Immune Responses in Parasitic Infections. CRC Press, Boca Raton, USA.Spicer, W.J. (2000) Clinical Bacteriology, Mycology and Parasitology. Harcourt, London, UK.Stephenson, L.S. (1987) The Impact of Helminth Infections on Human Nutrition: Schistosomes and
Soil-transmitted Helminths. Taylor and Francis, London, UK.Strickland, T. (ed.) (2000) Hunter’s Tropical Medicine and Emerging Infectious Diseases, 8th edn.
Saunders, Philadelphia, USA.Sun, T. (1993) Progress in Clinical Parasitology, Vol. 3. Field and Wood, New York, USA.Taylor, A.E.R. and Baker, J.R. (eds) (1987) In vitro Methods for Parasite Cultivation. Academic Press,
London, UK. Thienpont, D., Rochette, F. and Vanparijs, O.F.G. (1979) Diagnosing Helminthiasis through
Coprological Examination. Janssen Research Foundation, Beerse, Belgium.Ubelaker, J.E. (1993) Stedman’s ASP Parasite Names. Williams and Wilkins, Baltimore, USA.Urquhart, G.M., Armour, J., Duncan, J.L., Dunn, A.M. and Jennings, F.W. (1987) Veterinary
Parasitology. Churchill Livingstone, New York, USA.Van den Bossche, H. (1995) Principles of anthelmintic chemotherapy. In: Farthing, M.J.G., Keusch,
G.T. and Wakelin, D. (eds) Enteric Infections 2: Intestinal Helminths. Chapman and Hall, London,UK, pp. 267–286.
van Lichtenberg, F. (1991) Pathology of Infectious Diseases. Raven Press, New York, USA.Wakelin, D.M. (1988) Helminth infections. In: Wakelin, D.M. and Blackwell, J.M. (eds) Genetics of
Resistance to Bacterial and Parasitic Infection. Taylor and Francis, London, UK, pp. 83–100.Walls, K. and Schantz, P. (eds) (1986) Immunodiagnosis of Parasitic Diseases. I. Helminthic Diseases.
Academic Press, Orlando, USA.Whitfield, P.J., Molyneux, D.H., Anderson, R.M., Bryant, C., Hart, D.T., Chappell, L.H., Gutteridge,
W.E. and Cox, F.E.G. (1993) Modern Parasitology: a Textbook of Parasitology, 2nd edn. BlackwellScientific Publications, Oxford, UK.
WHO (1987) Prevention and Control of Intestinal Parasitic Diseases. World Health Organization,Geneva, Switzerland.
WHO (1990) WHO Model Prescribing Information: Drugs Used in Parasitic Diseases. World HealthOrganization, Geneva, Switzerland.
Working Group on Parasite Control – The Hashimoto Intitiative (1998) The Global Parasite Controlfor the 21st Century. Government of Japan, Kobe, Japan.
Wyler, D.J. (1990) Modern Parasite Biology: Cellular, Immunological and Molecular Aspects. W.H.Freeman, New York, USA.
Trematodes
Kumar, V. (1999) Trematode Infections and Diseases of Man and Animals. Kluwer, Dordrecht, TheNetherlands.
Smyth, J.D. and Halton, D.W. (1983) The Physiology of Trematodes, 2nd edn. Cambridge UniversityPress, Cambridge, UK.
Cestodes
Adams, A.M. and Rausch, R.L. (1999) Diphyllobothriasis. In: Connor, D.H., Chandler, F.W., Schwartz,D.A., Manz, H.J. and Lack, E.E. (eds) Pathology of Infectious Diseases. Vol. 2. Appleton andLange, Norwalk, Connecticut, USA, pp. 1377–1390.
Arai, H.P. (ed.) (1980) Biology of the Tapeworm Hymenolepis diminuta. Academic Press, New York, USA.
13W. & H.D. - Further Reading 14/11/01 4:31 PM Page 284

Further Reading 285
Smyth, J.D. and McManus, D.P. (1989) The Physiology and Biochemistry of Cestodes. CambridgeUniversity Press, Cambridge, UK.
Wardle, R.A., McLeod, J.A. and Radinovsky, S. (1974) Advances in the Zoology of Tapeworms,1950–1970. University of Minnesota Press, Minneapolis, USA.
Nematodes
Anderson, R.C. (2000) Nematode Parasites of Vertebrates – their Development and Transmission, 2ndedn. CAB International, Wallingford, UK.
Bird, A.F. and Bird, J. (1991) The Structure of Nematodes. Academic Press, New York, USA.
Immunology
Allen, J.E. and Maizels, R.M. (1997) Th1–Th2 paradigm: reliable paradigm or dangerous dogma?Immunology Today 18, 387–392.
Bancroft, A.J., Grencis, R.K. and MacDonald, T.T. (1998) Th1 and Th2 cells and immunity to intesti-nal helminths. Chemical Immunology 71, 198–208.
Capron, A. (1998) Schistosomiasis: forty years’ war on the worm. Parasitology Today 14, 379–384.Finkelman, F.D., Shea-Donahue, T., Goldhill, J., Sullivan, C.A., Morris, S.C., Madden, K.B., Gause,
W.C. and Urban, J.F., Jr (1997) Cytokine regulation of host defense against parasitic gastrointestinalnematodes: lessons from studies with rodent models. Annual Review of Immunology 15, 505–533.
Janeway, C. and Travers, P. ( 1999) Immunobiology, 4th edn. Churchill-Livingstone, London.Kennedy, M.W. and Harnett, W. (eds) (2001) Parasitic Nematodes – Molecular Biology, Biochemistry
and Immunology, CAB International, Wallingford, UK.Kierszenbaum, F. (ed.) (1994) Parasitic Infections and the Immune System. Academic Press, San
Diego, USA.Liew, F.Y. and Vickerman, K. (eds) (1997) Immune effector mechanisms in parasitic infections.
Philosophical Transactions of the Royal Society B 352, 1293–1394.Moqbel, R. (ed.) (1992) Allergy and Immunity to Helminths: Common Mechanisms or Divergent
Pathways. Taylor and Francis, London, UK.Parham, P. (2000) The Immune System. Elsevier Science, London, UK.Prichard, D.I. (1995) The survival strategies of hookworms. Parasitology Today 11, 255–259.Roitt, I., Brostoff, J. and Male, D. (1998) Immunology, 5th edn. Mosby, London, UK.Röllinghof, M. and Rommel, M. (1994) Immunologische und molekulare Parasitologie. Gustav
Fischer, Berlin, Germany.Wakelin, D. (1994) Host populations: genetics and immunity. In: Scott, M.E. and Smith, G. (eds)
Parasitic and Infectious Diseases. Academic Press, London, UK, pp. 83–100.Wakelin, D. (1996) Immunity to Parasites: How Parasitic Infections are Controlled. Cambridge
University Press, Cambridge, UK.Wakelin, D. (1997) Parasites and the immune system: conflict or compromise? Bioscience 47, 32–40.Wang, C.C. (ed.) (1991) Molecular and Immunological Aspects of Parasitism. American Association
for the Advancement of Science, Washington, USA.Warren, K.S. (ed.) (1993) Immunology and Molecular Biology of Parasitic Infections, 3rd edn.
Blackwell Scientific Publications, Boston, USA.
History
Cox, F.E.G. (ed.) (1996) The Wellcome Trust Illustrated History of Tropical Diseases. Wellcome Trust,London, UK.
Dunlop, R.H. and Williams, D.I. (1996) Veterinary Medicine: an Illustrated History. Mosby Year Book,St Louis, USA.
Farley, J. (1991) Bilharzia. A History of Imperial Tropical Medicine. Cambridge University Press,Cambridge, UK.
13W. & H.D. - Further Reading 14/11/01 4:31 PM Page 285

286 Further Reading
Foster, W.D. (1965) A History of Parasitology. Livingstone, Edinburgh, UK.Garnham, P.C.C. (1971) Progress in Parasitology. Athlone Press, London, UK.Grove, D.I. (1990) A History of Human Helminthology. CAB International, Wallingford, UK (available
on-line at www.red-e2.com). Hoeppli, R. (1959) Parasites and Parasitic Infections in Early Medicine and Science. University of
Malaya Press, Singapore.Kean, B.H., Mott, K.E. and Russell, A.J. (eds) (1978) Tropical Medicine and Parasitology: Classic
Investigations, Vols 1 and 2. Cornell University Press, Ithaca, USA.
13W. & H.D. - Further Reading 14/11/01 4:31 PM Page 286

Abbreviata (= Physaloptera) caucasia 187Acanthocephalans 106–107Acanthocephalus rauschi 107Accidental filarial infections 226–228Achillurbainia
nouveli 37–38recondita 37–38
Acuaria sp. 187Adhesive (= Scotch/Sellotape) tape technique
75, 162–163ADCC (antibody dependent cell–mediated
cytotoxicity) 23, 30, 198, 217, 244,250
Aedes as vectors of bancroftian filariasis 200as vectors of dirofilariasis 226as vectors of malayan filariasis 205
African Programme for Onchocerciasis(APOC) 220
Agamomermishominis 184restiformis 184
Agar plate culture method for larvae 123–125Aggregation of helminth infections 122, 132,
168AIDS 23, 83, 119, 200Alaria
alata 58americana 58, 156marcianae 58, 156
Albendazolecestodes 82, 92, 96, 97intestinal nematodes 123–124, 125,
132–133, 140, 142, 147, 152, 153, 159,167
tissue nematodes 174, 175, 181, 186,199, 201, 210, 222, 232
trematodes 51–52Allergic responses to 241, 248
cestodes 94
intestinal nematodes 120, 156tissue nematodes 179, 181, 196, 209,
218, 222, 226, 230Amoebiasis 53, 141, 146, 166Amphibians as hosts 56, 58, 70, 71, 107, 145,
184, 186Anaemia and helminths 21, 47, 57, 68,
130–132, 138, 166Anatrichosoma cutaneum 156, 175Ancylostoma
braziliense 160caninum 137, 160ceylanicum 137differential features from Necator 130duodenale 134–137
clinical manifestations 135epidemiology 136geographical distribution 134immunology 120–122life cycle 135morphology 134pathogenesis 136prevention and control 136
japonica 137malayanum 137secreted polypeptide (ASP) 132tubaeforme 160
Angiostrongylus see ParastrongylusAnisakis simplex 154–156
clinical manifestations 155diagnosis 155epidemiology 155–156geographical distribution 154life cycle 154location in host 154morphology 154pathogenesis 155prevention 156treatment 155
Annelids as hosts 157, 147, 184
Index
287
14Worms & H.D. - Index 20/11/01 1:46 PM Page 287

Anophelesas vectors of bancroftian filariasis
199–200as vectors of malayan filariasis 205 as vectors of timoran filariasis 205
Anorexia 58, 68, 129, 137, 150, 166, 173, 175
Antibiotics and lymphatic filariasis 199Antibodies, monoclonal 24, 36, 44, 75, 145,
155, 198–199Antigenic disguise 23Antigens
coproantigens 48, 75, 93excretory/secretory (ES) 30, 75,109, 132,
155, 159, 180, 204, 251Ants as hosts 46, 103, 139Aonchotheca (= Capillaria) philippinensis
173–174clinical manifestations 173–174diagnosis 174epidemiology 174geographical distribution 174life cycle 173location in host 173morphology 173pathogenesis 174treatment 174
Apophallus donicus 54Appendicitis and helminths 74, 141, 150, 155,
162, 167, 184Armillifer
agkistrodontus 240armillatus 240
Artemisins (Artemether and Artesunate) inschistosomiasis 25
Artyfechinostomum araoni 57malayanum 57mehrai 57sufratyfex 57
Ascaris lumbricoides 147–153
clinical manifestations 141–150diagnosis 151epidemiology geographical distribution 148life cycle 148location in host 148morphology 148pathogenesis 151prevention and control 152–153treatment 151–152zoonotic aspects 153
suum 153Aspiration of cysts of Echinococcus 92Australobilharzia terrigalensis 32, 156Autoinfection 120, 153, 162, 173
B cells 30, 244Bacteria 3, 41, 119, 158, 160, 166, 196, 202, 230Baermann technique 122Bancroftian filariasis see Wuchereria bancroftiBats as hosts 159, 188Baylisascaris procyonis 153–154, 156Bears as hosts 71, 137, 154, 182–183, 228Beetles as hosts 103, 106, 107, 188Bertiella
mucronata 103studeri 102
Bile ducts and helminths 41, 44, 45, 47, 150,187
Bilharziella polonica 32Biomphalaria
alexandria group 27, 29choamphala group 27, 29glabrata 27, 29pfeifferi group 27, 29spp. 5, 27sudanica group 27, 29
Biopsy 24, 47, 81, 137, 146, 155, 175, 180, 209,227
Birdsas hosts of
cestodes 69, 70, 71, 103nematodes 153, 173, 182, 186, 188trematodes 31, 47, 54, 58
in transmission of Taenia 76Bithionol 48Blindness see Eye lesionsBolbosoma sp. 107Brain involvement 22, 35–36, 53, 71, 81, 85, 86,
90, 96, 144, 154, 158, 159Brugia
beaveri 206guyanensis 206 malayi 202–206
clinical manifestations 204diagnosis 204epidemiology 204–205geographical distribution 202life cycle 204location in host 202morphology 202pathogenesis 204prevention and control 205treatment 204zoonotic aspects 205
pahangi 205timori 205
Buffaloes as hosts 49, 92Buginetta alouattae 103Bulimus fuchsianus 38Bulinus
africanus group 27camerunensis 27
288 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 288

forskali group 27reticulatus group 27spp. 5, 11, 27truncatus group 27
Bunostomum phlebotomum 160
Calabar swellings 207Calodium (= Capillaria) hepaticum 156, 175Camels as hosts 49, 140Cancer and helminths 18–19, 41, 44, 45, 83,
155, 226Capillaria
aerophila see Eucoleus aerophilushepatica see Calodium hepaticumphilippinensis see Aonchotheca
philippinensisCardiac damage 22, 53, 84, 97, 120, 143, 147,
179, 226, 241Carneophallus brevicaeca 59Carnivores (wild) as hosts 37, 45, 59, 71, 84, 85,
92, 98, 99, 108, 154Cathaemasia cabreri 59Cats as hosts of
cestodes 69, 71, 94intestinal nematodes 153, 157, 160tissue nematodes 187, 205, 234trematodes 45, 54, 56
Cattle as hosts 31, 46, 49, 56, 75–76, 86, 92, 139,187
Central nervous system (CNS) involvement 22,83, 84, 90, 120, 125, 159,179, 186, 209
Centrocistus armatus 54caninus 54cuspidatus 54formosanus 54kurokawai 54longus 54
Cerithidea cingulata 52Cervical smear 162Cestodes 63–105
classification 64–66recognition in sections 267
Ceviche 186Cheilobus quadrilabiatus 125Cheilospirura 187Chemotherapy, mass or universal 28, 133–134,
201, 220Chordodes capensis 108Chrysomyia bezziana 242Chrysops spp. as intermediate hosts of loiasis
centurioni 206, 210dimidiatus 206, 210distinctipennis 206, 210langi 206, 210silicea 206, 210
Cirrhosis 22, 41Classification of
cestodes 64–65nematodes 109–113trematodes 6–9
Clinostomum complanatum 59Clonorchis sinensis 38–43
clinical manifestations 39control and prevention 42diagnosis 42epidemiology 42–43geographical distribution 38immunology 41–42life cycle 38location in host 38morphology 38pathogenesis 41treatment 42
Coatimundi as host 146, 206Cochliomyia hominovorax 242Codiella leachi 5, 44Coenurus larva 83Cognitive ability and nematodes 130, 151, 166Computerized axial tomography (CT) 36, 48, 81, 91Concentration methods for faeces examination
260–262Concomitant immunity 23, 198Contracaecum osculatum 155Copepods as intermediate hosts of
cestodes 66–67, 70, 101nematodes 185, 229
Corbicula sp. 57Cordylobia anthropophaga 242Corticosteroids 119, 159, 181, 210Corynosoma strumosum 107Cotylurus japonicus 59Counting techniques for microfilariae 264–265Creeping eruption see Cutaneous larva migransCrustaceans as hosts 145, 154, 156, 184
crabs and crayfish 34, 36, 37, 219see also Copepods
Cryptocotyle lingua 54Culex as vectors of bancroftian filariasis
annulirostris 200bitaeniorhynchus 200pipiens molestus 200pipiens pallens 200quinquefasciatus 200
Culicoides as vectors of Mansonella ozzardi
furens 225paraensis 225phlebotomus 225
as vectors of Mansonella perstans and M.streptocerca
grahami 223, 225milnei 223, 225
Index 289
14Worms & H.D. - Index 20/11/01 1:46 PM Page 289

Cultivation of first-stage nematode larvae122–123, 262–263
Cutaneous larva migrans 159–160, 187Cyclops see CopepodsCyclodontostomum purvisi 137Cyclophyllidea 64, 71–103Cyclosporin 25, 124Cysticercosis 80–83Cysticercus
bovis 74cellulosae 78, 80–83longicollis 83
Cystitis 18Cytokines 23, 197, 209, 217, 244, 245, 249
Dams and schistosomiasis 29Dermatitis 32, 156, 160, 214–215Dermatobia hominis 241–242 Diagnostic techniques for
eggs in faeces 258–260microfilariae in blood 263–264
Diaptomus see CopepodsDicrocoelium
dendriticum 45–46hospes 46
Diethylcarbamazine 147, 159, 199, 201, 210,218, 222, 224, 226
Dioctophyma renale 184Diorchitrema
amplicaecale 55formosanum 55pseudocirratum 55
Diphyllobothrium cameroni 69cordatum 69dalliae 69dendriticum 69elegans 69giljacicum 69hians 69klebanovskii 69lanceolatum 69latum 65–68
clinical manifestations 68diagnosis 68epidemiology 69epidemiology 69geographical distribution 65life cycle 66location in host 65morphology 65pathogenesis 68prevention and control 68treatment 68zoonotic aspects 69
minus 69
nihonkaiense 69orcini 69pacificum 69scoticum 69skjrabini 69tangussicum 69ursi 69yonagoense 69
Diplogonoporus brauni 69fukuokaensis 69grandis 69
Diploscapter coronata 125Diplostomum spathaceum 58Dipylidium caninum 102Dirofilaria
immitis 226louisianensis 226magalhaesi 226repens 226–227spectans 226striata 227–228tenuis 227–228ursi 227–228
Dogs as hosts ofcestodes 69, 71, 83, 84, 85, 94, 97,
102intestinal nematodes 137, 145, 153, 157,
160tissue nematodes 175, 183, 184, 187, 188,
234trematodes 31, 43, 45, 54, 57, 58
Dracunculus insignis 234medinensis 228–234
clinical manifestations 230–231diagnosis 231–232epidemiology 232geographical distribution 228life cycle 229location in host 228morphology 228–229pathogenesis 231prevention and control 232–233treatment 232zoonotic aspects 234
Drepanidotaenia lanceolata 101
Ecdysteroids 119Echinochasmus
fujianensis 57japonicus 57jiufoensis 57liliputanus 57perfoliatus 57
Echinococcosis 85–87
290 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 290

Echinococcus equinus 92granulosus 85–93
clinical manifestations 87–90diagnosis 90–91epidemiology 87, 93–94geographical distribution 85immunology 93life cycle 85–87location in host 85morphology 85pathogenesis 90prevention and control 92–93treatment 91vaccines 93
multilocularis 94–97clinical manifestations 94–96diagnosis 96epidemiology 96–97geographical distribution 94life cycle 94morphology 94pathogenesis 96prevention and control 97treatment 96
oligarthrus 97 ortleppi 92spp. differentiation 88vogeli 97
Echinoparyphium paraulum 57recurvatum 57
Echinostomaangustitestis 57cinetorchis 57echinatum (= lindoense) 57hortense 57ilocanum 56japonicum 57macrorchis 57malayanum 57melis 57revolutum 57
Eggcounting techniques 260–262identification 256–257production
cestodes 66intestinal nematodes 129, 135, 148, 161, 165trematodes 13, 16, 38, 50
Elephantiasis 194–195, 204Endoscopy for anisakids 155Enterobius
gregorii 161vermicularis 160–164
clinical manifestations 162diagnosis 162–163
epidemiology 163–164geographical distribution 160life cycle 161–162location in host 160morphology 160–161pathogenesis 162prevention and control 164treatment 163zoonotic aspects 164
Entero-test 123Eosinophilia and
cestodes 74intestinal nematodes 144, 146, 149, 158,
166tissue nematodes 175, 178, 186, 193, 195,
214, 226trematodes 19, 40, 47, 53, 59
Eosinophils, staining for 255Epidemiology of helminths 252–254Episthmium caninum 57Eucoleus (= Capillaria)
aerophilus 175boehmi 175
Eurytrema pancreaticum 46Eustrongylides
ignotus 184tubifex 184
Eye lesions andacanthocephalans 241cestodes 71intestinal nematodes 144, 154, 158leeches 242tissue nematodes 178, 208, 211, 216–217,
226
Facilitation in mosquitos 200Faecal examination
for diagnosis of larvae 261–263for eggs 255–262quantitative techniques 260–262
Fasciola gigantica 49hepatica 46–49
clinical manifestations 47diagnosis 48epidemiology 48geographical distribution 46immunology 41–42life cycle 46–47location in host 46morphology 46pathogenesis 47prevention and control 45treatment 48zoonotic aspects 48
indica 49
Index 291
14Worms & H.D. - Index 20/11/01 1:46 PM Page 291

Fasciolopsis buski 49–51clinical manifestations 50diagnosis 50epidemiology 51geographical distribution 49life cycle 50location in host 49morphology 50pathogenesis 50prevention and control 51treatment 50–51zoonotic aspects 51
Fibrosis 18, 21–22, 41, 48, 120. 141, 178, 196,208
Filarial infections, accidental 226–228infective larvae
mounting 265staining before dissection 265
vectors, dissection of 265Fish
freshwater as intermediate hosts 42–43, 53,54, 56, 58–59, 69, 174, 185, 186, 187
marine as final hosts 155marine as intermediate hosts 71, 154–155,
156, 187Fish dishes, in transmission 45, 89, 156, 186,
187Fischoederius elongatus 56Fleas as hosts 102Flies as hosts 187Flubendazole 82Formol–ether egg concentration method
259–260Foxes as hosts 94, 97, 145, 182
Gasterophilus sp. 242Gastrodiscoides hominis 55–56
geographical distribution 55life cycle 55location in host 55morphology 55zoonotic aspects 55–56
Geohelminths 115–168Geophagia 153, 168Gigantobilharzia
huttoni 32sturniae 32
Gnathostoma doloresi 186hispidum 156, 186nipponicum 187spinigerum 184–186
clinical manifestations 186diagnosis 186epidemiology 86
geographical distribution 185life cycle 185–186location in host 184pathology 187
Goats as hosts 31, 76, 92Gongylonema pulchrum 187Gordius
aquaticus 108chilensis 108gesneri 108inesae 108ogatae 108perronciti 108reddyi 108robustus 108setiger 108skorikowi 108
Granulomas 20, 22–23, 36, 48, 153, 155, 159,227
‘Ground itch’ see Cutaneous larva migransGymnophalloides seoi 59Gyraulis chinensis 50
Haematuria in schistosomiasis 18Haemonchus contortus 139Hairworms see NematomorphaHalicephalus (= Micronema) deletrix 125Halzoun 241‘Hanging groin’ in onchocerciasis 215Haplorchis
microchis 55pleurolophocerca 55pumilio 55tachui 55vanissimus 55yokogawai 55
Harada–Mori larval cultivation technique262–263
Heart involvement 22, 53, 54, 125, 179, 226Helminths
preservation 205–207recognition in sections 267–269
Heliosoma sp. 58Hepatitis 18, 40, 45, 46, 159, 175Hepatomegaly 22, 44, 96, 146, 158, 159, 175Herbivores in general as hosts 46, 49, 56, 70, 76,
86, 92, 139, 140, 187Heterobilharzia americana 32Heterophyes
dispar 55equalis 55heterophyes 51–53
clinical manifestations 53diagnosis 54epidemiology 54geographical distribution 51
292 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 292

life cycle 52location in host 51morphology 52pathogenesis 53prevention and control 53treatment 53zoonotic aspects 53
heterophyes nocens 55 katsuradai 55
Heterophyopsis continua 57Himasthla muehlensi 57Hookworms 126–127, 160Horses as hosts 92, 125, 154HTLV-1 and Strongyloides 119Human leucocyte antigen (HLA) 17, 217Hydatid 64, 85, 86, 87, 89, 90, 91, 97Hydrocele 194, 195, 199, 201Hydronephrosis 18Hymenolepis
diminuta 101geographical distribution 101life cycle 101location in host 101morphology 101
nana 98–101clinical manifestations 99diagnosis 99epidemiology 99–100geographical distribution 98life cycle 98location in host 98morphology 98pathogenesis 99prevention and control 100treatment 100zoonotic aspects 100–101
Hyperinfection 119, 124Hypodereum conoideum 57 Hypoderma bovis 242Hysterothylacium aduncum 155
Immunodiagnostic techniquescomplement fixation (CF) 42, 132coelectrosyneresis (Co–ES) 209DNA probes 218, 227enzyme linked immunosorbent assay
(ELISA) andcestodes 71, 75, 79, 93intestinal nematodes 123, 132, 145, 146,
154, 155, 159tissue nematodes 174, 180, 186, 199, 209,
218, 232trematodes 24, 36, 42, 44, 48
immunoblotting techniques 24, 48, 75, 93,145, 155, 159, 180, 186, 209
immunochromatographic test (ICT) 198
immunodiffusion 146, 174immunoelectrophoresis 146indirect fluorescent antibody (IFA) test 232indirect haemagglutination (IHA) 93, 132,
174intradermal (ID) test 36, 42, 48latex agglutination 146radioimmunoassay (RIA) 132sodium dodecyl sulphate – polyacrylamide
gel electrophoresis (SDS–PAGE) 49, 75,159, 232
Immunoglobulins 244IgA 31, 41, 121, 149, 244IgD 243IgE 23, 41, 121, 123, 198, 217, 244, 249IgG 24, 41, 123, 159, 180, 244, 248
IgG1 121, 155, 198, 209, 217IgG2 31, 198IgG4 23, 142, 198, 209, 217, 232, 251
IgM 42, 59, 198, 217, 244, 248Immunology of
cestodes 93helminths 243–251intestinal nematodes 120–122liver flukes 41–42lymphatic filariae 196–198 Onchocerca 217schistosomes 23
Immunomodulation 23, 119, 180, 198Inermicapsifer madagascariensis 102–103Inflammation due to
cestodes 74, 84intestinal nematodes 120, 144, 146, 147,
153, 160, 162, 167tissue nematodes 179, 193, 244 trematodes 18, 41, 47, 53
Insects, in general, as hosts 46, 58, 102, 103,157, 188
Interleukins 96, 197, 214, 248Intestinal obstruction and
cestodes 68, 74intestinal nematodes 119, 150trematodes 20, 50
Isoparorchis hypselobagri 59Ivermectin 124, 153, 154, 163, 167, 175, 199,
201, 210, 218, 222, 224, 232
Katayama syndrome 19–20Kato–Katz faecal technique 258–259Key to nematodes in sections 269Kidney worm see DioctophymaKidneys and
cestodes 78, 90intestinal nematodes 119, 157tissue nematodes 184, 178, 193, 210, 222trematodes 20, 50
Index 293
14Worms & H.D. - Index 20/11/01 1:46 PM Page 293

Lagochilascaris minor 153, 156Larva currens 119, 122Larva migrans 156–160
cutaneous 159–160ocular 158visceral (VLM) 159
Larvaecultivation of 122–124, 261–263filarial, in insects, staining and mounting
265Leeches 241Levamisole 133, 152, 153, 163Ligula intestinalis 69Limitation in mosquito infections 200Limnatis nilotica 241Linguatula serrata 240–241Liver flukes, immunology of 41–42Liver involvement and
cestodes 87, 91, 96, 97intestinal nematodes 150, 155, 157tissue nematodes 175, 186trematodes 21–22, 40–41, 43–44, 45, 46, 47
Loa loa 206–211clinical manifestations 207–208diagnosis 209epidemiology 210geographical distribution 206life cycle 206–207location in host 206morphology 206pathogenesis 208–209prevention and control 210treatment 210zoonotic aspects 210–211
Loaina sp. 211Loeffler’s syndrome 149–150, 158Lung involvement and
cestodes 71, 90, 97intestinal nematodes 119–120, 129, 149,
153, 157–158, 179tissue nematodes 186, 195–196, 208trematodes 19, 35, 37, 58
Lymphatic filariasis 190–206Lymphoscintigraphy 193Lymphatic involvement 43, 193–196, 198, 204,
206, 214, 222, 226
Macdonald pipette for egg counts 262Macranthorhynchus
hirudinaceus 107ingens 107
Magnetic Resonance Imaging (MRI) 81, 91Malabsorption syndrome 120, 151, 174Malnutrition 119, 132, 161, 166Mammals, marine, as hosts 69–70, 107,
155–156, 182
Mammomonogamuslaryngeus 142–143
clinical manifestations 142diagnosis 143geographical distribution 142life cycle 142morphology 142pathogenesis 142treatment 143zoonotic aspects 143
nasicola 143Mansonella
digitatum 223perstans 221–223ozzardi 225–226rodhaini 225semiclarum 222streptocerca 223–225
Mansoniaas vectors of bancroftian filariasis
200as vectors of malayan filariasis 205as vectors of subperiodic malayan filariasis
205Marisa cornuarietus in schistosomiasis control
29Marrara syndrome 241Marshallagia marshalli 139Marsupials as hosts 38, 92, 153Mathematical models 253Mathevotaenia symmetrica 102–103Mazzotti (provocation) test 217McMaster egg counting chamber 262Mebendazole 92, 123, 133, 138, 153, 163, 174,
181, 186, 232Mecistocirrus digitatus 139Melanoides tuberculata in schistosomiasis
control 29Meningitis 35, 119, 144, 145, 178Meningonema peruzii 223Mermis nigrescens, M. sp. 184Mesocestoides
lineatus 103variabilis 103
Metagonimus minutis 55takahashi 55yokogawai 53–55
clinical manifestations 54diagnosis 54epidemiology 54geographical distribution 54life cycle 54location in host 54morphology 54pathogenesis 54
Metastrongylus elongatus 147
294 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 294

Metorchisalbidus 45conjunctus 45
Metrifonate 25Metronidazole 232Meyers–Kouwenaar’s syndrome 196Mice as hosts
experimental 23, 71, 83, 120, 159, 250natural 37, 100, 153, 157, 164, 175
Microbilharzia variglandis 32Microfilariae
counting techniques 164–165differentiation 189staining 263–264
Microphallus minus 59Microtriches 63–64Millipore membrane filter 264Mites
as hosts 64, 65, 102Molluscicides 28Molluscs, bivalves, as intermediate hosts 56, 57,
59, 158, 184Moniliformis moniliformis 106–107
clinical manifestations 107life cycle 106location in host 106morphology 106pathology 107treatment 107
Monkeys as hosts 31, 56, 55, 103, 125, 140, 146,164, 168, 175, 187, 203, 211, 223
Mortality rate 24, 92, 96, 144, 150, 174, 231Mosquitoes
control 201–202, 205as intermediate hosts 199, 200, 205,
226Myiasis 241–242
Nanophyetus salmincola, N. s. schikhobalowi58–59
Natural killer (NK) cells 244Necator
americanus 126–134clinical manifestations 129–130diagnosis 132differentiation from Ancylostoma 130epidemiology 133geographical distribution 126immunology 120–122life cycle 127–128location in host 126morphology 126–127pathogenesis 130–132prevention and control 133–134treatment 132–133zoonotic aspects 134
argentinus 137suillus 137
Necrosis due to cestodes 96, 99intestinal nematodes 132, 144, 155tissue nematodes 179, 196trematodes 22, 36, 47, 53, 57
Nematodes 109–239classification of 109–115identification key for sections of 269key to those parasitic in humans
113–114life cycles 110mode of transmission 113recognition in sections 267–269
Nematodirus abnormalis 139Nematomorpha 108Neochordades columbianus 108Neodiplostomum seoulensis 58Neorickettsia helminthoeca 58 Neotricula aperta 27‘Nevinny’s basophilic halo’ 179Niclosamide 28, 53, 69, 75, 80, 99Nippostrongylus brasiliensis 120Nodulectomy 218Nodules 184, 186, 214, 218, 226Nuclear DNA reduction 154
Oedema due tocestodes 90intestinal nematodes 120, 132, 141, 167tissue nematodes 178, 179, 186, 194, 209,
215, 224, 230trematodes 50, 57
Oesophagostomumaculeatum 140, 141 apiostomum 142bifurcum 140–142
clinical manifestations 140–141diagnosis 141epidemiology and control 142geographical distribution 140life cycle 140morphology 140pathogenesis 140–141treatment 142
stephanostomum 140Oestrus ovis 242Oligochaetes see AnnelidsOnchocerca
armillatus 220cervicalis 220gibsoni 220gutturosa 220ochengi 220reticulata 220
Index 295
14Worms & H.D. - Index 20/11/01 1:46 PM Page 295

Onchocerca continuedvolvulus 211–220
clinical manifestations 213–217diagnosis 217–219epidemiology 218–219immunology 217geographical distribution 211life cycle 213location in host 211morphology 211pathogenesis 213–217prevention and control 219–220treatment 218zoonotic aspects 220
Onchocerciasis Control Programme (OCP)219–220
Oncomelania formosana 34hupensis 27lindoensis 27nosophora 34quadrasi 27spp. 5, 11–12, 27
Opisthorchis felineus 44–45
clinical manifestations 45diagnosis and treatment 45epidemiology 45geographical distribution 44immunology 41–42life cycle 44location in host 44morphology 44zoonotic aspects 45
guayaquitensis 45 noverca 45sinensis see Clonorchis sinensis viverrini 43–44
clinical manifestations and pathogenesis43
diagnosis 44epidemiology 44geographical distribution 39, 43immunology 41–42life cycle 43location in host 43morphology 43treatment 44zoonotic aspects 44
Orientobilharzia turkestanica 32Ornithobilharzia turkestanicum 32Ostertagia
cicumcincta 139 ostertagi 139
Oxamniquine in schistosomiasis 25Oxantel pamoate in trichuriasis 167
Palm Island mystery disease 159Pancreatitis 41, 150Pangolins as hosts 205Parachordodes
alpestris 108pustulosus 108raphaelis 108tolusanus 108violaceus 108wolterstoffi 108
Parafossarulus manchouricus 5, 38Paragonimus
africanus 37bangkokensis 37caliensis 37heterotremus 36, 37kellicotti 37mexicanus 37miyazaki 37ohirai 37philippinensis 37 pulmonalis 37 sadoensis 37skrjabini 37uterobilateralis 37westermani 32–37
clinical manifestations 35diagnosis 36epidemiology 36–37geographical distribution 32, 33life cycle 34–35location in host 32morphology 32pathogenesis 35prevention and control 37treatment 36zoonotic aspects 37
Paragordius cinctus 108esavianus 108tanganyikae 108tricuspidatus 108varius 108
Paralecithodendriumglandulosum 58obtusum 58
Parascaris equorum 154, 156Parastrongylus (= Angiostrongylus)
cantonensis 143–145, 156clinical manifestations 143–144diagnosis 144–145epidemiology 145geographical distribution 143life cycle 143location in host 143mophology 143treatment 145
296 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 296

costaricensis 145–147, 156clinical manifestations 146diagnosis 146geographical distribution 143life cycle 146location in host 145morphology 145–146treatment 147
mackerrasae 145malaysiensis 145vasorum 145
Paratenic hostsacanthocephalans 107cestodes 67, 70intestinal nematodes 145, 155tissue nematodes 184, 186, 188, 234trematodes 37, 58
Paryphostomumbangkokensis 59sufrartyfex 59
PCR (polymerase chain reaction) amplification30, 199, 200, 209, 218, 227
Peloderastrongyloides 126teres 126
Peritonitis 141, 150Phagicola
longa 55ornamenta 55ornata 55
Phaneropsolusbonnei 58spinicirrus 58
Pharyngostomum flapi 55Philophthalmus lacrymosus, P. sp. 59Physaloptera transfuga 187Pica see GeophagiaPigs as hosts
domestic ofcestodes 78, 80, 92intestinal nematodes 137, 140, 149,
153tissue nematodes 182, 183, 187trematodes 31, 43, 45, 54, 55
wild 80, 182, 183Pila sp. 56, 57Pirenella conica 5, 52Plagiorchis
harinasutai 58javensis 58muris 58philippinensis 58
Planarians as hosts 145Planorbis sp. 57Platyhelminthes 3Poikilorchis congolensis 38Polypylis (= Segmentina) hemisphaerula 50
Portal hypertension 21–22, 40, 41Praziquantel
cestode infections 68, 75, 80, 82, 84, 92, 99,101, 102
trematode infections 24, 36, 42, 44, 50, 53,57, 59
Prehistoric helminth infections 106, 164, 168Preservation of helminths 165–167Primates (non human) as hosts 23, 84, 125, 153,
168, 209, 211, 226Procerovum
calderoni 55varium 55
Prohemistomum vivax 59Prosthodendrium molenkampi 58Proteases 26, 119, 127Protozoans (except amoebiasis) 132, 249Pruritis 74, 119, 120, 129, 135, 209, 214, 222,
224, 230pruritis ani and pruritis vulvae 162
Pseudamphistomumaethiopicum 45truncatum 45
Pseudoacanthocephalus bufonis 107Pseudoapolysis 68Pseudophyllidea 64, 65–71Pseudoterranova decipiens 155Pseudotubercle 20, 35Psilorchis hominis 59Pygidiopsis summa 55Pyramicocephalus anthocephalus 69–70Pyrantel pamoate 133, 152, 163, 181
Rabbits and hares as hosts 85, 175, 211Radiography 18, 36, 81, 91, 146, 149, 151, 155,
158, 229Raillietina
asiatica 103celebensis 103garrisoni 103madagascariensis 103siviraji 103
Rats as hosts 37, 43, 54, 57Recognition of helminths in sections 267–269Rectal prolapse 166Reptiles as hosts 70, 71
snakes 187, 240Rhabditis
axei 126elongata 126inermis 126niellyi 126pellio 126sp. 126taurica 126terricola 126
Index 297
14Worms & H.D. - Index 20/11/01 1:46 PM Page 297

Rhinoestrus purpureus 242Rictularia spp. 188‘Ridley–Hissette’ fundus 216Rodents (apart from mice and rats) as hosts 31,
41, 55, 94, 97, 100, 101, 120, 146, 153,164, 188
Robertsiella sp. 27Ruminants see Herbivores
Sanitation andgeohelminths 133–134lymphatic filariasis 201–202schistosomiasis 29
‘Sashimi’ in transmission 45, 69, 156, 186Schistocephalus solidus 69Schistosoma
bovis 32curassoni 32haematobium 9–17intercalatum 10, 11, 13, 16, 17japonicum 10–17, 156malayensis 10, 11mansoni 10, 12–13, 15–17margrebowiei 32mattheei 31mekongi 10, 16, 31rodhaini 32sinensium 10, 31spindale 32
Schistosomatium douthitti 32, 156Schistosomes 9–32
clinical manifestations 17–23differential features 10–11, 13diagnosis 23–4epidemiology 25–27geographical distribution 10immunology 23life cycles 11–17morphology 10–11pathogenesis 17–23prevention and control 27–31snail hosts 27treatment 24–25zoonotic aspects 31–32
Schistosomula 12–14Semisulcospira
amurensis 5, 34, 54libertina 5, 34, 54
Sections, recognition of helminths in 267–269Sheep as hosts
acanthocephalans 241cestodes 85, 93nematodes 125, 139, 187trematodes 30, 31, 46, 47, 48, 49
Simulium spp.as vectors of onchocerciasis in Africa 219as vectors of onchocerciasis in the
Americas 219as vector of Mansonella ozzardi 225as vector of Dirofilaria ursi 228
Skin snip 218, 224Slugs as hosts 143, 146Snails
control of 28–29hosts of intestinal trematodes 5, 50, 52, 54,
55, 56, 57, 58hosts of liver flukes 38, 43, 44, 46, 47, 49,
50hosts of lung flukes 34hosts of nematodes 145hosts of schistosomes 27infected with larval trematodes
Snakes see ReptilesSoluble egg antigen (SEA) 22, 24Sowda 215Sparganosis 70–71Sparganum proliferum 71Spirocerca lupi 188Spirometra
erinacei 71houghtoni 71mansoni 71mansonoides 71 spp. 70–71, 156theileri 71
‘Splendore–Hoeppli’ phenomenon 23Splenomegaly 19, 21, 40, 97Sputum 35, 36, 90, 129, 138, 142, 149, 187, 211 Staining of
filarial larvae in insects 265microfilariae 263–264trematodes and cestodes 265
Stellantchasmusamplicaecalis 55falcatus 55
Stichosome 165, 177Stictodora
fuscata 55manilensis 55
Stoll egg counting technique 161–162‘Strogonina’ in transmission 71Strongyloides
canis 125cebus 125felis 125fuelleborni 125fuelleborni kellyi 125myopotami 125papillosus 125 planiceps 125procyonis 125ransomi 125ratti 120simiae 125stercoralis 115–126
298 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 298

clinical manifestations 119diagnosis 122–123epidemiology 124geographical distribution 115immunology 120–122life cycle 117–119location in host 115morphology 115–117pathogenesis 119–120prevention and control 124treatment 124–125zoonotic aspects 125–126
westeri 125venezuelensis 120
Stunting of growth 130, 137Suramin 218‘Swimmer’s itch’ 18, 32, 156Swine see PigsSwollen belly syndrome 125Symmer’s pipestem fibrosis see FibrosisSyphacia obvelata 164
T cells 20, 23, 30, 31, 96, 120, 198, 209, 217,244–245
Taeniabrauni 83crassiceps 83glomeratus 84longihamatus 85multiceps 83
clinical manifestations 83life cycle 83morphology 83pathogenesis 83
ovis 93saginata 71–76
clinical manifestations 74diagnosis 75epidemiology 75–76geographical distribution 72immunology 93life cycle 72–74location in host 72morphology 72pathogenesis 74prevention and control 76treatment 75zoonotic aspects 76
saginata asiatica 76serialis 84solium 76–82
cysticercosis 80–82clinical manifestations 79diagnosis 79epidemiology 80geographical distribution 76–77
immunology 93life cycle 78–79location in host 76morphology 78pathogenesis 79prevention and control 80treatment 79–80zoonotic aspects 80
taeniaeformis 250Tarebria granifera 29T cells 20, 23, 30, 31, 96, 120, 198, 209, 217,
244–245Temephos 220, 233Ternidens deminutus 139–140
clinical manifestations 139diagnosis 140epidemiology 140geographical distribution 139life cycle 139location in host 139treatment 140zoonotic aspects 140
Test tube culture of larvae (Harada–Mori)262–263
Tetrathyridium larva 103Thelazia
californiensis 187callipaeda 187
Thiabendazole 123, 147, 153, 154, 159, 167,175, 181, 232
Thiara granifera 5, 34‘Tonle sap’ schistosome 31Toxascaris leonina 157Toxocara
canis 157cati 157pteropodis 159
Trematodes 3–62classification 6–9life cycle stages 6medically important species 4mode of infection 8morphology 5, 7preservation of 265–266recognition in sections 267snail hosts of 5staining for 266
Trichinella britovi 182–183murrelli 182nativa 182–183nelsoni 182–183papuae 183pseudospiralis 182–183spiralis 120, 176–183
clinical manifestations 177–179diagnosis 180
Index 299
14Worms & H.D. - Index 20/11/01 1:46 PM Page 299

Trichinella continuedspiralis continued
epidemiology 182–184geographical distribution 176life cycle 176–177location in host 176morphology 176pathogenesis 179–180prevention and control 182treatment 181zoonotic aspects 182–183
Trichobilharzia brevis 32, 156maegraithi 32, 156ocellata 32, 156stagnicolae 32, 156
Trichocephalus see TrichurisTrichostrongylus
affinis 139axei 139brevis 139calcaratus 139capricola 139colubriformis 139lerouxi 139orientalis 139probolurus 139skjrabini 139vitrinus 139
Trichurismuris 120, 168ovis 168suis 168trichiura 164–168
clinical manifestations 165–166diagnosis 167epidemiology 167–168geographical distribution 164life cycle 165location in host 164morphology 164–165pathogenesis 166–167prevention and control 168treatment 167zoonotic aspects 168
vulpis 168Triclabendazole 48Tropical pulmonary eosinophilia 195–196Tuberculosis 20, 36, 119‘Tumeur de Dapaong’ 141Tumours in
cestodes 83, 89, 90–91intestinal nematodes 96, 99myiasis 242tissue nematodes 132, 144, 155trematodes 36
Tumour necrosis factor (TNF) 217
Tunga penetrans 240
Ulceration 20, 50, 53, 70, 119, 120, 137, 139,151, 155, 162, 230
Ultrasound (US) 18, 49, 81, 92, 96, 143, 151,194, 198, 199, 217
Uncinaria stenocephala 160Urine 16, 18, 24, 29, 31, 110, 123
Vaccines 30–31, 48, 96, 251Ventilated improved pit (VIP) latrine 134Viruses 3, 30, 71, 158, 166Visceral larva migrans 156–159Vitamins
A 215B12 68, 132 C 151
Vivipara sp. 57
‘Wakana disease’ 135Watch glass culture method for larvae 123Water plants and
fascioliasis 48fasciolopsiasis 51malayan filariasis 205
Watsonius watsoni 56Whipworms see TrichurisWHO (World Health Organization) initiatives 1,
200, 219, 233Wohlbachia in filariae 196–197, 218Wuchereria
bancrofti 190–202clinical manifestations 193–196diagnosis 198–199epidemiology 199–200geographical distribution 190immunology 196–198life cycle 192–193location in host 190morphology 190–191pathogenesis 196prevention and control 200–202treatment 199zoonotic aspects 202
bancrofti var. vauceli 192kalimantani 202lewisi 202
X-rays see Radiography
Zinc sulphate flotation for eggs 260
300 Index
14Worms & H.D. - Index 20/11/01 1:46 PM Page 300




















