
Women's sleep in health and disease
22
Women’s sleep in health and disease Andrea Dzaja a , Sara Arber b , Jenny Hislop b , Myriam Kerkhofs c , Caroline Kopp d , Thomas Pollm€ acher a ,P€ aivi Polo-Kantola e , Debra J. Skene f , Patricia Stenuit c , Irene Tobler d , Tarja Porkka-Heiskanen g, * a Max Planck Institute of Psychiatry, Munich, Germany b Department of Sociology, University of Surrey, Guildford, UK c Sleep Laboratory Vesale Hospital, Free University of Brussels, Belgium d Institute of Pharmacology and Toxicology, University of Zurich, Switzerland e Department of Physiology, University of Turku, Finland f School of Biomedical and Molecular Sciences, University of Surrey Guildford, UK g Institute of Biomedicine, University of Helsinki, Finland Received 19 January 2004; received in revised form 29 April 2004; accepted 8 May 2004 Abstract A huge amount of knowledge about sleep has accumulated during the last 5 decades following the discovery of rapid eye movement (REM) sleep. Nevertheless, there are numerous areas of considerable ignorance. One of these concerns the particularities of sleep in women. Most basic and clinical studies have been performed in male subjects, and only very recently research groups around the world have addressed women’s sleep in health and disease. In this review, we summarize the present knowledge on the influence of oestrogens on the brain and on the distinctive changes of sleep across the menstrual cycle, during pregnancy and menopause. In addition, studies in female rodents are reviewed as well as the knowledge on female peculiarities regarding the interactions between sleep regulation and age-related changes in circadian rhythms. We also address specific aspects of sleep loss and sleep disorders in women. Finally, very recent studies on the sociology of sleep are summarized and future directions in the field are discussed. Ó 2004 Elsevier Ltd. All rights reserved. Keywords: Women; Sleep; Hormones; Circadian; Menstrual cycle; Pregnancy; Sex steroids 1. Introduction In sleep surveys, women report considerably more sleep problems than men (e.g., Akerstedt et al., 2002a,b; Hublin et al., 1996; Lindberg et al., 1997), while poly- somnographically measured sleep quality in elderly men appears to be worse than in women (Buysse et al., 1991). But does this necessarily imply differences in sleep be- tween men and women? As most studies on sleep and sleep loss have been conducted in males, it is not yet clear which factors contribute most to the sleep distur- bances reported by women. Changes in sleep patterns are often associated with hormonal factors, particularly oestrogen levels, with women typically suffering from disturbed sleep in connection with the menstrual cycle, pregnancy and menopause. Yet explaining disturbances in women’s sleep is more complex. What is the role of the genome and other biological factors as opposed to psychological, sociological and socio-economic factors? Men and women, even when sharing a common home, mostly live very different lives, with different responsi- bilities and rights. Taking care of an infant or elderly relatives, for example, also disturbs sleep (Driver and Shapiro, 1992; Santiago et al., 2001). In this context, women’s sleep patterns, as well as health and well-being, may be determined not only by physiology but rather by social roles and interactions. Using a multidisciplinary approach, this review aims to summarise what is pres- ently known about different aspects of sleep and sleep disturbances in women. * Corresponding author. Tel.: +358-9-1918533; fax: +358-9-1918681. E-mail address: [email protected].fi (T. Porkka-Heiskanen). 0022-3956/$ - see front matter Ó 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.jpsychires.2004.05.008 Journal of Psychiatric Research 39 (2005) 55–76 J OURNAL OF P SYCHIATRIC RESEARCH www.elsevier.com/locate/jpsychires
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Women's sleep in health and disease

JOURNALOF
PSYCHIATRIC
Journal of Psychiatric Research 39 (2005) 55–76RESEARCH
www.elsevier.com/locate/jpsychires
Women’s sleep in health and disease
Andrea Dzajaa, Sara Arberb, Jenny Hislopb, Myriam Kerkhofsc, Caroline Koppd,Thomas Pollm€achera, P€aivi Polo-Kantolae, Debra J. Skenef, Patricia Stenuitc,
Irene Toblerd, Tarja Porkka-Heiskaneng,*
a Max Planck Institute of Psychiatry, Munich, Germanyb Department of Sociology, University of Surrey, Guildford, UK
c Sleep Laboratory Vesale Hospital, Free University of Brussels, Belgiumd Institute of Pharmacology and Toxicology, University of Zurich, Switzerland
e Department of Physiology, University of Turku, Finlandf School of Biomedical and Molecular Sciences, University of Surrey Guildford, UK
g Institute of Biomedicine, University of Helsinki, Finland
Received 19 January 2004; received in revised form 29 April 2004; accepted 8 May 2004
Abstract
Ahuge amount of knowledge about sleep has accumulated during the last 5 decades following the discovery of rapid eye movement
(REM) sleep. Nevertheless, there are numerous areas of considerable ignorance. One of these concerns the particularities of sleep in
women.Most basic and clinical studies have been performed inmale subjects, and only very recently research groups around the world
have addressed women’s sleep in health and disease. In this review, we summarize the present knowledge on the influence of oestrogens
on the brain and on the distinctive changes of sleep across the menstrual cycle, during pregnancy and menopause. In addition, studies
in female rodents are reviewed as well as the knowledge on female peculiarities regarding the interactions between sleep regulation and
age-related changes in circadian rhythms. We also address specific aspects of sleep loss and sleep disorders in women. Finally, very
recent studies on the sociology of sleep are summarized and future directions in the field are discussed.
� 2004 Elsevier Ltd. All rights reserved.
Keywords: Women; Sleep; Hormones; Circadian; Menstrual cycle; Pregnancy; Sex steroids
1. Introduction
In sleep surveys, women report considerably more
sleep problems than men (e.g., Akerstedt et al., 2002a,b;
Hublin et al., 1996; Lindberg et al., 1997), while poly-
somnographically measured sleep quality in elderly men
appears to be worse than in women (Buysse et al., 1991).
But does this necessarily imply differences in sleep be-tween men and women? As most studies on sleep and
sleep loss have been conducted in males, it is not yet
clear which factors contribute most to the sleep distur-
bances reported by women. Changes in sleep patterns
are often associated with hormonal factors, particularly
* Corresponding author. Tel.: +358-9-1918533; fax: +358-9-1918681.
E-mail address: [email protected] (T. Porkka-Heiskanen).
0022-3956/$ - see front matter � 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jpsychires.2004.05.008
oestrogen levels, with women typically suffering from
disturbed sleep in connection with the menstrual cycle,
pregnancy and menopause. Yet explaining disturbances
in women’s sleep is more complex. What is the role of
the genome and other biological factors as opposed to
psychological, sociological and socio-economic factors?
Men and women, even when sharing a common home,
mostly live very different lives, with different responsi-bilities and rights. Taking care of an infant or elderly
relatives, for example, also disturbs sleep (Driver and
Shapiro, 1992; Santiago et al., 2001). In this context,
women’s sleep patterns, as well as health and well-being,
may be determined not only by physiology but rather by
social roles and interactions. Using a multidisciplinary
approach, this review aims to summarise what is pres-
ently known about different aspects of sleep and sleepdisturbances in women.

56 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
2. Basics about sleep and circadian rhythms
2.1. Sleep
Sleep is a behavioural state characterised by a typicalbody position, usually the eyes are closed and respon-
siveness to external stimuli is diminished. Sleep is most
accurately defined by electroencephalography (EEG)
measures which allow, together with the electromyo-
gram (EMG) and electrooculogram (EOG) define, the
classification into different sleep stages according to a set
of criteria (Rechtschaffen and Kales, 1968). In addition,
spectral analysis of the sleep EEG has led to importantinsights into the sleep regulatory processes (e.g., Daan
et al., 1984). Spectral power, particularly in the low
frequency band, the ‘‘delta’’ band (0.75–4.5 Hz), has
proven to be a reliable marker of sleep homeostasis and
is thought to reflect sleep intensity (Borb�ely, 1982a,b).Sleep is not merely a passive state characterised by the
absence of wakefulness. During sleep the cortical ac-
tivity of the brain undergoes regular, cyclic changesbetween the two main sleep states rapid eye movement
(REM) sleep (also called active sleep or paradoxical
sleep) and non-REM sleep, which in humans and other
primates comprises 4 stages (S1–S4). Stages S3 and S4
are defined as deep sleep or slow wave sleep (SWS) in
humans (Rechtschaffen and Kales, 1968). Sleep begins
with S1, proceeds to S2 and usually through S3 and S4
to REM sleep, before the cycle starts again. During non-REM sleep physiological variables such as heart rate,
respiration and cortical activity slow down, body tem-
perature decreases and energy consumption is lower.
When REM sleep begins, cortical activity resembles the
waking pattern. During this state, activity of the motor
neurones at the level of the brain stem/spinal cord is
inhibited, so that body muscles cannot move.
SWS is generally regarded as the deeper, refreshing,restorative part of sleep, though it is not clear what
substrate in the brain and/or in the body is restored
(Rechtschaffen, 1998). The role of REM sleep is even
less clear. Despite many theories including a role in
brain plasticity and learning, experimental evidence for
any of these theories is still scarce (Siegel, 2001; Tononi
and Cirelli, 2001).
2.2. Circadian rhythms
The two main components of physiological sleep
regulation are the homeostatic and the circadian com-
ponent, described in the two process model of sleep
regulation (Borb�ely, 1982a,b). The circadian component
regulates the timing of sleep: for humans sleeping is
easier during the night than during the day. The ho-meostatic component keeps track of the duration of the
previous wakefulness period: the longer we stay awake,
the longer and deeper will be the following sleep. These
components normally work together to ensure correctly
timed and sufficient sleep. However, it is important for
the diagnosis of sleep disturbances to understand the
relation of the two components and to be able to assess
them separately. Both ageing and gender may differe-nentially affect the components of sleep regulation.
Circadian rhythms are endogenously generated by
the pacemaker localised in the hypothalamic suprach-
iasmatic nuclei (SCN). Efferent signals from the SCN
drive circadian rhythms in physiology (melatonin, core
body temperature, cortisol) and behaviour (sleep/wake
cycle, alertness, performance) that have a periodicity of
approximately (circa) 24 h. The circadian SCN clockand its output rhythms are synchronised (entrained) to
the 24-h day by environmental light which is transmitted
from the retina to the SCN primarily via the retinohy-
pothalamic tract (RHT). Ocular light is the major time
cue (also called ‘‘zeitgeber’’) for synchronisation of the
human circadian clock (e.g., Lockley et al., 1997; Skene
et al., 1999), the resetting effect of light being dependent
upon intensity (Boivin et al., 1996; Zeitzer et al., 2000)and circadian time of exposure (Czeisler et al., 1989;
Honma and Honma, 1988; Khalsa et al., 2003; Minors
et al., 1991; Van Cauter et al., 1994).
The two main sleep states, non-REM sleep and REM
sleep, are at least in part regulated separately, and un-
dergo different changes during development and ageing.
2.3. Contribution of the circadian system to age-related
changes in sleep
Studies investigating the role of the circadian system
in age-related sleep changes indicate an inability to
sustain sleep at particular circadian phases (Dijk et al.,
1999; Duffy et al., 1998, 1999a,b; Duffy and Czeisler,
2002). In strictly controlled laboratory studies, older
subjects (64–74 years) exhibited an earlier wake time andan advance (by approximately 1 h) in the rhythms of
melatonin and core body temperature compared to
younger subjects (Dijk et al., 1999; Duffy et al., 1998,
1999a,b). The circadian phase occurred later relative to
the habitual wake up time in the older subjects com-
pared to the young subjects suggesting that older people
wake up earlier in the circadian cycle.
2.4. Age-related changes in sleep
A prominent change with ageing is the reduction in
stages 3 and 4 sleep (SWS) (Feinberg, 1974; Lee and
DeJoseph, 1992; Schweiger, 1972), particularly in men.
This decline coincides with the reduction in growth
hormone (GH) secretion (Van Cauter et al., 2000). As
GH is involved in the regulation of non-REM sleep, ithas been speculated that the decrease in GH could at
least partly explain the decline in SWS. It is interesting
to note that aged women have better preserved SWS

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 57
than aged men (Webb, 1982; Reynolds et al., 1985).
Moreover, the difference in SWS appears to take place
after the age of 30 years, young women and men had
equal amounts of SWS as measured by spectral power
(Ehlers and Kupfer, 1997).Reduced sleep efficiency is also a frequent finding in
the elderly, while stage 1 sleep increases. The number of
nocturnal awakenings increases, as does nocturnal sleep
latency (Carskadon et al., 1982); fragmented sleep ap-
pears to be the main reason for subjectively poor sleep.
An additional frequent finding is an increase in early
morning awakenings and difficulty falling asleep again
(e.g., Feinsilver, 2003). REM sleep appears to be pre-served better in the course of ageing, though shorter
REM latencies have been reported, and REM latencies
are longer in old women than men (Rediehs et al., 1990).
It should be kept in mind that difficulties with sleep in
the elderly are often associated with poor health (for
example, pain, breathing problems and restless legs) and
depression. Side effects of medication may also con-
tribute to poor sleep quality.
2.5. Age-related changes in circadian rhythms
Both endogenous and exogenous factors can con-
tribute to age-related changes in circadian rhythms.
Ageing of the retina, the SCN oscillator, the SCN af-
ferent and efferent neural connections or the down-
stream target nuclei or glands such as the pineal glandare possible endogenous factors (Skene and Swaab,
2003), while changes in environmental light exposure
such as reduced daytime illumination or changes in be-
haviour including early morning awakening or daytime
napping are exogenous factors.
Changes in both the amplitude (amount) and phase
(timing) of circadian rhythms have been reported with
ageing in humans. The reduction in the amplitude of themelatonin rhythm has been widely reported (Bojkowski
and Arendt, 1990; Ferrari et al., 2000; Iguichi et al.,
1982; Kennaway et al., 1999; Luboshitzky et al., 1998;
Sack et al., 1986; Skene et al., 1990a,b; Waldhauser
et al., 1988; Young et al., 1988), as well as a reduction in
the amplitude of the core body temperature rhythm
(Carrier et al., 1996; Dijk et al., 2000; Duffy et al., 1998).
Although many reports indicate that melatonin levelsdecline with age, some recent studies do not support a
reduction (Zeitzer et al., 1999; Fourtillan et al., 2001).
The constant routine conditions (dim light, semi-
recumbent, isocaloric, sleep deprived) and the strict se-
lection criteria for the elderly subjects (non-medicated,
abstinence from alcohol, caffeine, nicotine) the subjects
underwent in these studies may explain the discrepancy
(Skene and Swaab, 2003).Another commonly reported feature in the elderly is
the earlier timing (phase advance) of circadian rhythms
in melatonin (Dijk et al., 1999; Duffy et al., 1999a,b),
core body temperature (Carrier et al., 2002; Duffy et al.,
1998) and cortisol (Sherman et al., 1985). Changes in
circadian rhythms may be due to global neurodegener-
ation with ageing, which includes the SCN (Swaab et al.,
1985), producing a general disturbance of all SCN-dri-ven circadian rhythms. Changes in the timing and
amount of light exposure either as a result of changes in
behaviour, for example, early morning awakening or
age-related changes in the photic input pathway (retina-
RHT-SCN) may also explain the age-related changes in
circadian rhythmicity. Reduced photic entrainment
could also be a result of environmental factors. For
example, there is evidence to suggest that elderly peopleare exposed to reduced illumination levels in their daily
lives (Campbell et al., 1988; Mishima, 2001; Shochat
et al., 2000; Van Someren et al., 1997). Some studies
have shown an inverse relationship between light in-
tensity and sleep disturbances in the elderly (Mishima
et al., 2001; Shochat et al., 2000).
3. Sex steroids and sleep
Sex steroids, e.g., oestrogen, progesterone and tes-
tosterone, not only exert peripheral effects on female sex
organs such as the gonads, but also have powerful ac-
tions on the brain (Baulieu, 1998; Fink et al., 1998;
McEwen, 1999a,b). It is remarkable that sex steroids are
not only involved in the regulation of gonadal hormonesecretion and reproductive behaviours, but also affect
functions such as cognitive performance and sleep
(Sherwin, 2003; Thomson and Oswald, 1977a,b; Manber
and Armitage, 1999; Empson and Purdie, 1999). Apart
from the knowledge derived from the effects of hormone
replacement therapies (HRT) and oral contraceptives on
sleep, data on the influence of steroid hormones on
human sleep are limited. While the effects of oestrogenon brain function have been intensively studied in hu-
mans, the effects of progesterone, that acts in the brain
as a neurosteroid, have been studied mainly in rodents,
and they are less well understood in humans. Thus for
example, the effects of progesterone on sleep and the
sleep EEG were investigated in a double-blind placebo-
controlled crossover study in nine male subjects (Friess
et al., 1997). Progesterone led to an increase in non-REM sleep, a decrease in slow frequency range EEG
activity (0.4–4.3 Hz), and an increase in the higher fre-
quency range activity (>15 Hz) during non-REM sleep.
Furthermore, pregnenolone, a precursor of progester-
one acting as a c-amino-butyric acid (GABA A) recep-
tor agonist, enhanced SWS and reduced EEG activity
in higher frequency bands (Steiger et al., 1993). The role
of physiological oestrous-related changes in progester-one levels on sleep as well as the consequences of pro-
gesterone treatment in females needs to be further
investigated.

58 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
Oestrogen has recently been established as a multi-
purpose brain messenger (Halbreich and Kahn, 2000;
Toran-Allerand, 2000). Through manifold interactions
with the most important neurotransmitter systems at
critical brain nuclei (e.g., the basal forebrain and theraphe nuclei) oestrogen can regulate a multitude of
brain functions, including cognitive performance, mood,
movement co-ordination, pain and sleep (Toran-Aller-
and, 2000; Hutchison and Beyer, 1994; Natale et al.,
2001). It has been shown that many of these interactions
are gender related (McEwen, 1988).
3.1. Menstrual cycle and sleep
Female sex steroids have various effects on central
and peripheral systems. The fluctuating character of
female sex hormones, which is essential for the men-
strual cycle, is reflected in changes in cognitive function,
mood, appetite and sexual activity as well as in physi-
ological changes in temperature and sexual organs such
as the uterus, ovaries and mammae. These complex in-fluences of female sex hormones on different systems
may contribute to the sleep changes that have been
observed in the course of the menstrual cycle.
3.1.1. Hormonal changes across the menstrual cycle
In the central nervous system, the menstrual cycle is
regulated by the pulsatile secretion of the hypothalamic
gonadotropin-releasing hormone (GnRH), which regu-lates the secretion of the pituitary hormones – luteiniz-
ing hormone (LH) and follicle stimulating hormone
(FSH). These hormones in turn regulate the secretion of
oestrogen. The release of GnRH is regulated by neuro-
transmitters (e.g., GABA, DA and serotonin), neuro-
peptides, endogenous opioids and steroid hormones
(oestrogen, progesterone and testosterone) (Jaffe et al.,
1990; Levine et al., 1995; Silberstein and Merriam,2000).
45
30
15
Day 2 4 6 8 10 12 14
Oestrogenpg/ml
FSHmlE/ml
LH
FSH
Oe
Fig. 1. Female sex hormone concentrations across the me
The menstrual cycle lasts, on average, 28 days and
consists of the follicular (before ovulation) and the luteal
(after ovulation) phases; ovulation takes place 14 days
before menstruation begins. Precise and co-ordinated
changes in the plasma concentration of the reproductivehormones oestrogen, progesterone, LH and FSH are
required to create the conditions needed for the matu-
ration and ovulation of a single oocyte each month from
puberty to menopause, the induction of menstruation or
the implantation of the conceptus (Chabbert et al., 1998)
(Fig. 1).
Highest values of oestrogen occur shortly before
ovulation in the follicular phase, with a second increasein the luteal phase, whereas progesterone concentrations
rise in the luteal phase. At the end of the follicular
phase, the LH peak induces ovulation leading to a rise in
body temperature of about 0.5 �C.
3.1.2. Menstrual cycle effects on circadian timing
Female sex hormones, the menstrual cycle, oral
contraceptive use and HRT may all have confoundingeffects on the measurement of circadian rhythms. Tra-
ditionally women have been excluded from most circa-
dian studies, but the recent demonstration of sex
hormone receptors (ERa, ERb and progesterone re-
ceptors) in the human SCN with increased ERa ex-
pression in premenopausal women compared to young
males (Kruijver and Swaab, 2002) suggests the possi-
bility of a direct effect of sex hormones on the humancircadian system.
A gender difference in the phase of the circadian body
temperature rhythm has been reported in young people.
Compared to men, naturally cycling women have been
shown to exhibit phase-advanced circadian temperature
rhythms (Baehr et al., 1999; Baker et al., 2001; Lee,
1988), whereas others have found no gender differences
in circadian phase (Kattapong et al., 1995; Winget et al.,1977). The amplitude of the temperature rhythm may
16 18 20 22 24 26 28
45
30
15
Progesteroneng/ml
LHmlE/ml
P
nstrual cycle (Schmidt-Matthiesen, 1992, modified).

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 59
also be reduced in cycling women compared with men
(Kattapong et al., 1995; Rogacz et al., 1988).
In general, the melatonin rhythm (phase, amplitude
and total amount produced) does not appear to change
throughout the menstrual cycle in normally cyclingwomen (Berga and Yen, 1990; Brzezinski et al., 1988;
Parry et al., 1997). This lack of effect of the menstrual
cycle on melatonin rhythmicity requires confirmation in
dim light and posture-controlled experiments. In addi-
tion, studies designed to assess gender differences in the
melatonin profile are lacking. The large inter-individual
variation in melatonin production (for review, see
Arendt, 1995) may mask observation of any genderdifferences.
3.1.3. Sleep measurements across the menstrual cycle
There are few systematic studies addressing changes
in sleep during the menstrual cycle. Driver and col-
leagues (1996) obtained polysomnographic recordings at
2-day intervals throughout the menstrual cycle in nine
healthy young women. Subjective ratings of sleep qual-ity and mood did not show significant variations across
the menstrual cycle. No differences were observed in
total sleep time, sleep efficiency, sleep latency, REM
sleep latency or SWS. However, there was a consistent
variation in EEG power density in the 14.25–15.0 Hz
band corresponding to the upper frequency range of
sleep spindles, with a maximum of power density ex-
pressed in the luteal phase, parallel with the rise in bodytemperature. A reduction in the power density of the
same frequency band has been reported during preg-
nancy (Brunner et al., 1994), suggesting that this par-
ticular frequency band is sensitive to modulation by sex
steroids and/or body temperature. It is notable that
delta power (0.75–4.5 Hz) did not show changes during
the menstrual cycle, suggesting that the homeostatic
regulation of sleep is not affected by the hormone fluc-tuation during the menstrual cycle. REM sleep, ex-
pressed as a percentage of total sleep time, tended to be
higher in the early follicular phase (27.4%) compared to
the late luteal phase (22.9%), and it was inversely cor-
related with body temperature.
Changes in sleep spindles across the menstrual cycle
were also observed in another study: the lowest spindle
frequency appeared in the follicular phase about 18days before the onset of menstruation, whereas the
highest frequency was measured in the luteal phase 3
days before the onset of menstruation (Ishizuka et al.,
1994).
A significant increase in sleepiness and occurrence of
SWS during the daytime in the luteal phase was ob-
served by Shibui et al. (2000). A decrease in the ampli-
tude of core body temperature, TSH and cortisolrhythms, as well as a decrease in melatonin secretion in
the luteal phase was reported. Also, REM sleep latency
was shorter in the luteal phase compared to the follic-
ular phase, while the percentages of various sleep stages
remained unchanged (Lee et al., 1990).
In summary, in healthy women total sleep time, sleep
efficiency, sleep latency and subjective sleep quality do
not appear to be strongly modulated by the menstrualcycle (Baker et al., 2001; Driver et al., 1996; Ishizuka
et al., 1994; Lee et al., 1990; Parry et al., 1989). In the
high frequency spindle range EEG power density un-
dergoes a marked increase in the luteal phase, coinciding
with an increase in body temperature (Driver et al.,
1996). Some changes in REM sleep and REM sleep la-
tency have been reported (Driver et al., 1996; Lee et al.,
1990), but the findings have not been consistent acrossstudies (Baker et al., 2001).
3.1.4. Sleep complaints associated with the menstrual
cycle
Sleep disturbances perceived by women in associa-
tion with the menstrual cycle often occur in parallel
with several other symptoms. The exacerbation of cer-
tain medical conditions including migraine, epilepsy,asthma, rheumatoid arthritis, irritable bowel syndrome
and diabetes, at specific phases of the menstrual cycle is
well documented (Case and Reid, 1998). Also symp-
tomatic intensification of psychiatric disorders such as
schizophrenia, bipolar disorder, depression, anxiety
disorders, bulimia nervosa and substance abuse have
been reported during the pre-menstrual and menstrual
phases (Hendrick et al., 1996). As the symptoms usuallycluster in the late luteal phase, the condition is referred
to the ‘‘pre-menstrual dysphoric disorder’’ (PMDD),
which involves various mood disturbances including
depressed mood, irritability, dysphoria, affect lability,
anxiety and changes in appetite as well as physical
symptoms such as breast tenderness, headache, acne
and cramps (Andersch and Hahn, 1981; Vanselow,
1998; Gotts et al., 1995; Hurt et al., 1992; West, 1989).Women with PMDD complain about sleep distur-
bances such as hypersomnia, insomnia, awakenings
during the night, tiredness in the morning, unpleasant
dreams, a marked lack of energy and concentration
difficulties, especially in the late luteal phase of the
menstrual cycle (Eriksson et al., 1990; Smith et al.,
2003; Cohen et al., 2002).
The subjective experience of poor sleep quality hasbeen verified in several studies by polysomnographic
recordings. When women with severe pre-menstrual
depression were compared to healthy controls, an in-
crease in stage 2 sleep and a decrease of REM sleep were
measured (Parry et al., 1989). Stage 3 sleep and the
number of intermittent awakenings varied significantly
with phases of the menstrual cycle. The number of
awakenings reached its maximum in the late luteal phasewith lowest values for all the sex hormones involved and
showed a minimum in the early luteal phase shortly after
the peak of LH and FSH when progesterone rises. The

60 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
number of awakenings was associated with LH levels.
EEG alterations similar to those associated with major
depressive disorders, such as a reduced REM sleep la-
tency and an increased REM density (Benca et al., 1992;
Parry et al., 1989).Women with negative affect symptoms (verified with
the profile of moods state scale, POMS) demonstrated
during the pre-menstruum, significantly less delta sleep
during both the follicular and luteal phases, in com-
parison to healthy controls. REM sleep latency was
shown to be significantly shorter during the luteal phase
compared to the pre-ovulatory phase, while there was
no difference in sleep latency or the percentage of REMsleep (Lee et al., 1990; Parry et al., 1989).
Subjective sleep quality, sleep efficiency and REM
sleep of women suffering from primary dysmenorrhoea
decreased in the presence of dysmenorrhoic pain, while
SWS was not affected. Even in the absence of pain the
dysmenorrhoic women had different sleep patterns to
controls: in the mid-luteal, mid-follicular and menstrual
phases, the dysmenorrhoics expressed less REM sleepcompared to the controls. The negative correlation be-
tween body temperature and the amount of REM sleep
was observed in both dysmenorrhoic and control sub-
jects (Baker et al., 1999).
In conclusion, both subjective and objective sleep
quality are impaired in women suffering from dysm-
enorrhoea and/or PMS/PMDD.
3.2. Pregnancy and sleep
Pregnancy is accompanied by dramatic changes in
hormonal levels: oestrogen, progesterone and prolactinlevels increase, as well as human chorionic gonadotropin
(HCG), that is produced by the placenta. A rapid
withdrawal of most of these hormones at the time of
childbirth is accompanied by an additional increase in
circulating prolactin (Darling and Hawkins, 1981; Go-
ebelsmann, 1979; Chrousos et al., 1998).
The subjective sleep quality of most pregnant women
is disturbed starting as early as during the first trimesterof pregnancy (Hedmann et al., 2001; Santiago et al.,
2001). According to sleep surveys, the most frequent
subjective reasons for disturbed sleep during pregnancy
are restless legs and leg cramps, low back pain, foetal
movement and frequent micturation (Hertz et al., 1992).
The prevalence of restless legs syndrome increases dur-
ing pregnancy (Hedmann et al., 2002) and has been
found to be a major factor reducing sleep (McParlandand Pearce, 1988). Periodic leg movements occurred in
all 10 women who had multiple pregnancies (Nikkola
et al., 1996).
During the first trimester of pregnancy the total
amount of sleep increased significantly, decreased in the
second trimester and was shortest three months after
delivery (Hedmann et al., 2001). Sleep became more
restless and fragmented towards the end of pregnancy
accompanied by reduced sleep quality, but was nor-
malised within approximately three months after de-
livery. Although total sleep time increased in the firsttrimester, subjective sleep quality was worse. Due to
decreased depth of sleep, woman in the third trimester
were more likely to be awakened by noise in the envi-
ronment (Hedmann et al., 2002). At the same time
mechanical effects induced a more frequent urge to
urinate, resulting in more awakenings (Baratte-Beebe
and Lee, 1999).
Polysomnographic studies during pregnancy haveshown some inconsistent findings. A decrease in REM
sleep (Brunner et al., 1994; Hertz et al., 1992), but also
no change in REM sleep was observed (Driver and
Shapiro, 1992; Lee et al., 2000a,b). Increased awaken-
ings after sleep onset as well as decreased sleep efficiency
are consistent findings (Brunner et al., 1994; Driver and
Shapiro, 1992; Hertz et al., 1992; Lee et al., 2000a,b).
SWS has been found to be decreased, increased or un-changed. Spectral analysis of the EEG in non-REM
sleep revealed a progressive reduction of power density
during the course of pregnancy; the largest decrease
(30%) occurred in the 14.25–15.0 Hz band (Brunner
et al., 1994). Sleep continues to be disturbed after de-
livery. In addition to nocturnal awakenings for breast-
feeding, hormonal changes may play a role in sleep
disturbances during the postpartum period (Lee et al.,2000a,b; Swain et al., 1997; Lee et al., 1992; Schweiger,
1972; Hertz et al., 1992).
Possible influences of prolactin on sleep were inves-
tigated in a study comparing 12 breast-feeding women
with controls and with women who bottle-fed their in-
fants. Polysomnography showed a marked increase in
SWS in lactating women as compared to the other two
groups that may be attributed to increased levels ofcirculating prolactin (Blyton et al., 2002). Postpartum
women reported more time awake after retiring and
more naps than control women while overall sleep time
was equal between the groups (Swain et al., 1997). In
summary, pregnancy- and postpartum-related sleep
disturbances have a complex background that includes
hormonal, mechanical, emotional and sociological
factors.
3.3. Menopause and sleep
3.3.1. Menopausal transition
A cessation of ovarian endocrine function at meno-
pause leads to a marked decrease in endogenous oest-
rogen and progesterone secretion. This change results in
a more than threefold increase in the secretion of FSH
from the pituitary gland. An elevation of FSH over 30
IU/L confirms the menopause. Alterations in several

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 61
other hormones, including an increase in LH or a
decrease in androgens and prolactin may also occur
(Speroff, 1994). During the menopausal transition, sev-
eral biological functions, including sleep, are affected.
During the menopause, women seek medical assis-tance for the following symptoms: vasomotor instability
(hot flashes and sweating), disturbances in the menstrual
pattern, somatic symptoms (insomnia, headache, dizzi-
ness, palpitation, numbness, myalgia, vaginal and
urinary tract symptoms), as well as psychological
symptoms (anxiety, depression, decline in libido, lack of
concentration, and memory impairment) (Erkkola et al.,
1991; Holte and Mikkelsen, 1991; Oldenhave et al.,1993). Vasomotor symptoms are observed in 68–85% of
symptomatic menopausal women, whereas insomnia is
present in 51–77% (Brincat and Studd, 1988; Erkkola
et al., 1991; Anderson and Falestiny, 2000).
3.3.2. Sleep quality during the menopausal transition
According to a study conducted in France, 73% of
12,778 subjects had experienced sleeping problems dur-
ing the month preceding the survey and 29% of them
had continuous sleeping problems (Leger et al., 2000).
Women more often reported sleep disturbances than
men (Leger et al., 2000; Ohayon, 1996). Insomnia wasreported by 25% of the women and severe insomnia by
15% of the women between 50 and 64 years of age; in the
age group over 65 years the prevalence was 25% and
16%, respectively (Leger et al., 2000). However, epide-
miological data indicating which proportion of sleep
disturbances in middle-aged women is directly linked to
the menopause is lacking.
Sleep disturbances can either be caused by the men-opause or just coincide with the menopausal period. The
clinical picture of menopausal insomnia is indistin-
guishable from common insomnia, which manifests it-
self as difficulty in falling asleep, frequent awakenings or
awakening too early in the morning (Roffwarg, 1979).
Frequent awakenings may suggest that insomnia is
secondary to vasomotor events inducing awakenings
(Woodward and Freedman, 1994). Although vasomotorsymptoms correlate strongly with sleep complaints
(Polo-Kantola et al., 1999a,b), insomnia may occur in
the absence of vasomotor symptoms and can be the
exclusive climacteric symptom. Early morning awaken-
ing may indicate menopausal depression. In the morn-
ing, women may have a feeling of insufficient and
non-refreshing sleep and complain about fatigue, tired-
ness, lack of initiative and impaired memory (Brincatand Studd, 1988).
3.3.3. Circadian rhythm changes with ageing
A few studies have investigated the circadian systemin the elderly and the influence of gender. Campbell and
colleagues showed that the acrophase of body temper-
ature was phase-advanced (by an average of 1.25 h) in
older women compared to age-matched men (Campbell
et al., 1989). The older women also woke up earlier and
had a shorter sleep duration. Similar advanced temper-ature rhythms have also been observed in healthy elderly
women compared to elderly men (Moe et al., 1991). In
this study, the older women also exhibited a larger
amplitude and a higher peak of core body temperature
compared with the men.
3.3.4. Effect of hormone replacement therapy on sleep
quality
Hormone replacement therapy (HRT) is widely used
to control climacteric symptoms (Erkkola et al., 1991;
Wiklund et al., 1992, 1993). It is also an effective treat-
ment to control menopausal sleeping complaints (Polo-Kantola et al., 1998). It facilitated falling asleep and
decreased nocturnal restlessness and awakenings. Wo-
men reported less tiredness in the morning and during
the daytime. The degree of improvement in vasomotor
symptoms was an important predictor of the degree of
improvement in sleep disturbance, providing further
evidence that these two complaints are linked. However,
the subset of menopausal women who reported insom-nia in the absence of vasomotor symptoms, also mark-
edly benefit from HRT (Polo-Kantola et al., 1998). This
was also demonstrated in a recent large randomised,
placebo-controlled study evaluating the long-term ef-
fects of HRT (Women’s Health Initiative, WHI) where
participants were mainly climacterically asymptomatic
(Hays et al., 2003). Two explanations are possible:
firstly, women regarding themselves as asymptomaticmay nevertheless have hot flushes. If this is the case,
alleviation of the vasomotor symptoms again plays an
important role in improving sleep quality. Secondly,
sleep disturbances may be a direct consequence of
changing hormone levels associated with menopause.
There are contradictory findings about the effects of
HRT on objective sleep quality as measured by all night
polysomnography. The main outcome of previousstudies in healthy women is presented in Table 1.
Menopausal insomnia is not characterised by specific
abnormalities in a polysomnographic sleep recording
(Polo-Kantola et al., 1999a,b). However, intensive hot
flashes may cause arousals (Erlik et al., 1981) and have
been found to be associated with increases in stage 4
sleep, a shortened first REM sleep period and disrupted
sleep with reduced sleep efficiency (Woodward andFreedman, 1994).
Some authors have reported fewer awakenings, a
decrease in nocturnal wakefulness and an increase in
REM sleep during HRT (Antonijevic et al., 2000;
Thomson and Oswald, 1977a,b). Moreover, a shorten-
ing of sleep latency, an increase in REM sleep (Schiff
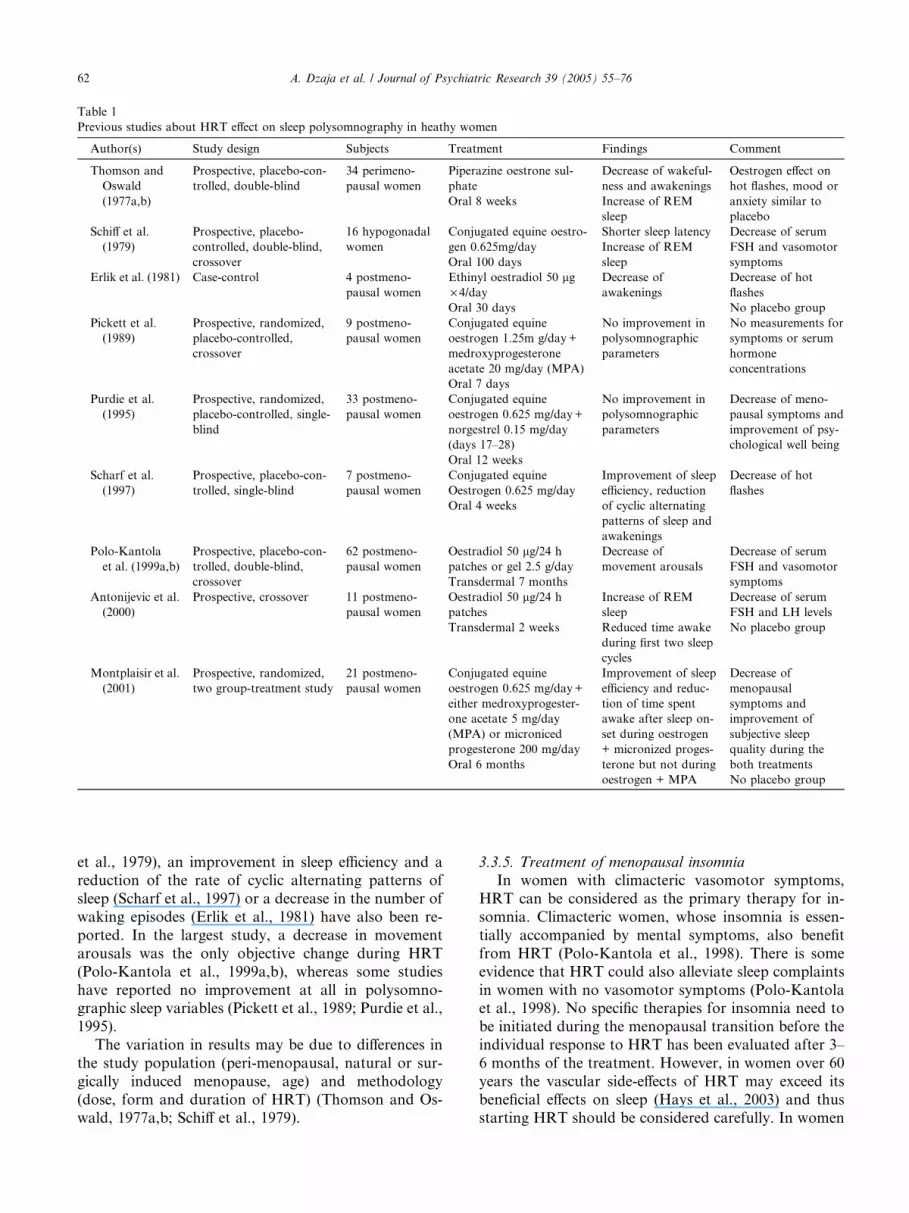
Table 1
Previous studies about HRT effect on sleep polysomnography in heathy women
Author(s) Study design Subjects Treatment Findings Comment
Thomson and
Oswald
(1977a,b)
Prospective, placebo-con-
trolled, double-blind
34 perimeno-
pausal women
Piperazine oestrone sul-
phate
Oral 8 weeks
Decrease of wakeful-
ness and awakenings
Increase of REM
sleep
Oestrogen effect on
hot flashes, mood or
anxiety similar to
placebo
Schiff et al.
(1979)
Prospective, placebo-
controlled, double-blind,
crossover
16 hypogonadal
women
Conjugated equine oestro-
gen 0.625mg/day
Oral 100 days
Shorter sleep latency
Increase of REM
sleep
Decrease of serum
FSH and vasomotor
symptoms
Erlik et al. (1981) Case-control 4 postmeno-
pausal women
Ethinyl oestradiol 50 lg· 4/day
Decrease of
awakenings
Decrease of hot
flashes
Oral 30 days No placebo group
Pickett et al.
(1989)
Prospective, randomized,
placebo-controlled,
crossover
9 postmeno-
pausal women
Conjugated equine
oestrogen 1.25m g/day+
medroxyprogesterone
acetate 20 mg/day (MPA)
No improvement in
polysomnographic
parameters
No measurements for
symptoms or serum
hormone
concentrations
Oral 7 days
Purdie et al.
(1995)
Prospective, randomized,
placebo-controlled, single-
blind
33 postmeno-
pausal women
Conjugated equine
oestrogen 0.625 mg/day+
norgestrel 0.15 mg/day
(days 17–28)
No improvement in
polysomnographic
parameters
Decrease of meno-
pausal symptoms and
improvement of psy-
chological well being
Oral 12 weeks
Scharf et al.
(1997)
Prospective, placebo-con-
trolled, single-blind
7 postmeno-
pausal women
Conjugated equine
Oestrogen 0.625 mg/day
Oral 4 weeks
Improvement of sleep
efficiency, reduction
of cyclic alternating
patterns of sleep and
awakenings
Decrease of hot
flashes
Polo-Kantola
et al. (1999a,b)
Prospective, placebo-con-
trolled, double-blind,
crossover
62 postmeno-
pausal women
Oestradiol 50 lg/24 h
patches or gel 2.5 g/day
Transdermal 7 months
Decrease of
movement arousals
Decrease of serum
FSH and vasomotor
symptoms
Antonijevic et al.
(2000)
Prospective, crossover 11 postmeno-
pausal women
Oestradiol 50 lg/24 h
patches
Increase of REM
sleep
Decrease of serum
FSH and LH levels
Transdermal 2 weeks Reduced time awake
during first two sleep
cycles
No placebo group
Montplaisir et al.
(2001)
Prospective, randomized,
two group-treatment study
21 postmeno-
pausal women
Conjugated equine
oestrogen 0.625 mg/day+
either medroxyprogester-
one acetate 5 mg/day
(MPA) or microniced
progesterone 200 mg/day
Oral 6 months
Improvement of sleep
efficiency and reduc-
tion of time spent
awake after sleep on-
set during oestrogen
+ micronized proges-
terone but not during
oestrogen + MPA
Decrease of
menopausal
symptoms and
improvement of
subjective sleep
quality during the
both treatments
No placebo group
62 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
et al., 1979), an improvement in sleep efficiency and a
reduction of the rate of cyclic alternating patterns of
sleep (Scharf et al., 1997) or a decrease in the number of
waking episodes (Erlik et al., 1981) have also been re-
ported. In the largest study, a decrease in movement
arousals was the only objective change during HRT
(Polo-Kantola et al., 1999a,b), whereas some studies
have reported no improvement at all in polysomno-graphic sleep variables (Pickett et al., 1989; Purdie et al.,
1995).
The variation in results may be due to differences in
the study population (peri-menopausal, natural or sur-
gically induced menopause, age) and methodology
(dose, form and duration of HRT) (Thomson and Os-
wald, 1977a,b; Schiff et al., 1979).
3.3.5. Treatment of menopausal insomnia
In women with climacteric vasomotor symptoms,
HRT can be considered as the primary therapy for in-
somnia. Climacteric women, whose insomnia is essen-
tially accompanied by mental symptoms, also benefit
from HRT (Polo-Kantola et al., 1998). There is some
evidence that HRT could also alleviate sleep complaints
in women with no vasomotor symptoms (Polo-Kantolaet al., 1998). No specific therapies for insomnia need to
be initiated during the menopausal transition before the
individual response to HRT has been evaluated after 3–
6 months of the treatment. However, in women over 60
years the vascular side-effects of HRT may exceed its
beneficial effects on sleep (Hays et al., 2003) and thus
starting HRT should be considered carefully. In women

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 63
with concerns or contraindications for HRT (previous
breast and endometrium cancer, tromboembolic event,
liver disease) other treatment options, including anti-
depressants (e.g., selective serotonin reuptake inhibitors,
SSRI), gabapentin, dietary isofavones and soy foods, aswell as relaxation therapies should be considered. Al-
though menopause often causes or worsens sleep dis-
turbances, it should be remembered that part of the
disturbances may just coincide with the menopausal
period rather than being of endocrinological origin.
Thus, symptoms and signs that would normally lead to a
full sleep evaluation in premenopausal women should
also be taken seriously in postmenopausal women andnot automatically be considered as a normal conse-
quence of the menopause.
4. Contribution of animal models: influence of age and the
oestrus cycle on sleep
Animal studies, mostly conducted with rodents, have
provided essential contributions to the understanding of
the neurobiological mechanisms underlying the age- and
hormone-related changes in sleep and circadian rhythms.
4.1. Age and sleep in animals
Age-related modifications in daily and circadian
sleep–wake patterns have been investigated foremost inmale rodents. Early studies reported a shorter period of
the circadian locomotor activity rhythm (Pittendrigh
and Daan, 1974) and sleep (Van Gool et al., 1987) in
ageing hamsters and rats. However, more recent studies
performed in hamsters showed that the circadian period
does not slow down systematically with age (Davis and
Viswanathan, 1998; Duffy et al., 1999a,b). In aged mice
no change or a lengthening of the circadian period wasfound (reviewed by Weinert, 2000). These inconsistent
findings could be related to species and strain variability
and to different illumination conditions. An amplitude
decrease both in the daily sleep–wakefulness rhythm as
well as in the circadian sleep rhythm was reported in
27- to 35-month old C57BL mice compared to 3- to 15-
month old mice (Welsh et al., 1986). In the rat, an age-
dependent reduction of the sleep–wake amplitude wasfound in animals maintained under a controlled light/
dark cycle, but not under constant conditions (Men-
delson and Bergmann, 1999a,b; Van Gool and Mirmi-
ran, 1983; Van Gool et al., 1987).
Changes in sleep architecture have been reported in
ageing rodents. Early studies described a decline of total
sleep time and a shortening of individual sleep episodes
in 22- to 30-month old rats compared to 4- to 8-monthold rats (Van Gool and Mirmiran, 1983; Zepelin et al.,
1972). Exposure to a higher light intensity led to a re-
versal of these age-related changes (Witting et al., 1993).
An age-dependent reduction of sleep was found also in
mice (Eleftheriou et al., 1975). Although this result was
based on 6-h sleep records performed at an unspecifiedtime of day, the results suggested that the age at which
these modifications appeared was strain-dependent. In
C57BL/6 mice the changes had already occurred at 6
months while in DBA mice they appeared only later, at
12–23 months. Sleep was more fragmented in 27- to 35-
month old C57BL mice compared to younger, 3- to 12-
month old mice (Welsh et al., 1986). In rats and mice
recorded under constant environmental conditionssubtle age-related sleep changes were indicated by the
larger sleep fragmentation in old animals, while the
overall amount of vigilance states did not vary with age
(Van Gool et al., 1987; Welsh et al., 1986; Mendelson
and Bergmann, 1999a).
Studies investigating the effects of age on the sleep
EEG in rodents are rare and the results showed some
inconsistencies. Thus, non-REM sleep EEG power inthe delta range was either reduced or unchanged in ap-
proximately 24-month old rats (Tani and Ishihara, 1988;
Van Gool and Mirmiran, 1983). Both studies are limited
by the small amount of non-REM sleep used for the
EEG analysis that consisted either in a single 10 min
period per animal or in only a few 60–90 s periods
preceding the onset of a REM sleep episode. Delta
power based on 24 h and expressed relative to total EEGpower in non-REM sleep showed no age-related changes
between 3-, 12- and 24-month old Fischer-344 rats
(Mendelson and Bergmann, 1999a,b). On the other
hand, 24-h delta EEG power computed by a software
system was drastically reduced in 20-month-old Fischer-
344 rats, while in young and old Sprague–Dawley rats it
remained similar, approximately in the range found in
the old Fischer-344 rats (Shiromani et al., 2000).The effect of age on the homeostatic regulation of
sleep in rodents was addressed recently. The sleep re-
bound following sleep deprivation was reduced in mid-
dle-aged and old rats compared to young adults
(Mendelson and Bergmann, 2000). The typical en-
hancement of delta EEG power in non-REM sleep after
sleep deprivation was still present in rats at the age of 12,
20 and 24 months, but the effect of age on this responseremains controversial (Mendelson and Bergmann,
1999a,b; Shiromani et al., 2000).
4.2. Potential neurobiological mechanisms
The neurobiological mechanisms underlying age-de-
pendent changes in sleep remain unknown. The modifi-
cation of the daily sleep–wake pattern might be related to
a deterioration of the circadian clock itself, suggested by
a reduction of the mean peak of the neuronal firing

64 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
within the SCN in aged rats (Satinoff et al., 1993).
Moreover, the reduction of the sleep–wake amplitude
may be related to a dysfunction of the hypocretin/orexin
system. Thus, a decrease in hypocretin receptor 2 mRNA
expression in the hippocampus, thalamus and brainstemwas found in 12- to 24-month old C57BL/6 mice (Terao
et al., 2002) and a decrease in both prepro-orexin mRNA
as well as in the concentrations of the orexin peptides was
found in the ageing (12–14 months) rats (Porkka-He-
iskanen et al., 2003). A recent study claimed that the
transgenetically induced over-expression of the human
mutant b-amyloid precursor protein might enhance the
age-related disturbances in the sleep–wake distributionof 20- to 26-month old mice (Huitron-Resendiz et al.,
2002). The effects of age on sleep, however, were not
assessed statistically. A deficiency in c-fos induction by
waking in the hypothalamus and cingulate cortex might
be involved in the smaller sleep rebound after sleep de-
privation in old rats (Basheer and Shiromani, 2001).
4.3. Sleep and ageing in female rats and mice
Sleep studies in ageing female rodents are scarce. In
an old population of female Long–Evans rats Li and
Satinoff (1995) distinguished between individuals whichstill displayed a circadian rhythm of body temperature
and a sleep pattern that was similar to the one obtained
in young rats, and individuals which had no temperature
rhythm or an unstable rhythm and a concomitant
damping in their daily sleep–wake amplitude. The in-
fluence of the oestrus cycle on these patterns was not
specifically addressed.
Female C57BL/6 mouseFemale rat
Days
Days20 8 20 88
1
5
10
15
20 8 20 881
5
10
15
(a)
(b)
Fig. 2. Individual running wheel activity plots in a 12 h light–12 h dark
in a female mouse of the strain C57BL/6 (a) and a female Sprague
Dawley rat (b). Records are double-plotted, i.e. the horizontal axis
represents 48 h and each 24-h interval is represented twice. The black
and white bars at the top indicate the timing of the LD cycle.
4.4. Sleep and the oestrus cycle in rodents
A few studies focusing on young adults examined the
influence of oestrus cyclicity on sleep regulation in ro-dents. The oestrous cycle is defined by the succession of
four stages: proestrus, oestrus, metoestrus and diestrus.
Each stage can be determined according to the following
criteria based on histology of vaginal smears (Michael,
1976): proestrus, nucleated epithelial cells predominant;
oestrus, only cornified cells; metoestrus, leukocytes and
cornified cells; diestrus, leukocytes predominant. In mice
as in the rat, ovulation is preceded by a sharp rise incirculating LH and FSH (Murr et al., 1973). In outbred
mice a major surge of plasma progesterone in the late
afternoon of proestrus and a second peak in metoestrus
following a pattern similar to the rat were found (Mi-
chael, 1976). The timing of the main peak of proges-
terone was shown to depend on the mouse strain
(DeLeon et al., 1990).
Young female rats are well known to exhibit regular4- or 5-day oestrus cycles. In mice the cycle-length is
more variable, between 4 and 9 days, and cycling is less
regular. Early studies in young adult mice suggested a
complex interaction between hereditary and environ-
mental factors in the regulation of the length of the
oestrus cycle (Barkley and Bradford, 1981; Champlin,1971; De Leon and Barkley, 1987). Comparisons of
three inbred strains, C57BL/6, DBA and C3H/He, sug-
gested that the frequency of regular oestrus cycles and
their duration are differentially genetically regulated
(Nelson et al., 1992a,b).
A recent study of inbred mice reported a minor oes-
trus-dependent modulation of sleep that depended on
the genetic background (Koehl et al., 2003). In C57BL/6mice, the proestrus night was accompanied by a decrease
in REM sleep only, while in C3H/He mice a reduction of
non-REM and REM sleep was observed during diestrus.
On the other hand, BALB/c mice showed no sleep
changes across the oestrous cycle. Although infradian
rhythms of body temperature and locomotor activity
had been suggested to reflect the ovarian cycle in out-
bred mice (Weinert, 1996), such a relationship could notbe detected in C57BL/6 and C3H/He mice by Koehl
et al. (2003). Another study showed that C57BL/6 mice
recorded continuously during several oestrus cycles are
often more active in the second half of the proestrus
night compared to metoestrus and diestrus (Fig. 2), but
the increase of motor activity could not be attributed
specifically and consistently to proestrus days (Kopp
et al., 2002). In contrast to mice, female rats show amarked and consistent increase in motor activity and a
concomitant reduction in sleep during the proestrus

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 65
night (Fig. 2; Colvin et al., 1968; Kent et al., 1991;
Kleinlogel, 1983; Schwierin et al., 1998; Yokoyama
et al., 1966). This period of intense activity associated
with increased receptivity and mating behaviour is fol-
lowed by an enhancement of sleep on the oestrus day(Yokoyama et al., 1966).
Only minor changes in sleep were induced by ovari-
ectomy in the rat. Non-REM sleep was not or only
slightly increased while REM sleep was consistently
enhanced (Fang and Fishbein, 1996; Li and Satinoff,
1996). An inhibitory action of gonadal steroids on sleep,
particularly on REM sleep has been proposed (Colvin
et al., 1968; Fang and Fishbein, 1996). Sleep EEGspectral analysis revealed that the sleep rebound occur-
ring in the oestrus light period after the spontaneous
partial sleep deprivation of the proestrus night in the rat
is accompanied by an enhancement of delta activity in
non-REM sleep (Schwierin et al., 1998). Interestingly,
the study showed that homeostatic sleep response to 6 h
sleep deprivation was similar in proestrus and in oestrus.
The relation between peripheral and central con-centrations of steroids across the oestrous cycle was
investigated in young adult mice from the HsdOla
strain (Corpechot et al., 1997). The levels of proges-
terone in the brain followed a clear daily pattern, with a
peak in the first half of the night and reaching maximal
values in metoestrus and oestrus, but no correlation
was found between brain and plasma progesterone
levels. However, there was a positive correlation be-tween the concentration of pregnenolone sulphate in
the brain and the oestrous-dependent pattern of plasma
progesterone. Interestingly, exogenous progesterone
and its metabolite allopregnanolone as well as preg-
nenolone are known to affect sleep and/or the sleep
EEG in the rat (Lancel et al., 1994, 1996, 1997). These
neuroactive steroids exert their influence upon an al-
losteric modulation of the GABA-A receptors (Lam-bert et al., 1995; Lancel et al., 1999). Complex
interactions between sleep regulation, oestrous-related
patterns of steroids, and the GABAergic system can be
expected in ageing females. Further investigations
should help to clarify the mechanisms involved in the
effects of HRT in postmenopausal women.
5. Sleep disorders in women
Sleep disorders are among the most frequent prob-
lems affecting people’s health. Insomnia, for example,
affects more than 10% of the general Western popula-
tion at any given time point (e.g., Hajak, 2001). Hy-
persomnia is less frequent, but still 1–2% of the
population suffers from obstructive sleep apnoea (OSA),which, in addition to causing marked excessive daytime
sleepiness, leads to serious pulmonary and cardiovas-
cular morbidity (Young and Peppard, 2000).
Due to the aforementioned specific features of female
physiology, it is not surprising to find prominent gender
differences in the frequency of some sleep disorders. The
most important ones will be described below.
5.1. Insomnia
Insomnia is a subjective and non-specific complaint
that is related to the quantity or quality of night sleep.
Insomniacs might complain about difficulties falling
asleep, maintaining sleep or of premature morning
awakenings resulting in a subjectively reduced total
sleep time. Moreover, even the hours these people sub-jectively sleep might be perceived as superficial and
fragile and closer to dozing than to consolidated deep
sleep. Complaints of poor sleep have a myriad of causes
that encompass almost every aspect of sleep medicine
and some of them occur about twice as frequently in
women than in men (Harris, 2003).
Unfortunately, the reasons for the strong prepon-
derance of women among insomniacs have not beensystematically studied and reviewed, and in the present
context we can only examine some very obvious un-
derlying circumstances. Among these are the endocrine
peculiarities related to the menstrual cycle and to age-
ing that have been described in detail earlier in this
review article. However, it is interesting to note here
that the direct influence of hormonal changes on sleep
in women might be less pertinent for understandinginsomnia in women than expected. For example, ho-
meostatic sleep regulatory mechanisms, as indexed by
the time course of EEG slow wave activity, are not
substantially affected by the menstrual cycle (Driver,
1996). Changes in subjectively perceived and objectively
measured night sleep are only significant in women with
additional complaints summarised as the PMDD. This
suggests that hormonal changes across the menstrualcycle per se have only a minor direct and specific im-
pact on sleep in women, but in a subgroup affect sleep
and physical and psychological well-being simulta-
neously. It seems, though, that disturbed premenstrual
sleep is part of a complex of complaints rather than
reflecting a primary problem of disturbed sleep initia-
tion and maintenance.
Sleep complaints are particularly frequent in peoplesuffering from major psychiatric disorders and in par-
ticular affective disorders. Because these are about twice
as frequent in women compared to men (Harris, 2003),
psychiatric morbidity is likely to contribute significantly
to the preponderance of females among insomniacs.
Interesting sleep EEG gender differences have been re-
ported in depressed subjects (reviewed by Manber and
Armitage, 1999). Whether these differences reflect clini-cally relevant gender differences in the neurobiological
basis of the interactions between sleep and mood is an
important, but still unanswered question.

66 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
The gender differences in the EEG that have been
reported in healthy people (e.g., Carrier et al., 2001)
suggest that there may be differences in basic sleep reg-
ulating mechanisms. It is interesting to speculate that
these could lead to an increased general vulnerability ofwomen’s sleep. This should be reflected in an increased
prevalence of ‘‘pure’’ insomnia (primary and/or psy-
chophysiological insomnia) that is not related to hor-
monal changes or psychiatric morbidity. However, to
our knowledge, this has not been clearly demonstrated.
5.2. Restless legs syndrome and periodic leg movements
The restless legs syndrome (RLS) is a clinical entity
characterised by a combination of unpleasant sensations
in the legs (much less frequently also in the arms) and a
strong urge to move (Wetter and Pollm€acher, 1997).
Typically, symptoms start in the evening or upon retiring
to bed and have a major negative impact on sleep onset.
Following sleep onset the symptoms might resume dur-
ing periods of intermittent wakefulness, thereby dis-turbing sleep maintenance. During sleep itself the vast
majority of subjects affected display periodic limb
movements (PLMDs), which might or might not be ac-
companied by arousals (Pollm€acher and Schulz, 1993).
Prevalence estimates for RLS range from 5% to 10%
and, overall, no consistent gender difference has been
reported. Most researchers discriminate between an idi-
opathic form with a strong genetic background and asecondary, symptomatic RLS. Among these secondary
forms is RLS during pregnancy and, consequently, a
significant number of pregnant women experience RLS
symptoms. Recent epidemiological data suggest a prev-
alence of about 25% with an incidence peak during the
second and third trimester (Lee et al., 2001). Idiopathic
RLS has been reported to worsen during pregnancy
(Winkelmann et al., 2000). The causes of the emergenceor worsening of RLS symptoms during pregnancy are
unknown. As various lines of evidence suggest a link
between RLS and anaemia it has been hypothesised that
signs of iron deficiency during pregnancy might correlate
with the appearance of RLS symptoms. However, nei-
ther haemoglobin nor iron or ferritin predicted the oc-
currence of RLS in pregnant women, whereas folate
levels appeared to be reduced. In the same study it wasshown that RLS symptoms might persist after delivery,
but appear to cease in the majority of women (Leger
et al., 2000). This area of women’s sleep disorders is
clearly under-investigated. The epidemiological database
is small and we know almost nothing about risk factors,
cause, and specific options for treatment.
5.3. Obstructive sleep apnoea syndrome
The most common and widely studied of the sleep-
disordered breathing syndromes, affecting about 2%
of middle-aged women and about 4% of middle-aged
men (Young, 1993), is obstructive sleep apnoea syn-
drome (OSAS). Hallmarks of risk for OSAS are
overweight with snoring and excessive daytime sleep-
iness. Because clinical reports in the 1970s and 1980ssuggested that OSAS primarily affected men, women
were excluded from general epidemiological studies
until 1993 (Young et al., 1993; Kapsimalis and Kry-
ger, 2002a,b). The early studies in clinical populations
reported male:female prevalence ratios for OSAS at
10:1, while more recent studies in general population
samples report prevalence between 2:1 and 4:1
(Young et al., 1993; Kripke et al., 1997; Bixler et al.,2001; Kapsimalis and Kryger, 2002a,b). If a variant
mainly characterised by heavy snoring but few ap-
noeas is included, this ratio even goes down to 1:1.5
(Mohsenin, 2001).
One reason for the long-standing underestimation of
the prevalence of OSAS in women might be a difference
in the symptom profile. In particular, patient’s self-re-
ports (which are critical for patient’s help-seeking be-haviour and doctor’s appreciation of the complaint)
suggest that loud snoring and gasping are much more
prevalent in male OSAS patients than in females (Red-
line et al., 1994).
Obesity is typically associated with OSAS. However,
there is compelling evidence that female OSAS patients
are much more obese than males (Kapsimalis and
Kryger, 2002a,b). Interestingly, two groups of femaleOSAS patients are particularly prone to obesity: pre-
menopausal women and postmenopausal women on
HRT. Given that obesity is a strong risk factor for
OSAS, these findings suggest that female sex hormones
might be protective at least in non-obese women. This
idea is further supported by epidemiological evidence
showing that the prevalence of OSAS in premenopau-
sal and postmenopausal women on HRT is similar(<1%) (Bixler et al., 2001) and much lower than in
postmenopausal women without HRT (2.7%) (Bixler
et al., 2001). Finally, pregnancy might be protective
against OSAS even in obese women (Maasilta et al.,
2001).
Furthermore, there seem to be slight differences in the
medical consequences of OSAS between males and fe-
males. Whereas females more often display depressionand chronic obstructive lung disease, men are more
frequently diagnosed with coronary artery disease.
It is beyond the scope of the present article to dis-
cuss in detail the reasons for the prominent gender
differences in the occurrence of OSAS and in the fea-
tures of this disease. Numerous contributing factors are
likely to play a role, ranging from gender differences in
fat distribution across upper airway anatomy to en-docrine aspects. Readers are referred to a recent com-
prehensive review on this issue (Kapsimalis and
Kryger, 2002a,b).

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 67
5.4. Narcolepsy
Narcolepsy is a rare sleep disorder (prevalence
<1:2000) characterised by excessive daytime sleepiness
and cataplexy, which is manifested as a sudden short-lasting loss of muscle tone triggered by emotions (Ove-
reem et al., 2001). Further symptoms include automatic
behaviour, sleep paralysis and hypnagogic hallucina-
tions. The key pathophysiological feature of narcolepsy
is a disturbance in the transitions between wakefulness,
REM and non-REM-sleep. The causes of narcolepsy are
multifactorial, including a genetic component associated
with a human leukocyte antigen (HLA-DQB1*0602)and unknown environmental factors. Possibly through
an autoimmune mechanism these factors trigger a loss
of orexin production in the lateral hypothalamus.
Orexins, also called hypocretins, are involved in the
regulation of arousal, sleep, and appetite. Orexin defi-
ciency occurring through a number of steps that are not
yet fully understood, leads to the symptoms of narco-
lepsy (Dalal et al., 2002).Studies on the epidemiology of narcolepsy consis-
tently report a male gender predominance. The ratio of
female to male patients is around 1:1.5. The reason for
this gender difference is unknown and it has been a long-
standing assumption that the rate of diagnosed female
patients is artificially low due to a lower tendency of
women to seek medical care. It has been argued that
different social settings, in particular with respect to jobrequirements, lead to an increased (relative) disability in
males and, as a consequence, to an increased liability to
seek medical help.
However, recent epidemiological evidence points in
an interesting different direction. Tafti and colleagues
have analysed functional polymorphisms of catechol-
O-methyl transferase in narcoleptic subjects, because
this enzyme is critically involved in the metabolism ofbiogenic amines and, hence, important for vigilance
regulation. Their findings indicate a complex gender/
genotype/drug response interaction in narcolepsy: fe-
male patients with this disorder, compared to males,
have a higher probability of carrying a polymorphism
associated with low COMT activity (suggesting higher
availability of biogenic amines). These women are less
severely affected than males and respond more favour-ably to treatment that enhances DA activity (for review
see Tafti and Dauvilliers, 2003). Although these very
novel findings deserve independent replication, they
provide the first data supporting the hypothesis that
gender-related genetic peculiarities are important de-
terminants of sleep disorders in women.
5.5. REM sleep behaviour disorder
REM sleep behaviour disorder (RSBD) is a para-
somnic sleep disturbance characterised by impaired
REM sleep-related active inhibition of muscle tone
(Schenck andMahowald, 2002). This inhibition prevents
major movements during REM sleep which could occur
as a behavioural correlate of dreaming. Hence, normal
individuals might display minor face or limb movementsduring REM sleep, but they neither sit up nor leave the
bed. RSBD patients, in contrast, show major, often vi-
olent movements while vividly dreaming. This kind of
behaviour is one of the major causes of sleep-related
injury to the sleeping subjects themselves or to their bed
partners. RSBD is rare and typically occurs during the
second half of life with a peak incidence in the 50s and
60s. There is a remarkable association with Parkinson’sdisease and other neurodegenerative disorders. Surpris-
ingly, RSBD occurs almost exclusively in men. The ratio
of female to male patients is about 1:9. The reasons for
this striking gender difference are not known.
6. Effects of sleep loss in women
Chronic sleep curtailment is now a common feature
in our society both for men and particularly for women.
Average sleep duration has decreased from 9 h in 1910
to 7.5 h (Broman et al., 1996; Webb and Agnew, 1975)
and 6.22 h (Jean-Louis et al., 2000). The 24/7 focus of
modern life results in shift work for about 20% of
workers. As a consequence, sleep duration is reduced to
5 h per work day in these workers (Bliwise, 1996). Theacute and chronic effects of this sleep reduction for
women are poorly documented.
6.1. Shift work
Shift work, particularly at night, is problematic for
the worker. It represents a serious risk factor for sleep
disorders such as insomnia and daytime sleepiness(Akerstedt, 1998, 2003). The frequency of sleep com-
plaints in this population is twofold higher than in the
general population. These problems are mainly due to a
disruption of the normal sleep/wake pattern but also
involve other factors such as age, gender, stress at work,
health problems and social and family factors.
Loss of sleep is a general complaint in shift workers
of both genders (H€arm€a et al., 1998). Objective sleepmeasures indicate that 1–4 h less sleep is obtained during
daytime sleep than during a normal night (Czeisler et al.,
1980). This loss of sleep tends to accumulate with suc-
cessive night shifts. As a consequence, there is an in-
crease in daytime sleepiness and a tendency to sleep
more and to take more naps on holidays and weekends
(Webb and Agnew, 1975).
High levels of sleepiness during night and morningshifts have been documented. Moreover, involuntary
sleep episodes at work have been shown in several
studies based on questionnaire data and on continuous

68 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
EEG recordings (Kecklund and Akerstedt, 1993; Kogi
and Ohta, 1975). According to Partinen and colleagues
(1984) sleep disturbances are more frequent among
manual workers than among physicians or managers.
It is now well established that sleep disturbances aremore frequent in women and in older individuals
(Akerstedt, 2002; Avidan, 2002; Feinsilver, 2003; Ford
and Cooper-Patrick, 2001). In a representative sample of
58,115 individuals Akerstedt and colleagues found more
sleep disturbances and fatigue in women (Akerstedt,
2002). The reasons for this increased frequency is not yet
clear. It is possible that family duties and/or hormonal
factors could play a role. Several studies have analysedthe effects of night shifts and early shifts in nurses (Barak
et al., 1996; Gold et al., 1992; Kogi and Ohta, 1975;
Wootten, 2000). A recent study of health professionals
by Ohayon et al. (2002) showed that working on a ro-
tating daytime shift induces significant sleep distur-
bances, more sick leave and more work-related accidents
compared to fixed daytime and night time work.
In addition to fatigue and sleep disturbances, shiftwork, particularly at night, may also increase the risk of
some forms of cancer in women. One proposed expla-
nation is that exposure to light at night suppresses
melatonin production which has potential oncostatic
action. According to Davis and colleagues the risk of
breast cancer is higher among women who frequently do
not sleep at night with an increased risk among subjects
working in the brightest places (Davis et al., 2001). Therisk was particularly high in women who had worked 30
or more years at night (Schernhammer et al., 2001).
Moreover, a recent study indicates that working a ro-
tating night shift at least three nights per month for 15
or more years increases the risk of colorectal cancer in
women (Schernhammer et al., 2003).
6.2. Sleep deprivation studies
Total sleep deprivation and its effects on performance
have focussed on studies in men (Barak et al., 1996;
Harrison et al., 2000). The symptoms include increased
tiredness, decrease in psychomotor performance and
vigilance, mood changes, memory deficits and during a
prolonged sleep deprivation, also hallucinations
(Drummond and Brown, 2001). Functions which requireintact frontal lobe activity (e.g., motivation and creative
thinking) are particularly sensitive to the effects of sleep
deprivation (for a review see Muzur, 2002). Several hy-
potheses have been advanced to explain performance
decrements after sleep deprivation, including reduction
in the capacity to process information, overall changes in
arousal and decreased capacity to sustain attention due
to involuntary micro sleeps. During these brief sleep ep-isodes the subjects are not able to react to stimuli re-
sulting in ‘‘response lapses’’. According to this ‘‘lapse
hypothesis’’ subjects are more affected by sleep loss in
tasks involving work-paced stimuli rather than self-paced
stimuli. Furthermore, it is performance speed rather than
performance accuracy that is compromised (Koslowsky
and Babkoff, 1992). In a sleep restriction study on 8
young women impaired performance was found in asimple reaction time test (Tilley and Wilkinson, 1984).
Long term sleep restriction studies for 7 (Dinges et al.,
1997) or 14 (Van Dongen et al., 2003) consecutive days
have been performed in men and women and indicate
that even moderate sleep restriction can seriously impair
waking behavioural functions in both genders. Gender
differences were not addressed in these studies.
Sleep restriction and sleep deprivation have also beenshown to have a negative impact on carbohydrate me-
tabolism (VanHelder et al., 1993; Gonzales-Ortiz et al.,
2000), endocrine function (Spiegel et al., 1999) and on
immunity (Dinges et al., 1995; Spiegel et al., 2002) in
healthy men. However, such findings remain to be
demonstrated in women.
One important issue related to sleep deprivation
concerns the recovery process. Typically, there is anincrease in SWS during the first recovery night after
total sleep deprivation in young and elderly men and
women (e.g., Reynolds et al., 1986). Furthermore, wo-
men showed a more dramatic increase in slow wave
activity after 40 h of total sleep deprivation than age
matched men indicating a greater response to sleep de-
privation in women (Armitage et al., 2001).
In depressed patients, total, partial and selective sleepdeprivation has consistently been shown to improve
mood in men and women independent of the diagnostic
subtype of depression (Leibenluft et al., 1992; Wu and
Bunney, 1990; Wehr, 1992; Parry et al., 2000). This
short-lasting improvement of mood is followed by a
relapse after a recovery sleep episode even if it is of short
duration (Holsboer-Trachsler and Seifritz, 2000). Be-
sides these acute laboratory studies, few data exist onthe consequences of long term sleep restriction on wo-
men’s health. A recent study in 71,617 women health
professionals aged 45–65 years in the USA that was
based on questionnaires, found after a ten year follow
up, an increased risk of cardiac disease in women
sleeping both less than or more than 8 h per night (Ayas
et al., 2003). Despite their epidemiological interest, such
large population data based only on questionnaires needto be interpreted with caution. Several confounding
factors that could in turn affect sleep duration and
health status must be taken into account.
7. Sociological insights
7.1. The social context of women’s sleep
The sociology of sleep offers an exciting new dimen-
sion in sleep research. Rather than focussing exclusively

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 69
on the physiology of sleep, sociologists contend that
sleep takes place within a social context which influences
the patterning of sleep throughout the life course. As a
mirror of everyday life, sleep reflects the roles and re-
sponsibilities, gender divisions, health patterns andtransitions which characterise people’s lives. In the con-
text of lifestyle factors, women are more likely to expe-
rience and admit to tiredness than men (Cox et al., 1987;
Brannen and Moss, 1988; Blaxter, 1990; Popay, 1992).
Distinct from sleepiness, this tiredness may be linked to
women’s roles and responsibilities as they try to balance
the demands of motherhood, work and relationships.
While studies by Aubert and White (1959a,b), Sch-wartz (1970), Taylor (1993), Williams and Bendelow
(1998) and Williams (2002) have established the theo-
retical underpinnings and research agenda for the soci-
ology of sleep, there has until now been a lack of
empirical studies which focus specifically on an exami-
nation of the relationship between social context and
women’s sleep. Hislop and Arber’s (2003a,b,c) empirical
research on sleep in women aged 40 and over addressesthis gap.
7.2. Women’s sleep at mid-life
Hislop and Arber (2003a,b,c) suggest that sleep has
become a disposable resource for women in mid-life; a
low order priority which competes unfavourably with
the demands of motherhood, career, marriage and car-ing for ageing parents. While women accept that sleep is
essential to good health, the reality of their lives dictates
the extent to which they can access adequate sleep to
meet their needs. Women’s sleep may be affected by
expectations, responsibilities and stress associated with
their roles as partners, mothers, daughters, and em-
ployees, as well as by life crises and transitions such as
divorce, menopause, retirement, illness and widowhood.In a society in which gendered divisions of labour
ensure that women ‘continue to bear primary responsi-
bility for the completion of housework and caring work’
(Pilcher, 2000), many women find that rather than being
a place of rest and recuperation, the bedroom becomes
an ‘invisible workplace’ in which they remain on duty
throughout the night available to provide the physical
and emotional support needed to ensure the well-beingof their family. Over time, the sense of commitment to
family may mask sleep disruption to such an extent that
poor sleep patterns become the norm; an expected,
though undesired, outcome of the reality of women’s
lives. As Popay (1992) states in her study of women’s
experiences of ill health, women may accept poor sleep
and daytime fatigue as a ‘normal’ feature of everyday
life and ‘part of the price to be paid for combining paidwork and domestic labour’. For women, a good night’s
sleep may be one which follows an expected pattern of
disruption reflecting the constraints imposed by their
social roles and responsibilities. As such, a woman’s
right to sleep free from interference from others (Wil-
liams and Bendelow, 1998), is effectively challenged by
the social parameters surrounding women’s sleep.
7.3. Women’s sleep in later life
A pattern of sleep disruption may accumulate over
the life course establishing a fertile environment for poor
sleep patterns in older women. Sleep disruption, with its
consequent impact on well-being and quality of life, is
the product of the interplay of social factors as much as
it is the outcome of the biological effects of ageing.From a sociological viewpoint, later life is character-
ised by a series of transitions and changes in identity and
status as women move from work to retirement, from an
active to more sedentary lifestyle, from caring for chil-
dren and elderly parents to caring for their partner, and
from the role of spouse to widow. As lifestyles in later life
are restructured, so too are sleep patterns. The transition
from work to retirement, for example, may be charac-terised by shifts in sleep–wake patterns that were once
structured by alarm clocks and work routines. Increased
napping during the day, lack of exercise and boredom
can have adverse effects on sleep at night. Hislop and
Arber (2003a,b,c) suggest that it is the changes associated
with life course transitions during this period, acting in
combination with the physiological effects of ageing on
sleep, which may explain the increased potential for sleepdisruption in later life.
For partnered women, the quality of sleep in later life
also reflects changes in their relationship. Women’s sleep
may be affected by their partner’s increased restlessness,
with his snoring, prostate problems, and the pain and
discomfort of poor health interfering with a women’s
ability to achieve a good night’s sleep. Increased caring
responsibilities for partners as their health declines maylead to further sleep disruption.
Women’s sleep in widowhood highlights the extent to
which partners continue to influence women’s sleep
patterns even in absentia. Beyond the initial period of
grieving, women may find it difficult to regain the sleep
patterns they enjoyed during their marriage. The feelings
of loss widows experience as they contemplate a future
of empty nights without the companionship of theirpartner are a constant reminder of the shift in identity
from wife to widow. The loss of their partner’s gendered
role as protector may further erode sleep in widowhood
as women adjust to sleeping in an empty house without
the sense of security once provided by their partner.
7.4. Women’s management of sleep disruption
Hislop and Arber’s research shows that women are
proactive in seeking ways to improve their sleep. For
most women, dealing with sleep problems involves

70 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
choosing from a range of self-help strategies, over-the-
counter remedies and alternative therapies. These
strategies, however, are also subject to social and cul-
tural constraints. While lone women faced with sleep
disruption during the night may get up, move aroundthe house, watch television or read, these options are not
always available to women with partners and/or chil-
dren. Out of commitment to their family, women often
choose to tolerate sleep disruption rather than disturb
the sleep of others in the house. While moving to an-
other room may seem like a logical solution to sleep
disruption caused by a restless or snoring partner, this is
not always a viable option. As Taylor (1993) suggested,the right to sleep privately may be linked to socio-
economic status, reflected in the availability of spare
rooms or beds. Moreover, cultural taboos associated
with the symbolic relationship between marriage and
sleeping in a double bed may prevent many women
adopting this strategy. With the double bed symbolising
the marital bond, women may be reluctant to challenge
the basis of their relationship or invite social stigma bymoving to another room, preferring to remain in an
unfavourable sleeping environment despite its adverse
effects on their sleep.
Hislop and Arber found that women continue to see
sleep disruption as an inevitable part of their roles as
mothers, wives, partners, workers, carers, and widows,
despite the efforts of the medical profession, pharma-
ceutical companies, and alternative therapists to en-courage women to regard sleep as a health issue. While
women often try to improve their diet and exercise as
part of a healthy lifestyle programme, sleep continues to
be considered an expendable luxury. It is only when
sleep disruption extends beyond ‘acceptable normality’
and seriously impacts on the enactment of roles and
responsibilities that women seek help from their doctor.
While mid-life and older women continue to accept poorsleep as a product of their social reality, the ‘true’ extent
of sleep disruption in the community may go unnoticed.
8. Summary
While many aspects of sleep and sleep disorders share
similarities across gender, there are important differ-ences which need to be recognised and acknowledged by
clinicians, general practitioners and researchers. The
quality and amount of sleep women obtain is not only
influenced by gender differences in physiology but also
by a number of sociological factors linked to the role a
woman has in society and in the family. There are cur-
rently few research studies which focus on women. Al-
most all we know about sleep and sleep debt is based onexperiments on male subjects.
While survey studies clearly point out changes in
women’s sleep quality in association with hormonal
levels during the menstrual cycle, pregnancy and men-
opause, studies specifically addressing the relationship
between hormonal levels and sleep are rare. Moreover,
although common causes of disturbed sleep such as shift
work, stress, excessive use of caffeine or alcohol, pain, anoisy environment and inappropriate temperature or
lighting in the bedroom affect both men and women, the
consequences of chronic sleep loss remain unclear. We
do not know whether women are more or less vulnerable
to the effects of sleep loss than men, and/or whether age
modulates the effects of sleep loss differently across
gender. Many women appear to accept sleep disruption
or loss as part of their natural everyday life: taking careof an infant during the first year(s) of life usually in-
volves frequent night-time awakenings, leading to
chronic sleep debt in the mother. A snoring partner, the
social habits of a teenager that stays out late, and care-
taking of parents needing night-time assistance in later
life are common circumstances in women’s lives. Nev-
ertheless, virtually no attention has been paid to the
consequences of this ‘‘natural’’ sleep loss in society. Atired mother, for example, may have less patience with
her child; a tired commuter is at greater risk in traffic
and at the work place.
Finding ways to improve sleep quality in women in-
volves addressing a range of issues. Should women use
HRT to alleviate sleep disturbances? Would hormone
treatment help alleviate sleep disturbances appearing
during specific phases of the menstrual cycle? Whatremedies and strategies do women currently use to cope
with ‘‘natural’’ and inevitable sleep loss experienced as
part of everyday life? Is it in fact possible to overcome
the disturbances which contribute to this loss of sleep?
What information should women receive and how
should this information be presented?
New methods of imaging, such as proton spectros-
copy, the fast development of molecular biology, andthe application of sociological research methods to sleep
research will open new avenues for future research on
sleep. Multidisciplinary research, which incorporates
basic research conducted both on humans and animals,
clinical research and sociological research is needed to
assess the complicated relationship between different
factors in sleep regulation. This will allow the develop-
ment of strategies which can help women fight thenegative consequences of sleep loss and lead to an
overall improvement of their health.
References
Arendt J. Melatonin and the mammalian pineal gland. London:
Chapman & Hall; 1995.
Akerstedt T. Sleepiness as a consequence of shift work. Sleep 1988;11:
17–34.
Akerstedt T. Shift work and disturbed sleep/wakefulness. Occupa-
tional Medicine (Oxford) 2003;53:89–94.

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 71
Akerstedt T, Fredlund P, Gillberg M, Jansson B. Work load and work
hours in relation to disturbed sleep and fatigue in a large
representative sample. Journal of Psychosomatic Research 2002;
53:585–8.
Akerstedt T, Knutsson A, Westerholm P, Theorell T, Alfredsson L,
Kecklund G. Sleep disturbances, work stress and work hours: a
cross-sectional study. Journal of Psychosomatic Research 2002;53:
741–8.
Andersch B, Hahn L. Premenstrual complaints. II. Influence of oral
contraceptives. Acta Obstetricia et Gynecologica Scandinavica
1981;60:579–83.
Anderson WM, Falestiny M. Women and sleep. Primary Care Update
of Obstetric and Gynecology 2000;7:131–7.
Antonijevic IA, Stalla GK, Steiger A. Modulation of the sleep
electroencephalogram by estrogen replacement in postmenopausal
women. American Journal of Obstetrics and Gynecology 2000;182:
277–82.
Armitage R, Smith C, Thompson S, Hoffmann R. Sex difference in
slow wave activity in response to sleep deprivation. Sleep Research
Online 2001;4:33–41.
Aubert V, White H. (I) Sleep: a sociological interpretation 1. Acta
Sociologica 1959;4:46–54.
Aubert V, White H. (II) Sleep: a sociological interpretation 11. Acta
Sociologica 1959;4:1–16.
Avidan AY. Sleep changes and disorders in the elderly patient. Current
Neurology and Neuroscience Reports 2002;2:178–85.
Ayas NT, White DP, Manson JE, Stampfer MJ, Speizer FE, Malhotra
A, Hu FB. A prospective study of sleep duration and coronary
heart disease in women. Archives of Internal Medicine 2003;163:
205–9.
Baehr E, Revelle W, Eastman C. Individual differences in morning-
ness-eveningness and the phase and amplitude of the circadian
temperature rhythm. Sleep 1999;22:S164.
Baker FC, Driver HS, Rogers GG, Paiker J, Mitchell D. High
nocturnal body temperatures and disturbed sleep in women with
primary dysmenorrhea. American Journal of Physiology 1999;277:
E1013–21.
Baker FC, Waner JI, Vieira EF, Taylor SR, Driver HS, Mitchell D.
Sleep and 24 hour body temperatures: a comparison in young men,
naturally cycling women and women taking hormonal contracep-
tives. Journal of Physiology 2001;530:565–74.
Barak Y, Achiron A, Kimh R, Lampl Y, Gilad R, Elizur A, Sarova-
Pinhas I. Health risks among shift workers: a survey of female
nurses. Health Care for Women International 1996;17:527–33.
Baratte-Beebe KR, Lee K. Sources of midsleep awakenings in
childbearing women. Clinical Nursing Research 1999;8(4):386–97.
Barkley MS, Bradford GE. Oestrus cycle dynamics in different strains
of mice. Proceedings of the Society for Experimental Biology and
Medicine 1981;167:70–7.
Basheer R, Shiromani PJ. Effects of prolonged wakefulness on c-fos
and AP1 activity in young and old rats. Brain Research and
Molecular Brain Research 2001;89:153–7.
Baulieu EE. Neurosteroids: a novel function of the brain. Psychoneu-
roendocrinology 1998;23:963–87.
Benca RM, Obermeyer WH, Thisted RA, Gillin JC. Sleep and
psychiatric disorders. A meta-analysis. Arch Gen Psychiatry 1992;
49:651–68.
Berga SL, Yen SS. Circadian pattern of plasma melatonin concentra-
tions during four phases of the human menstrual cycle. Neuroen-
docrinology 1990;51:606–12.
Bixler EO, Vgontzas AN, Lin HM, Ten Have T, Rein J, Vela-Bueno
A, Kales A. Prevalence of sleep-disordered breathing in women:
effects of gender. American Journal of Respiratory and Critical
Care Medicine 2001;163:608–13.
Blaxter M. Health and lifestyles. London: Routledge; 1990.
Bliwise DL. Historical change in the report of daytime fatigue. Sleep
1996;19:462–4.
Blyton DM, Sullivan CE, Edwards N. Lactation is associated with an
increase in slow-wave sleep in women. Journal of Sleep Research
2002;11:297–303.
Boivin DB, Duffy JF, Kronauer RE, Czeisler CA. Dose-response
relationships for resetting of human circadian clock by light.
Nature 1996;379:540–2.
Bojkowski CJ, Arendt J. Factors influencing urinary 6-sulphatoxyme-
latonin, a major melatonin metabolite, in normal human subjects.
Clinical Endocrinology (Oxford) 1990;33:435–44.
Borb�ely AA. Sleep regulation. Introduction. Human Neurobiology
1982;1:161–2.
Borb�ely AA. A two process model of sleep regulation. Human
Neurobiology 1982;1:195–204.
Brannen J, Moss P. Mothers and daughters: a three generational study
of health attitudes and behaviour. London: Heinemann; 1988.
Brincat M, Studd JW. Menopause-a multi system disease. Baillieres
Clinical Obstetrics and Gynaecology 1988;2:289–316.
Broman JE, Lundh LG, Hetta J. Insufficient sleep in the general
population. Neurophysiologie Clinique 1996;26:30–9.
Brunner DP, Munch M, Biedermann K, Huch R, Huch A, Borb�elyAA. Changes in sleep and sleep electroencephalogram during
pregnancy. Sleep 1994;17:576–82.
Brzezinski A, Lynch HJ, Seibel MM, Deng MH, Nader TM, Wurtman
RJ. The circadian rhythm of plasma melatonin during the normal
menstrual cycle and in amenorrheic women. Journal of Clinical
Endocrinology and Metabolism 1988;66:891–5.
Buysse DJ, Reynolds 3rd CF, Monk TH, Hoch CC, Yeager AL,
Kupfer DJ. Quantification of subjective sleep quality in healthy
elderly men and women using the Pittsburgh Sleep Quality index
(PSQI. Sleep 1991;14:331–8.
Campbell SS, Gillin JC, Kripke DF, Erikson P, Clopton P. Gender
differences in the circadian temperature rhythms of healthy elderly
subjects: relationships to sleep quality. Sleep 1989;12:529–36.
Campbell SS, Kripke DF, Gillin JC, Hrubovcak JC. Exposure to light
in healthy elderly subjects and Alzheimer’s patients. Physiology
and Behavior 1988;42:141–4.
Carrier J, Land S, Buysse DJ, Kupfer DJ, Monk TH. The effects of age
and gender on sleep EEG power spectral density in the middle
years of life (ages 20–60 years old). Psychophysiology 2001;38:232–
42.
Carrier J, Monk TH, Buysse DJ, Kupfer DJ. Amplitude reduction of
the circadian temperature and sleep rhythms in the elderly.
Chronobiology International 1996;13:373–86.
Carrier J, Paquet J, Morettini J, Touchette E. Phase advance of sleep
and temperature circadian rhythms in the middle years of life in
humans. Neuroscience Letters 2002;320:1–4.
Carskadon MA, Brown ED, Dement WC. Sleep fragmentation in the
elderly: relationship to daytime sleep tendency. Neurobiology of
Aging 1982;3:321–7.
Case AM, Reid RL. Effects of the menstrual cycle on medical
disorders. Archives of International Medicine 1998;158:1405–
12.
Chabbert BN, Djakoure C, Maitre SC, Bouchard P. Regulation of the
human menstrual cycle. Front Neuroendocrinology 1998;19:151–
86.
Champlin AK. Suppression of oestrus in grouped mice: the effects of
various densities and the possible nature of the stimulus. Journal of
Reproduction and Fertility 1971;27:233–41.
Chrousos GP, Torpy DJ, Gold PW. Interactions between the
hypothalamic-pituitary-adrenal axis and the female reproductive
system: clinical implications. Annals of Internal Medicine 1998;
129:229–40.
Cohen LS, Soares CN, Otto MW, Sweeney BH, Liberman RF, Harlow
BL. Prevalence and predictors of premenstrual dysphoric disorder
(PMDD) in older premenopausal women. The Harvard Study of
Moods and Cycles. Journal of Affective Disorders 2002;70:125–
32.

72 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
Colvin GB, Whitmoyer DI, Lisk RD, Walter DO, Sawyer CH.
Changes in sleep–wakefulness in female rats during circadian and
oestrus cycles. Brain Research 1968;7:173–81.
Corpechot C, Collins BE, Carey MP, Tsouros A, Robel P, Fry JP.
Brain neurosteroids during the mouse oestrous cycle. Brain
Research 1997;766:276–80.
Cox B, Blaxter M, Buckle A. The health and lifestyle survey:
preliminary report. London: The Health Promotion Research
Trust; 1987.
Czeisler CA, Kronauer RE, Allan JS, Duffy JF, Jewett ME, Brown
EN, Ronda JM. Bright light induction of strong (type 0)
resetting of the human circadian pacemaker. Science 1989;244:
1328–33.
Czeisler CA, Weitzman E, Moore-Ede MC, Zimmerman JC, Knauer
RS. Human sleep: its duration and organization depend on its
circadian phase. Science 1980;210:1264–7.
Daan S, Beersma DG, Borb�ely AA. Timing of human sleep: recovery
process gated by a circadian pacemaker. American Journal of
Physiology 1984;246:R161–83.
Dalal M, Schuld A, Beitinger P, Bubendorff V, Pollm€acher T.
Neuroendocrine and metabolic aspects of narcolepsy. Somnologie
2002;6:95–100.
Darling MR, Hawkins DF. Sex hormones in pregnancy. Clinics in
Obstetrics and Gynaecology 1981;8(2):405–19.
Davis FC, Viswanathan N. Stability of circadian timing with age in
Syrian hamsters. American Journal of Physiology 1998;44:R960–8.
Davis S, Mirick DK, Stevens RG. Night shift work, light at night, and
risk of breast cancer. Journal of National Cancer Institute 2001;93:
1557–62.
De Leon DD, Barkley MS. Male and female genotype mediate
pheromonal regulation of the mouse oestrus cycle. Biology of
Reproduction 1987;37:1066–74.
DeLeon DD, Zelinski-Wooten MB, Barkley MS. Hormonal basis of
variation in oestrous cyclicity in selected strains of mice. Journal of
Reproduction and Fertility 1990;89:117–26.
Dijk DJ, Duffy JF, Czeisler CA. Contribution of circadian physiology
and sleep homeostasis to age-related changes in human sleep.
Chronobiol 2000;17:285–311.
Dijk DJ, Duffy JF, Riel E, Shanahan TL, Czeisler CA. Ageing and the
circadian and homeostatic regulation of human sleep during forced
desynchrony of rest, melatonin and temperature rhythms. Journal
of Physiology 1999;516:611–27.
Dinges DF, Douglas SD, Hamarman S, Zaugg L, Kapoor S. Sleep
deprivation and human immune function. Advances in Neuroim-
munology 1995;5:97–110.
Dinges DF, Pack F, Williams K, Gillen KA, Powell JW, Ott GE,
Aptowicz C, Pack AI. Cumulative sleepiness, mood disturbance,
and psychomotor vigilance performance decrements during a week
of sleep restricted to 4–5 h per night. Sleep 1997;20:267.
Driver HS. Sleep in women. Journal of Psychosomatic Research
1996;40(3):227–30.
Driver HS, Dijk DJ, Werth E, Biedermann K, Borb�ely AA. Sleep and
the sleep electroencephalogram across the menstrual cycle in young
healthy women. Journal of Clinical Endocrinology and Metabo-
lism 1996;81(2):728–35.
Driver HS, Shapiro CM. A longitudinal study of sleep stages in young
women during pregnancy and postpartum. Sleep 1992;15:449–53.
Drummond SP, Brown GG. The effects of total sleep deprivation on
cerebral responses to cognitive performance. Neuropsychophar-
macology 2001;25:S68–73.
Duffy JF, Czeisler CA. Age-related change in the relationship between
circadian period, circadian phase, and diurnal preference in
humans. Neuroscience Letters 2002;318:117–20.
Duffy JF, Dijk DJ, Hall EF, Czeisler CA. Relationship of endogenous
circadian melatonin and temperature rhythms to self-reported
preference for morning or evening activity in young and older
people. Journal of Investigative Medicine 1999;47:141–50.
Duffy JF, Dijk DJ, Klerman EB, Czeisler CA. Later endogenous
circadian temperature nadir relative to an earlier wake time in older
people. American Journal of Physiology 1998;275:R1478–87.
Duffy JF, Viswanathan N, Davis FC. Free-running circadian period
does not shorten with age in female Syrian hamsters. Neuroscience
Letters 1999;271:77–80.
Ehlers CL, Kupfer DJ. Slow-wave sleep: do young adult men and
women age differently. Journal of Sleep Research 1997;6:211–5.
Eleftheriou BE, Zolovick AJ, Elias MF. Electroencephalographic
changes with age in male mice. Gerontologia 1975;21:21–30.
Empson JA, Purdie DW. Effects of sex steroids on sleep. Annals of
Medicine 1999;31:141–5.
Eriksson E, Lisjo P, Sundblad C, Andersson K, Andersch B, Modigh
K. Effect of clomipramine on premenstrual syndrome. Acta
Psychiatrica Scandinavica 1990;81:87–8.
Erkkola R, Holma P, Jarvi T, Nummi S, Punnonen R, Raudaskoski T,
Rehn K, RyynanenM, Sipila P, Tunkelo E. Transdermal oestrogen
replacement therapy in a Finnish population. Maturitas 1991;13:
275–81.
Erlik Y, Tataryn IV, Meldrum DR, Lomax P, Bajorek JG, Judd HL.
Association of waking episodes with menopausal hot flushes.
JAMA 1981;245:1741–4.
Fang J, Fishbein W. Sex differences in paradoxical sleep: influences of
oestrus cycle and ovariectomy. Brain Research 1996;734:275–85.
Feinberg I. Changes in sleep cycle patterns with age. Journal of
Psychiatric Research 1974;10:283–306.
Feinsilver SH. Sleep in the elderly. What is normal? Clinics in Geriatric
Medicine 2003;19:177–88.
Fink G, Sumner BE, McQueen JK, Wilson H, Rosie R. Sex steroid
control of mood, mental state and memory. Clinical and Exper-
imental Pharmacology and Physiology 1998;25:764–75.
Ferrari E, Arcaini A, Gornati R, Pelanconi L, Cravello L, Fioravanti
M, Solerte SB, Magri F. Pineal and pituitary-adrenocortical
function in physiological aging and in senile dementia. Experimen-
tal Gerontology 2000;35:1239–50.
Ford DE, Cooper-Patrick L. Sleep disturbances and mood disorders:
an epidemiologic perspective. Depression Anxiety 2001;14:3–6.
Fourtillan JB, Brisson AM, Fourtillan M, Ingrand I, Decourt JP,
Girault J. Melatonin secretion occurs at a constant rate in both
young and older men and women. Am J Physiol Endocrinol Metab
2001;280:E11–22.
Friess E, Tagaya H, Trachsel L, Holsboer F, Rupprecht R. Proges-
terone-induced changes in sleep in male subjects. American Journal
of Physiology 1997;272:E885–91.
Goebelsmann U. Protein and steroid hormones in pregnancy. Journal
of Reproductive Medicine 1979;23(4):166–77.
Gold DR, Rogacz S, Bock N, Tosteson TD, Baum TM, Speizer FE,
Czeisler CA. Rotating shift work, sleep, and accidents related to
sleepiness in hospital nurses. American Journal of Public Health
1992;82:1011–4.
Gonzales-Ortiz M, Martinez-Abundis E, Balcazar-Munoz BR, Pascoe-
Gonzales S. Effect of sleep deprivation on insulin sensitivity and
cortisol concentration in healthy subjects. Diabetes, Nutrition and
Metabolism – Clinical and Experimental 2000;13:80–3.
Gotts G, Morse CA, Dennerstein L. Premenstrual complaints: an
idiosyncratic syndrome. Journal of Psychosomatic Obstetrics and
Gynecology 1995;16:29–35.
Hajak G. Epidemiology of severe insomnia and its consequences in
Germany. European Archives of Psychiatry and Clinical Neuro-
science 2001;251:49–56.
Halbreich U, Kahn LS. Selective oestrogen receptor modulators–
current and future brain and behaviour applications. Expert
Opinion on Pharmacotherapy 2000;1:1385–98.
H€arm€a M, Tenkanen L, Sj€oblom T, Alikoski T, Heinsalmi P.
Combined effects of shift work and life-style on the prevalence of
insomnia, sleep deprivation and daytime sleepiness. Scandinavian
Journal of Work, Environment and Health 1998;24:300–7.

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 73
Harris T. Depression in women and its sequelae. Journal of Psycho-
somatic Research 2003;54:103–12.
Harrison Y, Horne JA, Rothwell A. Prefrontal neuropsychological
effects of sleep deprivation in young adults-a model for healthy
aging. Sleep 2000;23:1067–73.
Hays J, Ockene JK, Brunner RL, Kotchen JM, Manson JE, Patterson
RE, Aragaki AK, Shumaker SA, Brzyski RG, LaCroix AZ,
Granek IA, Valanis BG. Effects of estrogen plus progestin on
health-related quality of life. NEJM 2003;348:1839–54.
Hedmann C, Pohjasvaara T, Tolonen U, Suhonen-Malm AS, Myllyl€a
VV. Effects of pregnancy on mothers’sleep. Sleep Medicine
2002;3:37–42.
Hendrick V, Altshuler LL, Burt VK. Course of psychiatric disorders
across the menstrual cycle. Harvard Review of Psychiatry 1996;4:
200–7.
Hertz G, Fast A, Feinsilver SH, Albertario CL, Schulman H,
Fein AM. Sleep in normal late pregnancy. Sleep 1992;15(3):
246–51.
Hislop J, Arber S. (I) Sleepers wake! the gendered nature of sleep
disruption among mid-life women. Sociology 2003;37(4):695–711.
Hislop J, Arber S. (II) Understanding women’s sleep management:
beyond medicalisation-healthisation. Sociology of Health and
Illness 2003;26(6):815–37.
Hislop J, Arber S. (III) Sleep as a social act: A window on gender roles
and relationships. In: Arber S, Davidson K, Ginn J, editors.
Gender and ageing: changing roles and relationships. Maidenhead:
McGraw Hill/Open University Press; 2003. p. 186–205.
Holsboer-Trachsler E, Seifritz E. Sleep in depression and sleep
deprivation: a brief conceptual review. World Journal of Biological
Psychiatry 2000;1:180–6.
Holte A, Mikkelsen A. The menopausal syndrome: a factor analytic
replication. Maturitas 1991;13:193–203.
Honma K, Honma S. Circabidian rhythm: its appearance and
disappearance in association with a bright light pulse. Experientia
1988;44:981–3.
Hublin C, Kaprio J, Partinen M, Heikkil€a K, Koskenvuo M. Daytime
sleepiness in an adult, Finnish population. Journal of Internal
Medicine 1996;239:417–23.
Huitron-Resendiz S, Sanchez-Alavez M, Gallegos R, Berg G, Craw-
ford E, Giacchino JL, Games D, Henriksen SJ, Criado JR. Age-
independent and age-related deficits in visuospatial learning, sleep–
wake states, thermoregulation and motor activity in PDAPP mice.
Brain Research 2002;928:126–37.
Hurt SW, Schnurr PP, Severino SK, Freeman EW, Gise LH, Rivera-
Tovar A, Steege JF. Late luteal phase dysphoric disorder in 670
women evaluated for premenstrual complaints. American Journal
of Psychiatry 1992;149:525–30.
Hutchison JB, Beyer C. Gender-specific brain formation of estrogen in
behavioural development. Psychoneuroendocrinology 1994;19:
529–41.
Iguichi H, Kato KI, Ibayashi H. Age-dependent reduction in serum
melatonin concentrations in healthy human subjects. Journal of
Clinical Endocrinology and Metabolism 1982;55:27–9.
Ishizuka Y, Pollak CP, Shirakawa S, Kakuma T, Azumi K, Usui A,
Shiraishi K, Fukuzawa H, Kariya T. Sleep spindle frequency
changes during the menstrual cycle. Journal of Sleep Research
1994;3:26–9.
Jaffe RB, Plosker S, Marshall L, Martin MC. Neuromodulatory
regulation of gonadotropin-releasing hormone pulsatile discharge
in women. American Journal of Obstetrics and Gynecology 1990;
163:1727–31, 1990.
Jean-Louis G, Kripke DF, Ancoli-Israel S, Klauber MR, Sepulveda
RS. Sleep duration, illumination and activity patterns in a
population sample: effects of gender and ethnicity. Biology of
Psychiatry 2000;47:921–7.
Kapsimalis F, Kryger MH. (I) Gender and obstructive sleep apnea
syndrome, part 1: Clinical features. Sleep 2002;25:412–9.
Kapsimalis F, Kryger MH. (II) Gender and obstructive sleep apnea
syndrome, part 2: mechanisms. Sleep 2002;25:499–506.
Kattapong K, Fogg L, Eastman C. Effects of gender, menstrual cycle
phase and hormonal contraceptive use on circadian body temper-
ature rhythms. Chronobiology International 1995;12:257–66.
Kecklund G, Akerstedt T. Sleepiness in long distance truck driving: an
ambulatory EEG study of night driving. Ergonomics 1993;36:
1007–17.
Kennaway DJ, Lushington K, Dawson D, Lack L, van den Heuvel C,
Rogers N. Urinary 6-sulfatoxymelatonin excretion and aging: new
results and a critical review of the literature. Journal of Pineal
Research 1999;27:210–20.
Kent S, Hurd M, Satinoff E. Interactions between body temperature
and wheel running over the estrous cycle in rats. Physiology and
Behavior 1991;49:1079–84.
Khalsa SB, Jewett ME, Cajochen C, Czeisler CA. A phase response
curve to single bright light pulses in human subjects. Journal of
Physiology 2003;549:945–52.
Kleinlogel H. The female rat’s sleep during oestrous cycle. Neuropsy-
chobiology 1983;10:228–37.
Koehl M, Battle SE, Turek FW. Sleep in female mice: a strain
comparison across the oestrus cycle. Sleep 2003;26:267–72.
Kogi K, Ohta T. Incidence of near accidental drowsing in locomotive
driving during a period of rotation. Journal of Human Ergology
1975;4:65–76.
Kopp C, Ressel V, Tobler I. Oestrus cycle and motor activity in inbred
mouse strains. Journal of Sleep Research 2002;11:126.
Koslowsky M, Babkoff H. Meta-analysis of the relationship between
total sleep deprivation and performance. Chronobiology Interna-
tional 1992;9:132–6.
Kripke DF, Ancoli-Israel S, Klauber MR, Wingard DL, Mason WJ,
Mullaney DJ. Prevalence of sleep-disordered breathing in ages 40–
64 years: a population-based survey. Sleep 1997;20(1):65–76.
Kruijver FP, Swaab DF. Sex hormone receptors are present in the
human suprachiasmatic nucleus. Neuroendocrinology 2002;75:
296–305.
Lambert JJ, Belelli D, Hill-Venning C, Peters JA. Neurosteroids and
GABAA receptor function. Trends in Pharmacology Science
1995;16:295–303.
Lancel M, Cronlein TA, Muller-Preuss P, Holsboer F. Pregnenolone
enhances EEG delta activity during non-rapid eye movement sleep
in the rat, in contrast to midazolam. Brain Research 1994;646:85–
94.
Lancel M, Faulhaber J, Holsboer F, Rupprecht R. Progesterone
induces changes in sleep comparable to those of agonistic GABAA
receptor modulators. American Journal of Physiology
1996;271:E763–72.
Lancel M, Faulhaber J, Holsboer F, Rupprecht R. The GABA (A)
receptor antagonist picrotoxin attenuates most sleep changes
induced by progesterone. Psychopharmacology 1999;141:
213–9.
Lancel M, Faulhaber J, Schiffelholz T, Romeo E, Di Michele F,
Holsboer F, Rupprecht R. Allopregnanolone affects sleep in a
benzodiazepine-like fashion. Journal of Pharmacology and Exper-
imental Therapeutics 1997;282:1213–8.
Lee KA. Circadian temperature rhythms in relation to menstrual cycle
phase. Journal of Biological Rhythms 1988;3:255–63.
Lee KA, DeJoseph JF. Sleep disturbances, vitality, and fatigue among
a select group of employed childbearing women. Birth
1992;19:208–13.
Lee KA, McEnany G, Zaffke ME. REM sleep and mood state in
childbearing women: sleepy or weepy. Sleep 2000;23:877–85.
Lee KA, Shaver JF, Giblin EC, Woods NF. Sleep patterns related to
menstrual cycle phase and premenstrual affective symptoms. Sleep
1990;13:403–9.
Lee KA, Zaffke ME, Baratte-Beebe K. Restless legs syndrome and
sleep disturbance during pregnancy: the role of folate and iron.

74 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
Journal of Womens Health and Gender-Based Medicine
2001;10:335–41.
Lee KA, Zaffke ME, McEnany G. Parity and sleep patterns during and
after pregnancy. Obstetrics and Gynecology 2000;95:14–8.
Leger D, Guilleminault C, Dreyfus JP, Delahaye C, Paillard M.
Prevalence of insomnia in a survey of 12,778 adults in France.
Journal of Sleep Research 2000;9:35–42.
Leibenluft E, Noonan BM, Wehr TA. Diurnal variation: reliability of
measurement and relationship to typical and atypical symptoms of
depression. Journal of Affect Disorders 1992;26:199–204.
Levine JE. Chappell LM. Bauer-Dantoin AC. Wolfe AM. Porkka-
Heiskanen T. Urban JH. Amplitude and frequency modulation of
pulsatile luteinizing hormone-releasing hormone release. Cellular
and Molecular Neurobiology 1995;15(1):117–39.
Li H, Satinoff E. Changes in circadian rhythms of body temperature
and sleep in old rats. American Journal of Physiology 1995;269:
R208–14.
Li H, Satinoff E. Body temperature and sleep in intact and ovariec-
tomized female rats. American Journal of Physiology 1996;271:
R1753–8.
Lindberg E, Janson C, Gislason T, Bjornsson E, Hetta J, Boman G.
Sleep disturbances in a young adult population: can gender
differences be explained by differences in psychological status.
Sleep 1997;20:381–7.
Lockley SW, Skene DJ, Arendt J, Tabandeh H, Bird AC, Defrance R.
Relationship between melatonin rhythms and visual loss in the
blind. Journal of Clinical Endocrinology and Metabolism 1997;82:
3763–70.
Luboshitzky R, Yanai D, Shen-Orr Z, Israeli E, Herer P, Lavie P.
Daily and seasonal variations in the concentration of melatonin in
the human pineal gland. Brain Research Bulletin 1998;47:
271–6.
Maasilta P, Bachour A, Teramo K, Polo O, Laitinen LA. Sleep-related
disordered breathing during pregnancy in obese women. Chest
2001;120:1448–54.
Manber R, Armitage R. Sex, steroids, and sleep: a review. Sleep 1999;
22:540–55.
McEwen BS. Actions of sex hormones on the brain: ‘organization’ and
‘activation’ in relation to functional teratology. Progress in Brain
Research 1988;73:121–34.
McEwen BS. Permanence of brain sex differences and structural
plasticity of the adult brain. Proceedings of the National Academy
of Sciences of the United States of America 1999;96:7128–30.
McEwen BS, Alves SE. Estrogen actions in the central nervous system.
Endocrine Reviews 1999;20(3):279–307.
McParland P, Pearce JM. Restless leg syndrome in pregnancy. BMJ
1988;297(6662):1543.
Mendelson WB, Bergmann BM. Age-related changes in sleep in the
rat. Sleep 1999;22:145–50.
Mendelson WB, Bergmann BM. EEG delta power during sleep in
young and old rats. Neurobiology Aging 1999;20:669–73.
Mendelson WB, Bergmann BM. Age-dependent changes in recovery
sleep after 48 h of sleep deprivation in rats. Neurobiology Aging
2000;21:689–93.
Michael SD. Plasma prolactin and progesterone during the oestrus
cycle in the mouse. Proceedings of the Society for Experimental
Biology and Medicine 1976;153:254–7.
Minors DS, Waterhouse JM, Wirz-Justice A. A human phase-response
curve to light. Neuroscience Letters 1991;133:36–40.
Mishima K, Okawa M, Shimizu T, Hishikawa Y. Diminished
melatonin secretion in the elderly caused by insufficient environ-
mental illumination. Journal of Clinical Endocrinology and Me-
tabolism 2001;86:129–34.
Moe KE, Prinz PN, Vitiello MV, Marks AL, Larsen LH. Healthy
elderly women and men have different entrained circadian temper-
ature rhythms. Journal of the American Geriatrics Society 1991;39:
383–7.
Mohsenin V. Gender differences in the expression of sleep-disordered
breathing : role of upper airway dimensions. Chest 2001;120:1442–7.
Montplaisir J, Lorrain J, Denesle R, Petit D. Sleep in menopause:
differential effects of two forms of hormone replacement therapy.
Menopause 2001;8:10–6.
Murr SM, Geschwind II, Bradford GE. Plasma LH and FSH during
different oestrous cycle conditions in mice. Journal of Reproduc-
tion and Fertility 1973;32:221–30.
Muzur A, Pace-Schott EF, Hobson JA. The prefrontal cortex in sleep.
Trends in Cognitive Sciences 2002;6(11):475–81.
Natale V, Albertazzi P, Zini M, Di Micco R. Exploration of cyclical
changes in memory and mood in postmenopausal women taking
sequential combined oestrogen and progestogen preparations.
International Journal of Obstetrics and Gynaecology 2001;108:
286–90.
Nelson JF, Felicio LS, Osterburg HH, Finch CE. Differential
contributions of ovarian and extraovarian factors to age-related
reductions in plasma estradiol and progesterone during the oestrus
cycle of C57BL/6J mice. Endocrinology 1992;130:805–10.
Nelson JF, Karelus K, Felicio LS, Johnson TE. Genetic influences on
oestrous cyclicity in mice: evidence that cycle length and frequency
are differentially regulated. Journal of Reproduction and Fertility
1992;94:261–8.
Nikkola E, Ekblad U, Ekholm E, Mikola H, Polo O. Sleep in multiple
pregnancy – breathing patterns, oxygenation, and periodic leg
movements. American Journal of Obstetrics and Gynecology 1996;
174:1622–5.
Ohayon M. Epidemiological study on insomnia in the general
population. Sleep 1996;19:S7–S15.
Ohayon MM, Lemoine P, Arnaud-Briant V, Dreyfus M. Prevalence
and consequences of sleep disorders in a shift worker population.
Journal of Psychosomatic Research 2002;53:577–83.
Oldenhave A, Jaszmann LJ, Haspels AA, Everaerd WT. Impact of
climacteric on well-being. A survey based on 5213 women 39 to 60
years old. American Journal of Obstetrics and Gynecology 1993;
168:772–80.
Overeem S, Mignot E, van Dijk JG, Lammers GJ. Narcolepsy: clinical
features, new pathophysiologic insights, and future perspectives.
Journal of Clinical Neurophysiology 2001;18:78–105.
Parry BL, Berga SL, Mostofi N, Klauber MR, Resnick A. Plasma
melatonin circadian rhythms during the menstrual cycle and after
light therapy in premenstrual dysphoric disorder and normal
control subjects. Journal of Biological Rhythms 1997;12:47–64.
Parry BL, Curtan ML, Stuenkel CA, Yokimozo M, Tam L, Powell
KA, Gillin JC. Can critically timed sleep deprivation be useful in
pregnancy and postpartum depressions. Journal of Affect Disor-
ders 2000;60:201–12.
Parry BL, Mendelson WB, Duncan WC, Sack DA, Wehr TA.
Longitudinal sleep EEG, temperature, and activity measurements
across the menstrual cycle in patients with premenstrual depression
and in age-matched controls. Psychiatry Research 1989;30:285–
303.
Partinen M, Eskelinen L, Tuomi K. Complaints of insomnia in
different occupations. Scandinavian Journal of Work, Environment
and Health 1984;10:467–9.
Pickett CK, Regensteiner JG, Woodard WD, Hagerman DD, Weil JV,
Moore LG. Progestin and estrogen reduce sleep-disordered breath-
ing in postmenopausal women. Journal of Applied Physiology
1989;66:1656–61.
Pilcher J. Women in comtemporary Britain. London: Routledge; 2000.
Pittendrigh CS, Daan S. Circadian oscillations in rodents: a systematic
increase of their frequency with age. Science 1974;186:548–50.
Pollm€acher T, Schulz H. Periodic leg movements (PLM): their
relationship to sleep stages. Sleep 1993;16:572–7.
Polo-Kantola P, Erkkola R, Helenius H, Irjala K, Polo O. When does
estrogen replacement therapy improve sleep quality. American
Journal of Obstetrics and Gynecology 1998;178:1002–9.

A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76 75
Polo-Kantola P, Erkkola R, Irjala K, Helenius H, Pullinen S, Polo O.
Climacteric symptoms and sleep quality. Obstetrics and Gynecol-
ogy 1999;94:219–24.
Polo-Kantola P, Erkkola R, Irjala K, Pullinen S, Virtanen I, Polo O.
Effect of short-term transdermal estrogen replacement therapy on
sleep: a randomized, double-blind crossover trial in postmeno-
pausal women. Fertility and Sterility 1999;71:873–80.
Popay J. My health is all right, but I’m just tired all the time: Women’s
experience of ill health. In: Roberts H, editor. Women’s health
matters. London: Routledge; 1992.
Porkka-Heiskanen T, Alanko L, Kalinchuk A, Heiskanen S, Stenberg
D. The effect ageing of on prepro-orexin gene expression and
contents of Orexin A and B in the rat brain et al. Neurobiology of
ageing 2004;25:231–8.
Purdie DW, Empson JA, Crichton C, Macdonald L. Hormone
replacement therapy, sleep quality and psychological wellbeing.
British Journal of Obstetrics and Gynecology 1995;102:735–9.
Rechtschaffen A, Kales A. A manual of standardized terminology,
techniques and scoring system for sleep stages of human subjects.
Bethesda, MD: National Institute of Neurological Diseases and
Blindness; 1968.
Rechtschaffen A. Current perspectives on the function of sleep.
Perspectives in Biology and Medicine 1998;41:359–90.
Rediehs MH, Reis JS, Creason NS. Sleep in old age: focus on gender
differences. Sleep 1990;13:410–24.
Redline S, Kump K, Tishler PV, Browner I, Ferrette V. Gender
differences in sleep disordered breathing in a community-based
sample. American Journal of Respiratory and Critical Care
Medicine 1994;149:722–6.
Reynolds 3rd CF, Kupfer DJ, Taska LS, Hoch CC, Sewitch DE,
Spiker DG. Sleep of healthy seniors: a revisit. Sleep 1985;8(1):
20–9.
Reynolds 3rd CF, Kupfer DJ, Hoch CC, Stack JA, Houck PR,
Berman SR. Sleep deprivation in healthy elderly men and women:
effects on mood and on sleep during recovery. Sleep 1986;9:492–
501.
Roffwarg H. Diagnostic classification of sleep and arousal disorders.
Sleep 1979;2:1–137.
Rogacz S, Duffy JF, Ronda JM, Czeisler C. The increase in body
temperature during the luteal phase of the menstrual cycle is only
observed during the subjective night and is independent of sleep.
Sleep Research 1988;17:395.
Sack RL, Lewy AJ, Erb DL, Vollmer WM, Singer CM. Human
melatonin production decreases with age. Journal of Pineal
Research 1986;3:379–88.
Santiago JR, Nolledo MS, Kinzler W, Santiago TV. Sleep and sleep
disorders in pregnancy. Annals of Internal Medicine
2001;134(5):396–408.
Satinoff E, Li H, Tcheng TK, Liu C, McArthur AJ, Medanic M,
Gillette MU. Do the suprachiasmatic nuclei oscillate in old rats as
they do in young ones. American Journal of Physiology 1993;265:
R1216–22.
Scharf MB, McDannold MD, Stover R, Zaretsky N, Berkowitz DV.
Effects of estrogen replacement therapy on rates of cyclic alternat-
ing patterns and hot-flush events during sleep in postmenopausal
women: a pilot study. Clinical Therapeutics 1997;19:304–11.
Schenck CH, Mahowald MW. REM sleep behavior disorder: clinical,
developmental, and neuroscience perspectives 16 years after its
formal identification in SLEEP. Sleep 2002;25:120–38.
Schernhammer ES, Laden F, Speizer FE, Willett WC, Hunter DJ,
Kawachi I, Colditz GA. Rotating night shifts and risk of breast
cancer in women participating in the nurses’ health study. Journal
of National Cancer Institute 2001;93:1563–8.
Schernhammer ES, Laden F, Speizer FE, Willett WC, Hunter DJ,
Kawachi I, Fuchs CS, Colditz GA. Night-shift work and risk of
colorectal cancer in the nurses’ health study. Journal of National
Cancer Institute 2003;95:825–8.
Schiff I, Regestein Q, Tulchinsky D, Ryan KJ. Effects of estrogens on
sleep and psychological state of hypogonadal women. JAMA
1979;242:2404–5.
Schmidt-Matthiesen. Gyn€akologie und Geburtshilfe. ISBN 3-7945-
1470-X, Schattauer 1992.
Schwartz B. Notes on the sociology of sleep. Sociological Quarterly
1970;11(Fall):485–99.
Schweiger MS. Sleep disturbance in pregnancy. A subjective survey.
American Journal of Obstetrics and Gynecology 1972;114:879–82.
Schwierin B, Borb�ely AA, Tobler I. Sleep homeostasis in the female rat
during the oestrus cycle. Brain Research 1998;811:96–104.
Sherman BM, Pfohl B, Winokur G. The correspondence of plasma
ACTH and cortisol before and after dexamethasone in healthy and
depressed subjects. Psychiatric Medicine 1985;3:41–52.
Sherwin BB. Estrogen and cognitive functioning in women. Endocrine
2003;24(2):133–51.
Shibui K, Uchiyama M, Okawa M, Kudo Y, Kim K, Liu X, Kamei Y,
Hayakawa T, Akamatsu T, Ohta K, Ishibashi K. Diurnal
fluctuation of sleep propensity and hormonal secretion across the
menstrual cycle. Biology of Psychiatry 2000;48:1062–8.
Shiromani PJ, Lu J, Wagner D, Thakkar J, Greco MA, Basheer R,
Thakkar M. Compensatory sleep response to 12 h wakefulness in
young and old rats. American Journal of Physiology-Regulatory
Integrative and Comparative Physiology 2000;278:R125–33.
Shochat T, Martin J, Marler M, Ancoli-Israel S. Illumination levels in
nursing home patients: effects on sleep and activity rhythms.
Journal of Sleep Research 2000;9:373–9.
Siegel JM. The REM sleep-memory consolidation hypothesis. Science
2001;294:1058–63.
Silberstein SD, Merriam GR. Physiology of the menstrual cycle.
Cephalalgia 2000;20:148–54.
Skene DJ, Bojkowski CJ, Currie JE, Wright J, Boulter PS, Arendt J. 6-
sulphatoxymelatonin production in breast cancer patients. Journal
of Pineal Research 1990;8:269–76.
Skene DJ, Lockley SW, Thapan K, Arendt J. Effects of light on human
circadian rhythms. Reproduction Nutrition Development 1999;39:
295–304.
Skene DJ, Swaab DF. Melatonin rhythmicity: effect of age and
Alzheimer’s disease. Experimental Gerontology 2003;38:199–206.
Skene DJ, Vivien-Roels B, Sparks DL, Hunsaker JC, Pevet P, Ravid
D, Swaab DF. Daily variation in the concentration of melatonin
and 5-methoxytryptophol in the human pineal gland: effect of age
and Alzheimer’s disease. Brain Research 1990;528:170–4.
Smith MJ, Schmidt PJ, Rubinow DR. Operationalizing DSM-IV
criteria for PMDD: selecting symptomatic and asymptomatic
cycles for research. Journal of Psychiatric Research 2003;37:
75–83.
Speroff L. The menopause. A signal for the future. In: Lobo R, editor.
Treatment of the postmenopausal women: basic and clinical
aspects. New York: Raven Press Ltd; 1994.
Spiegel K, Leproult R, Van Cauter E. Impact of sleep debt on
metabolic and endocrine function. Lancet 1999;354:1435–9.
Spiegel K, Sheridan JF, Van Cauter E. Effect of sleep deprivation on
response to immunization. JAMA 2002;288:1471–2.
Steiger A, Trachsel L, Guldner J, Hemmeter U, Rothe B, Rupprecht
R, Vedder H, Holsboer F. Neurosteroid pregnenolone induces
sleep-EEG changes in man compatible with inverse agonistic
GABAA-receptor modulation. Brain Research 1993;615:
267–74.
Swaab DF, Fliers E, Partiman TS. The suprachiasmatic nucleus of the
human brain in relation to sex, age and senile dementia. Brain
Research 1985;342:37–44.
Swain AM, O’Hara MW, Starr KR, Gorman LL. A prospective study
of sleep, mood, and cognitive function in postpartum and
nonpostpartum women. Obstetrics and Gynecology 1997;90:381–6.
Tafti M, Dauvilliers Y. Pharmacogenomics in the treatment of
narcolepsy. Pharmacogenomics 2003;4:23–33.

76 A. Dzaja et al. / Journal of Psychiatric Research 39 (2005) 55–76
Tani Y, Ishihara T. Changes in EEG associated with sleep-awake
behavior in young adult versus aged adult Fischer-344 rats.
Physiological Behaviour 1988;44:389–92.
Taylor B. Unconsciousness and Society: The Sociology of Sleep.
Journal of Politics, Culture and Society 1993;6:463–71.
Terao A, Apte-Deshpande A, Morairty S, Freund YR, Kilduff TS.
Age-related decline in hypocretin (orexin) receptor 2 messenger
RNA levels in the mouse brain. Neuroscience Letters 2002;332:
190–4.
Thomson J, Oswald I. Effect of oestrogen on the sleep, mood, and
anxiety of menopausal women. British Medical Journal 1977;2:
1317–9.
Thomson J, Oswald I. Effect of oestrogen on the sleep, mood, and
anxiety of menopausal women. British Medicine Journal 1977;2:
1317–9.
Tilley AJ, Wilkinson RT. The effects of a restricted sleep regime on the
composition of sleep and on performance. Psychophysiology
1984;21:406–12.
Tononi G, Cirelli C. Modulation of brain gene expression during sleep
and wakefulness: a review of recent findings. Neuropsychophar-
macology 2001;25:S28–35.
Toran-Allerand CD. Novel sites and mechanisms of oestrogen action
in the brain. Novartis Foundation Symposium 2000;230:56–69.
Van Cauter E, Leproult R, Plat L. Age-related changes in slow wave
sleep and REM sleep and relationship with growth hormone and
cortisol levels in healthy men. JAMA 2000;284:861–8.
Van Cauter E, Sturis J, Byrne MM, Blackman JD, Leproult R, Ofek
G, L’Hermite-Baleriaux M, Refetoff S, Turek FW, Van Reeth O.
Demonstration of rapid light-induced advances and delays of the
human circadian clock using hormonal phase markers. American
Journal of Physiology 1994;266:E953–63.
Van Dongen HP, Maislin G, Mullington JM, Dinges DF. Investigator:
Van Dongen HP, Mullington JM, Dinges DF. – The cumulative
cost of additional wakefulness: dose-response effects on neurobe-
havioral functions and sleep physiology from chronic sleep
restriction and total sleep deprivation. Sleep 2003;26:117–126.
Van Gool WA, Mirmiran M. Age-related changes in the sleep pattern
of male adult rats. Brain Research 1983;279:394–8.
Van Gool WA, Witting W, Mirmiran M. Age-related changes in
circadian sleep–wakefulness rhythms in male rats isolated from
time cues. Brain Research 1987;413:384–7.
VanHelder T, Symons JD, Radomski MW. Effects of sleep deprivation
and exercise on glucose tolerance. Aviation, Space and Environ-
mental Medicine 1993;64(6):487–92.
Van Someren EJ, Kessler A, Mirmiran M, Swaab DF. Indirect bright
light improves circadian rest-activity rhythm disturbances in
demented patients. Biology of Psychiatry 1997;41:955–63.
Vanselow W. A comprehensive approach to premenstrual complaints.
Australian Family Physician 1998;27:354–61.
Waldhauser F, Weiszenbacher G, Tatzer E, Gisinger B, Waldhauser
M, Schemper M, Frisch H. Alterations in nocturnal serum
melatonin levels in humans with growth and aging. Journal of
Clinical Endocrinology and Metabolism 1988;66:648–52.
WebbWB. Sleep in older persons: sleep structures of 50- to 60-year-old
men and women. Journal of Gerontology 1982;37:581–6.
Webb WB, Agnew Jr HW. Sleep efficiency for sleep–wake cycles of
varied length. Psychophysiology 1975;12:637–41.
Wehr TA. Improvement of depression and triggering of mania by sleep
deprivation. JAMA 1992;267:548–51.
Weinert D. Lower variability in female as compared to male
laboratory mice: Investigations on circadian rhythms. Journal of
Experimental Animal Sciences 1996;37:121–37.
Weinert D. Age-dependent changes of the circadian system. Chrono-
biology International 2000;17:261–83.
Welsh DK, Richardson GS, Dement WC. Effect of age on the
circadian pattern of sleep and wakefulness in the mouse. Journal of
Gerontology 1986;41:579–86.
West CP. The characteristics of 100 women presenting to a gyneco-
logical clinic with premenstrual complaints. Acta Obstetricia et
Gynecologica Scandinavica 1989;68:743–7.
Wetter TC, Pollm€acher T. Restless legs and periodic leg movements in
sleep syndromes. Journal of Neurology 1997;244:S37–45.
Wiklund I, Berg G, Hammar M, Karlberg J, Lindgren R, Sandin K.
Long-term effect of transdermal hormonal therapy on aspects of
quality of life in postmenopausal women. Maturitas 1992;14:225–
36.
Wiklund I, Karlberg J, Mattsson LA. Quality of life of postmeno-
pausal women on a regimen of transdermal estradiol therapy: a
double-blind placebo-controlled study. American Journal of Ob-
stetrics and Gynecology 1993;168:824–30.
Williams S. Sleep and Health: Sociological reflections on the dormant
society. Health 2002;6:173–200.
Williams S, Bendelow G. The lived body. Sociological themes,
embodied issues. London: Routledge; 1998.
Winget C, DeRoshia C, Vernikos-Danellis J, Rosenblatt W, Hethe-
rington N. Comparison of circadian rhythms in male and female
humans. Waking and Sleep 1977;1:359–63.
Winkelmann J, Wetter TC, Collado-Seidel V, Gasser T, Dichgans M,
Yassouridis A, Trenkwalder C. Clinical characteristics and fre-
quency of the hereditary restless legs syndrome in a population of
300 patients. Sleep 2000;23:597–602.
Witting W, Mirmiran M, Bos NP, Swaab DF. Effect of light intensity
on diurnal sleep–wake distribution in young and old rats. Brain
Research Bulletin 1993;30:157–62.
The Women’s Health Initiative (WHI).
Woodward S, Freedman RR. The thermoregulatory effects of meno-
pausal hot flashes on sleep. Sleep 1994;17:497–501.
Wootten N. Evaluation of 12-hour shifts on a cardiology nursing
development unit. British Journal of Nursing 2000;9:2169–74.
Wu JC, Bunney WE. The biological basis of an antidepressant
response to sleep deprivation and relapse: review and hypothesis.
American Journal of Psychiatry 1990;147:14–21.
Yokoyama A, Ramirez V, Sawyer C. Sleep and wakefulness in female
rats under various hormonal and physiological conditions. General
and Comparative Endocrinology 1966;7:10–7.
Young IM, Francis PL, Leone AM, Stovell P, Silman RE. Constant
pineal output and increasing body mass account for declining
melatonin levels during human growth and sexual maturation.
Journal of Pineal Research 1988;5:71–85.
Young T. Analytic epidemiology studies of sleep disordered breathing–
what explains the gender difference in sleep disordered breathing.
Sleep 1993;16(8 Suppl):S1–2.
Young T, Peppard P. Sleep-disordered breathing and cardiovascular
disease: epidemiologic evidence for a relationship. Sleep 2000;23/
4:S122–6.
Zeitzer JM, Daniels JE, Duffy JF, Klerman EB, Shanahan TL, Dijk
DJ, Czeisler CA. Do plasma melatonin concentrations decline with
age? Am J Med JID – 0267200 1999;107:432–6.
Zeitzer JM, Dijk DJ, Kronauer R, Brown E, Czeisler C. Sensitivity of
the human circadian pacemaker to nocturnal light: melatonin
phase resetting and suppression. Journal of Physiology 2000;526(Pt
3):695–702.
Zepelin H, Whitehead WE, Rechtschaffen A. Aging and sleep in the
albino rat. Behaviour 1972;7:65–74.




















