
Voluntary exercise prevents the obese and diabetic metabolic syndrome of the melanocortin-4 receptor...
14
The FASEB Journal • Research Communication Voluntary exercise prevents the obese and diabetic metabolic syndrome of the melanocortin-4 receptor knockout mouse Carrie Haskell-Luevano,* ,1 Jay W. Schaub,* Amy Andreasen,* Kim R. Haskell,* Marcus C. Moore,* Lorraine M. Koerper,* Francois Rouzaud,* Henry V. Baker, † William J. Millard,* Glenn Walter, ‡ S. A. Litherland, § and Zhimin Xiang* *Department of Pharmacodynamics, College of Pharmacy, † Department of Molecular Genetics and Microbiology, ‡ Department of Physiology and Functional Genomics, and § Department of Pathology, Immunology, and Laboratory Medicine, University of Florida, Gainesville, Florida, USA ABSTRACT Exercise is a mechanism for maintenance of body weight in humans. Morbidly obese human pa- tients have been shown to possess single nucleotide poly- morphisms in the melanocortin-4 receptor (MC4R). MC4R knockout mice have been well characterized as a genetic model that possesses phenotypic metabolic disorders, including obesity, hyperphagia, hyperinsulin- emia, and hyperleptinemia, similar to those observed in humans possessing dysfunctional hMC4Rs. Using this model, we examined the effect of voluntary exercise of MC4R knockout mice that were allowed access to a running wheel for a duration of 8 wk. Physiological parameters that were measured included body weight, body composition of fat and lean mass, food consump- tion, body length, and blood levels of cholesterol and nonfasted glucose, insulin, and leptin. At the termina- tion of the experiment, hypothalamic mRNA expres- sion levels of neuropeptide Y (NPY), agouti-related protein (AGRP), proopiomelanocortin (POMC), cocaine- and amphetamine-regulated transcript (CART), orexin, brain-derived neurotropic factor (BDNF), phosphatase with tensin homology (Pten), melanocortin-3 receptor (MC3R), and NPY-Y1R were determined. In addition, islet cell distribution and function in the pancreas were examined. In the exercising MC4R knockout mice, the pancreatic islet cell morphology and other physiological parameters resembled those observed in the wild-type littermate controls. Gene expression profiles identified exercise as having a significant effect on hypothalamic POMC, orexin, and MC3R levels. Genotype had a signif- icant effect on AGRP, POMC, CART, and NPY-Y1R, with an exercise and genotype interaction effect on NPY gene expression. These data support the hypothesis that volun- tary exercise can prevent the genetic predisposition of melanocortin-4 receptor-associated obesity and diabe- tes.—Haskell-Luevano, C., Schaub, J. W., Andreasen, A., Haskell, K. R., Moore, M. C., Koerper, L. M., Rouzaud, F., Baker, H. V., Millard, W. J., Walter, G., Litherland, S. A., Xiang, Z. Voluntary exercise prevents the obese and diabetic metabolic syndrome of the melanocortin-4 recep- tor knockout mouse. FASEB J. 23, 642– 655 (2009) Key Words: energy homeostasis leptin insulin -melano- cyte stimulating hormone Obesity and its associated diseases are becoming more prevalent in society. To combat this growing epidemic, scientists are looking at a myriad of treat- ment possibilities to both prevent and correct obesity. One well-recognized mechanism is the combination of diet and exercise. Although positive changes in diet are associated with proper cellular nutrition and changes in energy homeostasis, very little is know about the molecular mechanism by which exercise can both prevent and correct increased body weights. Energy and body weight homeostasis are extremely complex physiological mechanisms that rely on endogenous mechanisms as well as genetic and environmental in- puts. Herein, we have examined how voluntary exercise can bypass the genetic predisposition for overeating, obesity, and increased serum insulin and leptin levels associated with type 2 diabetes found in the melanocor- tin-4 receptor (MC4R) knockout (KO) mouse model (1). MC4R gene polymorphisms also have been identi- fied in morbidly obese humans (2–17), providing trans- lational potential between humans and the genetic MC4R KO mouse model. The melanocortin pathway consists of the endoge- nous agonists derived from the proopiomelanocortin (POMC) gene and the endogenous antagonist agouti- related protein (AGRP) interacting with the melano- cortin-3 receptor (MC3R) and MC4R of hypothalamic neurons. Stimulation of the MC3R and/or MC4R path- ways by endogenous agonists such as -melanocyte- stimulating hormone results in activation of the cAMP signal transduction pathway (18 –23) to generate intra- and intercellular physiological responses that can be blocked or regulated by AGRP (24 –26). An under- 1 Correspondence: Department of Pharmacodynamics, University of Florida, PO Box 100487, Gainesville, FL 32610, USA. E-mail: [email protected]fl.edu doi: 10.1096/fj.08-109686 642 0892-6638/09/0023-0642 © FASEB
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Voluntary exercise prevents the obese and diabetic metabolic syndrome of the melanocortin-4 receptor...

The FASEB Journal • Research Communication
Voluntary exercise prevents the obese and diabeticmetabolic syndrome of the melanocortin-4 receptorknockout mouse
Carrie Haskell-Luevano,*,1 Jay W. Schaub,* Amy Andreasen,* Kim R. Haskell,*Marcus C. Moore,* Lorraine M. Koerper,* Francois Rouzaud,* Henry V. Baker,†
William J. Millard,* Glenn Walter,‡ S. A. Litherland,§ and Zhimin Xiang**Department of Pharmacodynamics, College of Pharmacy, †Department of Molecular Genetics andMicrobiology, ‡Department of Physiology and Functional Genomics, and §Department of Pathology,Immunology, and Laboratory Medicine, University of Florida, Gainesville, Florida, USA
ABSTRACT Exercise is a mechanism for maintenanceof body weight in humans. Morbidly obese human pa-tients have been shown to possess single nucleotide poly-morphisms in the melanocortin-4 receptor (MC4R).MC4R knockout mice have been well characterized as agenetic model that possesses phenotypic metabolicdisorders, including obesity, hyperphagia, hyperinsulin-emia, and hyperleptinemia, similar to those observed inhumans possessing dysfunctional hMC4Rs. Using thismodel, we examined the effect of voluntary exercise ofMC4R knockout mice that were allowed access to arunning wheel for a duration of 8 wk. Physiologicalparameters that were measured included body weight,body composition of fat and lean mass, food consump-tion, body length, and blood levels of cholesterol andnonfasted glucose, insulin, and leptin. At the termina-tion of the experiment, hypothalamic mRNA expres-sion levels of neuropeptide Y (NPY), agouti-relatedprotein (AGRP), proopiomelanocortin (POMC), cocaine-and amphetamine-regulated transcript (CART), orexin,brain-derived neurotropic factor (BDNF), phosphatasewith tensin homology (Pten), melanocortin-3 receptor(MC3R), and NPY-Y1R were determined. In addition,islet cell distribution and function in the pancreas wereexamined. In the exercising MC4R knockout mice, thepancreatic islet cell morphology and other physiologicalparameters resembled those observed in the wild-typelittermate controls. Gene expression profiles identifiedexercise as having a significant effect on hypothalamicPOMC, orexin, and MC3R levels. Genotype had a signif-icant effect on AGRP, POMC, CART, and NPY-Y1R, withan exercise and genotype interaction effect on NPY geneexpression. These data support the hypothesis that volun-tary exercise can prevent the genetic predisposition ofmelanocortin-4 receptor-associated obesity and diabe-tes.—Haskell-Luevano, C., Schaub, J. W., Andreasen, A.,Haskell, K. R., Moore, M. C., Koerper, L. M., Rouzaud,F., Baker, H. V., Millard, W. J., Walter, G., Litherland, S.A., Xiang, Z. Voluntary exercise prevents the obese anddiabetic metabolic syndrome of the melanocortin-4 recep-tor knockout mouse. FASEB J. 23, 642–655 (2009)
Key Words: energy homeostasis � leptin � insulin � �-melano-cyte stimulating hormone
Obesity and its associated diseases are becomingmore prevalent in society. To combat this growingepidemic, scientists are looking at a myriad of treat-ment possibilities to both prevent and correct obesity.One well-recognized mechanism is the combination ofdiet and exercise. Although positive changes in diet areassociated with proper cellular nutrition and changesin energy homeostasis, very little is know about themolecular mechanism by which exercise can bothprevent and correct increased body weights. Energyand body weight homeostasis are extremely complexphysiological mechanisms that rely on endogenousmechanisms as well as genetic and environmental in-puts. Herein, we have examined how voluntary exercisecan bypass the genetic predisposition for overeating,obesity, and increased serum insulin and leptin levelsassociated with type 2 diabetes found in the melanocor-tin-4 receptor (MC4R) knockout (KO) mouse model(1). MC4R gene polymorphisms also have been identi-fied in morbidly obese humans (2–17), providing trans-lational potential between humans and the geneticMC4R KO mouse model.
The melanocortin pathway consists of the endoge-nous agonists derived from the proopiomelanocortin(POMC) gene and the endogenous antagonist agouti-related protein (AGRP) interacting with the melano-cortin-3 receptor (MC3R) and MC4R of hypothalamicneurons. Stimulation of the MC3R and/or MC4R path-ways by endogenous agonists such as �-melanocyte-stimulating hormone results in activation of the cAMPsignal transduction pathway (18–23) to generate intra-and intercellular physiological responses that can beblocked or regulated by AGRP (24–26). An under-
1 Correspondence: Department of Pharmacodynamics,University of Florida, PO Box 100487, Gainesville, FL 32610,USA. E-mail: [email protected]
doi: 10.1096/fj.08-109686
642 0892-6638/09/0023-0642 © FASEB

standing of the importance of the melanocortin path-way in energy homeostasis developed from humangenetic studies (27–29), KO and transgenic mousemodels (1, 24, 30–34), and the discovery of humanpolymorphisms (2–17). Central administration of mela-nocortin agonists led to decreased food intake, whereassimilarly administered melanocortin antagonists in-creased food intake and the sustained increases in foodintake (days) on administration of AGRP (34–40).Melanocortin agonist POMC-expressing neurons havebeen identified as coexpressing cocaine- and amphet-amine-regulated transcript (CART) (41), the MC3R(42, 43), leptin receptors (44, 45), and the neuropep-tide Y (NPY) Y1 receptor (46). Melanocortin antagonistAGRP-expressing neurons have been identified as co-expressing NPY (47, 48), the MC3R (42, 43), and leptinreceptors (44, 45). The POMC/CART and AGRP/NPYneurons innervate several brain nuclei containing theMC4R that have been suggested to be important forfeeding and energy homeostasis, but the exact site ofthe MC4R feeding behavior “center” remains to beidentified, although it is thought to be within themacrostructure of the hypothalamus. Leptin and insu-lin are adiposity factors that regulate POMC/CART andAGRP/NPY neuronal signaling and associated physio-logical effects. The leptin and insulin hormones signalin the hypothalamic neurons through the phosphati-dylinositol 3-kinase pathway that is inhibited by phos-phatase with tensin homology (Pten) (49, 50). Pten hasalso been identified as an important factor for pancre-atic �-cell homeostasis (51). Brain-derived neurotropicfactor (BDNF) is expressed in the hypothalamus (ven-tromedial hypothalamus, dorsomedial hypothalamus,lateral hypothalamus, and paraventricular nucleus) andhas been demonstrated to reduce food intake, decreasebody weight gain, modify activity (52), and functiondownstream of the melanocortin receptors in the ag-outi Ay genetic mouse model (53). The receptor forBDNF is the tyrosine kinase B (TrkB) protein that iswidely expressed in the hypothalamus and brain, in-cluding the arcuate nucleus, which also contain thePOMC/CART- and AGRP/NPY-expressing neurons(52). However, TrkB does not appear to colocalize witheither population (53). Thus, the melanocortin path-way is a “key” neuroendocrine pathway responding toseveral endocrine signals (directly and indirectly) andparticipating in the complex physiological process ofingestive behavior and energy homeostasis.
In this study, we have examined the effects of 8 wk ofvoluntary exercise on the genetic MC4R KO mousemodel in age- and weight-matched littermate controls.We evaluated several physiological parameters includ-ing food intake, body weight, body length, fat mass,lean mass, blood cholesterol, glucose, insulin, andleptin. To begin to search for a putative underlyingmolecular mechanism, we evaluated mRNA expressionlevels of NPY, AGRP, POMC, CART, MC3R, NPY-Y1R,orexin, BDNF, and Pten in the hypothalamus of thebrain. In addition, we evaluated insulin immunohisto-chemical staining of pancreatic �-cells and insulin
secretion to determine whether voluntary exercise af-fects insulin secretion capacity of the pancreas in theexercised vs. sedentary MC4R KO mice.
MATERIALS AND METHODS
All studies were performed in accord with accepted standardsof humane animal care and were approved by the institu-tional animal care and use committee at the University ofFlorida.
Experimental animals
MC4R�/� heterozygous mice were generously provided byDr. Dennis Huszar (Millennium Pharmaceuticals, Cam-bridge, MA, USA). The MC4R�/� homozygous KO andwild-type controls used in this experiment were obtainedusing a heterozygous breeding strategy and were not con-trolled for litter size (the observed average litter size for theseheterozygous MC4R breeding pairs was similar to that of thewild-type breeding pairs in our colony). Genotyping wasperformed as described previously (1). Mice were housed ina 12-h light/dark cycle (lights on at 11 PM and off at 11 AM)and had ad libitum access to rodent chow and water for theduration of this study. At the initial experimental setup, thegroups were littermate-matched for genotype, age, and bodyweight. No statistically significant differences were observedbetween the sedentary vs. exercised experimental groups atthe initiation of these experiments. The data presented at wk7 are after 1 wk of experimental treatment.
Running wheel vs. conventional housing experiments
Conventionally housed mice were housed in standard mouseshoebox-size cages singly or in a group of 2 mice/cage asindicated with the corresponding data. Exercising male micewere housed individually in cages containing a running wheel(Mini Mitter Co., Sunriver, OR, USA). Wheel revolutionswere quantified by recording the magnetic switch closures ofa magnet placed on the revolving wheel and recorded viacomputer as described previously (54, 55).
Cumulative food intake
Cumulative food intake was determined for singly housedmice by measuring the weight of standard chow (HarlanTeklad 8604 diet; 24% crude protein, 4% crude fat, and 4.5%maximum crude fiber, with digestible energy of 3.30 kcal/g)remaining in the cage top wire feeder 3 times weekly. Thebedding was inspected for large intact pellets �0.1 g todecrease the error associated with determining food intake.Feed efficiency (amount of body weight gained relative to thecaloric intake) was calculated as the total body weight changeeach week divided by food intake (kcal) for the respectiveweek with the entire data averaged over the entire experi-ment for each group.
Body length
Body length (nose to anus, measured in cm) was determined3 times weekly with a linear ruler after the animals were lightlyanesthetized with isoflurane gas.
643VOLUNTARY EXERCISE AND THE MC4R KO MOUSE

Nonfasted glucose
Nonfasted blood glucose values were determined using aglucometer (Glucometer Elite XL, Bayer Corporation,Deerfield, IL, USA) with a sensitivity of 20 mg/dl and arange of detection to 600 mg/dl. A whole-blood sample of1 �l was measured twice weekly. To avoid compoundingerror in our metabolic and molecular measurement, fast-ing glucose measurements were not used to prevent apotential effect on expression levels of AGRP, NPY, andother neuroendocrine pathways involved in the regulationof energy homeostasis (56).
Cholesterol
Cholesterol was determined from a 15-�l plasma sample oncea week using a strip meter manufactured by Polymer Tech-nology Systems Inc. (Indianapolis, IN, USA). We used PTSPanels Total Cholesterol test strips with a detection range of100–400 mg/dl.
MRI
Magnetic resonance spectroscopy and imaging of the bodycomposition of the mice in vivo were performed at theUniversity of Florida, McKnight Brain Institute advanced MRIand spectroscopy facility. All animals were imaged on a 4.7-TOxford magnet using a Bruker Avance console and BrukerParaVision software (Bruker, Karlsruhe, Germany). The micewere anesthetized by reflexive inhalation of 1.5–2% isoflu-rane and 1 L of oxygen. A small-animal instrument monitor-ing and gating system was used for monitoring of and gatingon respiratory rate. The mice were centered in a custom-builtquadrature transmit/receive birdcage with a radiofrequencycoil 5 cm in diameter and 12.5 cm in length. After shimmingand rapid acquisition with relaxation enhancement (RARE)encoded pilot scans to confirm position, a single-pulse spec-trum was acquired with repetition time (TR) of 4.0 s, sweepwidth (SW) of 8000, acquisition size of 4000, and 16 averages.The water and lipid peaks were integrated using BrukerXWinNMR software, and lipid was standardized to an integralvalue of 1.000. After spectral acquisition, a multislice multie-cho (MSME)_TOMO RARE 3-dimensional scan was acquiredwith the following parameters: field of view (FOV), 7.0 �4.0 � 2.5 cm; matrix, 256 � 128 � 96; TR, 350.0 ms; echotime (TE), 6.8 ms; number of averages (NEX), 1; and RAREfactor, 16. These parameters resulted in a resolution of 273 �313 � 260 �m inplane. Serial transverse slices were collectedwith a RARE-BIO T2-weighted sequence with the followingparameters: FOV, 4.5 � 3.6 cm; thickness, 2.0 mm; matrix,256 � 256; TR, 3500.0 ms; TE, 7.0 ms; and NEX, 4. Theseparameters resulted in an inplane resolution of 176 � 141�m. Liver-localized spectroscopy was performed using stimu-lated echo acquisition mode (STEAM). Voxel location andsize validation were performed on phantoms and via imagingof the voxel in vivo. Voxel size was kept at �40 mm3. Alllocalized spectroscopy scans were respiratory gated. Thesequence was triggered at expiration, and TR was keptbetween 4 and 4.5 s. TE and mixing time were 20 and 10 ms,respectively; 6-kHz SW with 4000 points was collected with 16averages.
Fat and lean mass determination
The individual mice were placed weekly into a acrylic tube(polymethyl methacrylate) that was then inserted into themouse EchoMRI-100 instrument (Echo Medical Systems LLC,Houston TX, USA) (57, 58) to determine the mass (g) of the
fat vs. lean body composition. Total body weight was mea-sured twice weekly using a standard top-loading laboratorybalance.
Hormone assays in blood plasma
Blood-drop samples were drawn from the anterior facial veininto vials containing K2EDTA to prevent coagulation. Plasmawas collected by centrifugation and stored at �20°C untilanalyzed. Plasma hormone levels (insulin and leptin) weremeasured from a 10-�l sample (in duplicate) using commer-cially available Endocrine Lincoplex kits (Linco Research, St.Charles, MO) according to the manufacturer’s instructions.
Quantitative real-time polymerase chain reaction(qRT-PCR)
At the time of sacrifice, the hypothalamus of the brain wasdissected (excised with a razor blade by trimming out all ofthe brain structures, including the midbrain, cerebellum, andcortex; average hypothalamus weight 17.70.6 mg) fromeach mouse and placed immediately in RNAlater (AmbionInc., Austin, TX, USA) and held at 4°C for 24 h with transferto �20°C until RNA extraction. RNA was extracted usingTRIzol reagent and quantified, and integrity was confirmedby gel electrophoresis. cDNA was generated from 2 �g of totalRNA per the manufacturer’s instructions (Applied Biosys-tems, Foster City, CA, USA). qRT-PCR reactions were per-formed using 100 ng of cDNA as template with TaqManprimers and reagents in a ABI 7300 system (Applied Biosys-tems). Samples were run in duplicate reactions on a single96-well plate for each gene probe to confirm consistency inthe amount of PCR products. The hypoxanthine guaninephosphoribosyltransferase 1 (Hprt1) housekeeping gene wasused to correct for the amount of input mRNA and for datanormalization. We selected this housekeeping gene because1) it did not show any significant differences between geno-type or treatment from our microarray experiments, and 2) itis recommended for a mouse housekeeping gene by AppliedBiosystems (application note “Using TaqMan® EndogenousControl Assays to Select an Endogenous Control for Experi-mental Studies,” available at http://www.appliedbiosystems.com). Levels of mRNA are expressed as fold differencescompared with the wild-type sedentary group. The folddifference over the control Hprt1 gene is calculated as 2�Ct,where Ct � Ct(gene) � Ct(Hprt1 gene). Ct is the PCR cyclein which the fluorescent signal associated with the exponen-tial growth exceeds the threshold (10 times noise level).
Statistical analysis
Data are expressed as means se. Statistical differencesbetween groups of mice (genotype and exercise) over timewere determined using repeated-measures analysis of vari-ance (ANOVA) with the Bonferroni post hoc test. If a signifi-cant time � treatment interaction term was observed, indi-vidual comparisons between groups at given time points wereperformed using simple main effects post hoc testing with aBonferroni correction for the number of comparisons per-formed to set the experiment-wise probability at P � 0.05(59). Simple regression analysis was used to examine therelation between phenotypes, and results were consideredstatistically significant if P � 0.05. For RT-PCR, a two-wayANOVA with the Bonferroni post hoc test using the Ct valuesfor genotype effects, exercise effects, and genotype � exerciseinteractions was used. A simple Pearson correlation regres-sion analysis was used to examine the relation between
644 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal

different experimental parameters. Values of P � 0.05 wereconsidered significant.
RESULTS
Exercise overrides genetically induced obesity inMC4R KO mice
Heterozygous (�/�) breeding of the MC4R KO mice(1), resulted in the generation of the homozygous KOmice (�/�) and wild-type (�/�) littermate mice usedin these experiments. Starting at 6 wk of age, 10–12MC4R KO (�/�) and wild-type male mice (weight-and genotype-matched littermates) were singly housedin a Nalgene cage equipped with a running wheel andallowed free access to voluntary exercise for the exper-iment duration of 8 wk. At 6 wk of age and theinception of the experiment, the MC4R KO mice hadaverage body weight of 23 g, and no statistically signif-icant difference was observed for the two groups estab-lished as conventionally housed and running wheel-housed mice. The average body weight of the wild-typelittermate mice was 19 g; again no statistically signifi-cant difference was observed for the two groups estab-lished as conventionally housed and running wheelhoused mice. However, there was a statistically signifi-cant difference (P�0.001) between the wild-type andMC4R KO littermate body weights, consistent withprevious studies (1). Figure 1 summarizes the averageactivity between the exercising wild-type and MC4R KOlittermates in which no statistically significant differ-ences were observed. Tables 1 and 2 summarize andanalyze the physiological parameters measured in thisstudy.
Body weight of the mice in the 4 experimental groupswas measured twice weekly with averages of the 8-wkexperiment. During the initial experimental setup, boththe MC4R KO and wild-type littermate group bodyweights were not statistically different between the con-
ventionally housed vs. running wheel-housed groups.However, as already evident at 6 wk of age, the MC4R KOmice (23.10.5 g) were heavier than the wild-type mice(19.90.3 g) (P�0.001). No statistical difference wasobserved between the two MC4R KO groups after 1 wk ofexercise. Differences in body weight between the MC4RKO groups become apparent after 2 wk of running wheelexercise and persisted throughout the remainder of the8-wk trial. No statistically significant differences wereobserved between the wild-type conventionally housed vs.running wheel-housed mice at any time. Despite theobserved reduction in body weight in exercised MC4R KOmice, their body weights remained above those of thewild-type animals throughout the study (Table 1).
Exercise normalizes fat to lean body mass in MC4RKO mice
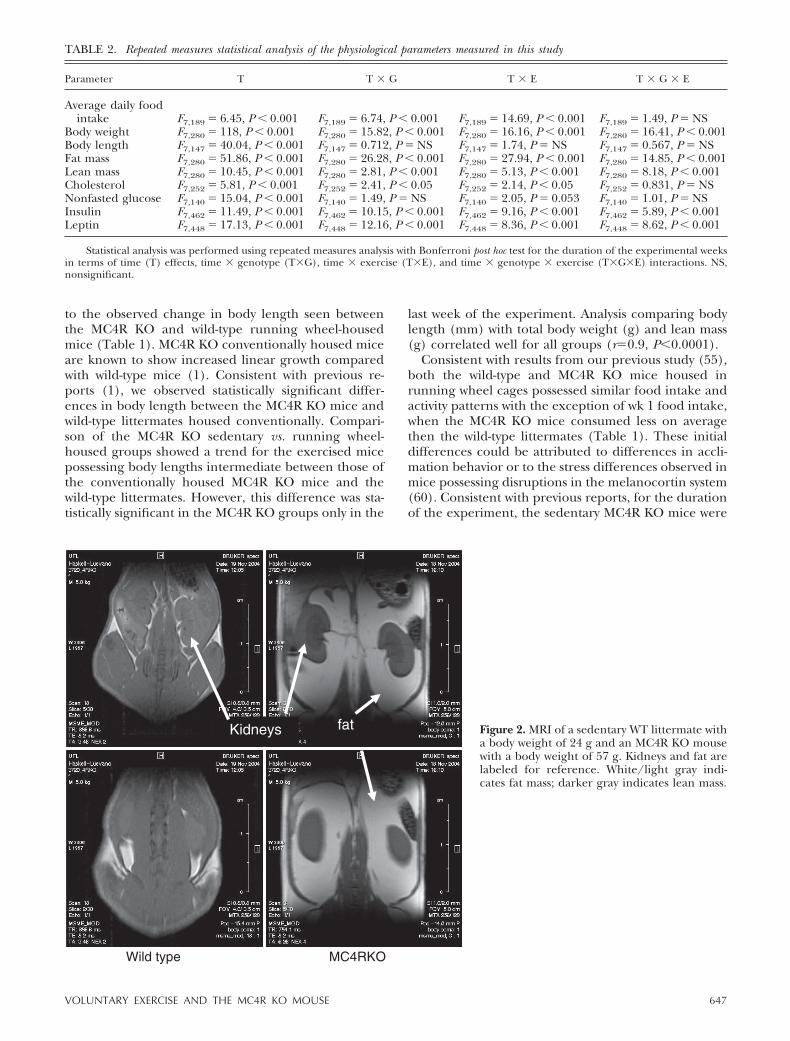
Changes in body weight could be due to a variety offactors. To visualize fat vs. lean body mass changes inmice, we performed MRI. Figure 2 indicates the com-parison between a representative wild-type (24 g) andMC4R KO (57 g) sedentary mouse. Figure 3 illustratesa snapshot of representative mice from each group overtime of treatment, starting at age 7 wk, after 1 wk ofinclusion in the experiment, at age 11 wk, and at age 14wk, the final week of the experiment after 8 wk oftreatment. Increased body weight in the MC4R KOmice appears to be predominately associated withincreased fat mass throughout the body, as postu-lated. MRI is a valuable technique for the visualiza-tion and distribution of fat tissue throughout thebody; however, it is difficult to quantify such changesusing this approach. Therefore, we used low-resolu-tion nuclear magnetic resonance techniques andequipment for the quantitation of live animal fat vs.lean mass. Using the EchoMRI-100 system (57, 58),we analyzed the body composition of our experimen-tal mice weekly, as summarized in Table 1. Statisti-cally significant differences in fat mass between theMC4R KO sedentary housed mice vs. the exercisingMC4R KO mice emerged after wk 2 of the experi-ment, correlating with total body weight (r�0.9,P�0.001). However, there were no statistically signif-icant differences between the MC4R exercised micevs. the wild-type conventionally housed mice, whichis different from the results that were observed fortotal body weight. Comparison of lean mass showedstatistically significant differences between the con-ventionally housed MC4R KO and wild-type litter-mate mice, the exercised MC4R KO and wild-typegroups, and the MC4R KO exercised and the wild-type sedentary housed groups. However, both theMC4R KO and wild-type groups housed either in tradi-tional conventional housing or in cages equipped with arunning wheel possessed the same relative lean mass forthe duration of the experiment. Interestingly, compar-ison of the ratio between lean and fat mass between thedifferent groups revealed that although, as postulated,no significant changes were observed between the
Figure 1. Summary of the average daily running wheel (RW)activity between the wild-type (WT) and MC4R KO litter-mates. Values are averages se; n � 10–12 mice/group. �,MC4R KO RW mice; E, wild-type littermate RW mice. Nostatistical difference was observed between the two groups.
645VOLUNTARY EXERCISE AND THE MC4R KO MOUSE

wild-type groups and the exercising MC4R KO mice, forthe sedentary MC4R KO mice, it appears that while theyincrease body weight and fat mass, they are also de-creasing relative lean mass (Fig. 4).
In contrast to total body weight, there were nostatistically significant differences in body fat massbetween the MC4R KO exercised mice vs. the wild-typesedentary mice. This discrepancy could be attributable
TABLE 1. Summary of the physiological parameters measured in this study
Week of experiment (age)
Parameter 1 (7 wk) 2 (8 wk) 3 (9 wk) 4 (10 wk) 5 (11 wk) 6 (12 wk) 7 (13 wk) 8 (14 wk)
Average daily foodintake (g)
WT Conv 3.7 0.1 3.5 0.1 3.4 0.1 3.4 0.1 3.3 0.1 3.2 0.1 3.2 0.03 3.2 0.1WT RW 3.9 0.2 4.2 0.1de 4.4 0.1de 4.3 0.1de 4.2 0.1de 4.1 0.1d 4.1 0.1de 4.0 0.1d
MC4R KO Conv 3.8 0.1 3.8 0.1a 3.9 0.02a 3.6 0.03a 3.7 0.1a 3.9 0.1a 3.6 0.1 3.7 0.1a
MC4R KO RW 3.1 0.2bcf 4.2 0.1bc 4.4 0.1bc 4.3 0.1bc 4.1 0.2bc 4.2 0.2c 4.3 0.1bc 4.2 0.1bc
Body weight (g)WT Conv 20.8 0.3 21.2 0.3 21.3 0.3 21.6 0.3 22.0 0.3 22.2 0.3 22.5 0.3 22.8 0.3WT RW 20.6 0.4e 20.9 0.4e 21.5 0.4e 22.0 0.4e 22.2 0.4e 22.3 0.4e 22.7 0.4e 22.6 0.3e
MC4R KO Conv 25.2 0.7a 25.5 0.6ab 26.4 0.6ab 27.5 0.7ab 28.1 0.7ab 29.2 0.8ab 30.3 0.8ab 31.0 0.8ab
MC4R KO RW 23.6 0.7cf 23.0 0.6cf 23.8 0.6cf 24.3 0.7cf 23.9 0.8c 24.4 0.7c 24.9 0.8cf 25.2 0.8cf
Body length (noseto anus, mm)
WT Conv 83.2 0.5 82.5 0.4 83.3 0.5 85.2 0.6 85.6 0.6 86.1 0.6 87.0 0.5 89.3 0.4WT RW 81.6 0.5e 83.1 0.5e 83.3 0.6e 84.4 0.5 85.2 0.5 85.7 0.4 86.4 0.4e 88.0 0.7e
MC4R KO Conv 86.3 0.8a 85.6 0.4a 86.8 0.5a 86.6 0.6 87.6 1.0 88.2 0.5 89.9 0.5a 92.9 0.4ab
MC4R KO RW 84.3 1.3f 84.6 0.7 86.0 0.9 85.9 0.8 86.0 1.2 86.6 0.8 88.7 0.8 89.4 0.8Fat mass (% body
weight)WT Conv 7 1 8 1 8 1 8 1 8 1 9 1 9 1 9 1WT RW 7 1e 7 1e 7 1e 7 1e 7 1e 8 1e 7 1e 7 1e
MC4R KO Conv 11 1a 14 2ab 15 2ab 18 2ab 19 2ab 21 2ab 23 2ab 23 2ab
MC4R KO RW 8 1 10 1 10 1 10 1 9 1 11 1 11 1f 11 1Lean mass (%
body weight)WT Conv 79 1 80 1 79 1 79 1 79 1 80 1 81 1 78 1WT RW 78 2e 81 2e 79 1e 80 1e 79 1e 80 1e 77 1e 78 1e
MC4R KO Conv 74 2a 75 2ab 72 2ab 70 1ab 70 1ab 69 1ab 68 2ab 66 2ab
MC4R KO RW 74 1cf 79 1 77 1 77 1 79 1 79 1 78 1 76 1Cholesterol
(mg/dl)WT Conv 169 8 156 7 169 9 148 8 158 8 163 7 157 8 176 9WT RW 137 8de 132 9de 137 10de 134 9e 126 8de 130 8de 126 8de 140 11de
MC4R KO Conv 184 10b 192 13ab 185 8 180 12a 197 15ab 200 14ab 190 13a 207 11a
MC4R KO RW 156 9 158 5 173 5f 167 10 165 7 172 9f 167 7f 183 12f
Nonfasted glucose(mg/dl)
WT Conv 92 3 99 4 102 2 94 2 97 3 107 2 96 3 113 5WT RW 90 7 107 3 108 4 109 2d 109 2 107 5 100 2 115 4e
MC4R KO Conv 98 3 108 3 116 7 112 5a 108 10 108 5 108 5 144 7ab
MC4R KO RW 83 3 114 6 108 7 105 2 104 3 104 3 99 4 117 5Insulin (pM)
WT Conv 104 6 103 5 86 4 82 5 83 4 122 7 90 6 98 9WT RW 136 16 140 12e 118 16e 96 10e 135 14e 143 8e 110 10e 106 8e
MC4R KO Conv 167 14ab 208 13ab 180 16ab 181 15ab 238 22ab 250 20ab 308 28ab 327 45ab
MC4R KO RW 110 8 131 6c 117 6c 112 9c 132 12c 140 12 121 11 131 13Leptin (pM)
WT Conv 61 5 56 7 65 6 56 6 61 4 99 8 64 5 60 10WT RW 50 6e 60 9e 47 10e 48 7e 56 7e 80 3e 54 6e 64 6e
MC4R KO Conv 227 29ab 322 36ab 345 42ab 324 32ab 407 40ab 500 54ab 454 45ab 602 85ab
MC4R KO RW 89 11 114 11 78 9 88 8 97 8 140 11 104 17 138 19
Values indicate the average se of n � 10–12 mice/group. Conventionally housed mice were housed 2 mice/cage. Statistical analysis wasperformed using repeated-measures analysis with the Bonferroni post hoc test for the duration of the experimental weeks (see Table 2).Statistically significant (P�0.05) differences between groups were observed as indicated by footnotes. Conv, conventional housing; RW, runningwheel housing; WT, wild type. aWT Conv vs. MC4R KO Conv. bMC4R KO Conv vs. MC4R KO RW. cMC4R KO RW vs. WT Conv. dWT Conv vs.WT RW. eMC4R KO Conv vs. WT RW. fMC4RKO RW vs. WT RW.
646 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal
to the observed change in body length seen betweenthe MC4R KO and wild-type running wheel-housedmice (Table 1). MC4R KO conventionally housed miceare known to show increased linear growth comparedwith wild-type mice (1). Consistent with previous re-ports (1), we observed statistically significant differ-ences in body length between the MC4R KO mice andwild-type littermates housed conventionally. Compari-son of the MC4R KO sedentary vs. running wheel-housed groups showed a trend for the exercised micepossessing body lengths intermediate between those ofthe conventionally housed MC4R KO mice and thewild-type littermates. However, this difference was sta-tistically significant in the MC4R KO groups only in the
last week of the experiment. Analysis comparing bodylength (mm) with total body weight (g) and lean mass(g) correlated well for all groups (r�0.9, P�0.0001).
Consistent with results from our previous study (55),both the wild-type and MC4R KO mice housed inrunning wheel cages possessed similar food intake andactivity patterns with the exception of wk 1 food intake,when the MC4R KO mice consumed less on averagethen the wild-type littermates (Table 1). These initialdifferences could be attributed to differences in accli-mation behavior or to the stress differences observed inmice possessing disruptions in the melanocortin system(60). Consistent with previous reports, for the durationof the experiment, the sedentary MC4R KO mice were
TABLE 2. Repeated measures statistical analysis of the physiological parameters measured in this study
Parameter T T � G T � E T � G � E
Average daily foodintake F7,189 � 6.45, P � 0.001 F7,189 � 6.74, P � 0.001 F7,189 � 14.69, P � 0.001 F7,189 � 1.49, P � NS
Body weight F7,280 � 118, P � 0.001 F7,280 � 15.82, P � 0.001 F7,280 � 16.16, P � 0.001 F7,280 � 16.41, P � 0.001Body length F7,147 � 40.04, P � 0.001 F7,147 � 0.712, P � NS F7,147 � 1.74, P � NS F7,147 � 0.567, P � NSFat mass F7,280 � 51.86, P � 0.001 F7,280 � 26.28, P � 0.001 F7,280 � 27.94, P � 0.001 F7,280 � 14.85, P � 0.001Lean mass F7,280 � 10.45, P � 0.001 F7,280 � 2.81, P � 0.001 F7,280 � 5.13, P � 0.001 F7,280 � 8.18, P � 0.001Cholesterol F7,252 � 5.81, P � 0.001 F7,252 � 2.41, P � 0.05 F7,252 � 2.14, P � 0.05 F7,252 � 0.831, P � NSNonfasted glucose F7,140 � 15.04, P � 0.001 F7,140 � 1.49, P � NS F7,140 � 2.05, P � 0.053 F7,140 � 1.01, P � NSInsulin F7,462 � 11.49, P � 0.001 F7,462 � 10.15, P � 0.001 F7,462 � 9.16, P � 0.001 F7,462 � 5.89, P � 0.001Leptin F7,448 � 17.13, P � 0.001 F7,448 � 12.16, P � 0.001 F7,448 � 8.36, P � 0.001 F7,448 � 8.62, P � 0.001
Statistical analysis was performed using repeated measures analysis with Bonferroni post hoc test for the duration of the experimental weeksin terms of time (T) effects, time � genotype (T�G), time � exercise (T�E), and time � genotype � exercise (T�G�E) interactions. NS,nonsignificant.
Figure 2. MRI of a sedentary WT littermate witha body weight of 24 g and an MC4R KO mousewith a body weight of 57 g. Kidneys and fat arelabeled for reference. White/light gray indi-cates fat mass; darker gray indicates lean mass.
647VOLUNTARY EXERCISE AND THE MC4R KO MOUSE

generally hyperphagic compared with the sedentarywild-type littermate mice (P�0.05). The exercisingwild-type and MC4R KO littermate groups consumedmore food on average then the sedentary housedcontrol groups throughout the experiment (P�0.05).Feeding efficiency averaged over the 8-wk experimentwas not statistically different for the exercising MC4RKO (14.31.2 g/kcal�103) and wild-type (18.20.5g/kcal�103) littermate groups. However, feeding effi-ciency over the duration of the experiment was signif-icantly different for the conventionally housed MC4RKO (68.80.7 g/kcal�103) and wild-type (25.10.4g/kcal�103) (P�0.001) groups.
Blood glucose and cholesterol levels
In these studies, nonfasted whole-blood glucose levelswere examined in the different experimental groups.
Blood glucose data showed statistically significantchanges during wk 4 and 8 of the experiment (Table1). Plasma cholesterol values were also monitored forthe duration of the experiment. Cholesterol levels werereduced from those of their sedentary controls in bothexercised groups to a similar degree, and the choles-terol levels of the sedentary MC4R KO mice weresimilar to those of the sedentary wild-type littermatecontrol mice.
Blood insulin and leptin concentrations arenormalized with exercise in MC4R KO mice
We evaluated the average weekly plasma levels of thedifferent experimental groups in attempts to identifythe duration of exercise that becomes critical for pre-venting increased insulin and leptin levels. Significant
Figure 3. MRI of a representative littermate mouse from each experimental group over time to compare body fat distributionpatterns (white) and the effect of exercise on genotype. Indicated wk 7 age time point is after exposure to 1 wk of experimentaltreatment (sedentary vs. exercise). Week 11 is after exposure to 5 wk of treatment, and wk 14 is the age of the mice at thetermination of the experiment and exposure to experimental treatment for 8 wk.
648 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal

differences were observed after only 1 wk of exercisebetween the MC4R KO mice housed conventionally orin the presence of a running wheel (P�0.01). A secondnotable difference in insulin and leptin values for theconventionally housed MC4R KO mice occurred at wk5 of the experiment. Voluntary exercise by the MC4RKO mice prevented increased plasma insulin and leptinlevels and maintained values similar to those observedin the wild-type littermate mice. Plasma leptin (pM)values correlated with fat mass (g) (r�0.8 for MC4R KOmice, r�0.6 for wild-type mice, P�0.0001) for allgroups and with total body weight (g) (r�0.8 for MC4RKO conventionally housed, r�0.7 for MC4R KO exer-cised, and r�0.5 for wild-type exercised, P�0.0001;r�0.3 for the wild-type sedentary housed group,P�0.011). Plasma leptin (pM) values correlated well tolean mass (g) for the MC4R KO groups (r�0.5,P�0.0001) but not for the wild-type groups (P�0.13and 0.58). Plasma insulin (pM) also correlated withtotal body weight (g) for the MC4R KO groups (r�0.6for conventionally housed and r�0.5 for running wheelhoused mice, P�0.0001) but not for the wild-typegroups (P�0.42 and 0.76). Moreover, plasma insulin(pM) values correlated with fat mass (g) for both theMC4R KO groups (r�0.6 for conventionally housedand r�0.3 for running wheel housed mice, P�0.0001)and for the wild-type sedentary group (r�0.3, P�0.014)but not for the wild-type exercised group (P�0.82). Inaddition, comparison of plasma insulin (pM) and leanmass (g) correlated well for the MC4R KO sedentary
group (r�0.3, P�0.0018) and for the MC4R KO exer-cised group (r�0.4, P�0.0001) but not for the wild-typesedentary (P�0.84) or wild-type exercised (P�0.43)groups. An overall correlation was observed, comparingthe plasma insulin and leptin values of all groups(r�0.7, P�0.0001).
MC4R KO liver weights are normalized with exercisein MC4R KO mice
At the termination of the experiment, the heart, liver,brain, and kidneys were harvested and weighed. Onlyliver weights of the sedentary housed MC4R KO micewere found to be significantly different from those ofthe exercised MC4R KO mice and the wild-type litter-mate control groups (P�0.001) (Fig. 5). It is welldocumented for both humans and the MC4R KOmouse that increased body fat also results in increasedfatty livers (61).
MC4R KO prediabetic increase in pancreatic isletarea and �-cell mass is normalized by exercise
Because the MC4R KO mouse eventually developscharacteristic insulin resistance and hyperinsulinemiasimilar to human type 2 diabetes (1), we examined theeffect of voluntary exercise on the islet cell distributionand function in the pancreas of MC4R KO and wild-type littermate mice in our study. We examined thepancreatic islet cell histology and identified �-cell insu-lin (n�3–4 mice/group). Insulin staining indicatedthat all four experimental groups possessed similar�-cell numbers (data not shown) but total islet areadiffered between the MC4R KO conventionally housedmice (P�0.004) and the MC4R KO exercised and bothwild-type groups (Fig. 6). These data support thehypothesis that voluntary exercise of the MC4R KOmouse can prevent/delay the onset of type 2 diabetes.These data are consistent with the onset of a predia-betic state with increased circulating insulin levels
Figure 5. Collected tissue weights (mg) at the termination ofthe experiment. f, MC4R KO sedentary group; �, MC4R KORW group, s, WT littermate sedentary group; `, WT RWgroup. Values are averages se; n � 10–12 mice/group.***P � 0.001; two-way ANOVA with Bonferroni post hoc tests.
Figure 4. Effect of voluntary exercise on the ratio of lean to fatmass over time. Values are averages se; n � 10–12mice/group. f, MC4R KO sedentary group; �, MC4R KORW group; F, WT littermate control sedentary group; E, WTlittermate RW group. Statistical analysis was performed usingrepeated-measures analysis with a Bonferroni post hoc testfor the duration of the experimental weeks in terms oftime (F7,208�11.7, P�0.0001) effects, time � genotype(F7,208�1.69, NS), time � exercise (F7,208�3.93, P�0.0001),and time � genotype � exercise (F7,208�0.65, NS) interac-tions. NS, nonsignificant. Statistically significant differenceswere observed between the sedentary MC4R KO mice andMC4R KO exercising mice as well as both the wild-type groupsas indicated. *P � 0.05, **P � 0.01, ***P � 0.001.
649VOLUNTARY EXERCISE AND THE MC4R KO MOUSE
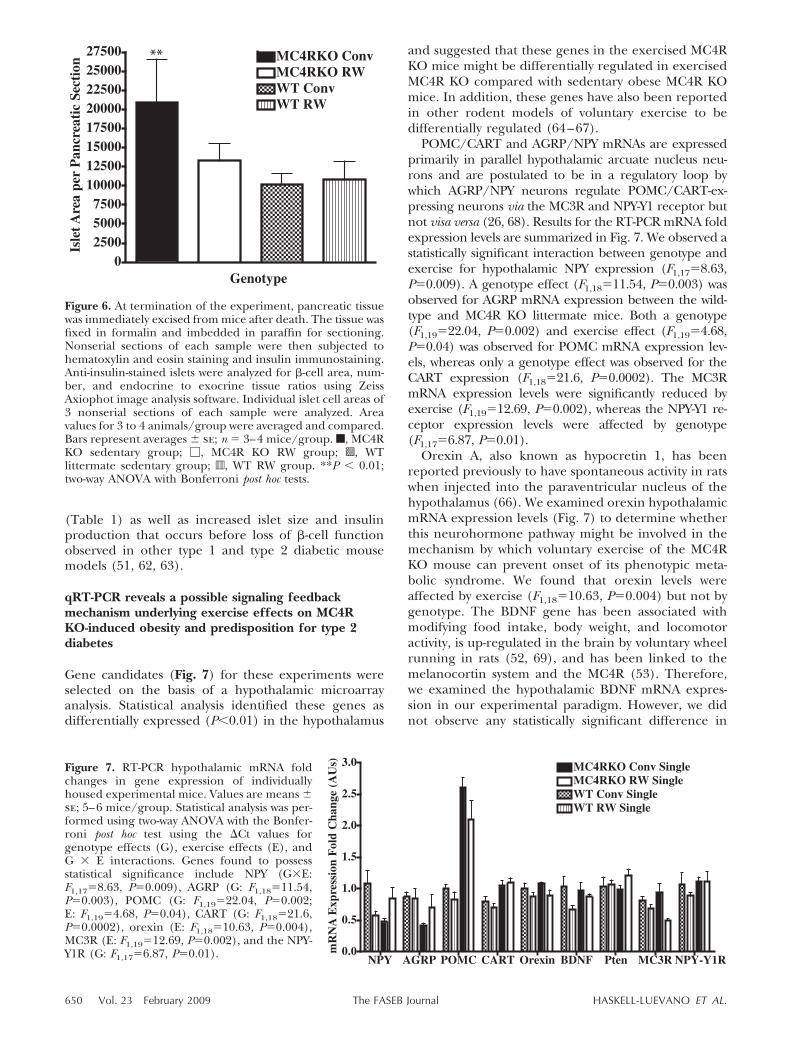
(Table 1) as well as increased islet size and insulinproduction that occurs before loss of �-cell functionobserved in other type 1 and type 2 diabetic mousemodels (51, 62, 63).
qRT-PCR reveals a possible signaling feedbackmechanism underlying exercise effects on MC4RKO-induced obesity and predisposition for type 2diabetes
Gene candidates (Fig. 7) for these experiments wereselected on the basis of a hypothalamic microarrayanalysis. Statistical analysis identified these genes asdifferentially expressed (P�0.01) in the hypothalamus
and suggested that these genes in the exercised MC4RKO mice might be differentially regulated in exercisedMC4R KO compared with sedentary obese MC4R KOmice. In addition, these genes have also been reportedin other rodent models of voluntary exercise to bedifferentially regulated (64–67).
POMC/CART and AGRP/NPY mRNAs are expressedprimarily in parallel hypothalamic arcuate nucleus neu-rons and are postulated to be in a regulatory loop bywhich AGRP/NPY neurons regulate POMC/CART-ex-pressing neurons via the MC3R and NPY-Y1 receptor butnot visa versa (26, 68). Results for the RT-PCR mRNA foldexpression levels are summarized in Fig. 7. We observed astatistically significant interaction between genotype andexercise for hypothalamic NPY expression (F1,17�8.63,P�0.009). A genotype effect (F1,18�11.54, P�0.003) wasobserved for AGRP mRNA expression between the wild-type and MC4R KO littermate mice. Both a genotype(F1,19�22.04, P�0.002) and exercise effect (F1,19�4.68,P�0.04) was observed for POMC mRNA expression lev-els, whereas only a genotype effect was observed for theCART expression (F1,18�21.6, P�0.0002). The MC3RmRNA expression levels were significantly reduced byexercise (F1,19�12.69, P�0.002), whereas the NPY-Y1 re-ceptor expression levels were affected by genotype(F1,17�6.87, P�0.01).
Orexin A, also known as hypocretin 1, has beenreported previously to have spontaneous activity in ratswhen injected into the paraventricular nucleus of thehypothalamus (66). We examined orexin hypothalamicmRNA expression levels (Fig. 7) to determine whetherthis neurohormone pathway might be involved in themechanism by which voluntary exercise of the MC4RKO mouse can prevent onset of its phenotypic meta-bolic syndrome. We found that orexin levels wereaffected by exercise (F1,18�10.63, P�0.004) but not bygenotype. The BDNF gene has been associated withmodifying food intake, body weight, and locomotoractivity, is up-regulated in the brain by voluntary wheelrunning in rats (52, 69), and has been linked to themelanocortin system and the MC4R (53). Therefore,we examined the hypothalamic BDNF mRNA expres-sion in our experimental paradigm. However, we didnot observe any statistically significant difference in
Figure 6. At termination of the experiment, pancreatic tissuewas immediately excised from mice after death. The tissue wasfixed in formalin and imbedded in paraffin for sectioning.Nonserial sections of each sample were then subjected tohematoxylin and eosin staining and insulin immunostaining.Anti-insulin-stained islets were analyzed for �-cell area, num-ber, and endocrine to exocrine tissue ratios using ZeissAxiophot image analysis software. Individual islet cell areas of3 nonserial sections of each sample were analyzed. Areavalues for 3 to 4 animals/group were averaged and compared.Bars represent averages se; n � 3–4 mice/group. f, MC4RKO sedentary group; �, MC4R KO RW group; s, WTlittermate sedentary group; `, WT RW group. **P � 0.01;two-way ANOVA with Bonferroni post hoc tests.
Figure 7. RT-PCR hypothalamic mRNA foldchanges in gene expression of individuallyhoused experimental mice. Values are means se; 5–6 mice/group. Statistical analysis was per-formed using two-way ANOVA with the Bonfer-roni post hoc test using the Ct values forgenotype effects (G), exercise effects (E), andG � E interactions. Genes found to possessstatistical significance include NPY (G�E:F1,17�8.63, P�0.009), AGRP (G: F1,18�11.54,P�0.003), POMC (G: F1,19�22.04, P�0.002;E: F1,19�4.68, P�0.04), CART (G: F1,18�21.6,P�0.0002), orexin (E: F1,18�10.63, P�0.004),MC3R (E: F1,19�12.69, P�0.002), and the NPY-Y1R (G: F1,17�6.87, P�0.01).
650 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal

comparing genotype or exercise in our experiment.Similar to BDNF, hypothalamic Pten mRNA expressionwas not found to be significantly different betweengenotype or exercise in this study.
Comparisons of conventionally group housed vs.singly housed phenotypes
To investigate whether differences between paired orsingly housed sedentary MC4R KO or wild-type litter-mate mice possessed similar or distinctly different phys-iological parameters or gene expression patterns in ourexperimental paradigm, we compared physiologicalparameters as well as hypothalamic gene expressionpatterns (Fig. 8). We observed that although the abso-lute values were slightly different, comparisons such aschanges in body weight (and other measured physio-logical parameters discussed in our experimental para-digm) over time between singly or paired convention-ally housed mice resulted in the same trends and werenot statistically significantly different among the samegenotype. These data are consistent with previous re-ports of the MC4R KO phenotype (1, 55, 70). Compar-isons of hypothalamic mRNA gene expression levelsalso were not statistically different between the singly orgroup conventionally housed mice.
DISCUSSION
Obesity, including obesity linked to genetic factors andhuman single nucleotide polymorphisms, is becomingmore prevalent in human societies. Exercise is a well-established method to prevent and/or reduce bodyweight and its effect is enhanced in the presence of aproper diet. Herein, we examined the MCR4R KO
mouse genetic model for obesity and type 2 diabetesand discovered that voluntary exercise at a young agecan prevent the physiological metabolic syndrome ob-served under sedentary housing conditions (1, 55).Several models for studying voluntary exercise of malerodents have been reported and reviewed (71). Thesemodels include the agouti Ay heterozygous mouse (72,73), which ectopically overexpresses the MC3R andMC4R antagonist agouti protein (74), the �-endorphinknockout mouse (which is derived from the POMCgene transcript) (75), the obese carboxypeptidease Eknockout mice (Cpefat/fat) mouse (76, 77), diet-inducedobesity or diet-resistant Sprague-Dawley rats (67, 78),Otsuka Long-Evans Tokushima fatty (OLETF) rats thatlack CCK-1 receptors (64, 79), wild-type C57BL/6Nmice (80), wild-type Sprague-Dawley rats (65, 78, 81),and Syrian Golden hamsters (82).
Exercise has been previously reported to slow theweight gain of the Ay mouse (72, 73, 83). Chiu et al.(73) examined the effects of diet restriction and volun-tary exercise for the obese agouti Ay [B6.Cg-Ay (Ay)]genetic mouse model starting at 15 wk of age for up to11 wk. Plasma leptin concentrations were �3-foldhigher in the sedentary Ay mice compared with thesedentary B6 control mice but were not decreased onexercise in either the Ay or control groups. After 8 wk ofdifferent experimental housing conditions, as antici-pated, the sedentary MC4R KO mice possessed 10-foldincreased plasma leptin values compared with thesedentary wild-type littermate controls (P�0.001) andpossessed 4-fold higher leptin plasma levels comparedwith the exercised MC4R KO mice (P�0.001), whichpossessed 2-fold higher levels than the sedentary con-trol group (Table 1). No difference was observedbetween the sedentary and exercised wild-type litter-mate control groups. Thus, the agouti Ay mouse, which
Figure 8. Comparison of Conv sedentary mice housed either in pairs or singly. A) Changes in body weight during theexperiment, starting at age 7 wk after exposure to 1 wk of experimental treatment. Statistical analysis was performed usingrepeated-measures analysis with the Bonferroni post hoc test . No statistical significance was observed between the paired or singlyhoused mice per genotype. B) Summary of the RT-PCR hypothalamic mRNA fold changes in gene expression of paired or singlyhoused Conv mice. Statistical analysis was performed using two-way ANOVA with the Bonferroni post hoc test. No statisticalsignificance was observed between the paired or singly housed mice per genotype.
651VOLUNTARY EXERCISE AND THE MC4R KO MOUSE

is arguably the same as an MC3R/MC4R double knock-out mouse by the ectopic expression of the agoutiantagonist for both these receptors (34, 74, 84) has adifferent response of leptin to exercise. Ay mice arelonger (anal-nasal length) then control B6 mice. Exer-cise was reported to increase body length in bothmodels. At the conclusion of this study, the sedentaryMC4R KO mice were 4% longer (nasal-anal length)then the sedentary wild-type littermate mice (P�0.001)and 4% longer then the exercised MC4R KO mice(P�0.01), which had the same body length as thesedentary wild-type control group (Table 1). SedentaryAy mice had higher total cholesterol levels then B6mice (64%) and the exercised Ay mice (14%). After 8wk of voluntary exercise, the sedentary MC4R KO micehad 18% increased total cholesterol levels comparedwith the sedentary wild-type littermate mice and 12%decreased cholesterol levels, similar to the sedentarywild-type control level, compared with the exercisedMC4R KO mice (Table 1). Exercised Ay and wild-typeB6 mice had similar running wheel activity profiles andconsumed approximately equivalent amounts of food,similar to the findings observed in this study for theMC4R KO and wild-type littermate control mice.
Levin et al. (78) studied the effects of voluntaryexercise (4 wk) as well as dietary fat composition onSprague-Dawley rats that developed diet-induced obe-sity (DIO) or diet resistance (DR). In this study, theyreported that exercising rats ate the same amounts asthe sedentary rats for both the DIO and DR phenotypesbut that exercising rats gained less weight over the 4-wkperiod in which they were allowed access to runningwheels and the exercising DIO rats had a significant63% reduction in weight gain over this period. Exercis-ing decreased leptin levels by 12–18% and insulin levelsby 21 and 14%, respectively in the DR and DIOphenotypes and resulted in decreased total fat padweights. In the studies reported herein with the MC4RKO mouse model, we observed similar food intakequantities and kilocalories between the exercising wild-type and MC4R KO littermate groups and a decrease inbody weight gain, comparing the exercised MC4R KOvs. the sedentary MC4R KO mice. Consistent withprevious reports (1), at the conclusion of this study,sedentary MC4R KO mice increased body weight by 36%.Exercise in the MC4R KO mice resulted in a 74% reduc-tion in weight gain compared to sedentary controls,whereas both the exercised and sedentary wild-type micehad identical weight gain profiles. Compared with thesedentary wild-type mice at the termination of the 8-wkexperiment, the sedentary MC4R KO mice had a 10-fold,900% increase (P�0.001) in plasma leptin concentrationand exercising MC4RKO mice had 4-fold, 77% (P�0.001)reduced leptin levels compared with those of the seden-tary MC4R KO mice. Comparison of the exercising wild-type and MC4R KO littermate mice showed a 54%(P�0.001) increase in plasma leptin values. In anotherstudy, Patterson et al. (67) examined the effects of selectedgene expression in several hypothalamic nuclei, studying
either voluntary exercise or food restriction in diet-in-duced obese rats. These authors, using the experimentalparadigm of 3 wk of voluntary exercise (and sedentarycontrols) postweaning, reported 27% lower MC3R and28% lower NPY-Y1R arcuate expression levels associatedwith exercising. With use of the experimental paradigm of8 wk of voluntary exercise postweaning, decreases of 16%(wild-type) and 46% (MC4R KO) were observed forMC3R hypothalamic gene expression (Fig. 8). The hypo-thalamic NPY-Y1R expression levels were modified bywild-type vs. MC4R KO genotypes (4–19% increase) be-tween the sedentary and exercising littermate mice.
Voluntary running wheel exercise of the obese andhyperphagic OLETF rats that lack CCK-1 receptors hasbeen reported to normalize food intake and bodyweight compared with sedentary control animals (64,79). These studies resulted in the discovery that inresponse to exercise, hypothalamic arcuate NPYmRNA levels were increased and arcuate POMClevels were not elevated (64). Interestingly, foodintake and body weight are normal in CCK-1 receptorknockout mice (85). An interesting difference be-tween the OLETF rats and CCK-1R knockout mice isthat the mice do not have increased dorsomedialhypothalamus NPY mRNA expression whereas theOLETF rats do (79), indicating the possibility of bothspecies-specific and genetic types of differences be-tween these two models.
Thus, there are several similar trends among thedifferent genetic and diet-induced obesity rat modelsand the MC4R KO genetic mouse model used herein inthat several common and general observations can beseen with the different models. However, using theagouti Ay and MC4R KO obese genetic models, as wellas other genetic rodent models such as the OLETF rats,genotype-specific differences have also been revealed.These data provide supporting experimental evidencethat genetic rodent models, possessing phenotypessimilar to the same human genetic modifications, areimportant for studying the effects of voluntary exerciseand changes in physiology and gene expression profilesthat might be genotype-specific and important fordevelopment of human therapeutic agents. In addi-tion, because the voluntary exercise field is relativelyyoung and research is increasing at a fast pace withrapidly improving technologies, the determination ofmechanisms associated with exercise cause and/oreffect are still being identified and the parametersthat define these factors are still being developed.Toward these goals, the study presented herein pro-vides further physiological characterization of thegenetic MC4R KO obese mouse model and theeffects of voluntary exercise over a 2-month durationand supports the hypothesis that voluntary exercisecan prevent the genetic predisposition of MC4R-associated obesity and diabetes.
This work was supported by an American Diabetes Associ-ation Research Award and National Institutes of Health grantRO1DK057080 (C.H.L.). We thank Dr. Dennis Huszar and
652 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal

Millennium Pharmaceuticals (Cambridge, MA, USA) for pro-viding the MC4RKO mice. We thank Dr. Maureen Keller-Wood, Dr. Charles Wood, and Dr. Hendrik Luesch for theexpert technical assistance and discussions regarding theRT-PCR experiments, Dr. Maureen Keller-Wood for assis-tance with the statistical analysis, and Dr. Martha Campbell-Thompson and the Pathology Molecular Core for help withpancreatic islet cell staining and quantitation.
REFERENCES
1. Huszar, D., Lynch, C. A., Fairchild-Huntress, V., Dunmore, J. H.,Smith, F. J., Kesterson, R. A., Boston, B. A., Fang, Q., Berkemeir,L. R., Gu, W., Cone, R. D., Campfield, L. A., and Lee, F. (1997)Targeted disruption of the melanocortin-4 receptor results inobesity in mice. Cell 88, 131–141
2. Vaisse, C., Clement, K., Guy-Grand, B., and Froguel, P. (1998) Aframeshift mutation in human MC4R is associated with adominant form of obesity. Nat. Genet. 20, 113–114
3. Yeo, G. S., Farooqi, I. S., Aminian, S., Halsall, D. J., Stanhope,R. G., and O’Rahilly, S. (1998) A frameshift mutation in MC4Rassociated with dominantly inherited human obesity. Nat. Genet.20, 111–112
4. Krude, H., Biebermann, H., Luck, W., Horn, R., Brabant, G.,and Gruters, A. (1998) Severe early-onset obesity, adrenalinsufficiency and red hair pigmentation caused by POMCmutations in humans. Nat. Genet. 19, 155–157
5. Krude, H., and Gruters, A. (2000) Implications of proopiomel-anocortin (POMC) mutations in humans: the POMC deficiencysyndrome. Trends Endocrinol. Metab. 11, 15–22
6. Hinney, A., Schmidt, A., Nottebom, K., Heibult, O., Becker, I.,Ziegler, A., Gerber, G., Sina, M., Gorg, T., Mayer, H., Siegfried, W.,Fichter, M., Remschmidt, H., and Hebebrand, J. (1999) Severalmutations in the melanocortin-4 receptor gene including a non-sense and a frameshift mutation associated with dominantly inher-ited obesity in humans. J. Clin. Endocrinol. Metab. 84, 1483–1486
7. Hinney, A., Remschmidt, H., and Hebebrand, J. (2000) Candi-date gene polymorphisms in eating disorders. Eur. J. Pharmacol.410, 147–159
8. Sina, M., Hinney, A., Ziegler, A., Neupert, T., Mayer, H.,Siegfried, W., Blum, W. F., Remschmidt, H., and Hebebrand, J.(1999) Phenotypes in three pedigrees with autosomal dominantobesity caused by haploinsufficiency mutations in the melano-cortin-4 receptor gene. Am. J. Hum. Genet. 65, 1501–1507
9. Farooqi, I. S., Yeo, G. S., Keogh, J. M., Aminian, S., Jebb, S. A.,Butler, G., Cheetham, T., and O’Rahilly, S. (2000) Dominantand recessive inheritance of morbid obesity associated withmelanocortin 4 receptor deficiency. J. Clin. Invest. 106, 271–279
10. Vaisse, C., Clement, K., Durand, E., Hercberg, S., Guy-Grand, B.,and Froguel, P. (2000) Melanocortin-4 receptor mutations are afrequent and heterogeneous cause of morbid obesity. J. Clin.Invest. 106, 253–262
11. Mergen, M., Mergen, H., Ozata, M., Oner, R., and Oner, C.(2001) A novel melanocortin 4 receptor (MC4R) gene mutationassociated with morbid obesity. J. Clin. Endocrinol. Metab. 86,3448–3451
12. Gu, W., Tu, Z., Kleyn, P. W., Kissebah, A., Duprat, L., Lee, J.,Chin, W., Maruti, S., Deng, N., Fisher, S. L., Franco, L. S., Burn,K., Yagaloff, K., Nathan, J., Heymsfield, S., Albu, J., Pi-Sunyer,F. X., and Allison, D. B. (1999) Identification and functionalanalysis of novel human melanocortin-4 receptor variants. Dia-betes 48, 635–639
13. Dubern, B., Clement, K., Pelloux, V., Froguel, P., Girardet, J. P.,Guy-Grand, B., and Tounian, P. (2001) Mutational analysis ofmelanocortin-4 receptor, agouti-related protein, and �-melano-cyte-stimulating hormone genes in severely obese children.J. Pediatr. 139, 204–209
14. Vink, T., Hinney, A., van Elburg, A. A., van Goozen, S. H.,Sandkuijl, L. A., Sinke, R. J., Herpertz-Dahlmann, B. M., Hebe-brand, J., Remschmidt, H., van Engeland, H., and Adan, R. A.(2001) Association between an agouti-related protein genepolymorphism and anorexia nervosa. Mol. Psychiatry 6, 325–328
15. Yeo, G. S., Lank, E. J., Farooqi, I. S., Keogh, J., Challis, B. G., andO’Rahilly, S. (2003) Mutations in the human melanocortin-4
receptor gene associated with severe familial obesity disruptsreceptor function through multiple molecular mechanisms.Hum. Mol. Genet. 12, 561–574
16. Lubrano-Berthelier, C., Durand, E., Dubern, B., Shapiro, A.,Dazin, P., Weill, J., Ferron, C., Froguel, P., and Vaisse, C.(2003) Intracellular retention is a common characteristic ofchildhood obesity-associated MC4R mutations. Hum. Mol.Genet. 12, 145–153
17. Farooqi, I. S., Keogh, J. M., Yeo, G. S., Lank, E. J., Cheetham, T.,and O’Rahilly, S. (2003) Clinical spectrum of obesity andmutations in the melanocortin 4 receptor gene. N. Engl. J. Med.348, 1085–1095
18. Roselli-Rehfuss, L., Mountjoy, K. G., Robbins, L. S., Mortrud,M. T., Low, M. J., Tatro, J. B., Entwistle, M. L., Simerly, R. B., andCone, R. D. (1993) Identification of a receptor for mela-notropin and other proopiomelanocortin peptides in the hypo-thalamus and limbic system. Proc. Natl. Acad. Sci. U. S. A. 90,8856–8860
19. Mountjoy, K. G., Mortrud, M. T., Low, M. J., Simerly, R. B., andCone, R. D. (1994) Localization of the melanocortin-4 receptor(MC4-R) in neuroendocrine and autonomic control circuits inthe brain. Mol. Endocrinol. 8, 1298–1308
20. Gantz, I., Konda, Y., Tashiro, T., Shimoto, Y., Miwa, H., Munzert,G., Watson, S. J., DelValle, J., and Yamada, T. (1993) Molecularcloning of a novel melanocortin receptor. J. Biol. Chem. 268,8246–8250
21. Gantz, I., Miwa, H., Konda, Y., Shimoto, Y., Tashiro, T., Watson,S. J., DelValle, J., and Yamada, T. (1993) Molecular cloning,expression, and gene localization of a fourth melanocortinreceptor. J. Biol. Chem. 268, 15174–15179
22. Konda, Y., Gantz, I., DelValle, J., Shimoto, Y., Miwa, H., andYamada, T. (1994) Interaction of dual signal transductionpathways activated by the melanocortin-3 receptor. J. Biol. Chem.269, 13162–13166
23. Daniels, D., Patten, C. S., Roth, J. D., Yee, D. K., and Fluharty,S. J. (2003) Melanocortin receptor signaling through mitogen-activated protein kinase in vitro and in rat hypothalamus. BrainRes. 986, 1–11
24. Ollmann, M. M., Wilson, B. D., Yang, Y.-K., Kerns, J. A., Chen, Y.,Gantz, I., and Barsh, G. S. (1997) Antagonism of centralmelanocortin receptors in vitro and in vivo by agouti-relatedprotein. Science 278, 135–138
25. Haskell-Luevano, C., and Monck, E. K. (2001) Agouti-relatedprotein (AGRP) functions as an inverse agonist at a constitu-tively active brain melanocortin-4 receptor. Regul. Pept. 99, 1–7
26. Proneth, B., Xiang, Z., Pogozheva, I. D., Litherland, S. A.,Gorbatyuk, O. S., Shaw, A. M., Millard, W. J., Mosberg, H. I., andHaskell-Luevano, C. (2006) Molecular mechanism of the con-stitutive activation of the L250Q human melanocortin-4 recep-tor polymorphism. Chem. Biol. Drug Des. 67, 215–229
27. Barsh, G. S., Farooqi, S., and O’Rahilly, S. (2000) Genetics ofbody-weight regulation. Nature 404, 644–651
28. Commuzzie, A. G., Hixson, J. E., Almasy, L., Mitchell, B. D.,Mahaney, M. C., Dyer, T. D., Stern, M. P., MacCluer, J. W., andBlangero, J. (1997) A major quantitative trait locus determiningserum leptin levels and fat mass is located on human chromo-some 2. Nat. Genet. 15, 273–276
29. Hager, J., Dina, C., Francke, S., Dubois, S., Houari, M., Vatin, V.,Vaillant, E., Lorentz, N., Basdevant, A., Clement, K., Guy-Grand,B., and Froguel, P. (1998) A genome-wide scan for humanobesity genes reveals a major susceptibility locus on chromo-some 10. Nat. Genet. 20, 304–308
30. Yaswen, L., Diehl, N., Brennan, M. B., and Hochgeschwender,U. (1999) Obesity in the mouse model of pro-opiomelanocortindeficiency responds to peripheral melanocortin. Nat. Med. 5,1066–1070
31. Chen, A. S., Marsh, D. J., Trumbauer, M. E., Frazier, E. G.,Guan, X. M., Yu, H., Rosenblum, C. I., Vongs, A., Feng, Y., Cao,L., Metzger, J. M., Strack, A. M., Camacho, R. E., Mellin, T. N.,Nunes, C. N., Min, W., Fisher, J., Gopal-Truter, S., MacIntyre,D. E., Chen, H. Y., and Van Der Ploeg, L. H. (2000) Inactivationof the mouse melanocortin-3 receptor results in increased fatmass and reduced lean body mass. Nat. Genet. 26, 97–102
32. Butler, A. A., Kesterson, R. A., Khong, K., Cullen, M. J., Pelley-mounter, M. A., Dekoning, J., Baetscher, M., and Cone, R. D.(2000) A unique metabolic syndrome causes obesity in the mela-nocortin-3 receptor-deficient mouse. Endocrinology 141, 3518–3521
653VOLUNTARY EXERCISE AND THE MC4R KO MOUSE

33. Graham, M., Shutter, J. R., Sarmiento, U., Sarosi, I., and Stark,K. L. (1997) Overexpression of AGRT leads to obesity intransgenic mice. Nat. Genet. 17, 273–274
34. Lu, D., Willard, D., Patel, I. R., Kadwell, S., Overton, L., Kost, T.,Luther, M., Chen, W., Yowchik, R. P., Wilkison, W. O., andCone, R. D. (1994) Agouti protein is an antagonist of themelanocyte-stimulating-hormone receptor. Nature 371, 799–802
35. Fan, W., Boston, B. A., Kesterson, R. A., Hruby, V. J., and Cone,R. D. (1997) Role of melanocortinergic neurons in feeding andthe agouti obesity syndrome. Nature 385, 165–168
36. Rossi, M., Kim, M. S., Morgan, D. G., Small, C. J., Edwards, C. M.,Sunter, D., Abusnana, S., Goldstone, A. P., Russell, S. H.,Stanley, S. A., Smith, D. M., Yagaloff, K., Ghatei, M. A., andBloom, S. R. (1998) A C-terminal fragment of agouti-relatedprotein increases feeding and antagonizes the effect of �-mela-nocyte stimulating hormone in vivo. Endocrinology 139, 4428–4431
37. Hagan, M. M., Rushing, P. A., Pritchard, L. M., Schwartz, M. W.,Strack, A. M., Van Der Ploeg, L. H., Woods, S. C., and Seeley,R. J. (2000) Long-term orexigenic effects of AGRP-(83–132)involve mechanisms other than melanocortin receptor block-ade. Am. J. Physiol. Regul. Integr. Comp. Physiol. 279, R47–R52
38. Marsh, D. J., Hollopeter, G., Huszar, D., Laufer, R., Yagaloff,K. A., Fisher, S. L., Burn, P., and Palmiter, R. D. (1999)Response of melanocortin-4 receptor-deficient mice to anorec-tic and orexigenic peptides. Nat. Genet. 21, 119–122
39. Marsh, D. J., Miura, G. I., Yagaloff, K. A., Schwartz, M. W., Barsh,G. S., and Palmiter, R. D. (1999) Effects of neuropeptide Ydeficiency on hypothalamic agouti-related protein expressionand responsiveness to melanocortin analogues. Brain Res. 848,66–77
40. Grill, H. J., Ginsberg, A. B., Seeley, R. J., and Kaplan, J. M.(1998) Brainstem application of melanocortin receptor ligandsproduces long-lasting effects on feeding and body weight.J. Neurosci. 18, 10128–10135
41. Elias, C. F., Lee, C., Kelly, J., Aschkenasi, C., Ahima, R. S.,Couceyro, P. R., Kuhar, M. J., Saper, C. B., and Elmquist, J. K.(1998) Leptin activates hypothalamic CART neurons projectingto the spinal cord. Neuron 21, 1375–1385
42. Bagnol, D., Lu, X. Y., Kaelin, C. B., Day, H. E., Ollmann, M.,Gantz, I., Akil, H., Barsh, G. S., and Watson, S. J. (1999)Anatomy of an endogenous antagonist: relationship betweenagouti-related protein and proopiomelanocortin in brain.J. Neurosci. 19, RC26
43. Jegou, S., Boutelet, I., and Vaudry, H. (2000) Melanocortin-3receptor mRNA expression in pro-opiomelanocortin neurons ofthe rat arcuate nucleus. J. Neuroendocrinol. 12, 501–505
44. Baskin, D., Breininger, J., and Schwartz, M. W. (1999) Leptinreceptor mRNA identifies a subpopulation of neuropeptide Yneurons activated by fasting in rat hypothalamus. Diabetes 48,828–833
45. Cheung, C. C., Clifton, D. K., and Steiner, R. A. (1997)Proopiomelanocortin neurons are direct targets for leptin inthe hypothalamus. Endocrinology 138, 4489–4492
46. Broberger, C., Landry, M., Wong, H., Walsh, J. N., and Hokfelt,T. (1997) Subtypes Y1 and Y2 of the neuropeptide Y receptorare respectively expressed in pro-opiomelanocortin- and neu-ropeptide-Y containing neurons of the rat hypothalamic arcuatenucleus. Neuroendocrinology 66, 393–408
47. Hahn, T. M., Breininger, J. F., Baskin, D. G., and Schwartz,M. W. (1998) Coexpression of AGRP and NPY in fasting-activated hypothalamic neurons. Nat. Neurosci. 1, 271–272
48. Broberger, C., Johansen, J., Johansson, C., Schalling, M., andHokfelt, T. (1998) The neuropeptide Y/agouti gene-relatedprotein (AGRP) brain circuitry in normal, anorectic, andmonosodium glutamate-treated mice. Proc. Natl. Acad. Sci.U. S. A. 95, 15043–15048
49. Ning, K., Miller, L. C., Laidlay, H. A., Burgess, L. A., Perera,N. M., Downes, C. P., Leslie, N. R., and Ashford, M. L. J. (2006)A novel leptin signalling pathway via PTEN inhibition in hypo-thalamic cell lines and pancreatic �-cells. EMBO J. 25, 2377–2387
50. White, M. F. (2003) Insulin signaling in health and disease.Science 302, 1710–1711
51. Nguyen, K. T., Tajmir, P., Lin, C. H., Liadis, N., Zhu, X. D.,Eweida, M., Tolasa-Karaman, G., Cai, F., Wang, R., Kitamura, T.,Belsham, D. D., Wheeler, M. B., Suzuki, A., Mak, T. W., and
Woo, M. (2006) Essential role of Pten in body size determina-tion and pancreatic �-cell homeostasis in vivo. Mol. Cell. Biol. 26,4511–4518
52. Kernie, S. G., Liebl, D. J., and Parada, L. F. (2000) BDNFregulates eating behavior and locomotor activity in mice. EMBOJ. 19, 1290–1300
53. Xu, B., Goulding, E. H., Zang, K., Cepoi, D., Cone, R. D., Jones,K. R., Tecott, L. H., and Reichardt, L. F. (2003) Brain-derivedneurotrophic factor regulates energy balance downstream ofmelanocortin-4 receptor. Nat. Neurosci. 6, 736–742
54. Butler, A. A., Marks, D. L., Fan, W., Kuhn, C. M., Bartolome, M.,and Cone, R. D. (2001) Melanocortin-4 receptor is required foracute homeostatic responses to increased dietary fat. Nat. Neu-rosci. 4, 605–611
55. Irani, B. G., Xiang, Z., Moore, M. C., Mandel, R. J., andHaskell-Luevano, C. (2005) Voluntary exercise delays monoge-netic obesity and overcomes reproductive dysfunction of themelanocortin-4 receptor knockout mouse. Biochem. Biophys. Res.Commun. 326, 638–644
56. Li, J. Y., Lescure, P. A., Misek, D. E., Lai, Y. M., Chai, B. X.,Kuick, R., Thompson, R. C., Demo, R. M., Kurnit, D. M.,Michailidis, G., Hanash, S. M., and Gantz, I. (2002) Fooddeprivation-induced expression of minoxidil sulfotransferase inthe hypothalamus uncovered by microarray analysis. J. Biol.Chem. 277, 9069–9076
57. Taicher, G. Z., Tinsley, F. C., Reiderman, A., and Heiman, M. L.(2003) Quantitative magnetic resonance (QMR) method forbone and whole body composition analysis. Anal. Bioanal. Chem.377, 990–1002
58. Tinsley, F. C., Taicher, G. Z., and Heiman, M. L. (2004)Evaluation of a quantitative magnetic resonance method formouse whole body composition analysis. Obes. Res. 12, 150–160
59. Kirk, R. E. (1968) Experimental Design: Procedures for the BehavioralSciences, Brooks and Cole, Belmont, CA, USA
60. De Souza, J., Butler, A. A., and Cone, R. D. (2000) Dispropor-tionate inhibition of feeding in A(y) mice by certain stressors: acautionary note. Neuroendocrinology 72, 126–132
61. Sutton, G. M., Trevaskis, J. L., Hulver, M. W., McMillan, R. P.,Markward, N. J., Babin, M. J., Meyer, E. A., and Butler, A. A.(2006) Diet-genotype interactions in the development of theobese, insulin-resistant phenotype of C57BL/6J mice lackingmelanocortin-3 or -4 receptors. Endocrinology 147, 2183–2196
62. Baetens, D., Stefan, Y., Ravazzola, M., Malaisse-Lagae, F.,Coleman, D. L., and Orci, L. (1978) Alteration of islet cellpopulations in spontaneously diabetic mice. Diabetes 27, 1–7
63. Garris, D. R., and Garris, B. L. (2004) Cytochemical analysis ofpancreatic islet hypercytolipidemia following diabetes (db/db)and obese (ob/ob) mutation expression: influence of genomicbackground. Pathobiology 71, 231–240
64. Bi, S., Scott, K. A., Hyun, J., Ladenheim, E. E., and Moran, T. H.(2005) Running wheel activity prevents hyperphagia and obesityin Otsuka Long-Evans Tokushima fatty rats: role of hypotha-lamic signaling. Endocrinology 146, 1676–1685
65. Kawaguchi, M., Scott, K. A., Moran, T. H., and Bi, S. (2005)Dorsomedial hypothalamic corticotropin-releasing factor medi-ation of exercise-induced anorexia. Am. J. Physiol. Regul. Integr.Comp. Physiol. 288, R1800–R1805
66. Kiwaki, K., Kotz, C. M., Wang, C., Lanningham-Foster, K., andLevine, J. A. (2004) Orexin A (hypocretin 1) injected intohypothalamic paraventricular nucleus and spontaneous phys-ical activity in rats. Am. J. Physiol. Endocrinol. Metab. 286,E551–E559
67. Patterson, C. M., Bunn-Meynell, A. A., and Levin, B. E. (2008)Three weeks of early onset exercise prolongs obesity-resistancein DIO rats after exercise cessation. Am. J. Physiol. Regul. Integr.Comp. Physiol. 294, R290–R301
68. Cowley, M. A., Smart, J. L., Rubinstein, M., Cerdan, M. G.,Diano, S., Horvath, T. L., Cone, R. D., and Low, M. J. (2001)Leptin activates anorexigenic POMC neurons through a neuralnetwork in the arcuate nucleus. Nature 411, 480–484
69. Oliff, H. S., Berchtold, N. C., Isackson, P., and Cotman, C. W.(1998) Exercise-induced regulation of brain-derived neurotro-phic factor (BDNF) transcripts in the rat hippocampus. BrainRes. Mol. Brain Res. 61, 147–153
70. Chen, A. S., Metzger, J. M., Trumbauer, M. E., Guan, X., Yu, H.,Fraizer, E. G., Marsh, D., Forrest, M. J., Gopal-Truter, S., Fisher,J., Camacho, R. E., Strack, A., Mellin, T. N., MacIntyre, D. E.,
654 Vol. 23 February 2009 HASKELL-LUEVANO ET AL.The FASEB Journal

Chen, H. Y., and Van der Ploeg, L. (2000) Role of themelanocortin-4 receptor in metabolic rate and food intake inmice. Transgenic Res. 9, 145–154
71. Dishman, R. K., Berthoud, H. R., Booth, F. W., Cotman, C. W.,Edgerton, V. R., Fleshner, M. R., Gandevia, S. C., Gomez-Pinilla,F., Greenwood, B. N., Hillman, C. H., Kramer, A. F., Levin, B. E.,Moran, T. H., Russo-Neustadt, A. A., Salamone, J. D., VanHoomissen, J. D., Wade, C. E., York, D. A., and Zigmond, M. J.(2006) Neurobiology of exercise. Obesity 14, 345–356
72. Goodrick, C. L. (1978) Effect of voluntary wheel exercise onfood intake, water intake, and body weight for C57BL/6J miceand mutations which differ in maximal body weight. Physiol.Behav. 21, 345–351
73. Chiu, S., Fisler, J. S., Espinal, G. M., Havel, P. J., Stern, J. S., andWarden, C. H. (2004) The yellow agouti mutation alters some butnot all responses to diet and exercise. Obes. Res. 12, 1243–1255
74. Bultman, S. J., Michaud, E. J., and Woychick, R. P. (1992) Molec-ular characterization of the mouse agouti locus. Cell 71, 1195–1204
75. Koehl, M., Meerlo, P., Gonzales, D., Rontal, A., Turek, F. W., andAbrous, D. N. (2008) Exercise-induced promotion of hippocampalcell proliferation requires �-endorphin. FASEB J. 22, 2253–2262
76. Che, F. Y., Yuan, Q., Kalinina, E., and Fricker, L. D. (2005)Peptidomics of Cpefat/fat mouse hypothalamus: effect of food depri-vation and exercise on peptide levels. J. Biol. Chem. 280, 4451–4461
77. Yuan, Q., Fontenele-Neto, J. D., and Fricker, L. D. (2004) Effectof voluntary exercise on genetically obese Cpefat/fat mice: quan-titative proteomics of serum. Obes. Res. 12, 1179–1188
78. Levin, B. E., and Dunn-Meynell, A. A. (2006) Differential effectsof exercise on body weight gain and adiposity in obesity-proneand -resistant rats. Int. J. Obes. (Lond.) 30, 722–727
79. Moran, T. H., and Bi, S. (2006) Hyperphagia and obesity inOLETF rats lacking CCK-1 receptors. Philos. Trans. R. Soc. Lond.B Biol. Sci. 361, 1211–1218
80. Droste, S. K., Gesing, A., Ulbricht, S., Muller, M. B., Linthorst,A. C., and Reul, J. M. (2003) Effects of long-term voluntaryexercise on the mouse hypothalamic-pituitary-adrenocorticalaxis. Endocrinology 144, 3012–3023
81. Narath, E., Skalicky, M., and Viidik, A. (2001) Voluntary andforced exercise influence the survival and body composition ofageing male rats differently. Exp. Gerontol. 36, 1699–1711
82. Coutinho, A. E., Fediuc, S., Campbell, J. E., and Riddell, M. C.(2006) Metabolic effects of voluntary wheel running in youngand old Syrian Golden hamsters. Physiol. Behav. 87, 360–367
83. Carpenter, K., and Mayer, J. (1958) Physiologic observations onyellow obesity in the mouse. Am. J. Physiol. 193, 499–504
84. McNulty, J. C., Jackson, P. J., Thompson, D. A., Chai, B., Gantz,I., Barsh, G. S., Dawson, P. E., and Millhauser, G. L. (2005)Structures of the agouti signaling protein. J. Mol. Biol. 346,1059–1070
85. Kopin, A. S., Mathes, W. F., McBride, E. W., Nguyen, M.,Al-Haider, W., Schmitz, F., Bonner-Weir, S., Kanarek, R., andBeinborn, M. (1999) The cholecystokinin-A receptor mediatesinhibition of food intake yet is not essential for the maintenanceof body weight. J. Clin. Invest. 103, 383–391
Received for publication March 21, 2008.Accepted for publication September 25, 2008.
655VOLUNTARY EXERCISE AND THE MC4R KO MOUSE




















