
Shape and volume of craniofacial cavities in intentional skull deformations
Upload
independentCategory
view
0download
0
Original Research Article
Variation and Causal Factors of Craniofacial Robusticityin Patagonian Hunter-Gatherers from the Late Holocene
VALERIA BERNAL,* S. IVAN PEREZ, AND PAULA N. GONZALEZCONICET, Division Antropologıa, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata,La Plata 1900, Argentina
ABSTRACT Fueguian-Patagonian skulls have been characterized as some of the most ro-bust of any modern crania. However, the causal factors of such robusticity remain unsettled. Weassess within- and among-sample cranial robusticity of seven samples from continental Patago-nia and Tierra del Fuego, using geometric morphometric techniques. In addition, the biome-chanical, phylogenetic, and climatic hypotheses proposed to account for robusticity in such sam-ples are discussed. Two Amerindian samples of farmers and two early middle Holocene samplesfrom South America were included. The results show: 1) large variation in craniofacial robustic-ity among Patagonian samples, with the highest robusticity in samples from south continentalPatagonia and Isla Grande of Tierra del Fuego, whereas central and north Patagonian samplesdisplay the same degree of robusticity as farmer samples; 2) that early middle Holocene samplesdisplay lower levels of robusticity than South Patagonian samples; and 3) strong associationbetween latitude and craniofacial robusticity, with the most robust craniofacial morphologiesoccurring at the highest latitudes. In consequence, neither masticatory stress nor retention ofancestral features is supported by the morphological evidence analyzed. Hence it is hypothe-sized that endocrine changes related to cold climate may be a plausible explanation for severalcraniofacial features found in Fueguian and south continental Patagonian samples, such astheir large masticatory component, and pronounced supraorbital ridge and glabellar region.Am. J. Hum. Biol. 18:748–765, 2006. ' 2006 Wiley-Liss, Inc.
The craniofacial morphology of late Holo-cene (ca. 3,000 years 14C BP to historicaltimes) hunter-gatherers from continental Pa-tagonia and Tierra del Fuego presents somefeatures that depart from the majority ofAmerindian samples (Gonzalez-Jose, 2003;Lahr, 1995; Lalueza et al., 1997; Sardi, 2002).In terms of cranial dimensions, the remainsfrom this area have been characterized ashaving a very tall and broad face, very tallorbits, and a very broad cranial base (Lahr,1995). Likewise, the Fueguian-Patagonianshave been considered a relatively homogene-ous group in morphological terms (Lahr andWright, 1996), and their skulls have been con-sidered among the most robust of any moderncrania, due to the pronounced development ofthe supraorbital ridge, glabellar region, andsuperciliary arches (Gonzalez-Jose, 2003; Lahrand Wright, 1996). Several hypotheses wereproposed to explain this pattern of robusticity,of which the most frequently mentioned arethe biomechanical and phylogenetic hypothe-ses (Hernandez et al., 1997; Lahr, 1995; Lahr
and Wright, 1996; Lalueza et al., 1997). Thebiomechanical hypothesis argues that cranio-facial robusticity in the morphology of thesepopulations is a phenotypic response duringontogeny (i.e., phenotypic plasticity) to high-magnitude forces due to masticatory and para-masticatory activity resulting from hard dietsand the use of the mouth as a tool. The phylo-genetic hypothesis states that the highly dis-tinct pattern of robusticity of Fueguian-Patago-nian samples is the product of long-standingisolation, which, through random genetic drift,resulted in the retention of ‘‘ancestral’’ features
Grant sponsor: Antorchas Foundation; Grant number:14116-111.
*Correspondence to: Valeria Bernal, Division Antropolo-gıa, Facultad de Ciencias Naturales y Museo, Museo de LaPlata, Paseo del Bosque s/n, La Plata 1900, Argentina.E-mail: [email protected]
Received 26 December 2005; Revision received 29 March2006; Accepted 31 March 2006
Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/ajhb.20546
AMERICAN JOURNAL OF HUMAN BIOLOGY 18:748–765 (2006)
VVC 2006 Wiley-Liss, Inc.

that had been common in Pleistocene crania.However, these hypotheses have not been sys-tematically evaluated, so the causal factors ofPatagonian craniofacial robusticity remainunsettled. On the other hand, numerous fea-tures that result from adaptation to a cold cli-mate characterize populations living at lowtemperatures. For instance, the shape of thenasal aperture (Gonzalez-Jose, 2003; Hernan-dez et al., 1997), the body proportions, and thepostcranial robusticity (Pearson and Millones,2005) of Fueguians were hypothesized to be cli-matic responses. Considering the relevance ofclimatic factors to explain morphological varia-tion, the climatic hypothesis is proposed hereas a causal factor of craniofacial robusticity inFueguian-Patagonian populations.The term robusticity is used in the study of
craniofacial variation to describe a diversearray of morphological traits (Pearson, 2000).This quality is attributed to crania possessinglarge superstructures, such as brow ridges,occipital crests, occipital tori, mastoid pro-cesses, nuchal crests (Lahr and Wright, 1996),large external dimensions, thick vault bones(Lieberman, 1996), rugged muscle markings,or some nonspecific combination of these fea-tures. Thus defined, the term robusticity re-fers to form (i.e., size and shape). The relation-ship between increased robustness and largersize is unclear, as some authors found strongrelationships between both variables (e.g., Lahrand Wright, 1996), whereas others found thatpopulations with larger crania are not alwaysthe more robust (e.g., Hawks et al., 2000).The characters employed to describe cranial
robusticity have been evaluated mainly bymeans of linear measurements and nonmetrictraits such as the degree of development oftubercles, ridges, and tori (e.g., Brown andMaeda, 2004; Lahr and Wright, 1996; O’Con-nor et al., 2005), with the last being more com-monly used to describe cranial robusticity; thedifferences are expressed in terms of ‘‘more’’or ‘‘less’’ robust morphologies (e.g., Ahernet al., 2004). One problem with approachesbased on nonmetric traits is the high degree ofinter- and intraobserver subjectivity of thesequalitative variables; therefore, a comprehen-sive overview of methods for quantifying cra-nial robusticity is still necessary (Petersen,2000). Pearson (2000) proposed the calculationof a ‘‘common robusticity factor’’ by means ofmultivariate analyses, such as principal com-ponent or factor analysis, to analyze skeletalrobustness. However, the use of traditionalmorphometric techniques (Marcus, 1990; Rey-
ment et al., 1984) has numerous limitations,e.g., the spatial relationships between varia-bles are usually lost, and both components ofform (i.e., size and shape) cannot be satis-factorily analyzed. In this sense, geometricmorphometric techniques are superior to tra-ditional ones for finding axes of cranial ro-busticity, because they combine methods thatcapture the geometry of morphological struc-tures of interest, and preserve this informa-tion throughout multivariate analyses (Adamset al., 2004; Oxnard, 1978). Thus, geometricmorphometric techniques allow a better de-scription of shape than traditional morpho-metric techniques (Rohlf and Marcus, 1993),because they use powerful statistical methodsbased on models for shape variation of anentire configuration of points corresponding tothe locations of morphological landmarks andsemilandmarks (Bookstein, 1991, 1997; Rohlf,1999, 2000).1 Likewise, this approach permitsus to quantify the variation and covariationbetween traits, allowing an integrated analy-sis of human robustness, in contrast to ap-proaches that tend to segregate the organisminto discrete units affected by different evolu-tionary forces (Churchill, 1998).The purpose of this paper is to assess the
cranial robusticity (defined as the degree ofdevelopment of cranial vault superstructuresand muscle attachment sites) of several sam-ples from continental Patagonia and Tierradel Fuego, using geometric morphometric tech-niques. Our general goals are to study the var-iation of craniofacial robusticity within andamong Fueguian-Patagonian samples, and todiscuss the influence of hypothesized causesof robusticity such as biomechanical, phylo-genetic, and climatic factors. More particulargoals include: 1) analysis of variation in cra-niofacial robusticity within and among Fue-guian-Patagonian samples in order to assessits homogeneity for this region; 2) assessmentof correlation between robustness and size, atintra- and intersample levels; and 3) evalua-tion of biomechanical factors by means of com-parison of samples from populations with dif-ferent diets: hunter-gatherers from Patagonia,who consumed harder foods, and farmers fromcentral-west and northwestern Argentina, whoconsumed softer foods. If the robustness werecaused by high masticatory stress, a greater
1Landmarks refer to two- or three-dimensional coordi-nates of biologically definable points. Semilandmarks referto point coordinates distributed evenly according to a givenalgorithm along a contour (Bookstein, 1991, 1997).
749CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

degree of craniofacial robustness would be ex-pected to occur in samples from hunter-gath-erers than in those from farmers. It is importantto note that masticatory stress from chewingdepends on the mechanical properties and prep-aration of the foods consumed, which may notalways be related to broad categories of subsist-ence such as ‘‘farmers’’ or ‘‘hunter-gatherers.’’However, for the populations analyzed here,there is evidence to support the distinction be-tween hard and soft diets (see below). Furthergoals are: 4) assessment of the phylogenetic factorthrough the comparison of Patagonian sampleswith an early-middle Holocene sample fromColombia (Tequendama, ca. 7,300–5,800 years14C BP; Correal Urrego and van der Hammen,1977) and a middle Holocene Patagonian sample(Cerro Sota and Palli Aike, ca. 4,500–3,500 years14C BP; Bird, 1988; Hedges et al., 1992) (If cra-niofacial robustness were due to the fact thatpopulations from Patagonia retained differentlevels of robusticity, such samples should besimilar to earlier samples.); and 5) evaluationof the climatic factor by comparison of samplesfrom latitude 25–558 South, using this vari-able as an indirect estimator of temperature.If craniofacial robustness were related to tem-perature, the samples should display an asso-ciation between morphology and latitude.
MATERIALS AND METHODS
Samples
Patagonia is represented by seven late Holo-cene samples from north Patagonia to Tierra delFuego (Fig. 1, Table 1). For comparative pur-poses, one sample from west central Argentina(Cuyo region) and one from northwest Argen-tina were analyzed (Fig. 1, Table 1). Thesesamples belonging to prehistoric populationsof the late Holocene come from different geo-graphic and ecological regions, distributedalong 3,000 km (from latitude 25–558 South;Fig. 1). In addition, they comprise differentsubsistence systems: 1) Patagonian populationscharacterized by subsistence, based mainly onhunting of guanaco (Lama guanicoe) and sealion (Otaria flavescens) and gathering of somefruits, roots, and seafood (Borrero, 1997); and2) west-central and northwestern Argentinepopulations from the later late Holocene,based principally on agriculture (Canals Frau,1950; Marquez Miranda, 1950). Likewise, twoearly-middle Holocene samples from SouthAmerica (Table 1) were included to test thephylogenetic hypothesis. A detailed descriptionof all samples is presented in the Appendix.
Because the same cranial features used forsex estimation are used in assessing robustness(Buikstra and Ubelaker, 1994; Walrath et al.,2004), and because no estimation of sex in-dependent of cranial morphology could be ac-hieved for most samples, males and femaleswere pooled in the comparisons. However, beforecomparing robusticity among samples, the sex ofindividuals was estimated in order to know if thesex ratio was unbalanced in any of them. Theresults show that only the sample from southernPatagonia (Magallanes) is unbalanced, havingone female and 10 males (Table 1).
Morphometric analysis
In geometric morphometrics, the notion ofshape is restricted to boundary informationonly (Bookstein, 1982). Thus, our analysisincluded craniofacial features that can be cap-
Fig. 1. Map shows locales of samples of crania analyzed.Abbreviations in all figures are expanded in Table 1.
750 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb
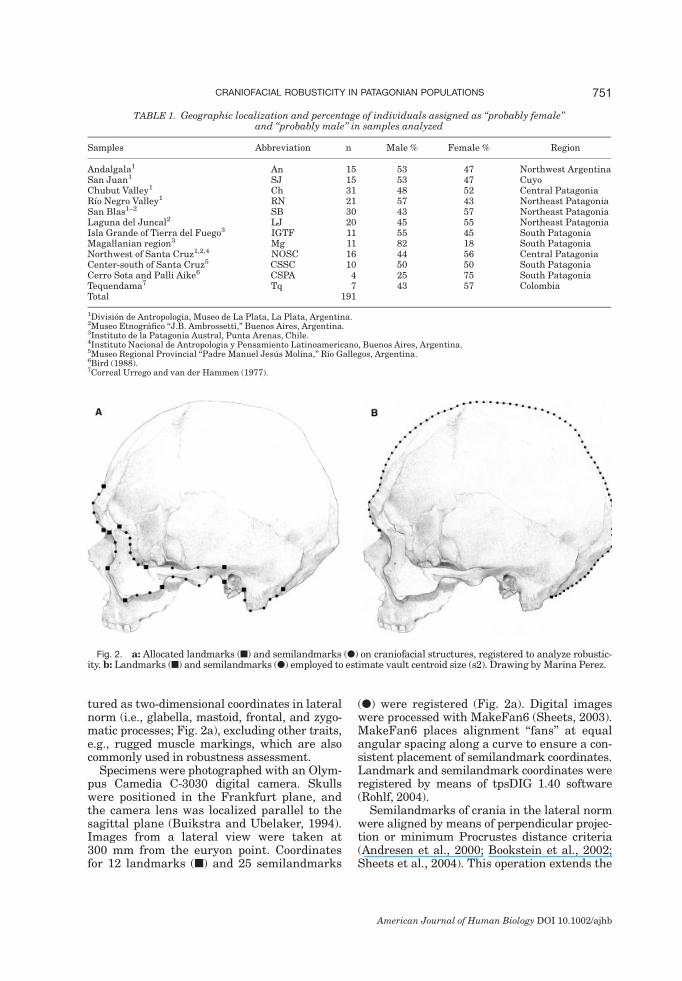
tured as two-dimensional coordinates in lateralnorm (i.e., glabella, mastoid, frontal, and zygo-matic processes; Fig. 2a), excluding other traits,e.g., rugged muscle markings, which are alsocommonly used in robustness assessment.Specimens were photographed with an Olym-
pus Camedia C-3030 digital camera. Skullswere positioned in the Frankfurt plane, andthe camera lens was localized parallel to thesagittal plane (Buikstra and Ubelaker, 1994).Images from a lateral view were taken at300 mm from the euryon point. Coordinatesfor 12 landmarks (n) and 25 semilandmarks
(l) were registered (Fig. 2a). Digital imageswere processed with MakeFan6 (Sheets, 2003).MakeFan6 places alignment ‘‘fans’’ at equalangular spacing along a curve to ensure a con-sistent placement of semilandmark coordinates.Landmark and semilandmark coordinates wereregistered by means of tpsDIG 1.40 software(Rohlf, 2004).Semilandmarks of crania in the lateral norm
were aligned by means of perpendicular projec-tion or minimum Procrustes distance criteria(Andresen et al., 2000; Bookstein et al., 2002;Sheets et al., 2004). This operation extends the
TABLE 1. Geographic localization and percentage of individuals assigned as ‘‘probably female’’and ‘‘probably male’’ in samples analyzed
Samples Abbreviation n Male % Female % Region
Andalgala1 An 15 53 47 Northwest ArgentinaSan Juan1 SJ 15 53 47 CuyoChubut Valley1 Ch 31 48 52 Central PatagoniaRıo Negro Valley1 RN 21 57 43 Northeast PatagoniaSan Blas1–2 SB 30 43 57 Northeast PatagoniaLaguna del Juncal2 LJ 20 45 55 Northeast PatagoniaIsla Grande of Tierra del Fuego3 IGTF 11 55 45 South PatagoniaMagallanian region3 Mg 11 82 18 South PatagoniaNorthwest of Santa Cruz1,2,4 NOSC 16 44 56 Central PatagoniaCenter-south of Santa Cruz5 CSSC 10 50 50 South PatagoniaCerro Sota and Palli Aike6 CSPA 4 25 75 South PatagoniaTequendama7 Tq 7 43 57 ColombiaTotal 191
1Division de Antropologia, Museo de La Plata, La Plata, Argentina.2Museo Etnografico ‘‘J.B. Ambrossetti,’’ Buenos Aires, Argentina.3Instituto de la Patagonia Austral, Punta Arenas, Chile.4Instituto Nacional de Antropologia y Pensamiento Latinoamericano, Buenos Aires, Argentina.5Museo Regional Provincial ‘‘Padre Manuel Jesus Molina,’’ Rıo Gallegos, Argentina.6Bird (1988).7Correal Urrego and van der Hammen (1977).
Fig. 2. a: Allocated landmarks (n) and semilandmarks (l) on craniofacial structures, registered to analyze robustic-ity. b: Landmarks (n) and semilandmarks (l) employed to estimate vault centroid size (s2). Drawing by Marina Perez.
751CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

generalized least square (GLS) Procrustessuperimposition (Gower, 1975; Rohlf, 1990;Rohlf and Slice, 1990): in addition to optimallytranslating, scaling, and rotating landmarks,the semilandmarks are slid along the outlinecurve until they match as well as possible thepositions of corresponding points along the out-line of a reference specimen (Adams et al.,2004). This is done because individual points inthe curves are not claimed to be homologous inall subjects. Consequently, the variation alongtangent directions is not informative, and onlythe coordinate normal to the outline bears in-formation about differences between specimensor groups (Bookstein, 1997; Bookstein et al.,2002). In this study, tpsRelw 1.40 (Rohlf, 2004)was used to align the semilandmarks alongtheir respective curves, sliding them along soas to minimize the Procrustes distance betweenthe subject and the reference (Sampson et al.,1996; Sheets et al., 2004).Shape differences between specimens were
studied by thin-plate spline analysis (Book-stein, 1989), using centroid size (s) as sizemeasurement (Bookstein, 1991). Two meas-urements of centroid size were used: oneobtained from the coordinates recorded onrobusticity traits (i.e., glabella, mastoid, fron-tal, and zygomatic processes; s1), and anotherfrom 80 points describing the sagittal plane ofthe vault (s2) (Fig. 2b). The latter centroid sizeis considered here as an overall estimate ofcranial size. Both uniform (global changeamong landmarks and semilandmarks) andnonuniform (localized change among land-marks and semilandmarks) components ofshape variation were analyzed in this study(Rohlf, 1993, 1996). The alpha parameter,which determines the relative weighting ofthe principal warps (eigenvectors of the bend-ing energy matrix; Bookstein, 1991; Rohlf,1993, 1996) at different scales, was set at zero,as suggested by Rohlf (1993). Partial warps(shape variables generated by thin-plate sp-line transformations; Bookstein, 1989, 1991)and centroid size values were obtained withtpsRelw 1.40 (Rohlf, 2004).Principal component analysis (PCA) was
performed on partial warps as well as on par-tial warps plus centroid size, in order todescribe major trends in shape and form,respectively. The principal components (PCs)obtained from the covariance matrix of partialwarp variables are termed ‘‘relative warps’’(RW; Bookstein, 1991; Rohlf, 1993). The shapechanges of each specimen with respect to thereference configuration (i.e., consensus or
mean form) can be visualized as deformationgrids based on each relative warp axis, andwere obtained with tpsRelw 1.40 (Rohlf,2004). The PCA from partial warps plus cent-roid size (which represent the form of speci-mens) was obtained from the correlation ma-trix, because the measurement scale of thecentroid size differs from that of the partialwarps. The deformation grids along the firstPC axis were obtained with tpsRegr 1.28(Rohlf, 2004). PCA was done to evaluate bothwithin-sample and among-sample variation,in order to find a ‘‘common robusticity factor’’(Pearson, 2000; Petersen, 2000). Among-sam-ples comparisons were made using the consen-sus configurations of each sample. We calcu-lated Spearman’s correlation between latitudeand PC1 and RW1 scores of the sample con-sensus to assess whether there is an associa-tion between this ‘‘common robusticity factor’’and latitude.
To address the problem of estimating sexualdimorphism in skeletal samples where the sexof individuals cannot be reliably determined,several studies suggested using techniquesthat correlate sample variability with sexualdimorphism (Fleagle et al., 1980; Leuteneggerand Shell, 1987; Plavcan, 1994). Therefore,besides calculating the ‘‘common robusticityfactor,’’ the morphological diversity or varia-tion within each sample was evaluated bymeans of the measurement of disparity estab-lished by Foote (1993; also see discussion inZelditch et al., 2004). This measurement isdefined as morphological disparity D ¼ S(di
2)/(n � 1), where di represents the distance fromeach specimen to the centroid of its group. Dis-parity measurement was calculated by meansof Disparity Box 6 software (Sheets, 2003),which uses partial Procrustes distance as thedi measure. Size variation within each samplewas assessed using the standard deviation(SD) of centroid size. Finally, we calculatedthe association between the first score of rela-tive warps analysis and the Foote measure-ment of disparity, as well as the correlationbetween mean and SD of the centroid size, inorder to find out if dimorphism covaried withthe robusticity of samples.
RESULTS
Craniofacial robusticity among lateHolocene samples
Figure 3 shows the first two relative warpsfor the consensus shape of Patagonian, SanJuan, and Andalgala samples, which account
752 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

for 61.23% of the explained variance, and thedeformation grids of the first axis. The maindifferences along this axis, which explain 40.15%of the variance, are located in the glabella, themalar, and the frontal and zygomatic pro-cesses of the malar, whereas the mastoid pro-cess does not show changes. As seen in the de-formation grids (Fig. 3b), the samples with thehighest negative values for the first RW axis(i.e., SJ, An, NOSC, RN, and SB; abbreviationsare expanded in Table 1) show a less prominentglabella and narrower malar, frontal, and zygo-matic processes than samples with the highestpositive values (i.e., CSSC, IGTF, Mg, and LJ).
The samples from southernmost Patagonia(i.e., CSSC, IGTF, and Mg) are smaller thansamples from north Patagonia, consideringboth vault-based and robusticity-based meancentroid size (Table 2). The correlation be-tween RW1 and mean centroid size estimatedfor robusticity traits is low (rs ¼ �0.030). Thesamples occupying positive values of RW1 con-tain individuals characterized by a prominentglabella and wider malar, frontal, and zygo-matic process. Some of these samples includethe smallest individuals (Mg, IGTF, and LJ),whereas others comprise the greatest sizes(e.g., CSSC). On the other hand, the samples
Fig. 3. a: First two relative warps for consensus shape of late Holocene samples. b: Deformation grids representvariation along first relative warp axis, which represents common factor of robusticity. Grids show transformationfrom mean into less (RW1�) and more (RW1þ) robust morphologies.
753CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

characterized by a less prominent glabellaand narrower malar, frontal, and zygomaticprocesses contain both large and small indi-viduals. Similar results (rs ¼ �0.115) wereobtained using the vault-based centroid size(s2).The results of PCA performed on shape and
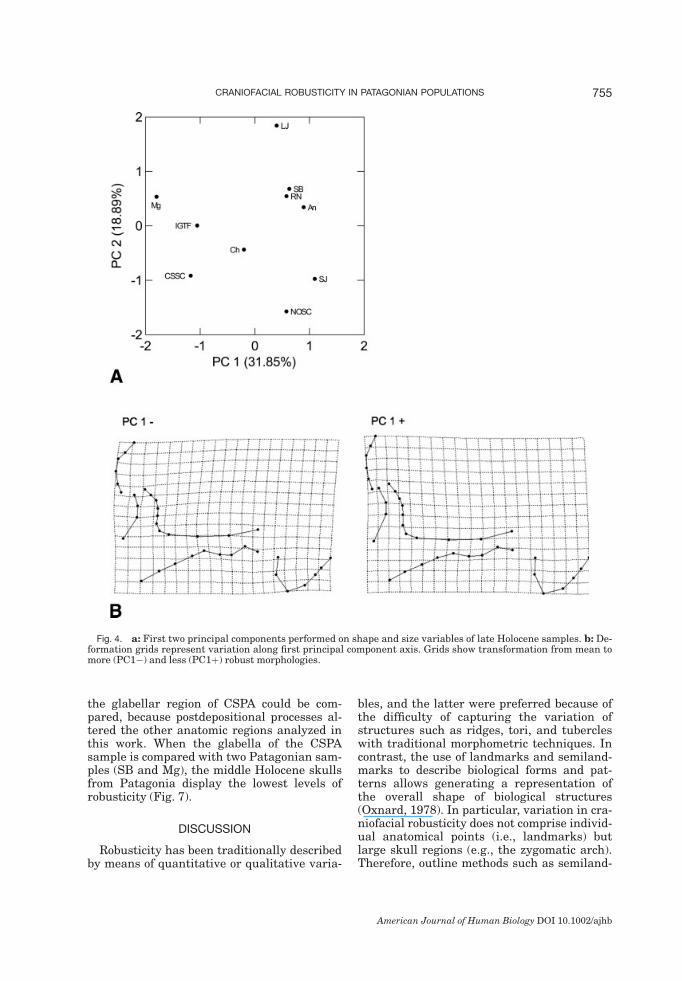
size variables are shown in Figure 4a. The re-sults obtained in this analysis agree with thoseobtained by relative warps analysis. The defor-mation grids for the first component presentsimilar variation to those corresponding to thefirst RW (Fig. 4b). In the PCA, the componentloading of centroid size was low for the firstaxis (�0.286); hence almost all variation in thisaxis corresponds to shape variation.
Relationship between latitude andmorphometric variables
When the Patagonian, San Juan, and An-dalgala samples are compared, our resultssupport a strong correlation between latitudeand shape (rs ¼ 0.76), as well as between lati-tude and form (rs ¼ �0.91). The samples fromhigh latitudes present ‘‘strongly formed’’ mor-phologies (corresponding to positive values ofRW1 and negative values of PC1), whereassamples from low latitudes present ‘‘weaklyformed’’ morphologies (corresponding to nega-tive values of RW1 and positive values of PC1).On the contrary, the association between lati-tude and size, in terms of both robusticity-based and vault-based values, is weak (rs ¼0.23 and rs ¼ 0.006, respectively).When only samples from the later late Holo-
cene of Patagonia are analyzed, the associa-tion between latitude and shape is rs ¼ 0.68,and between latitude and form is rs ¼ 0.82,whereas latitude and size measured on robust
traits (s1) and vault (s2) are negatively corre-lated (rs¼ �0.429 and rs ¼ �0.61, respectively).
Craniofacial robusticity within lateHolocene samples
Despite the fact that the association bet-ween the measurement of disparity and RW1(r ¼ 0.364) is low, there is a slight tendency inthe samples with ‘‘strongly formed’’ morpholo-gies to present higher disparity (IGTF, LJ, andCSSC), with the exception of the Mg sample(Table 3). The fact that this is the only sex-unbalanced sample could account for its lowdisparity.
With respect to size, the highest SD andlowest SD values are found both in small-sizedand large-sized samples (Table 2). In sum-mary, the association between SD and size,both robusticity-based and vault-based, is notsignificant (r ¼ �0.449, P ¼ 0.193, and r ¼�0.052, P ¼ 0.888, respectively).
Our results show differences in the patternof within-group variation among samples.Within each sample, the glabella, the malar,frontal, and zygomatic process, and mastoidprocess range from ‘‘strongly formed’’ to ‘‘weak’’morphologies (Fig. 5). The most dimorphicsamples are from Patagonia (Mg, CSSC, LJ,Ch, and NOSC; Fig. 5). The morphologicalvariation observed along the first PC is asmall fraction of the total variation withineach sample (the variance explained by thefirst axis ranges from 15–25%; Table 3). Inmost samples, differences in form between in-dividuals can be interpreted as shape differen-ces, because the loading of this variable ishigher than the loading for size in the first PC(Table 3). In only two samples (SJ and LJ) wasthis pattern inverted, with size being the vari-able with higher loading in the first PC. Inthese samples, there was a significant associa-tion between size and shape, whereas this waslow in the other samples (Table 3).
Craniofacial robusticity within early-middleHolocene samples
The Tequendama deformation grids showthat neither the mastoid process nor the gla-bella vary along the first RWaxis, whereas themalar process changes slightly from ‘‘strongly’’to ‘‘weakly formed’’ (Fig. 6). The degree ofrobusticity observed in the middle HolocenePatagonian sample (Cerro Sota and Palli Aike)is similar to that of samples from north Pata-gonia and northwest Argentina, but differs fromthe samples from southernmost Patagonia. Only
TABLE 2. Means and standard deviations of twocentroid sizes calculated for late Holocene samples1
Sample s1 mean s1 SD s2 mean s2 SD
An 31.14 2.21 71.56 4.38Mg 33.22 1.59 71.86 3.50IGTF 33.14 1.89 74.33 3.99SB 33.38 0.98 75.06 3.02SJ 33.32 2.28 75.83 3.65CSSC 35.99 1.46 76.29 3.02NOSC 34.37 2.04 76.52 4.09LJ 32.04 2.13 77.17 4.29Ch 35.04 1.70 78.53 3.45RN 35.14 1.69 79.51 4.05
1s1, centroid size obtained from coordinates recorded on robus-ticity traits; s2, centroid size obtained from 80 points describingsagittal plane of vault.
754 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb
the glabellar region of CSPA could be com-pared, because postdepositional processes al-tered the other anatomic regions analyzed inthis work. When the glabella of the CSPAsample is compared with two Patagonian sam-ples (SB and Mg), the middle Holocene skullsfrom Patagonia display the lowest levels ofrobusticity (Fig. 7).
DISCUSSION
Robusticity has been traditionally describedby means of quantitative or qualitative varia-
bles, and the latter were preferred because ofthe difficulty of capturing the variation ofstructures such as ridges, tori, and tubercleswith traditional morphometric techniques. Incontrast, the use of landmarks and semiland-marks to describe biological forms and pat-terns allows generating a representation ofthe overall shape of biological structures(Oxnard, 1978). In particular, variation in cra-niofacial robusticity does not comprise individ-ual anatomical points (i.e., landmarks) butlarge skull regions (e.g., the zygomatic arch).Therefore, outline methods such as semiland-
Fig. 4. a: First two principal components performed on shape and size variables of late Holocene samples. b: De-formation grids represent variation along first principal component axis. Grids show transformation from mean tomore (PC1�) and less (PC1þ) robust morphologies.
755CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

mark-based techniques are more adequatewhen analyzing the contours that describesuch regions. The combination of multivariatestatistics with quantitative analysis of mor-phology by means of semilandmarks used in
this work allowed the quantification of cranialrobusticity and the calculation of a ‘‘commonrobusticity factor’’ (Pearson, 2000; Petersen,2000). The latter was obtained by means of aPCA that converts the complex relationships
TABLE 3. Results of within-sample analysis of late Holocene samples
% explainedvariance, PC1
Component loadingof size in PC1
% explainedvariance, RW1
Correlation,s1 vs. RW1
Foote disparitywithin group
An 22.62 �0.27 29.88 0.07 0.0021Mg 23.69 �0.05 27.84 0.26 0.0020Ch 15.83 0.02 26.45 0.003 0.0018NOSC 17.65 0.36 25.27 �0.19 0.0019CSSC 24.27 0.27 40.04 �0.41 0.0025RN 20.52 �0.51 29.32 0.34 0.0020LJ 17.98 �0.69 26.08 0.59** 0.0026SB 15.47 �0.17 24.57 �0.02 0.0019SJ 19.64 0.85 28.60 �0.52* 0.0021IGTF 21.27 �0.11 32.61 0.13 0.0033
*P < 0.05.
Fig. 5. Deformation grids represent variation within each late Holocene sample along first principal componentaxis.
756 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

of morphometric trends into fewer and sim-pler factors (Sokal and Rohlf, 1969). Likewise,geometric morphometric techniques allow usto visualize the differences in shape along thefirst PC.The first PC obtained using the consensus
configurations of late Holocene samples reflectstheir variation in robusticity (Figs. 3, 4). Inthese samples, the first axis largely describesdifferences in shape, whereas size has lowloading; thus, the differences in robusticitycan be essentially expressed as shape varia-tion. Likewise, there is no association betweenthe size and shape variations summarized bythe first axis; this implies that the differencesin robusticity among samples are not relatedto allometric processes (Atchley and Hall,1991; Gould, 1977; Klingenberg, 1998). In thisstudy, the concept of allometry implies the ex-istence of some shape change associated withincrease in size (Mosimann, 1970). Hence, therelationship between cranial robusticity andsize among populations follows a pattern thatdoes not differ from isometry (Mosimann,1970). The lack of association between sizeand shape is manifest in two robust samplesfrom Patagonia (i.e., IGTF and Mg) whosecentroid sizes are smaller than those of two ofthe most gracile samples (i.e., Ch and RN)from the same region (Table 2). At least forthe cranial traits considered here, no correla-tion was found between size and robusticity.Our results differ from those of Lahr andWright (1996), who found a positive associa-tion between both variables in a study includ-ing samples from around the world. As ourstudy includes exclusively South Americansamples, the different scale of analysis couldaccount for the contrasting results. In thissense, Hawks et al. (2000) asserted that thequestion of whether robustness is a function of
size has different answers at different levels ofanalysis. Within populations, there is usuallya clear relationship between size and robust-ness, because males are on average larger andmore robust than females, but among popu-lations this relationship can vary according towhich populations are compared (Hawkset al., 2000; Wolpoff et al., 2001). Furtheranalyses including larger and more numeroussamples from different geographical regionscould help elucidate this issue.In the late Holocene samples, the increase
in robusticity is related to an increase inwithin-sample shape variation (Table 3). Asshown in Figure 5, such within-sample varia-tion is represented by a great dispersionalong the first PC axis (i.e., common robustic-ity factor). These results can be attributed tothe greater expression of sexual dimorphismin most robust samples. This argument issupported by the fact that sexual dimorphismis one of the most noticeable causes of varia-tion in biological populations (Hanken andHall, 1993; Plavcan, 1994; Wood and Lynch,1996). In addition, the features analyzed cor-respond to sexually diagnostic characters inmodern human crania (Buikstra and Ube-laker, 1994), most of which are related tomuscle insertion sites such as the mastoidprocesses or zygomatic arch. In two samples,a significant association between the first axisthat described shape differences (RW1) andcentroid size was found (Table 3), in whichthe most robust individuals had larger cent-roid sizes of skulls (the negative associationfound for SJ is due to the position of the mostrobust individuals in negative values alongthe first RW). Such results contrast with thelack of association found between size andshape when among-sample robusticity wasconsidered.
Fig. 6. Deformation grids represent variation within Tequendema sample along first relative warp axis.
757CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

When the variation in craniofacial robustic-ity among Fueguian-Patagonian samples isassessed, the results show large variation.The most robust samples are those from southcontinental Patagonia and Isla Grande ofTierra del Fuego, whereas the degree of robus-
ticity of samples from north and central Pata-gonia is similar to farmer samples from westcentral and northwest Argentina (Figs. 3, 4).Previously, other authors considered Fue-guian-Patagonian samples to be compara-tively robust among Amerindian recent popu-lations (Lahr, 1995). However, our resultsshow that the samples from this region areheterogeneous with respect to craniofacialrobusticity, and some of them exhibit the samelevel of robusticity as other non-PatagonianAmerindian samples.
As pointed out above, several hypotheseswere suggested to account for Patagonian cra-niofacial robusticity. The biomechanical hy-pothesis (Hernandez et al., 1997; Lahr, 1995;Lalueza et al., 1997) is supported by evidencefrom observational and experimental studiesshowing that mechanical loading has an im-portant effect on skeletal morphology, withespecially marked effects during the adoles-cent growth spurt (Hanken and Hall, 1993;Herring, 1993; Pearson and Lieberman, 2004).Mechanical loading on osseous structures isproduced by high muscle-force magnitudesresulting from muscle development. In thissense, Vogl et al. (1993) suggested that varia-bility in bone form is determined by intrinsicgenetic and nonheritable environmentaleffects, as well as epigenetic interactions aris-ing from muscles. Simultaneously, the devel-opment of cranial muscles is controlled mainlyby environmental factors such as masticatorystress and epigenetic factors, including hor-monal action (e.g., growth hormone) (Lieber-man, 1996; Vogl et al., 1993). In consequence,when only the areas of muscle attachment areanalyzed, it is difficult to discriminate whichof all these result in increased muscle develop-ment. For that reason, we consider both mor-phometric data and evidence independent ofmorphology in order to discuss the biomechan-ical hypothesis. If Patagonian robusticity re-sults from high levels of masticatory stressdue to hard-food diets, an association betweenthe types of food consumed by a populationand its craniofacial robusticity would be ex-pected. However, our results indicate thatsome samples of hunter-gatherers and farm-ers, belonging to populations that consumedhard and soft diets, respectively, display simi-lar levels of robusticity (e.g., samples from cen-tral-west and northwest Argentina, and north-east Patagonia). In contrast, several samples ofPatagonian hunter-gatherers, who consumedthe same food types, differ markedly in theircraniofacial robusticity (Figs. 3, 4). In addition,
Fig. 7. Deformation grids represent variation in gla-bellar region within middle Holocene Patagonian sam-ple (CSPA) along first relative warp axis. For compara-tive purposes, two other Patagonian samples areincluded (Ao and SB).
758 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

the biomechanical hypothesis is not supportedeither by the evidence of dietary pattern (fromstable-isotopic analyses, and zooarchaeological,ethnographic, and documentary evidence) orby the dental-wear evidence (Barberena, 2001;Bernal et al., 2005; Gomez Otero et al., 1999;Guichon et al., 2001). Numerous studies indi-cate that subsistence from north Patagonia toIsla Grande of Tierra del Fuego was mainlybased on guanaco (Lama guanicoe), with a neg-ligible proportion of marine resources (e.g.,mollusks and pinnipeds) and vegetables (Bar-berena et al., 2001; Gomez Otero et al., 1999;Guichon, 2002; Guichon et al., 2001). Due tothe fact that prehistoric Patagonian diets arehighly homogeneous, they cannot account forthe variation in robusticity among these sam-ples. Bernal et al. (unpublished findings) ana-lyzed the relationship between rate of dentalwear and craniofacial robusticity in severalPatagonian samples, considering dental wearas indirect evidence of masticatory stress. Thisassumption is based on observational and ex-perimental data that found a strong associa-tion between dental wear and diet hardness(see discussion in Scott, 1979; Teaford andOyen, 1989). Their results show that craniofa-cial robusticity is not associated with dentalwear; particularly, two of the most robust sam-ples from south Patagonia show the lowestdental-wear values obtained for this region. Inconsequence, assuming that the rate of dentalwear results principally from food hardness,the morphological differences among Patago-nian samples cannot be directly attributed tovariations in masticatory stress. Previously, itwas noted that although biomechanical stressinfluences skeletal development, the patternsof robusticity among populations cannot beexplained by dental loading alone. For in-stance, Eskimos’ skulls are under higher bio-mechanical stress than other modern popula-tions’ skulls, but they are not among the mostrobust (Turner, 1985).On the other hand, the phylogenetic hypoth-
esis was established on the basis of an assem-bly of cranial features, some related to robus-ticity, which are shared by certain fossils (e.g.,Upper Cave Zhoukoudian) and recent popula-tions from Australia and Patagonia (e.g.,Gusinde, 1939; Imbelloni, 1937; Lahr, 1995;Neves and Pucciarelli, 1990; Pucciarelli, 2004;Sergi, 1911; von Eickstedt, 1934; see discus-sion in Marcellino and Colantonio, 1985;Munizaga, 1976). Such similarity among thesesamples was interpreted in terms of differen-tial retention of the distinctive level of robus-
ticity of Pleistocene populations (Lahr, 1995).These characteristics, along with the fact thatnumerous authors indicate that late Holocenepopulations from Tierra del Fuego share mor-phological traits with the first inhabitants ofAmerica (e.g., Deniker, 1900; Hooton, 1933;Imbelloni, 1937; Lahr, 1995; Pucciarelli, 2004;de Quatrofages, 1879; Soren, 1888; ten Kate,1885), termed Paleoamericans (Deniker, 1900;Pucciarelli, 2004) or Paleoindians (Neves andPucciarelli, 1989, 1990), led to the descriptionof such populations as ‘‘primitive’’ or ‘‘ances-tral.’’ If the robust features of Patagoniansamples are ‘‘ancestral,’’ earlier Americansamples (particularly those from Patagonia)would be expected to show such features. Al-though South American fossils from the earlyHolocene are relatively few and Paleoameri-cans are a heterogeneous group (Barrientoset al., 2003; Jantz and Owsley, 2001), someconclusions can be drawn from the analysis ofthe two early samples analyzed in this workand from the cranial morphology of otherearly samples from South America. The phylo-genetic hypothesis is not supported by mor-phological evidence, either from early-middleHolocene South American (i.e., Tequendema;ca. 7,300–5,800 years 14C BP; Correal Urregoand van der Hammen, 1977) or from middleHolocene Patagonian samples (Cerro Sota andPalli Aike; ca. 4,500–3,500 years 14C BP; Bird,1988; Hedges et al., 1992) studied here (Fig. 6).In addition, the craniofacial morphology of skel-etons from Arroyo Seco 2 (Southeast Pampa,Argentina; ca. 7,800–6,300 years 14C BP;Barrientos, 1997; Politis and Madrid, 2001),Lagoa Santa (Brazil; ca. 9,000–6,000 years14C BP; Dillehay, 2000), and Bano Nuevo(Patagonia, Chile; ca. 9,000–8,500 years 14CBP; Mena and Reyes, 2001) displays notice-ably lower levels of robusticity than late Holo-cene south Patagonian samples. The robustic-ity of these early samples is comparable tothat in the majority of late Holocene Amerin-dians. The presence of slight superciliaryarches in the skulls of Cerro Sota was notedby Munizaga (1976), who considered themmodern features. Therefore, crania exhibitinghigh levels of robusticity seem to be typical oflater late Holocene south Patagonian popula-tions. In view of this evidence, it is more parsi-monious to hypothesize that this patternappeared sometime during the Holocene thanto assume the loss of this feature in the rest ofthe American populations. Thus, such fea-tures would not be homologous to those foundin other populations such as Australian Aborigi-
759CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

nes. Instead, they represent homoplasies thatdo not satisfy the conditions for being consid-ered phylogenetic traits (Lieberman, 1999).Under the cold climatic hypothesis, an asso-
ciation between latitude and craniofacialrobusticity would be expected. Our resultsshow a strong correlation between both varia-bles, with the most robust craniofacial mor-phologies occurring at the highest latitudes(some studies, based on traditional craniomet-ric variables, of samples of Patagonia andTierra del Fuego also found a high associationbetween morphology and geographical spread;Guichon, 2002; Guichon et al., 1989–1990;Hernandez, 1992). However, it is important topoint out that this correlation should be inter-preted cautiously, because the robust shapesdo not change along a continuous gradient.Rather, the most robust samples are fromsouth continental Patagonia and Isla Grandeof Tierra del Fuego, whereas the degree ofrobusticity of samples from northernmost andcentral Patagonia is similar to that of samplesfrom lower latitudes (Figs. 3, 4). Thus themost robust crania belong to populationswhich were exposed to extremely cold temper-atures. In contrast, skull size is negativelyassociated with latitude, with the south Pata-gonian samples being smallest. For Patagoniaand Tierra del Fuego, cold climate was pro-posed as a causal factor of variation in nasalshape (Gonzalez-Jose, 2003; Hernandez, 1992;Lalueza et al., 1997) and postcranial robustic-ity (Pearson and Millones, 2005; Stock, 2004),but not in cranial robusticity. Pearson and Mil-lones (2005) found that long bones of Fueguianpopulations have large epiphyses in relation tobone length, and their robusticity indexes aresimilar to those of Arctic (Inuit) or North Euro-pean (Sami) populations. Such proportionsreflect a large amount of body mass relative tostature in all these populations.The fact that human climatic adaptation is
reflected in external body morphology, partic-ularly in general body size and shape includ-ing the trunk and limb proportions and longbone robusticity (Pearson, 2000; Ruff et al.,2002), as well as in nasal form and generalhead shape (Beals et al., 1984; Churchill,1998), is widely recognized. The relationshipbetween cold climate and skeletal robusticitywas pointed out by Bulbeck (2001), Churchill(1998), Pearson (2000), Stock (2002), Trinkaus(1981), and others. Pearson (2000) addressedthe influences of climate and behavior on longbone robusticity. His results suggest that thepredominant systemic influence on postcra-
nial robusticity may be climatic rather thanbehavioral. However, Pearson (2000) proposedthat ‘‘proximate influence on long bone robus-ticity is probably not climate itself but bodymass and body mass relative to stature, bothof which are elevated in high latitude popula-tions.’’ The influence of climate on overall bodysize, body mass, and shape in homeothermicspecies has been recognized for over a century(Allen, 1877; Bergmann, 1847). Studies of liv-ing human populations show that the rules ofBergmann (1847) and Allen (1877) apply verywell to living humans, with cold-adapted pop-ulations characterized by a broad body withrelatively short limbs (Holliday, 1997; Ruff,1994; Trinkaus, 1981). On the other hand, ex-perimental data derived from studies of ani-mals raised under different environmental con-ditions provide some support to the hypothesisthat environmental temperature can affectgrowth patterns and resulting body propor-tions. For example, Lee et al. (1969) found thatrats growing in cold temperatures showedslower growth rates and shorter but relativelythicker long bones than rats grown at highertemperatures. Most studies focused on thepostcranial skeleton, but little is known aboutthe influence of climate on other features ofskeletal variability.
Several authors proposed that skeletalrobusticity in cold-adapted populations canresult from endocrine action (see discussion inChurchill, 1998). This is partially supportedbecause the influence of climate on severalphysiological characteristics is well-documented(Leonard et al., 2002). For instance, humanpopulations living in cold climates have ele-vated basal metabolic rates. This elevationwas postulated as a physiological adaptationto chronic and severe stress (Leonard et al.,2002; Snodgrass et al., 2005). Studies on cir-cumpolar populations found that metabolicrates appear to be at least partially related toupregulation of thyroid hormones, and thatboth acclimatization and genetic adaptationsseem to play a role in these metabolic re-sponses (Leonard et al., 2002). The systemicphysiological control over robusticity through-out the skeleton was demonstrated by experi-mental studies (Reisenfeld, 1975; Vogl et al.,1993). Lieberman (1996) demonstrated that inthe growing skeleton, exercised animals devel-oped thicker cortical bone throughout the skel-eton, including the cranium, at locations notdirectly loaded during locomotion. In summary,these studies documented the stimulating effectof hormones on craniofacial robusticity, cra-
760 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

nial vault thickening, brow-ridge growth (Lie-berman, 1996), development of major areas ofmuscle attachment (e.g., malar bone; Voglet al., 1993), and muscle development (Reisen-feld, 1975; Vogl et al., 1993). Such changes inmuscle might cause simultaneous changes inskeletal development among different mor-phogenetic components within a complex mor-phological structure, or between differentcomplex structures. In light of this evidence, itseems probable that low temperature actingon hormonal factors can result in increasedlevels of robusticity in human populations.Variations in endocrine factors acting on
skeletal robusticity during development andgrowth can result from either acclimatization(i.e., phenotypic plasticity) or genetic adapta-tions. Thus both processes may account forthe craniofacial robusticity found in late Holo-cene Fueguian and south Patagonian samples.Due to the fact that Patagonian populationsfrom the middle Holocene (e.g., CSPA) wholived in an extremely cold climate do not dis-play the robust features that characterize lateHolocene populations from the same region,the hypothesis of acclimatization may be con-sidered to have lesser support than geneticadaptations. However, this issue requires fur-ther research.In summary, our results show that the mor-
phological evidence does not support eitherthe biomechanical or phylogenetic hypothesesas the main causes of craniofacial robusticityin Patagonian populations. In contrast, theaction of climate on endocrine factors mayaccount for the features observed. However,further research into the influence of cold cli-mate on hormones is required to establish theexact mechanisms by which climate may acton skeletal robusticity.
CONCLUSIONS
The present work demonstrated that semi-landmark-based methods combined with mul-tivariate statistics allow an objective analysisof craniofacial robusticity. This approach al-lowed us to compare the pattern of robusticityin several samples, indicating a large varia-tion among Patagonian samples in both cra-niofacial robusticity and size, with the mostrobust and smallest size samples from southcontinental Patagonia and Isla Grande ofTierra del Fuego. In addition, the objectiveand complete description achieved by meansof geometric morphometrics allowed us to
assess the causal factors (i.e., masticatorystress, retention of ancestral characters, andclimatic adaptation) that could account for theparticular pattern of craniofacial robusticityfound in Fueguian-Patagonia populations.Given that robust features may result from avariety of biological processes, it is necessaryto establish the main factor that could accountfor the pattern observed in each population.Particularly in the case of populations consid-ered in this work, the biomechanical hypothe-sis is not supported by the evidence analyzed.No association between consumed diet anddegree of robusticity was found; some skulls ofhunter-gatherers showed the same develop-ment of robust features as farmers’ skulls.Therefore, the difference in biomechanicalloading sustained by individuals does notseem to be the main cause of variation inrobusticity among these populations. On theother hand, the craniofacial morphology ofearly and late Holocene samples studied herealso does not support the phylogenetic hypoth-esis. While the later late Holocene samplesfrom south Patagonia exhibit high levels ofcraniofacial robusticity, the earlier samplesfrom this region, as well as those from otherregions of South America, show less develop-ment of robust features. Considering this evi-dence, the robust morphologies of late popula-tions from south Patagonia do not seem toresult from the retention of ancestral charac-ters. In contrast, we found a significant associ-ation between latitude and craniofacial robus-ticity, with the most robust craniofacial mor-phologies occurring at the highest latitudes.In this sense, a combination of research strat-egies is required to understand the way tem-perature can influence cranial robusticity.Experimental studies, designed to assess thehormonal changes related to exposure toextremely cold temperatures, can be useful indiscerning whether skeletal growth and devel-opment are altered under such conditions,and which mechanisms are implied. Anequally important approach is the analysis ofcraniofacial robusticity variation in a widerrange of populations by means of quantitativetechniques such as those proposed in the cur-rent work, in order to establish how this pat-tern varies under different environmentalconditions. A large amount of data regardingpostcranial robusticity in cold-climate popula-tions exists, but craniofacial robusticity in thesame groups has received little attention.Thus, an integrated analysis is needed todetermine whether systemic factors acting on
761CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

the skeleton can result in increased cranialand postcranial robusticity.
ACKNOWLEDGMENTS
We are grateful to Hector M. Pucciarelli (Di-vision Antropologıa, Facultad de CienciasNaturales y Museo, La Plata, Argentina), InesBaffi and Leandro Luna (Museo Etnografico‘‘J.B. Ambrosetti,’’ Buenos Aires, Argentina),Rafael Goni (Instituto Nacional de Antropolo-gıa y Pensamiento Latinoamericano, BuenosAires, Argentina), Mateo Martinic (Institutode la Patagonia Austral, Punta Arenas,Chile), and the staff of the Museo RegionalProvincial ‘‘Padre Manuel Jesus Molina’’ (RıoGallegos, Argentina) for granting access to thehuman skeletal collections under their care.Marina Sardi provided helpful comments onan earlier version of the manuscript. We alsothank Cecilia Morgan for help with the Eng-lish-language version of the manuscript, andMarina Perez for illustrations.
LITERATURE CITED
Adams DC, Rohlf FJ, Slice DE. 2004. Geometric morpho-metrics: ten years of progress following the ‘‘revolution.’’Ital J Zool 71:5–16.
Ahern JCM, Karavanic I, Paunovic M, Jankovic I, SmithFH. 2004. New discoveries and interpretations of homi-nid fossils and artifacts from Vindija Cave, Croatia.J Hum Evol 46:25–65.
Allen JA. 1877. The influence of physical conditions on thegenesis of species. Radical Rev 1:108–140.
Andresen PR, Bookstein FL, Conradsen K, Ersbøll B,Marsh J, Kreiborg S. 2000. Surface-bounded growth mod-eling applied to human mandibles. IEEE Trans Med Imag19:1053–1063.
Atchley WR, Hall BK. 1991. A model for development andevolution of complex morphological structures. Biol Rev66:101–157.
Barberena R, Borrero LA, Guichon RA. 2001. Circulacionde poblaciones humanas e isotopos estables en Patagoniameridional. Rev Arg Antropol Biol 3:34.
Barrientos G. 1997. Nutricion y dieta de las poblaciones abor-ıgenes prehispanicas del sudeste de la region pampeana.Ph.D. dissertation, Universidad Nacional de La Plata.
Barrientos G, Pucciarelli HM, Politis G, Perez I, Sardi M.2003. The craniofacial morphology of the early/middle Hol-ocene human populations from the Pampean region(Republica Argentina): getting a new insight into the mor-phological variability of early Americans. In: Miotti L,Salemme M, Flegenheimer N, editors. Where the southwinds blow: ancient evidence for Paleo South Americans.College Station: Center for the Studies of the First Ameri-cans and Texas A&MUniversity Press. p 69–75.
Beals KL, Smith CL, Dodd S. 1984. Brain size, cranial mor-phology, climate and time machine. Curr Anthropol 23:301–330.
Bergmann C. 1847. Ueber die Verhaltnisse der Warmeoko-nomie der Thiere zu ihrer Grosse. Gottinger Stud 3:595–708.
Bird J. 1988. Travels and archaeology in south Chile, edi-tado por J. Hyslop. Iowa City: University of Iowa Press.
Bookstein FL. 1982. Foundations of morphometrics. AnnuRev Ecol Syst 13:451–470.
Bookstein FL. 1989. Principal warps: thin-plate splines andthe decomposition of deformations. IEEE Trans Patt AnalMach Intell 11:67–585.
Bookstein FL. 1991. Morphometric tools for landmark data:geometry and biology. Cambridge: Cambridge UniversityPress.
Bookstein FL. 1997. Landmark methods for forms withoutlandmarks: localizing group differences in outline shape.Med Image Anal 1:225–243.
Bookstein FL, Streissguth AP, Sampson PD, Connor PD,Barr HM. 2002. Corpus callosum shape and neuropsy-chological deficits in adult males with heavy fetal alcoholexposure. Neuroimage 15:233–251.
Borrero LA. 1997. The origins of ethnographic subsistencepatterns in Fuego- Patagonia. In: McEwan C, Borrero LA,Prieto A, editors. Patagonia. Natural history, prehistoryand ethnography at the uttermost end of the earth. London:British Museum Press. p 60–81.
Brown P, Maeda T. 2004. Post-Pleistocene diachronicchange in East Asian facial skeletons: the size, shape andvolume of the orbits. Anthropol Sci 112:29–40.
Buikstra J, Ubelaker D. 1994. Standards for data collectionfrom human skeletal remains. Arkansas ArchaeologicalSurvey research series 44. Fayetteville: Arkansas Ar-chaeological Survey.
Bulbeck FD. 2001. Robust and gracile Australian Pleisto-cene crania: the tale of the Willandra Lakes. In: Siman-juntak T, Prasetyo B, Handini R, editors. Sangiran: man,culture and environment in Pleistocene times. Jakarta:Yayasan Obor Indonesia. p 60–106.
Canals Frau S. 1950. The Huarpes. In: Steward JH, editor.Handbook of South American Indians. Washington, DC:US Government Printing Office. p 169–176.
Churchill SE. 1998. Cold adaptation, heterochrony, andNeandertals. Evol Anthropol 7:46–60.
Correal Urrego G, van der Hammen T. 1977. Investiga-ciones arqueologicas en los abrigos rocosos del Tequen-dana. Bogota: Banco Popular.
Deniker J. 1900. Races et peuples de la terre. Paris: Masson.de Quatrofages A. 1879. L’homme fossile de Lagoa-Santa
au Bresil el ses descendants actuels. Moscow: CongresAnthropologique de Moscou.
Dillehay TD. 2000. The settlement of the Americas: a newprehistory. New York: Basic Books.
Fleagle JG, Kay RF, Simons EL. 1980. Sexual dimorphismin early anthropoids. Nature 287:328–330.
Foote M. 1993. Contributions of individual taxa to overallmorphological disparity. Paleobiology 19:403–419.
Gomez Otero J, Belardi JB, Sunico A, Taylor R. 1999.Arqueologıa de cazadores-recolectores en penınsula deValdes (costa central de Patagonia): primeros resultados.In: Goni R, editor. Soplando en el viento. Actas de las Ter-ceras Jornadas de Arqueologıa de la Patagonia. BuenosAires: Neuquen. p 393–417.
Gonzalez-Jose R. 2003. El poblamiento de la Patagonia.Analisis de la variacion craneofacial en el contexto delpoblamiento Americano. Ph.D. dissertation, Universidadde Barcelona.
Gower JC. 1975. Generalized Procrustes analysis. Psycho-metrika 40:33–51.
Gould SJ. 1977. Ontogeny and phylogeny. Cambridge, MA:Harvard University Press.
Guichon RA. 2002. Biological anthropology in Fuego-Pata-gonia. In: Briones C, Lanata JL, editors. Archaeologicaland anthropological perspectives on the native peoples ofPampa, Patagonia, and Tierra del Fuego to the nine-teenth century. Westport, CT: Bergin & Garvey. p 13–29.
Guichon RA, Martı I, Aspillaga A, Cocilovo JA, RothhammerF. 1989–1990. Contribucion al conocimiento de las rela-
762 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

ciones biologicas entre las poblaciones aborıgenes de Pata-gonia austral y Tierra del Fuego. Runa 19:27–39.
Guichon RA, Borrero LA, Prieto AI, Cardenas P, Tykot R.2001. Nuevas determinaciones de isotopos estables paraTierra del Fuego. Rev Arg Antropol Biol 3:113–126.
Gusinde M. 1939. Die Feuerland-Indianer. Volume III/2.Die Anthropologie der Feuerland-Indianer. Vienna: Mod-ling.
Hanken J, Hall BK. 1993. The skull, volume 3. Functionaland evolutionary mechanisms. Chicago: University ofChicago Press.
Hawks J, Oh S, Hunley K, Dobson S, Cabana G, Dayalu P,Wolpoff MH. 2000. An Australasian test of the recentAfrican origin theory using the WLH-50 calvarium.J Hum Evol 39:1–22.
Hedges REM, Housley RA, Bronk CR, Van Klinken GJ.1992. Radiocarbon dates from the Oxford AMS system:archaeometry datelist 15. Archaeometry 34:337–357.
Hernandez M. 1992. Morfologıa craneal de las etnıas de laTierra del Fuego: diferencias sexuales e intergrupales.An Inst Patagonia Ser Cienc Soc 21:81–98.
Hernandez M, Lalueza C, Garcıa-Moro C. 1997. Fueguiancranial morphology: the adaptation to a cold, harsh envi-ronment. Am J Phys Anthropol 103:103–117.
Herring SW. 1993. Epigenetic and functional influences onskull growth. In: Hanken J, Hall BK, editors. The verte-brate skull, volume 1, development. Chicago: Universityof Chicago Press. p 153–206.
Holliday TW. 1997. Postcranial evidence of cold adaptationin European Neanderthal. Am J Phys Anthropol 104:245–258.
Hooton E. 1933. Racial types in America and their relationto Old World types. In: Jenness D, editor. The Americanaborigines. Toronto: Toronto Press. p 315–355.
Imbelloni J. 1937. Fueguidos y Laguidos. Posicion actual dela raza Paleo-Americana o de Lagoa Santa. An Mus ArgCienc Nat 39:6–103.
Jantz RL, Owsley DW. 2001. Variation among early NorthAmerican crania. Am J Phys Anthropol 114:146–155
Klingenberg CP. 1998. Heterochrony and allometry: theanalysis of evolutionary change in ontogeny. Biol Rev 73:79–123.
Lahr MM. 1995. Patterns of modern human diversification:implications for Amerindian origins. Yrbk Phys Anthro-pol 38:163–198.
Lahr MM, Wright RVS. 1996. The question of robusticityand the relationship between cranial size and shape inHomo sapiens. J Hum Evol 31:157–191.
Lalueza C, Hernandez M, Garcıa-Moro C. 1997. La morfolo-gıa facial de las poblaciones fueguinas: reflejo de unaadaptacion al frıo? An Inst Patagon Ser Cienc Hum 25:45–58.
Lee MM, Chu PC, Chan HC. 1969. Effects of cold on theskeletal growth of albino rats. Am J Anat 124:239–249.
Leonard WR, Sorenson MV, Galloway VA, Spencer GJ,Mosher MJ. 2002. Climatic influences on basal metabolicrates among circumpolar poplulations. Am J Hum Biol14:609–620.
Leutenegger W, Shell B. 1987. Variability and sexualdimorphism in canine size of Australopithecus and ex-tant hominoids. J Hum Evol 16:359–367.
Lieberman DE. 1996. How and why humans grow thinskulls: experimental evidence for systemic cortical robus-ticity. Am J Phys Anthropol 101:217–236.
Lieberman DE. 1999. Homology and hominid phylogeny:problems and potential solutions. Evol Anthropol 7:142–151.
Marcellino AJ, Colantonio SE. 1983. Relaciones morfologi-cas de los aborıgenes prehispanicos del territorio argen-tino. I: con aborıgenes australianos. Publ Inst Antropol41:7–17.
Marcus LF. 1990. Traditional morphometrics. In: Rohlf FJ,Bookstein FL, editors. Proceedings of the Michigan Mor-phometrics Workshop, special publication number 2. AnnArbor: University of Michigan Museum of Zoology. p 77–122.
Marquez Miranda F. 1950. The Diaguita of Argentina. In:Steward JH, editor. Handbook of South American Indi-ans. Washington, DC: US Government Printing Office.p 169–176.
Mena F, Reyes O. 2001. Montıculos y cuevas funerarias enPatagonia. Una vision desde cueva Bano Nuevo 1, XIregion. Chungara 33:21–30.
Mosimann JE. 1970. Size-allometry: size and shape variableswith characterizations of the log normal and generalizedgamma distributions. J Am Stat Assoc 65:930–945.
Munizaga JR. 1976. Paleoindio en Sudamerica (restos oseoshumanos de las cuevas de Palli Aike y Cerro Sota, pro-vincia de Magallanes, Chile). In homenaje al Dr. Gustavole Paige. Universidad del Norte. Santiago: AlfabetoImpresores, Ltda.
Neves WA, Pucciarelli HM. 1989. Extra-continental biologi-cal relationships of early South American human re-mains: a multivariate analysis. Cienc Cult 41:566–575.
Neves WA, Pucciarelli HM. 1990. The origin of the firstAmericans: an analysis based on the cranial morphology ofearly South American remains. Am J Phys Anthropol 81:274.
O’Connor CF, Franciscus RG, Holton NE. 2005. Bite forceproduction capability and efficiency in Neandertals andmodern humans. Am J Phys Anthropol 127:129–151.
Oxnard CE. 1978. One biologist’s view of morphometrics.Annu Rev Ecol Syst 9:219–241.
Pearson OM. 2000. Activity, climate, and postcranial robus-ticity implications for modern human origins and scenar-ios of adaptive change. Curr Anthropol 41:569–207.
Pearson OM, Lieberman DE. 2004. The aging of ‘‘Wolff ’slaw’’: ontogeny and responses to mechanical loading incortical bone. Yrbk Phys Anthropol 47:63–99.
Pearson OM, Millones M. 2005. Rasgos esqueletales deadaptacion al clima y a la actividad entre los habitantesaborıgenes de Tierra del Fuego. Magallania 33:37–50.
Petersen HC. 2000. Comment on ‘‘Activity, climate, andpostcranial robusticity implications for modern humanorigins and scenarios of adaptive change,’’ by O.M. Pear-son. Curr Anthropol 41:596–597.
Plavcan JM. 1994. Comparison of four simple methods forestimating sexual dimorphism in fossils. Am J PhysAnthropol 94:465–476.
Politis G, Madrid P. 2001. Arqueologıa pampeana: estadoactual y perspectivas. In: Berberian E, Nielsen A, editors.Historia Argentina prehispanica. Cordoba: Editorial Bru-jas. p 737–814.
Pucciarelli HM. 2004. Migraciones y variacion craneofacialhumana en America. Complutum 15:225–248.
Reisenfield A. 1975. Endocrine control of skeletal robustic-ity. Acta Anat (Basel) 91:481–499.
Reyment RA, Blackith RE, Campbell NA. 1984. Multivari-ate morphometrics. London: Academic Press.
Rohlf FJ. 1990. Rotational fit (Procrustes) methods. In:Rohlf FJ, Bookstein FL, editors. Proceedings of the Mich-igan Morphometrics Workshop. Special publication no. 2.Museum of Zoology. Ann Arbor: University of Michigan.p 227–236.
Rohlf FJ. 1993. Relative warps analysis and an example ofits application to mosquito wings. In: Marcus LF, Bello E,Garcıa-Valdecasas A, editors. Contributions to morpho-metrics. Madrid: Monografıas del Museo Nacional deCiencias Naturales. p 132–159.
Rohlf FJ. 1996. Mophometric spaces, shape componentsand the effects of linear transformations. In: Marcus LF,Corti M, Loy A, Slice D, Naylor G, editors. Advances inmorphometrics. New York: Plenum Press. p 117–129.
763CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb

Rohlf FJ. 1999. Shape statistics: Procrustes superimposi-tions and tangent spaces. J Classif 16:197–223.
Rohlf FJ. 2000. Statistical power comparisons among alter-native morphometric methods. Am J Phys Anthropol111:463–478.
Rohlf FJ. 2004. Tps software. Stony Brook: Department ofEcology and Evolution, SUNYat Stony Brook.
Rohlf FJ, Marcus LF. 1993. A revolution in morphometrics.Tree 8:129–132.
Rohlf FJ, Slice DE. 1990. Extensions of the Procrustesmethod for the optimal superimposition of landmarks.Syst Zool 39:40–59.
Ruff CB. 1994. Morphological adaptation to climate in mod-ern and fossil hominids. Yrbk Phys Anthropol 37:65–107.
Ruff CB, Trinkaus E, Holliday TW. 2002. Body proportionsand size. In: Zilhao J, Trinkaus E, editors. Portrait of theartist as a child: the Gravettian human skeleton from theAbrigo do Lagar Velho and its archaeological context.Trabalhos de Arqueologia, Vol. 22 Lisbon: Instituto Por-tugees de Arqueologia. p 365–391.
Sampson PD, Bookstein FL, Sheehan H, Bolson EL. 1996.Eigenshape analysis of left ventricular outlines from con-trast ventriculograms. Adv Morphometrics 284:131–152.
Sardi ML. 2002. Diferenciacion craneofacial en aborıgenesde la Patagonia y su relacion con grupos americanos yextra-americanos. Ph.D. dissertation, Universidad Nacio-nal de La Plata.
Scott GR. 1979. Dental wear scoring technique. Am J PhysAnthropol 51:213–218.
Sergi G. 1911. L’uomo secondo le origini, l’antichite, la var-iazioni e distribuzione geografica. Turin: Pisa-Nistri.
Sheets HD. 2003. IMP—integrated morphometrics package.Buffalo, NY: Department of Physics, Canisius College.
Sheets HD, Keonho K, Mitchell CE. 2004. A combined land-mark and outline-based approach to ontogenetic shapechange in the Ordovician trilobite Triarthrus becki. In:Elewa A, editor. Applications of morphometrics in pale-ontology and biology. New York: Springer. p 67–81.
Snodgrass JJ, Leonard WR, Tarskaia LA, Alekseev VP, Kri-voshapkin VG. 2005. Basal metabolic rate in the Yakut(Sakha) of Siberia. Am J Hum Biol 17:155–172.
Sokal RR, Rohlf FJ. 1969. Biometry. San Francisco: Freeman.Soren H. 1888. Lagoa-Santa racen. E Museo Lundii 1:28–
34.Stock JT. 2002. Climatic and behavioural influences on
postcranial robusticity among Holocene foragers. Ph.D.dissertation, University of Toronto.
Stock JT. 2004. Differential constraints on the pattern ofskeletal robusticity in human limbs relative to climaticand behavioral influences on morphology. Am J PhysAnthropol [Suppl] 38:188–189.
Teaford MF, Oyen OJ. 1989. Differences in the rate of molarwear between monkeys raised on different diets. J DentRes 68:1513–1518.
ten Kate H. 1885. Le crane de Lagoa Santa. Bull SocAnthropol Paris 8:240–244.
Trinkaus E. 1981. Neanderthal limb proportions and coldadaptation. In: Stringer C, editor. Aspects of human evo-lution. London: Taylor and Francis. p 187–224.
Turner CG II. 1985. Comment on ‘‘The supraobital torus: amost remarkable peculiarity’’ by M.D. Russel. CurrAnthropol 26:355.
Vogl C, Atchley WR, Cowley DE, Crenshaw P, Murray JD,Pomp D. 1993. The epigenetic influence of growth hor-mone on skeletal development. Growth Dev Aging 57:163–183.
von Eickstedt F. 1934. Rassenkunde und Rassengeschichteder Menschheit. Stuttgart: Enke.
Walrath DE, Turner P, Bruzek J. 2004. Reliability test ofthe visual assessment of cranial traits for sex determina-tion. Am J Phys Anthropol 125:132–137.
Wolpoff MH, Hawks J, Frayer DW, Hunley K. 2001. Mod-ern human ancestry at the peripheries: a test of thereplacement theory. Science 291:293–297.
Wood CG, Lynch JM. 1996. Sexual dimorphism in the cra-niofacial skeleton of modem humans. In: Marcus LF,Corti M, Loy A, Slice D, Naylor G, editors. Advances inmorphometrics. New York: Plenum Press. p 407–414.
Zelditch ML, Swiderski DL, Sheets HD, Fink WL. 2004.Geometric morphometrics for biologists: a primer. Lon-don: Elsevier Academic Press.
APPENDIX: LATE HOLOCENE SAMPLES
Northeast Patagonia
Samples include Rıo Negro Valley (RN;408490 South, –638010 West); Laguna del Jun-cal (LJ; 408490 South, –638010 West); and SanBlas (SB; 408340 South, –628140 West). Thesesamples include individuals dated to the laterlate Holocene (RN and SB; ca. 1,500–500years 14C BP) and early late Holocene (LJ; ca.3,000–1,500 years 14C BP). They belong toarchaeological sites in the Rıo Negro RiverValley (Rn and LJ) near the coast, and siteslocated on the Peninsula San Blas, also nearthe coast. These samples come from a flat anddry region with extensive steppe grasslands.Subsistence was based mainly on huntingguanaco (Lama guanicoe), with a lesser pro-portion of rheas, and gathering vegetables(e.g., Prosopis sp. and Arjona tuberosa) andseafood (Mytilus edulis, Alaucomya ater ater,and others).
Central Patagonia
Samples include Chubut Valley (Ch; 438200South, –658030 West) and northwest of SantaCruz (NOSC; 478280 South, –718310 West).These samples include individuals dated tothe later late Holocene (ca. 1,500–500 years14C BP), and come from sites in the ChubutRiver Valley near Trelew City (Ch) and sitesnear Salitroso Lake (NOSC). The former sam-ple comes from a flat and dry region withextensive steppe grasslands. Subsistence wasbased mainly on hunting guanaco (Lama gua-nicoe), with a lesser proportion of rheas, andgathering fruits, roots, and seafood (Mytilusedulis, Alaucomya ater ater, and others). Thesecond sample comes from a glacial valleynear the Andes Mountains. The economy wasbased mainly on hunting guanaco (Lama gua-nicoe), complemented with rheas, and gather-ing fruits (e.g., Berberis australis).
764 V. BERNAL ET AL.
American Journal of Human Biology DOI 10.1002/ajhb

South Patagonia
Samples include south continental Patago-nia (Mg and CSSC; from 518360 South to528440 South and 688530 West to 728300 West)and Isla Grande de Tierra del Fuego (IGTF;538160 South, –688570 West). These samplesinclude individuals dated to the later late Hol-ocene, and come from archaeological siteslocated to the north of the Straits of Magellan(Mg and CSSC) and northeast Tierra delFuego (IGTF). This region has extensivegrasslands, and was inhabited by terrestrialhunter-gatherers who consumed mainly gua-naco (Lama guanicoe), sea lions (Otaria flaves-cens), and other marine resources (Mytilus
edulis, Alaucomya ater ater, and others). Dueto the scarcity of vegetables in south Patago-nia, populations from this region incorporateda very low proportion of such resources.
Central-north Argentina
Samples include northwest Argentina (An;278340 South, 668190 West) and Cuyo (SJ;318300 South, 698250 West). The economy wasbased on domesticated plants and animals.Some of the cultivated species were maize (Zeamays), beans (Phaseolus vulgaris), and Cucur-bita sp. Pastoralism was important mainly fortransport and clothes.
765CRANIOFACIAL ROBUSTICITY IN PATAGONIAN POPULATIONS
American Journal of Human Biology DOI 10.1002/ajhb
Copyright © 2022 FDOKUMEN




















