
Movilidad Endógena y Variaciones Demográficas: Una Aplicación Para Ecuador
Upload
khangminh22Category
view
2download
0
Variaciones morfológicas en poblaciones de jutía conga capromys piloride (rodentia, capromyidae), y su relación con la conservación
José Luis de la Fuente Arzola

TESIS DOCTORAL
JUNIO 2019
Jose Luis de la Fuente Arzola
VARIACIONES MORFOLÓGICAS EN POBLACIONES DE JUTÍA CONGA,
CAPROMYS PILORIDE (RODENTIA, CAPROMYIDAE), Y SU RELACIÓN CON LA
CONSERVACIÓN
Universidad de Alicante


DEPARTAMENTO DE ECOLOGÍA
FACULTAD DE CIENCIAS
VARIACIONES MORFOLÓGICAS EN POBLACIONES DE JUTÍA CONGA, CAPROMYS
PILORIDE (RODENTIA, CAPROMYIDAE), Y SU RELACIÓN CON LA
CONSERVACIÓN
Jose Luis de la Fuente Arzola
PROGRAMA DE DOCTORADO
Conservación y Restauración de Ecosistemas
Tesis presentada para aspirar al grado de
DOCTOR POR LA UNIVERSIDAD DE ALICANTE
DIRECTORES
Dr. Vicente Berovides Álvarez
Universidad de La Habana
Cuba
Dra. Josabel Belliure Ferrer
Universidad de Alcalá de Henares
España
VARIACIONES MORFOLÓGICAS EN POBLACIONES DE JUTÍA CONGA, CAPROMYS PILORIDE
(RODENTIA, CAPROMYIDAE), Y SU RELACIÓN CON LA CONSERVACIÓN.


A mi madre Consuelo Arzola
A mis hijos Pepe y Camila


Agradecimientos
Al DrC. Vicente Berovides Alvarez, por su valiosa guía y apoyo durante el desarrollo de la tesis y
sus continuas enseñanzas en el transcurso de mi vida profesional
A la DrC. Jasabel Belliure Ferrer, por su valiosa contribución y oportunas recomendaciones durante
el desarrollo y la revisión final de la tesis
Al DrC. Antonio Escarre por su contribución en el desarrollo de la tesis y por ser ejemplo de
constancia científica
A los DrC. Andreu Bonet, Germán López, Jordi Cortina, Rubén Chamizo y DrC Nicasio Viña,
que contribuyeron como profesores al enriquecimiento de nuestro acervo científico.
A las Universidades de Alicante y de Pinar del Río, por apoyar este doctorado cooperado.
A todos los trabajadores de BIOECO Santiago de Cuba, por su hospitalidad durante el desarrollo de
los cursos recibidos
A todos los trabajadores de las Áreas Protegidas de Las Picúas, Najasa , Isla, Jardines de la Reina,
Ciénaga de Zapata, Península de Guanahacabibes, Cayo Saetía y Hatibonico por su apoyo en
trabajo de campo de la tesis, en especial a MsC. José Luis Linares y los Lic Jesús Pascua y José
Luis Collazo.
A mis compañeros del Instituto de Investigaciones Porcinas en especial a los del Centro de
Promoción y Desarrollo del Biogás.
A todos, muchas gracias.


Resumen
Las poblaciones de jutía conga ocupan un amplio espectro de hábitas (formaciones vegetales) en
Cuba, pero no existen estudios sobre variaciones morfológicas entre poblaciones que incluyan los
posibles factores determinantes de las mismas, a efectos de inferir caracteres adaptativos que
pudieran considerarse para delimitar diferentes unidades de significación evolutiva para la
conservación de la especie. El prsente trabajo tiene como objetivo general determinar la relación
entre variables morfológicas adaptativas, y factores climáticos y de estructura de la vegetación, en
poblaciones de jutía conga que viven en las formaciones vegetales de manglar, bosque y vegetación
xerofítica en Cuba. Consideramos que si las condiciones de los distintos hábitats en cuanto al clima
y tipo de vegetación son suficientemente diferentes entre ellos, entonces la morfometria y
morfofisiología de las poblaciones de jutía conga que los habitan presentarán adaptaciones locales
diferenciadas en relación con dichos hábitats, y estos representarían unidades de manejo
diferenciadas para la conservación de la especie.
Se estudiaron las variaciones morfométricas y morfofisiológicas en ocho poblaciones de jutía conga
(Capromys pilorides) de hábitats diferentes (manglar, vegetación de costa de cayo, bosque de
ciénaga, matorral xeromorfo y bosque semideciduo) en los meses de febrero, marzo y abril (finales
de la seca) y agosto, septiembre y octubre (finales de la lluvia). Para ello se capturaron 768
individuos adultos de jutía conga, 417 hembras y 351 machos. Las poblaciones estudiadas fueron:
Área Protegida de Recursos manejados Las Picuas, Najasa e Isla, Parque Nacional Jardines de la
Reina, Ciénaga de Zapata y Península de Guanahacabibes, Refugio de Fauna Cayo Saetía y Reserva
Ecológica Hatibonico. A cada ejemplar capturado se efectuó un grupo de mediciones que
constituyen variables adaptativas; estas variables se utilizaron para elaborar índices con el fin de
determinar el grado de adaptación local y el grado de diferenciación entre las poblaciones.
Posteriormente, a partir de esta información se establecieron categorías infraespecíficas para la
conservación. Se analizaron 22 variables morfométricas y morfofisiológicas. Nueve variables se
consideraron en sus valores absolutos: peso corporal (PC), peso del hígado (PH), largo corporal total
(LT), largo cabeza tronco (LCT), largo de la cola (Co), pie izquierdo con uña (Pi), grosor de la
médula (GM), corteza (GC) renal y largo de la última porción del intestino grueso (LIG). Trece
variables se consideraron en sus valores relativos, ponderadas con respecto al LT y LCT para
obtener los siguientes índices: Robustez (Ro), condición física (CF), peso por milímetro lineal (PL),
pie relativo (PiR), cola relativa (CoR), índice de absorción de agua, dos índices de reabsorción de
agua, tres índices de nutrición, índice largo del pie/ largo de la cola y el índice de dimorfismo

sexual. Para todas las variables consideradas se estudió también la respuesta diferencial adaptativa
entre sexos dentro de cada población. Para identificar procesos de adaptación se utilizó un índice
climático de Martonne como indicador de las condiciones locales. Los resultados mostraron
diferencias estadísticamente significativas entre las poblaciones de los hábitats bosque y manglar en
cuanto a las variables absolutas y relativas, con valores de incremento del LCT entre poblaciones a
medida que se pasaba del manglar al bosque. Se detectó una fuerte asociación entre LCT, PC y Pi,
así como de los índices de absorción, reabsorción y nutrición con respecto al índice climático de
Martonne, lo que refleja un proceso de adaptación local. Se discuten los resultados en relación a la
protección y uso sostenible de la especie sobre la base de la determinación de cinco grupos de
unidades de manejo para la conservación, conformados por las poblaciones de los siguientes
hábitats: manglar, ciénaga y vegetación xerofítica, bosque con individuos de hábitos arborícolas y
bosque con individuos de hábitos terrestres.

ÍNDICE
CAPÍTULO I. Introducción general ................................................................................................ 1
I.1.- Variación fenotípica y aproximaciones metodológicas para su estudio............................... 3
I.2.- La adaptación local ........................................................................................................... 5
I.3.- Caracteres morfológicos ................................................................................................... 6 I.3.1.- El hígado.................................................................................................................. 8
I.3.2.- El intestino ............................................................................................................. 10 I.3.3.- El riñón .................................................................................................................. 13
I.4. Funadmentos de la investigación utilizando caracteres morfológicos ................................ 15
I.5. Selección natural y varaciones morfológicas adaptativas .................................................. 16
I.6. La especie objeto de estudio: La jutía conga (Capromys pilorides) ................................... 20 I.6.1.- Taxonomía ............................................................................................................. 20
I.6.2.- Distribución y abundancia ...................................................................................... 22 I.6.3.- Alimentación y coprofagia ..................................................................................... 22
I.6.3.- Depredadores y parásitos ....................................................................................... 24 I.6.4.- Reproducción ......................................................................................................... 25
I.6.5.- Hábitos .................................................................................................................. 26 I.6.6.- Conservación y designaciones no taxonómicas ....................................................... 27
I.6. Uso de la jutía conga como recurso natural del bosque (recurso forestal no maderero) ..... 29
I.6. Factores reguladores de las poblaciones de la jutía conga ................................................. 31
I.7. Fundamentación teórica de la presente tesis ..................................................................... 34 I.1.7.1.- Novedad científica y valor teórico de la tesis ....................................................... 35
CAPÍTULO II. Materiales y Métodos Generales ........................................................................... 36
II.1. Poblaciones muestreadas: Caracteristicas de las áreas de estudio ..................................... 36 II.1.1.- Parque Nacional Jardínes de la Reina (Jardínes) .................................................... 36
El grupo insular Jardines de la Reina se extiende desde el Golfo de
Guacanayabo hasta la Bahía de Casilda, en la parte meridional de la Isla de
Cuba. Su extensión es de 360 km y está formado por 661 cayos. ...................37 II.1.2.- Área Protegida de Recursos Manejadas Sabana-Camaguey (Sabana). ................... 38
II.1.3.- Parque Nacional Guanahacabibes (Guanahacabibes). ............................................ 39 II.1.4.- Área Protegida de Recursos Manejados Sierra del Chorrillo (Najasa). ................... 41
II.1.5.- Área Protegida de Recursos Manejados Sur de la Isla de la Juventud (Isla). .......... 42 II.1.6.- Área Protegida de Recursos Manejados Cienaga de Zapata (Zapata). .................... 43
II.1.7.- Reserva Ecológica Hatibonico (Hatibonico). ......................................................... 44 II.1.8.- Paisaje Natural Protegido Cayo Saetía .................................................................. 46
II.2. Muestra utilizada ............................................................................................................ 47
II.2. Variables e índices medidos ............................................................................................ 48
II.3. Efectos a considerar ........................................................................................................ 50

II.4. Análisis estadísticos ........................................................................................................ 50
CAPÍTULO III. Variaciones morfométricas en poblaciones de jutía conga (Capromys pilorides) .. 52
III.1. Introducción .................................................................................................................. 52
III.2. Materiales y métodos ..................................................................................................... 53
III.3. Resultados y discusión ................................................................................................... 58
III.4. Conclusiones ................................................................................................................. 83
CAPÍTULO IV. Análisis comparativo de los índices de reabsorción de agua en poblaciones de jutía
conga (Capromys pilorides) .......................................................................................................... 84
IV.1. Introducción................................................................................................................... 84
IV.2. Materiales y métodos ..................................................................................................... 85
IV.3. Resultados y discusión ................................................................................................... 88
CAPÍTULO V. Comparación de un índice de longitud intestinal en poblaciones de jutía conga
Capromys pilorides (Rodentia: Capromydae) de diferentes hábitats. ............................................101
V.1. Introducción .................................................................................................................. 101
V.2. Materiales y métodos .................................................................................................... 102
V.3. Resultados y discusión .................................................................................................. 104
CAPÍTULO VI. Análisis comparativo de tres índices nutricionales en poblaciones de jutía conga
(Capromys pilorides). .................................................................................................................. 114
VI.2. Materiales y métodos................................................................................................... 114
VI.1. Introducción ................................................................................................................ 118
VI.3. Resultados y discusión ................................................................................................ 118
CAPÍTULO VII. Variaciones morfológicas locales de dos poblaciones de jutía conga (Capromys
pilorides) en el área geográfica Peninsula de Guanahacabibes ......................................................134
VII.1. Introducción ............................................................................................................... 134
VII.2. Materiales y métodos ................................................................................................. 135
VII.3. Resultados y discusión ............................................................................................... 137
CAPÍTULO VIII. Determinación de unidades de manejo para la conservación en poblaciones de jutía
conga (Capromys pilorides) .........................................................................................................147
VII.1. Introducción ............................................................................................................... 147
VII.2. Materiales y métodos ................................................................................................. 147
VII.3. Resultados y discusión ............................................................................................... 149

CAPÍTULO IX. Discusion general ...............................................................................................159
CAPÍTULO X. Conclusiones .......................................................................................................160
Bibliografía ..................................................................................................................................162


1
CAPÍTULO I. Introducción general
La jutía conga (Capromys pilorides) constituye una especie endémica de Cuba (Gundlach, 1877;
Lora, 1895; Bucher, 1937; Morh, 1939; Barbour, 1945; Varona, 1980; Wilson y Reeder, 19993; Silva
et a., 2007) y posiblemente clave en los ecosistemas cubanos (Berovides et al., 2009). Pertenece a la
familia Capromyidae del orden Rodentia (Silva et al., 2007), el cual constituye la tercera parte de
todas las especies de mamíferos del mundo. En Cuba existen en la actualidad siete especies dentro
de la familia Capromyidae, pertenecientes a tres géneros (Capromys, Mesocapromys y Mysateles),
pero anteriormente la familia estuvo representada por otros cuatro géneros más con 14 especies, que
se han extinguido (Silva et al., 2007).
Dentro de los roedores caviomorfos de Cuba, la jutía conga es la especie de más amplia distribución
geográfica (Varona, 1980; Silva et al., 2007) y la más generalista dentro de las especies
macrolocalizadas. Ocupa una gran variedad de hábitats diferentes (Smith y Berovides, 1984;
Berovides et al., 1990a, b; Sánchez et al., 1992; Berovides, 2005; de la Fuente y Berovides, 2009),
como muestra de una posible gran plasticidad fenotípica o de adaptaciones genéticas a diferentes
presiones selectivas. Se trata de una especie que no modifica ni acondiciona el sustrato (Silva et al.,
2007); en este sentido, la disponibilidad de refugios en los bosques es uno de los factores que limita
su densidad poblacional (Comas et al., 1989a; Berovides y Comas, 1997b; Cañizares y Berovides,
2007; Linares, 2005; 2009). No obstante, es una especie que no se encuentra en ninguna de las
categorías de amenaza de la IUCN (2006), con densidades de 1,3 a 4 individuos adultos por hectárea
(ha) para las poblaciones de bosque, 130 individuos por ha para las poblaciones de manglar y 20
individuos por ha para las poblaciones de las costas sur y norte (Berovides y Comas, 1997b).
También se trata de la especie más politípica, con cinco subespecies descritas, algunas de las cuales
aún se encuentran pendientes de estudios posteriores que ratifiquen su estatus subespecífico (Varona,
1983; Silva et al., 2007).
En Cuba es conocido el uso que hacen de la jutía conga los campesinos y pescadores para el
autoconsumo (Pimentel, 2006; Berovides et al., 2007), así como el uso que, desafortunadamente,
hacen algunos sectores sociales con fines lucrativos. La especie posee un rendimiento en canal del
45 al 50 % y un alto valor proteico en su carne (Berovides, 1987; Berovides y Comas, 1993;
Gutierrez y Berovides, 1995), todo lo cual implica que es una especie sometida a una intensa

2
explotación. Por todo lo planteado, se deben establecer planes de manejo que involucren criterios de
conservación sobre la base particular de las características de las poblaciones y su adaptación a las
diferentes presiones selectivas a las que están sometidas, como son las precipitaciones, humedad,
tipo de suelo, formación vegetal, actividad humana, depredadores y patógenos (Berovides y Comas,
1997b). Todos estos agentes contribuyen a establecer diferencias nada despreciables entre
poblaciones desde el punto de vista medioambiental, sobre todo en lo referente a la diversidad de
especies de plantas generadoras de la productividad primaria bruta, las cuales constituyen la fuente
principal de nutrientes para la jutía conga (Manojina et al., 1989; Manojina y Abreu, 1990; Comas et
al., 1994a; Pimentel y Hernández, 2007).
En los tiempos recientes la conservación de las especies al nivel infraespecífico trata de distinguir
las denominadas inicialmente Unidades de Significación Evolutivas (USE) como poblaciones que
poseen atributos genéticos y fenotípicos significativos que determinan el potencial evolutivo, con el
propósito de preservar la biodiversidad, los procesos evolutivos y los ecosistemas (Ryder, 1986;
Waples, 1991; Moritz, 1994). Estos criterios son aplicables a especies bajo algún grado de amenaza
o que se encuentran en uso por las comunidades humanas, como es el caso de la jutía conga.
La determinación de estas unidades es de suma importancia tanto para la conservación de la especie
como para su mantenimiento en cautiverio (Crandall, 2000; 2009; Fraser y Bernatzchez, 2001), ya
que se diferencian linajes genéticos con distintas historias evolutivas. En el caso de la conservación,
resulta necesario conocer si existe variabilidad genética y fenotípica entre sus poblaciones, con el
objetivo de determinar si estas deben ser trabajadas como unidades de manejo para la conservación
de forma independiente, o si el manejo debe realizarse al nivel de especie, es decir, aplicar un plan
de manejo común para todas las poblaciones. Para el caso de la cría en cautiverio, evita mezclas de
diferentes linajes genéticos en los que se rompe la coadaptación de los genes específicos de cada
población (Templeton, 1986; Barraowclough et al., 1996).
Los patrones y procesos de la adaptación local que conllevan a una correspondencia organismo
ambiente son reconocidos como un importante mecanismo de variación genética, y se les asigna un
papel crucial en la divergencia de incipientes especies. El bajo flujo genético o fuertes presiones
selectivas del hábitat, fuerte selección contra genotipos óptimamente adaptados a otros hábitats, pero
moderada selección contra genotipos intermedios y pequeñas variaciones temporales en las fuerzas
de selección, constituyen algunos de los factores ecológicos que promueven la adaptación local. La

3
adaptación local puede ser limitada por el flujo genético, confundida por la deriva genética,
modulada por la selección natural e impedida por la falta de variación genética, siendo la selección
natural diversificadora la fuerza conductora de este proceso (Kaweki y Ebert, 2004; Hoeskstra et al.,
2005; Novoa et al., 2005).
Independientemente de su utilidad para la conservación de especies, los estudios de adaptación
fenotípica local proporcionan una importante información sobre la acción de la selección natural. Es
por ello que constituyen un paradigma para probar hipótesis evolutivas sobre los caracteres
favorecidos o no por factores medioambientales particulares (Kaweki y Ebert, 2004).
I.1.- Variación fenotípica y aproximaciones metodológicas para su estudio
La variación es una condición indispensable para que ocurra el proceso de evolución, representa la
materia prima que es modelada por las fuerzas evolutivas, todo lo cual conduce a un cambio. La
definición más simple es la que afirma que la variación es un fenómeno por el cual no existen dos
organismos exactamente iguales para ninguno de sus caracteres. La variación a nivel de organismo,
y de acuerdo al carácter que varía, se clasifica en: variación fisiológica, morfológica y conductual.
En su conjunto constituyen el fenotipo del individuo, el cual es el resultado de la interacción de sus
genes con los factores ambientales donde se desarrolló y donde habita. La existencia de variación
entre poblaciones de muchos caracteres conjuntamente puede indicar que son rasgos adaptativos, si
bien en algunos casos las variaciones se encuentran asociadas por ligamiento o pleiotropía, y en
otros casos no se trata de variaciones adaptativas y su diferenciación entre poblaciones se debe al
azar (Berovides, 2006).
Una premisa central en la teoría sintética de la evolución es que la variabilidad fenotípica dentro de
las poblaciones es el material para la diferenciación y, potencialmente, de la especiación (Mayr,
1963). Es por ello que la variación ha sido objeto de una gran cantidad de estudios en todas sus
modalidades desde la época de Darwin hasta nuestros días. La variación se considera como una de
las fuerzas principales de la evolución (Santos y Hortelano, 1997), mientras, que Gould (1980)
reconoce a la evolución como un proceso jerárquico con modos complementarios pero diferentes en
tres niveles principales: variación entre poblaciones, especiación y patrones de macroevolución
(Mayr, 1988). Nespolo (2003) reconoce a la selección natural como un segundo estado de un

4
proceso de dos pasos, el primero de los cuales consiste en la generación de variación en cada
generación susceptible de servir como material para la selección, la cual actúa posteriormente como
segundo paso, y concluye que esta última no pueda ser posible sin una continua regeneración de
variación.
Por otra parte, se considera que la magnitud de esta variabilidad determina la tasa de la
diferenciación y la covarianza de los caracteres, así como la trayectoria o sentido en que esta se
verifica, por lo que es el elemento de trascendental importancia para comprender el fenómeno de la
evolución (Lande, 1979). Las hipótesis actuales del origen y mantenimiento de la variabilidad dentro
de las poblaciones invocan procesos microevolutivos de selección, deriva, flujo genético y mutación
(Dennison y Baker, 1991; Nespolo, 2003; Martin y Moestrup, 2003), muy bien estudiados en
roedores de islas (Adler y Levins, 1994; Pergams y Ashley, 2001). Responsables en gran medida de
los bajos niveles de variación son los efectos cuellos de botella, por los que pasan cíclicamente las
poblaciones (Gaines, 1985). Mayr (1963) considera que existen dos grandes tipos o clases de
variación, la intrapoblacional o no geográfica (se refiere a diferencias observadas entre miembros de
una misma población) y la variación geográfica, la cual comprende diferencias observadas entre
miembros de distintas poblaciones.
El estudio de la variación ha estado basado en el avance tanto de aspectos tecnológicos como
conceptuales. Entre los enfoques y aproximaciones metodológicas que se han usado están el estudio
de la variación morfológica, morfométrica, cromosómica, genética y molecular. La variación
morfológica en sus primeros estudios para las descripciones de los taxa estaba basada
fundamentalmente en el material preservado en fluidos. Así, las diferencias dentro o entre los taxa se
reconocían y describían como cuestiones de grado (color, tamaño y forma), o bien por la presencia o
ausencia de caracteres anatómicos. Estudios clásicos de este tipo lo constituyen las revisiones de
finales del siglo XIX y principio del siglo XX, publicadas en la serie North American Fauna (Santos
y Hortelano, 1997). Este periodo está caracterizado por cierto grado de subjetividad en la
interpretación y conclusión de las observaciones, las cuales dependían básicamente del grado de
experiencia del investigador de cada taxón. Sin embargo, los caracteres estudiados eran directamente
observables y su elección estaba basada en un profundo conocimiento de la anatomía y biología
básica del taxón en estudio. La variación morfométrica se sustenta en el análisis simultáneo de
varios caracteres cuantitativos, definida como descripción cuantitativa, y en el análisis y la

5
interpretación de la variación del tamaño y la forma en biología. Sí el objetivo que se persigue es el
de comprender los procesos de adaptación de los seres vivos a su medio ambiente, la mejor elección
es la de estudiar los elementos estructurales de las especies, pues estos están sujetoa a las presiones
selectivas del medio ambiente (Santos y Hortelano, 1997).
Los roedores representan el grupo de mamíferos vivientes más diverso, con más de 2200 especies
descritas (Samuels y Van Valkenburgh, 2008). En los roedores, por su tamaño relativamente
pequeño, cambios sutiles en la morfología de sus huesos y tejidos blandos pueden tener relevantes
consecuencias funcionales, haciendo de ello un buen ejemplo de especialización ecológica, con o sin
cambios morfológicos radicales (Shephed y Digten, 2012). Alimentación y uso de sustrato (dieta y
locomoción) son importantes atributos morfológicos que han sido relacionados con la morfología en
este grupo (Carrizo et al., 2014), en especial en su dentición (Croft, et al., 2011).
Los roedores representan un grupo clave dentro de los ecosistemas, dada su posición en las cadenas
tróficas y su importante papel en la aireación del suelo, dispersión de semillas y polinización
(Gómez, 2015). Es por ello que son considerados prioritarios para la conservación (Amori y
Gippoliti, 2003), y en especial el grupo de los caviomorfos (Vasallo y Antenucci, 2015)
I.2.- La adaptación local
Los estudios de la adaptación local proporcionan conocimientos importantes del valor de la
selección natural en relación al flujo genético y otras fuerzas evolutivas. Constituyen ejemplos de
pruebas de las hipótesis evolucionistas de los caracteres favorecidos por factores medioambientales
particulares (Kawecki y Ebert, 2004). Respuestas plásticas al ambiente pueden influenciar
fuertemente la divergencia entre poblaciones locales que varían en las características de sus hábitats
(Patton y Yang, 1971: Bozinovic et al., 2011)
La adaptación local ha sido reconocida como un importante mecanismo de variación genética, y los
escenarios posibles para la especiación alopátrica y simpátrica asignan a la adaptación local un papel
crucial en la iniciación de la divergencia de incipientes especies (Rogovin y Surov, 1990; Schluter,
2001; Turelli, 2001; Hoekstra et al., 2005; Novoa et al., 2005). Las fuerzas de la selección natural a
menudo varían en el espacio, como resultado de la interacción genotipo x ambiente. En ausencia de
otras fuerzas, la selección natural diversificadora puede causar en cada población local la expresión

6
de caracteres que proporcionan una ventaja a las condiciones ambientales locales, con un patrón
donde los genotipos residentes de cada población tendrán un promedio relativo de oportunidades
más alto en su hábitat local que genotipos de otros hábitats. Este patrón guía a la adaptación local
(Williams, 1996).
Generalmente, una adaptación local es un rasgo del fenotipo el cual es funcionalmente modulado
por la selección natural pasada, mejorando el rasgo o carácter en dependencia de la localidad y entre
los rasgos alternativos existentes. Los caracteres adaptativos tienden a ser fijados en la población, y
una vez que son genéticamente fijados continúan expresándose, aun si cambian las causas que
originaron el carácter y dan como resultado una pérdida de ventaja para el nuevo medio (Stearns,
1994: Ojeda et al. 1999: Morris, 2011).
La adaptación local puede ser limitada por el flujo genético, confundida por la deriva genética,
opuesta por la selección natural debida a variabilidad ambiental temporal, y limitada por la falta de
variación genética o por la arquitectura genética de los caracteres (Kawecki y Ebert, 2004). Los
factores ecológicos que promueven la adaptación local (Kawecki y Ebert, 2004) son:
Bajo flujo genético entre poblaciones
Fuerte selección contra genotipos óptimamente adaptados a otros hábitats, pero moderada
selección contra genotipos intermedios.
Pequeña variación temporal en las fuerzas de selección
Pequeñas diferencias entre hábitat en tamaño y calidad
La adaptación es un fenómeno biológico íntimamente relacionado con la variación, y de la misma
forma que la variación afecta a caracteres morfológicos, fisiológicos y conductuales, igualmente la
adaptación ocurre en cualquiera de estos tipos de caracteres.
I.3.- Caracteres morfológicos
El estudio de la diversidad fisiológica es fundamental para entender cómo los organismos se
enfrentan a la variación ambiental abiótica y biótica en el tiempo y el espacio (Priersman y Drent,
2003). Pero el estudio directo de la fisiología de cualquier órgano puede ser sumamente difícil, por

7
la dificultad de conseguir equipamientos ambientales que simulen el medio natural lo máximo
posible y de mantener a los animales. Sin embargo, existe una forma de medir la eficiencia
fisiológica de un órgano sobre la base de que ciertas medidas lineales o de peso en el órgano de
interés, de forma absoluta o relativa a otra variable (Ojaeti y Dallneier, 2000), pueden reflejar un
buen o un mal funcionamiento del mismo. Usualmente esta variable es el peso de un órgano
completo, el peso de partes asociadas de un órgano, o un índice construido a partir de alguna de las
partes del órgano. Estos índices se denominan entonces morfofisiológicos (Ignatova y Khritoforova,
2003), y muchos de ellos se utilizan en el estudio y manejo de las especies de la fauna (Bookhout,
1994; Braun, 2005).
En el siguiente cuadro se indican algunos de los órganos o sistemas de órganos utilizados como
índices morfofisiológicos, y la fisiología no reproductiva con la que se relacionan, en estudios de
roedores y de otros mamíferos.
Fisiología Variables morfofisiológicas
Nutrición
Grasa total corporal
Peso de la canal
Relación longitud corporal/peso
Grasa renal
Grasa en médula ósea
Peso del hígado
Respuesta al estrés Peso de las glándulas suprarrenales
Metabolismo de reabsorción del
agua
Largo, ancho y alto del riñón
Peso del riñón
Grosor o área de médula renal y corteza renal
Flexibilidad, plasticidad del
desarrollo, respuesta
morfofisiológica
Longitud y peso de las partes del tracto digestivo
(estómago, intestino delgado, ciego y colon)
Para el presente estudio sobre las poblaciones de jutía conga en Cuba se han seleccionado tres
órganos específicos, el hígado, el riñón, y la última porción del intestino grueso (colon y recto).

8
Estos órganos están relacionados, como se puede consultar en el cuadro anterior, con la nutrición y
la reabsorción del agua, y vinculados a respuestas morfofisiológicas plásticas del desarrollo.
I.3.1.- El hígado
Las principales funciones del hígado, la mayor glándula de los vertebrados, quedan reflejadas en el
siguiente cuadro (Pough et al., 1999; Tzamakakis, 2000; Nadal, 2001; Kardong, 2002).
Homeostasis Glucosa Proteína Grasa y colesterol
Hormonas Vitaminas
Síntesis
Proteínas Ácidos biliares Heparina
Colesterol Estrógenos Angiotesinógeno
Proteínas de fase aguda Hematíes (vida fetal)
Almacenamiento Vitaminas Colesterol Glucógeno
Hierro Cobre Grasas
Excreción Ácidos biliares Colesterol Bilirrubina Drogas
Hormonas Venenos Fosfolípidos
Filtración Venenos Nutrientes Bilirrubina Ácidos biliares
IgA Drogas Células muertas o dañadas
Inmune Excreción de IgA en el tracto digestivo
Macrófagos filtradores de antígenos
La función de excreción de sustancias dañinas (desintoxicación) es muy importante en
monogástricos que no tienen un rumen para esta función. El hígado es uno de los órganos con más
alta demanda de flujo sanguíneo, compitiendo con el riñón, el corazón y el cerebro. La alometría
general en mamíferos para el hígado, en relación al peso corporal, es:
Peso del hígado= 0.82 · (Peso corporal) exp 0.87
lo que significa que el aumento del peso del hígado no es directamente proporcional al del cuerpo y
que pesa relativamente poco en animales grandes.
Hume (1989) destaca el papel desintoxicante del hígado señalando que este depende de la
composición química de las plantas que se ingieren; básicamente influye si son especies ricas en

9
antinutrientes como los taninos, o ricas en sustancias tóxicas como los aceites esenciales. Los
mangles (sobre todo el mangle rojo, Rhizophora mangle) son ricos en las primeras y los árboles de
los bosques semideciduos en las segundas. Esta particularidad es esencial tenerla en cuenta a la hora
de realizar comparaciones que consideren el peso del hígado de poblaciones de ambos tipos de
hábitats.
Gosling et al. (1980) utiliza como índice del estado nutricional en la especie coipu (Myocastor
coypus) la relación entre el peso del hígado y el cubo de la longitud de la pata trasera (g/cm3)
2. Este
índice aproximadamente no dimensional se empleó en vez del peso corporal para evitar sesgo
cuando en el denominador se utiliza otra variable que esté asociada a la utilizada como numerador,
como es el caso del peso del hígado y el peso corporal. Se recomienda solo utilizar en animales
adultos y tener en cuenta los cambios estacionales y ciclos reproductivos cuando se hacen las
mediciones. Una comparación de dicho índice, entre tres poblaciones de coipu, dos costeras y otra
de vida libre, produjo para las primeras unas medias de 9.78 ± 2.16 (buenas condiciones
nutricionales) y 6.59 ± 2.41 (malas condiciones nutricionales), y para la de vida libre, 8.50 ± 2.70;
las diferencias entre ellas fueron significativas. Otros estudios con grandes roedores tratan de
relacionar el peso del hígado con la flexibilidad del tracto digestivo (revisado con mayor detalle en
el siguiente apartado), con el costo energético de la lactancia, y con la contaminación ambiental
(este último en pequeños roedores).
Sabat y Bozinovic (2008) probaron la hipótesis de que el tipo de dieta y la flexibilidad fisiológica
del tracto digestivo (incluyendo el hígado) están correlacionados en el degus (Octodon degus).
Estudiaron el efecto de diferentes dietas artificiales en cuanto a su composición química (altas o
bajas en almidón y proteínas desde el destete hasta adulto) sobre el comportamiento digestivo
medido como peso (gramos) seco del hígado. Los animales con dieta de proteína (alta o baja) y baja
en almidón presentaron pesos del hígado mayores y significativamente diferentes a los de dietas
ricas en almidón. Este resultado puede relacionarse con la alta demanda para la desintoxicación y
eliminación de metabolitos, derivados del metabolismo de los aminoácidos.
Trabajando con la misma especie, Naya et al. (2008) estudiaron de nuevo la flexibilidad del tracto
digestivo, pero esta vez en relación a la lactancia, un periodo del ciclo de vida de los mamíferos con
una alta demanda de energía. Estos autores encontraron que los valores del contenido del tracto

10
digestivo, su peso seco, el peso del hígado, y la tasa metabólica de reposo, fueron todos mayores en
los lactantes que en los no lactantes.
Ignatova y Khristoforova (2003), trabajando con pequeños roedores murinos y utilizando el índice
peso hígado/peso corporal, detectaron diferencias de este índice en poblaciones sometidas a
presiones tecnogénicas (contaminación con plomo y zinc), comparándolos con los que vivían en
ecosistemas naturales. Los primeros presentaron en otoño un decremento del peso relativo del
hígado de entre 39.0-49.0mg/g por debajo del límite de la norma. Dicho decremento fue indicativo
de agotamiento físico por baja disponibilidad de energía y consecuente mortalidad masiva.
Comprobaron además que la población se mantenía por migración de juveniles de ecosistemas
próximos no contaminados.
En relación a la jutía conga, Lora (1985) señala que el hígado es la víscera más interesante de esta
especie, porque además de la división en cuatro lóbulos, como en el resto de las especies del género,
los presenta divididos en lobulillos independientes, lo que asemeja una aglomeración de pequeños
hígados. Ellos fueron los primeros en dar datos sobre el peso relativo del hígado de la jutía conga
(Peso del órgano/Peso corporal) para seis poblaciones en diferentes hábitats, con los menores pesos
relativos en las jutías de cayos (2.4-3.0) y los mayores en las de ciénagas y bosques (3.2-4.1).
Posteriormente, este patrón fue confirmado con más detalle en dos poblaciones (una de mangle y
otra de bosque), utilizando la relación peso/peso y el índice de Gosling (Sánchez et al., 1992).
Similar comparación se hizo para otras dos poblaciones, una de bosque y otra de manigua costera
(hábitat xerofítico), por Berovides (2005), utilizando solo la relación peso/peso, siendo
significativamente menor el índice en la población de hábitat xerofítico.
I.3.2.- El intestino
Las principales funciones del intestino grueso son (Kardong, 2002):
Mecánica. Mezcla residuos alimentarios y produce movimientos de grandes cantidades de
alimento.
Digestión química. Efectuada por bacterias que lo colonizan, que producen vitaminas K y B.
Absorción. Vitaminas, electrolitos y agua remanente.
Defecación. Eliminación de heces fecales.

11
La absorción del agua remanente se logra porque su epitelio está cubierto con las aberturas de las
glándulas intestinales tubulares, que penetran profundo en la mucosa y contienen células absortivas
del agua. Los estudios de la flexibilidad fisiológica del tracto digestivo y sus consecuencias son
importantes a causa de que ello puede ser un limitante que influye en la historia de vida de la
especie, en especial sobre el crecimiento, la supervivencia y la reproducción (Mcwillian y Karasov,
2001).
Usualmente, las variables de longitud de alguna parte del tracto digestivo (estómago, intestino
delgado, intestino grueso o sus partes) se procesan de forma relativa al peso corporal (Sabat y
Bocinovic, 2008) o al largo corporal (Schauenberg, 1977; Hansson, 1985), para evitar el efecto
asociado de estas otra svariables. Freehling y Moore (1987) ofrecen los detalles de cómo medir con
exactitud dichas longitudes.
Las investigaciones en roedores, en relación a la variación del peso o longitud absoluta o relativa de
varias partes del tracto digestivo, se han realizado para determinar su flexibilidad en relación con los
hábitos alimentarios (Vorontsov, 1962; Karasov y Diamond, 1983; Gross et al., 1985; Ellis et al.,
1994; Wang et al., 2003; Sassi et al., 2007), así como frente a diferentes tipos de dietas (Sabat y
Bocinivic, 2008) y diferencias geográficas (Hansson, 1985). Este último estudio demuestra, al
menos para Clethrionomys glareolus de Suecia, que la variación de la longitud intestinal es
hereditaria, pero con una gran plasticidad del desarrollo. Las diferencias encontradas en intestino
delgado, el ciego y el colon, entre localidades geográficas, se relacionan con las reglas de
Bergmann, con diferencias en estrategias demográficas, y con los modos de alimentación.
Naya (2008) realizó una revisión sobre estos enfoques, que aunque no son aplicables a todas las
especies, se refieren mayormente a pequeños roedores. Las conclusiones principales fueron:
existe una clara correspondencia entre ajustes morfológicos del tracto digestivo y cambios de
las condiciones ambientales, de tal modo que la flexibilidad en el ajuste de su tamaño puede
considerarse un mecanismo fisiológico generalizado, demostrado bajo condiciones de
laboratorio y vida libre;
en animales de laboratorio, los factores experimentales más investigados fueron la calidad de
la dieta, el estado reproductivo, la temperatura ambiental y el ayuno, mientras que para
especies silvestres, los más analizados fueron calidad de la dieta y temperatura;

12
para especies silvestres, no se han detectado diferencias en flexibilidad del intestino delgado
con diferentes tratamientos;
los cambios en longitud pueden estar relacionados con un decremento en el tiempo de
retención del alimento, mientras que los cambios en peso lo estarían con la necesidad de una
alta absorción específica;
referidos específicamente al intestino grueso, estos estudios han demostrado su incremento
de tamaño con dietas de alto contenido de material indigestible, sobre todo el ciego y el
colon;
las variaciones estacionales en el tamaño del tracto digestivo son comunes y están
relacionadas sobre todo con cambios en la calidad de la dieta y la temperatura;
las hembras exhiben mayor tamaño del tracto intestinal que los machos, lo que es más
marcado durante la estación reproductiva;
las diferencias del tracto intestinal entre poblaciones para una misma especie se relacionan
sobre todo con la composición de la dieta;
animales que consumen dietas pobres, deben tener grandes cámaras digestivas;
un incremento en demanda energética y una dieta de baja calidad, determinan un aumento en
longitud y peso del tracto digestivo;
los principales factores que incrementan el tamaño de los órganos digestivos son: dietas con
alto contenido en fibra, bajas temperaturas, la lactancia, y la interacción entre estos y otros
factores (como parasitismo y contenido de taninos en la dieta).
Para la jutía conga, la longitud relativa al largo corporal de la última porción del intestino grueso
(colon y recto) fue utilizada como un índice de la capacidad de absorción de agua por Sánchez
(1992). Berovides (2005) utilizó este índice en dos poblaciones de esta especie, en hábitat de bosque
y xerofítico. La población en este último hábitat mostró una longitud relativa de 5.29 en relación a
4.24 del hábitat de bosque, lo cual fue una diferencia altamente significativa. De la Fuente (2008)
estableció una comparación para este índice intestinal en seis poblaciones de jutía conga de
diferentes hábitats, donde se evidenció altos valores para las poblaciones de manglar y vegetación
xerofítica y bajos valores para las poblaciones de bosque.

13
I.3.3.- El riñón
Las funciones principales del riñón, en relación con el metabolismo del agua, son (Schmidt- Nielsen,
1977; Pough et al., 1999):
Su conservación por reabsorción.
Produce orina muy concentrada por ultrafiltración del agua; las asas de Henle son la clave de
este proceso, dado el arreglo estructural en la médula de sus segmentos descendentes
(extracción de agua) y ascendente extracción de sodio. La concentración final de la orina en
los mamíferos está determinada por la cantidad de sodio acumulado en los fluidos de la
médula.
La relación alométrica del peso del riñón en relación al peso corporal indica no isometría. El peso
del riñón es relativamente pequeño en animales grandes.
Las variables medidas en el riñón (usualmente se toma uno solo, por la alta correlación de los
valores medidos entre ambos) utilizadas como índices de reabsorción de agua son el peso, el ancho,
el largo y el alto del órgano, así como el área o grosor de la corteza y la médula. Las fórmulas
utilizadas establecen relaciones entre médula y corteza, ya sea utilizando su área o su grosor (Yabe,
1982; 1983; Studier et al., 1983; Ojeda et al., 1999); no obstante, otros autores han utilizado el
grosor de la médula en relación a la raíz cúbica del volumen del riñón (Sperber, 1944; Greegor,
1974; Ojeda et al., 1999; Al-Kahtani et al., 2004).
El fundamento fisiológico para mamíferos del uso de estos índices renales fue establecido por los
trabajos de Sperbe (1944), Schmidt- Nielsen y Dell (1961), Heisinger et al., (1973), Brownfield y
Wunder (1976), Beuchat (1990, 1996) y Al-Kahtani et al., (2004), e incluyen varias especies de
roedores. Estos trabajos establecieron que el grosor relativo de la médula renal se correlaciona con
el peso del animal, la concentración de orina (explica el 59% de su variación) y la aridez del hábitat
en términos de altas temperaturas y baja precipitación y productividad primaria. La habilidad para la
concentración de orina por la reabsorción del agua es debida a la relación directamente proporcional
entre el grosor de la médula y la longitud relativa máxima de las asas de Henle. El riñón de los
mamíferos es único al tener todas sus nefronas con asas de Henle (Bankir y De Reuffignac, 1985).

14
Los roedores sudamericanos de los que se derivan los caprómidos antillanos (Davalos, 2004) poseen
sistemas estructurales y funcionales (al nivel celular, de sistemas de órganos y de organismos) para
la conservación de agua semejantes a las de otros roedores más estudiados de Norteamérica,
presentando plasticidad fenotípica y diversidad fisiológica entre sus especies. A estos se añaden
conductas y estrategias ecológicas que también permiten la conservación del agua (Bozinovic y
Gallardo, 2005).
Según diversos autores (Reaka y Armintage, 1976; Yabe, 1982; 1983; Beuchat, 1990; 1996; Díaz y
Ojeda, 1999; Díaz, 2001; Al-Kahtani et al., 2004; Jackson et al., 2004; Díaz et al., 2006), pequeños
mamíferos (mayormente roedores) de tierras áridas de Argentina, Chile y Sudáfrica, ratas y ratones
de Japón, y roedores en general, los factores que más inciden en las variables morfométricas
medidas en los riñones son:
El tipo de dieta, con una médula gruesa en insectívoros y una corteza reducida en herbívoros
solo en la estación de seca.
La estación del año; la seca interacciona con el tipo de dieta.
El hábitat, con mayores valores de los índices renales en hábitats xéricos.
La temperatura.
El peso corporal.
La historia evolutiva o filogenia.
Sin embargo, no siempre se dan estas relaciones. Díaz et al (2006), trabajando con especies de
roedores sudamericanos, no encontraron relación entre el grosor relativo de la médula y la
temperatura, precipitación y productividad primaria como efecto del hábitat entre las especies
estudiadas, pero admiten que la relación puede existir. Igualmente, Ntshotsho et al. (2004), en dos
especies de roedores no encontraron relación de un índice renal con la precipitación, pero reconocen
que el agua es un factor limitante para las poblaciones de roedores. La relación con la temperatura y
otros factores climáticos podría explicarse porque en las especies de roedores estudiados los ajustes
para estos factores no se hacen por mecanismos fisiológicos, sino conductuales, como el uso de
refugio (Jackson et al., 2004).

15
El otro grupo de especies de mamíferos trabajado para los índices renales, aparte de los roedores, es
el de los murciélagos neotropicales (Geluso, 1978; Studier et al., 1983; Schondube et al., 2001).
Estos estudios encontraron los mismos efectos sobre los índices que en los roedores, pero destacaron
como mayor efecto el peso corporal y otros no señalados para los roedores, como el agua libre en los
alimentos, la proteína en la dieta, la concentración de iones específicos y la deshidratación
ambiental. En este grupo existe la distinción entre médula interna y externa, que se distinguen mejor
cuando mayor es la proteína en la dieta. Otro grupo trabajado para los índices renales fue el de los
armadillos sudamericanos (Greegor, 1975), encontrándose la misma relación de valores altos de
índice renal en hábitat xéricos.
En la jutía conga, los primeros estudios que utilizaron un índice renal, fueron los de Sánchez et al.
(1992) y Berovides (2005). En ambos trabajos se utilizó el índice renal de Yabe (1983), el primero
en una población de bosque y otra de mangle, y el segundo en una población de bosque y otra de
vegetación xerofítica. En las poblaciones con mayor carencia de agua (manglar y vegetación
xerofítica), se tuvieron valores promedios mayores y estadísticamente significativos del índice en
comparación con los de bosque, resultado concordante con lo planteado en la literatura.
Posteriormente, de la Fuente y Berovides (2013) confirmaron estos resultados.
I.4. Funadmentos de la investigación utilizando caracteres morfológicos
La morfología general de una especie puede limitar la posible gama de todos los rasgos de su
historia de vida, siendo el tamaño el más importante (Thornigton y Heaney, 1981; Saugr y Slade,
1985; Taylor et al., 1985; Campbell y Slad, 1993; Wainwright y Reilly, 1994; Santos y Hortelano,
1997). La historia de vida constituye un criterio relevante (el más relevante podríamos decir) a
considerar para la conservación de la especie (Gould, 1966; Boyce, 1988; Lindstedt y Swin, 1988;
Starns, 1992); que resulte o no favorecida por la selección natural, esto es, que sea adaptativa, va
ligado al hábitat donde vive la población (Begon et al., 1998; Dobson, 1988; Travis, 1994;
Mousseaur, 2000). De esta forma, analizar caracteres morfológicos relacionados con la historia de
vida permite entender el grado de adaptación de las poblaciones, su potencial para evolucionar, y la
necesidad de su conservación.

16
El mantenimiento de la variabilidad genética de caracteres adaptativos (como en nuestro caso los
morfológicos), debe ser un objeto primario en la conservación de la biodiversidad (Ericsson et al.,
1993). Ello implica el análisis de un número adecuado de poblaciones que habiten en diferentes
condiciones ambientales (Humphries et al., 1995). La evolución de caracteres adaptativos puede
ocurrir en una escala temporal de años o décadas, lo que resulta de particular importancia para la
conservación, y está asociado sobre todo a poblaciones bajo explotación humana (Stockwell et al.,
2003) y de islas (Millien, 2006), elementos presentes en el caso de las poblaciones de jutía conga en
Cuba. Los estudios bajo la condición insular, sobre todo en pequeños roedores, dieron los primeros
ejemplos de microevolución, demostrando que estas poblaciones difieren de sus parientes
continentales en conductas, demografía, genética, fisiología, morfología, ecología y alozimas (Adler
y Levine, 1994; Pergamon y Ashley, 2001).
I.5. Selección natural y varaciones morfológicas adaptativas
La consecuencia de la selección natural casi siempre es la adaptación, tanto en su significado como
proceso operativo o como proceso histórico (Nespolo, 2003: Moreno, 2008). Ello supone entonces
que si estos caracteres han respondido a la selección natural es porque tienen una base genética
(Mousseaur et al., 2000; Charmantier y Garant, 2005). Pero no siempre un cambio adaptativo es
producto solamente de la selección natural, ya que la deriva genética también puede contribuir a ello
(Gould y Jonson, 1972). Por otra parte, el proceso de adaptación que más interesa al investigador
evolutivo es la adaptación local (Kawecki y Eberst, 2004), que se origina básicamente por selección
natural disruptiva o sexual (Van Doon et al., 2009), y puede tener como base genética para los
caracteres morfológicos las mutaciones de genes específicos, sobre todo reguladores del desarrollo,
estructurados en módulos (Klingenberg, 2008), o ser producto de la plasticidad fenotípica o norma
de reacción específica de cada gen (Travis, 1994; Trussell y Elter, 2001). Más recientemente, Yom-
Tov et al. (1999), Pergam y Achley (2001), Millien (2006) y Millien y Damuth (2004), trabajando
con roedores y mamíferos, demostraron que en poblaciones de islas (como es el caso de la jutía
conga), las variaciones morfológicas se aceleran en sus cambios evolutivos.
Los agentes selectivos que más parecen influir sobre la morfología de los roedores según Fleming
(1975), son el clima (sus variaciones estacionales) y la alimentación (a través de la productividad

17
primaria de la vegetación, influida a su vez por el tipo de suelo y el clima). Dos de los mejores
trabajos al respecto son los de Nevo et al. (1998), con dos especies en Israel, y el de Wolf et al.
(2009) en dos especies de nuevo México. Los efectos climáticos (temperatura, precipitación y
radiación solar) afectaron a todos los caracteres morfométricos (peso, tamaño, cola, pata y oreja), al
menos en algún periodo del año, pero no se especifica si fueron cambios genéticos o plasticidad
fenotípica. Otros estudios se han enfocado en las tasas metabólicas en relación a las variables
climáticas (Rezende et al., 2004; Novoa et al.; 2005), o en esta y la productividad del hábitat
(Bozinoviv et al., 2009). Este último trabajo emplea un índice climático (índice de Martonne), que
afecta a la tasa metabólica basal, que depende en parte del peso corporal.
Los cambios temporales del hábitat afectan los fenotipos de los individuos y la demografía de las
poblaciones. De acuerdo a la duración de estos cambios, se clasifican en cambios cíclicos repetibles
(estacionales), cambios direccionales (con una dirección en los cambios por largos periodos de
tiempo), y cambios erráticos (no predecibles). De acuerdo a la duración de las condiciones
favorables y desfavorables a los organismos en cada etapa del cambio, se clasifican en constantes,
estacionales, impredecibles y efímeros (Begon et al., 1998).
Las variaciones de temperatura y de recursos alimentarios en los trópicos son menos drásticas que
en las regiones templadas, a excepción de los periodos de seca y lluvia, lo que hace que la estrategia
de la historia de vida (y sus cambios morfológicos asociados fundamentalmente al tamaño) sean
muy diferentes (Cervantes, 1991). El cambio en uso de los recursos alimentarios en mamíferos no
especialistas constituye una respuesta común a los cambios estacionales en los trópicos; estos
cambios pueden darse dentro de la misma localidad, o como consecuencia de un cambio por
migraciones a otra localidad (Janzen y Wilson, 1983).
La estación de seca tiene un fuerte efecto en el patrón reproductivo de mamíferos neotropicales y en
la varianza de la supervivencia (Fleming, 1975; Smyth et al., 1982). El efecto de los cambios
climáticos durante la estación de seca, en poblaciones de roedores neotropicales, se ha estudiado por
Smythe et al. (1982), Janzen y Wilson (1983), Cervantes (1991) y Krisher (2006). Se ha constatado
que se recurre a la migración para escapar de los efectos negativos de la estación de seca.
Estudios morfométricos de variación en relación al tamaño, peso corporal, largo de la cola y largo
del pie de forma comparativa entre poblaciones de jutía conga, han sido efectuados en Cuba por
varios autores (Smith y Berovides, 1984; Berovides et al., 1990; Sánchez et al., 1992; Berovides y

18
Comas, 1993; Berovides, 2005; de la Fuente y Berovides, 2009, 2015), los cuales concluyen
coincidentemente la incidencia de valores altos de la cola y el pie con la actividad trepadora de la
jutía conga, así como diferencias de tamaño y peso en relación al hábitat. Sin embargo, en ninguno
de los trabajos se abordó la variación de los caracteres desde el punto de vista estacional.
En vertebrados, la variación en atributos ecológicos, tales como alimentación y uso de sustratos, está
casi siempre asociada con variaciones en morfología (Ricklefs y Miles, 1944). Esta posee, además,
elementos de relaciones filogenéticas, azar y respuesta adaptativa común, que a menudo no se
consideran en los estudios (Verde et al., 2017)
Desde el punto de vista adaptativo, la ecología y la morfología están relacionadas por demandas
funcionales comunes, pero la morfología puede reflejar también estructuras ancestrales retenidas,
más que adaptación a las presentes condiciones, de ahí la necesidad del enfoque filogenético,
aunque esto necesariamente, no siempre ocurre (Tulli et al, 2015)
Una vez identificadas variables morfológicas ecológicamente relevantes, podemos interpretar
similaridades entre especies y variables ecológicas y utilizar este conocimiento para interpretar otros
problemas ecológicos de la conservación (Verde et al., 2016).
Los roedores representan el grupo de mamíferos vivientes más diverso, con más de 2200 especies
descritas (Samuels y Van Valkenburgh, 2008). En los roedores, por su tamaño relativamente
pequeño, cambios sutiles en la morfología de sus huesos y tejidos blandos pueden tener relevantes
consecuencias funcionales, haciendo de ello un buen ejemplo de especialización ecológica, con o sin
cambios morfológicos radicales (Shephed y Digten, 2012). Alimentación y uso de sustrato (dieta y
locomoción) son importantes atributos morfológicos que han sido relacionados a la morfología en
este grupo (Carrizo et al., 2014), en especial en su dentición (Croft et al., 2011).
Dentro de los caracteres morfológicos, el tamaño corporal determina la historia de vida de la especie
según Begon et al. (1998). Estos autores plantean que la morfología general de un organismo puede
limitar la posible gama de todos los rasgos de su ciclo de vida. De ellos, el tamaño es quizás el
aspecto más evidente del ciclo vital de un organismo, y varía entre especies, poblaciones e
individuos.
Un tamaño grande permite aumentar la capacidad competitiva, incrementar el éxito como
depredador, reducir la vulnerabilidad como presa, mantener constantes funciones corporales, y

19
aumentar la eficacia reproductiva. El término tamaño no solo incluye el tamaño en sí, sino también
las reservas acumuladas, el estado corporal, etc (Begon et al. 1998). Los primeros trabajos que de
forma general establecen relaciones entre tamaño y otros aspectos de la biología del animal son los
de Cuthill y Houston (1997), que señalan mayor éxito reproductivo en animales grandes,
especialmente hembras de mamíferos, y los de Taylor et al. (1982), que señalan que animales
grandes requieren menos energía para transportar un peso dado sobre una determinada distancia, que
los pequeños. El trabajo clásico para mamíferos es el de Sterns (1983), donde estableció que la
historia de vida de cada especie covaría con el tamaño y su historia filogenética, pero también puede
hacerlo con las estrategias locomotoras (Calder, 1984). Otros estudios se refieren a la relación del
tamaño como limitante de los diseños y funciones de órganos (Lindstedt y Swain, 1988): la
selección de alimento y espacio (Brown y Mawcer, 1989; Van Soest, 1996), la ventaja fisiológica de
un gran tamaño bajo condiciones de estrés (Brown. 1995), y el gigantismo de pequeños mamíferos
en islas (Millien y Damuth (2004).
Para roedores en específico, los estudios de la variación del tamaño en relación con agentes
selectivos se han realizado para la rata de Nueva Guinea y la altitud, donde el tamaño disminuye con
esta (Taylor et al., 1985), para la rata del algodón de Estados Unidos en relación a la alimentación
(aumenta el tamaño en juveniles) y los cambios climáticos (lo disminuye en invierno; Donan y
Slade, 1995). En esta misma especie, la selección natural favorece un rápido crecimiento en invierno
entre individuos pequeños (Saber y Slade, 1985); en la ardilla común europea, el tamaño se
relaciona con la alimentación y el éxito reproductivo (Warters et al., 2007). Otros estudios abarcan
tres especies de roedores de islas del Pacífico (Yom-Tow et al., 1999) en relación a la densidad (lo
aumenta), competencia intraespecífica y depredación (en ambos casos lo disminuye); para varias
especies de roedores sudamericanos, Ebensperger y Cofre (2001) encontraron el tamaño asociado al
grupo social (lo aumenta), y Martin y Moestrup (2003) relacionan el tamaño corporal de 25 especies
de roedores con el grado de fragmentación del paisaje, provocando que las especies de mediano y
gran tamaño disminuyan su talla.
Con relación al peso o masa corporal, también se han hecho numerosos estudios en roedores, que
señalan los efectos de ser un animal poco o muy poco pesado. Cuthill y Houston (1997) señalan, por
ejemplo, que animales más pesados son más propensos a reproducirse exitosamente (en especial
hembras de mamíferos) y tienden a ser mejores competidores. Saeur y Slade (1985), y Campbell y

20
Slad (1993), analizaron el efecto del peso en la supervivencia de Sigmodon hispidus de
Norteamérica, viendo que la supervivencia de los adultos dependió de su peso corporal y de la
estación del año (mayor supervivencia de individuos pesados en primavera y menor en otoño).
Igualmente, tanto para roedores como para otros mamíferos, diferencias en el peso corporal explican
mucha de la variación en el costo mínimo de transporte, que lógicamente será mayor para un animal
pesado (Bogas y Frappell, 2000). Otros estudios demuestran la influencia del peso en la tasa de
metabolismo basal (en ratones, Speakmaqn et al., 2004), en la economía del agua (en el ratón
oliváceo de Chile, Bozinovic et al., 2011), y en la amplitud del nicho ecológico ( Jenning et al.,
2007).
Índices que relativizan el peso con relación al tamaño del animal (como condición física o índice de
masa corporal), también pueden ser indicativos de adaptación, aunque no se han encontrado trabajos
al respecto en roedores. En aves, Van der Veen (1999) establece que animales en buenas condiciones
físicas escapan más de los depredadores y son menos vulnerables a parásitos y otras enfermedades.
Por su parte, Schulte-Hostedde (2001) señala que, en muchos vertebrados, un peso relativamente
alto en relación al tamaño, es indicador de buenas condiciones y de mayor probabilidad de
supervivencia en tiempo de escasez de los alimentos y/o condiciones climáticas extremas.
Los factores más influyentes en el peso corporal son, además del tamaño del cual depende, el clima
(Wolf et al., 2009) y la alimentación (Warters et al., 2007), y con el tamaño, estos cambios pueden
ser por causas genéticas o ambientales (plasticidad fenotípica). En el carnero de grandes cuernos
(Ovis canadensis) está demostrado que los cambios estacionales de peso son debidos a plasticidad
fenotípica (Pelletier et al., 2007).
I.6. La especie objeto de estudio: La jutía conga (Capromys pilorides)
I.6.1.- Taxonomía
En Cuba existen tres géneros vivientes de jutías (Berovides y Pimentel, 2000) y en la actualidad
existen siete especies vivientes (Silva et al., 2007). La jutía conga (Capromys pilorides), dentro de
estas siete especies, es la de más amplia distribución geográfica (Smith y Berovides, 1984;
Berovides et al., 1990), se encuentra representada a todo lo largo y ancho de la isla principal y en

21
gran parte de los subarchipiélagos que la rodean. Hasta la fecha hay cinco subespecie descritas:
Capromys pilorides pilorides (Say, 1822) presente en la isla principal, Capromys pilorides relictus
(Allen, 1911) localizada al norte de la Isla de la Juventud, Capromys pilorides ciprianoi (Borroto,
Camacho y Ramos, 1992) presente al sur de la Isla de la Juventud, Capromys pilorides doceleguas
(Varona, 1980) localizada en el Archipiélago Jardines de la Reina, Capromys pilorides
gundlachianus (Varona, 1983) localizada en el archipiélago Sabana-Camaguey (Silva et al., 2007).
A continuación se muestra la clasificación taxonómica (Silva et al., 2007):
Reino: Animalia
Filo: Chordata
Subfilo: Vertebrata
Clase: Mammalia
Subclase: Theria
Infraclase: Eutheria
Orden: Rodentia
Suborden: Hystricognatha
Infraorden: Hystricognathi
Familia: Capromydae
Subfamilia: Capromynae
Género: Capromys
Especie: Capromys pilorides
Subespecie: C.p.pilorides
C.p.relictus
C.p.doceleguas
C.p.gundlachianus
C.pciprianoi
Los estudios referentes a la ecología de esta especie tienen gran significación para el diseño de una
metodología de su manejo, que propicie al mismo tiempo su conservación, enfatizando en los
aspectos que se discuten a continuación.

22
I.6.2.- Distribución y abundancia
Ha sido uno de los temas más estudiados, debido al interés que despierta la jutía conga, el mamífero
terrestre de mayor tamaño que constituye la mayor reserva de carne silvestre de la fauna autóctona
de Cuba (Berovides y Comas, 1993). La jutía conga es la más generalista de los caprómidos y puede
encontrarse en casi todas las regiones de Cuba, viviendo en hábitats tan disímiles como manglares,
pinares, cuabales, ciénagas y matorral xeromórfo, bosque montano y todo tipo de bosque
semideciduo (Berovides y Comas, 1990; Comas y Berovides, 1990). Su abundancia en términos de
densidad (individuos/ha) es muy variable (Comas y Berovides, 1989; Berovides y Comas, 1997a).
Las mayores densidades (90 individuos/ ha) se registran en los manglares pertenecientes a las
cayerias Jardines de la Reina (Berovides y Comas, 1991) y las menores (menos de 10 individuos/ha)
en las áreas de bosques (Berovides y Comas, 1990; Berovides y Comas, 1997a, b; Suárez, 2001;
Linares et al., 2010). Hernández (2003) determinó una densidad media de 2,3 individuos/ha en
bosques semideciduos y 5,7 individuos/ha en mogotes del valle de San Andrés.
Es muy importante determinar las causas de la reducción de las poblaciones de jutías. Al respecto, la
mayoría de los autores coincide en atribuirla a la actividad antrópica. Oliver (1977) planteó que para
la jutía de Jamaica, a su reducción poblacional han contribuido varios factores, entre los cuales
pueden citarse: la acción antrópica, cambios drásticos en el ecosistema natural en el que habitan,
depredación y competición con animales introducidos. Berovides y Comas (1993) reafirmaron estos
criterios para la jutía conga y precisaron que las densidades medias de 2 a 4 jutías/ha en los hábitats
boscosos parecen ser las normales bajo las condiciones de actividad humana en forma de caza
furtiva o presencia de otras especies de caprómidos. Cañizares (2003) determinó la densidad de jutía
conga en la localidad La Sabina de hábitat bosque siempreverde, con una densidad de 2.5 a 4.3
jutías/ha. Hernández (2005), en la Península de Guanahacabibes, determinó la densidad en
dependencia de la formación vegetal y el grado de antropización, reportando de 5 a 10
individuos/ha.
I.6.3.- Alimentación y coprofagia
Estos aspectos han sido evaluados por diferentes autores, entre ellos Manójina et al., (1989), Comas
et al., (1989b), Comas et al., (1989c), Hernández (2003), Rosa (2004), Pimentel y Hernandez
(2007), y Linares et al., (2010), quienes demostraron el amplio espectro de especies vegetales que

23
consumen las jutías, así como la no selectividad de sus partes, aunque en ocasiones prefieren corteza
u hojas, en dependencia del hábitat y de la densidad poblacional. Estos resultados sugieren que en
lugares de alta densidad las jutías son menos selectivas, ocurriendo lo inverso en sit ios de una
densidad baja, como es el caso del macizo forestal central de la cordillera de Guaniguanico.
La jutía conga es un fitófago generalista, con una actividad alimentaria polifásica en las 24 horas del
día. En Guanacahabibes, Manójina et al. (1989); determinaron que de 51 especies vegetales solo
rechazaron 12 y consumieron productos de origen animal; más recientemente, en esta misma área,
Linares (2005) en un estudio acerca de la utilización de las diferentes especies de plantas por la jutía
identificó un total de 40 especies, las cuales son incluidas en su dieta. De ellas, 34 son árboles (85%)
y 6 son arbustos (15%). Las diferencias en cantidad de las especies utilizadas se deben
probablemente a que el estudio de Manójina et al. (1989, 1990) se realizó con jutías en cautiverio, y
el de Linares (2005) en vida libre. Por su parte, Hernández (2003) determinó en el área del valle de
San Andrés que las jutías no consumían productos de origen animal; esto pudiera estar relacionado
con las diferencias en la disponibilidad de materiales alimenticios en distintas áreas. Se reportan
hasta la fecha un total de 105 especies de plantas que puede consumir la jutía conga, así como el
consumo en cautiverio de dos especies de lagartos anolis (Comas et al., 1994a; Silva et al., 2007).
Los requerimientos de agua son bajos para todas las jutías. En Guanacahabibes, Manójina et al.
(1989) demostraron que pueden vivir sin agua durante meses, aunque en condiciones normales el
consumo promedio diario es de 118 ml, oscilando entre 20 y 455 ml. En general se satisfacen de este
líquido con materiales alimenticios suculentos y lamiendo el rocío de las hojas y el que se deposita
en las oquedades de las rocas, ya que la estructura de los riñones les permite la máxima reabsorción
de agua a partir de la filtración glomerular (Sánchez et al., 1992). Johnson et al. (1975) expresaron
que los riñones de la jutía conga presentan grandes asas de Henle, típico de los riñones capaces de
realizar una abundante reabsorción del agua contenida en la orina.
Estudios realizados en vida libre y en cautiverio han reportado la coprofagía (ingestión de bolos
fecales tomados directamente del ano) conducta a la que los animales recurren para la reabsorción
de minerales y vitaminas del complejo B sintetizadas en el intestino por la acción microbiana. La
coprofagía es practicada por las jutías en algún momento de su periodo de descanso, sentadas en el
suelo o sobre la rama de un árbol donde los bolos fecales son tomados directamente con la boca,
aunque en ocasiones son tomados con la mano y llevados a la boca con una duración media de 6.04

24
minutos para las hembras y 7.07 minutos para los machos y una ingestión media de 8.9 bolos las
hembras y 8.4 los machos. Los neonatos practican la coprofagía a los dos o tres días de nacidos y
pueden ingerir hasta ocho bolos fecales seguidos (Silva et al., 2007).
I.6.3.- Depredadores y parásitos
La depredación es un factor importante que incide sobre la decadencia de las poblaciones de jutías,
pero para muchos es aun más importante la depredación directa por el hombre, ya que la carne de las
jutías es nutritiva y sabrosa, y estas han sido consistentemente cazadas por hombres y perros desde
los tiempos precolombinos. Se supone que la cacería excesiva dio como resultado la exterminación
de varias especies de jutías, cuyos fósiles son muy conocidos, por haber sido hallados en los
residuarios aborígenes, que marcan el lugar donde se asentaron las comunidades primitivas (Varona,
1974).
Entre las principales especies depredadoras de la jutía conga se encuentra el perro jíbaro (Canis
lupus familiaris), que en las Bahamas es señalado por Clough (1972) como depredador natural.
Pimentel (1987) señala que solo en una noche llegan a eliminar decenas de jutías atacando en
manadas o en solitario, aunque utilicen solo unas pocas para alimentarse. El majá de Santa María
(Epicrates angulifer), considerado uno de los depredadores naturales en Cuba (Hernández, 2005), y
el cocodrilo cubano (Crocodylus rohombifer) en la ciénaga de Zapata (Soberón et al., 2001), se
consideran también importantes depredadores. Otras especies como el gavilán del monte (Buteo
jamaicenis solitudinis), la lechuza (Tito alba furcata) y la mangosta (Herpestes javanicus
auropunctatus) han sido implicadas también en la depredación de jutías (Silva et al., 2007)
En diferentes estudios realizados por varios autores se han encontrado cuatro especies de protozoos,
nueve especies de helmintos, cinco especies de ácaros y una especie de insecto parasitando a la jutía
conga. En cría en cautiverio se han reportado como enfermedades más frecuentes la teniasis,
coccidiosis, sarna, leptospirosis y toxoplasmosis (Silva et al., 2007). A partir de órganos internos de
la jutía conga se aislaron cepas pertenecientes a Mycobacteriun tuberculsis, subtipos del complejo
avium- intracelllulare (Cornide et al., 1987; Mederos et al., 1992). Merino y Bouza (1988)
detectaron microfilaria en la sangre de la jutía conga, donde comprobaron sus efectos en los tejidos
pulmonares y esplénico con neumonía intersticial y esplenitis aguda.

25
I.6.4.- Reproducción
La reproducción ha sido profundamente estudiada por diversos autores. Smith y Berovides (1984a)
plantean que en la jutía conga la pareja en edad reproductiva realiza la cópula en horas de la tarde, la
gestación dura 142 días, las hembras presentan antes del parto las mamas inflamadas, con un rango
de embriones o neonatos de 1.9 a 3.3; las crías por lo general a los dos meses aún maman y no
toman alimentos, aunque en cautiverio se ha observado la ingestión de alimentos sólidos con unos
cuantos días de nacidos (Comas et al., 1994b; Berovides, 2008). La reproducción tiene lugar en
cualquier época del año, y generalmente presentan un parto al año con un rango de 1.5 a 1.7 por la
duración de la lactancia (tres a seis meses).
Comas et al. (1994a) afirmaron haber observados manifestaciones claras de estro en jutías congas
cautivas. Taylor (1970), mediante exudados vaginales, estableció que el ciclo estral de la especie es
de 16.3 días con una amplitud de 13 a19 días. La jutía es un animal con una gestación de 17-18
semanas, con más de un ciclo estral al año, y el parto dura tres horas como máximo. Los picos de
gestaciones ocurren en la estación de seca y los picos de partos en mayo y julio, estableciéndose que
depende de la disponibilidad de alimentos (Manójina, 1984; Manójina y Abreu, 1987a). En la jutía
conga un macho puede tener varias hembras, y los más jóvenes que no han formado sus familias, así
como los sub-adultos desplazados del grupo al llegar a su madurez sexual, tienden a desplazarse
solos hasta encontrar hembras jóvenes con las cuales formar un nuevo grupo familiar. Los grupos
familiares son muy filopatricos. Este comportamiento puede verse también con los viejos
reproductores que son desplazados por otros machos más fuertes (Hernández, 1985; Berovides,
1998), como ocurre en otras especies parecidas, como Myocastor coypus (Dagault y Suboreau,
1990).
Las crías recién nacidas tienen los ojos y el pabellón de las orejas abiertas, son capaces de una
locomoción normal al cabo de una hora de nacidas y son expulsadas del grupo familiar al producirse
un nuevo parto (Smith y Berovides, 1984a; Frías et al., 1987). Otras variables reproductivas de la
especie, tanto en vida libre como en cautiverio, como la duración de la gestación, número de
camadas por año, crías por parto y edad al destete fueron estudiadas por Bucher (1937) y Taylor
(1970). Manójina y Abreu (1987b) describieron un parto de Capromys pilorides: dos crías nacieron
con un intervalo de 20 minutos, de posición cefálica; la madre se comió la placenta y lamió a los
recién nacidos.

26
Resultados relevantes fueron obtenidos por Gutiérrez et al., (1999) al estudiar la dinámica
reproductiva de dos poblaciones de jutía conga (Jardines de la Reina y Najasa). Durante un año (de
noviembre de 1991 hasta octubre de 1992), investigaron los cambios trimestrales de los índices
reproductivos y compararon dicha dinámica entre estas poblaciones con diferentes hábitats: una de
bosque y otra de manglar. En ambas analizaron 118 machos y 334 hembras. A los machos se les
midió el largo corporal y el largo del testículo, para determinar el índice reproductivo (IRM), según
la formula:
IRM= (largo del testículo/largo del cuerpo) x 100.
A las hembras se les tomó el estado reproductivo por observación de los cuernos uterinos y las
mamas, definiendo los estados reproductivos: hembras vacías, hembras lactantes y lactantes-
gestantes. Al analizar estadísticamente los resultados (frecuencia, valores medios y coeficiente de
variación), los compararon entre trimestres y entre poblaciones, y concluyeron que los índices
reproductivos son siempre más altos en las estaciones de mayor disponibilidad de alimentos y los
hábitats más ricos en estos recursos. Miguel (2008) demuestra que los parámetros reproductivos en
los machos no varían de forma estacional, a diferencias de las hembras que cambian de acuerdo a la
variedad y disponibilidad de alimentos. Además, el potencial reproductivo en las poblaciones
estudiadas es más elevado en la población de bosque que en la población de manglar.
I.6.5.- Hábitos
La jutía conga es una de las más terrestres de todas las especies de jutías que habitan en Cuba (Silva
et al., 2007). Berovides et al., (1990) clasifica los hábitos de la jutía conga en: terrestre (cuando los
animales viven exclusivamente en el suelo), semiterrestre (cuando los animales viven en el suelo,
pero suben ocasionalmente a los árboles), semiarborícola de árboles (cuando frecuentan los árboles),
y semiarborícola de mangle (si viven en manglares).
La cola de la jutía conga es la única dentro de las especies de jutía que no es prensil y no
experimenta autotomía caudal. Este apéndice puede moverse lateral y verticalmente; al caminar por
la rama de los árboles mantiene el extremo de la cola diagonalmente sobre la rama con el ápice algo
curvado hacia abajo, lo cual ayuda al animal a sostenerse (Bucher, 1937).

27
La actividad de la jutía conga es crepuscular nocturna, aunque presenta ciclos de actividad polifásico
(alimentación y descanso) en las 24 horas de día. Bucher (1937) tuvo razón al afirmar que la conga
es más activa de día que la carabalí o andaraz. Milisnikov (1993) en un estudio de la retina a niveles
altos de iluminación encontró que la conga presenta la retina más adaptada a niveles altos de
iluminación que la carabalí.
I.6.6.- Conservación y designaciones no taxonómicas
Históricamente, se ha tratado de definir la conservación como el estado de armonía entre el hombre
y la tierra, entendiéndose por armonía el balance y la estabilidad que deben de tener todas las
acciones del hombre hacia la naturaleza (Soulé, 1985). Esta concepción se ha modificado con el
paso del tiempo. En su concepción moderna, la conservación de un recurso natural biótico implica
tanto su protección como su explotación racional (Primack, 1995), basados ambos aspectos en el
conocimiento de la ecología de las especies (Heywood y Watson, 1995; Prescot-Allen y Prescot-
Allen, 1996).
Los estudios de manejo de recursos y condiciones en vida libre (in situ), sientan las bases para
decidir el uso sostenible de la especie en aspectos tales como: en qué época extraer los animales,
cantidad a extraer, y factores claves del hábitat que no deben alterarse durante las extracciones
(Prescot-Allen y Prescot-Allen, 1996; Cawghley y Gunn, 1996). El manejo in situ de la fauna
silvestre se realiza a través de medidas biotécnicas, que constituyen un aspecto de una disciplina
conocida como biocinegética, que trata de resolver de forma científica la protección y el aumento
cualitativo y cuantitativo de la fauna silvestre (McNeely, 1994). La mejor estrategia para conservar
la biodiversidad es mantenerla en su ambiente natural donde se ha desarrollado evolutivamente
(conservación in situ), ya que también es importante conservar sus interacciones, los procesos
ecológicos en que participa, así como sus procesos de evolución natural (Primack, 1995).
La biología de la conservación está encaminada al estudio de los impactos humanos sobre las
especies y a la elaboración de aproximaciones metodológicas que aminoren dichos impactos. Es una
ciencia multidisciplinaria que surge como una necesidad a la crisis de la biodiversidad, aspectos que
no son ajenos a la fauna cubana (Berovides y Gerhartz, 2007).
En la actualidad, la conservación de las especies al nivel infraespecífico trata de distinguir las
denominadas Unidades de Significación Evolutivas (USEs) como poblaciones que poseen atributos

28
genéticos significativos que determinan el potencial evolutivo, con el propósito de preservar la
biodiversidad, los procesos evolutivos y los ecosistemas (Ryder, 1986). Esta categoría se le otorga,
por lo general, a poblaciones bajo algún grado de amenaza que se encuentran aisladas
geográficamente de poblaciones conespecíficas, y que presentan características ecológicas y
genéticas únicas (Ryder, 1986).
Con posterioridad se ha reelaborado el concepto de USE, redefiniéndolo como Unidades
Operacionales de Conservación que combina información biológica con la socioeconómica
(Dodson et al., 1998), Unidades de Manejo=Poblaciones Demográficamente Independientes
(Paterson, 2005), poblaciones con flujo genético nulo o escaso y nicho ecológico diferenciable
(Crandall, 2000, 2009 ), y Unidades Designables (Green, 2005), que son poblaciones que presentan
un estatus de conservación diferente, son genéticamente diferenciables y poseen disyunción de
rango y variación geográfica. Todas ellas reciben el nombre general de unidades de manejo para la
conservación (Vogler, 1994, Fraser y Bernatchez, 2001).
Los estudios en especifico de la variación morfológica entre poblaciones pudieran ser indicativos de
diferenciación filogenética y adaptaciones diferenciales a la ecología local, y por consiguiente,
proporcionar indicios de la existencia de unidades de manejo para la conservación (Bush y Adams,
2007). La morfología general de una especie puede limitar la posible gama de todos los rasgos de su
historia de vida, siendo el tamaño el más importante (Begon et al., 1998).
Para la conservación de las especies resulta necesario conocer si existe variabilidad genética y
fenotípica entre sus poblaciones, con el objetivo de determinar si estas deben ser manejadas como
unidades de manejo para la conservación de forma independiente, o si el manejo debe realizarse al
nivel de especie, es decir, aplicar un plan de manejo común para todas las poblaciones. El
mantenimiento de la variabilidad genética y fenotípica de caracteres adaptativos (incluyendo los
caracteres morfológicos) se plantea que debe ser un objetivo primario en la conservación de las
especies (Erickson et al., 1993). Ello implica el análisis de un número adecuado de poblaciones que
habiten en diferentes condiciones ambientales, utilizando la interacción de genotipo por ambiente
con variables ambientales climáticas y edáficas (Humphries et al., 1995).
La evolución de caracteres adaptativos puede ocurrir en una escala de años o décadas, lo que resulta
de particular importancia para la conservación, sobre todo en lo que se refiere a poblaciones bajo
explotación humana (Stockwell et al., 2003) y de islas (Millien, 2006), situación que se corresponde

29
con la especie estudiada en esta investigación. Esta rápida adaptación a menudo está asociada a los
mismos factores antropogénicos responsable de la actual crisis de extinción de especies, que incluye
en especial sobreexplotación (Conover y Munich, 2002) y alteración del hábitat, que son en Cuba
los principales factores de amenaza para la jutía conga.
Según Stockwell y Ashley (2004), la incorporación de caracteres adaptativos en los planes de
conservación de especies rompería el pensamiento tradicional de que las especies cambian en largos
periodos de tiempo, incorporaría a su arsenal metodológico no solo los marcadores moleculares (en
su mayoría neutrales), sino también importante caracteres cuantitativos adaptativos, y por último, se
podrían medir las fuerzas selectivas y predecir los cambios genéticos y fenotípicos influenciados por
las actividades humanas, que son hoy una potente fuerza evolutiva. Independiente de su utilidad
para la conservación de especies, los estudios de adaptación fenotípica local proporcionan una
importante información sobre el poder de la selección natural sobre el flujo del gen y otras fuerzas
evolutivas. Ellos son un paradigma para probar hipótesis evolutivas sobre los caracteres favorecidos
o no por factores medioambientales particulares (Kaweki y Ebert, 2004).
La jutía conga ha sido estudiada en cuanto a su variación morfométrica entre poblaciones, y dichos
caracteres pudieran ser indicativos de diferenciación filogenética y adaptaciones diferenciales a la
ecología local, y por consiguiente, proporcionar indicios de la existencia de unidades de manejo para
la conservación La determinación de estas unidades es de suma importancia tanto para la
conservación de la especie (Crandall, 2000: Fraser y Bernalchez, 2001) como para su mantenimiento
en cautiverio, ya que se diferencian linajes genéticos con distintas historias evolutivas. En el caso de
la conservación, dichas unidades pudieran necesitar diferentes acciones de manejo, y el caso de la
cría en cautiverio, evitar mezclas de diferentes linajes genéticos en los que se rompe la coadaptación
de los genes específicos de cada población (Templeton, 1986; Barraowclough et al., 1996).
I.6. Uso de la jutía conga como recurso natural del bosque (recurso forestal no maderero)
La jutía conga sin duda ha constituido y constituye aún hoy un valioso recurso natural faunístico,
más concretamente, un recurso forestal no maderable desde los aborígenes hasta el actual uso de
subsistencia por pescadores y campesinos. Esta especie, además de su valor intrínseco como especie
peculiar de la fauna cubana, posee valores materiales reales (Berovides y Comas, 1993) que, de

30
forma general o localizada, utilizan las poblaciones de campesinos y pescadores, pero posee también
valores potenciales que pueden ser utilizados por el sector estatal. Su uso principal es alimentario
(carne) y también pueden utilizarse su piel y grasa (medicamento) y el propio animal como mascota
o para el ecoturismo.
La jutía conga es considerada un “microganado” (NRC, 1991), como otros muchos pequeños
mamíferos. Los roedores están en mayoría en este grupo, y por esta razón muchas de sus especies se
estudian (Jorri, 2001; Wilson, 2011) y se protegen (Amori y Gippoliti, 2003).
El uso de las jutías por las comunidades de campesinos y pescadores ya se cita desde el siglo XIX
(Gurdlach, 1877) y por muchas otras fuentes anteriores. Para la jutía conga en específico, su valor
como recurso natural fue evaluado por Berovides y Comas (1993) y Berovides et al. (2009). Linares
(2008) evaluó en la península de Guanahacabibes el potencial cárnico por hectárea de bosque que
produce la jutía conga, encontrando una producción de 1.5 a 1.9 kg de carne por hectárea por año.
Berovides et al. (2007) demuestra el uso sostenible, pero en el límite de esta especie por parte de los
pescadores del poblado de Carahata, y Pimentel (2009) analizó las causas de su extirpación en la
cordillera de Guaniguanico por sobreexplotación de la especie. Tradicionalmente, la jutía conga ha
sido criada por los campesinos cubanos y ya existen normas para este fin (Pimentel, 2008).
Las características de la biología individual y poblacional de la jutía conga que resultan de
relevancia para su utilización sostenible como recurso natural (Berovides et al., 2009), son:
Tamaño adulto (longitud cabeza tronco) 460 a 520 mm
Peso adulto 3 (manglar) a 4 (bosque) kg
Madurez sexual 1 año
Actividad sexual Todo el año, pico de partos en mayo y junio
Camadas/año 1
Hijos/camada 1 a 3
Duración ciclo estral 13 a 19 días, poliéstrica
Duración gestación 96 a 157 días
Duración lactancia 3 a 6 meses
Longevidad máxima (cautiverio) 9 a 10 años
Peso al nacer 150 g

31
Organización familiar Poliginia (un macho con 3 a 4 hembras)
Eficacia reproductiva 0.20 kg de neonatos/kg de la hembra
Coloración Agutí (bosque), variada (manglar)
Alimentación Fitófaga generalista (105 especies de plantas), coprofagía
Refugios Cuevas en rocas y tierra, raíces y copa de árboles
Actividad Crepuscular nocturna
Distribución geográfica Toda Cuba, incluyendo isletas (cayos)
Hábitat Bosque, manglares, ciénagas y matorrales secos
Densidades 1.1 a 7.4 indiv/ha (bosques)
17.6 a 130.0 indiv/ha (manglar)
Tasa de incremento anual 20 a 30 %
Productos útiles Carne, piel y grasa
Rendimiento en canal 45% (manglar) a 50% (bosque)
Eficiencia en la producción de carne 0.83 kg de canal /animal/año
Algunos autores e instituciones han señalado a los grandes roedores como una de las fuentes
alimentarias principales a explotar dentro de los países en vía de desarrollo (FAO-PNUMA, 1985;
Kyle, 1987; National Researcha Council, NCR 1991; Jorri, 201; Wilson, 2011).
I.6. Factores reguladores de las poblaciones de la jutía conga
Las alteraciones del hábitat, la disminución de recursos limitantes (reguladores), y el uso directo por
el hombre, son las causas involucradas en la disminución numérica o extirpación de muchas
poblaciones de jutía conga por toda Cuba. Concretamente para esta especie, y en dependencia de su
hábitat, se ha descrito:
Hábitats
Bosques Manglares
Para alteraciones del hábitat.
Disminución o fragmentación
por deforestación.
Cambios de composición y
Disminución por enfermedad y
muerte del mangle.

32
estructural.
Por la disminución de
recursos limitantes. Perdida de refugios y alimento. Pérdida de refugios.
Por el uso directo por el
hombre.
Perdida de reproductores y
reclutas.
Perdida de reproductores y
reclutas.
Al igual que para muchas otras especies de vertebrados cubanos, para esta especie los factores
naturales reguladores claves son los refugios, al menos en el hábitat de bosque, donde presentan un
amplio nicho trófico (Comas et al., 1994). Hairston et al. (1960), Beck (1995), y Johson et al.
(1975) reconocen que para esta especie en cautiverio, el suministro de refugio es muy importante.
Las evidencias en vida libre para este planteamiento provienen de:
- Un análisis de regresión múltiple de la densidad de jutía conga sobre siete variables de la
vegetación y el sustrato en el área protegida Sierra del Chorillo, Camagüey (Comas et al.
1989), donde se determinó que el grado de rocosidad (fuente de refugios) fue el factor que
más influyó sobre la densidad.
- El gran aumento de la densidad de jutía conga en el área de la base naval de Guantánamo se
considera relacionado con el abandono de las trincheras (montículos de tierra con cavidades), que
utilizaron las jutías como refugio.
- La mayor densidad que se da para subadultos en refugios conformados por sitios con vegetación
tipo bejuqueras y zarzales.
- Al análizar los factores naturales reguladores potenciales de las poblaciones de jutía conga
(alimento, refugio, clima, competidores, depredadores y patógenos), para los cuales la especie o es
generalista (alimento) o resistente (patógenos), se aprecia que sólo los refugios quedan entonces
como factores naturales potencialmente reguladores.
- A pesar del intenso uso que hacen de esta especie de jutía los campesinos y pescadores en toda
Cuba, aún persiste en todos sus variados hábitats, desde los manglares de puro mangle rojo,
pasando por las maniguas costeras, hasta los diferentes tipos de bosque casi puros o degradados
que aún quedan en Cuba. Los estudios más recientes sobre su abundancia son los de Berovides y
Comas (1997a, b) y Berovides (1998b), que resumen y amplían estudios anteriores (Berovides y
Alfonso, 1987; Comas y Berovides, 1990). El resultado básico es que rara vez las densidades de

33
las jutías de bosques superan los 10 individuos/ha. ¿Qué factores determinan este límite? Al
parecer, la respuesta está en los refugios y la depredación humana. En los hábitats de bosques, la
jutía conga posee dos tipos de refugios básicos, cuevas y árboles (Gundlach, 1877); de estos
últimos, al parecer no aprovechan los huecos de los troncos (pero sí los que se forman entre las
raíces), y en su lugar utilizan como protección las epífitas o las innumerables lianas y bejucos
(bejuqueras) que se encuentran en las ramas y copas de dichos árboles. En otros hábitats (ciénagas,
manglares y maniguas costeras) los refugios son otros (cayos de mangle prieto, raíces de mangle
rojo, árboles huecos y arbustos).
En determinadas localidades boscosas se pueden dar las siguientes situaciones con respecto al uso
de los refugios cueva y árbol:
1. Solo se utilizan las cuevas y los individuos no suben a los árboles. Ejemplo: sur de la Isla de la
Juventud.
2. Solo se utilizan los árboles pues no hay cuevas o éstas son muy escasas. Ejemplo: Cayo Saetía.
3. Existen árboles y cuevas y ambos se utilizan. Ejemplo: Área protegida Sierra del Chorrillo,
Najasa.
Las cuevas evidentemente se utilizan para el reposo diurno y la reproducción, y al parecer debe
ocurrir lo mismo con los árboles cuando no hay cuevas; en este último caso, dichos árboles también
pudieran ser sitios de alimentación. Cuando concurren árboles y cuevas, es posible que se prefieran
las cuevas como refugio, pero si estas son limitadas, se utilizarían entonces los árboles. La brillante
observación de Gundlach (1877) de que la jutía conga se encontraba en árboles gruesos, sugirió la
hipótesis de que, cuando se da la condición de refugio solo en los árboles, el desarrollo de estos
(medido por su diámetro a la altura de 1.3 m o DBH) y su abundancia podrían ser factores limitantes
claves de la distribución y abundancia de jutía conga en los hábitats boscosos. Si, además, estos
mismos grandes árboles son utilizados preferentemente como sitios de alimentación nocturna,
entonces el efecto limitante será más marcado, independientemente de la existencia o no de cuevas.
Esta hipótesis fue confirmada por Cañizares y Berovides (2007).

34
I.7. Fundamentación teórica de la presente tesis
El problema sobre el cual se fundamentala presente investigación es que:
Las poblaciones de jutía conga ocupan un amplio espectro de hábitas (formaciones vegetales) en
Cuba, pero no existen estudios sobre variaciones morfológicas entre poblaciones que incluyan los
posibles factores determinantes de las mismas, a efectos de inferir caracteres adaptativos que
pudieran considerarse para delimitar diferentes unidades de significación evolutiva para la
conservación de la especie.
Asumiendose como hipótesis que:
Si las condiciones de los distintos hábitats en cuanto al clima y tipo de vegetación son
suficientemente diferentes entre ellos, entonces la morfometria y morfofisiología de las poblaciones
de jutía conga que los habitan presentarán adaptaciones locales diferenciadas en relación con dichos
hábitats, y estos representarían unidades de manejo diferenciadas para la conservación de la especie.
Tiene como objetivo general:
determinar la relación entre variables morfológicas adaptativas, y factores climáticos y de estructura
de la vegetación, en poblaciones de jutía conga que viven en las formaciones vegetales de manglar,
bosque y vegetación xerofítica en Cuba.
Como Objetivos específicos se plantean:
1. Comparar las variables morfológicas adaptativas refereridas a la morfometría (peso, medidas
lineales y sus índices) y la morfofisiología (índices nutricionales y de retención de agua)
entre poblaciones de jutía conga que viven en diferentes formaciones vegetales.
2. Relacionar las variables morfológicas con un índice climático y de estructura de la
vegetación.
3. Determinar la variación de la estructura de tamaño corporal y el grado de dimorfismo sexual
entre poblaciones.
4. Evaluar el grado de variabilidad fenotípica entre caracteres y entre poblaciones.
5. Determinar unidades de significación evolutiva para el manejo y la conservación de las
poblaciones estudiadas, sobre la base de los resultados obtenidos.

35
I.1.7.1.- Novedad científica y valor teórico de la tesis
La novedad científica de la tesis es que por primera vez se realiza un estudio comparativo de
variables morfométricas y morfofisiológicas adaptativas considerando:
Varias poblaciones de jutía conga en diferentes hábitats.
Su dinámica espacio- temporal
La adaptación local dentro y entre las metapoblaciones estudiadas.
La designación de unidades de manejo para la conservación, como categorías
infraespecíficas, lo cual permitirá establecer planes de gestión diferenciados para las
poblaciones de jutía conga.

36
CAPÍTULO II. Materiales y Métodos Generales
II.1. Poblaciones muestreadas: Caracteristicas de las áreas de estudio
Las poblaciones estudiadas corresponden a diferentes áreas geográficas que pertenecen al Sistema
Nacional de Áreas Protegidas. Se analizaron ocho poblaciones (Figura 1) caracterizadas por
diferentes tipos de hábitat (formaciones vegetales) y categorías de manejo
(CNAP/CITMA/GEF/PNUD, 2002). A continuación, se describen las principales características de
las áreas de estudio, de acuerdo a la información contenida en los planes de manejo de las mismas,
que se encuentran en el Centro Nacional de Áreas Protegidas (CNAP). Se tomaron en cuenta cuatro
aspectos básicos que se considera pueden afectar a las poblaciones de jutía conga: localización,
clima, vegetación y actividades humanas. Las formaciones vegetales bajo el acápite Vegetación se
describen con detalles en Capote y Berazain (1985) y Borhidi (1996).
Figura 1. Distribución de las áreas geográfica donde se encontraban las poblaciones de jutía conga
(Capromys pilorides) estudiadas. En cursivas los nombres de las poblaciones. APRM = Área
Protegida de Recursos Manejados. PN = Parque Nacional. RE = Reserva Ecológica. PNP =
Paisaje Natural Protegido.
II.1.1.- Parque Nacional Jardínes de la Reina (Jardínes)
Localización

37
El grupo insular Jardines de la Reina se extiende desde el Golfo de Guacanayabo hasta la Bahía de
Casilda, en la parte meridional de la Isla de Cuba. Su extensión es de 360 km y está formado por
661 cayos.
Clima
Las temperaturas oscilan entre los 26 y 27 oC por la mañana y entre los 29 y 31
oC por la tarde, con
excepción de los días afectados por los frentes fríos (24 oC en la mañana y hasta 27
oC en la tarde).
La dirección predominante de los vientos está entre el Sureste y el Sur. En horas de la mañana
aparece el cielo con pocas nubes, que van aumentando a medida que pasa el día, ocurriendo
tormentas y chubascos con bastante frecuencia. En verano la humedad relativa está entre 85 y 90 %,
mientras que en invierno es menor, entre 70 y 75 %.
Vegetación
Desde el punto de vista fitogeográfico estos cayos pertenecen al Sector Cuba Central, Subsector
Cuba Centro Occidental y al distrito Cayería Sur.
Se determinaron en el área de estudio seis tipos de formaciones vegetales, siendo la de interés para
este trabajo el bosque de mangle. Esta formación vegetal está constituida por una franja mixta de
mangle rojo (Rhizophora mangle) con mangle prieto (Avicennia germinans), Laguncularia
racemosa (patabán) y Conocarpus erecta (yana) a manera de sucesión desde la interfase agua - tierra
hacia tierra firme.
En algunos cayos, esta formación puede estar representada solo por Rhizophora mangle, que se
encuentra con mayor frecuencia en la línea costera sobre un sustrato turboso, llegando a ser la
formación más representativa y de mayor extensión en todo el archipiélago. Su altura es variable
según las características del sustrato, y puede llegar a alcanzar los 8 m. En estos cayos se tomaron
todas las jutías conga para el estudio.
Actividades humanas
Jardines de la Reina carece de asentamientos poblacionales. Las comunidades que se vinculan con
este Parque Nacional son Júcaro (1696 pobladores), Playa Florida (430) y Santa Cruz del Sur (17
300). Las principales actividades humanas en la zona son la pesca de plataforma, industria pesquera,
camaronicultura, actividad agropecuaria, el desarrollo del turismo para la explotación de Jardines de
la Reina, y la actividad forestal.

38
Actualmente se realiza una tala selectiva de mangle al año, con lo que se extrae corteza para la
extracción de taninos usados para curtir pieles y como lubricantes para la explotación petrolera.
Además de esta extracción comercial, se realiza tala ilícita, así como pesca y caza ilícita de especies.
II.1.2.- Área Protegida de Recursos Manejadas Sabana-Camaguey (Sabana).
Localización
Esta región fue clasificada como área protegida de recursos manejados. Contiene sistemas naturales
y seminaturales y es objeto de actividades de manejo para garantizar la protección y el manejo de la
diversidad biológica, así como para proporcionar al mismo tiempo un uso sostenible de productos
naturales y servicios para satisfacer necesidades locales o nacionales.
El estudio se llevó a cabo dentro de la zona que alcanza Cayo Blanquizal a Cayo Cristo, que al sur
limita con la línea de costa de los municipios Corralillo, Quemado de Güines y Sagua la Grande.
Dicha zona se destaca por sus altos valores y su estado de conservación, y la conforman un conjunto
de cayos y áreas marinas ubicadas en la parte central del archipiélago, formando el Refugio de
Fauna Las Picuás.
Clima
El clima de la zona presenta una influencia marítima predominante. Por esta causa, el contenido de
humedad del aire es muy elevado, superior al 70 % y el régimen térmico es también elevado y
uniforme, con una temperatura media mensual que oscila entre 22 ºC y 27 ºC. La lluvia se
caracteriza por totales anuales entre 1000 y 1200 mm, concentrada mayormente en los meses de
mayo hasta octubre; en los meses del período menos lluvioso (noviembre hasta abril) se observa
entre 300 y 400 mm de lluvia.
Los vientos en la zona tienen un comportamiento muy estable, con dirección predominante de
región Este. En la mayoría de los meses del año la velocidad media del viento oscila alrededor de los
15 km /h. Los ciclones tropicales constituyen el fenómeno meteorológico más peligroso que puede
afectar a la biota del lugar.
Vegetación
De forma general, los cayos y cayuelos del noroeste de Villa Clara, pertenecientes al archipiélago de
Sabana, se caracterizan por presentar un sustrato areno fangoso. De ahí que la vegetación en la

39
mayoría de ellos esté dominada por el bosque de mangle y, en algunos casos donde se presentan
formaciones de dunas, existe un tipo de formación arbustiva con una composición florística
homogénea.
El bosque de mangle (manglar) donde predominan las jutías conga, está formado por las cuatro
especies existentes en Cuba: mangle rojo (Rhysophora mangle), mangle prieto (Avicennia
germinans), Patabán (Laguncularia racimosa) y Yana (Conocarpus erectus). Están presentes
también Batis maritima y Salicornia perennis.
Actividades humanas
El primero de enero de 1959 se crea en el área una Cooperativa Pesquera, definiéndose los renglones
fundamentales que son captura de langosta y peces. Actualmente la población de Carahata, la que
más influye en el área, ha ascendido a un total de 666 habitantes y además de la pesca se desarrolla
la ganadería y la agricultura. Las actividades humanas que pueden afectar a las poblaciones de jutía
conga son la tala de especies de la flora (Coccotrinax litoralis, Eugenia sp) para su utilización en la
confección de artes de pesca y la caza ilícita por parte de pescadores.
II.1.3.- Parque Nacional Guanahacabibes (Guanahacabibes).
Localización
Este Parque Nacional se ubica en la porción más occidental del municipio Sandino, ocupando un
área de 39 830 ha. Geográficamente está comprendido dentro de la Región Cuba Occidental, en la
sub-región Penínsulas Cársicas, en el Distrito Pinareño, Sub-Distrito de la Llanura Costera
Meridional y Occidental, formando el grupo de paisajes de la Llanura Cársica y Pantanosa de
Guanahacabibes
El Parque Nacional Guanahacabibes constituye el núcleo de la Reserva de la Biosfera Península de
Guanahacabibes, el mismo cuenta con un área terrestre de 23 880 hectáreas y un área marina de 15
950 hectáreas, para un total de 39 830 hectáreas.
Clima
En la península las temperaturas oscilan entre 21.5 °C de promedio de mínimas y 29.3 °C de promedio
de máximas, siendo 25.8 °C la media anual. Las lluvias se comportan con un acumulado promedio de 1
443.1 mm/año. La humedad relativa promedio anual en la península es del 81%. Los vientos

40
predominan fundamentalmente del Este y del Noreste, presentándose en ambas direcciones en el
80% de los casos. El predominio de las bajas presiones extratropicales, principalmente en el mes de
marzo, genera fuertes vientos del Sur con altas probabilidades de penetraciones del mar. Las
mayores probabilidades de afectaciones por tormentas locales severas y ciclones tropicales se
observan entre los meses de junio y noviembre.
Vegetación
Se presentan nueve tipos de vegetación en el Parque Nacional Guanahacabibes, siendo las de
nuestro interés el bosque semideciduo y el manglar, ya que una de la población de jutía estudiada
habitaba el ecotono de estas formaciones vegetales, en específicos el bosque con el manglar de
Conocarpus erecta (yana) y la otra solo el bosque semideciduo. Este bosque es la formación
predominante dentro del territorio del Parque y la más importante por sus potencialidades para la
conservación. Ocupa el núcleo cársico principal, sobre todo en la porción central, desarrollado sobre
afloramientos calizos. Generalmente forma dos estratos arbóreos, uno inferior de 6 a 10 m de alto y
otro superior que alcanza de 10 a 14 m de alto, destacándose además algunos emergentes de hasta 20
m de alto.
En el estrato inferior predominan las especies siempre verdes como Drypetes alba, Cordia
gerascanthus, Oxandra lanceolada, entre otras. En el estrato arbóreo superior predominan especies
deciduas como Cedrela odorata, Cedrela cubensis y Bursera simaruba. El estrato arbustivo es
escaso y casi nunca está presente el herbáceo.
El bosque de mangles se localiza en las zonas cenagosas de la costa Norte de la península. Se
forman diferentes variantes como manglar de franja de Rizophora mangle, manglar achaparrado de
esta especie o de Conocarpus erecta y manglar mixto donde predominan Avicennia nitida y
Laguncularia racemosa.
Actividades humanas
Los asentamientos humanos en áreas del Parque resultan escasos y dispersos. El principal núcleo
poblacional en el interior del área protegida es la comunidad de La Bajada con 28 viviendas, que se
ubica en la franja litoral del Parque. Las principales actividades económicas de los pobladores locales
del Parque son: la forestal y la apícola. También realizan actividad de cría intensiva de puercos en las
áreas más internas del Parque. Algunos pobladores están vinculados al trabajo dentro del sector
turístico.

41
En la actualidad no existe población asociada a las áreas terrestres incluidas en las Zonas de
Conservación del Parque Nacional Guanahacabibes. Las Zonas de Conservación de El Veral y Cabo
Corrientes han permanecido sin interacción humana por más de 40 años, al ostentar la categoría de
Reservas Naturales. La principal amenaza para las poblaciones de jutía conga del área es la presión
permanente por parte de la población sobre los ecosistemas, generando actividades de pesca, caza
furtiva y colecta de especies de la flora y la fauna, principalmente en la zona de amortiguamiento y
de uso público del Parque.
II.1.4.- Área Protegida de Recursos Manejados Sierra del Chorrillo (Najasa).
Localización
La Unidad “La Belén” se encuentra situada al centro del municipio de Najasa, al sudeste de la
provincia de Camagüey; rodeada por entidades estatales. Por los valores naturales y las actividades
socioeconómicas que se desarrollan, fue declarada Área Protegida de Recursos Manejados
(APRM.), la zona comprendida de la Sierra del Chorrillo en el territorio La Belén.
Clima
Las temperaturas corresponden al régimen térmico muy cálido con medias anuales entre 24 - 26 ºC;
las temperaturas diarias sobrepasan los 30 ºC como es característico de un clima tropical.
La precipitación media anual es de 1200 - 1400 mm, concentrada de un 80 - 82 % en el período
lluvioso de mayo - octubre, con énfasis especial en mayo y junio.
El viento predominante es del norte – este, con valores de 3,6 - 4,4 m/s, aunque también se aprecian
significativamente los del sur - este.
Vegetación
La Sierra del Chorrillo, perteneciente a la Sierra de Najasa, se ubica fitogeográficamente en la
subprovincia Cuba Central. Presenta seis formaciones vegetales siendo de nuestro interés el bosque
semideciduo mesófilo secundario. Esta formación vegetal se halla tanto en las laderas menos
abruptas con algo de suelo y en algunas cimas. Posee tres estratos: uno arbóreo con emergentes, uno
arbustivo, que en algunos lugares se hace denso y otro herbáceo, limitado al sotobosque. Dentro del
estrato dominante se observan, entre los emergentes: Calophyllum antillanum, Ceiba pentandra,
Hibiscus elatus y Cordia gerascanthus, que alcanzan hasta 30.0 m de altura. En el dosel abundan

42
además: Albizia saman, Bursera simaruba, Cedrela odorata, Swietenia mahagonii y Licaria
trianda.
Actividades humanas
El municipio Najasa abarcan una población aproximada de 16 324 habitantes, lo que origina una
densidad de 18.1 habitantes por km2.La actividad económica fundamental es la ganadería, actividad
esta que surge con la fundación del territorio por los primeros habitantes, aprovechando las
características del relieve, las que son muy favorables para la cría y fomento de esta actividad
productiva. La amenaza principal para las poblaciones de jutía conga son las actividades de caza y
tala furtivas, los incendios y la introducción de especies exóticas de la flora y la fauna.
II.1.5.- Área Protegida de Recursos Manejados Sur de la Isla de la Juventud (Isla).
Localización
El área esta en el sur de la Isla de la Juventud, limitada al norte por la Cienaga de Lanier que
atraviesa la isla centralmente con rumbo oeste- este- nordeste desde la ensenada de Siguanea hasta la
boca oriental de Sanjuán convirtiéndose en una barrera geográfica natural que divide al norte de la
Isla de la llanura carsica del sur.
Clima
El clima se caracteriza por fluctuar su temperatura entre los 22.4 y 28.7 grados Celsius, las
precipitaciones anuales entre 1056 y 1400 mm y el viento predominante de componente Este, con
velocidad promedio de 9.4 Km/h
Posee rasgos particulares en su clima expresado en la poca influencia que ejercen sobre ella los
frentes fríos en la temporada invernal y la marcada influencia de los vientos del sur en el periodo de
la primavera. La humedad relativa general se comporta en el orden del 80%
Vegetación
Se reconocen nueve formaciones vegetales en el área, pero la jutía conga de nuestro estudio habita
en el bosque semideciduo. Es característica de estos bosques la presencia de dos estratos arbóreos, el
superior formado por árboles caducifolios y el inferior, por siempre verdes esclerofilos. Hay
abundancia de arbustos u lianas. Las epifitas son muy escasas. Se encuentran a veces elementos

43
xerofíticos, espinosos y suculentas. Es la formación vegetal más representativa, se desarrolla sobre
suelos esqueléticos de piedra hueca, pobres, formados por rendzinas tropicales pardas y negras.
Actividades humanas
En el año 1946 sus boques sufrieron un incendio forestal conocido como el más grande del
territorio, lo que en la actualidad repercute en sus valores económicos. Del total de hectáreas de
bosque firmes solo 92.6 ha aproximadamente han sido objeto de construcciones sociales,
encaminadas a fines científicos, económicos y de uso publico.
Esta región históricamente ha tenido una baja población. En la actualidad existe un poblado,
Cocodrilo, formado por trescientos habitantes y que debe su origen a la migración de personas de la
Isla de Gran Caimán a principios del siglo XX. La actividad de los pobladores del lugar siempre ha
estado dirigida históricamente a la actividad forestal y a la pesca.
II.1.6.- Área Protegida de Recursos Manejados Cienaga de Zapata (Zapata).
Localización
La Ciénaga de Zapata con una extensión de 4 520 km2 (CNNG, 2000), constituye una unidad
ecológica con valores naturales muy importantes, de significativa diversidad biológica, por lo que
dentro de ella han sido declaradas o propuestas cuatro áreas protegidas de diferentes categorías,
entre ellas el Parque Nacional, llamado con su mismo nombre y que abarca 136 890 ha, con un área
terrestre de108 200 ha y un área marina de 28 690 ha. Este Parque Nacional, ocupa toda la porción
occidental de la Ciénaga de Zapata.
Clima
La temperatura media en enero es de 21 °C, y en julio de 27 °C. La temperatura media anual es de
23 °C para la zona central de la Península de Zapata y 25 °C para el resto del territorio. El viento
predominante es del este, generalmente débil, y eventualmente asociado a anticiclones migratorios
al norte de Cuba.Las precipitaciones tienen valores que oscilan entre 1200 y 1 300 mm en el período
lluvioso y 250 y 300 mm en el período seco La humedad relativa en la región presenta valores altos
debidos a la disponibilidad del elemento agua en gran parte de su extensión. El valor promedio es de
un 85 %, que tiende a aumentar hacia el sur. Entre los eventos hidrometeorológicos extremos, los
más importantes son los huracanes y las tormentas locales severas.

44
Vegetación
La Península de Zapata se considera un distrito fitogeográfico por la individualidad de su flora y
vegetación: su mayor relación florística está dada con la Península de Guanahacabibes y el sur de la
Isla de la Juventud.
De las 17 formaciones vegetales presentes en la Ciénaga de Zapata se reportan 14 para el Parque
Nacional. La formacions donde se estudiaron las jutias fué el bosque de ciénaga, próximo al maglar
de Conocarpus erecta (yana). Dicho bosque se caracteriza por la presencia de un estrato arbóreo de
5-15m (puede llegar hasta 20m) de altura con la mayor parte de los árboles perennifolios. Presenta
estrato arbustivo, algunas hierbas, lianas y generalmente abundantes epífitas y algunas palmas. Estos
tipos de bosques se desarrollan en suelos mayormente turbosos, que pueden permanecer
temporalmente inundados con agua dulce entre cuatro y seis meses del año. Esta formación vegetal
es de las más ricas en especies del área. Las especies características son: Bucida palustris (júcaro de
ciénaga), Bucida buseras (Júcaro negro), Tabebuia angustata (Roble blanco), Taliparites elatus
(Majagua), Calophyllum antillanum (Ocuje), Salíx caroliniana (Clavellina), Sabal marítima (Guano
de cana), Roystonea regia (Palma real), Annona glabra Bagá), Chrysobalanus icaco (Icaco), Myrica
cerifera (Arraigan), Erythroxylum confusum (Arabo carbonero).
Actividades humanas
Aunque en el área del Parque no existen asentamientos humanos, hay varios asentamientos
colindantes que su área de influencia es el Parque mismo (Santo Tomás, Vínculo, Pálpite y Playa
Larga). Entre las prácticas negativas las más reiteradas son la pesca; la caza y la tala furtivas. Sin
embargo, en este asunto se distinguen enfoques, los que señalan que estas prácticas en pequeñas
proporciones no afectan el medio ambiente, pero cuando estas actividades se utilizan con fines
comerciales, por su intensidad pueden perjudicar la naturaleza y aún más, los que reconocen el
efecto nocivo, las justifican y legitiman como estrategias de sobrevivencia.
II.1.7.- Reserva Ecológica Hatibonico (Hatibonico).
Localización
La Reserva Ecológica Hatibonico, ubicada en la porción más occidental del valle de Guantánamo,
situada al E de Santa María Loreto-Sierra Larga, Sierra Maestra Oriental (Acevedo, 1989), es el

45
límite occidental del distrito fitogeográfico Guantanamense. Constituye la región más seca del país y
el extremo Oriental de la Sierra Maestra principal cadena de montañas de Cuba. Está comprendida
dentro de la región físico geográfica de las montañas de la Gran Piedra. Posee una superficie total de
6274 ha de las cuales 5390 ha son terrestres y las restantes 884 ha son marinas.
Clima
Las variables meteorológicas se comportan de la siguiente forma:
Temperatura media anual: 25 - 27 ºC
Temperatura mínima media anual: 21 - 23 ºC
Temperatura máxima media anual: 30 - 31 ºC
Precipitación anual: 400 - 600mm
Meses de mayor precipitación: Septiembre - Noviembre
Humedad relativa media anual: 76%
Humedad máxima media anual: 84-86%
Humedad mínima media anual: 65-75%
Vientos del norte (influyen por las noches): 5 - 10 Km
Vegetación
La flora del territorio responde a las características litológicas y climáticas del área, propiciando un
xerofísmo casi generalizado. Se cuenta con un inventario florístico con 372 especies.
En el área se pueden observa cinco formaciones vegetales. Las jutías en nuestro estudio se
encontraban en el Matorral Xeromorfo Costero y Subcostero (manigua costera). Este ocupa toda la
franja estrecha del litoral y es muy bien definida, constituye una vegetación de bajo porte con
presencia de arbustos y escasos emergentes, suculentas espinosas. Mezclándose muchas veces con
vegetación arbustiva semidesértica en las que se acentúa más la presencia de cactus y agaves.
Algunas de las especies de este ecosistema fueron: Cammiphora gluca (almaciguillo), Capparis
flexuosa (palo barba de indio), Capparis grisebachii (olivo), Dendrocereus nudiflorus (Aguacate
Cimarrón), Coccotrinax argantea (guano blanco), Opuntia militaris (patana), Opuntia stricta (Tuna
brava), Agave sp (maguey), Consolea sp (tuna de la cruz), Mamilaria prolifera.

46
Actividades humanas
Se aprecia una alteración del medio natural en un rango de 15 % - 20 % sobre todo en las áreas
llanas y onduladas, donde la riqueza desde el punto de vista económico de las especies arbóreas es
mayor y generalmente de fácil acceso para su explotación.
El uso del suelo está dirigido mayoritariamente a las actividades forestales con fines de
conservación y protección de la flora, fauna, suelo, el aire, las cuencas entre otros factores. El área
que ocupa esta Reserva ha estado históricamente sometida a presiones de uso agropecuarios por
parte de los pobladores de la región y organismos estatales, fundamentalmente dirigidos a la
realización de cultivos de subsistencia, caza y tala para la obtención del carbón vegetal y quemas de
pastos para la ganadería extensiva.
A pesar de las regulaciones existentes persisten en el área protegida a pequeña escala problemáticas
como la caza furtiva, talas ilegales, libre pastoreo dentro y en la zona de amortiguamiento con
fuertes afectaciones en los ecosistemas influyendo en la actividad económica del área.
II.1.8.- Paisaje Natural Protegido Cayo Saetía
Localización
Cayo Saetía está ubicado en la costa norte de Cuba, a la entrada de la bahía de Nipe provincia de
Holguín. Debe su condición al canal de Dumois construido para facilitar el tráncito de personas. En
la época de nuestro estudio era considerado un Paisaje Natural Protegido, pero actualmente ya no lo
es.
Clima
La temperatura fluctúa entre un máximo absoluto de 36 y un mínimo de 11 grados centígrados,
estando la media anual alrededor de los 26 grados centígrados. Las precipitaciones oscilan entre
1300 y 1400 mm anuales
Vegetación
En Cayo Saetía existen cuatro formaciones vegetales que se suceden en franja unas a otra de la
costa norte a la sur. En la costa sur predominan los manglares y a continuación el bosque
semideciduo micrófilo o monte seco. Le sigue el bosque semideciduo típico, donde se relizaron las

47
capturas de jutía conga para la presente investigación, y finalmente vegetación cultural (pastos,
cultivos, vegetación secundaria, etc). Las características del bosque semideciduo típico son
similares a las descritas para las otras localidades.
Actividades Humanas
Las actividades humanas fundamentales del Cayo hasta la fecha han sido la de forestal y la turística
cinegética. Dentro del Cayo solo existe una pequeña población humana y los suelos se emplean en
pastos para el ganado ovino y bovino.
II.2. Muestra utilizada
Estas poblaciones pueden considerarse como metapoblaciones (Hanski y Gilpin, 1991; Hanski y
Gorgiotti, 2004), ya que los muestreos se realizaron en localidades relativamente alejadas dentro de
las áreas geográficas, por lo que podrían estar siendo muestreadas una o varias de las
subpoblaciones que componen la metapoblación. Para evitar los efectos de muestrear
subpoblaciones que pudieran estar diferenciadas por el habitat, los muestreos dentro de estas áreas
se hicieron siempre en la misma formación vegetal, excepto para el caso de Guanahacabibes, donde
sí se distinguieron dos subpoblaciones ubicadas en diferentes habitats.
Atendiendo a la experiencia en el estudio de la especie, se distinguieron cuatro hábitos en cada una
de las poblaciones estudiadas:
Semiarboricola. Animales siempre capturados en árboles, en formaciones de mangle rojo (Jardines)
o bosque semideciduo (Najasa). Poco móviles.
Semiterrestres. Animales capturados en formaciones vegetales no boscosas (Sabana) o boscosas
pero muy móviles (Zapata, Guanahacabibes y Saetía).
Terrestres. Animales capturados en zona xerofítica sin árboles (Hatibonico) o en bosque pero
ocupando solo cuevas, sin subir a los árboles (Isla).
La muestra total ascendió a 768 ejemplares, 417 hembras y 351 machos, capturados en los años
2007 y 2008 durante algunos o todos los trimestres del año. Todos los ejemplares capturados fueron
adultos utilizando el criterio de Smith y Berovides (1984), que consiste en la inspección de los
genitales para determinar la evaginación del pene en los machos y la presencia de vagina abierta en
las hembras. Las muestra óptimas por población fueron calculadas según Morrison et al. (2000), y

48
resultó entre 10 a 40 individuos por población teniendo en cuenta la variabilidad de los caracteres
(entre 10 a 20 %) y un 80 % de certeza, para detectar un 5 % de diferencias entre medias, a un nivel
de confianza del 0.05.
II.2. Variables e índices medidos
Se utilizaron variables morfológicas e índices derivados de ellas, que se relacionan con funciones
adaptativas en la especie. Dichas funciones se describen en detalle en capítulos aparte. Las
consideraciones teóricas para el estudio de variables morfológicas se encuentran en Wainwright y
Reilly (1994) de forma general, y en Santos y Hortelano (1997) para mamíferos.
Se trabajaron tres tipos de variables morfológicas para comparar entre poblaciones:
Variables morfométricas. Medidas lineales del cuerpo y los apéndices, y peso o masa corporal
(Lundrigan, 1996; Monroy et al., 2005; Silva et al., 2007). Las medidas lineales fueron: el largo
Cabeza tronco (LCT) y largo de la cola (Co), medidas con una cinta métrica; el largo de la pata
izquierda con uña (Pa), medida con un pie de rey. Todas las medidas con precisión de 0.01 mm. La
variable peso corporal se determinó con una balanza dinamómetro expresándose en gramos, con una
precisión de 0.01 gramos.
Variables morfofisiológicas. Nombradas así porque se basan en medidas lineales o pesos de órganos
con funciones fisiológicas vitales y bien conocidas (Pough, 1999; Smith-Nielsen, 1999), que
transformadas en índices, indican la eficacia funcional del órgano en cuestión. Incluimos estas
variables porque el estudio de la diversidad fisiológica es fundamental para entender cómo los
organismos se enfrentan a la variación ambiental en el espacio y el tiempo (Priersma y Drent, 2003).
Se analizaron el grosor de la corteza (GC) y médula renal (GM), medidas con un pie de rey, y la
longitud de la última porción del intestino grueso (LIG), que comprende el colon y recto, medida
con una cinta métrica. En todos los casos las medidas se expresaron en milímetros (mm), con una
precisión de 0.01 milímetros. Para el caso del hígado, se tomó su peso en gramos con una balanza
analítica.
Los índices derivados de todas las variables morfológicas antes señaladas fueron:
Variables morfométricas:
Robustez (Davis y Hall, 1951)

49
Condición física (Sievert y Keith, 1985)
Peso relativo a largo total del cuerpo (Presente estudio)
Cola relativa a LCT (Lim, 1970; Fleming, 1974; Thompson, 1990)
Pata relativa a LCT (Fleming, 1974; Thompson, 1990)
Índice de dimorfismo sexual (Frayer, 1980)
Largo cola/largo pata (Anderson et al., 1983)
Los mismos utilizados por Verde et al. (2016)
Variables morfofisiológicas:
Índice de nutrición I (Gosling, 1980)
Índice de nutrición II (Presente estudio)
Índice retención de agua I (Yabe, 1982, 1983; Diaz et al, 2006 )
Índice retención de agua II (Presente estudio)
Índice reabsorción de agua (Schauenberg, 1977; Hanson, 1985)
Todos estos índices se describen en detalle en los capítulos correspondientes.
Además, se comparó entre poblaciones:
- la variabilidad de cada carácter, medido por el coeficiente de variación y una variabilidad
promedio considerando todos los caracteres.
- las correlaciones paramétricas lineales simples de todos los caracteres, tomados dos a dos, y
las regresiones paramétricas lineales simples del peso y longitudes de pie y cola absolutas
(variables dependientes) en el largo cabeza-tronco (variable independiente).
- el dimorfismo sexual, medido como la diferencia: valor medio del macho-valor medio de la
hembra. Aquí se considerarán valores destacados, los que se encuentren muy por encima del
valor promedio absoluto de todos los valores de cada población.

50
II.3. Efectos a considerar
El interés central de esta investigación fue determinar diferencias estadísticamente significativas
entre poblaciones, teniendo en cuenta además el sexo de los animales y los cambios climáticos en
Cuba por trimestres del año (Diaz Cisnero, 1989; Gutierrez y Rivero, 1999). Por esta razón, las
variables objeto de estudio se compararon entre poblaciones en tres análisis por separado,
considerando el número de poblaciones muestreadas y los trimestres del año en que se hicieron
dichos muestreos, ya que no todas las poblaciones estaban representadas en todos los trimestres.
Para los caracteres morfométricos, un primer análisis consideró las ocho poblaciones y solo el
trimestre febrero, marzo y abril (FMA), que marca los finales de la estación de la seca o poco
lluviosa en Cuba, considerando los efectos población, sexo y su interacción, eliminando el posible
efecto climático de los trimestres.
En un segundo análisis se consideraron cinco poblaciones y los trimestres FMA y agosto, septiembre
y octubre (ASO), que constituye este último los finales de la estación de lluvia en Cuba, para así
considerar los efectos de estos trimestres tan diferentes en cuanto a condiciones climáticas, además
de los efectos población y sexo y sus posibles interacciones.
Un último análisis consideró los cuatro trimestre del año FMA, mayo junio y julio (MJJ), ASO y
noviembre diciembre y enero (NDE) solo en dos poblaciones (Najasa y Jardines).
Para las variables morfofisiológicas se hicieron análisis similares, pero solo los dos primeros con un
número menor de poblaciones.
Para evidenciar la presencia de adaptación local dentro de una misma área geográfica y trimestre
(FMA), se seleccionó el Parque Nacional Guanahacabibes, donde se compararon dos poblaciones
para las variables morfométricas y morfofisiológicas.
II.4. Análisis estadísticos
Para el análisis estadístico de los datos morfológicos seguimos la metodología empleada por
Schmidly (1971; 1973) y Martinez et al. (1991), que centran su atención en la variación entre
poblaciones o variación geográfica, como es el caso del presente trabajo. Se analizaron 23 variables
morfológicas, diez en sus valores absolutos y 13 en sus valores relativos (Índices).

51
A las variables solas o en conjunto se le realizaron los siguientes análisis, empleando el programa
SPSS versión 11.5 y el programa STATISTC versión 6.0:
- Determinación de los tamaños efectivos de muestras (N) por población.
- Prueba de normalidad Kolmogorov-Smirnov
- Prueba para la homogeneidad de varianza de Levene
- Cálculo de los estadísticos: media aritmética (X), desviación típica (S) y coeficiente de
variación para cada población, trimestre del año y sexo.
- Análisis de varianza bifactorial con los efectos población, sexo e interacción población/sexo.
- Análisis de varianza trifactorial con los efectos población, sexo, trimestre del año y sus
interacciones.
- Prueba t para las diferencias entre trimestres o sexo dentro de cada población.
- Regresión lineal simple (modelo I) de los valores promedios de las variables morfométricas
y morfofisiológicas y sus coeficientes de variación (variables dependientes), en el rango de
valores de un índice climático (variable independiente), que refleja la disponibilidad de agua
y el grado de productividad primaria en las localidades de estudio.
- Regresión lineal simple (modelo II) del largo de la cola, largo de la pata, altura de la oreja,
largo del intestino, peso corporal y el peso del hígado (variables dependientes) en el largo
cabeza tronco (variable independiente), para cada población (Sokal y Rolhf, 1995; Painter et
al., 1999; Dytham, 2003; Salvador Figueras y Gargallos, 2003, Whoilock y Schluter, 2009
y Zar, 2009). También se siguieron recomendaciones más específicas para el uso de los
intervalos de confianza ( Cherry, 1996; Steidl et al., 1997; Johnson, 1999; McPherson y De
Stefano, 2003), una probabilidad de rechazo de la hipótesis nula no siempre del 5 %
(Johnson, 1999), análisis de varianza con varianzas heterogéneas (Steel y Torrie, 1980),
perfiles de variabilidad con los coeficiente de variación (Sokal y Braumann, 1980) y la
prevalencia del criterio biológico sobre el estadístico, a la hora de interpretar los datos
(Johnson, 1999; McPherson y De Stefano, 2003).

52
CAPÍTULO III. Variaciones morfométricas en poblaciones de jutía conga
(Capromys pilorides)
III.1. Introducción
La variabilidad de caracteres dentro de las poblaciones, es el material potencial para la
diferenciación y para la especiación (Santos y Hortelano, 1997). Este se ha estudiado
constantemente y con distintas aproximaciones metodológicas: morfológicas, morfométricas,
cariológicas y moleculares, inicialmente asociado a problemas taxonómicos y actualmente con una
tendencia a problemas diversos como la biogeografía y la conservación tanto in situ como ex situ. La
conservación tiene entre sus objetivos la de preservar la biodiversidad y el potencial genético de las
poblaciones, más aun en poblaciones endémicas que se encuentran en algunas de las categorías de
amenaza o con un alto grado de explotación, algunas de las características que se ajustan a la jutía
conga, ya que es sabido que de este recurso hacen uso los campesinos y pescadores (Berovides,
1987).
En Cuba se han realizado estudios de variación morfométrica en la jutía conga, siendo el primero el
de Smith y Berovides (1984), que compara cinco poblaciones de hábitat de bosque y manglar (no
incluyó el hábitat de vegetación xerofítica). Se analizaron además los índices relativos derivados del
peso, y las muestran no se efectuaron en un solo trimestre, ignorando posibles efectos estacionales.
Otro trabajo se realizó en dos poblaciones con hábitat de bosque y manglar (Sánchez et al., 1992),
con un número reducido de individuos y la carencia también de hábitat con vegetación xerofítica.
Berovides (2005) incorpora el hábitat de vegetación xerofítica, comparando este con el de bosque;
de la Fuente y Berovides (2009) consideraron dos poblaciones en manglares de diferentes
localidades, y Linares et a.l (2010) caracteriza la morfología de las poblaciones de Guanahacabibes.
En todos estos estudios, limitados por las pocas poblaciones en estudio y la realización solo en
determinadas épocas del año, las poblaciones de bosque siempre presentaron mayores valores
promedios de las medidas morfométricas, seguidos de la vegetación xerofítica, presentándose los
menores valores en las poblaciones de mangle.
En relación a los cambios estacionales, estos pueden hacer semejantes ciertas diferencias entre
hábitats, atenuarse o desaparecer durante la estación de lluvia o en relación a la temperatura,
humedad relativa y la precipitación; esto no ocurre con la composición taxonómica y estructural de
la vegetación, que siempre muestra diferencias estacionales (Janzen, 1976). Los cambios

53
estacionales más marcados para las zonas neotropicales se dan en las especies de hábitats más
xéricos, sobre todo en relación a los cambios de la disponibilidad de agua (Bozionovic y Gallardo,
2006).
Los hechos anteriores, hasta donde conocemos, no se han considerado en los estudios sobre
morfología de la jutía conga, donde o bien se han ignorado o solo se han considerado unos pocos
meses de estudio (de la Fuente y Berovides, 2015). En Cuba existe para casi todo el territorio
nacional una estación de lluvia (Mayo a Octubre) y otra de seca (Noviembre a Abril), con cambios
marcados en precipitaciones y, sobre todo, en la estructura y fisionomía de la vegetación, que con
seguridad afectan a la morfología, supervivencia y reproducción de las poblaciones de jutía conga,
pero de lo cual no se tiene información.
El objetivo general de este capítulo fue determinar el grado de variación morfométrica en ocho
poblaciones de jutía conga en diferentes hábitats.
Los objetivos específicos son:
Comparar nueve variables morfométricas entre ocho poblaciones de jutía conga, que viven
en hábitats de bosque, manglar y vegetación xerofítica, muestreadas en el trimestre febrero,
marzo y abril (finales de la estación de seca).
Relacionar las variables morfométricas con un índice climático, asociado a la precipitación,
temperatura y la productividad primaria del hábitat.
Determinar el grado de dimorfismo sexual para los caracteres estudiados entre dichas
poblaciones.
Determinar el grado de variación estacional de la morfometría en cinco poblaciones de jutía
conga en diferentes hábitats, entre los trimestres de febrero, marzo y abril (finales de la
época de seca) y los trimestres de agosto, septiembre y octubre (finales de la época de
lluvia). Dos de estas cinco poblaciones disponían también de datos para el año completo, lo
que fue analizado por separado.
III.2. Materiales y métodos
Estudio para la estación de seca

54
Se analizaron ocho poblaciones de jutía conga con una muestra total de 389 ejemplares capturados
(203 hembras y 186 machos). Las muestras por sexo, formación vegetal y categorías de manejo para
cada localidad se pueden observan en la Tabla I.
Tabla I. Localidad de las poblaciones, categoría de manejo, tamaño de muestra por sexo y hábitat
(formación vegetal) para el estudio morfométrico de la Jutía conga (Capromys pilorides). El
nombre abreviado de cada población de jutía se da entre paréntesis.
Localidad y categoría de manejo Sexo N Hábitat
Parque Nacional Jardines de la Reina (Jardines) H 27 Manglar (Rizophora
mangle) M 20
Are protegida de recursos manejados
Sabana Camaguey (Sabana)
H 23 Manglar y Vegetación
de costa de cayo M 22
Parque Nacional Cienaga de Zapata
(Zapata)
H 18 Bosque de ciénaga y
manglar M 16
Reserva ecológica Hatibonico
(Hatibonico)
H 21 Manigua costera
M 33
Parque Nacional de Guanahacabibes (Guanacahabibes)
H 27 Bosque semideciduo
M 20
Area protegida de recursos manejados
Najasa (Najasa)
H 48 Bosque semideciduo
secundario M 42
Refugio de Fauna Saetía (Saetía)
H 16 Bosque semideciduo M 14
Area protegida de recursos manejados
Isla Sur (Isla Sur)
H 23 Bosque semideciduo
M 19
En la Tabla II se indican las precipitaciones y temperaturas medias anuales que permitieron calcular
un índice climático de los hábitats en cada localidad, denominado índice climático de Martonne
(Bozinovic et al., 2009), por medio de la fórmula:
IC=Precipitación media anual/Temperatura media anual +10 0 C
Este índice está asociado a la productividad vegetal de los hábitats y a la tasa metabólica basal
(independiente del peso) en los individuos de la población (Bozinovic et al., 2009). Los valores de
este índice fueron convertidos en rangos (Al-Kahtani et al., 2004), donde el rango 1 representó la
más baja disponibilidad de nutrientes (bajos valores del índice) y el 8 la más alta disponibilidad.
Estos rangos para cada población fueron los siguientes: Jardines 1, Sabana 2, Zapata 3, Hatibonico
4, Najasa 5, Guanahacabibes 6, Saetía 7 e Isla 8.
Tabla II. Valores de precipitación y temperatura de ocho localidades donde vive la jutía conga, para
la confección de un índice climático.

55
Localidad Precipitación (mm) Temperatura (ºC)
Jardines de la Reina 1000 27
Archipiélago Sabana Camaguey 1000 26.5
Cienaga de Zapata 950 23.5
Hatibonico 600 28 Najasa 1300 25
Guanahacabibes 1350 25.8
Saetía 1400 26 Isla sur 1400 25
En relación a la cola y pie relativos, se clasificó cada población por sus hábitos, ya que de acuerdo a
estos, las poblaciones semiarborícolas (utilizan con frecuencia los árboles) deben tener los mayores
valores promedios de cola y pata relativos, por sus respectivas funciones adaptativas para equilibrio
y trepa (Fleming, 1974; Elissamburu y Vizcaino, 2004; Rivas y Linares, 2006), en comparación con
las semiterrestres (uso poco frecuente de los árboles) y las terrestres. Esta clasificación se ofrece en
la Tabla III.
Tabla III. Agrupamiento de poblaciones de jutía conga según sus hábitos.
Grupo Población
Habitat Vegetación
Terrestre V. xerofítica Hatibonico
Terrestre Bosque Isla/Guanahacabibes
Semiterrestre V. costera Sabana/Zapata
Semiterrestre Bosque Saetía
Semiarburícola Manglar Jardines
Semiarburícola Bosque Najasa
Los ejemplares capturados y analizados fueron adultos según el criterio de Smith y Berovides
(1984), y a cada uno se les determinó el peso (PC) en gramos (g) y el largo cabeza tronco (LCT),
largo del pie izquierdo con uña (Pi) y largo de la cola (Co) en milimétro (mm), de los cuales se
derivaron seis índices que se indican en la Tabla IV.
A las variables absolutas e índices derivados de las mismas (variables dependientes), se les
determinó el tipo de distribución estadística por la prueba de normalidad de Kolmogorov –Smirnov
y para los diferentes tratamientos, la prueba de homogeneidad de varianza de Levene. Los valores
medios de las variables se compararon estadísticamente mediante un análisis de varianza (ANOVA)
bifactorial, para los efectos (variables independientes) localidad, sexo y su interacción, así como una
prueba de Tukey para determinar las diferencias de valores medios entre poblaciones específicas. Se

56
realizaron también análisis de regresión lineal simple, para determinar la dependencia de las
variables medidas con el índice de Martonne (Modelo I) y de estas mismas variables con el LCT
(Modelo II). El dimorfismo sexual se determinó por la simple diferencia de: valor promedio de los
macho-valor promedio de las hembras, para cada carácter.
Tabla IV. Índices derivados de las variables absolutas medidas en poblaciones de jutía conga, así como su
función adaptativa.
ÍNDICES MORFOMÉTRICOS FÓRMULAS FUNCIÓN ADAPTATIVA
Robustez, peso relativo al volumen del cuerpo (Ro)
PC/LCT3*100000 Supervivencia y reproducción (Stearns,1992)
Condición Física (CF), Raíz cubica del peso relativo al largo de la pata
LCT/Pi Supervivencia y reproducción
(Sievert y Keith, 1985)
Peso por milímetro lineal relativo al largo total (LT)
PC/LT Supervivencia y reproducción (presente estudio)
Cola Relativa a LCT (CoR)
LC/LCT*100 Termorregulación (Millien et al,
2006), arboricidad (Fleming,
1974)
Pie Relativo a LCT (PiR)
Pi/LCT *1000000 Termorregulación (Millien et al.,
2006), arboricidad (Fleming,
1974)
Pie/cola ( relación Pie entre cola) Pi/Co Arboricidad (Fleming,1974)
Estudio de los cambios estacionales
Para este estudio se compararon los valores medios de los datos morfométricos entre los trimestres
de febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO). El estudio se realizó con las
poblaciones de los Parques nacionales (PN) Jardines de la Reina, y Guanahacabibes, las Areas
Protegidas de Recursos Manejados (APRM) Sabana Camaguey, Sierra del Shorrillo e Isla, con una
muestra total de 650 ejemplares de jutía conga (362 hembras y 288 machos). Además, para APRM
Sierra del chorrillo y PN de Jardines de la Reina, se efectuó un análisis similar para cada uno de los
trimestres del año, al contar con datos de todo el año. Las muestras por localidad de las poblaciones,
sexo, trimestres del año y categorías de manejo, se muestran en la Tabla V.
Tabla V. Localidad de las poblaciones, categoría de manejo, tamaño de muestra por sexo, hábitat y trimestres
del año para el estudio morfométrico de la Jutía conga (Capromys pilorides). El nombre abreviado de
cada población de jutía conga se da entre paréntesis.
57
Localidad y categoría de manejo
Sexo
NDE FMA MJJ ASO
N N N N
Parque Nacional Jardines de la Reina
(Jardines)
H 14 27 21 35
M 9 20 11 32
Are protegida de recursos manejados Sabana Camaguey (Sabana)
H - 23 - 26
M - 22 - 29
Parque Nacional de Guanahacabibes
(Guanahacabibess)
H - 27 - 28
M - 20 - 20
Area protegida de recursos manejados Najasa (Najasa)
H 23 48 32 23
M 10 42 28 12
Area protegida de recursos manejados
Isla sur (Isla Sur)
H - 23 - 12
M - 19 - 14
Los ejemplares capturados y analizados fueron todos adultos según el criterio de Smith y Berovides
(1984), y a cada uno se les determinó el peso en gramos (g) y el largo cabeza tronco (LCT), largo
total (LT), largo del pie izquierdo con uña (Pi) y largo de la cola (Co) en milímetros (mm). De estas
medidas se derivaron seis índices, ya descritos anteriormente.
Para la comparación de las cinco localidades y los dos trimestres (FMA y ASO), a las variables
absolutas e índices derivados de las mismas se les determinó el tipo de distribución por la prueba de
normalidad de Kolmogorov –Smirnov y la prueba de Levene para la homogeneidad de varianza y
se compararon las medias estadísticamente, mediante un análisis de varianza trifactorial, con los
efectos localidad, estación, sexo y sus interacciones, para determinar las diferencias entre estos
efectos. Un análisis de varianza bifactorial también se llevo a cabo con los efectos localidad, sexo e
interacción, dentro de cada uno de los trimestres, para determinar los cambios estacionales
específicos de cada localidad y dentro de cada localidad, pruebas de t para detectar cambios
específicos entre trimestres dentro de cada población. Se utilizó la prueba de Tukey y la prueba de
Dunnett en dependencia de la homogeneidad de las varianza, para determinar las diferencias de
valores medios para cada una de las poblaciones en específico. Los coeficientes de variación
promedio se compararon entre las cinco poblaciones dentro de cada trimestre y entre los mismos,
por la prueba de Kruskal-Wallis. Los coeficientes de variación por variables entre los trimestres
dentro de cada población se compararon de forma indirecta, utilizando las comparaciones de las
varianzas por la prueba F. Igual procedimiento se siguió para el análisis de las dos poblaciones con
datos de los cuatro trimestres.

58
III.3. Resultados y discusión
Proporción de sexos (% de hembras) y cociente de edad (% de no adultos)
La proporción de sexos, como % de hembras, fluctuó entre las poblaciones con un valor de
alrededor del 50 %, que es el valor esperado. Solo Zapata tuvo un valor algo inferior (38.9 %), pero
en todos los casos estos valores no difieren estadísticamente entre poblaciones, ni con respecto a la
proporción esperada de 50 % de hembras.
Respecto a la edad, la proporción de individuos no maduros sexualmente en cada población se
ofrece en la Tabla VI. Considerando el valor límite de 440 mm para el largo cabeza-tronco para
ambos sexos como criterio promedio de madurez sexual (Smith y Berovides, 1984), se observa un
claro gradiente de manglar a no bosque y a bosque de individuos de menor a mayor edad. Además,
se observan los porcentajes más altos entre los primeros grupos. Las diferencias fueron
estadísticamente significativas.
Varias hipótesis podrían explicar estos resultados:
- El criterio del límite de 440 mm no es válido para todas las poblaciones.
- En las poblaciones de no bosque, al ser más abiertas, existe mayor probabilidad de detectar y
capturar a los no adultos, que generalmente están más ocultos, ocurriendo lo contrario en los
bosques.
- En las poblaciones de no bosque, al ser de menor tamaño, posiblemente muchos individuos
menores de 440 mm ya son maduros sexualmente, se comportan como tales y son más
capturados.
Tabla VI. Proporción (%) de individuos no adultos, en ocho poblaciones de jutía conga. N=Muestra
total; n= cantidad de individuos no adultos
Población N n %
Zapata 35 12 34,3
Jardines 73 20 27,4
Sabana 34 7 20,6
Hatibonico 59 8 13,6
Guanahacabibes 53 6 11,3
Najasa 108 7 6,5
Saetía 31 1 3,2
Isla 44 0 0

59
Largo cabeza-tronco y peso
En la Tabla VII se muestran los valores promedios y la desviación típica del largo cabeza tronco
(LCT) en las ocho poblaciones de jutía conga, observándose la formación de cinco grupos con
diferencias estadísticamente significativas y superpuestos, estructurados de la siguiente manera:
grupo 1, integrado por las poblaciones de hábitat de manglar (Jardines);
grupo 2, integrado por las poblaciones de Sabana, Zapata y Hatíbonico con hábitat de
manglar más vegetación de costa de cayo, ecotono bosque-ciénaga y manigua costera,
respectivamente;
grupo 3, conformado por las poblaciones de Hatíbonico, Najasa y Guanahacabibes, las dos
últimas con hábitat de bosque semideciduo;
grupo 4, que lo integra Najasa, Guanahacabibes y Saetía, este último de hábitat bosque
semideciduo;
grupo 5, conformado por las poblaciones de Saetía e Isla, esta con hábitat de bosque
semideciduo.
Esta segregación de los grupos en cuanto al LCT pudiera corresponderse con la disponibilidad de
nutrientes en los diferentes hábitats, lo cual facilitaría la máxima expresión de este carácter para las
poblaciones de bosque, con más variedad de nutrientes (de la Fuente, 2008), expresiones
intermedias para las poblaciones de hábitat de manigua costera y más extremas para las poblaciones
de hábitat de manglar.
Tabla VII. Valores promedios (X) y desviación típica (S) del largo cabeza tronco (LCT) en ocho
poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes difieren a
p<0.05 por una prueba de Tukey
Población X S
Jardines 458,70 a 33,40
Sabana 456,77 a 30,17
Zapata 465,14 ab 44,83
Hatibonico 481,50 bc 40,75
Najasa 488,84 cd 35,51
Guanahacabibes 489,51 cd 39,31
Saetía 512,16 de 37,90
Isla 525,61 e 32,68

60
La Tabla VIII refleja los pesos promedios y la desviación típica de las ocho poblaciones de jutía
conga estudiadas, donde se observan diferencias estadísticamente significativas con la formación de
seis grupos superpuestos. Los grupos conformados quedaron integrados de la siguiente forma:
grupo1, estructurado por la población de Jardines y Sabana;
grupo 2, (Sabana y Zapata);
grupo3, (Zapata y Hatibonico);
grupo 4, (Hatibonico, Najasa y Guanahacabibes);
grupo 5, (Najasa, Guanahacabibes y Saetía);
grupo 6, (Saetía e Isla).
El peso corporal (el cual muestra un patrón semejante al LCT, lo que era de esperar dada la alta
correlación entre ambas variables) posee mayor nivel de discriminación, lo cual indica que pudiera
estar afectado no solamente por la disponibilidad de nutrientes en los diferentes hábitats (Bozinovic
et al., 2011) y la influencia del LCT, sino también por el tipo de actividad preferencial que desarrolla
la especie en su ecosistema (semiarborícola, semiterrestre y terrestre) facilitando así su adaptación
(Elisamburo y Vizcaíno, 2004: Riva y Linares, 2006 ).
Tabla VIII. Valores promedios (X) y desviación típica (S) del peso corporal (PC) en ocho
poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes difieren a
p<0.05 por una prueba de Tukey
Población X S
Jardines 2405,31 a 532,56
Sabana 2811,44 ab 576,84
Zapata 3142,35 bc 931,79
Hatibonico 3883,33 cd 845,35
Najasa 3509,11 de 816,19
Guanahacabibes 3853,31 de 1007,49
Saetía 4141,16 ef 918,78
Isla 4592,90 f 912,76
Los índices derivados del peso y el largo cabeza tronco, que expresan de forma relativa condición
física (CF), robustez (Ro) y peso por milímetro lineal (PC/LT), se exponen en la Tabla IX. Todos los
índices presentaron diferencias estadísticamente significativas entre poblaciones, reflejando los

61
resultados siguientes. Para la CF, los menores valores para este índice se muestran en las
poblaciones de hábitat manglar, manglar más vegetación de costa de cayo y bosque de ciénaga y
manglar (Jardines, Sabana y Zapata), fundamentalmente, incluyendo además a la población de
Najasa, que es una población de hábitat de bosque. Ocupan posiciones intermedias todas las
poblaciones de hábitat de bosque semideciduo, además de poblaciones como Sabana (hábitat
manglar) y Zapata (bosque de ciénaga y manglar), y ocupan posiciones extremas algunas
poblaciones de hábitat de bosque semideciduo (Isla y Guanahacabibes), con el valor máximo para la
población de Hatibonico, que es una población de hábitat de manigua costera.
Este patrón observado para la CF presenta ciertas incongruencias ecológicas, lo cual se debe al
factor de corrección utilizado (largo del pie), que varía indistintamente en las poblaciones de
acuerdo al habito, y hace que poblaciones de hábitat de bosque semideciduo se encuentren entre las
poblaciones de menor índice, y poblaciones de hábitat manglar ocupen también, posiciones
intermedias, así como un bajo nivel diferencial entre poblaciones.
El índice Ro muestra un mayor grado de discriminación entre poblaciones de diferentes hábitats, con
el menor valor del índice para la población de Jardines, de hábitat de manglar exclusivamente, y
posiciones intermedias sin diferenciar estadísticamente en las poblaciones de hábitat de manglar más
bosque-ciénaga (Sabana y Zapata) en conjunto con poblaciones de hábitat de bosque semideciduo
(Saetía, Isla y Najasa). El valor mayor para este índice se observa en la población de Hatíbonico,
con diferencias estadísticamente significativas con respecto al resto de las poblaciones. Estos
resultados, a pesar de tener un mayor poder diferencial entre las poblaciones con respecto a la CF,
tienen como desventaja que en la confección del índice no se incluye el largo de la cola de los
ejemplares analizados; este rasgo, se sabe que muestra diferencias entre poblaciones de forma
absoluta y relativa.
El peso por milímetro lineal expresó diferencias estadísticamente significativas, dando el mayor
nivel de diferenciación entre las poblaciones, y diferenciando las poblaciones de hábitat de manglar
y manigua costera de las poblaciones de hábitat de bosque semideciduo. Los valores más bajos de
este índice en orden creciente corresponden a las poblaciones de Jardines, Sabana y Zapata, que son
poblaciones de hábitat manglar, manglar más vegetación de costa de cayo y bosque-ciénaga.
Muestra posiciones intermedias para Najasa y Guanahacabibes, y los valores mayores en orden
creciente corresponden a Hatibonico, Saetía e Isla. Todas, a excepción de Hatíbonico, son

62
poblaciones de hábitat de bosque semideciduo, mostrando una concordancia ecológica con la
diversidad y disponibilidad de nutrientes que muestran los bosques con respecto a los manglares.
Tabla IX. Valores promedios (X) y desviación típica (S) de los índices de robustez, condición física y
el peso por milímetro lineal, en ocho poblaciones de jutía conga (Capromys pilorides).
Medias con índices diferentes difieren a p<0.05 por una prueba de Tukey.
Población
Robustez Condición Física Peso por milímetro lineal
X S X S X S
Jardines 2,47 a 0,34 14,79 a 1,035 4,65 a 0,81
Sabana 2,94 b 0,48 15,73 ab 1,468 5,52 b 0,95 Zapata 3,06 bc 0,48 15,61 ab 1,317 6,00 bc 1,34
Hatibonico 3,44 d 0,44 17,84 c 1,126 7,17 ef 1,21
Najasa 2,96 bc 0,35 15,82 ab 1.210 6,40 cd 1,19 Guanahacabibes 3,24 cd 0,48 16,56 bc 1,136 7,09 de 1,43
Saetía 3,03 bc 0,191 16,28 b 1,084 7,25 ef 1,21
Isla 3,13 bc 0,339 16,59 bc 1,273 7,96 f 1,24
Pie y cola
Los valores de la cola absoluta y relativa se reflejan en la Tabla X. Se observaron diferencias
estadísticamente significativas entre poblaciones. La variable cola absoluta muestra un patrón que
no es el esperable con el hábito descrito para la especie por otros autores. Esto puede deberse al
efecto del tamaño corporal en el largo de este apéndice para esta especie, lo cual introduce un error a
la hora de establecer un patrón diferencial entre poblaciones con el fin de evidenciar la adaptación
ecológica a la vida terrestre, semiterrestre o semiarborícola.
La variable cola relativa reveló un patrón diferencial entre poblaciones de hábitos terrestres y
semiterrestres (con los menores valores) y hábito semiarborícola (con los valores mayores). Este
resultado es el esperado de acuerdo a la función adaptativa de este apéndice, es decir, un órgano de
equilibrio para moverse en los árboles.
Las poblaciones de Guanahacabibes e Isla mostraron los valores más bajos del índice cola relativa,
siendo al parecer poblaciones terrestres o semiterrestres que, además, presentan los refugios entre las
piedras y cuevas de sus localidades (Linares, 2005). Las poblaciones de Sabana y Zapata, con los
valores intermedios, poseen hábitos semiterrestres (Smith y Berovides, 1984), y las poblaciones de
Najasa, Jardines y Saetía, de hábito semiarborícola, poseen los mayores valores.
La población de Hatibonico presentó un alto valor de la cola relativa. Sin embargo, se sabe que de
acuerdo al hábitat en que se desarrolla, el poseer una larga cola relativa no representa una adaptación

63
a la vida arborícola, por la no existencia de árboles dentro de la localidad en que habita. No obstante,
cabe señalar que es precisamente Hatibonico la región de más alta temperatura, lo cual sugiere que
pudiera estar relacionado con posibles funciones termoreguladoras según el principio de Allee, que
establece que en las zonas de altas temperaturas los apéndices en los mamíferos tienden a ser más
largos. Otras posibles explicaciones serían que se trate de una población de bosque con un reciente
arribo a esta localidad, o que la cola sea utilizada como reservorio de grasa para enfrentar periodos
de escasez alimentaria, lo que debe ser frecuente en este hábitat.
Tabla X. Valores promedios (X) y desviación típica (S) del largo de la cola absoluta y relativa en
ocho poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes difieren
a p<0.05 por una prueba de Tukey.
Población Cola absoluta Cola relativa
X S X S
Jardines 248,12 b 17,93 54,29 d 4,79
Sabana 224,66 a 15,72 49,30 ab 3,60 Zapata 235,91 a 17,82 50,92 bc 3,43
Hatibonico 264,64 c 22,90 55,11 d 4,25
Najasa 264,13 c 20,96 54,23 d 4,85
Guanahacabibes 233,14 a 23,14 47,77 a 4,66 Saetía 272,60 c 17,56 53,33 cd 2,81
Isla 250,78 b 16,67 47,77 a 2,78
En la Tabla XI se muestran los valores del pie y pie relativo, con diferencias estadísticamente
significativas entre poblaciones. Los valores absolutos del pie muestran un patrón semejante a los
valores de la cola absoluta ya analizados anteriormente, debido a que en ambos casos estos
apéndices están influenciados por el tamaño corporal (al presentar asociación significativamente
estadística con esta variable). El pie relativo agrupó las poblaciones en tres grupos con un alto nivel
de solapamiento, según se refleja a continuación:
grupo 1, integrado por la población de Hatíbonico, Isla, Saetía y Guanahacabibes;
grupo 2, integrado por todas las poblaciones menos Hatíbonico y Zapata;
grupo 3, conformado por todas las poblaciones menos Hatíbonico e Isla.
Esto significa que solo Hatíbonico y la Isla conforman las poblaciones de más bajo índice versus
Zapata, que es la población de más alto índice.

64
Tabla XI. Valores promedios (X) y desviación típica (S) del largo del pie absoluto y relativo en ocho
poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes difieren a
p<0.05 por una prueba de Tukey.
Población Pie absoluto Pie relativo
X S X S
Jardines 90,09 ab 3,125 19,72 bc 1,41
Sabana 90,06 ab 5,772 19,76 bc 1,36 Zapata 92,95 bc 5,244 20,11 c 1,71
Hatibonico 87,60 a 4,958 18,27 a 1,25
Najasa 96,44 cde 8,670 19,80 bc 2,01 Guanahacabibes 94,13 cd 6,818 19,22 abc 1,44
Saetía 98,11 de 4,860 19,21 abc 1,21
Isla 99,99 e 6,449 19,06 ab 1,28
Los valores del índice pie/cola para ocho poblaciones de jutía conga se muestran en la Tabla XII. Se
observaron diferencias estadísticamente significativas entre las poblaciones, agrupándose de la
forma siguiente:
grupo 1, con valores muy bajos del índice (mas cola que pata) y constituido por las
poblaciones de Hatibonico, de hábito terrestre;
grupo 2, representado por las poblaciones de Jardines, Saetía y Najasa con valores medios
del índice, de hábito semiarborícola;
grupo 3, formado por las poblaciones de Sabana, Zapata, Isla y Guanahacabibes con valores
altos (alrededor del 40 %) en el índice, de hábitos terrestres y semiterrestres.
El grupo 1, compuesto por la población de Hatibonico, presenta una peculiaridad que contradice el
hábito obtenido con los valores del índice pie/cola. Esta característica se observó también en el
índice cola relativa, y posiblemente se debe a cambios adaptativos de la población respecto a varios
efectos ya discutidos. El patrón observado en el índice pie/cola se corresponde con el patrón del
índice cola relativa, pero en sentido contrario, debido a que en este primer índice el largo de la cola
se utiliza en el denominador para relativizar el valor de la variable pie.
Tabla XII. Valores promedios (X) y desviación típica (S) del índice pata/cola en ocho poblaciones
de jutía conga (Capromys pilorides). Medias con índices diferentes difieren a p<0.05 por una
prueba de Tukey.
Población Pie/cola
65
X S
Jardines 36.45 b 0.42
Sabana 39.94 c 0.42 Zapata 39.51 c 0.49
Hatibonico 33.34 a 0.39
Najasa 36.95 b 0.30 Guanahacabibes 40.62 c 0.42
Saetía 36.08 b 0.52
Isla 39.98 c 0.45
Índice climático
Los valores de regresión para cada una de las variables estudiadas respecto al índice climático de
Martonne se muestran en la Tabla XIII. Se observan resultados estadísticamente significativos para
las variables LCT, PC, PC//LT y Pi, lo cual significa que estas variables están asociadas
estadísticamente con características de la localidad como la temperatura, precipitación y producción
primaria, con un mayor grado de dependencia con respecto al peso corporal, variable que se
encuentra más condicionada de forma directa por estos aspectos.
Los apéndices cola absoluta y relativa y pie relativo, por sus características adaptativas, se
encuentran más relacionados con los hábitos de la población dentro de la localidad que con la propia
localidad y las características del hábitat.
Otros autores han encontrado efectos similares sobre el tamaño corporal en poblaciones de roedores.
Pergamon y Ashley (2001) lo mostraron para tres especies de roedores en islas, y Martin y Moestrup
(2003) lo mostraron en relación a la disminución del tamaño en poblaciones de grandes y medianos
roedores, por la fragmentación del hábitat.
Tabla XIII. Regresiones de nueve variables morfométricas en un índice climático para ocho
poblaciones de jutía conga (Capromys pilorides). * p<0.05; **p<0.01; ***p<0.001
Variables F R2 b Sb
LCT 89.3*** 93.7 9.81 1.03
PC 37.4** 86.2 274.19 44.80
Ro 1.75 ns 22.6 0.05 0.04
CF 1.86 ns 23.7 0.18 0.13
PC/LT 8.01* 57.2 0.27 0.09
Co 2.17 ns 14.4 3.72 2.52
CoR 0.2 ns 3.3 -0.24 0.53
Pi 15.06* 71.5 1.42 0.37
PiR 0.9 ns 12.8 -0.09 0.09

66
Hábitos en relación a pie y cola
La Tabla XIV muestra la relación entre los hábitos terrestres, semiterrestres y semiarborícolas, y los
valores medios de pie y cola relativos. Se observa claramente cómo las medias de cola relativa se
incrementan según se va de terrestre a semiarborícola, de acuerdo con lo esperado para la función
adaptativa de este índice, pero el pie relativo prácticamente no cambia, debiendo esperarse una
disminución de su valor medio de terrestre a semiarborícola.
Para la cola relativa, solo Hatibonico se sale de la tendencia esperada (posee una cola relativamente
larga a pesar de ser terrestre), lo que podría deberse a que esta población vive en un ambiente
xerofítico y la cola serviría como órgano de reserva de grasa, como ocurre en otras especies de
roedores, o que obedece a la regla de Allee, como ya se discutió. La no concordancia del pie con los
hábitos podría ser debida a otros factores, no relacionados ni con el clima ni con los hábitos.
Tabla XIV. Relación de los hábitos con los valores promedios de pie y cola relativos, en ocho
poblaciones de jutía conga.
Hábitos Población Cola relativa Pie relativo
Terrestre Hatibonico 55.1 18.3
Isla 47.8 19.1
Guanahacabibes 47.8 19.2
Semiterrestre
Sabana 49.3 19.8
Zapata 50.9 20.1
Setía 53.3 19.2
Semiarborícola
Jardines 54.3 19.7
Najasa 54.2 19.8
En la tabla se pude observar que los mismos hábitos pueden darse en diversas formaciones
vegetales, por ejemplo, semiarborícola en Jardines (manglar) y en Najasa (bosque), lo que explica su
no relación con el índice climático. Esto sugiere que no son las formaciones vegetales las que de
forma directa y actual influyen en dichos hábitos. Por ejemplo, una población de hábito terrestre que
ha migrado recientemente a una formación vegetal con árboles, podría mantener en este su hábito
terrestre por algún tiempo.
Para la cola relativa, estos resultados coinciden con los de Elissamburu y Vizcaino (2004), que
demuestran el valor adaptativo de nueve índices funcionales relativos a las proporciones de las
extremidades en 10 especies de roedores caviomorfos. También con los de Rivas y Linares (2006),

67
que estudiando roedores sigmodontidos, pudieron distinguir tres tipos de forma de pata según sus
adaptaciones: terrestre de vegetación boscosa, terrestre de vegetación abierta y trepadores-arbóreos.
Regresión en el LCT
Los valores de regresión para tres variables absolutas PC, Co y Pa (variables dependientes) con
respecto al LCT (como variable independiente) se exponen en la Tabla XV.
El peso corporal mostró valores altamente significativos para cada uno de las poblaciones, con una
tendencia de presentar los valores menores en las poblaciones de hábitat de cayos (Jardines y
Sabana) con respecto a las poblaciones de hábitat de bosque y manigua costera.
Esto sugiere que por cada unidad de aumento de la variable independiente (LCT) existe menor
aumento de la variable dependiente (PC) en las poblaciones de hábitat de cayo, con respecto a las de
otros hábitats.
Las variables cola y pie fueron significativas estadísticamente en cada uno de las poblaciones, con
valores de incrementos más discretos para el pie con respecto a la cola, lo cual obedece al grado de
alometría diferencial que presentan estos apéndices.
Tabla XV. Regresiones de tres variables morfométricas en el largo cabeza tronco (LCT) para ocho
poblaciones de jutía conga (Capromys pilorides). * p<0.05; **p<0.01; ***p<0.001
Población PC Co Pi
Jardines 12.83*** 0.155* 0.027*
Sabana 13.54*** 0.221** 0.092***
Zapata 18.72*** 0.281*** 0.059**
Hatibonico 17.73*** 0.343*** 0.080***
Najasa 20.09*** 0.181** 0.061*
Guanahacabibes 20.63*** 0.267*** 0.081***
Saetía 23.05*** 0.322*** 0.079***
Isla 23.10*** 0.219** 0.082**
Se observa una clara tendencia del PC a presentar un aumento de su dependencia con el LCT, a
medida que se pasa de habitats de no bosque a bosque. Para Co y Pi, solo Jardines difiere
notablemente del resto de las poblaciones, con valores menores de regresión.
Variabilidad de los caracteres
La Tabla XVI muestra el coeficiente de variación (CV) para algunas de las variables estudiadas, así
como el coeficiente de variación promedio para todas las variables por población.

68
El CV promedio fue muy similar entre todas las poblaciones (entre 9.6 y 12.7). Las variables PC, Ro
y PC/LT mostraron los mayores valores del CV en todas las poblaciones.
Tabla XVI. Coeficiente de variación (CV) para las variables largo cabeza tronco (LCT), peso
corporal (PC), robustez (Ro), condición física (CF), peso relativo al largo total (PC/LT), cola
relativa (Co), pie relativo (PiR) e índice pie/cola (Pi/Co) en ocho poblaciones de jutía conga
(Capromys pilorides).
Población LCT PC Ro CF PC/LT CoR PiR Pi/Co CV
promedio
Jardines 7.28 22.14 13.52 6.96 17.41 8.82 7.15 6.44 10.86
Sabana 6.60 20.51 16.32 9.28 17.21 7.28 6.88 7.08 10.74
Zapata 9.63 29.65 15.55 8.39 22.16 6.73 8.45 5.6 12.77 Hatibonico 8.46 21.77 12.79 6.27 16.87 7.71 6.78 7.4 1058
Najasa 7.26 23.26 11.82 7.64 18.43 8.92 10.10 8.3 11.38
Guanahacabibes 8.03 26.14 14.51 6.82 20.02 9.75 7.49 8.8 12.69 Saetía 7.40 22.18 6.27 6.63 16.62 5.26 6.29 6.24 9.61
Isla 6.22 20.02 10.54 7.65 15.45 5.81 6.73 7.5 10.00
Dimorfismo sexual
El dimorfismo sexual para las ocho variables estudiadas se muestran en la tabla XVII, donde se
refleja para el LCT un dimorfismo sexual en cada una de las poblaciones, con una tendencia a
incrementar los valores destacados por encima de la media, en la medida que en la población
aumenta el tamaño corporal (regla de Rensh, Dale et. al, 2007) como primer aspecto, e influenciados
por el habito (terrestre, semiterrestre y semiarborícola) como segundo aspecto. Todo esto permite en
un menor o mayor grado, hacer un uso diferencial del microhabitat por los sexos en cada localidad
(segregación morfológica sexual), de acuerdo a las características del hábitat y al tipo de hábito
(Emlen y Oring, 1977; Ruckstuhl, 2007).
El peso corporal presento una tendencia semejante que el LCT, debido a la influencia que este
presenta con respecto al peso corporal.
Los índices derivados del peso Ro y CF, no mostraron un patrón relacionado con las variable PC y
LCT, pero si un claro patrón para tres de las poblaciones de habitat no boscoso, ya que en este caso
en dichas poblaciones, las diferencias en estos índices siempre fueron a favor de las hembras. El
PC/LT sin embargo mantuvo un patrón semejante a las variables absolutas PC y LCT, debido a que

69
este índice se encuentra más directamente relacionado con dichas variables, por constituir el peso
por unidad lineal de medida.
Los valores de la cola relativa mostraron dimorfismo sexual destacado en las poblaciones de Sabana,
Hatibonico e Isla, lo que parece estar asociado a sus hábitos terrestres o semi terrestres.
El pie relativo expresó poco dimorfismo sexual entre poblaciones, con solo Sabana y
Guanahacabibes con valores destacados.
Tabla XVII. Índice de dimorfismo sexual (media de los machos menos media de las hembras) para ocho
variables morfométricas en ocho poblaciones de jutía conga (Capromys pilorides). Diferencias con
asteriscos señalan valores por encima de la media de las diferencias, en valores absolutos.
Población LCT PC Ro CF PC/LT CoR PaR
Jardines 6.02 344.11* 0.25* 0.20 0.59* 0.44 0.43
Sabana 2.20 -130 -0.22* -1.38* -0.34 2.77* 1.10*
Zapata 0.9 -190.97 -0.07 -0.78* -0.43* -0.02 0.45
Hatibonico 25.86* 409.09* -0.18* -0.67* 0.39 2.12* 0.4 Najasa 5.2 98.10 -0.04 0.16 0.07 1.71 0.35
Guanahacabibes 4.6 343.94* 0.18* -0.59* 0.51* 1.58 0.90*
Saetía 15.76* 285.77 -0.08 -0.23 0.30 -1.19 0.09 Isla 27.37* 777.23* 0.06 0.02 0.97* 4.7* 0.12
Si analizamos de forma general los datos de la tabla XVII, teniendo en cuenta los hábitats de las
poblaciones, vemos que las de no bosque presentaron 14 casos de dimorfismo destacado, contra
nueve para las poblaciones de bosques, influyendo sobre todo los valores de Ro y CF. Esto pudiera
explicarse por el hecho de que en los hábitats más adversos la segregación morfológica de los sexos
es mas marcada, ocupando las hembras microhabitat mas benignos, lo que produce pesos absolutos
mayores y relativos negativos, reflejados en Ro y CF. Tal situación no se daría en los habitat de
bosque, al tener éste condiciones más benignas.
Superimpuesto al anterior patrón esta el de los hábitos de las poblaciones. En zonas de no bosque,
las diferencias destacadas promedios por población no cambian según se pasa del hábito
semiarborícola al semiterrestres. Jardines (semiarborícola) solo tiene tres diferencias destacadas, y
Sabana y Zapata (ambas semiterrestres) tiene en promedio lo mismo. Para las poblaciones de
bosques si ocurren cambios. Najasa (semiarborícola) no presenta diferencias destacadas,
Guanahacabibes y Saetía tienen una media de 2.5 de diferencias destacadas y Haibonico e Isla
(terrestres) una media de 4.5. En los bosques (al menos en los semideciduos), el hábito
semiarborícola parece reducir o eliminar la segregación morfológica de los sexos, posiblemente al

70
explotarse por ambos un espacio bidimensional limitado. Para las poblaciones terrestres el espacio a
explotar sería tridimensional y por tanto con más posibilidades de segregación. La condición
semiterretre sería una situación intermedia.
Fig. 1. Diferencias sexuales (Valor de los machos-Valor de las hembras) en poblaciones de jutia
conga de bosques (arriba) y no bosques (abajo), para largo cabeza-tronco (LCT) y peso
corporal (PESO).
En las figuras se observa que la regla de Rensh Depende del gradiente arborícola (Najasa/Jardines)-
terrestre (Isla/Hatibonoco) y del tipo de habitat (Bosque, vs no bosque).
De los siete factores que discute Isaac (2005), y que pueden producir dimorfismo sexual de tamaño
en los mamíferos (selección sexual, latitud, clima, alimentos, densidad, crecimiento diferencial y
segregación del nicho ecológico), para el caso de la jutía conga solo nos hemos referidos a la
segregación del nicho ecológico de hábitat y hábitos, pero algunos de los otros aspectos antes
mencionados, también pudieran estar jugando su papel.
Grado de diferenciación de las poblaciones
Nuestros resultados confirman con mas poblaciones y muestras los trabajos preliminares de Smith y
Berovides (1984b), Berovides et al (1990a y b) y Berovides (2005) en relación a la existencia de
ecotipo en la jutía conga, o sea poblaciones con caracteres morfológicos (además de fisiológicos y
conductuales) perfectamente adaptables a las condiciones del hábitat de cada población. La
existencia de dichos ecotipos apoya fuertemente el reconocimiento de cada uno de ellos como una
unidad de manejo para la conservación, ya que poseen uno de los dos criterios de Crandall et al
(2000) para el reconocimiento de dichas entidades, o sea la ocupación por cada población de un
nicho ecológico diferente. El otro criterio, el aislamiento genético parcial, esta por investigar, pero
observaciones anecdóticas señalan su existencia.
0
10
20
30
40
50
60
70
80
90
Najasa Guanaha Saetía Isla
poblaciónes
dif
ere
ncia
s
LCT
PESO
-30
-20
-10
0
10
20
30
40
50
Jardines Sabana Zapata Hatibonico
poblaciones
dif
ere
ncia
s
LCT
PESO

71
Efecto de los trimestres sobre las diferencias entre localidades
Largo cabeza-tronco y peso
La tabla XVIII muestra los valores medios y desviación típica del largo cabeza-tronco (LCT) en
cinco poblaciones de jutía conga en los trimestres febrero, marzo y abril (FMA) y agosto,
septiembre y octubre (ASO), así como la dinámica estacional para las poblaciones de Jardines y
Najasa en los cuatro trimestres del año.
Se observo diferencias estadísticamente significativas entre poblaciones dentro de cada trimestre con
el siguiente patrón. En el trimestre FMA las poblaciones de manglar y manglar más vegetación de
costa de playa (Jardines y Sabana) mostraron valores promedios del LCT inferiores a las
poblaciones de bosque semideciduo (Najasa, Guanahacabibes e Isla). En el trimestre ASO (lluvia)
hubo un mayor solapamiento estadístico entre las poblaciones, donde solo la población de la Isla se
diferenció estadísticamente del resto de las poblaciones, lo cual pudiera estar relacionado con
posibles cambios estratégicos en la reproducción (más estrategas r que K para las poblaciones de
manglar) que conllevarían a la inclusión de forma diferencial en el tiempo de grupos de juveniles
maduros sexualmente en las poblaciones de manglar ( Gutierrez et al., 1999 ), con un aumento del
LCT en el trimestre ASO donde alcanzan su valor máximo. En las poblaciones de manglar los
cambios de seca a lluvia al parecer son más acentuados, lo que conllevaría a una estrategia
reproductiva diferente, sustentada por el hecho de que en los meses de lluvia apenas se observan
gestantes en estas poblaciones, lo cual evitaría el nacimiento en los meses de seca y utilizar los
meses de lluvia para recuperarse.
Las poblaciones de Jardines y Najasa presentaron diferencias estadísticamente significativas entre
poblaciones para los trimestres noviembre, diciembre y enero (NDE), FMA y mayo, junio y julio
(MJJ), con valores promedios más bajos para la población de Jardines con respecto a Najasa. En el
trimestre de ASO no se mostraron diferencias estadísticamente significativas lo cual coincide con la
mayoría de las poblaciones analizadas para este trimestre.
Tabla XVIII. Valores promedios (X) y desviación típica (S) del largo cabeza tronco (LCT) en los trimestres
noviembre, diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y
agosto, septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril
(FMA) y agosto, septiembre y octubre (ASO) para cinco poblaciones de jutía conga (Capromys
pilorides). Medias con índices diferentes difieren a p<0.05 por una prueba de Tukey
72
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 465.56a 31.40 458.70b 33.40 445.84a 21.01 469.77a 30.37
Sabana ---- ---- 456.77a 30.17 ---- ---- 479.12a 37.53 Najasa 503.20b 30.65 488.84c 35.51 476.74b 35.42 488.91a 37.85
Guanahacabibes ---- ---- 489.51c 39.31 ---- ---- 488.92a 28.57
Isla ---- ---- 525.61d 32.68 ---- ---- 528.23b 29.41
Los valores promedios y desviación típica del peso corporal (PC) en cinco poblaciones de jutía
conga para los trimestres de FMA y ASO, así como los cuatros trimestres NDE, y MJJ para las
poblaciones de Jardines y Najasa se expresan en la tabla XIX.
Los resultados obtenidos muestran diferencias estadísticamente significativas entre las poblaciones
tanto en el trimestre FMA como en el trimestre ASO. En el trimestre ASO también se observó un
mayor solapamiento estadístico entre las poblaciones al igual que con la variable LCT, influenciado
por el aumento del LCT en el trimestre ASO para las poblaciones de manglar y la estabilidad para
esta variable en los dos trimestres (FMA y ASO) en las poblaciones de bosque semideciduo y la alta
correlación entre el PC y el LCT.
Diferencias estadísticamente significativas en los trimestres NDE, FMA Y MJJ entre las poblaciones
de jardines y Najasa con valores promedios más altos para Najasa.
El peso como variable altamente correlacionada con el LCT manifestó un patrón semejante a este,
separando a las poblaciones de hábitat manglar y manglar más vegetación de costa de playa de las
poblaciones de bosque semideciduo.
El PC no solo se encuentra influenciado por el tamaño, los parámetros como la precipitación,
humedad y temperatura que inciden en la productividad primaria y que cambian de un trimestre a
otro son aspectos ambientales que influyen en el PC también de forma independiente.
Tabla XIX. Valores promedios (X) y desviación típica (S) del peso corporal (PC) en los trimestres
noviembre, diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y
agosto, septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril
(FMA) y agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys
pilorides). Medias con índices diferentes difieren a p<0.05 por una prueba de Tukey.
73
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 2448.69a 373.43 2405,31a 532,55 2288.75a 336.75 2925.14a 573.81
Sabana ---- ---- 2811,44a 576,86 ---- ---- 3592.25bc 986.47 Najasa 2963.33b 687.24 3509,11b 816,19 3418.26b 909.42 3451.42ab 858.99
Guanahacabibes ---- ---- 3853,31b 1007,48 ---- ---- 3675.00bc 672.18
Isla ---- ---- 4592,90c 912,75 ---- ---- 4276.92c 946.40
La tabla XX expresan los valores promedios y desviación típica del índice de robustez (Ro) para los
trimestres FMA y ASO en cinco poblaciones de jutía conga y los trimestres NDE y MJJ para dos
poblaciones.
Diferencias estadísticamente significativas entre las poblaciones se observan, para los trimestres de
FMA y ASO, agrupándose en tres niveles estructurados de la siguiente manera, grupo 1 Jardines e
Isla, grupo 2 Sabana con Najasa y el grupo 3 Guanahacabibes, (para el trimestre FMA) y en dos
niveles de grupo para el trimestre ASO conformados de la siguiente manera, grupo 1 Jardines, Isla y
Najasa; grupo 2 Sabana y Guanahacabibes. Existe un patrón semejante entre ambos trimestres auque
con un mayor solapamiento entre las poblaciones para el trimestre de ASO asociado a las variable
LCT y PC las cuales se utilizan para elaborar este índice.
El índice de Ro distribuye los gramos de peso por volumen cúbico, pero no tiene en cuenta el largo
total pues omite el largo de la cola que si queda incluida en el PC, si tomamos en consideración que
el largo de la cola (Co) varia de forma absoluta y relativa con el hábito de la especie y que por ende
representa porcientos diferentes del LCT y del peso corporal para las diferentes poblaciones, los
valores de Ro pudieran estar influenciado por:
1. Inclusión de juveniles maduros sexualmente con una larga cola relativa (CoR).
2. 2 Población de hábitat semiarborícola como Najasa y un alto valor de cola relativa, mostrar
un valor de Ro bajo.
3. 3 Poblaciones como Sabana de hábito semiterrestre y una CoR baja, presentar un valor alto
de Ro
En las poblaciones de Jardines y Najasa para los cuatro trimestres del año (NDE, FMA, MJJ y ASO)
se mostraron diferencias estadísticamente significativas en los trimestres FMA, MJJ entre
poblaciones, con valores promedios más altos para la población de Najasa, observándose
solapamiento estadístico en los trimestres de NDE y ASO.

74
Tabla XX. Valores promedios (X) y desviación típica (S) índice de Robustez, en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de Tukey.
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 2,44a 0,41 2,47a 0,33 2,44a 0,41 2,47a 0,33
Sabana ---- ---- 2,94b 0,48 ---- ---- 2,94b 0,48
Najasa 2,32a 0,50 2,96b 0,35 2,32a 0,50 2,96b 0,35 Guanahacabibes ---- ---- 3,24c 0,47 ---- ---- 3,24c 0,47
Isla ---- ---- 3,13ab 0,33 ---- ---- 3,13ab 0,33
Los valores promedios y desviación típica de la condición física (CF) en cinco poblaciones de jutía
conga para los trimestres de FMA y ASO, así como los trimestres NDE y MJJ para las poblaciones
de Jardines y Najasa se exponen en la tabla XXI.
Se observan diferencias estadísticamente significativas entre las poblaciones para el trimestre FMA.
No hubo diferencias estadísticamente significativas entre las poblaciones en el trimestre de ASO, ni
en los trimestres NDE y MJJ entre las poblaciones de Jardines y Najasa. La variable CF se
mantuvo constante dentro de cada población en los diferentes trimestres.
El índice de condición física (CF) distribuye el peso a una medida lineal como el largo de la pata,
siendo influenciado este índice por el peso corporal y una variable como la pata que se encuentra
asociada al hábito de la población en relación al hábitat, lo cual hizo posible que poblaciones como
Najasa de hábitat bosque semideciduo y hábito semiarborícola se agrupen en el trimestre FMA
(donde son más marcadas las diferencias de tamaño) con poblaciones de hábitat manglar.
Tabla XXI. Valores promedios (X) y desviación típica (S) índice de Condición Física, en los trimestres
noviembre, diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y
agosto, septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril
(FMA) y agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys
pilorides). Medias con índices diferentes difieren a p<0.05 por una prueba de TuKey

75
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 15,25a 0,93 14,79a 1,03 15,50a 0,76 15,93a 1,06
Sabana ---- ---- 15,73a 1,46 ---- ---- 16,37a 1,51 Najasa 15,49a 0,94 15,82a 1.21 14,83a 1,43 16,79a 1,66
Guanahacabibes ---- ---- 16,56b 1,13 ---- ---- 16,43a 0,83
Isla ---- ---- 16,59b 1,27 ---- ---- 15,99a 0,72
La tabla XXII muestra los valores promedios y desviación típica del índice peso por milímetro lineal
(PC/LT) para los trimestres FMA y ASO en cinco poblaciones de jutía conga y los trimestres NDE y
MJJ para las poblaciones de Jardines y Najasa.
Se observaron diferencias estadísticamente significativas entre poblaciones en los Trimestres FMA y
ASO con un mismo patrón que los índices de Ro y CF, solo que en este caso el índice PC/LT al ser
una variable que depende del Largo total y del PC además de no tomar en cuenta los apéndices
(cola y pata) para relativizar el índice, se mostró como la variable que más diferenció las
poblaciones en los trimestres de FMA y ASO por el hecho que no utiliza variables adaptativa con
una fuerte influencia con el hábito como la cola y la pata, haciendo más dependiente la distribución
del peso en dependencia del hábitat y de la relación propia del PC con el hábito de la población.
Las poblaciones de hábitat manglar mostraron valores promedios más bajos versus las poblaciones
de bosques con valores promedios más altos, en ambos grupos las poblaciones de hábitos
semiarborícola mostraron los valores promedios más bajos como se aprecia dentro de las
poblaciones de mangle en Jardines que es una población de mangle con hábito semiarborícola y en
Najasa que es población de bosque semideciduo con hábito semiarborícola
Las poblaciones de Jardines y Najasa mostraron diferencias estadísticamente significativas entre
poblaciones en los trimestres NDE, FMA y MJJ, auque en el trimestre ASO no hubo diferencias
entre las dos poblaciones, la población de Najasa mostró valores promedios más altos
Tabla XXII. Valores promedios (X) y desviación típica (S) índice de peso por milímetro lineal, en
los trimestres noviembre, diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo
junio y julio (MJJ) y agosto, septiembre y octubre (ASO) para dos poblaciones y en los
trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO) para cinco
poblaciones jutía conga (Capromys pilorides). Medias con índices diferentes difieren a
p<0.05 por una prueba de TuKey.

76
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 3,39a 0,43 4,65a 0,81 3,30a 0,36 4,01a 0,65
Sabana ---- ---- 5,52b 0,95 ---- ---- 4,94bc 1,11 Najasa 3,86b 0,81 6,40c 1,18 4,56b 1,03 4,55ab 0,93
Guanahacabibes ---- ---- 7,09d 1,42 ---- ---- 5,08bc 0,80
Isla ---- ---- 7,96e 1,23 ---- ---- 5,38c 0,83
El LCT, PC e índices derivados del largo y el peso, expresan diferencias estadísticamente
significativas entre poblaciones para los trimestres FMA y ASO en mayor o menor medida de
acuerdo a la variable utilizada en relativizar el índice y el valor adaptativo de la misma, dependiendo
en todos los casos del hábitat y el hábito de la población. Las condiciones medianbientales más
favorables en los hábitat de bosque (disponibilidad de alimentos) permiten una estrategia
reproductiva más prolongada, lo cual incide en la inclusión diferencial de jóvenes maduros
sexualmente en el conjunto de las poblaciones estudiadas, teniendo en cuenta además que las
poblaciones de manglar la actividad reproductiva cesa en determinados trimestres del años, siendo
menos frecuente la inclusión de jóvenes maduros sexualmente durante la mayoría de los trimestres
del año, al parecer una de las causas del aumento del LCT y PC en el trimestre de ASO respecto a
FMA para las poblaciones de manglar y una estabilidad en la poblaciones de bosque en relación a
estos trimestres con su respectiva incidencia en los índices derivados del LCT y el PC.
El análisis de las diferencias estadísticamente significativas entre trimestres dentro de las
poblaciones para las variables LCT, PC, Ro, CF y PC/LT se muestran a continuación:
1. Diferencias estadísticamente significativas en las poblaciones de Jardines y Sabana para
todas las variables y con mayores valores promedios en el trimestre ASO, donde las
condiciones del hábitat se hacen más favorables.
2. Similitud para las variables LCT, PC y PC/LT entre los trimestres analizados para las
poblaciones de Najasa, Guanahacabibes e Isla, las cuales son poblaciones de bosque
semideciduo donde los cambios de seca a lluvia no son tan drásticos en cuanto a la
disponibilidad de nutrientes como en las poblaciones de hábitat xérico.
3. Diferencias estadísticamente significativas para la Ro entre trimestres para la población de la
Isla asociado a diferencias estadísticamente significativas de la cola absoluta y la cola
relativa.

77
4. Diferencias estadísticamente significativas para la CF entre trimestres para la población de
Najasa, asociado a diferencias estadísticamente significativas de la pata absoluta y el pie
relativo.
Cola y pie
Los valores promedios y desviación típica del pie absoluto (Pi) en cinco poblaciones de jutía conga
para los trimestres de FMA y ASO, así como los cuatros trimestres del año (NDE, FMA, MJJ y
ASO) para las poblaciones de Jardines y Najasa se observan en la tabla XXIII.
Se observaron diferencias estadísticamente significativas entre las poblaciones para los trimestres de
FMA y ASO, así como diferencias estadísticamente significativas en los trimestres NDE y MJJ entre
poblaciones de Jardines y Najasa.
La variable pata absoluta se mostró influenciada por el LCT debido a la correlación existente entre
ambas variables, separando a las poblaciones en función del tamaño e influenciado también auque
en menor medida por el hábito de la población, sobre todo en el trimestre FMA, efecto que cambió
en el trimestre ASO donde se observó un mayor solapamiento estadístico para la variable pata
asociado a un mayor solapamiento estadístico del LCT, lo cual favoreció que para este trimestre
influenció algo más el hábitat que el hábito en la separación de los grupos.
Tabla XXIII. Valores promedios (X) y desviación típica (S) del pie absoluto, en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de TuKey
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 88,21a 4,12 90,09a 3,12 85,31a 3,33 89,40a 4,14
Sabana ---- ---- 90,06a 5,77 ---- ---- 92,75a 5,21
Najasa 92,20b 5,93 96,44c 8,67 98,43b 5,13 89,68ab 5,47 Guanahacabibes ---- ---- 94,13b 6,81 ---- ---- 93,67b 5,92
Isla ---- ---- 99,99d 6,44 ---- ---- 99,94c 4,95

78
La tabla XXIV muestra los valores promedios y desviación típica del índice pie relativo (PiR) para
los trimestres FMA y ASO en cinco poblaciones de jutía conga y los trimestres NDE y MJJ para las
poblaciones de Jardines y Najasa.
Al eliminar el efecto del tamaño, los valores promedios de la PiR no mostraron diferencias
estadísticamente significativas entre las cinco poblaciones para los trimestres de FMA y ASO, ni en
el trimestre NDE entre las poblaciones de Jardines y Najasa.
Las diferencias estadísticamente significativas entre las poblaciones de Jardines y Najasa para el
trimestre de MJJ pudieran estar relacionado con la inclusión en el muestreo de juveniles maduros
sexualmente de forma diferencial en la población de bosque con respecto a las de manglar, lo cual
ocasionó el incrementó del índice en Najasa para este trimestre y el valor promedio mayor para lo
cuatro trimestres estudiados en el trimestre FMA para la población de Jardines.
Tabla XXIV. Valores promedios (X) y desviación típica (S) del pie relativo, en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de TuKey
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 19,00a 1,18 19,72a 1,41 19,15a 0,83 19,09a 1,26
Sabana ---- ---- 19,76a 1,31 ---- ---- 19,44a 1,51 Najasa 18,37a 1,50 19,80a 2,00 20,72b 1,36 18,43a 1,63
Guanahacabibes ---- ---- 19,22a 1,44 ---- ---- 19,17a 0,89
Isla ---- ---- 19,06a 1,28 ---- ---- 18,94a 0,84
Los valores promedios y desviación típica de la cola absoluta (Co) en los trimestres de FMA y ASO
para cinco poblaciones de jutía conga, así como los valores promedios y desviación típica de los
trimestres NDE y MJJ para las poblaciones de Jardines y Najasa se observan en la tabla XXV.
Se observaron diferencias estadísticamente significativas entre las poblaciones en los trimestres
FMA y ASO. La separación de estos grupos en el trimestre FMA se asocian a la influencia que
presenta el LCT sobre la variable cola debido a la correlación existente entre ambas variables,
separando a las poblaciones en función del tamaño y en menor medida por el hábito de la población,
lo cual hace que poblaciones terrestres coma la Isla y semiarborícola como Najasa queden en el

79
mismo grupo y separadas de otras poblaciones semiterrestres (Guanahacabibes y Najasa) y
semiarborícolas (Najasa). En el trimestre ASO se observó un mayor solapamiento estadístico, lo cual
se asocia a una mayor igualdad entre los LCT de las poblaciones ocasionando la separación de los
grupos más por el habito que por la influencia del tamaño sobre esta variable.
En el trimestre FMA en las poblaciones de Jardines y Najasa se observaron diferencias
estadísticamente significativas, igualmente asociadas a la influencia del LCT y a las diferencias para
este trimestre y el efecto que este ejerce sobre el largo de la cola, no habiendo diferencias
estadísticamente significativas en los trimestres NDE, MJJ y ASO lo cual resulta concordante con
los hábitos de estas poblaciones ya que son poblaciones de hábito semiarborícola.
TablaXXV. Valores promedios (X) desviación típica (S) de la cola absoluta en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de TuKey.
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 254,04a 17,04 248,12b 17,92 245,25ª 20,39 255,44b 14,66 Sabana ---- ---- 224,66a 15,71 ---- ---- 238,12a 20,01
Najasa 260,40a 15,68 264,13c 20,95 267,00a 18,55 262,77b 16,75
Guanahacabibes ---- ---- 233,14a 23,14 ---- ---- 231,07a 13,32 Isla ---- ---- 250,78b 16,67 ---- ---- 238,84a 23,64
Los resultados de los valores promedios y desviación típica en cinco poblaciones para la cola
relativa (CoR) en los trimestres FMA y ASO así como los valores promedios y desviación típica en
los trimestres NDE y MJJ en las poblaciones de Jardines Y Najasa , se muestran en la tabla XXVI.
Se observaron diferencias estadísticamente significativas entre poblaciones para los trimestres de
FMA y ASO con un patrón semejante en ambos trimestres y separando las poblaciones de acuerdo al
hábito debido a que se elimina el efecto del tamaño en la elaboración del índice, mostrando valores
promedios mayores para las poblaciones de hábito semiarborícola como Jardines y Najasa y valores
promedios menores en las poblaciones de hábitos semiterrestres y terrestes como las poblaciones de
Sabana, Guanahacabibes e Isla respectivamente.

80
En el trimestre MJJ auque no se mostró diferencias estadísticamente significativas sí se evidenció la
tendencia hacia un valor promedio mayor en la población de Najasa respecto a Jardines relacionado
posiblemente por las mismas causas explicadas anteriormente para este mismo trimestre en el índice
PaR, lo cual se hace constante para los apéndices, aspecto que se evidenció de forma inversa en el
trimestre NDE.
TablaXXVI. Valores promedios (X) y desviación típica (S) de la cola relativa, en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de TuKey.
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 54,70b 3,93 54,29b 4,79 54,98a 3,43 54,51c 3.495 Sabana ---- ---- 48.77a 4.55 ---- ---- 49,91b 4.900
Najasa 52,92a 4,48 54,23b 4,84 56,21a 4,53 54,00c 4,696
Guanahacabibes ---- ---- 47,77a 4,66 ---- ---- 47,34ab 2.850 Isla ---- ---- 47,47a 3.27 ---- ---- 44.60a 3.210
La tabla XXVII refleja los valores promedios y desviación típica del índice pata/cola (Pi/Co) para
los trimestre FMA y ASO en cinco poblaciones de jutía conga y los trimestres NDE y MJJ para las
poblaciones de Jardines y Najasa.
Los valores promedios del índice mostraron diferencias estadísticamente significativas entre las
poblaciones para los trimestres FMA y ASO diferenciándose las poblaciones de hábitos
semiarborícola (Jardines y Najasa) de las poblaciones de hábitos semiterrestres y terrestres (Sabana,
Guanahacabibes e Isla respectivamente).
Las poblaciones de Jardines y Najasa de hábito semiarborícola no mostraron diferencias
estadísticamente significativas.
El índice pata cola representa la proporción de la pata en la cola, por lo tanto los valores promedios
mayores significan una pata más grande en relación a la cola o una cola más pequeña en relación a
la pata lo cual indica que las poblaciones de hábitos semiarborícolas se representaron con los
valores promedios más bajos y en las poblaciones terrestres y semiterrestres se observaron los
valores más altos.

81
El índice pata cola reflejó de forma eficiente el hábito de la población y sugiere el mayor efecto de
la cola con respecto a la pata en la actividad trepadora de la especie.
Tabla XXVII. Valores promedios (X) y desviación típica (S) del índice pie/cola, en los trimestres noviembre,
diciembre y enero (NDE); febrero, marzo y abril (FMA); mayo junio y julio (MJJ) y agosto,
septiembre y octubre (ASO) para dos poblaciones y en los trimestres febrero, marzo y abril (FMA) y
agosto, septiembre y octubre (ASO) para cinco poblaciones jutía conga (Capromys pilorides). Medias
con índices diferentes difieren a p<0.05 por una prueba de TuKey
Población NDE FMA MJJ ASO
X S X S X S X S
Jardines 34,81a 2,00 36,45a 2,35 34,98a 2,79 35,09a 2,431
Sabana ---- ---- 40,72b 3,22 ---- ---- 39,16b 3,412 Najasa 35,49a 2,59 36,95a 3,10 36.99b 2,53 34,23a 2,729
Guanahacabibes ---- ---- 40,55b 3,47 ---- ---- 40,59bc 2,470
Isla ---- ---- 40,29b 3,28 ---- ---- 42,80c 2,456
Los índices PiR y CoR no presentan el efecto del tamaño debido a que en la elaboración de los
mismos se utiliza como factor de corrección el LCT. El índice PiR no mostró diferencias
estadísticamente significativas entre poblaciones para los trimestres FMA y ASO, lo cual sugiere no
ser un buen indicador de la actividad trepadora, a diferencia deL índice CoR que si separó a las
poblaciones por el hábito de las mismas.
Los apéndices pie y cola absoluta mostraron diferencias estadísticamente significativas entre
poblaciones para los trimestres FMA y ASO. Debido a la correlación existente con el LCT,
separando a las poblaciones en el trimestre FMA (donde las diferencias en cuanto al LCT entre
poblaciones son más acentuadas) en función del tamaño, característica que disminuye en el
trimestre ASO donde al disminuir los efectos del tamaño la separación se efectúa más en función del
hábito que por la influencia del LCT, fundamentalmente para la variable cola absoluta ya que los
grupos que se forman en el trimestre ASO para la pata absoluta, no son concordantes con el índice
PiR para este mismo trimestre, que no mostró diferencias estadísticamente significativas,
ratificando el poco valor indicativo para la actividad trepadora de este índice.
El índice pata cola no presentó la influencia del LCT y si mostró diferencias estadísticamente
significativas entre poblaciones en función del hábito de la población en los trimestres FMA y ASO,
dando muestras de ser un buen indicador da la actividad trepadora.

82
El análisis de las variables cola y pata de forma absoluta y relativa, así como el índice pie/cola
dentro de las poblaciones en los trimestres analizados mostró diferencias estadísticamente
significativas, cumpliendo con las siguientes condiciones:
1. Los mayores valores promedios de la pata relativa y la cola relativa entre trimestres dentro
de una misma población se corresponden con cambios en la estructura de edad por la
inclusión de juveniles maduros sexualmente que presentan los mayores valores promedios
de estos apéndices de forma relativa, asociado a que los apéndices cola absoluta y pata
absoluta alcanzan valores próximos a su máximo valor en una etapa temprana de su
desarrollo ontogenético antes de alcanzar el máximo valor del LCT, condición que se
observó en la población de Jardines para la pata relativa.
2. Diferencias estadísticamente significativas de cola y pata absoluta pero no de forma relativa
entre trimestres dentro de una misma población se asocian a un incremento de los apéndices
absolutos en correspondencia con el aumento del tamaño, el cual se observó en la población
de Sabana.
3. Las diferencias estadísticamente significativas de la cola absoluta y de la cola relativa entre
trimestres dentro de una población, corresponden al aumento de la cola absoluta en un
trimestre diferente al trimestre en el que aumentó la cola relativa, condición que se evidenció
en la población de la Isla.
4. Diferencias estadísticamente significativas de la pata absoluta y de la pata relativa entre
trimestres dentro de una población, corresponden al aumento de la pata absoluta en un
trimestre diferente al trimestre en el que aumentó la pata relativa, aspecto que se observó en
la población de Najasa
Efecto de la población sobre las diferencias entre trimestres
Las diferencias entre la estación de seca (FMA) y la estación de lluvia (ASO), para las variables
LCT, PC, Ro, CF, PC/LT, CoR, PiR y Pi/Co se muestran en la tabla XXVIII. Los resultados
reflejaron diferencias para la variable LCT en las poblaciones de Jardines y Sabana y diferencias en
todas las poblaciones con respecto a la variable PC, siendo esta última, la variable que más
diferencias mostró en todas las poblaciones con un mayor valor para la población de Sabana.
En las poblaciones de bosque se observa una estabilidad entre los trimestres FMA y ASO para esta
variable , aquí los cambios de seca a lluvia son al parecer menos acentuados, lo que sugiere el hecho

83
de que estas poblaciones se demoran más en alcanzar la madures sexual y que como estrategias
reproductivas para estas poblaciones, su reproducción es durante todo el año, con una disminución
al 50% en los trimestres ASO y NDE pero mantenida y no cesa abruptamente como en las
poblaciones de manglar ( Gutierrez et al., 1999).
Tabla XXVIII. Valores promedios de las diferencias (estación de lluvia menos la estación de seca) dentro
poblaciones para las variables largo cabeza tronco (LCT), peso corporal (PC), robustez (Ro),
condición física (CF), peso por milimetro lineal (PC/LT), cola relativa (CoR) y pie/cola (Pi/Co) en
cinco poblaciones de jutía conga (Capromys pilorides). += p< 0.10; *= p < 0.05
Población LCT PC Ro CF PC/LT CoR PiR Pi/Co
Jardines 11.04+ 519.83* 0.34* 1.13* 0.75* 0.22+ -0.63+ -1.36
Sabana 22.35* 725.25* 0.16 0.07 0.61* 1.14+ -0.32* -1.56
Najasa 0.07 -57.69 -0.05 0.01 -0.07 -0.23 -1.51 -2.62 Guanahacabibes -0.59 -178.31 -0.11 -0.04 -0.03 -0.43 -0.10 0.04
Isla 2.72 -315.98 -0.39 -0.14 -0.55 -2.86 -0.12 2.51
III.4. Conclusiones
1. Las poblaciones de hábitat bosque presentan un mayor largo cabeza tronco, peso corporal, e
índices derivados del peso como la Robustez, Condición Física y peso por milímetro lineal,
con respecto a las poblaciones de hábitat manglar.
2. Los índices pata y cola relativos se relacionaron con los hábitos de la especie dentro de cada
localidad con un mayor o menor valor de los mismos de acuerdo al grado de menor o mayor
arboricidad respectivamete.
3. Las variables LCT, Pc y PC/LC mostraron mayor dimorfismo sexual dentro de cada
población con una tendencia al incremento del dimorfismo sexual en las poblaciones de
mayor tamaño corporal.
4. Las variables LCT, PC, PC/LT y Pi presentaron una dependencia estadísticamente
significativa con el índice climático de Martonne que está relacionado con la precipitación,
temperatura y la productividad primaria de los hábiat.

84
5. Las variables PC, Co y Pi presentaron dependencias estadísticamente significativas con
respecto al LCT con una tendencia a valores mayores para las poblaciones de hábitat de
bosque.
CAPÍTULO IV. Análisis comparativo de los índices de reabsorción de agua en
poblaciones de jutía conga (Capromys pilorides)
IV.1. Introducción
Los cambios estacionales de seca (febrero, marzo y abril) a lluvia (agosto, septiembre y octubre)
representan cambios relevantes en cuanto a la disponibilidad de agua en los ecosistemas estudiados,
haciendo estas diferencias más apreciables de acuerdo a las características de los hábitats.
La retención de agua en los animales terrestres, es una de las adaptaciones a la escases de este
recurso en los hábitats con ambientes predominantemente secos En los mamíferos es la presencia de
asas de Henle largas la encargada de concentrar la orina e impedir una pérdida excesiva de agua,
demostrándose que el grosor relativo de la médula renal es directamente proporcional a la longitud
de las asas de Henle (Al-kahtani et. al, 2004).
Estudios realizados por Díaz ()001) han evidenciado en mamíferos argentinos de tierras áridas, los
cambios estacionales del grosor de la médula y corteza renal, los cuales cambian en función de la
dieta y la estación del año. Como este autor y otros (Reaka y Armintage, 1976; Ojeda et al., 1999;
Ntshotsho et al., 2004; Bosonovic y Gallardo, 2005) establecen que las diferencias entre poblaciones
se relacionan con la disponibilidad de agua y como este recurso cambia de forma estacional, es de
esperar que también lo hagan las medidas e índices de los órganos relacionado con la retención del
agua.
Priersma y Drent (2003) han afirmado que el estudio de la diversidad fisiológica es fundamental
para entender como los organismos se enfrentan a las variaciones ambientales en espacio y tiempo.

85
De igual forma, Al-kahtani et. al (2004), llevaron a cabo un análisis de la capacidad de retención de
agua en varias especies de roedores de Chile y Argentina. En el mismo compararon poblaciones de
hábitat mésicos y áridos a través de un índice renal, que tuvo en cuenta el grosor de la médula del
riñón relativo a su volumen y obtuvieron que las poblaciones de hábitat áridos presentaron mayores
valores de este índice que las de hábitat mésicos. Por su parte, Díaz y Ojeda (2006), realizaron un
estudio para determinar la asociación entre el tipo de hábitat y la morfología del riñón en varias
especies de roedores histricomorfos sudamericanos. Para ello, midieron numerosas variables de los
riñones y crearon índices renales a partir de estas. Los resultados obtenidos sugieren que las especies
de hábitat con bajas precipitaciones, poseen riñones más grandes para impedir una mayor pérdida de
agua.
En la jutía conga también se han realizado estudios del índice renal de Yabe donde se compara la
capacidad de retención de agua entre poblaciones con diferentes tipos de hábitat (bosque y manglar)
evidenciando que las jutías que viven en los manglares presentaron un grosor relativo de la médula
renal mayor que las adaptadas a ambientes más húmedos, como bosques secundario (Sánchez et
al.,1992), no obstante no se analizaron los cambios estacionales y las posibles variaciones en el
índice utilizado en relación a la precipitación.
Estos estudios morfofisiológicos podrían servir para diferenciar unidades de manejo para la
conservación entre las poblaciones estudiadas, como se ha demostrado en relación a la longitud
relativa a la última porción del intestino grueso (de la Fuente y Berovides, 2009)
El objetivo de nuestro trabajo fue analizar comparativamente, poblaciones de jutía conga en
diferentes hábitats (manglar y bosque) mediante dos índice de retención de agua (índice de
reabsorción) utilizando el índice propuesto por Yabe y un índice propuesto por el autor, donde se
relaciona el grosor de la médula con una medida lineal que elimine el posible efecto del tamaño
(LCT) en dos trimestres del año.
IV.2. Materiales y métodos
Análisis dentro de una estación del año.
Los muestreos se llevaron a cabo durante un año en el trimestre febrero, marzo y abril, que se
corresponde con los finales del periodo de seca o poco lluvioso en Cuba. Esto se hizo para evitar el
posible efecto estacional de los índices estudiados, ya que nuestro interés se centraba en el análisis

86
de la variación de los índices entre las poblaciones, como medidas de adaptación local a la escasez o
abundancia de agua en los hábitats de dichas poblaciones.
Las poblaciones fueron seleccionadas sobre la base de diferencias en cuanto a formación vegetal,
precipitaciones y disponibilidad de agua (Tabla I).
Fueron capturados un total de 178 individuos utilizando perros y escopeta, Todos los animales
fueron adultos según la metodología de Smith y Berovides (1984) y se les determinó el sexo y el
largo cabeza tronco. Con posterioridad se les extrajeron los riñones y con un bisturí quirúrgico se le
aplicó un corte longitudinal, midiendo el ancho de la médula (M) y tres medidas de ancho de corteza
(C1, C2 y C3). Las mediciones fueron tomadas con un pie de rey con error de 0.01 mm. Con estas
variables se confeccionó el índice renal propuesto por Yabe (1983), el cual mide el grosor de la
médula renal relativo al grosor del riñón y el índice propuesto por el autor, que mide el grosor de la
médula renal relativo al largo cabeza tronco (LCT).Los índices se elaboraron a partir de las
siguientes formulas
IRRY =M / (M+∑(Ci)/3)
IRRL=M / LCT
Donde: IRRY es el índice renal de Yabe e IRRL es el índice renal de los autores, M es el grosor de la
médula renal, C es el valor promedio de las medidas de la corteza renal y LCT es el largo cabeza
tronco.
En la tabla I se dan las precipitaciones y temperaturas medias anuales que nos permitieron calcular
el grado de humedad de los hábitats en cada localidad por el índice climático de Martonne
(Bozinovic et al., 2009), por medio de la fómula :
IC=Precipitación media anual/Temperatura media anual +10 0 C
Este índice está asociado a la productividad vegetal de los hábitats y a la tasa metabólica basal
(independiente del peso) en los individuos de la población (Bozinovic et al., 2009).
Este índice realmente sólo tiene significado para las poblaciones de jutía conga de tierra firme, ya
que para los manglares de Juan Clarito (Sabana) y Jardines, la limitación está en la carencia total de
agua dulce en los manglares y cayos, independiente de la precipitación. Por esta razón no se
tomaron los valores del índice como tal, sino que estos fueron convertidos a rango, donde el rango 1
representó alta disponibilidad de agua dulce y el 6 poca o nula disponibilidad.

87
Tabla I. Formaciones vegetales, precipitaciones y temperaturas medias anuales, índices climáticos y sus
rangos (en orden decreciente de disponibilidad de agua), en seis localidades donde viven poblaciones
de jutía conga.
Localidad N Formación
vegetal
Precipitación
/
Temperatura
Madia anual
Indice
climático /
rango
Cayo Saetías (norte Holguín)
28 Bosque semideciduo
secundario
1400/26 38.89/1
Península de
Guanahacabibe (Guanahacabibes, P. Río)
47 Bosque
semideciduo sobre Carso
1350/25.8
37.7/2
Najasa (sur Camaguey) 26 Bosque
semideciduo
secundario
1300/25.0 37.14/3
Hatibonico (Guantánamo) 29 Vegetación
xerofítica
600/28 15.79/4
Cayo Juan Clarito (Archipiélago Sabana-Camaguey)
23 Manglar (varias especies)
1000/26.5 -/5
Jardines
(sur Camaguey)
25 Manglar
(mangle rojo)
1000/27 -/6
Los datos se ajustaron a la distribución normal y cumplieron con la homogeneidad de varianza, lo
cual se comprobó mediante las pruebas estadísticas Kolmogorov-Smirnov y Test de Levene,
respectivamente.
Para determinar las diferencias en valores promedios entre poblaciones con respecto a los índices
renales, se efectuó un análisis de varianza (ANOVA) bifactorial , tomando en cuenta los efectos
localidad y sexo (estando estos equilibrados en su proporción sexual).Posteriormente se realizó la
prueba Tukey de comparación de medias.. Se llevaron a cabo dos regresiones lineales simples,
tomando como variable independiente el rango de disponibilidad de agua por hábitat y como
variable dependiente el valor promedio de los índices renales por población y su coeficiente de
variación.

88
Como el índice de Yabe se ve afectado tanto por el grosor absoluto de la médula como de la corteza,
se realizaron también análisis de varianzas para ambas variables, teniendo en cuenta los efectos
población y sexo.
Análisis entre estaciones del año. Se analizaron tres poblaciones para el trimestre Febrero, Marzo y
Abril (FMA, finales de la estación seca) y el trimestre Agosto, Septiembre y Octubre (ASO, finales
de la estación de lluvia), las muestras por trimestres, sexos y localidad, así como el tipo de hábitat y
la disponibilidad de agua, fueron las siguientes:
Localidad de Jardines de la Reina, Trimestre FMA, 13 hembras y 12 machos, trimestre ASO, 22
hembras y 18 machos. Hábitat de mangle rojo con carencia total de agua dulce.
Localidad Area protegida Najasa, Trimestre FMA, 14 hembras y 12 machos, trimestre ASO, 20
hembras y 11 machos. Hábitat de Bosque semideciduo degradado, con disponibilidad de agua
(precipitación media anual: 1300 mm).
Localidad Parque nacional Guanahacabibes, Trimestre FMA 27 hembras y 20 machos, trimestre
ASO, 29 hembras y 19 machos. Hábitat de bosque semideciduo relativamente conservado, con
disponibilidad de agua limitada (precipitación media anual: 1350 mm).
Todos los ejemplares capturados fueron adultos según criterio de Smith y Berovides (1984).Las
variables utilizadas largo cabeza tronco (LCT), grosor de la médula (GM) y corteza renal (GC)
fueron expresadas en milímetro (mm) y medidas con una cinta métrica para el LCT y un pie de rey
para las variables GM y GC.
Para el análisis se utilizaron el índice renal propuesto por Yabe (1983), el cual mide el grosor de la
médula renal relativa al grosor del riñón y el índice propuesto por el autor, que mide el grosor de la
médula renal relativo al largo cabeza tronco (LCT).
Las variables obtenidas y los índices de reabsorción (IRR) elaborados se le efectuó la prueba de
distribución normal Kolmogorov-Smirnov, la prueba de homogeneidad de varianza de Levene y un
análisis de varianza bifactorial entre localidad en cada trimestre por separados además de un análisis
de varianza trifactorial considerando los efectos trimestres, sexos y localidad
IV.3. Resultados y discusión
Análisis dentro de una estación del año

89
El análisis de varianza del índice renal de Yabe por poblaciones (Tabla II) mostró diferencias entre
localidad y sexo, agrupando las poblaciones en cuatros subgrupos superpuestos entre si, ocupando
los valores intermedios las poblaciones de hábitat de bosque como Saetía y Guanahacabibes y
posiciones extremas las poblaciones de hábitat de manglar y manigua costera (Jardines, Sabana Y
Hatibonico) con los mayores valores promedios. La población de Najasa tuvo el menor valor
promedio. Las diferencias entre sexos fueron provocadas mayormente por el hecho de que en las
poblaciones de bosque, las hembras siempre presentaron mayores valores que los machos (74,44 vs
73,05), evidenciando un dimorfismo sexual para este índice, dependiente el tipo de hábitat.
Tabla II. Número de individuos por población (N), valores promedios (X), desviaciones estándar (S) y
coeficientes de variación (CV) para el índice renal de Yabe, en seis poblaciones de jutía conga
(Capromys pilorides). IC. Indice climático. Medias con índices diferentes difieren a p<0.05 por una
prueba de TuKey
Población N X S CV IC
Najasa 26 70.47ª 6.07 8.61 2
Saetía 28 71.66ab 4.51 6.29 1
Guanahacabibes 47 73.78b 4.03 5.46 3 Jardines 25 75.86bc 1.66 2.19 6
Sabana 23 75.97c 1.53 2.01 5
Hatibonico 29 78.66c 3.06 3.89 4
Las diferencias entre poblaciones, para el índice renal relativo al LCT (tabla III) mostró diferencias
estadísticamente significativas, diferenciando las poblaciones de manglar y manigua costera con
respecto a las de bosque. Las diferencias entre sexo no fueron significativas
Tabla III. Número de individuos por población (N), valores promedios (X), desviaciones estándar (S),
coeficientes de variación (CV) e índice climático (IC) para el índice renal relativo al lago cabeza
tronco, en seis poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes
difieren a p<0.05 por una prueba de TuKey
Población N X S CV IC
Najasa 28 23.53ª 2.93 12.45 1
Saetía 26 27.07b 5.10 18.84 2
Guanahacabibes 47 27.61b 4.49 16.26 3 Jardines 29 29.14bc 2.89 9.92 4
Sabana 23 30.72cd 3.57 11.62 5

90
Hatibonico 25 32.53d 2.34 7.19 6
Los valores de la regresión para cada uno de los índices y coeficientes de variación (Tabla IV) en el
índice climático, mostraron un grado de asociación lineal elevado para el índice renal relativo al
LCT, con resultados estadísticamente significativos y un incremento de 1.64 del índice renal por
cada unidad que aumenta el rango del índice climático. A pesar de que el índice de Yabe no reflejo
resultados estadísticamente significativos, si mostró un valor alto de asociación lineal con un
incremento de 1.21 por unidad de aumento del rango del índice, mucho menor que el índice anterior.
El coeficiente de variación reflejó resultados estadísticamente significativos para la regresión en el
índice renal de Yabe y no significativo estadísticamente para el índice renal relativo al LCT. En
ambos casos la magnitud de incremento de la variable dependiente (coeficientes de variación) tuvo
valores negativos en los dos índices y mayor efecto modular en el índice renal relativo al largo.
Tabla IV. Regresión (b), coeficiente de determinación (R2), F del ANOVA y error de la regresión (Sb) para
los valores promedios y el coeficiente de variación de un índice climático en seis poblaciones de jutía
conga (Capromys pilorides), para los índices renales relativo al grosor del riñón (Yabe) y relativo al
largo cabeza tronco (IRRL). *p<0.05; **p<0.01
Valores Promedios
Índices F R2 % b Sb
Yabe 5.03 ns 55.70 1.21 0.54 IRRL 99.84** 96.15 1.64 0.16
Coeficiente de Variación
Yabe 12.23* 75.36 -1.22 0.35
IRRL 3.54 ns 46.96 -1.55 0.82
El análisis de varianza para la comparación de las diferencias entre los valores promedios de la
corteza renal para las seis poblaciones estudiadas (Tabla V), reveló diferencias estadísticamente
significativas tanto entre población como entre sexo, lo cual muestra una variación entre las
poblaciones independiente del hábitat xérico, mésico o hídrico. Las hembras presentaron menores
valores que los machos (4.62 vs 4.82), lo que explica el mayor valor del índice de Yabe en las
primeras.

91
Tabla V. Valores promedios (X), desviación típica (S) y coeficiente de variación (CV) del grosor de la corteza
renal en seis poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes difieren
a p<0.05 por una prueba de TuKey
Población N X S CV
Hatibonico 29 3.80ª 0.78 20.53
Saetía 28 4.74b 0.79 16.67
Jardines 25 4.76b 0.56 11.76
Guanahacabibes 44 4.78b 0.90 18.83
Najasa 26 5.16b 1.01 19.57
Sabana 23 6.35c 0.78 12.28
Los valores promedios de la médula renal (Tabla VI) mostraron diferencias estadísticamente
significativas por localidad con un patrón de secuencia entre las poblaciones muy semejantes a la
secuencia conformada por los índices renales, con la diferencia de que el valor de la médula
absoluta no separa con la misma precisión las poblaciones de manglar y manigua costera con
respecto a las poblaciones de bosque, posiblemente por la falta de ponderación de su valor absoluto
,el cual pudiera estar influenciado por otras variables morfológicas como el tamaño y peso del riñón
y el peso y largo corporal entre otras variables (Díaz, 2001).
Tabla VI. Valores promedios (X), desviación típica (S) y coeficiente de variación (CV) del grosor de la
médula renal en seis poblaciones de jutía conga (Capromys pilorides). Medias con índices diferentes
difieren a p<0.05 por una prueba de TuKey
Población N X S CV
Saetía 28 12.02a 1.29 10.73
Guanahacabibes 44 13.47ab 2.01 14.92
Najasa 26 13.82bc 2.23 16.34 Hatibonico 29 13.84bc 1.25 9.03
Jardines 25 14.82bc 1.36 9.18
Sabana 23 15.23c 1.45 9.52
La tabla VII muestra los valores de regresión (b), F del ANOVA, coeficiente de determinación (R2)
y el error típico (Sb.) de la médula renal (variable dependiente) en el largo cabeza tronco (LCT
variable independiente) para seis poblaciones de jutía conga, en donde se evidenció valores de la
regresión estadísticamente significativos en todas las poblaciones. Resultados que señalan que a
mayor LCT mayor será la médula renal.
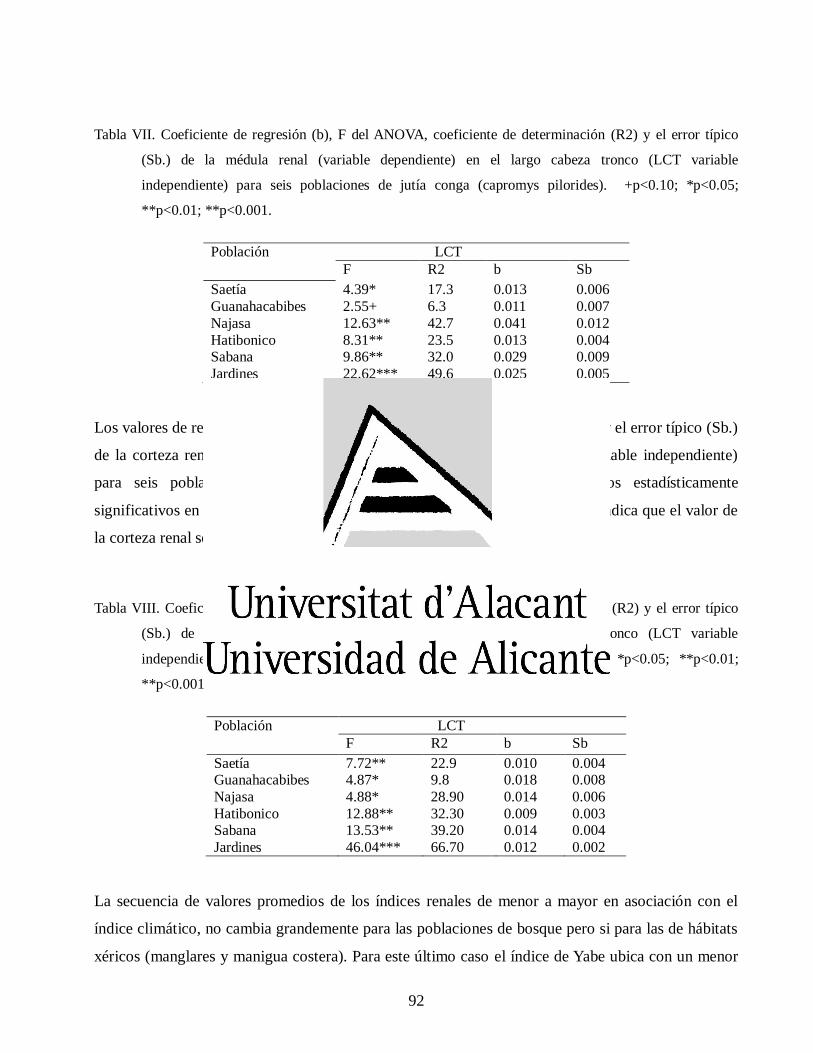
92
Tabla VII. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la médula renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) para seis poblaciones de jutía conga (capromys pilorides). +p<0.10; *p<0.05;
**p<0.01; **p<0.001.
Población LCT
F R2 b Sb
Saetía 4.39* 17.3 0.013 0.006
Guanahacabibes 2.55+ 6.3 0.011 0.007
Najasa 12.63** 42.7 0.041 0.012
Hatibonico 8.31** 23.5 0.013 0.004 Sabana 9.86** 32.0 0.029 0.009
Jardines 22.62*** 49.6 0.025 0.005
Los valores de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico (Sb.)
de la corteza renal (variable dependiente) en el largo cabeza tronco (LCT variable independiente)
para seis poblaciones de jutía conga (Tabla VIII), mostraron resultados estadísticamente
significativos en los valores de la regresión para todas las poblaciones, lo cual indica que el valor de
la corteza renal será mayor a medida que aumenta el LCT
Tabla VIII. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la corteza renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) en seis poblaciones de jutía conga (capromys pilorides). *p<0.05; **p<0.01;
**p<0.001
Población LCT
F R2 b Sb
Saetía 7.72** 22.9 0.010 0.004 Guanahacabibes 4.87* 9.8 0.018 0.008
Najasa 4.88* 28.90 0.014 0.006
Hatibonico 12.88** 32.30 0.009 0.003 Sabana 13.53** 39.20 0.014 0.004
Jardines 46.04*** 66.70 0.012 0.002
La secuencia de valores promedios de los índices renales de menor a mayor en asociación con el
índice climático, no cambia grandemente para las poblaciones de bosque pero si para las de hábitats
xéricos (manglares y manigua costera). Para este último caso el índice de Yabe ubica con un menor

93
valor a la población de Jardines y con el máximo valor a la de Guantánamo, ocurriendo lo contrario
para el índice relativo al LCT. A nuestro juicio el índice relativo al LCT está reflejando de manera
más eficiente el grado de disponibilidad de agua en estas localidades, ya que en Jardines hay
carencia absoluta de ella mientras que en Guantánamo los animales pueden obtenerla de la
vegetación xerofítica que la acumula. El alto índice de Yabe en Guntánamo podía estar influido por
los cambios que sufre la corteza del riñón, la cual se reduce en esta época de seca (Diaz, 2001).
Los resultados evidenciaron una relación entre los valores medios de los índices renales de las
diferentes poblaciones de jutía conga y la disponibilidad de agua de los ecosistemas donde habita, es
decir, que a hábitats más xéricos, mayores valores de estos índices renales. Estos resultados amplían
los obtenidos por Sánchez et. al (1992) y Berovides (2005), que trabajaron con solo cuatro
poblaciones de esta especie. Otros autores han obtenido resultados similares, pero con murciélagos
(Studier et. al, 1983) y roedores (Yabe, 1982, 1983; Ojeda et al; 1999; Al-Kathani, 2004). Los
valores de los índices renales representan variaciones adaptativas a la ecología local de carácter
fisiológico, que le permiten a esta especie desarrollarse en ambientes con diferente disponibilidad de
agua.
El análisis de la variación de los índices renales medidos a través del coeficiente de variación,
también mostró valor adaptativo al presentar una relación lineal de pendiente negativa con respecto
al rango de disponibilidad de agua. Este resultado era de esperar, ya que en un ambiente xérico, las
variaciones extremas del índice renal no se tolerarían y serían eliminadas por la selección natural
estabilizante.
Teniendo en cuenta los resultados anteriores opinamos que el índice IRRL puede ser utilizado como
un carácter adaptativo más en los estudios de poblaciones de jutía conga, para determinar unidades
de manejo para la conservación.
Según nuestros resultados, se puede concluir que los índices renales empleado (grosor de la medula
con respecto al grosor del riñón y grosor de la médula con respecto al LCT) miden la capacidad de
reabsorción diferencial de agua entre poblaciones con hábitats distintos, con una mayor exactitud y
correspondencia entre los valores promedio del índice y las características del hábitat para el índice
renal relativo al LCT, el cual solo refleja diferencias entre localidad. Las diferencias
estadísticamente significativas entre sexos para el índice renal de Yabe se encuentra influenciado por
las diferencias estadísticamente significativas de los valores de la corteza renal con independencia

94
del hábitat, la cual afecta también la magnitud de discriminar con exactitud las diferencias entre
localidades para este índice
Diferencias entre estaciones.
En las tablas IX y X se resume la estadística de los índices estudiados por localidad, trimestre
(estación) y sexo. Para el índice IRRY, el ANOVA trifactoral reveló diferencias estadísticamente
significativas entre localidades (F=8.08; p <0.001), trimestres (F=7.99; p<0.001)y la interacción
localidad por trimestre (F=3.75;p<0.05).Dada esta interacción, se analizaron los trimestres por
separados.
Para el índice IRRL sus valores fueron poco variables entre trimestres y sexo, solo hubo diferencias
estadísticamente significativas entre las localidades (F=26.76; p 0.05). La población de manglar
(Jardines) presentó valores mayores con respecto a las poblaciones de bosque. Aunque para este
índice no hubo interacción localidad por estación, también se analizaron los trimestres por
separados, a fin de comparar estos resultados con los del índice IRRY.
Tabla IX. Población, trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO), sexo
macho (M) y Hembra (H) así como las medias (X) y desviaciones típicas (S) para el índice de
reabsorción de Yabe (IRRY) en la jutía conga (Capromys pilorides).
Población Trimestre Sexo N X S
Jardines FMA
H 13 75.81 1.46
M 12 75.89 1.92
ASO H 22 73.92 2.32 M 18 73.40 2.53
Najasa FMA H 14 72.35 5.19
M 12 74.24 5.62
ASO H 20 74.88 2.86 M 11 72.98 2.7
Guanaha FMA H 27 74.40 4.27
M 20 72.90 3.58
ASO H 29 69.94 6.34
M 19 70.11 4.7
Tabla X. Población, trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO), sexo
macho (M) y Hembra (H) así como las medias (X) y desviaciones típicas (S) para el índice de
reabsorción relativo al largo cabeza tronco (IRRL) en la jutía conga (Capromys pilorides).
Población Trimestre Sexo N X S
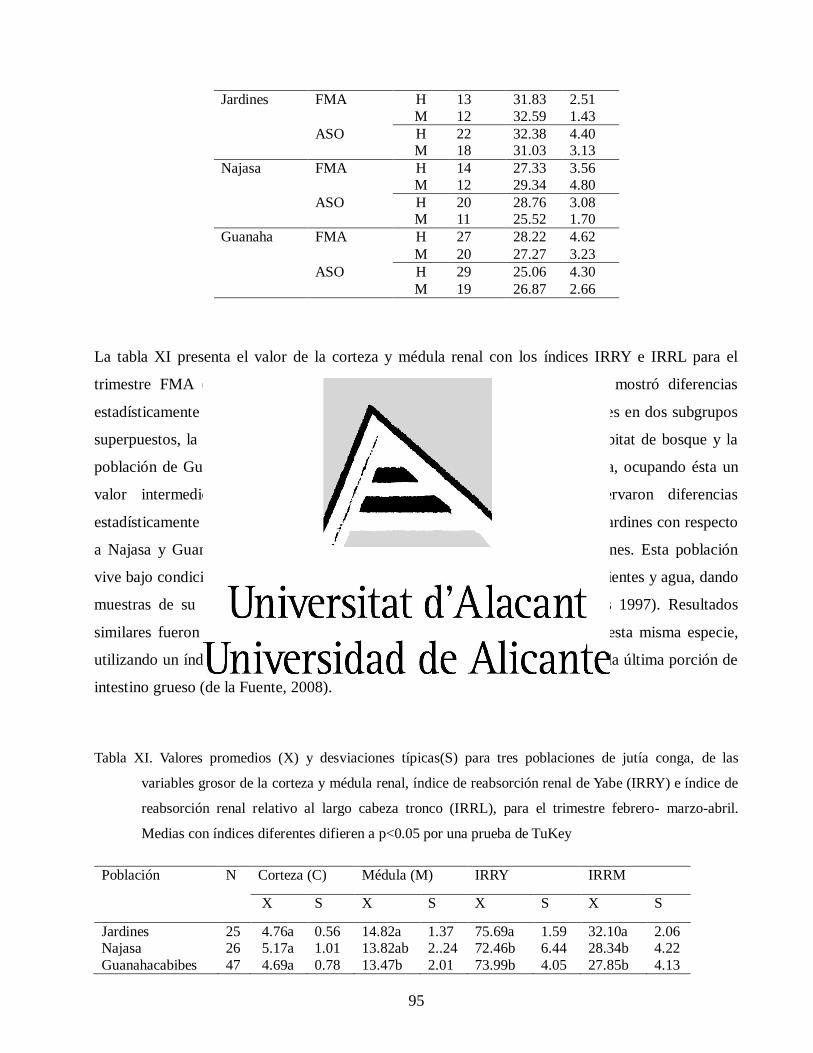
95
Jardines FMA
H 13 31.83 2.51
M 12 32.59 1.43
ASO H 22 32.38 4.40 M 18 31.03 3.13
Najasa FMA H 14 27.33 3.56
M 12 29.34 4.80
ASO H 20 28.76 3.08 M 11 25.52 1.70
Guanaha FMA H 27 28.22 4.62
M 20 27.27 3.23
ASO H 29 25.06 4.30
M 19 26.87 2.66
La tabla XI presenta el valor de la corteza y médula renal con los índices IRRY e IRRL para el
trimestre FMA (seca). El valor de la corteza para las tres localidades no mostró diferencias
estadísticamente significativas. El grosor de la médula diferenció las poblaciones en dos subgrupos
superpuestos, la localidad de Jardines de hábitat de manglar con Najasa de hábitat de bosque y la
población de Guanahacabibes de hábitat de bosque con la población de Najasa, ocupando ésta un
valor intermedio. Al analizar ambos índices de reabsorción se observaron diferencias
estadísticamente significativas y coincidentes, discriminando a la localidad de Jardines con respecto
a Najasa y Guanahacabibes, con máximos valores para la población de Jardines. Esta población
vive bajo condiciones más crítica respecto a la variedad y disponibilidad de nutrientes y agua, dando
muestras de su grado de adaptación al hábitat manglar (Berovides y Comas 1997). Resultados
similares fueron obtenidos al comparar poblaciones de manglar y bosque de esta misma especie,
utilizando un índice de absorción de agua, considerando la longitud relativa de la última porción de
intestino grueso (de la Fuente, 2008).
Tabla XI. Valores promedios (X) y desviaciones típicas(S) para tres poblaciones de jutía conga, de las
variables grosor de la corteza y médula renal, índice de reabsorción renal de Yabe (IRRY) e índice de
reabsorción renal relativo al largo cabeza tronco (IRRL), para el trimestre febrero- marzo-abril.
Medias con índices diferentes difieren a p<0.05 por una prueba de TuKey
Población N Corteza (C) Médula (M) IRRY IRRM
X S X S X S X S
Jardines 25 4.76a 0.56 14.82a 1.37 75.69a 1.59 32.10a 2.06 Najasa 26 5.17a 1.01 13.82ab 2..24 72.46b 6.44 28.34b 4.22
Guanahacabibes 47 4.69a 0.78 13.47b 2.01 73.99b 4.05 27.85b 4.13

96
Los valores de la corteza y médula renal con los índices de reabsorción IRRY e IRRL para el
trimestre ASO (lluvia) se exponen en la tabla XII. Se observan diferencias estadísticamente
significativas entre localidades para las cuatro variables estudiadas, siendo necesario destacar que
solo el grosor de la médula y el índice IRRL coincidieron en sus resultados, discriminando a la
población de Jardines de hábitat manglar con respecto a Najasa y Guanahacabibes con hábitat de
bosque, con un patrón consistente cuando se compara con los resultados del trimestre FMA. El
índice de reabsorción IRRY agrupó a una población de manglar (Jardines) con una de bosque
(Najasa), con falta de concordancia con respecto al trimestre FMA, debido a los cambios para el
grosor de la corteza renal de un trimestre al otro, estadísticamente significativos para las poblaciones
de Jardines y Najasa. Estos cambios afectan el IRRY, por ser la corteza una variable que se toma en
cuenta para la elaboración de este índice. El índice de reabsorción IRRL mantiene un mismo patrón
en los dos trimestres, con valores discriminativos y consistentes entre las poblaciones de manglar y
bosque.
Tabla XII. Valores promedios(X) y desviaciones típicas (S) para tres localidades en la jutía conga, de las
variables grosor de la corteza y médula renal, índice de reabsorción renal de Yabe (IRRY) e índice de
reabsorción renal relativo al largo cabeza tronco (IRRL), para el trimestre agosto- septiembre-
octubre. Medias con índices diferentes difieren a p<0.05 por una prueba de TuKey
Población N Corteza(C) Médula(M) IRRY IRRM
X S X S X S X S
Jardines 40 5.25a 0.48 14.83a 1.45 73.78a 2.36 31.77a 3.89
Najasa 31 4.62b 0.55 13.42b 1.35 74.31a 2.98 27.53b 3.07
Guanahacabibes 48 5.10a 0.62 12.48b 1.77 70.87b 4.00 25.65b 4.02
Los análisis por separado para cada trimestre confirmaron el patrón obtenido considerando ambos
trimestres (ANOVA trifactorial).Este análisis por el índice IRRY agrupó a las poblaciones de
Jardines +Najasa y Guanahacabibes + Najasa, con valores ligeramente mayores en el trimestre
FMA con respecto al trimestre ASO, la interacción localidad por estación se fundamenta por una
disminución del índice en las poblaciones de Jardines y Guanahacabibes en ambos sexos hacia el
trimestre ASO con respecto al trimestre FMA y un aumento de la variable para el trimestre ASO en

97
las hembras de Najasa influenciado fundamentalmente por los cambios estacionales en la corteza
renal. Díaz (2001) registra que durante la estación de seca la corteza se reduce en roedores
herbívoros de hábitat xerofítico en Argentina, cambio que se observa en Jardines y Guanahacabibes,
pero no en Najasa. En esta última población, con un hábitat más benigno, podrían estar actuando
otros factores distintos de la disponibilidad de agua, como la dieta o el tamaño, que también influyen
en la estructural renal (Díaz, 2001).
Los resultados obtenidos entre las comparaciones de las poblaciones en cuanto al grosor de
lacorteza, solo mostraron diferencias estadísticamente significativas por localidad (tabla XIII),
cuando se analizaron en su conjunto los dos trimestres, diferenciando las poblaciones de bosque de
las poblaciones de manglar.
Tabla XIII. Valores promedios (X), desviación típica (S), coeficiente de variación (CV) y pruebas t entre los
trimestres febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO) para el grosor de la
corteza renal en tres poblaciones de jutías conga (Capromys pilorides).
Población Trimestre N X S CV t
Jardines FMA 25 4.7 0.57 12.13 4.07***
ASO 40 5.24 0.49 9.35
Najasa FMA 26 5.22 1.01 19.35 2.43**
ASO 31 4.61 0.54 11.71
Guanahacabibes FMA 47 4.74 0.82 17.30 1.48 NS
ASO 48 5.1 0.73 14.31
Los coeficientes de variación de los índices renales (tabla XIV.) mostraron un aumento en el
trimestre ASO con respecto a FMA para las poblaciones de Guanahacabibes y Jardines, con una
disminución para el trimestre ASO en la población de Najasa. Los coeficientes de variación
mostraron una tendencia al aumento a partir de la población de jardines con hábitat de manglar,
seguido de las poblaciones de Guanahacabibes y Najasa de hábitat bosque, lo cual pudiera estar
relacionado con el accionar de la selección natural estabilizante que permite la supervivencia y
reproducción de organismos con rangos más amplios del índice de reabsorción para las poblaciones
de bosque, que mantienen mejores condiciones en cuanto a la disponibilidad del agua.

98
Tabla XIV. Valores de los coeficientes de variación de los índices renal de Yabe (IRRY) y renal relativo al
largo cabeza tronco (IRRL), en los trimestres febrero, marzo y abril (FMA) y agosto septiembre y
octubre (ASO) en tres poblaciones de jutías conga (Capromys pilorides).
Localidad IRRY IRRL
FMA ASO FMA ASO
Jardines 2.90 3.20 6.40 12.24
Najasa 8.89 4.01 14.89 11.15 Guanahacabibes 5.49 5.64 14.83 15.67
La tabla XV y XVI muestran los valores de regresión de la médula renal (variable dependiente) con
respecto al LCT (variable independiente) en los trimestres FMA y ASO respectivamente.
En el trimestre FMA se observó resultados estadísticamente significativos para la F del ANOVA en
todas las poblaciones, con un mayor incremento (b) de la variable dependiente por unidad de
incremento de la variable independiente para la población de Najasa.
Para el trimestre ASO se observaron resultados no significativos estadísticamente en todas las
poblaciones, lo cual refiere que el incremento de la variable dependiente por unidad de incremento
de la variable independiente no es significativo en este trimestre.
Tabla XV. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la médula renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) en tres poblaciones de jutía conga (Capromys pilorides), en los trimestres febrero,
marzo y abril (FMA).
Población LCT
F R2 b Sb
Jardines 22.62*** 49.6 0.025 0.005
Najasa 12.63** 42.7 0.041 0.012 Guanahacabibes 2.55+ 6.3 0.011 0.007
Tabla XVI. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la médula renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) en tres poblaciones de jutía conga (capromys pilorides) en los trimestres agosto,
septiembre y octubre (ASO).
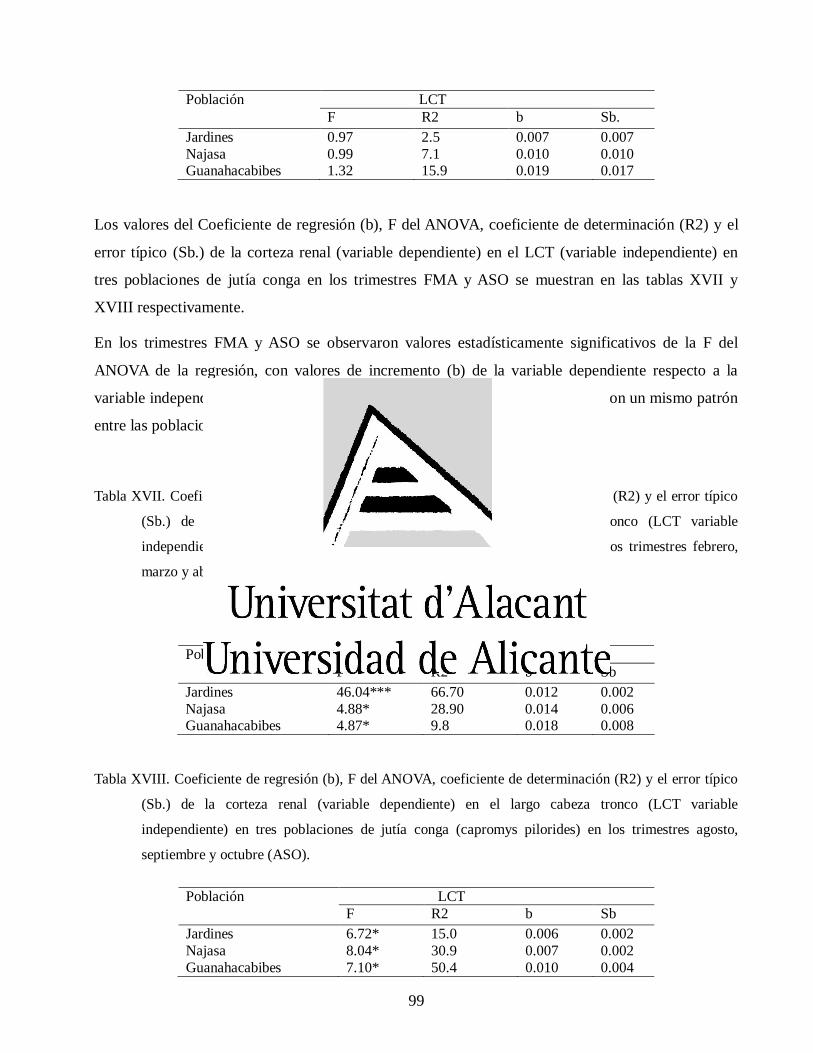
99
Población LCT
F R2 b Sb.
Jardines 0.97 2.5 0.007 0.007
Najasa 0.99 7.1 0.010 0.010 Guanahacabibes 1.32 15.9 0.019 0.017
Los valores del Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el
error típico (Sb.) de la corteza renal (variable dependiente) en el LCT (variable independiente) en
tres poblaciones de jutía conga en los trimestres FMA y ASO se muestran en las tablas XVII y
XVIII respectivamente.
En los trimestres FMA y ASO se observaron valores estadísticamente significativos de la F del
ANOVA de la regresión, con valores de incremento (b) de la variable dependiente respecto a la
variable independiente semejantes entre poblaciones dentro de cada trimestre y con un mismo patrón
entre las poblaciones en cada uno de los trimestres
Tabla XVII. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la corteza renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) en tres poblaciones de jutía conga (Capromys pilorides), en los trimestres febrero,
marzo y abril (FMA).
Población LCT
F R2 b Sb
Jardines 46.04*** 66.70 0.012 0.002
Najasa 4.88* 28.90 0.014 0.006 Guanahacabibes 4.87* 9.8 0.018 0.008
Tabla XVIII. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico
(Sb.) de la corteza renal (variable dependiente) en el largo cabeza tronco (LCT variable
independiente) en tres poblaciones de jutía conga (capromys pilorides) en los trimestres agosto,
septiembre y octubre (ASO).
Población LCT
F R2 b Sb
Jardines 6.72* 15.0 0.006 0.002
Najasa 8.04* 30.9 0.007 0.002
Guanahacabibes 7.10* 50.4 0.010 0.004

100
Nuestros resultados coinciden plenamente con otros autores., sobre todo con los de Al-kahtani et al.
(2004), que llevaron a cabo un análisis de la capacidad de retención de agua en varias especies de
roedores de Chile y Argentina. En el mismo compararon poblaciones de hábitat mésicos y áridos a
través de un índice renal, que tuvo en cuenta el grosor de la médula del riñón relativo a su volumen
y obtuvieron que las poblaciones de hábitat áridos presentaron mayores valores de este índice que
las de hábitat mésicos. Por su parte, Díaz et. al (2006), realizaron un estudio para determinar la
asociación entre el tipo de hábitat y la morfología del riñón en varias especies de roedores
histricomorfos sudamericanos. Para ello, midieron numerosas variables de los riñones y crearon
índices renales a partir de estas. Los resultados obtenidos sugieren que las especies de hábitat con
bajas precipitaciones, poseen riñones más grandes para impedir una mayor pérdida de agua.
En la jutía conga también se han realizado estudios de índices renales que comparan la capacidad de
reabsorción de agua entre poblaciones con diferentes tipos de hábitat. En poblaciones de jutía conga
de bosque y manglar, Sánchez et al. (1992), encontraron una estrecha relación entre un índice renal
de Yabe y las características del hábitat. Hallaron que las jutías que viven en los manglares
presentaron un grosor relativo de la médula renal mayor que las adaptadas a ambientes más
húmedos, como bosques secundarios y montanos. Resultados similares obtuvo Berovides (2005)
para hábitat de bosque y manigua costera.
En conclusión, el índice renal relativo al largo (IRRL) contribuye a diferenciar las poblaciones de
hábitat manglar de las poblaciones de hábitat bosque con un alto valor discriminativo y consistente
independiente del efecto estacional, resultados que no se evidenciaron en el índice de Yabe ni en el
grosor de la médula renal como variable absoluta.

101
CAPÍTULO V. Comparación de un índice de longitud intestinal en poblaciones de
jutía conga Capromys pilorides (Rodentia: Capromydae) de diferentes
hábitats.
V.1. Introducción
Los ecosistemas en que hábitat la jutía conga presentan diferencias entre si de acuerdo a la
formación vegetal y la disponibilidad de agua entre otros factores bióticos y abióticos, siendo
necesario establecer una comparación de acuerdo al nivel xérico que nos permita diferenciar el
grado de adaptación a la disponibilidad de agua. Yabe (1983) propone un índice renal basado en la
relación entre el grosor de la médula renal y el grosor total del riñón. Este índice se fundamenta en
la función fisiológica de las asas de Henle ubicadas en la médula renal, que se encarga de reabsorber
el agua para concentrar la orina y así mantener un equilibrio hídrico necesario de acuerdo a la
disponibilidad de agua (Smith-Nielsen, 1997). Estudios realizados en varias especies de caviomorfos
no han encontrado asociación entre este índice y la disponibilidad de agua (Díaz et al., 2006). Si
embargo para la jutía conga si se encontró esta asociación (Sánchez et al., 1992; Berovides, 2005).

102
No obstante, se hace necesario elaborar un índice que refleje el mecanismo de absorción
de agua utilizando para ello un índice derivado de la longitud de la última porción del intestino
grueso, tomando en cuenta la importante función fisiológica que desempeña esta en la absorción de
agua proveniente de la ingestión directa o indirecta a través de los alimentos (Smith-Nielsen, 1997).
Estudios realizados en mamíferos para el largo del intestino grueso han reflejado cambios
estacionales relacionados con la dieta, principalmente asociado con un incremento del ciego (Díaz et
al., 2006)
Pocos son los estudios relacionados con los cambios estacionales asociados a variables
morfofisiológicas en roedores en el mundo y ninguno se ha llevado a cabo en los roedores
endémicos de Cuba.
Por todo lo anteriormente planteado el objetivo de este trabajo fue establecer una comparación entre
poblaciones de jutia conga de hábitat diferentes en cuanto a su disponibilidad de agua, mediante un
índice de absorción derivado de la longitud de la última porción del intestino grueso.
V.2. Materiales y métodos
Análisis dentro de una estación del año. En la tabla I se muestran las características climáticas y de
vegetación de los hábitats de seis poblaciones estudiadas, según Díaz Cisneros (1989) para las
variables climáticas y Capote y Berazain (1995) para las formaciones vegetales. Así como el índice
climático de Martonne por medio de la fórmula:
IC=Precipitación media anual/Temperatura media anual +10 0 C
Este índice está asociado a la productividad vegetal de los hábitats y a la tasa metabólica basal
(independiente del peso) en los individuos de la población (Bozinovic et al., 2009).
No se tomaron los valores del índice como tal, sino que estos fueron convertidos a rango (Al-
Kahtani et al., 2004), donde el rango 1 representó alta disponibilidad de agua dulce y el 6 poca o
nula disponibilidad. Estos rangos para cada población fueron los siguientes: Saetía 1,
Guanahacabibes 2, Najasa 3, Hatibonico 4, Sabana 5, Jardines 6.

103
Tabla I. Localidad, formación vegetal, precipitación, temperatura promedio anual e índice climático/ rango en
seis poblaciones de jutía conga (Capromys pilorides).
Localidad Población Formación vegetal Temperatura
(oC)
Precipitación
(mm)
Indice
Climático
/rango
Parque nacional
Jardines de la Reina
Jardines Manglar (mangle
rojo)
27 1000 -/6
Refugio de Fauna
Las Picuas
Sabana Manglares y
manigua costera
26.5 1000 -/5
Reserva Ecológica
Hatibónico
Hatibonico Matoraral
xeromorfo
28 600 15.79/4
Area Protegida de
Recursos Manejados La
Belén
Najasa Bosque
semideciduo mesófilo(BSM)
secundario
25 1300 37.14/3
Parque Nacional Guanahaca-bibes
Guanahacabibes BSM y yanal 25.8 1350 37.7/2
Paisaje protegido
Saetía
Saetía BSM 26 1400 38.89/1
Los ejemplares capturados fueron medidos en milímetros (mm) para el largo cabeza tronco y para la
longitud de la última porción del intestino grueso (desde su inserción del ciego al ano). Las muestras
fueron obtenidas en el segundo trimestre de la estación seca en Cuba (febrero, marzo y abril) con un
total de 174 de animales (92 hembras y 82 machos). Todos los individuos fueron adultos según
criterio de tamaño, peso y desarrollo de los genitales (Smith y Berovides 1984).
El índice utilizado (índice de absorción de agua) fue calculado como la longitud de la última porción
del intestino grueso relativo al largo cabeza tronco, para eliminar el posible efecto del tamaño
(Schauenberg, 1977)
Se calcularán las medias, desviaciones típicas y coeficientes de variación para los valores absolutos
y su índice derivado, se determinó el tipo de distribución por la prueba de Kolmogorov-Smirnov y la
prueba de homogeneidad de varianza de Levene comparándose mediante un análisis de varianza
bifactorial, considerando los efectos localidad y sexo y la interacción de estos factores. Se realizó
también un análisis de regresión lineal simple entre la longitud cabeza tronco como variable
independiente y la longitud última porción del intestino grueso como variable dependiente para la
muestra total y para cada una de las poblaciones estudiadas.
Análisis entre estaciones del año.

104
Las muestras de jutía conga fueron obtenidas en el segundo trimestre de la estación de seca febrero,
marzo y abril (FMA) y en el segundo trimestre de la estación de lluvia agosto septiembre y octubre
(ASO). Se capturaron un total de 227 ejemplares, de las cuales 128 pertenecieron al trimestre FMA
y 119 al trimestre ASO (Tabla II).
Tabla II. Tipos de ecosistema y muestra poblacional por sexo, en los trimestres febrero, marzo y abril (FMA)
y agosto septiembre y octubre (ASO) en tres poblaciones de jutía conga (Capromys pilorides)
Población N Ecosistema
FMA ASO
H M H M
Jardines 27 16 23 18 Manglar Najasa 18 20 18 12 Bosque semideciduo
Guanahacabibes 27 20 29 19 Bosque semideciduo
Los individuos analizados fueron todos adultos según criterio de Smith y Berovides (1984) y
medidos en milímetro (mm) para el largo cabeza tronco y la longitud última porción del intestino
grueso (desde su inserción del ciego al ano). El índice utilizado (índice de absorción de agua) fue
calculado como la longitud de la última porción del intestino grueso relativo al largo cabeza tronco
(LIU/LCT), para eliminar el posible efecto del tamaño (Schauenberg, 1977; Sánchez, 1992).
Las variables utilizadas fueron analizadas por la prueba de normalidad de Kolmogorov –Smirnov y
la prueba de homogeneidad de varianza de Levene. Se calcularán las medias, y coeficientes de
variación para los valores absolutos y su índice derivado, comparándose mediante un análisis de
varianza (ANOVA) trifactorial, considerando los efectos localidad, sexo y trimestre del año (como
estaciones del año), así como un ANOVA bifactorial con los efectos sexo y localidad para
determinar las diferencias entre las poblaciones dentro de cada trimestre, así como la prueba F de
Snedecor para la homogenidad de varianza.
V.3. Resultados y discusión
Análisis dentro de una estación del año.
La tabla III muestra las localidades, hábito, muestras, F del ANOVA y los coeficientes de regresión
en seis poblaciones de jutía conga, donde se mostró que:

105
La correlación lineal simple del largo cabeza tronco con el largo de la última porción del intestino
grueso y su regresión, no mostraron asociación (r = 0.072; b= -0.638; p> 0.05) lo cual nos indica que
esta longitud no esta influenciada por el largo cabeza tronco, tomando en cuenta los valores totales.
Sin embargo, por poblaciones las regresiones resultaron estadísticamente significativas para las
poblaciones de bosque (Najasa, Guanahacabibes y Saetía). Al parecer las condiciones de hábitat
xérico impiden el establecimiento de una asociación entre el largo cabeza tronco y el largo intestinal,
pero este si puede establecerse en los hábitats de bosque, donde además el valor de la regresión va
aumentando en dependencia de los hábitos de la especie
Tabla III. Hábito, coeficiente de regresión (b), F del ANOVA y error típico (Sb) del largo de la última porción
del intestino grueso (variable dependiente) en el largo cabeza tronco (variable independiente), para
seis poblaciones de jutía conga (Capromys pilorides).
Localidad Hábito N F ANOVA b Error típico
Jardines SM 43 3.83 NS 1.23 0.63
Sabana ST 20 4.47 NS 4.12 1.94
Hatibonico TE 21 1.25 NS 1.47 1.31 Najasa S 19 18.11 *** 2.80 0.66
Guanahacabibes ST 47 5.41 * 3.42 1.47
Saetía S 24 3.67 * 4.65 2.43
SM= Semiarborícola de mangle * p< 0.05
ST= Semiterrestre ***p<0.001
TE= Terrestre
S= Semiarborícola
Los coeficientes de regresión de la longitud última porción del intestino grueso (variable
dependiente) en el peso corporal (variable independiente), por localidades, hábito, muestras y F del
ANOVA en seis poblaciones de jutía conga, se muestran en la tabla IV. Se destaca el alto valor e la
regresión en Jardines, única población con carencia total de agua dulce.
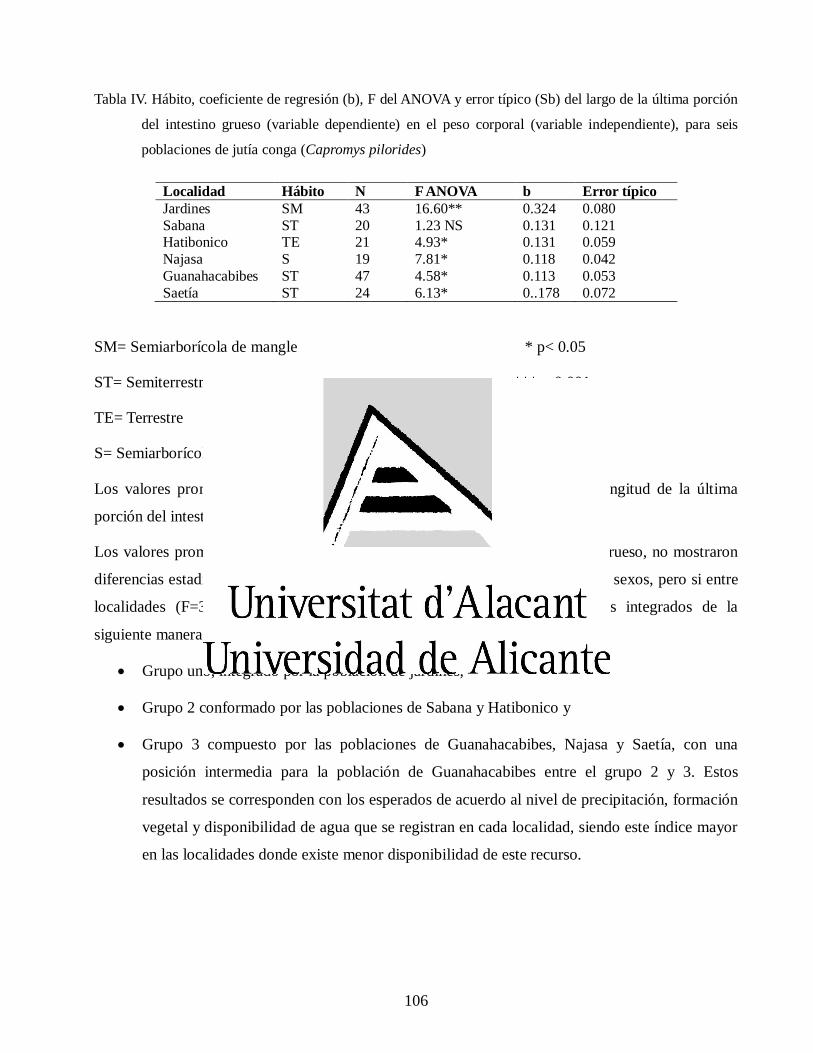
106
Tabla IV. Hábito, coeficiente de regresión (b), F del ANOVA y error típico (Sb) del largo de la última porción
del intestino grueso (variable dependiente) en el peso corporal (variable independiente), para seis
poblaciones de jutía conga (Capromys pilorides)
Localidad Hábito N F ANOVA b Error típico
Jardines SM 43 16.60** 0.324 0.080
Sabana ST 20 1.23 NS 0.131 0.121 Hatibonico TE 21 4.93* 0.131 0.059
Najasa S 19 7.81* 0.118 0.042
Guanahacabibes ST 47 4.58* 0.113 0.053
Saetía ST 24 6.13* 0..178 0.072
SM= Semiarborícola de mangle * p< 0.05
ST= Semiterrestre ***p<0.001
TE= Terrestre
S= Semiarborícola
Los valores promedios, coeficientes de variación y desviación típica de la longitud de la última
porción del intestino grueso se muestran en la tabla V.
Los valores promedios absolutos obtenidos de la última porción del intestino grueso, no mostraron
diferencias estadísticamente significativas entre sexos ni interacción localidad x sexos, pero si entre
localidades (F=37.019; p<0.001), separando las poblaciones en tres grupos integrados de la
siguiente manera:
Grupo uno, integrado por la población de jardines,
Grupo 2 conformado por las poblaciones de Sabana y Hatibonico y
Grupo 3 compuesto por las poblaciones de Guanahacabibes, Najasa y Saetía, con una
posición intermedia para la población de Guanahacabibes entre el grupo 2 y 3. Estos
resultados se corresponden con los esperados de acuerdo al nivel de precipitación, formación
vegetal y disponibilidad de agua que se registran en cada localidad, siendo este índice mayor
en las localidades donde existe menor disponibilidad de este recurso.

107
Tabla V. Valores promedio (X), desviación típica (S) y coeficiente de variación (CV) del largo de la última
porción del intestino grueso, en seis poblaciones de jutía conga (capromys pilorides). Medias con
índices diferentes difieren a p<0.05 por una prueba de TuKey
Población N X S CV
Jardines
(Mangle rojo)
43 3149.53 a
324.45 10.30
Sabana(Manglar y manigua) 20 2616.50 b
323.40 12.36
Hatibonico
(V. xerofítica)
21 2556.19 b
251.88 9.85
Najasa (BSM)
19 2213.74 c
181.40 8.19
Guanahacabibes
(BSM y manglar)
47 2311.87 bc
412.86 17.86
Saetía
(BSM)
24 2197.08 c
308.13 14.02
La tabla VI refleja los valores promedios, desviación típica y coeficiente de variación del largo de la
última porción del intestino grueso relativo al largo cabeza tronco, en seis poblaciones de jutía conga
donde se observa que:
Los valores del índice de absorción no mostraron diferencias estadísticamente significativas entre
sexos, pero si entre localidad (F= 72.25; p<0.001) e interacción entre localidad x sexo (F= 2.36;
p<0.05). Este índice de absorción, con resultados semejantes a los valores absolutos de la longitud
última del intestino grueso, diferencia aun más los tres grupos de poblaciones formados, sin posición
intermedia desde el punto de vista estadístico para la población de Guanahacabibes. Por todo lo
antes expuesto, opinamos que el uso de este índice es mejor que el de los valores absolutos.
Nuestros resultados con respectos al índice, concuerdan con lo esperado fisiológicamente, ya que el
desarrollo de mecanismos compensadores a las condiciones adversas entre otras adaptaciones,
consistiría en aprovechar la máxima cantidad de agua posible de los alimentos, cuya extracción
fundamentalmente se realiza en los mamíferos en la última porción del intestino grueso, sitio de
regulación del balance hídrico, absorbiendo el agua de las heces fecales (Smidt-Nielsen, 1997), pero
debe de tenerse en cuenta la existencia de coprofagía en la especie bajo estudio y en algunas de sus
poblaciones, también la ingestión de plantas ricas en taninos, que afectan la longitud del tracto
intestinal (Hladik y Picq, 2001).

108
El análisis de la variabilidad del índice (medido por el coeficiente de variación) indicó que las
localidades de Sabana y Guanahacabibes poseen los mayores coeficientes de variación, lo que
pudiera estar asociado a que estas poblaciones experimentan los hábitas más heterogéneos en cuanto
a vegetación y disponibilidad de agua, mientras que Jardines experimenta la menor variabilidad.
Estos resultados reflejan que:
En las poblaciones de Sabanay Guanahacabibes existen valores extremos más distantes del valor
central, debido a que estos no comprometen la supervivencia y reproducción de las poblaciones,
dada las características del hábitat y su abundancia relativa a la diponibilidad del agua, no obstante,
la permanencia relativamente cerca de un manglar en las poblaciones muestreadas de
Guanahacabibes, pudiera influenciar en que los coeficientes de variación de esta población sean
más dispersos que los coeficientes de variación de otras poblaciones.
La población de Jardines exhibió los coeficientes de variación menos dispersos, asociado a que
valores más distantes y muy inferiores al valor promedio pudieran comprometer la supervivencia y
reproducción de la población debido a que la disponibilidad relativa de agua es muy baja.
Tabla VI. Valores promedio (X), desviación típica (S) y coeficiente de variación (CV) del largo de la última
porción del intestino grueso relativo al largo cabeza tronco, en seis poblaciones de jutía conga
(Capromys pilorides). Medias con índices diferentes difieren a p<0.05 por una prueba de TuKey.
Población N X S CV
Jardines (Mangle rojo) 43 6.87 a
0.44 6.40
Sabana(Manglar y manigua) 20 5.55 b
0.84 15.13
Hatibonico (V. xerofítica) 21 5.33 b
0.59 11.07
Najasa (BSM) 19 4.66 c
0.31 6.65
Guanahacabibes (BSM y manglar) 47 4.73 c
0.77 16.28
Saetía (BSM) 24 4.24 c
0.49 11.56

109
La Tabla VII muestra el coeficiente de regresión (b), F del ANOVA, coeficiente de determinación
(R2) y el error típico para las variables última porción del intestino grueso absoluto y relativo al
largo cabeza tronco como variables dependientes, en relación a un índice climático (variable
independiente) en seis poblaciones de jutía conga, donde se observa:
Valores estadísticamente significativos en las dos variables.
Mayor porcentaje de relación lineal (madido por R2) de la variable 2
Tabla VII. Valores de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error típico de b
(Sb), de la última porción del intestino grueso absoluto (variable 1) y relativo al largo cabeza
tronco(variable 2) como variable dependiente, en relación a un índice climático (variable
independiente), en seis poblaciones de jutía conga (Capromys pilorides)
Variable F R2 b Sb
Variable 1 15.93* 79.9 171.96 43.08
Variable 2 26.53** 86.9 0.47 0.09
**P<0.01
*p<0.05
Tanto para el valor absoluto del largo de la última porción del intestino grueso, como para el índice
derivado del mismo, se encontraron diferencias estadísticamente significativas entre los tres grupos
de poblaciones de la jutía conga, un primer grupo lo formó la población de Jardines, un segundo
estuvo compuesto por la población de Sabana, Hatibonico y Guanahacabibes y un tercer grupo que
incluyó a las poblaciones de Najasa y Saetía. Los resultados obtenidos muestran una estrecha
relación entre la longitud del intestino (valores absolutos o como índices relativos) con la carencia
de agua en los hábitats donde viven cada población, lo cual índica la adaptación de la especie a la
disponibilidad de este recurso
Análisis entre estaciones del año
Se analizan primero las diferencias en media y variabilidad entre poblaciones dentro de cada
estación Los valores promedios y desviación típica del largo absoluto de la última porción del
110
intestino grueso para los trimestres FMA Y ASO, en tres poblaciones de jutía conga se muestran en
la tabla VIII. Se observaron diferencias estadísticamente significativas entre poblaciones dentro del
trimestre FMA (resultado presentado en el capítulo anterior), donde las poblaciones de bosques
semideciduos presentaron igualdad estadística entre ellas con valores promedios más bajos y con
diferencias estadísticamente significativas con respecto a la población de Jardines. Sin embargo, no
hubo diferencias estadísticamente significativas entre poblaciones para ASO, resultado similar al
obtenido para los cambios estacionales de los caracteres morfométricos, lo que puede explicarse por
las mismas causas.
Se registra también un coeficiente de variación mucho menor en Jardines que en las otras dos
poblaciones de bosque. Resultado similar al obtenido con los índices renales y que se explica por las
mismas causas.
La tabla IX refleja los valores promedios y desviaciones típicas del índice relativo del intestino en
los trimestres FMA y ASO, en tres poblaciones de jutía conga. Los resultados mostraron un patrón
semejante en los trimestres estudiados (a diferencia del valor absoluto, que solo detecto diferencias
en FMA), con las ya conocidas diferencias estadísticamente significativas, entre las poblaciones de
bosque semideciduo con respecto a la población de hábitat manglar.
Tabla VIII. Valores promedios (X), muestras (N) y coeficiente de variación (CV) del largo última porción del
intestino grueso (valor absoluto), en tres poblaciones de jutía conga (Capromys pilorides) en los
meses de febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO). Medias y CV
con letras diferentes, difieren a p<0.05 entre poblaciones dentro trimestres, por las pruebas
de Tukey y Levene respectivamente
Población N X CV
FMA ASO Total FMA ASO Total
Jardines 84 3149.53 b 3192.14 b 3170.33 10.30a 6.85a 8.74
Najasa 68 2213.73 a 2306.00 b 2270.22 8.19a 12.56a 11.24
Guanahacabibes 95 2267.08 a 2417.14 b 2301.53 16.50b 13.33a 15.87

111
Tabla IX. Valores promedios (X), muestras (N) y desviación típica (DS) del largo última porción del intestino
grueso (relativo) en tres poblaciones de jutía conga (Capromys pilorides) en los meses de febrero,
marzo y abril (FMA) y agosto septiembre y octubre (ASO). Medias y CV con letras diferentes,
difieren a p<0.05 entre poblaciones dentro trimestres, por las pruebas de Tukey y Levene
respectivamente
Población N X CV
FMA ASO Total FMA ASO Total
Jardines 84 6.87 b 6.83 b 6.85 6.40a 0.44a 4.66
Najasa 68 4.66 a 4.69 a 4.68 6.78a 9.19b 8.27
Guanahacabibes 95 4.64 a 4.94 a 4.71 15,52b 12.65c 14.98
En cuanto a la variabilidad, el patrón encontrado en FMA, de menor CV en Jardines para las
longitudes absolutas y relativas en comparación con las poblaciones de bosques, se mantiene por
igual en el semestre ASO, indicando las importancias de mantener esta baja variabilidad, aún frente
a los cambios estacionales.
Al analizar las diferencias de valores promedios entre estaciones dentro de cada población por
pruebas de t, encontramos que para el largo absoluto del intestino, este tiende a valores menores y
estadístcamente significativos en el trimestre ASO en todas las poblaciones, pero las diferencias
entre trimestres (valores medios de ASO- FMA) son mas marcadas en las poblaciones de bosques
(42.61 para Jardines vs 92.27 y 150.06 para Najasa y Guanahacabibes respectivamente).Sin
embargo, al relativizar el largo del intestino con relación al tamaño, no se dan diferencias
estadísticas entre estaciones en ninguna población. Esto sugiere que las diferencias entre
poblaciones para esta variable en términos relativos, se corresponde con una adaptación local estable
en el genotipo y no como un producto de la norma de reacción de éste.
Las Tablas X y XI muestran los valores del coeficiente de regresión de la longitud última del
intestino grueso en valores absolutos y relativos (variable dependiente) con respecto al largo cabeza
tronco y el peso corporal (variables independientes) respectivamente. Se observó que para el
incremento tanto de los valores absolutos como relativos, con respecto al aumento en una unidad de
las variables independientes LCT y peso corporal, se obtuvieron los máximos valores en la

112
población de Jardines, en ambos trimestres, pero las diferencias entre trimestres dentro de cada
población, para las longitudes absolutas presentaron mayores diferencias.
Las poblaciones de bosque semideciduo (Najasa y Guanahacabibes) presentaron valores semejantes
de incrementos entre las poblaciones para los trimestres FMA y ASO.
Estos resultados manifestaron constancia entre las poblaciones con similitud en cuanto a la
disponibilidad de agua, con un patrón muy semejante dentro de todas las poblaciones y trimestres,
independientemente del empleo de la variable LCT o peso corporal como variables independientes.
Tabla X. Coeficiente de regresión (b) de tres poblaciones de jutía conga (Capromys pilorides) en los meses de
febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO) para la última porción del
intestino en relación al largo cabeza tronco.
Población N b.
FMA ASO
Jardines 84 7.12*** 6.44***
Najasa 68 2.80** 4.35**
Guanahacabibes 95 2.92* 3.26 NS
Tabla XI. Coeficiente de regresión (b) de tres poblaciones de jutía conga (Capromys pilorides) en los meses
de febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO) para la última porción del
intestino en relación al peso corporal.
Población N b.
FMA ASO
Jardines 84 0.324*** 0.311***
Najasa 68 0.118* 0.188***
Guanahacabibes 95 0.113* 0.137 NS

113
Se vuelven a mostrar las diferencias en variabilidad del largo absoluto y relativo del intestino
respectivamente (Tablas XII y XIII), pero ahora referido a las diferencias entre trimestres dentro de
cada población. Solo para Jardines estas diferencias fueron estadísticamente significativas. Este
resultado pudiera estar asociado a cambios drásticos en la calidad de los alimentos en relación a las
estaciones de seca y lluvia, (se conoce que los alimentos de más baja calidad pueden aumentar el
tamaño de la cámara digestiva, situación que no se daría para los bosques.
Tabla XII. Coeficiente de variación (CV) de tres poblaciones de jutía conga (Capromys pilorides) en los
meses de febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO) para el valor absoluto
de la última porción del intestino grueso.
Población N CV F
FMA ASO
Jardines 84 10.30 6.86 2.20 *
Najasa 68 8.19 12.56 2.55 NS
Guanahacabibes 95 16.50 13.33 1.34 NS
F de los cocientes de varianza para las diferencias entre estaciones dentro de cada localidad
Tabla XIII. Coeficiente de variación (CV) de tres poblaciones de jutía conga (Capromys pilorides) en los
meses de febrero, marzo y abril (FMA) y agosto septiembre y octubre (ASO) para el índice del
intestino
Población N CV F
FMA ASO
Jardines 84 6.4 0.44 211 *
Najasa 68 6.87 9.17 1.93 NS
Guanahacabibes 95 15.52 12.55 1.35 NS
F de los cocientes de varianza para las diferencias entre estaciones dentro de cada localidad

114
No hemos encontrado en la literatura extranjera trabajos que estudien los cambios de tamaño del
intestino grueso en relación con su función de absorción de agua, que es lo que se analiza en esta
tesis. Además, los resultados antes señalados, acerca de la flexibilidad de esta variable frente a
varios factores ambientales, para el caso del intestino grueso, solo se refieren al colon y ciego y
excluyen la última porción o recto. Por consiguiente, dichos resultados deben ser tomados con
cautela, porque podrían ser o no válidos para esta última porción del intestino grueso, que es la mas
involucrada en la función antes mencionada
CAPÍTULO VI. Análisis comparativo de tres índices nutricionales en poblaciones
de jutía conga (Capromys pilorides).
VI.2. Materiales y métodos
Análisis para una sola estación del año.
Se analizaron poblaciones de jutía conga pertenecientes a sietes localidades del Sistema Nacional de
Areas Protegidas, con una muestra total de 252 ejemplares capturados todos en el trimestre febrero,

115
marzo y abril, para eliminar la variación estacional reportada para las variables que aquí se estudian
(de la Fuente y Berovides, 2009).
La tabla I releja las localidades de las poblaciones con sus categorías de manejo, tipo de hábitat y
las muestras por localidades y sexo de las siete poblaciones estudiadas.
Tabla I. Localidades de las poblaciones con sus categorías de manejo, muestras por localidad y sexo y tipo de
hábitat, en siete poblaciones de jutía conga (Capromys pilorides)
Localidad y categoría de manejo Sexo N Hábitat
Area protegida de recursos manejado
Jardines de la Reina
H 27 Manglar (Rizophora mangle)
M 16
Are protegida de recursos manejados
Archipiélago Sabana Camaguey
H 19 Manglar y
Manigua costera M 13
Reserva ecológica Hatibonico H 20 Manigua costera
M 33
Area protegida de recursos manejados
Parque Nacional de Guanahacabibes
H 27 Bosque semideciduo
M 20
Area protegida de recursos manejados
Najasa
H 18 Bosque semideciduo
M 20
Refugio de Fauna
Saetía
H 16 Bosque semideciduo
M 14
Area protegida de recursos manejados
Isla sur
H 14 Bosque semideciduo
M 10
Los ejemplares estudiados fueron todos adultos( Smith y Berovides, 1984); cada uno de los
animales capturados fueron sexados y se le determinó el largo cabeza tronco (LCT) y largo del pie
izquierda con uña (Pi) en milímetro (mm), así como el peso corporal ( PC) y peso del hígado ( PH)
en gramos (gr).Con el peso del hígado se confeccionó el índice nutricional de Gosling (1980), que

116
relaciona PH/Pi al cubo; un segundo índice que relaciona el peso del hígado con el peso corporal
(PH/PC) y un índice nutricional propuesto por los autores (Gosling modificado), utilizando la
relación PH/LCT al cubo. A las variables absolutas e índices nutricionales se les efectuó la prueba de
normalidad de Kolmogorov- Smirnov. Los valores promedios de los índices de nutrición se
compararon entre poblaciones mediante un análisis de varianza (ANOVA) bifactorial con los efectos
población y sexo y los valores promedios de los coeficientes de variación por un análisis de varianza
no paramétrico de Kruskall-Wallis
Las localidades de las poblaciones se organizaron a criterio de un índice climático (IC) o índice de
Martonne (Bozinovic et al., 2009), que relaciona la precipitación media anual con la temperatura
medial anual en cada localidad, según la fórmula:
IC=Precipitación media anual/Temperatura media anual +10 0 C
y que esta asociado a su vez a la producción primaria de los hábitats. Los valoras de este índice
fueron convertidos en rangos (Al-Kahtani et al., 2004), donde el rango 1 representó la más baja
disponibilidad de nutrientes (bajos valores del índice) y el siete la más alta disponibilidad. Estos
rangos para cada población fueron los siguientes: Jardines 1, Sabana 2, Hatibonico 3, Najasa 4,
Guanahacabibes 5, Saetía 6 e Isla 7.
A los valores promedios de los índices nutricionales (variable dependiente) se le determinó el grado
de dependencia con respecto a los rangos (variable independiente) mediante una regresión lineal
simple. Para cada uno de los análisis estadísticos se utilizo el sistema SPSS11.5.
Análisis para las diferencias entre estaciones. Se estudiaron cuatros poblaciones de jutía conga, dos
pertenecientes a ecosistemas de bosques (Parque Nacional Guanahacabibes y Area Protegida
Najasa) y dos pertenecientes a ecosistemas de manglar (Refugio de Fauna Las Picúas, archipiélago
Sabana-Camaguey y Area Protegida Jardines de la Reina). Se capturaron un total de 301
ejemplares, con una muestra por localidad, sexo y trimestre del año de la siguiente manera:
Población de Guanahacabibes:
Trimestre FMA N=27 hembras y 20 machos
Trimestre ASO N= 29 hembras y 19 machos.
Población de Najasa:

117
Trimestre FMA N= 18 hembras y 20 machos
Trimestre ASO N= 18 hembras y 19 machos
Población de Sabana:
Trimestre FMA N= 19 hembras y 13 machos
Trimestre ASO N= 18 hembras y 12 machos.
Población de Jardines:
Trimestre FMA N= 27 hembras y 16 machos
Trimestre ASO N= 12 hembras y 14 machos.
El trimestre FMA se corresponde con la estación de seca en Cuba y el ASO con la de lluvia.
Todos los ejemplares analizados fueron adultos y se les determino el peso corporal (PC) y el peso
del hígado en gramos, así como el largo cabeza tronco (LCT) en mm. El índice nutricional se
confeccione dividiendo el peso del hígado entre LCT al cubo, según Gosling et al (1980)
modificado, sustituyendo el largo del pie por LCT, ya que este presenta menor variación en la jutía
conga y elimina el efecto del tamaño y el peso. Al peso del hígado (PH) y al índice analizado se les
determinó la distribución normal mediante la prueba de Kolmogorov-Smirnov y a la prueba de
homogeneidad de varianza de Levene, las variables se procesaron estadísticamente mediante un
análisis de varianza (ANOVA) trifactorial, considerando los efectos trimestres, población y sexo.
Las diferencias entre trimestres o sexos dentro de cada población se compararon por pruebas t. Se
calcularon además las regresiones lineales simples del peso del hígado en el peso corporal y el LCT.
Las poblaciones de Jardines y Najasa poseían datos de las actividades reproductivas de machos y
hembras durante los trimestres estudiados (Gutierrez et al., 1999) y dichos datos fueron asociados a
la dinámica del índice nutricional en esos trimestres, al considerar la gran dependencia de las
actividades reproductivas del factor nutricional.
Las variables reproductivas consideradas fueron:
DT: Descenso testicular
IR: Indice reproductivo (peso testículo/peso corporal)
HR: Hembras en reproducción (lactancia, gestación o ambas)
EA: Estadio embrionario A (el mas temprano)

118
VI.1. Introducción
La disponibilidad de nutrientes y la adaptación de los organismos dentro del ecosistema, se ve
reflejada, entre otros aspectos, en su estado nutricional, el cual ha sido estudiado por diversos
métodos, entre los que se encuentran el peso relativo del hígado (Gosling et al., 1980 en el
coipo;Smith y Berovides, 1984; Sánchez et al., 1992 y Berovides, 2005 en la jutía conga).Ello se
debe a que este órgano es quien dirige el destino de cada uno de los nutrientes según las demandas
fisiológicas en cada momento, como almacén de glucógeno, carbohidratos (la primera fuente de
energía de los organismos) y aminoácidos, entre otras funciones de importancia vital para los
organismos(Aundesirk y Aundesirk, 1996).
Una peculiaridad morfológica del hígado de la jutía conga, es que además de poseer varios lóbulos
como en las otras especies de jutías, cada lóbulo esta subdividido en lobulillos (Lora, 1895). El
índice de Gosling fue utilizado por Sánchez et al (1992), para comparar poblaciones de jutia conga
en hábitat de manglar y bosque y compararlo también con un índice similar del peso del hígado,
pero relativo al peso corporal. Otros autores (Smith y Berovides, 1984) también utilizaron este
último índice. En otra especie de caviomorfo (Octodon degus, Sabat y Bozinivic, 2008) se han
estudiado los cambios del peso del hígado en relación con la dieta y el desarrollo del animal.
En Cuba las dos estaciones más marcadas y de mas influencia en los ecosistemas cubanos lo son la
seca y la lluvia, por lo que el presente trabajo tuvo como objetivo analizar los cambios poblacionales
y estacionales de un indices nutricional dentro y entre poblaciones de jutía conga de diferentes
hábitat, lo cual puede proporcionar información acerca de la naturaleza adaptativa de dicho índice,
de su relación con la dinámica reproductiva de la población y de si sus cambios son producto o no
de la norma de reacción de la especie, elementos que pueden servir de base para determinar
unidades de significación evolutiva o sus equivalentes (Green, 2005), para implementar futuros
planes de conservación y manejo de la especie basados en dichas unidades.
VI.3. Resultados y discusión
Análisis para una estación del año.

119
Las diferencias entre poblaciones para el índice peso del hígado relativo al peso corporal, se
muestran en la tabla II. Se observaron diferencias estadísticamente significativas, separándose las
poblaciones en tres grupos, con valores crecientes del índice. Grupo 1 Sabana, Hatibonico,
Guanahacabibes y Saetía, de hábitat de manglar con manigua costera, manigua costera y bosque
semideciduo respectivamente; grupo 2 Jardines de la Reina y Najasa, con hábitat de manglar y
bosque semideciduo respectivamente y por último el grupo tres, representado por la Isla, con hábitat
de bosque semideciduo. El primer grupo tiene un estado nutricional inferior al segundo grupo y este
a su vez inferior al tercero. Es de señalar que en los tres grupos se encuentran representadas
poblaciones de bosque, que poseen altos índices de disponibilidad de nutrientes, aspecto que se
asocia perfectamente para los grupos 2 y 3 de mayores índices nutricionales, pero no para el grupo
1, donde se encuentran los valores más bajos, los cuales deben de corresponderse con las localidades
de menor disponibilidad de nutrientes como en los manglares. Este resultado se pudiera explicar por
el grado de asociación entre el peso del hígado y el peso corporal, con correlaciones para cada
población positivas, altas y estadísticamente significativas. Los mayores valores de correlación se
encontraron en las poblaciones de Guanahacabibes (r=0.81) y Najasa (r=0.78), lo cual incide
negativamente en el índice, provocando la falta de concordancia ecológica en los resultados, siendo
de esperar cuando se utiliza factores de corrección altamente asociados como el peso del hígado y el
peso corporal (r=0.743, con amplitud de valores para cada población entre 0.66 a 0.81).
Es de señalar que nuestros resultados en relación a este índice se corresponden con los obtenidos
por Smith y Berovides (1984) en cuatro poblaciones (Jardines 2.4 ,Sabana 3.00, Najasa 3.1 e Isla
4.1) con resultados muy semejante para las poblaciones de bosque y diferencias no marcadas en las
poblaciones de manglar, debido al trimestre de muestreo para estas poblaciones en los meses de
mayo, junio y julio para Jardines y agosto, septiembre y octubre en la población de Sabana, lo cual
pudiera estar influenciado por los cambios estacionales para este índice.
Sánhez et al (1992) en dos poblaciones (Jardines 3.29 y Najasa 3.32) reflejo resultados muy
similares con los obtenidos en nuestro estudio para estas mismas poblaciones.
El índice de nutrición peso del hígado relativo al peso corporal, mostró además diferencias
estadísticamente significativas entre sexos, con mayores valores promedios en las hembras con
respecto a los machos, posiblemente por las mismas causas explicadas anteriormente, debido a que
los machos presentan mayor valor de correlación con el peso corporal (r=0.775) que las hembras
(r=0.687).

120
Tabla II. Valores promedios (X), desviaciones típicas (DS) y coeficientes de variación (CV) del índice peso
del hígado relativo al peso corporal, en sietes poblaciones de jutía conga (Capromys pilorides)
Población N X DS CV
Jardines 43 3.45b 0.605 17.53
Sabana 32 2.86ª 0.469 16.39
Hatibonico 53 2.57ª 0.418 16.26
Guanahacabibes 45 2.65ª 0.426 16.07
Najasa 25 3.75b 0.660 17.60
Saetía 30 2.80ª 0.417 14.89
Isla 24 4.16c 0.695 16.70
Medias con letras diferentes, difieren a p<0.05 por una prueba de Tukey
El análisis entre las poblaciones para el índice del peso del hígado relativo al largo de la pata al cubo
(Tabla III), muestra también diferencias estadísticamente significativas entre las poblaciones,
conformándose tres grupos con el siguiente orden creciente. Grupo 1 Guanahacabibes, Isla y Saetía,
de hábitat bosque semideciduo; grupo 2 Jardines de la Reina, Sabana y Hatibonico, con hábitat
manglar, manglar más manigua costera y manigua costera respectivamente y el grupo 3 integrado
por Najasa con hábitat de bosque semideciduo. Estos resultados son discrepantes en su mayoría con
respecto a los resultados del índice de nutrición anterior y con los resultados esperados según el
índice climático (valores altos del índice nutricional en poblaciones de bosques). Las poblaciones de
la Isla, Saetía y Guanahacabibes, que quedan enmarcadas en el primer grupo con los índices mas
bajos, son poblaciones de bosque que presentaron los valores promedios mayores de pata, siendo
esta la causa de la discrepancia. Como dato adicional se observa en la población de Najasa el menor
valor promedio para esta misma variable, ocupando el grupo 3 (grupo de mayor índice nutricional).
El grado de asociación entre el largo de la pata y el peso del hígado para todas las poblaciones fue de
r=0.38 p<0.05 con valores positivos y estadísticamente significativos en las poblaciones de Sabana,
Hatibonico, Guanahacabibes y Saetía y un valor negativo en la población de la Isla. Las poblaciones
de Jardines y Najasa no mostraron asociación estadísticamente significativa.

121
Estos resultados indican que además de no utilizar factores de corrección altamente asociados al
peso del hígado para el índice de nutrición, es necesario también aumentar en lo posible el número
de poblaciones en un estudio que muestren el comportamiento de las variables en su conjunto dentro
de todas las poblaciones en general, para no incurrir en falsos resultados por las variaciones
absolutas y relativas de dichas variables.
El análisis para el índice de nutrición de Gosling mostró además diferencias significativas
estadísticamente entre sexos, con valores promedios mayores para las hembras con respecto a los
machos, resultado similar al anterior y explicado por las mismas causas.
Tabla III. Valores promedios (X), la desviación típica (DS) y coeficientes de variación (CV) del índice peso
del hígado relativo al largo de la pata al cubo, en siete poblaciones de jutía conga (Capromys
pilorides).
Población N X DS CV
Jardines 43 1.23b 0.081 6.5
Sabana 32 1.316b 0.148 11.29
Hatibonico 53 1.315b 0.161 12.29
Guanahacabibes 45 1.097ª 0.133 12.12
Najasa 25 1.493c 0.194 12.99
Saetía 30 1.048ª 0.121 11.54
Isla 24 1.057a 0.142 13.43
Medias con letras diferentes, difieren a p<0.05 por una prueba de Tukey
La comparación entre poblaciones para el índice de nutrición relativo al largo cabeza tronco (Tabla
IV), produjo diferencias estadísticamente significativas entre las poblaciónes estudiadas. Las
poblaciones se diferenciaron en dos grupos que se muestran en orden creciente. Grupo 1 Jardines,
Sabana, Guanahacabibes, Hatibonico, Saetía y Najasa de hábitat manglar, manglar más manigua
costera, bosque semideciduo, manigua costera y bosque semideciduo respectivamente y el grupo 2
conformado por la Isla con habitat de bosque semideciduo.

122
Estos resultados muestran un gradiente entre poblaciones de hábitat de manglar, manigua costera, y
bosque semideciduo, registrándose los menores valores dentro del grupo 1 del índice de nutrición
para las poblaciones de hábitat de manglar y los mayores valores para las poblaciones de hábitat de
bosque, resultados esperados en relación a la disponibilidad de nutrientes según el índice climático.
Los resultados para este índice de nutrición muestran además concordancia con los resultados del
índice de nutrición relativo al peso en relación a la población de la Isla, que mostró el valor mayor
del índice estadísticamente significativo con respecto al resto de las poblaciones.
El índice nutricional del peso del hígado relativo al largo cabeza tronco no mostró diferencias entre
sexos estadísticamente significativas, lo cual constituye un aspecto diferencial más con respectos a
los índices anteriores.
Tabla IV, Valores promedios (X), la desviación típica (DS) y coeficiente de variación (CV) del
índice peso relativo del hígado al largo cabeza tronco en siete poblaciones de jutía conga
(Capromys pilorides).
Población N X DS CV
Jardines 43 0.845ª 0.301 35.62
Sabana 32 0.859ª 0.322 37.48
Hatibonico 53 0.996ª 0.356 35.74
Guanahacabibes 45 0.931ª 0.334 35.87
Najasa 25 1.048ª 0.176 16.79
Saetía 30 1.027ª 0.369 35.92
Isla 24 1.896b 0.806 42.51
Medias con letras diferentes, difieren a p<0.05 por una prueba de Tukey
El coeficiente de variación mostró el comportamiento siguiente en cada uno de los índices: Para el
índice nutricional relativo al peso corporal se obtuvieron valores semejantes entre las poblaciones
con un promedio de 16.49.El índice nutricional relativo al largo de la pata reflejó un patrón
semejante entre las poblaciones, con un valor promedio de 11.45 y un menor valor en la población
de Jardines de 6.5.En el índice nutricional relativo al largo cabeza tronco, se observaron valores

123
semejantes entre las poblaciones para un promedio de 34.27 y un valor inferior de 16.79 en la
población de jardines y de 42.51 como valor máximo para la población de la Isla.
El coeficiente de variación entre los índices nutricionales aumentó del índice relativo al largo cabeza
tronco respecto al índice relativo al peso corporal y de este en relación al índice relativo al largo de
la pata, aumento estadísticamente significativo. Este resultado puede estar indicando que el primer
índice es más sensible a los cambios naturales de disponibilidad de alimentos, que se dan dentro de
casa población, lo que señala una ventaja mas para el uso de dicho índice.
La tabla V muestra los valores de regresión de forma creciente, para los tres índices nutricionales
utilizados, en relación al ordenamiento de las poblaciones de acuerdo al incremento favorable del
hábitat según el índice climático. Estos resultados reflejan que solo el índice de nutrición relativo al
largo cabeza tronco muestra una asociación entre el valor del índice de nutrición y el índice
climático, estadísticamente significativa, además de presentar el valor mayor de incremento de la
variable dependiente (índice de nutrición) por cada unidad de aumento de la variable independiente
(rangos asignados para el índice climático). Estos resultados sugieren que es precisamente el índice
de nutrición del peso del hígado relativo al largo cabeza tronco. el índice más adecuado para reflejar
el estado nutricional en las poblaciones estudiadas.
Tabla V. Valores de regresión (b), F del NOVA, error típico (Sb) y coeficiente de determinación (R2)
de los índices nutricionales relativo al largo cabeza tronco (PH/LCT3), peso corporal
(PH/PC) y al largo del pie (PH/Pi3) en relación a un índice climático en siete poblaciones de
jutía conga (Capromys pilorides).
Indice F R2 b Sb
PH/LCT3 6.472* 56.40 0.128 0.05
PH/PC 0.959 NS 16.10 0.114 0.116
PH/Pi3 1.027 NS 17.0 0.032 0.031
* p<0.05 NS= No significativo

124
Las regresiones entre las variables peso del hígado (variable dependiente) en relación al LCT Y PC
(variables independientes) se muestran en la Tabla VI. Se observaron valores estadísticamente
significativos para la regresión del peso del hígado con respecto al peso corporal, con incrementos
(b) aproximadamente iguales de la variable dependiente respecto a la variable independiente, en
cada una de las poblaciones. Este era un resultado a esperar, y evidencia que las condiciones de
disponibilidad de alimentos, tan disímiles entre esas poblaciones, no afecta la relación peso hígado-
peso corporal, posiblemente por tratarse de un mecanismo homeostático.
La regresión del peso del hígado (variable dependiente) respecto al LCT (variable independiente)
solo mostró significación estadística para la población de Jardines, localidad con la poblacions de
jutías en habitat mas extremo (manglar de mangle rojo). Esto implica que alimentos de baja calidad
(como suponemos que son los de estos habitats), si influyen en el tamaño del animal, cosa que no
sucede en los bosques.
La no dependencia del peso del hígado en el largo corporal (excepto Jardines), añade otro punto más
de ventaja, para el uso de este índice, en los estudios ecológicos de la jutía conga.
Tabla VI. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error
típico (Sb.) del peso del hígado (variable dependiente) en el largo cabeza tronco (LCT) y
peso corporal (variables independientes) en siete poblaciones de jutía conga (capromys
pilorides).
Población LCT Peso corporal
F R2 b Sb. F R2 b Sb.
Jardines 87.63*** 6.8 0.470 0.051 48.69*** 54.3 0.028 0.004
Sabana 0.33 1.8 -0.720 -0.128 24.38*** 57.5 0.024 0.005
Hatibonico 3.31 6.1 -0.142 0.080 66.61*** 56.6 0.022 0.003
Guanahacabibes 0.16 0.4 0.057 0.138 27.21*** 61.6 0.030 0.006
Najasa 0.46 2.7 0.159 0.232 87.82*** 66.0 0.024 0.003
Saetía 1.18 4.0 0.134 0.123 38.53*** 57.9 0.019 0.003
Isla 0.29 1.6 0.100 0.184 14.02*** 43.8 0.025 0.007

125
La tabla VII, representa el dimorfismo sexual (media de los machos – medias de las hembras) para
los índices relativo al LCT (H/LCT3), al PC (H/PC) y al largo del pie (H/Pi3). Todos los índices
mostraron entre dos a tres diferencias notables (valores muy por encimes de la media de las
diferencias en valores absolutos) y no coincidentes. La mayoría de las diferencias son negativas, lo
que señala que independiente de la localidad, las hembras tienden a tener una mejor condición
nutricional, lo que esta de acuerdo con Emlen y Oring (1977) y Ruckstuhl (2007) para mamíferos en
general y Berovides et a, (1990) para la jutía conga en particular, de que las hembras ocupan habitas
con mejores condiciones de recursos (entre ellos los alimentarios) que los machos. Si esta
segregación sexual por habitat se da realmente, entonces las poblaciones terrestres y semiterrestres
de jutía conga de bosques (las que tienen la mayor posibilidad de hacer tal segregación) deben de
presentar las diferencias entre sexos mas marcadas. Esto es lo que precisamente indica el índice del
peso del hígado en relación al largo corporal.
Tabla VII. Dimorfismo sexual (D.S.) para los índices peso del hígado relativo al largo cabeza tronco
al cubo (H/LCT3), relativo al peso corporal (H/PC) y relativo al largo del la pie al cubo
(H/Pi3) en siete poblaciones de jutía conga (capromys pilorides)
Población H/LCT3 H/PC H/Pi3
Jardines 0.13 -0.52* -0.07
Sabana -0.01 0.20 -0.11
Hatibonico 0.15* -0.09 -0.19*
Guanahacabibes -0.09 -0.07 -0.10
Najasa -0.10 -0.27 -0.15*
Saetía 0.15* -0.13 -0.08
Isla -0.20* -0.46* 0.00
Diferencias con asteriscos señalan valores por encima de la media de las diferencias, en valores
absolutos

126
Los resultados anteriores evidencian que el índice del peso del hígado relativo al largo corporal,
refleja un gradiente que separa a las poblaciones de hábitat bosque de las poblaciones de hábitat de
manigua costera y manglar, así como la segregación ecológica entre los sexos en dependencia del
hábito y el habita, todo lo cual no se evidenció en los índices nutricionales relativo al peso corporal
y al largo de la pata. Estos resultados sugieren, que, para utilizar un índice de nutrición basado en el
peso del hígado, es necesario no utilizar variables altamente correlacionadas (peso corporal) para
ponderar los índices y es necesario además utilizar el mayor número posible de poblaciones de
diferentes localidades, que demuestren las características de las variables de ponderación antes de
ser utilizadas
Efectos de las estaciones del año.
La tabla VIII muestra la variable peso del hígado en los cuatros poblaciones estudiadas. Dada la alta
correlación con el peso corporal, esta variable siguió el mismo patrón que el peso corporal,
separando poblaciones de manglares y bosques, pero con diferencia en cuanto a los cambios por
trimestres, que aquí fueron más marcados. Los ANOVAS produjeron diferencias estadísticamente
significativas (F=4.57, p<0.01) por población (Sabana y Jardines con respecto a Najasa y
Guanahacabibes) con interacción población por trimestre (F=9.43, p<0.001). Dicha interacción se
produce por el aumento de la variable para los meses de ASO en las poblaciones de Cayos (Jardines
y Sabana) y su disminución en las poblaciones de bosque (Najasa y Guanahacabibes), ocurriendo o
contrario en FMA. Este resultado podría estar relacionado con la actividad reproductiva más activa
para las poblaciones de bosque (Tabla IV), como una estrategia adaptativa a las mejores condiciones
alimenticias en este ecosistema, lo cual implicaría un mayor gasto energético. Los cambios
estacionales para la variable peso del hígado dentro de cada localidad, solo fueron significativos
para las poblaciones de Jardines y Guanahacabibes, asociado a una tendencia de aumentar en peso
corporal para los trimestres de ASO en las poblaciones de manglar (Tabla I).
Tabla VIII. Valores promedios (X), desviación típica (S), y coeficiente de variación (CV)) en los
trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO), del peso del
hígado, en cuatro poblaciones de jutía conga (capromys pilorides.

127
Población Trimestre N X S CV t
Jardines FMA 43 82.68 19.83 23.98 29.50*
ASO 26 108.40 25.13 23.18
Sabana FMA 32 81.08 21.88 26.98 0.94 NS
ASO 30 90.20 21.75 24.11
Najasa FMA 38 105.37 30.54 28.98 0.272 NS
ASO 37 100.70 17.28 17.16
Guanahacabibes FMA 47 110.47 24.55 22.22 4.89*
ASO 48 93.78 17.70 18.87
Las regresiones del peso del hígado (variable dependiente) en el tamaño corporal y el peso corporal,
considerando el efecto de los trimestres, se muestran en las tablas IX y X respectivamente. En todas
las poblaciones, hubo una fuerte dependencia lineal estadísticamente significativa, del peso del
hígado en relación al peso corporal y solo para la población de Jardines en relación al LCT en los
dos trimestres (FMA y ASO) y Guanahacabibes para un solo trimestre (ASO). Este es el mismo
patón encontrado cuando solo se analizó FMA.
Tabla IX. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error
típico (S.) del hígado (variable dependiente) en el largo cabeza tronco (LCT) y el peso
corporal (variables independientes) en cuatro poblaciones de jutía conga (Capromys
pilorides), en el trimestre febrero, marzo y abril (FMA).
Población LCT Peso corporal
F R2 b E.S. b F R2 b E.S. b
Jardines 87.63*** 6.8 0.470 0.051 48.69*** 54.3 0.028 0.004
Sabana 0.33 1.8 -0.720 -0.128 24.38*** 57.5 0.024 0.005
Najasa 0.46 2.7 0.159 0.232 87.82*** 66.0 0.024 0.003
Guanahacabibes 0.16 0.4 0.057 0.138 27.21*** 61.6 0.030 0.006

128
Tabla X. Coeficiente de regresión (b), F del ANOVA, coeficiente de determinación (R2) y el error
típico (S.) del hígado (variable dependiente) en el largo cabeza tronco (LCT) y el peso
corporal (variables independientes) en cuatro poblaciones de jutía conga (capromys
pilorides), en el trimestre agosto, septiembre y octubre (ASO)
Población LCT Peso corporal
F R2 b E.S. F R2 b E.S.
Jardines 18.32*** 32.50 0.166 0.109 44.07*** 53.7 0.033 0.005
Sabana 0.003 0.00 0.007 0.137 2.57+ 10.0 -0.008 0.005
Najasa 17.04 37.80 0.271 0.066 8.56** 23.4 0.009 0.003
Guanahacabibes 3.41+ 22.20 0.171 0.097 5.14* 30.0 0.014 0.006
De forma general, el patrón observado en FMA se mantiene en ASO para la dependencia del peso
del hígado en el LCT y PC. En relación a la dependencia del peso del hígado con el LCT, se
mantiene la significación estadística solo en Jardines, lo que puede estar indicando un alto valor
adaptativo el mantener esta relación, independiente de los cambios estacionales. Las dos
poblaciones de habitat no boscoso, disminuyeron la magnitud del valor de la regresión y aunque no
son significativas, las regresiones de las poblaciones de bosques, por el contrario, tendieron a
aumentar en magnitud. Esto sugiere que las condiciones benignas del trimestre ASO en este habitat,
parecen propiciar una cierta dependencia del peso del hígado con el LCT, pero muy débil.
Excepto para Jardines, la dependencia del peso del hígado en el peso corporal, aunque se mantuvo
significativa, tuvo la tendencia a disminuir su magnitud en todas las poblaciones. Es como si la
mayor energía disponible y limitada contenida en el hígado en ASO y en el habitat de bosque, se
dedicará mas al aumento de tamaño que al aumento de peso.
Dinámica reproductiva en manglares y bosques y su relación con los índices nutricionales.
El grado de actividad reproductiva (AR) en dos de las poblaciones estudiadas presentó diferencias
marcadas entre trimestres y poblaciones (Tabla XI). Para los machos, Najasa siempre presentó

129
mayores valores de AR en ambos trimestres y la actividad fue mayor en FMA. Para las hembras, el
porcentaje en reproducción siempre fue mayor en Najasa y estable entre los dos trimestres, lo que no
se dio en Jardines, además, las etapas tempranas de estadios embrionarios disminuyen de FMA a
ASO, pero con diferentes valores. Todos estos datos sugieren que la dinámica reproductiva de las
hembras en estas poblaciones es muy diferente, lo que podría influir en la dinámica del índice
nutricional.
Tabla XI. Actividad reproductiva en febrero, marzo y abril (FMA) y agosto, septiembre y octubre
(ASO) en dos poblaciones de jutía conga (Capromys pilorides)
Machos Hembras
Trimestre Población DT% IR HR% EA%
FMA Jardines 100 4.89 68.75 43
Najasa 100 5.48 72.91 29
ASO Jardinaes 25 4.65 37.87 33
Najasa 46 5.17 73.08 15
DT: Descenso testicular
IR: Indice reproductivo (peso testículo/peso corporal)
HR: Hembras en reproducción (lactancia, gestación o ambas)
EA: Estadio embrionario A (el mas temprano)
El análisis de varianza para el índice nutricional peso hígado/LCT, mostró que solo fueron
estadísticamente significativa las interacciones poblaciones por trimestre (F=6.46, p<0.001) y
trimestre por sexo (F=7.14, p<0.01). Este índice parece mas sensible a las interacciones población
por trimestre y por sexo que el peso del hígado, que si detectaba diferencias entre poblaciones
independiente de dicha interacción y no detectó diferencias entre sexos. En el trimestre FMA
aparecen altos valores del índice en las poblaciones de bosque en comparación con las de manglares,
ocurriendo lo contrario en el trimestre ASO, que es el mismo patrón observado para el peso del
hígado, con la misma explicación. La interacción población por trimestre significativa, para el peso
del hígado tanto absoluto como relativo, evidencia fuertemente que esta variable es producto de la

130
norma de reacción del genoma de la jutía conga, ya que respuestas plásticas al ambiente pueden
influenciar fuertemente la divergencia entre poblaciones locales, que varían en las caracteristicas de
sus hábitats (Patton y Yang, 1971), como en este caso.
Cuando se analizan las anteriores diferencias por separados para cada trimestre, resultaron
estadísticamente significativas (Figura 1). Este patrón se explica teniendo en cuenta tanto el ciclo
reproductivo de la jutía conga como la disponibilidad del alimento, que cambia con los trimestres
(mas abundante en el trimestre ASO).
Figura 1. Indice nutricional en cuatro poblaciones de jutía conga, por trimestre.
El índice nutricional por población y trimestre (Tabla XII) muestra los siguientes resultados. La
población de Jardines se ve favorecida en el trimestre ASO, lo que posiblemente este relacionados
con cambios nutricionales favorables en la época de lluvia, más evidentes para ecosistemas de
manglar con respecto a ecosistemas de bosques. Además, debe de considerarse la estrategia
reproductiva de la población de Jardines (Gutiérrez et al., 1999), ya que en esta población disminuye
en un 46% la actividad reproductiva en las hembras y 75% en los machos durante el trimestre ASO,
Indice nutricional
0
0,2
0,4
0,6
0,8
1
1,2
FMA ASO
Trimestres
Ind
ice
Jardines
Sabana
Najasa
Guanaha

131
lo que a su vez disminuye el gasto energético y prepara la población para la próxima etapa
reproductiva, como muestra de una población bien adaptada al ecosistema de manglar.
La población de Sabana con hábitat de cayo y manigua costera representa una posición intermedia
entre manglar y bosque, donde el índice nutricional también es favorecido en el trimestre ASO,
aunque estratégicamente la población mantiene una actividad reproductiva( datos no presentados)
más elevada ya que disminuye un 75% de gestantes en comparación con la población de Jardines
que disminuye en un 85% para el trimestre de ASO (Gutierrez, et al.1991).Esto redunda en un
mayor gasto energético y un aumento no semejante, al parecer por la falta de adaptación total a este
ecosistema, influenciado por el flujo genético con las poblaciones de bosque o por un reciente arribo
geológico o ambas (Berovides y Comas, 1997) )
Las poblaciones de Najasa y Guanahacabibes, con hábitat de bosque, son nutricionalmente más
favorecidas por la variedad de alimentos vegetales (Comas et al., 1994), cuyas proteínas son mas
accesibles que en los manglares. Sabat y Bozinovic, (2008) en el caviomorfo Octodon degus,
demostraron que el peso del hígado se incrementaba con una dieta rica en proteínas. Este hecho
permite en ambas poblaciones de jutías, una estrategia reproductiva más estable, pero a su vez un
mayor gasto energético en los dos trimestres, ejemplificado para la población de Najasa, donde se
observa el mismo por ciento de hembras reproductivas en ambos trimestres, con una fase de meseta
mas prolongada para los partos hacia el trimestre ASO (Gutierrez et al., 1999). Por otra parte, Naya
et al., (2008) demostraron en el Degus que con la lactancia (una de las etapas de alto gasto
energético) aumentaba el peso del hígado.
Tabla XII. Valores medios (X), desviación típica (S), prueba t y coeficiente de variación (CV) por
población en los trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre
(ASO) para el índice nutricional, en cuatro poblaciones de jutía conga (Capromys pilorides).
Poblaciones Estación N X S CV t
Jardines FMA 43 0.835 0.119 14.23 5.57***
ASO 26 1.046 0.212 20.27
Sabana FMA 32 0.861 0.297 34.49 0.82 NS
ASO 30 0.931 0.275 29.54

132
Najasa FMA 38 0.911 0.243 26.67 0.85 NS
ASO 37 0.847 0.129 15.23
Guanahacabibes FMA 47 0.931 0.301 30.25 0.83 NS
ASO 48 0.898 0.252 31.58
Las diferencias del índice por trimestres entre los sexos fueron significativas para FMA (estación de
seca con escasez de alimentos), con valores mayores para las hembras (Tabla XIII ), resultado que
puede explicarse por la estructura social que conforman las poblaciones de jutía conga, compuestas
por grupos familiares con un macho y cuatro hembras aproximadamente (sistema de apareamiento
poligínico), ocurriendo la selección de los mejores lugares por parte de las hembras (Berovides et
al., 1990), que se encuentran en una fase temprana de su ciclo reproductivo, al contrario de los
machos que en este trimestre poseen una elevada actividad reproductiva pero bajo las condiciones
desfavorables en este trimestre . El trimestre ASO (época de lluvia con abundancia de alimentos) no
muestra diferencias estadísticamente significativas entre los sexos, posiblemente porque los machos,
ahora con menor actividad reproductiva, se recuperan, lo que parece válido en ambos hábitats. Pero
las hembras, mas influidas que los machos por la disponibilidad de alimentos (Emlen y Oring, 1977;
Ruckstuhl, 2007), varían en su actividad reproductiva en dependencia del hábitat. En los manglares
(Jardines) se disminuye la actividad reproductiva en este trimestre, lo que no ocurre en el bosque
(Najasa), en este último ecosistema se mantiene igual la actividad reproductiva que en FMA. Dada
esta mayor actividad, debía esperarse una disminución del índice, pero Naya et al., (2008)
demostraron en el Degus que con la lactancia (una de las etapas de alto gasto energético), aumentaba
el peso del hígado.
Tabla XIII. Valores medios (X), desviación típica (S), y coeficiente de variación (CV) por sexo en
los trimestres febrero, marzo y abril (FMA) y agosto, septiembre y octubre (ASO), para el
índice nutricional peso hígado/LCT, en cuatro poblaciones de jutía conga (Capromys
pilorides).
Estación Sexo N Media S CV t
133
FMA H 91 0.931 0.278 29.86 2.61*
M 69 0.815 0.230 28.22
ASO H 66 0.861 0.218 24.47 1.08 NS
M 55 0.937 0.225 24.01
Los resultados con el índice nutricional del peso del hígado relativo al LCT, evidencian diferencias
entre las poblaciones de manglares y bosques, pero dependientes de la estación del año. Estudios
comparativos realizados por otros autores, entre una población de cayo y una de bosque (Sánchez et
al., 1992) no hallaron diferencias significativas, siendo de señalar que en estos casos el índice de
nutrición utilizado fue diferente, pues el peso del hígado lo dividieron entre el peso total del cuerpo
y el largo de la pata al cubo. Nuestros resultados indican que las poblaciones de jutía conga han
sufridos adaptaciones locales bajo diferentes presiones selectivas.
Nuestros resultados señalan un comportamiento bastante parecido de la variable peso del hígado
absoluto y relativizado al LCT (índice nutricional), pero recomendamos el uso del segundo por su
mejor comportamiento discriminatorio, en relación al efecto total de las diferencias entre sexos, no
detectado por el peso absoluto.
De los resultados anteriores se concluye que el índice nutricional aquí empleado, no mostró
diferencias estadísticamente significativas entre poblaciones con hábitat diferentes (manglar y
bosque) al analizar los efectos trimestre, población y sexo, pero si una interacción cruzada
significativa localidad por trimestre del año, lo cual explica que, en un análisis independiente para
cada trimestre, si se detectaran diferencias significativas entre localidades. Este patrón diferencial
entre poblaciones de bosque y manglar, tiene como causas más probables, la alta relación del índice
con la actividad reproductiva propia de cada población, y con la disponibilidad de recursos
alimenticios.

134
CAPÍTULO VII. Variaciones morfológicas locales de dos poblaciones de jutía
conga (Capromys pilorides) en el área geográfica Peninsula de
Guanahacabibes
VII.1. Introducción
En las metapoblaciones, como conjunto de poblaciones interconectadas por flujo genético con efecto
homogenizador (Hanski y Gogiotti, 2004), pueden presentarse variaciones locales entre sus
subpoblaciones por efecto de la selección natural. Ambos procesos evolutivos se reconocen como
uno de los elementos fundamentales en la diferenciación de las poblaciones (Kawecki y Ebert,2004)
y son estudiados analizando la variación de caracteres morfológico, fisiológicos, conductuales,
cariológico y moleculares, estando inicialmente asociados a problemas mayormente taxonómicos.
Actualmente se enfoca además hacia problemas de la conservación, evolución y biogeogrfía (Santos
y Hortelano, 1997).
Las metapoblaciónes que conforman las especies en la naturaleza, presentan diferentes categorías
de agrupamiento de sus subpoblaciones, siendo las subespecies de los taxonomistas los grupos más
superiores, aunque desde el punto de vista genético y conservacionista, existen ahora agrupaciones
que pueden o no coincidir con las subespecies tradicionales de los taxónomos. Las subespecies
tradicionales se clasifican en geográficas y ecológicas, siendo estas últimas, subespecies geográficas
que viven bajo condiciones más específicas del hábitat local (Mayr, 1969), como son la diversidad
en la formación vegetal, humedad, precipitación, temperatura y sustrato entre otros efectos, que
conllevan a variaciones adaptativas entre poblaciones.
La jutía conga (Capromys pilorides) con una amplia distribución geográfica y diversos hábitats
ocupacionales, ha sido estudiada en cuanto a su variación geográfica considerando variables
morfológicas, morfométricas, y morfofisiológicas (Lora,1895; Mohr,.1939; Abreu y Manójina,1989;
Smith y Berovides,1984; Berovides et al., 1990a;1990b; Sánchez et al,1992; Berovides,2005; Silva
et al., 2007; de la Fuente y Berovides, 2008; de la Fuente y Berovides,2009). Estos estudios se han
llevado cabo en poblaciones de bosques, manglares, ciénagas, vegetación xerofítica y vegetación
secundaria, pero geográficamente muy distantes entre sí, quedando pendiente un análisis de la
variación en una misma área geográfica, donde se puedan distinguir poblaciones cercanas
pertenecientes a la misma metapoblación pero que puedan responder de manera diferencial a las
fuerzas evolutivas predominantes, el flujo genético (que mantiene la homogeneidad entre las
poblaciones) o las presiones de selección natural ( que conducen a la diferenciación local).

135
El objetivo de nuestro trabajo fue analizar comparativamente mediante variables morfométricas,
morfofisiológicas e índices derivados de estas variables absolutas, diferencias locales entre dos
poblaciones de jutía conga que pertenecen al área geográfica de Guanahacabibes, las que forman
parte de una metapoblación en dicha área.
VII.2. Materiales y métodos
En el área geográfica perteneciente a la península de Guanahacabibes, donde se ubica el Parque
Nacional de igual nombre, se estudiaron dos poblaciones pertenecientes a dos localidades de hábitat
de bosque semideciduo, Bolondrón y Carabelita, separadas entre si por unos 20 Km de distancia. En
la tabla I se dan las cantidades de animales capturados por localidad y sexo y las características de
los bosques de estas dos localidades.
Tabla I. Localidad de las poblaciones, sexo, tamaño de muestra (N) y formaciones vegetales, en dos
poblaciones de jutía conga (Capromys pilorides) del Parque Nacional Guanahacabibes.
Localidad N Sexo Formación vegetal
Bolondrón 21 H Bosque semideciduo sobre suelo de resina roja
(acumulación de materia orgánica en el suelo calizo) que
favorece el desarrollo de la vegetación
19 M
Carabelita 34 H Bosque semideciduo sobre suelo calizo, que forma un
ecotono con el manglar, mas pobremente desarrollado
que en Bolodrón 21 M
Todos los ejemplares capturados fueron adultos según el criterio de Smith y Berovides (1984)
y se les determinó el peso corporal (PC) y el peso del hígado (PH) en gramos (g) así como el largo
cabeza tronco (LCT), Largo total (LT), largo de la cola (LC), largo de la pata izquierda con uña (Pi),
grosor de la corteza renal (GC), grosor de la médula renal (GM) y longitud última porción del
intestino grueso (LIU) en mm. A cada una de las variables absolutas obtenidas se les realizó la
prueba de normalidad Kolmogorov-Smirnov, la prueba de homogeneidad de varianza de Levene y se

136
confeccionaron los índices morfométricos y morfofisiológicos derivados de las variables absolutas,
que aparecen en la Tabla II.
Tabla II. Variables e índices morfométricos y morfofisiológicos con sus valores adaptativos,
utilizados en el estudio de dos poblaciones de jutía conga (Capromys pilorides) del Parque
Nacional Guanahacabibes.
INDICES MORFOMÉTRICOS FÓRMULAS FUNCIÓN ADAPTATIVA
Robustez (Peso relativo al volumen
del cuerpo)
PC/LCT3*100000 Supervivencia y reproducción
(Stearns,1992)
Peso corporal relativo al largo total
(PC/LT)
PC/LT*1000 Supervivencia y reproducción
(Stearns,1992)
Cola Relativa a LCT (CoR) LC/LCT*100 Termorregulación (Millien et
al,2006), arboricidad (Fleming,1974)
Pie Relativo a LCT (PiR) Pi/LCT *1000000 Termorregulación (Millien et, 2006),
arboricidad (Fleming,1974)
INDICES mORFOFISIOLÓGICOS
Indice de nutrición. Peso del hígado
relativo al volumen del cuerpo
(PH/LCT)
PH/LCT3*1000000 Estado nutricional (Indice de gossling
et al., 1980, modificado por de la
Fuente y Berovides, 2009)
Grosor medula renal relativo al grosor
del riñón
(IRRY)
GM/GM+GC*100 Capacidad de reabsorber agua (Yabe,
1983)
Grosor medula renal relativo al LCT
(IRRL)
GM/LCT*1000 Capacidad de reabsorber agua (de la
Fuente y Berovides ,en prensa)
Longitud última porción intestino
grueso relativo al LCT
LIU/LCT Capacidad de absorber agua
(Sánchez et al , 1992)
Los valores medios de los índices elaborados y las variables absolutas se compararon entre
poblaciones mediante un análisis de varianza (ANOVA) bifactorial, considerando los efectos

137
población y sexo, descartando el efecto estación, ya que todos los animales fueron capturados en el
trimestre febrero, marzo, abril que se corresponde con la estación de seca en Cuba. También se
analizó la variabilidad de los caracteres estudiados (medida por el coeficiente de variación) para las
poblaciones y los sexos.
VII.3. Resultados y discusión
Las variables absolutas largo cabeza tronco (LCT) y peso corporal (PC) representadas en la tabla III,
mostraron un mismo patrón, donde se observaron mayores pesos corporales y LCT en los machos
con respecto a las hembras en las dos poblaciones estudiadas, con diferencias estadísticamente
significativas. El peso corporal independiente del sexo, mostró diferencias estadísticamente
significativas entre las poblaciones, con valores promedios mas altos en la población de Bolondrón
con respecto a la población de Carabelita.
El LCT reflejó una tendencia de valores promedios más altos en la población de Bolondrón con
respecto a Carabelita, no significativo estadísticamente.
Esto resultado sugieren que las diferencias de peso corporal entre poblaciones no se deben solo a las
diferencias con respecto al LCT, sino a otros aspectos como nutricionales y o conductuales,
diferenciados entre las localidades
Tabla III. Valores medios (X) y desviación típica (S) por sexo y población, para el largo cabeza
tronco (LCT) y peso corporal (PC), en dos poblaciones de jutía conga (Capromys pilorides)
del Parque Nacional Guanahacabibes.
LCT (mm) PC (g)
Población Sexo X S X S X S X S
Bolondrón H 485.25 44.74 499.62 40.80 3829.06 851.53 4315.87 975.73
M 515.07 30.20 4835.13 838.72
Carabelita H 476.53 27.94 483.40 30.94 3365.99 473.86 3556.53 559.95
M 494.52 32.98 3865.14 560.62

138
Diferencias entre poblaciones (F del ANOVA):
No significativa F=23.7 **
Diferencias entre sexos dentro de poblaciones:
Bolondrón t =2.15* t = 3.31** * p< 0.05
Carabelita t = 2.16* t = 3.53** ** p < 0.01
Los índices de Robustez e índice relativo al largo total (tabla IV), mostraron resultados semejantes
entre sí en cuanto a las diferencias entre poblaciones, donde en ambos índices la población de
Bolondrón presentó mayores valores promedios con diferencias estadísticamente significativas con
respecto a la población de Carabelita, lo cual señala una mayor distribución del peso por unidad de
tamaño ,sugiriendo que el hábitat en la localidad de Bolondrón es más favorable o permite posibles
cambios conductuales que conlleven a estos resultados.
En los dos índices y en ambas localidades, los valores promedios de los machos fueron mayores que
en las hembras, siendo estadísticamente significativo para el índice de peso relativo al largo total.
Tabla IV. Valores medios (X) y desviación Típica (S) por sexo y población, para el índice de
robustez y el índice del peso relativo al largo total (PC/LT), en dos poblaciones de jutía
conga (Capromys pilorides) del Parque Nacional Guanahacabibes.
Robustez PC/LT
Población Sexo X S X S X S X S
Bolondrón H 3.35 0.55 3.44 0.53 5.24 0.81 5.69 0.96
M 3.53 0.52 6.17 0.89
Carabelita H 3.12 0.45 3.16 0.45 4.79 0.58 4.99 0.62
M 3.21 0.45 5.32 0.54
Diferencias entre poblaciones (F del ANOVA):
F=6.2 * F=18.0 **

139
Diferencias entre sexos dentro de poblaciones:
Bolondron t = No significativa t = 2.44* * p< 0.05
Carabelita t = No significativa t = 2.11* ** p < 0.01
Los valores promedios de la cola absoluta y relativa (tabla V) expresaron diferencias por población
estadísticamente significativas y diferencias entre sexos e interacción población por sexo, para la
variable cola absoluta. El resultado en relación al sexo está posiblemente relacionado con los valores
promedios mayores del LCT en los machos con respecto a las hembras en ambas poblaciones, lo que
por alometría se reflejaría en la cola. La interacción población por sexo viene dada por el hecho de
que la diferencia entre sexos en cola absoluta es mayor en Bolondrón (estadísticamente sigificativa)
con respecto Carabelita (no significativa).
La variable cola relativa, como elimina el efecto del tamaño, no presentó diferencias entre
poblaciones independiente del sexo, ni interacción, auque sí diferencias apreciables entre sexos para
la población de Bolondrón, con resultados estadísticamente significativos. Esto indica que, para esta
población, la diferencia en longitud de cola entre sexos es independiente en parte del tamaño del
animal. Como este carácter indica habilidad trepadora, al parecer los machos se segregan de las
hembras en árboles más altos, a donde pueden trepar mejor. Al menos para nuestra muestra, esto fue
lo que observamos.
Las variables pata absoluta y pata relativa (tabla VI) mostraron el mismo patrón que la cola absoluta
y relativa, con diferencias estadísticamente significativas de ambas variables entre poblaciones y
entre sexo e interacción población por sexo solo para la pata absoluta. Las mismas causas dadas para
el patrón observado en la cola, son las que posiblemente estén actuando aquí y refuerzan la hipótesis
de la mayor arboricidad de los machos de Bolondrón.
Tabla V. Valores medios (X) y desviación Típica (S) por sexo y población, para la longitud de la cola
absoluta y relativa en dos poblaciones de jutía conga (Capromys pilorides) del Parque
Nacional Guanahacabibes.
Población Cola Cola relativa
Sexo X S X S X S X S
Bolondron H 238.50 18.94 251.25 22.22 43.33 3.52 50.41 3.97
M 264.87 17.03 51.56 4.21
Carabelita H 225.18 19.68 226.41 18.47 47.48 5.58 47.01 4.72

140
M 228.43 16.56 46.26 2.83
Diferencias entre poblaciones (F del ANOVA):
F=35.5 *** F=12.6 ***
Diferencias entre sexos dentro de poblaciones:
Bolondron t =4.07*** t = 5.92*** *** p< 0.001
Carabelita t = No significativa t = No significativa
Tabla VI. Valores medios (X) y desviación Típica (S) por sexo, para la longitud del pie absoluto y
relativo, en dos poblaciones de jutía conga (Capromys pilorides) del Parque Nacional
Guanahacabibes.
Población Pie Pie Relativo
Sexo X S X S X S X S
Bolondron H 92.82 4.92 97.13 6.93 19.22 1.33 19.50 1.44
M 101.73 5.76 19.81 1.53
Carabelita H 89.14 4.27 90.70 5.73 18.74 0.98 18.80 0.97
M 93.21 6.10 18.87 0.97
Diferencias entre poblaciones (F del ANOVA):
F=27,2 *** F=7.2** * p< 0.05
Diferencias entre sexos dentro de poblaciones:
Bolondrón t = 4.64*** t = No significativa * p< 0.01
Carabelita t = 2.68* t = No significativa *** p< 0.001
La tabla VII presenta los índices de reabsorción renal relativos al grosor del riñón (IRRY) y al LCT
(IRRL), donde se muestran diferencias estadísticamente significativas entre poblaciones solo para el
índice IRRL. Estos resultados ratifican el alto nivel discriminativo del índice IRRL con respecto al
IRRY, cuando se analizan poblaciones con diferencias en cuanto a la disponibilidad de agua.

141
Igualmente deben considerarse las afectaciones del índice IRRY producto de los cambios en la
corteza renal entre sexo y por estación, lo que posiblemente produjo la deferencia estadísticamente
significativa entre sexos en Bolondrón.
Los resultados de Carabelita con respecto a Bolondrón sugieren una localidad más xérica a causa de
la humedad, tipo de suelo, temperatura o por la influencia que representa la presencia de un manglar
cercano, de donde posiblemente provienes muchos de los animales capturados para este estudio.
Tabla VII. Valores medios (X) y desviación Típica (S) por sexo y localidad, para un índice de
reabsorción relativo al grosor del riñón (IRRY) y un índice de reabsorción relativo al largo
cabeza tronco (IRRL), en dos poblaciones de jutía conga (Capromys pilorides) del Parque
Nacional Guanahacabibes.
Población IRRY IRRL
Sexo X S X S X S X S
Bolondron H 73.26 3.12 72.11 2.84 27.26 3.76 26.25 3.57
M 70.81 1.91 25.17 3.11
Carabelita H 74.11 4.36 73.45 4.37 29.70 5.02 29.00 4.36
M 72.38 4.28 27.90 2.75
Diferencias entre poblaciones (F del ANOVA):
No significativa F=7.9**
Diferencias entre sexos dentro de poblaciones:
Bolondrón t =2.65* t = No significativa * p< 0.05
Carabelita t = No significativa t = No significativa * * p< 0.01
El índice de nutrición no mostró diferencias estadísticas entre poblaciones (Tabla VIII), pero si una
interacción cruzada estadísticamente significativa, presentando los machos de Bolondrón mayores
valores que las hembras y ocurriendo lo contrario en Carabelita, aunque no se evidenció diferencias
estadísticas entre sexos dentro de cada población. El peso del hígado esta influido mayormente por
la nutrición y el estado reproductivo del animal, y ambos factores pudieran ser diferentes en las dos

142
poblaciones, afectando de forma diferencial a machos y hembras. Al menos para la reproducción, ya
existen evidencias de esta diferencia (Linares, en prensa).
Tabla VIII. Valores medios (X) y desviación típica (S) por sexo y población, para un índice de
nutrición relativo al largo cabeza tronco (PH/LCT), en dos poblaciones de jutía conga
(Capromys pilorides) del Parque Nacional Guanahacabibes.
PH/LCT
Población Sexo X S X S
Bolondrón H 0.92 0.18 0.96 0.18
M 1.00 0.18
Carabelita H 0.94 0.23 0.91 0.21
M 0.85 0.16
Diferencias entre poblaciones (F del ANOVA):
No significativa
Diferencias entre sexos dentro de poblaciones:
Bolondron t = No significativa
Carabelita t = No significativa
La tabla IX refleja el valor relativo de la última porción del intestino grueso, donde se observaron
diferencias estadísticamente significativas entre poblaciones, con valores promedios mayores para la
población de Carabelita. Resultados similares se reportan para este índice al analizar poblaciones de
hábitat de manglar (Sánchez et al., 1992). Esto sugiere que la población de Carabelita se encuentra
mejor adaptada a condiciones más extremas de carencia de agua, lo que concuerda con los valores
del índice IRRL, en ambos casos relacionan a la población de Carabelita con un hábitat más xérico o
con la influencia del manglar en esta localidad.

143
Tabla IX. Valores medios (X) y desviación Típica (S) por sexo para el índice intestinal relativo al
largo cabeza tronco (LIU/LCT), en dos poblaciones de jutía conga (Capromys pilorides) del
Parque Nacional Guanahacabibes.
Población Sexo X S X S
Bolondrón H 4.08 0.51 4.14 0.65
M 4.20 0.80
Carabelita H 4.98 0.69 5.01 0.61
M 5.07 0.46
Diferencias entre poblaciones (F del ANOVA):
F=37.4***
Diferencias entre sexos dentro de poblaciones:
Bolondron t = No significativa ***p<0.001
Carabelita t = No significativa
Los valores del coeficiente de variación para los caracteres morfométricos (LCT, PC, Co y Pi) y
morfofisiolóicos (IRRY, IRRL, PH/LCT e LIU/LCT), se muestran en la tabla X.
Para ambas poblaciones, el grado de variación del conjunto de caracteres morfométricos y
morfofisiolóicos (medidos por el coeficiente de variación) fue diferente. Los primeros fueron más
variables en Bolondrón y los segundos en Carabelita. Para las variables morfométricas, este patrón
se puede explicar en términos de una mayor heterogeneidad estructural de la vegetación en
Bolondrón, la que como ya señalamos explica también las diferencias en valores promedios entre
machos y hembras en esta localidad, así como la mayor variabilidad de las hembras (CV
promedio=11.17 vs 8.82 para los machos), al ser estas menos arborícolas. Para los caracteres
morfofisiológicos, el patrón observado también podría explicarse en términos de la disponibilidad de
agua, mas heterogénea en Carabelita, por tratarse de una población que utiliza dos hábitats
diferentes en cuanto a este recurso, pero aquí son los machos los que poseen mayor coeficiente de
variación promedio (CV=8.74 vs 8.36 para las hembras), posiblemente por su mayor movilidad.

144
TablaX. Coeficientes de variación (CV) para los caracteres largo cabeza tronco, peso corporal, largo
de la cola, largo del pie, índice renal Yabe (IRRY), índice renal relativo al largo cabeza
tronco (IRRRL), índice nutrición e índice intestinal en dos poblaciones de jutía conga
(Capromys pilorides), del Parque nacional Guanahacabibes.
Caracteres Bolondrón Carabelita
Largo cabeza tronco 8.17 6.40
Peso corporal 22.61 15.73
Largo cola 8.84 8.16
Largo pie 7.13 6.32
CV Promedio 11.69 9.15
Indice IRRY 3.94 5.95
Indice IRRL 13.60 15.03
Indice nutrición 18.75 23.08
Indice intestinal 15.70 12.17
CV Promedio 13.00 14.06
Nuestros resultados en general son de la misma magnitud que los registrados para poblaciones de
jutía conga muy separadas geográficamente, conviviendo en varios hábitats (Berovides et al., 1990a;
1990b; Sánchez et al., 1992; Berovides, 2005), lo que demuestra de nuevo la existencia de ecotipos,
pero a una escala espacial mas reducida. Bajo estas condiciones, el flujo genético manifestaría su
efecto homogenizador entre las poblaciones y si este es elevado, elimina o enlentece la adaptación
local entre poblaciones de hábitat diferentes (Lenormand, 2002), lo que aparentemente no ocurre en
las dos poblaciones estudiadas.
Las diferencias morfométricas y morfofisiológicas entre las localidades de Bolondrón y Carabelitas,
separadas por una distancia aproximada de 20 Km sin la presencia de barreras física entre ellas,
muestran o bien que las presiones selectivas locales disminuyen o anulan el flujo genético, al
eliminar a los inmigrantes inadaptados, o que entre ellas existe poco flujo genético o ninguno,
debido a la filopatría de la especie, que hace que solo migren muy pocos individuos. Ambos

145
procesos podrían estar aquí en operación, ya que los organismos genéticamente más favorecidos, no
solo son los responsables del flujo genético entre las poblaciones, en ocasiones organismos
marginales que no pueden competir por los recursos, son también responsables de la migración
(López-Sepulcre, 2009). Si este es el caso para la jutía conga, entonces los emigrantes entre
poblaciones serian estos pocos individuos marginales, que, dada la adaptación a su localidad de
origen y su baja competitividad, tendrían poca probabilidad de supervivencia y reproducción en su
nueva localidad y serian eliminados por la selección natural.
Al menos para la variación local del color del pelaje, en 11 del roedor ratón de bolsillo (Chaetodipus
intermedius, Hoesktra et al (2005) encuentran una fuerte correlación entre dicha variación y el color
de fondo del hábitat, reconociéndola como adaptación local sometida a una fuerte selección natural,
a pesar del marcado flujo genético entre las poblaciones.
Similar resultado encuentra Laurence (2013) en poblaciones de la rata almizclera (Ondatra
zibethicus) de Canadá, caracteres craneales y microsatélites.
Nuestros hallazgos señalan la necesidad de especificar con exactitud la localidad de las muestras
cuando se hacen los estudios morfométricos con la jutía conga, lo que no ocurrió en el trabajo de
Abreu y Manójina (1989), que sólo refieren la localidad como península de Guanahacaibes, aunque
si se refieren al tipo de hábitat (bosque semideciduo), pero queda demostrado en este trabajo que la
sola referencia al tipo de bosque sin mas especificaciones, no garantiza la repetibilidad de los
resultados. Comparando sus datos morfométricos con los nuestros, se evidencia que las muestras
fueron extraídas de un bosque similar al de Bolondrón.
Todos los resultados anteriores muestran, que en el área geográfica de la península de
Guanahacabibes, las poblaciones de Bolondrón y Carabelita presentaron variaciones entre localidad
estadísticamente significativas, en correspondencia con el hábitat, con valores promedios mayores
con respecto al peso corporal e índices de robustez y masa corporal en la población de Bolondrón y
valores promedios de los índices de reabsorción renal relativo al largo y longitud última porción del
intestino grueso asociado a un hábitat más xérico, en la población de Carabelita, además, de
diferencias en los índices pie relativo y cola relativa. Estos resultados evidencian que las presiones
selectivas en estas localidades superan al efecto del flujo genético entre ellas de existir éste,
produciendo las adaptaciones locales observadas. Este resultado sugiere fuertemente, que estas
poblaciones de jutía conga en el área de la península de Guanahacabibes y quizás en muchas otras
áreas de Cuba, están sufriendo un proceso de especiación ecológica, o sea , un proceso incipiente de

146
formación de especie por respuesta adaptativa a la selección natural estabilizante (dentro de la
población) y disruptiva (entre las poblaciones) lo que logra una mejor adaptación a las condiciones
locales, independiente del efecto homogenizador del flujo genético (Schluter, 2001). La
independencia de flujo genético, según un modelo, se logra por selección sexual (las hembras eligen
machos con el fenotipo local), dimorfismo sexual y filopatría, todo lo cual refuerza la acción de la
selección natural (Sander et al., 2009). Para la jutía conga, lo primero esta por demostrar, pero las
dos últimas condiciones si se dan para la especie (Berovides et a, 1990).

147
CAPÍTULO VIII. Determinación de unidades de manejo para la conservación en
poblaciones de jutía conga (Capromys pilorides)
VII.1. Introducción
Los caracteres adaptativos como adecuación organismo- ambiente y producto del modelaje de la
selección natural, representa un aspecto de gran interés para diferenciar poblaciones dentro de la
misma especie, sometida a diferentes presiones selectivas, con disyunción de rango y hábitat
diferentes. Aquí la variación de caracteres adaptativos constituye el material inicial para la
especiación ecológica y la evolución dentro y entre poblaciones. Preservar esta es garantizar el
potencial genético evolutivo que puede constituir en mayor o menor grado, unidades de manejo para
la conservación, como una designación infraespecífica, para lograr una protección y uso sostenible
más objetivo aplicable a poblaciones bajo algún grado de amenaza o bajo explotación
(Crandall, 2000.).
Hemos comentado con anterioridad que la jutía conga no se encuentra en ninguna de las categorías
de amenaza de la UICN, pero que sin embargo es utilizada como recurso por parte de campesinos y
pescadores, haciendola perfectamente ajustable al criterio de categoria infraespecífica no
taxonómica, en este caso, de unidades de manejo para la conservación, como parte de un plan de
gestión integral, que contribuya a una mejor conservación del potencial evolutivo de la especie.
Tomando en consideración lo anteriormente planteado, nuestro objetivo fue crear, sobre la base de
las variaciones morfológicas (morfométricas y morfofisiológicas) entre las poblaciones de jutía
conga, un conjunto de criterios que contribuyan a conformar unidades de manejo para la
conservación en esta especie.
VII.2. Materiales y métodos
Se utilizó una muestra total de 768 ejemplares (417 hembras y 351 machos) de jutía conga,
pertenecientes a nueve poblaciones correspondientes a ocho localidades con disyunción de rango
geográfico (excepto en el área de Guanahacabibes) y hábitats diferentes. Los muestreos se realizaron
en los trimestres febrero, marzo y abril (FMA); mayo, junio y julio (MJJ); agosto, septiembre y
octubre (ASO) y noviembre, diciembre y enero (NDE).

148
En el área de la península de Guanahacabibeslas las capturas de jutía conga se realizaron en las áreas
de Bolondrón y Carabelita, considerándose como poblaciones independientes. Los datos de intestino
absoluto y grosor de la médula en las poblaciones de Zapata e Isla fueron obtenidos de trabajos
anteriores independientes a este estudio. Todos los ejemplares muestreados fueron adultos siguiendo
los criterios establecidos por Smith y Berovides (1984)
En el estudio se emplearon 16 variables morfológicas (10 morfométricas y 6 morfofisiológicas) que
se muestran a continuación:
1) Variables morfométricas 2) Variables morfofisiológicos
Largo cabeza tronco (LCT) Peso del hígado (PH)
Largo total (LT) Grosor de la médula renal (GM)
Peso corporal (PC) Largo del intestino (LI)
Indice de Robustez (Ro) Indice de nutrición relativo al LCT
Largo del pie (Pi) Indice intestinal relativo al LCT
Largo de la cola (Co) Indice renal relativo al LCT
Pie relativo al largo (PiR)
Cola relativa al largo (CoR)
Indice pie relativo la cola (Pi/Co)
Peso corporal relativo al LT
Las variables de medidas lineales se midieron con una cinta métrica y un Pie de Rey en milímetros
(mm) y las variables de peso con una balanza en gramos (g).
A todas las variables se les realizó la prueba de Kolmogorv –Smirnov para determinar el tipo de
distribución y la prueba de Levene de homogeneidad de varianza. A todas las variables se les efectuó
un criterio de selección, para determinar cuales eran las más idóneas para diferenciar poblaciones
como unidades de manejo para la conservación, que incluyó los siguientes aspectos:
1. Variables relativas que diferencian a las poblaciones por el hábito o el hábitat, que no se
encuentren influenciadas por el efecto del LCT de forma directa y que separen a las
poblaciones en función del valor adaptativo de las mismas.
149
2. Variables que muestren diferencias estadísticamente significativas entre poblaciones en cada
uno de los trimestres, así como que mantienen un mismo patrón cuando se consideran todos
los trimestres juntos.
Los ANOVAS de cada variable seleccionada se dieron en el capítulo 7, de los que se derivaron los
diagramas de caja para conformar grupos de poblaciones que no incluyen a sus medianas dentro de
los cuarteles (Whitlock y Schluter, 2009).
VII.3. Resultados y discusión
Las variables seleccionadas se exponen en las Tabla I (largos y peso), II (apéndices) y III
(morfofisiológico), con sus valores promedios y desviación típica, ignorando el efecto estación
(FMA y ASO). Según los resultados de los Anovas (Capítulo anteriores), las variables LCT, PC,
PC/LT, IRRL y LIU/LCT difieren entre poblaciones según el hábitat (bosque, vegetación xerofítica
y de manglar) con diferentes disponibilidades de nutrientes y de recursos hídricos. Las variables
CoR y Pi/Co también difieren entre poblaciones según los hábitos (terrestres, semiterrestres y
semiarborícola). Las variables en su conjunto mostraron un nivel elevado de concordancia entre los
valores promedios obtenidos por población, las agrupaciones poblacionales formadas en relación a
cada una de las variables y las condiciones ecológicas y hábitos de las poblaciones
La Tabla I. Valores promedios (X) y desviación típica (S) para los trimestres febrero, marzo y abril y
agosto, septiembre y octubre para el largo corporal, peso corporal e índice derivado del largo
y el peso en nueve poblaciones de jutía conga (Capromys pilorides).
Población Variables
LCT PC PC/LT
X S X S X S
Jardines 458.74 ab 33.40 2405.31a 532.55 3.38a 0.60
Sabana 456.77a 30.17 2811.44ab 576.83 4.09b 0.69
Zapata 465.14abc 44.83 3142.35bc 931.79 4.41b 0.98
150
Hatibonico 481.50bcd 40.74 3883.33de 845.35 5.15cde 0.85
Najasa 488.84cde 35.51 3509.11cd 818.19 4.62bc 0.86
Guanahacabibes I 499.67de 40.80 4315.87ef 975.73 5.69ef 0.96
Guanahacabibes II 483.40cd 30.94 3556.32cd 559.95 4.99cd 0.62
Saetía 512.16ef 37.93 4141.16ef 918.77 5.22de 0.87
Isla 525.61f 32.68 4592.90f 912.75 5.88f 0.90
Letras diferentes difieren estadísticamente a P<0.05 por una prueba de Tukey
La Tabla II. Valores promedios (X) y desviación típica (S) para los trimestres febrero, marzo y abril
y agosto, septiembre y octubre para los índices cola relativa y pie/cola en nueve poblaciones
de jutía conga (Capromys pilorides).
Población Variables
CoR Pi/Co
X S X S
Jardines 54.29e 4.79 36.45b 2.35
Sabana 49.60abc 3.30 39.94d 2.83
Zapata 50.92cd 3.43 39.51d 2.22
Hatibonico 55.11e 4.25 33.34a 2.47
Najasa 54.23e 4.84 36.95bc 3.10
Guanahacabibes I 50.41bc 3.97 38.78cd 2.44
Guanahacabibes II 47.01a 4.72 40.32d 4.10
Saetía 53.33de 2.81 36.08b 2.25
Isla 47.77ab 2.78 39.98d 3.00
Letras diferentes difieren estadísticamente a P<0.05 por una prueba de Tukey

151
La Tabla III. Valores promedios (X) y desviación típica (S) para los trimestres febrero, marzo y abril
y agosto, septiembre y octubre para los índices intestino (LIU/LCT) y riñón (IRRL) en nueve
poblaciones de jutía conga (Capromys pilorides)
Población IRRL LIU/LCT
X S X S
Jardines 32.23d 3.59 6.83e 0.06
Sabana 30.60cd 3.11 5.64d 0.75
Zapata 29.00 2.92 5.10 0.33
Hatibonico 28.41bc 2.75 5.31cd 0.61
Najasa 27.34b 3.95 4.68b 0.38
Guanahacabibes I 26.24b 3.56 5.04c 0.43
Guanahacabibes II 29.01bc 4.36 5.20c 0.34
Saetía 22.61a 2.56 4.24a 0.49
Isla 26.90 3.10 4.70 0.50
Letras diferentes difieren estadísticamente a P<0.05 por una prueba de Tukey
La distribución del largo cabeza tronco (LCT), peso corporal (PC) y el índice de distribución del
peso con respecto al largo total (PC/LT) se muestra en las Fig. 1A, 1C y 1E, para nueve poblaciones
de jutía conga. Las variables LCT, PC y PC/LT, presentaron un alto grado de asociación en todas las
poblaciones, lo cual ocasionó que las mismas se agruparan de forma similar (Figura 1B, 1D y 1F)
La variable LCT separó las poblaciones en tres grupos diferentes, que no incluyen a sus medianas
dentro de los cuartiles:
1) Poblaciones de tamaños pequeños como Jardines (hábito semiarborícola), Sabana (hábito
semiterrestre) y Zapata (hábito semiarborícola) de hábitat manglar, manglar mas vegetación
de costa de cayo y manglar de ciénaga con ecotono de bosque respectivamente.
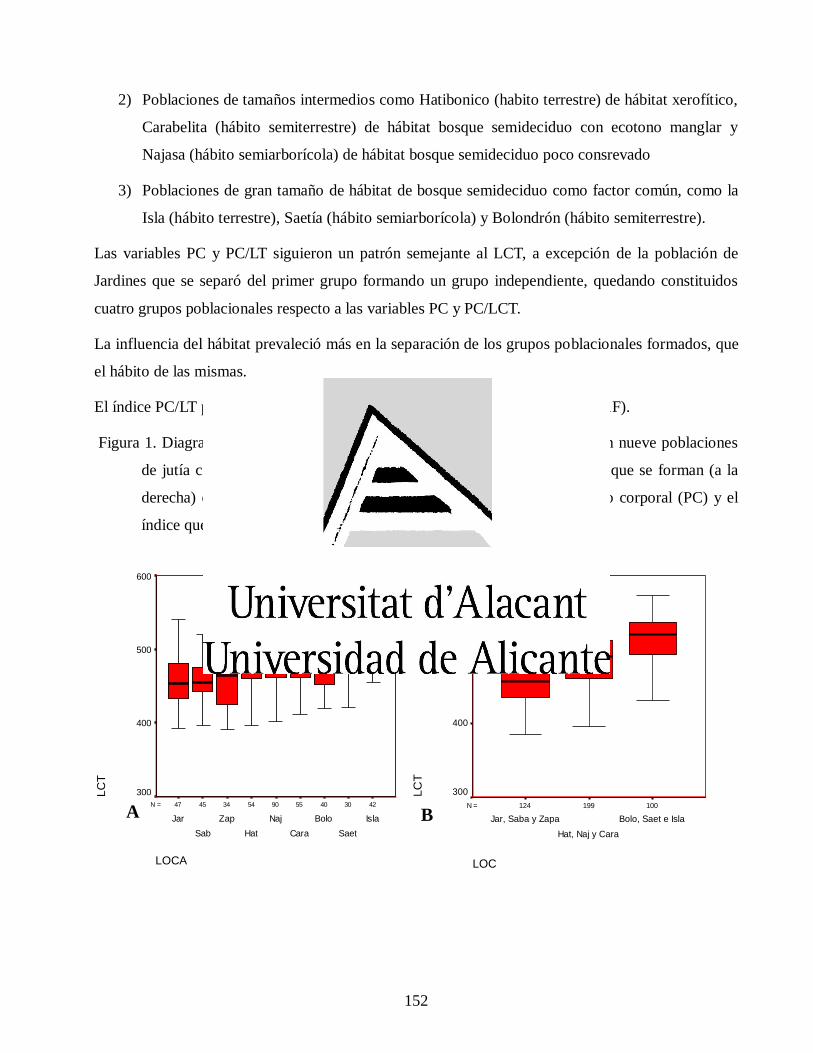
152
2) Poblaciones de tamaños intermedios como Hatibonico (habito terrestre) de hábitat xerofítico,
Carabelita (hábito semiterrestre) de hábitat bosque semideciduo con ecotono manglar y
Najasa (hábito semiarborícola) de hábitat bosque semideciduo poco consrevado
3) Poblaciones de gran tamaño de hábitat de bosque semideciduo como factor común, como la
Isla (hábito terrestre), Saetía (hábito semiarborícola) y Bolondrón (hábito semiterrestre).
Las variables PC y PC/LT siguieron un patrón semejante al LCT, a excepción de la población de
Jardines que se separó del primer grupo formando un grupo independiente, quedando constituidos
cuatro grupos poblacionales respecto a las variables PC y PC/LCT.
La influencia del hábitat prevaleció más en la separación de los grupos poblacionales formados, que
el hábito de las mismas.
El índice PC/LT presentó mayor similitud con el PC que con el LCT (Fig. 1D y 1F).
Figura 1. Diagrama de caja con la mediana, los cuartiles y valores extremos en nueve poblaciones
de jutía conga (Capromys pilorides) así como los grupos poblacionales que se forman (a la
derecha) en función de las variables largo cabeza tronco (LCT), el peso corporal (PC) y el
índice que relaciona peso corporal con respecto al largo total (PC/LT)
423040559054344547N =
LOCA
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
LC
T
600
500
400
300
100199124N =
LOC
Bolo, Saet e Isla
Hat, Naj y Cara
Jar, Saba y Zapa
LC
T
600
500
400
300
A B
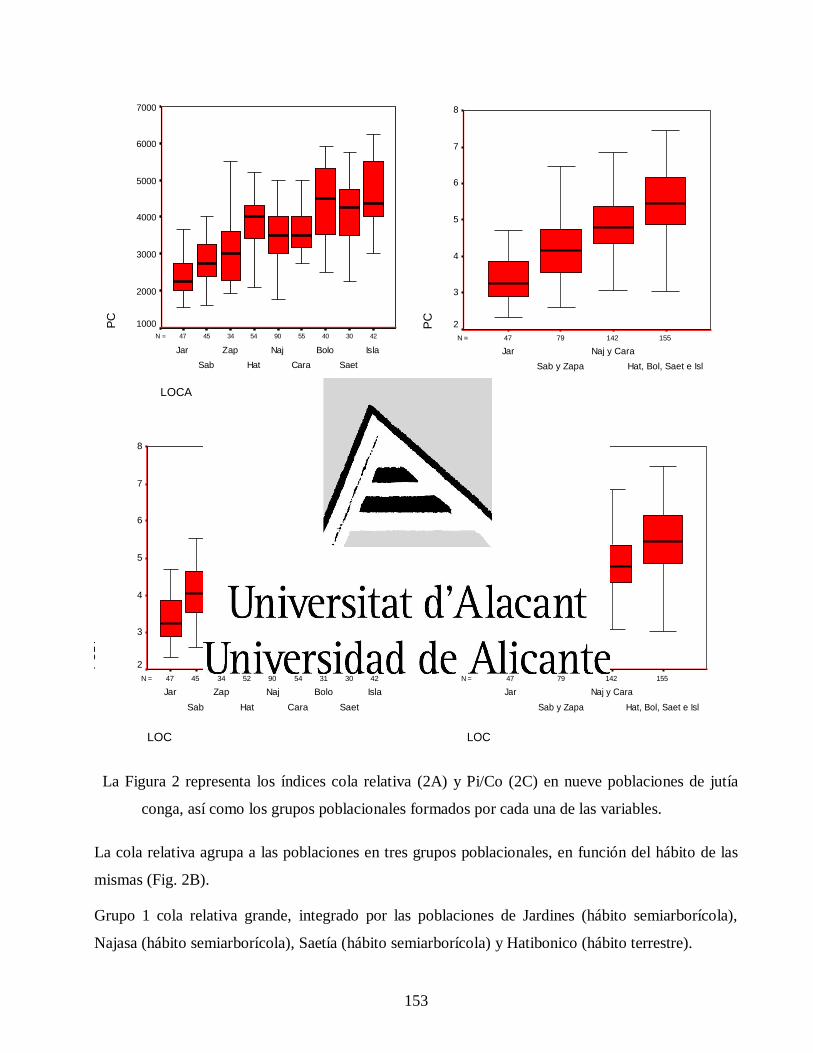
153
423040559054344547N =
LOCA
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
PC
7000
6000
5000
4000
3000
2000
1000
1551427947N =
LOC
Hat, Bol, Saet e Isl
Naj y Cara
Sab y Zapa
Jar
PC
8
7
6
5
4
3
2
423031549052344547N =
LOC
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
PC
LT
8
7
6
5
4
3
2
1551427947N =
LOC
Hat, Bol, Saet e Isl
Naj y Cara
Sab y Zapa
Jar
PC
LT
8
7
6
5
4
3
2
La Figura 2 representa los índices cola relativa (2A) y Pi/Co (2C) en nueve poblaciones de jutía
conga, así como los grupos poblacionales formados por cada una de las variables.
La cola relativa agrupa a las poblaciones en tres grupos poblacionales, en función del hábito de las
mismas (Fig. 2B).
Grupo 1 cola relativa grande, integrado por las poblaciones de Jardines (hábito semiarborícola),
Najasa (hábito semiarborícola), Saetía (hábito semiarborícola) y Hatibonico (hábito terrestre).
C D
E F

154
Grupo 2 cola relativa intermedia integrado por las poblaciones de Zapata (hábito semiterrestre) y
Bolondrón (hábito semiterrestre), cuya mediana no se separa de los cuarteles del próximo grupo.
Grupo3 cola relativa pequeña integrado por las poblaciones de la Isla (hábito terrestre), Carabelita
(hábito semiterrestre) y Sabana (hábito semiterrestre).
La población de Hatibonico a pesar de ser una población de hábito terrestre integra el grupo 1,
posiblemente relacionado con un reciente arribo geológico de la población en la localidad o una
adaptación a las altas temperaturas de la zona como apéndice termorregulador, según el principio de
Allen.o como órgano de reserva.
El índice pie/ cola, relativiza el pie en función de la cola, mostrando que a menor valor del índice,
más grande es la cola respecto al pie y por tanto es más elevada la actividad trepadora, agrupando a
las poblaciones con un patrón parecido al patrón de la cola relativa, pero a la inversa en cuanto al
valor numérico del índice y tres grupos poblacionales integrado de la siguiente manera.
Grupo 1. Integrado por la poblacion de Hatibonico, con bajo valor del índice
Grupo 2. Poblaciones de Jardines, Najasa y Saetía, con valores intermedios del índice
Grupo 3. Poblaciones de Sabana, Zapata, Carabelita, Bolondrón e Isla, con valores altos del índice
Figura 2. Diagrama de caja con la mediana, los cuartiles y valores extremos en nueve poblaciones de
jutía conga (Capromys pilorides), así como los grupos poblacionales que se forman (a la
derecha) en función de las variables cola relativa (CoR) e índice pie cola (Pi/Co)
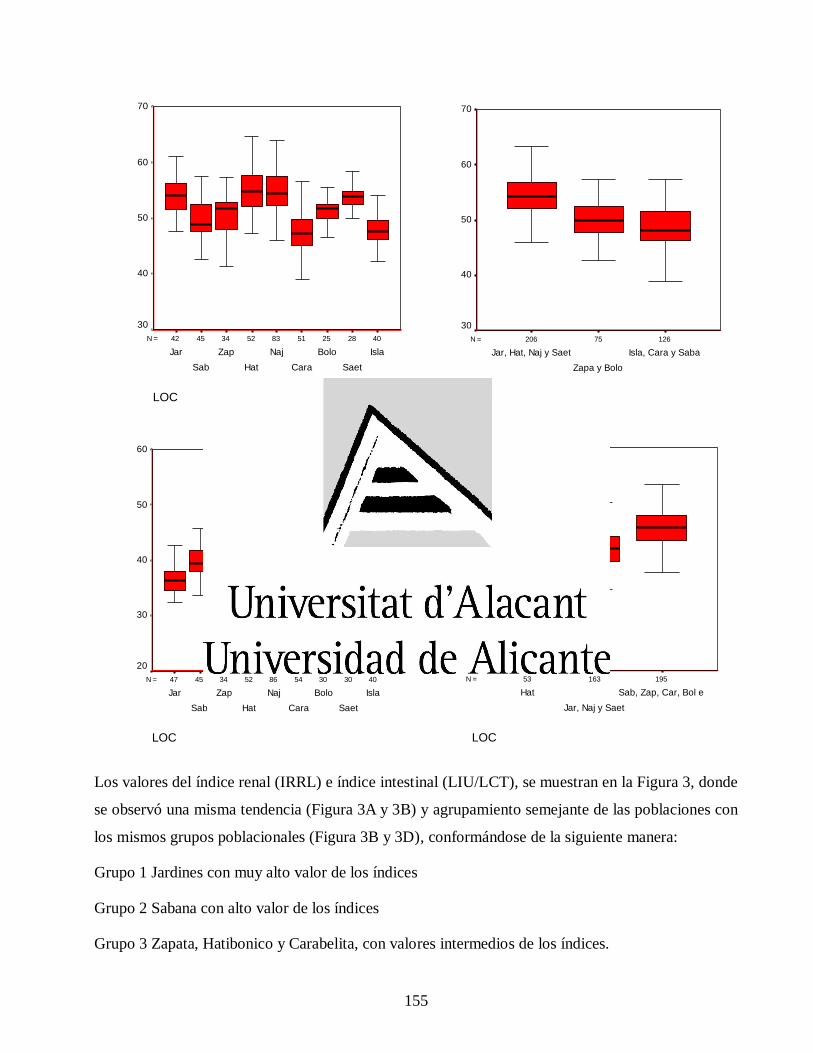
155
402825518352344542N =
LOC
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
CO
RE
LA
70
60
50
40
30
12675206N =
LOC
Isla, Cara y Saba
Zapa y Bolo
Jar, Hat, Naj y Saet
CO
RE
LA
70
60
50
40
30
403030548652344547N =
LOC
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
PIC
OLA
60
50
40
30
20
19516353N =
LOC
Sab, Zap, Car, Bol e
Jar, Naj y Saet
Hat
PIC
OLA
50
40
30
20
Los valores del índice renal (IRRL) e índice intestinal (LIU/LCT), se muestran en la Figura 3, donde
se observó una misma tendencia (Figura 3A y 3B) y agrupamiento semejante de las poblaciones con
los mismos grupos poblacionales (Figura 3B y 3D), conformándose de la siguiente manera:
Grupo 1 Jardines con muy alto valor de los índices
Grupo 2 Sabana con alto valor de los índices
Grupo 3 Zapata, Hatibonico y Carabelita, con valores intermedios de los índices.
A B
C D
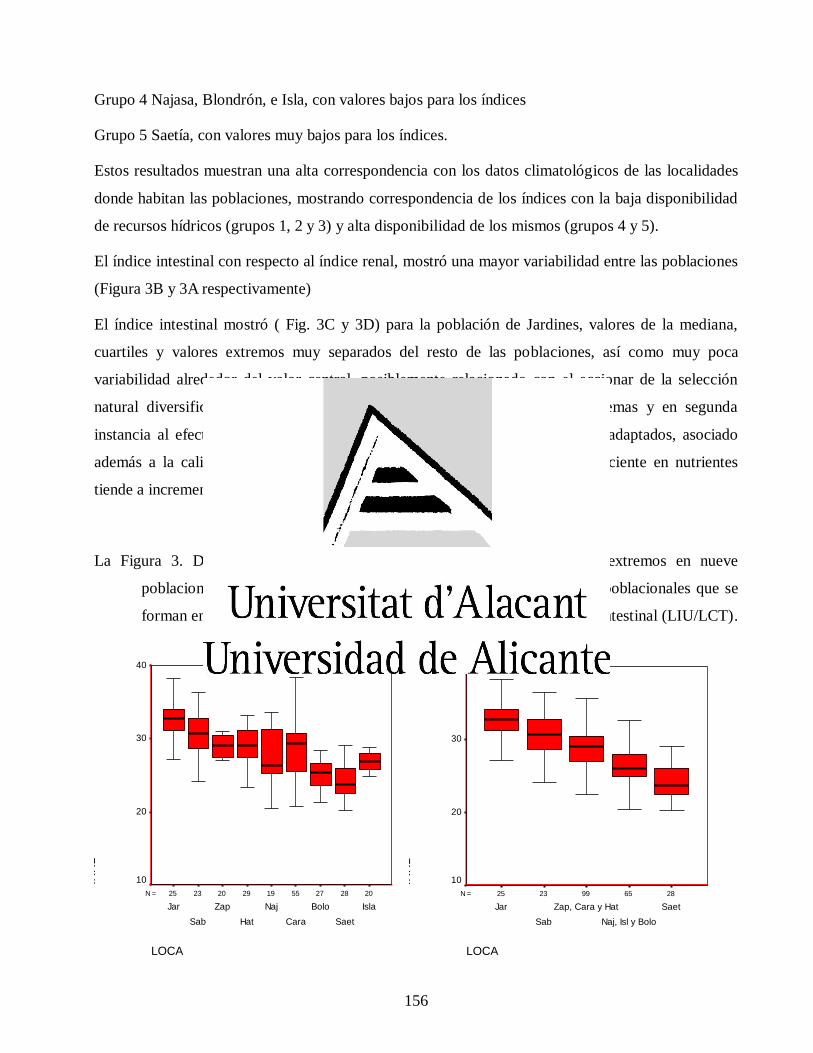
156
Grupo 4 Najasa, Blondrón, e Isla, con valores bajos para los índices
Grupo 5 Saetía, con valores muy bajos para los índices.
Estos resultados muestran una alta correspondencia con los datos climatológicos de las localidades
donde habitan las poblaciones, mostrando correspondencia de los índices con la baja disponibilidad
de recursos hídricos (grupos 1, 2 y 3) y alta disponibilidad de los mismos (grupos 4 y 5).
El índice intestinal con respecto al índice renal, mostró una mayor variabilidad entre las poblaciones
(Figura 3B y 3A respectivamente)
El índice intestinal mostró ( Fig. 3C y 3D) para la población de Jardines, valores de la mediana,
cuartiles y valores extremos muy separados del resto de las poblaciones, así como muy poca
variabilidad alrededor del valor central, posiblemente relacionado con el accionar de la selección
natural diversificadota en primera instancia, en hábitat de condiciones extremas y en segunda
instancia al efecto de la selección natural estabilizante sobre estos fenotipos adaptados, asociado
además a la calidad de la alimentación que como se conoce, cuando es deficiente en nutrientes
tiende a incrementar el tamaño de la cámara digestiva.
La Figura 3. Diagrama de caja con la mediana, los cuartiles y valores extremos en nueve
poblaciones de jutía conga (Capromys pilorides) así como los grupos poblacionales que se
forman en función del índice renal relativo al largo (IRRL) y del índice intestinal (LIU/LCT).
202827551929202325N =
LOCA
Isla
Saet
Bolo
Cara
Naj
Hat
Zap
Sab
Jar
IRR
L
40
30
20
10
2865992325N =
LOCA
Saet
Naj, Isl y Bolo
Zap, Cara y Hat
Sab
Jar
IRR
L
40
30
20
10
A B
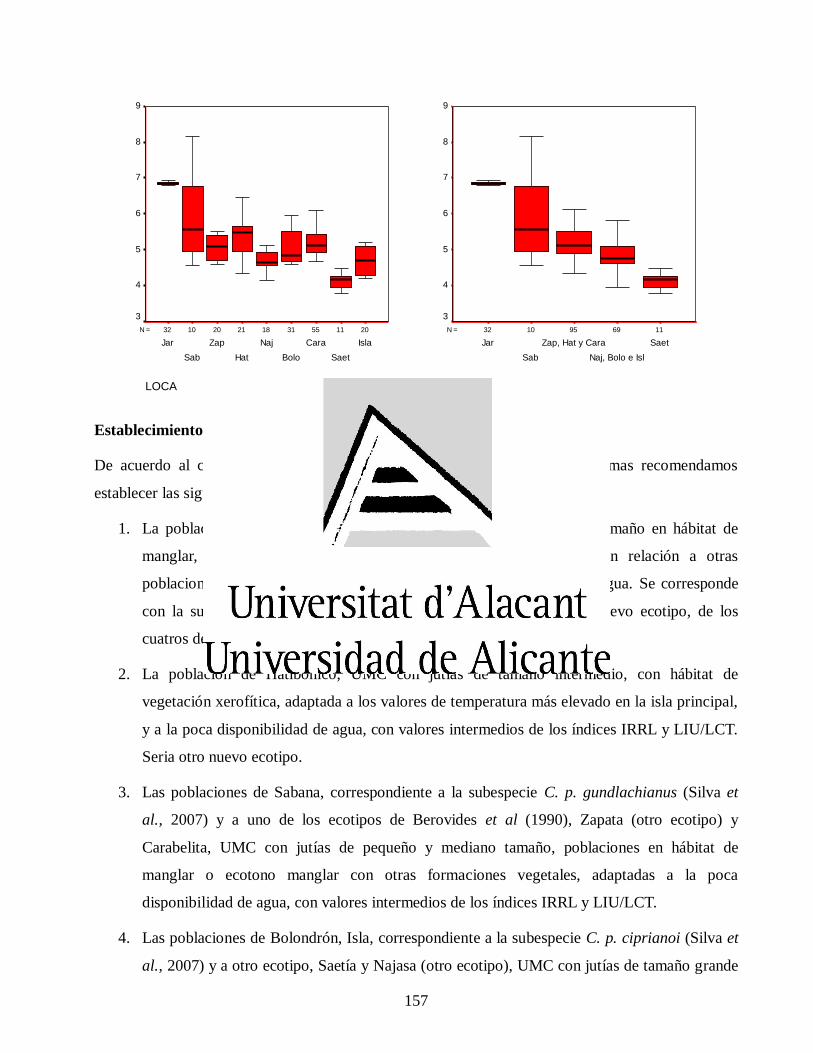
157
201155311821201032N =
LOCA
Isla
Saet
Cara
Bolo
Naj
Hat
Zap
Sab
Jar
IND
INT
ES
9
8
7
6
5
4
3
1169951032N =
LOCA
Saet
Naj, Bolo e Isl
Zap, Hat y Cara
Sab
Jar
IND
INT
ES
9
8
7
6
5
4
3
Establecimiento de las unidades de manejo para la conservación.
De acuerdo al conjunto de variables analizadas y los resultados de las mismas recomendamos
establecer las siguientes unidades de manejo para la conservación (UMC):
1. La población de Jardines de la Reina, UMC con jutías de pequeño tamaño en hábitat de
manglar, adaptada a la carencia de agua con valores extremos, en relación a otras
poblaciones de mangle, para los índices de retención y absorción de agua. Se corresponde
con la subespecie C. p. doceleguas (Silva et al., 2007) y sería un nuevo ecotipo, de los
cuatros descritos por Berovides et al (1990).
2. La población de Hatibonico, UMC con jutías de tamaño intermedio, con hábitat de
vegetación xerofítica, adaptada a los valores de temperatura más elevado en la isla principal,
y a la poca disponibilidad de agua, con valores intermedios de los índices IRRL y LIU/LCT.
Seria otro nuevo ecotipo.
3. Las poblaciones de Sabana, correspondiente a la subespecie C. p. gundlachianus (Silva et
al., 2007) y a uno de los ecotipos de Berovides et al (1990), Zapata (otro ecotipo) y
Carabelita, UMC con jutías de pequeño y mediano tamaño, poblaciones en hábitat de
manglar o ecotono manglar con otras formaciones vegetales, adaptadas a la poca
disponibilidad de agua, con valores intermedios de los índices IRRL y LIU/LCT.
4. Las poblaciones de Bolondrón, Isla, correspondiente a la subespecie C. p. ciprianoi (Silva et
al., 2007) y a otro ecotipo, Saetía y Najasa (otro ecotipo), UMC con jutías de tamaño grande
C D

158
o mediano, en hábitat de bosque, adaptadas a una mayor disponibiliudad de agua, con
valores bajos en los índices IRRL y LIU/LCT.
Nuestro agrupamiento que es solo preliminar pues se refiere solo a caracteres morfológicos,
coincides parcialmente con las subespecies reconocidas para la jutía conga (Silva et al., 2007) y para
los ecotipos de Berovides et al. (1990). Según el criterio de Crandall et al (2000) que es el que
seguimos, estas UMC se definen en términos de aislamiento ecológico (nichos ecológicos distintos,
traducidos en diferencias en adaptación local entre poblaciones) y genético. Nuestros resultados
apoyan la hipótesis que para la jutía conga se da la primera situación, pero la segunda, sólo se podrá
confirmar utilizando marcadores moleculares neutrales. En un análisis preliminar, utilizando ocho
marcadores moleculares proteicos, Berovides et al (1990) definieron tres grupos sobre esta base:
Isla, Sabana y Zapata-Najasa, lo que coincides parcialmente con nuestros resultados, pues el ultimo
grupo, en nuestros resultados esta separado.
Como señalan Fraser y Bernatchez (2001), el reconocimients de las UMC, solo será definitivamente
válido, cuando se consideren muchas variables de forma conjunta, entre ellas las morfológicas, pero
también las ecológicas y moleculares.

159
CAPÍTULO IX. Discusion general
Esta tesis ha tenido un enfoque evolutivo, utilizando como especie modelo la jutia conga Capromys
pilorides (Rodentia: capromidae) y siguiendo los planteamientos generales de Hendry et al (2011),
en relación a la aplicación de los principios evolutivos en la teoría y la práctica de la conservación
de especies.
Como señala Hendry et al (2011), la variación fenotípica determina como los organismos
interactúan con su ambiente y responden a las resultantes presiones selectivas, dando lugar en
muchos casos a la adaptación. Este fue el objetivo central de la presente tesis, analizando caracteres
fenotípicos relativo a la morfología (caracteres morfométricos) y fisiología (caracteres
morfofisiológicos), en relación a sus respuestas a la selección natural al nivel de individuo y en
específicos a los agentes selectivos clima, composición y estructura a la vegetación y disponibilidad
de agua, respuestas que fueron positivas en función de los tipos de hábitat, que diferían en la
intensidad de la acción de dichos agentes selectivos, provocando adaptación local. Sin embargo, no
queda descartado por completo, el efecto aleatorio de la deriva genética en algunos casos.
La adaptación local registrada en los resultados, en esencia entre hábitats de bosque y hábitats de
manglares, puede ser causada por diferencias genéticas, mayormente por muchos genes estructurales
y reguladores, específicos para cada adaptación, así como plasticidad fenotípica (también de origen
genético), cambios epigenéticos, efecto materno y otras formas de herencia no genéticas
(Bundurianky y Day, 2009), aspecto no estudiado en la presente tesis, pero puede postularse como
hipótesis, que las adaptaciones aquí registrada son mayormente producidas por plasticidad
fenotípica, dado lo rápido que este especie de jutia se adapta a variadas condiciones naturales no
típica y al cautiverio. Dicha rapidez indica más Plasticidad fenotípica que cambios en la
constitución genética, pues esta última es un proceso que requiere más tiempo. (Hendry et al.,
2011).
En relación a la conservación de species, Hendry et al (2011) destacan como la reducción del
tamaño de las poblaciones naturales reduce la variabilidad genética de estas y su adaptación, luego
el conocimiento de que caracteres son adaptativos, como se hace en esta tesis, es de relevancia para
la conservación de las especies. Pero además como se demostró al revelar adaptación local entre las
poblaciones estudiadas, se pueden elaborar planes de conservación que tengan en cuenta dicha
adaptación, en específicos definiendo que población o conjunto de población, conforman unidades

160
de manejo para dicha conservación u otras equivalentes, como realmente se pudo hacer con la
especie estudiada.
CAPÍTULO X. Conclusiones
1.1.1. Las poblaciones de hábitat de bosque presentaron un mayor largo cabeza tronco
(LCT), peso corporal (PC), e índices derivados del peso como la Robustez, Condición
Física y peso por milímetro lineal, con respecto a las poblaciones de hábitat de manglar.
1.1.2. Los índices pie y cola relativos se relacionaron con los hábitos de la especie dentro de
cada localidad, con un mayor o menor valor de los mismos de acuerdo al grado mayor o
menor de arboricidad respectivamente, con un mejor indicador de la actividad trepadora
para el índice cola relativa. Los índices cola relativa y pie/ cola constituyen las mejores
variables que diferencian a las poblaciones asociadas a la actividad trepadora.
1.1.3. Las variables LCT, PC y peso/mm lineal mostraron el mayor dimorfismo sexual
dentro de cada población.
1.1.4. Las variables LCT, PC, peso/mm lineal y Pie relativo presentaron una dependencia
estadísticamente significativa con el índice climático de Martonne, que está relacionado
con la precipitación, temperatura y la productividad primaria del hábitat.
1.1.5. Los índices renales empleados (grosor de la medula con respecto al grosor del riñón y
grosor de la médula con respecto al largo cabeza-tronco LCT) tuvieron mayores valores
promedios para las poblaciones de hábitat manglar y vegetación xerofítica, con una
mayor exactitud y correspondencia entre los valores promedio del índice y las
características del hábitat para el índice renal relativo al LCT.
1.1.6. El índice nutricional no mostró diferencias estadísticamente significativas entre
poblaciones con hábitat diferentes, pero si una interacción cruzada significativa
localidad por trimestre del año.
1.1.7. El índice del peso del hígado relativo al largo corporal, reflejo un gradiente que
separa a las poblaciones de hábitat bosque con respecto a las poblaciones de hábitat
manigua costera y manglar, así como una alta asociación entre este índice de nutrición y
el índice climático de Martonne.

161
1.1.8. El largo absoluto de la última porción del intestino grueso y su índice derivado
diferenciaron a las poblaciones de bosque con respecto a las de hábitat xérico.
1.1.9. En el área geográfica de la península de Guanahacabibes, las poblaciones de
Bolondrón y Carabelita presentaron variaciones entre localidad estadísticamente
significativas en correspondencia con el hábitat.
1.1.10. Las poblaciones estudiadas a través de las variables seleccionadas se diferenciaron en
cinco grandes grupos que pudieran ser catalogadas como unidades de manejo para la
conservación.

162
Bibliografía
Adler, G.H; R. Levins.1994. The island syndrome in rodent populations Quart. Rev. Biol. 69:473-
490.
Abreu, R. M., y N. Manojina. 1989. Caracterización morfológica de Capromys pilorides (Rodentia:
Capromydae) de la Península de Guanahacabibesen Pinar del Río.Peyama, 372:1-14.
Al-Kahtani, M., C. Zuleta and E. Caviedes. 2004 ; Kidney mass and relative medullary thickness of
rodents in relation to habitat, body size and phylogeny ; Physiol.Bioch. Zoo ; Vol. 77 ; 346-
365.
Amori, G; S. Geppoliti 2003. A higher taxon approvch to rodent conservations prioritres for the 21st
century. Animal Biodiversity Conservation 26 :1-18.
Anderson, S., Ch. A. Woods, and G. S. Morgan.1983. Geocapromys brownii. Mammalian Species.
The Amer. Soc. Mammalogists, 201: pp. 1-5.
Aundesirk, T., G. Aundesirk.1996. Biología. La Vida en la Tierra. Prentice-Hall Hispanoamericana,
S. A. Mexico. 947 pp.
Barbour, T.1945. Appedix. En A naturalist in Cuba, Little, Brown, Boston, pp. 287-302.
Barraowclough, G. F., and N. R. Flesness. 1996. Species, Subspecies, and Races : The problem of
Units of Management in Conservation. En: Wild Mammals in Captivity. Kliman, G., M. E.
Allen, K. V. Thompson, and S. Lumpkin. The Univ. Chicago Press, Chicagopp., 247-254.
Beck. M.W. 1995. Size-specific shelter limitation in stone crabr: a test of the demographic
bottleneck hypothesis. Ecology 76- 968-980.
Begón, M., J. L. Harper and C.R. Townsend. 1998. Ecology. Individuals, Populations and
Community. Blackwell Sci. Puvl., Boston, 1068 pp.
Berovides; V., O. Pimentel, M. Cañizares, J. Castillo, y M. Morales. 2009. Estudios de la Jutía
Conga (Capromys pilorides) como Recurso Natural. Publicaciones Univ. Alicante, España,
143 pp.
Berovides, V. y A. Comas. (1997a). Variaciones de la coloración del pelaje en la jutía conga
Capromys pilorides. Ciencias Biológicas 11:3-9.

163
Berovides, V. y A. Comas. (1990). Patrones conductuales de la jutía conga Capromys pilorides en
cautividad I. Conductas individuales y actividad. Biología IV.2:123-121.
Berovides, V. y A. Comas. (1991). The critical condition of hutias in Cuba. Oryx 25:7-14.
Berovides, V. y A. Comas. (1993). Valoración de la jutía conga, Capromys pilorides (Rodentia,
Capromidae), como recurso natural. Biología 7(2-3): 125-138.
Berovides, V. (1987). Evaluación de los caprómidos de Cuba como recurso natural. Rev. Flora,
Fauna y Áreas silvestres. 2,5:3-14.
Berovides, V. (1998). Estructura familiar y abundancia de la Jutía Conga (Capromys pilorides). Rev.
Biología. 12:81-82.
Berovides, V. (2005) Variaciones morfofisiológicas en poblaciones de jutía conga, Capromys
pilorides, en hábitat de bosque y manigua costera. CUBAZOO, 13: 11-15.
Berovides, V. 2006. Biología Evolutiva. Pueblo y Educación, La Habana, 407 pp.
Berovides, V. 2008. Patrones conductuales de la jutía conga (Capromys pilorides) observadas en
cautiverio. CUBAZOO, 18: 57-59.
Berovides, V. y Gutieres, A. (1999). Grado de heterocigocidad y peso corporal en la jutía conga,
Capromys pilorides, (Rodentia, Capromyidae). Biología, 13.1: 59-60.
Berovides, V., A. Comas. 1997. Abundancia de la jutía conga (Capromys pilorides) en varios
hábitats de Cuba. Rev. Biología 11: 25-30.
Berovides, V., R. Borroto, A. Camacho. 1990: Biología sexual del género capromys (Rodentia,
Capromyidae). Revista biología 1: 21-32.
Berovides, V., y J. L. Gerhartz .2007. Diversidad de la Vida y su Conservación Editorial Científico
Técnica, La Habana, 98 pp.
Berovides, V., y M. A. Alfonso. 1987. Densidad y biomasa en poblaciones de jutía (Capromys sp.)
Cien. Biol., 18:127-130.
Berovides, V., y O. Pimentel. 2000. Densidad y coexistencia de dos especies de roedores
caviomorfos en el área protegida Mil Cumbres, Pinar del Río, Cuba. Rev. Biol., 14(1): 22-26.
Berovides, V: Pimental, O: Cañizares, M: Castillo, J: Morales, M: 2009. Estudios de La jutís conga
(Capromys pilorides) como recurso natural. Publicaciones Univerdidad de Alicante. 143 pp.

164
Berovides, V; A. Camacho; A. Comas; R. Borroto. 1990b. Variación ecológica en poblaciones de la
jutía conga, Capromys pilorides (Rodentia capromyidae). Ciencias Biológicas 23: 44-58.
Berovides, V; A. Comas. 1997a. Densidad y productividad de la jutía conga (Capromys pilorides)
en manglares cubanos. Caribb. J. Sci. 33: 121-123.
Berovides, V; M. A. Alfonso; A. Camacho. 1990ª. Variabilidad morfológica de la jutía conga
Capromys pilorides (Rodentia, Capromyidae) de Cuba. Doñana Acta Vertebrata. 17: 122-127.
Berovides, V; M. Morales; J. Castillo.2007. Estudio del uso de la jutía conga (Capromps pilorides)
por los pescadores de Carahata, Villa Clara. CUBAZOO 17:59-64.
Beuchat, C. A. 1996. Structure and concentrating ability of the mammalias kidney: correlations with
habitat. Am. J. Physiol. 271: 157-179.
Blake, B. H. 1997. The effects of the kidney Structure and the animal cycle on water requirements
in golden-mantled ground squirrels and chipmunks. Comp. Biochem. Physiol. 584: 413-419.
Boggs, D. F., and P. B. Frappell. 2000. Unifying principles of locomotion: Fareword. Phisiol.Bioch.
Zool. 73: 647-650.
Book hout, T. A., (Editor) 1994. Research and Management Techniques for Wildlife and Habitats.
The Wildlife Soc. Bethesda, Maryland. 740pp.
Borhidi, A. 1996. Phytogeografy and Vegetation Ecology of Cuba. Akademia Kiado, Budapest. 923
pp.
Bozinovic, F; J.M. Rojas; P.A. Gallardo; R.E: Palma; E Gianoli .2011. Body mass and water
economy in the South American olivaceus field mouse alohg a latitudinal gradient:
Implications for climate change. J. Arid Enviroment 75: 411-415.
Braun, C. E., (Editor) 2005. Research and Management Techniques for Wildlife and Habitats. The
Wildlife Soc. Bethesda, USA.. 974pp.
Brown, J. H. 1995. Macroecology. Univ. Chicago Press, Chicago, 250pp.
Brown, J. H., and B. A. Mawcer. 1989. Macroecology: the division of food and space among
species on continents. Science 243: 1145-1150.
Brownfield, M. S., and B. A. Wunder .1976. Relative medullary area; a new index for estimating
urinary concentrating capacity of mammals. Comp Bioch. Phisiol. 55 A: 64-75.

165
Bucher, G. C. (1937). Notes of life history and habits of Capromys. Mem. Soc. Cubana Hist.
Bunduriansky, R; T. Day. 2009. Nongenetic inheritance and its evolutionary implications. Annual
Review Ecol. Evol. Systematic 40: 103-125.
Bush, V., and C. E. Adams. 2007. Using phenotypic variation to determine conservation value:
application of a novel approach to artic charr. Ecology of freshwater fish. 16: 29-33.
Calder, W. A. 1984. Size Function and Life History. Harvard Univ. Press, Cambridge, Mass, 564pp.
Campbell, M. T., and N. A. Slad. 1993. Effect of mass seasonal survivorship of northern cotton rats.
J. Mamm. 74: 921-981.
Cañizares, M. 2003. Selección de microhábitat por la jutía con (Capromys pilorides) y su uso como
recurso natural en la Reserva Ecológica Alturas de Banao. Tesis en opción al Grado
Académico de Master en Ciencias, Instituto de Ecología y Sistemática, CITMA, 90pp.
Cañizares, M., Berovides, V. 2007. Uso de refugio diurno como factores reguladores naturales de
las poblaciones jutía conga. CUBAZOO, 17: 9-16.
Capote, R. Berazaín. 1985. Clasificación de las formaciones vegetales de Cuba. Rev. Jardín
Botánico 2: 27-56.
Carrizo, L.V; Tilli, M.J; Dos Santos, D.A. 2014. Interplay between postcraneal morphology and
locomotor types in Neotropicalsigmodontinae rodents. J. Anat. 224: 469-481.
Carrizo, L.V; Tilli, M.J; Dos Santos, D.A. 2014. Interplay between postcraneal morphology and
locomotor types in Neotropicalsigmodontinae rodents. J. Anat. 224: 469-481.
Cawghley, G., and C, J. Gunn. 1996. Are big mammals simply little mammals writ large?
Oecologia, 59: 7-17.
Cervantes, F. A. 1991. Supporting the study of the intraspecific variation life-history strategies of
tropical mammals. Anales Inst. Biol. Univ. Auton. Mexico. Sec-Zool., 62:365-372.
Charmantier, A., and D. Garant. 2005. Envioronmental quality and evolutionary potential: lessons
from wild populations potential: Proc. R. Soc. London, Ser. B. 272: 1415-1421.
Cherry, S. 1996. A comparson of confidence interval methods for habitat use-availabiliity studies. J.
Wild Mang., 60: 653-658.
Clough, G. C. 1972. Biology of the Bahamas hutia. Geocapromys ingrahami.

166
Comas A., F. Rosales y R. Gonzáles. 1989b. Datos sobre la alimentación de la jutía conga en el
Área Protegida Sierra del Chorrillo. Biología 1: 3-28.
Comas A., R. González, G. Cepero, y V. Berovides. 1989a. Densidad de la jutía conga Capromys
pilorides (Rodentia: Capromydae), en el área protegida Sierra del Chorrillo, Camaguey. Cien.
Biol., 21/22: 115-129.
Comas, A. y V. Berovides. 1990. Densidad de la Jutía Conga en los cayos del grupo insular Jardines
de la Reina, Cuba. Biología 1: (11-18).
Comas, A., y V. Berovides. 1994. Estudio de la jutía conga como Capromys pilorides, recurso
natural. 3er Simposio de Zoología, La Habana, Resumenes, pp. 145-146.
Comas, A.; F. Rosales, R. Gonzáles y M. Peláez. 1989c. Ecología trófica de la jutía conga
Capromys pilorides, (Rodentia; Capromyidae); en el área protegida Sierra del Chorrillo,
Camaguey, Cuba. Biología 8 (75-79).
Comas, A.; R. Gonzáles, U. Peláez, y V. Berovides. 1994b. Patrones conductuales de la jutía conga
Capromys pilorides, (Rodentia; Capromyidae), en cautividad. II. Conductas sociales y
reproductivas. Rev. Biol. 8: 65-74.
Cornide, R. I., C. Ferrera, C. Jiménez, J. A. Valdivia, y A. M. Gutiérrez. 1987. Hallazgo de
Mycobacterium (M. Tuberculosis y complejo avium-intracellulare) en la jutía conga
(Capromys pilorides). Misc. Zool., 35: 1-2.
Crandall, K.A. ; U.R, Bininda-Emonds.; G. M. Mace; R.K. Wayne. 2000. Considering evolutionary
processes in conservation biology. Trends Ecol.Evol, 15: 290-295.
Crandall, K. A. 2009. A multifaceted approach to species conservation. Anim. Conserv., 12: 105-
106.
Croft ; D.A; Niemi, K; Franco, A. 2011. Incisor morphology reflects diet in caviomorph rodents. J.
Mammal.92: 871-879.
Croft; D.A; Niemi, K; Franco, A. 2011. Incisor morphology reflects diet in caviomorph rodents. J.
Mammal.92: 871-879.
Cuthill, I. C., and I. A. Houston. 1997. Manoging time and energy. En: Kiebs, J. R., and N. Davies
(Ed.). Beharvior ecology: evolutionary approach. Blackwell, pp 97-120.

167
Dagault, N. y M. Suboreau. 1990. Characteristic of the reproductive function of the Male Coypu
(Miocastor coypus) in the area of the Poitou marshlands. Can. J. Zool: 219:1584-1589.
Dale, J; P.O. Duna; J. Figuerola. 2007. Sexual selection explain Rench rule of allometry for sexual
size dimorfism. Proc. R. Soc. B. 274: 2971-2979.
Davis, D. E., and O. Hall.1951. The seasonal reproductive condition of female Norway (Brown)
rats in Baltimore, Maryland. Physiol. Zool., 24: 9-20.
Dennison, M. D., and A. J. Baker. 1991. Morphometric variability in continentaland atlantic island
population of chaffinches (Fringilla coelebs). Evolution 45: 29-39.
Díaz Cisneros, L. R. 1989. Regionalización climática general. VI. 44 (Mapa 55). En: Nuevo Atlas
Nacional de Cuba. Inst. geografía, Academia de Ciencias de Cuba.
Diaz, G. 2001. Ecofisiología de pequeños mamíferos de las tierras áridas de Argentina:
adaptaciones renales. Mastozoología Neotropical 8:173-174.
Díaz, G. B., y R. A. Ojeda. 1999.Kidney structure and allometry of argentine desrt rodents. J. Arid.
Environ. 41:453-461.
Diaz, G.B., R.A. Ojeda, E.L. Rezende. 2006. Renal morphologi, phylogenetic history an desert
adaptation of South American Hystricognath Rodents. Functional Ecology; Vol. 20; No 609-
620.
Dytham, C. 2003. Choosing and Using Statistics. A Biologist´s Guide. Blackwell Publ. Malden.
MA., USA.
Ebensperger, L. A., and H. Cofre. 2001. On the evolution of group-living in the new Word cursorial.
Behav. Ecol. 12: 227-236.
Elissamburu, A; S.F.Vizcaíno 2004. Limb proportions and adaptations in caviomorph rodents. J.
Zoology 262: 145-159.
Ellis, B. A., J. N. Mills, and Ej. T. Kennedy.1994. The relationship among diet, alimentary tract
morphology, and life history for five species of rodents from the central Argentina. Acta
Theriol. 39: 345-355.
Emlen, S. T; L. W. Oring. 1977. Ecology, sexual selection, and the evolution of mating system.
Science. 197: 215-223.

168
FAO-PNUMA. 1985. Manejo de la fauna silvestre y desarrollo rural. Oficina regional de la FAO
para América Latina y el Caribe. Doc. Técnico 2. 161 pp.
Fleming, T. H. 1975. The role of the small mammals in tropical ecosystems. En: Small mammals
Golley F. B., K. Petrosewicz, and L.Ryszkowski (Eds.). Cambridge Univ Press, Cambridge,
pp., 269-298.
Fleming, Th. 1974. the population ecology of the species of Costa Rican Heteroyid rodents.
Ecology 55: 493-510.
Fraser, D.J; L.Bernatchez. 2001. Adaptive evolutionary conservations: towards a unified concept for
defining conservations units. Molecular Biology 10: 2741-2752.
Freehling, M., and J. Moore. 1987. A comparison of two techniques for measuring intestinal length.
J. Wildl. Manage. 51: 108-111.
Frías, A., N. Hernández, y R. Carnero. 1987. Datos reproductivos de dos formas de jutía conga,
Capromys pilorides (Rodenthia: Caviomorpha). Poeyana 345:1-6.
Fuente de la, J. L., V. Berovides. 2009. Morfometría comparada en dos poblaciones de jutía conga
(Capromys pilorides) que viven en hábitat de manglares. CUBAZOO 1: 63-66.
Fuente de la, J. L., V. Berovides. En prensa. Variación Estacional de un Indice Nutricional en Cuatro
Poblaciones de Jutía Conga (Capromys pilorides). CUBAZOO.
Fuente de la, J. L; V. Berovides. 2008. Comparación de un índice de longitud intestinal en seis
poblaciones de jutía conga capromys pilorides (Rodentia: capromyidae). Mesoamericana 12:
33-37.
Fuente de la, J. L; V. Berovides. 2009. Morfometría comparada en dos poblaciones de jutía conga
(Capromys pilorides) que viven en hábitat de manglares. CUBAZOO 1: 63-66.
Fuente de la, J. L; V. Berovides. 2011. Variación Estacional de un Indice Nutricional en Cuatro
Poblaciones de Jutía Conga (Capromys pilorides) que viven en hábitat de manglares.
CUBAZOO 23: 75-80.
Fuente de la; V. Berovides. 2013. Comparación de dos índices de reabsorción de agua en tres
poblaciones de Capromys pilorides (Rodentia: Capromydae) Rev. Colombiana Cienc.Anim.
5(2): 259-269.

169
Fuente, de la; V. Berovides. 2015. Variaciones morfológicas en poblaciones de jutía conga
(capromys pilorides), durante la estación de seca CUBAZOO 28: 44-55.
Fuente.de la, A.; V. Berovides y A. Camacho. (1992). Variación intrapoblacional de la jutía conga,
(Capromys pilorides) en el área protegida Sierra del Chorrillo, Camaguey, Cuba. Biología
6:3:183-189.
Gaines, M. S. 1985. Genetic. In: R.H. Tamarin (edit.). Biology of New World Microtus. Spec. Publ.
No. 8, Soc. Mamm. Pp. 845-883.
Geluso, K. N. 1978. Urine concentrating ability and renal structure of insectivorous bats. J.
Mammal 59: 312-323.
Gomes, H.2015. The great variety of dental structure in rodents: new insight into their ecological
diversity. In: Evolution in the rodents (eds. P.G. Cox, L Hauitier; PP 424-447) Cambridge
Univ. Press. Uk.
Gomes, H.2015. The great variety of dental structure in rodents: new insight into their ecological
diversity. In: Evolution in the rodents (eds. P.G. Cox, L Hauitier; PP 424-447) Cambridge
Univ. Press. Uk..
Gosling, L. M; Huson, G. C. Addison. 1980: Age estimation of coypus (Myocastor coypus) fom eye
lens weight. J. Applied Ecology 17: 641-647.
Gould, S. J. 1980. Is e new and general theory of evolution emerging. Paleantology 6: 19-130.
Gould, S. J. And R. F. Jonson. 1972 Geographic variation. Ann.Rev. Ecol. Syst. 3: 457-498.
Green, D. M. 2005. Designable uniits for estatus Assessment of endangered species. Consevation
Biology 19: 1813-1820.
Gregor, D. H. 1975. Renal capabilities of an argentine desert armadillo. J. Mammalogy 56: 626-
632.
Gross, J. E., Z. Wang, and B. A. Wunder. 1985. Effects of food quality and energy needs: changes in
gut morphology and capacity of Microtus ochrogaster. J. Mammal. 66: 661-667.
Gurdlach J, 1877. Contribución a la Mamalogía Cubana. Imprenta Mortrel, La Habana, 53 pp.
Gutierrez, A., A. Coma, V. Berovides. 1999. Dinámica reproductora de la jutía conga Capromys
pilorides (Rodentia, Capromyidae) en diferentes hábitats. Rev. Biología 13: 125-128.

170
Gutierrez, A., y V. Berovides. 1995. Efectos morfométricos, genéticos ambientales, sobre
rendimiento en canal de la jutía conga (Capromys pilorides). Vida Silvestre Neotropical, 4(I):
28-31.
Hairston, N.G; F.E. Smith; L. B. Slobodkin. 1960. Community structure, population control and
competition. Am. Nat. 94: 421-425.
Hanski, I. A., O. E. Gogiotti. 2004. Ecology, Genetic and Evolution of Metapopulations. Academic
Pres, Londres. 650 pp .
Hansson, L.1985. Geographic differences in bank voles Clethriomomys glareolus in relation to
ecogeographical rules and possible demographic and nutritive strategies. Ann. Zool. Fennic.,
22: 319-328.
Heisinger, J. F., T. S. King, and H. W. Halling. 1973. Renal adaptations to macro-and microhabitats
in the family cricetidae. Comp. Bioch. Phisiol. 44ª 767-774.
Hendry, A. P; M. T. Kinnison; M. Heino. 2011. Evolutionary principles and their practical
application. Evolutionary Applications, Blackwell Pub: 159-183.
Hernández, E. 2003. Elementos de manejo en vida libre y cautiverio de las jutías conga en el valle
de San Andrés. Tesis en opción al titulo de ingeniero forestal. Universidad de Pinar del Río.
87 pp.
Hernández, F. R., J. L. Linares, R. Sotolongo. 2005. Densidad y distribución de la jutía conga
Capromys pilorides (Rodentia: Capromidae) en la Península de Guanahacabibes, Cuba. Rev.
Electrón. Veter. REDVET, 6(9): 1-8.
Hernández, F. R., y O. Pimentel 2005. Enfermedades, prásitos y depredadores de la jutía conga
(Capromys pilorides) en el macizo forestal central de la cordillera de Guaniguanico. Rev.
Electrón. Veter. REDVET, 6 (8): 7 páginas no numeradas.
Hernández, N. 1985. Primeras observaciones sobre conducta sexual en machos de Capromys
pilorides. Misc. Zoológicas. 4p.
Heywood, V. H; y R. T. Watson. 1995. Global Biodiversity assessment. UNEP, Cambridge Univ.
Press. 17-22.

171
Hoekstra, H. E; J.G.Krenz; M.W. Nashman.2005. Local adaption in the rock poket mouse
(chaetodipus intermedius); natural selection and phylogenetic history of populations. Heredity
94: 217-218.
Hune, I. 1989. Reading the entrails of evolution. New. Scientist1203: 43-47.
Isaac, J.L. 2005. Potencial cause and life-history consequences of sexual size domorfhism in
manmals. Manmal rew. 1: 101-115.
IUCN (International Union for the Conservation of Nature and Natural Resources) 2006 Red list of
threatened species. The IUCN Species Survival Commission (CD), Cambridge. IUCN/SSCV
12, Gland, Zuiza. 135 pp.
Jackson, T. P., N. C. Bennett, and A. C. Spinks. 2004. Is the distribution of the arid occurring
otomyne rodents of southern Africa related to physiological adaptation or refuge type?
Zoology 264: 1-10.
Janzen, D. H., and D. E. Wilson 1983. Mammals. En: Costa Rican Natural History. Janzen, D. H.,
(Eds.). Univ. Chicago Press, Chicagopp., 426-442.
Jenning, S., J. A. Oliveira, and K. J. Warr. 2007. Measurement of body size and abundance in test
of macrocological and food web theory. J. Animal Ecol. 76: 72-82.
Johnson, D. H. 1999. Insignificance of statistical significance testing. J. Wild Manage., 63: 763-
772..
Johnson, M. L., R. H. Taylor and W. Winnick. 1975. The breeding and exhibition of Capromys
rodent at Tacoma zoo. International Zool. Yearbook: 15:53-56.
Jorri, F. 2001. La cría de roedores tropicales (Thryonomys swinderianus y Atherurus africanus)
como fuente de alimento en Gabón, Africa Central. Tesis doctoral, Fac. Veterinaria, Univ.
Autónoma de Barcelona, España. 21pp.
Karasov, W. H., and J. M. Diamond. 1983. Adaptative regulation of sugar and amino acid transport
by vertebrate intestine. Am. J. Physiol., 245G: 443-462.
Kardong, K. V.2002. Vertebrate Comparative Anatomy, Function, Evolution. Mc graw Hill, Boston,
763pp.
Kawecki, T. J., D. Ebert. 2004. Conceptual issues in local adaptation. Ecology Letters 7: 1225–
1241.

172
Lenormand, T. 2002. Gene flowand the limits to natural selection. Trends Ecol. Evol. 17: 183-
189.
Kile, R. 1987. Rodents under the carving knife. New scientist, 58-62.
Krisher, J. 2006. Un Compañero Neotropical. American Birding Assoc., Colorado, 437pp.
Lande, R. 1979. Quantitative genetic of multivariate evolution, Appliedto brain: body size
allometry. Evolution 33: 402-416.
Lawrence, S. 2013 Genetic variation and populatros genetic structure of muskrat, ondatra zilethicus.
Laurentian Univ. Sudbury, Ontario.154pp.
Lim, B.1970. Distribution relative abundance, food habits and parasite patterns of giant rats (Rattus)
in west Malaysia. J. Mammal., 51: 730-740.
Linares, J. L. 2005. Influencia del grado de antropización y del tipo de formación vegetal sobre la
densidad de jutía conga (Capromys pilorides Say) en la reserva de la Biosfera Península de
Guanacahabibes. Tesis en opción al Grado Académico de Master en Ciencias Forestales.
UPR. 73 pp.
Linares, J. L., y V. Berovides. 2008. La jutía conga (Capromys pilorides) como recurso natural
alimentario, en el Parque Nacional Guanahacabibes. CUBAZOO, 18: 29-31
Linares, J. L., y V. Berovides. 2009. Abundancia y selección de refugio de la jutía conga (Capromys
pilorides) de manglares. CUBAZOO. 19: 41-45.
Linares, J.L. 2014. Ecología reproductiva de la jutía conga (Capromys pilorides) en dos poblaciones
del Parque Nacional Guanahacabibes. CUBAZOO 26: 53-62.
Linares, J. L; Berovides; A.Rojas; L.Márquez, D. Cobián; J.A.Camejo; A.Sosa. 2010. Morfometría,
densidad y alimentación de la jutía conga (Capromys pilorides), en la reserva de Biosfera
Península de Guanahacabibes. CUBAZOO 21: 61-65.
Lindstedt, S., and S. D. Swain. 1988. Body size as a constrint of design and function. En: M- S.
Óbice (Ed). Evolution of Life Histories of Mammals. Yala Univ. Press, Londres. pp 93-105.
Lora, A. 1895. Contribucón al estudio del Género Capromys. Tesis de Doctorado. Real Univ.
Literaria de la Habana. 68 pp.

173
Lundrigan, B.1996. Standard methods for measurig mammals. En: Kleiman D. G., M. E. Allen, and
K. V. Thompson (Ed.) Wild Mammals in Captivity. Univ. Of Chicago Press, Chicago, pp.
566-570.
Manójina N. 1984. Observaciones etoecológicas sobre la reproducción en Cautiverio de la jutía
conga (Capromys pilorides) en territorio cubano. 2do Cogreso de biología, La Habana,
Resumenes, p. 461.
Manójina, N. y R. Abreu. (1987). Características reproductivas de jutía conga Capromys pilorides
en la península de Guanacahabibes. Poeyana: 348: 1-8.
Manójina, N., R. Abreu y A. González. 1989. Datos sobre la alimentación de la Jutía Conga
(Capromys pilorides) en Guanacahabibes. Poeyana 369:1-13.
Manójina, N., y R. Abreu. 1990. Utilización de algunos moluscos y reptiles en la dieta de la jutía
conga (Rodentia: Capromydae). Cien. Biol., 23: 127-129, I lám.
Martín, N; P. Moestrup.2003. Changes in mamnalian body lengh over 175 años- Adaptation to
fragmentedn landscape. Conservation Ecology. 7: 1-7.
Mayr, E. 1963. Population Species, and Evolution. Belknap Press of Harvard Univer. Pres,
Cambridge, Massashusetts 260 pp.
Mayr, E.1988. toward a new philosophy of biology. Observations of an evolucionist. Harvard
Univer. Pres.
McNeely, J. 1994. Áreas protegidas para el siglo XXI: trabajando para proporcionar beneficios a la
sociedad. Unasylva. 1:11-18.
McPherson, G. R., and S. De Stefano. 2003. Applied Ecology and Natural Resourse Mangement.
Cambridge Univ. Press, Cambridge, Uk. 165pp.
Mcwillian, S. R., W. H. Karasov. 2001. Phenotypic flexibility in digestive system structure and
function in mygratory birds. Comp. Bioch. Physiol., A 128: 579-593.
Mederos, L. M., C. Jimenez, A. M. Gutiérrez, C. Ferrá, y J. A. Valdivia. 1992. Identificación de
cepas pertenecientes al complejo MAI. aisladas en Capromys pilorides (jutía conga). 2do
Eencuentro de la Sociedad Cubana de Medicina Veterinaria, La Habana (sin paginación).
Merino, N., y M. Bouza. 1988. Primer reporte de Cuba de filaria en la jutía conga (Capromys
pilorides). Misc. Zool., 41: 2.

174
Miguel, D. 2008. Patrones estacionales y espaciales de la reproducción en dos poblaciones de
capromys pilorides (Rodentia: Capromidae) Tesis en opción al Grado Académico de Master
en Ciencias. Universidad de la Habana, 52 pp.
Milisnikov, A. N. 1993. Variabilidad proteica, divergencia, y evolución de las jutías cubanas (en
ruso) En investigaciones sobre la fauna cubana, Academia de Ciencia de Rusia, Nauka,
Moscú, pp 97-110.
Miljutin, A.1997. Ecomorpholy of the Baltic rodents: body form, ecological strategies, and adaptive
evolution. Estomian Acad. Publ.45 pp.
Miljutin, A.1997. Ecomorpholy of the Baltic rodents: body form, ecological strategies, and adaptive
evolution. Estomian Acad. Publ.45 pp.
Millien, V., and J. Damuth. 2004. Climate change and size evolution in an island rodents species:
new perspectives on the island rule. Evolution, 58: 1353-1360.
Millien, V., S. K. Lyons; L. Olson. 2006. Ecottypic variation in the context of global climate
change. Ecology Letters 9: 853-869.
Monroy, A. G., A. Hureña, and L. A. Espinosa. 2005. Variación morfométrica de Peroyscus
maniculatus fulvus y Rothrodontomys megalotis saturatus de la ciudad de Mexico DF. Rev.
Mex. Maztozoologia. 9 : 72-84.
Moreno, J. 2008. Los Retos actuales del Darwinismo. ¿Una Teoría en Crisis? Editorial Síntesis,
Madrid, 381pp.
Morh, E.1939. Die Baum-und ferkel-ratten-gattungen Capromys Desmarest (sens. ampl.) und
Plagiodontia Cuvier. Mitt. Hamburg. Zool. Mus. Inst., 48: 48-118.
Morris, P. W. 2011. Adaptation and habitat selection in the eco-evolutionary process. Proc. R.
Soc.B. 278: 2401-2410.
Nadal, J.2001. Vertebrados. Orígen, Organización, Diversidad y Biología. Ediciones Omega,
Barcelona, 707pp.
Naya, D. E. 2008. Gut size flexibility in rodents what we know, and don´t know, after a century of
research. Rev. Chilena Hist. Nat., 81: 599-612.
Naya, D., L. A. Ebensperger; P. Sabat; F. Bozinovic. 2008. Digestive and metabolic flexibility allow
female Degus to cope with lactation costs. Physiol. Bioch. Zoo. 81 : 186 194.

175
Nespolo, R.F.2003. Evolution by natural selection: more evidence than ever before. Rev.chilena
Hist. Natural 76: 699-716.
Novoa, F., A. Rivera, and M. Rosenmann. 2005. Intraespecific differences in metabolic rate of
Chroeomys olivaceus (Rodentia: Muridae): the effect of thermal acclimation in arid and mesie
habitats. Rev. Chilena, Hist. Nat., 78: 207-214.
NRC. 1991. Microlivestock. Nat. Academy Press. Washington. 499 pp.
Ntshotho, P., J. Van Aarde; S. W. Nicolson; T. P. Jackson. 2004. Renal physiology of two southern
african mastomys species (Rodentia; Muridae): A salt-loading experiment to assess
concentrating ability. Comp. Bioch. Physiol. 139: 444-447.
Ojaeti, J., y F. Dallneier. 2000. Manejo de Fauna Silvestre Neotropical. Smithsonian
Institution/MAB Biodiversity Program. Washington DC, 290 pp.
Ojeda, R; C.E. Boghi, G. B. Díaz; S. M. Giannori, M. A. Mares; J.K.Braun 1999. Evolutionary
convergence of the highly adapted desert rodents Tympanoctomys barrerae (Octodontidae) J.
Ard. Enviroments. 41: 443-452.
Oliver, W. L. R. (1977). The hutias of West Indies. International Zool. Yearb., 17:14-21.
Pelletier, F., D. Reale, D. Garant, and D. W. Coltman. 2007. Selection on hereditable seasonal
phenotypic plasticicity of body mass. Evolution, 61: 1969-1979 et al., 2007.
Pergamo, O. R. W; M.V.Ashley.2001. Microevolución in island rodents. Genetics 112-113: 245-
256.
Pimentel, O, 2006. Fundamentos Biológicos de la Jutía Conga (Capromys pilorides) para su Manejo
en Vida Libre en la cordillera de Guaniguanico. Tesis de Doctorado Univ. De Montaña, San
Andrés, Pinar del Río, 100pp.
Pimentel, O. (1987). Medidas biotécnicas para las áreas protegidas cubanas. Sistemas de normas
E.N.P.R.F.F. La Habana. 48 pp.
Pimentel, O. 2009. Factores que influyen en la extirpación de la jutía conga (capromp pilorides) en
el macizo forestal central de la cordillera de Guaniguanico. CUBAZOO 20: 73-78.
Pimentel, O., F. Hernández. 2007. Estudio de la utilización de las especies de vegetales para la
alimentación de la jutía conga (Capromys pilorides say). CUBAZOO, 16: 75-79.

176
Pimentel. O.2008. LA cría en cautiverio de la jutía conga (Capromys pilorides), como aporte a la
sostenibilidad de la pequeña economía rural del valle de San Andrés. CUBAZOO 18: 33-36.
Pough, F. H., Ch. M. Janis, and J. B. Heiser. 1999. Vertebrate Life. Prentice Hall, New Jersey,
733pp.
Prescott-Allen, R; C. Prescott-allen 1996. Assessing the Sustainability of Uses of Wild Species.
IUCN/SSCV 12, Gland, Zuiza. 135 pp.
Priersman, T., I. Drent. 2003. Phenotypic Flexibility and the evolution of organismal design. Trends.
Ecol. Evol. 18: 228-233.
Primack, R.B. (1995). Essentials of Conservation Biology. Sinauer -Sunderland. USA, 277pp.
Reaka M. L., and K. B. Armintage. 1976. The water economy of harvest mice from xeric and mesic
environment. Physiol. Zoolo. 49: 307-327.
Rezende, E. L., F. Bozinovic, and T. Garland. 2004. Climatic adaptation and the evolution of basal
and maximum rates of metabolism in rodents. Evolution: 1361-1474.
Riclefs, R. E; Miles, D.B.1994.Ecological and evolutionary inferences from morphology.In:
Ecological morphology (ed: P.C Wainwright; S.M.Reilly,pp 13-41. Univ. Chicago Press,
Chicago.
Rivas, B.A;O.J.Linares. 2006. Cambios en la forma de la pata posterior entre roedores
sigmodontinas según su locomoción y hábitat. Mastozoología Neotropical 31: 205-215.
Rosa, R. (2004). Estudio para el manejo en vida libre y cautiverio de la jutía conga (Capromys
pilorides), en la península de Guanacahabibes. Tesis en opción al grado de Master en
Ciencias. Universidad de Pinar del Río. 159 pp.
Ruckstuhl, K.E. 2007. Sexual segregation in vertebrate. Integ. Comp. Biol. 47: 245-257.
Ryder, O.A. 1986. Species; conservation and systematics: the dilemma of subspecies. Trends Ecol.
Evol. 1:9-10.
Sabat, P.; F. Bozinovic. 2008. Do changes in dietary chemistry during ontogeny affect digestive
performance in adults of the herbivourus rodent Octodon Degus. Comp. Bioch. Physiol. 151:
455-460.

177
Samuels, J. X; Van Valkenburgh, B. 2008.Skeletal indicators of locomotor adaptations in living and
extints rodents.J.Morphol. 269: 1387-1411.
Samuels, J. X; Van Valkenburgh, B. 2008.Skeletal indicators of locomotor adaptations in living and
extints rodents.J.Morphol. 269: 1387-1411.
Sánchez, J. A., A. Comas, V. Berovides. 1992. Indices morfofisiológicos en poblaciones de jutía
conga (Rodentia, Capromyidae). Rev. Biología 3: 174-182.
Santos, J. A., Y. Hortelano. 1997. La variación en mamíferos: una revisión de los enfoques
metodológicos actuales. Acta Zoo. Mex. 70: 13-34.
Sassi, P., C. Borghi, and F. Bozinovic. 2007. Spatial and seasonal plasticity in digestive morphology
of cavies (Microcavia australis) inhabiting habitats with different plantquality. J. Mammal.,
88: 165-172.
Schauenberg, P. 1977. Longuer de le intestin du chat foriester Felis silvestris Shreber. Mammalia
41: 357-360.
Schluter, D. 2001. Ecology and the origin of especies. Trends Ecol. Evol., 16: 372-380.
Schmidt-Nielsen, B; R. O’Deell. 1961. Structure and concentrating mechanism in the mammalian
kidney. Am. J. Physiol. 200: 1119-1124.
Schondube, J. E., M. L. Herrera, and C. Martinez del Río. 2001. Diet and evolution of digestión and
renal funtion in phillostomid bats. Zoology 104: 59-73.
Schulte-Hostedde, A. I. 2001. Sexual dimorphism in body composition of small mammals. Can. J.
Zoo 79: 1016-1020.
Shepherd, J.D; Ditgen, R.S. 2012. Predation by Rattusnovergicusan a natural small mammals in an
Araucaria forest. Rev. Chil. Hist. Nat.85: 155-159.
Shepherd, J.D; Ditgen, R.S. 2012. Predation by Rattusnovergicusan a natural small mammals in an
Araucaria forest. Rev. Chil. Hist. Nat.85: 155-159.
Sievert, P. R., and L. B. Keith. 1985. Survival of snowshoe hares at a geographic range boundary. J.
Wildl. Manage., 49: 854-865.
Silva, G; W. Suarez; S. Diaz. 2007 Compendio de los mamíferos terrestres autótonos de Cuba
vivientes y extinguidos. Museo Nac. Hist. Nat. La Habna, Cuba. 465 pp.

178
Smith, R., y V. Berovides. 1984a. Reproducción y ecología de la jutía conga. Poeyana, 280:1-20.
Smith, R; V. Berovides. 1984b. Ecomorfología y rendimiento de la jutía conga (Capromys
pilorides). Poeyana 279: 1-19.
Smith-Nielsen, K. 1997. Animal Phisiology. Cambridge Univ. Press, UK.612 pp.
Smythe, N., W. E. Glanz, and E. G. Leigh.1982. Population regulation in some terrestrial
frugivores. En: The Ecology of a Tropical Forest. Leigh, E. G., A. S. Rand, and D. M.
Windsor (Eds.). Smithsonian Inst. Washington Dc. 292 pp.
Soberón, R., R. Ramos, y B. Barr. 2001. Population survey and stomach content analysis. Crocodile
Specialist Group Newletter, 20 (I): 13-14.
Sokal, R. R., and F. J. Rolhf. 1995.Biometry. W-11Freeman and Co., S. Fco., 205 pp.
Soulé, M. (1991). Conservation: Tactic for a constant crisis. Science 253: 744-750.
Soulé, M. 1985.What is Conservation biology?. Bioscience, 35 (11): 727-734 .
Speakman, J. R., E. Krol, and M. S. Johmson. 2004. The functional synificance of individual
variation in basal metabolic rate. Physiol. Bioch Zool 77: 900-915 .
Sperber, I .1944. Studies on the mammalian kidney. Zool. Bidr Upp. 22: 249-432.
Stearns, S. C. 1983. The influency of size and phylogeny on patterns of covariation amo life history
traits in mammals. OIKOS, 41: 173-187.
Stearns, S. C. 1992. The Evolutionary links between fixed and variable traits. Acta Paleontol.
Polon., 38: 215-232.
Stearns, S. C. The Evolution of Life Histories. Oxford. Univ. Press, Oxford. 249pp.
Steel, R. G. D., and J. H. Torrie. 1980. Principles and Procedures of Statistic. McGraw-Hill, New
York, 633 pp.
Studier, E.H; S. J. Wisniewsky; A. T. Feloman; 1983. Kidney structure in neotropical bats; J.
Mammal. Vol.64; 445-452.
Suárez, J. 2001. Manejo y uso de la fauna silvestre en el valle de San Andrés. Tesis en opción al
título de Ingeniero Forestal. Universidad de Pinar del Río. 97 pp..

179
Taylor, C. R., N. C. Heglurd, G. M. O. Maloiy. 1982. Energetics and mechanics of terrestrial
locomotion. J. Exp. Biol. 97: 1-21.
Taylor, J. M., S. C. Smith, and J. H. Calaby. 1985.Altitudinal distribution and body size among New
Guinea rattus. J. Mamm 66: 353-358.
Taylor, R. H. 1970. Reproduction development and behaviors of the Cuban hutia conga (Capromys
pilorides) in captivity. Unpublished Thesis. Univ. Of Puget Sound, Tacoma. Washington. 190
pp.
Thompson, D. B.1990. Different spatial scales of adaptation in jhe climbing beharvior of
Peromyscus maniculatus. Evolution, 44: 952-965.
Thornigton, R. W., and L. R. Heaney. 1981. Body proportions and glading adaptations of flyng
squirrels. J. Mamm. 62: 101-104.
Tulli, M; Carrizo, L; Samuels, J.2015 Morphological variation of the forelimb and claw in
neotropicalsigmodontinae rodents. J. Mamm Evol. 23: 1-11.
Tulli, M; Carrizo, L; Samuels, J.2015 Morphological variation of the forelimb and claw in
neotropicalsigmodontinae rodents. J. Mamm Evol. 23: 1-11.
Turelli, M., N. H. Barton y J. A. Coyne. 2001. Theory and especiation. Trends Ecol. Evol, 16: 330-
343.
Tzamakakis, T. A. 2000. Liver assist devices. Animal Rev. Biomed. Engeneering, 2: 607-632.
Van Soest, P. J. 1996. Allometry and ecology of feeding beharvior and digestive capacity in
herbivores: A review. Zoo. Biology, 15: 455-479.
Varona, L. S. 1974. Catalogo de mamíferos vivientes y extinguidos de las Antillas. Academia de
Ciencias de Cuba. La Habana. 139pp.
Varona, L. S. 1980a. Una nueva especie de Capromys pilorides (Rodentia: Capromydae). Mem.
Soc. Cien. Nat. La Salle, 40 (114): 141-148.
Varona, L. S. 1980b. Mamíferos de Cuba. Gente Nueva, La Habana, 111 pp.
Varona, L. S. 1983. Nueva subespecie de jutía conga Capromys pilorides (Rodentia: Capromydae).
Caribbean J. Sci., 19 (3/4): 77-79.

180
Vasallo, A. y D. Antenicci (eds). 2015. Bilogy of caviomorph rudents: Diversity and evolution.
Mastozoología Neotropical 23: 54-63.
Verde, L.D., D. O. Fisher y M. Schweizer. 2016. Morphology captures diet and locomotor type in
rodents. Doj 10:5281.
Vorontsov, N. 1962. The way of food specialization and evolution of the alimentary system in
murieidea. In: Kratochvil, J., J. Pelikan (Ed.). Internac. Sympo. Method Mammalogical
Investigation. Publ. House Acad, Praha, pp. 360-377.
Wainwright, P. C. and S. M. Reilly. 1994. Ecological Morphology. Unive. Chicago Press, Chicago,
367pp.
Wang, D., Y. Pei, J. Yang y Z. Wang. 2003. Digestive tract morphology and food habit in six species
of rodents. Folia Zool., 52: 51-55.
Warters, L. A., M. Vermeulen y S. Van Dongen. 2007. Efects of spatio- temporal variation in food
supply on red squirrel Sciurus vulgaris body size and body mass and its consequences for
some yitness components. Ecography 30: 51-65.
Whitlock, M.C. y D. Schluter. 2009 The Analysis of Biological Data. Robert Co. Pub. Colorado.
799 pp.
Williams, G. C. 1996. Adaptation and Natural Selection. Princeton University Press
Wilson, D. E. y D. M. Reeder. 1993. Mammal Species of the World. Smithsorian Inst. Press,
Washington, 800 pp.
Wilson, R.T. 2011. Small Animals for Small Farms.FAO, Roma 92pp.
Wolf, M., M. Friggens y J. Salazar-Bravo. 2009. Does weather shape rodents? Climate related
changes in morphology of two heteromyd species. Naturwissenschaften, 96: 93-101.
Yabe, T. 1982. Influense of dehydrations on breeding, body size and kidney structure in norway rats
(Rattus norvegicus) on an islet. Physiol. Ecol. Japan; Vol. 19; 3-7.
Yabe, T. 1983. Renal structural indices for the ability to conserve water in rodents Mus molossinus,
Rattus norvegicus and Rattus rattus. Physiology Ecol Japan; Vol. 20; 53-57.
Yom-Tov, Y., S. Yom-Tov y H. Moller. 1999. Competitiom, coexistence, and adaptation among
rodent invader to pacific and New Zealad island. J. Biogeografy 26: 947-958.

181
Zar, H. J. 2009. Bioestatistical Analysis. Prentice Hall, New Jersey. 460 pp.
Copyright © 2022 FDOKUMEN




















