
Particle separation and fractionation by microfiltration - WUR ...

Variability in amino acid composition ofalpine crustacean zooplankton and itsrelationship with nitrogen-15fractionation
MARC VENTURA 1,2* AND JORDI CATALAN 1
1BIOGEODYNAMICS AND BIODIVERSITY GROUP (CSIC-UB), CENTRE D’ESTUDIS AVANCATS DE BLANES (CEAB), ACCES A LA CALA SANT FRANCESC, 14, 17300,
BLANES, CATALONIA, SPAIN AND2
INSTITUT DE RECERCA DE L’AIGUA, UNIVERSITAT DE BARCELONA AV. DIAGONAL, 684, 08034 BARCELONA, CATALONIA, SPAIN
*CORRESPONDING AUTHOR: [email protected]
Received February 10, 2010; accepted in principle April 7, 2010; accepted for publication May 11, 2010
Corresponding editor: Beatrix E. Beisner
Amino acids (AAs) are critical biochemical compounds for living organisms.Because of the limited capacity for their de novo synthesis in many animals, thenutritional value of food largely depends on its AA composition relative to theanimal’s requirements. To improve present knowledge on AA variability in fresh-water crustaceans, we studied the inter- and intraspecific variability in three con-trasting species from an oligotrophic alpine lake (Daphnia pulicaria, cladocera;Cyclops abyssorum, cyclopoid copepod and Diaptomus cyaneus, calanoid copepod).Inter-species differences were larger than intraspecific variation, confirming a non-strict homeostasis in freshwater crustacean zooplankton. The intraspecific variabil-ity differed for each species: in Daphnia, it was mainly related with ontogeneticchanges rather than reproduction; in Cyclops, both factors were equally important;and reproduction was the most relevant in Diaptomus. Reproduction changes wereassociated with serine and phenylalanine in the three species, while the AAsresponsible for ontogenetic changes differed in each species. There were nogender differences in AA composition in any of the two copepod species. FreeAAs formed a very low percentage of total AA pool (,2.7%). Taking advantageof the fact that Daphnia is the main prey for Cyclops in the lake studied, we furtherinvestigated to what extent the AA composition is related with Cyclops–Daphnia
nitrogen stable isotope fractionation. Only those AAs that are both essential andare not trans-aminated during protein synthesis had a significant correlation withnitrogen stable isotope fractionation, supporting the hypothesis that an AA imbal-ance can be responsible for a variable nitrogen stable isotope fractionation.
KEYWORDS: amino acid composition; homeostasis; food quality; nitrogen frac-tionation; stoichiometry
I N T RO D U C T I O N
Amino acids (AAs) are critical biochemical compoundsfor living organisms (i.e. as building blocks of proteins,
as part of coenzymes and as precursors for the biosyn-thesis of some molecules) (Strayer, 1988). Because of thelimited capacity for their de novo synthesis in many
doi:10.1093/plankt/fbq066, available online at www.plankt.oxfordjournals.org. Advance Access publication June 8, 2010
# The Author 2010. Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected]
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010 by guest on June 22, 2016
http://plankt.oxfordjournals.org/D
ownloaded from

animals, the AA composition of food becomes veryimportant in animal nutrition. This has been demon-strated in different types of terrestrial organisms such asbutterflies (O’Brien et al., 2003), birds (Ramsay andHouston, 1998), farm animals (D’mello, 1994) andhumans (Reeds, 2000), and also for aquatic organismssuch as farmed fish (Conceicao et al., 2003; Ronnestadet al., 2003), rotifers (Boechat and Adrian, 2006),mussels (Kreeger et al., 1996) and marine copepods(Kleppel and Burkart, 1995; Guisande et al., 1999,2000). This knowledge contrasts with the few ecologicalstudies focusing on the relative importance of AAs foranimal growth and reproduction (Conklin andProvasoli, 1977; Boechat and Adrian, 2006).Stoichiometry studies focusing on elements, mainlyphosphorus, but also carbon and nitrogen (reviewed inSterner and Elser, 2002) or fatty acids, mainly highlyunsaturated fatty acids such as eicosapentanoic acid ordocosahexanoic acid (e.g. Muller-Navarra et al., 2000),have received more attention than AAs. Since most ofanimal’s nitrogen content is in the form of AAs, com-parison between organisms and their food AA compo-sition has been suggested as a step forward in usingstoichiometric arguments for a better understanding ofnitrogen limitation (Anderson et al., 2004). This sugges-tion is based on two main assumptions, namely that theconsumer is unable to synthesize some AAs in signifi-cant quantities (i.e. essential AAs) and that it is homeo-static with respect to the composition in essential AAs.The former assumption is supported by numerousstudies, including in crustaceans (Claybrook, 1983),whereas the latter has only been partially demonstratedand further studies are required (Cowgill et al., 1986;Anderson et al., 2004).
An imbalance between the AA composition of foodand those of the consumer has been proposed as one ofthe main mechanisms explaining nitrogen stable isotopefractionation (i.e. change in isotope ratios, D15N)between diet and animal tissues (Martınez del Rio andWolf, 2005). Nitrogen stable isotopes together with thoseof carbon have become a common tool for describingtrophic relationships, although the physiological mech-anism explaining D15N (crucial for estimating dietsources) is still not resolved (Martınez del Rio et al.,2009). The physiological mechanism could be causedby the deficiency in one or several essential AAs leadingto greater protein intake and eventually to higher cata-bolism of the AAs in excess for protein synthesis favour-ing a preferential body elimination of lighter nitrogen(Martınez del Rio and Wolf, 2005). Few studies haveprovided evidence for this hypothesis with the exceptionof Robbins et al. (Robbins et al., 2005), in a comparativestudy, and Ventura and Catalan (Ventura and Catalan,
2008), in a field study, that reported a significant nega-tive correlation between D15N and the protein qualityof the diet (e.g. AA composition). An important unre-solved question is if there are some particular AAs thatare more relevant for D15N. The essential part of anAA (and what differentiates them biochemically) is theircarbon skeleton. All the nitrogen of AAs is in the aminogroup, which can be trans-aminated by animals in,a priori, all AAs. However, it has been observed thatsome AAs appear to retain approximately the samenitrogen isotopic composition as their food source (arenot trans-aminated; “source AAs”), whereas othersbecome enriched in 15N by the animals’ metabolism(undergo trans-amination; “trophic AAs”) (McClellandand Montoya, 2002; Popp et al., 2007). Different pro-portions of “trophic” and “source” AAs explain whyanimal tissues differ in 15N (Martınez del Rio et al.,2009). Since the trans-amination is not related to the AAcarbon skeleton, some trophic/source AAs are essentialwhile others are nonessential, making a complicatedphysiological puzzle. What particular AAs or AA groupis directly related to D15N has not been described andtherefore needs to be explored more in depth. One mayhypothesize that among the essential AAs, only thosethat are not trans-aminated would have a higher rel-evance than the others.
Zooplankton has played a central role in stoichi-ometry studies, particularly Daphnia, which has beenextensively used as a model organism. Investigationsexamining the AA composition are scarce, however.Most past studies of the AA composition of zooplanktonhave focused on the variability in the total AA (TAA)composition of adult marine or freshwater zooplanktoncompared to their food, the results showing that adultshave a very strict AA composition (Cowey and Corner,1963; Cowgill et al., 1986; Guisande et al., 2000). Allthose studies are indicative of AA composition homeo-stasis in adults of crustaceans. In contrast, there are fewstudies focusing on the TAA intraspecific variability.Significant differences in AA composition during devel-opment have been observed in Artemia spp. and lobsters(Helland et al., 2000; Mente et al., 2001). In cladoceransand copepods, Dabrowski and Rusiecki (Dabrowski andRusiecki, 1983) found differences in TAA between latedevelopmental stages of different species, although theydid not examine the degree of these TAA changes foreach species. Changes in TAA were reported for twoCalanus species during development from egg to thesecond naupliar stage (Laabir et al., 1999) and betweennauplii of different seasons (Helland et al., 2003a).Ontogenetic variability in species of cladocerans, cala-noid and cyclopoid copepods were studied in differentcoastal lagoons and wetlands by Brucet et al. (Brucet
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1584
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

et al., 2005). They analysed all the lake-populations sim-ultaneously, and found a more pronounced variabilityin the TAA composition related with ontogeny in cope-pods, while Daphnia variability was mostly related withlake type and not with ontogeny. However, the relativeimportance of ontogeny, reproduction and gender inAA composition changes has not been evaluated forthese animal groups.
In this context, with the aim of testing the hypoth-esis that interspecific differences among the mainfreshwater groups of crustacean zooplantkton aregreater than their intraspecific variation, we studiedthe relative proportions of the AA pools and its vari-ation in crustacean zooplankton species of an alpinelake throughout a whole seasonal cycle. We selected alake that had one species of each of the main fresh-water crustacean groups, a cladoceran (Daphnia pulicaria
Forbes), a cyclopoid copepod (Cyclops abyssorum Sars)and a calanoid copepod (Diaptomus cyaneus Gurney). Ina previous study, these three species were found tohave substantial changes in their elemental content ofcarbon and phosphorus during their life cycles associ-ated with reproduction (Ventura and Catalan, 2005).In contrast, nitrogen was the only compound whichdid not change significantly. In addition, Cyclops wasdescribed as being a predatory copepod feeding pri-marily on Daphnia, and the D15N of both species wasstrongly correlated with their protein quality, whichwas interpreted as evidence of consumer AA (nitrogen)limitation (Ventura and Catalan, 2008). Therefore,these two species also present an ideal framework forinvestigating the specific role of AAs inD15N. Contrasting with Cyclops, the other two species,Diaptomus and Daphnia, are more generalistic consu-mers (an omnivore and herbivore, respectively), andtherefore they are not suitable for this particular com-parison. The specific hypotheses driving the studywere (i) interspecific variability of AA composition isgreater than intraspecific AA variability; (ii) reproduc-tion, gender and ontogenetic changes are importantintraspecific sources of AA variability; and (iii) essentialAAs that are not trans-aminated during tissue incorpor-ation are those most closely related with the variabilityin D15N between Cyclops and Daphnia.
M E T H O D
Study site and sample collection
Lake Redon (formerly lake Redo) is an oligotrophicglacial cirque lake located at 2240 m a.s.l. in the centralPyrenees (428380N, 08460E). It is relatively large (24 ha)
and it is one of the deepest lakes (73 m) in the Pyrenees.It is dimictic, covered by ice during half of the year,usually from mid-December until late May or thebeginning of June. The lake has average microbialbiomass ratios of 10:2:2:1 for phytoplankton:bacteria:-heterotrophic nanoflagelates:ciliates during the ice-freeperiod (Felip et al., 1999). Rotifers are usually less abun-dant than crustaceans (the ratio between Daphnia androtifer biomass is approximately 10:1 during the ice-freeperiod; Camarero et al., 1999).
The lake was sampled on 14 occasions fromDecember 1998, just after the lake was completely icecovered, to December 1999, after it was ice-coveredagain. Samples were collected at the deepest point ofthe lake either after drilling the ice-cover or from a plat-form anchored throughout the summer at the samepoint. Zooplankton samples were collected by verticalhauls from 65 m to the surface with a 200 mm net.Individuals were kept alive and transported cold (48C)until they were frozen (2208C) in the laboratory a fewhours after collection.
The three crustacean zooplankton species studiedhad contrasting life histories, although all produced asingle cohort per year. A complete description of theirlife cycles during the study period can be found inVentura and Catalan (Ventura and Catalan, 2005) andtheir trophic relationships in Ventura and Catalan(Ventura and Catalan, 2008). Cyclops is a predatorycopepod feeding primarily on Daphnia in this lake(Ventura and Catalan, 2008). Its life cycle was synchro-nized with that of Daphnia being the only two planktoniccrustacean species inhabiting the lake during the wholeyear. Adult females of Daphnia from the previoussummer survived below the ice-cover, reproducedduring the end of the ice-covered period and the first2 months of the ice-free period, and disappeared fromthe lake afterwards, leaving the juveniles growing upduring the summer (Fig. 1) (Ventura and Catalan,2005). Adult females (termed “first cohort females” inour study) were therefore clearly distinguishable fromthe juveniles born during the ice-free period (termed“second cohort females”). We therefore compared theAA composition of the two cohorts, of the reproducingand non-reproducing first cohort females, and theireggs. Similar to Daphnia, adult females of Cyclops sur-vived below the ice-cover in order to reproduce duringthe end of the ice-covered period. Adult males werepresent in the lake from December to February belowthe ice-cover. Nauplii appeared the following summer,just after the ice-cover melted (copepods have six nau-pliar, and five copepodite stages before adults develop)and coincided with the spring production maximum.Copepodites CI–CIII dominated Cyclops biomass
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1585
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
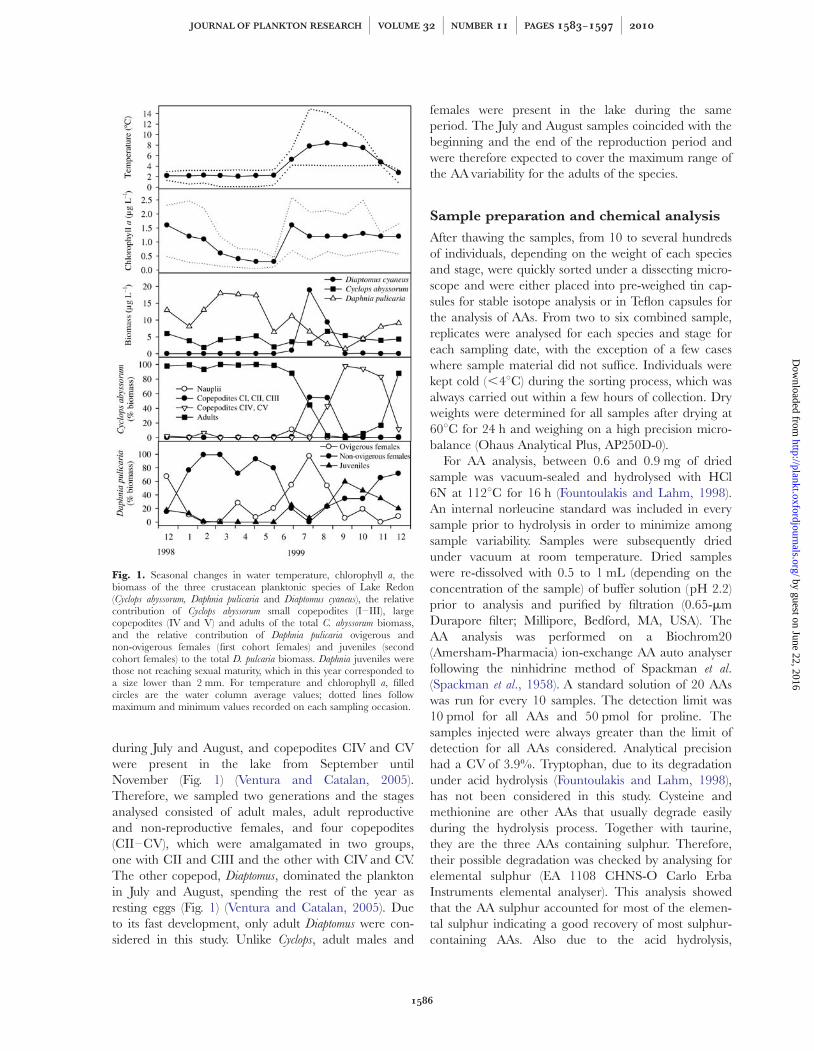
during July and August, and copepodites CIV and CVwere present in the lake from September untilNovember (Fig. 1) (Ventura and Catalan, 2005).Therefore, we sampled two generations and the stagesanalysed consisted of adult males, adult reproductiveand non-reproductive females, and four copepodites(CII–CV), which were amalgamated in two groups,one with CII and CIII and the other with CIV and CV.The other copepod, Diaptomus, dominated the planktonin July and August, spending the rest of the year asresting eggs (Fig. 1) (Ventura and Catalan, 2005). Dueto its fast development, only adult Diaptomus were con-sidered in this study. Unlike Cyclops, adult males and
females were present in the lake during the sameperiod. The July and August samples coincided with thebeginning and the end of the reproduction period andwere therefore expected to cover the maximum range ofthe AA variability for the adults of the species.
Sample preparation and chemical analysis
After thawing the samples, from 10 to several hundredsof individuals, depending on the weight of each speciesand stage, were quickly sorted under a dissecting micro-scope and were either placed into pre-weighed tin cap-sules for stable isotope analysis or in Teflon capsules forthe analysis of AAs. From two to six combined sample,replicates were analysed for each species and stage foreach sampling date, with the exception of a few caseswhere sample material did not suffice. Individuals werekept cold (,48C) during the sorting process, which wasalways carried out within a few hours of collection. Dryweights were determined for all samples after drying at608C for 24 h and weighing on a high precision micro-balance (Ohaus Analytical Plus, AP250D-0).
For AA analysis, between 0.6 and 0.9 mg of driedsample was vacuum-sealed and hydrolysed with HCl6N at 1128C for 16 h (Fountoulakis and Lahm, 1998).An internal norleucine standard was included in everysample prior to hydrolysis in order to minimize amongsample variability. Samples were subsequently driedunder vacuum at room temperature. Dried sampleswere re-dissolved with 0.5 to 1 mL (depending on theconcentration of the sample) of buffer solution (pH 2.2)prior to analysis and purified by filtration (0.65-mmDurapore filter; Millipore, Bedford, MA, USA). TheAA analysis was performed on a Biochrom20(Amersham-Pharmacia) ion-exchange AA auto analyserfollowing the ninhidrine method of Spackman et al.(Spackman et al., 1958). A standard solution of 20 AAswas run for every 10 samples. The detection limit was10 pmol for all AAs and 50 pmol for proline. Thesamples injected were always greater than the limit ofdetection for all AAs considered. Analytical precisionhad a CV of 3.9%. Tryptophan, due to its degradationunder acid hydrolysis (Fountoulakis and Lahm, 1998),has not been considered in this study. Cysteine andmethionine are other AAs that usually degrade easilyduring the hydrolysis process. Together with taurine,they are the three AAs containing sulphur. Therefore,their possible degradation was checked by analysing forelemental sulphur (EA 1108 CHNS-O Carlo ErbaInstruments elemental analyser). This analysis showedthat the AA sulphur accounted for most of the elemen-tal sulphur indicating a good recovery of most sulphur-containing AAs. Also due to the acid hydrolysis,
Fig. 1. Seasonal changes in water temperature, chlorophyll a, thebiomass of the three crustacean planktonic species of Lake Redon(Cyclops abyssorum, Daphnia pulicaria and Diaptomus cyaneus), the relativecontribution of Cyclops abyssorum small copepodites (I–III), largecopepodites (IV and V) and adults of the total C. abyssorum biomass,and the relative contribution of Daphnia pulicaria ovigerous andnon-ovigerous females (first cohort females) and juveniles (secondcohort females) to the total D. pulcaria biomass. Daphnia juveniles werethose not reaching sexual maturity, which in this year corresponded toa size lower than 2 mm. For temperature and chlorophyll a, filledcircles are the water column average values; dotted lines followmaximum and minimum values recorded on each sampling occasion.
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1586
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
asparagine and glutamine were measured together withaspartic acid and glutamic acid, respectively. AAs areexpressed as relative mass percentage of TAA through-out this paper. These units were chosen rather thanabsolute concentrations (e.g. mg AA/mg C or DW)since a description of the biochemical variability of thepresent species and study period showed that there wasan important seasonal variability in the lipid and carbo-hydrate content (up to 50% of their DW) (Ventura andCatalan, 2005), which were much greater than thechanges in the AA composition. Using the relativeunits, we focus on the variability in the AA compositionrather than the factors affecting the gross biochemicalcomposition. However, we provide the TAA concen-tration (in % dry weight) which allows for calculation ofthe absolute concentration also used in the literature.
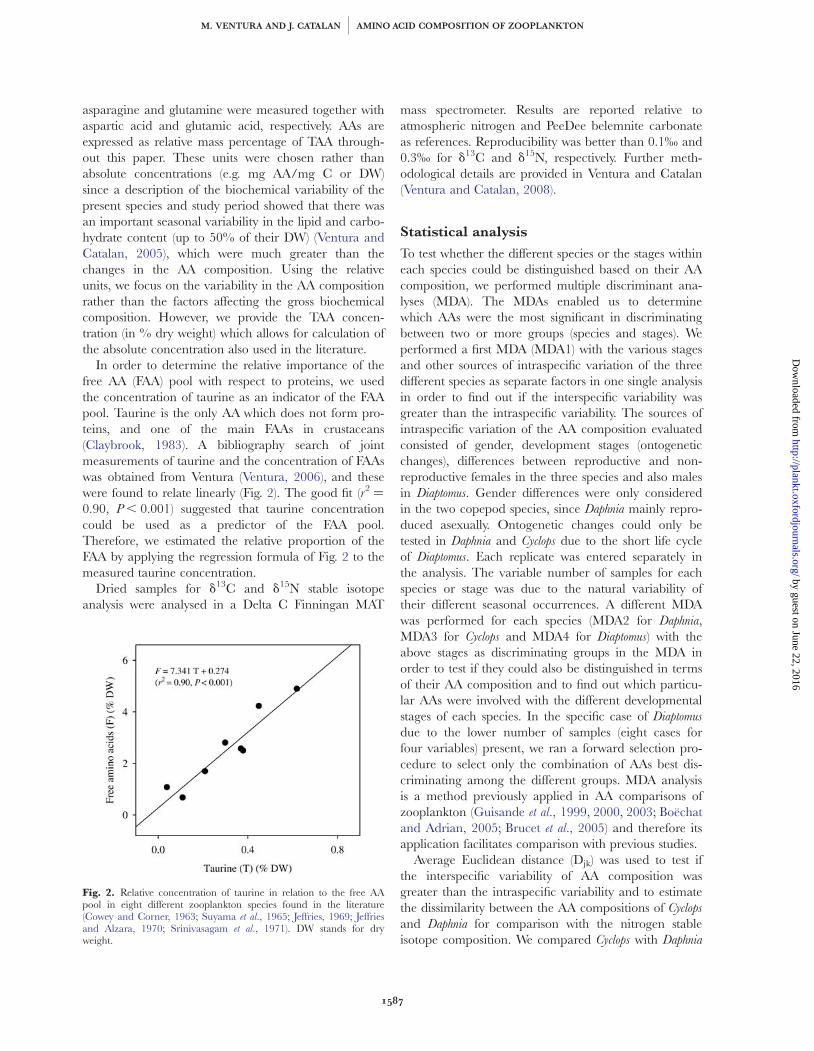
In order to determine the relative importance of thefree AA (FAA) pool with respect to proteins, we usedthe concentration of taurine as an indicator of the FAApool. Taurine is the only AA which does not form pro-teins, and one of the main FAAs in crustaceans(Claybrook, 1983). A bibliography search of jointmeasurements of taurine and the concentration of FAAswas obtained from Ventura (Ventura, 2006), and thesewere found to relate linearly (Fig. 2). The good fit (r2 ¼
0.90, P , 0.001) suggested that taurine concentrationcould be used as a predictor of the FAA pool.Therefore, we estimated the relative proportion of theFAA by applying the regression formula of Fig. 2 to themeasured taurine concentration.
Dried samples for d13C and d15N stable isotopeanalysis were analysed in a Delta C Finningan MAT
mass spectrometer. Results are reported relative toatmospheric nitrogen and PeeDee belemnite carbonateas references. Reproducibility was better than 0.1‰ and0.3‰ for d13C and d15N, respectively. Further meth-odological details are provided in Ventura and Catalan(Ventura and Catalan, 2008).
Statistical analysis
To test whether the different species or the stages withineach species could be distinguished based on their AAcomposition, we performed multiple discriminant ana-lyses (MDA). The MDAs enabled us to determinewhich AAs were the most significant in discriminatingbetween two or more groups (species and stages). Weperformed a first MDA (MDA1) with the various stagesand other sources of intraspecific variation of the threedifferent species as separate factors in one single analysisin order to find out if the interspecific variability wasgreater than the intraspecific variability. The sources ofintraspecific variation of the AA composition evaluatedconsisted of gender, development stages (ontogeneticchanges), differences between reproductive and non-reproductive females in the three species and also malesin Diaptomus. Gender differences were only consideredin the two copepod species, since Daphnia mainly repro-duced asexually. Ontogenetic changes could only betested in Daphnia and Cyclops due to the short life cycleof Diaptomus. Each replicate was entered separately inthe analysis. The variable number of samples for eachspecies or stage was due to the natural variability oftheir different seasonal occurrences. A different MDAwas performed for each species (MDA2 for Daphnia,MDA3 for Cyclops and MDA4 for Diaptomus) with theabove stages as discriminating groups in the MDA inorder to test if they could also be distinguished in termsof their AA composition and to find out which particu-lar AAs were involved with the different developmentalstages of each species. In the specific case of Diaptomus
due to the lower number of samples (eight cases forfour variables) present, we ran a forward selection pro-cedure to select only the combination of AAs best dis-criminating among the different groups. MDA analysisis a method previously applied in AA comparisons ofzooplankton (Guisande et al., 1999, 2000, 2003; Boechatand Adrian, 2005; Brucet et al., 2005) and therefore itsapplication facilitates comparison with previous studies.
Average Euclidean distance (Djk) was used to test ifthe interspecific variability of AA composition wasgreater than the intraspecific variability and to estimatethe dissimilarity between the AA compositions of Cyclops
and Daphnia for comparison with the nitrogen stableisotope composition. We compared Cyclops with Daphnia
Fig. 2. Relative concentration of taurine in relation to the free AApool in eight different zooplankton species found in the literature(Cowey and Corner, 1963; Suyama et al., 1965; Jeffries, 1969; Jeffriesand Alzara, 1970; Srinivasagam et al., 1971). DW stands for dryweight.
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1587
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

since the former was described as being a predatorycopepod feeding primarily on Daphnia in the same lakeand study period (Ventura and Catalan, 2008). TheEuclidean distance is a metric distance given by thePythagorean formula (Legendre and Legendre, 1998):
Dij ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiXn
i¼1
ðXij � XikÞ2s
where Xij and Xik are the percentages of the AA i ofspecies 1 ( j) or species 2 (k), and n is the number of AAsconsidered. The distance was calculated with all theAAs when evaluating interspecific versus intraspecificAA variability. For comparison of the distance betweenCyclops and Daphnia, we calculated the distance of allAAs or separately for the essential AAs (lysine,phenylalanine þ tyrosine, leucine, valine, threonine, iso-leucine, histidine, cysteine þ methionine), non-essentialAAs (glutamic acid, aspartic acid, alanine, arginine,glycine, proline and serine), trophic AAs (alanine,asparagine þ asparctic acid, glutamine þ glutamic acid,isoleucine, leucine, proline and valine) and source AAs(arginine, glycine, lysine, serine, threonine andphenilalanine þ tyrosine). The average Euclidean dis-tance between Cyclops and Daphnia using the differentAA groups was compared with the D15N of bothspecies with linear regression analysis.
In order to determine which particular AAs could bemost imbalanced in Daphnia in comparison with thepredator, Cyclops, we calculated the AA imbalance asfollows (Conceicao et al., 2003):
AAi ¼ dietAAi �consumer AAi
consumerAAi� 100
where dietAA and consumerAA are the contributions of agiven AA (i) to the AA profiles of the consumer.
Differences in individual AAs were tested withone-way ANOVA analysis (Underwood, 1997) withspecies or stage as independent variable and the percen-tage of the AAs as the dependent variable. AA percen-tage data were previously normalized with arcsine rootsquare transformation when required, and a test ofhomogeneity of variances was performed beforeANOVA. To test if Euclidean distances of interspeciesversus intraspecies comparison were significantly differ-ent, we used one-way ANOVA with the Euclidean dis-tance between all the pairs of samples analysed asdependent variable and if the comparison was fromamong species comparison (three cases) or withinspecies (three more cases) as independent variables. Weused Tukey’s post hoc test for detecting significant
differences among or within species. All statistical ana-lyses were performed with SPSS V17.
R E S U LT S
Interspecies differences
The three species had different TAA content. Daphnia
had the lowest relative concentration, followed byCyclops and Diaptomus (27.5% on dry weight+ 4.6 SD,45.4+ 8.4, 59.7+ 13.4, respectively) (F2,22 ¼ 42.3, P ,
0.001; Tukey’s post hoc test P , 0.0001) (Table I). Mostof the AAs were forming proteins, the FAA pool beinga small proportion of the TAA pool of each species.Daphnia FAA relative concentration (2.7+ 1.5% ofTAA) was slightly higher than Diaptomus (1.2+ 0.5% ofTAA, F2,22 ¼ 4.2, P , 0.05; Tukey’s P ¼ 0.014) and notstatistically different from Cyclops (1.9+ 0.8% of TAA,Tukey’s P ¼ 0.162).
The average Euclidean interspecific distance was twotimes greater than the average intraspecific distance(3.7+ 1.1 and 1.7+ 0.6, respectively; F5,1080 ¼ 706,P , 0.001; Tukey’s P , 0.001; Fig. 3). The two cope-pods had a more similar composition to each otherthan to the cladoceran, while the three species had asimilar intraspecific distance (Fig. 3).
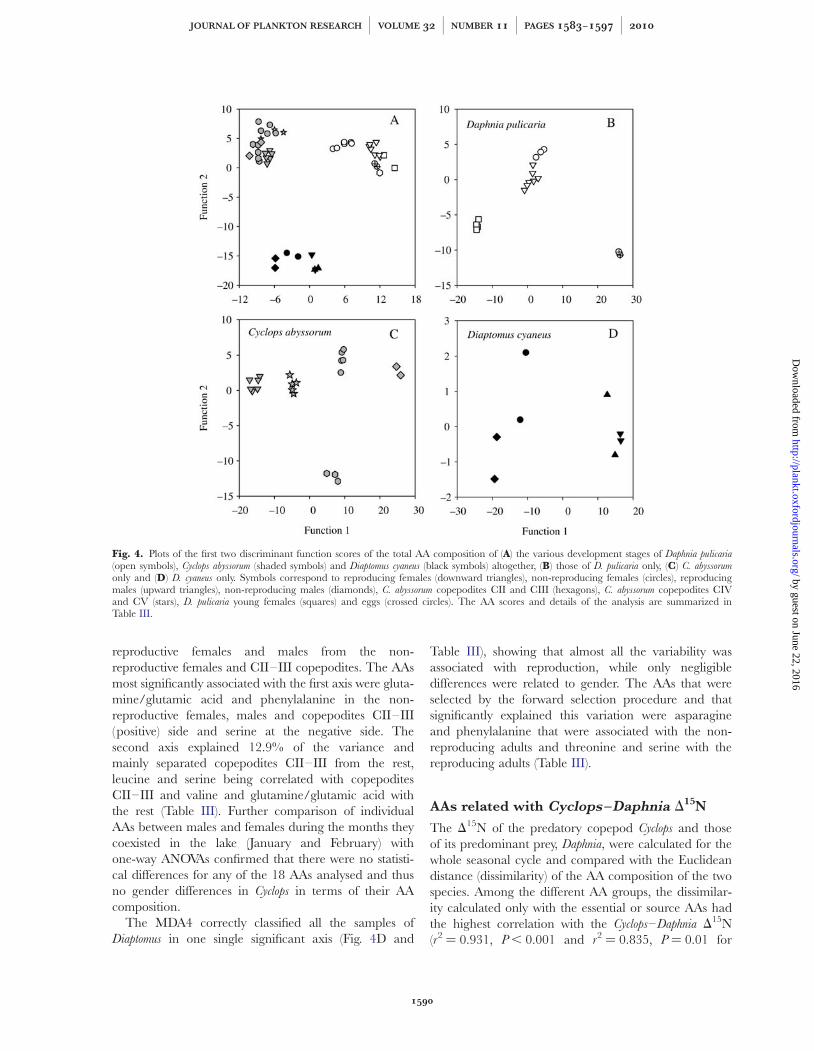
The AA composition of the three species studied wasspecies specific (Fig. 4A). The most abundant AAs forthe two copepods (in order of importance and nottaking into account asparagine/aspartic acid and gluta-mine/glutamic acid which were analysed together) werealanine, leucine, tyrosine, lysine and arginine, whichaltogether accounted for 37% of the TAA pool. For thecladoceran, the most abundant AAs were leucine, argi-nine, alanine, valine, threonine and lysine, accountingfor 40% of the TAA pool (Table II). The MDA1,including all the stages of the three species, was able toclassify the stages of the three species based on their AAcomposition (100% of the samples being correctlyclassified with the exception of half of the Diaptomus
non-reproductive females that were identified as males;Fig. 4A and Table III). The first axis of the MDA1explained 47% of the variance and mainly segregatedthe two copepods from the cladoceran. The AAs signifi-cantly correlated with this first axis were proline, serineand threonine, their relative concentrations beinghigher in Daphnia, and glycine and taurine, with higherrelative concentrations in the two copepods (Table III).The second axis explained 39% of the variance andsegregated Diaptomus from the other two species.Diaptomus had a higher alanine and serine relative
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1588
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

concentration, while Cyclops and Daphnia had higherglycine concentration.
Intraspecific variation
Within each species, there were no differences in therelative proportion of TAA, protein AA (PAA), FAA,essential AA and nonessential AA pools between mostof the different stages, with the exception of Cyclops
males that had a lower relative concentration than thefemales in the lake during the same period (2.9+ 0.1and 1.6+ 0.5% dry weight of TAA for females andmales, respectively; F1,3 ¼ 15.8, P ¼ 0.028) and Daphnia
eggs that had higher TAA relative concentration thanthe first cohort females (F1,8 ¼ 2.8, P ¼ 0.1 Tukey’s post
hoc test P ¼ 0.013) (Table I). In contrast, there was adecrease from non-reproducing females to reproducingfemales of FAA and essential AA pools of Cyclops (F1,9 ¼
20.5, P ¼ 0.001 and F1,9 ¼ 7.2, P ¼ 0.025 respectively),a slight decrease in the females of Daphnia (F1,8 ¼ 2.8,P ¼ 0.1 and F1,8 ¼ 3.3, P ¼ 0.1 respectively) and inboth males and females of Diaptomus (F1,6 ¼ 3.8, P ¼
0.1 and F1,6 ¼ 2.8, P ¼ 0.1, respectively).The MDA2 correctly classified 100% of the samples
of Daphnia (Fig. 4B and Table III). The first axes sum-marized 74.9% of the variance and separated the threestages analysed: first cohort females, second cohortfemales and eggs. The AAs most strongly associatedwith the axes were proline and arginine, more abundantin the eggs (positive) side, and glycine and leucine,more abundant in second generation females (negative)side. The second axis explained the remaining 22.4%of the variation and separated the second generationfemales and eggs (negative) side from the reproductiveand non-reproductive first generation females (positive)side. The AAs most strongly associated with the axeswere positively, threonine and glutamine/glutamic acidand negatively, arginine. Further comparison of individ-ual AAs between reproducing and non-reproducing firstcohort females with one-way ANOVAs indicated thatreproducing females had higher histidine, serine andcystine (P ¼ 0.003, P ¼ 0.003 and P ¼ 0.007, respect-ively) and lower phenylalanine (P ¼ 0.021).
For Cyclops, results of the MDA3 analysis showed that100% of the cases were correctly classified (Table III)(Fig. 4C). The first axes explained 78.2% of the varianceand mainly segregated the CIV–V copepodites, the
Fig. 3. Average Euclidean distance (with 10th and 25th percentiles)of the interspecific (shaded box plots) and intraspecific (white boxplots) distance in the amino acid composition. Distances werecomputed among all samples analysed and therefore the intraspecificdistances include both the distances between stages and within stages.Abbreviations stand for Cyclops abyssorum (C.a.), Daphnia pulicaria (D.p.)and Diaptomus cyaneus (D.c.). Letters above each box-plot denotesignificantly different distances from the one-way ANOVA Tukey’s posthoc test (P , 0.001).
Table I: Relative proportions of the different amino acid (AA) pools in the different species/stagesanalysed (mean + SD)
Species Stage/gender TAA (% DW) FAA (% TAA) PAA (% TAA) EAA (% TAA) NEAA (% TAA)
Cyclops abyssorum CII– III 47.5+13.4a 1.7+1.0a 98.3+1.0a 45.6+0.4a 54.4+0.4a
CIV–V 41.9+4.8a 2.4+0.9a 97.6+0.9a 45.9+2.4a 54.1+2.4a
Non-reproducing females 37.7+4.3a,b 2.7+0.4a,b 97.3+0.4a,b 45.4+0.5a,b 54.6+0.5a,b
Reproducing females 53.6+4.7a,c 1.4+0.5a,c 98.6+0.5a,c 44.7+0.3a,c 55.3+0.3a,c
Males 47.6+7.3a 1.6+0.5a 98.4+0.5a 45.2+1.6a 54.8+1.6a
Daphnia pulicaria First-cohort non-reproducing females 25.3+2.3a,b* 2.6+0.7a,b* 97.4+0.7a,b 45.5+1.2a,b* 54.1+0.5a,b*First-cohort reproducing females 27.3+1.9a,c* 1.9+0.4a,c* 98.1+0.4a,b 44.6+2.0a,c* 55.4+2.0a,c*Second-cohort females 29.3+5.0a,d 3.3+2.6a 96.7+2.6a 46.2+1.0a 53.8+1.0a
Eggs 34.1+6.6d 2.3+0.3a 97.7+0.3a 47.7+2.2a 52.3+2.2a
Diaptomus cyaneus Non-reproducing females 45.8+1.3a,b 1.6+0.6a,b 98.4+0.6a,b 45.1+0.0a,b 54.9+0.0a,b
Reproducing females 71.4+0.9a,c 1.0+0.1a,c 99.0+0.1a,b 45.3+0.4a,b 54.7+0.4a,b
Non-reproducing males 49.0+0.0a,b 1.3+0.1a,b 98.7+0.1a,b 45.1+0.4a,b 54.9+0.4a,b
Reproducing males 72.6+4.9a,c 1.0+0.1a,c 99.0+0.1a,b 45.4+0.5a,b 54.6+0.5a,b
Total AA (TAA) concentration is expressed as mass percentage of dry weight (DW) and is subdivided in the protein AA pool (PAA) and the free aminopool (FAA) that are expressed as the mass percentage of TAA. The relative proportions of essential and nonessential AA pools (EAA and NEAA,respectively) are also calculated. Letters after SDs denote ANOVA statistically significant stages (a, d) or reproducing versus non-reproducing females(b, c) at P , 0.05 or *P , 0.1.
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1589
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
reproductive females and males from the non-reproductive females and CII–III copepodites. The AAsmost significantly associated with the first axis were gluta-mine/glutamic acid and phenylalanine in the non-reproductive females, males and copepodites CII–III(positive) side and serine at the negative side. Thesecond axis explained 12.9% of the variance andmainly separated copepodites CII–III from the rest,leucine and serine being correlated with copepoditesCII–III and valine and glutamine/glutamic acid withthe rest (Table III). Further comparison of individualAAs between males and females during the months theycoexisted in the lake (January and February) withone-way ANOVAs confirmed that there were no statisti-cal differences for any of the 18 AAs analysed and thusno gender differences in Cyclops in terms of their AAcomposition.
The MDA4 correctly classified all the samples ofDiaptomus in one single significant axis (Fig. 4D and
Table III), showing that almost all the variability wasassociated with reproduction, while only negligibledifferences were related to gender. The AAs that wereselected by the forward selection procedure and thatsignificantly explained this variation were asparagineand phenylalanine that were associated with the non-reproducing adults and threonine and serine with thereproducing adults (Table III).
AAs related with Cyclops–Daphnia D15N
The D15N of the predatory copepod Cyclops and thoseof its predominant prey, Daphnia, were calculated for thewhole seasonal cycle and compared with the Euclideandistance (dissimilarity) of the AA composition of the twospecies. Among the different AA groups, the dissimilar-ity calculated only with the essential or source AAs hadthe highest correlation with the Cyclops–Daphnia D15N(r2 ¼ 0.931, P , 0.001 and r2 ¼ 0.835, P ¼ 0.01 for
Fig. 4. Plots of the first two discriminant function scores of the total AA composition of (A) the various development stages of Daphnia pulicaria(open symbols), Cyclops abyssorum (shaded symbols) and Diaptomus cyaneus (black symbols) altogether, (B) those of D. pulicaria only, (C) C. abyssorumonly and (D) D. cyaneus only. Symbols correspond to reproducing females (downward triangles), non-reproducing females (circles), reproducingmales (upward triangles), non-reproducing males (diamonds), C. abyssorum copepodites CII and CIII (hexagons), C. abyssorum copepodites CIVand CV (stars), D. pulicaria young females (squares) and eggs (crossed circles). The AA scores and details of the analysis are summarized inTable III.
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1590
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
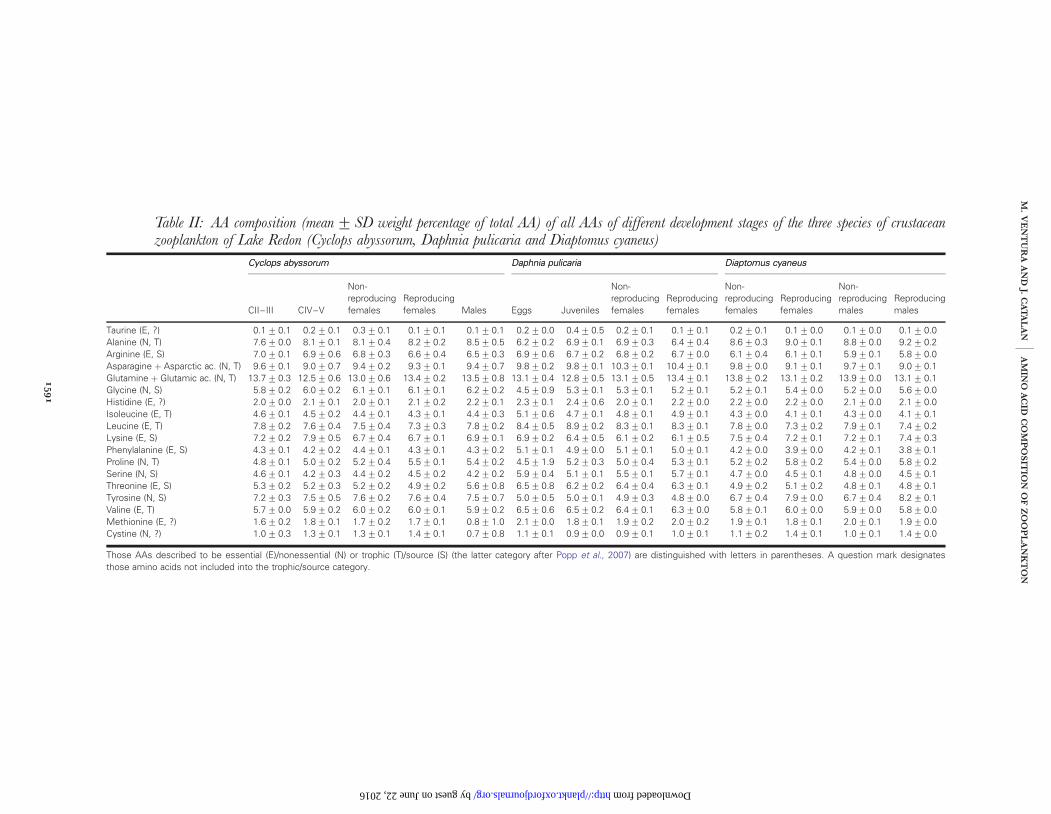
Table II: AA composition (mean+ SD weight percentage of total AA) of all AAs of different development stages of the three species of crustaceanzooplankton of Lake Redon (Cyclops abyssorum, Daphnia pulicaria and Diaptomus cyaneus)
Cyclops abyssorumCyclops abyssorum Daphnia pulicariaDaphnia pulicaria Diaptomus cyaneusDiaptomus cyaneus
CII–III CIV–V
Non-reproducingfemales
Reproducingfemales Males Eggs Juveniles
Non-reproducingfemales
Reproducingfemales
Non-reproducingfemales
Reproducingfemales
Non-reproducingmales
Reproducingmales
Taurine (E, ?) 0.1+0.1 0.2+0.1 0.3+0.1 0.1+0.1 0.1+0.1 0.2+0.0 0.4+0.5 0.2+0.1 0.1+0.1 0.2+0.1 0.1+0.0 0.1+0.0 0.1+0.0Alanine (N, T) 7.6+0.0 8.1+0.1 8.1+0.4 8.2+0.2 8.5+0.5 6.2+0.2 6.9+0.1 6.9+0.3 6.4+0.4 8.6+0.3 9.0+0.1 8.8+0.0 9.2+0.2Arginine (E, S) 7.0+0.1 6.9+0.6 6.8+0.3 6.6+0.4 6.5+0.3 6.9+0.6 6.7+0.2 6.8+0.2 6.7+0.0 6.1+0.4 6.1+0.1 5.9+0.1 5.8+0.0Asparagine þ Asparctic ac. (N, T) 9.6+0.1 9.0+0.7 9.4+0.2 9.3+0.1 9.4+0.7 9.8+0.2 9.8+0.1 10.3+0.1 10.4+0.1 9.8+0.0 9.1+0.1 9.7+0.1 9.0+0.1Glutamine þ Glutamic ac. (N, T) 13.7+0.3 12.5+0.6 13.0+0.6 13.4+0.2 13.5+0.8 13.1+0.4 12.8+0.5 13.1+0.5 13.4+0.1 13.8+0.2 13.1+0.2 13.9+0.0 13.1+0.1Glycine (N, S) 5.8+0.2 6.0+0.2 6.1+0.1 6.1+0.1 6.2+0.2 4.5+0.9 5.3+0.1 5.3+0.1 5.2+0.1 5.2+0.1 5.4+0.0 5.2+0.0 5.6+0.0Histidine (E, ?) 2.0+0.0 2.1+0.1 2.0+0.1 2.1+0.2 2.2+0.1 2.3+0.1 2.4+0.6 2.0+0.1 2.2+0.0 2.2+0.0 2.2+0.0 2.1+0.0 2.1+0.0Isoleucine (E, T) 4.6+0.1 4.5+0.2 4.4+0.1 4.3+0.1 4.4+0.3 5.1+0.6 4.7+0.1 4.8+0.1 4.9+0.1 4.3+0.0 4.1+0.1 4.3+0.0 4.1+0.1Leucine (E, T) 7.8+0.2 7.6+0.4 7.5+0.4 7.3+0.3 7.8+0.2 8.4+0.5 8.9+0.2 8.3+0.1 8.3+0.1 7.8+0.0 7.3+0.2 7.9+0.1 7.4+0.2Lysine (E, S) 7.2+0.2 7.9+0.5 6.7+0.4 6.7+0.1 6.9+0.1 6.9+0.2 6.4+0.5 6.1+0.2 6.1+0.5 7.5+0.4 7.2+0.1 7.2+0.1 7.4+0.3Phenylalanine (E, S) 4.3+0.1 4.2+0.2 4.4+0.1 4.3+0.1 4.3+0.2 5.1+0.1 4.9+0.0 5.1+0.1 5.0+0.1 4.2+0.0 3.9+0.0 4.2+0.1 3.8+0.1Proline (N, T) 4.8+0.1 5.0+0.2 5.2+0.4 5.5+0.1 5.4+0.2 4.5+1.9 5.2+0.3 5.0+0.4 5.3+0.1 5.2+0.2 5.8+0.2 5.4+0.0 5.8+0.2Serine (N, S) 4.6+0.1 4.2+0.3 4.4+0.2 4.5+0.2 4.2+0.2 5.9+0.4 5.1+0.1 5.5+0.1 5.7+0.1 4.7+0.0 4.5+0.1 4.8+0.0 4.5+0.1Threonine (E, S) 5.3+0.2 5.2+0.3 5.2+0.2 4.9+0.2 5.6+0.8 6.5+0.8 6.2+0.2 6.4+0.4 6.3+0.1 4.9+0.2 5.1+0.2 4.8+0.1 4.8+0.1Tyrosine (N, S) 7.2+0.3 7.5+0.5 7.6+0.2 7.6+0.4 7.5+0.7 5.0+0.5 5.0+0.1 4.9+0.3 4.8+0.0 6.7+0.4 7.9+0.0 6.7+0.4 8.2+0.1Valine (E, T) 5.7+0.0 5.9+0.2 6.0+0.2 6.0+0.1 5.9+0.2 6.5+0.6 6.5+0.2 6.4+0.1 6.3+0.0 5.8+0.1 6.0+0.0 5.9+0.0 5.8+0.0Methionine (E, ?) 1.6+0.2 1.8+0.1 1.7+0.2 1.7+0.1 0.8+1.0 2.1+0.0 1.8+0.1 1.9+0.2 2.0+0.2 1.9+0.1 1.8+0.1 2.0+0.1 1.9+0.0Cystine (N, ?) 1.0+0.3 1.3+0.1 1.3+0.1 1.4+0.1 0.7+0.8 1.1+0.1 0.9+0.0 0.9+0.1 1.0+0.1 1.1+0.2 1.4+0.1 1.0+0.1 1.4+0.0
Those AAs described to be essential (E)/nonessential (N) or trophic (T)/source (S) (the latter category after Popp et al., 2007) are distinguished with letters in parentheses. A question mark designatesthose amino acids not included into the trophic/source category.
M.
VE
NT
UR
AA
ND
J.C
AT
AL
ANj
AM
INO
AC
IDC
OM
PO
SIT
ION
OF
ZO
OP
LA
NK
TO
N
1591
by guest on June 22, 2016 http://plankt.oxfordjournals.org/ Downloaded from
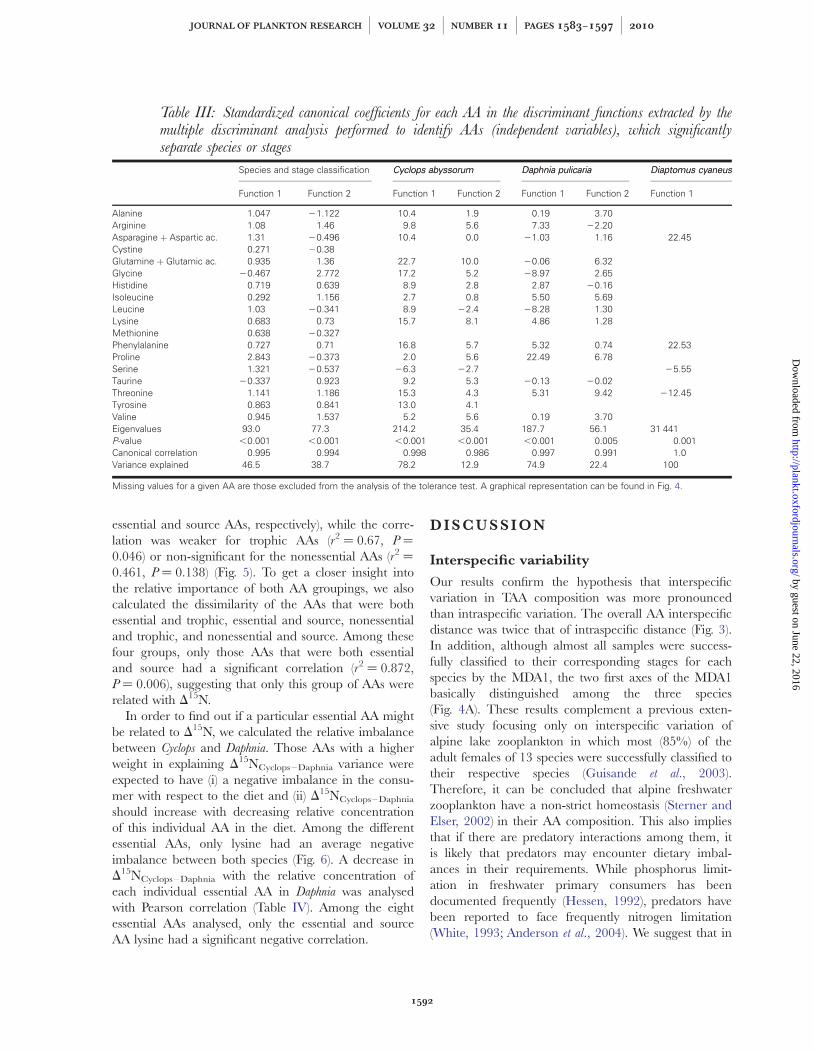
essential and source AAs, respectively), while the corre-lation was weaker for trophic AAs (r2 ¼ 0.67, P ¼
0.046) or non-significant for the nonessential AAs (r2 ¼
0.461, P ¼ 0.138) (Fig. 5). To get a closer insight intothe relative importance of both AA groupings, we alsocalculated the dissimilarity of the AAs that were bothessential and trophic, essential and source, nonessentialand trophic, and nonessential and source. Among thesefour groups, only those AAs that were both essentialand source had a significant correlation (r2 ¼ 0.872,P ¼ 0.006), suggesting that only this group of AAs wererelated with D15N.
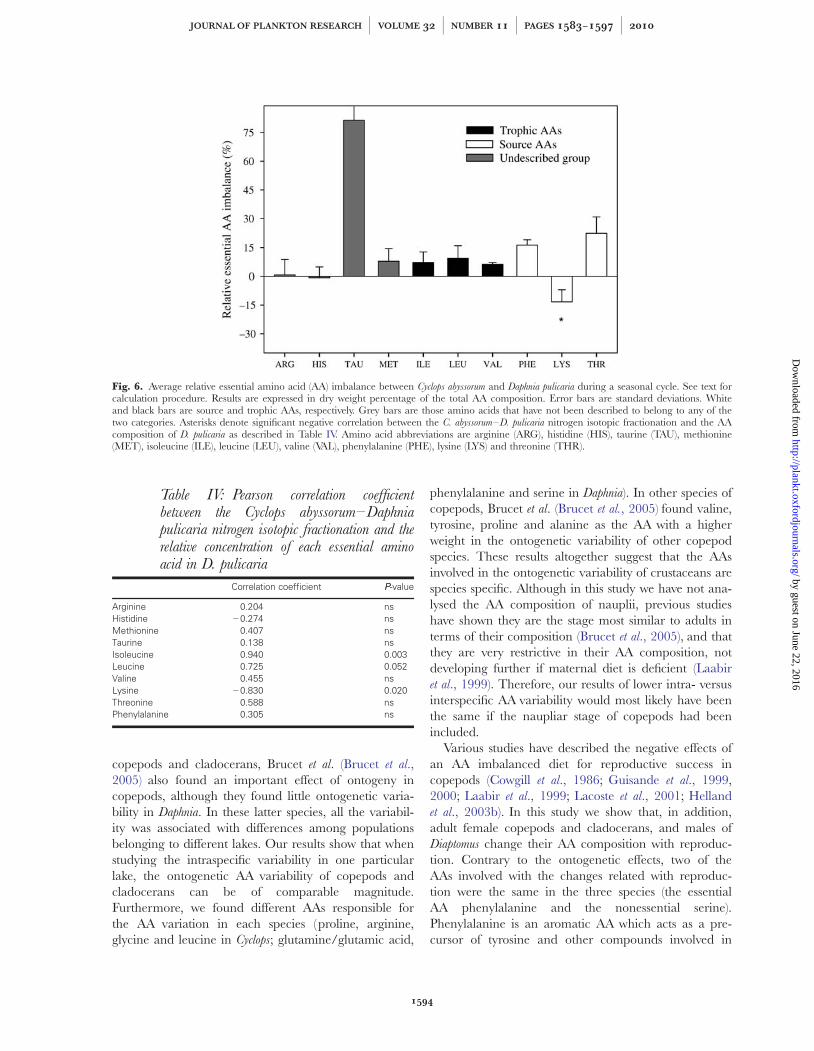
In order to find out if a particular essential AA mightbe related to D15N, we calculated the relative imbalancebetween Cyclops and Daphnia. Those AAs with a higherweight in explaining D15NCyclops – Daphnia variance wereexpected to have (i) a negative imbalance in the consu-mer with respect to the diet and (ii) D15NCyclops – Daphnia
should increase with decreasing relative concentrationof this individual AA in the diet. Among the differentessential AAs, only lysine had an average negativeimbalance between both species (Fig. 6). A decrease inD15NCyclops – Daphnia with the relative concentration ofeach individual essential AA in Daphnia was analysedwith Pearson correlation (Table IV). Among the eightessential AAs analysed, only the essential and sourceAA lysine had a significant negative correlation.
D I S C U S S I O N
Interspecific variability
Our results confirm the hypothesis that interspecificvariation in TAA composition was more pronouncedthan intraspecific variation. The overall AA interspecificdistance was twice that of intraspecific distance (Fig. 3).In addition, although almost all samples were success-fully classified to their corresponding stages for eachspecies by the MDA1, the two first axes of the MDA1basically distinguished among the three species(Fig. 4A). These results complement a previous exten-sive study focusing only on interspecific variation ofalpine lake zooplankton in which most (85%) of theadult females of 13 species were successfully classified totheir respective species (Guisande et al., 2003).Therefore, it can be concluded that alpine freshwaterzooplankton have a non-strict homeostasis (Sterner andElser, 2002) in their AA composition. This also impliesthat if there are predatory interactions among them, itis likely that predators may encounter dietary imbal-ances in their requirements. While phosphorus limit-ation in freshwater primary consumers has beendocumented frequently (Hessen, 1992), predators havebeen reported to face frequently nitrogen limitation(White, 1993; Anderson et al., 2004). We suggest that in
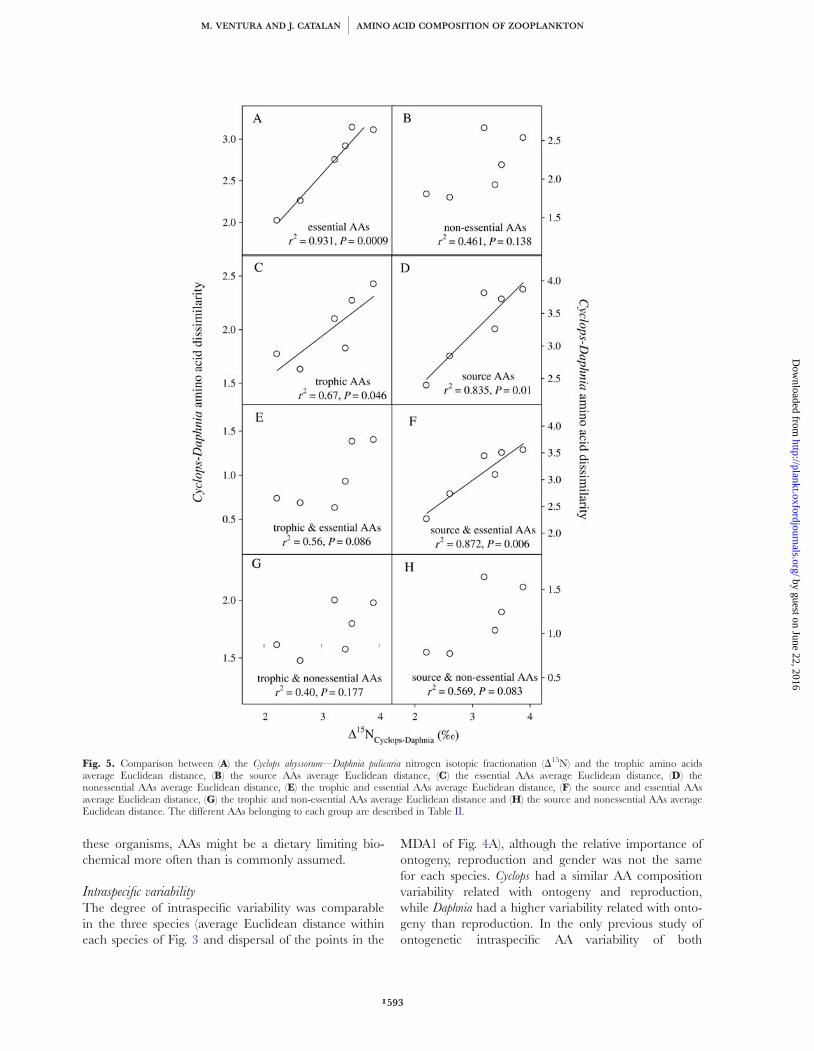
Table III: Standardized canonical coefficients for each AA in the discriminant functions extracted by themultiple discriminant analysis performed to identify AAs (independent variables), which significantlyseparate species or stages
Species and stage classification Cyclops abyssorumCyclops abyssorum Daphnia pulicariaDaphnia pulicaria Diaptomus cyaneusDiaptomus cyaneus
Function 1 Function 2 Function 1 Function 2 Function 1 Function 2 Function 1
Alanine 1.047 21.122 10.4 1.9 0.19 3.70Arginine 1.08 1.46 9.8 5.6 7.33 22.20Asparagine þ Aspartic ac. 1.31 20.496 10.4 0.0 21.03 1.16 22.45Cystine 0.271 20.38Glutamine þ Glutamic ac. 0.935 1.36 22.7 10.0 20.06 6.32Glycine 20.467 2.772 17.2 5.2 28.97 2.65Histidine 0.719 0.639 8.9 2.8 2.87 20.16Isoleucine 0.292 1.156 2.7 0.8 5.50 5.69Leucine 1.03 20.341 8.9 22.4 28.28 1.30Lysine 0.683 0.73 15.7 8.1 4.86 1.28Methionine 0.638 20.327Phenylalanine 0.727 0.71 16.8 5.7 5.32 0.74 22.53Proline 2.843 20.373 2.0 5.6 22.49 6.78Serine 1.321 20.537 26.3 22.7 25.55Taurine 20.337 0.923 9.2 5.3 20.13 20.02Threonine 1.141 1.186 15.3 4.3 5.31 9.42 212.45Tyrosine 0.863 0.841 13.0 4.1Valine 0.945 1.537 5.2 5.6 0.19 3.70Eigenvalues 93.0 77.3 214.2 35.4 187.7 56.1 31 441P-value ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 0.005 0.001Canonical correlation 0.995 0.994 0.998 0.986 0.997 0.991 1.0Variance explained 46.5 38.7 78.2 12.9 74.9 22.4 100
Missing values for a given AA are those excluded from the analysis of the tolerance test. A graphical representation can be found in Fig. 4.
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1592
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
these organisms, AAs might be a dietary limiting bio-chemical more often than is commonly assumed.
Intraspecific variabilityThe degree of intraspecific variability was comparablein the three species (average Euclidean distance withineach species of Fig. 3 and dispersal of the points in the
MDA1 of Fig. 4A), although the relative importance ofontogeny, reproduction and gender was not the samefor each species. Cyclops had a similar AA compositionvariability related with ontogeny and reproduction,while Daphnia had a higher variability related with onto-geny than reproduction. In the only previous study ofontogenetic intraspecific AA variability of both
Fig. 5. Comparison between (A) the Cyclops abyssorum—Daphnia pulicaria nitrogen isotopic fractionation (D15N) and the trophic amino acidsaverage Euclidean distance, (B) the source AAs average Euclidean distance, (C) the essential AAs average Euclidean distance, (D) thenonessential AAs average Euclidean distance, (E) the trophic and essential AAs average Euclidean distance, (F) the source and essential AAsaverage Euclidean distance, (G) the trophic and non-essential AAs average Euclidean distance and (H) the source and nonessential AAs averageEuclidean distance. The different AAs belonging to each group are described in Table II.
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1593
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
copepods and cladocerans, Brucet et al. (Brucet et al.,2005) also found an important effect of ontogeny incopepods, although they found little ontogenetic varia-bility in Daphnia. In these latter species, all the variabil-ity was associated with differences among populationsbelonging to different lakes. Our results show that whenstudying the intraspecific variability in one particularlake, the ontogenetic AA variability of copepods andcladocerans can be of comparable magnitude.Furthermore, we found different AAs responsible forthe AA variation in each species (proline, arginine,glycine and leucine in Cyclops; glutamine/glutamic acid,
phenylalanine and serine in Daphnia). In other species ofcopepods, Brucet et al. (Brucet et al., 2005) found valine,tyrosine, proline and alanine as the AA with a higherweight in the ontogenetic variability of other copepodspecies. These results altogether suggest that the AAsinvolved in the ontogenetic variability of crustaceans arespecies specific. Although in this study we have not ana-lysed the AA composition of nauplii, previous studieshave shown they are the stage most similar to adults interms of their composition (Brucet et al., 2005), and thatthey are very restrictive in their AA composition, notdeveloping further if maternal diet is deficient (Laabiret al., 1999). Therefore, our results of lower intra- versusinterspecific AA variability would most likely have beenthe same if the naupliar stage of copepods had beenincluded.
Various studies have described the negative effects ofan AA imbalanced diet for reproductive success incopepods (Cowgill et al., 1986; Guisande et al., 1999,2000; Laabir et al., 1999; Lacoste et al., 2001; Hellandet al., 2003b). In this study we show that, in addition,adult female copepods and cladocerans, and males ofDiaptomus change their AA composition with reproduc-tion. Contrary to the ontogenetic effects, two of theAAs involved with the changes related with reproduc-tion were the same in the three species (the essentialAA phenylalanine and the nonessential serine).Phenylalanine is an aromatic AA which acts as a pre-cursor of tyrosine and other compounds involved in
Fig. 6. Average relative essential amino acid (AA) imbalance between Cyclops abyssorum and Daphnia pulicaria during a seasonal cycle. See text forcalculation procedure. Results are expressed in dry weight percentage of the total AA composition. Error bars are standard deviations. Whiteand black bars are source and trophic AAs, respectively. Grey bars are those amino acids that have not been described to belong to any of thetwo categories. Asterisks denote significant negative correlation between the C. abyssorum–D. pulicaria nitrogen isotopic fractionation and the AAcomposition of D. pulicaria as described in Table IV. Amino acid abbreviations are arginine (ARG), histidine (HIS), taurine (TAU), methionine(MET), isoleucine (ILE), leucine (LEU), valine (VAL), phenylalanine (PHE), lysine (LYS) and threonine (THR).
Table IV: Pearson correlation coefficientbetween the Cyclops abyssorum–Daphniapulicaria nitrogen isotopic fractionation and therelative concentration of each essential aminoacid in D. pulicaria
Correlation coefficient PP-value
Arginine 0.204 nsHistidine 20.274 nsMethionine 0.407 nsTaurine 0.138 nsIsoleucine 0.940 0.003Leucine 0.725 0.052Valine 0.455 nsLysine 20.830 0.020Threonine 0.588 nsPhenylalanine 0.305 ns
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1594
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

important physiological functions, while serine isinvolved in lipid biosynthesis and also a precursor ofmelanine. The results of this study suggest that they arerelevant during the reproduction of crustaceans. Inaddition to the changes in individual AAs, we alsofound a tendency for the FAA content to decrease fromnon-reproductive to reproductive females. This agreeswith other studies showing that eggs contain a largerproportion of FAAs and that the AA composition ofeggs is decisive for the successful development of theembryos (Guisande et al., 1999; Laabir et al., 1999;Helland et al., 2003a, b).
Contrasting with the ontogenetic and reproductioneffects, we found no changes in the AA compositionassociated with gender in the two copepod species. Theonly difference attributed to gender was a higher pro-portion of the FAA pool in Cyclops females comparedwith males. This contrasts with the equal proportionbetween both sexes in Diaptomus, which might reflect thedifferent reproductive strategies described for freshwatercalanoids and cyclopoids. This result also agrees withthe findings of a parallel study (Ventura and Catalan,2005), in which Diaptomus males were found to followsimilar elemental composition changes with reproduc-tion as females, while the elemental content of Cyclops
males did not change with time. This was attributed tothe requirement of Diaptomus to mate every egg clutch,while Cyclops females can produce multiple clutches ofeggs with only one impregnation. Therefore, the presentresults on the changes in the FAA pool also support thehypothesis that the different reproductive strategies ofcopepods have a relevant effect on the elemental andbiochemical homeostasis of males.
AAs involved in D15N
There are different physiological factors that mightinfluence D15N, including those related with fasting,growth and diet quality in terms of protein quantity orquality (reviewed by Martınez del Rio and Wolf, 2005).While most of these factors have been demonstratedeither experimentally or through field studies, the effectsof varying AA composition have scarcely been studied(Martınez del Rio et al., 2009). In a previous study, weshowed that the carbon isotopic variation in Cyclops
juveniles could be explained by fitting an isotopicgrowth model based on feeding entirely on Daphnia.However, this was not the case for nitrogen isotopicvariability. Cyclops nitrogen isotopic compositionvariation and the Cyclops to Daphnia D15N wereclosely correlated to the dissimilarity in the AA compo-sition between the two species (Ventura and Catalan,2008).
In this study, we show that among the different AAs,only those that are both essential and source AAs had ahigh and significant correlation with D15N. Amongthese, only lysine had a negative imbalance in the con-sumer as it increased its relative concentration in thediet when the D15N decreased. This suggests that theimbalance of only one AA can be responsible for a highmetabolic turnover, which can ultimately be responsiblefor a higher D15N. Our results therefore are in agree-ment with the hypothesis that an AA deficiency can beresponsible for a variable D15N (Martınez del Rio andWolf, 2005).
Although the recognition of a variable D15N mightseem a difficulty for a widespread use of 15N as atrophic position estimator, it opens new perspectives forits interpretation, since it could be used as an indicatorof the relative AA imbalance of an animal and as a con-sequence, an indicator of the relative degree of AAlimitation and as a result of the overall nitrogen limit-ation. Although this might seem difficult to apply togeneral field studies, future studies using individual AAstable isotope composition of carbon or nitrogen (e.g.Larsen et al., 2009) might substantially help interpret-ations in stable isotope trophic ecology.
AC K N OW L E D G E M E N T S
We thank LL. Camarero, B. Claramunt, A. Miro,P. Renom, T. Buchaca and G. Cots for their assistance infield, and R. Franco for laboratory assistance. The TAAanalyses were performed in the Serveis CientıficoTecnics of the University of Barcelona. A.M. Poulsenassisted in linguistic/editorial corrections. T. Buchaca,S. Pla, T. Larsen and anonymous reviewers are acknowl-edged for their helpful comments on the manuscript.
F U N D I N G
This is a study of the Biogeodynamics and BiodiversityGroup (CSIC-UB) and the group of Ecology ofEnvironmental Changes (2005SGR 00510). Theresearch was supported by the research projectTRAZAS (CGL2004-02989) and the EuropeanCommission projects MOLAR (ENV4-CT95-0007) andEUROLIMPACS (GOCE-CT-2003-505540). M.V. waspartially supported by a Ramon y Cajal grant.
R E F E R E N C E S
Anderson, T. R., Boersma, M. and Raubenheimer, D. (2004)Stoichiometry: linking elements to biochemicals. Ecology, 85, 1193–1202.
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1595
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

Boechat, I. G. and Adrian, R. (2005) Biochemical composition of algi-vorous freshwater ciliates: you are not what you eat. Fems. Microbiol.
Ecol., 53, 393–400.
Boechat, I. G. and Adrian, R. (2006) Evidence for biochemical limit-ation of population growth and reproduction of the rotifer Keratella
quadrata fed with freshwater protists. J. Plankton Res., 28, 1027–1038.
Brucet, S., Boix, D., Lopez-Flores, R. et al. (2005) Ontogenic changesof amino acid composition in planktonic crustacean species. Mar.
Biol., 148, 131–139.
Camarero, L., Felip, M., Ventura, M. et al. (1999) The relative impor-tance of the planktonic food web in the carbon cycle of an oligo-trophic mountain lake in a poorly vegetated catchment (Redo,Pyrenees). J. Limnol., 58, 203–212.
Claybrook, D. L. (1983) Nitrogen metabolism. In Mantel, L. H. (ed.),The biology of crustacea. Internal anatomy and physiological regulation.Academic Press, New York, pp. 163–213.
Conceicao, L. E. C., Grasdalen, H. and Ronnestad, I. (2003) Aminoacid requirements of fish larvae and post-larvae: new tools andrecent findings. Aquaculture, 227, 221–232.
Conklin, D. and Provasoli, L. (1977) Nutritional requirements of thewater flea Moina macrocopa. Biol. Bull., 152, 337–350.
Cowey, C. B. and Corner, E. D. S. (1963) On the nutrition andmetabolism of zooplankton II. The relationship between the marinecopepod Calanus helgolandicus and particulate material in playmouthsea water, in terms of amino acid composition. J. Mar. Biol. Assoc.
UK, 43, 495–511.
Cowgill, U. M., Emmel, H. W., Hopkins, D. L. et al. (1986) Variation inchemical composition, reproductive success and body weightof Daphnia magna in relation to diet. Int. Rev. Gesamten Hydrobiol., 71,79–99.
D’mello, J. P. F. (1994) Amino Acids in Farm Animal Nutrition. CABInternational, Guildford.
Dabrowski, K. and Rusiecki, M. (1983) Content of total and freeamino acids in zooplanktonic food of fish larvae. Aquaculture, 30,31–42.
Felip, M., Bartumeus, F., Halac, S. et al. (1999) Microbial planktonassemblages, composition and biomass, during two ice-free periodsin a deep high mountain lake (Estany Redo, Pyrenees). J. Limnol.,58, 193–202.
Fountoulakis, M. and Lahm, H. W. (1998) Hydrolysis and amino acidcomposition analysis of proteins. J. Chromatogr. A, 826, 109–134.
Guisande, C., Maneiro, I. and Riveiro, I. (1999) Homeostasis in theessential amino acid composition of the marine copepod Euterpina
acutifrons. Limnol. Oceanogr., 44, 691–696.
Guisande, C., Riveiro, I. and Maneiro, I. (2000) Comparisons amongthe amino acid composition of females, eggs and food to determinethe relative importance of food quantity and food quality tocopepod reproduction. Mar. Ecol. Prog. Ser., 202, 135–142.
Guisande, C., Bartumeus, F., Ventura, M. et al. (2003) Role of foodpartitioning in structuring the zooplankton community in mountainlakes. Oecologia, 136, 627–634.
Helland, S., Triantaphyllidis, G. V., Fyhn, H. J. et al. (2000) Modulationof the free amino acid pool and protein content in populations of thebrine shrimp Artemia spp. Mar. Biol., 137, 1005–1016.
Helland, S., Nejstgaard, J. C., Fyhn, H. J. et al. (2003a) Effects ofstarvation, season, and diet on the free amino acid andprotein content of Calanus finmarchicus females. Mar. Biol., 143,297–306.
Helland, S., Nejstgaard, J. C., Humlen, R. et al. (2003b) Effects ofseason and maternal food on Calanus finmarchicus reproduction, withemphasis on free amino acids. Mar. Biol., 142, 1141–1151.
Hessen, D. O. (1992) Nutrient element limitation of zooplankton pro-duction. The Am. Nat., 140, 799–814.
Jeffries, H. P. (1969) Seasonal composition of temperate plankton com-munities: free amino acids. Limnol. Oceanogr., 14, 41–52.
Jeffries, H. P. and Alzara, L. (1970) Dominace-diversity relationshipsof the free amino acids in coastal zooplankton. Comp. Biochem.
Physiol., 37, 215–223.
Kleppel, G. S. and Burkart, C. A. (1995) Egg production and thenutritional environment of Acartia tonsa: the role of food quality incopepod nutrition. ICES J. Mar. Sci., 52, 297–304.
Kreeger, D. A., Hawkins, A. J. S. and Bayne, B. L. (1996) Use ofdual-labeled microcapsules to discern the physiological fatesof assimilated carbohydrate, protein carbon, and protein nitrogen insuspension-feeding organisms. Limnol. Oceanogr., 41, 208–215.
Laabir, M., Poulet, S. A., Cueff, A. et al. (1999) Effect of diet on levelsof amino acids during embryonic and naupliar development of thecopepod Calanus helgolandicus. Mar. Biol., 134, 89–98.
Lacoste, A., Poulet, S. A., Cueff, A. et al. (2001) New evidence of thecopepod maternal food effects on reproduction. J. Exp. Mar. Biol.
Ecol., 259, 85–107.
Larsen, T., Taylor, D. L., Leigh, M. B. et al. (2009) Stable isotope fin-gerprinting: a novel method for identifying plant, fungal or bac-terial origins of amino acids. Ecology, 90, 3526.
Legendre, P. and Legendre, L. (1998) Numerical Ecology. ElsevierScience, Amsterdam.
Martınez Del Rio, C. and Wolf, B. O. (2005) Mass-balance modelsfor animal isotopic ecology. In Starck, J. M. and Wang, T. (eds),Physiological and ecological adaptations to feeding in vertebrates. SciencePublishers, Enfield, New Hampshire, pp. 141–174.
Martınez Del Rio, C. M., Wolf, N., Carleton, S. A. et al. (2009)Isotopic ecology ten years after a call for more laboratory exper-iments. Biol. Rev., 84, 91–111.
Mcclelland, J. W. and Montoya, J. P. (2002) Trophic relationships andthe nitrogen isotopic composition of amino acids in plankton.Ecology, 83, 2173–2180.
Mente, E., Houlihan, D. F. and Smith, K. (2001) Growth, feeding fre-quency, protein turnover, and amino acid metabolism in Europeanlobster Homarus gammarus L. J. Exp. Zool., 289, 432.
Muller-Navarra, D. C., Brett, M. T., Liston, A. M. et al. (2000) Ahighly unsaturated fatty acid predicts carbon transfer betweenprimary producers and consumers. Nature, 403, 74–77.
O’brien, D. M., Boggs, C. L. and Fogel, M. L. (2003) Pollen feedingin the butterfly Heliconius charitonia: isotopic evidence for essentialamino acid transfer from pollen to eggs. Proc. R. Soc. B Biol. Sci.,270, 2631–2636.
Popp, B. N., Graham, B. S., Olson, R. J. et al. (2007) Insight into thetrophic ecology of yellowfin tuna, Thunnus albacares, from compound-specific nitrogen isotope analysis of proteinaceous amino acids. InDawson, T. and Seigwolf, R. (eds), Isotopes as Tracers in Ecological
Change. Elsevier, New York, pp. 173–190.
Ramsay, S. L. and Houston, D. C. (1998) The effect of dietary aminoacid composition on egg production in blue tits. Proc. R. Soc. B Biol.
Sci., 265, 1401–1405.
Reeds, P. J. (2000) Dispensable and Indispensable amino acids forHumans. J. Nutr., 130, 1835–1840.
JOURNAL OF PLANKTON RESEARCH j VOLUME 32 j NUMBER 11 j PAGES 1583–1597 j 2010
1596
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from

Robbins, C. T., Felicetti, L. A. and Sponheimer, M. (2005) The effectof dietary protein quality on nitrogen isotope discrimination inmammals and birds. Oecologia, 144, 534–540.
Ronnestad, I., Tonheim, S. K., Fyhn, H. J. et al. (2003) The supply ofamino acids during early feeding stages of marine fish larvae: areview of recent findings. Aquaculture, 227, 147–164.
Spackman, D. H., Stein, W. H. and Moore, S. (1958) Automaticrecording apparatus for use in the chromatography of amino acids.Anal. Chem., 30, 1190–1206.
Srinivasagam, R. T., Raymont, J. E. G., Moodie, C. F. and Raymont,K. K. B. (1971) Biochemical studies on marine zooplankton X.The amino acid composition of Euphausia superba, Meganyctiphanes
norvegica and Neomysis integer. J. Mar. Biol. Assoc. U.K., 51, 917–925.
Sterner, R. W. and Elser, J. J. (2002) Ecological Stoichiometry. PrincetonUniversity Press, Princeton.
Strayer, L. (1988) Biochemistry, W.H. Freeman and company, New York.1084 pp.
Suyama, M., Nakajima, K. and Nonaka, J. (1965) Studies on theprotein and non-protein nitrogenous constituents of Euphausia. Bull.
Japan. Soc. Sci. Fish., 31, 302–306.
Underwood, A. J. (1997) Experiments in Ecology: Their Logical Design and
Interpretation using Analysis of Variance. Cambridge University Press,Cambridge.
Ventura, M. (2006) Linking biochemical and elemental compositionof freshwater and marine crustacean zooplankton. Mar. Ecol. Prog.
Ser., 327, 233–246.
Ventura, M. and Catalan, J. (2005) Reproduction as one ofthe main causes of temporal variability in the elementalcomposition of zooplankton. Limnol. Oceanogr., 50, 2043–2056.
Ventura, M. and Catalan, J. (2008) Incorporating life histories anddiet quality in stable isotope interpretations of crustacean zooplank-ton. Freshwater Biol., 53, 1453–1469.
White, T. C. R. (1993) The Inadequate Environment: Nitrogen and the
Abundance of Animals. Springer-Verlag, New York.
M. VENTURA AND J. CATALAN j AMINO ACID COMPOSITION OF ZOOPLANKTON
1597
by guest on June 22, 2016http://plankt.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















