
UTILIZACIÓN DE VERTEBRADOS DURANTE EL PERÍODO FORMATIVO EN PUNTA TAMES, COSTA ARREICA DE...
197
UNIVERSIDAD INTERNACIONAL SEK FACULTAD DE ESTUDIOS DEL PATRIMONIO CULTURAL ARQUEOLOGÍA UTILIZACIÓN DE VERTEBRADOS DURANTE EL PERÍODO FORMATIVO EN PUNTA TAMES, COSTA ARREICA DE ANTOFAGASTA. MEMORIA PARA OPTAR AL TÍTULO DE ARQUEÓLOGA ALUMNA: ROCÍO MAC-LEAN PROFESORA GUÍA: VERÓNICA BAEZA TUTORA: VICTORIA CASTRO FONDECYT 1100951 SANTIAGO, MARZO 2014
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of UTILIZACIÓN DE VERTEBRADOS DURANTE EL PERÍODO FORMATIVO EN PUNTA TAMES, COSTA ARREICA DE...

UNIVERSIDAD INTERNACIONAL SEK FACULTAD DE ESTUDIOS DEL PATRIMONIO CULTURAL ARQUEOLOGÍA
UTILIZACIÓN DE VERTEBRADOS DURANTE
EL PERÍODO FORMATIVO EN PUNTA TAMES,
COSTA ARREICA DE ANTOFAGASTA.
MEMORIA PARA OPTAR AL TÍTULO DE ARQUEÓLOGA
ALUMNA: ROCÍO MAC-LEAN
PROFESORA GUÍA: VERÓNICA BAEZA
TUTORA: VICTORIA CASTRO
FONDECYT 1100951
SANTIAGO, MARZO 2014

A mis padres,
Consuelo Mac-Lean y Jorge Robotham.
Gracias por su ayuda incondicional y por alentar el desarrollo de mis sueños.

AGRADECIMIENTOS
Quisiera expresar mi más profundo agradecimiento a aquellas personas e instituciones
que hicieron posible el desarrollo de éste trabajo: al equipo Fondecyt 1100951, en
especial a la profesora Victoria Castro por su paciencia y confianza. A las profesoras
Verónica Baeza e Isabel Cartagena por su apoyo constante, correcciones y comentarios.
A mis amigos, Carolina Caro, Consuelo Iglesis, Francisco Vera, Catalina Rodillo, Gonzalo
Messina y a todos quienes me apoyaron y acompañaron en este largo proceso que al fin
se ve materializado, éste trabajo está dedicado a ustedes.

CONTENIDOS:
INTRODUCCIÓN: 1
CAPÍTULO 1 5
Antecedentes geográficos 5
1.1. Geografía del Norte Árido: 5
1.2. Geografía de la región de Antofagasta: 6
1.3. Geografía de Punta Tames: 10
1.4. Ecosistema marino: 12
1.4.1. Fenómeno del Niño: 15
Antecedentes Zoogeográfícos: 15
1.5. Vertebrados: 15
1.6. Vertebrados endémicos de la región de Antofagasta: 16
1.6.1 Herpetofauna: 16
1.6.2. Ictiofauna: 17
1.6.3. Avifauna: 18
1.6.4. Mastofauna: 19
Antecedentes Arqueológicos: 21
1.7. Antecedentes previos al Período Formativo: 21
1.8. Período Formativo: 23
1.8.1. Período Formativo en el Norte Árido: 23
1.8.2. Período Formativo en la costa arreica: 25
1.8.3. Intercambio a larga distancia en la región de Antofagasta: 27
1.8.4. Período Formativo en el tramo Punta Atala - Punta Tames: 29
1.8.5. Período Formativo en Punta Tames: 31
1.9. Utilización de vertebrados: 42
1.9.1. Consumo: 42
1.9.2. Industria ósea: 45
1.10. Utilización de invertebrados marinos: 47
CAPÍTULO 2 50
2.1. Problemática y justificación: 50
2.2. Objetivos: 52
2.2.1. Objetivo general: 52
2.2.2. Objetivos específicos: 52
2.3. Marco teórico: 52
2.3.1. Grupos litoraleños y adaptación: 53
2.3.2. Arqueología de los animales: 56

2.3.3. Procesos de formación de sitios arqueológicos e historias tafonómicas: 60
CAPÍTULO 3 64
3. Marco metodológico: 64
3.1. Trabajo de campo: 64
3.2. Trabajo de laboratorio: 64
3.2.1. Análisis: 65
3.2.2. Variables: 65
3.3. Trabajo de gabinete: 71
3.3.1. Gráficos y tablas: 71
3.3.2. Métodos para calcular la abundancia taxonómica: 73
3.3.3. Métodos para calcular la abundancia anatómica: 73
3.3.4. Articulación de datos: 74
CAPÍTULO 4 75
4. Resultados: 75
4.1. Muestra: 75
4.2. CF3: 77
4.3. P3: 84
4.4. PC8: 95
4.5. Chungungo 6: 99
4.6. Perfil etario: 102
4.7. Análisis de resultados: 103
4.7.1. Diversidad taxonómica: 103
4.7.2. Historias tafonómicas: 105
4.7.3 Presencia taxonómica y anatómica: 107
4.7.4. Modificaciones culturales: 112
CAPÍTULO 5 116
5. Discusión y conclusiones: 116
CAPÍTULO 6 134
6.1. Referencias citadas: 134
6.2. Anexos: 151
Anexo 1. Vertebrados endémicos de la región de Antofagasta: 151
Anexo 2. Fichas de los sitios arqueológicos: 159
Anexo 3. Variables y Códigos para tabular los datos: 167
Anexo 4. Lista general de partes esqueletarias: 170
Anexo 5. Fichas de taxones identificados en la muestra: 176
Anexo 6. Imágenes: 189

LISTA DE TABLAS:
Tabla 1: Divisiones del litoral Chileno entre Arica y el Canal de Chacao. 06
Tabla 2: Cronología y períodos culturales del Norte Grande de Chile. 21
Tabla 3: Sitios y unidades que componen la muestra. 32
Tabla 4: Distancia entre los sitios que componen la muestra. 34
Tabla 5: Distancia de los sitios en relación a la línea de costa. 34
Tabla 6: Fuentes de consumo, origen y producto. 43
Tabla 7: NISP y MNI, sitio CF3. 78
Tabla 8: Estadios de meteorización, sitio CF3. 79
Tabla 9: Estado de fractura, sitio CF3. 80
Tabla 10: Porcentajes de fragmentación, sitio CF3. 80
Tabla 11. Termoalteraciones, sitio CF3. 81
Tabla 12: Descripción e imágenes de artefacto óseo. Sitio CF3, Túmulo 1, Nivel 3. 82
Tabla 13: Descripción e imágenes de artefacto óseo. Sitio CF3, Alero 1, Nivel 1. 83
Tabla 14: NISP y MNI, sitio P3. 85
Tabla 15: MAU Lama sp, sitio P3. 86
Tabla 16: MAU Avifauna marina, sitio P3. 86
Tabla 17: Estado de fractura, sitio CF3. 88
Tabla 18: Porcentajes de fragmentación, sitio P3. 89
Tabla 19: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3. 91
Tabla 20: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3. 92
Tabla 21: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3. 92
Tabla 22: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3. 93
Tabla 23: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3. 93
Tabla 24: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3. 94
Tabla 25: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3. 94
Tabla 26: NISP y MNI, sitio PC8. 95
Tabla 27: Estado de fractura, sitio PC8. 96
Tabla 28: Porcentajes de fragmentación, sitio PC8. 97
Tabla 29: Descripción e imágenes de pieza ósea con huellas de corte. Sitio PC8. 99
Tabla 30: NISP y MNI, sitio Chungungo 6. 100
Tabla 31: Porcentajes de fragmentación, sitio Chungungo 6. 101

LISTA DE FIGURAS:
Figura 1: II Región de Antofagasta, Chile. 07
Figura 2: Unidades y subunidades naturales de la II Región de Antofagasta. 08
Figura 3: Clima de la II región de Antofagasta. 10
Figura 4: Ubicación de Punta Tames, II región de Antofagasta. 11
Figura 5: Punta Tames, II región de Antofagasta. 11
Figura 6: Esquema físico de las zonas del medio marino. 12
Figura 7: Esquema físico de las zonas del medio marino. 13
Figura 8: Esquema físico de las zonas del medio marino. 14
Figura 9: Ubicación de los sitios CF3, P3, PC8 y Chungungo 6. 32
Figura 10: Detalle de la ubicación de los sitios CF3, P3 y PC8. 33
Figura 11: Detalle de la ubicación del sitio Chungungo 6. 33
Figura 12: Emplazamiento del sitio CF3. 36
Figura 13: Emplazamiento del sitio P3. 38
Figura 14: Emplazamiento del sitio PC8. 39
Figura 15: Emplazamiento del sitio Chungungo 6. 40
Figura 16: Caleta el Fierro. 42
Figura 17: Isla guanera frente a Caleta el Fierro. 42
Figura 18: Piezas óseas identificadas y no identificadas. 75
Figura 19: Piezas óseas identificadas a nivel de orden, familia, género o especie. 76
Figura 20: Piezas óseas identificadas a nivel de clase. 76
Figura 21: Frecuencia de taxones en cada nivel de Túmulo1 y Unidad 4, Sitio CF3. 77
Figura 22: Frecuencia de taxones en nivel 1 de Alero 1, Sitio CF3. 78
Figura 23. Marcas naturales, sitio CF3. 81
Figura 24: Frecuencia de taxones en cada nivel del sitio P3. 84
Figura 25: MAU ictiofauna de la clase Actinopterygii, sitio P3. 87
Figura 26: Segmentos del esqueleto según clase, sitio P3. 87
Figura 27: Estadios de meteorización, sitio P3. 88
Figura 28: Otras marcas, sitio P3. 90
Figura 29: Termoalteración, sitio P3. 90
Figura 30: Frecuencia de taxones en cada nivel del sitio PC8. 95
Figura 31: Estadios de meteorización, sitio PC8. 96
Figura 32: Otras marcas, sitio PC8. 98
Figura 33: Frecuencia de taxones en cada nivel de Alero 1, sitio Chungungo 6. 99
Figura 34: Estadios de meteorización, sitio Chungungo 6. 100

Figura 35: Otras marcas, sitio Chungungo 6. 101
Figura 36: Perfil etario. 102
Figura 38: Distancia entre los sitios CF3, P3 y PC8 129

1
INTRODUCCIÓN:
La presente memoria para optar al título de arqueóloga lleva por nombre „Utilización de
vertebrados durante el Período Formativo en Punta Tames, Costa Arreica de
Antofagasta‟. Este estudio fue patrocinado y financiado por el proyecto Fondecyt 1100951
„Del Período Formativo al Tardío en la costa de Antofagasta. Cronología e interrelaciones
con las tierras altas‟ cuya arqueóloga responsable es Victoria Castro. En este marco, se
pretende contribuir a la comprensión respecto de la utilización de vertebrados, en cuanto
al consumo alimentario y al uso de huesos como materia prima por parte de los grupos
litoraleños que habitaron Punta Tames durante el Período Formativo.
Múltiples aspectos de la vida cotidiana de nuestros antepasados son posibles de
comprender mediante el estudio de cómo estos se desenvolvieron en su entorno; las
interrogantes que hoy surgen respecto a los eventos ocurridos durante el pasado, pueden
ser parcialmente resueltas a través de vestigios arqueológicos, siendo los restos
faunísticos un material con gran potencial informativo.
La zooarqueología, en tanto disciplina enfocada al estudio de restos faunísticos
procedentes de sitios arqueológicos, nos permite reconstruir de manera virtual procesos
culturales del pasado; el registro arqueofaunístico constituye una herramienta capaz de
aproximarnos al comportamiento humano, en cuanto a las estrategias de subsistencia y a
la adaptación al medio ambiente (Davis 1989, Reiz y Wing 1999, Chaix y Méniel 2006,
Gamble 2008).
Es importante considerar que los animales han protagonizado un rol fundamental en el
desarrollo de la vida humana. A lo largo de nuestra historia, hemos explotado sus
recursos en post de nuestro beneficio (Mengoni 1988), es por esta razón que el análisis y

2
la comprensión de restos arqueofaunísticos nos permite acceder a información inédita
sobre nuestro propio pasado.
El período cultural en el que se enfocará la presente investigación, corresponde al
Período Formativo, ubicado cronológicamente entre el 1000 a.C. y el 400 d.C. En
términos generales, durante este período fueron implementados cambios a nivel
sociopolítico, económico y tecnológico, entre los cuales se encuentra la transformación
del modo de subsistencia de apropiación a producción de recursos, mediante la
implementación de una economía agropastoralista (Muñoz 1989, Núñez L 1989).
El escenario de esta memoria se sitúa en la segunda región de Antofagasta,
específicamente en el litoral árido de arreismo absoluto. La aridez que reviste a este
paisaje desértico conlleva a la escasez de recursos hídricos superficiales, generando un
ambiente terrestre con insuficientes recursos bióticos, razón por la cual no fue posible
realizar actividades agrícolas y pastoriles (López 1982, Niemeyer y Cereceda 1984,
Carvallo 1990).
Dichos factores geoambientales no permitieron que los grupos litoraleños de la costa
arreica implementaran el modo de vida formativo, razón por la cual, los habitantes de esta
área continuaron con los patrones de subsistencia arcaicos, vale decir, una economía
basada en la caza, pesca y recolección.
A pesar de la escases de recursos terrestres, la costa desértica constituye una fuente de
abundante biomasa que permitió la explotación de variados recursos bióticos (Núñez y
Varela 1969, Llagostera 1982, 1989, Núñez P 2003, Núñez y Santoro 2011). A partir de
esto, los grupos litoraleños especializaron las estrategias de extracción de recursos
marinos, desarrollando una alta eficiencia adaptativa (Llagostera 1982).

3
Entre los paisajes de la costa arreica se encuentra la localidad de Punta Tames, la cual
presenta escasos antecedentes respecto del Período Formativo, siendo la principal fuente
de información cuatro sitios recientemente excavados: P3 y Chungungo 6 –conchal-, CF3
–cementerio- y PC8 –habitacional-. Producto de tales excavaciones fueron recuperadas
muestras de osteofauna, es decir restos óseos de vertebrados (Aldunate 2011, Castro V
2011, López 2011, Varas 2012).
Los estudios realizados hasta la fecha, no han abarcado la temática sobre el uso de
vertebrados por parte de los habitantes de Punta Tames durante el Período Formativo.
Esta información en tanto actividad cotidiana, es necesaria para la comprensión de los
eventos ocurridos y, por lo tanto, del desarrollo cultural.
Sobre la base de lo anterior, esta memoria tiene por objetivo general caracterizar la
utilización de los recursos faunísticos, específicamente de vertebrados -respecto al
consumo alimentario y al uso de huesos como materia prima- durante el Período
Formativo en Punta Tames.
Acorde a lo anterior, serán planteados una serie de objetivos específicos, siendo estos: la
contextualización del área de estudio en términos geográficos, zoológicos y
arqueológicos, analizar las muestras de osteofauna procedentes de los sitios CF3, P3,
PC8 y Chungungo 6; para posteriormente establecer el consumo alimentario y
procedencia de los taxones e identificar y caracterizar instrumentos óseos.
En consecuencia, se pondrán en dialogo los datos extraídos tanto del análisis como de la
bibliografía. Para de este modo generar una conclusión capaz de contribuir a la
comprensión respecto de la utilización de vertebrados en cuanto al consumo alimentario y

4
al uso de huesos como materia prima, por parte de los grupos litoraleños que habitaron
Punta Tames durante el Período Formativo.

5
CAPÍTULO 1
Antecedentes geográficos
1.1. Geografía del Norte Árido:
El Norte Árido de Chile se ubica en el extremo septentrional del país y limita con las
repúblicas del Perú, Argentina y Bolivia. Se caracteriza por poseer un 70% de superficie
de desierto absoluto y por la presencia de cuatro distintas franjas orientadas de forma
longitudinal. En primer lugar, de oriente a poniente, se ubica la línea litoral, posteriormente
se presenta la Cordillera de la Costa, la cual posee un ancho aproximado de 40 kilómetros
y una altura media de 1500 m.s.n.m. Continuando hacia el este, se ubica la pampa
(estepa desértica), la que posee un ancho promedio de 30 kilómetros y una altura
aproximada de 1000 m.s.n.m. Finalmente, se encuentran la precordillera, alcanzando una
altura entre 1600 y 3000 m.s.n.m. y la Cordillera de los Andes (altiplano andino), la que
supera los 4000 m.s.n.m. (Niemeyer 1989). Las costas que bañan el norte de Chile se
dividen en dos litorales áridos: al norte se encuentra el litoral árido de valles y quebradas,
que abarca desde Arica hasta Pisagua y al sur se ubica el litoral árido de arreismo
absoluto (costa arreica), el que contempla desde Pisagua a Chañaral (Llagostera 1989;
Ver tabla 1).

6
Tabla 1: Divisiones del litoral Chileno entre Arica y el Canal de Chacao. Tomado y
modificado de Llagostera (1989).
1.2. Geografía de la región de Antofagasta:
La segunda región de Antofagasta se ubica en el litoral árido de arreismo absoluto o costa
arreica y se extiende entre los 20°56‟ y 26 °05‟ S, los 67 °00 W y el Océano Pacifico,
abarcando una superficie de 125.911 km². La principal característica de esta región es el
paisaje desértico, lo que provoca una fuerte reducción de los recursos hídricos
superficiales (Carvallo 1990). Siendo el rio Loa, la única fuente hidrográfica permanente
del área, recorriendo 440kms desde su nacimiento en la falda norte del volcán Miño en
Ojos del Miño hasta su desembocadura en Caleta Huelén. Este río atraviesa el desierto
de Atacama fecundando oasis, valles y proveyendo de agua potable al área (Niemeyer y
Cereceda 1984). Producto de tales condiciones de aridez, Antofagasta posee escasos

7
suelos para actividades agrícolas y reducidos recursos bióticos (flora y fauna) (Carvallo
1990; Ver figura 1).
Figura 1: II Región de Antofagasta, Chile (Google Earth).
En cuanto a la geomorfología de la región, de oriente a poniente, se presentan cuatro
principales unidades de relieve: macizo andino, depresión intermedia, cordillera de la
Costa y planicie litoral, las que a su vez se dividen en subunidades (Carvallo 1990; Ver
figura 2).

8
Unidad Subunidad
1. Macizo andino 1.1. Altiplano
1.2. Cordillera de los Andes
1.3. Fosas prealtiplánicas
1.4. Precordillera
2. Depresión intermedia 2.1. Pampa del Tamarugal
2.2. Pampa Ondulada
2.3. Desierto de Atacama
3. Cordillera de la Costa 3.1. Sector norte
3.2. Sector sur
4. Planicie litoral 4.1. Franja litoral norte
4.2. Sector Península de Mejillones
4.3. Franja litoral sur
Figura 2: Unidades y subunidades naturales de la II Región de Antofagasta. Tomado y
modificado de Carvallo (1990).
El macizo andino sobrepasa los 4.000 m.s.n.m. se constituye por relieves volcánicos
sobrepuestos y se divide en cuatro subunidades: altiplano, cordillera de los Andes, fosas

9
prealtiplánicas y precordillera. La depresión intermedia consiste en un plano
depositacional continuamente interrumpido por cordones montañosos y cerros isla. Esta
gran depresión se presenta a lo largo de todo el perfil longitudinal de la región y se divide
en tres subunidades: pampa de Tamarugal, pampa Ondulada y desierto de Atacama. La
Cordillera de la Costa está compuesta por un conjunto de cordones de cerros, posee un
ancho promedio de 50 kilómetros y una altura que supera los 1.800 m.s.n.m, su extensión
es asimétrica ya que en algunas zonas -norte de Tocopilla y sur de Antofagasta- limita
abruptamente con el mar en forma de grandes acantilados. Finalmente, la planicie litoral
se caracteriza por la escasa presencia de playas de arena, debido a la interrupción de la
Cordillera de la Costa, razón por la que el litoral se divide en tres subunidades: franja
litoral norte, sector Península de Mejillones y franja litoral sur (Carvallo 1990).
El clima de la región presenta variaciones típicas de la aridez casi absoluta, ya que
cambia según su cercanía con la costa debido a la humedad y en la Cordillera de los
Andes debido a la altura. A medida que ésta aumenta, se producen lluvias las cuales
fluctúan entre 25 y 50 mm anuales, permitiendo la presencia de oasis al borde del desierto
(Carvallo 1990; Ver figura 3).

10
A Clima desértico con nublados abundantes
B Clima desértico normal
C Clima de desierto marginal de altura
Figura 3: Clima en la II región de Antofagasta. Tomado y modificado de Carvallo (1990).
1.3. Geografía de Punta Tames:
La localidad de Punta Tames se ubica en la II región de Antofagasta y está limitada por el
Océano Pacífico y la Cordillera de la Costa. Presenta bajos niveles de precipitaciones y
una rica biomasa marina (Núñez y Varela 1967), el clima es templado y la presencia de
mar posibilita las condiciones ecológicas adecuadas para la subsistencia de organismos
vivos (López C 1982; Ver figura 4 y 5).

11
Figura 4: Ubicación de Punta Tames, II región de Antofagasta (Google Earth).
Figura 5: Punta Tames, II región de Antofagasta (Google Earth).

12
1.4. Ecosistema marino:
El ecosistema marino se divide en cinco zonas: litoral, sublitoral (superior e inferior),
batial, abisal y hadal. (Moreno y Castillo 1976; Ver figura 6).
Figura 6: Esquema físico de las zonas del medio marino. Tomado y modificado de
Nautilus-ecosistemas.blogspot.com.
El sublitoral superior corresponde al área entre los 20 y 40 metros de profundidad. El
sublitoral inferior se sitúa entre los 40 y 200 metros de profundidad, siendo la zona que
posee la mayor extensión de plataforma continental -en Chile Central logra un ancho de
entre 15 y 30 kilómetros, se amplía hacia el sur-. La zona batial se ubica entre los 200 y
3000 a 4000 metros de profundidad, lo que provoca que la penetración de luz sea menor,
lo que dificulta el proceso de fotosíntesis de algas, reduciendo la variedad de especies. En
la zona abisal se presentan peces propios de una gran profundidad -entre 4000 y 6000
metros de profundidad-. Finalmente, la zona hadal corresponde a las aguas más

13
profundas del océano, en donde se presentan grandes fosas oceánicas (Moreno y Castillo
1976).
Por otra parte, se denomina sistema pelágico a la zona marina próxima a la superficie
(sublitoral). Los peces que habitan esta área son denominados peces pelágicos o peces
de la columna de agua y tienen como característica principal que su ciclo biológico no
tiene relación con los fondos oceánicos. El sistema pelágico se divide en dos regiones:
nerítica -cerca de la costa- y oceánica -mar adentro- (Moreno y Castillo 1976; Ver figura
7).
Figura 7: Esquema físico del de las zonas del medio marino. Tomado y modificado de
Nautilus-ecosistemas.blogspot.com.
La región nerítica corresponde a la columna de agua ubicada por sobre la plataforma
continental, en donde predominan las aguas de la corriente de Humboldt. Esta corriente
se caracteriza por no poseer una gran variedad de especies. Sin embargo, presenta un
alto número de ejemplares, los que tienen una vida relativamente corta, un crecimiento

14
rápido y una elevada fecundidad. Conforme a lo anterior esta zona se encuentra entre las
más productivas del océano (Moreno y Castillo 1976, Llagostera 1989).
La región oceánica se divide en tres zonas dispuestas estratigráficamente: las zonas
epipelágica, mesopelágica, batipelágica y abisopelágica, corresponden a la columna de
agua ubicada por sobre el talud continental (Ver figura 8). La zona más fértil corresponde
a la epipelágica, la que se extiende desde la superficie hasta el punto de extinción de la
luz solar -alrededor de los 200 metros de profundidad-. Dicho de otro modo, se extiende a
la misma altura que la región nerítica pero mar adentro. En la región oceánica la densidad
de materia viva por unidad de volumen es inferior que en la región nerítica (Moreno y
Castillo 1976).
Figura 8: Esquema físico de las zonas del medio marino. Tomado y modificado de
Nautilus-ecosistemas.blogspot.com.

15
1.4.1. Fenómeno del Niño:
El fenómeno del Niño constituye una alteración en el sistema de la corriente de Humboldt,
provocada por aguas más calientes que se desplazan desde Ecuador hacia el sur.
Consiste en un calentamiento ocasional, irregular y aperiódico de las aguas (Zavala
2008:17). Las temperaturas sufren una elevación de alrededor de 2°C, lo que provoca
cambios en los patrones climáticos, alejando a ciertas especies marinas (Zavala 2008).
Antecedentes Zoogeográfícos:
1.5. Vertebrados:
La gran diversidad de individuos que componen el reino animal es clasificada
principalmente en las categorías filo, clase, orden, familia, género y especie, las que
consisten en agrupaciones de organismos con características en común, siendo la
categoría de especie la más específica (Simpson 1961, Grayson 1984, Reitz y Wing
1999,).
En esta investigación nos centraremos únicamente en restos osteofaunísticos, es decir,
en huesos de animales vertebrados procedentes de sitios arqueológicos.
Los vertebrados pertenecen al filo Cordados y se caracterizan por poseer estructura ósea
y alta adaptabilidad al medio ambiente (Padilla y Cuesta 2003). Entre las clases de
Cordados se encuentran reptiles, anfibios, peces, aves y mamíferos, los que se
distribuyen en distintos órdenes, familias, géneros y especies (Padilla y Cuesta 2003).
La fauna endémica chilena se encuentra regionalmente diferenciada, según las
condiciones ecológicas de los ecosistemas (López C 1982). La II Región de Antofagasta
posee tres eco-regiones: el litoral, la estepa desértica y el macizo andino; en cada una de

16
ellas habitan diversas especies de vertebrados, ya que cada nicho ecológico constituye
una frontera vital, donde las especies ejercen tuición (Donoso-Barrios 1966:26, Ver anexo
1).
1.6. Vertebrados endémicos de la región de Antofagasta:
1.6.1 Herpetofauna:
La herpetofauna se constituye por reptiles y anfibios. Los reptiles están provistos de una
piel seca y cubierta de escamas, poseen la capacidad de regular su temperatura corporal
según la del ambiente y están adaptados a la vida terrestre y/o acuática (Núñez H 2008).
Por su parte los anfibios se caracterizan por desarrollar una vida acuática hasta llegar a
edad adulta en la cual pasan la mayor parte del tiempo fuera del agua (Donoso-Barrios
1966). Los anfibios se clasifican en tres órdenes Anura (ranas y sapos), Gymnophiona
(cecilias) y Urodela (salamandras), en Chile solo habitan anuros, siendo el grupo menos
numeroso de vertebrados presentes en el país (Méndez y Correa 2008).
Herpetofauna del Litoral:
La hepertofauna del litoral se encuentra representada por tres especies de los géneros
Tropidurus, Liolaemus y Phyllodactylus, que consisten en pequeñas lagartijas.
Adicionalmente dos especies del orden Testudines, correspondientes a tortugas marinas
(Donoso-Barrios 1966, López C 1982).
Herpetofauna de la estepa desértica:
Entre la fauna de la estepa desértica habitan tres especies de los géneros Tropidurus y
Liolaemus, todos correspondientes a lagartos del orden Squamata (Donoso-Barrios 1966,
López 1982) y un anuro del género Bufo (Méndez y Correa 2008).

17
Herpetofauna del macizo andino:
En el macizo andino se encuentran nueve especies de lagartos del género Liolaemus
(Donoso-Barrios 1966, López C 1982) y cuatro especies de anuros de los géneros
Telmatobius, Pleurodema y Bufo (Méndez y Correa 2008).
Cabe mencionar que las culebras chilenas se reúnen en dos géneros, los que se
distribuyen por casi todo el país, pero los antecedentes sobre su biología son sumamente
escasos (Núñez H 2008).
1.6.2. Ictiofauna:
Los peces o ictiofauna viven en el agua, respiran a través de branquias y poseen
extremidades con forma de aleta, que facilitan su desplazamiento por el medio acuático
(Moreno y Castillo 1976).
Ictiofauna del litoral:
En el litoral se presentan peces osteíctios y condrictios. Los primeros se caracterizan por
poseer un esqueleto óseo y están representados por la clase Actinopterygii, es decir
peces con aletas radiadas. Por otra parte, los peces condrictios se caracterizan por
poseer un esqueleto cartilaginoso y se encuentran representados por Elasmobranquios,
tales como rayas y tiburones (Moreno y Castillo 1976).
La ictiofauna del área se encuentra distribuida por las distintas zonas del ecosistema
marino, siendo el sublitoral superior el área de mayor diversidad, con dieciséis especies,
seguido la región nerítica con nueve especies, el sublitoral inferior con ocho especies y
finalmente, la región oceánica con seis especies (Moreno y Castillo 1976).

18
1.6.3. Avifauna:
Las aves o avifauna son plumíferos, bípedos, de mandíbula con forma de pico córneo, se
reproducen por huevos, la mayoría posee una adaptación completa al vuelo y la mitad de
las especies presenta algún tipo de migración (Araya y Millie 1992, Vilina y Cofré 2008a).
Existen tres tipos de aves: marinas, acuáticas y terrestres. Se denomina aves marinas a
las especies que viven en el mar, que se alimentan en forma directa del medio marino o
bien que utilizan este hábitat en algún ciclo de su vida, como por ejemplo, las épocas
reproductivas (Vilina y Pizarro 2008). Las aves acuáticas habitan ambientes húmedos
tales como humedales (Vilina y Cofré 2008b). Finalmente, las aves terrestres realizan su
ciclo de vida sin la necesidad de acceder a ambientes acuáticos y/o marinos (Vilina y
Cofré 2008a).
Avifauna del litoral:
La avifauna del litoral se compone por cincuenta y nueve especies correspondientes a
nueve órdenes. Siendo Charadriiformes, Sphenisciformes, Procellariiformes,
Pelecaniformes y Suliformes las aves marinas más abundantes, seguido de aves
terrestres como lo son gorriones Passeriformes, aves nocturnas Strigiformes, aves
rapaces Falconiformes y una especie de picaflor Apodiforme (Jaramillo 2005, Vilina y
Cofré 2008b).

19
Avifauna de la estepa desértica:
En la estepa desértica habitan siete especies Passeriformes, dos Falconiformes y una
Strigiforme, todas correspondientes a aves terrestres. (Jaramillo 2005, Vilina y Cofré
2008a).
Avifauna del macizo andino:
En el macizo andino se encuentran aves terrestres y acuáticas que suman un total de
cincuentaisiete especies. Las especies terrestres se distribuyen en los órdenes
Passeriforme, Falconiforme y Apodiforme. Mientras que las especies acuáticas se
encuentran representadas por los órdenes Phoenicopteriforme, Charadriiforme,
Anseriforme, Gruiforme, Podicipediformes y Suliforme (Jaramillo 2005)
1.6.4. Mastofauna:
Los mamíferos o mastofauna se caracterizan por la presencia de un olfato altamente
desarrollado, una mayor complejidad del cerebro, sangre caliente y hembras provistas de
glándulas mamarias, las que producen leche para alimentar a las crías (Iriarte 2008).
Los mamíferos se agrupan en dos subclases Prototheria y Theria. Los Prototherios son
ovíparos, es decir, se reproducen por huevos y no poseen glándulas mamarias
completamente desarrolladas. Por otro lado, los Therios son la única subclase de
mamíferos presentes en Chile. Son vivíparos, es decir, se reproducen por crías vivas y se
dividen en dos infraclases: Metatheria y Euteria. Los Metatherios o marsupiales poseen
poca o nula placenta, mientras que los Euterios o placentarios poseen reproducción
vivípara placentaria y glándulas mamarias plenamente desarrolladas, constituyendo el
infraorden más numeroso de mamíferos vivientes (Iriarte 2008).

20
Mastofauna del litoral:
En el litoral habitan una gran variedad de Euterios. Los terrestres se encuentran
representados por trece especies procedentes de las familias Canidae (cánidos) y Felidae
(félidos) y de los órdenes Chiroptera (murciélagos), Rodentia (roedores) y Artiodactyla
(ungulados) el cual contempla a Hippoamelus antisensis (taruca) y Lama guanicoe
(guanaco) actualmente ausentes en el área (Núñez P 2003, Iriarte 2008). Entre los
mamíferos marinos se encuentran un Mustelidae (chungungo), dos Otariidae (lobo
marino) y veinticinco especies de cetácea (cetáceos) representados por las familias
Balaenidae (ballena), Delphinidae (delfín), Ziphiidae (zifo), Physeteridae (cachalote) y
Phocoenidae (marsopa) (Iriarte 2008).
Mastofauna de la estepa desértica:
En la estepa desértica solo se encuentra una especie de mamífero terrestre
correspondiente al orden Rodentia (Iriarte 2008).
Mastofauna del macizo andino:
Los mamíferos del macizo andino están representados por veinte especies,
correspondientes a los órdenes Didelphimorpia (marsupiales), Chiropteda (murciélagos),
rodentia (roedores), Artiodactyla (ungulados), mustelidae (hurón) y Carnívora -de las
familias Felidae (félidos) y Canidae (cánidos)- (Iriarte 2008).

21
Antecedentes Arqueológicos:
1.7. Antecedentes previos al Período Formativo:
Con la finalidad de comprender los procesos ocurridos durante el pasado, los datos
arqueológicos son distribuidos en su dimensión temporal constituyéndose períodos
culturales. La prehistoria del Norte Grande de Chile se divide en los Períodos Arcaico,
Formativo, Medio, Intermedio Tardío y Tardío.
Tabla 2: Cronología y períodos culturales del Norte Grande de Chile. Tomado y
modificado de Berenguer (1997).
Previo a abordar el estudio del Período Formativo es necesario comprender el Período
cronológico anterior, ya que en el Formativo fueron consolidados procesos desarrollados
durante el Arcaico (Núñez L 1989, Núñez y Santoro 2011).
La mayor parte de los antecedentes arqueológicos del litoral árido de arreismo absoluto
han sido asignados al Período Arcaico, los eventos ocurridos durante dicho período han
sido sintetizados por Llagostera (2005), quien los caracteriza en cinco fases:

22
En la primera fase (10.700 a 9.500 a.p.), la economía estaba basada en la caza, pesca y
recolección marina, siendo peces y mariscos las principales fuentes de alimentación. En
menor medida eran consumidos camélidos, pinnípedos, cánidos, roedores y aves. Estos
grupos manejaban una tecnología específica y eficiente con una alta adaptación a la vida
en el litoral. La segunda fase (9.500 a 9.000 a.p.) se caracterizó por la adopción de
nuevas manifestaciones culturales pertenecientes al complejo Huentelauquén, las pautas
de alimentación continuaron siendo las mismas de la fase anterior. Sobre los
acontecimientos ocurridos durante la tercera fase (9.000 a 6.000 a.p.) no se poseen
mayores antecedentes (Llagostera 2005). Las manifestaciones de la cuarta fase (6.000 a
5.000 a.p.) son consideradas como una extensión hacia el sur de la Fase II del litoral árido
de valles y quebradas, esta fase se divide en dos sub-fases: una Temprana caracterizada
por el anzuelo de concha circular y una Tardía, en donde el anzuelo de concha es
reemplazado por el de espinas de cactáceas (Llagostera 1989). En la quinta fase (5.000 a
4.000 a.p.), se dio inicio a la arquitectura (Schaedel 1957). En la sexta fase (4.000 a 3.000
a.p.) fueron incorporados pisos selladores de argamasa de ceniza de algas con las que se
cubría a los difuntos, los cuales eran inhumados al interior de las construcciones,
agregando una funcionalidad funeraria a los sitios habitacionales (Bittmann 1984).
Durante esta fase también se estableció contacto con algunos grupos que habitaban al
interior de la región (Núñez et al. 2008).

23
1.8. Período Formativo:
1.8.1. Período Formativo en el Norte Árido:
El Período Formativo se ubica cronológicamente entre el 1000 a.C. y el 400 d.C. y se
caracterizó por una serie de cambios a nivel sociopolítico, económico y tecnológico, lo que
conllevo una serie de logros que ampliaron las expectativas de vida (Núñez L 1989).
La concepción que inicialmente se tenía sobre este período estaba fundada sobre el
historicismo cultural, en el que la secuencia cultural fue basada en evidencias materiales
tales como tipologías cerámicas y contextos fúnebres, entre otros. Actualmente la
transición hacia el modo de vida Formativo es comprendida como un hecho sumamente
variable, estas distinciones están relacionadas a factores locales, regionales y ecológicos
(Edward 2006).
Los componentes que caracterizan al Período Formativo se desarrollaron bajo la
influencia de poblaciones de tierras altas. Estos se dieron en forma paulatina y
consistieron en la transformación del modo de subsistencia de apropiación a producción
de recursos mediante la implementación de una economía agropastoralista -la caza y
recolección continuaron como auxiliar-, la generación de excedentes de producción, el
consumo de sustancias alucinógenas y el desarrollo de nuevas tecnologías, tales como:
alfarería, textilería y prácticas minero-metalúrgicas. En consecuencia, surgieron
especialistas de tiempo completo y el intercambio a larga distancia. (Núñez L 1989).
Dichas innovaciones, transformaron el patrón de residencia, intensificando el uso del
territorio, promoviendo la vida sedentaria y la construcción de recintos de piedra y barro,
lo que llevó posteriormente al aumento demográfico, produciendo la emergencia de clases

24
sociales diferenciadas (Muñoz I 1989). La nueva ideología del Período Formativo se
iniciaba a partir de la premisa de repartir los bienes a modo tal de generar autosuficiencia
en cada comunidad (Núñez L 1989, Pimentel et al. 2011).
Mediante la interacción entre grupos lejanos, algunas influencias de tierras altas fueron
enclavadas en la costa, aportando diversos componentes culturales que fueron adoptados
y empleados por los grupos litoraleños tanto del litoral árido de valles y quebradas como
del litoral árido de arreismo absoluto (Núñez y Santoro 2011).
En el litoral árido de valles y quebradas las relaciones costa-oasis datan del Período
Arcaico, esto sumado a la presencia de recursos fluviales permitió, para el Período
Formativo el desarrollo de una economía agro-marítima. Las sociedades pescadoras-
recolectoras de la subárea Valles Occidentales experimentaron cambios hortícola-
agrícolas que permitieron la conformación de patrón de asentamiento aldeano con
estructuras construidas de materiales livianos (Núñez y Santoro 2011).
Dichas aldeas eran emplazadas en torno a vertientes de los tramos inferiores de los
valles. El consumo de productos agrícolas era complementado con recursos proteicos
marítimos, al margen de aportes cárneos continentales ante la carencia de caza y crianza
de camélidos en estas tierras bajas (Núñez y Santoro 2011:510) y al igual que en las
tierras altas fueron incorporados nuevas tecnologías tales como alfarería, metalurgia de
oro y cobre, y uso de materias primas exóticas (Núñez y Santoro 2011).
Debido a factores geográfico-ambientales en algunas áreas el modo de vida Formativo
no fue implementado. Un ejemplo de ello, es el caso de los grupos que habitaron el litoral
árido de arreismo absoluto de la región de Antofagasta. Puesto que las condiciones
ambientales de dicha área no facilitaron el desarrollo de labores agrícolas, ni pastoriles

25
(Carvallo 1990), por lo que se mantuvieron los patrones Arcaicos (Núñez P 2003). Es
decir, ya que no fue posible la implementación de una economía agraria, se conservó una
encomia marítima.
Durante el Período Formativo, en la costa arreica se conservó el modo de subsistencia
desarrollado durante el Período Arcaico, basado en la caza, pesca y recolección marina.
De este modo, las estrategias de apropiación de recursos se especializaron al nivel de
lograr objetivos socioculturales y alimenticios tan óptimos como los recurrentes en los
asentamientos complejos circumpuneños (Núñez y Santoro 2011:502).
Estos grupos se organizaban bajo un régimen relativamente igualitario; desarrollaron una
alta movilidad en torno a espacios productivos, con un patrón residencial disperso y de
discreta densidad. En cuanto a la funebrita, los difuntos eran inhumados en compañía de
sus implementos costeros de uso cotidiano (Núñez y Santoro 2011), en ocasiones con
elementos obtenidos mediante el intercambio con grupos del interior –tales como
cerámica y artefactos para el consumo de psicotrópicos- (Núñez L 1979) o del litoral árido
de valles y quebradas –como el patrón de entierro de la Fase Alto Ramírez de Arica-
(Moragas 1982).
1.8.2. Período Formativo en la costa arreica:
En cuanto a las ocupaciones en el litoral árido de arreismo absoluto durante el Período
Formativo, se presenta una concentración de sitios en la desembocadura del rio Loa. Los
sitios Caleta Huelén 7 (2030±80 ap.), Caleta Huelén 10 (735±100 ap. y 2000±70 ap.),
Caleta Huelén 10A. (2320±80 ap.) y Caleta Huelén 43 (2400±90 ap.) corresponden a
cementerios de túmulos, en los cuales los cuerpos eran cubiertos con esteras de fibra
vegetal y plumas. Los ofertorios se componían de conchas de molusco, instrumentos

26
líticos, artefactos de pesca y recolección marítima, tales como: anzuelos de espina de
quisco y bolsas de red entre otros; también elementos propios de las tierras altas, como lo
son objetos de cobre, vasijas de cerámica, cestería, cordelería, implementos para la
inhalación de alucinógenos, quínoa y lana de camélidos (Núñez L 1979).
A lo largo de toda la costa norte del país, durante el Período Formativo los contextos de
explotación marítima son mayoritarios (Núñez L 1989). Probablemente la alimentación de
estas poblaciones se constituyó principalmente de recursos marítimos, ya que basta mirar
el mapa de Chile para darse cuenta de la importancia que asume el litoral en relación a
esta angosta franja de tierra (Llagostera 1989:57). La costa de Chile está bañada por la
corriente de Humboldt, compuesta por corrientes subantárticas frías, caracterizadas por
ser sumamente nutritivas, por lo que la productividad del litoral del Norte de Chile resulta
altamente favorecida (Llagostera 1989).
Entre los vertebrados que habitan las costas de la región se encuentran los peces, los
cuales constituyen un caso excepcional ya si bien poseen una vida corta, crecen en cortos
periodos de tiempo y poseen una elevada fecundidad (Llagostera 1989), convirtiéndolos
en un eficiente recurso. La hipótesis de predominio en el consumo de peces es avalada
por los anzuelos de concha y de espina de cactus que han sido hallados en sitios 1
asignados a los Períodos Arcaico Tardío y Formativo Temprano de la costa de
Antofagasta (Llagostera 1982, 1989, 2005). Llagostera (1990) plantea que la presencia
del congrio en los contextos arqueológicos del sitio Punta Blanca, al sur de Tocopilla,
1 Cerro Colorado, Punta Morada, Cobija 13, Los Canastos, Montenegro y Huelén 42.

27
sería un bio-indicador de navegación. Este impulso en la economía marítima se habría
desarrollado hacia el 230 d.C.
1.8.3. Intercambio a larga distancia en la región de Antofagasta:
Con la finalidad de abastecerse de recursos variados y/o foráneos, los habitantes de la
región generaron redes de contacto mediante viajes temporales a territorios lejanos
(Núñez L 1989). Estas redes de intercambio probablemente ya se encontraban en
funcionamiento durante el Período Arcaico Tardío, pero fueron consolidadas en el
Formativo (Berenguer 2004, Núñez P 2003).
Para explicar tal movilidad, Núñez y Dillehay (1979) plantearon el modelo de „movilidad
giratoria‟. El cual consiste en un tráfico a larga distancia en el que mediante el apoyo de
llamas cargueras, eran intercambiados bienes de zonas ecológicas tan lejanas como los
son la costa, la puna y la selva. Para realizar estos intercambios fueron trazados un
conjunto de rutas fijas, las cuales eran determinadas según necesidades logísticas, la
equivalencia entre los recursos intercambiados y la jerarquía del asentamiento con el cual
intercambiar. Cabe mencionar que este modelo otorga protagonismo a los grupos de
tierras altas postulándolos como gestores de esta articulación.
Pimentel y colaboradores (2011), han generado un modelo en el que los grupos costeros
adquieren mayor relevancia. Esta propuesta se realizó sobre la base de senderos y
evidencias asociadas, mediante las cuales los autores analizaron las posibles estrategias
de movilidad y postulan que en el desierto de Atacama, durante el Período Formativo, se
establecieron relaciones en forma relativamente fluida y estable a través de dos
modalidades: caravanera y costera.

28
La modalidad caravanera es definida como circuitos por los que grupos de las tierras altas
junto a recuas de llamas cargueras bajaban a la costa. A lo largo de estos senderos
fueron construidos campamentos de descanso compuestos por precarias estructuras,
muros a ras de piso e hilada simple; el material cultural asociado a estas estructuras son
principalmente líticos y cerámicas de los tipos Quillagua café amarillo, Grupo los Morros,
Sequitor negro pulido y Loa café alisado. Esta modalidad habría llegado al litoral tras
recorrer 160 kilómetros desde Calama o 250 kilómetros desde San Pedro de Atacama
(Pimentel et al. 2011).
Por otro lado, la modalidad costera es definida como recorridos peatonales, sin el apoyo
de animales de carga. Estos senderos eran transitados por grupos litoraleños que
visitaban a distintos oasis de Atacama, hasta 70 kilómetros al interior. Los campamentos
de descanso no presentan mayor inversión en el ámbito de la arquitectura, se presentan
escasos geoglifos, material lítico y cerámica Quillagua café amarillo, Grupo los Morros,
Sequitor negro pulido y Loa café alisado (Pimentel et al. 2011).
La presencia de bienes foráneos -marinos y andinos- en diversos sitios de la región ha
permitido confirmar el contacto e intercambio entre habitantes de tierras lejanas, como
también los productos en circulación, siendo principalmente materias primas, alimentos y
bienes de prestigio (Mostny 1952, Núñez y Santoro 2011 y Pimentel et al. 2011).
Desde la costa al interior eran llevados: peces, moluscos, sal, óxido de hierro y guano,
entre otros productos (Mostny 1952, Núñez y Santoro 2011 y Pimentel et al. 2011). La
abundancia de estos recursos atrajo el interés de las sociedades que vivían en los oasis y
quebradas altas del interior, provocando movimientos poblacionales a lo largo de la
transecta altitudinal (Aldunate et al. 2010:342).

29
Por otra parte, desde el interior hacia la costa eran transportados: poroto, ají, zapallo,
calabaza, algarrobo, maíz, cebil, quínoa, materias primas líticas, lana de camélido, objetos
de cobre, láminas de oro, artefactos para el consumo de psicotrópicos, sustancias
alucinógenas y contenedores de cerámica de diversos tipos2 (Llagostera 1989, Uribe et al.
2007, Núñez y Santoro 2011, Pimentel et al. 2011).
Cabe mencionar que la región de Antofagasta presenta una gran distancia entre la costa y
las tierras altas, estimulando una mayor independencia entre los grupos, lo que pudo
conducir al desarrollo independiente de estos; hecho que posteriormente debió ser
favorable al momento de ejercer relaciones complementarias a nivel vertical (Llagostera
1989, Núñez L 1989).
1.8.4. Período Formativo en el tramo Punta Atala - Punta Tames:
El tramo entre Punta Atala y Punta Tames contempla 35 kilómetros; lamentablemente
para esta zona y período, la información arqueológica es sumamente escasa (Castro V
2010).
La primera evidencia del Formativo en la zona corresponde al sitio Cobija 10, el que con
fechas entre el 320 a.C y el 350 d.C se sitúa a un kilómetro del pueblo de Cobija, a unos
300 metros de la línea litoral y consiste en un cementerio de túmulos con alrededor de 90
tumbas; es propuesto por Moragas (1982) como la expresión más meridional de la fase
Alto Ramírez de Arica.
Este extenso cementerio presenta un patrón de entierro con variaciones mínimas. Cada
túmulo fue compuesto por maderos de quisco, los que fueron dispuestos en forma
2 QCA: Quillagua Café Amarillo, QRP: Quillagua Rojo Pulido, QTC: Quillagua Tarapacá Café Amarillento, TNP: Topater Negro Pulido, LRA: Loa Rojo Alisado, LCA: Loa Café Alisado, LMS: Grupo los Morros, SGP: Grupo San Pedro Grueso Pulido, SRP: San Pedro Rojo Pulido, SEQ: Negro Pulido Sequitor.

30
horizontal y cubiertos por esteras sostenidas con rocas sobre las que el difunto era
depositado en posición decúbito lateral con las extremidades flectadas. Luego, el cuerpo
era cubierto con argamasa la que posteriormente era tapada con sedimento y conchuela
(Moragas 1982).
Los ofertorios eran sumamente escasos y se componían de artefactos líticos -cuchillo,
punta de proyectil, dardo, entre otros-, conquiológicos –cuchillo- y óseos -punzón,
cabecera de arpón, entre otros-, pieles de aves con sus plumas -fueron identificados
Phalacrocorax sp. (cormorán). y Sula variegata (piquero)-, vellones de lana de camélido
teñidos rojos, cordelería -fibra vegetal, lana de camélido y pelo humano-, bolsas de fibra
vegetal, cestería y anzuelos de raíz de cactácea (Moragas 1982).
Según los análisis químicos, la argamasa empleada para cubrir los cuerpos era elaborada
con sal extraída el mar, materiales obtenidos de la arena, principalmente sílice y calcio
proveniente de las conchas de moluscos, sedimentos terrestres entre otros elementos,
capaces de lograr una alta compactación. Este sellador era utilizado únicamente para la
elaboración de túmulos y era depositado en estado fresco (Moragas 1982).
Los túmulos eran rellenados con una serie de materiales tales como cerámica -la cual
solo era depositada como relleno y no como ofertorio- lascas, conchas, cueros y pelos de
Lama guanicoe (guanaco), Lontra felina (chungungo) y Otaria flevescens (lobo marino de
un pelo), mechones de pelo humano, plumas, coprolitos de camélido, hojas y fibras
vegetales. Cabe mencionar la presencia de túmulos simbólicos, en lo que no se
inhumaron restos humanos, sin embargo se realizaron eventos de quema (Moragas
1982).

31
Durante la última década, han sido registrados nuevos sitios con fechas formativas en el
área, en el marco del proyecto Fondecyt 1050991 „El desierto costero y sus vinculaciones
con las tierras altas de Cobija a Calama‟, a cargo de Victoria Castro desarrollado entre los
años 2005 y 2009 fueron excavados los sitios: Mantos del Pacífico 5, Mantos del Pacífico
29, Copaca 5, Copaca 7, Punta Grande 2, Bandurrias 3, Chungungo 3, Chungungo 4,
Tames 2, Guasilla 9 y Guasilla 19, Caleta el Fierro 2, 3 y 4 (Castro V et al. 2010).
1.8.5. Período Formativo en Punta Tames:
Los únicos antecedentes con los que contamos sobre ocupaciones humanas durante el
formativo en la localidad de Punta Tames provienen de los sitios Caleta el Fierro 3, P3,
PC8 y Chungungo 6, lo que fueron excavados en el marco del proyecto Fondecyt
1100951 „Del Período Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟ a cargo de Victoria Castro desarrollado entre 2010 y
2014 (Castro V 2010).
Los restos de osteofauna a analizar para cumplir los objetivos propuestos en esta
memoria proceden de excavaciones estratigráficas realizadas en dichos sitios.
1.8.5.1. Sitios que componen la muestra:
Los sitios Caleta el Fierro 3 (en adelante CF3), P3, PC8 y Chungungo 6, fueron
seleccionados por la presencia de restos osteofaunísticos y por estar asignados al
Período Formativo, según fechado radio carbónico o, según su contexto en el caso del
sitio P3 y la unidad 4 de CF3 (Ver anexo 2).

32
Sitio Unidad Fechas (ap.)
CF3 Alero 1 1580 ± 20
Túmulo 1 1810 ± 25
1880 ± 25
2170 ± 25
Unidad 4
P3 Unidad 1
PC8 Unidad 1 1840 ± 25
Chungungo 6 Alero 1 2570 ± 25
Tabla 3: Sitios y unidades que componen la muestra, con sus respectivos fechados radio
carbónicos (Castro V 2012).
Figura 9: Ubicación de sitios CF3, P3, PC8 y Chungungo 6 (Moreno 2013).

33
Figura 10: Detalle ubicación sitios CF3, P3 y PC8 (Moreno 2013).
Figura 11: Detalle ubicación sitio Chungungo 6 (Moreno 2013).

34
Chungungo 6 P3 CF3
P3 2008 m
CF3 2011 m 35 m
PC8 2152 m 158 m 179 m
Tabla 4: Distancia entre los sitios que componen la muestra (Moreno 2013).
Chungungo 6 P3 CF3 PC8
126 m 172 m 203 m 226 m
Tabla 5: Distancia de los sitios en relación a la línea de costa (Moreno 2013).
a. CF3:
Consiste a un cementerio de túmulos de aproximadamente 5000 m2 fuertemente
disturbado producto de excavaciones ilegales (Castro V 2011), en el que fueron
exhumados los restos óseos humanos correspondientes a seis adultos jóvenes, dos
juveniles y dos infantes (Constanzo 2012)
Según las enfermedades degenerativas articulares e inserciones musculares presentes
en el registro bioantropológico procedente de este sitio, Constanzo (2012) identificó
patologías producidas por la extensión y flexión de brazos y piernas asociadas al
lanzamiento de elementos arrojadizos, tales como: lanzas, estólicas y arpones, al
lanzamiento de redes de pesca, a la acción de remar, a la constante posición de estar
arrodillado en actividades de molienda o marisqueo y el transporte o arrastre de cargas
pesadas. Posiblemente estas actividades eran realizadas en forma constante y
prolongada, lo cual generó huellas de estrés mecánico asociadas a un sobre esfuerzo
muscular (Constanzo 2012).

35
Alero 1:
Consiste en un pequeño alero rocoso escasamente saqueado en el que fueron
exhumados restos óseos pertenecientes a una mujer de aproximadamente 30 años de
edad (Andrade Com. Pers. 2011), acompañada de su ajuar y ofertorio compuesto de:
tejidos, cordelería, madera, trozos de cuero de Lobo marino (Otariidae), un artefacto óseo
de forma tubular, numerosos clastos redondeados, algunos con pigmento rojo, variados
líticos, trozos de mineral de cobre, cerámica indeterminada, osteofauna y malacofauna
(Aldunate 2011). Los resultados obtenidos a partir de los análisis de isotopos estables,
realizado a los restos bioantropológicos revelaron un predominio del consumo de dieta
marina (Castro V Com. Pers. 2013).
Al exterior del alero fueron registrados variados instrumentos líticos y uno conquiológico
correspondiente a un cuchillo manufacturado sobre valva de choro zapato (Choromytilus
chorus) (Castro P 2011a), pigmento rojo, un pequeño rollo de piel de Lobo marino
(Otariidae), cerámica Loa café alisada y algunos fragmentos atribuibles al Periodo
Intermedio Tardío, una cuenta de concha posiblemente de bivalvo, una cuenta de
picoroco (Megabalanus psittacus) (Castro P 2011a) un anzuelo de cobre y un fogón
(Aldunate 2011).
Túmulo 1:
Consiste en un montículo funerario con un agujero en el centro producto de saqueo. Este
túmulo presenta elementos similares a los descritos por Moragas (1982) en el sitio Cobija
10, ya que posee restos bioantropológicos, trozos de cactáceas, argamasa, rocas,
fragmentos de esteras y cordelería, además de ofertorios tales como, variadas taxa de
moluscos, un artefacto óseo, guijarros (Castro V 2011) una pluma de Petrel moteado

36
(Daption cápense) (Peña-Villalobos y Fibla 2011), material de relleno como cerámica Loa
café alisada (Varela 2011), lascas y conchuela. Cabe destacar la presencia de dos
elementos -algas y osteofauna- que no son mencionados por Moragas (1982) en la
descripción del material recuperado en el sitio Cobija 10.
Unidad 4:
Consiste en el perfil de saqueo de un túmulo funerario con similares características al
Túmulo 1. En él fueron hallados restos bioantropológicos, fragmentos de textiles y estera,
ofertorios tales como un pulidor, un instrumento lítico indeterminado, restos de osteofauna
y malacofauna, además de relleno compuesto por cerámica Sequitor negro pulido y
conchuela (Aldunate 2011).
Figura 12: Emplazamiento del sitio CF3.

37
b. P3:
Consiste en un basural; los basurales arqueológicos presentes en las costas son
denominados conchales (Núñez P 2003). Según la definición de Orquera y Piana (1992)
los conchales son producto de la acumulación de residuos de alimentación humana. Se
caracterizan por el predominio de conchas de moluscos, además de la presencia de
huesos de animales de distintas especies, lentes calcinadas y carbonosas, utensilios
abandonados, residuos de su confección y proporciones diversas de matriz (Orquera y
Piana 1992:21).
Unidad 1:
Consiste en una unidad en la que se excavaron diversos líticos, tales como: lascas,
microlascas y guijarros, fragmentos de cerámica Loa café alisada, restos de osteofauna y
malacofauna, plumas de Gaviota dominicana (Larus dominicanus) (Peña-Villalobos y Fibla
2011), un artefacto conquiológico de Choro zapato (Choromytilus chorus) (Castro P
2011b), escasos restos de planta silvestre identificada como Flor de viuda (Zephyra
elegans) (Varas 2011), fragmentos de estera, una malla tejida, madera, madera trabajada
y carbón. Dos rasgos; el primero consistió en un evento de quema en el cual se
encontraban restos faunísticos no quemados, y el segundo consistió en un piso de ceniza
(López M 2011). Cabe mencionar que, si bien para este sitio no se realizaron fechados
radiocarbónicos, se atribuye al Período Formativo debido a la presencia de cerámica Loa
café alisada (Uribe et al. 2007) y de argamasa similar a la descrita por Moragas (1982).

38
Figura 13: Emplazamiento del sitio P3.
c. PC8:
Este sitio se compone de seis grandes depresiones de carácter habitacional. Hacia el
este, dentro del mismo sitio se ubica una gran estructura de piedra con cuatro hiladas
(Aldunate 2011).
Unidad 1
Consiste en una unidad excavada al interior de una de las habitaciones. En superficie se
registró cerámica Turi rojo alisado y Turi gris alisado (Varela 2011). En la capa 1 se
registró cerámica de los tipos Ayquina y Dupont, ambas asignadas al Periodo Intermedio
Tardío (Uribe et al. 2007, Varela 2011) y una preforma de punta de proyectil (Aldunate
2011). Mientras que en la capa 2 fue registrado un cuchillo manufacturado sobre valva de
ostión (Argopecten purpuratus) (Castro P 2011b), un fragmento de cerámica
indeterminado (Varela 2011), restos botánicos correspondientes a plantas cultivadas de
maíz (Zea mays), plantas silvestres de recolección como algarrobo (Prosopis sp.), plantas

39
silvestres como flor de viuda (Zephyra elegans) y espinas de cactácea, entre las cuales
una presenta evidencias de corte en el extremo opuesto a la punta (Varas 2011).
Cordelería sobre pelo de camélido y cabello humano, material osteofaunístico y
malacológico (Aldunate 2011), plumas de piquero (Sula variegata) y pelicano (Pelecanus
thagus) (Peña-Villalobos y Fibla 2011).
Figura 14: Emplazamiento del sitio PC8.
d. Chungungo 6:
Alero 1
Consiste en las afueras de un abrigo rocoso en el que fueron recuperados mineral de
cobre, variados líticos, fragmentos de cerámica indeterminada, una posible cuenta de
collar, pigmento rojo, material osteofaunístico y malacológico. En el tercer nivel fueron
registradas rocas que parecen haber sido acomodadas en diferentes tiempos de la
ocupación. Bajo estas rocas se halló una gran cantidad de valvas enteras, las cuales

40
corresponden a eventos de desconche de loco (Concholepas), chitón (Chitoniadae) y lapa
(Fisurella sp.) (Varas 2012).
Figura 15: Emplazamiento del sitio Chungungo 6.
1.8.5.2. Uso actual de Punta Tames:
Punta Tames es una localidad sumamente aislada con escasa presencia de pescadores,
siendo Caleta el Fierro el lugar con mayor actividad de la localidad. Esta caleta está
formada por seis casas construidas con material ligero (madera, lona, malla Raschel y
planchas de Zinc) y dos casas emplazadas sobre los cimientos de antiguas
construcciones de piedras y cemento. Algunas de estas casas poseen pequeños jardines
provistos de plantas y flores. La principal actividad desarrollada en esta caleta es la
extracción de guano de aves marinas con fines comerciales (Castro V et al. 2010).

41
Figura 16: Caleta el Fierro.
Figura 17: lsla guanera frente a Caleta el Fierro.

42
1.9. Utilización de vertebrados:
Por utilización de vertebrados se entiende a todos los recursos que el humano puede
aprovechar de peces –ictiofauna-, anfibios y reptiles –herpetofauna-, aves –avifauna- y
mamíferos –mastofauna-. Estos beneficios se reflejan en el área de la alimentación, las
materias primas, servicios y prácticas funerarias, así como en ámbitos de la eficacia
simbólica (Chaix y Méniel 2006).
En términos alimenticios, los vertebrados son aprovechados por su carne, menudillos o
viseras, sangre, grasa, leche, médula y huevos. Entre las materias primas posibles de
extraer de vertebrados se encuentran hueso, cuero, lana, pelo, plumas, tendones, viseras,
excremento, cuernos y astas. Algunos servicios de animales presentes en actividades
humanas son: el transporte de carga, el tiro de carros, asistentes de caza y vigilancia del
hogar. Respecto a la utilización de vertebrados en rituales mortuorios, se encuentran
sacrificios y entierros (Chaix y Méniel 2006).
Para los efectos de esta investigación sólo se considerarán el consumo alimentario y el
uso de huesos como materia prima, ya que la muestra se compone únicamente de restos
óseos.
1.9.1. Consumo:
Dentro de las necesidades vitales del humano se encuentra la alimentación. Las
principales fuentes de consumo son el agua, los productos terrestres y los marinos
(Núñez P 2003).

43
Fuentes de consumo Origen Producto
Agua Cursos de agua, aguadas, vertientes, torrentes
de agua.
Agua dulce o
semidulce
Productos terrestres Caza terrestre Vertebrados
Recolección terrestre Vegetales
Productos marítimos Caza y pesca en alta mar Vertebrados
Pesca en sector intermedios Vertebrados
Pesca y recolección en la orilla del mar Vertebrados
Mariscadores recolectores buceadores Invertebrados
Mariscadores recolectores en marea Invertebrados y
algas
Tabla 6: Fuentes de consumo, origen y producto (Elaboración propia basado en Núñez P
2003).
Según la tabla 6, durante el Período Formativo los vertebrados fueron obtenidos por
diversos medios, tales como: caza terrestre, caza y pesca de alta mar, pesca en sector
intermedio y pesca y recolección en la orilla del mar (Núñez P 2003). Sin embargo, dada
la ausencia de cursos fluviales los habitantes del litoral árido de arreismo absoluto
mantuvieron una dieta focalizada principalmente en productos marinos (Núñez y Santoro
2011).
Vertebrados consumidos en la costa arreica y su modo de obtención:
- Caza terrestre: los productos terrestres eran obtenidos mediante la caza comunitaria, en
ella se empleaban redes, trampas, piedras y artefactos como Waracas (hondas),
boleadoras, arcos y proyectiles. Los principales recursos terrestres eran guanaco (Lama
guanicoe) y taruca (Hippocamelus antisensis), además de variados taxones de roedores y
aves. Cabe mencionar que debido al intercambio con grupos de tierras altas eran
adquiridos productos terrestres exóticos (Núñez P 2003).

44
- Caza y pesca en alta mar: después del 200 d.C. algunos productos marinos eran
extraídos mediante el uso de embarcaciones de cuero de lobo en las que era posible
navegar en alta mar (Llagostera 1982). Mediante el empleo de artefactos tales como
arpones, anzuelos y palos eran obtenidos mamíferos marinos y peces de gran tamaño,
como lo son lobo marino (Otariidae), cetáceos (Cetácea), albacora (Xiphias gladius) y
congrios (Genypterus sp.) (Núñez P 2003).
- Pesca en sector intermedio: los productos del sector intermedio eran obtenidos mediante
la navegación por las orillas, en ella se utilizaban anzuelos, tridentes, arpones y redes.
Los principales recursos extraídos eran congrios (Genypterus sp), dorados (Coryphaena
sp.), atunes (Thunnus sp.), jurel (Trachurus murphyi), anchoa (Engralrs ringens) y sardina
(Sardinops sagax) (Llagostera 1982, Núñez P 2003).
- Pesca y recolección en la orilla del mar: mediante el uso de chopes y anzuelos eran
obtenidos vieja negra (Graus nigra), tollo (Mustelus mento), tomoyo (Auchenionchus
microcirrhis), lenguado de ojos chicos (Paralichthys microps), lisa (Mugil cephalus), jurel
(Trachurus murphyi) y pejeperro (Pimelometopon maculates) (Núñez P 2003).
En cuanto al consumo de vertebrados en el tramo ubicado entre Punta Atala y Punta
Tames, contamos con los análisis de osteofauna procedente de los sitios Mantos de la
luna 4 y Copaca 1, correspondientes al Período Arcaico y del sitio Guasilla 2,
correspondiente al Periodo Intermedio Tardío.
En el sitio Mantos de la Luna 4, se registró un predominio por parte de camélidos
(Camelidae); en menor medida se presentaron restos de lobos marinos (Otariidae) y
delfines (Delphinidae). Por otro lado, la menor frecuencia de los taxones cormorán
(Phalacrocorax sp.), piquero (Sula variegata), pelícano (Pelecanus thagus), gaviota (Larus

45
sp.) y pingüino (Spheniscidae) sugieren que el recurso aviar fue explotado en forma
oportunista y/o casual (Olguín 2011).
En el sitio Copaca 1 se registró una alta presencia de delfines (Delphinidae) seguidos de
lobos marinos (Otariidae) y camélidos (Camelidae). A juzgar por su gran aporte cárnico y
calórico además de encontrarse distribuidos homogéneamente a lo largo de toda la
secuencia cultural del sitio, es probable que dichos taxones fueran mayormente
explotados. Por otro lado, se detectó la presencia de un ejemplar de Chungungo (Lontra
felina) lo cual sugiere la caza oportunista de éste. Adicionalmente se registró la
explotación de avifauna, en la que se identificó la tendencia por fárdelas y petreles
(Procellariiformes), cormoranes y piqueros (Suliformes) y pelicanos (Pelecaniformes). En
menor medida fueron registrados gaviota (Charadriiformes) y pingüino (Sphenisciformes)
(Olguín et al. 2012, Peña-Villalobos et al. 2013).
En el sitio Guasilla 2, se detectó el predominio por parte de lobos marinos (Otariidae),
seguido de delfines (Delphinidae), camélidos (Camelidae) y cormoranes
(Phalacrocoracidae). En menor medida se identificaron piquero (Sula variegata), pelícano
(Pelecanus thagus), gaviota (Larus sp) y pingüino (Spheniscidae) (Olguín 2011).
1.9.2. Industria ósea:
Se denomina industria ósea a los conjuntos de utensilios elaborados con materiales de
origen animal, sobre los que el humano aplica determinadas técnicas de intervención con
la finalidad de modificarlos y producir útiles de trabajo (Eiroa et al. 1999). El hueso es una
materia prima de fácil obtención, puesto que la práctica de la caza de los animales
salvajes se ha llevado a cabo desde los orígenes de la vida humana (Eiroa et al. 1999).

46
Por ello, el material óseo constituyó un soporte comúnmente utilizado ya que es un tejido
duro y resistente, rígido y fácil de trabajar con los medios adecuados. La manufactura de
artefactos óseos debió ser tarea de artesanos especializados y dichas técnicas de trabajo
se resumen en percusión, incisión, pulimento o abrasión y perforación (Eiroa et al. 1999).
Llagostera (1989) menciona que durante el Período Arcaico en la costa Norte de Chile se
emplearon una serie de artefactos óseos para la explotación marítima, pero la mayor
variedad de estos se encuentra en el litoral árido de valles y quebradas -al norte de
nuestra área de estudio-. Estos artefactos fueron el arponcillo con cabezal de hueso; el
Chope, consistente en un instrumento de hueso enmangado, utilizado para extraer
moluscos univalvos y desconchar bivalvos; el anzuelo de hueso, constituido por un hueso
atado a una pesa de roca empleado para la pesca; y la Potera conformada por una pesa
de roca, un eje con garfios rectos de hueso, empleado para capturar pulpos y peces por
arrastre (Llagostera 1989, Standen 2003, Berenguer 2009). En cuanto al litoral de
arreismo absoluto, se atribuye el uso de anzuelos de hueso con cabezal de retención
(Llagostera 1989). Cabe mencionar que el tamaño de estos artefactos variaba de acuerdo
a la presa, y para su manufactura se utilizaban huesos de camélidos o de lobos marinos
(Dauelsberg y Álvarez 1968).
En cuanto a la explotación de recursos terrestres, el único artefacto óseo utilizado en el
Norte de Chile corresponde a los ganchos de estólica. La estólica consiste en un arma
propulsora que actúa como una prolongación del brazo y tiene por finalidad portar
efectividad y alcance al lanzamiento de un proyectil. Su utilización data del Período
Arcaico (Rivera y Zlatar 1982, Standen 2003).

47
También han sido hallados en calidad de ajuares y/u ofertorio, variados artefactos óseos,
tales como: cuentas de collar tubulares, espátulas, punzones, recipientes o cubiletes que
consistían en una diáfisis de ave sellada con piel de camélido y/o ave en sus dos
extremos. Estos recipientes probablemente eran utilizados para el almacenaje de
pigmentos (Standen 2003).
En el área de estudio se han realizado hallazgos y análisis de artefactos óseos, pero
ninguno de ellos asignado al Período Formativo. En el sitio Mantos de la Luna 4,
correspondiente al Período Arcaico fueron extraídos dos fragmentos de diáfisis pulidas
posiblemente asociadas al trabajo textil. En Guasilla 2, correspondiente al Período
Intermedio Tardío fueron hallados 29 artefactos óseos, entre los que se registraron
agujas, tubos, ganchos de potera, punzones y otros sin una función determinada. En
términos tafonómicos, en general, los artefactos poseen un estado de conservación de
regular a malo, ya que en general presentaron pérdida de tejido compacto y adhesiones
de sal (Santander 2011).
1.10. Utilización de invertebrados marinos:
Los moluscos componen un registro recurrente en las costas ya que son explotados como
alimento, como materia prima para fabricar instrumentos y como elementos simbólicos
(Corrales 1988-89). A partir del análisis de los restos malacológicos procedentes de los
sitios CF3, PC8 y P3 Castro P (2011a, 2011b) identificó taxones que fueron utilizados en
cada uno de estos ámbitos.
En términos alimenticios los invertebrados más consumidos corresponden a lapa
(Fisurella sp.), loco (Concholepas concholepas) y chitón (Chitonidae), seguidos de almeja
(Veneridae) choro zapato (Choromytilus chorus), picorocos (Megabalanus psittacus),

48
ostión (Argopecten purpuratus), erizo (Loxechinus albus) y jaiba mora (Homolaspis plana),
dichos taxones poseen una alta importancia económica (Castro P 2011a, 2011b).
La mayoría de las especies identificadas en el análisis proceden del intermareal rocoso
(Fisurellidae, Concholepas, Chitonidae, Loxechinus albus y Veneridae) y constituyen
taxones característicos del litoral árido de arreismo absoluto. Por otro lado, se detectó la
presencia de moluscos bivalvos de hábitat arenosos como ostión (Argopecten purpuratus)
y almeja (Veneridae), por lo cual Castro P (2011) postula que estas poblaciones
excepcionalmente realizaban recolecciones en playas arenosas (Castro P 2011a, 2011b).
En cuanto al uso simbólico de los invertebrados, fueron hallados restos de pequeños
caracoles, tales como: Turritela cingulata, Oliva peruviana, Crasilabrum crasilabrum,
Prisogaster niger, Nasarius gayi, Priene rude, Priene scabrum, Littorina peruviana,
Bulimulus sp., Acanthina monodon, Aenator fontainei, Sinum cymba y Cyclocardia
compressa en contextos fúnebres (Castro P 2011a, 2011b).
Finalmente, fueron registrados artefactos manufacturados sobre concha. Entre los
ornamentales se encuentran una cuenta de collar posiblemente de bivalvo y una pre
forma de cuenta de picoroco (Megabalanus psittacus), ambos procedentes de CF3. Entre
los artefactos utilitarios, en CF3 se encontró un cuchillo de choro zapato (Choromytilus
chorus) (Castro P 2011a), en P3 se registró un cuchillo de choro zapato (Choromytilus
chorus) y en PC8 un cuchillo de ostión (Argopecten purpuratus) (Castro P 2011b).
Tras el análisis de los restos malacológicos, Castro P (2011a) concluye que durante el
Período Formativo los grupos litoraleños de Punta Tames tuvieron preferencia por la
explotación de especies del intermareal rocoso. Explotando taxones de fácil obtención, a
excepción de algunas especies que eran extraídas mediante buceo como lo son el

49
picoroco (Megabalanus psittacus), el choro zapato (Choromytilus chorus) y en algunos
casos el erizo (Loxechinus albus).
Debido al gran tamaño de los individuos de lapa (Fisurellas) y loco (Concholepas)
analizados, Castro P (2011b) propone que los grupos litoraleños planificaban estrategias
de recolección, desarrollando una tendencia en la selección de individuos de mayor
tamaño, esto con la finalidad de permitir el crecimiento de los moluscos pequeños para
ser recolectados al alcanzar su talla de madurez. Esta especialización en las estrategias
de recolección de moluscos implicó la posterior realización de tareas específicas como el
buceo.

50
CAPÍTULO 2
2.1. Problemática y justificación:
En el marco del proyecto Fondecyt 1050991 3 , el equipo de trabajo encabezado por
Victoria Castro logró detectar evidencias bioantropológicas, orgánicas y culturales propias
de poblaciones de tierras altas del Loa Medio, quebradas y oasis de Atacama en el litoral,
desde el Período Formativo hasta el Tardío (con fechas entre el 315 d.C. y el 1450 d.C.).
Dichos hallazgos dieron lugar al desarrollo del proyecto Fondecyt N 11009514 , el que
surgió con la finalidad de esclarecer el contacto entre grupos circumpuneños y litoraleños,
específicamente en el tramo ubicado entre Punta Tames y Punta Atala, costa arreica de
Antofagasta. Cabe mencionar que esta área presenta un escaso registro de información
arqueológica entre los Períodos Formativo y Tardío (Castro V 2010).
Para inferir el interés de las poblaciones de tierras altas por el litoral, en este tramo, se
han realizado excavaciones estratigráficas, en las que se han recuperado una serie de
vestigios que han permitido la realización de diversos análisis: de restos
bioantropológicos, con el objetivo de constatar la presencia de individuos provenientes de
grupos étnicos diferentes y de restos culturales y bioindicadores, los que pudieron resultar
como producto de caza, pesca y recolección (Castro V 2010).
Por otra parte, los estudios realizados hasta la fecha no han abarcado la temática sobre el
uso de vertebrados en Punta Tames durante el Período Formativo. Las únicas evidencias
registradas en cuanto a este tema corresponden a los sitios: Cerro Colorado, Punta
Morada, Cobija 13, Los Canastos, Montenegro y Caleta Huelén 42, los que se encuentran
3 El desierto costero y sus vinculaciones con las tierras altas: de Cobija a Calama.
4 Del Período Formativo al Tardío en la costa de Antofagasta: cronología e interrelaciones con las tierras altas.

51
emplazados cercanos al área y poseen evidencias de consumo de peces entre el Período
Arcaico Tardío y Formativo Temprano (Llagostera 1989, 2005).
Si bien dentro del proyecto se han realizado análisis de osteofauna, ninguna de las
muestras proceden de sitios asignados al Período Formativo (Mantos de la luna 4 y
Copaca 1 con fechas arcaicas y Guasilla 2 correspondiente al Período Intermedio Tardío)
(Olguín 2011, Olguín et al. 2012).
Numerosos autores se han referido al modo de vida de las sociedades pescadoras de la
costa arreica durante diversos períodos (Moragas 1982, Llagostera 1982, 1989, 1990,
2005, Núñez L 1989, Núñez P 2003). Sin embargo, existe un vacío de información sobre
la utilización de vertebrados -en cuanto al consumo alimentario y al uso de huesos, como
materia prima-, por parte de los habitantes de Punta Tames durante el Período Formativo.
La carencia de esta información en tanto actividad cotidiana, es necesaria para la
comprensión de los eventos ocurridos y, por lo tanto, del desarrollo cultural. Cabe
mencionar, la presencia de restos arqueofaunísticos en sitios arqueológicos generalmente
son resultado de las actividades económicas y constituyen una evidencia mediante la cual
es posible aproximar patrones de conducta, a modo tal de comprender procesos
regionales y de interacción con tierras altas.
En consecuencia, se propone caracterizar la utilización de recursos faunísticos,
específicamente de vertebrados -respecto al consumo alimentario y al uso de huesos,
como materia prima- por parte de los grupos litoraleños que se asentaron en Punta
Tames durante el Periodo Formativo.

52
2.2. Objetivos:
2.2.1. Objetivo general:
Caracterizar la utilización de vertebrados durante el Período Formativo en la localidad de
Punta Tames, costa arreica de Antofagasta.
2.2.2. Objetivos específicos:
Para cumplir el objetivo general se comprenderán muestras de osteofauna procedentes
de excavaciones estratigráficas en los sitios CF3, P3, PC8 y Chungungo 6.
Contextualizar el área de estudio en términos geográficos, zoológicos y
arqueológicos.
Analizar el registro osteofaunístico de los sitios.
Establecer el consumo alimentario.
Detectar la presencia de taxones eventualmente procedentes de otros pisos
ecológicos.
Identificar y caracterizar instrumentos óseos.
2.3. Marco teórico:
El interés por el estudio de restos animales y vegetales surge en el marco de la
arqueología procesual con Clarke (1984), quien postula que muchos aspectos de las
sociedades antiguas pueden ser comprendidos mediante el estudio de cómo estas se
adaptaron a su entorno.
Desde esta perspectiva, la Ecología Cultural principalmente a través de J. Steward,
establece que la adaptación al medio ambiente promueve al cambio cultural (Renfrew y

53
Bahn 1998). De esta manera el estudio de los restos faunísticos procedentes de sitios
arqueológicos constituye una herramienta que permite inferir sobre la relación del ser
humano con el medio ambiente en el que se desenvuelve, y cómo se generan los
cambios culturales (Mengoni 1988).
En este contexto se proponen los siguientes lineamientos teóricos:
2.3.1. Grupos litoraleños y adaptación:
En tiempos prehispánicos, el litoral árido de arreismo absoluto fue poblado por grupos
humanos que se caracterizaron por una alta movilidad y una economía basada en la caza,
pesca y recolección (Núñez P 2003). Estos grupos no estuvieron exentos del contacto
con viajeros del viejo mundo. La presencia colonial trajo consigo el surgimiento de
variadas formas de denominación para los habitantes de dicho litoral (Letelier 2011), entre
las que han persistido los términos: Uros, Puquinas, Pro-anches, Camanchacas,
Camanchangos y Changos (Bittmann 1980, Castro V 2009).
Los apelativos Uros y Puquinas probablemente eran empleados para segmentos
poblacionales que se asentaban en el litoral, pero que originalmente provenían del
altiplano, específicamente de islas y/u orillas del lago Titicaca (Rostworowski 1986).
El término Camanchacas se referiría a las poblaciones que se asentaban a lo largo de la
costa de Tarapacá. El seudónimo Pro-anches, correspondía a poblaciones de la costa de
Antofagasta (Núñez P 2003) y pudo ser aplicado a Camanchacas no sometidos
(Casassas 1992). Por otra parte, Camanchangos hacía referencia a una clasificación
exclusiva para aquellos grupos que intercambiaban productos desde Cobija al interior de
Atacama (Martínez 1985a).

54
Los términos Camanchacas y Camanchangos, habrían sido utilizados como sinónimos, y
a raíz de ambos derivó el concepto de Chango (Rostworowski 1986), esta designación fue
acuñada por cronistas el siglo XVII y actualmente es empleado como un apelativo
genérico para referirse a dichos grupos (Bittmann 1984, Castro V 2009).
Compartimos con Martínez (1985b) que los variados términos registrados por los
cronistas pudieron ser aplicados como meras denominaciones para la distinción de
actividades económicas y/o estados ocupacionales especializados y no corresponder a
entidades étnicas vinculadas al parentesco. Letelier (2011) plantea que aunque estos
grupos presenten diferencias en la extracción de recursos, es prácticamente imposible
reconocer y/o establecer el carácter étnico de cada grupo. En base a lo expuesto y
considerando que la presente memoria se enmarca cronológicamente en el período
Formativo, el término Chango resulta excluyente ya que solo se refiere a los grupos de
tiempos históricos.
Otro término apelativo aplicado a estos grupos es el mencionado por Llagostera (1982)
quien se refiere a sociedades pescadoras, contemplando únicamente a la pesca y
aislando otros recursos importantes para la subsistencia. Para efectos de esta memoria
se adoptará el termino de grupos litoraleños (Llagostera 1982, Núñez P 2003) ya que
excluye especificaciones étnicas, ocupacionales, espaciales y temporales.
Los grupos litoraleños desarrollaron una alta eficiencia adaptativa gracias a su óptimo
modo de producción (Llagostera 1982). Por adaptación entendemos un proceso
compuesto por fases sucesivas, mediante las cuales es posible acceder a nuevos niveles
de energía. En cada uno de estos niveles se aplican estrategias de adaptación, cada vez
más eficaces con la finalidad de optimizar la extracción de recursos y de energía

55
disponibles (Cohen 1974). Para Perlman (1980) la adaptación consiste en que los
recursos adquiridos proporcionan mayor energía que la gastada en el proceso de
obtención, considerando las oportunidades que ofrece el medioambiente habitado.
Entonces, la adaptación al medio ambiente motiva al cambio cultural (Renfrew y Bahn
1998), en ese sentido; incorporamos la propuesta de Binford (1988) quien define el
concepto de cultura como un medio extra somático de adaptación.
Los grupos litoraleños ejecutaron un óptimo modo de producción, basado en la extracción
de recursos litorales. Llagostera (1982, 1989) propuso un modelo teórico para la
comprensión de la apropiación de recursos costeros por parte de estos grupos. Este
modelo postula tres etapas sucesivas denominadas dimensiones. La primera etapa
corresponde a la dimensión longitudinal, cuyo inicio data del 8000 a.C. aproximadamente
y consistió en la recolección de recursos en la franja intermareal. Se denomina dimensión
batitudinal a la segunda etapa situada desde el 6500 a.C. aproximadamente. La principal
característica de esta etapa radica en la anexión de nuevas tecnologías como lo son el
uso de anzuelos y arpones, estas herramientas generaron un cambio favorable; el acceso
a recursos ictiológicos de profundidad. Finalmente, la dimensión latitudinal se inició el 200
d.C y consistió en la incorporación del uso de embarcaciones, las que funcionaron como
medios de transporte a una mayor extensión, permitiendo el acceso mar a dentro y, en
consecuencia, a nuevos recursos (Llagostera 1982, 1989).
Cabe mencionar que el área de estudio se encuentra sumamente limitada respecto de
recursos alimenticios terrestres, por este motivo el mar debió actuar como fuente
proteínica por excelencia (Llagostera 1982, 1989). Todo esto, derivó en la imperante
necesidad de producir tecnología especializada para la extracción de recursos costeros.
Esta optimización de los medios de producción permitió la conquista sucesiva de las

56
fracciones tanto físicas como energéticas del mar, logrando eficiencia adaptativa
(Llagostera 1982). Compartimos con la propuesta de Perlman (1980) quien señala que
basta que un grupo obtenga parte de sus recursos del mar para hablar de adaptaciones
costeras.
En consecuencia, desde esta perspectiva, el cambio cultural está condicionado al medio y
es así como las fases del proceso de adaptación suceden en tanto sea requerido. Prueba
de ello, es la eficiente adaptación costera desarrollada por los grupos litoraleños.
2.3.2. Arqueología de los animales:
La zooarqueología, en tanto disciplina enfocada al análisis de restos faunísticos
procedentes de sitios arqueológicos, permite reconstruir de manera virtual los procesos
culturales de los grupos humanos de tiempos pretéritos (Davis 1989, Reitz y Wing 1999,
Chaix y Méniel 2006, Gamble 2008). Esta disciplina se focaliza únicamente en aquellos
animales que se relacionaron con las personas. Sin embargo es necesario destacar que
su objetivo es el estudio de los humanos, ya que la información que puede adquirirse a
través de los restos arqueofaunísticos es de gran utilidad para comprender a las
sociedades humanas del pasado (Davis 1989).
Los restos faunísticos procedentes de sitios arqueológicos poseen un alto potencial
informativo sobre aspectos culturales y bilógicos; a partir de ellos es posible conocer la
relación entre el humano y la fauna a través del tiempo, determinar el rol de los animales
al interior de una sociedad, medir el impacto que el humano generó sobre los animales,
aproximarse a las estrategias adaptativas, como también a las consecuencias que
pudieron conducir a la extinción de algunas especies de animales, los cambios culturales
producidos por el proceso de extinción y de domesticación, las modificaciones somáticas

57
y morfológicas producto de la domesticación de ciertas especies, por último, los restos
arqueofaunísticos también aportan a la comprensión de las variaciones en la distribución
espacial y temporal de los animales (Mengoni 1988).
La mayoría de los restos faunísticos registrados en sitios arqueológicos corresponden a
fragmentos óseos de mastofauna, avifauna, herpetofauna, ictiofauna y malacofauna
(Davis 1989). Considerando este registro, White (1953) estableció la relevancia de
analizar restos faunísticos con la finalidad de aproximar las estrategias de caza, los
patrones de trozamiento y de distribución, además de la presencia relativa de especies en
los sitios. A partir de esto, se sistematizó el análisis faunístico en arqueología y
comenzaron a inferirse las actividades que desarrollaban las personas al interior de
determinados contextos (Mengoni 1988).
El procesamiento de los animales involucra una serie de actividades que son desarrollas
en forma secuencial, las principales son: la evisceración, la extracción de sangre, órganos
internos y cuero, la reducción de la carcasa, el descarne de los huesos, la separación de
grasa, la extracción de medula ósea y la selección de unidades anatómicas para la
confesión de artefactos óseos (Lyman 1994). El trozamiento primario es ejecutado en el
lugar de obtención de la presa y tiene la finalidad de facilitar el transporte de la misma, en
estas situaciones quienes integraron la partida de caza pueden consumir algunas partes
anatómicas. El trozamiento secundario consiste en la división de unidades de trozamiento
primario y tiene por finalidad generar porciones menores para repartirlas entre los
integrantes de un grupo y el trozamiento terciario se realiza durante el consumo final, es
decir cuando se preparan las comidas o durante la ingesta (De Nigris 1994).

58
Los conjuntos arqueofaunísticos se caracterizan por presentar restos óseos con altos
grados de fragmentación, asociados a artefactos y en ocasiones, a evidencias de fuego.
La variación de abundancia y dispersión de los conjuntos arqueofaunísticos permite
identificar las actividades que se realizaron en dicho lugar, diferenciando los sitios de
matanza –trozamiento primario-, procesamiento –trozamiento secundario-, y acabado
final –trozamiento terciario- (Butzer 1989).
Los sitios de matanza se caracterizan por la presencia de restos óseos del esqueleto
axial, articulaciones óseas, bajos índices de fragmentación, dispersión y diversidad de
especies. Por otro lado, los sitios habitacionales o de acabado final se caracterizan por la
presencia de restos óseos del esqueleto apendicular, sin articulaciones óseas y altos
índices de fragmentación, dispersión y diversidad de especies. Los sitios de
procesamiento y descuartizamiento poseen una conjunción de características entre sitios
de matanza y de acabado final, siendo los artefactos asociados un recurso fundamental
para la interpretación de su funcionalidad (Butzer 1989).
Es necesario considerar que los huesos de animales presentes en basurales
probablemente correspondieron a restos de comida, así como los huesos asociados a
fogones, sugieren patrones culinarios asociados a la cocción (Davis 1989), cabe
mencionar que para este último caso, los huesos que presentan termoalteraciones
también pudieron ser empleados como combustible (Perlés 1977 en Mengoni 1988) y por
lo tanto, no estar estrictamente relacionados al consumo.
Cabe mencionar que la acumulación de resto faunísticos no siempre tiene un origen
antrópico, ya que los lechos de huesos pueden también ser producidos por agentes
animales y/o climáticos. Las acumulaciones por agentes animales consisten en lugares de

59
depositación de restos óseos faunísticos trasportados por carnívoros para su consumo o
por roedores para roer sus incisivos. Es por esto que suelen escasear elementos pesados
y/o voluminosos, como cráneos, siendo huesos largos los elementos mayormente
representados. Por otro lado, las acumulaciones geológicas consisten en lechos naturales
de huesos compuestos por restos de individuos muertos en épocas de stress climático
(Butzer 1989).
Otros elementos dignos de considerar respecto de los análisis zooarqueológicos son: los
taxones, el perfil etario y los cálculos de abundancia. La identificación de taxa revela la
conducta económica que desarrolló el grupo que originó el conjunto arqueofaunístico, la
edad de los individuos permite deducir el manejo que los grupos humanos tendrían sobre
los animales o bien la estación del año en el que sitio fue ocupado, finalmente, la
cuantificación de los restos óseos permite aproximar el número de personas que
habitaron cierto lugar (Davis 1989).
El enfoque de la presente memoria se centrará en restos arqueofaunísticos de
vertebrados, específicamente en sus restos óseos. Por lo tanto, en adelante el término
para referirnos a este registro será el de osteofauna. Los animales vertebrados se
caracterizan por poseer estructura ósea y por la presencia de un sistema nervioso central
que coordina las masas musculares y que se origina como un tubo hueco que se expande
por la región cefálica, dando la posibilidad de que prácticamente todas las neuronas
pueden acceder directamente a los nutrientes, lo que les confiere a un gran valor
adaptativo (Padilla y Cuesta 2003:13). Los vertebrados pertenecen al filo Cordados, el
que se constituye de 45.000 especies dentro de las cuales se encuentra el humano y los
animales directamente relacionados a su alimentación: ictiofauna, herpetofauna, avifauna
y mastofauna (Padilla y Cuesta 2003).

60
Cabe destacar que la taxonomía animal clasifica los diferentes tipos de individuos que se
encuadran dentro del reino animal en una serie de categorías concretas (Padilla y Cuesta
2003). Estas categorías se componen por organismos reconocidos como una unidad
formal a cualquier nivel de clasificación jerárquica siendo filo, clase, orden, familia, género
o especie, las principales categorías; cada uno de estos grupos es denominado taxón
(Simpson 1961, Grayson 1984, Reitz y Wing 1999). Se debe tener en consideración que
si bien cada especie posee características diagnósticas, no siempre es posible establecer
dicha categoría, puesto que las historias tafonómicas deterioran las piezas óseas. En
estos casos se denomina a un taxón más amplio, ya que la precisión no es estrictamente
necesaria, basta con asignar la familia (Becker 2004).
De acuerdo a lo expuesto, se tendrán en consideración las herramientas provistas por la
zooarqueología para la identificación de conjuntos osteofaunísticos, en tanto aporte a la
comprensión de los grupos litoraleños.
2.3.3. Procesos de formación de sitios arqueológicos e historias tafonómicas:
En arqueología el contexto implica una trama espaciotemporal en un medio cultural y/o no
cultural, factible de aplicar tanto a un sólo artefacto como a toda una constelación de sitios
(Butzer 1989). Existen múltiples definiciones para el término sitio arqueológico, entre ellas
se encuentra la de Binford (1964), quien lo define como un lugar en el que se hallan
características y/o artefactos de un grupo cultural. Por otro lado; Deetz (1967) se refiere a
sitio arqueológico como un lugar con concentración de material producido por actividades
humanas. House y Schiffer (1975) definen sitio arqueológico como un área dotada de
evidencias de comportamiento de grupos culturales del pasado.

61
Según Berenguer (1987a) un sitio arqueológico corresponde a aquel lugar que contiene
restos de una o más ocupaciones humanas y cuyos límites se definen según el suelo
estéril. Esta definición excluye temas tales como los procesos de formación, profundidad
de los depósitos y presencia de asociaciones, entre otros. La definición de sitio
arqueológico debe ser lo suficientemente flexible como para incluir hasta las más mínimas
transformaciones del ambiente provocadas por el hombre (Cornejo y Gallardo 1988). Por
ello, en el marco de esta memoria, se comprenderá al sitio arqueológico como el resultado
de actividades humanas posibles de observar a través de las evidencias, tanto culturales
como no culturales (Shiffer 1990). Cabe mencionar que la pobreza o riqueza relativa de
elementos arqueológicos no puede ser un criterio para definir un lugar como sitio
(Berenguer 1987a:73).
Por otra parte, Schiffer (1990) postula que los elementos presentes en los sitios
arqueológicos no son estáticos y poseen un historial. En base a ello, propone un modelo
en donde los artefactos forman parte de dos contextos distintos. El primer contexto es el
sistémico, en el cual los elementos son participes de un sistema de comportamiento.
Posteriormente, al ser abandonados, estos elementos forman parte del contexto
arqueológico, siendo olvidados y con el pasar del tiempo comienzan a adquirir
componentes no culturales y a sufrir cambios post-depositacionales (Schiffer 1990).
Compartimos la propuesta de Schiffer (1991) en tanto que los restos presentes en un sitio
arqueológico sufren transformaciones en forma sucesiva desde su participación en el
contexto sistémico, pasando por el contexto arqueológico, hasta su hallazgo y posterior
empleo como objeto de estudio por parte de los arqueólogos. Los procesos de formación
y transformación de sitios arqueológicos consisten en eventos, actividades y/o procesos
capaces de afectar a los restos arqueológicos posterior a su participación en el contexto

62
sistémico (Schiffer 1991). Estas transformaciones pueden deberse a agentes humanos,
geomorfológicos, biogénicos y/o fisiogénicos (Butzer 1989). La comprensión de los
procesos de formación permite esclarecer los factores influyentes en la conformación del
registro arqueológico, permitiendo estimar su acción particular (Jackson et al. 2005). Cabe
considerar que las particularidades del entorno de los sitios arqueológicos son claves en
los procesos de formación y transformación de los restos, puesto que la combinación de
los procesos culturales y no culturales determina su estado de preservación (Butzer
1989).
El estudio de los procesos de formación del registro arqueológico puede desarrollarse
desde variadas disciplinas como lo son la arqueología experimental (Baena 1997:3), la
etnoarqueología (Binford 1978) y la tafonomía (Gifford 1981, Lyman 1987, 1994), entre
otras.
La arqueología experimental es un modelo de contrastación de hipótesis a través de la
experimentación que de forma rigurosa admite la validez, para fases de la Prehistoria, de
un proceso técnico desarrollado en la actualidad (Baena 1997:3). La etnoarqueología
consiste en la aplicación del método etnográfico para el estudio de datos arqueológicos.
Vale decir, intenta reconstruir el pasado sobre la base de datos empíricos mediante la
observación participante (Binford 1978, 1967, Politis 2010). Para efectos de esta memoria
serán consideradas las herramientas contribuidas por la tafonomía, en tanto disciplina
enfocada en el estudio de la transición de los organismos desde la biosfera a la litósfera.
Se constituye como la ciencia de las leyes de integración o enterramiento que estudia los
procesos de fosilización, permitiendo reconstruir causas dinámicas a partir de registros
estáticos (Efremov 1940).

63
Los restos faunísticos procedentes de sitios arqueológicos constituyen un registro fósil, en
un espacio geográfico y un contexto geológico definidos (Lyman 1987). Los modelos e
hipótesis tafonómicos son entonces necesarios, entre otras cosas, para dar cuenta de la
formación del registro arqueofaunístico (Cruz et al. 1993-1994:94), ya que permiten
identificar procesos selectivos de acumulación y preservación de los restos óseos
depositados por diversas causas, tales como: la acreción mecánica, muerte natural de los
animales, caza por parte de carnívoros o intervención antrópica (Butzer 1989).
En general, la composición de los conjuntos fósiles sufre modificaciones que los diferencia
del conjunto original (Gifford 1981, Lyman 1994). El transporte y la destrucción diferencial
de los restos constituyen las principales causas de transformación. Ambos procesos
pueden estar originados en la acción de animales (carnívoros, roedores e insectos),
vegetales y/o fuerzas abióticas (Aire, fuego, tierra y agua) (Gifford 1981).
A partir de esto, serán consideradas las propuestas sobre los estudios de procesos de
formación de sitios arqueológicos para la comprensión de las historias tafonómicas de los
sitios que componen la muestra.

64
CAPÍTULO 3
3. Marco metodológico:
Para lograr los objetivos previamente planteados, el plan de trabajo de esta memoria ha
sido dividido en tres etapas: trabajo en campo, trabajo en laboratorio y trabajo en
gabinete. El estudio de recursos bibliográficos fue una actividad desarrollada en forma
permanente.
3.1. Trabajo de campo:
El trabajo de campo consistió en dos campañas de terreno, desarrolladas por el equipo
del proyecto Fondecyt 1100951. La primera fue realizada entre el 18 y el 28 de agosto del
2011, en ella se excavaron los sitios CF3, PC8 y P3. La segunda campaña fue realizada
entre el 12 y 24 agosto 2012, en ella se excavó el sitio Chungungo 6.
3.2. Trabajo de laboratorio:
En el trabajo de laboratorio se desarrolló la limpieza, identificación y análisis de los restos
osteofaunísticos procedentes de los sitios antes mencionados. Esta actividad se dividió en
dos etapas: data primaria y data secundaria.
La data primaria consistió en la identificación de las piezas, su lateralidad, completitud,
estado de fusión, articulación, meteorización, termoalteraciones, modificación culturales y
no culturales, Por otro lado, la data secundaria consistió en productos analíticos, tales
como el perfil etario, los modelos de faenamiento, la contribución dietaria, las estrategias
de procesamiento y el cálculo de medidas de estimación taxonómica y anatómica (Reitz y
Wing 1999).

65
Con respecto a la muestra, no se realizó selección, siendo analizado el universo total de
los restos osteofaunísticos recuperados en las excavaciones.
3.2.1. Análisis:
Para la ejecución del análisis fueron realizados los siguientes pasos: en primer lugar, se
realizó la limpieza de los restos que lo ameritaban. Para dicho procedimiento fueron
utilizados pinceles y brochetas, luego se procedió al análisis macroscópico en el cual se
identificaron la mayoría de las variables que serán descritas posteriormente. Tras esto, las
piezas óseas con huellas antrópicas fueron analizadas con lupa binocular de 20x.
Finalmente, se elaboró el registro visual de piezas relevantes; este registro consistió en la
toma de fotografías con cámara digital y dibujos de los artefactos para lo que se requirió
de lápiz, papel milimetrado y papel transparente.
Durante el análisis, los datos fueron relevados individualmente, es decir, se extrajo
información de cada una de las piezas óseas. Estos datos fueron introducidos en el
programa computacional Microsoft Excel, con la finalidad de generar una base de datos
capaz de incorporar toda la información registrada (Becker 2004, Velásquez 2004), la que
fue procesada durante el trabajo de gabinete.
Este procedimiento se llevó a cabo en el laboratorio de zooarqueología de la UISEK, entre
los meses de octubre y noviembre del 2012 (Mac-Lean 2012) y de junio del 2013.
3.2.2. Variables:
Para la realización del análisis fueron seleccionadas variables; cada una de ellas
constituyó un elemento distintivo que aportó al orden y comparación de la muestra
(Tamayo 2003).

66
Las variables fueron seleccionadas debido a su relevancia en tanto aporte a la discusión
final. La mayoría de ellas fueron propuestas por Mengoni (1999) en lo que dicho autor
denomina „Sistema para el registro de información arqueofaunística‟ donde señala la
importancia de considerar: taxón, unidad anatómica, porción, fusión, fracción, lateralidad,
fractura primaria, estado de fractura, marcas antrópicas –distribución, orientación y
frecuencia-, otras marcas, termoalteración y meteorización (Mengoni 1999). Por otro lado,
fueron agregadas variables consideradas relevantes tales como: sitio, unidad, nivel, capa,
rasgo, clase de Cordado, segmento, ubicación de la fractura, edad y observaciones. Cada
una de las variables posee códigos mediante los cuales es posible tabular los datos (Ver
anexo 3).
Variables y su relevancia:
Sitio, unidad, nivel, capa y rasgo: esta información es sumamente necesaria, ya
que consiste en el lugar del que fue extraída la unidad anatómica.
Taxón: consiste en asignar un taxón al nivel que sea posible.
Los taxones son grupos de organismos reconocidos como una unidad dentro de los
niveles de clasificación jerárquica: filo, clase, orden, familia, género o especie. Cabe
mencionar que la nomenclatura binominal da nombre a cada especie para lograr
identificarlas. Este nombre está compuesto por dos palabras: la primera indica el género,
el cual agrupa a determinadas categorías taxonómicas según las similitudes estructurales
y/o funcionales de las especies. La segunda palabra corresponde a una denominación
específica de la especie (Simpson 1961, Grayson 1984, Reitz y Wing 1999, Padilla y
Cuesta 2003).

67
Unidad anatómica 1: consiste en la identificación anatómica a nivel particular por
ejemplo: vértebra cervical (Mengoni 1999; Ver anexo 4).
Unidad anatómica 2: consiste en la identificación anatómica a nivel general por
ejemplo: vértebra (Mengoni 1999).
La asignación taxonómica y anatómica de las piezas analizadas se ejecuta mediante el
uso de criterios de identificación, tales como: forma, espesor, curvatura, forma y ubicación
de las superficies articulares, ubicación, tamaño y forma de los forámenes, ubicación y
forma de las zonas de fijación de ligamentos y tendones (Berwick 1975). Esta
identificación se realizó mediante un estudio comparativo con colecciones de referencias
pertenecientes al laboratorio de zooarqueología de la UISEK y colecciones privadas,
además de consultas a manuales de identificación de especies (Pacheco et al. 1979,
Cohen y Serjeantson 1986, Benavente et al. 1993, Falabella et al. 1995, Ericson y Stora
1999, Galotta y Márquez 2004, Hillson 2005, Adams y Crabtree 2008).
Segmento: se refiere a la zona del esqueleto a la que corresponde la unidad
anatómica. Por zona del esqueleto se entiende a las mayores porciones en el que
es segmentado (Mengoni 1988). Axial o apendicular, (craneal, caudal y precaudal
en el caso de la ictiofauna).
Lateralidad: consiste en el lado del cuerpo del que proviene la pieza ósea
(Mengoni 1999).
La asignación de lateralidad de las piezas óseas pares es necesaria para el cálculo de la
estimación del número mínimo de individuos (MNI) (Grayson 1984, Mengoni 1988, Lyman
2008).

68
Porción: consiste en la parte de la pieza ósea que se encuentra representada
(Mengoni 1999).
Esta información resulta útil para el cálculo de las unidades anatómicas mínimas (MAU),
el que permite detectar las piezas óseas mayormente representadas en la muestra
(Binford 1978, 1984).
Fusión: consiste en el estado de la fusión de la pieza ósea (Mengoni 1999), esto
permite estimar la edad del individuo al momento de su muerte (Mengoni 1988).
Edad: consiste en asignar un rango etario entre juvenil y adulto.
La fusión de las piezas óseas permite conocer las edades en las que se concentra la
sobrematanza, como así también, identificar el tipo de estrategia de subsistencia. La
dentición también posee un potencial informativo, ya que el reemplazo de los dientes
deciduos por los permanentes, el brote de molares y el desgaste también son aplicables a
la estimación del perfil etario (Mengoni 1988).
Marcas:
Las marcas presentes en las unidades anatómicas son testimonios de su historia. Es
posible identificar dos tipos de marcas: de origen natural y de origen antrópico (Binford
1984, Davis 1989, Chaix y Méniel 2006, Mengoni 1999, Reitz y Wing 1999).
Marcas antrópicas: consisten en las huellas realizadas por instrumentos (Mengoni
1999).
Las marcas de origen antrópico se dividen en dos categorías: la primera consiste en las
huellas originadas producto de la caza del animal, dicho de otro modo, son las heridas

69
que provocaron su muerte. La segunda categoría consiste en las huellas producidas por
la preparación para el consumo del animal (Chaix y Méniel 2006, Davis 1989). El análisis
de las modificaciones producto del procesamiento, preparación, consumo y descarte
permite esclarecer la etapa de explotación a la que estaban siendo sometidas las piezas
óseas (Mengoni 1999, Muñoz S 1997).
Para inferir la etapa de explotación a la que fue sometida la unidad anatómica son
utilizadas, a modo de complemento, las siguientes variables (Mengoni 1999):
Distribución.
Frecuencia.
Orientación, profundidad y largo.
Ubicación.
Estos datos permiten reconstruir los procedimientos culturales de reducción,
desollamiento, desarticulación, descarne y procesamiento de los animales (Mengoni
1988:93).
Marcas naturales: consisten en huellas producidas por agentes no humanos, tales
como plantas y/o animales (Mengoni 1999).
Las marcas naturales se producen debido a que las unidades anatómicas se encuentran
expuestas a una serie de agentes que participan en la degradación. Como por ejemplo,
los procesos tafonómicos, el crecimiento de raicillas y las marcas dentarias de carnívoros
o de roedores (Chaix y Méniel 2006, Grifford 1981).
Meteorización: consiste en la identificación del estadio de meteorización en el que
se encuentra la pieza ósea (Mengoni 1999).

70
La meteorización es un proceso que ocurre en el contexto arqueológico, en el que los
restos óseos son atacados por agentes físicos y/o químicos. Esto produce la separación y
destrucción de sus componentes microscópicos orgánicos e inorgánicos, por lo que
Behrensmeyer (1978) define seis estadios de meteorización, los que corresponden a
etapas de deterioro progresivo (Ver anexo 3).
Las marcas naturales y la meteorización permiten reconstruir las historias tafonómicas de
los sitios (Lyman 1987).
Fracción: consiste en el porcentaje de la pieza ósea representada (Mengoni 1999),
este cálculo se realiza proyectando el porcentaje ausente.
Fractura primaria: consiste en la morfología general de la fractura (Mengoni 1999).
Estado de fractura: consiste en el estado en el que se encontraba la unidad
anatómica al momento de ser fracturada (Mengoni 1999).
Las fracturas se originan por diversos factores en distintos momentos, ya que pueden ser
producidas por factores biológicos no humanos, como también pueden ser resultado de
actividades humanas relacionadas con el procesamiento del animal (Mengoni 1988) o
bien, pueden generarse al momento de la excavación estratigráfica. El análisis de la
morfología y el estado de la fractura resultan útiles para esclarecer su origen.
Termoalteración: consiste en las alteraciones térmicas presentes en la unidad
anatómica (Mengoni 1999).
El fuego puede estar relacionado a la cocción de carne, aunque no puede ser descartada
la posibilidad de que las piezas óseas quemadas hayan sido arrojadas a un fogón por
descarte o bien empleadas como combustible (Perlés 1977 en Mengoni 1988). Las

71
alteraciones térmicas son identificadas de acuerdo al color y modificación de la superficie
de la pieza ósea (Mengoni 1988, 1999).
Observaciones: consiste en un apartado para realizar comentarios con respecto a
la unidad anatómica analizada.
3.3. Trabajo de gabinete:
El Trabajo de gabinete consistió en la realización de gráficos, tablas y estimación del
número de especímenes identificados (NISP), del número mínimo de individuos (MNI), del
número mínimo de elementos esqueletales (MNE) y de las unidades anatómicas mínimas
(MAU). Esto con la finalidad de articular la información mediante una metodología
cuantitativa. De este modo, generar datos objetivos a través de la interpretación de los
gráficos y tablas, además de su posterior vinculación a la bibliografía.
3.3.1. Gráficos y tablas:
La identificación de las variables estuvo sujeta al potencial informativo de los restos
osteofaunísticos. Por ello, no todas las piezas óseas fueron identificadas con la misma
precisión, en algunos casos fue posible establecer el orden, familia, género o especie,
mientras que para otros solo es posible referirse a la clase. Lo mismo ocurre con la
asignación de la unidad anatómica, algunas piezas fracturadas o meteorizadas no pueden
ser identificadas (Mengoni 1988).
Mediante las variables registradas en el programa computacional Microsoft Excel fue
posible contabilizar los resultados mediante la realización gráficos y tablas.

72
Los gráficos y tablas realizados fueron los siguientes:
Filtro de informe Rótulos de comuna Rótulos de fila
Sitio Unidad Clase
Sitio Unidad Taxón
Sitio - Unidad Nivel Taxón
Sitio Unidad Perfil etario
Sitio Unidad Estado de fractura
Sitio Unidad Termoalteración
Sitio – Unidad Unidad Meteorización
Sitio Unidad Fragmentación
Sitio Unidad Marcas naturales
Sitio Clase Segmento
En el caso de los instrumentos óseos y las piezas óseas con huellas de origen antrópico
fueron realizadas tablas con variables específicas.
Tabla de descripción de unidades anatómicas con marcas de origen antrópico:
Sitio Unidad Nivel Pieza Taxón Tipo Distribución Orientación Frecuencia Ubicación
ósea de profundad
marca largo
Tabla de descripción de instrumentos óseos:
Sitio Unidad Nivel Pieza Taxón Función Forma Etapa de Conservación Trat. de
ósea
Prod. superficie
Posterior a la realización de dichos gráficos y tablas fueron realizadas medidas de
abundancia taxonómica mediante la estimación del número de especímenes identificados
(NISP) y del número mínimo de individuos (MNI). Además de medidas de abundancia

73
anatómica mediante la estimación del número mínimo de elementos esqueletales (MNE) y
de las unidades anatómicas mínimas (MAU).
3.3.2. Métodos para calcular la abundancia taxonómica:
El NISP consiste en la cantidad de huesos por taxón; puede estar determinado a nivel de
orden, familia, género o especie (Grayson 1984, Lyman 2008). El MNI es calculado a
partir de las unidades anatómicas mayormente representadas. En esta ocasión fue
considerada la lateralidad -izquierda o derecha- para los huesos pares (Grayson 1984,
Lyman 2008) y la cresta supraoccipital para la especie Trachurus murphyi, ya que
consiste en un hueso del neurocráneo sumamente distintivo y de buena preservación
(Falabella et al. 1995). Junto a la lateralidad de la pieza ósea se considera la edad del
individuo, la cual es estimada por la fusión de los huesos. Estos cálculos se aplican para
medir la importancia relativa de cada especie al interior de un conjunto óseo (Mengoni
1988).
3.3.3. Métodos para calcular la abundancia anatómica:
El MNE consiste en la determinación de la abundancia en que cierta unidad anatómica se
encuentra representada en la muestra -independiente de su lateralidad- (Lyman 1994).
Por su parte, el MAU permite establecer cuál es la unidad anatómica más representada
en la muestra. Este cálculo se realiza dividiendo el MNE de cada unidad anatómica por
las veces que dicha unidad se encuentra en el esqueleto (Binford 1978, 1984, Lyman
2008).
Cabe mencionar que para el cálculo del MAU no se contabilizan todas las unidades
anatómicas, sino que se consideran las zonas diagnosticas de las mismas. Dicho de

74
otro modo, el MAU es calculado en base a porciones de las piezas óseas más
representativas debido a sus características, intrínsecas, como por ejemplo la alta
densidad (De Nigris 1994).
3.3.4. Articulación de datos:
El tipo de estudio que define a esta investigación es del tipo correlacional, ya que se
busca explicar el comportamiento y valor de las variables. Vale decir, que se intentará
percibir cómo se relacionan o vinculan diversos fenómenos entre sí, o si es que estos no
se vinculan. Destacamos que la investigación correlacional posee, en alguna medida, un
valor explicativo. En base a ello, se pretende esclarecer qué provoca el fenómeno
(Hernández Sampieri et al. 1998).
Tras la interpretación de los gráficos fue recopilada información sobre la taxonomía,
distribución, longitud, etología, forma de obtención y relación con los humanos de cada
uno de los taxones identificados durante el análisis.
Estos datos sumados a los antecedentes y al marco teórico explicitado, fueron vinculados
con la finalidad de generar una conclusión y cumplir los objetivos de la presente memoria.

75
CAPÍTULO 4
4. Resultados:
4.1. Muestra:
De un total de 327 piezas óseas, un 45% (n = 148) fue adscribidle a la categoría
„identificado‟ -asignados a nivel de orden, familia, género o especie-. El 55% (n = 178)
restante fue asignados a la categoría „no identificado‟ -adscribidles a nivel de clase-.
Figura 18: Piezas óseas identificadas y no identificadas.
Las piezas óseas identificadas correspondieron a Egretta sp. 24% (n = 36), Otariidae 22%
(n = 33), Phalacrocorax sp. 15% (n = 22), Lama sp. 8% (n = 12), Trachurus murphyi 8% (n
= 12), Pelecanus thagus 8% (n = 11), Larus dominicanus 7% (n = 10), Cilus gilberti 6% (n
= 9), Sula variegata 1% (n = 2) y Spheniscus humboldti 1% (n = 1).

76
Figura 19: Piezas óseas identificadas a nivel de orden, familia, género o especie.
Dentro de la categoría „no identificado‟ las piezas óseas fueron asignadas a una clase.
Resultando un 58% (n = 103) avifauna marina, un 28% (n = 49) ictiofauna de la clase
Actinopterygii, un 12 % (n = 23) mastofauna terrestre y finalmente un 2% (n = 4)
mastofauna marina.
Figura 20: Piezas óseas identificadas a nivel de clase.

77
Cabe mencionar que para la mayor parte de la muestra no fue posible dar asignación
taxonómica a nivel de especie dada la baja tasa de identificación producto del deterioro de
las piezas.
4.2. CF3:
Identificación taxonómica:
Figura 21: Frecuencia de taxones en cada nivel de Túmulo1 y Unidad 4, Sitio CF3.
En las unidades Túmulo 1 y Unidad 4 del sitio CF3 registraron un total de 42 piezas
óseas. Los vertebrados con mayor representatividad fueron de la familia Otariidae,
seguido de mastofauna terrestre indeterminada y, en menor medida fueron registrados:
avifauna indeterminada, ictiofauna indeterminada y Pelecanus thagus.

78
Figura 22: Frecuencia de taxones en nivel 1 de Alero 1, Sitio CF3.
En la unidad Alero 1 del sitio CF3 fueron registradas 64 piezas entre las cuales se detectó
un fuerte predominio por parte de la avifauna. Los taxones mayormente representados
fueron Egretta sp. e ictiofauna indeterminada, seguido de avifauna indeterminada, Larus
dominicanus y Phalacrocorax sp. En menor medida se registraron Otariidae, mastofauna
terrestre indeterminada, Lama sp. y Pelecanus thagus.
Taxón NISP % MNI
Egretta sp. 16 25 1
Lama sp. 2 3 1
Otariidae 30 47 1
Phalacrocorax sp. 5 8 1
Larus dominicanus 9 14 2
Pelecanus thagus 1 2 1
Total 63 100
Tabla 7: NISP y MNI, sitio CF3.
El cálculo del MNI indicó la presencia de al menos dos individuos de Larus dominicanus,
del resto de las taxas se identificó la presencia de al menos un individuo.

79
Tafonomía:
Meteorización:
CF3 Total general
Meteorización Alero 1 Túmulo 1 Unidad 4
0 58 7 65
1 4 3 1 8
2 26 1 27
3 1 4 5
4 1
1
Total general 64 36 6 106
Tabla 8: Estadios de meteorización, sitio CF3.
En cuanto a los niveles de meteorización, en la unidad Alero 1 el 90.5% (n=58) no
presentó meteorización, el 6.5% (n = 4) presentó fisuras paralelas a la estructura fibrosa,
el 1.5% (n=1) presentó aspecto fibroso y/o destrucción del tejido compacto y el 1.5% (n =
1) presentó agrietamientos profundos y abiertos, desprendimiento de astillas. Por su
parte, en la unidad Túmulo 1 el 72% (n = 26) presentó descascaramiento y exfoliación del
tejido externo, el 20% (n = 7) presentó superficie lisa sin agrietamientos y el 8% (n = 3)
presentó fisuras paralelas a la estructura fibrosa. Finalmente en la Unidad 4 el 68% (n = 4)
presentó aspecto fibroso y/o destrucción del tejido compacto, el 16% (n = 1) presentó
descascaramiento y exfoliación del tejido más externo y el 16% (n = 1) presentó fisuras
paralelas a la estructura fibrosa.

80
Estado de fractura:
CF3 Total general
Estado de fractura Alero 1 Túmulo 1 Unidad 4
Fresca 3 3
No fresca 47 36 6 89
Sin fractura 14
14
Total general 64 36 6 106
Tabla 9: Estado de fractura, sitio CF3.
El 2% (n = 3) de las piezas presentaron fracturas realizadas mientras el hueso se
encontraba fresco, esto puede ser resultado de actividades humanas relacionadas con el
procesamiento del animal. Cabe mencionar que las piezas con fracturas en estado fresco
proceden de la unidad Alero 1. El 85% (n = 89) de las piezas sufrieron fractura en estado
no fresco, vale decir, fueron fracturadas al interior del contexto arqueológico o durante la
excavación. El 13% (n = 14) de las piezas se presentó completa.
Fragmentación:
CF3 Total general
Fracción Alero 1 Túmulo 1 Unidad 4
10% 12 33 5 50
20% 1 1 2
30% 3 1 1 5
40%
50% 1 1
60%
70% 2 2
80% 1 1
90% 31 31
100% 14 14
Total general 64 36 6 106
Tabla 10: Porcentajes de fragmentación, sitio CF3.
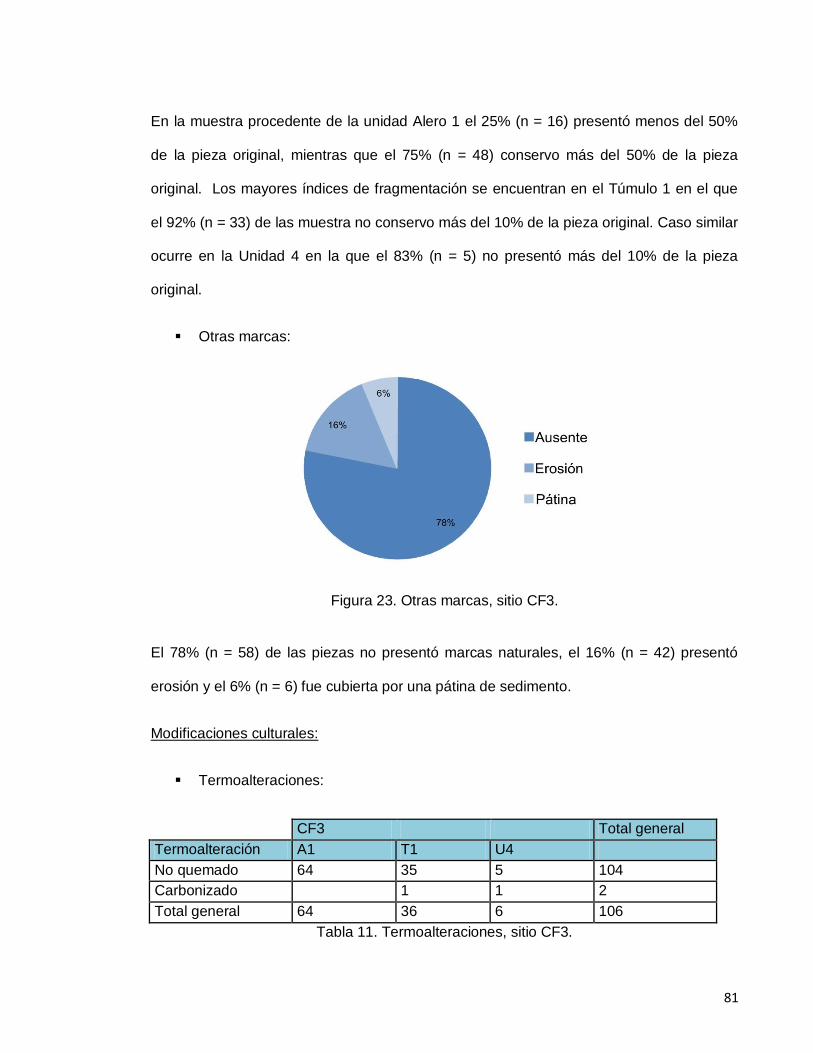
81
En la muestra procedente de la unidad Alero 1 el 25% (n = 16) presentó menos del 50%
de la pieza original, mientras que el 75% (n = 48) conservo más del 50% de la pieza
original. Los mayores índices de fragmentación se encuentran en el Túmulo 1 en el que
el 92% (n = 33) de las muestra no conservo más del 10% de la pieza original. Caso similar
ocurre en la Unidad 4 en la que el 83% (n = 5) no presentó más del 10% de la pieza
original.
Otras marcas:
Figura 23. Otras marcas, sitio CF3.
El 78% (n = 58) de las piezas no presentó marcas naturales, el 16% (n = 42) presentó
erosión y el 6% (n = 6) fue cubierta por una pátina de sedimento.
Modificaciones culturales:
Termoalteraciones:
CF3 Total general
Termoalteración A1 T1 U4
No quemado 64 35 5 104
Carbonizado 1 1 2
Total general 64 36 6 106
Tabla 11. Termoalteraciones, sitio CF3.

82
Del total de la muestra, el 1% (n = 2) de las piezas se hallaron carbonizadas, mientras que
el 99% (n = 104) de las piezas no presentaron alteración térmica. La primera pieza
carbonizada procede de CF3, unidad T1, nivel 4 y corresponde a un fragmento de cráneo
de mastofauna terrestre, la segunda pieza procede de CF3, unidad 4, nivel 2 y
corresponde a un hueso largo indeterminado de mastofauna terrestre.
Artefactos óseos:
Sitio Unidad Nivel
Pieza
ósea Taxón Función Forma
Etapa de
producción Conservación
Trat.
Superficie
CF3 T1 2 Indet.
Mastofauna Gancho
estólica Ovalada Terminado Incompleto Alisado Indet.
Fotografía:
Dibujo:
Tabla 12: Descripción e imagen de artefacto óseo. Sitio CF3, Túmulo 1, Nivel 3.

83
Sitio Unidad Nivel
Pieza
Taxón Función Forma Etapa de
producción Conservación
Trat.
ósea Superficie
CF3 A1 1
Hueso
Avifauna
Indet Inhalar Tubular Terminado Incompleto
Alisado e
Importa de
embarrilado
Largo
Indet
Fotografías:
Dibujo:
Tabla 13: Descripción e imagen de artefacto óseo. Sitio CF3, Alero 1, Nivel 1.

84
4.3. P3:
Identificación taxonómica:
Figura 24: Frecuencia de taxones en cada nivel del sitio P3.
En todos los niveles del sitio P3 la avifauna ostenta la mayor presencia, a excepción del
nivel dos en el que las piezas óseas corresponden principalmente a ictiofauna. El primer
nivel se caracterizó por su abundancia y heterogeneidad. En dicho nivel predominó la
avifauna indeterminada, seguida en forma decreciente de ictiofauna indeterminada, Cilus
gilberti, Lama sp., Trachurus murphyi, Egretta sp., Pelecanus thagus y Phalacrocorax sp.

85
En el segundo nivel, la presencia de avifauna disminuyó drásticamente, siendo restos
ictiológicos los principales componentes del nivel. En el tercer nivel, la avifauna volvió a
aumentar. Aunque para a la mayoría de las piezas óseas no fue posible asignar especie.
En este nivel fueron registradas piezas óseas correspondientes a mastofauna terrestre
indeterminada y a lama sp -casi ausentes en el nivel anterior-. La anteriormente
abundante ictiofauna se ausenta en éste y los siguientes niveles. En el cuarto nivel, se
produjo una fuerte reducción de material, en el que destacan escasos retos de avifauna,
mastofauna terrestre y una pieza correspondiente a Otariidae, constituyendo la única
presencia de mastofauna marina en el sitio. El resto del material de este nivel
corresponde principalmente a avifauna seguida de Lama sp y mastofauna terrestre
indeterminada. Finalmente, el nivel cinco presentó un predominio de avifauna,
representado en las especies: Pelecanus thagus, Phalacrocorax sp, Egretta sp, Sula
variegata, Larus dominicanus, Spheniscus humboldti y avifauna indeterminada, además
de mastofauna terrestre indeterminada y Lama sp.
Taxón NISP % MNI
Cilus gilberti 9 16 1
Egretta sp. 4 7 1
Lama sp. 10 18 1
Otariidae 1 2 1
Pelecanus thagus 8 14 1
Phalacrocorax sp. 10 18 2
Spheniscus humboldti 1 2 1
Sula variegata 2 3 2
Trachurus murphyi 10 18 10
Larus dominicanus 1 2 1
Total 56 100
Tabla 14: NISP y MNI, sitio P3.

86
Fue detectada la presencia de al menos 10 individuos de Trachurus murphyi, dos al
menos dos individuos de Phalacrocorax sp. y Sula variegata. Para el resto de los taxones
solo pudo establecerse la presencia de al menos un individuo.
Abundancia anatómica:
El sitio P3 fue el único que reunió los requisitos para realizar la estimación de abundancia
anatómica. De este modo, fue posible la realización de los cálculos MNE y MAU de Lama
sp, avifauna marina indeterminada e ictiofauna de la clase Actinopterygii.
NISP MNE MAU %MAU
1ª Falange 3 3 0.3 15
Escapula 1 1 0.5 25
Radioulna 2 2 2 100
Astrágalo 1 1 0.5 25
Calcáneo 1 1 0.5 25
Tabla 15: MAU Lama sp, sitio P3.
NISP MNE MAU %MAU
Carpo metacarpo 6 6 3 100
Coracoides 3 3 1.5 50
Fémur 1 1 0.5 1.6
Húmero 4 4 2 66
Pelvis 3 1 0.5 1.6
Radio 4 3 1.5 1.6
Escapula 4 4 2 66
Tibia 3 3 1.5 1.6
Tarso metatarso 4 4 2 66
Ulna 6 4 2 66
Tabla 16: MAU Avifauna marina, sitio P3.

87
Figura 25: MAU ictiofauna de la clase Actinopterygii, sitio P3.
Figura 26: Segmentos del esqueleto según clase, sitio P3.

88
Tafonomía:
Meteorización:
Figura 27: Estadios de meteorización, sitio P3.
En general, las piezas óseas se encontraban en buen estado de conservación. El 79% (n
= 136) no presentó alteraciones producidas por meteorización, en menor medida, el 13%
(n = 23) de las piezas presentaron descascaramiento y exfoliación del tejido más externo,
el 6% (n = 11) presentó fisuras paralelas a la estructura fibrosa, el 1% (n = 2) presentó
desprendimiento de astillas, agrietamientos profundos y abiertos. Finalmente, el 1% (n =
1) correspondió al unidades anatomías con aspecto fibroso y destrucción del tejido
compacto.
Estado de fractura:
Fractura Cantidad
Fresca 16
No fresca 145
Sin fractura 12
Total 173
Tabla 17: Estado de fractura, sitio CF3.

89
El 9% (n = 16) de las piezas presentaron fracturas en estado fresco, el 84% (n = 145) de
las piezas sufrieron fractura en estado no fresco y el 7% (n = 12) de las piezas se
presentó completa.
Fragmentación:
Fracción Cantidad
10% 102
20% 14
30% 7
40% 2
50% 3
60%
70% 3
80% 7
90% 23
100% 12
Total general 173
Tabla 18: Porcentajes de fragmentación, sitio P3.
El 59% (n = 102) de las piezas no conservó más del 10 % de la pieza original, el 15% (n =
26) conservó entre el 20% y el 50% de la pieza original. El 19% (n = 33) conservó entre el
70% y el 90% de la pieza original y el 7% (n = 12) corresponde a piezas conservadas en
un 100%.

90
Otras marcas:
Figura 28: Otras marcas, sitio P3.
El 73% (n = 126) de las piezas no presentó marcas naturales, el 13% (n = 19) presentó
diversos grados de erosión, el 11% (n = 19) se encontró cubierto por una pátina de
sedimento, el 2% (n = 4) presentó marcas de pisoteo y el 1% (n = 2) marcas modernas las
que pudieron ser producidas durante la excavación.
Modificaciones culturales:
Termoalteraciones:
Figura 29: Termoalteración, sitio P3.

91
El 1% (n = 2) de las piezas se presentan carbonizadas, el 99% (n = 171) restante, no
presentó alteraciones térmicas. La primera pieza procede del nivel 2 y corresponde a una
vértebra de ictiofauna de la clase Actinopterygii, la segunda pieza procede del nivel 3 y
corresponde a un hueso largo indeterminado de mastofauna terrestre.
Marcas de Procesamiento y/o consumo:
Sitio Unidad Nivel
Taxón
Tipo de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 3 Húmero
Sula
variegata Corte Aislada
Transversal
superficial
corta
Numerosas
(6 o más) Proceso
Imagen:
Tabla 19: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3, Unidad 1, nivel 3.

92
Sitio Unidad Nivel
Taxón
Tipo
de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 3
Tarso
Meta
Tarso
Phalacrocorax
sp. Corte
Pocos
Grupos
Transversal
superficial
corta
Pocas
(2 a 5) Proceso
Imagen:
Tabla 20: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, nivel 3.
Sitio Unidad Nivel
Taxón
Tipo
de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
Ósea
P3 U1 3 Falange
Lama
sp. Corte
Concentradas
en varios
grupos
Transversal
superficial
corta
Numerosas
(6 o más) Diáfisis
Imagen:
Tabla 21: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, Nivel 3.

93
Sitio Unidad Nivel
Taxón
Tipo de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 3 Astrágalo
Lama
sp. Corte Aisladas
Transversal
superficial
larga
Pocas
(2 a 5) Proceso
Imagen:
Tabla 22: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, Nivel 3.
Sitio Unidad Nivel
Taxón
Tipo de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 3 Calcáneo
Lama
sp. Corte Aisladas
Transversal
superficial
corta
Pocas
(2 a 5) Proceso
Imagen:
Tabla 23: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, Nivel 3.

94
Sitio Unidad Nivel
Taxón
Tipo de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 5 Escápula
Lama
sp. Corte Aisladas
Transversal
superficial
corta
Pocas
(2 a 5) Proceso
Imagen:
Tabla 24: Descripción e imagen de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, Nivel 5.
Sitio Unidad Nivel
Taxón
Tipo
de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
P3 U1 5
Hueso
largo
indet.
Lama
sp. Corte
Concentrado
en varios
grupos
Transversal
superficial
corta
Numerosas
(6 o más) Diáfisis
Imagen:
Tabla 25: Descripción e imágenes de unidad anatómica con huellas de corte. Sitio P3,
Unidad 1, Nivel 5.

95
4.4. PC8:
Identificación taxonómica:
Figura 30: Frecuencia de taxones en cada nivel del sitio PC8.
En el sitio PC8 se registró el predominio de Egretta sp, seguido de avifauna indeterminada
y Phalacrocorax sp. En menor medida se registraron Pelecanus thagus y Otariidae,
seguido de Trachurus murphyi y mastofauna terrestre indeterminada. En el segundo nivel
se observa un predominio de Egretta sp., seguido de avifauna indeterminada y
Phalacrocorax sp.
Taxón NISP % MNI
Egretta sp. 16 57 2
Otariidae 2 7 1
Pelecanus thagus 2 7 1
Phalacrocorax sp. 7 25 1
Trachurus murphyi 1 4 1
Total 28 100
Tabla 26: NISP y MNI, sitio PC8.

96
El cálculo del MNI indico la presencia de al menos dos individuos de Egretta sp. del resto
de las taxas se identificó la presencia de al menos un individuo.
Tafonomía:
Meteorización:
Figura 31: Estadios de meteorización, sitio PC8.
En general, las piezas óseas se encontraban en buen estado de conservación. El 77% (n
= 31) no presentó alteraciones producidas por meteorización, el 15% (n = 6) de las piezas
presentó fisuras paralelas a la estructura fibrosa, el 5% (n = 2) presentó agrietamientos
profundos y abiertos, además de desprendimiento de astillas, finalmente el 3% (n = 1)
correspondió al unidades anatomías con aspecto fibroso y destrucción del tejido
compacto.
Estado de fractura:
Fractura Cantidad
Fresca 19
No fresca 9
Sin fractura 12
Total 40
Tabla 27: Estado de fractura, sitio PC8.

97
El 47.5% (n = 19) de las piezas presentaron fracturas en estado fresco, el 22.5% (n = 9)
de las piezas sufrieron fractura en estado no fresco y el 30% (n = 12) de las piezas se
presentó completa.
Fragmentación:
Fracción Cantidad
10% 11
20% 2
30% 1
40% 1
50% 3
60%
70% 2
80% 2
90% 6
100% 12
Total general 40
Tabla 28: Porcentajes de fragmentación, sitio PC8.
El 27% (n = 11) de las piezas no conservó más del 10 % de la pieza original, el 18% (n =
7) conservó entre el 20% y el 50% de la pieza original. El 25% (n = 10) conservó entre el
70% y el 90% de la pieza original y el 30% (n = 12) corresponde a piezas conservadas en
un 100%.

98
Otras marcas:
Figura 32: Otras marcas, sitio PC8.
El 77% (n = 31) de las piezas no presento marcas naturales, el 20% (n = 8) presentó una
cobertura de pátina parcial y el 3% (n = 1) presentó erosión.
Modificaciones culturales
Termoalteraciones:
No fueron registradas piezas con alteraciones térmicas.
Marcas de Procesamiento y/o consumo:

99
Sitio Unidad Nivel
Taxón
Tipo
de
marca Distribución
Orientación
Profundidad
Largo Frecuencia Ubicación
Pieza
ósea
PC8 U1 1 Radio
Avifauna
índet Corte Aislada
Transversal
superficial
corta
Numerosas
(6 o más)
Epífisis Px
y diáfisis
Imagen:
Tabla 29: Descripción e imágenes de pieza ósea con huellas de corte. Sitio PC8, Unidad
1, Nivel 1.
4.5. Chungungo 6:
Identificación taxonómica:
Figura 33: Frecuencia de taxones en cada nivel de Alero 1, sitio Chungungo 6.

100
En la unidad alero 1 del sitio Chungungo 6 fue registrada una muestra escasa. El nivel
uno, en forma decreciente se presentó: mastofauna marina indeterminada, mastofauna
terrestre indeterminada y Trachurus murphyi. En el nivel dos fue recuperada una pieza
ósea, correspondiente a ictiofauna indeterminada.
Taxón NISP % MNI
Trachurus murphyi 1 100 1
Total 1 100
Tabla 30: NISP y MNI, sitio Chungungo 6.
Tafonomía:
Meteorización:
Figura 34: Estadios de meteorización, sitio Chungungo 6.
El 62% (n = 5) de las piezas no presentó alteraciones producidas por meteorización,
mientras que el 38% (n = 3) presentó descascaramiento y exfoliación del tejido externo.

101
Estado de fractura:
El 100% de la muestra presento fracturas en estado no fresco.
Fragmentación:
Fracción Cantidad
10% 3
20% 3
30%
40% 1
50% 1
60%
70%
80%
90%
100%
Total general 8
Tabla 31: Porcentajes de fragmentación, sitio Chungungo 6.
El 75% (n = 6) de la muestra no conservo más del 20% de la pieza original, el 25% (n = 2)
restante conservo entre el 40% y el 50% de la pieza original.
Otras marcas:
Figura 35: Otras marcas, sitio Chungungo 6.
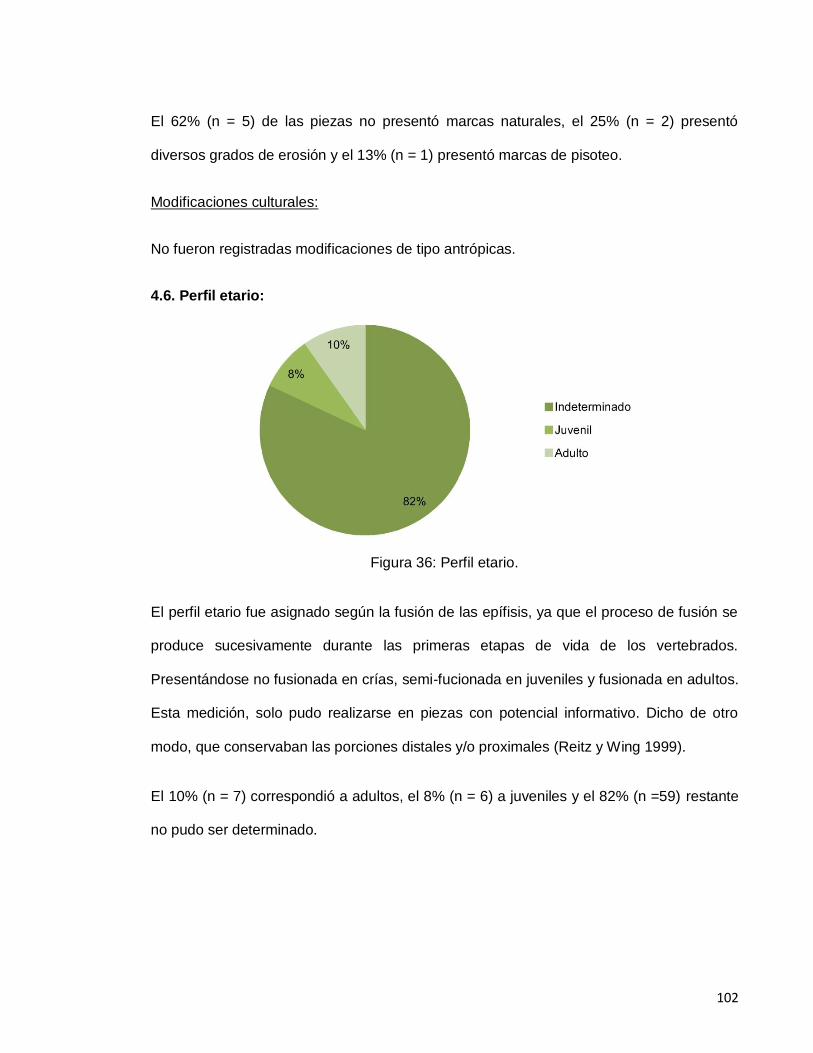
102
El 62% (n = 5) de las piezas no presentó marcas naturales, el 25% (n = 2) presentó
diversos grados de erosión y el 13% (n = 1) presentó marcas de pisoteo.
Modificaciones culturales:
No fueron registradas modificaciones de tipo antrópicas.
4.6. Perfil etario:
Figura 36: Perfil etario.
El perfil etario fue asignado según la fusión de las epífisis, ya que el proceso de fusión se
produce sucesivamente durante las primeras etapas de vida de los vertebrados.
Presentándose no fusionada en crías, semi-fucionada en juveniles y fusionada en adultos.
Esta medición, solo pudo realizarse en piezas con potencial informativo. Dicho de otro
modo, que conservaban las porciones distales y/o proximales (Reitz y Wing 1999).
El 10% (n = 7) correspondió a adultos, el 8% (n = 6) a juveniles y el 82% (n =59) restante
no pudo ser determinado.

103
4.7. Análisis de resultados:
4.7.1. Diversidad taxonómica:
Los resultados del análisis de la muestra de osteofauna, revelan la presencia de taxones
de avifauna marina, ictiofauna, mastofauna marina y mastofauna terrestre endémica de la
costa arreica. Siendo descartada la presencia de herpetofauna, fauna de hábitos
nocturnos y especies domesticadas (Iriarte 2008, Jaramillo 2005)
Entre las clases presentes en la muestra, la de mayor frecuencia resulto ser la avifauna
(n=185), seguida de ictiofauna (n=70), mastofauna marina (n=37) y mastofauna terrestre
(n=35).
Los taxones identificados fueron los siguientes:
Avifauna:
A nivel de especie fueron identificados Pelecanus thagus (pelicano), Sula variegata
(piquero), Larus dominicanus (gaviota dominicana) y Spheniscus humboldti (pingüino de
Humboldt). A nivel de género fue identificado Phalacrocorax sp. (cormorán) y Egretta sp.
(garza).
Ictiofauna:
A nivel de especie fueron identificados Trachurus murphyi (jurel) y Cilus gilberti (corvina),
ambos correspondientes a la clase Actinopterygii.

104
Mastofauna marina:
Fue identificada la familia Otariidae, pero no fue posible dar asignación taxonómica a nivel
especie, debido a que los otáridos del área de estudio se encuentran representada por
dos especies: Arctocephalus australis (lobo fino de dos pelos) y Otaria flavescens (lobo
marino de un pelo). Ambos taxones poseen una gran semejanza a nivel osteométrico
(Borella et al. 2010), adicionalmente, poseen un marcado dimorfismo sexual en el que la
hembra Otaria flavescens es sumamente similar al macho Arctocephalus australis (Iriarte
2008). A juzgar por lo anteriormente planteado, y debido a la ausencia de unidades
anatómicas clave, solo fue posible dar asignación taxonómica a nivel de familia.
Mastofauna terrestre:
Fue registrada la presencia del género Lama, debido a la estado de preservación de las
unidades anatómicas no fue posible realizar mediciones osteométricas para dar
asignación taxonómica a nivel de especie. Sin embargo, es probable que las unidades
anatómicas asignadas a este género correspondan a Lama guanicoe (guanaco) ya que
constituye una especie endémica (Iriarte 2008) con registros de consumo en el área
(Núñez P 2003).
En consecuencia y debido a que cada taxón posee características propias, fue recopilada
información sobre la taxonomía, distribución, longitud, etología, forma de obtención y
relación con los humanos de cada uno de ellos (Ver anexo 5).

105
4.7.2. Historias tafonómicas:
CF3, Túmulo 1 y Unidad 4:
Fueron registrados bajos niveles de preservación, ya que el 64% de la muestra presentó
descascaramiento y exfoliación del tejido externo, dicha meteorización podría deberse a la
deshidratación progresiva que sufre el material óseo previo a su enterramiento
(Behrensmeyer 1978, Laborde 1986, Mengoni 1988). Considerando que se trata de un
sitio saqueado es posible que estos restos se encontraran en superficie y fueran
agregados al sitio durante la excavación ilegal. Los estragos del saqueo también son
posibles de apreciar en los altos grados de fragmentación, ya que el 90% de la muestra
no conserva más de un 10% de la pieza original. Con respecto de las marcas naturales el
100% de la muestra presentó erosión. Por otro lado, fueron registradas unidades
anatómicas con fracturas en estado fresco.
CF3, Alero 1:
Se observó una buena conservación de la muestra, ya que el 90.5% no presento
meteorización. El 70% de las unidades anatómicas conservo más del 80% de la pieza
original. La presencia de marcas naturales fue casi nula, salvo escasas piezas que
presentaron pátina y el 6% de la muestra fue fracturado en estado fresco.
Cabe considerar que esta unidad se ubica al interior de un alero rocoso, los cuales se
caracterizan por permitir la acumulación de restos de animales producto de causas
naturales como lo son la acción de los carnívoros y las muertes naturales (Cruz 2011).

106
P3, Unidad 1:
El 79% de la muestra no presento meteorización. El 73% no presento marcas naturales.
En cuanto a la fragmentación, el 59% de la muestra conserva el 10% de la pieza original.
Estos altos índices de fragmentación podrían deberse a la calidad de conchal del sitio, en
donde los desechos son descartados y acumulados permitiendo la fractura de las piezas.
Cabe considerar que el 9% de las unidades anatómicas fueron fracturadas en estado
fresco.
Fue detectada la presencia de eventos de quema y lentes de ceniza con piezas óseas no
quemadas, esto podría deberse a que a medida de que el espacio se veía reducido -
próximo a colapsar- la basura era quemada para re ocupar el conchal.
PC8, Unidad 1:
En términos generales, se mantuvo un buen estado de conservación, donde el 77% de la
muestra no presento meteorización. El 77% no presento marcas naturales. Por otro lado
llama la atención que el 30% de las unidades anatómicas conservan el 100% de la pieza
original, sin embargo, un 27% conserva solo el 10%. En cuanto al estado de las fracturas,
el 47.5% presento fractura en estado fresco.
Chungungo 6, Alero 1:
El 62% no presento meteorización ni marcas naturales, ninguna unidad anatómica
conservo más del 50% de la pieza original y no fueron identificadas fracturas en estado
fresco.

107
4.7.3 Presencia taxonómica y anatómica:
CF3, Túmulo 1 y Unidad 4:
Identificación taxonómica:
Ambas unidades poseen escasa y homogénea presencia de mastofauna marina,
mastofauna terrestre, avifauna e ictiofauna. Con un fuerte predominio por parte de
Otariidae en superficie.
Según los antecedentes recopilados sobre cementerios de túmulos este tipo de sitios
carece de la presencia de vertebrados –a excepción de artefactos óseos- (Moragas 1982).
Esta información sumada a los altos grados de fragmentación y meteorización, además
de los antecedentes de saqueo. Permiten suponer que la presencia de osteofauna en este
sector del sitio se debe a eventos post-depositacionales disturbados por excavadores
ilegales.
En consecuencia, resulta impreciso considerar este registro, por lo que los resultados del
análisis de las unidades Túmulo 1 y Unidad 4 –a excepción de un artefacto óseo- no
serán incluidos en las conclusiones.
CF3, Alero 1:
Identificación taxonómica:
Fue registrada la presencia de ictiofauna, avifauna marina, mastofauna terrestre y marina,
esta unidad presenta un fuerte predominio por parte de avifauna marina, siendo Egretta
sp. el taxón mayormente representado. Es de suma importancia considerar que no
contamos con registros de explotación de individuos de este género, esto podría deberse

108
a que presenta bajos índices de masa utilizable. Sin embargo, no debe ser descartado su
aprovechamiento oportunista o en épocas de estrés.
La alta presencia de restos óseos de avifauna marina podría deberse a variados factores,
en primer lugar: según los estudios realizados por Cruz (1999, 2000, 2007, 2009, 2011)
en Patagonia, durante el período de nidificación y cría de aves marinas, en las costas son
depositados numerosos restos, ya que en estas instancias los índices de mortalidad se
elevan. De este modo las probabilidades de contaminación de depósitos arqueológicos
por la muerte natural de avifauna marina son muy altas. En segundo lugar, esta alta
mortalidad constituye una importante fuente de alimento disponible para carnívoros
locales (Cruz 2011:162) los cuales al transportar de los restos participan en los procesos
de transformación de sitios arqueológicos (Cruz 2011). En tercer lugar, los abrigos
rocosos constituyen puntos en el paisaje que generalmente presentan altas tasas de
sedimentación o condiciones que atemperan la acción de los procesos destructivos, lo
que facilita la conservación de huesos de aves (Cruz 2011:156).
Cabe mencionar que si bien no fueron identificadas marcas dentarias de carnívoros, no
debe descartarse su participación en el origen del conjunto óseo, ya que como lo indican
Fernández y colaboradores (2010), ocasionalmente en aleros rocosos con depósitos
arqueológicos, los carnívoros incorporan unidades anatómicas sin la presencia de huellas
que permitan establecer su procedencia.
En base a los antecedentes recopilados sobre la alimentación de los vertebrados que
habitan el área (Jaramillo 2005, Iriarte 2008, Vilina y Cofré 2008b), a las altas
posibilidades de contaminación de restos faunísticos en aleros (Cruz 2011) y a la
ausencia de piezas óseas con modificaciones antrópicas. Es posible inferir que la el

109
conjunto osteofaunístico presente en la unidad alero 1 del sitio CF3, sea producto de
eventos post-depositacionales y no a una acumulación cultural. Ya que la presencia de
avifauna marina, mastofauna marina y mastofauna terrestre podría deberse tanto a
muertes naturales como al transporte por parte de carnívoros, mientras que la presencia
de ictiofauna podría deberse al transporte por parte de avifauna marina piscívora.
Abundancia anatómica:
La mayor parte de las unidades anatómicas registradas proceden del esqueleto
apendicular. Sobre la base del cálculo del MNI, fueron identificados dos individuos de
Larus dominicanus, para el resto de los taxones solo pudo establecerse la presencia de
un individuo. Cabe considerar que la función anatómica de un hueso determina sus
propiedades mecánicas, las que a su vez median sobre los efectos que los procesos
tafonómicos tendrán sobre ese hueso (Cruz 2011:152) ya que la forma y tamaño sumado
a la microestructura del tejido óseo influyen en la destrucción o preservación de los restos
de cada taxón (Behrensmeyer 1991, Lyman 1994). En base a esto es posible inferir que la
presencia de cresta supraoccipital de Trachurus murphyi y las unidades anatómicas
procedentes del esqueleto apendicular de aves voladoras se debe que constituyen un
registro con mayores índices de preservación (Falabella et al. 1995, Cruz 2011). En
consecuencia, la ausencia de partes esqueletales no siempre implica intervención
antrópica (Cruz 2011).

110
P3, Unidad 1:
Identificación taxonómica:
Los resultados del análisis y los antecedes, permiten inferir que P3 corresponde a un
conchal, no saqueado por lo que constituye un eficiente medio para la interpretación.
Lamentablemente, debido a la ausencia de indicadores clave, no fue posible discernir
lapsos de abandono de la ocupación, razón por la que la muestra procedente de los cinco
niveles excavados será interpretada como una totalidad.
En esta unidad fue registrado un fuerte predominio por parte de la avifauna marina,
seguido de ictiofauna, mastofauna terrestre y finalmente mastofauna marina. Los taxones
con mayor frecuencia resultaron ser Phalacrocorax sp. y Lama guanicoe. En menor
medida fueron registrados Cilus gilberti, Trachurus murphyi y Pelecanus thagus.
Finalmente fueron identificados escasos restos de Sula variegata, Larus dominicanus,
Spheniscus humboldti, Egretta sp. y Otariidae.
Abundancia anatómica:
En el caso de Trachurus murphyi se detectó la presencia de al menos 10 individuos. Para
Phalacrocorax sp. y Sula variegata se detectó la presencia de al menos dos individuos.
Del resto de los taxones solo pudo establecerse la presencia de al menos un individuo.
El segmento del esqueleto mayormente representado fue el apendicular, para el caso de
la ictiofauna las unidades anatómicas representadas consistieron huesos del
neurocráneo, vertebras de las zonas precaudal y caudal, no fueron registradas espinas.

111
PC8, Unidad 1:
Identificación anatómica:
El taxón mayormente representado fue Egretta sp, como ha sido mencionado
anteriormente este taxón no constituye un recurso explotado por el humano. También
fueron identificadas taxas de ictiofauna, avifauna marina, mastofauna terrestre y
mastofauna marina.
Considerando que la muestra procede del interior de una estructura de piedras es
altamente probable que este registro tenga un origen no antrópico ya que la construcción
genera un efecto similar al ocurrido en los Aleros. Los cuales según lo señalado por Cruz
(2011) poseen condiciones propicias para la acumulación natural de restos faunísticos.
Adicionalmente, esta unidad posee cerámica correspondiente al Período Intermedio
Tardío, por lo que es posible que estos recintos hayan sido re utilizados durante dicho
periodo. En consecuencia, resulta impreciso considerar este registro.
Abundancia anatómica:
Fue detectada la presencia de al menos dos individuos de Egretta sp, para el resto de los
taxones solo pudo establecerse la presencia de al menos un individuo. Las unidades
anatómicas registradas proceden principalmente del esqueleto apendicular. Cabe
recordar cómo fue señalado anteriormente la conservación diferenciada de partes
esqueletaria no implica intervención humana (Cruz 2011).

112
Chungungo 6:
Identificación taxonómica:
Esta unidad presentó escasos y fragmentados restos de mastofauna marina, mastofauna
terrestre e ictiofauna. La única especie identificada fue Trachurus murphyi. Dadas las
características de este registro y considerando lo propuesto por Lyman (1994), se torna
altamente probable que la presencia de vertebrados en esta unidad se deba a eventos
post-depositacionales producidos por acreción mecánica, al traslado por parte de
carnívoros y/o a la muerte natural de los animales (Butzer 1989, Cruz 2011).
Abundancia anatómica:
Fue detectada la presencia de al menos un individuo de Trachurus murphyi.
4.7.4. Modificaciones culturales:
CF3, Túmulo 1 y Unidad 4:
Artefacto óseo:
En el nivel 2 del túmulo 1 fue registrado un artefacto óseo identificado como un gancho de
estólica Este artefacto fue confeccionado sobre un hueso de mastofauna, se encuentra
terminado y fragmentado en tres partes, en cuanto al tratamiento de superficie, este
gancho fue alisado.
La estólica consiste en un arma propulsora que actúa como una prolongación del brazo y
tiene por finalidad portar efectividad y alcance al lanzamiento de un proyectil. Esta arma
se compone por una varilla de madera con o sin acanaladura central, en la parte proximal

113
posee un mango en el que se ubica un gancho, un asa de cuero o un agujero; mientras
que en su parte distal se ubica un proyectil (Rivera y Zlatar 1982).
Técnicamente esta arma antecede al uso del arco y flecha en el sentido que los principios
de fuerza involucrados son más sencillos, aunque menos eficientes en términos de
energía empleada. (Rivera y Zlatar 1982:14). Su uso data del periodo arcaico y se
atribuye a actividades de caza terrestre (Standen 2003).
CF3, Alero 1:
Artefacto óseo:
En el nivel 1 fue registrado un artefacto óseo identificado como un tubo para insuflar
narcóticos. Este artefacto fue confeccionado sobre diáfisis de ave, se encuentra terminado
y fragmentado en un extremo. En cuanto al tratamiento de superficie, este tubo fue
alisado y posee un embarrilado en el extremo que se conserva.
Los tubos para insuflar narcóticos consisten en cilindros finos y huecos. Confeccionados
con madera o huesos de aves y en ocasiones son decorados con figuras antropomorfas
y/o zoomorfas (Torres 1986).
El uso de alucinógenos data del Período Formativo (Muñoz I 1989) y se asocia
generalmente con una especializada parafernalia destinada a preparar, contener,
manipular e inhalar polvos psicoactivos, en la que manos de morteros, morteros,
cubiletes, espátulas, tabletas y tubos. Son al parecer, sus componentes más conspicuos
(Berenguer 1987b:33). Estos implementos empleados para la inhalación de alucinógenos
son comúnmente hallados en el desierto de atacama (Oyarzún 1931, Mostny 1956, Núñez
L 1963).

114
El principal componente de las sustancias inhaladas provenía del cebil, un árbol en el que
crecen vainas que contienen de 8 a 16 semillas, las cuales eran tostadas y pulverizadas
para su consumo (Torres 1986). El cebil es una especie endémica del noroeste argentino
y a partir del formativo constituyo un recurso de intercambio (Berenguer 1997).
P3, Unidad 1:
Marcas de procesamiento y/o consumo:
En el nivel tres fue detectada la presencia de huellas antrópicas, en un húmero de Sula
variegata y en un tarso metatarso de Phalacrocorax sp. Ambos casos corresponden a
cortes transversales, superficiales y cortos, ubicados en el proceso. Adicionalmente
fueron identificadas marcas en una falange, un astrágalo y un calcáneo de Lama
guanicoe. Estas unidades anatómicas fueron halladas asociadas in situ. Las huellas
consisten en cortes superficiales de orientación transversal. Finalmente en el nivel cinco
fue detectada la presencia de huellas de corte en el proceso de una escapula y en la
diáfisis de un hueso largo indeterminado, ambos correspondientes a Lama guanicoe. La
escapula presentó cortes transversales, superficiales y cortos, de distribución aislada y
frecuencia única, mientras que el hueso largo indeterminado presentó cortes
transversales, superficiales y cortos, numerosos y concentrados en varios grupos.
En base a lo descrito por De Nigris (1994) es posible inferir que dichos cortes fueron
hechos con la intención de desarticular las carcasas para generar porciones menores,
apropiadas para su consumo. Cabe mencionar que solo fueron recuperadas dos piezas
óseas con termoalteraciones por lo cual no es posible aproximarnos a los patrones
culinarios propuestos por De Nigris (1994) y Davis (1989).

115
PC8, Unidad 1:
Marcas de procesamiento y/o consumo:
Fue registrado un radio de avifauna indeterminada, con pequeños cortes aislados de
orientación transversal y baja profundidad, ubicados desde la epífisis proximal hasta la
diáfisis. Tomando en consideración las descripciones de De Nigris (1994) estas incisiones
fueron hechas durante la preparación del animal con la intención de reducir las porciones
para facilitar su consumo.
Si bien anteriormente se señaló que debido a las características de esta unidad resulta
impreciso considerar el conjunto osteofaunístico, la unidad anatómica previamente
descrita si será considerada, ya que presentas huellas de tipo antrópicas, adicionalmente,
fue hallada en asociación a cerámica Loa café alisada y constituye una evidencia
equivalente a las registradas en el sitio P3.
Chungungo 6, Alero 1:
No fueron registradas modificaciones culturales.

116
CAPÍTULO 5
5. Discusión y conclusiones:
Sitio Chungungo 6:
Los restos osteofaunísticos recuperados en el sitio Chungungo 6 no cuentan con el
potencial informativo capaz de contribuir a la comprensión de la utilización de vertebrados
durante el Período Formativo en Punta Tames; si consideramos la propuesta de Lyman
(1994) y las características del conjunto osteofaunístico recuperado en cuanto a la función
de sitio, a lo efímero de la muestra y a su historia tafonómica, se torna altamente probable
que la presencia de vertebrados se deba a eventos post-depositacionales. Si bien las
causas no lograron ser determinadas, estas pueden corresponder a la acreción mecánica,
el traslado de carcasas por parte de carnívoros y/o la muerte natural de los animales
(Butzer 1989, Cruz 2011). En consecuencia, el sitio Chungungo 6 corresponde a un alero
rocoso ubicado a 126 metros de la línea de costa en el que fueron desarrolladas
actividades de desconche durante el Formativo medio (Varas 2012, Moreno 2013).
Por otro lado, el conjunto de sitios constituido por CF3 -a excepción de Alero 1-, P3 y PC8
contaron con el potencial informativo capaz de aportar a la comprensión sobre la
utilización de vertebrados durante el formativo tardío en Punta Tames. Cabe mencionar
que si bien Alero 1 se encuentra emplazado al interior del sitio CF3, presenta fechas más
tardías y un patrón de entierro sumamente distinto al registrado en el resto del sitio,
motivo por lo que esta unidad será interpretada independientemente.

117
Sitios CF3, P3 y PC8:
CF3:
El sitio CF3 consiste en un cementerio de túmulos con un emplazamiento de
aproximadamente 5000 m2 (Castro V 2011), en el que fueron exhumados restos
bioantropológicos; materiales empleados para la cobertura de los cuerpos; variados
ajuares/ofertorios y material de relleno (Castro V 2011, Constanzo 2011, Peña-Villalobos y
Fibla 2011, Varela 2011).
Debido a que el sitio CF3 se encuentra saqueado, no es posible establecer un patrón de
entierro. Sin embargo, este cementerio es análogo al sitio Cobija 10 descrito por Moragas
(1982), puesto que ambos han sido asignados al Período Formativo, se encuentran
emplazados en el área de estudio y comparten prácticamente el mismo registro material.
El sitio Cobija 10 fue interpretado como la expresión más meridional de la Fase Alto
Ramírez y consiste en un extenso cementerio de túmulos con un complejo patrón de
entierro (Moragas 1982). Si consideramos que CF3 posee un patrón de entierro similar al
del sitio Cobija 10, es posible relacionar a los grupos litoraleños de Punta Tames con la
tradición fúnebre de la Fase Alto Ramírez desarrollada en Arica durante el Período
Formativo.
Moragas (1982) no hace mención con respecto de la presencia de restos osteofaunísticos
en Cobija 10 -a excepción de artefactos óseos-. Este antecedente sumado a la historia
tafonómica del sitio, nos indica que la presencia de osteofauna en CF3 se debe a eventos
post-depositacionales posiblemente originados por el saqueo.

118
La única evidencia de utilización de vertebrados presente en el cementerio de túmulos de
CF3 consiste en un gancho de estólica, el cual debió ser depositado en calidad de
ofrenda. La presencia de dicho artefacto indica la realización de caza terrestre (Rivera y
Zlatar 1982, Standen 2003), según Núñez P (2003) la estólica era empleada para la caza
de mastofauna -principalmente Lama guanicoe (guanaco)- y avifauna, lo cual concuerda
con la muestra de osteofauna procedente del sitio P3, ya que en dicho sitio fueron
registrados fragmentos óseos de Lama guanicoe (guanaco) y diversas taxas de aves.
P3:
El sitio P3 consiste en un basural en el que fue registrada la presencia de malacofauna,
osteofauna, ceniza, carbón, artefactos descartados y residuos de su confección. Sobre la
base de las definiciones de Orquera y Piana (1992) es posible afirmar que dicho registro
corresponde a un conchal, los cuales consisten en basurales costeros en los que son
acumulados desechos, principalmente residuos de alimentación humana (Butzer 1989,
Orquera y Piana 1992, Núñez P 2003). Por lo que la presencia de restos osteofaunísticos
en estos contextos revela conductas alimenticias en cuanto a la explotación de
vertebrados.
Desde esta perspectiva y considerando que el sitio no se encuentra disturbado, la
muestra de osteofauna procedente de la unidad 1 del sitio P3, nos aproximará a la
utilización de vertebrados por parte de los habitantes de Punta Tames durante el
Formativo Tardío.
La muestra de osteofauna estuvo compuesta por 173 piezas óseas en base a las que
fueron identificadas seis taxa de avifauna, dos taxa de ictiofauna, una taxa de mastofauna
terrestre y una taxa de mastofauna marina.

119
Entre el registro aviar, fue identificado Pelecanus thagus (pelicano), Phalacrocorax sp.
(cormorán), Spheniscus humboldti (pingüino de Humboldt), Sula variegata (piquero),
Larus dominicanus (gaviota Dominicana), y Egretta sp (garza). Estas taxas corresponden
a aves guaneras con antecedentes de consumo por parte de humanos –a excepción de
Egretta sp. (garza)- (Olguín 2011, Olguín et al. 2012, Peña-Villalobos et al. 2013). Los
taxones de avifauna con mayor frecuencia resultaron ser Phalacrocorax sp. (cormorán) y
Sula variegata (piquero).
En cuanto a la presencia de Phalacrocorax sp. (cormorán), es posible que la mayor parte
de las unidades anatómicas asignadas a esta familia correspondan a la especie
Phalacrocorax brasilianus (yeco), dado que posee una característica etológica que la
convierte en un recurso de fácil obtención.
Phalacrocorax brasilianus (yeco) es una especie piscívora que obtiene su alimento a
través del buceo, el que se ve facilitado por su plumaje permeable. Sin embargo, al salir
del agua debe secar sus plumas con el sol y el viento, adoptando una postura
característica en forma de „M‟ la que mantiene por prolongados períodos de tiempo (Araya
y Millie 1992, Jaramillo 2005; Ver anexo 6). Dicha conducta pudo aumentar la
vulnerabilidad de esta especie, permitiendo capturas sorpresivas por la espalda,
convirtiéndola en un recurso de fácil obtención. Dicha característica etológica sumado a
los altos índices de masa utilizable esta especie (Smith 2011 en Peña-Villalobos et al.
2013), pudieron convertir al Phalacrocorax brasilianus (yeco) en el Phalacrocoracidae
(cormorán) mayormente consumido, dado que constituye una óptima fuente de alimento,
siendo un recurso que proporcionaba mayor energía que la invertida durante el proceso
de obtención.

120
La especie Sula variegata (piquero), posee una conducta etológica que podría convertirla
en un recurso de fácil obtención. Esta característica radica en su particular forma de
alimentación, en la captura ictiofauna precipitándose en picada hacia el agua. Dicha
estrategia es realizada desde 40 metros aproximadamente, de este modo logra
sumergirse hasta 8 metros de profundidad, en cada viaje de alimentación los piqueros
realizan hasta 68 inversiones (Stucchi 2011: Ver anexo 6). Esta característica etológica,
pudo permitir que durante los viajes de alimentación los individuos impactaran en
acantilados o islotes, permitiendo el aprovechamiento oportunista mediante la recolección
de ejemplares, siendo un recurso ocasional cuyo aporte energético es superior al gasto
invertido en su obtención. Cabe mencionar que estas muertes naturales pueden contribuir
a la formación post-depositacional de sitios arqueológicos.
En cuanto a la presencia de Pelecanus thagus (pelicano), su explotación podría deberse a
que constituye una especie que proporciona diversas materias primas, tales como diáfisis
de huesos largos para la confección de copunas (Niemeyer 1965-66, De Vivar 1952) y
plumas para la confesión de diademas (Horta 2000). Adicionalmente, posee un alto grado
de masa total utilizable (Smith 2011 en Peña-Villalobos et al. 2013). Dichas características
convierten a Pelecanus thagus (pelicano) en un recurso con alto potencial utilitario, lo cual
debió incentivar su explotación.
La especie Spheniscus humboldti (pingüino de Humboldt) posee una característica
etológica que la convierte en un recurso de fácil obtención. Dado que constituye un ave no
voladora que durante el período de nidificación y cría pasa prolongados períodos de
tiempo en la superficie terrestre, dichas características convierten a Spheniscus humboldti
(pingüino de Humboldt) en un ave cuya obtención no requiere de mayores dificultades.
Adicionalmente, esta especie posee un alto rendimiento económico, por lo que debió

121
constituir una óptima fuente de consumo ya que posiblemente proporcionó mayor energía
que la invertida durante el proceso de captura (Cruz 2005).
Por otra parte, Larus dominicanus (gaviota Dominicana) posee un bajo índice de masa
utilizable (Smith 2011 en Peña-Villalobos et al. 2013). Esta especie corresponde a la
familia Laridae (gaviotas), la cual no constituye un recurso rentable, sin embargo según
Peña-Villalobos y colaboradores (2013) los láridos pudieron ser explotados en forma
oportunista o en contextos específicos, como lo son la escases de recursos de mayor
rentabilidad.
Finalmente, fueron identificadas unidades anatómicas pertenecientes al género Egretta
sp. (garza), debido a que este género presenta bajos índices de masa utilizable, no
constituyó un recurso de interés. Sin embargo, no debe ser descartado su
aprovechamiento oportunista o en épocas de estrés.
Con respeto a la obtención del recurso aviar, este era adquirido mediante la caza
terrestre, en la que eran empleados instrumentos tales como redes; trampas; boleadoras
y estólicas, entre otros (Núñez P 2003, Standen 2003). Estos taxones –a excepción de
Egretta sp. (garza)- nidifican en acantilados o islotes, en colonias relativamente densas
(Jaramillo 2005), lo cual pudo facilitar su captura en épocas de cría. Cabe considerar que
debido a su tamaño relativamente grande, las aves guaneras pudieron ser una apreciada
fuente alimenticia que promovió la diversificación de la dieta humana (Peña-Villalobos et
al. 2013:302).
Recapitulando, los taxones Phalacrocorax sp. (cormorán), Sula variegata (piquero) y
Spheniscus humboldti (pingüino de Humboldt) constituyen una óptima fuente de consumo,
ya que proporcionan mayor energía que la invertida durante el proceso de obtención.

122
Pelecanus thagus (pelicano) constituyó un recurso altamente valioso debido a su alto
potencial utilitario. Finalmente, Larus dominicanus (gaviota Dominicana) y Egretta sp.
(garza) pudieron ser explotados en forma oportunista o como recursos de emergencia en
épocas de estrés.
En lo respecta a la ictiofauna, fueron registradas las especies Trachurus murphyi (jurel) y
Cilus gilberti (corvina), ambas taxa constituyeron parte de la alimentación de los grupos
litoraleños (Núñez P 2003), así como también un recurso intercambiable con grupos del
interior (Mostny 1952, Castro et al. 2010, Castillo 2011, Núñez y Santoro 2011, Pimentel
et al. 2011).
El cálculo del MNI de ictiofauna, Trachurus murphyi (jurel) constituye la taxa con mayor
frecuencia -al menos 10 individuos-. Según estudios en la actualidad, esta especie posee
una longitud de 65 cm, con un peso de 2,5 kilogramos aproximadamente, de lo cual cerca
del 55% es comestible (Vargas et al. 1993). Trachurus murphyi (jurel) habita en el sistema
pelágico entre las regiones nerítica y oceánica (Moreno y Castillo 1976), es obtenido
desde la orilla o desde el sector intermedio mediante la navegación y para su captura eran
empleados redes, anzuelos, tridentes o arpones (Núñez P 2003). La época del año más
apropiada para la obtención de este recurso es a fines del otoño e inicios del invierno,
dado que esta especie se aproxima a la costa en grandes cardúmenes en busca de
alimento, permitiendo que su obtención sea realizada principalmente desde la orilla
(Vargas et al. 1993).
La especie Cilus gilberti (corvina), habita en el sublitoral superior (Moreno y Castillo 1976)
y es obtenida desde la orilla mediante el uso de redes o anzuelos (Núñez P 2003). No
contamos con mayores antecedentes respecto su la etología y masa total utilizable. Sin

123
embargo, estamos al tanto de que esta especie posee una talla equivalente –y mayor- a la
de Trachurus murphyi (jurel), por lo que es posible inferir que ambas especies poseen un
grado de masa total utilizable similar, es decir de 55%.
En cuanto a las unidades anatómicas recuperadas, ambas especies estuvieron
representadas por piezas óseas procedentes del segmento craneal y vertebras, tanto
caudales como precaudales. En base a dicho registro es posible inferir que en el conchal
fueron depositados los desechos de preparación y consumo de Trachurus murphyi (jurel)
y Cilus gilberti (corvina). No obstante, debe tenerse en consideración la posibilidad de que
la ausencia de espinas sea producto de la conservación diferencial.
En consecuencia, el recurso ictio estuvo representado por las especies Trachurus murphyi
(jurel) y Cilus gilberti (corvina). Ambos taxones eran obtenidos principalmente desde la
orilla y constituían una óptima fuente de consumo y de intercambio con grupos del interior.
Por otro lado fue registrada mastofauna terrestre, la que estuvo representada por la
familia camelidae (camélidos), si bien no fue posible dar asignación taxonómica a nivel de
especie, es probable que las unidades anatómicas asignadas a esta familia correspondan
a Lama guanicoe (guanaco) esto es respaldado por su endemismo (Iriarte 2008) y
registros de consumo de esta especie (Núñez P 2003).
Las unidades anatómicas de Lama guanicoe (guanaco) presentes en el sitio
corresponden a un individuo y proceden del esqueleto apendicular -falanges primeras,
astrágalo, calcáneo, radioulna y escapula-. Frente a este tipo de registro vale considerar
el modelo de Otala (2010) en el que señala que los esqueletos de mamíferos de gran
tamaño poseen unidades anatómicas de alto potencial de supervivencia entre las que se
encuentran huesos largos, mandíbula, astrágalo, calcáneo, falanges primeras y algunos

124
huesos del cráneo tales como dientes y unidades anatómicas de bajo potencial de
supervivencia entre las que se encuentran vértebras, costillas, pelvis, escápulas,
tarsianos, carpianos y falanges segunda y tercera (Otala 2010).
Entonces, la presencia de huesos largos puede deberse a la conservación diferencial,
pero de ser así se habrían conservado otros elementos de alto potencial de supervivencia.
Por lo que estamos frente a una toma de decisiones con transporte selectivo. Durante el
trozamiento primario rara vez se separan aisladamente cada uno de los huesos que
integran las extremidades (Mengoni 1991:184), es por esto que en ocasiones las
unidades anatómicas de bajo rendimiento ingresan articuladas a unidades de mayor
rendimiento. A juzgar por lo anterior es posible inferir que la presencia de Lama guanicoe
(guanaco) en este sitio se deba al descarte de segmentos anatómicos con bajos índices
de utilidad.
Lama guanicoe (guanaco) posee varios recursos explotables para el beneficio humano:
carne, huesos, cuero y pelaje (Núñez L 1989). Esta especie era cazada mediante el
empleo de waracas, boleadoras, arcos y estólicas, entre otros (Núñez P 2003, Standen
2003). Cabe considerar que esta especie es el mamífero herbívoro endémico más grande
del área (Bonavia 1996), por lo que pudo constituir un recurso eficiente; ya que al ser de
gran tamaño, debió otorgar altos niveles de masa utilizable y al ser herbívoro suponemos
que no debió representar una amenaza para el humano al momento de la caza.
Finalmente, la mastofauna marina identificada estuvo representada por la familia Otariidae
(lobos marinos). Si bien no se logró dar asignación taxonómica a nivel de especie, es
posible que las unidades anatómicas analizadas correspondan a Otaria flavescens (lobo
marino de un pelo), ya que es el otárido más común en el área (Aguayo y Maturana

125
1973). Adicionalmente, el uso de cuero, tripas, sangre y el consumo de carne se atribuye
a esta especie (Niemeyer 1965-66, Llagostera 1982).
Los cueros y tripas eran empleados para la confección de balsas, las cuales constituyeron
un eficaz medio de transporte entre caletas que permitió la explotación de guano en islas,
además de la pesca y el arponeo en alta mar (Núñez L 1986). Por otro lado, la carne y la
sangre eran consumidas por su alto nivel proteico, constituyendo una de las principales
fuentes de proteínas presente en la costa arreica (Lizarraga 1968, Llagostera 1982).
Los otáridos (lobos marinos) eran obtenidos de diversas formas, los adultos eran cazados
mediante el uso de arpones y palos, mientras que los juveniles eran recolectados
(Llagostera 1982, Núñez P 2003). Según escritos de cronistas …en los días que no hace
aire andan los lobos marinos descuidados durmiendo, y llegan seguros los indios con sus
balsas. Tíranle un harpón de cobre y por la herida se desangran y muere (De Vivar
1987:10).
En cuanto a las marcas de procesamiento y/o consumo registradas en el sitio P3, fueron
detectadas huellas de cortes en unidades anatómicas procedentes del esqueleto
apendicular de Sula variegata (piquero), Phalacrocorax sp. (cormorán) y Lama guanicoe
(guanaco). Sobre la base de las descripciones de Butzer (1989), De Nigris (1994) y
Mengoni (2006-2010) estas unidades anatómicas habrían sido el resultado del
trozamiento secundario. Lo anterior, sumado a la presencia de unidades anatómica del
esqueleto apendicular, los altos índices de fragmentación, la gran diversidad de especies -
en comparación al resto de los sitios que componen la muestra- y la ocasional presencia
de articulaciones, permiten inferir que el conchal presente en el sitio P3 corresponde a un
área de procesamiento y descuartizamiento. Cabe mencionar que solo fueron

126
recuperadas dos piezas óseas con termoalteraciones por lo que no fue posible establecer
patrones culinarios relacionados con la cocción, sin embargo, no debe descartase la
posibilidad de que la carne haya sido cocinada por métodos que no generan alteraciones
térmicas, como por ejemplo en caldos (De Nigris 1994).
La presencia de vertebrados con bajos índices de masa total utilizable no siempre debe
ser asumida como eventos post-depositacionales o aprovechamientos alimenticios
inusuales. Ya que es posible que estas presas constituyan eficientes fuentes de materias
primas, tales como huesos, cueros y plumas.
PC8:
El sitio PC8 consiste en un emplazamiento con la presencia de seis recintos
habitacionales, la muestra procede del interior de uno de estos recintos. Según los
resultados del análisis, este registro posee un origen tanto de formación artificial, como
natural.
Las evidencias antrópicas radican en la presencia de una pieza ósea correspondiente a
un hueso largo de avifauna de taxa indeterminada con huellas corte. Según las
descripciones de De Nigris (1994), estas marcas posiblemente fueron hechas durante la
preparación del animal con la intención de reducir las porciones y facilitar su consumo.
Es posible que la ausencia de modificaciones culturales en el resto de la muestra se deba
a que proviene del interior de una estructura, dicho contexto posee características
similares a las de un alero, los cuales según Cruz (2011) otorgan condiciones propicias
para la acumulación de restos faunísticos.

127
En consecuencia, es posible que tras el abandono de estos recintos habitacionales,
además de la reocupación por parte de grupos del Periodo Intermedio Tardío -esto es
abalado por la presencia de fragmentos de cerámica asignado a dicho período-, se hayan
originado una serie de eventos post-depositacionales producto de causas naturales. Ya
que la presencia de avifauna marina, mastofauna marina y mastofauna terrestre podría
deberse tanto a muertes naturales como al transporte por parte de carnívoros, mientras
que la presencia de ictiofauna podría deberse al transporte por parte de avifauna marina
piscívora.
Sitio CF3, Alero 1:
El Alero 1 del sitio CF3 consiste en un reparo rocoso con la presencia de un contexto
fúnebre en su interior (Aldunate 2011), en el que fue inhumado el cuerpo de una mujer de
aproximadamente 30 años (Andrade Com. Pers. 2011). Los resultados obtenidos a partir
de los análisis de isotopos estables, revelaron un predominio del consumo de dieta marina
por sobre otros tipo de recursos (Castro V Com. Pers. 2013). Los restos bioantropológicos
se encuentran asociados a variados componentes propios del modo de vida litoraleño -
cordelería, anzuelo de cobre, trozos de cuero de Otariidae (lobo marino), pigmento rojo,
líticos y moluscos- (Moragas 1982, Llagostera 1982, 1989, Standen 2003, Berenguer
2009, Castro V 2009) y componentes propios del interior -cerámica Loa café alisada y un
tubo óseo para insuflar narcóticos- (Oyarzún 1931, Mostny 1952, Núñez L 1963, Torres
1986, Muñoz I 1989, Berenguer 1997, Núñez y Santoro 2011).
Si bien en términos espaciales este alero se encuentra asociado a un cementerio de
túmulos, hasta la fecha no contamos con la presencia de sitios con cronología ni patrones
de entierro equivalentes. Por lo que es posible inferir que el Alero 1 de CF3 consiste en un

128
entierro aislado y fortuito -En el que fue inhumado el cuerpo de un integrante de un grupo
litoraleño de mediana o alta movilidad que se encontraba de paso por el sector.
En cuanto al registro osteofaunístico procedente del alero, las evidencias dan cuenta de
un conjunto originado de forma natural. Según lo describe Cruz (2011), los aleros
constituyen lugares propicios para la concentración de restos faunísticos, esto sumado a
la ausencia de huellas de procesamiento y/o consumo, permite inferir que las unidades
anatómicas correspondientes a avifauna marina, mastofauna marina y mastofauna
terrestre fueron ingresadas al alero por carnívoros o por acreción mecánica. Por su parte
los restos de ictiofauna pudieron ser transportados al alero por avifauna marina piscívora.
La única evidencia de utilización de vertebrados en el alero consiste en un tubo óseo
para insuflar narcóticos, este artefacto fue manufacturado sobre diáfisis de ave, la cual fue
alisada y embarrilada en uno de sus extremos. Dicha registro podría interpretarse como
una evidencia de contacto entre grupos litoraleños y poblaciones de tierras altas o del
litoral árido de valles y quebradas (Oyarzún 1931, Mostny 1952, Núñez L 1963, Torres
1986, Muñoz I 1989, Berenguer 1997, Núñez y Santoro 2011). El complejo inhalatorio
debió ser un componente cultural que mediante la interacción con poblaciones lejanas, los
grupos litoraleños adoptaron y emplearon.
Los grupos litoraleños del Formativo Tardío en Punta Tames
Los antecedentes proporcionados por los sitios CF3 -a excepción de Alero 1-, P3 y PC8
revelan que a inicios del Período Formativo Tardío, la localidad de Punta Tames fue
habitada por grupos litoraleños. Los cuales delimitaron y ocuparon el espacio según la
realización de actividades específicas, siendo CF3 un cementerio, P3 un conchal y PC8
un área habitacional.

129
Figura 38: Distancia entre los sitios CF3, P3 y PC8. Tomado y modificado de Moreno
(2013).
Las distancias entre los sitios CF3, P3 y PC8, permiten inferir que por motivos sanitaros,
el área habitacional fue instalada al margen de las áreas fúnebre y basural, las cuales se
encuentran entre 158 m y 175 m al noreste del área habitacional, a poca distancia entre sí
-35 m-.
En consecuencia, el sitio CF3 constituyó un cementerio de túmulos con influencias
procedentes de la fase alto Ramírez de Arica o de Cobija 10, el cual según Moragas
(1982) ya habría recibido esta influencia. Por su parte el sitio PC8 constituyó un área de
carácter habitacional en la que debieron ser realizadas actividades cotidianas, tales como
la preparación y el consumo de alimentos. Finalmente, el sitio P3 funcionó como un
conchal en el que fueron acumulados residuos de alimentación y artefactos descartados.

130
Por otro lado, el registro procedente del el alero ubicado al costado del sitio CF3 consiste
en entierro aislado con la presencia de elementos del interior a fines del Período
Formativo Tardío. Estas evidencias permiten inferir que se trata de una inhumación
fortuita de un grupo litoraleño que se encontraba de paso por el área, del cual la única
evidencia de utilización de vertebrados es un tubo óseo para insuflar narcóticos.
Utilización de vertebrados durante el Período Formativo Tardío en Punta Tames:
La costa arreica posee escasos recursos terrestres, sin embargo, el mar constituye una
fuente de abundante biomasa que permite la explotación de recursos bióticos (Núñez y
Varela 1969, López 1982, Llagostera 1982, 1989, Carvallo 1990, Núñez P 2003, Núñez y
Santoro 2011). Entre ellos se encuentra una gran variedad de vertebrados, de los cuales
algunos taxones fueron explotados por los grupos litoraleños para la confección de
artefactos óseos y el consumo alimentario (Llagostera 1982, 1989, 1990, Núñez P 2003,
Olguín 2011, Santander 2011, Olguín et al. 2012, Peña-Villalobos et al. 2013).
Respecto al uso de huesos como materia primera, se identificó la presencia de un gancho
de estólica y un tubo para insuflar narcóticos, ambos en contextos fúnebres. La estólica
constituyó un arma propulsora utilizada para la caza de guanacos y aves. Por otro lado el
tubo para insuflar narcóticos constituye un elemento ritual de influencia procedente de las
tierras altas. En base a lo anterior, es posible inferir que los artefactos óseos elaborados
por los grupos litoraleños de Punta Tames eran empleados para la obtención de recursos
alimentarios y para la eficacia simbólica.
Eiroa y colaboradores (1999) define industria ósea como un conjunto de utensilios. Dado
que solo contamos con la presencia de dos artefactos óseos, no es posible referirnos a
una industria ósea en Punta Tames. Sin embargo, si consideramos los abundantes

131
antecedentes sobre artefactos óseos en la costa arreica (Llagostera 1989, Santander
2011), resulta difícil asumir la escasa presencia de este registro en la muestra. Dicha
ausencia podría deberse a su extracción durante excavaciones ilegales, ya que los
artefactos óseos eran principalmente depositados como ofertorios, contextos los cuales
poseen un mayor potencial para ser saqueados.
En consecuencia, según la muestra analizada no es posible establecer que en Punta
Tames se desarrolló una industria ósea, sin embargo este resultado puede estar sujeto a
modificación.
En cuanto al consumo de vertebrados, fueron identificadas huellas de procesamiento y
consumo en Lama guanicoe (guanaco), Sula variegata (piquero) y Phalacrocorax sp.
(cormorán). Sin embargo, a pesar de la ausencia de marcas antrópicas en otras taxas, las
características del registro permite inferir que se trata de una formación artificial, por lo
cual toda la muestra ha sido considerada como evidencia de utilización de vertebrados.
De acuerdo a lo anterior, es posible inferir que los grupos litoraleños de Punta Tames
explotaron vertebrados endémicos, la caza estaba focalizada sobre individuos adultos y
especies silvestres.
Los cálculos de abundancia taxonómica y anatómica indican que en orden decreciente fue
consumido Trachurus Murphyi (jurel), familia Phalacrocoracidae, especialmente la especie
Phalacrocorax brasilianus (yeco) y Sula variegata (piquero). En menor frecuencia fueron
consumidos Pelecanus thagus (pelicano), Lama guanicoe (guanaco), Cilus gilberti
(corvina), Larus dominicanus (gaviota Dominicana), otáridos, especialmente Otaria
flavescens (lobo marino de un pelo), Spheniscus humboldti (pingüino de Humboldt) y
ardeidas del género Egretta.

132
Recapitulando, la alimentación de estos grupos se constituyó principalmente de recursos
marinos, tales como aves guaneras, peces del sistema pelágico y otáridos. Estos recursos
eran complementados con el consumo de invertebrados marinos (Castro P 2011a,
2011b). Adicionalmente fue explotado Lama guanicoe (guanaco), el cual constituye al
único recurso terrestre identificado. Considerando la propuesta de Perlman (1980) quien
señala que la adaptación costera consiste en que parte de los recursos explotados
provengan del mar, proponemos que los grupos litoraleños que habitaron Punta Tames
durante el Formativo Tardío desarrollaron adaptaciones costeras con un óptimo modo de
producción, basado en la extracción de recursos litorales.
Complementando lo anterior, la presencia de patologías en restos óseos humanos y el
material cultural procedentes de los sitios que componen la muestra, nos permiten
aproximar la forma de obtención de vertebrados por parte de los habitantes de Punta
Tames.
En cuanto a los restos bioantropológicos Constanzo (2012) identificó la presencia de
patologías producidas por la extensión y flexión de brazos y piernas las que están
asociadas al lanzamiento de elementos arrojadizos, tales como: lanzas, estólicas y
arpones, al lanzamiento de redes de pesca, a la acción de remar, a la constante posición
de estar arrodillado en actividades de molienda o marisqueo y al transporte o arrastre de
cargas pesadas. Por otro lado, fueron identificados artefactos para la caza y pesca como
lo son un anzuelo de cobre, una pesa de pesca, un gancho de estólica y tres puntas de
proyectil (Aldunate 2011, Castro V 2011). Dichas evidencias revelan que los vertebrados
fueron obtenidos a través de la caza, la pesca y en ocasiones la recolección.

133
Sobre la base de dichas evidencias, compartimos con Núñez P (2003) y Núñez y Santoro
(2011) que los grupos litoraleños que habitaron la costa arreica durante el Período
Formativo, mantuvieron un modo de vida arcaico en el que los recursos eran obtenidos
mediante la caza, pesca y recolección.
En consecuencia, según la información otorgada por el análisis osteofaunístico de los
sitios CF3, P3 y PC8. Es posible afirmar que los grupos litoraleños que habitaron Punta
Tames durante el Período Formativo Tardío desarrollaron una eficiente adaptación
costera, en la que vertebrados eran explotados con fines alimenticios, utilitarios y
simbólicos.

134
CAPÍTULO 6
6.1. Referencias citadas:
Adams. B y P. Crabtree.
2008 Comparative skeletal anatomy: A photographic atlas for medical examines
coroners, forensic anthropologist, and archaeologist. Ed. Humana Press.
Nueva York, EE. UU.
Aguayo. A y R. Maturana.
1973 Presencia del lobo común (Otaria flavescens) en el litoral chileno, Arica
(18º10‟S) a Punta Maiquillahue (39º27‟S). Biología Pesquera 6: 45-75.
Aldunate. C.
2011 Apuntes de terreno 18-28 agosto 2011. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
Aldunate. C, V. Castro y V. Varela.
2010 Los Atacamas y el pescado de Cobija en homenaje al maestro John Víctor
Murra. Chungará 42(1):341-347.
Araya. B y G. Millie.
1992 Guía de campo de las aves de Chile. Ed. Universitaria. Santiago, Chile.
Baena. J
1997 Arqueología experimental, algo más que un juego. Boletín de arqueología
experimental 1:2-5.
Becker. C.
2004 Animales que cuentan historias. Chungará 36:359-364.
Behrensmeyer. A.
1978 Taphonomic and ecologic information from bone weathering. Paleobiology
4:150-162.

135
1991 Terrestrial vertebrate accumulations. En: Taphonomy: releasing the data
locket in the fossil record, editado por P.A. Allison y D.E.G. Briggs, pp 291-
335. Ed. Plenum press. Nueva York, EE.UU.
Benavente. A, L. Adaro, P. Gecele y C. Cunazza.
1993 Contribución a la determinación de especies animales en arqueología:
familia camelidae y taruca del norte. Departamento técnico de
investigación, Universidad de Chile.
Berenguer. J.
1987a Problemas con la definición de sitio arqueológico. En: Arqueología y
Ciencias: Segundas Jornadas, editado por Museo Nacional de Historia
Natural, pp 61-80. Ed. Museo Nacional de Historia Natural. Santiago, Chile.
1987b Consumo nasal de alucinógenos en Tiwanaku: una aproximación
iconográfica. Boletín del museo chileno de arte precolombino 2:33-53.
1997 El norte grande en la prehistoria. En: Chile antes de Chile. Editado por
Museo Chileno de Arte Precolombino, pp 16-31 Ed. Museo Chileno de Arte
Precolombino. Santiago, Chile.
2004 Caravanas, Interacción y Cambio en el Desierto de Atacama. Ed. Sirawi.
Santiago, Chile.
2009 La costa, un lugar para vivir. En: Pescadores de la niebla. Editado por
Museo Chileno de Arte Precolombino, pp 23-52. Ed. Museo Chileno de Arte
Precolombino. Santiago, Chile.
Berwik. D.
1975 Valoración del análisis sistemático de los restos de fauna en los sitios
arqueológicos. Chungará 5:125-140.
Binford. L.
1964 A consideration of archaeological research design. American antiquity
29(4):225.441.
1967 Smudged pits and hide smoking. The use of analogy in Archaeological
reasoning. American Antiquity 32:1-12.
1978 Nunamiut Ethnoarchaeology. Ed. Academic Press. Nueva York, EE.UU.

136
1984 Faunal remains from Klasies River Mouth. Orlando. Ed. Academic Press.
Nueva York, EE.UU.
1988 Una odisea arqueológica. En: En busca del pasado. Ed. Crítica. Barcelona,
España.
Bittmann. B.
1980 Proyecto de investigación interdisciplinaria en la Costa Centro-Sur Andina”.
En Proyecto de Investigación Interdisciplinaria en la Costa Centro-Sur
Andina, editado por B. Bittmann, M. T. Ahumada y C. Moragas, vol. I, pp:
11- 20. Universidad del Norte, Sede Antofagasta, Departamento de
Arqueología. Imprenta Universitaria, Antofagasta.
1984 El proyecto Cobija: Investigaciones Antropológicas en la costa del desierto
de Atacama (Chile)”. Actas del XLIV Congreso Internacional de
Americanistas, pp: 99-146. San Pedro de Atacama, Chile.
Bonavia. D.
1996 Los camélidos sudamericanos: Una introducción a su estudio. Ed. Instituto
francés de estudios andinos. Lima, Perú.
Borella. F, L. Heureux, G y V. Silva.
2010 Estudio Osteométrico de Muestras Arqueológicas de individuos subadultos
de Otaria flavescens. Análisis del error Intraobserbador (EIO) en La
Arqueometría en Argentina y Latinoamérica, editado por S. Bertolino, R.
Cattaneo, A. Izeta. Ed. Universidad Nacional de Córdoba, Facultad de
Filosofía y Humanidades. Córdoba, Argentina.
Butzer. K.
1989 El contexto en Arqueología. En: Arqueología: una ecología del hombre. Ed.
Ballaterra. Barcelona, España.
Carvallo. C.
1990 Geografía de Chile: Geografía de la II Región de Antofagasta. Ed. Instituto
Geográfico Militar. Santiago, Chile.
Casassas. M.J.
1992 Carta del Factor de Potosí Juan Lozano Machuca (al virrey del Perú Don
Martín Enríquez) en que da cuenta de cosas de aquella villa y de minas de
los Lipes (año 1581). Estudios Atacameños 10:30-34.

137
Castillo. C.
2011 La aldea San Salvador y la circulación del pescado en el Formativo Medio
(500 a.C – 100 d.C) en la región de Antofagasta. Memoria para optar al
título de arqueóloga. Universidad internacional SEK, Santiago, Chile.
Manuscrito.
Castro. P.
2011a Análisis de invertebrados marinos (Moluscos, Crustáceos y Equinodermos)
Sitio Caleta el Fierro 3. Proyecto Fondecyt 1100951 „Del Periodo Formativo
al Tardío en la costa de Antofagasta. Cronología e interrelaciones con las
tierras altas‟. Manuscrito.
2011b Análisis de invertebrados marinos (Moluscos, Crustáceos y Equinodermos)
Sitios PC8 y P3. Proyecto Fondecyt 1100951 „Del Periodo Formativo al
Tardío en la costa de Antofagasta. Cronología e interrelaciones con las
tierras altas‟. Manuscrito.
Castro. V.
2009 De ídolos a santos. Evangelización y religión andina en los andes del sur.
Colección de Antropología. Fondo de publicaciones americanas
Universidad de Chile. Santiago, Chile.
2010 Informe Proyecto Fondecyt 1100951 „Del Periodo Formativo al Tardío en la
costa de Antofagasta. Cronología e interrelaciones con las tierras altas‟.
Manuscrito.
2011 Apuntes en terreno 18-28 agosto 2011. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
2012 Informe Proyecto Fondecyt 1100951 „Del Periodo Formativo al Tardío en la
costa de Antofagasta. Cronología e interrelaciones con las tierras altas‟.
Manuscrito.
Castro. V, C. Aldunate y V. Varela.
2010 Informe final Proyecto Fondecyt 1050991 “El Desierto costero y sus
vinculaciones con las tierras altas de Cobija a Calama”. Manuscrito.
Chaix. L y P. Méniel.
2006 Manual de Arqueozoología. Ed. Ariel. Barcelona, España.

138
Clarke. D.
1984 Arqueología Analítica. Ed. Bellaterra. Barcelona, España.
Cohen. A y D. Serjeantson.
1986 A manual for the identification of bird bones from archaeological sites. Ed.
Jubiulee Printers. Londres, Inglaterra.
Cohen. Y.
1974 Culture as adaptation. En: Man in Adaptation. Editado por Y. Cohen. Ed.
Aldine Publishing Company. Chicago, EEUU.
Constanzo. D.
2012 Análisis bioantropológico de los restos óseos recuperados en los sitios
Caleta del Fierro II y Caleta del Fierro III. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
Cornejo. L. y Gallardo. F.
1988 Definiendo el sitio arqueológico: Metodología en acción. Boletín de la
Sociedad Chilena de Arqueología 7:6-10.
Corrales. J.
1988-89 Fauna malacológica en asentamientos humanos de la región de Arica:
influencia conductual para el hombre primitivo. Dialogo andino 7/8:136-144.
Cruz. I.
1999 Pingüinos de Cabo Vírgenes (Santa Cruz). Aspectos tafonómicos e
implicaciones arqueológicas. En: Actas del XIII Congreso Nacional de
Arqueología Argentina, tomo 4, pp. 95-108. Córdoba, Argentina.
2000 Líneas tafonómicas y ecológicas para evaluar la explotación prehistórica de
aves acuáticas en la zona cordillerana (Prov. de Santa Cruz)”. En Desde el
país de los gigantes. Perspectivas arqueológicas en Patagonia, tomo I, pp.
202-217. Río Gallegos, Argentina.
2005 Los restos de pingüinos (Spheniscidae) de los sitios de Cabo Blanco (Santa
Cruz, Patagonia Argentina). Análisis tafonómico y perspectivas
arqueológicas. Intervenciones en Antropología 7:15-26.

139
2007 Avian Taphonomy: Observations at Two Magellanic Penguin (Spheniscus
magellanicus) Breeding Colonies and Their Implications for the Fossil
Record”. Journal of Archaeological Science 34:1252-1261.
2009 Tafonomía de huesos de cormoranes en la costa patagónica. Primeros
resultados”. En Arqueología de la Patagonia. Una mirada desde el último
confín, pp. 729-741. Ed. Utopía. Ushuaia, Argentina.
2011 Tafonomía de huesos de aves. Estado de la cuestión y perspectivas desde
el sur del neotrópico. Antípoda 13:147-174.
Cruz. I, N. Mondini y A. Muñoz.
1993-1994 Causas y azares: la encrucijada tafonómica. Shincal 4:123-130
Dauelsberg. P y L. Álvarez.
1968 Anzuelos confeccionados en hueso y en espinas de cactáceas,
procedentes de faldas del morro de Arica. Departamento de investigaciones
Arqueológicas de la Universidad del Norte, sede Arica.
Davis. S.
1989 La Arqueología de los animales. Ed. Bellaterra. Barcelona, España.
De Nigris. M.
1994 Patrones de fragmentación de huesos largos en el sitio Cerro de los Indios
1 Lago Posadas, provincia de Santa Cruz, Argentina. Tesis de licenciatura
en Ciencias Antropológicas. Universidad de Buenos Aires. Buenos Aires,
Argentina. Manuscrito.
De Vivar. G.
1987 (1558) Crónica y relación capiosa y verdadera de los reinos de Chile. Ed.
Universitaria. Santiago, Chile.
Deetz. J.
1967 Invitation to Archaeology. Ed. American Museum Science Books. Nueva
York, EE.UU.

140
Donoso-Barrios. R.
1960 Reptiles de Chile. Ed. Universidad de Chile. Santiago, Chile.
Edward. J.
2006 La domesticación de paisajes: ¿Cuáles son los componentes primarios del
Formativo?. Estudios Atacameños 32:43-57.
Efremov, I.
1940 Taphonomy: a new branch of paleontology. Pan American Geologist
74: 81-93.
Eiroa. J, J. Bachiller, L. Castro y J. Lomba.
1999 Nociones de tecnología y tipología en Prehistoria. Ed. Ariel. Barcelona,
España.
Ericson. G y J. Stora.
1999 A manual to the skeletal measurements of the seal genera Halichoerus and
Phoca (Mamamlis: Pinnipedia). Departament of Vertebrate Zoology,
Swedish Museum of Natural History. Estocolmo, Suecia.
Falabella. F, R. Méndez y L. Vargas.
1995 Claves osteológicas para peces de Chile Central: Un enfoque Arqueológico.
Ed. Artegrama. Santiago, Chile.
Fernández. P, I. Cruz y A. Forlano.
2010 Sitio 37: una madriguera de carnívoro en el Norte de la Patagonia Andina
(Cholila, Provincia de Chubut, Argentina). En Zooarqueología a principios
del siglo XXI: aportes teóricos, metodológicos y casos de estudio, editado
por: M. A. Gutiérrez, M. De Nigris, P. Fernández, M. Giardina, A. Gil, A.
Izeta, G. Neme y H. Yacobaccio, pp. 409-417. Ed. Espinillo. Buenos Aires,
Argentina.
Galotta. J y S. Márquez.
2004 Excerta anatómica camelidae 2. Ed. Fundación camélidos Sudamericanos.
Buenos aires, Argentina.

141
Gamble. C.
2008 ¿Cuántas Arqueologías existen? En Arqueología Básica. Ed. Ariel.
Barcelona, España.
Gifford. D.
1981 Taphonomy and Paleoecology: a critical review of archaeology‟s sister
disciplines. En Advances in archaeological method and theory. Academic
Press. Nueva York, EE.UU.
Grayson. D.
1984 Quantitative zooarchaeology: topics in the analysis of Archaeological
faunas. Ed. Academic Press. Orlando, EE.UU.
Hernández Sampieri. R, C. Fernández y P. Baptista.
1998 Metodología de la investigación. Ed. McGraw-Hill. D.F, México.
Hillson. S.
2005 Manual in Archaeology: Teeth. Institute of Archaeology, University College
London. Londres, Inglaterra.
House. J y M. Schiffer.
1975 Archaeological survey in the Cache River Basin. En: The Cache River
Archaeological Project: an experiment in contract Archaeology editado por
J. House y M. Schiffer. Ed. Arkansas Archaeological Survey, reserch series
8. Arkansas, EE.UU.
Horta . H.
2000 Diademas de plumas en entierros de la costa del norte de Chile:
¿evidencias de la vestimenta de una posible parcialidad pescadora?.
Chungará 32:235-243.
Iriarte. A.
2008 Mamíferos de Chile. Ed. Lynx. Santiago, Chile.
Jackson. D, C. Méndez, P. López, D. Jackson y R. Seguel.
2005 Asentamiento arqueológico en el semiárido de Chile: procesos de
formación, fauna extinta y componentes culturales. Interpretaciones en
antropología 6:139-151.

142
Jaramillo. A.
2005 Aves de Chile. Ed. Lynx. Barcelona, España
Laborde. A.
1986 Conservación y restauración en yacimientos prehispánicos. Ed. Monografic.
Barcelona, España.
Letelier. J.
2011 Clasificando Indios, un ejemplo de la construcción de la categoría Changos
durante el siglo XVI y XVII en la zona de la actual provincia del Loa. Taltalia
4:103-115
Lizarraga. R.
1968 (1605) Descripción breve del Perú. Biblioteca de autores españoles, tomo 221. Ed.
Atlas. Madrid, España
Llagostera. A.
1982 Tres dimensiones en la conquista prehistórica del mar: un aporte para el
estudio de las formaciones pescadoras de la costa sur andina. En: actas del
VII Congreso de Arqueología Chilena, pp 217-245. Ed. Kultrun. Valdivia,
Chile.
1989 Caza y pesca marítima (9.000 a 1.000 a. C.). En: Culturas de Chile.
Prehistoria. Desde sus orígenes hasta los albores de la Conquista,
editado por J. Hidalgo, V. Schiappacasse, H. Niemeyer, C. Aldunate e I.
Solimano, pp. 57-79. Ed. Andrés Bello. Santiago, Chile.
1990 La navegación prehispánica en el Norte de Chile: bioindicadores e
inferencias teóricas. Chungará 24/25:37-51.
2005 Culturas costeras precolombinas en el norte chileno: secuencia y
subsistencia de las poblaciones arcaicas. En Biodiversidad marina:
valoración, usos y perspectivas ¿hacia dónde va chile?, editado por E.
Figueroa. Ed. Universitaria. Santiago, Chile.
López. C.
1982 Geografía de Chile: biogeografía. Tomo III. Ed. Instituto geográfico militar.
Santiago, Chile.

143
López. M.
2011 Apuntes en terreno 18-28 agosto 2011. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
Lyman. L.
1987 Zooarchaeology and taphonomy: a general consideration. Journal of
Ethnobiology 7(1):93-117.
1994 Vertebrate taphonomy. Ed. Cambridge University Press. Gran Bretaña.
2008 Quantitative paleozoology. Ed. Cambridge University Press. Gran Bretaña.
Mac-Lean. R.
2012 Informe osteofauna: CF3, PC8 y P3. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
Martínez. J.L.
1985a Información sobre el comercio de pescado entre Cobija y Potosí, hecha por
el corregidos de Atacama, Don Juan de Segura (19 de Julio de 1591).
Cuadernos de Historia 5:161-171. Universidad de Chile, Santiago, Chile.
1985b Dispersión y movilidad en Atacama colonial. En: Encuentro de
Etnohistoriadores editado por: O. Silva, E. Medina y E. Téllez. Serie nuevo
mundo: Cinco siglos 1:53-69.
Méndez. M y C. Correa.
2008 Anfibios. En: Biodiversidad de Chile: patrimonio y desafíos, editado por J.
Rovira, J. Ugalde y M. Stutzin. Pp 276-283. Ed. Ocho libros. Santiago,
Chile.
Mengoni. G.
1988 Análisis de materiales faunísticos de sitios Arqueológicos. Xama N 1: 71-
120.
1991 La llama y sus productos primarios. Arqueología 1: 179-196.
1999 Cazadores de guanacos en la estepa patagónica. Ed. Sociedad Argentina
de Antropología. Buenos Aires, Argentina.

144
Moragas. C.
1982 Túmulos funerarios en la costa sur de Tocopilla (Cobija) – II Región.
Chungará 9:152-173.
Moreno. C.
2013 Aplicación de Sistemas de identificación Geográfica (SIG) en un caso de
estudio arqueológico: Cartografía y georeferenciación Herramienta para
memoria de título: “Utilización de vertebrados durante el Periodo Formativo
en Punta Tames, Costa Arreica de Antofagasta. Ms.
Moreno. C. y J.C. Castillo.
1976 Guía para el reconocimiento y observación de peces de Chile. Ed Gabriela
Mistral. Santiago, Chile.
Mostny. G.
1952 Una tumba de Chiuchiu. Boletín del Museo Nacional de Historia Natural,
XXVI (1):1-55.
Muñoz. I.
1989 El Período Formativo en el norte grande (1000 a. C. a 500 d.C.) En:
Culturas de Chile. Prehistoria. Desde sus orígenes hasta los albores de la
Conquista, editado por J. Hidalgo, V. Schiappacasse, H. Niemeyer, C.
Aldunate e I. Solimano, pp. 129-180. Editorial Andrés Bello. Santiago, Chile.
Muñoz. S.
1997 Explotación y procesamiento de ungulados en Patagonia meridional y tierra
del fuego. Anales del instituto de la Patagonia 25:201-222.
Nautilus-ecosistemas.blogspot.com
[Consult. 12.04.2011]. Disponible en:
http://nautilusecosistemas.blogspot.com/2013/04/algunas-definiciones.html
Niemeyer. H.
1965-66 Una balsa de cueros de lobo de la Caleta de Chañaral de Aceitunas (Prov.
de Atacama, Chile). Revista Universitaria N*L, Tomo 2:257-269.

145
1989 El escenario geográfico. En: Culturas de Chile. Prehistoria. Desde sus
orígenes hasta los albores de la Conquista, editado por J. Hidalgo, V.
Schiappacasse, H. Niemeyer, C. Aldunate e I. Solimano, pp. 1-12. Ed.
Andrés Bello. Santiago, Chile.
Niemeyer. H. y P. Cereceda.
1984 Ríos de régimen esporádico en la zona árida de Chile. En: Geografía de
Chile: Hidrografía, editado por E. López. Pp 29-84. Ed. Instituto Geográfico
Militar. Santiago, Chile.
Núñez. H.
2008 Reptiles. En: Biodiversidad de Chile: patrimonio y desafíos, editado por J.
Rovira, J. Ugalde y M. Stutzin. Pp 276-283. Ed. Ocho libros. Santiago,
Chile.
Núñez. L.
1963 Problemas en Torino a la tableta de rape. Anales de la universidad del
norte 2:149-168.
1976 Registro regional de fechas radio carbónicas del Norte de Chile. Estudios
Atacameños 4:69-111.
1986 Balsas prehistóricas del litoral chileno: grupos funciones y secuencia.
Boletín del museo chileno de arte precolombino 1:11-35.
1989 Hacia la producción de alimentos y la vida sedentaria (5000 a. C. a 900
d.C.) En: Culturas de Chile. Prehistoria. Desde sus orígenes hasta los
albores de la Conquista, editado por J. Hidalgo, V. Schiappacasse, H.
Niemeyer, C. Aldunate e I. Solimano, pp. 81-106. Ed. Andrés Bello.
Santiago, Chile.
Núñez. L. y J. Varela.
1967 Sobre los recursos de agua y el poblamiento prehispánico de la costa del
Norte Grande de Chile. Estudios Arqueológicos 3/4:7-41. Universidad de
Chile. Antofagasta, Chile.
Núñez. L. y T. Dillehay
1979 Movilidad giratoria, armonía social y desarrollo en los Andes Meridionales:
patrones de tráfico e interacción económica. Ed. Universidad Católica del
Norte, Antofagasta, Chile.

146
Núñez. L, P. de Souza, I. Cartagena y C. Carrasco.
2008 Quebrada Tulán: Evidencias de Interacción circumpuneña durante el
Formativo Temprano en el Sureste de la cuenca de Atacama. Producción y
Circulación prehispánicas de Bienes en el sur andino. En Colección historia
social Precolombina 2. Editado por A. Nielsen, M. Clara Rivolta, V. Seldes,
María M. Vásquez y P. Mercolli, pp 287-304. Ed. Brujas. Córdoba,
Argentina.
Núñez. L. y C. Santoro.
2011 El tránsito arcaico-formativo en la circumpuna y valles occidentales del
centro sur andino hacia los cambios „neolíticos‟. Chungará 43:487-530.
Núñez. P.
2003 Vivir el mar. Ed. Universidad de Antofagasta. Antofagasta, Chile
Olguín. L.
2011 Informe arqueofauna: Guasilla 2 y Mantos de la Luna 4. Proyecto Fondecyt
1100951 „Del Periodo Formativo al Tardío en la costa de Antofagasta.
Cronología e interrelaciones con las tierras altas‟ Manuscrito.
Olguín. L, I. Peña-Villalobos, P. Fibla y M. Sallaberry.
2012 Informe arqueofauna: Copaca 1. Proyecto Fondecyt 1100951 „Del Periodo
Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟ Ms.
Orquera. L y Piana. E.
1992 Un paso hacia la revolución del palimpsesto. En análisis espacial en la
arqueología patagónica. Editado por L. Borrero y J.L. Lanata, pp 21-52. Ed.
Ayllú. Buenos Aires, Argentina.
Otala. C.
2010 Índices de utilidad económica en contextos tafonómicos variados. En:
Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y
casos de estudio. Editado por M.A. Gutiérrez, M. De Nigris, P. M.
Fernández, M. Giardina, A. Gil, A. Izeta, G. Neme y H. Yacobaccio. Pp 157-
166. Ed. Espinillo. Buenos Aires, Argentina.

147
Oyarzún. A.
1931 Las tabletas y tubos para aspirar la paricá en atacama. Revista chilena de
historia y geografía 72:68-76.
Pacheco. V, A. Altamirano y E. Guerra.
1979 Guía de camélidos sudamericanos. Ed. Universidad Nacional Mayor de San
Marcos. Lima, Perú.
Padilla. F y A. Cuesta.
2003 Zoología aplicada. Ed. Díaz de Santos. Madrid, España.
Peña-Villalobos. I. y P. Fibla.
2011 Identificación de plumas de diversos sitios arqueológicos, a través de la
comparación de microestructuras. Proyecto Fondecyt 1100951 „Del Periodo
Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟. Manuscrito.
Peña-Villalobos. I, L. Olguín, P. Fibla, V. Castro y M. Sallaberry.
2013 Aprovechamiento humano de aves marinas durante el Holoceno medio en
el litoral árido del norte de Chile. Revista chilena de Historia Natural 86:301-
313.
Pimentel G, C. Rees, P. De Souza y L. Arancibia.
2011 Viajeros costeros y caravaneros. Dos estrategias de movilidad en el
Período Formativo del desierto de atacama, Chile. En: En Ruta:
arqueología, historia y etnografía del trafico sur andino, editado por L.
Núñez y A. Nielsen, pp 43-82. Ed. Brujas. Córdoba, Argentina.
Perlman. S.
1980 An optimum diet model, coastal variability, and hunter-gatherer behavior.
En: Advances in archeological method and theory 3, editado por M. Shiffer,
pp 257-310. Academic Press. Nueva York, EE.UU.
Politis. G.
2010 Aplicaciones de la Etnoarqueología para interpretar el registro. En:
arqueología, etnología em etno-historia en Iberoamérica: cosmología,
antropología em aplicação, editado por E.C. Moretti. Pp 275-328 Ed. UFGD.
Dourados, Brasil.

148
Reiz. E. y E. Wing.
1999 Zooarchaeology. Ed. Cambridge University Press. Oxford, Inglaterra.
Renfrew. C. y P. Bahn.
1998 Arqueología: teorías, métodos y práctica. Ed Akal. Madrid, España.
Rivera. M. y V. Zlatar.
1982 Las estólicas en el desarrollo Cultural temprano Prehispánico del Norte de
Chile. En: Actas del IX Congreso Nacional de Arqueología Chilena pp 14-
34. La Serena, Chile.
Rostworowski. M.
1986 La región del Colesuyu. Chungará 16-17:127-135.
Santander. B.
2011 Informe de análisis de artefactos óseos de los sitios Guasilla 2 y Mantos de
la Luna 4. Proyecto Fondecyt 1100951 „Del Periodo Formativo al Tardío en
la costa de Antofagasta. Cronología e interrelaciones con las tierras altas‟
Manuscrito.
Schaedel R.
1957 Informe general sobre la expedición a la zona comprendida entre Arica y La
Serena. En Arqueología Chilena, Contribución al Estudio de la Región
Comprendida entre Arica y La Serena, editado por R. P. Schaedel, pp. 1-
42. Universidad de Santiago, Chile.
Schiffer. M.
1990 Contexto arqueológico y contexto sistémico. Boletín de Antropología
Americana 22:81-93.
1991 Los procesos de formación del registro arqueológico. Boletín de
Antropología Americana. 23:39-45.
Sielfeld. W.
1983 Mamíferos marinos de Chile. Ed. Universidad de Chile. Santiago, Chile.

149
1999 Estado del conocimiento sobre conservación y preservación de Otaria
flavescens (Shaw 1800) y Arctocephalus australis (Zimmermann 1783) en
las costas de Chile. Estud. Oceanol. 18:81-96.
Simpson. G.
1961 Principles of animal Taxonomy. Ed. Columbia University Press. Nueva York,
EEUU.
Standen. V.
2003 Bienes funerarios del cementerio Chinchorro Morro 1: descripción, análisis
e interpretación. Chungará 35:2 175-207.
Stucchi. M.
2011 Atlas osteológico del piquero peruano sula variegata. Ed. Asociación para
la investigación y conservación de la biodiversidad. Lima, Perú.
Tamayo. M.
2003 El proceso de la investigación científica. Ed. Limusa. D.F, México.
Torres. C.
1986 Tabletas para alucinógenos en Sudamérica: tipología, distribución y rutas
de difusión. Boletín del Museo Chileno de Arte Precolombino. 1:37-53.
Uribe. M, L. Sanhueza y F. Bahamondes.
2007 La cerámica prehispánica tardía de Tarapacá, sus valles interiores y costa
desértica, Norte de Chile (CA. 900-1450 d.C.): una propuesta tipológica y
cronológica. Chungará 39(2):143-170.
Varas. V.
2011 Análisis de restos vegetales presentes en los sitios P3, PC8 y Caleta el
Fierro 3. Proyecto Fondecyt 1100951 „Del Periodo Formativo al Tardío en la
costa de Antofagasta. Cronología e interrelaciones con las tierras altas‟
Manuscrito.
2012 Apuntes en terreno 12-24 agosto 2012. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟ Manuscrito.

150
Varela. V.
2011 Análisis cerámico sitios CF3 y PC8. Proyecto Fondecyt 1100951 „Del
Periodo Formativo al Tardío en la costa de Antofagasta. Cronología e
interrelaciones con las tierras altas‟ Manuscrito.
Vargas M. L, F. Falabella y R. Meléndez.
1993 Bases para el manejo de datos ictioarqueológicos del “jurel” (Trachurus
symmetricus AYRES, 1855) (PISCES: PERCIFORMES: CARANGIDAE).
En Actas del XII Congreso Nacional de Arqueología Chilena, pp. 355-371.
Temuco.
Velásquez. H.
2004 Método para estudiar huesos de animales en sitios arqueológicos: ventajas
y problemas. Chungará 36:349-359.
Vilina. Y y H. Cofré.
2008a Aves terrestres. En: Biodiversidad de Chile: patrimonio y desafíos editado
por J. Rovira, J. Ugalde y M. Stutzin. Pp 246-257. Ed. Ocho libros.
Santiago, Chile.
2008b Aves acuáticas continentales. En: Biodiversidad de Chile: patrimonio y
desafíos editado por J. Rovira, J. Ugalde y M. Stutzin. pp 266-275. Ed.
Ocho libros. Santiago, Chile.
Vilina. Y y C. Pizarro.
2008 Aves marinas. En: Biodiversidad de Chile: patrimonio y desafíos editado por
J. Rovira, J. Ugalde y M. Stutzin. pp 258-265. Ed. Ocho libros. Santiago,
Chile.
White. T.
1953 A method of calculating the dietary percentage of various food animals
utilized by various aboriginal peoples. American Antiquity 18(4):396-398.
Zavala. P.
2008 Chile, paraíso subacuático. Ed. Kactus. Santiago, Chile.

151
6.2. Anexos:
Anexo 1. Vertebrados endémicos de la región de Antofagasta:
Se presenta el nombre común y científico de los vertebrados endémicos de la II región de
Antofagasta, según su ubicación geográfica (litoral, estepa desértica y macizo andino).
a. Vertebrados endémicos del litoral árido de arreismo absoluto:
Herpetofauna:
Nombre común Nombre científico
Tortuga coriácea del pacifico Dermochelys coriácea coriácea
Tortuga olivácea Lepidochelys olivacea
Lagartija de las playas Tropidurus peruvianus
Salamanqueja Phyllodactylus gerrophigus
Lagarto Liolaenus chilensis
Especies de herpetofauna endémicas del litoral (Donoso-Barrios 1966, López 1982).
Ictiofauna:
Sublitoral superior:
Nombre común Nombre científico
Baunco Doydixodon laevifrons
Bilagay Cheilodactylus variegatus
Cherlo o Cherno Acanthistius pictus
Cojinoba Seriolella violácea
Pejeperro Pimelometopon maculates
Tomoyo Auchenionchus microcirrhis
Vieja negra Graus nigra
Torito Bovichtus chilensis
Rollizo Mugiloides chilensis
Apañado o Parguaya Hemilutjanus macrophtalmos
Pintaroja Schroederichthys chilensis
Loro o San Pedro Oplegnathus insignis
Lenguado de ojos chicos Paralichthys microps
Torpedo o Tembladera Discopyge tschu
Corvina Cilus gilberti
Pichihuén Menticirrhus ophicephalus
Sublitoral inferior:
Nombre común Nombre científico
Merluza común o Pescada Merluccius gayi

152
Congrio colorado Genypterus chilensis
Congrio negro Genypterus maculatus
Peje águila Myliobatis peruvianus
Bagre de mar Aphos porosus
Banquillo Prolatilus jugularis
Pequén o Raya puñete Psammobatis scobina
Cojinoba Seriolella violácea
Región nerítica :
Nombre común Nombre científico
Anchoa Engralrs ringens
Agujilla o puntofijo Scomberesox saurus
Pejerrey de mar Odontesthes regia
Cabinza Isacia conceptionis
Sierra Thyrsites atun
Reineta o Hacha Lepidotus acustralis
Jurel Trachurus murphyi
Sardina Sardinops sagax
Lisa Mugil cephalus
Región oceánica:
Nombre común Nombre científico
Albacora o Pez espada Xiphias gladius
Atún de aleta larga Thunnus alalunga
Barrilete o Cachurrea Ktsuwonus pelamis
Dorado de altamar Coryphaena hippurus
Pejezorro Alopias vulpinus
Azuelo o Tintorera Prionace glauca
Jurel Trachurus murphy
Especies de ictiofauna endémicas del sublitoral superior, sublitoral inferior, región nerítica
y región oceánica (Moreno y Castillo 1976).
Aves:
Nombre común Nombre científico
Pelicano Pelecanus thagus
Piquero Sula variegata
Lile Phalacrocorax gaimardi
Guanay Phalacrocorax bougainvillii
Yeco Phalacrocorax brasilianus
Pingüino de Humboldt Spheniscus humboldti
Garza grande Ardea alba
Garza chica Egretta thula

153
Garza boyera Bubulcus ibis
Garza azul Egretta caerulea
Petrel gigante atlántico Macronectes giganteus
Petrel moteado Daption cápense
Albatros de Buller Thalassarche bulleri sp.
Albatros de frente blanca Thalassarche cauta salvini
Albatros de ceja negra Thalassarche melanophris
Fárdela blanca de Masatierra Pterodroma defilippiana
Fárdela blanca Puffinus creaptopus
Fárdela negra Puffinus griseus
Fárdela negra grande Procellaria aequinoctialis
Fárdela de Nueva Zelanda Procellaria westlandica
Golondrina de mar Oceanites oceanicus
Golondrina de mar chica Oceanites gracilis
Golondrina de mar negra Oceanodroma markhami
Golondrina de mar de collar Oceanodroma hornbyi
Yunco Pelecanoide garnotii
Huairavo Nycticorax nycticorax
Jote de cabeza colorada Cathartes aura
Aguilucho Buteo polysoma
Traro (Carancho) Caracara plancus
Chorlo nevado Charadrius alexandrinus
Chorlo ártico Pluvialis squatarola
Pilpilén negro Haematopus ater
Pilpilén Haematopus palliatus
Zarapito Numenius phaeopus
Playero vuelvepiedras Arenaria interpres
Playero blanco Calidris alba
Salteador pomarino Stercorarius pomarinus
Salteador chico Stercorarius parasiticus
Saltador de cola larga Stercorarius longicaudus
Salteador chileno Stercorarius chilensis
Gaviota dominicana Larus dominicanus
Gaviota peruana Larus belcheri
Gaviota Larus modestus
Gaviota de Sabine Xema sabini
Gaviotín sudamericano Sterna hirundinacea
Gaviotín monja Larosterna inca
Gaviotín chico Sterna lorata
Gaviotín elegante Sterna elegans
Rayador Rynchops niger
154
Tucúquere Bubo magellanicus
Lechuza Tyto alba
Picaflor del norte Rhodopis vesper
Minero Geositta cunicularia
Churrete costero Cinclodes nigrofumosus
Tijeral Leptasthenura aegithaloides
Mero gaucho Agriornis montana
Golondrina bermeja Hirundo rustica
Chercán Troglodytes muscullus
Chincol Zonotrichia capensis
Especies de avifauna endémicas del litoral (Jaramillo 2005).
Mastofauna:
Nombre común Nombre científico
Murciélago cola de ratón Tararida brasiliensis
Murciélago orejudo mayor Histiotus macrotus
Murciélago orejudo menor Histiotus montanus
Murciélago de Atacama Myotis atacamensis
Ratón oliváceo Abrothrix olivaceus
Ratón de cola larga Oligoryzomys longicaudatus
Ratón orejudo de Darwin Phyllotis darwini
Ratón orejudo grande Phyllotis magister
Guanaco Lama Guanicoe
Taruca o Huemul del norte Hippoamelus antisensis
Puma Puma concolor
Zorro culpeo Pseudalopex culpeaus
Zorro gris o zorro chilla Lycalopex griseus
Chungungo Lontra felina
Lobo fino de dos pelos Arctocephalus australis
Lobo marino de un pelo Otaria flavescens
Ballena franca austral Eubalaena australis
Ballena sei Balaenoptera borealis
Ballena de Bryde Balaenoptera edeni
Ballena azul Balaenoptera musculus
Ballena fin Balaeroptida physalus
Ballena jorobada Megaptera novaeangliae
Delfín común costero Delphinus capensis
Delfín común Delphinus delphis
Orca pigmea Feresa attenuata
Calderón de aleta corta Globicephala macrorhyncha

155
Calderón de aleta larga Globicephala melas
Falso calderón o delfín de Risso Gramphus griseus
Delfín oscuro Lagenorhynchus obscurus
Delfín liso Lisodelphis peroni
Orca Orcinus orca
Delfín moteado Stenella attenuata
Delfín de pico largo Stenella longirostris
Delfín de diente áspero Steno bredanensis
Delfín nariz de botella Tursiops truncatus
Marsopa espinosa o marsopa negra Phocoena spinipinnis
Ballena picuda de Blainville Mesoplodon densirostris
Ballena picuda del Perú Mesoplodon peruvianus
Ballena picuda de Cuvier Ziphius cavirostris
Cachalote pigmeo Kogia breviceps
Cachalote Physeter macrocephalus
Especies de mastofauna endémicas del litoral (Iriarte 2008).
b. Vertebrados endémicos de la estepa desértica:
Herpetofauna:
Nombre común Nombre científico
Iguana Tropidurus tarapacensis
Lagartija Phrynosaura reichei
Lagarto Liolaenus chilensis
Sapo de Atacama Bufo atacamensis
Especies de herpetofauna endémicas de la estepa desértica (Donoso-Barrios 1966, López
1982).
Aves:
Nombre común Nombre científico
Zorzal Chiguanco Tordus chiguanco chiguanco
Aguilucho Buteo polysoma
Halcón perdiguero Flaco femoralis
Pequén Athene cunicularia
Minero chico Geositta marítima
Golondrina de dorso negro Pygochelidon cyanoleuca
Golondrina bermeja Hirundo rustica
Chercán Troglodytes musculus
Chincol Zonotrichia capensis
Especies de avifauna endémicas de la estepa desértica (Jaramillo 2005).

156
Mastofauna:
Nombre común Nombre científico
Ratón orejudo grande Phyllotis magister
Especies de mastofauna endémicas de la estepa desértica (Iriarte 2008).
c. Vertebrados endémicos del macizo andino:
Herpetofauna:
Nombre común Nombre científico
Sapo de Hall Telmatobius halli
Sapo de Danko Telmatobius dankoi
Sapo espinoso Bufo spinulosus
Sapo de cuatro ojos marmóreo Pleurodema marmorata
Lagarto Liolaemus chilenis
Iguana de Constanza Liolaemus constanzae
Iguana de Paulina Liolaemus paulinae
Lagarto Liolaemus alticolor walkeri
Lagarto Liolaemus pantherinus
Lagarto Liolaemus signifer
Lagarto Liolaemus multiformis
Lagarto Liolaemus ornatus
Lagarto Liolaemus nigroviridis nigroroseus
Especies de herpetofauna endémicas del macizo andino (Donoso-Barrios 1966, López
1982).
Aves:
Nombre común Nombre científico
Flamenco chileno Phoenicopterus chilensis
Parina chica Phoenicopterus jamesi
Parina grande Phoenicopterus andinus
Yeco Phalacrocorax brasilianus
Aguilucho Buteo polysoma
Cóndor Vultur gryphus
Kilua Tinamotis pentlandii
Blanquillo Podiceps occipitalis
Piuquén Chloephaga melanoptera
Pato juarjual Lophonetta specularioides
Pato jergón grande Anas geórgica
Pato jergón chico Anas foavirostris

157
Pato puna Anas puna
Pato colorado Anas cyanoptera
Pato rana de pico ancho Oxyura ferruginea
Carancho cordillerano Phalcoboenus megalopterus
Halcón perdiguero Falco femoralis
Halcón peregrino Falco peregrinus
Pidén Pardirallus sanguinolentus
Tagua andina Fulica ardesiaca
Tagua gigante Fulica gigantea
Tagua cornuda Folica cornuta
Queltehue de la puna Vallenus resplendens
Caití Recurvirostra andina
Chorlo de la puna Charadrius alticola
Chorlo de campo Oreopholus ruficollis
Chorlo cordillerano Phegornis mitchellii
Becacina Gallinago paraguaiae
Perdicita cordillerana Attagis gayi
Perdicita cojón Thinocorus orbignyianus
Perdicita Thinocorus rumicivorus
Pollito de mar tricolor Phalaropus tricolor
Gaviota andina Larus serranus
Tucúquere Bubo magellanicus
Picaflor de la puna Oreotrochilus estella
Minero Geositta cunicularia
Bandurrilla de pico recto Upucerthia ruficaudus
Churrete de alas blancas Cinclodes atacamensis
Churrete acanelado Cinclodes fuscus
Tijeral Leptasthenura aegithaloides
Canastero chico Asthenes modesta
Mero gaucho Agriornis montana
Mero de la puna Agriornis andicola
Mero de Tarapacá Agriornis microptera
Dormilona de nuca rojiza Muscisaxicola rufivertex
Dormilona chica Muscisaxicola maculirostris
Dormilona fraile Muscisaxicola flavinucha
Golondrina de dorso negro Pygochelidon cyanoleuca
Golondrina de los riscos Haplochelidon andecola
Chirihue cordillerano Sicalis uropygialis
Cometocino del norte Phrygilus atriceps
Yal Phrygilus fruticeti
Pájaro plomo Phygilus unicolor

158
Plebeyo Phrygilus plebejus
Cometocino de dorso castaño Phrygilus dorsalis
Chincol Zonotrichia capensis
Jilguero cordillerano Carduelis uropygialis
Jilguero negro Carduelis atrata
Especies de avifauna endémicas del macizo andino (Jaramillo 2005).
Mastofauna:
Nombre común Nombre científico
Yaca del norte Thylamys pallidior
Quirquincho de la puna Chaetophractus nación
Murciélago cola de ratón Tararida brasiliensis
Ratón andino Abrothrix andinus
Ratón de la puna Auliscomys sublimis
Lauchita peruana Calomys lepidus
Ratón de pie sedoso Eligmodontia puerulus
Ratón de hocico anaranjado Neotomys ebriosus
Ratón orejudo amarillento Phyllotis xanthopygus
Tuco tuco de Atacama Ctenomys fulvus
Chinchilla cordillerana Chinchilla brevicaudata
Vizcacha Lagidium viscacia
Ratón Chinchilla de cola corta Abrocoma cinérea
Vicuña Vicugna vicugna
Guanaco Lama guanicoe
Gato montés o gato pajero Leopardus colocolo
Gato montés andino Leopardus jacobita
Puma Puma concolor
Zorro culpeo Lycalopex culpeaus
Quique o hurón Galictis cuja
Especies de mastofauna endémicas del macizo andino (Iriarte 2008).

159
Anexo 2. Fichas de los sitios arqueológicos:
Se presenta información básica de los sitios arqueológicos que componen la muestra -
CF3, P3, PC8 y Chungungo 6-.
CF3:
Sitio CF3
Ubicación UTM 369292 - 7495592 datum WGS84-86 huso 19S
Función Cementerio
Descripción Sitio con un emplazamiento de 5000 m2 aprox., se encuentra altamente
disturbado producto de excavaciones ilegales.
El análisis bioantropológico de los restos humanos exhumados en este
sitio indicó la presencia de al menos 10 individuos: seis adultos jóvenes
-tres posiblemente femeninos-, dos individuos juveniles y dos infantes.
Cabe mencionar que debido a las limitaciones propias de la muestra, no
fue posible determinar la estatura de los individuos (Constanzo 2012).
Sobre la base de la identificación de enfermedades degenerativas
articulares e inserciones musculares presentes en los restos óseos,
fueron detectadas patologías asociadas a la extensión y flexión de
brazos y piernas asociadas al lanzamiento de elementos arrojadizos, a
la acción de remar, a la constante posición arrodillada y al transporte o
arrastre de cargas pesadas. Posiblemente estas actividades eran
realizadas en forma constante y prolongada lo cual genero huellas de
estrés mecánico asociadas a un sobre esfuerzo muscular (Constanzo
2012).
Responsable Victoria Castro (Castro V 2011).
Antecedentes principales del sitio CF3.
Unidad Alero 1
Descripción
Pequeño alero rocoso escasamente saqueado, ubicado al límite norte
del sitio CF3.
Materiales Fueron registradas osamentas humanas correspondiente a una mujer
de aproximadamente 30 años de edad (Andrade Com. Pers 2011), en

160
asociación a un ofertorio compuesto por: tejidos, cordeles, palitos de
madera, trozos de cuero de Otariidae, un tubo óseo, numerosos clastos
redondeados muy pulidos, algunos con pigmento rojo, líticos tales como
pulidores -algunos con huellas de uso-, un raspador y una pesa de
pesca de pizarra, fragmentos cerámica indeterminada y trozos de
mineral de Cu.
Al exterior del alero fue excavada una cuadrícula de 3x1, en la que se
recuperaron: un fogón, un anzuelo de Cu completo, fragmentos de
cerámica LCA y asignables PIT (Varela 2011), un pequeño rollo de piel
de Otariidae, dos cuentas de collar una de ellas posiblemente
manufacturada sobre concha de bivalvo, y la otra sobre Megabalanus
psittacus un instrumento conquiológico correspondiente a un cuchillo de
Choromytilus chorus. En cuanto al material lítico, se hallaron dos
puntas pedunculadas, un pulidor de andesita, dos manos de moler,
todos con pigmento rojo.
Carlos Aldunate (Aldunate 2011).
Antecedentes principales de la unidad Alero 1 del sitio CF3.
Unidad Túmulo 1
Descripción
Montículo funerario fuertemente saqueado con un agujero en el centro,
fue excavado por niveles artificiales de 10 cm.
Materiales En la superficie fueron recolectados restos arqueofaunísticos, trozos de
cactus, líticos y material cerámico indeterminado. En el nivel 1 se
extrajeron numerosos restos malacológicos, carboncillo, restos óseos
humanos y de animales, desbastes de microlascas, cerámica LCA,
pequeños fragmentos de estera y una pluma de Daption cápense
(Peña-Villalobos y Fibla 2011). El nivel 2 se caracterizó por la presencia
de material malacológico, microlascas, fragmentos de estera, cerámica
LCA, un pequeño fragmento de alga, conchuela, restos óseos humanos
y de animales, un instrumento óseo indeterminado -ovalado y pulido- y
dos rasgos correspondientes a eventos de quema. En el nivel 3 se
recuperaron restos malacológicos menos fragmentados que en los
niveles anteriores, también fueron halladas microlascas, fragmentos de

161
alga, de estera y de cerámica LCA, un óseo humano en mal estado y
osteofauna. En el nivel 4 fueron hallados fragmentos de cerámica,
fragmentos de estera, carbón, líticos, restos malacológicos, faunísticos y
orgánicos, además de un semicírculo de rocas y una especie de
conglomerado de arenisca con piedrecillas y conchuela de aspecto
limoso -el que parecía ser producto de una proceso natural por
humedad-. A la los 36 cm de profundidad se presentó un una argamasa
blanquecina sumamente compacta -similar al cemento-. En el nivel 5 se
registró cerámica LCA, un trozo de argamasa, restos malacológicos y
faunísticos, líticos tales como microlascas y guijarros, un diente
humano, un fragmento de cordelillo torcido a un solo lado y un esqueleto
de alacrán. En nivel 6 se extrajeron una cuerdita aparentemente hecha
de lana de camélido, un fragmento de cordelería (torcido a un solo lado),
cerámica, material malacológico, restos faunísticos, guijarros y un trocito
de papel.
Responsable Victoria Castro (Castro V 2011).
Imágenes
Antecedentes principales de la unidad Túmulo 1 del sitio CF3.

162
Unidad 4
Descripción
Cuadrícula de 1x1, excavada por niveles artificiales de 10 cm. Situada
sobre el perfil de saqueo de un túmulo.
Materiales El primer nivel se caracterizó por la presencia de un fragmento de
cerámica SEQ, restos óseos humanos y faunísticos, un instrumento de
roca roja, un pulidor, conchuela, restos malacológicos y trozos de textil
en mal estado. En el segundo nivel presentó carbones, restos óseos
faunísticos, trozos de estera y cerámica SEQ.
Responsable Carlos Aldunate (Aldunate 2011).
Antecedentes principales de la unidad 4 del sitio CF3.
P3:
Sitio P3
Unidad 1
Ubicación UTM 368995 – 7495203 datum WGS84-86 huso 19S
Función Conchal
Descripción Cuadrícula de 1x1, excavada por niveles artificiales de 10 cm. Situada al
costado de un camino vehicular
Materiales En superficie fueron hallados restos malacológicos. El nivel 1 presentó
una malla tejida en fibra, escasos restos de planta silvestre identificada
como Zephyra elegans (Vargas 2011), restos de osteofauna y
malacofauna, además de un evento de quema en el cual se encontraban
restos faunísticos no quemados (rasgo 1). El nivel 2 se caracterizó por la
presencia de restos de osteofauna, fragmentos de cerámica LCA,
microlascas, lascas, guijarros y madera. También, se presenta un rasgo
(2) compuesto de ceniza y concha bajo el cual se encontraba una mezcla
de arena con tierra café oscura. El nivel 3 se compuso de un piso de
ceniza (correspondiente al rasgo 2) donde se presentaron madera
trabajada, carbón, un artefacto conquiológico de Choromytilus chorus
(Castro P 2011b), restos óseos de animales, conchas de moluscos,
plumas de Larus dominicanus (Peña y Fibla 2011), cerámica LCA. En el
Nivel 4 continuó la presencia del rasgo 2, la cantidad de material cultural

163
se redujo a un fragmento de estera, lascas, cerámica indeterminada y
restos óseos faunísticos. En el nivel 5 se registraron restos óseos
faunísticos, fragmentos de cerámica indeterminada, material
malacológico y lascas. El nivel 6 se presentó con una matriz más
compacta que en los niveles anteriores y el material cultural se redujo
notablemente.
Responsable Macarena López (López 2011).
Imágenes
Antecedentes principales de la unidad 1 del sitio P3.

164
PC8:
Sitio PC8
Unidad 1
Ubicación UTM 368851 – 7495136 datum WGS84-86 huso 19S
Función Habitacional
Descripción Cuadrícula de 1x1, excavada por niveles artificiales de 10 cm. Este sitio
se compone de seis grandes depresiones que aparentemente
corresponden a habitaciones. Hacia el Este, dentro del mismo sitio se
ubica una gran estructura compuesta por cuatro hiladas de rocas.
Materiales En superficie fue identificada cerámica TRA y TGA. La capa 1 se definió
como orgánica y se caracterizó por la presencia de una preforma de
punta de proyectil y fragmentos de cerámica AIQ y DUP, ambas
correspondientes al PIT (Aldunate 2011, Varela 2011), restos de plantas
cultivadas (Zea mays), silvestres de recolección (Prosopis sp), silvestres
(Zephyra elegans) y espinas de Cactáceas, una de ellas con huellas de
corte (Varas 2011). En la capa 2 fueron registrados restos de osteofauna
y moluscos, cordelería sobre pelo de camélido y cabello humano
(Aldunate 2011), un artefacto conquiológico de Argopecten purpuratus
(Castro P 2011b), restos botánicos de Prosopis sp, Zea mays y Zephyra
elegans (Varas 2011) plumas de Sula variegata y Pelecanus thagus
(Peña-Villalobos y Fibla 2011) y un fragmento de cerámica la que debido
a la erosión no pudo ser determinada (Varela 2011).
Responsable Carlos Aldunate (Aldunate 2011).
Imágenes
Antecedentes principales de la unidad 1 del sitio PC8.

165
Chungungo 6:
Sitio Chungungo 6
Unidad Alero 1
Ubicación UTM 370271- 7496735 datum WGS84-86 huso 19S
Función Conchal
Descripción Cuadrícula de 1x1, excavada por niveles artificiales de 10 cm. Situada a
las afueras de un abrigo rocoso.
Materiales En la superficie fueron encontrados restos malacológicos, roca de mineral
de Cu, variados líticos y un fragmento de cerámica indeterminada. El
primer nivel se caracterizó por la presencia de carboncillo, dos
fragmentos de cerámica indeterminada, restos osteofaunísticos y
malacológicos. En el segundo nivel fueron hallados dos fragmentos de
mineral de Cu, tres puntas de proyectil, lascas líticas primarias y
secundarias, una posible cuenta de collar y material malacológico. En el
nivel 3 fueron detectadas rocas que parecen haber sido acomodadas en
diferentes tiempos de la ocupación, bajo estas rocas fue registrada la
presencia de una gran cantidad de malacofauna –Concholepas,
Fisurellas y Chitoniadae- las que parecen ser producto de actividades de
desconche, también fue registrado material lítico y restos de pigmento
rojo.
Responsable Valentina Varas (Varas 2012).
Imágenes

166
Antecedentes principales de la unidad Alero 1 del sitio Chungungo 6.

167
Anexo 3. Variables y Códigos para tabular los datos:
Se presentan las variables empleadas para el análisis y los códigos aplicados para la
óptima tabulación de los datos.
Variable Código
Taxón
Pieza ósea (Ver anexo 4)
Pieza ósea específica (Ver anexo 4)
Porción CO = Completo
FR = Fragmento
MI = Partido longitudinalmente por la mitad
EP = Epífisis
SH = Diáfisis
PX = Proximal
DS = Distal
Fusión O= Fusión indeterminada
F = Epífisis fusionada
N = Epífisis sin funcionar
M = Epífisis fusionándose o marcada
A = Diáfisis sin fusionar
Fracción %
Lateralidad D = Derecha
I = Izquierda
O = Indeterminado
A = Ambos
Fractura/forma primaria o fractura primaria L= Longitudinal
T = Transversal
E = Espiral
A = Ausente
Fractura/forma de borde o estado de
fractura
G = Fresca
N = No fresca
Marcas antrópicas A = Ausente
CO = Corte
RA = Raspado
HP = Hoyos de percusión
EP = Estrías de percusión
NE = Negativos de impacto
PE = Perforado
MA = Machacado
Distribución de marcas antrópicas A = Ausente
S = Aisladas
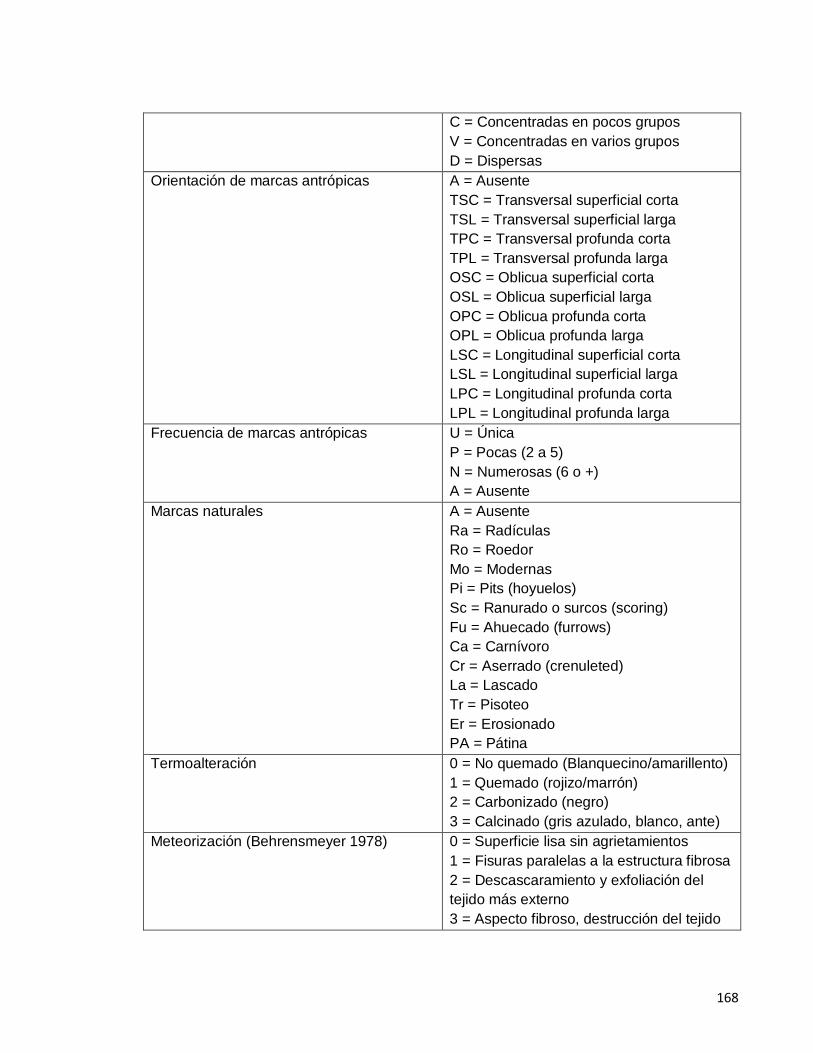
168
C = Concentradas en pocos grupos
V = Concentradas en varios grupos
D = Dispersas
Orientación de marcas antrópicas A = Ausente
TSC = Transversal superficial corta
TSL = Transversal superficial larga
TPC = Transversal profunda corta
TPL = Transversal profunda larga
OSC = Oblicua superficial corta
OSL = Oblicua superficial larga
OPC = Oblicua profunda corta
OPL = Oblicua profunda larga
LSC = Longitudinal superficial corta
LSL = Longitudinal superficial larga
LPC = Longitudinal profunda corta
LPL = Longitudinal profunda larga
Frecuencia de marcas antrópicas U = Única
P = Pocas (2 a 5)
N = Numerosas (6 o +)
A = Ausente
Marcas naturales A = Ausente
Ra = Radículas
Ro = Roedor
Mo = Modernas
Pi = Pits (hoyuelos)
Sc = Ranurado o surcos (scoring)
Fu = Ahuecado (furrows)
Ca = Carnívoro
Cr = Aserrado (crenuleted)
La = Lascado
Tr = Pisoteo
Er = Erosionado
PA = Pátina
Termoalteración 0 = No quemado (Blanquecino/amarillento)
1 = Quemado (rojizo/marrón)
2 = Carbonizado (negro)
3 = Calcinado (gris azulado, blanco, ante)
Meteorización (Behrensmeyer 1978) 0 = Superficie lisa sin agrietamientos
1 = Fisuras paralelas a la estructura fibrosa
2 = Descascaramiento y exfoliación del
tejido más externo
3 = Aspecto fibroso, destrucción del tejido

169
compacto
4 = Agrietamientos profundos y abiertos,
desprendimiento de astillas
5 = Astillamiento extensivo
Códigos del sistema para el registro de información arqueofaunística (Mengoni 1999).
Variable Código
Sitio, unidad, nivel y rasgo
Clase de cordado MM = Mastofauna marina
MT = Mastofauna terrestre
A = Avifauna
I = Ictiofauna
Segmento Apendicular
Axial
Craneal
Caudal
Precaudal
No identificado
Ubicación de la fractura D = Diáfisis
E = Epífisis
P = Proceso
A = Ausente
Edad 0 = Indeterminado
1 = Juvenil
2 = Adulto
Observaciones
Variables y códigos para la tabulación de datos (Elaboración propia).

170
Anexo 4. Lista general de partes esqueletarias:
Se presentan los códigos para la identificación de huesos en análisis de osteofauna de
mastofauna (Mengoni 1999).
Código Descripción
CRA Craneal indet.
BSC Caja craneal
MAXI Maxilar
MAXT Maxilar con dientes
PAR Parietal
OCC Occipital
BOC Basioccipital
OCN Occipital: cóndilo
FRN Frontal
MAN Mandíbula indet. o completa
MANT Mandíbula con dientes
SYM Sínfisis
SYMT Sínfisis con dientes
ANG Ángulo mandibular
RAM Ramus
CRN Proceso coronoide
ART Cóndilo articular
TTH Diente indet
ROOT Raíz
I0 Incisivo indet
DI0 Incisivo deciduo indet
P0 Premolar indet
DP0 Premolar deciduo indet
M0 Molar indet
CTH Molariforme indet
UP0 P superior indet.
UP1 P1 superior
UP2 P2 superior
UP3 P3 superior
DUP0 P superior deciduo indet
DUP1 P1 superior deciduo
DUP2 P2 superior deciduo
DUP3 P3 superior deciduo
UM0 Molar superior indet
UM1 M1 superior

171
UM2 M2 superior
UM3 M3 superior
DUM0 M superior deciduo indet
DUM1 M1 superior deciduo
DUM2 M2 superior deciduo
DUM3 M3 superior deciduo
UCH Molariforme superior indet
UTHR Serie dental superior
DUTR Serie dental superior decidua
LP0 P inferior indet
LP1 P1 inferior
LP2 P2 inferior
LP3 P3 inferior
DLP0 P inferior deciduo indet
DLP1 P1 inferior deciduo
DLP2 P2 inferior deciduo
DLP3 P3 inferior deciduo
LM0 M indet
LM1 M1 inferior
LM2 M2 inferior
LM3 M3 inferior
DLM0 M inferior deciduo indet
DLM1 M1 inferior deciduo
DLM2 M2 inferior deciduo
DLM3 M3 inferior deciduo
LCH Molariforme inferior indet
LTHR Serie dental inferior
DLTR Serie dental inferior decidua
HYO Hioides
AXL Axial indet
VRT Vértebra indet
VRTA Vértebra: arco
VRTC Vértebra: cuerpo
CER Vértebra cervical
ATL Atlas
AXI Axis
CER3 Cervical: vértebra 3
CER4 Cervical: vértebra 4
CER5 Cervical: vértebra 5
CER6 Cervical: vértebra 6
CER7 Cervical: vértebra 7

172
CERA Cervical: arco
CERC Cervical: cuerpo
CEF1 Cervical: proceso articular craneal (uno)
CEF2 Cervical: proceso articular craneal (dos)
CEH1 Cervical: proceso articular caudal (uno)
CEH2: Cervical: proceso articular caudal (dos)
CERT Cervical: proceso transverso
CERP Cervical: proceso espinoso
THO Torácica: vértebra indet
THO1 Torácica: vértebra 1
THO2: Torácica: vértebra 2
THO3 Torácica: vértebra 3
THO4 Torácica: vértebra 4
THO5 Torácica: vértebra 5
THO6 Torácica: vértebra 6
THO7 Torácica: vértebra 7
THO8 Torácica: vértebra 8
TH09 Torácica: vértebra 9
TH010 Torácica: vértebra 10
THO11 Torácica: vértebra 11
THO12 Torácica: vértebra 12
THO13 Torácica: vértebra 13
THO14 Torácica: vértebra 14
THO15 Torácica: vértebra 15
THO16 Torácica: vértebra 16
THO17 Torácica: vértebra 17
THO18 Torácica: vértebra 18
THO19 Torácica: vértebra 19
THO20 Torácica: vértebra 20
THOL Torácica: última vértebra
THOC Torácica: cuerpo
THOA Torácica: arco
THOP Torácica: proceso espinoso
LUM Lumbar: vértebra indet
LUM1 Lumbar: vértebra 1
LUM2 Lumbar: vértebra 2
LUM3 Lumbar: vértebra 3
LUM4 Lumbar: vértebra 4
LUML Lumbar: última vértebra
LUMC Lumbar: cuerpo
LUMA Lumbar: arco

173
LUMP Lumbar: proceso espinoso
LUMT Lumbar: proceso transverso
SAC Sacro o vértebra sacra
SAC1 Sacro: vértebra 1
SAC2 Sacro: vértebra 2
SAC3 Sacro: vértebra 3
SAC4 Sacro: vértebra 4
SAC5 Sacro: vértebra 5
SACL Sacro: última vértebra
SACC Sacro: cuerpo
SACA Sacro: proceso articular
CAU Vértebra caudal
CAUR Caudal: serie
TAIL Cola
RIB Costilla indet
RIB1 Costilla: primera
STE Esternón
SCP Escápula indet. o completa
SCPG Escápula: cavidad glenoidea
SCPA Escápula: acromion
SCPS Escápula: espina
SCPB Escápula: hoja
SCPC Escápula: coracoides
PEL Pelvis indet. o completa
ILI Ilion
ILIP Ilion: cuerpo
ILIT Ilion: cresta
ISC Isquion
ISCT: Isquion: tuberosidad
ISCI Isquion: incisura
ISCE Isquion: espina isquiática
PUB Pubis
ILIS Ilion e isquion
ILPB Ilion y pubis
ISPB Isquion y pubis
ACE Acetábulo
AILI Acetábulo: del ilion
AISC Acetábulo: del isquion
APUB Acetábulo: del pubis
AISI Acetábulo: isquion-ilion
APIL Acetábulo: pubis-ilion

174
APIS Acetábulo: pubis-isquion
LBN Hueso largo indet
HUM Húmero
HUME Húmero
HUM10 Húmero: diáfisis proximal
HUM11 Húmero: tuberosidad deltoidea
HUM12 Húmero: tuberosidad del redondo mayor
HUM15 Húmero: diáfisis media
HUM16 Húmero: foramen nutricio
HUM17 Húmero: cresta humeral y medial-anterior
HUM20 Húmero: diáfisis distal
HUM21 Húmero: cresta epicondilar
RAD Radio
RADE Radio: epífisis sin fusionar
ULN Ulna
ULNE Ulna: epífisis sin fusionar
MC0 Metacarpiano indet
MC1 Metacarpiano: primer dígito
MC2 Metacarpiano: segundo dígito
MC3 Metacarpiano: tercer dígito
MC4 Metacarpiano: cuarto dígito
MC5 Metacarpiano: quinto dígito
FEM Fémur
FEME Fémur: epífisis suelta
FEM40 Fémur: diáfisis proximal
FEM41 Fémur: trocánter menor
FEM42 Fémur: cresta intertrocanterica
FEM45 Fémur: diáfisis media
FEM46 Fémur: línea áspera
FEM47 Fémur: foramen nutricio
FEM50 Fémur: diáfisis distal
FEM51 Fémur: tuberosidad supracondilia lateral
TIB Tibia
TIBE Tibia: epífisis
TIB55 Tibia: diáfisis proximal
TIB56 Tibia: cresta tibial
TIB57 Tibia: foramen nutricio
TIB60 Tibia: diáfisis media
TIB61 Tibia: borde lateral y crestas del flexor digital
TIB62 Tibia: borde medial
TIB65 Tibia: diáfisis distal

175
FIB Fíbula
FIBE Fíbula: epífisis sin fusionar
MT0 Metatarsiano indet
MT1 Metatarsiano: primer dígito
MT2 Metatarsiano: segundo dígito
MT3 Metatarsiano: tercer dígito
MT4 Metatarsiano: cuarto dígito
MT5 Metatarsiano: quinto dígito
PAT Patella
PHA Falange indet
SES Sesamoideo indet.
CAR Carpiano indet
SCA Escafoides o radial
LUN Lunar o intermedio
CUN Cuneiforme o ulnar o triquetrum
MAG Magnum o 3º
UNC Unciforme o 4º
PIS Pisiforme o accesorio
TZD Trapezoides o 2º
TZM Trapecio o 1º
TAR Tarsiano indet.
AST Astrágalo
CAL Calcáneo
NAV Navicular
CUB Cuboides o 4º
CU1 Entocuneiforme o 1º
CU2 Cuneiforme intermedio o 2º
CU3 Ectocuneiforme o 3º
CALE Calcáneo: epífisis sin fusionar
MAL Hueso maleolar
SKEL Esqueleto completo
DER Huesos dérmicos
HOOF Pezuña
NID No identificable
CON Fragmento condiliar
EPI Epífisis: fragmento sin fusionar indet
ATS Superficie articular indet
CTC Cartílago
LAS Lasca ósea
Códigos para la tabulación de piezas óseas (Mengoni 1999).

176
Anexo 5. Fichas de taxones identificados en la muestra:
Se presentan las fichas con información respecto de los taxones identificados en la
muestra.
a. Avifauna
En el área de estudio habitan tres especies del genero Phalacrocorax: Lile (Phalacrocorax
gaimardi), Guanay (Phalacrocorax bougainvillii) y Yeco (Phalacrocorax brasilianus)
(Jaramillo 2005). Debido a que estas especies comparten grandes similitudes a nivel
osteométrico, no fue posible diferenciarlas, por lo que las unidades anatómicas fueron
asignadas taxonómicamente a nivel de género.
Phalacrocorax gaimardi:
Nombre común Lile
Nombre científico Phalacrocorax gaimardi
Género Phlacrocorax
Familia Phalacrocoracidae
Orden Suliforme
Clase Avifauna
Distribución Entre las costas de Arica y Aisén (Jaramillo
2005).
Longitud 76 cm (Jaramillo 2005).
Alimentación Peces, crustáceos, moluscos y algas
(Vilina y Cofré 2008b, Jaramillo 2005).
Etología Ave marina. No posee hábitos pelágicos,
se alimenta cerca de la costa. Nidifica en
colonias difusas en acantilados. Vuela
cerca del agua normalmente solo, no en
bandadas (Jaramillo 2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).

177
Uso por parte de los humanos Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Phalacrocorax gaimardi.
Phalacrocorax bougainvillii:
Nombre común Guanay
Nombre científico Phalacrocorax bougainvillii
Género Phalacrocorax
Familia Phalacrocoracidae
Orden Suliforme
Clase Avifauna
Distribución Entre las costas de Arica y Biobío
(Jaramillo 2005).
Longitud 76 cm (Jaramillo 2005).
Alimentación Piscívoro5 (Jaramillo 2005).
Etología Ave marina, pelágica y guanera. Se
alimenta en aguas profundas a muchos km
de la costa. De hábitos gregarios, cría en
colonias densas en islas planas.
Normalmente vuela en bandadas (Jaramillo
2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Explotación de guano (Jaramillo 2005,
Stucchi 2011).
- Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Phalacrocorax bougainvillii.
5 Piscívoro: que se alimenta de peces.

178
Phalacrocorax brasilianus:
Nombre común Yeco
Nombre científico Phalacrocorax brasilianus
Género Phalacrocorax
Familia Phalacrocoracidae
Orden Suliforme
Clase Avifauna
Distribución
Desde las costas de Arica a Punta Arenas,
también es común en el interior,
específicamente en la Cordillera entre Arica
y Antofagasta (Jaramillo 2005).
Longitud Entre 58 y 76 cm (Jaramillo 2005).
Alimentación Peces y pequeños cistáceos (Jaramillo
2005).
Etología Ave acuática, no estrictamente marina.
Nidifica en colonias en árboles,
generalmente asociados a masas de agua.
Vuela solo y en bandadas. Dado que no
posee plumas impermeables, tras salir del
agua, debe secarse en la superficie
extendiendo sus alas en forma de „M´
(Araya y Millie 1992, Jaramillo 2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Phalacrocorax brasilianus.

179
Pelecanus thagus:
Nombre común Pelicano
Nombre científico Pelecanus thagus
Género Pelecanus
Familia Pelecanidae
Orden Pelecaniforme
Clase Avifauna
Distribución Entre las regiones de Arica y Los Lagos
(Jaramillo 2005).
Longitud 152 cm (envergadura alas de punta a
punta: 228 cm) (Jaramillo 2005).
Alimentación Piscívoro (Jaramillo 2005).
Etología Ave marina y guanera. De hábitos
pelágicos y costeros. Anida en grandes
colonias (Jaramillo 2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Explotación de guano (Jaramillo 2005,
Stucchi 2011).
- Uso de plumas para la confección de
diademas (Horta 2000).
- Consumo de carne (Peña-Villalobos et al.
2013).
- Uso de huesos para la confección de
copunas -dispositivos para inflar balsas de
cuero de lobo- (Niemeyer 1965-66, De
Vivar 1952).
Ficha de identificación de Pelecanus thagus.

180
Egretta
En el área de estudio habitan dos especies del género Egretta: Garza chica (Egretta thula)
y Garza azul (Egretta caerulea). Debido a que ambas especies comparten grandes
similitudes a nivel osteométrico, las unidades anatómicas fueron asignadas
taxonómicamente a nivel de género.
Egretta thula:
Nombre común Garza chica
Nombre científico Egretta thula
Género Egretta
Familia Ardeidae
Orden Pelecaniforme
Clase Avifauna
Distribución Entre las regiones de Arica y Los Lagos.
Siendo escasas en el sector norte
(Jaramillo 2005).
Longitud 56 - 66 cm (Jaramillo 2005).
Alimentación Piscívoro (Jaramillo 2005).
Etología Ave acuática y marina. De hábitos
gregarios. Nidifica en árboles, riscos o
totorales. No presenta incomodidad al
compartir su hábitat con otras especies de
garzas (Jaramillo 2005).
Forma de obtención
Uso por parte de los humanos
Ficha de identificación de Egretta thula.
Egretta caerulea:
Nombre común Garza azul
Nombre científico Egretta caerulea
Género Egretta

181
Familia Ardeidae
Orden Pelecaniforme
Clase Avifauna
Distribución Desde Arica hasta la mitad de la II región
(Jaramillo 2005).
Longitud 56 cm (Jaramillo 2005).
Alimentación Piscívoro (Jaramillo 2005).
Etología Ave marina (Jaramillo 2005).
Forma de obtención
Uso por parte de los humanos
Ficha de identificación de Egretta caerulea.
Larus dominicanus:
Nombre común Gaviota Dominicana
Nombre científico Larus dominicanus
Género Larus
Familia Laridae
Orden Charadriiformes
Clase Avifauna
Distribución Desde Arica hasta Cabo de Hornos
(Jaramillo 2005).
Longitud 58 cm (envergadura alas de punta a punta:
entre 128 y 142 cm) siendo la gaviota más
grande de Sudamérica (Ayala y Millie 1992,
Jaramillo 2005).
Alimentación Piscívoro (Araya y Millie 1992, Jaramillo
2005).
Etología Ave marina, prefiere las costas rocosas
(Jaramillo 2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
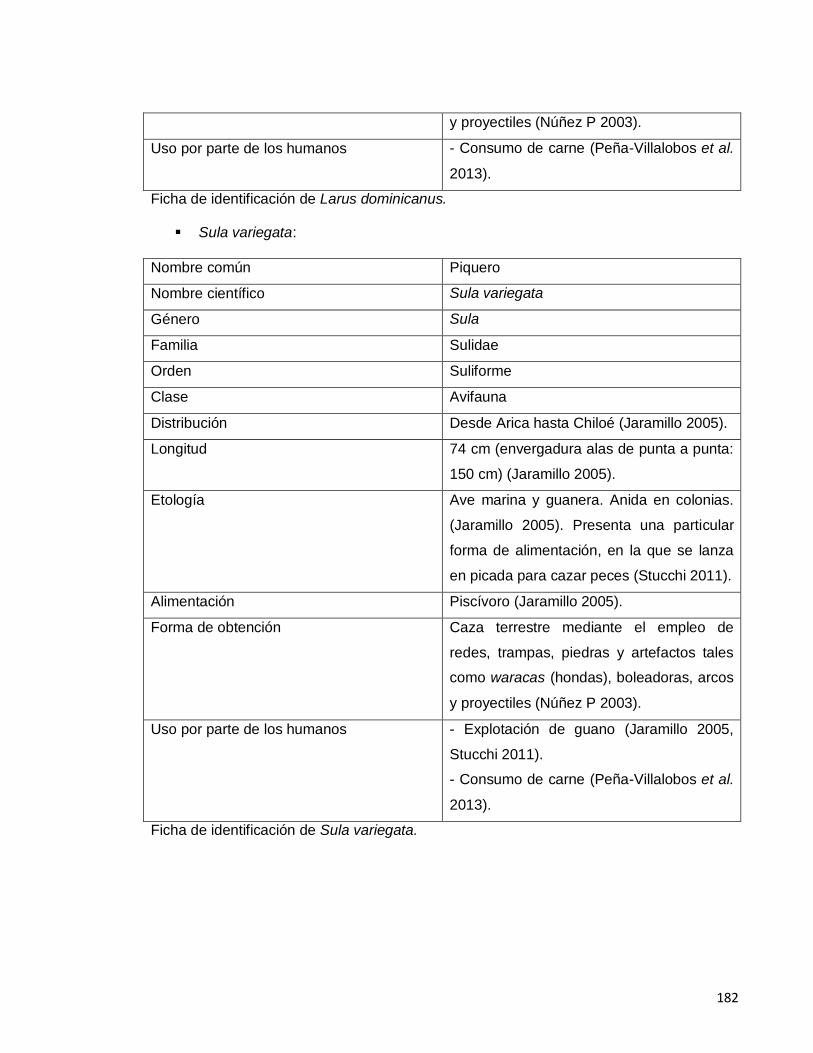
182
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Larus dominicanus.
Sula variegata:
Nombre común Piquero
Nombre científico Sula variegata
Género Sula
Familia Sulidae
Orden Suliforme
Clase Avifauna
Distribución Desde Arica hasta Chiloé (Jaramillo 2005).
Longitud 74 cm (envergadura alas de punta a punta:
150 cm) (Jaramillo 2005).
Etología Ave marina y guanera. Anida en colonias.
(Jaramillo 2005). Presenta una particular
forma de alimentación, en la que se lanza
en picada para cazar peces (Stucchi 2011).
Alimentación Piscívoro (Jaramillo 2005).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Explotación de guano (Jaramillo 2005,
Stucchi 2011).
- Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Sula variegata.

183
Spheniscus humboldti:
Nombre común Pingüino de Humboldt
Nombre científico Spheniscus humboldti
Género Spheniscus
Familia Spheniscidae
Orden Sphenisciforme
Clase Avifauna
Distribución Desde Arica hasta Chiloé (Jaramillo 2005).
Longitud 65 cm (Jaramillo 2005).
Etología Ave marina, pelágica y no voladora. Cría
en colonias durante todo del año.
Constituye la especie de ave mejor
adaptada al medio marino (Ayala y Millie
1992, Jaramillo 2005).
Alimentación Peces y cefalópodos 6 (Vilina y Cofré
2008b).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Consumo de carne (Peña-Villalobos et al.
2013).
Ficha de identificación de Spheniscus humboldti.
b. ictiofauna:
Trachurus murphyi:
Nombre común Jurel
Nombre científico Trachurus murphyi
Género Trachurus
Familia Carangidae
6 Cefalópodos: invertebrados marinos tales como pulpos y calamares.

184
Orden Perciforme
Clase Actinopterygii
Distribución De Arica a Valdivia (Moreno y Castillo
1976).
Longitud 65 cm (longitud máxima a los 12 años)
(Moreno y Castillo 1976). Puede lograr un
peso de 2,5 kg (Vargas et al. 1993).
Alimentación Microinvertebrados y peces (Moreno y
Castillo 1976).
Etología Pez pelágico de las regiones nerítica y
oceánica (Moreno y Castillo 1976). A fines
del otoño e inicios del invierno, busca
alimento, aproximándose a la costa en
grandes cardúmenes (Vargas et al. 1993).
Forma de obtención Pesca desde la orilla y en el sector
intermedio mediante el uso de anzuelos,
tridentes, arpones y redes (Llagostera
1982, Núñez P 2003).
Uso por parte de los humanos - Consumo de carne (Núñez P 2003).
- Producción de charquesillo para
intercambiarlo con grupos del interior
(Mostny 1952, Castro et al. 2010, Núñez y
Santoro 2011, Pimentel et al. 2011).
Ficha de identificación Trachurus murphyi.
Cilus gilberti:
Nombre común Corvina
Nombre científico Cilus gilberti
Género Cilus
Familia Sciaenidae
Orden Perciforme

185
Clase Actinopterygii
Distribución Desde Iquique a Chiloé (Moreno y Castillo
1976).
Longitud Entre 62 y 80 cm (Moreno y Castillo 1976).
Alimentación Microinvertebrados y peces (Moreno y
Castillo 1976).
Etología Sub litoral superior (Moreno y Castillo
1976).
Forma de obtención Pesca desde la orilla mediante el uso de
anzuelos y redes (Núñez P 2003).
Uso por parte de los humanos - Consumo de carne (Núñez P 2003).
- Intercambio con grupos del interior
(Castillo 2011).
Ficha de identificación de Cilus gilberti.
c. Mastofauna:
Mastofauna marina:
En el área de estudio habitan dos especies de la familia Otariidae: Arctocephalus australis
(Lobo fino de dos pelos) y Otaria flavescens (lobo marino de un pelo). Ambos poseen un
gran parecido a nivel osteométrico (Borella et al. 2010). Adicionalmente, poseen un
marcado dimorfismo sexual en el que la hembra Otaria flavescens es sumamente similar
al macho Arctocephalus australis (Iriarte 2008). Dichas semejanzas dificultan la
identificación a nivel de especie, por lo que las unidades anatómicas fueron asignadas a
nivel de familia.
Arctocephalus australis:
Nombre común Lobo fino de dos pelos
Nombre científico Arctocephalus australis
Género Arctocephalus
Familia Otariidae

186
Orden Carnívora
Clase Mammalia
Distribución Al norte: entre Punta Paquina (I región) y
Rocas Abato (II región). Al sur: entre la isla
de Chiloé (X región) y Cabo de Hornos (XII
región) (Iriarte 2008).
Longitud Machos: 2 m, hebras: 1,5 m (Iriarte 2008).
Alimentación Peces y krill (Iriarte 2008).
Etología Durante las horas de mayor calor
permanecen principalmente en el agua. En
la noche se alimentan en aguas poco
profundas. Forman grupos de hasta 13
hembras por macho (Iriarte 2008).
Forma de obtención Fue un recurso explotado desde tiempos
tempranos. Los adultos eran cazados
mediante el uso de arpones (Llagostera
1982) y palos (Núñez P 2003). Por otro
lado, los juveniles eran recolectados
(Llagostera 1982). Según escritos de
cronistas …en los días que no hace aire
andan los lobos marinos descuidados
durmiendo, y llegan seguros los indios con
sus balsas. Tíranle un harpón de cobre y
por la herida se desangran y muere (De
Vivar 1987:10).
Uso por parte de los humanos
- Consumo de carne y sangre debido a su
alto nivel proteico (Lizarraga 1968,
Llagostera 1982)
Ficha de identificación de Arctocephalus australis.

187
Otaria flavescens:
Nombre común Lobo marino de un pelo
Nombre científico Otaria flavescens
Género Otaria
Familia Otariidae
Orden Carnívora
Clase Mammalia
Distribución Desde Arica a Cabo de Hornos (Iriarte
2008)
Longitud Machos: 3,5 m . Hembras 2,5 m (Iriarte
2008)
Alimentación Peces, cefalópodos, crustáceos y
pingüinos (Sielfeld 1983).
Etología Forman grupos de hasta 13 hembras por
macho (Iriarte 2008, Sielfeld 1999).
Forma de obtención Fue un recurso explotado desde tiempos
tempranos. Los adultos eran cazados
mediante el uso de arpones (Llagostera
1982) y palos (Núñez P 2003). Por otro
lado, los juveniles eran recolectados
(Llagostera 1982). Según escritos de
cronistas …en los días que no hace aire
andan los lobos marinos descuidados
durmiendo, y llegan seguros los indios con
sus balsas. Tíranle un harpón de cobre y
por la herida se desangran y muere (De
Vivar 1987:10).
Uso por parte de los humanos
- Consumo de carne y sangre debido a su
alto nivel proteico (Lizarraga 1968,
Llagostera 1982)
- Uso de cuero y tripas para confección de
balsas (Niemeyer 1965-66, De Vivar 1987).

188
- Uso de grasa como combustible (siglos
XVI y XVII) (Aguayo y Maturana 1973).
Ficha de identificación de Otaria flavescens.
Mastofauna terrestre:
Lama guanicoe:
Nombre común Guanaco
Nombre científico Lama guanicoe
Género Lama
Familia Camelidae
Orden Artiodactyla
Clase Mammalia
Distribución Todo Chile en forma fragmentada, desde el
nivel del mar hasta sobre los 4.500 m.s.n.m
(Iriarte 2008).
Longitud 1,6 m (Iriarte 2008). Los camélidos
constituyen a los mamíferos herbívoros
nativos más grandes de Sudamérica
(Bonavia 1996).
Alimentación Herbívoro (Iriarte 2008).
Etología Nacimiento en verano, solo nace una cría
por hembra -11 meses de gestación-
(Iriarte 2008).
Forma de obtención Caza terrestre mediante el empleo de
redes, trampas, piedras y artefactos tales
como waracas (hondas), boleadoras, arcos
y proyectiles (Núñez P 2003).
Uso por parte de los humanos - Consumo de carne (Núñez L 1989).
- Uso de su cuero y pelaje (Núñez L 1989).
- Uso de huesos para confección de
artefactos (Núñez L 1989).
Ficha de identificación de Lama guanicoe.

189
Anexo 6. Imágenes:
Especie Phalacrocorax brasilianus en postura característica en forma de „M‟. Fotografía
de Juan Tassara tomada el 16/04/2012.
Esquema de Sula variegata precipitándose en picada hacia el agua (Stucchi 2011).




















