
Uptake and metabolism of allantoin and allantoate by cells of Chlamydomonas reinhardtii...
9
This article was downloaded by: [117.175.100.245] On: 25 March 2014, At: 11:01 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK European Journal of Phycology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tejp20 Uptake and metabolism of allantoin and allantoate by cells of Chlamydomonas reinhardtii (Chlorophyceae) Pedro Piedras , Miguel Aguilar & Manuel Pineda a Departamento de Bioquímica y Biología Molecular, and Instituto Andaluz de Biotecnología, Facultad de Ciencias, Universidad de Córdoba, 14071 Córdoba, Spain Published online: 03 Jun 2010. To cite this article: Pedro Piedras , Miguel Aguilar & Manuel Pineda (1998) Uptake and metabolism of allantoin and allantoate by cells of Chlamydomonas reinhardtii (Chlorophyceae), European Journal of Phycology, 33:1, 57-64, DOI: 10.1080/09670269810001736533 To link to this article: http://dx.doi.org/10.1080/09670269810001736533 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Uptake and metabolism of allantoin and allantoate by cells of Chlamydomonas reinhardtii...

This article was downloaded by: [117.175.100.245]On: 25 March 2014, At: 11:01Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
European Journal of PhycologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tejp20
Uptake and metabolism of allantoin and allantoate bycells of Chlamydomonas reinhardtii (Chlorophyceae)Pedro Piedras , Miguel Aguilar & Manuel Pinedaa Departamento de Bioquímica y Biología Molecular, and Instituto Andaluz de Biotecnología,Facultad de Ciencias, Universidad de Córdoba, 14071 Córdoba, SpainPublished online: 03 Jun 2010.
To cite this article: Pedro Piedras , Miguel Aguilar & Manuel Pineda (1998) Uptake and metabolism of allantoin andallantoate by cells of Chlamydomonas reinhardtii (Chlorophyceae), European Journal of Phycology, 33:1, 57-64, DOI:10.1080/09670269810001736533
To link to this article: http://dx.doi.org/10.1080/09670269810001736533
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Eur. J. Phycol. (1998), 33 : 57–64. Printed in the United Kingdom 57
Uptake and metabolism of allantoin and allantoate by cells of
Chlamydomonas reinhardtii (Chlorophyceae)
PEDRO PIEDRAS, MIGUEL AGUILAR AND MANUEL PINEDA
Departamento de BioquıUmica y BiologıUa Molecular, and Instituto Andaluz de BiotecnologıUa, Facultad de Ciencias, Universidad de CoU rdoba,14071 CoU rdoba, Spain
(Received 27 February 1997 ; accepted 30 September 1997)
The green alga Chlamydomonas reinhardtii can use the ureides allantoin and allantoate as sole nitrogen sources. Once the uptake systems
for allantoin and allantoate were induced, the uptake and growth rates were identical for the two ureides. However, the enzymatic
activities involved in the degradation of the two ureides (allantoinase and allantoicase) were regulated differently. Allantoinase seems to
be constitutive, since it was detected in all the nitrogen sources studied, while allantoicase behaved as an inducible enzyme, since it was
present only in cells cultured in ureides or any metabolic precursor of these compounds. Neither allantoinase nor allantoicase activities
were repressed by ammonium in the presence of ureides. Allantoicase activity was not induced under nitrogen starvation conditions,
while it was induced in cells that had been cultured with allantoin or allantoate in the dark. Allantoin uptake showed a pattern similar to
that of allantoate under all nutritional and environmental conditions tested. Inhibition of allantoin and allantoate uptake by N-
ethylmaleimide suggests that thiol (SH–) groups are involved in both uptake systems. The use of both allantoin and allantoate was
similarly inhibited by the metabolic poisons tested (cyanide, azide, 2,4-dinitrophenol and 3«-(3,4-dichlorophenyl)-1«,1«-dimethyl urea) but
only at very high concentrations. The possibility that uptake of allantoin and allantoate might take place through two independent
systems is discussed.
Key words : allantoate, allantoicase, allantoin, allantoinase, Chlamydomonas, nitrogen metabolism, uptake, ureides
Introduction
The presence of significant amounts of dissolved organicnitrogen (DON) in seawater and freshwater is wellestablished (Thurman, 1985). However, little is knownabout the nature of the components of DON, especiallythat of freshwater. A number of nitrogen compounds(amino acids, urea, purines and ureides among others) areknown to contribute to DON at variable concentrations(Antia et al., 1991). In spite of the low concentrations(micromolar) at which they are found, several authorshave proposed the relevance of these compounds asnitrogen sources for the growth of algae (Antia et al.,1980 ; Allison & Syrett, 1987 ; Oliveira & Huynh, 1990). Inparticular, the importance of purines and ureides as anitrogen source has been related to the existence ofnitrogen micro-cycles in which algae might be using thepurines and ureides excreted by animals and zooplankton(Antia et al., 1980). Some reports also suggest that in manyparts of the ocean most of the ammonium is taken up byheterotrophic bacteria and not by phytoplankton (Laws et
al., 1985 ; Kokkinakis & Wheeler, 1987).The few available data concerning the use of purines
and ureides by algae suggest that the degradation of thesecompounds follows the standard aerobic catabolism
Correspondence to : M. Pineda. Fax : 34 57 218606. e-mail :
bb1piprm!uco.es.
pathway (Scheme 1) proposed for other microorganisms(Vogels & van der Drift, 1976) and for higher plants(Reynolds et al., 1982). The first studies on purinecatabolism in Chlorella cells led to the identification ofsome intermediaries of the degradation pathway (Amman& Lynch, 1964). Later we found that Chlamydomonas cellscould grow in media with adenine, guanine, xanthine,hypoxanthine, urate, allantoin or allantoate as the solenitrogen source (Pineda et al., 1984). Uptake systems forurate (Pineda & Ca! rdenas, 1985 ; Pineda et al., 1987),xanthine and hypoxanthine (Pe! rez-Vicente et al., 1991),adenine and guanine (Lisa et al., 1995) have beencharacterized. In addition, xanthine dehydrogenase (Pe! rez-Vicente et al., 1988, 1992) and urate oxidase (Pineda et al.,1984 ; Alamillo et al., 1991, 1992) have been purified fromthis organism and their kinetic and molecular propertieshave been reported. The most relevant aspects of purineuptake and metabolism have been reviewed recently(Pineda & Ca! rdenas, 1996).
According to Antia et al. (1991), one would expect thatthose algae using purines for growth would also grow onureides as the sole nitrogen source. However, allantoinwas not used by the dozen freshwater microalgae tested,of which five species did grow both on xanthine and onuric acid (Birdsey & Lynch, 1962). Similarly, only fourspecies of marine microalgae from a survey of 18 speciesknown to use hypoxanthine proved to be allantoin
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014

58P. Piedras et al.
Scheme 1. Ureide biosynthesis and catabolism pathways in algae.
1, xanthine dehydrogenase (EC 1.1.1.204) ; 2, uricase (EC 1.7.3.3) ;
3, allantoinase (EC 3.5.2.5) ; 4 allantoicase (EC 3.5.3.4) ; 5, allantoate
deiminase (EC 3.5.3.9) ; 6, ureidoglycine aminohydrolase ;
7, ureidoglycine-oxalate transaminate ; 8, ureidoglycolate
dehydrogenase (EC 1.1.1.154) ; 9, ureidoglycolase (EC 4.3.2.3) ;
10, urease (EC 3.5.1.5) ; 11, allantoate deiminase (EC 3.5.3.9).
consumers (Antia et al., 1980). These authors suggest thatbenthic marine microalgae might be better adapted to useallantoin than the holoplanktonic types. Surprisingly, allsix freshwater chlorophycean algae tested could use bothhypoxanthine and allantoin, and none of them required asignificant acclimation period (Devi Prassad, 1983). Thereare even fewer reports on the use of allantoic acid byalgae. Apart from Chlamydomonas reinhardtii (Pineda et al.,1984), three prymnesiophytes, a chlorophyte, a cyano-bacterium and a eustigmatophyte from 11 species testedcould grow on allantoate, while two diatoms, a dino-flagellate, a chrysophyte and a prymnesiophyte showedno growth (Oliveira & Huynh, 1990).
Natural nitrogen sources are most frequently mixturesof different inorganic and organic compounds. Some ofthese mixtures have been studied in the laboratory(Pettersen, 1975 ; Ricketts, 1988). There has been atendency to consider urea, ammonium and nitrate as theonly nitrogen sources available to phytoplankton. How-ever, multiple nitrogen sources can function quiteeffectively for phytoplankton growth, particularly undernatural conditions, where single nitrogen sources are veryunlikely to occur. In this context, studying the uptake andmetabolism of ureides is of great interest. The enzymatic
activities involved in ureides metabolism – allantoinase(EC 3.5.2.5) and allantoicase (EC 3.5.3.4) or allantoatedeiminase (EC 3.5.3.9) – have been measured only rarely(Villeret, 1955, 1958 ; Oliveira & Huynh, 1990), and theirregulation has never been studied. Recently we havedeveloped a procedure for extracting both enzymes fromChlamydomonas cells, and found that they are associatedwith membranes (Piedras et al., 1995). In addition, theseenzymes have been purified and characterized (Piedras,1995), which has allowed us to demonstrate that allantoatedegradation is catalysed by allantoicase, but not byallantoate deiminase.
In this paper, we report on the physiological regulationof ureide uptake and allantoinase and allantoicase activitiesin the unicellular green alga Chlamydomonas reinhardtii
subjected to different nutritional and environmentalconditions. We believe that improving our knowledge ofthe regulatory aspects of ureide uptake and catabolism isessential for understanding the dynamics of nitrogenmicro-cycles in aquatic ecosystems, especially the con-nections between the nitrogen cycles of heterotrophs andphytoplankton. It will also help to re-evaluate the realrelevance of ureides for algal growth in natural eco-systems.
Materials and methods
Growth conditions and preparation of crude extracts
Cells of Chlamydomonas reinhardtii 6145c wild strain (fromDr R. Sager’s collection, Sidney Farber Center, New York)were cultured at 24 °C under continuous illumination(75–100 µm m−# s−", provided by daylight fluorescenttubes) in the liquid culture medium of Sueoka (1960) with8 mM ammonium chloride as the nitrogen source andunder a stream of CO
#-enriched air (4–5% v}v) as the
carbon source. Cells were harvested by centrifugation at4000 g for 10 min at 4 °C. Cell pellets were washed withdistilled water, then centrifuged as above, and transferredto minimum medium with allantoin or allantoate as thenitrogen source.
To prepare cell-free extracts, cells were harvested bycentrifugation at 14000 g for 10 min and stored at®20 °C until use. Frozen cells were disrupted by thawingand gentle shaking in 20 mM Tris-HCl, pH 8.3, 1 mMMnSO
%. The homogenate was then sonicated in a
Vibracell sonifer (Sonics and Materials, Danbury, CN) at40 W for 2 min (50% duty cycles), and the resultingsuspension used as cell extract for determination ofallantoinase and allantoicase activities. To avoid anylosses of enzymatic activities, cell extracts were preparedwithin 2–3 days after harvesting and activities measuredimmediately afterwards (Piedras et al., 1995).
Allantoin and allantoate uptake
The uptake of ureides was measured by following theirdisappearance from the media after sedimenting cells by
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014
Ureides uptake by Chlamydomonas 59
centrifugation at 14000 g for 10 min. Allantoin andallantoate in the supernatant were determined after Vogels& van der Drift (1970). The initial concentration of ureidewas 1 mM. In short-term experiments, aliquots of culturewere centrifuged at 6000 g for 2 min in a BeckmanMicrofuge E. Supernatants were frozen at ®20 °C untiluse, and determinations were performed in duplicate foreach sample.
Enzyme assays
Allantoinase was assayed by following the formation ofphenylhydrazone from the glyoxylate produced by acidhydrolysis of the reaction product, allantoate (Vogels &van der Drift, 1970). The standard assay mixture, in a finalvolume of 7 ml, was 20 mM Tris-HCl, pH 8.3, 10 mMallantoin and 1 mM MnSO
%. The reaction was carried out
at 30 °C and started by adding an adequate amount ofenzyme extract. The amount of allantoate formed wasdetermined at different times in 1 ml aliquots of thereaction mixtures. Enzymatic hydrolysis was stopped byadding 0.2 ml of 0.15 N HCl and immersing the glasstubes in boiling water for 10 min to transform allantoateinto glyoxylate. Then, 0.2 ml of 0.4 M phosphate buffer,pH 7.0, and 0.2 ml and 25 mM phenylhydrazine wereadded, and the mixture incubated for 10 min at roomtemperature. Finally, 1 ml of 12 N HCl and 0.2 ml of50 Mm potassium ferricyamide were added. After 15 minat room temperature, the absorbances of the samples weredetermined at 535 nm. Parallel controls to account fornon-enzymatic hydrolysis of allantoin to allantoate werecarried out along with the assays (Thomas et al., 1983).
Allantoicase activity was assayed by following theformation of phenylhydrazone from the glyoxylateproduced by alkaline hydrolysis of ureidoglycolate, theproduct of the enzymatic reactions. The standard assaymixture was 20 mM Tris-HCl, pH 8.3, and 5 mM allantoicacid in a final volume of 3 ml. The reaction was carried outat 30 °C and was started by adding an adequate amount ofenzyme extract. At different reaction times, the amount ofureidoglycolate formed was assayed in 0.2 ml aliquotsafter stopping hydrolysis by adding 0.2 ml of 0.5 NNaOH (which also transformed ureidoglycolate intoglyoxylate). To each aliquot of the reaction mixture,0.8 ml of 0.4 M phosphate buffer, pH 7.0, and 0.2 ml of25 mM phenylhydrazine were added. Samples were thentreated as above and their absorbances determined at535 nm. Controls were also included to correct for thenon-enzymatic breakdown of allantoate.
One unit of enzymatic activity is defined as the amountof enzyme that catalyses the formation of 1 micromole ofproduct per minute under the indicated assay conditions.
Analytical determinations
Protein concentration was determined after Bradford(1976) using bovine serum albumin as a standard.Allantoin and allantoate in the culture medium weredetermined after Vogels & van der Drift (1970), except
that the final volume was 2.6 ml. Density of cell cultureswas determined turbidimetrically at 660 nm, which is areliable index of cell density up to absorbances of 2. Ifrequired, exponential growth was verified by checking thelinearity of log(A
''!) versus time in cell cultures. Am-
monium and nitrate were determined by the phenol}hypochlorite and brucine methods, respectively (Pineda &Ca! rdenas, 1988).
All experiments were performed in triplicate. In theillustrations, the best of the three replicates is shown.
Chemicals
Allantoin, allantoic acid, 2,4-dinitrophenol (2,4-DNP), N-ethylmaleimide (NEM), 3«-(3,4-dichlorophenyl)-1«,1«-dimethyl urea (DCMU), ureidoglycolate and glyoxylatewere purchased from Sigma (St Louis, MO). Phenyl-hydrazine was from Merck (Darmstadt, Germany). Allother chemicals were of analytical grade.
Results and discussion
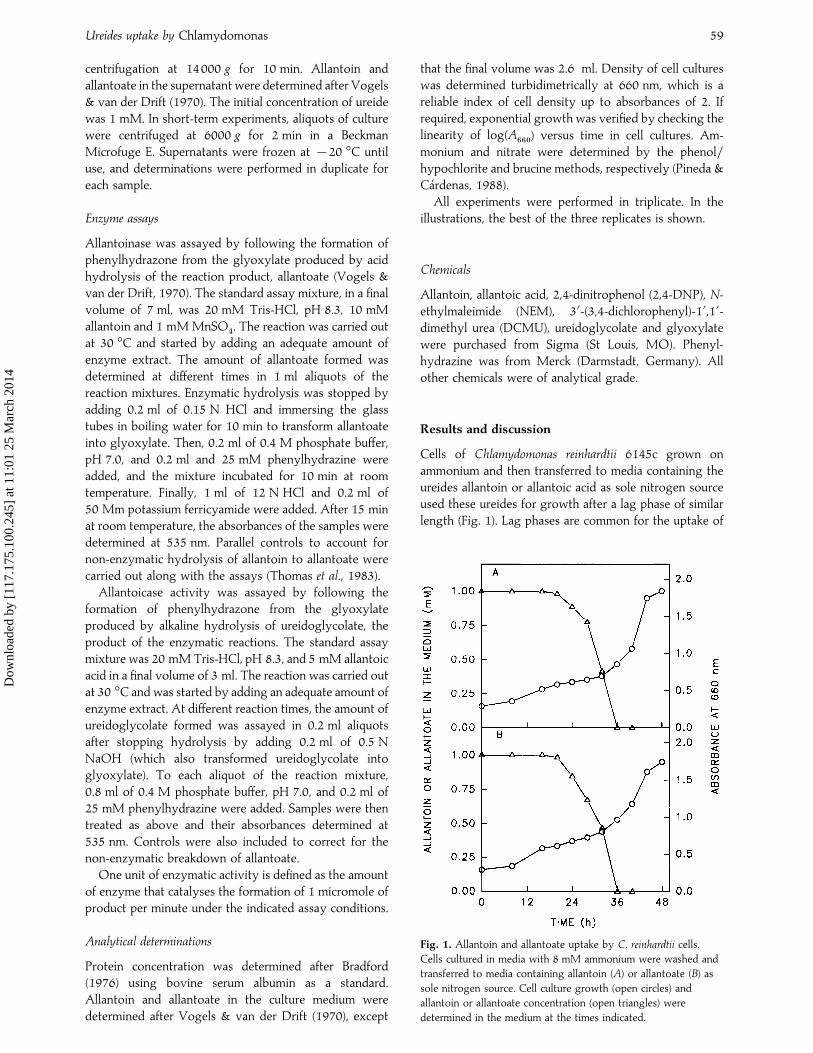
Cells of Chlamydomonas reinhardtii 6145c grown onammonium and then transferred to media containing theureides allantoin or allantoic acid as sole nitrogen sourceused these ureides for growth after a lag phase of similarlength (Fig. 1). Lag phases are common for the uptake of
Fig. 1. Allantoin and allantoate uptake by C. reinhardtii cells.
Cells cultured in media with 8 mM ammonium were washed and
transferred to media containing allantoin (A) or allantoate (B) as
sole nitrogen source. Cell culture growth (open circles) and
allantoin or allantoate concentration (open triangles) were
determined in the medium at the times indicated.
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014

60P. Piedras et al.
metabolic precursors of ureides by C. reinhardtii cells,though they are shorter than those for ureides (Pineda &Ca! rdenas, 1996). After these lag phases, the cells took upboth ureides linearly and at a similar rate, with maximumvalues of 46³9 fmol h−" cell−". These uptake rates arehigher than those found for purines in C. reinhardtii
(Pineda & Ca! rdenas, 1996).Growth rates and generation times in allantoin and
allantoic acid were also very similar (Fig. 1). Growth ratesin ureides are also similar to those in hypoxanthine andxanthine, higher than those in adenine, and lower thanthose in ammonium, nitrate, urea, guanine and urate(Pineda et al.. 1984 ; Pineda & Ca! rdenas, 1996). One wouldexpect that unicellular algae growing on purines as thesole nitrogen source could also use allantoin and allantoicacid for growth (Antia et al., 1991). However, five speciesof freshwater algae capable of growing on xanthine andurate could not grow on allantoin (Birdsey & Lynch,1962). In addition, only 4 of 18 marine species analysed,which could grow on hypoxanthine, grew on allantoin assole nitrogen source (Antia et al., 1980). On the otherhand, Devi Prasad (1983) found that all freshwater algaeable to grow on hypoxanthine could also grow in mediacontaining allantoate after an acclimation period. Exceptfor the results obtained in our laboratory (Pineda et al.,1984 ; this study), only Oliveira & Huynh (1990) haveproved that allantoic acid can be a good nitrogen sourceto support growth of microalgae. These authors suggestthat the utilization of allantoate is catalysed in mostspecies by allantoicase. In addition, they reinforcedprevious observations that allantoin is a poor nitrogensource, compared with purines, to support phytoplanktongrowth (Antia et al., 1980). As Amman & Lynch (1964)suggested, this inability to support growth could be dueto a barrier that prevents allantoin uptake or the absenceof a specific transporter.
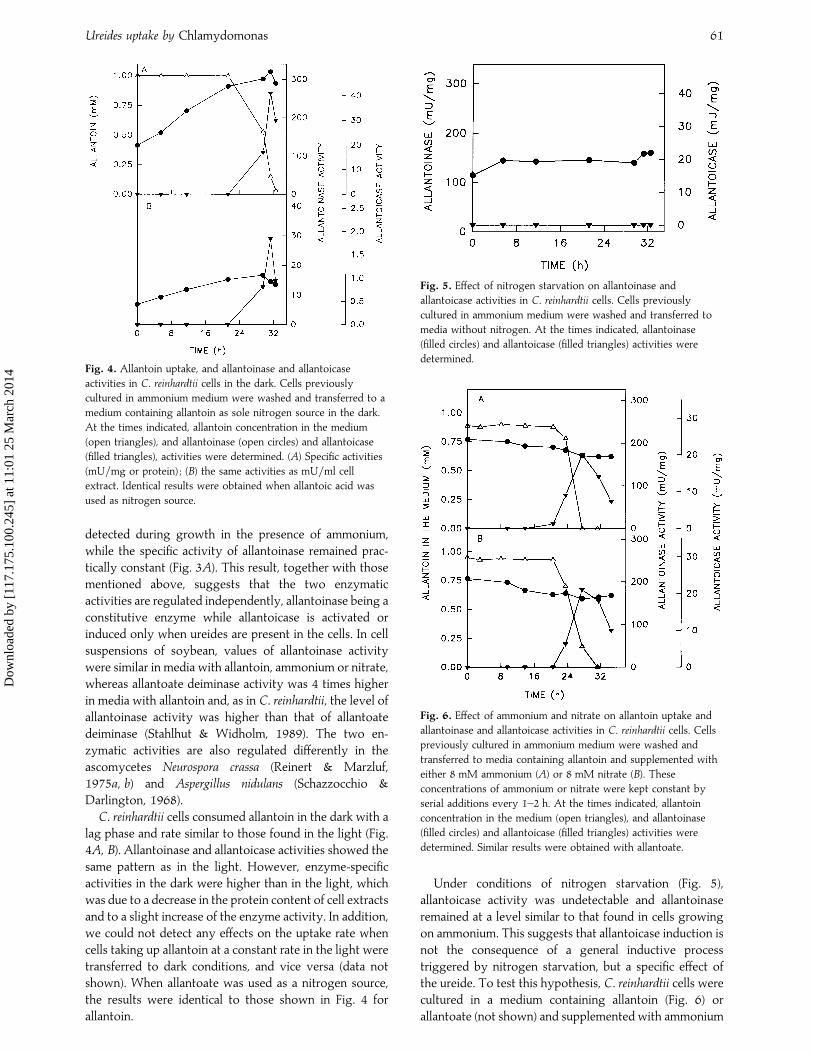
In C. reinhardtii cells, the enzymatic activities involvedin the catabolism of these ureides (allantoinase andallantoicase) changed in a similar way independently ofwhether the nitrogen source was allantoin or allantoate(Fig. 2). However, allantoinase behaved as a constitutiveenzyme whose specific activity increased when cells wereactively consuming allantoin (Fig. 2A) or allantoate (Fig.2B). Allantoicase behaved as an inducible enzyme whoseactivity was detectable only during the uptake of eitherureide (Fig. 2). These results demonstrate that allantointransport is not solely due to a metabolic drag promotedby allantoinase, since this enzyme is detectable at asignificant level without allantoin being taken from themedium (Fig. 2A). On the other hand, the two componentsof allantoate uptake (transport and intracellular enzymaticassimilation) seem to be perfectly coupled (Fig. 2B),making their experimental distinction difficult.
Allantoinase activity was also detected in cells culturedin media containing all the nitrogen sources studied –both inorganic (ammonium and nitrate) and the metabolicprecursors of ureides (adenine, xanthine, hypoxanthineand urate). Activity levels in these media were over 80%
Fig. 2. Allantoinase and allantoicase activities in C. reinhardtii
cells cultured in media containing allantoin or allantoate. Cells
cultured in media with 8 mM ammonium were washed and
transferred to media containing allantoin (A) or allantoate (B) as
sole nitrogen sources. At the times indicated, allantoin or
allantoate concentrations in the medium (open triangles), and
allantoinase (filled circles) and allantoicase (filled triangles)
activities were determined.
Fig. 3. Allantoinase and allantoicase activities in C. reinhardtii
cells cultured in media with ammonium as sole nitrogen source.
Cells were cultured with ammonium up to the late exponential
phase ; the media were then refreshed with sterile medium and
ammonium added at a final concentration of 8 mM. Ammonium
concentration in the medium (open triangles), and allantoinase
(filled circles) and allantoicase (filled triangles) activities were
determined at the times indicated.
of those found in cells grown with the ureides allantoinand allantoic acid. However, allantoicase activity was notdetected in cells that had been cultured in media withammonium or nitrate as sole nitrogen source, but it wasdetected when media contained a metabolic precursor ofureides as sole nitrogen source. To confirm these results,we studied the changes in allantoinase and allantoicaseactivities in cultures of C. reinhardtii with ammonium assole nitrogen source (Fig. 3). Allantoicase activity was not
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014
Ureides uptake by Chlamydomonas 61
Fig. 4. Allantoin uptake, and allantoinase and allantoicase
activities in C. reinhardtii cells in the dark. Cells previously
cultured in ammonium medium were washed and transferred to a
medium containing allantoin as sole nitrogen source in the dark.
At the times indicated, allantoin concentration in the medium
(open triangles), and allantoinase (open circles) and allantoicase
(filled triangles), activities were determined. (A) Specific activities
(mU}mg or protein) ; (B) the same activities as mU}ml cell
extract. Identical results were obtained when allantoic acid was
used as nitrogen source.
detected during growth in the presence of ammonium,while the specific activity of allantoinase remained prac-tically constant (Fig. 3A). This result, together with thosementioned above, suggests that the two enzymaticactivities are regulated independently, allantoinase being aconstitutive enzyme while allantoicase is activated orinduced only when ureides are present in the cells. In cellsuspensions of soybean, values of allantoinase activitywere similar in media with allantoin, ammonium or nitrate,whereas allantoate deiminase activity was 4 times higherin media with allantoin and, as in C. reinhardtii, the level ofallantoinase activity was higher than that of allantoatedeiminase (Stahlhut & Widholm, 1989). The two en-zymatic activities are also regulated differently in theascomycetes Neurospora crassa (Reinert & Marzluf,1975a, b) and Aspergillus nidulans (Schazzocchio &Darlington, 1968).
C. reinhardtii cells consumed allantoin in the dark with alag phase and rate similar to those found in the light (Fig.4A, B). Allantoinase and allantoicase activities showed thesame pattern as in the light. However, enzyme-specificactivities in the dark were higher than in the light, whichwas due to a decrease in the protein content of cell extractsand to a slight increase of the enzyme activity. In addition,we could not detect any effects on the uptake rate whencells taking up allantoin at a constant rate in the light weretransferred to dark conditions, and vice versa (data notshown). When allantoate was used as a nitrogen source,the results were identical to those shown in Fig. 4 forallantoin.
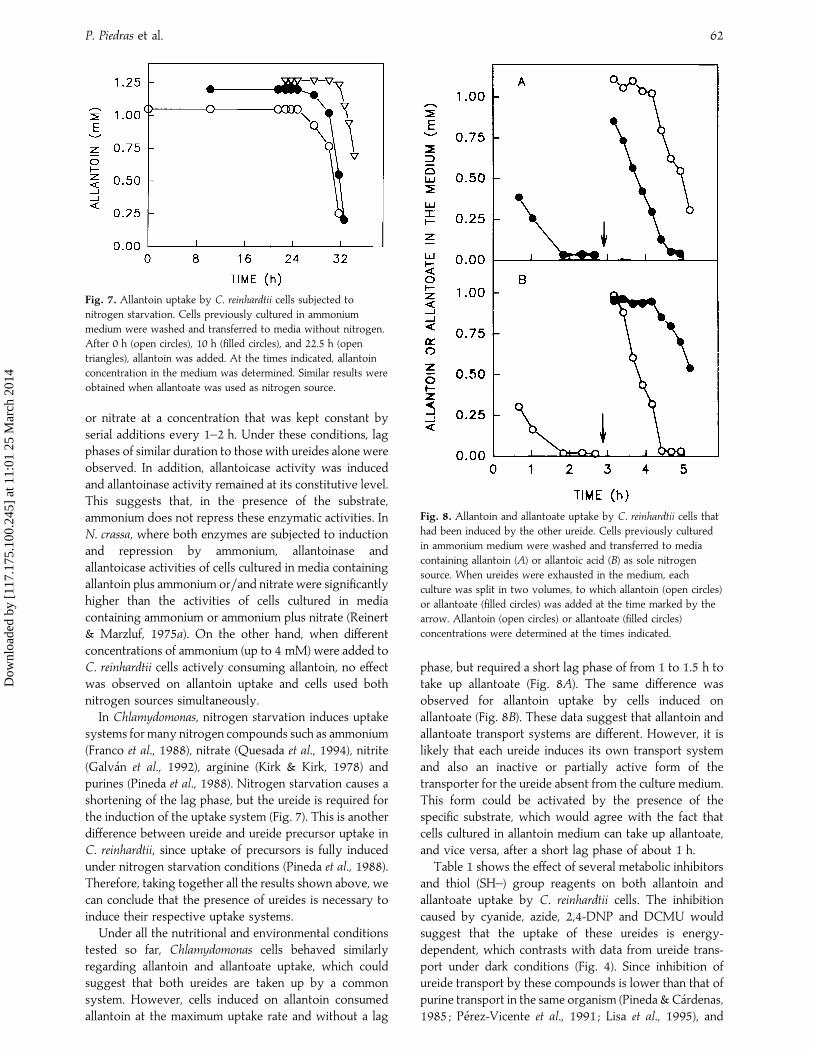
Fig. 5. Effect of nitrogen starvation on allantoinase and
allantoicase activities in C. reinhardtii cells. Cells previously
cultured in ammonium medium were washed and transferred to
media without nitrogen. At the times indicated, allantoinase
(filled circles) and allantoicase (filled triangles) activities were
determined.
Fig. 6. Effect of ammonium and nitrate on allantoin uptake and
allantoinase and allantoicase activities in C. reinhardtii cells. Cells
previously cultured in ammonium medium were washed and
transferred to media containing allantoin and supplemented with
either 8 mM ammonium (A) or 8 mM nitrate (B). These
concentrations of ammonium or nitrate were kept constant by
serial additions every 1–2 h. At the times indicated, allantoin
concentration in the medium (open triangles), and allantoinase
(filled circles) and allantoicase (filled triangles) activities were
determined. Similar results were obtained with allantoate.
Under conditions of nitrogen starvation (Fig. 5),allantoicase activity was undetectable and allantoinaseremained at a level similar to that found in cells growingon ammonium. This suggests that allantoicase induction isnot the consequence of a general inductive processtriggered by nitrogen starvation, but a specific effect ofthe ureide. To test this hypothesis, C. reinhardtii cells werecultured in a medium containing allantoin (Fig. 6) orallantoate (not shown) and supplemented with ammonium
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014
62P. Piedras et al.
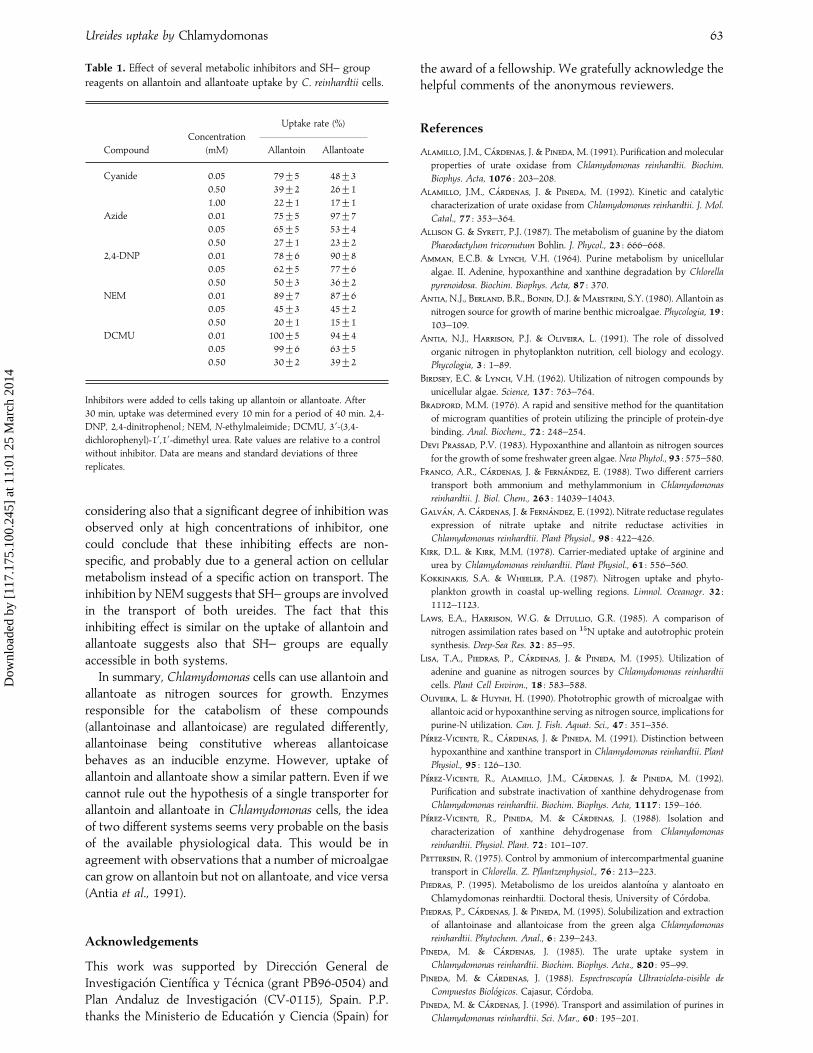
Fig. 7. Allantoin uptake by C. reinhardtii cells subjected to
nitrogen starvation. Cells previously cultured in ammonium
medium were washed and transferred to media without nitrogen.
After 0 h (open circles), 10 h (filled circles), and 22.5 h (open
triangles), allantoin was added. At the times indicated, allantoin
concentration in the medium was determined. Similar results were
obtained when allantoate was used as nitrogen source.
or nitrate at a concentration that was kept constant byserial additions every 1–2 h. Under these conditions, lagphases of similar duration to those with ureides alone wereobserved. In addition, allantoicase activity was inducedand allantoinase activity remained at its constitutive level.This suggests that, in the presence of the substrate,ammonium does not repress these enzymatic activities. InN. crassa, where both enzymes are subjected to inductionand repression by ammonium, allantoinase andallantoicase activities of cells cultured in media containingallantoin plus ammonium or}and nitrate were significantlyhigher than the activities of cells cultured in mediacontaining ammonium or ammonium plus nitrate (Reinert& Marzluf, 1975a). On the other hand, when differentconcentrations of ammonium (up to 4 mM) were added toC. reinhardtii cells actively consuming allantoin, no effectwas observed on allantoin uptake and cells used bothnitrogen sources simultaneously.
In Chlamydomonas, nitrogen starvation induces uptakesystems for many nitrogen compounds such as ammonium(Franco et al., 1988), nitrate (Quesada et al., 1994), nitrite(Galva! n et al., 1992), arginine (Kirk & Kirk, 1978) andpurines (Pineda et al., 1988). Nitrogen starvation causes ashortening of the lag phase, but the ureide is required forthe induction of the uptake system (Fig. 7). This is anotherdifference between ureide and ureide precursor uptake inC. reinhardtii, since uptake of precursors is fully inducedunder nitrogen starvation conditions (Pineda et al., 1988).Therefore, taking together all the results shown above, wecan conclude that the presence of ureides is necessary toinduce their respective uptake systems.
Under all the nutritional and environmental conditionstested so far, Chlamydomonas cells behaved similarlyregarding allantoin and allantoate uptake, which couldsuggest that both ureides are taken up by a commonsystem. However, cells induced on allantoin consumedallantoin at the maximum uptake rate and without a lag
Fig. 8. Allantoin and allantoate uptake by C. reinhardtii cells that
had been induced by the other ureide. Cells previously cultured
in ammonium medium were washed and transferred to media
containing allantoin (A) or allantoic acid (B) as sole nitrogen
source. When ureides were exhausted in the medium, each
culture was split in two volumes, to which allantoin (open circles)
or allantoate (filled circles) was added at the time marked by the
arrow. Allantoin (open circles) or allantoate (filled circles)
concentrations were determined at the times indicated.
phase, but required a short lag phase of from 1 to 1.5 h totake up allantoate (Fig. 8A). The same difference wasobserved for allantoin uptake by cells induced onallantoate (Fig. 8B). These data suggest that allantoin andallantoate transport systems are different. However, it islikely that each ureide induces its own transport systemand also an inactive or partially active form of thetransporter for the ureide absent from the culture medium.This form could be activated by the presence of thespecific substrate, which would agree with the fact thatcells cultured in allantoin medium can take up allantoate,and vice versa, after a short lag phase of about 1 h.
Table 1 shows the effect of several metabolic inhibitorsand thiol (SH–) group reagents on both allantoin andallantoate uptake by C. reinhardtii cells. The inhibitioncaused by cyanide, azide, 2,4-DNP and DCMU wouldsuggest that the uptake of these ureides is energy-dependent, which contrasts with data from ureide trans-port under dark conditions (Fig. 4). Since inhibition ofureide transport by these compounds is lower than that ofpurine transport in the same organism (Pineda & Ca! rdenas,1985 ; Pe! rez-Vicente et al., 1991 ; Lisa et al., 1995), and
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014
Ureides uptake by Chlamydomonas 63
Table 1. Effect of several metabolic inhibitors and SH– group
reagents on allantoin and allantoate uptake by C. reinhardtii cells.
Concentration
Uptake rate (%)
Compound (mM) Allantoin Allantoate
Cyanide 0.05 79³5 48³3
0.50 39³2 26³1
1.00 22³1 17³1
Azide 0.01 75³5 97³7
0.05 65³5 53³4
0.50 27³1 23³2
2,4-DNP 0.01 78³6 90³8
0.05 62³5 77³6
0.50 50³3 36³2
NEM 0.01 89³7 87³6
0.05 45³3 45³2
0.50 20³1 15³1
DCMU 0.01 100³5 94³4
0.05 99³6 63³5
0.50 30³2 39³2
Inhibitors were added to cells taking up allantoin or allantoate. After
30 min, uptake was determined every 10 min for a period of 40 min. 2,4-
DNP, 2,4-dinitrophenol ; NEM, N-ethylmaleimide ; DCMU, 3«-(3,4-dichlorophenyl)-1«,1«-dimethyl urea. Rate values are relative to a control
without inhibitor. Data are means and standard deviations of three
replicates.
considering also that a significant degree of inhibition wasobserved only at high concentrations of inhibitor, onecould conclude that these inhibiting effects are non-specific, and probably due to a general action on cellularmetabolism instead of a specific action on transport. Theinhibition by NEM suggests that SH– groups are involvedin the transport of both ureides. The fact that thisinhibiting effect is similar on the uptake of allantoin andallantoate suggests also that SH– groups are equallyaccessible in both systems.
In summary, Chlamydomonas cells can use allantoin andallantoate as nitrogen sources for growth. Enzymesresponsible for the catabolism of these compounds(allantoinase and allantoicase) are regulated differently,allantoinase being constitutive whereas allantoicasebehaves as an inducible enzyme. However, uptake ofallantoin and allantoate show a similar pattern. Even if wecannot rule out the hypothesis of a single transporter forallantoin and allantoate in Chlamydomonas cells, the ideaof two different systems seems very probable on the basisof the available physiological data. This would be inagreement with observations that a number of microalgaecan grow on allantoin but not on allantoate, and vice versa(Antia et al., 1991).
Acknowledgements
This work was supported by Direccio! n General deInvestigacio! n Cientı!fica y Te! cnica (grant PB96-0504) andPlan Andaluz de Investigacio! n (CV-0115), Spain. P.P.thanks the Ministerio de Educatio! n y Ciencia (Spain) for
the award of a fellowship. We gratefully acknowledge thehelpful comments of the anonymous reviewers.
References
A, J.M., C! , J. & P, M. (1991). Purification and molecular
properties of urate oxidase from Chlamydomonas reinhardtii. Biochim.
Biophys. Acta, 1076 : 203–208.
A, J.M., C! , J. & P, M. (1992). Kinetic and catalytic
characterization of urate oxidase from Chlamydomonas reinhardtii. J. Mol.
Catal., 77 : 353–364.
A G. & S, P.J. (1987). The metabolism of guanine by the diatom
Phaeodactylum tricornutum Bohlin. J. Phycol., 23 : 666–668.
A, E.C.B. & L, V.H. (1964). Purine metabolism by unicellular
algae. II. Adenine, hypoxanthine and xanthine degradation by Chlorella
pyrenoidosa. Biochim. Biophys. Acta, 87 : 370.
A, N.J., B, B.R., B, D.J. & M, S.Y. (1980). Allantoin as
nitrogen source for growth of marine benthic microalgae. Phycologia, 19 :
103–109.
A, N.J., H, P.J. & O, L. (1991). The role of dissolved
organic nitrogen in phytoplankton nutrition, cell biology and ecology.
Phycologia, 3 : 1–89.
B, E.C. & L, V.H. (1962). Utilization of nitrogen compounds by
unicellular algae. Science, 137 : 763–764.
B, M.M. (1976). A rapid and sensitive method for the quantitation
of microgram quantities of protein utilizing the principle of protein-dye
binding. Anal. Biochem., 72 : 248–254.
D P, P.V. (1983). Hypoxanthine and allantoin as nitrogen sources
for the growth of some freshwater green algae. New Phytol., 93 : 575–580.
F, A.R., C! , J. & F! , E. (1988). Two different carriers
transport both ammonium and methylammonium in Chlamydomonas
reinhardtii. J. Biol. Chem., 263 : 14039–14043.
G! , A. C! , J. & F! , E. (1992). Nitrate reductase regulates
expression of nitrate uptake and nitrite reductase activities in
Chlamydomonas reinhardtii. Plant Physiol., 98 : 422–426.
K, D.L. & K, M.M. (1978). Carrier-mediated uptake of arginine and
urea by Chlamydomonas reinhardtii. Plant Physiol., 61 : 556–560.
K, S.A. & W, P.A. (1987). Nitrogen uptake and phyto-
plankton growth in coastal up-welling regions. Limnol. Oceanogr. 32 :
1112–1123.
L, E.A., H, W.G. & D, G.R. (1985). A comparison of
nitrogen assimilation rates based on "&N uptake and autotrophic protein
synthesis. Deep-Sea Res. 32 : 85–95.
L, T.A., P, P., C! , J. & P, M. (1995). Utilization of
adenine and guanine as nitrogen sources by Chlamydomonas reinhardtii
cells. Plant Cell Environ., 18 : 583–588.
O, L. & H, H. (1990). Phototrophic growth of microalgae with
allantoic acid or hypoxanthine serving as nitrogen source, implications for
purine-N utilization. Can. J. Fish. Aquat. Sci., 47 : 351–356.
P! -V, R., C! , J. & P, M. (1991). Distinction between
hypoxanthine and xanthine transport in Chlamydomonas reinhardtii. Plant
Physiol., 95 : 126–130.
P! -V, R., A, J.M., C! , J. & P, M. (1992).
Purification and substrate inactivation of xanthine dehydrogenase from
Chlamydomonas reinhardtii. Biochim. Biophys. Acta, 1117 : 159–166.
P! -V, R., P, M. & C! , J. (1988). Isolation and
characterization of xanthine dehydrogenase from Chlamydomonas
reinhardtii. Physiol. Plant. 72 : 101–107.
P, R. (1975). Control by ammonium of intercompartmental guanine
transport in Chlorella. Z. Pflantzenphysiol., 76 : 213–223.
P, P. (1995). Metabolismo de los ureidos alantoı!na y alantoato en
Chlamydomonas reinhardtii. Doctoral thesis, University of Co! rdoba.
P, P., C! , J. & P, M. (1995). Solubilization and extraction
of allantoinase and allantoicase from the green alga Chlamydomonas
reinhardtii. Phytochem. Anal., 6 : 239–243.
P, M. & C! , J. (1985). The urate uptake system in
Chlamydomonas reinhardtii. Biochim. Biophys. Acta., 820 : 95–99.
P, M. & C! , J. (1988). EspectroscopıUa Ultravioleta-visible de
Compuestos BioloU gicos. Cajasur, Co! rdoba.
P, M. & C! , J. (1996). Transport and assimilation of purines in
Chlamydomonas reinhardtii. Sci. Mar., 60 : 195–201.
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014

64P. Piedras et al.
P, M., F! , E. C! , J. (1984). Urate oxidase of
Chlamydomonas reinhardtii. Physiol. Plant., 62 : 453–457.
P, M., C, P. & C! , J. (1987). Ammonium regulation of
urate uptake in Chlamydomonas reinhardtii. Planta, 171 : 496–500.
P, M., P! , R. & C! , J. (1988). Energy-dependent transport of
urate and xanthine in the unicellular green alga Chlamydomonas reinhardtii.
In Plasma Membrane Oxidoreductases in Control of Animal and Plant Growth
(Crane, F.L., Morre, D.J. & Low, H., editors), 209–217. Plenum Publishing,
New York.
Q, A., G! , A. & F! , E. (1994). Identification of nitrate
transporter genes in Chlamydomonas reinhardtii. Plant J., 5 : 407–419.
R, W.R. & M, G.A. (1975a). Regulation of the purine catabolic
enzymes in Neurospora crassa. Arch. Biochem. Biophys., 166 : 565–574.
R, W.R. & M, G.A. (1975b). Genetic and metabolic control of
the purine catabolic enzymes of Neurospora crassa. Mol. Gen. Genet., 139 :
39–55.
R, P.H.S., B, M.J., B, D.G., R, D.D. & S,
K.R. (1982). Ureide biogenesis in leguminous plants. Trends Biochem. Sci.,
7 : 366–368.
R, T.R. (1988). Homoeostasis in nitrogenous uptake}assimilation by
the green alga Platymonas (Tetraselmis) striata (Prasinphiceae). Ann. Bot.,
61 : 451–468.
S, C. & D, A.J. (1968). The induction and repression
of the enzymes of purine breakdown in Aspergillus nidulans. Biochim.
Biophys. Acta, 166 : 557–568.
S, R.W. & W, J.M. (1989). Ureide catabolism by soybean
(Glycine max (L.) Merrill) cell suspension cultures. II. Assimilation of
allantoin. J. Plant Physiol., 134 : 90–97.
S, N. (1960). Mitotic replication of deoxyribonucleic acid in
Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA, 46 : 83–91.
T, R.J., M, S.P. & S, L.E. (1983). Allantoinase from shoot
tissues of soybeans. Phytochemistry, 22 : 1117–1120.
T, E.M. (1985). Organic Geochemistry of Natural Waters. Nijhoff}Junk,
Dordrecht, The Netherlands.
V, S. (1955). Sur la pre! sence des enzymes des ure! ı$des glyoxyliques
chez les algues d’eau douce. C. R. Acad. Sci. Paris, 241 : 90–92.
V, S. (1958). Recherches sur la pre! sence des enzymes des ure! ı$ desglyoxyliques chez les algues marines. C. R. Acad. Sci. Paris, 246 :
1452–1454.
V, G.D. & D, C. (1970). Differential analysis of glyoxylate
derivatives. Anal. Biochem., 33 : 143–157.
V, G.D. & D, C. (1976). Degradation of purines and
pyrimidines by microorganisms. Bacteriol. Rev., 40 : 403–468.
Dow
nloa
ded
by [
117.
175.
100.
245]
at 1
1:01
25
Mar
ch 2
014




















