
The Chlamydomonas reinhardtii LI818 gene represents a distant relative of the cabI/II genes that is...
13
Plant Molecular Biology 32: 461-473, 1996. @ 1996 KluwerAcademic Publishers. Printed in Belgium. 461 The Chlamydomonas reinhardtii L1818 gene represents a distant relative of the cabl/ll genes that is regulated during the cell cycle and in response to illumination Fr6d6ric Savard, Christian Richard and Michel Guertin* Ddpartement de Biochimie, Facultd des sciences et de g~nie, Universitd Laval, Qudbec (Qudbec) G1K 7P4, Canada (*author for correspondence) Received 6 February 1996; accepted in revised form 17 June 1996 Key words: Chlamydomonas reinhardtii, chlorophyll a/b-binding protein gene, circadian rhythms, gene expression, light-regulated genes Abstract In the green unicellular alga Chlamydomonas reinhardtii, as in higher plants, the expression of the genes encoding the chlorophyll a/b-binding (CAB) polypeptides associated with photosystem I (PSI) and photosystem II (PSII) is regulated by endogenous (circadian clock) and exogenous signals (light and temperature). The circadian clock ensures that the oscillation in the levels of the different cab mRNAs is continuously kept in phase with light/dark (LD) cycles and is maximal by the middle of the day. On the other hand, light controls the amplitude of the oscillations. We report here the cloning and characterization of the C. reinhardtii LI818 gene, which identifies a CAB-related polypeptide and whose expression is regulated quite differently from the cab I/II genes. We show: (1) that in LD synchronized Chlamydomonas cells L1818 mRNA accumulation is subject to dual regulation that involves separable regulation by light and an endogenous oscillator; (2) that L1818 mRNA is fully expressed several hours before the cab I/II mRNAs and that the latter accumulate concomitantly; (3) that blocking the electron flow through PSII using DCMU prevents cells from accumulating cab I/II mRNAs but not L1818 mRNA and (4) that the accumulation of LI818 mRNA is abolished by blocking cytoplasmic protein synthesis, suggesting that these regulatory mechanisms are mediated by labile proteins. Introduction In Chlamydomonas, as in higher plants, the various types of CAB polypeptides assemble into the thylakoid membrane to form five light-harvesting complexes (LHC), (LHCI, LHCII, CP29, CP26 and CP24) which surround the two photochemical reaction centers of PSI and PSII and deliver captured light energy to them [4, 5]. Each of these complexes have different locations and roles in the two photosystems. LHCI polypeptides are exclusively associated with PSI while CP29, CP26 and CP24 polypeptides are specific to PSII. The most abundant ones, the LHCII polypeptides, serve as anten- nae for both photosystems; that is, under excess of light, a fraction of the LHCII dissociates from the PSII core and migrates close to the PSI to help to redistribute the light energy between PSI and PSII. CAB polypeptides show overall amino acid sequence homology in pairwise comparisons that ranges from 99% to 30%. However, all their hydro- pathy profiles show strong signals indicating the posi- tion in their sequences of three transmembrane helices. Of these, transmembrane helices I and III and stretches of residues preceding them are the most strongly con- served sequences among all CAB polypeptides. In fact, outside domains I and III, the different CAB poly- peptides have often little in common. The presence of these three transmembrane helices has now been con- firmed by the molecular structure of the pea LHCII [22]. Some of the highly conserved motifs are also found in other thylakoid membrane polypeptides of high- er plants and green algae, designated ELIE cbr and psbS [ 13, 23, 35]. These polypeptides represent distant
Transcript of The Chlamydomonas reinhardtii LI818 gene represents a distant relative of the cabI/II genes that is...

Plant Molecular Biology 32: 461-473, 1996. @ 1996 KluwerAcademic Publishers. Printed in Belgium.
461
The Chlamydomonas reinhardtii L1818 gene represents a distant relative of the cabl/ll genes that is regulated during the cell cycle and in response to illumination
Fr6d6r ic Savard , Chr i s t i an R i c h a r d and M i c h e l Guer t in* Ddpartement de Biochimie, Facultd des sciences et de g~nie, Universitd Laval, Qudbec (Qudbec) G1K 7P4, Canada (* author for correspondence)
Received 6 February 1996; accepted in revised form 17 June 1996
Key words: Chlamydomonas reinhardtii, chlorophyll a/b-binding protein gene, circadian rhythms, gene expression, light-regulated genes
Abstract
In the green unicellular alga Chlamydomonas reinhardtii, as in higher plants, the expression of the genes encoding the chlorophyll a/b-binding (CAB) polypeptides associated with photosystem I (PSI) and photosystem II (PSII) is regulated by endogenous (circadian clock) and exogenous signals (light and temperature). The circadian clock ensures that the oscillation in the levels of the different cab mRNAs is continuously kept in phase with light/dark (LD) cycles and is maximal by the middle of the day. On the other hand, light controls the amplitude of the oscillations. We report here the cloning and characterization of the C. reinhardtii LI818 gene, which identifies a CAB-related polypeptide and whose expression is regulated quite differently from the cab I/II genes. We show: (1) that in LD synchronized Chlamydomonas cells L1818 mRNA accumulation is subject to dual regulation that involves separable regulation by light and an endogenous oscillator; (2) that L1818 mRNA is fully expressed several hours before the cab I/II mRNAs and that the latter accumulate concomitantly; (3) that blocking the electron flow through PSII using DCMU prevents cells from accumulating cab I/II mRNAs but not L1818 mRNA and (4) that the accumulation of LI818 mRNA is abolished by blocking cytoplasmic protein synthesis, suggesting that these regulatory mechanisms are mediated by labile proteins.
Introduction
In Chlamydomonas, as in higher plants, the various types of CAB polypeptides assemble into the thylakoid membrane to form five light-harvesting complexes (LHC), (LHCI, LHCII, CP29, CP26 and CP24) which surround the two photochemical reaction centers of PSI and PSII and deliver captured light energy to them [4, 5]. Each of these complexes have different locations and roles in the two photosystems. LHCI polypeptides are exclusively associated with PSI while CP29, CP26 and CP24 polypeptides are specific to PSII. The most abundant ones, the LHCII polypeptides, serve as anten- nae for both photosystems; that is, under excess of light, a fraction of the LHCII dissociates from the PSII core and migrates close to the PSI to help to redistribute the light energy between PSI and PSII.
CAB polypeptides show overall amino acid sequence homology in pairwise comparisons that ranges from 99% to 30%. However, all their hydro- pathy profiles show strong signals indicating the posi- tion in their sequences of three transmembrane helices. Of these, transmembrane helices I and III and stretches of residues preceding them are the most strongly con- served sequences among all CAB polypeptides. In fact, outside domains I and III, the different CAB poly- peptides have often little in common. The presence of these three transmembrane helices has now been con- firmed by the molecular structure of the pea LHCII [22].
Some of the highly conserved motifs are also found in other thylakoid membrane polypeptides of high- er plants and green algae, designated ELIE cbr and psbS [ 13, 23, 35]. These polypeptides represent distant

462
relatives of CAB polypeptides and have recognizable homology only with the two central helices of CAB polypeptides. Remarkably, the psbS polypeptide is pre- dicted to have a fourth transmembrane helix [35]. ELIP polypeptides accumulate very rapidly in the thylakoid membrane during greening of etiolated plants where they may act as temporary substitutes for the LHC polypeptides [ 1 ]. Compelling evidence also exists for their participation in the cellular responses to light and osmotic stresses [ 1 ]. Interestingly, ELIP polypeptides show a high degree of homology with the cbr protein, a putative fl-carotene-binding protein from the green alga Dunaliella bardawil which accumulates concom- itantly with/3-carotenoids under conditions potentially engendering photooxydative damage, i.e. high light intensity and nutrient deprivation. Whether ELIP and cbr polypeptides bind chlorophyll has not been demon- strated. The psbS polypeptide associates with PSII, but is not involved in stress response. This 22 kDa protein represents a relative member to the CAB I/II and ELIP polypeptides and has been isolated in a complex with both chlorophyll a and b. It has been proposed a struc- tural role to bind extrinsic lumenal proteins.
Recently, we have shown that when the green unicellular alga Chlamydomonas eugametos is grown under a 12 h light/12 h dark regime, a restricted number of mRNAs accumulate transiently at the onset of the light period [7, 9]. One of these, the L1818e-1 mRNA, was further characterized and shown to be both under the control of a nonphotosynthetic photoreceptor and a circadian clock. By comparing the L1818e-1 predicted polypeptide sequence with proteins of known func- tion, we found that it has weak (ca. 30%) but signi- ficant overall homology with CAB polypeptides. Fur- ther amino acid sequence comparisons failed however to reveal significantly higher homology with any type of CAB potypeptides, suggesting that LI818e- l poly- peptide might represent a new type of CAB poly- peptide. Remarkably, it was found that L1818e-I and cabH mRNAs accumulate differently during the cell cycle. While the transcript level of LI8t8e-1 mRNA peaks after 1-2 h in the light phase, the cabll mRNA abundance increases over basal level after a 2 h lag and reaches a maximum towards the 6th hour.
We report here the presence of a small nuclear gene family in C. reinhardtii that encodes a polypeptide homologous to the C. eugametos LI818 protein. As C. reinhardtii and C. eugametos represent the two major lineages in a group of flagellates belonging to the highly diversified and polyphyletic genus Chlamydo- monas [6, 34], this strongly suggests that LI818 gene is
likely to be widely distributed in Chlamydomonas and possibly in other groups of green flagellates as well. It also indicates an evolutionarily conserved function.
Sequence comparisons indicated that L1818 genes encode polypeptides that are more related to CAB I/II polypeptides than to ELIE cbr and psbS polypeptides. However, despite their closer relationship to cabI/ll genes, L1818 genes are regulated quite differently dur- ing the cell cycle and in response to illumination, sug- gesting that LI818 polypeptides may play a function different from those played by CAB I / I I polypeptides.
Materials and methods
Strains and culture conditions
Synchronous liquid cultures of Chlamydomonas rein- hardtii mt + (CC620) were grown in minimal SG medi- um [ 14] with 1.0% CO2 on a 12h light/12h dark illumin- ation cycle in 4 liter bottles (internal diameter 15 cm). Cells were illuminated from both sides with cool-white fluorescent tubes (ca. 300 #E m -2 s-I) . Temperature inside the vessels varied between 19 and 23 °C.
Cell synchrony was checked by measuring cell density. In our conditions, cell population uniformely increases between the fourth and eight hour of the dark period. The first two L/D cycles produce fourfold divi- sions from 3 x 104 to 1.2 x 104 to 5 x 105 cells/ml. In the next cycle cell density doubles to reach l x 106 cells/ml and in the final cycle a last twofold division brings the cell density to ca. 2x 10 6 cells/ml.
Cells were harvested during the exponential phase of growth (ca. 5x 105 cells/ml) for analyses. In the experiments depicted in Figs. 6 and 8, cells were trans- ferred into 100 ml culture bottles (internal diameter 3 cm) just prior to the onset of the dark period.
Isolation of C. reinhardtii LI818r-3 cDNA clones
cDNA clones corresponding to C. reinhardtii L1818r-3 mRNA were isolated by screening a cDNA library pre- pared with poly(A) + RNA extracted from synchron- ized cells stimulated with light for 1 h. Construction of the cDNA library in the AZAP II vector (Strata- gene) was essentially as previously described [9]. A near full-length copy of L1818 mRNA was obtained and was thereafter named L1818r-3. The entire nucle- otide sequence of L1818r-3 cDNA was determined on both strands by the dideoxy method.

RNA extraction, northern blot analysis and mRNA quantification
RNA isolation and hybridizations with cDNA probes were done as previously described [9]. After autora- diography, the filters were exposed to a Fujix BAS IIIs imaging plate which was thereafter scanned with the Fujix BAS 1000 Bio-imaging Analyser System oper- ated with the Fujix MacBAS software. RNA samples were normalized on A260.
DNA isolation and Southern blot analysis
DNA isolation and Southern blot analysis were per- formed as previously described [9].
DNA probes
Lhcal*l cDNA [15] was a generous gift of Dr D. L. Herrin and represents a near full-length copy. cabll-1 cDNA [21 ] is a 300-350 bp DNA fragment which was kindly provided by Dr K. Kindle.
Results
Cloning of C. reinhardtii LI818 gene
To clone the C. reinhardtii LI818 gene, a AZAPII cDNA library was prepared with poly(A) RNA extrac- ted from cells harvested at the first hour of the light period, and probed with the C. eugametos LI818e- 1 cDNA [9]. From a primary screen of about 2x l0 s plaques, six positive and independent clones were iden- tified and further characterized. The size of the cDNA inserts was found to range from ca. 800 to 1500 bp. The largest insert, designated LI818r-3, appeared to represent a near full length copy as judged by northern blot analysis (not shown).
Sequencing of LI818r-3 cDNA indicated a 1542 bp insert containing a 5'-untranslated region (UTR) of 47 bp, a single large open reading frame (ORF) of 759 bp and a 3'UTR of 720 bp which includes the putative Chlamydomonas polyadenylation sequence, TGTAA, located 12 nucleotides upstream of a 16 bp poly(A) tail (Fig. 1A). Analysis of the LI818r-3 nucle- otide sequence predicted a polypeptide of 253 amino acids with a theoretical molecular mass of 27.5 kDa (Fig. I A). Inspection of the predicted amino acid sequence revealed that the first 30 residues would pos- sess the characteristic features of transit peptides of
463
chloroplastic precursors (Fig. IA). These are character- ized by a preponderance of positively charged amino acids and by a low content in acidic residues [19]. They are also enriched in hydroxylated amino acids and in small hydrophobic residues such as alanine [ 19]. In this regard, we noted that the sequence VSGKAG located at positions 30 to 35 in LI818r-3 polypeptide is very similar to the sequence VTMKAG found in the vicinity of the cleavage site of the C. reinhardtii Lhcal*l polypeptide precursor (positions 28 to 33) [15]. The Lhcal*l polypeptide precursor is cleaved between Met-30 and Lys-31.
Comparisons of sequence data revealed that LI818r-3 polypeptide shows ca. 60% overall identity to LI818e-1 polypeptide (Fig. 1B). This value rises to 64% when the putative transit peptides are excluded. Partial sequencing of the other cDNA clones revealed that they represent incomplete copies of the L1818r-3 mRNA.
At the nucleotide sequence level, L1818e-1 and L1818r-3 cDNAs show 64% identity over their cod- ing regions. No striking homology was noted when their 3'- and 5'-UTRs were compared.
LI818 polypeptides represent a new type of CAB polypeptides
Searching in protein data bases indicated that C. eugametos and C. reinhardtii polypeptides have weak but significant overall homology (<30%) with all known CAB polypeptides but failed to reveal signi- ficantly greater identity to any particular type of CAB polypeptides. In comparison, C. reinhardtii Lhcal* 1 and CABII-1 polypeptides [15, 17] show ca. 50% and ca. 75% overall identity respectively to their higher- plant counterparts.
Despite this weak overall identity to CAB poly- peptides, LI818 polypeptides are likely to be relatives of CAB polypeptides. As shown in the global align- ment depicted in Fig. 2A, they have two regions of strong homology with the different types of CAB poly- peptides, which correspond to the highly conserved transmembrane helices I and III and to a small segment preceding helix III. Furthermore, both polypeptides are predicted to possess six of the eight chlorophyll side- chain ligands (Fig. 2A) recently identified from the atomic model of the pea LHCII complex [22].
Finally, hydropathy plots of LI818 polypeptides predict a third hydrophobic region (Fig. 2B) that cor- responds to helix II. However, as shown in Fig. 2A, this region shows no significant homology at the amino

464
A agcttgcacct cct tgtgcacttcttcagccct tcaaccctaccaaaATGGCTATGATGA
M A M M M
TGCGCAAGGCCGCGGCTGTGCCCGCTTCTTCCCGCCGCTCGGTGGCTGTCAACTCCGTGA
R K A A A V P A S S R R S V A V N S V S
GCGGCAAGCGCACCGTGAGCGGCAAGGCCGGCGCCCCCGTGCCCGAGGACGTCCTGGCCT
G K R T V S G K A G A P V P E D V L A Y
ACGCCAAGACCCTGCCCGGTGTTACCGCTCCCTTCGACAACGTGTTCGACCCCGCTGGCT
A K T L P G V T A P F D N V F D P A G F
TCCTGGCCACTGCCTCCGTCAAGGATGTCCGTCGCTGGCGCGAGTCTGAGATCACCCACG
L A T A S V K D V R R W R E S E I T H G
GCCGCGTTGCTATGCTTGCCGCTCTGGGCTTCATCGTTGGCGAGCAGCTGCAGGACTTCC
R V A M L A A L G F I V G E Q L Q D F P
CCCTGTTCTTCAACTTCGACGGCCGCGTGTCTGGCCCCGCCATCTACCACTTCCAGCAGA
L F F N F D G R V S G P A I Y H F Q Q I
TCGGCCAGGGCTTCTGGGAGCCCCTGCTGATCGCCATCGGTGTGGCCGAGTCCTACCGTG
G Q G F W E P L L I A I G V A E S Y R V
TCGCCGTCGGCTGGGCCACCCCCACCGGCACCGGCTTCAACTCCCTGAAGGACGACTACG
A V G W A T P T G T G F N S L K D D Y E
AGCCCGGTGACCTGGGCTTCGACCCCCTGGGCCTCAAGCCCACCGACCCCGAGGAGCTGA
P G D L G F D P L G L K P T D P E E L K
AGAC C CTGCAGACCAAGGAGC TGAACAAC GGCC GC CTGGCCATGATC GC CATCGCCGCCT
T L Q T K E L N N G R L A M I A I A A F
TCGTGGCCCAGGAGCTGGTGGAGCAGAC CGAGATCTTCGAGCACCTGGTTCTGCGCTTCG
V A Q E L V E Q T E I F E H L V L R F E
AGAAGGAGGTTATCCTGGAGCTGGAGGACGTCGAGCGCGACCTGGGCCTGCCCCTCACCC
K E V I L E L E D V E R D L G L P L T P
CCCTGCCCGACAACCTCAAGGCCATCt aagtgcctct tcctcgatcgtgatct gc t t cac
L P D N L K A I
ggt t tggtcgcgggcggct tcggt tgc aatgtcgt tcatgggtggtacccggcgaacggt
gcggcgagcgcgggatggatggtgt agcgt tgt gtgagt cgctggtaat t t t t t t acaca
cgcgctgt tggcagaat tgtgtgacatgggatggggat tgacacgcgcgcacgtgcagca
cgcggcatgggcatgcggaaatgcgtgtcgcgggtgggtcctgggcagcgcaac tagaca
c tacggagcagagaat tct t t tgcgaacatatacaggagtgctgtgaatgtggttgagtg
c tgagcggaccgact tgcacgtgtgtggagtggtt agttgaagctgtggcgtc tc aggct
t tgcgatgcatccagcccggc t tgct tggctggcacggt tgcaagcgccaggc tgagagg
gccaaagaat aagagcgtgacagccggatgaggcgatgactgcaat t cgtgcatcgc tgt
acacacataagt t ttgacgctgtgagcgtgagtgcat cggatgtataagaaatgggctgc
tgt at at ttgct tgtgcgccctggt tgt t taacatgt at taagcatgtgct t t caaacac
atgctggat tcggat cgtatctt CCCCCt ctcatgggaagagaaagct caccacc t cagg
gtgatacgc t~t aacccggaaagatgaaaaaaaaaaaaaaaa
B
LI818e-i MAFTTKMQRSAVASRSSTVRVQAAGKTVSGSKTVSGGKTVGSASAESRRVAEVQAYLATL
LI818r-3 ---MAMMMRKAAAVPAS-SRRSVAVNSVSGKRTVSG ....... KAGAPVPEDVLAYAKTL
LISl8e-i PGCGVESGPFKGVWDPLSLAATATVGDVRRWRESEITHGRVAMLAALGFWGEQLEDFPA
L I 8 1 8 r - 3 PG---VTAPFDNVFDPAGFI~TASVKDV'RRWRESEITHGRV~LAALGFIVGEQLQDFPL
LI818e-I FFNFDGHITGQAIKQFDQVQQGFWEPLLIAIGLAESYRVSLGWATPTGTGFNNLKDEYDL
LI818r-3 FFNFDGRVSGPAIYHFQQIGQGFWEPLLIAIGVAESYRVAVGWATPTGTGFNSLKDDYEP ****** . ** . . ************ ****** *********** *** .
LI818e-i GNLYFDPLGLKPEDPEELRELQTKELNNGRLAMIAIAGFVLQEVAEPGVEIFQHLFFTIE
LIS18r-3 GDLGFDPLGLKPTDPEELKTLQTKELNNGRLAMIAIAAFVAQELVEQ-TEIFEHLVLRFE . . ******** ***** ***************** ** ** . *** ** .
LISlSe-i KDIVEEIDIIEKDLGLPPSFPVPTLPNLSSX
LISlSr-3 KEVILELEDVERDLGLP-LTPLP--DNLKAI . . . ***** . . **
Figure 1. Nucleotide sequence of L1818r-3 cDNA and deduced amino acid sequence. A. Nucleotide and deduced amino acid sequences of L1818r-3 cDNA. The putative polyadenylation site is underlined. B. Alignment of amino acid sequences encoding LI818r-3 and LI818e-I polypeptides. Analysis of the amino acid sequences was made using the Clustal program. * marks identical residues. Dot marks conservative
changes.

A
,~=,,~.~ . . . . . . . . . . . . . . . . . . . . . . . ~ . ~ , ~ , ~ i . . , . . . . . ~ ~ , ~ Lhca2* 1 SKY S TTPTARSATTVCVAADP~P - LWF PGSTP ..... -~Z~SL~P~P L~ILS! Lhca3* 1 - -RKASFVVRAASTP PV~QGAI~Q - LWFASKQS ...... L S ~ R I ~ P ~ - I Lhcb-m ......................... WLGPFSTN ..... ~ - ~ Y ~ ~ T ~ Lhcb-r ......................... WLGPYS~A- - --L -- ~ r ~ TA~AI r. ,hca~.*l . . . . . . . . . . . . . . . . . . . . . . r , ~ G ~ P ~ p ~ - : - : - : : - - ~ ~ L , lu
Lheb3*l ............. S ~ i D L W Y G P 1 3 R V K I ' L G P F S A Q ..... T ~ ~ A ~ _ _ _ Lhcb6* 1 ................ AAAAPKK~ - -WI PAVK LhcbS" 1 pAPVATS SAG I DI~LAKW~GP 13RR IYL P N G L ~ ~ ~
Z ....... LFSGP k ...... N L E R ~ i . . . . . . SLR ~GTGGFIEPKWLAY E ...... T
E ...... TFKRYRE k ...... SLKRFTR
m : : 2 : , :,r~,a,..~as
i ::::::
AL~ GRGD - - - I LG --- FI KI LN
~ G - - - • . . . . . . ~,~,~,,,~,,~,~,~,~,~ ~ ~
~ . : ~
ELIP-60 .............. E~LLSQLG .... ~G~ Lhcal*l WVKAQEWAAXP .... G~ATYLG - - QPVPI~ Lhcai* 1 TP---SWYT ...... EYFTD- - -TTT. Lhca3* 1 QETAIdKWF(~I~ P P ~ - - - NYT - Lhcb-m FGE PVWFKAGAQ I FSE~GLDYLGS pAL I ~A( Lhcb-r FGEAVWFKAGAQ I FSE~GLDYLGNP SLVII~ Lhcal*l ~LWA~/I~7 ~ ~ Z - ~ T I~F.
LhcblW 1 FG~A~KAGSQ I FS~GL D~LGNPNLV~ Lhcb3* 1 FK~pVWFKAGSQ I F S~GL DYLGNPNLVIIA~ Lhcb6*l ..... WFEAG - - -ADP~I~VAPFS .... FGT- LhcbS* 1 G PEAVWFKTGALLLDGNTLNYFGN - - - S I P I
I I
• - - QAWFAYTVAMLSX~L~ ~LPTILAIRFLAZ : : : : ~ , ~ w ~ ,,
IZVATLE
: : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : : :--s •
~ I ~ S M GKQYFLGL'GLGG . . . .
/NGGP .................... AGEG - e
/NGGP .................... AGGEG - e
/GGGP .................... LGEG [NGLPG ................... VG~ I
~/DFFDPDSQ SV~WAT p w ~ - S ~ N : G ~
~ , ~ . . . . . . . . . . . . . . . . . . . . . . ~ X L ~ W ~ - ~
RAG ....... ~SK ....... qGSG ...... ~GK- ...... hAD ....... hAD ....... F ~ - - .....
~ D . . . . . . .
ELIP-60 - -
Lhcal*l - -
Lhca2*l - -
Lhca3*l -- Lhcb-m - - Lhcb-r -- Lhcal*l --
Lhcbl* 1 -. Lhcb3*l Lhcb6*l lq] LhcbS*l
- - -AI
w
ZZX
~GLPEL~M~AATEIK~ ~ - -T~F I~V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
~ ---AI YLG GDVZIPKGIFPH ........ "-- . . . . . . . . . . . . . . . .
m~.loa~ - - - A I ~ ~ T S L K F H ................ I---SC TNNA~AAATKFTP SA ............
B F - - - ~ S ~ ,ATKFTPSA ............ i ---A ATIIGI SVPFF .............
: : - : . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ~ ' X T ~ : : - - - - - : , : - ± - - - ~ i ~ , ~ l - ~ ~ " ~ ~ x m r t , ca . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ~ - - - - .
~ - - - s m ~ r ~ , ~ Z ~ , Z m C ~ S ~ O S a ~ L L ~ V - ~ S a ~ S L . . . . . . . . .
465
B
lOO 2 0 0
I . . . . ' . . . . I . . . . ' . . . . I ' ' L 3 L1818r-3 I I 11 / I I I [ 3
W :: I I I I I I I I I I I . . . . I I I I I I~ - ' 3
I GRVAMLAALGFIVGEQL
II FWEPLLIAIGVAESYRVA
III GRLAMIAIAAFVAQEL
I O0 200
3 Lhcb3*l I , Ii III I- 3
A 0 A A A~A A , ~ "/" 0
I I I I I , , , , I , , , , , , , , , I , ,I -2
100 2 0 0
I GRWAMLGALGCIFDEVLE
II SILAVLGFQWLMGLVEGFR
III GRLAMFSMFGFFVQAI
Figure 2. Sequence analyses of LI818 polypeptides. A. Comparisons of the amino acid sequences of (1) ELIP-60 from Hordeum vulgare (barley); (2) Lhcal* 1 (Cab6A) from Lycopersicum esculentum (tomato); (3) Lhca2* 1 (Cab7) from Lycopersicum esculentum (tomato); (4) Lhca3* 1 (Cab8) from Lycopersicurn esculenturn (tomato); (5) Lhcb (Cab II) from C. moewusii; (6) Lhcb (Cab II-1) from C. reinhardtii; (7) Lhcal* 1 from C. reinhardtii; (8) LI818e-I from C. eugametos; (9) L1818r-3 from C. reinhardtii; (10) Lhcbl* 1 from Lemna gibba (swollen duckweed); (l 1) Lhcb3* 1 (Cab 13) from Lycopersicurn esculentum (tomato); (12) Lhcb6* 1 (CP24) from Spinacia oleracea (spinach) and (13) Lhcb5* 1 from Hordeurn vulgare (barley). The amino acid sequences of the pea Lhcb that span the thylakoid membrane are boxed [22]. Residues are shown in reverse type if nine of thirteen in a given match column are identically conserved. * marks chlorophyll side chain ligands and # marks charge-compensating arginines. LI818e-l and LI818r-3 sequences are shaded. Sequence data were taken directly from GenBank, EMBL and NBRF data libraries. B. Hydropathy plots of L1818r-3 (upper panel) and tomato Lhcb3* 1 (lower panel) polypeptides. Peaks I, II and III represent hydrophobic regions that may span the thylakoid membrane. The corresponding sequences are shown.

466
acid sequence level to helix II from other CAB poly- peptides.
LI818r-3 gene is part of a small multigene family
The genomic organization of the LI818r-3 gene in C. reinhardtii was investigated by first probing South- ern blots of nuclear DNA with the full-length LI818r-3 cDNA. As shown in Fig. 3A, under stringent condi- tions of hybridization and washing, numerous bands were obtained with restriction enzymes that do no cut the L1818r-3 cDNA (SacI, SalI and XhoI), indicating that LI818r-3 gene is part of a small multigene fam- ily. When a 450 bp EcoRI fragment covering part of the 3'UTR was used as a probe, unique bands were obtained, indicating that LI818r-3 gene can be discrim- inated amongst other LI818 genes (Fig. 3B). Recent experiments (our unpublished results) in which the 51UTR was used as a probe have shown that the LI818r- 3 gene is located in the 7 kb SacI fragment (Figs. 3A and B). The two additional faint bands of 9 and > 12 kb observed in the SacI digest are not always detected and presumably represent Ll818-related genes (Fig. 3B).
Under LD cycles, LI818r-3 mRNA level is controlled by light and an endogenous oscillator so as to reach maximal abundance towards the onset of the light period
As a first step in the characterization of the LI818r-3 gene expression, we measured LI818r-3 mRNA level in LD synchronized cells. As depicted in Fig. 4, under LD conditions L1818r-3 mRNA abundance starts to increase several hours prior to the end of the dark period, abruptly increases upon illumination and then decreases over the remaining light period to reach basal level soon after the onset of the dark period. Although Fig. 4 does not show a full 24 hour cycle, LI818 mRNA accumulation was found to recur at pre- cise intervals of 24 h (not shown). The amplitude of the L1818r-3 mRNA rhythm from peak to trough as determined by bio-imaging is about 50-100 fold. As also shown in Fig. 4, L1818r-3 mRNA level oscillates even when cells are kept in darkness at the onset of the normal light period. Note however that the amp- litude of the oscillation is considerably reduced, indic- ating a light-dependent increase. The magnitude of the light-dependent increase was found to vary from 5-20 fold between experiments. As the onset of the increase and decrease in L1818r-3 mRNA level do not coincide with that of the light or dark periods, this indicates that
Figure 3. Southern blot analyses of Chlamydomonas reinhardtii genomic DNA. A. DNA was cut with SacI, Sail, PstI, EcoRI and XhoI and hybridized to the L1818r-3 cDNA probe. B. The blot used in A was dehybridized and probed with an EcoRI fragment (Fig. 2A) covering the last 450 bp of the L1818r-3 cDNA. Position of DNA marker fragments (bp) are indicated at the left of panel A.
LI818r-3 mRNA oscillations are based on endogen- ous mechanisms. Identical results were obtained when the full-length cDNA was used as a probe (data not shown), indicating that all expressed LI818 genes are coordinately expressed.
The expression ofLI818 genes is not coordinated with that of the cabI/II genes
So far, two cab genes have been cloned and char- acterized in C. reinhardtii: the Lhcal*l gene which encodes a 20 kDa LHC I polypeptide and the cablI-1 gene which codes for a 28 kDa LHC II polypeptide [ 15, 17]. The Lhcal*l gene is present as a single-copy gene while the cablI-1 is part of a multigene family which comprises 7-8 members. Like their higher-plant coun- terparts [11, 20, 26, 27, 29, 32], Lhcal*l and cabH-1 mRNA levels oscillate with a periodicity of 24 h in LD

~o > m
t--
>
t~
100~
75
50-
25-
0
100-
75-
50
25
0
100
I
m ~ .,,.I.iTn~,..
50
25-
0 _;nil ,m ,l-i11!
I L I 8 1 8
- - L h c a l * l
m u m
- - c a b l l - 1
m
time (h)
Figure 4. Accumulation patterns of L1818r-3, Lhcal* 1 and cabH-1 mRNAs in LD synchronized cells. Cell samples were harvested at the indicated times from the eighth hour of the second dark period. At the onset of the light period, the culture was split and aliquots were retained in darkness (black bar) or exposed to light (white bar). Total RNA was isolated and analysed (5 /zg) by northern blotting using L1818r-3, Lhcal*l and cabH-I cDNAs as probes. The same blot was used for all probes. Radioactive signals were quantified by bio-imaging. For each gene, transcript levels are expressed relative to the highest point in the experiment.
conditions, reaching a maximum 6 to 8 h after the onset of the light period and decreasing thereafter [16, 18, 28, 30]. As this timing of expression with respect to the phase of the day is a hallmark of cab gene expression, we also examined the time scale of Lhcal*l and cabH- 1 mRNA accumulation in CC620 cells. As shown in
467
Fig. 4, cells entering the light period also accumu- late more Lhcal* 1 and cabH-1 mRNAs than darkened cells but after a lag period, at a time when the LI818 mRNA level has already started to decrease. There- after, both mRNAs increase concomitantly to reach a maximum around the 8th hour of the light period. As previously reported by others, we found that cabll- 1 mRNA abundance also accumulates when cells are kept in darkness at the onset of the normal light peri- od. The absence of continuous oscillation of Lhcal*l mRNA in the dark, however, differs from the data of Huang and Herrin [ 16]. This could be due to the growth of cells in minimal media in our experiments, which is expected to lead to energy deprivation in prolonged darkness, thereby inhibiting Lhcal*l gene expression. But, it could also be related to the fact that different strains were used (see Discussion).
C. reinhardtii LI818 mRNA level oscillates only in LD conditions
To see whether the oscillations in LI818 mRNA level are related to a circadian rhythm, we measured tran- script level when LD synchronized cells are main- tained in constant darkness (DD) during the normal light period. A property of circadian rhythms is that their oscillations persist with a periodicity close to 24 h in environmental conditions that do not provide time cues to cells, although more or less pronounced damp- ing is often observed [8]. As can be seen in Fig. 5A, we found that the LI818 mRNA level stops oscillating after one subjective day in darkness, suggesting that the fluctuations in LI818 mRNA level are not based on a circadian rhythm.
To ensure that circadian oscillations do occur in CC620 cells, we examined the accumulation of cabll-1 mRNA. Recent investigations have indeed shown that cabH-1 mRNA shows overt circadian oscillations in DD conditions [28]. As expected, we could detect free- running oscillations in cablI-1 mRNA with a period close to 24 h (Fig. 5A), which demonstrates that the circadian oscillator is operating.
We also measured LI818 mRNA level in LD syn- chronized cells maintained phototrophically in con- stant dim light (LL) (ca. 10/zE m -2 s-1 ); the rationale being that light could control the amplitude of the LI818 mRNA oscillations. We chose dim light, because under these conditions cell growth is greatly reduced (doubling time >48 h) so that oscillations related to cell cycle events can still be distinguished from those based on a circadian oscillator. Again and still in contrast to

468
A B
~/ ' / / . .~ / / / / / / . ; , - /Z . ( / / / / / /~
~° I ~ ~o I I I u818 oo I I I r oot I ' ' 4ol M i i i i i J J
~ / I I I I X 1 I ~ I I '
#a 0 ,
80
60
40
20
Joo o (=" I 0 0
> 8 0
~ 60
40
09 ¢-
20
cabl l - 1
I
r 4
0 0
D8 DI2 4 8 12 16 20 24 28 32 36 40 44 48 D8 D12 2 4 6 8 10 12 14 17 20 22 24 26 28 32 34 36
t ime (h) t ime (h)
Figure 5. In contrast to the cabH-I mRNA level, LI818 mRNA level oscillates only in LD conditions. A. At the end of the second dark period, a synchronized culture was maintained in darkness for 48 h. B. At the end of the second dark period, a synchronized culture was maintained in dim light (10 #E m -2 s -I ) for 36 h. Total RNA was isolated at the indicated times and analysed (5/~g) by northern blotting using L1818r-3 and cabH-I cDNAs as probes. The same blot was used for both probes. Radioactive signals were quantitfied by bio-imaging. The hatched bar indicates that cells were maintained in darkness during the normal light phase (A) or that they were illuminated during the normal dark period (B). For each gene, transcript levels are expressed relative to the highest point in the experiment.
cabH-1 mRNA, we could not detect persisting oscilla- tions in the L1818 mRNA level (Fig. 5B). The better oscillation of cabll-1 mRNA in DD versus continuous dim light may seem surprising but it should be stressed that damping varies between experiments either in DD or LL conditions, so that Fig. 5A and 5B should not be compared. Variation in damping of cabll-1 mRNA rhythm has also been observed by others [ 18, 28].
LI818 mRNA level is increased by light independently of the phase of the day
As the LI818 mRNA level starts to decrease at about the same time in cells kept in the dark or exposed to light, we hypothesized that the endogenous oscilla- tion might delineate a time frame during which LI818 mRNA accumulates and is responsive to light. In tomato leaves, light sensitivity of cab genes is negat- ively controlled by the circadian clock and is restricted
to a short period of the day, being maximal at the time point of the night/day transition [2, 25].
To evaluate this possibility, LD synchronized cells were maintained in DD conditions from the onset of the dark period. From that point and every three hours after that, a cell sample was exposed to a 1 h light pulse and RNA was extracted from cells either exposed to light or kept in darkness during the flash. As shown in Fig. 6, we found that L1818 gene expression is strongly photoinducible at every stage of the circadian period, indicating that the endogenous oscillator does not gate the light sensitivity of the LI818 gene. The increase in the magnitude of the induction observed at the end of the day is reproducible but levels off thereafter (data not shown).

The light-dependent increase in LI818 mRNA level does not require active photosynthesis
Although the preceding experiments revealed the exist- ence of a light responsive pathway controlling the LI818 mRNA level, they were not informative regard- ing the nature of the photoreceptors involved. Indeed, the increase in LI818 mRNA level could depend on the activity of functional components of photosyn- thetic electron transport or on the activation of light- responsive photoreceptors. To address this question, a synchronized culture was kept in darkness or exposed to light at the onset of the light period in the presence or absence of 3-(3,4-dichlorophenyl)- 1,1 -dimethylurea (DCMU), an inhibitor of the electron flow through PSII. Addition of the inhibitor to synchronized cul- tures of Chlamydomonas cells causes growth and cell cycle progression to stop unless the cells have become committed to complete the cell cycle, this commitment occurs at the middle of the light period [24, 31 ].
As shown in Fig. 7, blocking electron trans- port through PSII does not affect the light-dependent increase in L1818 mRNA level but it considerably slows down the decline in mRNA; in this experiment, tran- script level measured after 8 h of illumination is 5-fold higher in DCMU treated cells. Thus, it appears that L1818 mRNA accumulation is not dependent on active photosynthetic electron transport. In contrast, adding DCMU was found to prevent cells from accumulating both Lhcal*l and cablI-1 mRNAs,
Cytoplasmic protein synthesis is required to promote LI818 mRNA accumulation
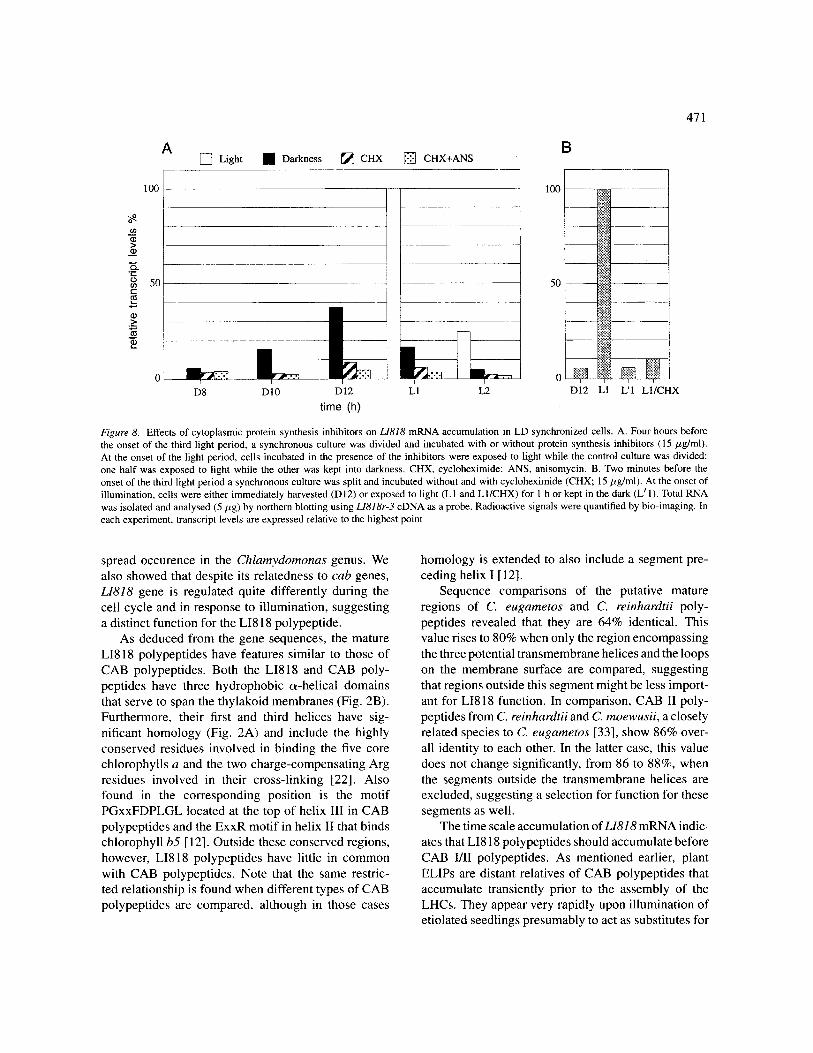
To determine whether the endogenous and light- dependent increases in LI818 mRNA abundance were dependent upon de novo protein synthesis, we inter- rupted the supply of protein molecules to the nucleo- cytoplasm and measured LI818 mRNA level. Cyclo- heximide, an inhibitor of the cytoplasmic ribosome, was added to a LD synchronized cell culture at the beginning of the induction period (at the 8th hour of the dark period) and RNA was extracted at different time points and analysed by northern blotting. The dose used (15 #g/ml) blocks protein synthesis by more than 95% within 5 min in C. reinhardtii [3].
As shown in Fig. 8, both the endogenous and light- dependent increases in L1818 mRNA level were almost completely suppressed in cells treated with cyclohex- imide. We repeated the experiment with a combination of two protein synthesis inhibitors, cycloheximide and
469
~ ' i f J i i l l l l l g i ~
co
(9
(9 o ) t--
>
100
80
60
40
20
i
I I
3 6 9 12 15 18 21 24
time (h)
Figure 6. L1818 mRNA level is increased by light independently of the phase of the circadian period, a. A LD synchronized culture was maintained in constant darkness from the onset of the second dark period. From this time and every 3 h after that, a cell sample was exposed to a 1 h light pulse (see Materials and methods). Total RNA was extracted from illuminated cells and from cells retained in the dark. RNA was analysed (5 #g) by northern blotting using L1818r-3 cDNA as a probe. Radioactive signals were quantified by bio-imaging. Transcript levels are expressed relative to the highest point in the experiment.
anisomycin. Both inhibitors block polypeptide chain elongation by interacting with different components of the ribosome [ 10] together causing nearly total inhibi- tion [3]. This experiment showed that the combination of anisomycin and cycloheximide slightly enhanced the inhibition of L1818 mRNA accumulation.
Thus the regulatory mechanisms controlling the timing of LI818 mRNA expression require de novo synthesis of labile proteins. However, because of the very long time that the cells have been in the presence of the inhibitor by the time they were exposed to light, one cannot distinguish between the turnover of a pro- tein that was present in the dark period or the failure to synthesize a protein in response to light. To distinguish between these possibilities, inhibitor was added at the onset of the light period. After 1 h, cellular RNA was extracted and analysed by hybridization. As shown in Fig. 8, blocking protein synthesis at the time cells enter the light period almost completely abolishes the light-

4 7 0
m
125
100
7 5 -
50 -
25
0 -
125
~ 100
*-" 75-
-.~
"~ o -
! ii~iii
iii
LI818 !,iii
ii
....... I i '?ig
! .......
:~'> i
125 -
100
75
50
25
0
Lhca 1 * 1
l c ; , ,
cab l l - 1
012 L1 L2L3 L4 L6 L8 L1 L2 L3 L4 L6 L8 LI L2 L3 L4L6 L8
time (h)
Figure 7. Effect of blocking electron flow through PSII on the light-dependent accumulation of L1818, Lhcal*l and cabH-I mRNAs in LD synchronized C. reinhardtii cells. Twenty minutes before the end of the second dark period a LD synchronized culture was split in three parts and one culture (No. 1) received DCMU (10/zM). At the onset of the light period, cultures 1 and 2 were exposed to light while culture 3 was maintained in darkness. Total RNA was isolated at the indicated times and analysed (5/~g) by northern blotting using L1818r-3, Lhcal* 1 and cabH-1 cDNAs as probes. The same blot was used for all probes. Radioactive signals were quantified by bio-imaging. Transcript levels are
expressed relative to the highest point in the experiment.
dependent increase, indicating that it depends on de
novo protein synthesis. Discussion
In this report we presented evidence for the existence of a new type of cab-re la ted gene that should be of wide-
471
A [ ] Light • Darkness [ ] CHX [ ] CHX+ANS
B
>
. B
O ffl
100
50
• . I I m
D8 D10 D12 L1 L2 time (h)
100 ~
5o iN
D12 L1 L'I LI/CHX
Figure 8. Effects of cytoplasmic protein synthesis inhibitors on L1818 mRNA accumulation in LD synchronized cells. A. Four hours before the onset of the third light period, a synchronous culture was divided and incubated with or without protein synthesis inhibitors (15 #g/ml). At the onset of the light period, cells incubated in the presence of the inhibitors were exposed to light while the control culture was divided: one half was exposed to light while the other was kept into darkness. CHX, cycloheximide; ANS, anisomycin. B. Two minutes before the onset of the third light period a synchronous culture was split and incubated without and with cycloheximide (CHX; 15/zg/ml). At the onset of illumination, cells were either immediately harvested (D 12) or exposed to light (L 1 and L 1/CHX) for 1 h or kept in the dark (L ~ 1). Total RNA was isolated and analysed (5 #g) by northern blotting using L1818r-3 cDNA as a probe. Radioactive signals were quantified by bio-imaging. In each experiment, transcript levels are expressed relative to the highest point
spread occurence in the Chlamydomonas genus. We also showed that despite its relatedness to cab genes, L1818 gene is regulated quite differently during the cell cycle and in response to illumination, suggesting a distinct function for the LIB 18 polypeptide.
As deduced from the gene sequences, the mature LI818 polypeptides have features similar to those of CAB polypeptides. Both the LI818 and CAB poly- peptides have three hydrophobic c~-helical domains that serve to span the thylakoid membranes (Fig. 2B). Furthermore, their first and third helices have sig- nificant homology (Fig. 2A) and include the highly conserved residues involved in binding the five core chlorophylls a and the two charge-compensating Arg residues involved in their cross-linking [22]. Also found in the corresponding position is the motif PGxxFDPLGL located at the top of helix III in CAB polypeptides and the ExxR motif in helix II that binds chlorophyll b5 [ 12]. Outside these conserved regions, however, LI818 polypeptides have little in common with CAB polypeptides. Note that the same restric- ted relationship is found when different types of CAB polypeptides are compared, although in those cases
homology is extended to also include a segment pre- ceding helix I [ 12].
Sequence comparisons of the putative mature regions of C. eugametos and C. reinhardtii poly- peptides revealed that they are 64% identical. This value rises to 80% when only the region encompassing the three potential transmembrane helices and the loops on the membrane surface are compared, suggesting that regions outside this segment might be less import- ant for LI818 function. In comparison, CAB II poly- peptides from C. reinhardtii and C. moewusii, a closely related species to C. eugametos [33], show 86% over- all identity to each other. In the latter case, this value does not change significantly, from 86 to 88%, when the segments outside the transmembrane helices are excluded, suggesting a selection for function for these segments as well.
The time scale accumulation ofLI818 mRNA indic- ates that LI818 polypeptides should accumulate before CAB I/II polypeptides. As mentioned earlier, plant ELIPs are distant relatives of CAB polypeptides that accumulate transiently prior to the assembly of the LHCs. They appear very rapidly upon illumination of etiolated seedlings presumably to act as substitutes for

472
the LHC polypeptides and to protect the newly synthes- ized photoreaction centers against photooxydation. In mature green leaves, ELIP synthesis is also observed but under light conditions that result in photoinhibition. Whether LI818 polypeptide could play such a role is not known, but it must be mentioned that ELIP poly- peptides are even more distant relatives of the LI818 polypeptides than are the LHC polypeptides.
Experiments in which darkened synchronized cul- tures of C. reinhardtii were exposed to light pulses revealed that L1818 gene represents an acute and posit- ive cellular response to illumination (Fig. 6). As these responses can be elicited throughout the circadian peri- od, this rules out the possibility that light regulation superimposes on the endogenous rhythm and indic- ates that LI818 gene is subject to dual regulation that involves separable regulation by light and an endogen- ous oscillator. This is in contrast with the situation observed in higher plants where light positively regu- lates the expression of the cab genes but is effective only during a restricted period of the day that is gated by the circadian oscillator [2, 25].
As shown in Fig. 4, cells entering the light phase also show a light-dependent increase in Lhcal*l and cabH-1 mRNAs but after a lag period. In this respect, it is noteworthy that under illumination conditions that do not support growth (i.e.darkness in minimal media) or support slow (dim light) phototrophic growth, cabll- 1 mRNA reaches maximal abundance more rapidly (Figs. 4, 5A and 5B), suggesting that light conditions favoring cell growth may delay the period of transcript accumulation.
Additional experiments (Fig. 7) showed that inter- rupting electron flow through PS II with DCMU pre- vents the Lhcal*l and cablI-1 mRNA accumulation associated with chloroplast development, suggesting that light induces increased cabI/II transcript abund- ance through the light reactions of photosynthesis. These results differ somewhat from those previously reported by Kindle who found that DCMU only par- tially inhibits the light-dependent increase in cabH-1 transcript [21 ]. However, those experiments were per- formed under lower light intensity (30 #E m -2 s - l). It is possible that strong light forces cells to adjust to their light environment by decreasing cabl/II mRNA levels and that DCMU prevents their daily increase by block- ing growth recovery. In contrast, the accumulation of L1818 mRNA remains unchanged or slightly increased when the electron flow is interrupted, indicating that it is regulated by nonphotosynthetic photoreceptors.
Is LI818 gene under the control of the circadian oscillator in C. reinhardtii?
To establish whether an endogenous rhythm is gener- ated by a circadian oscillator three phenomenological criteria must be met. Firstly, it must persist in constant environmental conditions (light or darkness and tem- perature) with a period that is close to 24 h. Secondly, its periodicity must be exactly 24 h when time cues such as LD cycles are provided. Thirdly, it must be temperature compensated which means that its period must remain relatively unchanged within a physiolo- gical range of temperature.
Although LI818 mRNA oscillations fulfill the second criterion, they fail to persist in DD or LL condi- tions (Fig. 5), suggesting (sensu stricto) that the endo- genous timing mechanism driving L1818 oscillations is not the circadian clock. This contrasts with the situation found in C. eugametos where LI818 mRNA accumu- lation shows persisting oscillations in DD conditions [9]. As we observed overt rhythms in cabll-1 mRNA, we can exclude the possibility that the circadian oscil- lator is not operating in C. reinhardtii CC620. It is possible, however, that in C. reinhardtii the L1818r-3 mRNA rhythm is controlled by the circadian clock but under permissive conditions (i.e., when cell growth and division is allowed). In this respect, it is note- worthy that in contrast to our observations no continu- ous oscillations in cabH-1 mRNA abundance have been measured in C. reinhardtii CC625 kept in darkness in minimal media. Persisting rhythms of cabll-1 mRNA in CC625 strain are detected either in LL conditions (dim light/minimal media) [18] or darkness (minimal media supplemented with acetate) [28]. These results may indicate that in different strains cell growth and division may be coupled with different strength to the circadian clock. This could also explain our failure to measure rhythms of Lhca 1" 1 mRNA in DD conditions.
Acknowledgements
This research was supported by a grant from the Nat- ural Sciences and Engineering Research Council of Canada. ES. is supported by a scholarship from the 'Le Fonds pour la Formation de Chercheurs et l 'Aide
la Recherche'.

473
References
I. Adamska I, Kloppstech K: The role of early light-induced proteins (ELIPS) during light stress. In: Baker NR, Bowyer JR (eds) Photoinhibition of Photosynthesis, pp. 205-219. Bios Scientific, Oxford (199~).
2. Anderson SL, Kay SA: Functional dissection of circadian clock- and phytochrome-regulated transcription of the Ara- bidopsis CAB2 gene. Proc Natl Acad Sci USA 92:1500-1504 (1995).
3. Baker E, Liggit P: Accelerated poly(A) loss and mRNA stabil- ization are independent effects of protein synthesis inhibition on c~-tubulin mRNA in Chlamydomonas. Nucl Acids Res 21: 2237-2246 (1993).
4. Bassi R, Soen S, Frank G, Zuber H, Rochaix JD: Characteriza- tion of chlorophyll-a/b binding proteins of photosystem I from Chlamydomonas reinhardtii. J Biol Chem 267:25714-25721 (1992).
5. Bassi R, Wollman FA: The chlorophyll-a/b binding proteins of photosystem II in Chlamydomonas reinhardtii. Planta 183: 423-433 (1992).
6. Buchheim MA, Turmel M, Zimmer EA, Chapman RL: Phylo- geny of Chlamydomonas (Chlorophyta) based on cladistic ana- lysis of nuclear 18S rRNA sequence data. J Phyco126:689-699 (1990).
7. Couture M, Chamberland H, St-Pierre B, Lafontaine J, Guertin M: Nuclear genes encoding chloroplast hemoglobins in the unicellular green alga Chlamydomonas eugametos. Mol Gen Genet 243:185-197 (1994).
8. Dunlap JC: Closely watched clocks: molecular analysis of circadian rhythms in Neurospora crassa. Trends Genet 6: 159- 165 (1990).
9. Gagn6 G, Guertin M: The early genetic response to light in the green unicellular alga Chlamydomonas eugametos grown under light/dark cycles involves genes that represent direct responses to light and photosynthesis. Plant Mol Biol 18: 429- 445 (1992).
10. Gale EF, Cundliffe E, Reynolds PE, Richmond MH, Waring MJ: Antibiotic inhibitors of ribosome function. In: Ltd JWS (ed) The Molecular Basis of Antibiotic Action, pp. 278-366. Wiley-lnterscience, London (1972).
11. Giuliano G, Hoffman NE, Ko K, Scolnik PA, Cashmore AR: A light-entrained circadian clock controls transcription of several plant genes. EMBO J 7:3635-3642 (1988).
12. Green BR, Kiihlbrandt W: Sequence conservation of light- harvesting and stress-response proteins in relation to the three- dimensional molecular structure of LHCII. Photosynth Res 44: 139-148 (1995).
13. Grimm B, Kruse E, Kloppstech K: Transiently expressed early light-inducible thylakoid proteins share transmembrane domains with light-harvesting chlorophyll binding proteins. Plant Mol Biol 13:583-593 (1989).
14. Harris E: The Chlamydomonas Sourcebook. Academic Press, San Diego, CA (1989).
15. Hwang S, Herrin DL: Characterization of a cDNA encod- ing the 20-kDa photosystem I light-harvesting polypeptide of Chlamydomonas reinhardtii. Curr Genet 23: 512-517 ( 1993 ).
16. Hwang S, Herrin DL: Control of Lhcal*l gene transcription by the circadian clock in Chamydomonas reinhardtii. Plant Mol Biol 26:557-569 (1994).
17. lmbault P, Wittemer C, Johanningmeier U, Jacobs JD, Howell SH: Structure of the Chlamydomonas reinhardtii cablI-I gene encoding a chlorophyll a/b-binding protein. Gene 73:397-407 (1988).
18. Jacobshagen S, Johnson CH: Circadian rhythms of gene expression in Chlamydomonas reinhardtii: circadian cycling of mRNA abundances of cab II, and possibly of beta-tubulin and cytochrome c. Eur J Cell Biol 64: 142-152.
19. Keegstra K, Olsen LJ, Theg SM: Chloroplastic precursors and their transport across the envelope membranes. Annu Rev Plant Physiol Plant Mol Biol 40:471-501 (1989).
20. Kellmann JW, Merforth N, Wiese M, Pichersky E, Piechulla B: Concerted circadian oscillations in transcripts levels of nine- teen Lha/b (Lhc) genes in Lycopersicon esculentum (tomato). Mol Gen Genet 237:439--448 (1993).
21. Kindle KL: Expression of a gene for a light-harvesting chloro- phyll a/b-binding protein in Chlamydomonas reinhardtii: effect of light and acetate. Plant Mol Biol 9:547-563 (1987).
22. Ktihlbrandt W, Wang DN, Fujiyoshi Y: Atomic model of plant light-harvesting complex by electron crystallography. Nature 367:614-621 (1994).
23. Lers A, Levy H, Zamir A: Co-regulation of a gene homologous to early light-induced genes in higher plants and 13-carotene biosynthesis in the alga Dunaliella bardawil. J Biol Chem 266:13698-13705 (1991).
24. McAteer M, Donnan L, John PCL: The timing of division in Chlamydomonas. New Phytol 99:41-56 ((1985).
25. Meyer H, Thienel U, Piechulla B: Molecular characteriza- tion of the diurnal/circadian expression of the chlorophyll a/b- binding proteins in leaves of tomato and other dicotyledonous and monocotyledonous plant species. Planta 180:5-15 (1989).
26. Millar AJ, Kay SA: Circadian control of cab gene transcription and mRNA accumulation in Arabidopsis. Plant Cell 3: 541- 55O (1991).
27. Nagy F, Kay SA, Chua NH: A circadian clock regulates tran- scription of the wheat Lhc-I gene. Genes Devel 2 :376-382 (1988).
28. Nikaido S, Locke CR, Weeks DP: Automated sampling and RNA isolation at room temperature for measurements of cir- cadian rhythms in Chlamydomonas reinhardtii. Plant Mol Biol 26:275-284 (1994).
29. Paulsen H, Bogorad L: Diurnal and circadian rhythms in the accumulation and synthesis of mRNA for the light-harvesting chlorophyll a/b-binding protein in tobacco. Plant Physiol 88: 1104-1109 (1988).
30. Shepherd HS, Ledoigt G, Howell SH: Regulation of light- harvesting chlorophyll-binding protein (LHCP) mRNA accu- mulation during the cell cycle in Chlamydomonas reinhardtii. Cell 32:99-107 (1983).
31. Spudich JL, Sager R: Regulation of the Chlamydomonas cell cycle by light and dark. J Cell Biol 85:136-145 (1980).
32. Taylor WC: Transcriptional regulation by a circadian rhythm. Plant Cell 1 : 259-264 (1989).
33. Turmel M, Bellemare G, Lemieux C: Physical mapping of dif- ferences between the chloroplast DNAs of the interfertile algae Chlamydomonas eugametos and Chlamydomonas moewusii. Curr Genet 11:543-552 (1987).
34. Turmel M, Gutell RR, Mercier JP, Otis C, Lemieux C: Analysis of the chloroplast large subunit ribosomal RNA gene from 17 Chlamydomonas taxa: three internal transcribed spacers and 12 group I intron insertion sites. J Mol Biol 232:446-467 (1993).
35. Wedel N, Klein R, Ljungberg U, Andersson B, Herrmann RG: The single-copy gene psbS codes for a phylogenetically intriguing 22 kDa polypeptide of photosystem II. FEBS Lett 314:61-66 (1992).




















