
Up-regulation of the vitamin C transporter SVCT2 upon differentiation and depolarization of myotubes
7
Up-regulation of the vitamin C transporter SVCT2 upon differentiation and depolarization of myotubes Marcela Low, Daniel Sandoval, Bryan Morales, Francisco Nualart, Juan Pablo Henríquez ⇑ Department of Cell Biology, Faculty of Biological Sciences, Universidad de Concepcion, Concepcion, Chile article info Article history: Received 1 September 2010 Revised 30 November 2010 Accepted 14 December 2010 Available online 19 December 2010 Edited by Berend Wieringa Keywords: Sodium-vitamin C co-transporter 2 Ascorbate Myogenesis Fiber-type Skeletal muscle Depolarization abstract In addition to its role as a strong antioxidant, vitamin C regulates the differentiation of several cell lineages. In vertebrate skeletal muscle, the vitamin C transporter SVCT2 is preferentially expressed in slow muscle fibers. To gain insights into the possible involvement of intracellular vitamin C on early myogenesis, we investigated the regulation of SVCT2 expression in cultures of chick fetal myo- blasts. SVCT2 expression increases in cultures of both, slow and fast muscle-derived myoblasts, as they fuse to form mainly fast myotubes. Interestingly, we found that SVCT2 could be positively mod- ulated by potassium-induced depolarization of myotubes. These findings suggest that SVCT2-medi- ated uptake of vitamin C could play diverse roles on skeletal muscle development and physiology. Ó 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction The formation of skeletal muscle during development involves a series of sequential events, including the commitment of somite- derived precursor cells to mononucleated myoblasts, and their subsequent fusion into multinucleated myotubes [1]. Mature skeletal muscles consist of variable proportions of fast and slow myofibers which, ultimately, account for their contractile activity [2]. Fiber types can be classified according to various parameters, such as the expression of type-specific protein isoforms and meta- bolic properties. Indeed, whereas most fast fibers are glycolytic, slow myofibers rely on oxidative metabolism for a permanent sup- ply of ATP [2]. Both mammalian and avian model systems have been em- ployed to identify the intrinsic properties and extrinsic factors that determine the characteristics of mature fibers [2,3]. Whereas mur- ine myoblasts seem preprogrammed to a particular phenotype [4], the definition of skeletal muscle fiber types in the chick relies on the patterning of muscle precursors, but also significantly on their innervation [5]. Vitamin C is the most important water-soluble antioxidant vita- min in the plasma and, as such, it has been characterized as one of the main non-enzymatic antioxidative systems in various tissues [6,7]. To accomplish intracellular functions, vitamin C must be effi- ciently incorporated into cells. Thus, whereas oxidized vitamin C, dehydroascorbic acid, is taken up by facilitative hexose transport- ers [8,9], two isoforms of high affinity sodium-dependent vitamin C co-transporters (SVCT1 and 2) mediate the uptake of ascorbic acid, the reduced form of vitamin C [10,11]. In addition to its role as a potent antioxidant, cumulative evi- dence supports that vitamin C positively regulates the differentia- tion of diverse cell lineages, such as osteoblasts, keratinocytes, and cells of the nervous system [12–15]. Interestingly, these effects of vitamin C are accompanied by dynamic expression patterns of their SVCT transporters [15–18], thus supporting the notion that transporter-mediated uptake of vitamin C is involved in cell differentiation. We have recently conducted the first detailed characterization of sodium-vitamin C co-transporter 2 (SVCT2) expression in skele- tal muscles [19]. Our data showed that SVCT2 is preferentially ex- pressed in slow skeletal myofibers in both developing chicks and adult mammalian species, including human [19]. These findings suggest that the uptake of vitamin C may help to counteract oxida- tion in mature slow muscle fibers. However, a possible role for 0014-5793/$36.00 Ó 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2010.12.020 Abbreviations: SVCT2, sodium-vitamin C co-transporter 2; PM, pectoralis major; MA, medial adductor; SERCA, slow sarco/endoplasmic reticulum calcium ATPase; ALD, anterior latissimus dorsi; PLD, posterior latissimus dorsi; NFAT, nuclear factor of activated T cells; NFjB, nuclear factor j of activated B cells ⇑ Corresponding author. Address: Laboratory of Developmental Neurobiology, Department of Cell Biology, Faculty of Biological Sciences, Universidad de Concep- cion, Casilla 160-C, Concepcion, Chile. Fax: +56 41 2245975. E-mail address: [email protected] (J.P. Henríquez). FEBS Letters 585 (2011) 390–396 journal homepage: www.FEBSLetters.org
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Up-regulation of the vitamin C transporter SVCT2 upon differentiation and depolarization of myotubes

FEBS Letters 585 (2011) 390–396
journal homepage: www.FEBSLetters .org
Up-regulation of the vitamin C transporter SVCT2 upon differentiation anddepolarization of myotubes
Marcela Low, Daniel Sandoval, Bryan Morales, Francisco Nualart, Juan Pablo Henríquez ⇑Department of Cell Biology, Faculty of Biological Sciences, Universidad de Concepcion, Concepcion, Chile
a r t i c l e i n f o a b s t r a c t
Article history:Received 1 September 2010Revised 30 November 2010Accepted 14 December 2010Available online 19 December 2010
Edited by Berend Wieringa
Keywords:Sodium-vitamin C co-transporter 2AscorbateMyogenesisFiber-typeSkeletal muscleDepolarization
0014-5793/$36.00 � 2010 Federation of European Biodoi:10.1016/j.febslet.2010.12.020
Abbreviations: SVCT2, sodium-vitamin C co-transpMA, medial adductor; SERCA, slow sarco/endoplasmiALD, anterior latissimus dorsi; PLD, posterior latissimusactivated T cells; NFjB, nuclear factor j of activated⇑ Corresponding author. Address: Laboratory of D
Department of Cell Biology, Faculty of Biological Sciecion, Casilla 160-C, Concepcion, Chile. Fax: +56 41 22
E-mail address: [email protected] (J.P. Henríquez
In addition to its role as a strong antioxidant, vitamin C regulates the differentiation of several celllineages. In vertebrate skeletal muscle, the vitamin C transporter SVCT2 is preferentially expressedin slow muscle fibers. To gain insights into the possible involvement of intracellular vitamin C onearly myogenesis, we investigated the regulation of SVCT2 expression in cultures of chick fetal myo-blasts. SVCT2 expression increases in cultures of both, slow and fast muscle-derived myoblasts, asthey fuse to form mainly fast myotubes. Interestingly, we found that SVCT2 could be positively mod-ulated by potassium-induced depolarization of myotubes. These findings suggest that SVCT2-medi-ated uptake of vitamin C could play diverse roles on skeletal muscle development and physiology.� 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
The formation of skeletal muscle during development involves aseries of sequential events, including the commitment of somite-derived precursor cells to mononucleated myoblasts, and theirsubsequent fusion into multinucleated myotubes [1]. Matureskeletal muscles consist of variable proportions of fast and slowmyofibers which, ultimately, account for their contractile activity[2]. Fiber types can be classified according to various parameters,such as the expression of type-specific protein isoforms and meta-bolic properties. Indeed, whereas most fast fibers are glycolytic,slow myofibers rely on oxidative metabolism for a permanent sup-ply of ATP [2].
Both mammalian and avian model systems have been em-ployed to identify the intrinsic properties and extrinsic factors thatdetermine the characteristics of mature fibers [2,3]. Whereas mur-ine myoblasts seem preprogrammed to a particular phenotype [4],the definition of skeletal muscle fiber types in the chick relies on
chemical Societies. Published by E
orter 2; PM, pectoralis major;c reticulum calcium ATPase;dorsi; NFAT, nuclear factor ofB cellsevelopmental Neurobiology,
nces, Universidad de Concep-45975.).
the patterning of muscle precursors, but also significantly on theirinnervation [5].
Vitamin C is the most important water-soluble antioxidant vita-min in the plasma and, as such, it has been characterized as one ofthe main non-enzymatic antioxidative systems in various tissues[6,7]. To accomplish intracellular functions, vitamin C must be effi-ciently incorporated into cells. Thus, whereas oxidized vitamin C,dehydroascorbic acid, is taken up by facilitative hexose transport-ers [8,9], two isoforms of high affinity sodium-dependent vitaminC co-transporters (SVCT1 and 2) mediate the uptake of ascorbicacid, the reduced form of vitamin C [10,11].
In addition to its role as a potent antioxidant, cumulative evi-dence supports that vitamin C positively regulates the differentia-tion of diverse cell lineages, such as osteoblasts, keratinocytes, andcells of the nervous system [12–15]. Interestingly, these effects ofvitamin C are accompanied by dynamic expression patterns oftheir SVCT transporters [15–18], thus supporting the notion thattransporter-mediated uptake of vitamin C is involved in celldifferentiation.
We have recently conducted the first detailed characterizationof sodium-vitamin C co-transporter 2 (SVCT2) expression in skele-tal muscles [19]. Our data showed that SVCT2 is preferentially ex-pressed in slow skeletal myofibers in both developing chicks andadult mammalian species, including human [19]. These findingssuggest that the uptake of vitamin C may help to counteract oxida-tion in mature slow muscle fibers. However, a possible role for
lsevier B.V. All rights reserved.
M. Low et al. / FEBS Letters 585 (2011) 390–396 391
vitamin C transport on early steps of myogenesis has not been ad-dressed. Here, we have analyzed the expression of SVCT2, and itsregulation by depolarization, in cultures of chick fetal myoblastsobtained from fast and slow muscles. Together with previous evi-dence, our data suggest that skeletal muscle development andphysiology could be regulated by intracellular vitamin C.
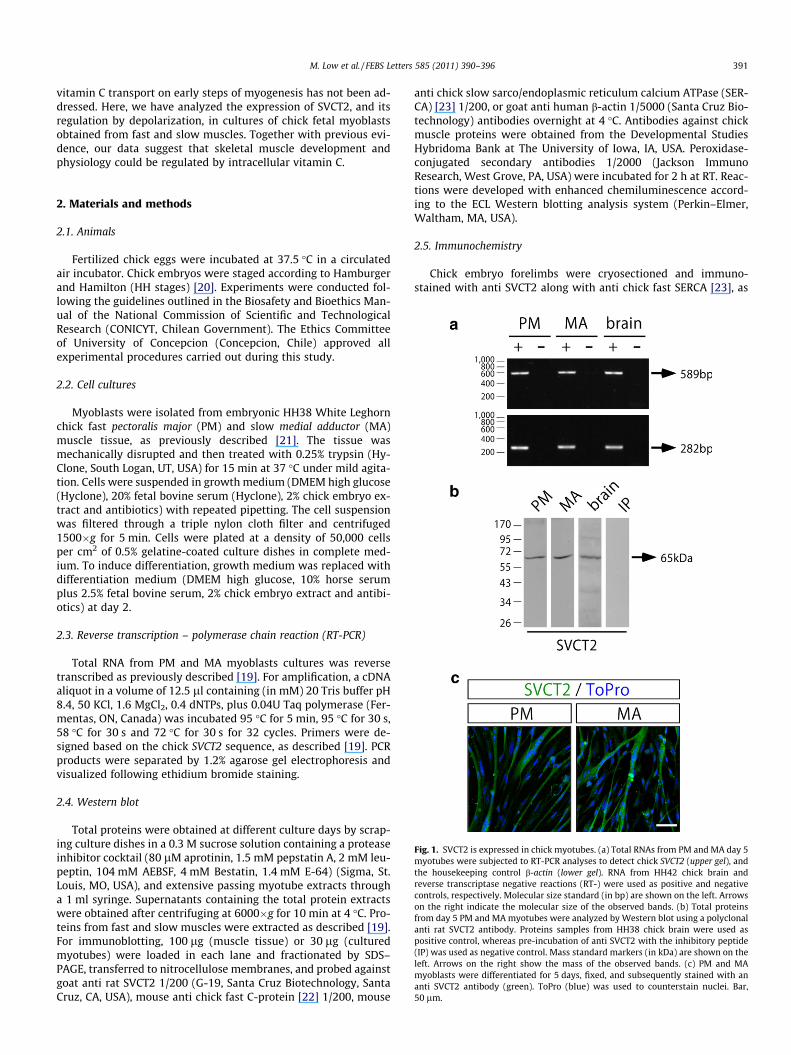
Fig. 1. SVCT2 is expressed in chick myotubes. (a) Total RNAs from PM and MA day 5myotubes were subjected to RT-PCR analyses to detect chick SVCT2 (upper gel), andthe housekeeping control b-actin (lower gel). RNA from HH42 chick brain andreverse transcriptase negative reactions (RT-) were used as positive and negativecontrols, respectively. Molecular size standard (in bp) are shown on the left. Arrowson the right indicate the molecular size of the observed bands. (b) Total proteinsfrom day 5 PM and MA myotubes were analyzed by Western blot using a polyclonalanti rat SVCT2 antibody. Proteins samples from HH38 chick brain were used aspositive control, whereas pre-incubation of anti SVCT2 with the inhibitory peptide(IP) was used as negative control. Mass standard markers (in kDa) are shown on theleft. Arrows on the right show the mass of the observed bands. (c) PM and MAmyoblasts were differentiated for 5 days, fixed, and subsequently stained with ananti SVCT2 antibody (green). ToPro (blue) was used to counterstain nuclei. Bar,50 lm.
2. Materials and methods
2.1. Animals
Fertilized chick eggs were incubated at 37.5 �C in a circulatedair incubator. Chick embryos were staged according to Hamburgerand Hamilton (HH stages) [20]. Experiments were conducted fol-lowing the guidelines outlined in the Biosafety and Bioethics Man-ual of the National Commission of Scientific and TechnologicalResearch (CONICYT, Chilean Government). The Ethics Committeeof University of Concepcion (Concepcion, Chile) approved allexperimental procedures carried out during this study.
2.2. Cell cultures
Myoblasts were isolated from embryonic HH38 White Leghornchick fast pectoralis major (PM) and slow medial adductor (MA)muscle tissue, as previously described [21]. The tissue wasmechanically disrupted and then treated with 0.25% trypsin (Hy-Clone, South Logan, UT, USA) for 15 min at 37 �C under mild agita-tion. Cells were suspended in growth medium (DMEM high glucose(Hyclone), 20% fetal bovine serum (Hyclone), 2% chick embryo ex-tract and antibiotics) with repeated pipetting. The cell suspensionwas filtered through a triple nylon cloth filter and centrifuged1500�g for 5 min. Cells were plated at a density of 50,000 cellsper cm2 of 0.5% gelatine-coated culture dishes in complete med-ium. To induce differentiation, growth medium was replaced withdifferentiation medium (DMEM high glucose, 10% horse serumplus 2.5% fetal bovine serum, 2% chick embryo extract and antibi-otics) at day 2.
2.3. Reverse transcription – polymerase chain reaction (RT-PCR)
Total RNA from PM and MA myoblasts cultures was reversetranscribed as previously described [19]. For amplification, a cDNAaliquot in a volume of 12.5 ll containing (in mM) 20 Tris buffer pH8.4, 50 KCl, 1.6 MgCl2, 0.4 dNTPs, plus 0.04U Taq polymerase (Fer-mentas, ON, Canada) was incubated 95 �C for 5 min, 95 �C for 30 s,58 �C for 30 s and 72 �C for 30 s for 32 cycles. Primers were de-signed based on the chick SVCT2 sequence, as described [19]. PCRproducts were separated by 1.2% agarose gel electrophoresis andvisualized following ethidium bromide staining.
2.4. Western blot
Total proteins were obtained at different culture days by scrap-ing culture dishes in a 0.3 M sucrose solution containing a proteaseinhibitor cocktail (80 lM aprotinin, 1.5 mM pepstatin A, 2 mM leu-peptin, 104 mM AEBSF, 4 mM Bestatin, 1.4 mM E-64) (Sigma, St.Louis, MO, USA), and extensive passing myotube extracts througha 1 ml syringe. Supernatants containing the total protein extractswere obtained after centrifuging at 6000�g for 10 min at 4 �C. Pro-teins from fast and slow muscles were extracted as described [19].For immunoblotting, 100 lg (muscle tissue) or 30 lg (culturedmyotubes) were loaded in each lane and fractionated by SDS–PAGE, transferred to nitrocellulose membranes, and probed againstgoat anti rat SVCT2 1/200 (G-19, Santa Cruz Biotechnology, SantaCruz, CA, USA), mouse anti chick fast C-protein [22] 1/200, mouse
anti chick slow sarco/endoplasmic reticulum calcium ATPase (SER-CA) [23] 1/200, or goat anti human b-actin 1/5000 (Santa Cruz Bio-technology) antibodies overnight at 4 �C. Antibodies against chickmuscle proteins were obtained from the Developmental StudiesHybridoma Bank at The University of Iowa, IA, USA. Peroxidase-conjugated secondary antibodies 1/2000 (Jackson ImmunoResearch, West Grove, PA, USA) were incubated for 2 h at RT. Reac-tions were developed with enhanced chemiluminescence accord-ing to the ECL Western blotting analysis system (Perkin–Elmer,Waltham, MA, USA).
2.5. Immunochemistry
Chick embryo forelimbs were cryosectioned and immuno-stained with anti SVCT2 along with anti chick fast SERCA [23], as
392 M. Low et al. / FEBS Letters 585 (2011) 390–396
previously described [19]. Myoblasts were seeded onto gelatine-coated 13-mm diameter glass coverslips, differentiated for 5 days,and fixed with cold methanol for 5 min. Cells were incubated withanti SVCT2 1/100 antibody diluted in Tris phosphate buffer con-taining 1% BSA, for 12–15 h at 4 �C. The corresponding alexa488-conjugated secondary immunoglobulin (Invitrogen, Carlsbad, CA,USA) was incubated for 2 h at RT, and the slides were subsequentlymounted with aqueous medium for fluorescence (Sigma). Imageswere acquired with a laser confocal Nikon D-Eclipse C1 micro-scope. Control experiments, performed in the absence of primaryantibodies and by co-incubation with the inhibitory peptide (SantaCruz Biotechnology), gave negative results (data not shown).Nuclei were stained by co-incubation with ToPRO-3 (Invitrogen),along with the secondary immunoglobulin.
2.6. Depolarization
Day 3 chick myotubes were incubated with Krebs–Ringermedium containing (in mM) 4.7 KCl, 20 HEPES-Tris, pH 7.4, 118NaCl, 3 CaCl2, 1.2 MgCl2, and 10 glucose, under resting conditionsfor 30 min. Depolarization was induced by incubation in a med-ium containing 84 mM KCl, whereas the sodium concentrationwas decreased proportionally to maintain the osmolarity of thesolution. After a 5 min depolarization period, cells were re-incu-bated under resting conditions for the indicated times. Both con-trol and experimental cells were subjected to semiquantitativeRT-PCR and Western blots analyses, as described [24,25]. Data
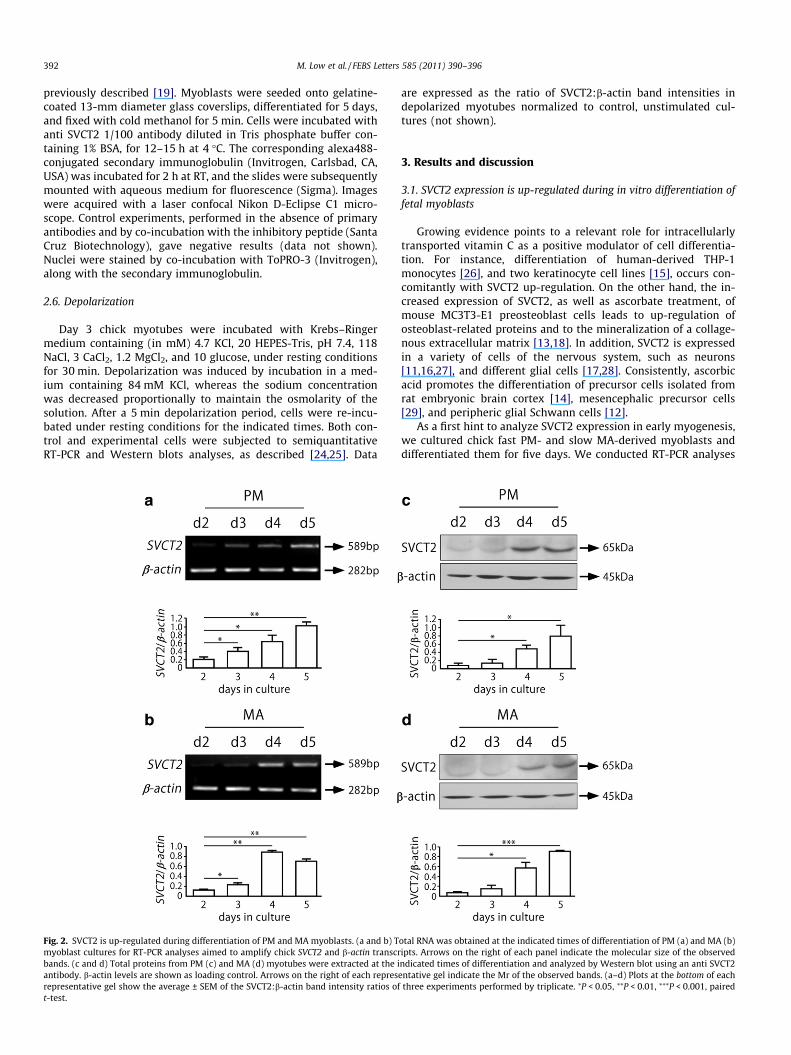
Fig. 2. SVCT2 is up-regulated during differentiation of PM and MA myoblasts. (a and b) Tmyoblast cultures for RT-PCR analyses aimed to amplify chick SVCT2 and b-actin transcrbands. (c and d) Total proteins from PM (c) and MA (d) myotubes were extracted at the iantibody. b-actin levels are shown as loading control. Arrows on the right of each represrepresentative gel show the average ± SEM of the SVCT2:b-actin band intensity ratios oft-test.
are expressed as the ratio of SVCT2:b-actin band intensities indepolarized myotubes normalized to control, unstimulated cul-tures (not shown).
3. Results and discussion
3.1. SVCT2 expression is up-regulated during in vitro differentiation offetal myoblasts
Growing evidence points to a relevant role for intracellularlytransported vitamin C as a positive modulator of cell differentia-tion. For instance, differentiation of human-derived THP-1monocytes [26], and two keratinocyte cell lines [15], occurs con-comitantly with SVCT2 up-regulation. On the other hand, the in-creased expression of SVCT2, as well as ascorbate treatment, ofmouse MC3T3-E1 preosteoblast cells leads to up-regulation ofosteoblast-related proteins and to the mineralization of a collage-nous extracellular matrix [13,18]. In addition, SVCT2 is expressedin a variety of cells of the nervous system, such as neurons[11,16,27], and different glial cells [17,28]. Consistently, ascorbicacid promotes the differentiation of precursor cells isolated fromrat embryonic brain cortex [14], mesencephalic precursor cells[29], and peripheric glial Schwann cells [12].
As a first hint to analyze SVCT2 expression in early myogenesis,we cultured chick fast PM- and slow MA-derived myoblasts anddifferentiated them for five days. We conducted RT-PCR analyses
otal RNA was obtained at the indicated times of differentiation of PM (a) and MA (b)ipts. Arrows on the right of each panel indicate the molecular size of the observedndicated times of differentiation and analyzed by Western blot using an anti SVCT2entative gel indicate the Mr of the observed bands. (a–d) Plots at the bottom of each
three experiments performed by triplicate. ⁄P < 0.05, ⁄⁄P < 0.01, ⁄⁄⁄P < 0.001, paired

M. Low et al. / FEBS Letters 585 (2011) 390–396 393
using specific primers to amplify an internal 589 bp fragment ofSVCT2 and a 282 bp fragment of chick b-actin mRNA, used hereas housekeeping control (Fig. 1a). Negative control experimentsperformed in the absence of reverse transcriptase did not showPCR products. Total RNA from HH42 chick brain was used as posi-tive control. As shown in Fig. 1a, a specific SVCT2 fragment wasamplified from PM and MA myotubes. To analyze SVCT2 expres-sion at the protein level, total myotube proteins were subjectedto Western blot using an antibody raised against a rat SVCT2 poly-peptide (Fig. 1b). Positive control experiments using chick brainextracts from stage HH38 embryos showed a single band of65,000 Dalton. A similar immunoreactive band was detected in to-tal protein extracts from PM and MA myotubes (Fig. 1b). As a fur-ther support of these findings, the expression of SVCT2 wasaddressed by immunocytochemistry in day 5 myotube cultures,using the same antibody (Fig. 1c). Together with our RT-PCR andWestern blot experiments, these immunodetection analyses showthat chick multinucleated myotubes do express SVCT2.
We next analyzed if SVCT2 expression is regulated during earlymyogenesis. First, total RNA samples from day 2 to day 5 myoblastscultures were subjected to RT-PCR analyses. As shown in Fig. 2,SVCT2 transcript is gradually increased as myogenesis proceeds.Quantification of the relative intensity of SVCT2 PCR product re-veals that, as compared to myoblasts cultured for 2 days, differen-tiated PM and MA myotubes up-regulate SVCT2 expression byabout five and sixfold, respectively (Fig. 2a and b). Consistent withthe expression profile of its transcript, the SVCT2 protein wasbarely detectable at early stages of myogenesis, and subsequentlyincreased with the course of differentiation. Quantification of bandintensity ratios shows that PM and MA myotubes significantly in-duce SVCT2 expression by about five and sixfold, respectively(Fig. 2c and d).
Up-regulation of SVCT2 during the fusion of chick fast- andslow-muscle derived myoblasts suggests that an increased uptakeof vitamin C could positively modulate early steps of myogenesis.
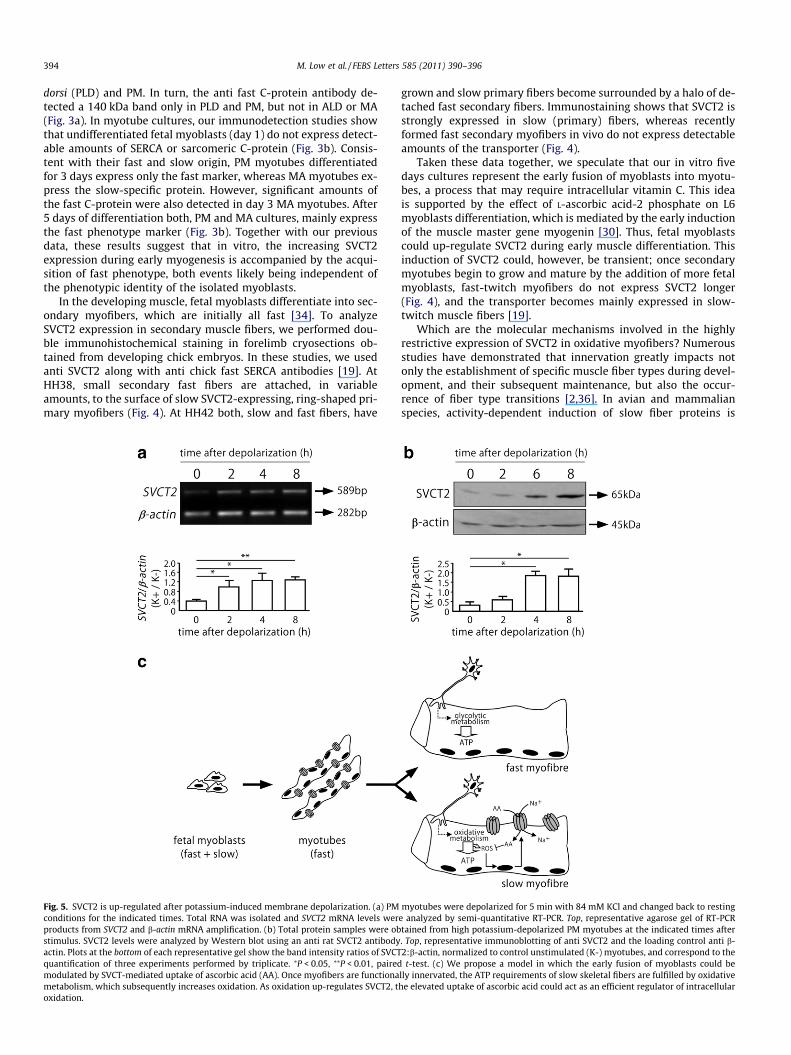
Fig. 3. Dynamic phenotypic profile of differentiating PM and MA myoblasts. Totalprotein extracts from HH39 chick embryo skeletal muscles were subjected toWestern blot analyses with fiber-type specific antibodies. (a) Extracts from the slowmuscles ALD and MA, as well as from the fast muscles PLD and PM, were probedagainst anti slow SERCA and anti fast C-protein antibodies. (b) Myoblasts obtainedfrom HH39 PM and MA muscles were differentiated for 5 days. Protein extractsobtained at various differentiation days (d1–d5) were immunoblotted against slowSERCA and fast C-protein. An anti b-actin antibody was used as loading control (aand b). Arrows indicate the Mr of the detected bands.
According to this notion, it has been shown that a stable form ofreduced vitamin C, L-ascorbic acid-2 phosphate, promotes the dif-ferentiation of rat-derived L6 myoblasts, as evidenced by their in-creased fusion into myotubes [30]. However, in contrast to ourfindings, SVCT2 mRNA and function is down-regulated throughoutin vitro myogenesis of L6 cells [31]. Even though murine and avianmyogenesis may differ in some aspects [3], an attractive interpre-tation of these seemingly contradictory results could be based onthe myoblasts employed in the different studies. It is well estab-lished that three populations of myoblasts – called embryonic, fetaland adult – predominate at different stages of muscle development[3,4,32]. L6 cells correspond to reliable models of adult myoblasts,those responsible of muscle growth and regeneration, as they wereisolated from newborn rats [33]. In turn, the fetal myoblasts usedin our studies were isolated from chick HH38 embryos, a develop-mental stage coinciding with secondary myogenesis, the processthat generates the vast majority of skeletal muscle fibers [34].Remarkably, vitamin C levels peak during secondary myogenesisin the chick, and strongly decay towards perinatal stages of devel-opment, when adult myoblasts predominate [35]. Therefore, eventhough further research has to be made to clarify this issue, it ispossible that embryonic, fetal and adult myoblasts may differ inthe mechanisms by which they use vitamin C to modulatemyogenesis.
3.2. Potassium-induced membrane depolarization up-regulates SVCT2in fast myotubes
Regardless of the phenotype of the prospective muscle theycome from, chick PM and MA fetal myoblasts give rise to fast myo-tubes in vitro [5]. To determine the phenotypic profile of differen-tiating PM and MA myotubes, we performed Western blot analysesusing monoclonal antibodies raised against slow SERCA and fast C-protein. We first ensured that these antibodies work as reliablemarkers of mutually exclusive populations of myotubes by per-forming immunoblot experiments of HH39 chick embryonic slowand fast skeletal muscles (Fig. 3a). Our results show that the antislow SERCA antibody recognizes a unique 110 kDa band only inprotein samples from the slow-twitch muscles anterior latissimusdorsi (ALD) and MA, but not in the fast muscles posterior latissimus
Fig. 4. SVCT2 is not expressed in chick fast secondary myofibers in vivo. Transversalforelimb cryosections from chick HH38 (left panels) and HH42 (right panels)embryos were double stained with a goat anti SVCT2 (green), together with amouse anti fast SERCA (red) antibody. Alexa488-conjugated anti goat and alexa546-conjugated anti mouse immunoglobulins were used as secondary antibodies.Magnified images are shown in the lower panels. Bars, 20 lm.
394 M. Low et al. / FEBS Letters 585 (2011) 390–396
dorsi (PLD) and PM. In turn, the anti fast C-protein antibody de-tected a 140 kDa band only in PLD and PM, but not in ALD or MA(Fig. 3a). In myotube cultures, our immunodetection studies showthat undifferentiated fetal myoblasts (day 1) do not express detect-able amounts of SERCA or sarcomeric C-protein (Fig. 3b). Consis-tent with their fast and slow origin, PM myotubes differentiatedfor 3 days express only the fast marker, whereas MA myotubes ex-press the slow-specific protein. However, significant amounts ofthe fast C-protein were also detected in day 3 MA myotubes. After5 days of differentiation both, PM and MA cultures, mainly expressthe fast phenotype marker (Fig. 3b). Together with our previousdata, these results suggest that in vitro, the increasing SVCT2expression during early myogenesis is accompanied by the acqui-sition of fast phenotype, both events likely being independent ofthe phenotypic identity of the isolated myoblasts.
In the developing muscle, fetal myoblasts differentiate into sec-ondary myofibers, which are initially all fast [34]. To analyzeSVCT2 expression in secondary muscle fibers, we performed dou-ble immunohistochemical staining in forelimb cryosections ob-tained from developing chick embryos. In these studies, we usedanti SVCT2 along with anti chick fast SERCA antibodies [19]. AtHH38, small secondary fast fibers are attached, in variableamounts, to the surface of slow SVCT2-expressing, ring-shaped pri-mary myofibers (Fig. 4). At HH42 both, slow and fast fibers, have
Fig. 5. SVCT2 is up-regulated after potassium-induced membrane depolarization. (a) PMconditions for the indicated times. Total RNA was isolated and SVCT2 mRNA levels werproducts from SVCT2 and b-actin mRNA amplification. (b) Total protein samples were obstimulus. SVCT2 levels were analyzed by Western blot using an anti rat SVCT2 antibodyactin. Plots at the bottom of each representative gel show the band intensity ratios of SVCTquantification of three experiments performed by triplicate. ⁄P < 0.05, ⁄⁄P < 0.01, pairedmodulated by SVCT-mediated uptake of ascorbic acid (AA). Once myofibers are functionametabolism, which subsequently increases oxidation. As oxidation up-regulates SVCT2, toxidation.
grown and slow primary fibers become surrounded by a halo of de-tached fast secondary fibers. Immunostaining shows that SVCT2 isstrongly expressed in slow (primary) fibers, whereas recentlyformed fast secondary myofibers in vivo do not express detectableamounts of the transporter (Fig. 4).
Taken these data together, we speculate that our in vitro fivedays cultures represent the early fusion of myoblasts into myotu-bes, a process that may require intracellular vitamin C. This ideais supported by the effect of L-ascorbic acid-2 phosphate on L6myoblasts differentiation, which is mediated by the early inductionof the muscle master gene myogenin [30]. Thus, fetal myoblastscould up-regulate SVCT2 during early muscle differentiation. Thisinduction of SVCT2 could, however, be transient; once secondarymyotubes begin to grow and mature by the addition of more fetalmyoblasts, fast-twitch myofibers do not express SVCT2 longer(Fig. 4), and the transporter becomes mainly expressed in slow-twitch muscle fibers [19].
Which are the molecular mechanisms involved in the highlyrestrictive expression of SVCT2 in oxidative myofibers? Numerousstudies have demonstrated that innervation greatly impacts notonly the establishment of specific muscle fiber types during devel-opment, and their subsequent maintenance, but also the occur-rence of fiber type transitions [2,36]. In avian and mammalianspecies, activity-dependent induction of slow fiber proteins is
myotubes were depolarized for 5 min with 84 mM KCl and changed back to restinge analyzed by semi-quantitative RT-PCR. Top, representative agarose gel of RT-PCRtained from high potassium-depolarized PM myotubes at the indicated times after. Top, representative immunoblotting of anti SVCT2 and the loading control anti b-2:b-actin, normalized to control unstimulated (K-) myotubes, and correspond to thet-test. (c) We propose a model in which the early fusion of myoblasts could be
lly innervated, the ATP requirements of slow skeletal fibers are fulfilled by oxidativehe elevated uptake of ascorbic acid could act as an efficient regulator of intracellular

M. Low et al. / FEBS Letters 585 (2011) 390–396 395
mediated by calcium signals inducing calcineurin-dependentdephosphorylation of the nuclear factor of activated T cells (NFAT),which translocates to the nucleus to activate target genes in coop-eration with other transcription factors [37,38]. Interestingly, highpotassium depolarization of rat myotubes results in transcriptionalregulation of more than 400 genes, including the up-regulation ofNFAT [25], and of the slow-specific isoforms of the sarcomeric tro-ponins I and C [24]. Therefore, as a first hint to analyze if innerva-tion of skeletal muscles could modulate SVCT2 expression, wechallenged chick fast PM myotubes to high (84 mM) KCl depolar-ization for 5 min, and changed them back to resting conditions.Samples were collected at different times after potassium stimula-tion and the expression of SVCT2 and b-actin transcripts was fol-lowed by a standardized semi-quantitative RT-PCR approach(Supplementary data Fig. 1). Our results show that myotube depo-larization resulted in a significant up-regulation of SVCT2 mRNA.Quantification of band intensities against control unstimulatedmyotubes show a threefold increase in SVCT2 expression 8 hoursafter potassium depolarization (Fig. 5a). Accordingly, immunode-tection experiments showed a fivefold increase of SVCT2 proteinafter the depolarizing stimulus, as compared to controls (Fig. 5b).These experiments suggest that specific patterns of electrical stim-ulation could somehow be involved in the preferential expressionof SVCT2 in slow myofibers. In this regard, well-known evidencedemonstrates: (a) that oxidation increases due to muscle activity,and (b) that oxidation itself stimulates a variety of antioxidant re-sponses in slow muscle fibers [39]. Interestingly, it has been shownthat hydrogen peroxide treatment of mouse C2C12 myotubes re-sults in up-regulation of SVCT2 via the nuclear factor j of activatedB cells (NFjB) [40]. Besides, NFjB expression is increased afterhigh potassium stimulation of rat myotubes [41]. Therefore, oxida-tion could somehow mediate the activity-dependent control ofSVCT2 expression in mature slow muscle fibers.
As our data suggest two levels of regulation of SVCT2 expres-sion, at early and later events of myogenesis, we propose a modelwhere vitamin C regulates different steps of skeletal muscle devel-opment (Fig. 5c). First, intracellular vitamin C could be required forearly muscle differentiation, as the fusion of chick fetal fast andslow-twitch muscle-derived myoblasts into fast myotubes isaccompanied by up-regulation of SVCT2. This notion is stronglysupported by cumulative evidence showing that SVCT2-mediateduptake of ascorbic acid potentiates the differentiation of severalcell lineages. On the other hand, our immunolocalisation studiesin developing chick muscles show that fast secondary myofibres,which are differentiated from fast and slow myoblasts in vivo, donot express SVCT2 longer, suggesting that other mechanismsshould account for SVCT2 expression in mature slow muscle fibres.We provide novel evidence to show that SVCT2 can be up-regu-lated by potassium-induced depolarization of fast myotubes. Alongwith previous findings, we propose that functional activity couldtrigger oxidation-dependent up-regulation of SVCT2. This mecha-nism would be particularly relevant in slow myofibers, as theyuse oxidative metabolism for energy production. In this regard, itis relevant to mention that no clear evidence exists to demonstratea protective role for vitamin C supplementation against exercise-induced oxidation [42]. It is then likely that up-regulation of theSVCT2 transporter could provide skeletal muscles, and perhapsother tissues, an efficient and sustained adaptive response againstoxidative stress.
Acknowledgements
Our work is supported by ANILLO PBCT ACT-02 (to J.P.H. andF.N.), FONDECYT 1100326 (to J.P.H.) and 1100396 (to F.N.) grants,and CONICYT ART-24091056 (to D.S.). M.L. and D.S. are CONICYTfellowship recipients. The authors thank Sylvain Marcellini, Nelson
Osses and members of our laboratory for useful discussion. We areindebted to the support of friends and colleagues after the damagecaused by the earthquake on February 27th, 2010.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2010.12.020.
References
[1] Bryson-Richardson, R.J. and Currie, P.D. (2008) The genetics of vertebratemyogenesis. Nat. Rev. Genet. 9, 632–646.
[2] Schiaffino, S. and Reggiani, C. (1996) Molecular diversity of myofibrillarproteins: gene regulation and functional significance. Physiol. Rev. 76, 371–423.
[3] Biressi, S., Molinaro, M. and Cossu, G. (2007) Cellular heterogeneity duringvertebrate skeletal muscle development. Dev. Biol. 308, 281–293.
[4] Biressi, S. et al. (2007) Intrinsic phenotypic diversity of embryonic and fetalmyoblasts is revealed by genome-wide gene expression analysis on purifiedcells. Dev. Biol. 304, 633–651.
[5] DiMario, J.X. and Stockdale, F.E. (1997) Both myoblast lineage and innervationdetermine fiber type and are required for expression of the slow myosin heavychain 2 gene. Dev. Biol. 188, 167–180.
[6] Frei, B., England, L. and Ames, B.N. (1989) Ascorbate is an outstandingantioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA 86, 6377–6381.
[7] Padayatty, S.J. et al. (2003) Vitamin C as an antioxidant: evaluation of its role indisease prevention. J. Am. Coll. Nutr. 22, 18–35.
[8] Rumsey, S.C., Daruwala, R., Al-Hasani, H., Zarnowski, M.J., Simpson, I.A. andLevine, M. (2000) Dehydroascorbic acid transport by GLUT4 in Xenopusoocytes and isolated rat adipocytes. J. Biol. Chem. 275, 28246–28253.
[9] Rumsey, S.C., Kwon, O., Xu, G.W., Burant, C.F., Simpson, I. and Levine, M. (1997)Glucose transporter isoforms GLUT1 and GLUT3 transport dehydroascorbicacid. J. Biol. Chem. 272, 18982–18989.
[10] Daruwala, R., Song, J., Koh, W.S., Rumsey, S.C. and Levine, M. (1999) Cloningand functional characterization of the human sodium-dependent vitamin Ctransporters hSVCT1 and hSVCT2. FEBS Lett. 460, 480–484.
[11] Tsukaguchi, H., Tokui, T., Mackenzie, B., Berger, U.V., Chen, X.Z., Wang, Y.,Brubaker, R.F. and Hediger, M.A. (1999) A family of mammalian Na+-dependent L-ascorbic acid transporters. Nature 399, 70–75.
[12] Eldridge, C.F., Bunge, M.B., Bunge, R.P. and Wood, P.M. (1987) Differentiationof axon-related Schwann cells in vitro. I. Ascorbic acid regulates basal laminaassembly and myelin formation. J. Cell Biol. 105, 1023–1034.
[13] Franceschi, R.T., Iyer, B.S. and Cui, Y. (1994) Effects of ascorbic acid on collagenmatrix formation and osteoblast differentiation in murine MC3T3-E1 cells. J.Bone Miner. Res. 9, 843–854.
[14] Lee, J.Y., Chang, M.Y., Park, C.H., Kim, H.Y., Kim, J.H., Son, H., Lee, Y.S. and Lee,S.H. (2003) Ascorbate-induced differentiation of embryonic cortical precursorsinto neurons and astrocytes. J. Neurosci. Res. 73, 156–165.
[15] Savini, I., Catani, M.V., Rossi, A., Duranti, G., Melino, G. and Avigliano, L. (2002)Characterization of keratinocyte differentiation induced by ascorbic acid:protein kinase C involvement and vitamin C homeostasis. J. Invest. Dermatol.118, 372–379.
[16] Castro, M. et al. (2001) High-affinity sodium-vitamin C co-transporters (SVCT)expression in embryonic mouse neurons. J. Neurochem. 78, 815–823.
[17] Gess, B., Lohmann, C., Halfter, H. and Young, P. (2010) Sodium-dependentvitamin C transporter 2 (SVCT2) is necessary for the uptake of L-ascorbic acidinto Schwann cells. Glia 58, 287–299.
[18] Wu, X., Itoh, N., Taniguchi, T., Hirano, J., Nakanishi, T. and Tanaka, K. (2004)Stimulation of differentiation in sodium-dependent vitamin C transporter 2overexpressing MC3T3-E1 osteoblasts. Biochem. Biophys. Res. Commun. 317,1159–1164.
[19] Low, M., Sandoval, D., Aviles, E., Perez, F., Nualart, F. and Henriquez, J.P. (2009)The ascorbic acid transporter SVCT2 is expressed in slow-twitch skeletalmuscle fibres. Histochem. Cell Biol. 131, 565–574.
[20] Hamburger, V. and Hamilton, H.L. (1992) A series of normal stages in thedevelopment of the chick embryo. Dev. Dyn. 195, 231–272.
[21] O’Neill, M.C. and Stockdale, F.E. (1972) A kinetic analysis of myogenesisin vitro. J. Cell Biol. 52, 52–65.
[22] Reinach, F.C., Masaki, T., Shafiq, S., Obinata, T. and Fischman, D.A. (1982)Isoforms of C-protein in adult chicken skeletal muscle: detection withmonoclonal antibodies. J. Cell Biol. 95, 78–84.
[23] Kaprielian, Z. and Fambrough, D.M. (1987) Expression of fast and slowisoforms of the Ca2+-ATPase in developing chick skeletal muscle. Dev. Biol.124, 490–503.
[24] Juretic, N., Urzua, U., Munroe, D.J., Jaimovich, E. and Riveros, N. (2007)Differential gene expression in skeletal muscle cells after membranedepolarization. J. Cell. Physiol. 210, 819–830.
[25] Valdes, J.A., Gaggero, E., Hidalgo, J., Leal, N., Jaimovich, E. and Carrasco, M.A.(2008) NFAT activation by membrane potential follows a calcium pathwaydistinct from other activity-related transcription factors in skeletal musclecells. Am. J. Physiol. Cell Physiol. 294, C715–C725.

396 M. Low et al. / FEBS Letters 585 (2011) 390–396
[26] Qiao, H. and May, J.M. (2009) Macrophage differentiation increases expressionof the ascorbate transporter (SVCT2). Free Radic. Biol. Med. 46, 1221–1232.
[27] Mun, G.H. et al. (2006) Immunohistochemical study of the distribution ofsodium-dependent vitamin C transporters in adult rat brain. J. Neurosci. Res.83, 919–928.
[28] Garcia Mde, L. et al. (2005) Sodium vitamin C cotransporter SVCT2 isexpressed in hypothalamic glial cells. Glia 50, 32–47.
[29] Yan, J., Studer, L. and McKay, R.D. (2001) Ascorbic acid increases the yield ofdopaminergic neurons derived from basic fibroblast growth factor expandedmesencephalic precursors. J. Neurochem. 76, 307–311.
[30] Mitsumoto, Y., Liu, Z. and Klip, A. (1994) A long-lasting vitamin C derivative,ascorbic acid 2-phosphate, increases myogenin gene expression and promotesdifferentiation in L6 muscle cells. Biochem. Biophys. Res. Commun. 199, 394–402.
[31] Savini, I., Catani, M.V., Duranti, G., Ceci, R., Sabatini, S. and Avigliano, L. (2005)Vitamin C homeostasis in skeletal muscle cells. Free Radic. Biol. Med. 38, 898–907.
[32] Stockdale, F.E. (1992) Myogenic cell lineages. Dev. Biol. 154, 284–298.[33] Richler, C. and Yaffe, D. (1970) The in vitro cultivation and differentiation
capacities of myogenic cell lines. Dev. Biol. 23, 1–22.[34] Fredette, B.J. and Landmesser, L.T. (1991) Relationship of primary and
secondary myogenesis to fiber type development in embryonic chickmuscle. Dev. Biol. 143, 1–18.
[35] Wilson, J.X. (1990) Regulation of ascorbic acid concentration in embryonicchick brain. Dev. Biol. 139, 292–298.
[36] Rubinstein, N.A. and Kelly, A.M. (1978) Myogenic and neurogeniccontributions to the development of fast and slow twitch muscles in rat.Dev. Biol. 62, 473–485.
[37] Chin, E.R. et al. (1998) A calcineurin-dependent transcriptional pathwaycontrols skeletal muscle fiber type. Genes Dev. 12, 2499–2509.
[38] Crew, J.R., Falzari, K. and DiMario, J.X. (2010) Muscle fiber type specificinduction of slow myosin heavy chain 2 gene expression by electricalstimulation. Exp. Cell Res. 316, 1039–1049.
[39] Palomero, J. and Jackson, M.J. (2010) Redox regulation in skeletalmuscle during contractile activity and aging. J. Anim. Sci. 88, 1307–1313.
[40] Savini, I., Rossi, A., Catani, M., Ceci, R. and Avigliano (2007) Redox regulation ofvitamin C transporter SVCT2 in C2C12 myotubes. Biochem. Biophys. Res.Commun. 361, 385–390.
[41] Valdes, J.A., Hidalgo, J., Galaz, J.L., Puentes, N., Silva, M., Jaimovich, E. andCarrasco, M.A. (2007) NF-kappaB activation by depolarization of skeletalmuscle cells depends on ryanodine and IP3 receptor-mediated calciumsignals. Am. J. Physiol. Cell Physiol. 292, C1960–C1970.
[42] McGinley, C., Shafat, A. and Donnely, A. (2009) Does antioxidantvitamin supplementation protect against muscle damage? Sport Med. 39,1011–1032.




















