
UNPREDICTABLE OFFSPRING SURVIVORSHIP IN THE DAMSELFLY, MEGALOPREPUS COERULATUS, SHAPES PARENTAL...
11
762 q 2001 The Society for the Study of Evolution. All rights reserved. Evolution, 55(4), 2001, pp. 762–772 UNPREDICTABLE OFFSPRING SURVIVORSHIP IN THE DAMSELFLY, MEGALOPREPUS COERULATUS, SHAPES PARENTAL BEHAVIOR, CONSTRAINS SEXUAL SELECTION, AND CHALLENGES TRADITIONAL FITNESS ESTIMATES OLA M. FINCKE 1 AND HEIKE HADRYS 2,3 1 Department of Zoology, University of Oklahoma, Norman, Oklahoma 73019 E-mail: [email protected] 2 Zoologisches Institut der Universitaet Frankfurt, D-60323 Frankfurt, Germany 3 Department of Ecology and Evolutionary Biology, Yale University, New Haven, Connecticut 06511 E-mail: [email protected] Abstract. Evolutionary biologists typically assume that the number of eggs fertilized or developing embryos produced is correlated with an individual’s fitness. Using microsatellite markers, we document for the first time estimates of realized fitness quantified as the number of offspring surviving to adulthood in an insect under field conditions. In a territorial damselfly whose males defend tree hole oviposition sites, patterns of offspring survivorship could not be anticipated by adults. Fewer than half of the parents contributing eggs to a larval habitat realized any reproductive success from their investment. The best fitness correlate was the span over which eggs in a clutch hatched. Among parents, female fecundity and male fertilization success were poor predictors of realized fitness. Although body size was correlated with female clutch size and male mating success, larger parents did not realize greater fitness than smaller ones. The uncoupling of traditional fitness surrogates from realized fitness provides strong empirical evidence that selection at the larval stage constrains selection on mated adults. Key words. Fitness, hatching asynchrony, larval survivorship, Odonata, selection. Received January 21, 2000. Accepted December 3, 2000. Many reproductive traits are thought to be shaped by sexual selection, resulting from variation among individuals in mating or fertilization success (Thornhill and Alcock 1983; Waage 1984; Clutton-Brock 1988; Andersson 1994). Evolutionary ex- planations for such traits depend on estimates of fitness, the precision of which determines the validity of selection models. For example, a male’s fitness is typically thought to depend primarily upon the number of mates he obtains (sexual selec- tion), fecundity of his mates whose eggs he fertilizes, and his life span (i.e., two components of natural selection) (e.g., Arnold and Wade 1984; Fincke 1988). Often, body size is viewed as the target of strong natural and/or sexual selection because it is commonly correlated with female clutch size or the number of mates or fertilizations that a male obtains (e.g., Howard 1988; Partridge 1988; Fincke 1992a; Andersson 1994; Choe 1994; Alcock and Houston 1996; Olsson and Shine 1996). Nevertheless, such conclusions about the relative strength of selective episodes and their effect on the evolution of specific traits remain premature for most species. This is because fitness of adults in natural conditions, particularly for animals lacking parental care, has been measured only indirectly, in terms of mating or fertilization success, or the number of eggs laid (e.g., Fincke 1986; Conner 1989; Moore 1990; McLain 1991; Barrowclough and Rockwell 1993; Had- rys and Siva-Jothy 1994). Whether these traditional fitness surrogates are proportional to the number of surviving off- spring a parent produces is typically unknown, due to the difficulty of following the fate of an individual’s offspring after its eggs have hatched. The Neotropical damselfly, Megaloprepus coerulatus (Odonata: Pseudostigmatidae) offers a rare opportunity to follow under field conditions the fate of a parent’s offspring from the egg stage to emergence as adult insects. Females mate and oviposit exclusively at water-filled tree holes, which are limiting reproductive resources for several sympatric odo- nate species (Fincke 1992b). Male M. coerulatus, the larger sex, defend large holes in light gaps that are usually created when a tree falls and depressions in its trunk fill with rain- water. Sexual selection favors large males, which are more successful than smaller ones in defending tree holes and con- sequently, in obtaining mates. A male requires a female to mate with him before permitting her to lay eggs in the hole he defends. The longer a male defends a site, the more fer- tilizations he obtains (Fincke 1992a). As is typical of odo- nates (Waage 1984, 1986; Fincke 1984a; Hadrys et al. 1993), the last male M. coerulatus to mate with a female before she oviposits fertilizes all the eggs in her clutch (H. Hadrys, T. Gerken, and O. Fincke, unpubl. data). During the eight to nine month breeding season at our study site in Panama, a large defended tree hole may be colonized by the offspring of more than a dozen territorial males and twice as many females, with a single female laying up to several hundred eggs in a hole. However, larval cannibalism reduces the density to about one final instar per 1–2 liter of water. Before dying up, even large tree holes rarely produce more than 15 emerging adult M. coerulatus annually (Fincke 1992a, 1994, 1998). Females also lay in small, undefended holes in forest understory, but holes smaller than one liter rarely produce more than a single adult (Fincke 1992a, 1998) and account for less than 10% of the yearly Megaloprepus adult recruitment. Me- galoprepus coerulatus larvae do not survive in dry tree holes. Adults aestivate during late dry season and females begin ovi- positing again about 1–2 months after the first tree hole filling rains of wet season (Fincke 1984b, 1992b). Here, we developed microsatellite markers to answer three major questions: (1) Within a tree hole, is there a predictable pattern of offspring survivorship to which parents can cue? (2) Do most parents realize at least some fitness from the
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of UNPREDICTABLE OFFSPRING SURVIVORSHIP IN THE DAMSELFLY, MEGALOPREPUS COERULATUS, SHAPES PARENTAL...

762
q 2001 The Society for the Study of Evolution. All rights reserved.
Evolution, 55(4), 2001, pp. 762–772
UNPREDICTABLE OFFSPRING SURVIVORSHIP IN THE DAMSELFLY,MEGALOPREPUS COERULATUS, SHAPES PARENTAL BEHAVIOR, CONSTRAINSSEXUAL SELECTION, AND CHALLENGES TRADITIONAL FITNESS ESTIMATES
OLA M. FINCKE1 AND HEIKE HADRYS2,3
1Department of Zoology, University of Oklahoma, Norman, Oklahoma 73019E-mail: [email protected]
2Zoologisches Institut der Universitaet Frankfurt, D-60323 Frankfurt, Germany3Department of Ecology and Evolutionary Biology, Yale University, New Haven, Connecticut 06511
E-mail: [email protected]
Abstract. Evolutionary biologists typically assume that the number of eggs fertilized or developing embryos producedis correlated with an individual’s fitness. Using microsatellite markers, we document for the first time estimates ofrealized fitness quantified as the number of offspring surviving to adulthood in an insect under field conditions. In aterritorial damselfly whose males defend tree hole oviposition sites, patterns of offspring survivorship could not beanticipated by adults. Fewer than half of the parents contributing eggs to a larval habitat realized any reproductivesuccess from their investment. The best fitness correlate was the span over which eggs in a clutch hatched. Amongparents, female fecundity and male fertilization success were poor predictors of realized fitness. Although body sizewas correlated with female clutch size and male mating success, larger parents did not realize greater fitness thansmaller ones. The uncoupling of traditional fitness surrogates from realized fitness provides strong empirical evidencethat selection at the larval stage constrains selection on mated adults.
Key words. Fitness, hatching asynchrony, larval survivorship, Odonata, selection.
Received January 21, 2000. Accepted December 3, 2000.
Many reproductive traits are thought to be shaped by sexualselection, resulting from variation among individuals in matingor fertilization success (Thornhill and Alcock 1983; Waage1984; Clutton-Brock 1988; Andersson 1994). Evolutionary ex-planations for such traits depend on estimates of fitness, theprecision of which determines the validity of selection models.For example, a male’s fitness is typically thought to dependprimarily upon the number of mates he obtains (sexual selec-tion), fecundity of his mates whose eggs he fertilizes, and hislife span (i.e., two components of natural selection) (e.g., Arnoldand Wade 1984; Fincke 1988). Often, body size is viewed asthe target of strong natural and/or sexual selection because itis commonly correlated with female clutch size or the numberof mates or fertilizations that a male obtains (e.g., Howard 1988;Partridge 1988; Fincke 1992a; Andersson 1994; Choe 1994;Alcock and Houston 1996; Olsson and Shine 1996).
Nevertheless, such conclusions about the relative strengthof selective episodes and their effect on the evolution ofspecific traits remain premature for most species. This isbecause fitness of adults in natural conditions, particularlyfor animals lacking parental care, has been measured onlyindirectly, in terms of mating or fertilization success, or thenumber of eggs laid (e.g., Fincke 1986; Conner 1989; Moore1990; McLain 1991; Barrowclough and Rockwell 1993; Had-rys and Siva-Jothy 1994). Whether these traditional fitnesssurrogates are proportional to the number of surviving off-spring a parent produces is typically unknown, due to thedifficulty of following the fate of an individual’s offspringafter its eggs have hatched.
The Neotropical damselfly, Megaloprepus coerulatus(Odonata: Pseudostigmatidae) offers a rare opportunity tofollow under field conditions the fate of a parent’s offspringfrom the egg stage to emergence as adult insects. Femalesmate and oviposit exclusively at water-filled tree holes, which
are limiting reproductive resources for several sympatric odo-nate species (Fincke 1992b). Male M. coerulatus, the largersex, defend large holes in light gaps that are usually createdwhen a tree falls and depressions in its trunk fill with rain-water. Sexual selection favors large males, which are moresuccessful than smaller ones in defending tree holes and con-sequently, in obtaining mates. A male requires a female tomate with him before permitting her to lay eggs in the holehe defends. The longer a male defends a site, the more fer-tilizations he obtains (Fincke 1992a). As is typical of odo-nates (Waage 1984, 1986; Fincke 1984a; Hadrys et al. 1993),the last male M. coerulatus to mate with a female before sheoviposits fertilizes all the eggs in her clutch (H. Hadrys, T.Gerken, and O. Fincke, unpubl. data).
During the eight to nine month breeding season at our studysite in Panama, a large defended tree hole may be colonizedby the offspring of more than a dozen territorial males and twiceas many females, with a single female laying up to severalhundred eggs in a hole. However, larval cannibalism reducesthe density to about one final instar per 1–2 liter of water. Beforedying up, even large tree holes rarely produce more than 15emerging adult M. coerulatus annually (Fincke 1992a, 1994,1998). Females also lay in small, undefended holes in forestunderstory, but holes smaller than one liter rarely produce morethan a single adult (Fincke 1992a, 1998) and account for lessthan 10% of the yearly Megaloprepus adult recruitment. Me-galoprepus coerulatus larvae do not survive in dry tree holes.Adults aestivate during late dry season and females begin ovi-positing again about 1–2 months after the first tree hole fillingrains of wet season (Fincke 1984b, 1992b).
Here, we developed microsatellite markers to answer threemajor questions: (1) Within a tree hole, is there a predictablepattern of offspring survivorship to which parents can cue?(2) Do most parents realize at least some fitness from the

763SURVIVORSHIP CHALLENGES FITNESS ESTIMATES
larval habitat they use? (3) Among mated individuals, whichtraits best predict parental fitness?
MATERIALS AND METHODS
The work was conducted in 1994–1997 on Barro ColoradoIsland, Panama (hereafter, BCI), where the population dy-namics of Megaloprepus coerulatus and the interactions ofthis species with other tree hole predators have been studiedextensively (Fincke 1992a,b, 1994, 1998, 1999). The island’slowland moist forest experiences a dry season from Decemberto April (for detailed site description, see Leigh 1999). Typ-ically, tree holes dry up completely by late March and refillby late May each year.
Natural Territory Tenure Duration and Tree Hole Use byMarked Adults
For each of three years, we monitored 5–18 defended sitesin the mature forest of the island’s central plateau, notingthe presence of marked adults. Male territories at tree-fallgaps were checked multiple times per day and multiple timesper week over the course of the wet season. We marked adultsby writing a number on the wing with an indelible marker.With forceps, we removed a single middle tibia for later DNAanalysis. Adults occasionally lose tibia naturally; our pro-cedure did not affect a male’s mating ability nor his territorytenure. Forewing length was measured as a correlate of bodysize. In sexually mature individuals, hind-wing length is sig-nificantly correlated with abdomen length (r 5 0.98, P 50.001, n 5 12 males; r 5 0.89, P 5 0.02, n 5 6 females)and wet weight (r 5 0.96, P 5 0.0001, n 5 12 males; r 50.86, P 5 0.03, n 5 6 females). Relative age was scored ona scale of 1–3. Young individuals (age 1) had a fresh ap-pearance and iridescent wings. Old individuals (age 3) haddull wings with signs of wear, pruniose (whitish) abdomens,and often had algae on the thorax or wings. Individuals be-tween these two extremes were scored as age 2. Unless oth-erwise noted, data refer to sexually mature individuals.
A territorial male was one that was known to defend atleast one site for three or more consecutive days. Males mayleave a territory voluntarily or because they are displacedafter an agonistic encounter, usually with a larger or similar-sized male (Fincke 1992a). When no such encounters werenoted before a male left a site, he was credited with leavingvoluntarily if no other males were known to defend the sitewithin two days from the last time he was seen on the ter-ritory.
Determining Patterns of Larval Survivorship
In 1994 and 1995, we measured the fitness of 48 uniquemated pairs by following the fate of all of their offspring inone of six identical, large larval habitats with abundant nu-trient input from which other major predators were excluded(i.e., the best case scenario for offspring survivorship). Thepairs represented a subset of 36 territorial males and their 48mates from the above field study. After mating with a markedterritorial male, a female was caught and a tibia was removedfor later analysis. She was then placed in a small, screenedoviposition cage supplied with moistened filter paper. Most
females began egg laying within 10 min. They were leftundisturbed in the containers until later that same afternoonor the next morning, when they were released. The eggs arelarge (circa 1.5 mm in length) and easy to count accurately.Within a day of being laid, eggs were counted and a clutchwas assigned to an artificial hole. Starting in September 1994a total of six such clutches, laid at roughly two-week intervals(i.e., the median time it took for a male to encounter a femaleat a territory), were added to each of three artificial tree holes.These holes simulated the case of new larval habitats createdby tree falls in mid-wet season. To minimize possible am-biguity in parentage determination of the surviving offspring,clutches from different females that were sired by the sameterritorial male were distributed among the holes.
Under natural conditions M. coerulatus insert eggs intobark just above the water line. In the experimental holes, thefilter paper containing the clutch was placed on a piece ofbark with the end of the paper touching the water so eggsremained moist. One such clutch was attacked by mold beforeany eggs could have hatched and consequently was excludedfrom analysis. To quantify the number of newly hatched lar-vae (‘‘neonates’’) actually entering the holes, in 1995 wemodified the above procedure to include a measure of hatch-ing success. Then, each clutch was kept separately, sub-merged in water in a 400 ml plastic jar. The jars were heldin an outdoor insectary, sheltered from rain and direct sun,where they experienced ambient temperature and photope-riod. The minimum time for egg hatch is 14 days. After twoweeks, each clutch was checked daily for neonates, whichwere deposited into their assigned hole. In 1995, starting inmid-July, each of three artificial holes received all neonatesfrom each of 10 clutches laid at roughly two-week intervalsover most of the wet season. These holes simulated naturallarval habitats that fill with water in the beginning of wetseason; the last egg inputs were in late December. Egg inputwas at the same approximate rate, but over a longer timespan than in 1994, with each hole receiving about twice thetotal eggs as in the previous year (Table 1). As the dry seasonprogressed, holes were topped off with water periodically toprevent any mortality due to hole drying.
The artificial larval habitats were made of nine-liter plastictubs containing two liters of loosely packed fallen leaves, allcollected from the same area, and a large piece of bark thatprovided a perch site for ovipositing prey species (primarilymosquitos). These artificial holes were larger by volume than90% of the natural tree holes on BCI (Fincke 1992b). Arti-ficial holes such as these are colonized by the same array ofprey species as the natural tree holes (Fincke et al. 1997).The tubs were positioned in one of two shaded sites at theedge of forest clearings and covered with chicken wire en-closures to permit the passage of prey species while pre-venting oviposition by predatory odonates or frogs. Predatorylarvae of Toxorhynchites theobaldi mosquitoes could not beexcluded but were removed when detected during a census.Final instar Toxorhynchites are able to kill small odonatelarvae (i.e., ,7 mm, Fincke 1999). Control tubs lacking M.coerulatus larvae were used to estimate prey abundance. Theabundance of mosquito larvae prey was kept consistent withbimonthly additions of 0.5 g yeast after day 170 (necessarybecause the enclosure prevented the natural input of detritus).

764 O. M. FINCKE AND H. HADRYS
TABLE 1. Mean (and range) of egg input, larval characteristics, hole productivity, and adult success in the larval habitats (n 5 3 each year).In 1994, egg input was spaced over a 12-week span, whereas in 1995, egg input was spaced over a 20-week span. * P , 0.05, t-tests.
1994 1995
Total clutches/hole 5.6 6 0.3 (5–6) 10 6 0 (10)*Total eggs/hole 304 6 42 (250–386) 776 6 98 (651–969)*Windows/hole 2.0 (2) 3.7 6 0.3 (3–4)*Final instars/hole 6.0 6 0.6 (5–7) 8.0 6 1.5 (6–11)Maximum estimated time to emergence (days) 143.8 6 8.6 (85–215) 161.7 6 7.8 (109–236)Final instar size (mm) 29.06 6 0.5 (27–34) 29.1 6 0.4 (26.5–33.0)Proportion of females successful 0.41 6 0.05 (0.33–0.50) 0.47 6 0.06 (0.4–0.6)Offspring/successful female 2.7 6 0.4 (2.0–3.5) 1.7 6 0.1 (1.50–1.75)*
At bimonthly intervals, the contents of the holes were cen-sused and all M. coerulatus larvae 6 mm or more in size weremeasured from the head to the end of the final abdominalsegment (excluding the leaflike caudal lamellae used in res-piration). When their wing pads became swollen, indicativethat feeding had ceased and emergence was imminent, finalinstar larvae were collected. They were permitted to emergein an insectary, after which a leg was collected. Adults weremarked and released. Maximum developmental time was thetime span between the day the clutch was laid day and theday the surviving offspring emerged. After the last final instarwas removed, a hole was checked weekly for two to threeweeks to insure that no other larvae were still present.Throughout, means are reported with 6 SE. A Pearson cor-relation is indicated by r; a Spearman rank correlation by rs.When P-values fall between 0.05 and 0.3, the power (1 2b) of a statistical test is also reported.
Identification of Parents and Their Surviving Offspring
Of the individuals monitored during the field study of1994–1995, a total of 200 individuals were genetically char-acterized by PCR typing. These included the individuals ofthe above-mentioned 48 mated pairs, all 42 of their survivingoffspring, and eight ‘‘test’’ families (parents and single lar-vae) of known pedigree. The latter were used to demonstratethat paternity could be correctly assigned using the four mi-crosatellite loci. Also typed were 50 unrelated individuals,which served as an exclusion control for the paternity as-signment and were used to estimate genotypic and allelicdiversity (H. Hadrys, W. Schroth, B. Streit, and O. Fincke,unpubl. ms.).
Total genomic DNA was isolated according to standardprocedures (Hadrys et al. 1992). Microsatellite loci were de-tected and isolated using the RAMS method (Randomly Am-plified Microsatellites; Ender et al. 1996; Siva-Jothy and Had-rys 1998). A total of 45 different RAPD profiles from threeunrelated individuals (average of 10–15 RAPD fragments/profile) were blotted onto nylon membrane (Boehringer,Roche Diagnostics, Mannheim, Germany) and checked forhybridization signals with the digoxigenin-labelled oligo-nucleotides (GA)10, (GT)10, and (AAT)9 (see Ender et al.for details). Positive DNA fragments (i.e. RAPD fragmentsthat gave a hybridization signal for any one of the labeledoligonucleotide sequences) were gel purified, cloned and se-quenced, each for three unrelated individuals. This way itwas possible to isolate not only the sequences but also todetect allelic polymorphisms of the microsatellite sequences.
For use in PCR typing, four microsatellite loci were chosenthat had revealed a high allelic and genotypic diversity. Prim-ers were designed from conserved flanking regions designedto amplify between 150 and 250bp of PCR products (For primersequences and complete microsatellite sequences see:http://www.rz.unifrankfurt.de/;hadrys/microsat.html. The observedheterozygosity values for the four loci were: MeAB3/11 50.75; MeM12/15 5 0.89; MeAB3/16 5 0.93 and MeAB5/195 0.72 (H. Hadrys, W. Schroth, B. Streit, and O. Fincke,unpubl. ms.).
The genetic characterization of all 200 individuals revealeda total of 42 different alleles, the distribution of which per-mitted paternity and maternity assignment for all but twooffspring. Parentage of the latter was resolved using date ofoviposition and estimated developmental rate.
Determination of Adult Fitness
The fitness that a female gained from laying a clutch in agiven hole was simply the number of her offspring thatemerged as adults. Our experiment was designed to detectany predictable patterns of offspring survivorship rather thanto measure variation in male fitness resulting from differencesamong males in territory tenure at a given hole. Thus, herewe use the data on offspring survivorship to compare thefitness of two male tactics: (1) males that stay at a site onlylong enough to get a single mating, and (2) males that stay60 days, the maximum observed tenure duration in the fieldstudy. The fitness of a male staying only long enough toobtain a single mating would depend only on his ability tofertilize his mate’s clutch. Thus, the fitness of tactic 1 wascalculated as the mean number of surviving offspring fromeach of the 47 clutches in the experimental holes. The fitnessof a male staying the maximum tenure duration would vary,depending on when the male began defending the hole. Forconvenience, we calculated the fitness of hypothetical malesbeginning their tenure at 10-day intervals from each other(about the average time a male spent at a site). For example,a male playing tactic 2 that arrives on day 1 would sire alloffspring from the clutches laid between days 1–60. There-after, fitness was calculated for hypothetical males arrivingat 10-day intervals and staying a total of 60 days (i.e., thesecond male stays from day 20–80, the third male from day30–90, etc.). The last male would be the one whose 60-daytenure includes the final clutch. Reiterating this calculationfor each of the six holes resulted in 51 hypothetical males,whose mean fitness was compared with the above 47 malesplaying tactic 1. Estimated male lifetime fitness, based on

765SURVIVORSHIP CHALLENGES FITNESS ESTIMATES
the field data on tenure duration, male life span, female en-counter rate, and larval growth and survivorship is presentedelsewhere (O. Fincke, O. Eikemeier, and H. Hadrys, unpubl.ms.).
Hatching Asynchrony over a Reproductive Season
Hatching span, independent of clutch size, was found tobe the best predictor of fitness that females realized from ourexperimental holes. We monitored egg hatching in a largersample of 111 clutches, each laid by a different female duringthe wet seasons of 1995–1997, to confirm that the span overwhich eggs hatched was not correlated with clutch size andto determine possible seasonal effects on hatching patterns.In each year, study day 1 was standardized to May 1, abouttwo weeks after the first tree hole-filling rains of wet season,and before most M. coerulatus start to oviposit (Fincke1992b). A female was collected at a defended site before shebegan laying eggs in a hole. After her clutch was collectedin a monolayer of filter paper, it was kept submerged in aseparate jar under ambient conditions as described above.Two weeks after it was laid, and up to 60 days thereafter,the clutch was checked daily and any neonates were removed.Thereafter, it was checked at least once every five days, untilall the eggs had hatched or until 150 days after the first hatch,whichever came first. After 150 days, any clutch that stillcontained unhatched eggs was checked at haphazard inter-vals; any larvae found were assigned a hatch date midwaybetween the day of the previous check and the day of de-tection.
RESULTS
Patterns of Offspring Survivorship
The average size of the clutches used in the larval survi-vorship experiment was 68.9 6 7.1 eggs (range 8–214). Den-sity of final instars in the experimental holes (Table 1) werewithin the ranges noted for similar-sized natural tree holesat territories frequented by ovipositing females throughoutthe wet season (Fincke 1992a, 1998). On 80% of the censuses,the density of mosquito prey available (as measured in con-trols) ranged from moderate to high (i.e., 50–150 mosquitoes/census). For 28% of the surviving larvae, even the maximumestimated time from egg to emergence as an adult was lessthan 130 days, as fast as the minimum rate known (Fincke1992a). The only evidence of colonization by other odonateswere three small Mecistogaster larvae removed from a controlhole after the piece of bark was mistakenly left in contactwith the screening, and a Triacanthagyna dentata dragonflylarva found in hole 5 at the end of the experiment.
Among the six larval habitats, periods favorable to off-spring survivorship varied considerably, despite the stan-dardized timing of egg input (Fig. 1). The only consistentpattern of M. coerulatus survivorship was that the first clutchlaid, regardless of its size, always produced at least one sur-viving offspring (range, 1–5 offspring). Subsequently, sur-vivorship was less predictable. Females that produced thenext surviving offspring laid a clutch as few as 15 days andas many as 78 days after the first female.
The first larva entering a hole enjoyed a preemptive ad-
vantage. Larger individuals ate and killed smaller conspe-cifics, evidenced by an occasional larval body part foundduring a census (see also Fincke 1994, 1996). Cannibalismreduced larval density within a hole to less than seven larvae($6 mm) on any given census. Multiple, small larvae werefound only shortly before final instars emerged, or when thelargest larva(e) present was 15 mm or smaller. The M. coe-rulatus that survived to emergence were thus a subset of thosehatching during a ‘‘window of opportunity,’’ either when nolarge larvae were present, or shortly before larger larvaeemerged (Fig. 1). The presence of a favorable window de-pended on larval growth rates coupled with the timing of egghatch in clutches laid previously and subsequently to a fe-male’s own. In 1994, when clutches were deposited over a12-week span, holes experienced fewer such windows thanin 1995, when egg input was over a 20-week span (Table 1).Using data from only the first six clutches in each hole (i.e.,controlling for between-year differences in the time span ofegg input), the number of final instars produced was notcorrelated with the number of eggs a hole received (rs 520.40, P 5 0.42, n 5 6 holes). Mean productivity of theholes, measured as the number of adults produced per dayover which holes were occupied, was nearly identical be-tween years, (0.030 6 0.00 vs. 0.027 6 0.00, 1994 and 1995respectively, t 5 0.47, P 5 0.66).
Female Fitness and Phenotypic Predictors of Fitness
On average, fewer than half of the pairs that contributedeggs to a hole produced any surviving offspring (Table 1).Survivorship of eggs in a clutch varied from 0 to 0.25 (x̄ 50.02 6 0.06, n 5 47 clutches, excluding the one attacked bymold). Of offspring known to hatch, survivorship varied from0 to 0.33 (x̄ 5 0.03 6 0.01, n 5 30 females). Successful andunsuccessful females did not differ significantly in the hatch-ing success of their clutch (t 5 0.42, n 5 47, P 5 0.67), timeto first hatch, clutch size, or the number of neonates produced(Fig. 2a-c).
Even though larger females produced larger egg clutches(Fig. 3), the number of surviving offspring was not a functionof a female’s body size (r 5 0.03, n 5 47, P 5 0.86) or thatof her mate (r 5 20.03, n 5 41, P 5 0.86). Indeed, repro-ductive success was not correlated significantly with the num-ber of eggs a female laid (r 5 0.20, n 5 47, P 5 0.17, 1 2b 5 0.39), or with the number of eggs that actually hatched(Fig. 4a). Even amongst females that produced at least onesurviving offspring, increasing clutch size was not correlatedwith the number of surviving offspring (rs 5 0.04, n 5 21,P 5 0.86). Rather, the best predictor of female reproductivesuccess was the span between hatching of the first and lastegg in a clutch (range 5 4–84 days; Fig. 4b). The hatch spanof successful clutches was about 1.5 times longer than un-successful ones (Fig. 2d). Hatching span was not a functionof clutch size (r 5 0.24, n 5 30, P 5 0.20, 1 2 b 5 0.37)in this experiment, nor in a larger sample described below.However, the time required for the first egg in a clutch tohatch was negatively correlated with clutch size (r 5 20.36,n 5 30, P 5 0.05).
The first clutch to be laid in a hole, even one as small aseight eggs, always produced some surviving offspring due
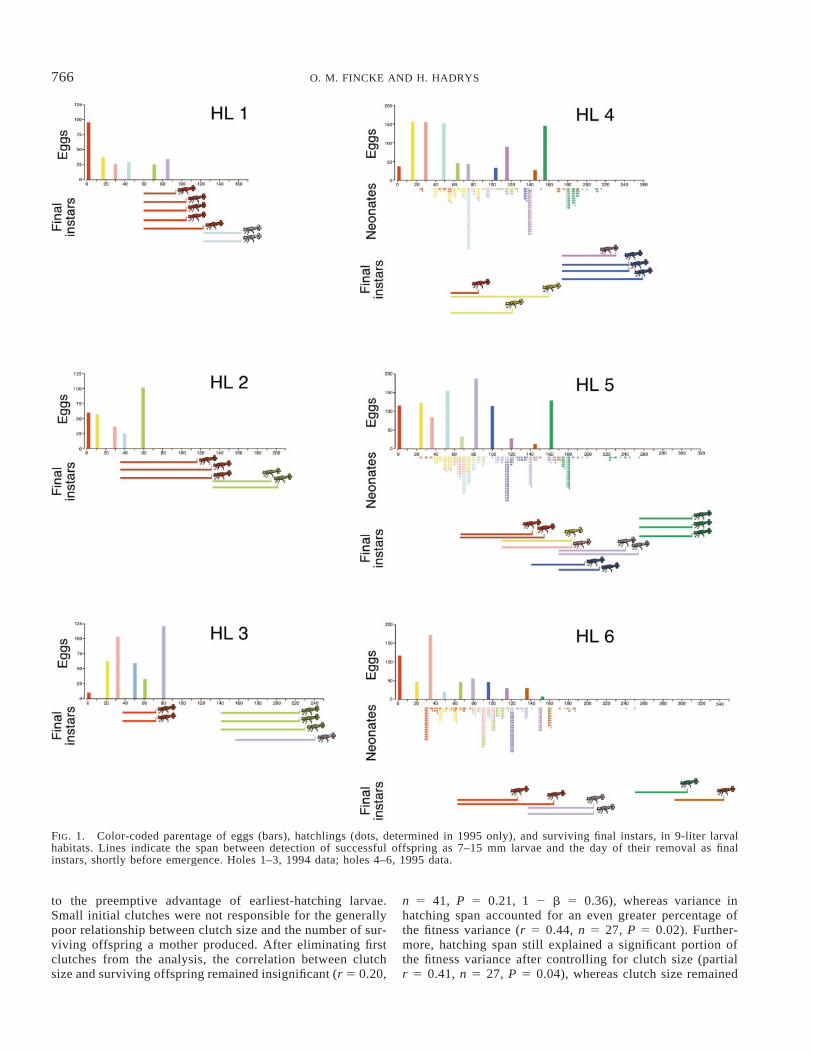
766 O. M. FINCKE AND H. HADRYS
FIG. 1. Color-coded parentage of eggs (bars), hatchlings (dots, determined in 1995 only), and surviving final instars, in 9-liter larvalhabitats. Lines indicate the span between detection of successful offspring as 7–15 mm larvae and the day of their removal as finalinstars, shortly before emergence. Holes 1–3, 1994 data; holes 4–6, 1995 data.
to the preemptive advantage of earliest-hatching larvae.Small initial clutches were not responsible for the generallypoor relationship between clutch size and the number of sur-viving offspring a mother produced. After eliminating firstclutches from the analysis, the correlation between clutchsize and surviving offspring remained insignificant (r 5 0.20,
n 5 41, P 5 0.21, 1 2 b 5 0.36), whereas variance inhatching span accounted for an even greater percentage ofthe fitness variance (r 5 0.44, n 5 27, P 5 0.02). Further-more, hatching span still explained a significant portion ofthe fitness variance after controlling for clutch size (partialr 5 0.41, n 5 27, P 5 0.04), whereas clutch size remained

767SURVIVORSHIP CHALLENGES FITNESS ESTIMATES
FIG. 2. Comparison of females that produced at least one surviving offspring from a clutch, with those producing none (a) clutch size,t 5 21.45, n 5 47, P 5 0.15, 1 2 b 5 0.41; (b) number of neonates, t 5 21.0, n 5 30, P 5 0.30, 1 2 b 5 0.25; (c) time to firsthatch, t 5 0.63, n 5 47, P 5 0.53; and (d) hatching span, t 5 22.16, n 5 30, P 5 0.04.
FIG. 3. Clutch size as a function of wing length of females usedin the experiment.
insignificantly correlated with surviving offspring after con-trolling for hatching span (partial r 5 0.11, n 5 27, P 50.59).
Seasonal Effects on Clutch Size, Hatching Patterns, andBody Size
One female caught on day 1 of the study in 1995 laid 514eggs (18 days to first hatch; hatching span 5 41 days), con-siderably larger than the second largest clutch of 344 eggs.We conservatively excluded this female from the followinganalysis; including her does not change any conclusion.
In the larger sample of 111 clutches collected throughoutthe wet season, hatching was highly asynchronous. The av-erage clutch size was 109.6 6 6.5 eggs (range 5 9–344).Eggs in a clutch hatched over a span of as little as four days,or as many as 184 days (x̄ 5 59.3 6 3.6 days). Hatchingspan was not correlated with clutch size in this larger sample(r 5 0.08, P 5 0.38). Limiting the analysis of hatching spanto the clutches with a documented hatching success of $0.70(x̄ 5 0.81 6 0.01) did not change this result (Fig. 5). Asshown in Table 2, the most variable portion of the hatchingsequence was the span over which the final quartile of theeggs in a clutch hatched. As was true for the smaller samplein our experiment above, the time required for the first eggin a clutch to hatch was negatively correlated with clutchsize (Fig. 6). In clutches of 50 or more eggs, the mean time
to first egg to hatch was 20.6 6 day (range 14–30 days),about four days earlier than the minimum hatch time of small-er clutches (x̄ 5 24.8 6 1.6 days, range 18–32 days, t 5 3.1,P 5 0.003). Clutch size (r 5 20.12, n 5 111, P 5 0.19, 12 b 5 0.35), hatching span (r 5 20.14, n 5 57, P 5 0.29,1 2 b 5 0.28), and the time to first hatch (r 5 20.14, n 5

768 O. M. FINCKE AND H. HADRYS
FIG. 4. Reproductive success of females as a function of (a) thenumber of larvae known to hatch and (b) the span between the timethe first and last eggs in a clutch hatched.
TABLE 2. Variation in hatching asynchrony (in days) of 57 clutcheshaving $ 0.70 hatching success, each laid by a different female. Thespan of a quartile could be zero if $25% of the eggs in a clutch hatchedon a single day.
x̄ 6 SE Range
Time to first hatchSpan of 1st quartileSpan of 2nd quartileSpan of 3rd quartileSpan of 4th quartile
20.7 6 0.465.4 6 0.094.1 6 0.057.1 6 1.2237.7 6 3.3
14–331–270–150–541–128
FIG. 6. Days to first egg hatch, as a function of clutch size.
FIG. 5. Hatching span as a function of the neonates known to hatchfrom clutches laid over the wet seasons of 1995–1997. Controllingfor laying date, partial r 5 0.15, n 5 57 clutches with $70% hatch-ing success, P 5 0.28, 1 2 b 5 0.30.
57, P 5 0.29, 1 2 b 5 0.28), all tended to decrease with theday of oviposition, though not significantly.
A negative correlation between wing length of individualsmarked at age 1 and the day of marking (r 5 20.25, n 5 79females, P 5 0.02; r 5 20.35, n 5 256 males, P 5 0.001)indicated that body size decreased as the wet season pro-gressed. Controlling for this seasonal effect, the age class ofthe mother had no detectable effect on her clutch size (partialrs 5 20.04, n 5 49, P 5 0.78), hatching span (rs 5 20.003,n 5 49, P 5 0.98) nor time to first hatch (rs 5 20.05, n 549, P 5 0.74). Controlling for the day of laying, clutch sizewas no longer correlated significantly with female size (par-tial r 5 0.07, n 5 98, P 5 0.51).
Controlling for day of marking, female body size was notcorrelated with relative life span (partial r 5 0.08, n 5 234,P 5 0.20), but only 15 females were seen more than once.For this subset, the span over which a female was seen (x̄ 521.1 6 7.0 days, range 5 2–88 days) was correlated withbody size (partial rs 5 0.58, P 5 0.03). Seven of these 15females were resighted at defended tree holes, on average11.7 6 4.3 days (range 3–35 days) after the first sightings.No female was seen to lay more than once in the same treehole.
Male Fitness and Phenotypic Predictors of Fitness
The mean span over which marked, five-legged males wereseen in this study was 12.8 6 1.3 days (range 5 1–155 days,

769SURVIVORSHIP CHALLENGES FITNESS ESTIMATES
FIG. 7. Length of residency at a site as a function of male size(47 territorial sites, 1994–1996 wet seasons). Controlling for dayof marking, partial r 5 0.15, n 5 116 males, P 5 0.10. Horizontalbar indicates males holding territories for #25 days, about the timerequired to encounter a female. For this group, partial r 5 0.2, n5 99, P , 0.05, see text).
n 5 365). This was not significantly different from the meanspan of 14.6 6 1.8 days (range 1–165 days, n 5 236, t 520.84, P 5 0.39) over which males were observed in a 1983–1984 study when no legs were removed (see Fincke 1992a).The longest span over which a marked M. coerulatus wasseen was 193 days. This male emerged in the insectary in1996 and was resighted as an old individual in 1997 (i.e., hewas not included in the above sample). Controlling for theseasonal decline in body size, wing length was correlatedwith the number of matings a male obtained (partial r 5 0.13,n 5 359, P 5 0.01). In contrast, the span over which a maturemale was seen was not a function of male wing length (partialr 5 0.06, n 5 359, P 5 0.26; partial r 5 20.01, n 5 178males seen more than once, P 5 0.92).
A male defending one of our experimental holes for themaximum time observed in the field (60 days) would have,on average, a realized fitness of only 2.6 6 0.19 survivingoffspring (range 5 0–7), regardless of the number of matings(r 5 0.13, n 5 51, P 5 0.35) or fertilizations obtained (r 50.10, n 5 51, P 5 0.46). At best, he would hit three windowsof opportunity for larval survivorship; at worst, none. In con-trast, a male staying at a site only long enough to obtain asingle mating would produce, on average, 0.89 6 0.18 off-spring (range 0–5). However, he would have a ninefold fitnesspayoff per day of defense (x̄ 5 0.35 6 0.14 offspring perday versus 0.04 6 0.00 for males staying 60 days, t 5 2.25,P 5 0.03). In other words, a male is not assured fitness simplyby maximizing the number of matings or fertilizations. Rath-er, the number of favorable windows to which he contributesoffspring is the best predictor of his success, explaining 74%of variation in realized fitness among mated males in thisexample (r 5 0.87, n 5 98, P 5 0.001).
Contrary to expectations if males were maximizing lifetimefertilization success, the largest territorial males did not de-fend holes for the longest time (Fig. 7). However, tenureduration was correlated positively with body size for terri-
torial males with the relatively short residency time of 25days or less (Fig. 7). Mean residency of a territorial male ata site was 11.4 6 14.1 days. Because the average time toencounter a female was 22.9 6 3.1 days (median 5 14 days,range 1–69 days), these results suggest that the largest, mostcompetitive males remained on a territory only long enoughto obtain one to two matings. Indeed, controlling for the daya male was first seen, body size was correlated with the num-ber of different tree hole sites a male frequented and leftvoluntarily (partial r 5 0.27, n 5 82 males seen at more thanone site, P 5 0.01).
DISCUSSION
Adult Fitness in Megaloprepus coerulatus
Because the experimental holes were designed as a bestcase scenario for larval survivorship, our results likely over-estimate survivorship of M. coerulatus in natural holes forseveral reasons. First, we limited egg input to one clutchevery two weeks. The number of surviving larvae was ef-fectively limited by the size of a tree hole and the time spanover which it received eggs, rather than by the total numberof eggs it received. At newly created larval habitats in a fallentree, adult activity can be five times higher than average, andconsequently, some holes receive multiple clutches per week,even though the total number of emerging adults remains low(Fincke 1992a). Secondly, we curtailed egg input at the endof wet season and kept holes filled, preventing mortality thatwould result from hole drying (see Fincke 1994). Finally,whereas other major intraguild predators were excluded fromall but one of our experimental holes, about half of the naturaltree holes on BCI that are defended by M. coerulatus are alsooccupied by Triacanthagyna or Gynacantha dragonflies.These predators reduce the total number of M. coerulatus thata hole can support, and in extreme cases, can eliminate allM. coerulatus offspring (Fincke 1992a, 1999).
Even under the favorable conditions for offspring survi-vorship in our experiment, variation in reproductive successamong parents was extreme, with typically less than half ofthe parents realizing any fitness from their reproductive in-vestment. Contrary to a common prediction of behavioralecologists, the fitness that a female realized from using aparticular larval habitat was not proportional to the numberof eggs she laid there. Larger sample sizes would have, inhindsight, provided more power for our statistical tests, butat considerable cost in terms of time and man power, giventhe low encounter rate with ovipositing females in this sys-tem. However, even if the low correlation between clutchsize and fitness were found to be significant with a largersample size, clutch size would still explain much less of thevariance than does hatching span. Larger than average clutch-es did not improve a female’s realized fitness because hatch-ing span was not correlated with clutch size, even in ourlarger sample where the power to detect a small but signif-icant trend was more robust. But more importantly, the win-dows of opportunity revealed by the microsatellite data offerthe explanation for the lack of strong correlation betweenclutch size and fitness. These unpredictable windows alsoexplain why the number of matings a male obtains is a poorindicator of the fitness he realizes from a defended hole. Not

770 O. M. FINCKE AND H. HADRYS
only are many of the females that use a given hole unsuc-cessful in producing a single surviving offspring, but in-creasing the number of mates that contribute embryos to ahole decreases the fitness gain per successful mate (see Table1). Females hedge their bets by producing highly asynchro-nous clutches, which effectively makes multiple matings ata given hole more redundant than they might be otherwise.
The maximum observed hatching span within a givenclutch in this study was 184 days, greater than that knownfor any odonate laying nondiapause eggs (see Miller 1992;Corbet 1999). Although under natural conditions they de-velop in bark above the water line, eggs in our study werein a monolayer, submerged under water, eliminating the pos-sibility of a moisture or oxygen gradient (the latter is a knownhatching cue in an odonate, Miller 1992). A sudden drop inambient temperature, which occurs during a heavy rain, is aproximate cue for hatching in M. coerulatus; neonates sufferrelatively low mortality before reaching the water when theycrawl over wet bark as opposed to dry bark (A. van’t Hofand O. Fincke, unpubl. data). However, eggs within a clutchdevelop at different rates, suggesting that females have someultimate control over the timing of egg hatch. Little is knownabout inherent control of the rate of embryogenesis, partic-ularly in odonates (see Corbet 1999), but female age can playa role (reviewed by Mousseau and Dingle 1991; Sternberg1995). However, in our study a female’s relative age had nodetectable effect on hatching span; other possible factors,such as egg size and laying order, are under investigation.
Aside from greater insurance against mortality betweenegg laying and the time neonates enter a hole, what mightfemales gain by laying a large clutch? In another study, (O.Fincke, unpubl. ms.), a clutch of 20 eggs developing in alarge hole with abundant prey produced, on average, threesurviving offspring, about four fewer than a clutch of 75 eggs.However, the latter produced no more offspring than a clutchof 100 eggs. Moreover, a clutch of 20 eggs produced largeroffspring than clutches of 75 or 100. In our experiment, eventhe largest clutches were ineffective at swamping cannibal-istic larvae already present in the holes; large clutches arealso ineffective in swamping preexisting dragonfly larvae aswell (O. Fincke, unpubl. ms.).
The first egg to hatch did so more quickly in clutches withmore than 50 eggs, although this apparent facilitation couldbe a larval response to increased competition, rather than aphenomenon under female control. This effect did not trans-late into a detectable advantage in our experiment, but mightbe advantageous when multiple females compete for an un-occupied hole. For example, if multiple clutches had beendeposited simultaneously into our experimental holes, largerclutches would be expected to have a preemptive advantageover smaller ones, at least for the initial egg input. Once ahole has been colonized by neonates however, a four-daylead on hatching from any subsequent clutch would be un-likely to improve the odds of hitting the next unpredictablewindow. Alternatively, a female might lay excess eggs if itis more difficult for her to find a tree hole than it is to amassthe energy needed to produce eggs. The female that laid anunusually large clutch did so early in wet season, after weeksduring which tree holes would have been dry. She may have‘‘dumped’’ her entire load of mature eggs simply because it
had been a long time since she encountered a water-filledtree hole. For example, the best predictor of clutch size inCoenagarion puella, a British damselfly constrained from lay-ing on rainy or overcast days, is the time since the last clutchwas laid (Banks and Thompson 1987).
Although studies on a variety of insects (often under labconditions) have demonstrated a positive correlation betweenfemale body size and fecundity (e.g., Boggs 1986; Wiklundand Karlsson 1988; Gwynne 1988; Honek 1993), for M. coe-rulatus, that relationship appears to be a statistical artifact.Body size declined as the season progressed, coupled with atendency for clutch size to decrease independently over thewet season. Rather, for the few females that were resighted,larger females were seen over a longer time span, althoughno such relationship was detected in males, which were re-sighted more often. However, after controlling for the sea-sonal decline in body size, larger males were still more likelyto mate. This result corroborates earlier data demonstratingthat the large size advantage in mating occurs because largerelative size best predicts the winners of territorial fights(Fincke 1992a).
Megaloprepus coerulatus has a maximum adult life spanthat is longer than that for any odonate species studied todate (reviewed by Corbet 1999). Adults roam widely overthe island (Fincke 1984b), and must use more than a singletree hole in order for the yearly replacement of the population,which, on BCI, appears to have been stable over the past 15years (Fincke 1998). Although our study was not designedto measure fitness of individuals over their life span, giventhe dynamics of larval survivorship, our results suggest thatamong parents, lifetime fitness is unlikely to be correlatedwith the total number of embryos produced. Rather, the num-ber of tree holes (or windows) over which both males andfemales distribute their embryos should be a better surrogateof lifetime fitness than the traditional ones such as female-clutch size or male-territory tenure duration or total fertil-izations. Indeed, females appear to rarely, if ever, lay multipleclutches in the same tree hole. Although staying at a territoryas long as possible maximizes a male’s fertilization successat a given territory (Fincke 1992a), that strategy is unlikelyan optimal one for the most competitive males. Upon leavingone site, the largest males have the best chance of securinganother territory, not because they are better at finding them,but because they can most easily oust any male defendinganother site. A simulation model incorporating field estimatesfor larval growth rates, male and female encounter rates, andtravel times between sites, demonstrated that fitness was max-imum for males that stayed at a site only long enough toobtain one to two matings (O. Fincke, O. Eikemeier, and H.Hadrys, unpubl. ms). This is consistent with our current find-ing that the largest males changed territorial sites after arelatively short time.
Our results have broad implications for studies of repro-ductive strategies, particularly in animals lacking parentalcare. First, understanding the pattern of offspring survivor-ship clarifies how larval ecology shapes adult reproductivestrategies. For M. coerulatus, unpredictable larval survivor-ship favors bet-hedging strategies by adult females and malesin ways that could not be predicted by focusing solely onthe dynamics of adult reproductive behavior. Females bet

771SURVIVORSHIP CHALLENGES FITNESS ESTIMATES
hedge by producing clutches with extreme hatch asynchrony,thereby increasing the chances that at least one offspringhatches during a window of opportunity. The timing of thesewindows cannot be anticipated by ovipositing females or ter-ritorial males, which explains why neither males nor femalesdiscriminate against tree holes that are already occupied bycon- or heterospecific odonate larvae (Fincke 1992a). Ourresults also explain the otherwise puzzling observations thatvery large males often abandon tree holes even though theyare not challenged by competitors, and why the newest larvalhabitats are those where male competition is greatest.
Secondly, uncertain patterns of offspring survivorship con-strains natural selection on female fecundity and sexual se-lection on males. Females and males that maximize the num-ber of larval habitats to which they contributed offspringwould have a selective advantage over those that maximizetheir input at a given site. Because neither clutch size norfertilization success was a good predictor of parental fitness,selection on mated individuals was constrained. If fitness isnot proportional to male fertilization success or female fe-cundity, then using these traditional fitness surrogates to es-timate the opportunity for selection or to calculate selectiondifferentials for adult traits, as is done often (e.g. Arnold andWade 1984; Brown 1988), can be misleading. For example,had we used territory tenure (or mating success) as a fitnesssurrogate, we would have incorrectly concluded that sexualselection on male size was stablilizing (Fig. 7). In M. coe-rulatus, both sexual and natural selection favors large males.In addition to a mating advantage, large males are best ableto maximize their chances of producing surviving offspringby distributing offspring among the greatest number of larvalhabitats. Jia and Greenfied (1997) provide a contrasting casein which a trait important for mating success is not neces-sarily the trait that makes a successful parent. Under poorlarval conditions, the offspring of male wax moths that werenot preferred mates experienced greater growth and survi-vorship than the offspring of preferred males.
Finally, quantifying fitness as the number of surviving off-spring is critical to understanding population processes. Theeffective population size (Ne), an important determinant ofpopulation stability and health (Soule and Wilcox 1980),would be significantly overestimated for M. coerulatus if cal-culated by simply noting the number of reproductive adults.Although most sexually mature females and 30% of adultmales are expected to produce embryos (Fincke 1992a), ourcurrent results suggest that fewer than half of all parentsactually contribute genes to the next generation. Mortalityby intraguild predators (Fincke 1992b, 1999) would furtherreduce our estimate of Ne. We are currently measuring theeffects of this low offspring survivorship on the genetic struc-ture of Megaloprepus coerulatus, whose habitat is restrictedto the increasingly fragmented forests of the Neotropics.
The uncoupling of clutch size or fertilization success fromrealized fitness at a site is likely to be common wheneverecological conditions that cannot be anticipated by adultsresult in windows that disproportionately favor offspring sur-vivorship. Whereas larval cannibalism is the mechanism thatmakes offspring survivorship unpredictable in tropical treeholes, in a bromeliad-breeding frog, snake predators that con-sume entire egg clutches produce a similar effect. Although
larger frogs produced larger egg clutches, Haugen (2001)found no correlation between the size of a female and thenumber of offspring surviving to metamorphosis. Even fornoncontainer breeders, the number of embryos entering ahabitat is usually far in excess of its carrying capacity, andlarval survivorship is typically low (e.g., Berryman 1988,Ohgushi 1991; Willis and Hendrick 1992; Tinkle and Dun-ham 1993; Duffy 1994; Dempster and McLean 1998; Dixonet al. 1999). Ecological factors such as deterioration of larvalhabitats or fluctuations in the density of food, predators, can-nibals, or parasites can result in unpredictable windows ofoffspring survivorship (e.g., Smith 1987; Newman 1989; Soand Dugeon 1989; Morin et al. 1990; Messina 1991; Anholt1994; Dixon et al. 1999). Our findings call for caution in thedesign of future studies aimed at obtaining evolutionarymeaningful estimates of fitness, particularly for species withcomplex life histories.
ACKNOWLEDGMENTS
We thank S. Yanoviak, T. Gerkin, B. Strauss, J. McFarland,and K. Montague for field assistance and W. Schroth formicrosatellite development. We are grateful to D. Mock, M.Kaspari, B. Schierwater, and two anonymous reviewers forhelpful comments on earlier drafts of the manuscript. Wethank C. McCallister for figures and the Smithsonian TropicalResearch Institute, Panama, for logistical support. The workwas supported by National Science Foundation grant IBN-9408143 to OMF and Deutsche Forschungsgemeinschaftgrants HA 1947/2-1 and 2-2 to HH.
LITERATURE CITED
Alcock, J., and Houston, T. F. 1996. Mating systems and male sizein Australian Hylaeine bees (Hymenoptera: Colletidae). Ethol-ogy 102:591–610.
Andersson, M. 1994. Sexual selection. Princeton Univ. Press,Princeton, NJ.
Anholt, B. R. 1994. Cannibalism and early instar survival in a larvaldamselfly. Oecologia 99:60–65.
Arnold, S. J., and Wade, M. J. 1984. On the measurement of naturaland sexual selection: applications. Evolution 38:720–734.
Banks, M. J., and Thompson, D. J. 1987. Lifetime reproductivesuccess of females of the damselfy Coenagrion puella. J. Anim.Ecol. 56:815–132.
Barrowclough, G. F., and R. F. Rockwell. 1993. Variance in lifetimereproductive success: estimation based on demographic data.Am. Nat. 141:281–295.
Berryman, A. A. ed., 1988. Dynamics of forest insect populations:patterns, causes, implications. Plenum Press, New York.
Boggs, C. 1986. Reproductive strategies of female butterflies: var-iation in and on fecundity. Ecol. Entomol. 11:7–15.
Brown, D. 1988. Components of lifetime reproductive success. Pp439–453 in T. H. Clutton-Brock, ed. Reproductive success: stud-ies of individual variation in contrasting breeding systems. Univ.of Chicago Press, Chicago, IL.
Choe, J. C. 1994. Sexual selection and mating systems in Zorotypusgueneyi Choe (Insecta: Zoraptera). I. Dominance hierarchy andmating success. Behav. Ecol. Sociobiol. 34:87–93.
Clutton-Brock, T. H., Ed. 1988. Reproductive success: studies ofindividual variation in contrasting breeding systems. Univ. ofChicago Press, Chicago, IL.
Conner, J. 1989. Field measurements of natural and sexual selectionin the fungus beetle, Boltitotherus conrutus. Evolution 42:736–749.
Corbet, P. S. 1999. Dragonflies: behavior and ecology of Odonata.Cornell Univ. Press, Ithaca, NY.

772 O. M. FINCKE AND H. HADRYS
Dixon, P. A., M. J. Milicich, and G. Sugihara. 1999. Episodic fluc-tuations in larval supply. Science 283:1528–1530.
Dempster, J. P., and I. F. G. McLean, Eds. 1998. Insect populations:in theory and in practice. Kluwer Academic Press, London.
Duffy, W. G. 1994. Demographics of Lestes disjunctus (Odonata:Zygoptera) in a riverine wetland. Can. J. Zool. 72:910–917.
Ender, A., K. Schwenk, T. Staedler, B. Streit, and B. Schierwater.1996. RAPD identification of microsatellites in Daphnia. Mol.Ecol. 5:437–441.
Fincke, O. M. 1984a. Sperm competition in the damselfly Enallagmahageni Walsh (Odonata: Coenagarionidae). Behav. Ecol. Socio-biol. 10:293–240.
———. 1984b. Giant damselflies in a tropical forest: reproductivebiology of Megaloprepus coerulatus with notes on Mecistogaster(Zygoptera: Pseudostigmatidae). Adv. Odonatol. 2:13–27.
———. 1986. Lifetime reproductive success and the opportunityfor selection in a nonterritorial damselfly (Odonata: Coenagrion-idae). Evolution 40:791–803.
———. 1988. Sources of variation in lifetime reproductive successin a non-territorial damselfly. Pp 24–42 in T. H. Clutton-Brock,ed. Reproductive success: studies of individual variation in con-trasting breeding systems. Univ. of Chicago Press, Chicago, IL.
———. 1992a. Consequences of larval ecology for territorialityand reproductive success of a neotropical damselfly. Ecology73:449–462.
———. 1992b. Interspecific competition for tree holes: conse-quences for mating systems and coexistence in neotropical dam-selflies. Am. Nat. 139:80–101.
———. 1994. Population regulation of a tropical damselfly in thelarval stage by food limitation, cannibalism, intraguild predationand habitat drying. Oecologia 100:118–127.
———. 1996. Larval behaviour of a tree hole-breeding damselfly:territoriality or size dominance? Anim. Behav. 51:77–87.
———. 1998. The population ecology of Megaloprepus coerulatusand its effect on species assemblages in water-filled tree holes.Pp. 391–416 in J. P. Dempster and I. F. G. McLean, eds. Insectpopulations: in theory and in practice. Kluwer Academic Press,London.
———. 1999. Organization of predator assemblages in Neotropicaltree holes: effects of abiotic factors and priority. Ecol. Entomol.24:13–23.
Fincke, O. M., S. P. Yanoviak, and R. D. Hanschu. 1997. Predationby odonates depresses mosquito abundance in water-filled treeholes in Panama. Oecologia 112:244–253.
Gwynne, D. T. 1988. Courtship feeding and the fitness of femalekatydids (Orthoptera: Tettigoniidae). Evolution 42:545–555.
Hadrys, H., and M. T. Siva-Jothy. 1994. Unravelling the compo-nents that underlie insect reproductive traits using a simple mo-lecular approach. Pp. 211–228 in B. Schierwater, G. P. Wagner,B. Streit, and R. DeSalle, eds. Molecular ecology and evolution:approaches and applications. Birkhaeuser, Basel, Switzerland.
Hadrys, H., M. Balick, and B. Schierwater. 1992. Applications ofrandom amplified polymorphic DNA (RAPD) in molecular ecol-ogy. Mol. Ecol. 1:55–63.
Hadrys, H., B. Schierwater, R. DeSalle, S. L. Dellaporta, and L.W. Buss. 1993. Determination of paternity in dragonflies byrandom amplified polymorphic DNA fingerprinting. Mol. Ecol.2:79–87.
Haugen, L. 2001. Privation and uncertainty in the small nursery ofPeruvian tadpoles: larval ecology shapes parental mating system.Ph.D. thesis, University of Oklahoma, Norman, OK.
Honek, A. 1993. Intraspecific variation in body size and fecundityin insects: a general relationship. Oikos 66:483–492.
Howard, R. D. 1988. Reproductive success in two species of an-urans. Pp. 99–113 in T. H. Clutton-Brock, ed. Reproductivesuccess: studies of individual variation in contrasting breedingsystems. Univ. of Chicago Press, Chicago, IL.
Jia, F.-Y., and Greenfield, M. D. 1997. When are good genes good?
Variable outcomes of female choice in wax moths. Proc. R. Soc.Lond. 264:1057–1063.
Leigh, E. G. 1999. Tropical forest ecology: a view from BarroColorado Island. Oxford Univ. Press, Oxford, U.K.
McLain, D. K. 1991. Components of variance in male lifetime cop-ulatory and reproductive success in a seed bug. Behav. Ecol.Sociobiol. 29:121–126.
Messina, F. J. 1991. Life-history variation in a seed beetle: adultegg-laying vs. larval competitive ability. Oecologia 85:447–455.
Miller, P. L. 1992. The effect of oxygen lack on egg hatching inan Indian dragonfly, Potamarcha congener. Physiol. Entomol.17:68–72.
Moore, A. J. 1990. The evolution of sexual dimorphism by sexualselection: the separate effects of intrasexual selection and in-tersexual selection. Evolution 44:315–331.
Morin, P. J., S. P. Lawler, and F. A. Johnson. 1990. Ecology andbreeding phenology of larval Hyla andersonii: the disadvantagesof breeding late. Ecology 71:1590–1598.
Mousseau, T. A., and H. Dingle. 1991. Maternal effects in insectlife histories. Annu. Rev. Entomol. 36:511–534.
Newman, R. A. 1989. Developmental plasticity of Scaphiopus cou-chii tadpoles in an unpredictable environment. Ecology 70:1175–1787.
Ohgushi, T. 1991. Lifetime fitness and evolution of reproductivepattern in the herbivorous lady beetle. Ecology 72:1110–2122.
Olsson, M., and R. Shine. 1996. Does reproductive success increasewith age or with size in species with indeterminant growth? Acase study using sand lizards (Lacerta agilis). Oecologica 105:175–178.
Partridge, L. 1988. Lifetime reproductive success in Drosophila.Pp. 11–23 in T. H. Clutton-Brock, ed. Reproductive success:studies of individual variation in contrasting breeding systems.Univ. of Chicago Press, Chicago, IL.
Siva-Jothy, M. T., and H. Hadrys. 1998. Molecular analysis ofindividuals, populations, species, and higher taxa. Pp. 37–53 inR. DeSalle, and B. Schierwater, eds, Molecular approaches toecology and evolution. Birkhaeuser, Basel, Switzerland.
Smith, D. C. 1987. Adult recruitment in chorus frogs: effects ofsize and date at metamorphosis. Ecology 68:344–350.
So, P.-M., and D. Dugeon. 1989. Life-history responses of larvip-arous Boettcherisca formosensis (Diptera: Sarcophagidae) to lar-val competition for food, including comparisons with oviparousHemipyrellia ligurriens (Calliphoridae). Ecol. Entomol. 14:349–356.
Soule, M. E., and B. A. Wilcox. 1980. (eds) Conservation biology.an evolutionary-ecological perspective. Sinauer Associates, Sun-derland, MA.
Sternberg, K. 1995. Influence of oviposition date and temperatureupon embryonic development in Somatochlora alpestris and S.arctica. J. Zool. 235:163–174.
Thornhill, R., and J. Alcock. 1983. The evolution of insect matingsystems. Harvard Univ. Press, Cambridge, MA.
Tinkle, D. W., A. E. Dunham, and J. D. Congdon. 1993. Life historyand demographic variation in the lizard Sceloporus graciosus: along-term study. Ecology 74:2413–2429.
Waage, J. K. 1984. Sperm competition and the evolution of odonatemating systems. Pp. 251–290 in R. L. Smith, ed. Sperm com-petition and the evolution of animal mating systems. AcademicPress,Orlando, FL.
Waage, J. K. 1986. Evidence for widespread sperm displacementability among Zygoptera (Odonata) and the means for predictingits presence. Biol. J. Linn. Soc. 28:285–300.
Wiklund, C., and B. Karlsson. 1988. Sexual size dimorphism inrelation to fecundity in some Swedish stayrid butterflies. Am.Nat. 131:132–138.
Willis, L. D., Jr., and Hendrick, A. C. 1992. Life history, growth,survivorship, and production of Hydropysche slossonae in MillCreek, Virginia. J. N. Am. Benthol. Soc. 11:290–303.
Corresponding Editor: C. Boggs




















