
UNIVERSIDADE FEDERAL DE MATO GROSSO INSTITUTO DE ...
111
UNIVERSIDADE FEDERAL DE MATO GROSSO INSTITUTO DE BIOCIÊNCIAS PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA E BIODIVERSIDADE REDE PRÓ CENTRO-OESTE Vandinelma de Oliveira Vieira POTENCIAL BIOTECNOLÓGICO DE MACRO BASIDIOMICETOS NO PROCESSO DE PRODUÇÃO DE MUDAS DE TOMATE E CONTROLE DE Sclerotinia sclerotiorum, AGENTE CAUSAL DO MOFO BRANCO CUIABÁ MATO GROSSO - BRASIL 2021
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of UNIVERSIDADE FEDERAL DE MATO GROSSO INSTITUTO DE ...

UNIVERSIDADE FEDERAL DE MATO GROSSO
INSTITUTO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA E BIODIVERSIDADE
REDE PRÓ CENTRO-OESTE
Vandinelma de Oliveira Vieira
POTENCIAL BIOTECNOLÓGICO DE MACRO BASIDIOMICETOS NO PROCESSO
DE PRODUÇÃO DE MUDAS DE TOMATE E CONTROLE DE Sclerotinia
sclerotiorum, AGENTE CAUSAL DO MOFO BRANCO
CUIABÁ
MATO GROSSO - BRASIL
2021

ii
VANDINELMA DE OLIVEIRA VIEIRA
POTENCIAL BIOTECNOLÓGICO DE MACRO BASIDIOMICETOS NO PROCESSO
DE PRODUÇÃO DE MUDAS DE TOMATE E CONTROLE DE Sclerotinia
sclerotiorum, AGENTE CAUSAL DO MOFO BRANCO
CUIABÁ
MATO GROSSO - BRASIL
2021
Tese apresentada à Universidade Federal de
Mato Grosso, como parte das exigências do
Programa de Pós-Graduação em
Biotecnologia e Biodiversidade rede Pró
Centro-Oeste, para obtenção do título de
Doctor Scientiae.
Orientador: Euziclei Gonzaga de Almeida.
Coorientador: Félix Gonçalves Siqueira.

iii
V658p Vieira, Vandinelma de Oliveira.
Potencial biotecnológico de macro basidiomicetos no processo de produção de mudas de tomate e controle de Sclerotinia sclerotiorum, agente causal do mofo branco / Vandinelma de Oliveira Vieira. -- 2021
110 f. : il. color. ; 30 cm.
Orientador: Euziclei Gonzaga de Almeida. Co-orientador: Félix Gonçalves Siqueira.
Tese (doutorado) – Universidade Federal de Mato Grosso, Rede Pró Centro-Oeste, Programa de Pós- Graduação em Biotecnologia e Biodiversidade, Cuiabá, 2021.
Inclui bibliografia.

iv
MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL DE MATO GROSSO
PRÓ-REITORIA DE ENSINO DE PÓS-GRADUAÇÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA E BIODIVERSIDADE -
REDE PRÓ CENTRO- OESTE
FOLHA DE APROVAÇÃO
TÍTULO : "Potencial biotecnológico de macro basidiomicetos no processo de produção
de mudas de tomate e controle de Sclerotinia sclerotiorum, agente causal do mofo branco"
AUTOR : Doutoranda Vandinelma de Oliveira Vieira
Tese de Doutorado defendida e aprovada em 26/11/2021.
Composição da Banca Examinadora:
Presidente Banca/Orientador Doutor: Euziclei Gonzaga de Almeida
Instituição : UNIVERSIDADE FEDERAL DE MATO GROSSO
Examinador Interno: Doutora Leonarda Grillo Neves
Instituição : UNIVERSIDADE DO ESTADO DE MATO GROSSO
Examinador Externo Doutor Eustáquio Dias
Instituição : UNIVERSIDADE FEDERAL DE LAVRAS
Examinador Externo Doutor Elias Alves
Instituição : EMBRAPA AGROENERGIA
Examinador Externo Doutora Daiana Wischral
Instituição: EMBRAPA AGROENERGIA
CUIABÁ, 26/11/2021.
Prof. Dr. Willian Krause
Coordenador Estadual PPGBB-MT

v
Dedico aos meus pais Manoel e Maria Aparecida, irmãos Osvaldo, Flávia, Wellington
e Mariana, sobrinhos Ana, Maria e Ricardo. Por nunca deixarem de me incentivar e
apoiar.
Ofereço ao meu filho Guilherme, por não me permitir desistir, minha fonte de amor e
compreensão.

vi
RESUMO GERAL
VIEIRA, Vandinelma de Oliveira, M.S., Universidade Federal de Mato Grosso,
novembro de 2021. Potencial biotecnológico de macromicetos no processo
de produção de mudas de tomate e controle de Sclerotinia sclerotiorum,
agente causal do mofo branco. Orientador: Euziclei Gonzaga de Almeida. Co-
Orientador: Felix Gonçalves Siqueira.
As tecnologias aplicadas na fungicultura podem ser uma das alternativas
implementada em áreas próximas de agroindústrias. O cultivo e a agroindústria de
óleo de palma (dendê) geram grandes quantidades de resíduos e efluentes, que
podem ser utilizados como matéria-prima em outros processos produtivos associados
aos macromicetos como forma de gerar renda e mitigar impactos ambientais. Com
base na perspectiva de expansão da fungicultura e geração de resíduos por parte da
dendeicultura, o objetivo foi realizar experimentos com macromicetos e resíduos da
dendeicultura para desenvolvimento de um substrato para produção de mudas de
tomate, testar um sistema de co-cultivo de tomate e cogumelos e por fim selecionar e
avaliar o potencial dos macromicetos no controle de Sclerotinia sclerotiorum. Os
resultados obtidos através da revisão de literatura demonstram o grande potencial
desses macrofungos em diferentes áreas, desde a indústria até o setor agropecuário.
Os testes para desenvolvimento de substratos para produção de mudas, por exemplo,
podemos ressaltar o alto potencial de indução de germinação e crescimento, aumento
da vida de prateleira das mudas no substrato inoculado com A. bisporus + resíduos
de dendê, enquanto que no sistema de co-cultivo de tomate e cogumelos ficou
evidente os bons rendimentos de cada cultura, antecipou a fase reprodutiva no
tomate, aumento do ciclo reprodutivo do cogumelo para 120 dias. Dos 17 fermentados
de macrofungos testados para a supressão de S. sclerotiorum os fungos O. canarii e
L. arvalis evidenciam pela primeira vez, excelentes fontes de metabólitos antifúngicos
capazes de inibir o crescimento do micélio de S. sclerotiorum, total ou parcialmente.
A alta atividade antifúngica do sobrenadante de O. canarii BRM-044600 foi
provavelmente devido à provável presença de composotos químicos como as
estrobilurinas. De modo geral pode-se afirmar que alguns macromicetos tem grande
potencialidade na geração de bioprodutos e bioinsumos para o setor agroindustrial.
Palavras-chave: fungicultura, promotor crescimento planta, macrofungos, controle
biológico, biofungicida.

vii
GENERAL ABSTRACT
VIEIRA, Vandinelma de Oliveira, M.S., Federal University of Mato Grosso, 2020.
Biotechnological potential of macromycetes in the tomato seedling
production process and control of Sclerotinia sclerotiorum, causal agent of
white mold. Advisor: Euziclei Gonzaga de Almeida. Co-Advisor: Felix Gonçalves
Siqueira.
The technologies applied in fungiculture can be one of the alternatives implemented in
areas close to agro-industries. The cultivation and agroindustry of palm oil (palm oil)
generate large amounts of waste and effluents, which can be used as raw material in
other production processes associated with macromycetes as a way of generating
income and mitigating environmental impacts. Based on the perspective of expansion
of fungiculture and generation of residues by oil palm cultivation, the objective was to
carry out experiments with macromycetes and oil palm residues to develop a substrate
for the production of tomato seedlings, test a tomato co-cultivation system and
mushrooms and finally to select and evaluate the potential of macromycetes to control
Sclerotinia sclerotiorum. The results obtained through the literature review
demonstrate the great potential of these macrofungi in different areas, from industry to
the agricultural sector. Tests for the development of substrates for seedling production,
for example, can highlight the high potential for inducing germination and growth,
increasing the shelf life of seedlings in the substrate inoculated with A. bisporus + palm
residues, while in the co-cultivation of tomato and mushrooms, it was evident the good
yields of each culture, anticipated the reproductive phase in tomato, increasing the
reproductive cycle of the mushroom to 120 days. Of the 17 fermented macrofungi
tested for S. sclerotiorum suppression, the fungi O. canarii and L. arvalis show for the
first time excellent sources of antifungal metabolites capable of inhibiting the growth of
the mycelium of S. sclerotiorum, totally or partially. The high antifungal activity of O.
canarii BRM-044600 supernatant was probably due to the probable presence of
chemical compounds such as strobilurins. In general, it can be said that some
macromycetes have great potential in the generation of bioproducts and bioinputs for
the agro-industrial sector.
Keywords: fungiculture, plant growth promoter, macrofungi, biological control,
biofungicide

6
SUMÁRIO
ESTRUTURA DA TESE ...................................................................................................................... 7
1. INTRODUÇÃO GERAL ............................................................................................................... 9
2. JUSTIFICATIVA ......................................................................................................................... 11
3. OBJETIVOS ................................................................................................................................ 12
3.1 Objetivo Geral ........................................................................................................................... 12
3.2 Objetivos Específicos .............................................................................................................. 12
4. REVISÃO BIBLIOGRÁFICA .................................................................................................... 13
4.1 Fungicultura de cogumelos comestíveis: aspectos gerais ................................................ 13
4.2 Brasil: Potencial integração entre produção de cogumelos comestíveis e a geração de
biofertilizantes e bioestimulantes ................................................................................................. 15
4.3 Brasil: Fungicultura de cogumelos comestíveis .................................................................. 18
4.4 Sistema produtivo de cogumelos no Brasil .......................................................................... 20
4.5 Biomassas vegetais residuais (BVRs) na produção de cogumelos................................. 25
4.6 SMS - Biomassa residual pós-colheita dos cogumelos ..................................................... 27
4.6.1 SMS - formulação de biofertilizantes e bioestimulantes ................................................. 28
4.6.2 SMS: biocontrole de fitopatógenos .................................................................................... 31
4.7 Fermentados fúngicos ou SMS e novos produtos/processos para agricultura Brasileira
........................................................................................................................................................... 35
4.8 Cultura do tomate ............................................................................................................... 36
4.9 Fitopatógeno do tomate – Sclerotinia sclerotiorum ....................................................... 37
Referências ...................................................................................................................................... 38
5 ARTIGO 01 – Artigo publicado na revista Saudi Journal of Biological Sciences (2022)
2756-2765. https://doi.org/10.1016/j.sjbs.2021.12.058 ................................................................ 51
6 ARTIGO 02 – Publicado na revista World Journal of Microbiology and Biotechnology
(2021) 37:203. https://doi.org/10.1007/s11274-021-03172-6 ...................................................... 81
7 CONSIDERAÇÕES FINAIS ............................................................................................ 105
8 REFERENCIAS GERAL ................................................................................................. 106
9 ANEXO............................................................................................................................. 108

7
ESTRUTURA DA TESE
Para facilitar a compreensão, o trabalho foi dividido em artigos, procurando responder
aos objetivos propostos.
*Artigos publicados:
Artigo 1: Vandinelma Oliveira Vieira, Aparecido Almeida Conceição, Joice Raisa Barbosa Cunha, Antony Enis Virginio Machado, Euziclei Gonzaga de Almeida, Eustáquio Souza Dias, Lucas Magalhães Alcantara, Robert Neil Gerard Miller, Félix Gonçalves de Siqueira. A new circular economy approach for integrated production of tomatoes and mushrooms. Saudi Journal of Biological Sciences, 2022. https://doi.org/10.1016/j.sjbs.2021.12.058.
Introdução Geral
Revisão Bibliográfica
Artigo 01 Artigo 02
Justificativa e Objetivos
*Desenvolvimento de substrato e cocultivo com macromicetos e residuos
da dendeicultura
Uso de sementes de tomate e fungicultura. Cocultivo de tomate e Pleurotus Ostreatus
Resultado promissor para: Substrato á base de colonizado
de A.bisporus + residuos de dendê; cocultivo com redução de ciclo do tomate e aumento
do ciclo do cogumelo.
*Supressão do crescimento de Sclerotinia
sclerotiorum por macromicetos
Screening de macromicetos com potencial de supressão de
S. sclerotiorum e AnáliseUHPLC-MS
O. canarii é promissor na produção de múltiplas
estrobirulinas capazes de controlar S. sclerotiorum

8
Artigo 2: de Oliveira Vieira, V., Conceição, A.A., de Oliveira Gorgulho Silva, C. et al. Characterization of extracellular secondary metabolites in Oudemansiella canarii BRM-044600 displaying antifungal activity against the phytopathogen Sclerotinia sclerotiorum. World J Microbiol Biotechnol 37, 203 (2021). https://doi.org/10.1007/s11274-021-03172-6

9
1. INTRODUÇÃO GERAL
O tratamento e aproveitamento de subprodutos agrícolas tem ganhado
destaque na agenda prioritária do setor do agronegócio, com vistas ao uso, no sentido
de dirimir passivos ambientais e aumentar a lucratividade com a geração de novos
bioprodutos, bioinsumos e bioprocessos. Os resíduos sólidos do processo de
industrialização, quando não adequadamente descartados causam impactos
significativos ao solo, ar e água. Resíduos lignocelulósicos, por exemplo, podem ser
usados como fonte de obtenção de monômeros de açúcares (glicose, xilose, dentre
outros) e compostos fenólicos derivados da quebra de lignina, que podem ser usados
como fonte de carbono e nitrogênio para o cultivo de microrganismos
industriais (Sánchez, 2009; Botelho et al., 2018).
Uma cadeia produtiva relevante é o óleo de palma, no Brasil conhecido como
óleo de dendê, uma cultura oleaginosa altamente produtiva e que apesar do alto
rendimento, gera milhões de toneladas de biomassa lignocelulósica anualmente
(Kurnia et al., 2016). Como exemplo dessa geração o maior produtor de óleo de dendê
no mundo a Indonésia com uma produção média de 37,753 milhões de toneladas e o
volume de biomassa (resíduos) gerada pela indústria pode chegar a 80% do total
(FAOSTAT, 2018; Sulaiman et al., 2011).
Após a retirada do óleo, diversos subprodutos como o cacho, palmiste
(prensagem da amêndoa), fibra do coco entre outros constituem um problema
ambiental-econômico para a agroindústria geradora, visto que tais subprodutos
tornam se resíduos quando não se têm um destino adequado. De acordo com Garcia-
Nunez et al. (2016) a biomassa vegetal sólida residual proveniente da agroindústria
do óleo de dendê é cerca de duas vezes ao óleo produzido.
Em virtude disso e das tendências mundiais em bioeconomia, tem-se procurado
tornar as cadeias produtivas sustentáveis, levando a buscar alternativas que possam
converter a biomassa lignocelulósica em produtos de valor agregado, existem estudos
sobre possível utilização da biomassa residual oriunda da dendeicultura como
combustível sólido (Nasrim et al., 2011), reforço da fibra em matrizes plásticas de
polietileno de alta densidade (Ishak et al., 1998; Sreekala e Sabu, 2011) carvão
ativado (Lua e Jia, 2009) agregados de concreto (Shafigh et al., 2011), alimentação
animal (Carvalho et al., 2012) produção de cogumelos comestíveis (Temu et al.,

10
2016), obtenção de enzimas quando utilizado como meio de crescimento para
microrganismos (Pal e Khanum, 2010). Tais bioativos e outros bioprodutos têm sido
investigados de biomassas pós-colheita de cogumelos (sigla em ingês SMS – Spent
Mushroom Substrate) que no que lhe concerne, usou BRVs (Hassan et al., 2018;
Beckers et al., 2019).
Os fungos filamentosos geradores de cogumelos comestíveis, têm sido
ferramentas biotecnológicas de relevância no desenvolvimento de bioprodutos,
quando cultivados em biomassas residuais de vegetais (BRVs), gerando resultados
como fontes de promotores de crescimento de plantas, biodefensivos, indutores de
resistência e inúmeros compostos produzidos (Nam et al., 2018; ANPC, 2019).
No segmento de desenvolvimento de produtos biotecnológicos associando a
dendeicultura e fungicultura podemos citar a produção de mudas de hortaliças, onde
substratos alternativos podem oferecer potencial tanto na redução de custos quanto
no desenvolvimento de sistemas de cultivo sustentáveis e ecologicamente corretos
(Kraska et al., 2018). O tomate (Solanum lycopersicum) apesar de ser uma planta
conhecida como de difícil cultivo e manejo devido a sua alta susceptibilidade e
exigência nutricional é uma planta modelo com características adequadas, já que
qualquer alteração de composição é expressa nos resultados, além de sua
expressividade de produção e consumo no Brasil.
Outra parte ainda pouco explorado e com grande potencial de crescimento são
as atividades antifugincas produzidas por macromicetos, os mesmos são
reconhecidos por produzirem metabólitos secundários e eliminar a competição com
outros fungos, estrobilurinas e oudemansinas, por exemplo, são produzidos por vários
gêneros Basidiomycota e apresentam atividade antifúngica forte e de amplo espectro
contra leveduras e fungos filamentosos (Schramm et al. 1978; Niego et al. 2021). A
doença do mofo branco causada por Sclerotinia sclerotiorum tem um amplo espectro
de plantas que são suceptiveis e com grande potencial de perdas se não manejada
corretamente e sem relatos na literatura de controle por macromicetos.
Associar resíduos sólidos vegetais a geração de processos tem como resultado
a ampliação do portifólio de produtos para o setor agroindustrial e/ou atrair empresas
de cunho biotecnológico para instalações próximo às agroindústrias de dendê (Morais
et al., 2017; Silva, 2019).

11
2. JUSTIFICATIVA
A agricultura ao longo dos anos vem buscando alternativas sustentáveis para
maximizar a produção, realizar de forma efetiva os tratos culturais e controle de pragas
e doenças. Estudos avaliam a utilização de recursos naturais para obtenção de
ferramentas eficientes na agricultura, mas que gerem menor impacto ambiental.
Considerando que o controle biológico tem se tornado prática comum em
cultivos comerciais sendo associadas a outros métodos, os chamados, manejo
integrado de pragas (MIP), e a crescente reutilização de resíduos nos processos de
produção agrícola a presente tese tem como foco o estudo e desenvolvimento de
produtos com base nos macromicetos.
A Embrapa Agroenergia possui estudos e resultados com uso de BRVs da
dendeicultura na produção de cogumelos comestíveis, moléculas bioativas oriundas
desses processos, entre outros. Uma parceria entre a Universidade Federal do Mato
Grosso e a Embrapa Agroenergia com a inteção de desenvolver um projeto de
reutilização dos resíduos da dendeicultura associados aos macromicetos com a
finalidade de gerar processos e/ou produtos com finalidade comercial. Seguindo em
duas linhas importantes para a agricultura que é a produção de mudas para o setor
da horticultura e no processo de supressão de doença.

12
3. OBJETIVOS
3.1 Objetivo Geral
Relacionar o potencial biotecnológico dos macromicetos no desenvolvimento
de bioinsumos como um susbtrato para produção de mudas de tomate (planta
modelo) a base de fermentados de macromicetos e biomassas residuais da
agroindustria do dendê, como também o desenvolvimento e obtenção de um
extrato bruto (biofungicida) obtidos dos fermentados de macromicetos, capaz
de controlar o Sclerotinia sclerotiorum (mofo-branco).
3.2 Objetivos Específicos
Avaliar os fermentados/colonizados e SMS oriundos da fungicultura na
produção de mudas da cultura do tomate;
Testar os os BVRs de dendê como possível insumo para composição dos
substrates (fertilizantes) para as mudas de tomate;
Verificar a viabilidade produtiva de um sistema de co-cultivo de cogumelos da
espécie Pleurotus ostreatus e tomate;
Averiguar extratos brutos de fermentados de macromicetos com capacidade de
impedir o crescimento e desenvolvimento de Sclerotinia sclerotiorum;
Elucidar o mecanismo de controle do fungo macromicetos no processo de
supressão de Sclerotinia sclerotiorum in vitro (metabolomica untarget).

13
4. REVISÃO BIBLIOGRÁFICA
4.1 Fungicultura de cogumelos comestíveis: aspectos gerais
O termo “cogumelo” é definido como o corpo de frutificação de macrofungos
filamentosos, principalmente distribuídos nos grupos taxonômicos do filo
Basydiomicota e alguns do Ascomytoca, com crescimento epígeo (acima do solo) ou
hipógeo (sob a terra), os quais podem ser classificados para aplicações humanas
como comestíveis, tóxicos e medicinais (Dias; De Brito, 2017). Os macromicetos
formadores de cogumelos desenvolvem-se em diversos habitats, sendo
categorizados em função de características metabólicas/ecologia-interação:
ectomicorrizicos, saprófitos e parasíticos. O maior interesse comercial está nas
espécies saprófitas, os quais possuem sistema de produção agrícola já desenvolvido,
enquanto as espécies ectomicorrizicas precisam de sistemas extrativistas de coleta
manual no meio ambiente natural – “mushroom hunters” (Grangeia et al., 2011; Kalač,
2013).
Dentre os cogumelos comestíveis e comercializados existe uma divisão quanto
aos processos de decomposição, os decompositores primários e secundários. Os
macromicetos decompositores primários são mais eficientes em degradar celulose,
hemicelulose e outros componentes vegetais, alguns exemplos são os Pleurotus spp.
e o Lentinula edodes. Os decompositores secundários geralmente estão presentes
em materiais compostados como o Agaricus spp. É possível notar uma continuação
no processo de transição metabólica de materiais lignocelulósicos e outros materiais
orgânicos para o solo, assim o processo de cultivo sucessivo de diferentes cogumelos
pode proporcionar um sistema completo de compostagem e utilização de resíduos
agrícolas. Isso, no entanto, dificilmente é aplicado em larga escala na produção de
cogumelos (Grimm; Wösten, 2018a).
A fungicultura voltada a produção de cogumelos comestíveis é um ramo da
agricultura que explora comercialmente o cultivo de macromicetos (fungos produtores
de cogumelos) que foram domesticados e com sistema agronômico estabelecido com
boas práticas de cultivos. Os cogumelos como alimento apresentam características
nutricionais interessantes em função de possuir teores de proteínas, carboidratos e
vitaminas elevados, quando desidratados. A prática de cultivo deste alimento remonta
à antiguidade, inclusive podem ter sido um dos alimentos consumidos nos primeiros

14
assentamentos humanos, principalmente em períodos de escassez de alimentos
(outono-inverno) para suprir a necessidade fisiológica de carboidratos e proteínas.
Atualmente, a fungicultura voltada a produção de cogumelos comestíveis e
medicinais é uma atividade praticada com produção em grande escala, principalmente
em países asiáticos e europeus. A produção e consumo no mundo tem aumentado de
forma significativa, principalmente em decorrência a maior divulgação de informações
sobre as qualidades nutricionais, sabor e classificação como alimento funcional por
possuir substâncias bioativas que auxiliam na regulação da microbiota animal. O setor
de produção de cogumelos pode ser classificado em três segmentos principais:
cogumelos comestíveis (com 54% do mercado), cogumelos medicinais (38%) e
cogumelos silvestres comestíveis (8%) (Chang; Wasser, 2018; Grimm; Wösten,
2018a; Royse; Baars; Tan, 2017).
A China é o maior produtor de cogumelos comestíveis no mundo. Em 2014, a
movimentação econômica deste setor neste país foi próxima ao de grãos, vegetais,
frutas e óleo. Os cogumelos comestíveis também auxiliam na economia agrícola de
outros países, tais como Japão, Coreia, Holanda, Espanha, Itália e Estados Unidos.
Em 2013 a produção mundial de cogumelos chegava em torno de 10 milhões de
toneladas, sendo a Ásia o maior produtor com 49,7% da produção, seguido pelas
Américas 25% (principalmente EUA e Canadá) e Europa 19%. Os principais tipos de
cogumelos produzidos na China são Lentinula edodes, Flammulina velutipes e
Pholiota nameko. Em 2009, a produção destes cogumelos chegou a mais de 80%, em
contraste com 5,7% registrado em 1978 (Miles; Chang, 2004; Mingyou et al., 2017).
No Brasil, a produção de cogumelos ainda não é expressiva, apesar do rápido
crescimento, principalmente no estado de São Paulo (Chang; Wasser, 2018;
FAOSTAT, 2014).
O aumento do consumo de cogumelos, em parte se dá devido a sua
composição nutricional (massa seca) na qual apresentam concentrações significativas
de proteínas, carboidratos (fibras alimentares), ácidos graxos poli-insaturados,
vitaminas (C, B1, B2, niacina e biotina) e baixo teor de lipídeos. Outra vantagem no
consumo de cogumelos são os bioativos (beta-glucanas, ergosteróis, metabólitos
secundários, entre outros) (Silva et al., 2022), conferindo-os como alimentos
funcionais, podendo ser inclusive utilizados para obtenção de extratos concentrados

15
de tais bioativos com potencial aplicação farmocológica-medicinal (Bach et al., 2019;
Mocan et al., 2018).
4.2 Brasil: Potencial integração entre produção de cogumelos comestíveis e a
geração de biofertilizantes e bioestimulantes
A indústria de cogumelos comestíveis, medicinais e silvestres, compreende um
mercado que gera mais de 63 milhões de dólares no mundo. O consumo de
cogumelos comestíveis teve grande progressão positiva, principalmente a partir de
1997, alcançando consumos de 4,7 kg per capita. A produção mundial de cogumelos
é concentrada na China devido aos hábitos culturais e culinários (Royse; Baars; Tan,
2017).
No entanto, também é observado o aumento da produção e consumo em
países desenvolvidos como os da América do Norte e Europeus, sendo os Estados
Unidos o segundo maior produtor mundial (Boin; Nunes, 2018). Embora os cogumelos
sejam considerados alimentos funcionais, sendo uma fonte alternativa e abundante
de proteínas, fibras e minerais (Bach et al., 2017), a produção e consumo dos mesmos
em países em desenvolvimento e subdesenvolvidos ainda são baixos, havendo
preferências pelo consumo de cogumelos selvagens (Adedokun; Okomadu, 2017;
Anchang, 2014; Dejene; Oria-De-Rueda; Martín-Pinto, 2017).
No Brasil é observado crescimento na produção e consumo, mas ainda com
maior concentração no estado de São Paulo e na região Sul do país (Dias, 2010). Um
dos maiores desafios da produção de cogumelos comerciais diz respeito a
necessidade de adaptar as condições de crescimento e frutificação fora do habitat
natural de cada espécie.
As tecnologias de cultivo de cogumelos comestíveis baseiam-se na biologia e
ecologia de cada espécie, que fora aprendida por observações na natureza e
adaptações/otimizações estabelecidas por pesquisas nas últimas décadas. A maioria
das espécies de cogumelos comestíveis são fungos saprofíticos, decompositores de
matéria orgânica na serrapilheira de diferentes biomas florestais. A ciclagem dos
nutrientes na serrapilheira por estes fungos é outra evidência sobre os benefícios
estabelecidos entre estes agrupamentos de organismos (Vieira; De Andrade, 2016;
Zied; Cunha et al., 2016).

16
Os mecanismos de decomposição dos fungos formadores de cogumelos, por
vezes encontrados no filo Badiomycota ou Ascomycota, ocorrem em função do
arsenal enzimático, tais como celulases, hemicelulases e ligninases. Estes fungos
podem ser classificados de acordo com o diferencial de produção das enzimas
lignocelulolíticas, quanto a desconstrução dos componentes da parede celular. Os
fungos da podridão branca possuem arsenal enzimático com capacidade de degradar
com mais facilidade a lignina, do que a holocelulose (celulose e hemicelulose). Os
fungos de podridão marrom ou parda é o inverso, preferencialmente degradam a
holocelulose em razão da lignina (Conceição et al., 2018).
O sistema de produção de cogumelos atende os mesmos princípios do habitat
natural do fungo sendo necessário a utilização de substrato que contenha fonte de
carbono e nitrogênio adequado, geralmente, biomassas vegetais base de
componentes vegetais capazes de suprir a necessidade fisiológicas da espécie a ser
cultivada. Além das variáveis nutricionais dos substratos a fungicultura comercial deve
lidar com o ambiente climatizado (umidade, temperatura e oxigenação controlados),
de modo a favorecer a colonização/miceliação do substrato vegetal e a frutificação
dos cogumelos, buscado maior produtividade e evitando a contaminação dos
substratos por outros fungos e bactérias oportunistas (Sánchez, 2010).
A fungicultura, geralmente, utiliza de material vegetal lignocelulósico residuais
(BVRs) ou coprodutos de outras cadeias produtivas agrícolas para formulação dos
substratos. Baseado em uma produção linear, o material residual utilizado para o
preparo do substrato gera um novo resíduo após o cultivo do cogumelo, o pós-cultivo
de cogumelo ou SMS. O destino do SMS pode ser a incorporação no solo como
suplemento a adubação, utilizado como material para compostagens, descartados em
aterros ou descartado de forma inadequado (passivo ambiental) (Meng et al., 2017;
Singh; Chauhan, 2016) (Fig. 01).

17
Figura 01. Produção de cogumelos a partir da biomassa residual vegetal baseado na
economia linear, com geração do resíduo SMS.
Os SMSs têm sido investigados como potenciais fontes de obtenção de
bioprodutos, por exemplo, como material de combustão para produção de energia,
como substrato para o cultivo de outros cogumelos ou como aditivo para ração animal
(Grimm; Wösten, 2018a). Ademais, as possibilidades de uso para estes resíduos são
extensas aplicando-se métodos de extração para obtenção de substâncias com alto
valor agregado: enzimas lignocelulolíticas como xilanases e lacases, ácidos
orgânicos, bioabsorventes, bioplasticos e biocombustíveis (Grimm; Wösten, 2018a;
Kulshreshtha, 2018; Sung et al., 2018; Tummo; Thammasittirong; Thammasittirong,
2018; Zhu; Hongji et al., 2018).
Em um modelo de bioeconomia circular, o resíduo da produção de cogumelos
pode ser inserido no setor agrícola como suplemento para o cultivo de plantas. A
composição da parede celular fúngica, formadora do micélio presente no SMS, possui
moléculas, como a quitina/quitosana que possui propriedades bioativas na indução de
defesa das plantas (Kumaraswamy et al., 2018). O SMS também pode ser incorporado
no solo, aumentando os teores de matéria orgânica e beneficiando os microrganismos
do solo, assim, os SMS têm sido estudados quanto aos efeitos biorreguladores e
bioestimulantes em cultivos agrícolas (Owaid; Abed; Al-Saeedi, 2017) (Fig. 2).

18
Figura 02. Produção de cogumelos a partir da biomassa vegetal residual (BVRs)
baseado na economia circular, com obtenção de moléculas bioativas
(biofertilizantes/bioestimulantes) e retorno para o beneficiamento dos cultivos
agrícolas.
4.3 Brasil: Fungicultura de cogumelos comestíveis
Aproximadamente 2000 espécies de cogumelos comestíveis são catalogadas
em diferentes biomas do mundo, dentre estas, apenas 25 espécies são popularmente
comercializadas apresentado sistema de produção agronômico definido. No Brasil, as
principais espécies comestíveis cultivadas são Agaricus bisporus, Lentinula edodes e
diferentes espécies do gênero Pleurotus spp. Também há produção da espécie
Agaricus subrufescens (alguns trabalhos citam erroneamente como sinonímia de
blazei e brasiliensis) que é produzida voltadas a exploração para processos
biotecnológicos ligados as suas propriedades medicinais, atribuídas alguns bioativos
como a beta-glucanas, por exemplo (Taofiq et al., 2019; Win; Ohga, 2018; Zied, Diego
C. et al., 2018; Silva et al., 2022).
A espécie A. bisporus foi a primeira cultivada comercialmente no Brasil, produto
da imigração italiana nos anos 50 nas cidades de Cabreúva e Atibaia no estado de
São Paulo, paralelamente com o assentamento de chineses em Mogi das Cruzes,

19
trazendo o cultivo de L. edodes e Pleurotus ostreatus (Eira, 1992). O cultivo de
cogumelos comestíveis no Brasil mostrou maior impacto no mercado nos últimos 20
anos, em função de melhores informações sobre o valor nutricional, técnicas de cultivo
e instrumentação (Dias, 2010).
Um dos fatores do aumento no consumo de cogumelos no Brasil foi a migração
e incorporação da cultura asiática, especialmente no estado de São Paulo, na qual foi
instalado técnicas de cultivo e adaptações para o clima da região. Por muito tempo
São Paulo deteve a posição do único produtor comercial de cogumelos no Brasil, no
entanto, mesmo ainda sendo o maior produtor, outros estados já possuem uma
produção significativa. Os cultivos nestes estados ainda são restritos a pequenas
propriedades rurais que utilizam mão-de-obra familiar e tecnologia rudimentar
passada de geração em geração.
A cultura do consumo de cogumelos é mais significativa em regiões onde
prevalecem núcleos de imigrantes asiáticos, como no estado de São Paulo,
caracterizados com hábitos alimentares alternativos (Dias, 2010; Girardi et al., 2018).
Acredita-se que por herança das primeiras colônias portuguesas, o Brasil não possui
como tradição ao consumo de cogumelos. Posto que o registro de uso de cogumelos
dos povos indígenas na alimentação ou propósitos medicinais é limitado, havendo
apenas relatos que indicam que os grupos Indígenas Sanema e Yanomami, na
Amazônia, são consumidores de uma grande variedade de cogumelos (Anpc, 2018;
Vargas-Isla; Ishikawa; Py-Daniel, 2013).
A Associação Nacional de Produtores de Cogumelos (ANPC) estima que o
consumo per capita/ano tem aumentado de 80 para 160 gramas de cogumelos no
Brasil. Porém, mesmo com este aumento o consumo ainda é pequeno, quando
comparado ao consumo na Europa, com média de 2 quilos/pessoa/ano e na Ásia com
média de 8 quilos/pessoa/ano. A ANPC atribui esse aumento principalmente ao
incremento das tendências e práticas alimentares, tais como veganismo e
vegetarianismo, e ao investimento em consumo in natura do produto (ANPC, 2018).
A produção de cogumelos no Brasil ainda é limitada pelo alto custo aplicado
nas técnicas usadas para a obtenção destes alimentos (Dias, 2010; Eira, 1992).
Gomes (2018) reitera que apesar de muitos brasileiros já conhecerem os benefícios
nutracêuticos dos cogumelos, esse alimento ainda não faz parte da dieta regular da

20
maioria da população devido aos preços ainda elevados, os quais oscilam de acordo
com a região do Brasil, chegando a superar R$15 reais por 200 gramas (fresco), sendo
ainda uma barreira à expansão do consumo.
Entretanto, há expectativas do maior conhecimento sobre este alimento e
adesão de mais produtores (fungicultores), com advento da internet/mídias sócias têm
sido veiculados em diferentes fontes e mecanismos de informações as vantagens de
consumo e técnicas de cultivo. Há na internet divulgações de cursos de produção de
cogumelos, tanto por instituições reconhecidas como a Embrapa, como também pelos
próprios produtores, que buscam aumentar a rede de produção, como também por
associações como a ANPC. A procura por cultivos de cogumelos também têm ocorrido
em função de veiculações de matérias jornalísticas descrevendo-os como alimentos
funcionais – chamando-os “super-foods”(Gomes, 2018).
A cultura de produção de cogumelos no Brasil ainda não consegue atender a
demanda do mercado consumidor, mesmo em regiões com produção consolidada,
como o estado de São Paulo. Assim, o país depende das importações de produtos
em conserva de países como a China, e os valores são comumente inferiores ao
produto nacional (Gomes, 2018). Diante disto, muitos fungicultores nacionais optaram
por reforçar a produção de cogumelos frescos, que apresentam vantagens como o
alto valor agregado e renumeração ao produtor, além de manter a maior parte das
características nutricionais, gastronômicas e nutracêuticas frente os produtos
importados (Gomes, et al., 2016).
4.4 Sistema produtivo de cogumelos no Brasil
A maior concentração de produção de cogumelos está no estado de São Paulo,
porém existem empresas em outros Estados e Distrito Federal como Minas Gerais,
Rio de Janeiro, Bahia, Pernambuco e Rio Grande do Sul. No estado de São Paulo, a
quantidade mensal de cogumelos frescos pode variar de 80 quilogramas a 60
toneladas, mostrando que no setor há presença de pequenos, médios e grandes
fungicultores (Gomes, et al., 2016; Ishikawa et al., 2017).
Os autores Gomes, et al., (2016) que descrevem os resultados através de
boletins de informações tecnológicas da Agência Paulista de Tecnologia dos
Agronegócios (APTA), ainda reiteram que consultaram 505 fungicultores na pesquisa,
alcançando uma produção global para o estado de São Paulo de 1.062 toneladas de

21
cogumelos por mês. O faturamento financeiro do setor, segundo os autores, gerou
uma na ordem de R$ 21.240.017,00 mensais, com a geração de aproximadamente
cinco mil (5000) empregos diretos. Deste modo, fica evidenciado o que se observa em
outros países, como Holanda, Espanha, Itália, Irlanda e outros que a fungicultura
apresenta potencial significativo para auxiliar a alocação de emprego e renda para
populações em volta dos centros produtivos (Gomes, Daniel; et al., 2016).
Segundo a ANPC (2018) no Brasil as espécies de macromicetos produtores de
cogumelos comestíveis mais cultivadas e comercializadas são Agaricus bisporus,
Lentinula edodes, Pleurotus sp. (algumas espécies do gênero Pleurotus, tendo como
principal a espécie P. ostreatus – diferentes cepas – com fenótipos distintos quanto a
coloração, por exemplo) e Agaricus subrufencens (erroneamente chamado de blazei
ou brasiliensis em outras publicações). Assim podemos descrever os cogumelos mais
cultivados da seguinte maneira:
Agaricus bisporus: Esta espécie é popularmente conhecida como
"Champignon de Paris” aqui no Brasil e em outros países, como também “Button
Mushroom”, em países de língua inglesa. Há pelo menos dois tipos de estirpes que
são comercializadas deste fungo que variam de acordo com a corpo do cogumelo
“white/branco e brown/marron, também conhecido como popularmente como
“portobello”. As propriedades nutricionais e medicinais são amplamente conhecidas,
desde os altos níveis de proteínas, aminoácidos, carboidratos, vitaminas e polifenóis;
como também as atividades biológicas antioxidantes, antimicrobianas, anti-
inflamatórias, anticancerígenas e imunomoduladoras (Muszyńska et al., 2015).
O Champignon é o cogumelo mais cultivado no mundo, especialmente em
Europa, América do Norte, China e Austrália. No Brasil, a produção atinge pelo menos
66% dos cogumelos in natura (Gomes, 2018). A composição do substrato para o
cultivo do A. bisporus inclui uma ampla variedade de resíduos agroindustriais, como
palhas de arroz e trigo, bagaço de cana como fonte de carboidratos/carbono e esterco
de galinhas, cavalos, como fontes de proteína/nitrogênio. Uma correta relação entre
carbono/nitrogênio (C/N) na composição do substrato e no processo de
compostagem/estabilização (pasteurização) é determinante na produtividade dos
cogumelos desta espécie de macrofungos (Muszyńska et al., 2015).

22
Para o estabelecimento de uma produção de cogumelos inicialmente temos o
desenvolvimento de uma tecnologia no processo de produção de sementes de
cogumelo, esse processo de colonização requer técnicas para se evitar
contaminações. Atualmente, existem diferentes cepas para os processos de produção
de sementes no mercado internacional.
No Brasil não se tem muitas informações sobre a origem das cepas usadas,
isso se deve ao fato de a maior parte ser reproduzido e comercializado por pequenas
empresas com baixos recursos tecnológicos e nem sempre dispõe de um controle de
qualidade genética e sanitária do material comercializadas. Assim, temos uma
importante fase da cadeia de produção necessitando de atestado de qualidade, para
se evitar que ocorra degeneração ou contaminação por algum outro agente infeccioso.
Portanto, no futuro, será importante que laboratórios especializados façam este
trabalho para evitar cepas que tenham sido degeneradas ou contaminadas por vírus
ou outros elementos infecciosos.
Outro aspecto que diferencia o cultivo de A. bisporus no Brasil do que em
países europeus e na América do Norte é a dificuldade associada ao uso de turfa de
alta qualidade como camada de revestimento isso porque após ocorrer o processo de
colonização do substrato é essencial que seja coberto por um material com
características físico-químicas específicas, microbiologicamente apropriado, para se
obter condições favoráveis de altos índices de produção e produtividade (Gomes,
2018).
Agaricus subrufescens: Este cogumelo é também chamado “Cogumelo do
Brasil”, em função de ser genuinamente brasileiro. Também é conhecido como
“Cogumelo do Sol”, “Royal Sun Agaricus” (Estados Unidos) e Himematsutake (Japão).
As características para os cogumelos desta espécie perpassam desde gastronômicas,
como sabor, textura e fragrância amendoada, como propriedades medicinais. Estas
últimas estão sendo atribuídas a alguns metabolitos deste fungo na prevenção de
câncer, diabetes, arterioscleroses e hepatites crônica, tendo também citações como
potencial componente para o tratamento de doenças do sistema imune e infeções
(Firenzuoli; Gori; Lombardo, 2008; Lima; Gris; Karnikowski, 2016).
O mercado brasileiro chegou a exportar no início dos anos 2000 (Rosa, 2006)
até 80% da produção nacional ao Japão, que por tradição consome quantidades

23
importantes de cogumelos. Atualmente a produção está voltada a atender o mercado
nacional, principalmente para obtenção de extratos com aplicações a produtos
nutracêuticas (Silva et al., 2022).
De acordo com Zied et al. (2018) estudos vêm sendo realizados para o
desenvolvimento de técnicas de melhor produção de A. subrufescens, mas de forma
geral o sistema de produção segue o mesmo dos demais cogumelos, sendo a etapa
de produção do substrato o mais importante para o sucesso no processo de produção,
uma prática aplicada aos processos de produção é a suplementação do substrato com
os nutrientes necessários para que haja uma redução de ciclo. Essa técnica de
suplementação já é aplicada em outros cogumelos e o mercado já disponibiliza as
formulações de acordo com o tipo de produção. Embora o suplemento comercial ainda
não esteja disponível em escala industrial para produção de A. subrufescens.
Pleurotus spp.: Dentro do gênero Pleurotus são cultivadas várias espécies de
cogumelos, que também podem receber nomes populares diferentes a depender do
paíse de produção/língua, desde “hiratake” (Japão) e “oyster mushroom” (Estados
Unidos e Europa). Além destes nomes populares, no Brasil também podem receber
outros nomes, em função de semelhança com outras espécies conhecidas no Japão,
por exemplo. O “shimeji branco” ou “shimeji preto” referem-se a estirpes de P.
ostreatus que possuem fenótipos quanto a coloração diferente em função das práticas
de cultivo (principalmente relacionados a coloração da estipe e maturidade – jovem
na forma de “boque de flores” ou mais velho na forma “ostra”). Além do P. ostreatus,
outras espécies também são cultivadas de forma comercial, tais como P.
ostreatoroseus (sinonímia djamor), P. eryngii, P. pulmonarius e P. citrinopileatus.
Os cultivos das espécies de Pleurotus podem ser feitos com distintos resíduos
da agroindústria como substrato (palha de cereais, serragem, bagaço de cana,
resíduos de algodão), com menor complexidade que o A. bisporus mas com as
condições de umidade e temperatura adequada, usando sacos ou diversos recipientes
de plástico. No Brasil, a produção atinge pelo menos 16% dos cogumelos in natura
(ANPC, 2018).
De acordo com Vieira et al. (2019) o cultivo de P. ostreatus segue em três fases
distintas que são a primeira preparação do substrato, nesse primeiro momento a
compostagem é realizada, esse é um processo de extrema importância para o

24
sucesso posterior visto que representa o principal custo associado à produção de
cogumelos. A segunda fase diz respeito ao processo de colonização do substrato por
micélio, as sementes geralmente advêm de outros produtores especializados em
produção de inóculos (sementes) de fungos produtores de cogumelos. Terceira fase
é a frutificação e colheita dos cogumelos e posterior comercialização (Vieira, et al.,
2019).
Existem outros trabalhos como o de Magalhães et al. (2018) que testou o
processo de cultivo de Pleurotus ostreatus em meio axênico com base em resíduos
de cana de açúcar (bagaço), suplementados com bioprodutos de cereais e obtiveram
como conclusão a definição de um método eficiente para a produção de alta taxa
biológica e valor de proteína pela reciclagem de resíduos agrícolas, sendo capaz de
se obter benefícios socioeconômicos, ambientais e uma cadeia produtiva rentável
para pequenos e médios produtores (Magalhães; Moreira; Zied, 2018).
Lentinula edodes: o cogumelo desta espécie é comumente conhecido como
“shiitake” ou “cogumelo do carvalho”. A produção de shiitake ocupa o segundo lugar
de produção globalmente, principalmente na China, Japão, Coreia e Taiwan (Miles;
Chang, 2004). Há qualidades gastronômicas acrescentadas com as propriedades
medicinais ao shiitake, como também, possibilidade de atividades antimicrobiana,
anticancerígena e antioxidantes de alguns bioativos encontrados nesta espécie
(Chowdhury; Kubra; Ahmed, 2015). No Brasil, a produção atinge pelo menos 12% dos
cogumelos in natura (ANPC, 2018).
Tem aumentado sua produção no Brasil devido ao retorno financeiro e
possibilidade de cultivo em pequenas áreas e baixo investimento. Possui
características de degradação e utilização de resíduos agrícolas, colheita mais rápida
e grande eficiência biológica (Souza, 2016). Esse fungo utiliza lignina, celulose e
hemicelulose como fonte de carbono e nutrientes, permitindo assim uma grande
diversidade de resíduos oriundos do sistema agrícola que podem ser utilizados como
exemplo o bagaço de cana (Eira, 1992; Souza, 2016).
Trabalho realizado no Estado do Paraná – Brasil, com L. edodes em dois
diferentes métodos de cultivo, sendo, toras de madeira (carvalho) e substrato axênico,
concluíram que a composição físico-química do meio de cultura influencia o a
composição nutricional do cogumelo (Bach et al., 2018).

25
4.5 Biomassas vegetais residuais (BVRs) na produção de cogumelos
O sistema de produção de cogumelos de modo geral é feito por meio do preparo
de substratos lignocelulósicos ou toras de madeira. Os substratos/toras são
preparados, acondicionados ou esterilizados, para posterior inoculação com
fragmentos de micélio da espécie fúngica (produtora do cogumelo). O inoculo (spawn,
inglês) previamente preparado por empresas especializadas ou de produção própria
contêm os propágulos prontos para colonização ou miceliação dos substratos. Após
a colonização total dos substratos/toras, estes são acondicionados em ambientes de
frutificação, controlados ou não, contendo umidade acima de 80%, como também
temperatura, oxigenação e luminosidade adequadas a cada tipo de espécie ou estirpe.
Durante a fase de furtificação, os cogumelos são colhidos rotineiramente,
armazenados e/ou levados ao mercado (Eira, 1992).
O uso de resíduos vegetais agroindustriais como substratos para o cultivo de
diferentes espécies de cogumelos é uma estratégia amplamente aplicada em vários
locais do mundo. O reaproveitamento de tais biomassas permite reduzir os custos de
produção, principalmente quando a biomassa vegetal está próxima ao fungicultor,
reduzindo o custo de frete. Estes resíduos vegetais podem ser combinados para
atender as necessidades nutricionais (C/N, por exemplo) de acordo com a espécie ou
estirpe fúngica. Na Tabela 2 são detalhados estudos da aplicação de cultivo de
cogumelos usando resíduos agroindustriais.
Tabela 2. Cultivo de cogumelos comestíveis em diferentes tipos e combinações com
resíduos vegetais agroindustriais.
Fungos/Cogumelos Substratos Referências
Pleurotus ostreatus e
Agaricus bisporus
Suplemento de farinha de amêndoa
desengordurada
(Pardo-Giménez et
al., 2018)
Pleurotus spp. Algodão, resíduos de espigas de milho. Poppe (2004)
Pleurotus florida (Mont.)
Singer Palha de soja, arroz e trigo
Abrar Ahmed et
al., (2009)

26
Pleurotus ostreatus
Casca de café enriquecida com selênio
(Se)
Da Silva et al.,
(2012)
Folhas de Dendê (Elaeis spp.) Ali et al., (2018)
Palha de arroz Das et al., (2015)
Serragem e farelo de arroz Owaid (2014)
Pleurotus citrinopileatus Resíduos de papel, resíduos industriais
de cartão
Kulshreshtha et
al., (2013)
Volvariella volvacea Cacho vazio de Dendê (Elaeis spp.) Triyono et al.,
(2019)
Lentinula edodes Palha de sorgo Chittaragi; Kumar;
Muniraju (2018)
Agaricus bisporus Bio-fortificação de substrato com Se,
Cu e Zn
(Rzymski et al.,
2017)
Agaricus subrufescens
(blazei)
Gramíneas (Brachiara sp., Cynodon
dactylon); palhas (trigo, arroz, milho,
cevada), bagaço de cana, casca de
girassol, esterco bovino e aves,
suplementos nitrogenados (semente de
algodão, farelo de trigo e soja); fibras de
soja e mandioca; suplementos como
ureia, sulfato de amônio, gesso e
calcário.
Mendonça et al.
2005; Minhoni et
al., 2005; Zied e
Minhoni 2009;
Matute et al. 2010;
Zaghi Junior et al.,
2010 citados por
(Largeteau et al.,
2011)
Ganoderma lucidum Poda vegetal urbana (Alquati et al.,
2016)
O ciclo de produção de cogumelos é renovado constantemente com novos
substratos colonizados com micélio fungico. Assim, a cada novo ciclo de produção se
faz necessário a retirada da biomassa residual pós-colheita dos cogumelos, o SMS),

27
que por sua vez precisará de uma destinação correta para descarte ou para geração
de outros produtos ou aplicações em processos agroindustriais, por exemplo.
4.6 SMS - Biomassa residual pós-colheita dos cogumelos
Anualmente, a indústria de cogumelos em países asiáticos, europeus ou norte-
americanos, geram milhões de toneladas SMS, que é uma mistura que pode conter
desde solos (produção de Agaricus), biomassas vegetais parcialmente degradadas e
massa micelial, com respectivos metabolitos microbianos. Embora contenha
ingredientes valiosos como carboidratos, lignina e diversas enzimas, o SMS ainda não
tem sido plenamente utilizado em outros processos biotecnológicos, seu destino vem
sendo o descarte, gerando custos financeiros e impactos ambientais.
A fim de valorizar o SMS, é essencial fracionar a mistura complexa em seus
componentes valiosos, o que é um desafio para as modelos de biorrefinarias. Grupos
de pesquisa europeus em consorcio com empresas estão trabalhando em modelos
de biorrefinarias utilizando o SMS para obter bioprodutos de valor agregado, tais
como: enzimas, nanocompositos, (Herawati; Istifadah, 2019), biofertilizantes,
bioplasticos, smartmushroom, biorescue (Funguschain, 2019 -
http://www.smartmushroom.eu; https://biorescue.eu/) entre outros (Siqueira et al.,
2017).
Em países com grande tradição na produção de cogumelos como a China, esse
resíduo (SMS) tem gerado grande preocupação, por ser um passivo ambiental (Zhu,
Jiying et al., 2015), pois para cada kg de cogumelo fresco colhido gera-se
aproximadamente 5 kg de SMS (Finney et al., 2009; MA et al., 2014). Entre as
estratégias atuais para disposição de SMS estão a queima, espalhar em terra,
compostagem com esterco de animais ou aterro (Chiu et al., 1998; Phan; Sabaratnam,
2012). No Brasil a maioria dos fungicultores descartam em solos agrícolas, fazem
compostagem ou cedem a pequenos horticultores para uso junto ao solo de cultivo
folhosos e outros.
Atualmente há vários estudos, em diferentes países, buscando alternativas de
uso do SMS para obtenção de produtos de valor agregado. Tem se estudado
estratégias de modelagem de biorrefinarias (Del Campo et al., 2018) com obtenção
de diversos bioprodutos, como também ações em processos de biorremediação,
nutriçãoanimal, matéria-prima energética (biogás ou bioetanol) e biofertilizantes

28
((Dale; Balan; Chundawat, 2007; Grimm; Wösten, 2018b; Mohd Hanafi et al., 2018;
Jordan; Mullen, 2007; Phan; Sabaratnam, 2012).
As características químicas e biológicas dos SMSs variam de acordo com a
espécie de fungo cultivada, os substratos e sistema de preparo, como também do
modelo de cultivo. Segundo Jordan et al (2008), a composição em média do SMS
recolhido de empresas produtoras de cogumelos contém celulose (38%),
hemicelulose (19%), lignina (15%) e matéria orgânica 645g/kg. Durante o crescimento
micelial, os fungos são capazes de produzir diversas enzimas lignocelulolíticas para a
quebra destas fibras estruturais vegetais, as quais podem ser extraídas e purificadas
para fins comerciais, por exemplo (Del Campo et al., 2018). O SMS também pode
servir de substrato para cultivo de outros microrganismos capazes de gerar
metabolitos com potencial uso como biofertilizantes, tais como Trichoderma
harzianum (Singh et al., 2018).
4.6.1 SMS - formulação de biofertilizantes e bioestimulantes
A utilização de reguladores vegetais pode mostrar-se altamente compensadora
em termos econômicos. Diversos estudos têm apontado o uso de fertilizantes
orgânicos oriundos dos resíduos de cadeias produtivas como uma prática de alto
potencial, sendo uma fonte sustentável e econômica para promover efeitos similares
aos reguladores comerciais.
Entre esses resíduos, já foi demostrado que os resíduos de SMS atuam nos
processos que incentivam a promoção de crescimento, proteção, aumento na
produção e produtividade dos vegetais, como descritas na Tabela 3 (Grimm; Wösten,
2018a; Okolie et al., 2019). O SMS ainda mantém altos níveis de matéria orgânica,
nitrogênio (N), fósforo (P), potássio (K) e outros nutrientes normalmente necessários
para o crescimento das culturas vegetais (Jordan; Mullen; Murphy, 2008; Roy et al.,
2015).
A transformação de resíduos orgânicos ricos em nutrientes, em valiosos
fertilizantes agrícolas e/ou substratos para crescimento e desenvolvimento vegetal,
pode contribuir para a conservação de materiais, reduzindo a geração de resíduos
(Paula et al., 2017). A aplicabilidade do alto conteúdo de matéria orgânica do SMS foi
registrada nos estudos de Siddhant e Singh (2009), no qual foi usado SMS do cultivo
de três espécies de cogumelos como fertilizante para o cultivo da planta Spinacea

29
oleracea (espinafre). Sendi et al., (2013), o SMS foi usado como substituto para musgo
de turfa para produzir Kai-lan/Kale (Brassica oleracea var. alboglabra) na Malásia. Nos
estudos de Ashrafi et al., (2014) o substrato utilizado para o cultivo de Pleurotus
ostreatus e P. florida, e posteriormente o SMS foi usado como fertilizante para
produção de mudas de tomate (Lycopersicon esculentum).
Tabela 3. SMS como um dos insumos para formulações e/ou aplicações destinados
a nutrição de plantas.
SMS Tipo de produto Tipos de usos para
nutrição de plantas Referências
SMS de
Agaricus bisporus
Bioestimulante Gestão de resíduos de
SMS
Medina et al.,
(2012)
SMS de
Agaricus bisporus
Fonte de nitrogênio Fertilizante orgânico e
condicionador de solo. Hackett, (2015)
SMS de
Flammulina
velutipes
Substrato de
crescimento de
mudas
Misturas hortícolas sem
solo.
Zhang; Duan; Li,
(2012)
SMS de Agaricus
bisporus/Pleurotus
ostreatus
Promoção do
crescimento
Fertilizante para plantas
pela quantidade de
carbono e nitrogênio.
Roy et al., (2015)
SMS
Biossorção e
produção de
componentes de
fertilizantes
Plantas biofortificadas com
maior teor de
micronutrientes (de 8 a
31%).
Tuhy et al.,
(2015)
SMS de Pleurotus
spp.
Fertilizante natural
e adubo
Disponibilização de N,
relação C/N
Owaid et al.,
(2017)

30
SMS de Agaricus
bisporus
Biofortificação:
SMS com
Trichoderma
harzianum e
minhocas
Melhoria das propriedades
antioxidantes e nutricionais
do tomate
Singh et al.,
(2018)
SMS de
Auricularia
auricula
SMS e resíduos de
biogás substituto
parcial ou total da
turfa
Substrato de crescimento
para mudas de tomate e
pimenta
Meng et al.,
(2017)
SMS de Pleurotus
floridanus
Biofertilizante
líquido com
resíduos orgânicos,
SMS e inoculantes
microbianos
Adubo líquido em milho e
quiabo
Okolie et al.,
(2018)
Fermentado de
Pleurotus,
Lentinus,
Ganoderma
e linhagens de
cogumelos
híbridos
Biofertilizante Crescimento de tomate e
trigo
Narh Mensah et
al., (2018)
SMS
Substrato com lodo
de esgoto, SMS e
bactéria
Acidithiobacillus
ferrooxidans
Liberação lenta de fósforo
em germinação de girassol
Saeid; Patel,
(2019)
SMS de Agaricus
subrufescens
SMS como
substrato para
Germinação e produção de
mudas de tomate
Lopes et al.,
(2015)

31
produção de
mudas
SMS de Pleurotus
eous
Promotor de
crescimento
Crescimento e produção de
pimenta e tomate em casa
de vegetação
Wiafe-Kwagyan;
Odamtten,
(2018)
SMS de Agaricus
bisporus e
Pleurotus spp.
Fertilizante
Rendimento e
nutrição de alface
Paredes et al.,
(2016)
SMS Fertilizante para
substituir nitrogênio
Produção de alface a alho
porró
Gobbi et al.,
(2016)
SMS
Biochar produzido
partir de SMS e
usado como
fertilizante
Melhoria qualidade físico-
química do solo
Mao et al.,
(2018)
SMS Pleurotus
spp.
Bioestimulante a
base de SMS
adicionado de
Brevibacillus
parabrevis
Germinação de
crescimento de tomate e
pepino
Oancea et al.,
(2017)
4.6.2 SMS: biocontrole de fitopatógenos
Na natureza, as plantas desenvolveram vários mecanismos de defesa contra
patógenos, cuja função está em impedir, diminuir ou neutralizar os efeitos prejudiciais
do agente infecioso. Estes mecanismos de resistência podem ser constitutivos, os
quais se caracterizam por atuar de forma passiva, com a presença de estruturas
defensivas (ex. tricomas, cutículas grossas e disposição de ceras) ou pela presença
e acumulação de substâncias toxicas nas células vegetais (ex. terpenoides, saponinas
e cianatos); ou mecanismos de resistência induzida, os quais atuam de forma ativa
(ativação de processos metabólicos por expressão gênica) em resposta ao ataque de
um fitopatógeno (Doughari, 2015).

32
No entanto, nas práticas agrícolas, os mecanismos naturais das plantas não
têm ação suficiente para impedir as perdas econômicas geradas como consequência
de infeções por fitopatógenos, os quais podem diminuir a produtividade, a qualidade,
ou até ocasionar a morte total dos cultivos. Por consequência, é comum observar o
uso de diversos tipos de pesticidas (agroquímicos, cobre, antibióticos) como estratégia
de controle, mas os impactos ambientais e à saúde humana do uso destes produtos
gera controvérsias.
Em vista disso, diversos autores têm procurado técnicas alternativas para
atenuar os impactos do uso de pesticidas. Atualmente, o maior conhecimento dos
mecanismos de indução a resistência aos fitopatógenos tem possibilitado, por meio
de programas de fitomelhoramento, o desenvolvimento, a detecção e incorporação de
biomoléculas para seleção de plantas menos susceptíveis as infeções (Silva, et al.,
2018). Tais tecnologias são capazes de produzir organismos geneticamente
modificadas (OGM) com resistência perdurável. Em 2019 já haviam 29 plantas OGM
aprovadas para serem comercializadas globalmente com resistência a infeção por
vírus e fungos (International Service for the Acquisition of Agri-biotech Applications,
2019).
Existem inúmeras pesquisas que abordam o controle biológico de fitopatógenos
em cultivos de plantas, com a finalidade da exploração de estratégias com
características sustentáveis. Extratos de cogumelos e/ou SMS tem sido investigados
como insumos potenciais para composição de formulações de fertilizantes com ação
inibitória de fitopatógenos (Tabela 4).
O SMS pode ser considerado como uma fonte facilmente disponível de
compostos ativos para proteger as plantas contra infecções fúngicas e bacterianas.
Prática que promove a redução do descarte de resíduos na cadeia produtiva de
cogumelos e levanta um método ambientalmente amigável para reduzir a incidência
de patógenos em cultivos agrícolas (Kang et al., 2017).
Tabela 4. Uso de espécies de fungos produtores de cogumelos e SMS no controle de
doenças e pragas de plantas agrícolas.

33
Microrganismo Processos utilizados Resultados Referência
SMS de Hericium
erinaceum, Hypsizigus
marmoreus, P. eryngii
Supressão de doenças de plantas por
extratos. Promissor como antioxidante e antibacteriano.
Lee et al.,
(2018)
SMS de Pleurotus eryngii,
Hericium erinaceus e
Lentinula edodes
Supressão da doença (Phythopthora
Blight e Fusarium oxysporum).
Promoveu o crescimento das plantas e
suprimiu a doença.
Kwak et al.,
(2015)
Fermentado de
Pycnoporus sanguineus (L.
ex Fr) Murr.
Ativa enzimas de defesa do feijoeiro
(Pseucocercospora griseola (sacc.)
Crous).
Obteve ativação das defesas da planta. Viecelli et al.,
(2009)
Fermentado Lentinula
edodes e A. blazei Indução a produção de fitoalexinas.
Eficientes no controle do oídio (Erysiphe
diffusa) no soja.
De Arruda et al,
(2012)
Fermentado de A. blazei Extrato. Inibiu a germinação de esporos de Puccinia
recondita f. Sp. Tritici.
Fiori-Tutida et
al. (2007)
Fermentado de L. edodes Filtrado do crescimento micelial.
Inibiu o crescimento micelial de Exserohilum
turcicum, agente causal da helmintosporiose
do sorgo.
Piccinin; Di
Piero;
Pascholati
(2010)

34
Cogumelos de A. blazei e
L. edodes
Extratos brutos, obtidos a partir do pó
seco do basidiocarpo.
Inibição germinação de esporos de Bipolaris
sorokiniana e Puccinia recôndita (mancha
marron e ferrugem da folha da cultura do
trigo).
Fiori-Tutida et
al., (2007)
SMS
Resíduos de biogás e SMS como
alternativa à turfa (100% de
substituição).
Maior crescimento morfológico em tomate e
menores concentrações de Fusarium.
Chen; Fan;
Meng, (2017)
SMS de Hericium erinaceus Extrato aquoso. Potencial supressão de murcha bacteriana de
tomate causada por Ralstonia solanacearum.
Kwak et al.,
(2015b)
SMS Esterco (adubo orgânico). Melhor desenvolvimento das culturas. Sagar et al.
(2009)
SMS Lyophyllum decastes e
Pleurotus eryngii Extrato aquoso autoclavado.
Efeito protetor contra 4 doenças fúngicas e
bacterianas.
Parada et al.
(2012)

35
4.7 Fermentados fúngicos ou SMS e novos produtos/processos para
agricultura Brasileira
O Brasil é considerado essencialmente agrícola, apresentando um área
cultivada de aproximadamente 75 milhões de hectares (IBGE, 2012). De acordo com
o Ministério de Agricultura (2015), após os Estados Unidos, é o segundo maior
produtor de grãos do mundo, principalmente soja. No entanto, outros produtos têm
grande destaque como o algodão, milho, cana de açúcar, plantas ornamentais, frutas
e olerícolas, assim em 2010 um de quatro produtos do agronegócio em circulação no
mundo teve sua origem no Brasil.
A alta produtividade está associada as inovações tecnológicas e eficiência dos
produtores. O manejo de doenças tem ganho foco nas empresas agrícolas, o
consumo de produtos fitossanitários (herbicidas, inseticidas, acaricidas, nematicidas,
etc) no Brasil tornou-se de grande importância, com crescimento de 190% (entre
2002-2012) e superando o crescimento mundial (93%) nesse período (Carneiro et al.,
2015). Entre os principais produtos agrotóxicos são os fungicidas, cujo mercado
movimentou US$2,91 bilhões no Brasil em 2014 (13,2% do produto comercial). Os
fungicidas foram empregados especialmente na aplicação foliar e tratamento de
sementes, e em função do favorecimento climático para o desenvolvimento de
doenças e a sua resposta cada vez mais agressiva, os fungicidas foram uns dos
produtos mais comercializados no país em 2016 (IEA, 2014).
A base da produção brasileira é a exportação, por tanto é necessária uma
produção agrícola de alta qualidade. Porém, o aumento da produção e padrões
elevados de qualidade implicam a utilização cada vez maior de agroquímicos, já que
as produções em larga escala exercem pressão de seleção sobre os organismos
desencadeando o surgimento de microrganismos patógenos resistente. As
consequências das doenças podem influenciar direta ou indiretamente na
rentabilidade do agronegócio, limitando a geografia de variedades de plantas,
reduzindo a qualidade e quantidade de produção.
O agronegócio brasileiro está em expansão, sendo um grande importador de
defensivos agrícolas. Em virtude da alta demanda de insumos e alta disponibilidade
de resíduos e subprodutos agrícolas, há potencial para se tornar um grande produtor
de biofertilizantes, bioestimulantes, biodefensivos de base biotecnológica e
microbiana. As BVRs podem ser utilizada como substrato ou “veículo” para

36
microrganismos ou compostos químicos microbianos (proteínas, peptídeos, ácidos
orgânicos, terpenóides e outros), base da formulação dos insumos biológicos para
agricultura voltada a bioeconomia circular. Neste sentido, os fermentados fúngicos
(antes da frutificação) ou SMS (após colheira dos cogumelos) produzidos pela cadeia
fungícola pode ser estudado quanto ao uso na fabricação de bioprodutos e/ou
bioinsumos com potencial utilização no agronegócio brasileiro e mundial. O modelo
de investigação e desenvolvimento de tais produtos já está amplamente divulgado nas
publicações cientificas e projetos de pesquisas de grandes países europeus
(Biorescue e Funguschain, EU-Horizon 2020 - https://funguschain.eu/news/read-
interview-mr-darragh-gaffney-monaghan-mushrooms-biorescue-project/attachment /
monaghanday/).
4.8 Cultura do tomate
O tomate (L. esculentum Mill) da família das solanáceas é uma hortaliça do tipo
fruto, com origem na América do Sul (parte ocidental), tendo países como Peru, Bolivia
e Equador como berço de origem. O cultivo comercial dessa hortaliça foi realizado por
tribos indígenas do México, sendo considerada planta venenosa por muitos anos na
Espanha e nessa época foi usada somente como ornamental. A utilização e consumo
do tomate ficou por conta dos italianos que descobriram o seu poder alimentar (Fontes
e Silva, 2002).
Existe atualmente no mrcado consumidor diversos tipos de tomate, com destaque
para o tipo cereja (L. esculentum var. Cerasiforme). No Brasil o tomate cereja foi
introduzido na década de 1990, tendo sabor mais adocicado que outras variedades
de tomates, o mesmo pode ser encontrado de formato arredondado ou tipo uva com
tamanhos pequenos. Outro fator de destaque é a coloração vermelha intensa ou
amarela para alguns híbridos, alta firmeza, boa consistência, resistência à doença e
valor nutricional (Souza, 2009; Junqueira, 2011).
O sistema de produção de tomate passa por um processo anterior que é a
produção de mudas, geralmente são transplantadas para o campo com 30 dias ou 4
folhas bem desenvolvidas. Um sistema essencial para o bom desenvolvimento das
mudas é o protegido, isso porque fatores adversos podem ser melhor controlados
como o ataque de insetos, doenças, controle de temperatura e umidade. Outros
fatores de igual relevância são a qualidade e procedência das sementes utilizadas,

37
água livre de contaminações e o substrato utilizado, este deve fornecer nutrientes e
aeração adequados para o bom desenvolvimento radicular e parte aérea (Collela, et.
al., 2019; Clemente et al., 2012; Bresolin et al., 2010; Alvarenga, 2013). As mudas de
qualidade garantem cerca de 60% do desempenho do ciclo produtivo da cultura,
inferindo assim que a fase de produção de mudas é indispensável para o êxito do
cultivo de tomate (Alvarenga, 2013).
Os sistemas protegidos têm sido utilizados em larga escala no cultivo de tomate,
essa preferência tem se dado por maior controle de irrigação, economizando água,
otimizando a aplicação de fertilizantes. No Brasil, a utilização de ambiente protegido,
principalmente para a produção de plantas olerícolas e ornamentais, vem se
destacando devido à maior proteção quanto a fenômenos climáticos, menor variação
de temperatura, diminuição de perdas de nutrientes e à redução dos custos com
fertilizantes e defensivos (Oliveira, 1995; Carrijo et al., 2004).
Segundo estudos de Beckmann e Cavalcante et al. (2007), a técnica do cultivo
protegido, diminui perdas em culturas sensíveis a variações como tomate, pimentão,
alface e ornamentais, minimiza os efeitos de intempéries climáticos e da variabilidade
ambiental, aperfeiçoa o desenvolvimento da cultura implantada, favorece o
crescimento das plantas e encurta o seu ciclo de produção, permite oferta o ano todo.
4.9 Fitopatógeno do tomate – Sclerotinia sclerotiorum
O tomateiro é uma cultura sensível a qualquer variável climática, nutricional e
ataque de fitopatógenos, isso tem maior criticidade no período chuvoso, limitando sua
produção e aumentando o custo da lavoura para o produtor, a adoção de métodos
preventivos é essencial, já que existe poucas medidas para minimizar as perdas pós
injuria na lavoura. Para evitar grandes perdas e aumento de custo de implantação e
condução da lavoura, faz-se necessário um manejo adequado de medidas de controle
envolvendo o uso de variedades resistentes e adoção de medida de exclusão,
erradicação e proteção da cultura do tomateiro (Kurozawa, 2005; Clemente, 2012;
Alvarenga, 2013).
Uma doença que tem histórico de instalação na cultura do temate é o mofo branco,
causado pelo ataque do fungo S. sclerotiorum. Esse fungo tem duas formas de
contaminação, por ascósporos que são levados pelo vento e por escleródios. Os
escleródios têm a capacidade de permanecer viáveis por longos períodos de tempo

38
porque são resistentes a condições adversas (controle químico e físico), bem como à
degradação biológica (Merriman 1976 Wu et al. 2008). Estudos como de Cosic et al.
(2012) e Adams e Ayers (1979) mostrou que a sobrevivência dos escleródios varia de
3 até 5 anos em condições favoráveis, solos não cultivados e acrescentam que a
viabilidade dos mesmos após tanto tempo pode chegar a 100%.
Os sintomas causados por S. sclerotiorum diferem entre as espécies hospedeiras
(Lumsden 1979; Morrall e Dueck 1982; Kora et al. 2003; McLaren et al. 2004). No
entanto, os sintomas mais comuns, são manchas irregulares embebidas e um micélio
branco semelhante ao algodão característico, presente nas folhas, caules, frutos e
pecíolos. Os sintomas secundários são lesões encharcadas, murcha, branqueamento
e destruição de plantas. O final do estágio é a formação do escleródio e com os
processos de decomposição das plantas os mesmos são transferidos para o solo
(Nelson et al. 1989; Kora et al. 2003).
A atividade dos agentes de controle biológico no solo é afetada por muitos fatores
ambientais abióticos e bióticos, temperatura, potencial hídrico, pH, pesticidas, matéria
orgânica, microrganismos do solo, espécies de plantas e assim por diante, tornando
esses agentes geralmente menos eficazes do que os pesticidas sintéticos. Porém,
devido ao efeito menos prejudicial ao meio ambiente e à falta de métodos químicos
eficazes, buscam-se métodos biológicos mais seguros para o controle e supressão de
S. sclerotiorum.
Referências
Abrar, A., Syed et al. (2009). Biological Efficiency And Nutritional Contents Of
Pleurotus florida (Mont.) Singer Cultivated On Different Agro-wastes. Nature
and Science, v. 1, n. 2, p. 127–129.
Adams, P. B., Ayers, W. A. (1979). Ecology of Sclerotinia species. Phytopathology,
69(8), 896-899.
Adedokun, O. M., Okomadu, C. A. (2017). Wild and domesticated mushroom
consumption in Nigeria. African Crop Science Journal, v. 25, n. 1, p. 123.
Ali, N et al. (2018). Cultivation of pleurotus ostreatus on oil palm fronds mixed with
rubber tree sawdust. Chemical Engineering Transactions, v. 63, p. 547–542.

39
Alquati, G. et al. (2016). Residues from urban vegetable pruning in the production of
the medicinal mushroom Ganoderma lucidum. African Journal of Agricultural
Research, v. 11, n. 38, p. 3664–3670.
Anchang, K. Y. (2014) Current Developments in Mushroom Biotechnology in Sub-
Saharan Africa. World Society for Mushroom Biology and Mushroom Products.
[S.l: s.n.].
ANPC. Associação Nacional de Produtores de Cogumelos. (2019). Disponível em:
<https://www.anpccogumelos.org/cogumelos>. Acesso em 15/06/2019.
Ashrafi, R. et al. (2014). Recycling of Spent Mushroom Substrate for the Production of
Oyster Mushroom. Research in Biotechnology, v. 5, n. 2, p. 13–21.
Bach, F. et al. (2017). Edible mushrooms: a potential source of essential amino acids,
glucans and minerals. International Journal of Food Science and Technology,
v. 52, n. 11, p. 2382–2392.
Bach, F. et al. (2018). Influence of cultivation methods on the chemical and nutritional
characteristics of Lentinula edodes. Emirates Journal of Food and Agriculture,
v. 30, n. 12, p. 1006–1013.
Bach, F. et al. (2019). Bio compounds of edible mushrooms: in vitro antioxidant and
antimicrobial activities. LWT, v. 107, p. 214–220.
Boin, E., Nunes, J. (2018). Mushroom Consumption Behavior and Influencing Factors
in a Sample of the Portuguese Population. Journal of International Food and
Agribusiness Marketing, v. 30, n. 1, p. 35–48. Disponível em:
<https://doi.org/10.1080/08974438.2017.1382420>. Acesso em: 22/06/2019.
Carneiro, F. F et al. (2015). Dossiê Abrasco. [S.l: s.n.]. Disponível em:
<http://abrasco.org.br/dossieagrotoxicos/>. Acesso em: 22/06/2019.
Chang, S. T., Wasser, S. P. (2018). Current and Future Research Trends in
Agricultural and Biomedical Applications of Medicinal Mushrooms and
Mushroom Products (Review). International Journal of Medicinal Mushrooms,
v. 20, n. 12, p. 1121–1133.

40
Chen, Y., Fan, H., Meng, F. (2017). Pleurotus ostreatus decreases cornstalk lignin
content, potentially improving its suitability for animal feed. Journal of the
Science of Food and Agriculture, v. 97, n. 5, p. 1592–1598.
Chittaragi, A., Kumar, A., Muniraju, K. M. (2018). Evaluation of Various Lignocellulosic
Products for the Cultivation of Shiitake Mushroom [Lentinula edodes (Berk.)
Pegler]. International Journal of Current Microbiology and Applied Sciences, v.
7, n. 4, p. 2319–7706.
Chiu, S. W. et al. (1998). Spent oyster mushroom substrate performs better than many
mushroom mycelia in removing the biocide pentachlorophenol. Mycological
Research, v. 102, n. 12.
Chowdhury, H. M. H.; Kubra, K., Ahmed, R. R. (2015). Screening of antimicrobial,
antioxidant properties and bioactive compounds of some edible mushrooms
cultivated in Bangladesh. Annals of Clinical Microbiology and Antimicrobials.
Conceicao, A. A et al. (2018). Bioconversion and Biotransformation Efficiencies of Wild
Macrofungi. In: Singh, Bhim; Lallawmsanga; Passari, Ajit (Org.). Biology of
Macrofungi. 1. ed. [S.l.]: Springer. p. 411. Disponível em:
<http://link.springer.com/10.1007/978-3-030-02622-6>. Acesso
em:06/09/2019.
Ćosić, J., Jurković, D., Vrandečić, K., Kaučić, D. (2012). Survival of buried Sclerotinia
sclerotiorum sclerotia in undisturbed soil. Helia, 35(56), 73-78.
Da Silva, M. C. S. et al. (2012). Enrichment of Pleurotus ostreatus mushrooms with
selenium in coffee husks. Food Chemistry.
Dale, B., Balan, V., Chundawat, S. (2007). Process For Conversion Of Mushroom
Lignocellulosc Waste To Useful Byproducts.
Das, N. et al. (2015). Comparative study of five pleurotus species cultivated in warm
temperature on non-sterilized rice straw. Emirates Journal of Food and
Agriculture.
De Arruda, R. S. et al. (2012). Efeito de extratos de cogumelos na indução de
fitoalexinas e no controle de oídio da soja em casa de vegetação. Bioscience
Journal.

41
Dejene, T., Oria-De-Rueda, J. A., Martín-Pinto, P. (2017). Wild mushrooms in Ethiopia:
A review and synthesis for future perspective. Forest Systems, v. 26, n. 1, p. 1–
7.
Dias, E. S. (2010). Mushroom cultivation in Brazil: Challenges and potential for growth
| Cultivo de cogumelos no Brasil: Desafios e potencialidades. Ciencia e
Agrotecnologia, v. 34, n. 4, p. 795–803.
Dias, E. S., De Brito, MR. (2017). Mushrooms: Biology and Life Cycle. In: Zied, Diego;
Pardo-Gimenez, Arturo (Org.). Edible and Medicinal Mushrooms: Technology
and Applications. 1. ed. Lavras: John Wiley & Sons Ltd. p. 15–33.
Doughari, J. H. (2015). An Overview of Plant Immunity. Journal of Plant Pathology &
Microbiology.
Eira, A. (1992). Cultivo de cogumelos (compostagem, condução e ambiente). Anais
da III Reunião Itinerante de Fitossanidade do Instituto Biológico.
FAOSTAT. (2014). Food and Agricultural Organization of the United Nations.
Finney, K. N. et al. (2009). The reuse of spent mushroom compost and coal tailings for
energy recovery: Comparison of thermal treatment technologies. Bioresource
Technology.
Fiori-Tutida, A. C. G et al. (2007). Extratos de Lentinula edodes e Agaricus blazei sobre
Bipolaris sorokiniana e Puccinia recondita f. sp. tritici, in vitro. Summa
Phytopathologica.
Firenzuoli, F., Gori, L., Lombardo, G. (2008). The medicinal mushroom Agaricus blazei
murrill: Review of literature and pharmaco-toxicological problems. Evidence-
based Complementary and Alternative Medicine. [S.l: s.n.].
Girardi, G. et al. Use of monitoring technologies in the cultivation of mushrooms
agaricus blazei for family agriculture. Brazilian Journal of Development, v. 4, n.
7, p. 3745–3754, 2018.
Gobbi, V. et al. (2016). Spent mushroom substrate as organic fertilizer: Vegetable
organic trials. Acta Horticulturae, v. 1146, p. 49–56.

42
Gomes, D. et al. (2016). Censo paulista de produção de cogumelos comestíveis e
medicinais. Pesquisa & Tecnologia. [S.l: s.n.].
Gomes, M. (2018) Agronegócio: Consumo e produção de cogumelos cresce no Brasil.
Disponível em:
<https://www.correiobraziliense.com.br/app/noticia/economia/2018/01/29/
internas_economia,656318/consumo-e-producao-de-cogumelos-no-
brasil.shtml>. Acesso em: 05/09/2019.
Grangeia, C. et al. (2011). Effects of trophism on nutritional and nutraceutical potential
of wild edible mushrooms. Food Research International, v. 44, n. 4, p. 1029–
1035.
Grimm, D.; Wösten, H. A. B. (2018a). Mushroom cultivation in the circular economy.
Applied Microbiology and Biotechnology, v. 102, p. 7795–7803.
Grimm, D.; Wösten, H. A. B. (2018b). Mushroom cultivation in the circular economy.
Applied Microbiology and Biotechnology, v. 102, n. 18, p. 7795–7803.
Herawati, L.; Istifadah, N. (2019). The potential of Spent Substrate of Oyster (Pleurotus
ostreatus) dan Shiitake (Lentinula edodes) Mushrooms to Control Damping-off
Disease (Rhizoctonia solani) in Tomato. Cropsaver, v. 1, n. 2, p. 93.
IBGE. (2012). Levantamento sistemático da produção agrícola. . [S.l: s.n.].
Ishikawa, N. et al. (2017). Principais Cogumelos Comestíveis Cultivados E Nativos Do
Estado De. Pesquisa & Tecnologia, v. 2, n. 14, p. 1–8. Disponível em:
<www.aptaregional.sp.gov.br>. Acesso em: 05/09/2019.
Jordan, S. N., Mullen, G. J.; Murphy, M. C. (2008). Composition variability of spent
mushroom compost in Ireland. Bioresource Technology.
Kalač, P. (2013). A review of chemical composition and nutritional value of wild-
growing and cultivated mushrooms. Journal of the Science of Food and
Agriculture, v. 93, n. 2, p. 209–218.
Kang, D. S et al. (2017). Defense response and suppression of Phytophthora blight
disease of pepper by water extract from spent mushroom substrate of Lentinula
edodes. Plant Pathology Journal, v. 33, n. 3, p. 264–275.

43
Kora, C.; McDonald, M. R.; Boland, G. J. (2003). Sclerotinia rot of carrot: an example
of phenological adaptation and bicyclic development by Sclerotinia
sclerotiorum. Plant Disease, 87(5), 456-470.
Kulshreshtha, S. (2018). Mushroom Biomass and Spent Mushroom Substrate As
Adsorbent to Remove Pollutants. [S.l: s.n.].
Kulshreshtha, S. et al. (2013). Cultivation of Pleurotus citrinopileatus on handmade
paper and cardboard industrial wastes. Industrial Crops and Products.
Kumaraswamy, R. V. et al. (2018). Engineered chitosan based nanomaterials:
Bioactivities, mechanisms and perspectives in plant protection and growth.
International Journal of Biological Macromolecules, v. 113, n. 2017, p. 494–506.
Disponível em: <https://doi.org/10.1016/j.ijbiomac.2018.02.130>. Acesso em:
05/09/2019.
Kwak, A. et al. (2015). Effect of spent mushroom substrates on Phythopthora Blight
disease and growth promotion of pepper. Journal of Mushroom.
Kwak, A. et al. (2015). Water extract from Spent Mushroom Substrate of Hericium
erinaceus suppresses bacterial wilt disease of tomato. Mycobiology2, v. 4, n. 3,
p. 311–318.
Largeteau, M. L. et al. (2011). The medicinal Agaricus mushroom cultivated in Brazil:
Biology, cultivation and non-medicinal valorisation. Applied Microbiology and
Biotechnology, v. 92, n. 5, p. 897–907.
Lima, C. U., Gris, E. F.; Karnikowski, M. G. (2016). Antimicrobial properties of the
mushroom Agaricus blazei–integrative review. Revista Brasileira de
Farmacognosia, 26, 780-786. Disponível em:
<http://dx.doi.org/10.1016/j.bjp.2016.05.013>. Acesso em: 05/09/2019.
Lopes, R. X. et al. (2015). Application of spent Agaricus subrufescens compost in
integrated production of seedlings and plants of tomato. International Journal of
Recycling of Organic Waste in Agriculture, v. 4, n. 3, p. 211–218.
Lumsden, R. D. (1979). Histology and physiology of pathogenesis in plant diseases
caused by Sclerotinia species. Phytopathology, 69(8), 890-895.

44
Ma, Y. et al. (2014). A Study on recycling of spent mushroom substrate to prepare
chars and activated carbon. BioResources.
Magalhães, A.C.; Moreira, B.R.A.; Zied, DC. (2018). Axenic cultivation of Pleurotus
ostreatus Var. Florida in supplemented sugarcane bagasse briquettes.
Engenharia Agricola, v. 38, n. 6, p. 835–843.
Mao, Y et al. (2018). Effects of biochar from spent mushroom substrate on the
physicochmical properties in a nutrient-poor soil. Adsabs.Harvard.Edu, v. 20, p.
3961. Disponível em:
<http://adsabs.harvard.edu/abs/2018EGUGA..20.3961M>. Acesso em
15/11/2019.
McLaren, D. L., Conner, R. L., Kutcher, H. R., Platford, R. G., Lamb, J. L., & Lamey,
H. A. (2004). Predicting diseases caused by Sclerotinia sclerotiorum on canola
and bean—a western Canadian perspective. Canadian journal of plant
pathology, 26(4), 489-497.
Medina, E. et al. (2012). Relationships between soil physico-chemical, chemical and
biological properties in a soil amended with spent mushroom substrate.
Geoderma.
Meng, L. et al. Feasibility of co-composting of sewage sludge, spent mushroom
substrate and wheat straw. Bioresource Technology, v. 226, p. 39–45, 2017.
Disponível em: <http://dx.doi.org/10.1016/j.biortech.2016.11.054>. Acesso em:
15/11/2019.
Merriman, P. R. (1976). Survival of sclerotia of Sclerotinia sclerotiorum in soil. Soil
Biology and Biochemistry, 8(5), 385-389.
Michereff, S. J. (2001). Proteção de plantas na agricultura sustentável. [S.l: s.n.].
Miles, P. G.; Chang, S-T. (2004). Mushroom: Cultivation, Nutritional Value, Medicinal
Effect, and Environmental Impact. [S.l: s.n.].
Mingyou, W. et al. (2017). Advances in Utilization of Agricultural Waste Based on
Production of Edible Fungi. Asian Agricultural Research, v. 09, p. 92–95.

45
Mocan, A. et al. (2018). Chemical composition and bioactive properties of the wild
mushroom: Polyporus squamosus (Huds.) Fr: A study with samples from
Romania.
Mohd Hanafi, F. H. et al. (2018). Environmentally sustainable applications of agro-
based spent mushroom substrate (SMS): an overview. Journal of Material
Cycles and Waste Management, v. 20, n. 3. Disponível em:
<http://dx.doi.org/10.1007/s10163-018-0739-0>. Acesso em: 15/11/2019.
Morrall, R. A. A.; Dueck, J. (1982). Epidemiology of Sclerotinia stem rot of rapeseed in
Saskatchewan. Canadian Journal of Plant Pathology, 4(2), 161-168.
Muszyńska, B. et al. (2015). In vitro cultures and fruiting bodies of culinary-medicinal
Agaricus bisporus (white button mushroom) as a source of selected biologically-
active elements. Journal of Food Science and Technology.
Narh Mensah, D. L. et al. (2018). Biochemical characterization and efficacy of
Pleurotus, Lentinus and Ganoderma parent and hybrid mushroom strains as
biofertilizers of attapulgite for wheat and tomato growth. Biocatalysis and
Agricultural Biotechnology, v. 16, p. 63–72. Disponível em:
<https://doi.org/10.1016/j.bcab.2018.07.021>. Acesso em: 15/11/2019.
Nelson, B. D.; Hertsgaard, D. M.; Holley, R. C. (1989). Disease progress of Sclerotinia
wilt of sunflower at varying plant populations, inoculum densities, and
environments. Phytopathology, 79(12), 1358-1363.
Oancea, F. et al. (2017). Enhancement of biostimulant activity of spent Pleurotus
substrate for seedling production. Acta Horticulturae, v. 1164, p. 55–61.
Okolie, O. C. et al. (2019). The Production of Liquid Biofertilizer from Cassava Peels
and Spent Mushroom Substrates Using Microbial Inoculants. Asian Journal of
Biotechnology and Bioresource Technology.
Owaid, M. N.; Abed, IA.; Al-Saeedi, SSS. (2017). Applicable properties of the bio-
fertilizer spent mushroom substrate in organic systems as a byproduct from the
cultivation of Pleurotus spp. Information Processing in Agriculture, v. 4, n. 1, p.
78–82. Disponível em: <http://dx.doi.org/10.1016/j.inpa.2017.01.001>.
Disponivel em: 22/06/2019.

46
Owaid, M. N. (2014). Growth Performance and Cultivation of Four Oyster Mushroom
Species on Sawdust and Rice Bran Substrates. Journal of advances in
biotechnology.
Parada, R. Y. et al. (2012). Suppression of fungal and bacterial diseases of cucumber
plants by using the spent mushroom substrate of lyophyllum decastes and
pleurotus eryngii. Journal of Phytopathology.
Pardo-Giménez, A. et al. (2018). Recycling of the biomass waste defatted almond meal
as a novel nutritional supplementation for cultivated edible mushrooms. Acta
Scientiarum. Agronomy, v. 40, n. 1, p. 39341.
Paredes, C. et al. (2016). Effects of spent mushroom substrates and inorganic fertilizer
on the characteristics of a calcareous clayey-loam soil and lettuce production.
Soil Use and Management, v. 32, n. 4, p. 487–494.
Phan, C. W.; Sabaratnam, V. (2012). Potential uses of spent mushroom substrate and
its associated lignocellulosic enzymes. Applied Microbiology and
Biotechnology.
Piccinin, E; Di Piero, R. M; Pascholati, S. F. (2010). (Lentinula edodes) mushroom
reduces growth of plant pathogens and leaf spot severity in sorghum. Summa
Phytopathologica.
Poppe, J. (2004). Agricultural Wastes As Substrates for Oyster Mushroom. Mushroom
Growers’ Handbook.
Rosa, L. H. (2006). Cultivo do “Cogumelo do sol”. Fundação Centro Tecnológico de
Minas Gerais, p. 1–18.
Roy, S. et al. (2015). Evaluation of Spent Mushroom Substrate as biofertilizer for
growth improvement of Capsicum annuum L. Journal of Applied Biology &
Biotechnology.
Royse, D. J.; Baars, J.; Tan, Q. (2017). Current Overview of Mushroom Production in
the World. In: Zied, Diego Cunha; Pardo-Gimenez, Arturo (Org.). Edible and
medicinal mushrooms: technology and applications. 1. ed. [S.l.]: y John Wiley &
Sons Ltd. p. 5–13.

47
Rzymski, P. et al. (2017). Cultivation of Agaricus bisporus enriched with selenium, zinc
and copper. Journal of the Science of Food and Agriculture, v. 97, n. 3, p. 923–
928.
Saeid, A.; Patel, A. (2019). Valorization of ash and spent mushroom substrate via solid-
state solubilization by Acidithiobacillus ferrooxidans. Waste Management, v. 87,
p. 612–620. Disponível em: <https://doi.org/10.1016/j.wasman.2019.02.048>.
Acesso em: 15/11/2019.
Sagar, M. P. et al. (2009). Indigenous technical knowledge about the use of spent
mushroom substrate. Indian Journal of Traditional Knowledge.
Sánchez, C. (2010). Cultivation of Pleurotus ostreatus and other edible mushrooms.
Applied Microbiology and Biotechnology, v. 85, n. 5, p. 1321–1337.
Sendi, H. et al. (2013). Spent Mushroom Waste as a Media Replacement for Peat
Moss in Kai-Lan (Brassica oleracea var. Alboglabra) Production. The Scientific
World Journal.
Siddhant, S. C. S. (2009). Recycling of spent oyster mushroom substrate to recover
additional value. Kathmandu University Journal of Science Engineering and
Technology.
Silva, M. S. et al. (2018). Review: Potential biotechnological assets related to plant
immunity modulation applicable in engineering disease-resistant crops. Plant
Science. [S.l: s.n.].
Silva, C. D. O. G., et al. (2022). Outdoor versus indoor cultivation: Effects on the
metabolite profile of Agaricus subrufescens strains analyzed by untargeted
metabolomics. Food Chemistry, 374, 131740.
Singh, R.; Chauhan, M. (2016). Effective Management of Agro-Industrial Residues as
Composting in Mushroom Industry and Utilization of Spent Mushroom Substrate
for Bioremediation. In: RATHOURE, ASHOK; DHATWALIA, VINOD K. (Org.).
Toxicity and Waste Management Using ioremediation. 1. ed. india: Engineering
Science Reference. p. 447.
Siqueira, F. et al. (2017). Bioeconomia: Resíduos Lignocelulósicos Agroindustriais
Pré-Tratados Por Basidiomicetos Para Nutrição Animal. [S.l: s.n.]. Disponível

48
em:
<https://www.alice.cnptia.embrapa.br/bitstream/doc/1083019/1/AnaisIXSICOG
VIIISNCCFelix2017.pdf>. Acesso em: 22/06/2019.
Souza, L. G. D. (2016). Cultivo de Lentinula edodes e Pleurotus ostreatus em bagaço
de cana-de-açúcar Cultivo de Lentinula edodes e Pleurotus ostreatus em
bagaço de cana-de-açúcar. p. 58.
Sung, H-J. et al. (2018). Physico-chemical, Nutritional, and Enzymatic Characteristics
of Shiitake Spent Mushroom Substrate (SMS). Korean Society of Life Science,
v. 28, n. 11, p. 1339–1346.
Taofiq, O. et al. (2019). Agaricus blazei Murrill from Brazil: An ingredient for
nutraceutical and cosmeceutical applications. Food and Function.
Triyono, S. et al. (2019). Cultivation of straw mushroom (Volvariella volvacea) on oil
palm empty fruit bunch growth medium. International Journal of Recycling of
Organic Waste in Agriculture.
Tuhy, Ł. et al. (2015). Conversion of spent mushroom substrate into micronutrient
fertilizer via biosorption in a pilot plant. Ecological Engineering, v. 84, p. 370–
374. Disponível em: <http://dx.doi.org/10.1016/j.ecoleng.2015.09.032>. Acesso
em: 20/06/2019.
Tummo, K.; Thammasittirong, A.; Thammasittirong, S. N. R. (2018). Agricultural waste
after cultivation of pleurotus ostreatus as a potential biomass resource for
ethanol production. Journal of the International Society for Southeast Asian
Agricultural Sciences.
Vargas-Isla, R.; Ishikawa, NK.; Py-Daniel, V. (2013). Contribuições etnomicológicas
dos povos indígenas da Amazônia. Biota Amazônia, v. 3, n. 1, p. 58–65.
Viecelli, C. A. et al. (2009). Indução de resistência em feijoeiro por filtrado de cultura
de Pycnoporus sanguineus contra Pseudocercospora griseola. Tropical Plant
Pathology.
Vieira, F. R.; De Andrade, M. C. N. (2016). Optimization of substrate preparation for
oyster mushroom (Pleurotus ostreatus) cultivation by studying different raw

49
materials and substrate preparation conditions (composting: phases I and II).
World Journal of Microbiology and Biotechnology, v. 32, n. 11.
Vieira, F. R. et al. (2019). Exploring oyster mushroom (Pleurotus ostreatus) substrate
preparation by varying phase I composting time: changes in bacterial
communities and physicochemical composition of biomass impacting
mushroom yields. Journal of Applied Microbiology, v. 126, n. 3, p. 931–944.
Wiafe-Kwagyan, M.; Odamtten, G.T. (2018). Use of Pleurotus eous Strain P-31 Spent
Mushroom Compost (SMC) as Soil Conditioner on the Growth and Yield
Performance of Capsicum annuum L. and Solanum lycopersicon L. Seedlings
under Greenhouse Conditions in Ghana. Tropical Life Sciences Research, v.
29, n. 1, p. 173–194.
Win, TT.; Ohga, S. (2018). Study on the Cultivation of <i>Agaricus blazei</i>
(Almond Mushroom) Grown on Compost Mixed with Selected Agro-Residues.
Advances in Microbiology.
Wu, B. M.; Subbarao, K. V.; Liu, Y. B. (2008). Comparative survival of sclerotia of
Sclerotinia minor and S. sclerotiorum. Phytopathology, 98(6), 659-665.
Zhang, RH.; Duan, ZQ.; LI, ZG. (2012). Use of Spent Mushroom Substrate as Growing
Media for Tomato and Cucumber Seedlings. Pedosphere.
Zhu, H. et al. (2018). Preparation, characterization and antioxidant activity of
polysaccharide from spent Lentinus edodes substrate. International Journal of
Biological Macromolecules.
Zhu, J. et al. (2015). Co-digestion of spent mushroom substrate and corn stover for
methane production via solid-state anaerobic digestion. Journal of Renewable
and Sustainable Energy.
Zied, DC. et al. (2016). Selection of strains for shiitake production in axenic substrate.
World Journal of Microbiology and Biotechnology, v. 32, n. 10, p. 2–7.
Zied, DC. et al. (2018). Using of Appropriated Strains in the Practice of Compost
Supplementation for Agaricus subrufescens Production. Frontiers in
Sustainable Food Systems.Abrar A., Syed et al. (2009). Biological Efficiency

50
And Nutritional Contents Of Pleurotus florida (Mont.) Singer Cultivated On
Different Agro-wastes. Nature and Science, v. 1, n. 2, p. 127–129.

51
5 ARTIGO 01 – Artigo publicado na revista Saudi Journal of Biological Sciences
(2022) 2756-2765. https://doi.org/10.1016/j.sjbs.2021.12.058
A new circular economy approach for integrated production of tomatoes and
mushrooms
Vandinelma Oliveira Vieiraa, Aparecido Almeida Conceiçãoa, Joice Raisa Barbosa Cunhab,
Antony Enis Virginio Machadoc, Euziclei Gonzaga de Almeidad, Eustáquio Souza Diase,
Lucas Magalhães Alcantaraf, Félix Gonçalves de Siqueirag*
aPhD student, Federal University of Mato Grosso, Biotechnology and Biodiversity of the Pro Centro Oeste Network, Cuiabá, Mato Grosso, Brazil.
bPhD student, Federal University of Lavras, Agricultural Microbiology, Lavras, Minas Gerais, Brazil.
cMaster in Biotechnology, Federal University of Tocantins, Guripi, Tocantins, Brazil.
dProfessor, Federal University of Mato Grosso, Institute of Biosciences, Department of Botany and Ecology, Cuiabá, Mato Grosso, Brazil.
eProfessor, Federal University of Lavras, Department of Biology, Lavras, Minas Gerais, Brazil.
fPhD student, University of Guelph, Department of Animal Biosciences, Guelph, Ontario, Canada.
gResearcher, Embrapa Agroenergy, Brasília, Distrito Federal, Brasil.
Resumo
Substrato de cogumelo usado é o subproduto gerado no final do ciclo de cultivo do
cogumelo. Pode ser usado na agricultura para diversos fins, incluindo produção de
mudas, condicionamento do solo ou aplicação como fertilizante orgânico. O tomate é
uma das culturas mais importantes do mundo, exigindo cuidados consideráveis, tanto
a nível nutricional como de controlo de doenças. O objetivo deste estudo foi investigar
a viabilidade do substrato de cogumelo usado como fonte de nutrientes para mudas
de tomate e desenvolver um sistema integrado de co-produção de tomate e cogumelo.
Para a produção de mudas, diferentes composições foram avaliadas com substrato
de cogumelo usado de Pleurotus ostreatus ou substrato colonizado com Agaricus
bisporus. Os parâmetros avaliados foram taxa de germinação, qualidade das mudas
e análises físico-químicas. Um sistema de produção integrado de tomate e cogumelo
foi desenvolvido usando um pote de 40 litros dividido em camadas superior (substrato
de cogumelo usado e solo), intermediário (substrato de cogumelo usado de P.
ostreatus) e inferior (cascalho). Para a produção de mudas, as plantas tratadas com

52
o substrato colonizado com A. bisporus apresentaram comprimento superior de raiz
(10,1 cm) e de parte aérea (6,6 cm). A coprodução de tomate e cogumelos também
se mostrou viável. Neste sistema de co-cultivo entre tomate e cogumelo, o tratamento
com o substrato colonizado por A. bisporus diferiu dos demais, apresentando este
tratamento elevados rendimentos de tomate (2,35 kg / vaso) e cogumelos (1,33 kg /
vaso) dentro o mesmo balde. Com este sistema de coprodução, o tempo de produção
de tomate foi reduzido em 60 dias e a produção contínua de cogumelos prolongada
em 120 dias. Esses achados mostram uma abordagem sustentável para o manejo de
diferentes resíduos agroindustriais, incentivando o uso desses resíduos para a
produção de olericultura e fungicultura.
Palavras-chave: Fungicultura, Co-cultivo de tomate e cogumelo, bioestimulantes
verdes, SMS.
Abstract
Spent mushroom Substrate is the by-product generated at the end of the mushroom
growing cycle. It can be used in agriculture for different purposes, including seedling
production, soil conditioning or application as an organic fertilizer. Tomato is one of the
world´ s most important crops, requiring considerable care, in terms of both nutrition
and disease control. The objective of this study was to investigate the viability of spent
mushroom substrate as a nutrient source for tomato seedlings and develop an
integrated tomato and mushroom co-production system. For seedling production,
different compositions were evaluated with spent mushroom substrate from Pleurotus
ostreatus or substrate colonized with Agaricus bisporus. The parameters evaluated
comprised germination rate, seedling quality and physicochemical analysis. A tomato
and mushroom integrated production system was developed using a 40-liter pot
divided into upper (spent mushroom substrate and soil), middle (spent mushroom
substrate from P. ostreatus) and lower (gravel) layers. For seedlings production, plants
treated with the substrate colonized with A. bisporus presented a superior root length
(10.1 cm) and aerial part length (6.6 cm). Co-production of tomato and mushrooms
was also shown to be viable. In this co-cultivation system between tomato and
mushroom, the treatment with the substrate colonized with A. bisporus differed from
others, with this treatment presenting high yields of tomato (2.35 kg/plant pot) and
mushrooms (1.33 kg/plant pot) within the same bucket. With this co-production system,

53
the tomato production time was reduced by 60 days and prolonged continuous
mushroom production by 120 days. These findings show a sustainable approach to
manage different agroindustrial residues, encouraging the use of these residues for
olericulture and fungiculture production.
Keywords: Fungiculture, Tomato and mushroom co-cultivation, green biostimulants,
SMS.
1. Introduction
Spent Mushroom Substrate (SMS) is a by-product of the mushroom industry.
During fungal cultivation, colonized biomass undergoes a series of alterations, often
considered as a pretreatment by the fungus, which, together with the mycelial mass
and its constituents (glucan, chitin, etc.), result in the production of SMS with a series
of characteristics that make it a product with high biotechnological value for
development of bioproducts for agriculture, fuels and livestock (Grimm and Wösten,
2018; Hassan et al., 2019; Siwulski et al., 2019).
Mushrooms can be cultivated on varied agroindustry residual green biomasses
(RGB). Such versatility enables different activities to be interconnected, such that a co-
product generated by one activity can serve as a raw material for a subsequent activity.
In this context, a number of co-products from Brazilian agriculture, including cereals,
oilseeds, straw and bagasse, can all be applied for mushroom production, which, in
turn, can subsequently generate SMS that can be employed as substrate for vegetable
and fruit seedling production (Collela et al., 2019). As such, SMS is a coproduct with
important biotechnological potential, suitable for connecting agricultural systems within
the precepts of the circular economy (Grimm et al., 2020; Meyer et al., 2020; Wan
Mahari et al., 2020; Zied et al., 2020). The palm oil (Elaeis guineensis) agribusiness,
for example, generates large amounts of RGB. In 2017, the State of Pará (Brazil)
produced approximately 480 thousand tons of oil, together with approximately 1.4
million tons of solid and liquid waste (Marlina et al., 2015). In addition to mushroom
production, these biomasses can have numerous applications, including thermal
elnergy, second generation fuels, chemicals, biomaterials and seedling production
(Abdeltawab and Khattab, 2018; Aziz and Hanafiah, 2020; Madusari, 2018; Mahidin et
al., 2020; Triyono et al., 2019).

54
For the production of vegetable seedlings, alternative substrates can offer
potential in both reduction of costs and in the development of sustainable and
environmentally friendly growing systems (Kraska et al., 2018). In this context, tomato
(Solanum lycopersicum) is an excellent model for investigation, given its economic
importance and phytosanitary particularities.
In order to improve efficiency in mushroom production, diversify sources of
vegetable biomass have been evaluated for the preparation of substrates, mainly
according to regional availability (de Siqueira et al., 2011; Gonçalves et al., 2010;
Martos, 1932). In this context, palm oil-producing regions, such as the state of Pará in
Brazil, offer potential for the implementation of circular economy models, integrating
production of oils (palm oil), mushrooms, olericulture (tomatoes) and substrates for
vegetable seedlings. The integration of production chains (palm oil culture and
fungiculture) has been shown to be possible through the use of byproducts or co-
products from palm oil industry (Kumla et al., 2020; Triyono et al., 2019).
The employment of substrates that are abundant and eficiente for seed
germination can reduce costs in the agricultural sector, especially for smallholder
producers (Madusari, 2018; Yau and Murphy, 2000). In this context, there has been a
significant increase in the use of sustainable and/or environmentally friendly substrates
as alternatives to inorganic or peat-based products for the production of greenhouse
vegetables.
The aim of this study was to investigate the feasibility of employing colonized
mushroom substrates (Agaricus bisporus), SMS (A. bisporus and Pleurotus ostreatus),
and palm oil residues as substrates for the production of tomato seedlings. The study
demonstrated a new system for integrated mushroom and tomato production in
acclimatized greenhouses, as a model of integration of palm oil production,
fungiculture and olericulture within the precepts of the circular economy.
2. Materials and methods
2.1 Biomaterials employed in substrate formulations
Experiments on tomato seedling production and integrated cultivation of tomato
and P. ostreatus were carried out in a controlled greenhouse facility at Support Center
for Energy Cultures (Núcleo de Apoio a Culturas Energéticas - NACE), an extension
of Embrapa Agroenergy.

55
The SMS of P. ostreatus (SMS-P), SMS of A. bisporus (SMS-A) and substrate
colonized by A. bisporus (CAB) were all acquired from local commercial mushroom
producers in the Federal District and the states of São Paulo and Paraná.
For the composting preparation, P. ostreatus SMS was mixed with grasses
(Brachiaria and Andropogon) and chicken manure in a proportion of 3:2:1 (v/v dried
base), moistened at 70%. This compost mixture was piled up and turned over every
two days, for a period of 3 weeks. The compost was further manipulated, as follows:
(1) drying at room temperature for production of Pleurotus ostreatus Phase I compost
(SC-1); (2) aeration of SC-1 in a mechanized forced air system (air compressor type)
for a period of seven days for preparation of Pleurotus ostreatus phase II compost (SC-
2). Both composts were crushed (0.5 mm) to obtain a granulometry compatible with
the seed germination tray cells. Agaricus bisporus was cultivated in a previously
composted substrate obtained directly from the mushroom farmer.
The bacterial strain Pseudomonas putida KT2440 (B) was selected based on
its potential in plant growth promotion (Carrasco et al., 2019; He et al., 2019; Li et al.,
2019). The strain was inoculated in Luria Bertani liquid medium and incubated under
agitation (180 rpm) at 30ºC for a 7-day period. The culture was then centrifuged,
resuspended in sterile distilled water, and adjusted to a concentration of 1.25x109
cells/mL. An aliquot of 1 mL was applied per cell tray containing tomato seeds.
2.2 Tomato seedling production treatments
Cherry tomato seedling production (Solanum lycopersicum Mill. var.
cerasiforme) was conducted under controlled environmental conditions of an average
temperature of 30 ºC, and micro aspersion irrigation three times a day for 15-minute
periods to maintain humidity at 70%. Two seeds were sown per cell in rigid polystyrene
trays previously filled with each treatment. After sowing, the seeds were covered with
1 cm of substrate. All experiments were conducted in triplicate.
Several formulations were prepared containing SMS as the main component of
the seedling substrate. Across the substrate formulations, eight different treatments
were assessed for seedling production that contained the following materials: soil
(dystroferric red latosol – sieved (0.45 mm)), substrate colonized with A. bisporus, A.
bisporus SMS, P. putida, SC-1 and SC-2. Proportions employed for each treatment
are described in Table 1 (T1 to T8). The commercial substrate Carolina Soil Class XVI

56
(Composed of peat, vermiculite, organic waste, class A agro-industrial organic waste
and limestone) was included as the control treatment for all experiments. Treatment 1
(T1-Table 1) was further examined for suitability in formulations, following addition of
different nutritional conditioners (NCs) in various combinations and proportions (T9 to
T12). The NCs employed in formulations were derived from residues from the palm oil
agroindustry. Finally, substrate formulation T10 was also further evaluated following
the addition of vermiculite (25% and 50%) (T13 to T14 - Table 1).
Table 1: Substrate formulations for cherry tomato seedling production
Treatments Substrate formulation description Substrate
components (%)
Control Carolina Soil Commercial Substrate 100
T1 Substrate colonized with A. bisporus 100
T2 SMS A. bisporus 100
T3 SMS A. bisporus + soil 75/25
T4 SMS P. ostreatus + P. putida 100
T5 Substrate for Pleurotus phase I 100
T6 Substrate for Pleurotus phase II 100
T7
SMS A. bisporus + SMS P. ostreatus/P.
putida + soil
25/25/50
T8
SMS A. bisporus + SMS P. ostreatus/P.
putida
50/50
T9 T1 + Palm kernel cake + palm oil ash + palm
oil fiber
50 + (15/15/20)
T10 T1 + Palm kernel cake + palm oil ash + palm
oil fruit bunch
50 + (15/15/20)
T11 T1 + Palm kernel cake + palm ash 50 + (25/25)

57
T12 T1 + Palm kernel cake + palm oil ash + palm
oil fiber + palm oil fruit bunch
50 + (15/15/10/10)
T13 T10 + vermiculite 75/25
T14 T10 + vermiculite 50/50
2.3 Mineralogical composition
The substrate from treatment T10 (substrate colonized with A. bisporus + Palm
kernel cake + palm oil ash + palm oil fruit bunch) and the control treatment (commercial
substrate) were further analysed in terms of pH, cation-exchange capacity (CEC),
humidity (at 60-65°C), percentage of organic Carbon, total organic matter, total
Nitrogen, total Phosphorus (P2O5), Potassium (K2O), Zinc, Manganese, Calcium,
Magnesium, Sulfur, Boron, Copper, and Iron. All mineralogical analyses were
performed by Nativa Laboratório de Agrícolas LTDA, on the basis of Inductively
coupled plasma atomic emission spectroscopy (ICP-OES) (Embrapa, 1997).
2.4 Germinative and quality parameters of seedlings
Evaluations relating to germination were conducted from sowing to thinning (20
days). Calculated variables comprised germination (G), germination speed index
(GSI), mean germination speed (MGS) and mean germination time (MGT). Analysis of
physical quality parameters was performed 30 days after sowing, with focus on root
length, aerial length, wet mass, and dry mass according to Collela et al. (2019). For
evaluation of variables in the seedling phase, three random plants were collected to
compose one experimental unit.
2.5 Integrated production of tomatoes and mushrooms (Pleurotus ostreatus)
For the integrated mushroom and tomato cultivation system, henceforth referred
to as co-cultivation, plastic pots with 40 L capacity were filled with three distinct layers
(Fig. 1). Since fresh SMS is composed of viable living fungal mycelium and plant
biomass with depleted nutrients, what incapacitate mushroom production, it is
hypothesized that adding more nutrients to this residue, especially nitrogen, may allow
the fungus to produce mushrooms a second time. Thus, the nitrogen present in the soil

58
supplement can provide the necessary nutrients for the fungal fructification
reactivation.
The co-cultivation experimental design consisted of five treatments with eight
repetitions each, varying in terms of the composition of the soil and substrate in the
upper layer (layer 3 – Fig. 1, supplementary fig. 2), as described in Table 2. The soil-
substrate content used in this layer were mixed proportionally (v/v). The volume of
SMS employed in the middle layer (layer 2 – Fig. 1, supplementary fig. 2) was also
proportional (v/v) to the upper layer volume. The lower layer (layer 1 – Fig. 1) consisted
of medium sized gravel, up to approximately 3 cm depth, to enable drainage of excess
water or slurry, while also allowing mushroom fruiting. After the pots were assembled
with all the layers, tomato seedlings were planted in the upper layer. For this, seedlings
originated from the substrate treatment T10 ((Substrate colonized with A. bisporus +
Palm kernel cake + palm oil ash + palm oil fruit bunch), given the positive results in the
previous tests.
Fig. 1. Layering scheme employed in the integrated cherry tomato and mushroom co-cultivation production system, conducted in 40-liter plastic pots inside an acclimatized greenhouse.

59
Fig. 2. Illustrative picture showing pots filled with a middle layer of SMS and an upper layer with soil mixed with diverse substrates employed in the integrated cherry tomato and mushroom co-cultivation production system.
Table 2. Composition of the upper layer treatments employed in cherry tomato and
mushroom (P. ostreatus - commercial SMS from two harvests) co-cultivation.
Treatment Upper layer composition
T1 Soil + Superficial fertilization with commercial fertilizer (5.7 g
Ammonium Sulfate, 19 g MAP (Monoammonium Phosphate), 7 g
KCl (Potassium Chloride)
T2 50% soil + 25% substrate colonized with A. bisporus + 12.5%
Palm kernel cake + 12.5% Palm fruit bunch fiber
T3 50% soil + 50% substrate for the cultivation of A. bisporus (same
used in experiment 2).
T4 50% soil + 50% substrate colonized with A. bisporus
T5 50% soil + 50% SMS P. ostreatus
For each of the treatments, after seedling planting, upper layers were irrigated
and maintained in the greenhouse under high humidity (85%) and a temperature range
between 25 and 35ºC. Nebulization was conducted twice a day for 5 minute periods
(at 6:00 a.m. and 6:00 p.m.).
After 60 days, the top layer from each treatments, with the exception of T1
(commercial substrate control), were supplemented (nutrient and soil replacement)

60
with 3 liters of a mixture containing 60% soil, 20% P. ostreatus SMS and 20% chicken
manure compost (Plante-Brasil - Super-Adubo®, Composition: pH 8, CTC-C 5.33, C/N
Ratio 5, humidity 30%, total P2O5 1%, total N 3%, total Mg 1%, H2O.K2O 2%, total Ca
4%, organic carbon 15% and CTC 80 mmol C/kg). After supplementation, co-
cultivation was extended for a further 60 days.
Mushrooms were harvested over 120 days during co-cultivation, across two
periods (i) 0-60 days and (ii) 61-120 days. Production was calculated according to the
average ratio of quantity (kg) of fresh mushrooms per pot and biological efficiency
(EB%) (ratio of fresh mushrooms harvested (grams) per dry substrate (grams)),
calculated on the basis of the following formula:
𝐸𝐵% = 𝑓𝑟𝑒𝑠ℎ 𝑤𝑒𝑖𝑔ℎ𝑡 𝑜𝑓 𝑚𝑢𝑠ℎ𝑟𝑜𝑜𝑚 (𝑔)
𝑑𝑟𝑦 𝑠𝑢𝑏𝑠𝑡𝑟𝑎𝑡𝑒 𝑤𝑒𝑖𝑔ℎ𝑡 (𝑔)∗ 100
Tomatoes were harvested from the initial stage of ripening (red spots) until ripe
(totally red). The yield of the tomato harvest was calculated according to the average
ratio of the quantity (kg) of tomatoes produced per pot.
2.6 Statistical analysis
Statistical analysis was performed using the program R, version 3.4.3. A
completely randomized design was employed in the experiments, with means
compared by the Scott Knott and Tukey test at the 5% significance level, with normality
and homogeneity verified on the basis of Shapiro Wilk, Bartlett and Poisson tests.
3. Results
3.1 Formulations for tomatoes seedlings
The qualitative parameters for the development of cherry tomato seedlings in
the different subtracts are presented in Table 3. We observed that the commercial
control presented parameters superior to the other treatments, thus being necessary
to improve the T1 to T8 formulations. Even if the index of germination speed and
germination speed of T1 is inferior to other treatments, the vigor of the seedlings in T1
by the set of parameters of root size and aerial part stood out from the others (Table
3). Among the treatments presented, the treatment T1 (Substrate colonized with A.
bisporus) presented percentage of germination statistically equal to the control (100%),
the parameters of root size were higher than the others treatments and presented size
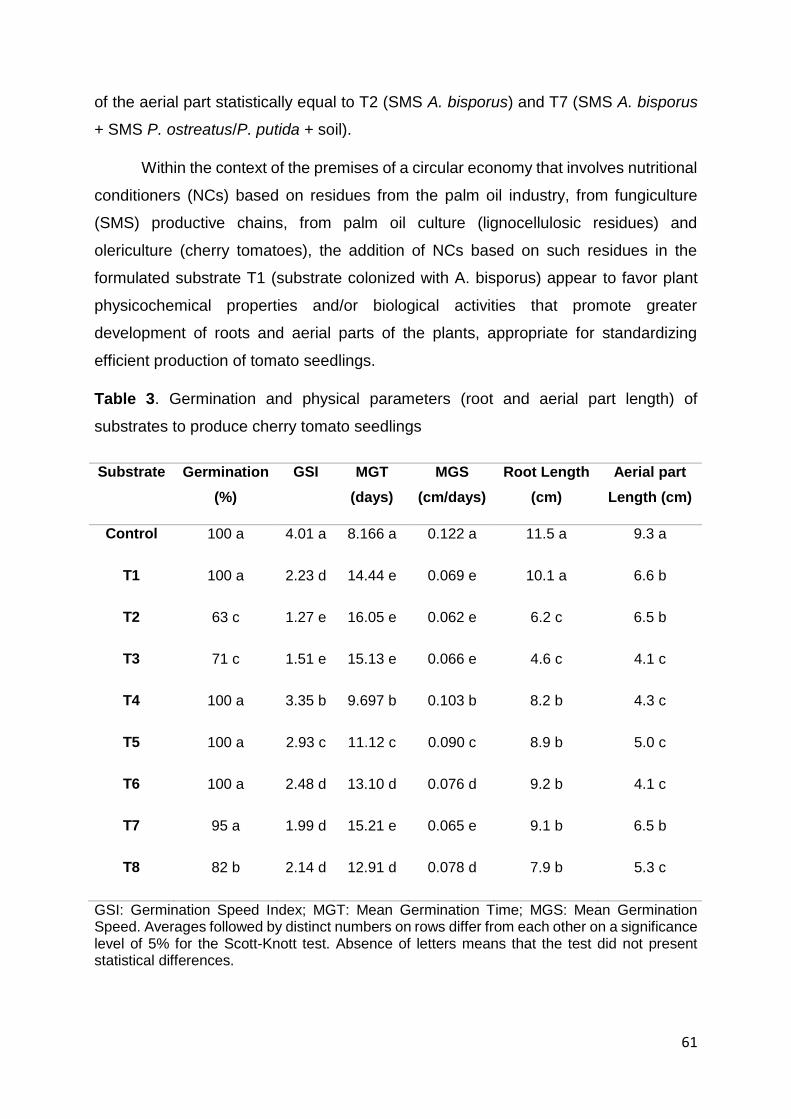
61
of the aerial part statistically equal to T2 (SMS A. bisporus) and T7 (SMS A. bisporus
+ SMS P. ostreatus/P. putida + soil).
Within the context of the premises of a circular economy that involves nutritional
conditioners (NCs) based on residues from the palm oil industry, from fungiculture
(SMS) productive chains, from palm oil culture (lignocellulosic residues) and
olericulture (cherry tomatoes), the addition of NCs based on such residues in the
formulated substrate T1 (substrate colonized with A. bisporus) appear to favor plant
physicochemical properties and/or biological activities that promote greater
development of roots and aerial parts of the plants, appropriate for standardizing
efficient production of tomato seedlings.
Table 3. Germination and physical parameters (root and aerial part length) of
substrates to produce cherry tomato seedlings
GSI: Germination Speed Index; MGT: Mean Germination Time; MGS: Mean Germination Speed. Averages followed by distinct numbers on rows differ from each other on a significance level of 5% for the Scott-Knott test. Absence of letters means that the test did not present statistical differences.
Substrate Germination
(%)
GSI MGT
(days)
MGS
(cm/days)
Root Length
(cm)
Aerial part
Length (cm)
Control 100 a 4.01 a 8.166 a 0.122 a 11.5 a 9.3 a
T1 100 a 2.23 d 14.44 e 0.069 e 10.1 a 6.6 b
T2 63 c 1.27 e 16.05 e 0.062 e 6.2 c 6.5 b
T3 71 c 1.51 e 15.13 e 0.066 e 4.6 c 4.1 c
T4 100 a 3.35 b 9.697 b 0.103 b 8.2 b 4.3 c
T5 100 a 2.93 c 11.12 c 0.090 c 8.9 b 5.0 c
T6 100 a 2.48 d 13.10 d 0.076 d 9.2 b 4.1 c
T7 95 a 1.99 d 15.21 e 0.065 e 9.1 b 6.5 b
T8 82 b 2.14 d 12.91 d 0.078 d 7.9 b 5.3 c

62
Results showed that the addition of NCs improved the performance of the
substrates colonized with A. bisporus to similar or even superior levels to those
obtained with the control for the production of tomato seedlings (Table 4). Treatments
T10 and T12 stood out from the others, with results equal to the control for all
parameters evaluated, with the exception of the length of aerial plant parts, for which
T10 and T12 showed results superior to the control. Therefore, these results show that
the by-products of the palm oil industry are excellent NCs for substrates employed in
tomato seedling production. T9 also showed promising results for all evaluated
parameters, with the exception of germination percentage, which was lower than
observed in the control. In comparison with T10 and T12, treatment T9 resulted in
plants with shorter aerial length, although this was similar to the control. The difference
between formulation T9 and formulations T10 and T12 was the absence of palm oil
fruit bunch fiber in the former, indicating a beneficial effect provided by this fiber.
Curiously, treatment T11 presented the worst results for all parameters
evaluated, with the exception of aerial part length, for which there was no significant
difference in relation to the control. The substrate from T11 showed higher compaction
compared to the other substrates, which may have negatively interfered with plant
germination and development. Moreover, this substrate received a higher proportion
of ash in relation to the others, which may also have raised the salinity of the substrate
to a higher level than that recommended for seedling production substrates.
Table 4. Germination and physical parameters (root and aerial part length) of
substrates for the production of cherry tomato seedlings using substrate colonized with
A. bisporus (T1) and nutritional conditioners
Treatments Germination
(%)
GSI MGT
(days)
MGS
(days)
Root
Length
(cm)
Aerial part
Length
(cm)
Control 97 a 5,04 a 6,06 a 0,16 9,9 a 8,4 b
T9
74 b 3,76 a 4,76 a 0,22 5,9 b 8,1 b
T10 80 b 4,19 a 4,94 a 0,20 9,0 a 14,6 a

63
T11
31 c 1,38 b 2,37 b 0,55 4,7 b 5,5 b
T12 91 a 4,66 a 5,89 a 0,17 8,5 a 11,9 a
GSI: Germination Speed Index; MGT: Mean Germination Time; MGS: Mean Germination Speed. Averages followed by distinct numbers on rows differ from each other on a significance level of 5% for the Scott-Knott test. Absence of letters means that the test did not present statistical differences. CO: Control (Commercial substrate); T9: 50% Substrate colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil + 20% Palm oil fiber; T10: 50% Substrate colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil + 20% Palm oil fruit bunch; T11: 50% Substrate colonized with A. bisporus + 25% Palm kernel cake + 25% Palm oil ash; T12: 50% Substrate colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil ash + 10% Palm oil fiber + 10% Palm oil fruit bunch.
Results of quantitative parameters from seedlings produced with the substrate
T10 are shown in Table 4. Although only enabling a qualitative analysis, which was not
statistically significant, a superiority in seedlings produced in the T10 substrate was
observed in terms of root growth and aerial part, which were well developed and ready
for transplantation (Fig. 3).
In order to improve the physicochemical parameters of the formulation T10
selected from experiment 2, the addition of vermiculite was tested in proportions of
25% or 50% of the total substrate. Although it was expected that the addition of
vermiculite would increase the substrate’s porosity and water retention capacity, this
did not result in improved germination rates or seedling quality (Table 6). For
germination percentage and GSI, the addition of vermiculite had a negative effect, with
values below the control.

64
Fig. 3. Tomato plants within 30 days cultivation, samples for evaluation of root growth
and aerial parts parameters. CO: Control (Commercial substrate); T9: 50% Substrate
colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil + 20% Palm oil
fiber; T10: 50% Substrate colonized with A. bisporus + 15% Palm kernel cake + 15%
Palm oil + 20% Palm oil fruit bunch; T11: 50% Substrate colonized with A. bisporus +
25% Palm kernel cake + 25% Palm oil ash; T12: 50% Substrate colonized with A.
bisporus + 15% Palm kernel cake + 15% Palm oil ash + 10% Palm oil fiber + 10% Palm
oil fruit bunch.
Table 6. Formulations of substrates for the production of cherry tomato seedlings from
experiment 3 and their respective parameters for germination and seedlings quality, in
the different substrates using the T10-CABDC (experiment 2) substrate as the base,
in addition to vermiculite in two proportions (25% and 50%)
Treatments G (%) GSI MGT
(days
)
MGS
(days
)
DW WW % H2O Root
lengt
h
(cm)
Aerial
part
lengt
h
(cm)
Co 93 a 3.8 a 6.6 0.15 0.030 a 0.31 a 89 10 a 5 a

65
GSI: Germination Speed Index; MGT: Mean Germination Time; MGS: Mean Germination
Speed; DW: Dry weight of tomato plants; WW: Wet weight of tomato plants; % H2O: The
difference between dry weight and wet weight; CO: Control (Commercial substrate); T13 = T10
+Vermiculite 25%; T14 = T10 + 50% vermiculite. Averages followed by distinct numbers on
rows differ from each other on a significance level of 5% for the Scott-Knott test. Absence of
letters means that the test did not present statistical differences.
3.2 Analysis of mineralogical composition
Analysis of primarily essential macronutrients in the T10 (50% substrate
colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil + 20% Palm oil
fruit bunch) substrate showed values of P and K higher than those observed in the
control treatment (commercial substrate), while the values of N were identical in both
substrates (Table 5).
Table 5. Comparison of the chemical characteristics of the substrate that presented
the best result (T10) and control (commercial substrate) in the production of cherry
tomato seedlings
Analysis Sample (%)
Control (Commercial Substrate) T10
pH CaCl2 (0.01M) 5.10 6.25
CEC (mmol L-1) 600.00 394.00
Humidity (60-65°C) 10.20 7.46
Organic Carbon 5.63 5.44
Total Organic Matter 9.72 9.38
Total Nitrogen 0.70 0.70
Total Phosphorus (P2O5) 1.60 5.02
Potassium (K2O) 0.60 1.20
T13 71 b 2.4 c 6.0 0.16 0.027 a 0.38 a 92 7.3 ab 4 a
T14 90 a 3.2 b 7.4 0.13 0.026 a 0.35 a 92 9.0 ab 4 a

66
Zinc 0.001 0.010
Manganese 0.020 0.020
Calcium 0.75 2.27
Magnesium 4.31 0.47
Sulfur 0.01 0.55
Boron 0.001 0.010
Copper 0.002 0.010
CEC: cation-exchange capacity
In addition to adequate mineral concentrations, results indicated that the
treatment T10 substrate presented a composition that allows good aeration and
maintenance of humidity, thus favoring the germination of tomato seeds and
development of seedlings for a longer period. Another interesting result was the
viability of tomato seedlings after 60 days of sowing. The substrate composed of T10
was able to maintain the vigor of tomato seedlings for a longer time compared to the
commercial substrate, as shown in Fig. 4.

67
Fig. 4. Cherry tomato seedlings with 60 days after sowing in the formulation T10 (50% Substrate colonized with A. bisporus + 15% Palm kernel cake + 15% Palm oil + 20% Palm oil fruit bunch) containing substrate colonized with A. bisporus and nutritional conditioners (lignocellulosic residues of the palm oil industry), and control formulation (commercial substrate).
3.3 Co-cultivation of tomatoes and mushrooms
Given the literature that shows the potential of integrating mushroom production
with agricultural production and the positive results obtained in this study with T10
substrate for production of tomato seedlings, a system of co-cultivation of tomato and
mushrooms (P. ostreatus) was examined in a controlled environment setup
(acclimatized greenhouse). The system was designed to enable both mushroom
harvesting after two production periods, together with a soil microenvironment
appropriate for eficiente cherry tomato production (Fig. 1). In this system, soil-substrate
enriched with nutrients met both the physiological demands of the plant and provided
nutrients (N and others) for the fungal substrate, promoting the physiological processes
of mushroom fruiting. It should be noted that both the tomatoes and mushrooms were
produced in a controlled environment that was not 100% ideal for either organism but
offering intermediate growth conditions to favor the co-cultivation.
Five different treatments were tested, consisting of various soilsubstrate
combinations for the upper layer (Table 2) and SMS from P. ostreatus in the middle
layer for all treatments. Mushrooms began to appear 10 days after the beginning of the
co-cultivation (Fig. 5) and continued to fruit until the end of the experimente (120 days).

68
Fig. 5. Illustration of the production of mushrooms within 10 days in the co-cultivation system with cherry tomatoes (still in the initial phase of seedling growth).
After 30 days co-cultivation, noticeable differences were apparent in tomato
plant development ascorss the treatments (Fig. 6), with T4 (soil + substrate colonized
with A. bisporus) the most noticeable in terms of growth promotion. This treatment was
also the first to result in flowering, approximately 50 days after seedling transplanting.
Fig. 6. Illustration of the growth of cherry tomato plants with 30 days in the co-cultivation system with P. ostreatus mushroom. Treatments differ on the content of the upper layer only. T1: Soil + Superficial fertilization with commercial fertilizer (5.7 g Ammonium Sulfate, 19 g MAP (Monoammonium Phosphate), 7 g KCl (Potassium Chloride); T2: 50% soil + 25% substrate colonized with A. bisporus + 12.5% Palm kernel cake + 12.5% Palm fruit bunch fiber; T3: 50% soil + 50% substrate for the cultivation of A. bisporus (same used in Step 2, but not inoculated); T4: 50% soil + 50% substrate colonized with A. bisporus; T5: 50% soil + 50% SMS P. ostreatus.
Results obtained following co-cultivation (Fig. 7) were evaluated in terms of
production (yield) of fresh mushrooms (Kg fresh mushrooms/pot) before and after
nutritional supplementation, biological efficiency (BE = g fresh mushrooms/g of dry
substrate), and tomato production (Kg per plant over a 60 days of harvest). Tomato
yields were highest at T3 and T4, with 2.6 and 2.4 Kg/pot, respectively.

69
Fig. 7. Production of fresh mushrooms (Pleurotus ostreatus) and cherry tomatoes in the integrated production system at 60 and 120 days. Treatments differ on the content of the upper layer only. T1: Soil + Superficial fertilization with commercial fertilizer (5.7 g Ammonium Sulfate, 19 g MAP (Monoammonium Phosphate), 7 g KCl (Potassium Chloride); T2: 50% soil + 25% substrate colonized with A. bisporus + 12.5% Palm kernel cake + 12.5% Palm oil fruit bunch fiber; T3: 50% soil + 50% substrate for the cultivation of A. bisporus (same used in experiment 2, but not inoculated); T4: 50% soil + 50% substrate colonized with A. bisporus; T5: 50% soil + 50% SMS P. ostreatus. BE – Biological efficiency. In order to compare both mushrooms and tomatoes yield, capital letters were used for the comparison of the same groups across different treatments (bars). Different letters differ from each other on a significance level of 5% for the Tukey test.
Mushroom yield over the 120 days of co-cultivation in T4 was significantly higher
than the others (1.3 Kg/pot), while also presenting the best biological efficiency
(66.4%). Mushroom production after supplementation of the plants at 60 days showed
better results only for T4, indicating that supplementation with N interfered positively in
the growth of P. ostreatus. For T1, T2, T3 and T5 was observed a biological efficiency
(%) mushroom of 24.4%, 12.8%, 45.3%, and 38.1%, respectively.
Discussion
A number of studies have reported that SMS contains high levels of organic
matter, carbohydrates, proteins, lipids, nitrogen, phosphorus, potassium and other
nutrients, with such characteristics suitable in applied in agricultural systems (Jordan
et al., 2008; Medina et al., 2012; Roy et al., 2015). The benefits of SMS as na organic
fertilizer and soil conditioner have also been reported (Courtney and Mullen, 2008;

70
Hackett, 2015). In addition to organic matter, SMS also have a population of
microorganisms, in addition to the original fungus, which are beneficial for plant
development, favoring competition with natural pathogens such as pathogenic fungi
and nematodes, as demonstrated in the work of de Moraes et al., (2020).
The use of SMS from mushroom production has demonstrated the effectiveness
of these materials in agriculture as substitutes for substrate components in the
production of seedlings of diferente plant cultivars. Zhang et al. (2012) described SMS
as a suitable co-product for the production of vegetable seedlings, using this substrate
for greenhouse production of cucumber and tomato seedlings. According to the
authors, SMS served as a potential substitute for peat, providing better
physicochemical properties that enabled good growth and development of cucumber
seedlings. Porto et al. (2018) observed a positive effect on germination time when SMS
from A. bisporus was added to substrate for the cultivation of bell pepper. This effect
may be due to the rapid release of a large number of ions which can be easily absorbed
by the plant, resulting in an increase in dry matter in a short period of time (Rashid et
al., 2018a, 2018b).
A number of benefits of SMS in agriculture have been described (Hanafi et al.,
2018), for example in the integration of fungiculture (SMS or colonized substrates) with
the development of products such as biofertilizers, using waste from two production
chains. The use of SMS has been indicated in circular economy models (Grimm and
Wösten, 2018), being applied as a substrate for substitution of peat usage in plant
production. In several countries, the use of natural peat (extraction from spring
environments) is now prohibited or restricted (Paula et al., 2017; Yau and Murphy,
2000; Zhang et al., 2012), given the negative environmental impact of it´s extraction
and resulting soil structure disturbance (Moxey and Moran, 2014).
Another important aspect of this type of substrate is the potential positive
interaction between plant and microorganisms present. In addition to the metabolites
produced by A. bisporus and its own cell wall constituents (chitin and b-glucans), these
substrates also present a rich microbiota which can influence the rhizosphere region
(de Moraes et al., 2020), favoring the establishment of a microbiota beneficial for
tomato seedling production. Tall and Meyling (2018) suggested that the microorganism
Beauveria bassiana is able to interact beneficially with corn plants depending on the

71
nutritional availability of the substrate. The authors described that the interaction could
vary from antagonista to mutualist and conclude that the plant-fungal relationship is
therefore complex, contradictory, and context dependent. Nonetheless, as described
herein, the inclusion of SMS from A. bisporus mushrooms in substrates for cultivation
or seedling production has shown positive results. The colonized substrate of A.
bisporus, also presents good qualities as a substrate for tomato. In the presente
experiment, the addition of a nutritional conditioner based on palm residue improves
the quality of the colonized substrate of A. bisporus for tomato seedlings.
For plant development, in addition to interactions with microorganisms, the
substrate needs to have an adequate balance of nutrients, which were mostly found in
the T10 substrate of this work such as Phosphorus and Potassium, both favoring the
development of the plant (Brito et al., 2018). Although phosphorus is na essential
element for plant growth, a is normally abundantly presente in soil, it cannot be
assimilated directly by plants. Vassilev et al. (2006) showed the potential of using
microorganisms in the processes of phosphorus solubilization in fermentative systems
as an alternative to fertilizer production, since organic acid producing microorganisms
are able to solubilize phosphates of aluminum, iron and calcium. According to Helm et
al. (2009), among other nutrients essential to the quality development of plants are
calcium and magnesium, where the first stimulates the absorption of other ions to
maintain the structure and normal function of cell membranes, and the later acts on
the composition of chlorophyll, proto-chlorophyll, pectin and phytin. Micronutrients,
such as copper, iron, manganese or zinc, are important as cofactors in the activation
of several plant enzymes, in the formation of chlorophyll and indoleacetic acid, nitrogen
absorption, and in enzymatic processes in general for plant growth (Brito et al., 2018).
Substrate pH is also important in seedling production, as it influences the availability
of nutrients and physiological balance of the plant. Shibata and Demiate (2003)
described an ideal pH between 6.0 and 7.0 for nutrient availability.
Vermiculite is a common component in several substrate formulations for
seedlings production, given its desirable physicochemical characteristics. However,
the results obtained in this study show that the addition of vermiculite is not always
necessary. A potential explanation for this may be that the A. bisporus substrate
already provides the same benefits as vermiculite, although this may vary according
to the components employed in the formulation of the A. bisporus mushroom substrate.

72
The tomato production yield in the co-culture conditions is in accord with
literature that describes alternative substrates such as vermicompost with coconut
fibers and rice husk ash (Truong et al., 2018), or mixtures of vermicompost, vermiculite
and cattle manure (Costa et al., 2018). Most tomato harvests observed in these
previous studies occurred in up to 80 days (Oliveira et al., 2020), in contrast to this
study, where an earlier harvest was observed at only 60 days in average.
SMS of Agaricus species has proven to be a good substrate componente for
fertigation of vegetables such as tomato, lettuce and cucumber (Stoknes et al., 2019).
This was also observed in the current work, with the treatment containing substrate
colonized with A. bisporus (T4) showing better results in terms of tomato growth and
productivity, when compared to the control and T5, which received SMS of Pleurotus
without the addition of any nutritional conditioners (Fig. 7). Other crops such as lettuce
and cucumber have also benefited from the use of biomass from fungiculture, such as
in the form of organic fertilizers (Elsakhawy and tawfik, 2019), demonstrating that there
is potential in the integration of production chains. In this work, it became evident that
in addition to the production of substrates for plant seedlings, it was also possible to
produce mushrooms (P. ostreatus) and tomatoes in the same environment, further
strengthening the principle of a circular economy.
The action of degradation of plant matter by fungi is widely studied and they also
develop important functions in symbiotic interactions with plants. Much has been
discussed about the importance of endophytic fungi for the development of plants and
vice versa (Kothe and Turnau, 2018). Basidiomycetes known as white-rot fungi are
highly related to decomposition processes due to their lignocellulolytic enzymes
(Sudarson et al., 2014), but little is discussed about the beneficial interactions with
living plants. Mushroom, such as those from Pleurotus Genera have been demonstrate
great potential against nematodes which can benefit the plant development
(Elkhateeb, Daba and Soliman, 2021). The interaction between plants and fungi is a
sophisticated strategy in which both benefit using a chemical language with the
production and secretion of several biomolecules, including phytohormones that can
modulate plant development (Rashid et al., 2018a, 2018b). Understanding this
relationship will allow for greater food productivity in an efficient and sustainable
manner. The cultivation of different mushroom species offers potential in different
agricultural production systems through the use of SMS as raw material for animal food

73
(birds, fish, and larvae), biofertilizers, bionematicide, biofungicide and others.
Industrialized nations could potentially include this in strategies to achieve self-
sustaining environments, while for developing nations the main incentive would be to
promote mushroom cultivation in circular food chains, likely achieving food security
and better public health. Smallholder farmers could also profit through mushroom
cultivation as an additional source of income and food (Grimm et al., 2020). This
circular economy model that integrates fungiculture with other sectors offers
considerable potential for implementation in different regions of Brazil, given the
availability of green biomasses for mushroom production, as well as the need to
generate alternative sources of food and income for smallholder farms.The oil palm
industry and the oil processing agroindustry generate many lignocellulosic residues,
which can be a source of raw material for the production of mushrooms, which can
then provide generated SMS applicable as a fertilizer for different crops. Grimm et al.
(2020) reviewed potential approaches to integrate mushroom production with other
food generating chains, reporting that the application of 100 tons of SMS from A.
bisporus per hectare resulted in a 59% increase in grain production as a result of higher
levels of soil phosphorus, potassium, nitrogen and organic matter.
Here we show, for the first time, how smallholder farmers may benefit from the
co-production of mushrooms and olericulture in regions that both require improved
living conditions and that generate large amounts of agroindustrial by-products, such
as those from the oil palm industry. In this way, results presented herein provide a
basis for the successful implementation of a circular economy model through the
integration of mushroom and cherry tomato production.
Conclusion
This work shows that it is possible to integrate productive chains in a sustainable
system such as fungiculture, olericulture, and palm oil culture through the use of spent
mushroom substrate (SMS) or colonized substrates from A. bisporus supplemented
with residues from the palm oil agroindustry for the production of cherry tomato
seedlings. Furthermore, an integrated and sustainable system of co-production of
tomato and mushrooms is proposed that is applicable to smallholder farmers.
References
Abdeltawab, A.M., Khattab, M.S.A., 2018. Utilization of Palm Kernel Cake as a

74
Ruminant Feed for Animal: A Review. Asian J. Biol. Sci. 11, 157–164.
https://doi.org/10.3923/ajbs.2018.157.164
Aziz, N.I.H.A., Hanafiah, M.M., 2020. Life cycle analysis of biogas production from
anaerobic digestion of palm oil mill effluent. Renew. Energy 145, 847–857.
https://doi.org/10.1016/j.renene.2019.06.084
Brito, T., Soares, V.N., Barreto, B., Almeida, S., Rodrigues, D.B., Tunes, L.M. De,
2018. Parâmetros de crescimento de plântulas de pimentão na semeadura em
diferentes substratos 117, 69–76.
Carrasco, J., Tello, M.L., De Toro, M., Tkacz, A., Poole, P., Péerez-Clavijo, M.,
Preston, G., 2019. Casing microbiome dynamics during button mushroom
cultivation: Implications for dry and wet bubble diseases. Microbiol. (United
Kingdom) 165, 611–624. https://doi.org/10.1099/mic.0.000792
Collela, C.F., Soares Costa, L.M.A., De Moraes, T.S.J., Zied, D.C., Rinker, D.L., Dias,
E.S., 2019. Potential utilization of spent agaricus bisporus mushroom substrate
for seedling production and organic fertilizer in tomato cultivation. Cienc. e
Agrotecnologia 43. https://doi.org/10.1590/1413-7054201943017119
Costa, E., Binotti, F.F. da S., Cardoso, E.D., Júnior, D.B.L., Zoz, T., Zuffo, A.M., 2018.
Cherry tomato production on different organic substrates under protected
environment conditions. Aust. J. Crop Sci. 12, 87–92.
https://doi.org/10.21475/ajcs.18.12.01.pne749
Courtney, R.G., Mullen, G.J., 2008. Soil quality and barley growth as influenced by the
land application of two compost types. Bioresour. Technol. 99, 2913–2918.
https://doi.org/10.1016/j.biortech.2007.06.034
de Moraes, T.S.J., Costa, L.M.A.S., Souza, T.P., Collela, C.F., Dias, E.S., 2020.
Fungal and bacterial population from spent mushroom substrate used to cultivate
tomato plants. Cienc. e Agrotecnologia 44, 1–8. https://doi.org/10.1590/1413-
7054202044010120
de Siqueira, F.G., Martos, E.T., da Silva, E.G., da Silva, R., Dias, E.S., 2011. Biological
efficiency of agaricus brasiliensis cultivated in compost with nitrogen
concentrations | Eficiência biológica de agaricus brasiliensis em composto com
concentrações de nitrogênio. Hortic. Bras. 29, 157–161.

75
Elsakhawy, T., tawfik, wael, 2019. Evaluation of Spent Mushroom Substrate extract
as biofertilizer for growth improvement of Rice (Oriza sativa). Egypt. J. Soil Sci. 0,
0–0. https://doi.org/10.21608/ejss.2019.18835.1320
Embrapa, 1997. Manual de Métodos de Análise de solo. Brazil. Centro Nacional de
Pesquisa de solos. 2, 1-212.
Gonçalves, C.C. de M., Paiva, P.C. de A., Dias, E.S., Siqueira, F.G. de, Henrique, F.,
2010. Avaliação do cultivo de Pleurotus sajor-caju (fries) sing. sobre o resíduo de
algodão da industria têxtil para a produção de cogumelos e para alimentação
animal. Ciência e Agrotecnologia 34, 220–225. https://doi.org/10.1590/s1413-
70542010000100028
Grimm, D., Kuenz, A., Rahmann, G., 2020. Integration of mushroom production into
circular food chains. Org. Agric. https://doi.org/10.1007/s13165-020-00318-y
Grimm, D., Wösten, H.A.B., 2018. Mushroom cultivation in the circular economy. Appl.
Microbiol. Biotechnol. 102, 7795–7803. https://doi.org/10.1007/s00253-018-9226-
8
Hackett, R., 2015. Spent mushroom compost as a nitrogen source for spring barley.
Nutr. Cycl. Agroecosystems 102, 253–263. https://doi.org/10.1007/s10705-015-
9696-3
Hanafi, F.H., Rezania, S., Mat Taib, S., Md Din, M.F., Yamauchi, M., Sakamoto, M.,
Hara, H., Park, J., Ebrahimi, S.S., 2018. Environmentally sustainable applications
of agro-based spent mushroom substrate (SMS): an overview. J. Mater. Cycles
Waste Manag. 20, 1383–1396. https://doi.org/10.1007/s10163-018-0739-0
Hassan, S.S., Williams, G.A., Jaiswal, A.K., 2019. Lignocellulosic Biorefineries in
Europe: Current State and Prospects. Trends Biotechnol. 37, 231–234.
https://doi.org/10.1016/j.tibtech.2018.07.002
He, Y., Pantigoso, H.A., Wu, Z., Vivanco, J.M., 2019. Co-inoculation of Bacillus sp. and
Pseudomonas putida at different development stages acts as a biostimulant to
promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 127, 196–
207. https://doi.org/10.1111/jam.14273
Helm, C.V., Coradin, J.H., Kestring, D.R., 2009. Avaliação da Composição Química

76
dos Cogumelos Comestíveis Agaricus bisporus, Agaricus brasiliensis, Agaricus
bisporus portobello, Lentinula edodes e Pleorotus ostreatus. Comun. Técnico -
Embrapa Florestas 1–7.
Hoffmann, R.B., 2018. Brazilian Journal of Animal and Environmental Research
Brazilian Journal of Animal and Environmental Research 168–178.
Jordan, S.N., Mullen, G.J., Murphy, M.C., 2008. Composition variability of spent
mushroom compost in Ireland. Bioresour. Technol. 99, 11–418.
https://doi.org/10.1016/j.biortech.2006.12.012
Klein, C., 2015. Substratos Alternativos Para Produção De Mudas. Rev. Bras.
Energias Renov. 4, 43–63. https://doi.org/10.5380/rber.v4i3.40742
Kothe, E., Turnau, K., 2018. Editorial: Mycorrhizosphere communication: Mycorrhizal
fungi and endophytic fungus-plant interactions. Front. Microbiol. 9, 1–4.
https://doi.org/10.3389/fmicb.2018.03015
Kraska, T., Kleinschmidt, B., Weinand, J., Pude, R., 2018. Cascading use of
Miscanthus as growing substrate in soilless cultivation of vegetables (tomatoes,
cucumbers) and subsequent direct combustion. Sci. Hortic. (Amsterdam). 235,
205–213. https://doi.org/10.1016/j.scienta.2017.11.032
Kumla, J., Suwannarach, N., Sujarit, K., Penkhrue, W., Kakumyan, P., Jatuwong, K.,
Vadthanarat, S., Lumyong, S., 2020. Cultivation of mushrooms and their
lignocellulolytic enzyme production through the utilization of agro-industrial waste.
Molecules 25, 1–41. https://doi.org/10.3390/molecules25122811
Li, T., Zhang, J., Shen, C., Li, H., Qiu, L., 2019. 1-Aminocyclopropane-1-carboxylate:
A Novel and Strong Chemoattractant for the Plant Beneficial Rhizobacterium
Pseudomonas putida UW4. Mol. Plant-Microbe Interact. 32, 750–759.
https://doi.org/10.1094/MPMI-11-18-0317-R
Madusari, S., 2018. The Effects of Soil Amended with Solid Fibrous Waste on the
Morphological Quality of Oil Palm ( Elaeis guineensis Jacq .) Seedling 15–22.
Mahidin, Saifullah, Erdiwansyah, Hamdani, Hisbullah, Hayati, A.P., Zhafran, M., Sidiq,
M.A., Rinaldi, A., Fitria, B., Tarisma, R., Bindar, Y., 2020. Analysis of power from
palm oil solid waste for biomass power plants: A case study in Aceh Province.

77
Chemosphere 253, 126714. https://doi.org/10.1016/j.chemosphere.2020.126714
Mamimin, C., Chanthong, S., Leamdum, C., O-Thong, S., Prasertsan, P., 2021.
Improvement of empty palm fruit bunches biodegradability and biogas production
by integrating the straw mushroom cultivation as a pretreatment in the solid-state
anaerobic digestion. Bioresour. Technol. 319, 124227.
https://doi.org/10.1016/j.biortech.2020.124227
Marlina, L., Sukotjo, S., Marsudi, S., 2015. Potential of Oil Palm Empty Fruit Bunch
(EFB) as Media for Oyster Mushroom, Pleurotus ostreatus Cultivation. Procedia
Chem. 16, 427–431. https://doi.org/10.1016/j.proche.2015.12.074
Martos, E.T., 1932. Cultivation of sisal. Nature 129, 62.
Medina, E., Paredes, C., Bustamante, M.A., Moral, R., Moreno-Caselles, J., 2012.
Relationships between soil physico-chemical, chemical and biological properties
in a soil amended with spent mushroom substrate. Geoderma.
https://doi.org/10.1016/j.geoderma.2011.12.011
Meyer, V., Basenko, E.Y., Benz, J.P., Braus, G.H., Caddick, M.X., Csukai, M., De
Vries, R.P., Endy, D., Frisvad, J.C., Gunde-Cimerman, N., Haarmann, T., Hadar,
Y., Hansen, K., Johnson, R.I., Keller, N.P., Kraševec, N., Mortensen, U.H., Perez,
R., Ram, A.F.J., Record, E., Ross, P., Shapaval, V., Steiniger, C., Van Den Brink,
H., Van Munster, J., Yarden, O., Wösten, H.A.B., 2020. Growing a circular
economy with fungal biotechnology: A white paper. Fungal Biol. Biotechnol. 7, 1–
23. https://doi.org/10.1186/s40694-020-00095-z
Moreno-Salazar, R., Sánchez-García, I., Chan-Cupul, W., Ruiz-Sánchez, E.,
Hernández-Ortega, H.A., Pineda-Lucatero, J., Figueroa-Chávez, D., 2020. Plant
growth, foliar nutritional content and fruit yield of Capsicum chinense biofertilized
with Purpureocillium lilacinum under greenhouse conditions. Sci. Hortic.
(Amsterdam). 261, 108950. https://doi.org/10.1016/j.scienta.2019.108950
Moxey, A., Moran, D., 2014. UK peatland restoration: Some economic arithmetic. Sci.
Total Environ. 484, 114–120. https://doi.org/10.1016/j.scitotenv.2014.03.033
Oliveira, R.C., Quintão Lana, R.M., Queiroz Luz, J.M., Queiroz, A.A., Bertoldo, D.L.,
2020. Biofertilizer in leaf and drip applications: An alternative to increase tomato
productivity. Comun. Sci. 11, 1–6. https://doi.org/10.14295/cs.v11i0.3376

78
Paula, F.S., Tatti, E., Abram, F., Wilson, J., O’Flaherty, V., 2017. Stabilisation of spent
mushroom substrate for application as a plant growth-promoting organic
amendment. J. Environ. Manage. 196, 476–486.
https://doi.org/10.1016/j.jenvman.2017.03.038
Rashid, H. M., Abed, I.A., Owaid, M.N., 2018. Mycelia growth performance of Agaricus
bisporus in culture media of composts supplemented with Sesbania sesban straw
and phosphate rock. Curr. Res. Environ. Appl. Mycol. 8, 323–330.
https://doi.org/10.5943/cream/8/3/4
Rashid, Husam Mahmod, Abed, I.A., Owaid, M.N., 2018. Effect of Sesbania sesban
on cultivation of Agaricus bisporus, Basidiomycota, and properties of spent
mushroom compost outcome. Open Agric. 3, 652–657.
https://doi.org/10.1515/opag-2018-0068
Roy, S., S, B., Chakraborty, U., Chakraborty, B., 2015. Evaluation of Spent Mushroom
Substrate as biofertilizer for growth improvement of Capsicum annuum L. J. Appl.
Biol. Biotechnol. https://doi.org/10.7324/JABB.2015.3305
Sharvini, S.R., Noor, Z.Z., Chong, C.S., Stringer, L.C., Glew, D., 2020. Energy
generation from palm oil mill effluent: A life cycle assessment of two biogas
technologies. Energy 191, 116513. https://doi.org/10.1016/j.energy.2019.116513
Shibata, C.K.R., Demiate, I.M., 2003. Cultivation and chemical analysis of the sun
mushroom (Agaricus blazei Murril). Publ. UEPG Ciencias Biol. e da Saude 9, 21–
32. https://doi.org/10.5212/publ.biologicas.v.9i2.0003
Siwulski, M., Rzymski, P., Budka, A., Kalač, P., Budzyńska, S., Dawidowicz, L.,
Hajduk, E., Kozak, L., Budzulak, J., Sobieralski, K., Niedzielski, P., 2019. The
effect of different substrates on the growth of six cultivated mushroom species and
composition of macro and trace elements in their fruiting bodies. Eur. Food Res.
Technol. 245, 419–431. https://doi.org/10.1007/s00217-018-3174-5
Stoknes, K., Wojciechowska, E., Jasinska, A., Noble, R., 2019. Amelioration of
composts for greenhouse vegetable plants using pasteurised agaricus mushroom
substrate. Sustain. 11. https://doi.org/10.3390/su11236779
Sudarson, J., Ramalingam, S., Kishorekumar, P., Venkatesan, K., 2014. Expeditious
Quantification of Lignocellulolytic Enzymes from Indigenous Wood Rot and Litter

79
Degrading Fungi from Tropical Dry Evergreen Forests of Tamil Nadu. Biotechnol.
Res. Int. 2014, 1–6. https://doi.org/10.1155/2014/127848
Tafner Jr, A.W., Silva, F.C., 2011. Colonização japonesa, história econômica e
desenvolvimento regional do Estado do Pará. Novos Cad. NAEA 13.
https://doi.org/10.5801/ncn.v13i2.437
Tall, S., Meyling, N. V., 2018. Probiotics for Plants? Growth Promotion by the
Entomopathogenic Fungus Beauveria bassiana Depends on Nutrient Availability.
Microb. Ecol. 76, 1002–1008. https://doi.org/10.1007/s00248-018-1180-6
Triyono, S., Haryanto, A., Telaumbanua, M., Dermiyati, Lumbanraja, J., To, F., 2019.
Cultivation of straw mushroom (Volvariella volvacea) on oil palm empty fruit bunch
growth medium. Int. J. Recycl. Org. Waste Agric. https://doi.org/10.1007/s40093-
019-0259-5
Truong, H.D., Wang, C.H., Kien, T.T., 2018. Effect of Vermicompost in Media on
Growth, Yield and Fruit Quality of Cherry Tomato (Lycopersicon esculentun Mill.)
Under Net House Conditions. Compost Sci. Util. 26, 52–58.
https://doi.org/10.1080/1065657X.2017.1344594
Vassilev, N., Medina, A., Azcon, R., Vassileva, M., 2006. Microbial solubilization of
rock phosphate on media containing agro-industrial wastes and effect of the
resulting products on plant growth and P uptake. Plant Soil 287, 77–84.
https://doi.org/10.1007/s11104-006-9054-y
Wan Mahari, W.A., Peng, W., Nam, W.L., Yang, H., Lee, X.Y., Lee, Y.K., Liew, R.K.,
Ma, N.L., Mohammad, A., Sonne, C., Van Le, Q., Show, P.L., Chen, W.H., Lam,
S.S., 2020. A review on valorization of oyster mushroom and waste generated in
the mushroom cultivation industry. J. Hazard. Mater. 400, 123156.
https://doi.org/10.1016/j.jhazmat.2020.123156
Yau, P.Y., Murphy, R.J., 2000. Biodegraded cocopeat as a horticultural substrate. Acta
Hortic. https://doi.org/10.17660/ActaHortic.2000.517.33
Zhang, R.H., Duan, Z.Q., Li, Z.G., 2012. Use of Spent Mushroom Substrate as Growing
Media for Tomato and Cucumber Seedlings. Pedosphere 22, 333–342.
https://doi.org/10.1016/S1002-0160(12)60020-4

80
Zied, D.C., Sánchez, J.E., Noble, R., Pardo-Giménez, A., 2020. Use of spent
mushroom substrate in new mushroom crops to promote the transition towards a
circular economy. Agronomy 10, 1–20.
https://doi.org/10.3390/agronomy10091239
Zou, Y., Du, F., Hu, Q., Yuan, X., Dai, D., Zhu, M., 2020. Integration of Pleurotus
tuoliensis cultivation and biogas production for utilization of lignocellulosic
biomass as well as its benefit evaluation. Bioresour. Technol. 317, 124042.
https://doi.org/10.1016/j.biortech.2020.124042

81
6 ARTIGO 02 – Publicado na revista World Journal of Microbiology and
Biotechnology (2021) 37:203. https://doi.org/10.1007/s11274-021-03172-6
Characterization of extracellular secondary metabolites
in Oudemansiella canarii BRM-044600 displaying antifungal activity
against the phytopathogen Sclerotinia sclerotiorum
Vandinelma de Oliveira Vieira1; Aparecido Almeida Conceição1; Caio de Oliveira
Gorgulho Silva2; Ruben Dario Romero-Pelaez2,3; Murillo Lobo Junior4; Patrícia Verardi
Abdelnur2; João Ricardo Moreira de Almeida2; Euziclei Gonzaga Almeida1; Félix
Gonçalves de Siqueira2.
1Universidade Federal do Mato Grosso, Avenue Fernando Corrêa da Costa, nº 2367, Boa Esperança, Cuiabá, MT 78060-900, Brazil.
2Brazilian Agricultural Research Corporation, Embrapa Agroenergy, W3 Norte, PqEB, Brasília, DF 70770-901, Brazil.
3Universidade de Brasília, Asa Norte, Brasília 70910-900, Brazil.
4Brazilian Agricultural Research Corporation, Embrapa Rice and Beans, Rodovia GO-462, Km 12, Fazenda Capivara, Zona Rural Caixa Postal: 179, Santo Antônio de Goiás, GO CEP: 75375-000, Brazil.
Resumo
A doença do mofo-branco, causada pelo fitopatógeno Sclerotinia sclerotiorum,
provoca graves perdas de produtividade em várias culturas economicamente
importantes. Agentes de biocontrole, especialmente fungos filamentosos
antagonistas, são alternativas ecologicamente corretas aos fungicidas químicos
usados no manejo do mofo branco. O objetivo deste estudo foi pesquisar fungos
basidiomicetos capazes de inibir S. sclerotiorum e investigar seus metabólitos
bioativos responsáveis por atividades antifúngicas. Dois dos 17 isolados de
basidiomicetos testados inibiram o crescimento micelial de S. sclerotiorum em
experimentos de cultura em pares em placas de ágar, nomeadamente Oudemansiella
canarii BRM-044600 e Laetisaria arvalis ATCC52088. O filtrado de cultura líquida de
O. canarii BRM-044600 exibiu a maior atividade antifúngica e foi selecionado para
investigação posterior. A análise UHPLC-MS sugere que seis estrobilurinas putativas,
incluindo estrobilurina A e / ou estereoisômeros deste composto (m / z 259,1299, [M

82
+ H] +) e três estrobilurinas putativas com m / z 257,1184 ([M + H] +) são
provavelmente responsáveis pela atividade antifúngica observada no filtrado da
cultura. Pela primeira vez, este trabalho demonstrou o potencial de O. canarii para
biocontrole branco e produção de estrobilurina.
Palavras-chave: Biocontrole, Basidiomicetos, Metabólitos secundários fúngicos,
Estrobilurinas, Fitopatógeno.
Abstract
White mold disease, caused by the phytopathogen Sclerotinia sclerotiorum, provokes
severe productivity losses in several economically important crops. Biocontrol agents,
especially antagonist filamentous fungi, are environmentally friendly alternatives to the
chemical fungicides used in white mold management. The objective of this study was
to screen for basidiomycete fungi capable of inhibiting S. sclerotiorum and investigate
their bioactive metabolites responsible for antifungal activities. Two out of 17 tested
basidiomycete isolates inhibited the mycelial growth of S. sclerotiorum in pair culture
experiments on agar plates, namely Oudemansiella canarii BRM-044600 and
Laetisaria arvalis ATCC52088. O. canarii BRM-044600 liquid culture filtrate exhibited
the greatest antifungal activity and was selected for further investigation. UHPLC-MS
analysis suggests that six putative strobilurins, including strobilurin A and/or
stereoisomers of this compound (m/z 259.1299, [M + H]+) and three putative
strobilurins with m/z 257.1184 ([M + H]+) are likely responsible for the antifungal activity
observed in the culture filtrate. For the first time, this work demonstrated the potential
of O. canarii for whitemold biocontrol and strobilurin production.
Keywords: Biocontrol, Basidiomycetes, Fungal secondary metabolites, Strobilurins,
Plant pathogen.
Introduction
White mold disease, also known as Sclerotinia stem rot, caused by the
phytopathogenic fungus Sclerotinia sclerotiorum, affects a wide spectrum of
economically importante crops, including soybean in which it causes a loss of up to
70% in productivity if not effectively managed (Lehner et al. 2017). S. sclerotiorum
infection can initiate either by myceliogenic germination of soil-borne sclerotia (highly
resistant survival structures of the fungus) or, more frequently, by air-borne ascospores

83
that originate from carpogenic germination of sclerotia (Smolińska and Kowalska
2018). S. sclerotiorum infection is characterized by an initial suppression of the host
defense system (biotrophic phase) followed by a necrotrophic phase involving the
killing of plant cells and subsequent nutrient uptake. Infection is mediated by secreted
plant cell wall degrading enzymes, metabolites such as oxalic acid, and small proteins
that act as pathogen effectors or cell death inducers (Liang and Rollins 2018; Wang et
al. 2019).
White mold is difficult to control for several reasons, including the lack of
cultivars with satisfactory resistance, the long‐term survival of sclerotia in soil, and the
airborne dissemination of ascospores. Successful control of White mold in soybean is
strongly dependent on synthetic fungicides, which pose environmental concerns
(Bolton et al. 2006; Lehner et al. 2017; Smolińska and Kowalska 2018). Biological
control of S. sclerotiorum using antagonistic microorganisms represents an
environmentally friendly alternative that can be integrated into other management
practices such as crop rotation, wider row spacing and resistant/tolerant cultivar
selection. Several species of bacteria and fungi capable of controlling S. sclerotiorum
have now been identified. Among the fungi, ascomycetes such as Trichoderma
harzianum, T. virens, Clonostachys rósea and Coniothyrium minitans have been well
characterized. These organisms employ a combination of biocontrol mechanisms that
includes mycoparasitism, synthesis of antifungal secondary metabolites (such as
polyketides and non-ribosomal peptides) and induction of plant systemic resistance
(Kamal et al. 2016; Smolińska and Kowalska 2018).
Even though certain basidiomycetes show potential for the biocontrol of plant
diseases, they are still poorly studied for white mold biocontrol. Antifungal activities in
this class of fungi rely on antifungal metabolites rather than mycoparasitism, with
basidiomycetes well recognized as producers of secondary metabolites that are
secreted to eliminate competition from other fungi. Strobilurins and oudemansins, for
example, are polyketides produced by various Basidiomycota genera that, while
presenting weak phytotoxicity, display strong and broad-spectrum antifungal activity
against yeasts and filamentous fungi, including several phytopathogens such as
Botrytis cinerea and Rhizoctonia solani (Anke et al. 1977, 1983; Schramm et al. 1978;
Anke 1995; Rosa et al. 2005; Niego et al. 2021). These compounds contain a (E)-β-
methoxyacrylate group, which acts as a toxophore and inhibit mitochondrial respiration

84
by blocking electron transfer in the bc1 complex of the electron transport chain,
preventing adenosine triphosphate (ATP) synthesis (Ypema and Gold 1999; Balba
2007).
Due to slow growth and specific nutritional requirements, the employment of
basidiomycetes as a source of bioactive compounds for biotechnological applications
has historically been limited (Karwehl and Stadler 2016; Sandargo et al. 2019).
However, evolution in fermentative processes and -omics technologies has increased
the recovery and discovery of their metabolites. As such, the development of
basidiomycete products has increased considerably in recente years (Spiteller 2015;
Sandargo et al. 2019).
This work objectives were to screen for basidiomycete species with antifungal
activities against S. sclerotiorum, investigate their metabolites responsible for
antifungal activity and provide a basis for research and development of biological
pesticides.
Methodology
Microorganisms
All fungi employed in this study, namely Amylosporus campbelli BRM-055617,
Fistulina hepatica BRM-047114, Fomis fasciatus BRM-055676, Ganoderma lucidum
BRM-055670, Hydnopolyporus fimbriatus BRM-055634, Laetisaria arvalis BRM-
055839 (ATCC52088), Lentinula edodes BRM- 055677, Lentinus sajor-caju BRM-
056047, Lepiota fusciceps BRM-050074, Oudemansiella canarii BRM-044600,
Panaeolus antillarum BRM-055673, Panus lecomtei BRM-044603, Pleurotus ostreatus
BRM-055505, Pleurotus pulmonarius BRM-055674, Pleurotus sapidus BRM-044252,
Pycnoporus sanguineus BRM-055516, Trametes versicolor BRM-047439 and S.
sclerotiorum BRM-063319 belong to the Collection of Microorganisms and Microalgae
Applied to Bioenergy (CMMAABio). Specific information can be accessed through
AleloMicro database (http://alelomicro.cenargen.embrapa.br/).
Paired culture tests
Paired culture tests were performed in triplicate using a dual culture method
(Carvalho et al. 2011) between 17 selected basidiomycete isolates and S. sclerotiorum
BRM-063319. Briefly, mycelial plugs (8 mm diameter) from colony borders of S.

85
sclerotiorum and each of the basidiomycete isolates were placed equidistantly on fresh
PDA plates (potato-dextrose-agar, KASVI, São José dos Pinhais, Brazil) and
incubated at 28 °C with a 12-h photoperiod (Quimis Q315M25 Incubator, Diadema,
São Paulo, Brazil). After 7 days incubation, the pathogen's colony diameter was
measured with a translucent ruler and used to calculate the percent of pathogen
mycelial growth inhibition (MGI%) caused by basidiomycetes isolates according to the
formula described by Nascimento et al. (2019):
Here, plate∅ is the plate diameterand PMG is the pathogen’s mycelial growth
(both in centimeters). MGI% values were used to select the most promising
basidiomycete isolates for biocontrol of S. sclerotiorum.
Preparation of lyophilized liquid culture filtrates from selected basidiomycetes
Oudemansiella canarii BRM-044600, L. arvalis ATCC52088 (the two isolates
with the highest MGI% values) and G. lucidum BRM-055670 (the isolate with the lowest
MGI% value) were subjected to submerged fermentation in potato dextrose broth
(PDB, KASVI, São José dos Pinhais, Brazil) following a method adapted from
Stracquadanio et al. (2020). Ten mycelial plugs (8 mm diameter) from 7-day-old PDA
cultures of each selected basidiomycete isolate were inoculated into 250 mL flasks
containing 100 mL of PDB and maintained at 28 °C and 180 rpm for 7 days using a
shaker-incubator (MAXQ5000, Thermo Scientific, Marietta, OH, USA). The liquid
cultures filtrates were recovered by vacuum filtration through filter paper and
lyophilized (LIOTOP K105, Liobras LTDA. São Carlos, São Paulo, Brazil).
Sclerotinia sclerotiorum growth inhibition assays using lyophilized filtrates of
selected basidiomycetes
Sclerotinia sclerotiorum growth inhibition assays using selected basidiomycete
culture filtrates were performed following a protocol adapted from Dutta et al. (2019)
and Fang et al. (2011). One gram of O. canarii BRM-044600, L. arvalis ATCC52088
and G. lucidum BRM-055670 lyophilized liquid culture filtrates were diluted in 50 mL of
sterile distilled water and then added to autoclaved 4% m/v PDA medium before agar
solidification (cooled down to approximately 60 °C). Two concentrations were

86
evaluated, namely 25% v/v (i.e., a mixture of 75 mL autoclaved PDA médium and 25
mL resuspended basidiomycete culture filtrate) and 50% v/v (i.e., a mixture of 50 mL
autoclaved PDA médium and 50 mL resuspended basidiomycete culture filtrate),
equivalent to final lyophilized filtrate concentrations of 5.0 and 10.0 mg/mL,
respectively. Sterile distilled water was mixed with autoclaved PDA medium at 25% or
50% (v/v) dilutions, as described above, in control experiments. One mycelial plug (8
mm diameter) from 7-day-old S. sclerotiorum cultures was placed at the center of the
PDA plates after agar solidification and incubated at 28 °C for 7 days (sufficient time
for growth of S. sclerotiorum in control experiments). All experiments were performed
in triplicate and repeated at least five times.
A solution of lyophilized liquid culture filtrate (at 20 mg/mL) was autoclaved at
121 °C for 15 min to verify the thermostability of antifungal compounds produced by O.
canarii BRM-044600, prior to being used in S. sclerotiorum growth inhibition assays.
Oudemansiella canarii BRM‐044600 liquid culture filtrate fractionation and
application in S. sclerotiorum growth inhibition assays
One gram of O. canarii BRM-044600 lyophilized liquid culture filtrate was
dissolved in 50 mL deionized water to which 50 mL ethyl acetate was added (Emsure®,
MERCK, Darmstadt, Germany). The organic phase was recovered after vortexing and
phase separation by centrifugation (10,000×g, 5 min) (Hermle Z360K, Labortechnik
GmbH, Wehingen, Germany). The extraction procedure was repeated three times. The
organic and aqueous fractions were vacuum-dried at room temperature (CentriVap
Concentrator/Coldtrap, Labconco, Kansas City, USA), dissolved in 50 mL of distilled
water and employed in growth inhibition assays with S. sclerotiorum. The dissolved
fractions were mixed with autoclaved PDA medium prior to agar solidification at two
concentrations, 25% and 50% (v/v). S. sclerotiorum was inoculated and incubated for
7 days, as described above. Aliquots from both aqueous and organic fractions obtained
after ethyl acetate fractionation were also retrieved for UHPLC-MS analysis.
Lyophilized PDB medium without O. canarii BRM-044600 inoculum was subjected to
the same ethyl acetate fractionation protocol in parallel as a negative control for
UHPLC-MS analysis.
A larger batch of ethyl acetate fractionation was also performed with 10 g of O.
canarii BRM-044600 lyophilized liquid culture filtrate. The resulting vacuum-dried

87
organic fraction was dissolved in 2 mL of acetonitrile 50% (v/v) (ACN, Biograde
Chemicals, Durham, USA) and subjected to further fractionation using Strata C18-E
cartridges (55 μm, 70 Å, 500 mg/3 mL, Phenomenex, Torrance, USA). Each 1 mL of
sample was applied onto a cartridge pre-equilibrated with ACN 50% and sequentially
eluted with 3 mL of ACN 50%, ACN 75% and ACN 100%. Fractions were vacuum-
dried at room temperature, dissolved in 50 mL of distilled water with the aid of
sonication (Branson 2800 ultrasonic bath, Branson, Brookfield, USA) for 30 min, then
applied in S. sclerotiorum growth inhibition assays. The dissolved fractions were mixed
with autoclaved PDA médium prior to agar solidification at 50% (v/v) concentration. S.
sclerotiorum was inoculated and incubated for 7 days at 28 °C, as described above.
UHPLC‐MS analysis of the O. canarii BRM‐044600 liquid culture filtrate
UHPLC-MS analyses were performed in a Shimadzu Nexera X2 UHPLC system
(Kyoto, Japan) equipped with an Acquity UPLC HSS T3 column (diameter 2.1 mm,
length 100 mm, particle size 1.8 μm) (Waters, Milford, USA), coupled to a maXis 4G™
mass spectrometer composed of an electrospray ionization (ESI) source and a
quadrupole time-of-flight (Q-TOF) analyzer (Bruker Daltonics, Bremen, Germany).
Samples were analyzed with elution gradients 1 or 2, as specified in the text.
Gradient 1 was comprised of eluents A [formic acid (FA) 0.1% v/v] and B (acetonitrile
with FA 0.1% v/v) as follows: 0.0 to 1.0 min (0% B), 1.0 to 3.0 min (0–5% B), 3.0 to
10.0 min (5–50% B), 10.0 to 13.0 min (50–100% B), 13.0 to 15.0 min (100% B) and
15.0 to 20.0 min (0% B). Gradient 2 was comprised of eluents A (10 mM ammonium
formate plus FA 0.1% v/v in methanol) and B (10 mM ammonium formate plus FA 0.1%
v/v in 2-propanol) as follows: 0.0 to 1.0 min (0% B), 1.0 to 3.0 min (0–5% B), 3.0 to
10.0 min (5–50% B), 10.0 to 13.0 min (50–100% B), 13.0 to 15.0 min (100% B) and
15.0 to 20.0 min (0% B). Flow rate, oven temperature and injection volumes were 0.4
mL/min, 45 °C and 1 μL (gradiente 1) or 0.5 μL (gradient 2). Formic acid, ammonium
formate and 2-propanol (for LC–MS Lichropur®) were acquired from Merck
(Darmstadt, Germany). LC–MS-grade methanol and acetonitrile were acquired from
Honeywell (Seelze, Germany) and Biograde (Durham, USA), respectively.
MS data acquisition was performed in both positive and negative ion modes.
Instrument parameters were as follows: end plate offset=500 V, capillary voltage=4000
V (negative mode) or 3800 V (positive mode), nebulizer pressure = 4.0 Bar, dry gas

88
flow = 9.0 L/min, dry temperature = 200 °C, spectra rate = 3.00 Hz, detection range =
m/z 75–1000. MS/MS parameters were as follows: collision energy = 20–50 v using a
basic stepping program; cycle time of precursor ion acquisition = 3.0 s; mass range =
75–1000 m/z; spectra rate = 3 Hz; pre pulse storage = 7.0 us; funnel 1 RF = 300.0
Vpp. Sodium formate (2 mM) was directly injected through a 6-port valve at the
beginning of each chromatographic run for external calibration. UHPLC-MS data were
acquired with otofControl (Bruker Daltonics, Bremen, Germany) and HyStar
(Shimadzu, Kyoto, Japan) software and analyzed with DataAnalysis (Bruker Daltonics,
Bremen, Germany).
Molecular formulas of the detected ions were inferred through SmartFormula
(DataAnalysis version 4.2, Bruker Daltonics). Metabolites were annotated by
comparison with the literature and by searches against the Kyoto Encyclopedia of
Genes and Genomes (KEGG) (Ogata et al. 2000), MetFrag (https://msbi.ipb-
halle.de/MetFrag/) (Ruttkies et al. 2016) and Metlin (https://metlin.scripps.edu) (Xue et
al. 2020) databases.
UHPLC-MS data were processed with XCMS Online
(https://xcmsonline.scripps.edu/) (Huan et al. 2017) using the centWave method for
feature detection, the orbiwarp method for retention time correction (profStep of 1 m/z),
mzwid = 0.015, minfrac = 0.5 and bw = 5 for retention time alignment. A bucket table
was generated containing the m/z values, retention times and intensities of the
detected ions. This table was used for unsupervised multivariate analysis (principal
component analysis, PCA) using MetaboAnalyst (https://www.metaboanalyst.ca/)
(Chong et al. 2018). Normalization was conducted by the sum method with the Pareto
scale employed as a data scaling method.
A molecular network was created using the online workflow (https://ccms-
ucsd.github.io/GNPSDocumentation/) on the GNPS website http://gnps.ucsd.edu)
(Wang et al. 2017). The data was filtered by removing all MS/MS fragment ions within
± 17 Da of the precursor m/z. MS/MS spectra were window filtered by choosing only
the top six fragment ions in the ± 50 Da window throughout the spectrum. The
precursor ion mass tolerance was set to 0.02 Da and a MS/MS fragment ion tolerance
of 0.2 Da. A network was then created where edges were filtered to have a cosine
score above 0.5 and more than three matched peaks. Edges between two nodes were

89
maintained in the network if and only if each node appeared in each other's respective
top 10 most similar nodes. Finally, the maximum size of a molecular family was set to
100, and the lowest scoring edges were removed from molecular families until the
molecular family size was below this threshold.
Results
Pair cultures of basidiomycete isolates versus S. sclerotiorum
Among 17 basidiomycete isolates paired cultured along with S. sclerotiorum on
PDA plates, L. arvalis BRM-055839 and O. canarii BRM-044600 showed the highest
pathogen mycelial growth inhibition (%MGI) values, at 70.0 and 63.3%, respectively,
while G. lucidum BRM-055670 exhibited the lowest %MGI value (i.e., none) (Table 1).
Basidiomycete isolate Basidiomycete growth diameter (cm)
S. sclerotiorum growth diameter (cm)
%MGI
Laetisaria arvalis BRM 055839 6.3 2.7 70.0
Oudemansiella canarii BRM 044600 5.7 3.3 63.3
Pleurotus sapidus BRM 044252 4.5 4.5 50.0
Panaeolus antillarum BRM 055673 4.5 4.5 50.0
Lepiota fusciceps BRM 050077 4.2 4.8 46.7
Panus lecomtei BRM 044603 4.2 4.8 46.7
Hydnopolyporus fimbriatus BRM 055634 4 5 44.4
Pycnoporus sanguineus BRM 055516 3.9 5.1 43.3
Amylosporus campbelli BRM 055624 3.9 5.1 43.3
Lentinus sajor-caju BRM 056047 3.9 5.1 43.3
Fistulina hepatica BRM 047114 3.8 5.2 42.2
Trametes versicolor BRM 047439 3.6 5.4 40.0
Pleurotus pulmonarius BRM 055674 3.4 5.6 37.8
Lentinula edodes BRM 055677 3.4 5.6 37.8
Pleurotus ostreatus BRM 055505 3.4 5.7 36.7
Fomis fasciatus BRM 055675 1 9 0.0
Ganoderma lucidum BRM 055670 0 9 0.0
Table 1. Percentage of Sclerotinia sclerotiorum mycelial growth inhibition (%MGI)
caused by basidiomycete isolates in paired culture experiments. Growth
measurements were taken seven days after inoculation.

90
Visually, L. arvalis ATCC52088 showed aggressive growth towards
S.sclerotiorum and caused the pathogen's hyphae to grow vertically towards the petri
dish lid during late growth stages, likely as a response to the biological stress induced
by L. arvalis (Fig. 1). O. canarii BRM-044600 also showed aggressive growth towards
the pathogen and a contact zone between both the species was formed, likely followed
by the release of metabolites from both fungi with potential growth inhibition activities.
In contrast, when G. lucidum BRM-055670 was tested, S. sclerotiorum grew faster and
overlapped G. lucidum mycelium. Based on these results, L. arvalis BRM-055839, O.
canarii BRM-044600 and G. lucidum BRM-055670 were selected for further
investigation.
Fig. 1 Paired cultures of basidiomycete isolates (a) along with S. sclerotiorum BRM-063319 (b) on PDA plates 144 h after inoculation. L. arvalis BRM-055839, O. canarii BRM-044600, G. lucidum BRM-055670.
Sclerotinia sclerotiorum mycelial growth inhibition by selected basidiomycete
liquid culture filtrates
To evaluate whether the apparent inhibition of S. sclerotiorum on PDA pair
cultures was caused by secreted antifungal metabolites, the lyophilized liquid culture
filtrates from the two most promising basidiomycete isolates were added to PDA plates
at two concentrations − 5.0 and 10.0 mg/mL— and tested for antifungal activity against
S. sclerotiorum. A complete inhibition of S. sclerotiorum mycelial growth was observed
on PDA plates containing O. canarii BRM-044600 lyophilized culture filtrate at both
concentrations (Fig. 2), indicating that O. canarii BRM-044600 secretes antifungal

91
compounds when grown on standard PDB medium, irrespective of the presence of the
pathogen. The lyophilized supernatant of O. canarii BRM-044600 also maintained
antifungal activity after autoclaving at 121 °C for 15 min (Figure S1, Online Resource
1), indicating that the bioactive compounds are more likely non-protein metabolites (not
susceptible to denaturation) and that these metabolites are heatstable. The lyophilized
filtrate of L. arvalis ATCC52088 liquid culture, on the other hand, only partially inhibited
S. sclerotiorum on PDA plates at either 5.0 or 10.0 mg/mL.
Fig. 2 Growth of S. sclerotiorum on PDA plates containing the liquid culture filtrates of
O. canarii BRM-044600 (plates c and d, at 5.0 and 10.0 mg/mL, respectively), L. arvalis
ATCC52088 (plates e and f, at 5.0 and 10.0 mg/mL, respectively) and G. lucidum BRM-
055670 (plates g and h, at 5.0 and 10.0 mg/mL, respectively). Control cultures on PDA
mixed with water at 25% (v/v) and 50% (v/v) dilutions are shown in (a) and (b),
respectively.
As expected, the lyophilized filtrate of G. lucidum BRM-055670 (tested as a
negative control) did not display any inhibitory activity against S. sclerotiorum (Fig. 2).
Considering the greater antifungal activity, O. canarii BRM-044600 liquid culture filtrate
was selected for further investigation by metabolomics analysis.
Metabolites from O. canarii BRM‐044600 liquid culture filtrate
UHPLC-MS analysis (using chromatographic gradient 1) of O. canarii BRM-
044600 culture filtrate revealed that the antifungal compound, strobilurin A (m/z
259.1330, [M + H]+, with predicted neutral molecular formula C16H18O3), and/or

92
stereoisomers of this compound, were putatively detected as three chromatographic
peaks, i.e. [email protected] (the notation corresponds to an ion detected using the
positive ion (PI) or negative ion (NI) mode having an m/z value of 259.1299 at a
retention time of 12.2 min), [email protected] (major peak) and [email protected]
(Fig. 3a). The MS/MS spectra of m/z 259 parent ions showed high similarity to that
previously reported for strobilurin A (Schramm et al. 1978), with the following matched
product ions: m/z 91 (putative ion formula [C7H7]+, corresponding to a benzyl ion), m/z
121 (putative ion formula [C8H9O]+), m/z 167 (putative ion formula [C13H11]+), m/z 199
(putative ion formula [C14H15O]+, corresponding to [M–CO2CH3]+) and m/z 227
(putative ion formula [C15H15O2]+, corresponding to [M–CH3OH]+) (Fig. 4a). Database
searches also suggested other putative annotations for the m/z 259 ions based on
MS1 data only (Table S1, Online Resource 1). However, based on the MS/MS
similarity to strobilurin A and the well-studied antifungal activity of strobilurins, these
ions most likely correspond to strobilurin A and/or stereoisomers of this compound.
Fig. 3 UHPLC-MS chromatograms of O. canarii BRM-044600 liquid culture filtrate (a), the aqueous (b) and organic (c) fractions obtained therefrom after ethyl acetate extraction. A scheme of the fractionation procedure is provided. Extracted ion chromatograms (EIC) are shown for m/z 167.0861 (the main in- source fragment

93
of its parent ion, m/z 259.1299, chosen due to its higher intensity compared to the parent ion), m/z 257.1160, m/z 209.1165 and m/z 169.0751, with color codes specified in the figure inset. The chromatograms of the filtrate (a) and the aqueous fraction extracted therefrom (b) were obtained with chromatographic gradient 1, while the chromatogram of the organic fraction (c) was obtained with the chromatographic gradient 2, as described in Materials and Methods. All UHPLC-MS chromatograms were obtained in the positive ion mode. The base peak chromatograms (BPC) of all samples are available in Figures S2-S4 (Online Resource 1).
The ion m/z 257.1179 ([M + H]+) with predicted neutral molecular formula
C16H16O3 was also detected in O. canarii BRM-044600 culture filtrates as three main
chromatographic peaks –[email protected], [email protected] (major peak) and
[email protected] (Fig. 3a). These ions appear to be structurally related to the putative
strobilurin A due to similarities in their MS/MS pattern, as shown in Fig. 4a. Molecular
Networking analysis corroborated their structural similarity, since the m/z 257 and m/z
259 ions were grouped in the same ion family with a cosine-score of 0.54 (Fig. 4b).
Several product ions are shared between these compounds, while some product ions
show a difference of 2.014 Da compared to their counterparts from the related
compound. The 2.014 Da mass difference between these compounds corresponds to
two hydrogen atoms, and possibly an unsaturation, although we could not predict the
localization of the unsaturation in the molecule. The results suggest that the m/z 257
([M + H]+) ions may correspond to hitherto unknown strobilurins since, as far as we
know, no strobilurin with m/z 257 is described in the literature. Despite this, database
searches also suggested other putative annotations for the m/z 257 ions based on
MS1 data only (Table S1, Online Resource 1). Complete isolation and chemical
characterization are required to unequivocally identify those compounds.
In addition to the putative strobilurins, other ions such as [email protected],
[email protected], [email protected] and [email protected] were also detected by
UHPLC-MS in O. canarii BRM044600 supernatant but not in potato dextrose broth
(control), as observed by manual inspection and principal component analysis of
UHPLC-MS chromatograms (Figures S5 and S6, Online Resource 1). These
compounds likely correspond to metabolites secreted by O. canarii BRM-044600 and
could also be involved in the antifungal activity exerted by the culture filtrate. Tentative
annotations for each of these ions are presented in Table S1 (Online Resource 1).

94
Ethyl acetate fractionation of O. canarii BRM-044600 culture filtrate was carried
out as a tentative approach to isolate and identify the compound(s) with antifungal
activity. None of the putative strobilurins (m/z 259 and m/z 257 ions) were detected in
the aqueous fraction by UHPLC-MS analysis (Fig. 3b, chromatographic gradient 1).
However, they were detected with high intensity in the organic fraction (Fig. 3c,
chromatographic gradient 2). The chromatographic gradient 2 (developed for the
analysis of samples analysis diluted in organic solvents) could not separate the
different isomers of m/z 259 and m/z 257, such that each of those ions appear as a
single chromatographic peak in Fig. 3c. However, it does not mean that isomers were
lost during the ethyl acetate fractionation (as shown later in the text). In addition to the
putative strobilurins, the ions m/z 209.1175 and m/z 169.0751 were also substantially
(but not completely) extracted by ethyl acetate (Fig. 3a–c).
Fig. 4 a Comparative MS/MS spectra of [email protected] and [email protected] ions detected in O. canarii BRM-044600 liquid culture filtrate. The product ions of [email protected] highlighted in bold correspond to those previously reported for strobilurin A isolated from Strobilurus tenacellus by Schramm et al. (1978). b The molecular network obtained from UHPLC-MS/MS data of O. canarii BRM-044600 liquid culture filtrate. Nodes and edges are labeled with m/z values and cosine scores, respectively. The m/z delta and annotation of the edge between the m/z 259 and m/z 257 ions are also given. C Structure of strobilurin A.

95
The vacuum-dried organic fraction showed high antifungal activity against S.
sclerotiorum on PDA plates, either completely inhibiting its growth at 50% v/v
concentration or causing a delay in mycelial growth at 25% v/v (Fig. 5d–e). The
aqueous fraction, on the other hand, exhibited low antifungal activity, causing only a
short delay in sclerotia formation when used at 25% or 50% v/v concentrations (Fig.
5b–c). A correlation between the presence of putative strobilurins (a class of
compounds with well-known antifungal activity) and the higher antifungal activity in the
organic fraction enable us to hypothesize that these compounds may be largely
responsible for the observed antifungal activity in O. canarii BRM-044600. The ions
m/z 209.1175 and m/z 169.0751 may also contribute to antifungal activity.
Fig. 5 Growth of S. sclerotiorum on PDA plates containing the aqueous fraction (plates b and c at 25% (v/v) and 50% (v/v) concentrations, respectively) and the organic fractions (plates d and e at 25% (v/v) and 50% (v/v) concentrations, respectively) extracted from O. canarii BRM-044600 liquid culture filtrate. The control culture on PDA lacking O. canarii extracts is shown in plate (a). All photographs were taken 7 days after inoculation.

96
The organic fraction from O. canarii BRM-044600 liquid culture filtrate was
further fractionated with a C18 cartridge yielding three fractions—I, II and III—eluted
with 50%, 75% and 100% acetonitrile, respectively. All fractions were tested for
antifungal activity against S. sclerotiorum and analyzed by UHPLC-MS. Fractions I and
II delayed the S. sclerotiorum growth compared to fraction III. S. sclerotiorum mycelia
did not reach the plate edges after 144 h on PDA plates containing fraction I. Fraction
II prevented sclerotia formation after 144 h, while full colonization and sclerotia
formation was observed on PDA plates containing faction III (Fig. 6e–j). Thus, fractions
could be ranked as I > II > III according to their antifungal activities against S.
sclerotiorum.
UHPLC-MS analysis (using chromatographic gradient 1) revealed that
[email protected] was the major component in fraction I, followed by
[email protected], [email protected], [email protected] and [email protected] (Fig.
6b). Fraction II contained all three m/z 259 ions (putative strobilurin A and/or isomers)
as well as [email protected] (in lower intensity than in fraction I) and
[email protected] (Fig. 6c). Fraction III did not contain any of the putative strobilurins
(m/z 257 and m/z 259 ions) (Fig. 6d), which was expected, since it did not inhibit growth
or sclerotia formation in S. sclerotiorum.

97
Fig. 6 UHPLC-MS analysis of fractions derived from O. canarii BRM-044600 culture filtrate (a–d) and their antifungal activity against S. sclerotiorum at 48 h and 144 h (e–j). Color codes for the extracted ion chromatograms (EIC) are specified in the figure inset. The base peak chromatograms (BPC) of all samples are available in Figure S7 (Online Resource 1).
Even though our results cannot conclusively indicate the causative compounds
of antifungal activity (since compounds were not isolated), the lower inhibitory effect of
fraction II compared to fraction I (Fig. 6c, d) allow us to cautiously suggest that
[email protected], rather than the m/z 259 ions, is the main contributor to the
antifungal activity in the supernatant of O. canarii BRM-044600. It is Worth noting that
strobilurin A is reported to be photo-unstable (Balba 2007), as it might have diminished
the antifungal activity of the O. canarii BRM-044600 culture filtrate fractions throughout
this study, especially fraction II. Additionally, the antifungal activity was lower in
fractions I and II when compared to that of original culture filtrate and its ethyl acetate
extract, inferring that antifungal activity is a result of synergistic action of multiple
metabolites.
Discussion
Basidiomycetes are natural reservoirs of secondary metabolites with antifungal
activity, conferring them na advantage in competitive environments. Although many
basidiomycete species have been reported as able to inhibit phytopathogenic fungi
(Sivanandhan et al. 2017), these have seldom been tested against S. sclerotiorum, the
causal agent of white mold disease. The screening for antagonist fungi against
phytopathogens is usually performed by confrontation assays, followed by fungal
extracts chemical analysis or fractions thereof, aiming to isolate and identify antifungal
compounds (Szekeres et al. 2006; Schalchli et al. 2017). Here, this approach was
applied to screen for basidiomycete species capable of inhibiting S. sclerotiorum and
identify the metabolites responsible for antifungal activities.
In this study, O. canarii BRM-044600 and L. arvalis ATCC52088 were selected
among 17 tested basidiomycete isolates for their abilities to secrete bioactive
metabolites capable of inhibiting S. sclerotiorum growth. It has been previously
reported that O. canarii CCB179 exhibits antifungal activity against the phytopathogen
Cladosporium sphaerospermum (Rosa et al. 2005) and several pathogenic Candida

98
species (Rosa et al. 2003). Likewise, antifungal activity of L. arvalis isolates against
the phytopathogens Pythium ultimum and R. solani has also been documented (Allen
et al. 1985; Martin et al. 1986; Lewis and Papavizas 1992; Murdoch and Leach 1993;
Larkin and Brewer 2020). Nevertheless, as far as we know, this is the first report
demonstrating that these two basidiomycete species can inhibit S. sclerotiorum
mycelial growth.
O. canarii BRM-044600 liquid culture filtrate showed the highest antifungal
activity against S. sclerotiorum, whereas L. arvalis ATCC52088 showed the highest
pathogen mycelial growth inhibition (%MGI) in PDA paired cultures. These results
indicate that O. canarii BRM-044600 secretes antifungal compounds regardless of the
presence of the pathogen S. sclerotiorum. Thus, despite the potential for further
investigation of L. arvalis ATCC52088, this work focused on the bio-guided
fractionation and metabolomics analysis of the supernatant of O. canarii BRM-044600.
The results suggest that the observed antifungal activity is mainly caused by putative
strobilurins, including three compounds annotated as strobilurin A, or stereoisomers of
this compound, and three other putative strobilurins with m/z values not previously
reported in the literature. One of these putative strobilurins ([email protected]) might
be the main contributor to the antifungal activity. Although the antifungal compound
oudemansin A (structurally related to strobilurins) had been previously identified in the
extract of another isolate of O. canarii (CCB179) (Rosa et al. 2005), such metabolite
was not detected in the present study. Further purification and chemical
characterization (such as nuclear magnetic resonance) are required to confirm the
identity and structure of the antifungal compounds. Once bioactive metabolites are
isolated, further characterization of their biological activities, such as their minimum
inhibitory concentrations, can be carried out and the potential of O. canarii BRM-
044600 cultures can be better assessed.
As far as we know, this is the first time that strobilurins have been reported to in
O. canarii. Natural strobilurins and oudemansins (strobilurin congeners) have been
previously isolated from the extracts of other Oudemansielloid/Xeruloid species, such
as O. melanotricha (former Xerula melanotricha), Mucidula mucida (former O. mucida),
Hymenopellis radicata (former O. radicata), X. longipes and X. pudens (von Jagow et
al. 1974; Anke et al. 1990; Niego et al. 2021), as well as other basidiomycete genera
such as Strobilurus, Cyphellopsis, Mycena, Hydropus, Filoboletus, Favolaschia and

99
Crepidotus (Anke et al. 1977, 1983; Schramm et al. 1978; Anke 1995). Several of these
species are reported to produce up to three strobilurins/oudemansins (except
Favolaschia sp. 87129, which synthetizes six of these compounds) (Anke 1995). The
detection of six putative strobilurins in O. canarii BRM-044600 liquid culture filtrate
reported here indicates the high potential of this isolate as a source of
biotechnologically relevant compounds and its potential to protect crops against
phytopathogenic fungi.
Strobilurins and oudemansins were among the first antifungal antibiotics
isolated from basidiomyces in the 1970’s (Ypema and Gold 1999; Bartlett et al. 2002;
Balba 2007). Since then, many other antifungal compound classes have been reported
in basidiomycetes, including phenolics, terpenes, acylcyclopentenediones, sterols,
peptides, and antifungal proteins, showing diverse modes of action (Sivanandhan et
al. 2017). Due to the simple structure and high antifungal activity of strobilurins and
oudemansins, together with their low toxicity towards plant and mammalian cells, they
have been selected as leading compounds for further synthetic modifications and
development of commercial fungicides (Sauter et al. 1999).
As many natural strobilurins are photolabile, several derivatives displaying
higher photo-stability and fungicidal efficacy have been chemically synthesized and
commercialized since the 1990s by agrochemical companies such as Syngenta
(former Zeneca) and BASF (Ypema and Gold 1999; Bartlett et al. 2002; Balba 2007).
This present work indicates that, even though many natural strobilurins have been
discovered so far (e.g. strobilurin A, B, C etc., named in the order of their discovery),
this class of natural products likely contains many antifungal compounds yet to be
discovered (such as the ones detected here). Such compounds deserve attention, as
they might present properties distinct to those of natural and synthetic strobilurins
known to date, such as superior stability or antimicrobial activities. Furthermore, this
work indicates that Oudemansielloid/Xeruloid taxa, exemplified here by O. canarii
BRM-044600, remain an unexhausted source of strobilurins not previous reported.
In summary, this study identified O. canarii and L. arvalis as potential biocontrol
agents for S. sclerotiorum. O. canarii BRM-04460 is capable of secreting growth
inhibiting molecules even in the absence of S. sclerotiorum. The characterization of O.
canarii BRM-044600 supernatants demonstrated that this fungus is an untapped

100
source of multiple putative strobilurins capable of inhibiting S. sclerotiorum mycelial
growth and, therefore, a promising agent for white mold biocontrol.
References
Allen MF, Boosalis MG, Kerr ED, et al (1985) Population Dynamics of Sugar Beets,
Rhizoctonia solani, and Laetisaria arvalis: Responses of a Host, Plant Pathogen, and
Hyperparasite to Perturbation in the Field †. Applied and Environmental Microbiology
50:1123–1127. https://doi.org/10.1128/aem.50.5.1123-1127.1985
Anke T (1995) The antifungal strobilurins and their possible ecological role. Canadian
Journal of Botany 73(Suppl. 1):S940–S945. https://doi.org/https://doi.org/10.1139/b95-
342
Anke T, Besl H, Mocek U, Steglich W (1983) Antibiotics from basidiomycetes. xviii)
strobilurin c and oudemansin b, two new antifungal metabolites from xerula species
(agaricales). The Journal of Antibiotics 36:661–666.
https://doi.org/10.7164/antibiotics.36.661
Anke T, Oberwinkler F, Steglich W, Schramm G (1977) The strobilurins - new antifungal
antibiotics from the basidiomycete strobilurus tenacellus. The Journal of Antibiotics
30:806–810. https://doi.org/10.7164/antibiotics.30.806
Anke T, Werle A, Bross M, Steglich W (1990) Oudemansin X, A new antifungal E-B-
methoxyacrylate from oudemansiella radicata (Relhan ex Fr.) Sing. The Journal of
antibiotics XLIII:1010–1011
Balba H (2007) Review of strobilurin fungicide chemicals. Journal of Environmental Science
and Health - Part B 42:441–451. https://doi.org/10.1080/03601230701316465
Bartlett DW, Clough JM, Godwin JR, et al (2002) The strobilurin fungicides. Pest
Management Science 58:649–662. https://doi.org/10.1002/ps.520
Bolton MD, Thomma BPHJ, Nelson BD (2006) Sclerotinia sclerotiorum (Lib.) de Bary:
biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology
7:1–16. https://doi.org/10.1111/j.1364-3703.2005.00316.x
Carvalho DDC, Mello SCM, Júnior ML, Silva MC (2011) Controle de fusarium oxysporum
f.sp. phaseoli in vitro e em sementes, e promoção do crescimento inicial do feijoeiro

101
comum por Trichoderma harzianum. Tropical Plant Pathology 36:28–34.
https://doi.org/10.1590/S1982-56762011000100004
Chong J, Soufan O, Li C, et al (2018) MetaboAnalyst 4.0: Towards more transparent and
integrative metabolomics analysis. Nucleic Acids Research 46:W486–W494.
https://doi.org/10.1093/nar/gky310
Dutta S, Woo EE, Yu SM, et al (2019) Control of Anthracnose and Gray Mold in Pepper
Plants Using Culture Extract of White-Rot Fungus and Active Compound Schizostatin.
Mycobiology 47:87–96. https://doi.org/10.1080/12298093.2018.1551833
Fang XL, Li ZZ, Wang YH, Zhang X (2011) In vitro and in vivo antimicrobial activity of
Xenorhabdus bovienii YL002 against Phytophthora capsici and Botrytis cinerea.
Journal of Applied Microbiology 111:145–154. https://doi.org/10.1111/j.1365-
2672.2011.05033.x
Huan T, Forsberg EM, Rinehart D, et al (2017) Systems biology guided by XCMS Online
metabolomics. Nature Methods 14:461–462. https://doi.org/10.1038/nmeth.4260
Kamal MM, Savocchia S, Lindbeck KD, Ash GJ (2016) Biology and biocontrol of Sclerotinia
sclerotiorum (Lib.) de Bary in oilseed Brassicas. Australasian Plant Pathology 45:1–14
Karwehl S, Stadler M (2016) Exploitation of fungal biodiversity for discovery of novel
antibiotics. Current Topics in Microbiology and Immunology 398:303–338.
https://doi.org/10.1007/82_2016_496
Larkin RP, Brewer MT (2020) Effects of crop rotation and biocontrol amendments on
rhizoctonia disease of potato and soil microbial communities. Agriculture (Switzerland)
10:1–20. https://doi.org/10.3390/agriculture10040128
Lehner MS, Pethybridge SJ, Meyer MC, del Ponte EM (2017) Meta-analytic modelling of
the incidence-yield and incidence-sclerotial production relationships in soybean white
mould epidemics. Plant Pathology 66:460–468. https://doi.org/10.1111/ppa.12590
Lewis JA, Papavizas GC (1992) Potential of Laetisaria arvalis for the biocontrol of
Rhizoctonia solani. Soil Biol Biochem 24:1075–1079.
https://doi.org/doi.org/10.1016/0038-0717(92)90055-3
Liang X, Rollins JA (2018) Mechanisms of broad host range necrotrophic pathogenesis in
sclerotinia sclerotiorum. Phytopathology 108:1128–1140

102
Martin SB, Abawi GS, Hoch HC (1986) The relation of population densities of the
antagonist, Laetisaria arvalis, to seedling diseases of table beet incited by Pythium
ultimum. Canadian Journal of Microbiology 32:156–159. https://doi.org/10.1139/m86-
031
Murdoch CW, Leach SS (1993) Evaluation of Laetisaria arvalis as biological control agent
of Rhizoctonia solani on white potato. American Potato Journal 70:625–634
Nascimento M da S, Nascimento M da S, Silva CE da, et al (2019) Antagonismo in vitro de
Thielaviopsis paradoxa e Fusarium oxysporum por fungos rizosféricos associados à
cactáceas do semiárido alagoano e eficiência de duas técnicas de avaliação. In:
Impactos das Tecnologias nas Ciências Agrárias 3. Atena Editora, pp 77–85
Niego AG, Raspé O, Thongklang N, et al (2021) Taxonomy, diversity and cultivation of the
oudemansielloid/xeruloid taxa hymenopellis, mucidula, oudemansiella, and xerula with
respect to their bioactivities: A review. Journal of Fungi 7:1–23
Ogata H, Goto S, Sato K, et al (2000) KEGG: Kyoto Encyclopedia of Genes and Genomes.
Nucleic Acids Research 28:27–30. https://doi.org/10.1093/nar/28.1.27
Rosa LH, Cota BB, Machado KMG, et al (2005) Antifungal and other biological activities
from Oudemansiella canarii (Basidiomycota). World Journal of Microbiology and
Biotechnology 21:983–987. https://doi.org/10.1007/s11274-004-7553-7
Rosa LH, Machado KMG, Jacob CC, et al (2003) Screening of Brazilian Basidiomycetes
for Antimicrobial Activity. Memorias do Instituto Oswaldo Cruz 98:967–974.
https://doi.org/10.1590/s0074-02762003000700019
Ruttkies C, Schymanski EL, Wolf S, et al (2016) MetFrag relaunched: Incorporating
strategies beyond in silico fragmentation. Journal of Cheminformatics 8:1–16.
https://doi.org/10.1186/s13321-016-0115-9
Sandargo B, Chepkirui C, Cheng T, et al (2019) Biological and chemical diversity go hand
in hand: Basidiomycota as source of new pharmaceuticals and agrochemicals.
Biotechnology Advances 37
Sauter H, Steglich W, Anke T (1999) Strobilurins: Evolution of a New Class of Active
Substances. Angewandte Chemie International Edition 38:.

103
https://doi.org/10.1002/(SICI)1521-3773(19990517)38:10<1328::AID-
ANIE1328>3.0.CO;2-1
Schalchli H, Hormazábal E, Rubilar O, et al (2017) Production of ligninolytic enzymes and
some diffusible antifungal compounds by white-rot fungi using potato solid wastes as
the sole nutrient source. Journal of Applied Microbiology 123:886–895.
https://doi.org/10.1111/jam.13542
Schramm G, Steglich W, Anke T, Oberwinkler F (1978) Strobilurin A und B, antifungische
Stoffwechselprodukte aus Strobilurus tenacellus. Chemische Berichte 111:2779–
2784. https://doi.org/doi.org/10.1002/cber.19781110806
Sivanandhan S, Khusro A, Paulraj MG, et al (2017) Biocontrol properties of
basidiomycetes: An overview. Journal of Fungi 3
Smolińska U, Kowalska B (2018) Biological control of the soil-borne fungal pathogen
Sclerotinia sclerotiorum –– a review. Journal of Plant Pathology 100:1–12
Spiteller P (2015) Chemical ecology of fungi. Natural Product Reports 32:971–993.
https://doi.org/10.1039/c4np00166d
Stracquadanio C, Quiles JM, Meca G, Cacciola SO (2020) Antifungal Activity of Bioactive
Metabolites Produced by Trichoderma asperellum and Trichoderma atroviride in Liquid
Medium. Journal of Fungi 6:263. https://doi.org/10.3390/jof6040263
Szekeres A, Leitgeb B, Kredics L, et al (2006) A novel, image analysis-based method for
the evaluation of in vitro antagonism. Journal of Microbiological Methods 65:619–622.
https://doi.org/10.1016/j.mimet.2005.09.014
von Jagow G, Gribble GW, Trumpower BL (1974) Mucidin and Strobilurin A Are Identical
and Inhibit Electron Transfer in the Cytochrome bcl Complex of the Mitochondrial
Respiratory Chain at the Same Site as Myxothiazolt. Academic Press
Wang M, Carver JJ, Phelan VV, et al (2017) Sharing and community curation of mass
spectrometry data with GNPS. Nature Biotechnology 34:828–837.
https://doi.org/10.1038/nbt.3597.Sharing
Wang Z, Ma LY, Cao J, et al (2019) Recent Advances in Mechanisms of Plant Defense to
Sclerotinia sclerotiorum. Frontiers in Plant Science 10:1314

104
Xue J, Guijas C, Benton HP, et al (2020) METLIN MS2 molecular standards database: a
broad chemical and biological resource. Nature Methods 17:953–954.
https://doi.org/10.1038/s41592-020-0942-5
Ypema HL, Gold RE (1999) Kresoxim-methyl: modification of a naturally occurring
compound to produce a new fungicide. Plant Disease 83:4–19.

105
7 CONSIDERAÇÕES FINAIS
A formulação de substratos para produção de mudas de tomate (planta modelo)
a partir de substratos pós-colheita de cogumelos usados (SMS) ou substratos
colonizados/fermentados de A. bisporus suplementados com resíduos da
agroindústria de óleo de dendê apresentou resultados significativos, criando
oportunidade para realização de estudos de viabilidade econômica desse bioinsumo.
A implantação de um sistema integrado de coprodução de tomate e cogumelos
(P. ostreatus) em ambiente com sistema controlado demonstrou ser eficaz no controle
de fatores adversos e evidenciou uma produção satisfatória comparada a outros
estudos. Esse sistema, também, abre uma oportunidade de estudo de viabilidade
econômica a partir dos dados alcançados neste primeiro tipo de trabalho relatado, na
literatura recente.
Os fermentados de macromicetos de O. canarii BRM-044600 e L. arvalis
ATCC52088 foram capazes de controlar o crescimento de Sclerotinia sclerotiorum em
uma taxa representativa. O. canarii BRM-044600 demonstrou ser capaz de produzir
multiplas estrobilurinas putativas capazes de inibir o crescimento micelial de S.
sclerotiorum, apresentando assim um potencial para o desenvolvimento de biofungida
para controle do mofo branco. Esse é um estudo inédito no contexto de controle
alternativo para doenças das culturas de interesse comercial.
Os estudos realizados macromicetoss geradores de cogumelos (fungicultura
comercial) e seus resíduos (SMS), como também os fermentados de outros
macramicetos não-comerciais indicam a possibilidade de obtenção de bioprodutos e
bioinsumos no cenáriosde uma economia circular, onde ocorre a geração de renda e
a mínima geração de resíduos.

106
8 REFERENCIAS GERAL
ANPC, Associação dos Produtores de Cogumelos. O setor de cogumelos. 2019.
Disponível em: http://www.anpc.org.br/. Acesso em: 09/03/2019.
Beckers, S. J.; Dallo, I. A.; del Campo, I.; Rosenauer, C.; Klein, K.; Wurm, F. R. From
Compost to Colloids—Valorization of Spent Mushroom Substrate. ACS
Sustainable Chemistry. Engineering. 7, 7, 6991-6998, 2019.
Botelho, R. V.; bennemann, G. D.; Torres, Y. R.; Sato, A. F. Potential for Use of the
Residues of the Wine Industry in Human Nutrition and as Agricultural Input.
Grapes and Wines - Advances in Production, Processing, Analysis and
Valorization.doi:10.5772/intechopen.73132. 2018.
Carvalho, J. S. O.; Azevedo, R. V.; Ramos, A. P. S.; Braga, L. G. T. Agroindustrial
byproducts in diets for Nile tilapia juveniles. Revista Brasileira de Zootecnia,
v. 41, n. 3, 479-484, 2012.
FAOSTAT. Agricultural Data, 2017. Production. Crops. Disponível em:
http://faostat.fao.org/default.aspx/. Acesso em: 10/03/2019.
Garcia-Nunez, J. A. Evolution of Palm Oil Mills into Biorefineries, Washington State
University. Ph.D. dissertation. 2015.
Harfouche, A. et al. Forest biotechnology advances to support global bioeconomy.
Annals of Silvicultural Research, v. 38, n. 2, p. 42-50, 2015.
Hassan, A. A. K.; Abdel-Jubouri, H. A. Production, Purification and Characterization of
Phytase from Fruit Bodies of local isolate of the basidiomycetes fungus and test
its impact on Some biological Pleurotus ostreatus (11L) variables in Albino Rats.
Global Proceedings Repository, p. 613 - 648, 2018.
Ishak, Z. A. M.; Aminullah, A.; Ismail, H.; Rozman, H. D. Effect of silane-based coupling
agents and acrylic acid based compatibilizers on mechanical properties of oil
palm empty fruit bunch filled high-density polyethylene composites. Journal of
Applied Polymer Science, v. 68, n. 13, p. 2189-2203, 1998.
Kurnia, J. C., Jangam, S. V., Akhtar, S., Sasmito, A. P., & Mujumdar, A. S Advances
in biofuel production from oil palm and palm oil processing wastes: A review.
Biofuel Research Journal. V. 9, p. 332-346, 2016.

107
Morais, K. S.; Marquez, A. F.; Gomes, T. G.; Machado, A. E. V.; Lemos, M.; Oliveira,
M. E. C.; Martins Jr, H.; Mendonça, S.; Siqueira, F. G. Subprodutos da
agroindústria do óleo de palma (dendê) para cultivo de cogumelos comestíveis
e nutrição de peixes. Revista Biotecnologia & Ciência. v.6, n.1, p.25-54,
2017.
Nasrin, A. B.; Choo, Y. M.; Lim, W. S.; Joseph, L.; Michael, S.; Rohaya, M. H.; Astimar,
A. A.; Loh, S. K. Briquetting of empty fruit bunch fibre and palm shell as a
renewable energy fuel. Journal of Engineering and Applied Sciences, v. 6,
n. 6, p. 446-451, 2011.
Pal, A.; Khanum, F. Production and extraction optimization of xylanase from
Aspergillus niger DFR-5 through solid-state-fermentation. Bioresource
Technology, v. 101, n. 19, p. 7563-7569, 2010.
Shafigh, P.; Jumaat, M. Z.; Mahmud, H. Oil palm shell as a lightweight aggregate for
production high strength lightweight concrete. Construction and Building
Materials, v. 25, n. 4, p.1848-1853, 2011.
Silva, E. A. Pré-tratamento biológico da biomassa lignocelulósica de cachos de
Dendê (Elaeis sp.) com macromicetos: Cultivo, seleção e caracterização
através de ferramentas ômicas. Tese de doutorado – UFLA Universidade
federal de Lavras MG. 2019.
Sreekala, M. S.; Sabu, T. Utilization of short oil palm empty fruit bunch fiber (OPEFB)
as a reinforcement in phenol-formaldehyde resins: studies on mechanical
properties. Journal of Polymer Engineering, v.16, n. 4, p.265-294, 2011.
Sulaiman, F. et al. An outlook of Malaysian energy, oil palm industry and its utilization
of wastes as useful resources. Biomass and bioenergy, v. 35, n. 9, p. 3775-
3786, 2011.

108
9 ANEXO
Publicação do artigo 01.

109
Publicação do artigo 02.




















