
Unconscious Processing of Body Actions Primes Subsequent Action Perception but Not Motor Execution
23
Unconscious Processing of Body Actions Primes Subsequent Action Perception but Not Motor Execution Sonia Mele, Alan D. A. Mattiassi, and Cosimo Urgesi University of Udine and Scientific Institute IRCCS Eugenio Medea, San Vito al Tagliamento, Pordenone, Italy Previous studies have shown that viewing body actions primes not only the visual perception of congruent versus incongruent actions, but also their motor execution. Here, we used a masked-priming paradigm to explore whether visuoperceptual and visuomotor action priming may also occur when the prime is not consciously perceived. In 5 experiments, healthy individuals were presented with masked implied-action primes and were then prompted to perceive congruent or incongruent implied-action stimuli or to execute congruent or incongruent finger movements. Results showed that implied-action primes affected subsequent action perception also when they were not consciously perceived. Uncon- scious visuoperceptual action priming effects were independent from spatial compatibility and reflected genuine action representation. Conversely, masked implied-action primes affected motor preparation and execution processes only when they were consciously perceived. The results provide evidence of unconscious visuoperceptual but not visuomotor action priming effects, suggesting that unconscious processing of actions affects perceptual, but not motor representations. Keywords: automatic imitation, masked priming, motor resonance, stimulus response compatibility, unconscious processing Common coding hypotheses claim that perceived and executed actions are represented in a common code (Hommel, Müsseler, Aschersleben, & Prinz, 2001; Prinz, 1997; Schütz-Bosbach & Prinz, 2007), suggesting a bidirectional, causal link between per- ceptual and motor processing (Avenanti, Candidi, & Urgesi, 2013; Avenanti & Urgesi, 2011). In keeping with these hypotheses, several studies have shown that action execution induces short- term (Casile & Giese, 2006; Cattaneo & Barchiesi, 2011; Hamil- ton, Wolpert, Frith, & Grafton, 2006) and long-lasting (Abernethy, Zawi, & Jackson, 2008; Aglioti, Cesari, Romani, & Urgesi, 2008; Urgesi, Savonitto, Fabbro, & Aglioti, 2012) effects on the way others’ actions are perceived. On the other hand, there is also evidence that action perception influences the execution of one’s own motor actions. Indeed, observing a moving body parts facil- itates not only the subsequent perception of similar actions (visuo- perceptual priming; Costantini, Committeri & Galati, 2008; Gül- denpenning, Koester, Kunde, Matthias, & Schack, 2011), but also the execution of congruent as compared with incongruent body movements (visuomotor priming or automatic imitation; Brass, Bekkering, Wohlschläger, & Prinz, 2000; Heyes, 2011; Kilner, Paulignan, & Blakemore, 2003; Stürmer, Aschersleben, & Prinz, 2000). The visuomotor action priming effect is found when view- ing either dynamic action sequences (Brass et al., 2000; Brass, Bekkering, & Prinz, 2001; Kilner et al., 2003; Stürmer et al., 2000) or single frames that imply actions (Brass et al., 2000; Craighero, Bello, Fadiga, & Rizzolatti, 2002; Craighero, Fadiga, Rizzolatti, & Umilta `, 1999; Craighero, Fadiga, Umilta `, & Rizzolatti, 1996; Vogt, Taylor, & Hopkins, 2003). Furthermore, the effect is con- sidered automatic because it is independent from the observer’s intent to imitate or even to process the action stimulus (Heyes, 2011) and it is not modulated by the predictability of the prime-cue association (e.g., manipulating the proportion of congruent and incongruent trials; Hogeveen & Obhi, 2013). It is unknown, how- ever, whether action priming effects require perceptual awareness or whether they occur also in response to actions that are not consciously perceived. The present study was aimed at testing the effects of actions that are not consciously perceived on subsequent action perception and execution processes. A typical paradigm to test automatic imitation requires partici- pants to provide finger responses that are congruent or not with the observed finger movements (e.g., Brass et al., 2000). The re- sponses to congruent movements are faster than those to incon- gruent movements, suggesting the observer is automatically en- gaged in the simulation of the observed movement, although processing the action stimulus is not explicitly required by the task and is even detrimental for task performance (Brass et al., 2000). Further studies (Bertenthal, Longo, & Kosobud, 2006; Brass et al., 2001; Catmur & Heyes, 2011; Jiménez et al., 2012; Wiggett, Sonia Mele, Alan D. A. Mattiassi, and Cosimo Urgesi, Department of Human Sciences, University of Udine, Udine, Italy and Scientific Institute IRCCS Eugenio Medea, San Vito al Tagliamento, Pordenone, Italy. The first and last authors thank Prof. G. Berlucchi for his generous support during their initial research training and for insightful discussion on topics related to the present study. The research was supported by grants from Istituto Italiano di Tecnologia SEED 2009 [Prot. no. 21538; to Cosimo Urgesi], from the Ministero Istruzione Universita ` e Ricerca [Progetti di Ricerca di Interesse Nazionale, PRIN 2009; Prot. no. 2009A8FR3Z; Futuro In Ricerca, FIR 2012, Prot. N. RBFR12F0BD; to Cosimo Urgesi], and from Istituto di Ricovero e Cura a Carattere Scientifico “E. Medea” [Ricerca Corrente 2012, Ministero Italiano della Salute; to Cosimo Urgesi,]. The authors declare no competing financial interests. Correspondence concerning this article should be addressed to Sonia Mele, Department of Human Sciences, University of Udine, via Margreth, 3, I-33100, Udine, Italy. E-mail: [email protected] This document is copyrighted by the American Psychological Association or one of its allied publishers. This article is intended solely for the personal use of the individual user and is not to be disseminated broadly. Journal of Experimental Psychology: Human Perception and Performance © 2014 American Psychological Association 2014, Vol. 40, No. 2, 000 0096-1523/14/$12.00 DOI: 10.1037/a0036215 1 AQ: 1 AQ: au AQ: 2 AQ: 7 tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S1 3/7/14 18:10 Art: 2013-1385 APA NLM
Transcript of Unconscious Processing of Body Actions Primes Subsequent Action Perception but Not Motor Execution

Unconscious Processing of Body Actions Primes Subsequent ActionPerception but Not Motor Execution
Sonia Mele, Alan D. A. Mattiassi, and Cosimo UrgesiUniversity of Udine and Scientific Institute IRCCS Eugenio Medea, San Vito al Tagliamento, Pordenone, Italy
Previous studies have shown that viewing body actions primes not only the visual perception ofcongruent versus incongruent actions, but also their motor execution. Here, we used a masked-primingparadigm to explore whether visuoperceptual and visuomotor action priming may also occur when theprime is not consciously perceived. In 5 experiments, healthy individuals were presented with maskedimplied-action primes and were then prompted to perceive congruent or incongruent implied-actionstimuli or to execute congruent or incongruent finger movements. Results showed that implied-actionprimes affected subsequent action perception also when they were not consciously perceived. Uncon-scious visuoperceptual action priming effects were independent from spatial compatibility and reflectedgenuine action representation. Conversely, masked implied-action primes affected motor preparation andexecution processes only when they were consciously perceived. The results provide evidence ofunconscious visuoperceptual but not visuomotor action priming effects, suggesting that unconsciousprocessing of actions affects perceptual, but not motor representations.
Keywords: automatic imitation, masked priming, motor resonance, stimulus response compatibility,unconscious processing
Common coding hypotheses claim that perceived and executedactions are represented in a common code (Hommel, Müsseler,Aschersleben, & Prinz, 2001; Prinz, 1997; Schütz-Bosbach &Prinz, 2007), suggesting a bidirectional, causal link between per-ceptual and motor processing (Avenanti, Candidi, & Urgesi, 2013;Avenanti & Urgesi, 2011). In keeping with these hypotheses,several studies have shown that action execution induces short-term (Casile & Giese, 2006; Cattaneo & Barchiesi, 2011; Hamil-ton, Wolpert, Frith, & Grafton, 2006) and long-lasting (Abernethy,Zawi, & Jackson, 2008; Aglioti, Cesari, Romani, & Urgesi, 2008;Urgesi, Savonitto, Fabbro, & Aglioti, 2012) effects on the wayothers’ actions are perceived. On the other hand, there is alsoevidence that action perception influences the execution of one’sown motor actions. Indeed, observing a moving body parts facil-itates not only the subsequent perception of similar actions (visuo-
perceptual priming; Costantini, Committeri & Galati, 2008; Gül-denpenning, Koester, Kunde, Matthias, & Schack, 2011), but alsothe execution of congruent as compared with incongruent bodymovements (visuomotor priming or automatic imitation; Brass,Bekkering, Wohlschläger, & Prinz, 2000; Heyes, 2011; Kilner,Paulignan, & Blakemore, 2003; Stürmer, Aschersleben, & Prinz,2000). The visuomotor action priming effect is found when view-ing either dynamic action sequences (Brass et al., 2000; Brass,Bekkering, & Prinz, 2001; Kilner et al., 2003; Stürmer et al., 2000)or single frames that imply actions (Brass et al., 2000; Craighero,Bello, Fadiga, & Rizzolatti, 2002; Craighero, Fadiga, Rizzolatti, &Umilta, 1999; Craighero, Fadiga, Umilta, & Rizzolatti, 1996;Vogt, Taylor, & Hopkins, 2003). Furthermore, the effect is con-sidered automatic because it is independent from the observer’sintent to imitate or even to process the action stimulus (Heyes,2011) and it is not modulated by the predictability of the prime-cueassociation (e.g., manipulating the proportion of congruent andincongruent trials; Hogeveen & Obhi, 2013). It is unknown, how-ever, whether action priming effects require perceptual awarenessor whether they occur also in response to actions that are notconsciously perceived. The present study was aimed at testing theeffects of actions that are not consciously perceived on subsequentaction perception and execution processes.
A typical paradigm to test automatic imitation requires partici-pants to provide finger responses that are congruent or not with theobserved finger movements (e.g., Brass et al., 2000). The re-sponses to congruent movements are faster than those to incon-gruent movements, suggesting the observer is automatically en-gaged in the simulation of the observed movement, althoughprocessing the action stimulus is not explicitly required by the taskand is even detrimental for task performance (Brass et al., 2000).Further studies (Bertenthal, Longo, & Kosobud, 2006; Brass et al.,2001; Catmur & Heyes, 2011; Jiménez et al., 2012; Wiggett,
Sonia Mele, Alan D. A. Mattiassi, and Cosimo Urgesi, Department ofHuman Sciences, University of Udine, Udine, Italy and Scientific InstituteIRCCS Eugenio Medea, San Vito al Tagliamento, Pordenone, Italy.
The first and last authors thank Prof. G. Berlucchi for his generoussupport during their initial research training and for insightful discussionon topics related to the present study. The research was supported by grantsfrom Istituto Italiano di Tecnologia SEED 2009 [Prot. no. 21538; to CosimoUrgesi], from the Ministero Istruzione Universita e Ricerca [Progetti diRicerca di Interesse Nazionale, PRIN 2009; Prot. no. 2009A8FR3Z; FuturoIn Ricerca, FIR 2012, Prot. N. RBFR12F0BD; to Cosimo Urgesi], andfrom Istituto di Ricovero e Cura a Carattere Scientifico “E. Medea”[Ricerca Corrente 2012, Ministero Italiano della Salute; to CosimoUrgesi,]. The authors declare no competing financial interests.
Correspondence concerning this article should be addressed to SoniaMele, Department of Human Sciences, University of Udine, via Margreth,3, I-33100, Udine, Italy. E-mail: [email protected]
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
Journal of Experimental Psychology:Human Perception and Performance
© 2014 American Psychological Association
2014, Vol. 40, No. 2, 0000096-1523/14/$12.00 DOI: 10.1037/a0036215
1
AQ: 1
AQ: auAQ: 2
AQ: 7
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

Downing, & Tipper, 2013) demonstrated that automatic imitationis not related to mapping the spatial compatibility between the sideof observed and executed movements (Simon, Hinrichs, & Craft,1970), but reflects genuine processing of the action depicted intask-relevant or task-irrelevant visual stimuli; such perceptual ac-tion representation converges onto the motor representation of theexecuted response (Catmur & Heyes, 2011).
Specific neural underpinnings for the convergence between per-ceptual and motor processing have been suggested by the discov-ery of a class of neurons, known as mirror neurons, in the mon-key’s premotor cortex that respond during both execution andobservation of goal-directed actions (di Pellegrino, Fadiga, Fo-gassi, Gallese, & Rizzolatti, 1992; Gallese, Fadiga, Fogassi, &Rizzolatti, 1996; Rizzolatti, Fadiga, Gallese, & Fogassi, 1996).The same mechanism may exist in the human fronto-parietalcortex (Cattaneo & Rizzolatti, 2009; Mukamel, Ekstrom, Kaplan,Iacoboni, & Fried, 2011), where largely overlapping activationshave been reported during action observation and execution (Fo-gassi & Ferrari, 2011; Rizzolatti & Craighero, 2004; Rizzolatti &Sinigaglia, 2010). For example, the observation of actions withdifferent effectors (e.g., mouth, hand, foot) activates differentregions of the premotor and parietal cortices, in accordance withthe classic motor homunculus (Buccino et al., 2001). In a similarvein, using single-pulse transcranial magnetic stimulation appliedover the primary motor cortex, it has been shown that actionobservation triggers a strictly congruent, muscle-specific facilita-tion of corticospinal excitability (Fadiga, Craighero, & Olivier,2005). In keeping with studies that manipulate the conditionsunder which automatic imitation effects occur (Heyes, 2011), thefacilitation of corticospinal excitability in response to observedactions takes place: a) independently of explicit instructions toimitate or rehearse the actions (Fadiga, Fogassi, Pavesi, & Rizzo-latti, 1995), thus being automatic; b) during passive viewing ofstatic images that imply body actions (Avenanti et al., 2013;Urgesi et al., 2010; Urgesi, Moro, Candidi, & Aglioti, 2006); c)independently of the spatial compatibility between the observedand the executed movements (Alaerts, Aggelpoel, Swinnen, &Wenderoth, 2009; Urgesi, Candidi, Fabbro, Romani, & Aglioti,2006).
Direct evidence of shared neural representations of observedand executed actions comes from recent neuroimaging studies thathave documented the adaptation of the neural activity in theinferior parietal cortex (Chong, Cunnington, Williams, Kanwisher,& Mattingley, 2008) and in the inferior frontal gyrus (Kilner, Neal,Weiskopf, Friston, & Frith, 2009) after repeated observations aswell as consecutive observation and execution (or vice versa) ofthe same action. Thus, previously seen actions affect not only theperceptual representation of observed actions, but also the motorrepresentation of executed actions. The overlap between the neuralpopulations involved in unimodal visuoperceptual and cross-modal visuomotor adaptation effects is in keeping with the pre-dictions of common-coding hypotheses, which point to a commonmechanism underlying visuoperceptual (perceptual) and visuomo-tor (i.e., automatic imitation) action priming. It is unclear, how-ever, whether the two behavioral effects are influenced in the sameway by the manipulation of the spatial compatibility between theprime stimulus and subsequent observed or executed actions andby the reduction of the visibility of the prime stimulus.
It is widely known that a stimulus can influence (prime) subse-quent perception and motor response even when it is not con-sciously perceived (Sumner, 2007). Varying the stimulus visibilityhas been widely used to manipulate perceptual awareness (Vanden Bussche, Van den Noortgate, & Reynvoet, 2009). For exam-ple, in the masked priming paradigm a prime stimulus is shortlypresented and its visibility is further reduced by presenting a mask(typically a scrambled version of the same stimulus) before and/orafter its presentation (respectively, forward and backward mask-ing). After a very short interval from this “sandwich” maskedprime presentation, the target stimulus is presented; results revealbetter processing of the target when it is related to the prime withrespect to a neutral condition in which the prime and the target arenot related (Sumner, 2007). Such facilitation reflects memoryprocesses that keep track of past perceptual experiences facilitatingthe perception of familiar attributes (Tulving & Schacter, 1990).The priming effect occurs at many different levels of informationprocessing, depending on the features of stimuli and tasks (seeKristjánsson & Campana, 2010 for a review), and may regard boththe perceptual similarity between the prime and the target or theirassociation with the same motor response. The effects related tothe perceptual attributes of the stimuli depend on the perceptualrepresentation system and seem to be driven in a bottom-upmanner (Campana, Cowey, & Walsh, 2002; Silvanto & Soto,2012; Tulving & Schacter, 1990). On the other hand, the effectsrelated to the association of the prime and target to the same motorresponse seem to depend on motor response selection processes(Eimer & Schlaghecken, 1998; Eimer & Schlaghecken, 2003).
In the present study, we used a masked priming paradigm toinvestigate how the perceptual and motor representations of ac-tions are affected by reducing the prime visibility and by reversingthe spatial and action compatibility between the prime and theprobe. We presented masked prime stimuli depicting a still hand ora finger-abduction implied action in conditions of low or highvisibility; the prime was followed by the presentation of a still orimplied-action probe hand. We first tested the effects of low andhigh visibility action primes on the detection of implied actions(Experiment 1) and on the discrimination of specific finger move-ments (Experiment 2) in the probe hand. Thus, we tested whethervisuoperceptual action priming effects occur independently ofperceptual awareness. As the perception of visual attributes, suchas color and shape, facilitates the processing of target stimuli thatshare the same attributes (Campana et al., 2002; Silvanto & Soto,2012; Tulving & Schacter, 1990), we expected that also implied-action primes should affect the processing of implied-actionprobes in conditions of low visibility. Furthermore, the actionpriming effects should be independent of the spatial compatibilitybetween the side of the space occupied by the prime and probefinger movements. Therefore, in Experiment 2 we tested the ef-fects of spatial compatibility on visuoperceptual action priming bypresenting the prime and probe hands with the fingers pointingupward or downward. This way, we opposed the correspondencebetween the moving finger and the space occupied by the move-ments in the prime and probe hand stimuli, disentangling the roleof action congruence and spatial compatibility in both low andhigh prime visibility conditions.
The main hypothesis of this study was derived from common-coding view of action observation and action execution (Hommelet al., 2001; Prinz, 1997). According to this hypothesis, a common
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
2 MELE, MATTIASSI, AND URGESI
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

representation is involved in the influence of action stimuli onsubsequent action perception and execution. Thus, if an impliedaction hand presented in conditions of low-visibility influencessubsequent action perception (visuoperceptual priming), it shouldhave the same effect on action execution (visuomotor priming). InExperiment 3, we used a forward and backward dynamic maskingprocedure and successfully prevented the conscious perception ofprime stimuli depicting a still hand or index or little finger abduc-tions. The masked prime presentation was followed by the presen-tation of a still or implied-action probe hand, to which participantsresponded performing congruent or incongruent finger movementsto indicate whether the probe hand implied or not a finger move-ment. We tested whether the reduction of prime visibility affectedin the same way its effects on the perception of the probe hand(visuoperceptual priming) and on the preparation of manual re-sponses that were congruent or incongruent with the observedfinger movements (visuomotor priming). Finally, in Experiment 4and 5 we tested the masked visuomotor action priming effectswhile participants executed index or little finger movements inresponse to neutral probes (colored squares) preceded by a primehand implying a congruent or incongruent movement. Therefore,this paradigm allowed us to test the modulation of reducing thevisibility of the prime on standard automatic imitation effects.
Overall, the results of the five experiments provided evidencethat action primes affected subsequent action perception also whenthey are not consciously perceived by the observer. This visuoper-ceptual action priming effect was independent from the spatialcompatibility between the moving finger in the two perceptualstimuli and it rather reflected genuine action-specific coding. Con-versely, action primes presented in conditions of low visibility didnot affect motor preparation and execution processes, becausecongruent or incongruent movements were executed the same way.A clear visuomotor effect was obtained only when the primingactions were presented in conditions of high visibility, thus sug-gesting that visuomotor action priming requires perceptual aware-ness.
Experiment 1
In Experiment 1, we tested whether the masked presentationof a static frame implying hand action influences the processingof the action implied by a consciously perceived probe hand.We also tested whether such perceptual influence is dependenton the specific processing participants are engaged with duringprobe observation. We thus compared the visuoperceptual prim-ing effects when participants were required to process theaction implied by the probe hand (action task) or its morphol-ogy (form task). The prime and probe stimuli could depict astatic hand or an implied-action hand. Furthermore, the primewas presented in conditions of low (17 ms duration) and high(50 ms duration) visibility. This way, we aimed to provideevidence of modulation of unconscious action processing on thevisual representation of subsequently presented actions. Weexpected faster response for congruent than incongruent probe-prime pairs, independently of the visibility of the prime; theeffect of action congruence, however, should be specific for theprocessing of body actions but not of body forms.
Method
Participants. Twenty-three right-handed students (10 men) ofthe University of Udine (mean age � 23.47 years, standard devi-ation � 2.93 years) participated in the experiment and gave theirwritten informed consent. All were native Italian speakers ofCaucasian ethnicity (as the models depicted in the stimuli) andreported having normal or corrected-to-normal visual acuity. Allparticipants were right-handed according to a standard handednessinterview (Briggs & Nebes, 1975). Participants were unaware ofthe purposes of the study and were debriefed at the end of theexperimental session. The procedures were approved by the EthicsCommittee of the Scientific Institute (IRCCS) Eugenio Medea(Bosisio Parini, Como, Italy) and were in accordance with theguidelines of the Declaration of Helsinki.
Stimuli. Stimuli were color pictures of the right hand of 8models (4 men) taken at rest or during maximal abduction of theindex finger. The pictures were taken with a Sony digital camerawith a resolution of 10.1 megapixels at a distance of 30 cm in frontof the hand. Lighting conditions were kept stable during picturetaking. The pictures were acquired in rapid sequence (multishotmode) to capture different phases of real index finger abductionmovements. The pictures were presented on a 60-Hz LCD screen(resolution 1024 � 768 pixels) and subtended a 9.5° � 12.7°region.
Procedure. Participants were administered two tasks, namelyan action task and a form task. In both tasks the following maskedpriming paradigm was used (Figure 1A): a picture of a still hand(sample) appeared on the screen for 250 ms and was followed bya scrambled version of the prime (forward mask) for 83 ms. Then,the prime hand appeared for 17 ms or 50 ms and was subsequentlymasked (backward mask); the duration of the backward maskcould be 100 ms or 67 ms and depended on the duration of theprime, so that the sum of prime and backward mask duration wasalways 117 ms. Lastly, a third hand (probe) was presented for 250ms and was followed by a blank screen. The prime and the probestimuli could depict a still hand (still) or an abduction of the indexfinger (implied-action). In the action task, the sample, the prime,and the probe depicted always the same model hand that could orcould not imply index finger abduction. In the form task, thesample and the probe could depict the hand of the same individualor of two different individuals of the same gender; the impliedaction was manipulated orthogonally to the identity of the model.Four 64-item blocks were presented, with alternating administra-tion of the action and form tasks. In blocks 1 and 2, the prime waspresented for 17 ms, thus below the threshold of conscious per-ception; in blocks 3 and 4, the prime was presented for 50 ms, thusabove the threshold of conscious perception (Sandberg, Martin,Timmermans, Cleeremans, & Overgaard, 2011). Subjectively, thepresentation of the sequence of forward mask, prime, and back-ward mask was experienced by the participants as a flickering ofthe mask. Participants were instructed to ignore the flickering ofthe mask and to focus on the sample and probe hands. They had toreport as fast and accurately as possible whether they perceived ornot a change in the visual appearance of the probe hand withrespect to the sample hand and, specifically a) an index fingermovement in the probe hand during the action task, or b) adifference in the morphology of the hand in the form task. The twotasks were administered to compare visuoperceptual action prim-
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
3UNCONSCIOUS PROCESSING AND ACTION PRIMING
F1
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

ing effects when the participants were explicitly asked to focus onaction or form cues (Urgesi, Candidi, Ionta, & Aglioti, 2007).Participants provided their responses by pressing on a two-keyresponse box with the right index and middle fingers (Figure 1B),
thus performing flexion movements different from those shownby the pictures. The order of the tasks and the assignment ofresponse types to the two keys were counterbalanced across par-ticipants, thus also controlling for the correspondence between the
Figure 1. Method and results for Experiment 1. A) Trial structure: participants were presented with a sequenceof three hands, namely the sample, always a still hand, and the prime and the probe that could depict either astill hand or an index finger abduction. The prime hand was presented in conditions of low (17 ms) and highvisibility (50 ms). In both the action and the form task, participants were instructed to report whether or not theyperceived a change of the visual appearance of the probe hand with respect to the sample hand and, specifically,a) an index finger movement in the probe hand during the action task, b) a difference in the morphology of thehand in the form task. B) Response configuration: participants provided their response by pressing on a two-keyresponse box with the index or middle finger. C) Results showed that the responses in the action, but not in theform task were facilitated for congruent versus incongruent prime-probe pairs when the prime was presented inconditions of both low and high visibility. Asterisks indicate significant comparisons at p � .05.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
4 MELE, MATTIASSI, AND URGESI
AQ: 8
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

finger abducted in the pictures and the finger used to report thepresence of implied-action probes. The whole procedure lastedabout 30 minutes. At the end of the experimental session partici-pants were requested to retrospectively report on the subjectiveperception of the prime for each block, thus allowing us to estab-lish whether the participants noticed the presentation of the prime.All participants reported that they did not see the prime in the17-ms prime duration blocks and saw it in the 50-ms ones.
Data analysis. We calculated the individual Error Rates (ERs)and mean Reaction Times (RTs) for the correct responses of eachexperimental condition (16 trials per cell). Three participants hadpoor performance levels (ERs � than 40%) in the action task,pointing to lack of task compliance, and were excluded from theanalysis; another two participants were excluded because they hadmore than 5% of anticipations (RTs � 120 ms). The final sampleentered in the analysis thus consisted of 18 participants (half withthe association of “moving” responses to index finger presses and“still” responses to middle finger presses and half with the oppo-site association). ERs and RTs data were entered into two four-waymixed-model Analyses of Variance (ANOVAs). The study used a2 � 2 � 2 design with Key as between-subjects factor and withthree repeated-measures variables: Task (action vs. form), Primeduration (low vs. high visibility), and Congruency (congruent vs.incongruent prime-probe pairs). All pairwise comparisons wereperformed using the Newman–Keuls post hoc sequential test,which reduces the size of the critical difference depending on thenumber of steps separating the ordered means; this procedure isoptimal for testing in the same design effects that may havedifferent sizes (Duncan, 1955; Dunnett, 1970; McHugh, 2011), asit was expected in our cases for low-visibility versus high-visibilityconditions. A significance threshold of p � .05 was set for allstatistical analyses. Effect sizes were estimated using the partial etasquared measure (�p
2). The data are reported as mean � standarderror of the mean (SEM).
Results
Reaction times. The ANOVA on RTs (Figure 1C) revealedsignificant main effect of Congruency (F1,16 � 30.785, p � .001,�p
2 � 0.658): in general, RTs for congruent trials (518.59 � 16.30ms) were faster than RTs for incongruent trials (545.81 � 17.60ms). The main effect of the between-subjects factor Key (F1,16 �1) and its two-way (all F1,16 � 3.3, p � .089, �p
2 � 0.4) andhigher-order (all F1,16 � 1.7, p � .214, �p
2 � 0.1) interactions withthe other variables were not significant. The two-way interactionsPrime duration � Congruency (F1,16 � 17.651, p � .006, �p
2 �0.524) and Task � Congruency (F1,16 � 9.493, p � .007, �p
2 �0.372) were significant and were further qualified by a significantthree-way interaction Prime duration � Task � Congruency(F1,16 � 6.072, p � .025, �p
2 � 0.275). To explore the significantthree-way interaction we ran two follow-up Prime duration �Congruency ANOVAs, one for each task. The ANOVA for theaction task showed nonsignificant main effect of Prime duration(F1,17 � 3.288, p � .087, �p
2 � 0.162), but significant main effectof Congruency (F1,17 � 63.895, p � .001, �p
2 � 0.79) andinteraction Prime duration � Congruency (F1,17 � 52.478, p �.001, �p
2 � 0.755). Post hoc comparisons indicated that RTs forcongruent trials were faster than RTs for incongruent trials in bothlow- (p � .01) and high-visibility (p � .001) conditions. Thus, in
both visibility conditions the presentation of congruent prime-probe pairs facilitated the responses as compared with incongruentpairs. However, the RTs for the congruent trials of the high-visibility condition were faster than those for the congruent trialsof the low-visibility condition (p � .001), whereas RTs for incon-gruent trials did not differ between the two visibility conditions(p � .269). This result points to a stronger facilitation effect for thehigh- than low-visibility primes. The ANOVA on RTs in the formtask revealed no main effects or interactions (all F1,17 � 2.25, p �.152; �p
2 � 0.117), suggesting that the congruency between theprime and probe actions did not affect the performance of partic-ipants when they had to focus on the morphology of the hand.
Error rates. Although ERs, in particular in the action task,were relatively low (see Table 1), we tested whether the samepattern obtained for RTs was found for ERs. A two-way Primeduration � Congruency ANOVA was thus run on the ERs for eachtask. The ANOVA on ERs in the action task revealed a significantmain effect of Congruency (F1,17 � 24.501, p � .001, �p
2 � 0.59),further qualified by a significant two-way interaction (F1,17 �11.713, p � .003, �p
2 � 0.408). Post hoc analyses indicated that, inthe low-visibility condition, participants committed a comparableproportion of errors for congruent (7.66 � 2.05%) and incongruenttrials (12.44 � 2.41%, p � .001); in contrast, in the high-visibilitycondition, they committed fewer errors in the congruent (3.61 �0.93%) than incongruent trials (12.44 � 2.41%, p � .001). Thus,the analysis of ERs revealed that subliminal action primes did notaffect the accuracy of participants in responding to congruentversus incongruent prime-probe pairs. In contrast, the performanceaccuracy was affected when the prime was presented in conditionsof high-visibility. The ANOVA on ERs in the form task revealedonly a significant main effect of Prime duration (F1,17 � 22.75,p � .001, �p
2 � 0.572), with better performance for the high-thanlow-visibility condition.
Discussion
The results of Experiment 1 showed that action primes canaffect action perception not only when they are consciously per-ceived (high-visibility condition) but also when their presentationis completely unnoticed (low-visibility condition). This result canbe interpreted at the perceptual level, because the action primefacilitated the perceptual processing of same stimulus probes (bothstatic hands or both implied action hands) and is in line with theperceptual priming literature (Sumner, 2007; Tulving & Schacter,1990). Importantly, however, the action priming effects were onlyobtained, independently of prime visibility, on the processing ofbody actions but not on the processing of body forms. The disso-ciation between the two tasks corroborates the action-specificity ofthe effects and is in keeping with the notion that the perceptualpriming depends on the specific features of the stimuli and task(Kristjánsson & Campana, 2010).
It is worth noting that participants responded with the index andmiddle fingers, with random allocation of the key to the tworesponses; therefore, half of them responded to the observation ofan index finger movement by moving the same finger. However,reversing the response-key association had no effects on the prim-ing effects, probably because of the difference between the fingermovements that were observed (abduction) and executed (fingerflexion), which triggered the access to different action representa-
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
5UNCONSCIOUS PROCESSING AND ACTION PRIMING
T1
AQ: 3
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

tions. This is in keeping with the findings that reducing theideomotor compatibility between observed and executed actionshinders the automatic imitation effects (Brass et al., 2000).
In sum, Experiment 1 provided evidence of perceptual primingeffects that were specific for the processing of actions and inde-pendent of the perceptual awareness of the participants and of thetype of motor response being prepared. Three possible limitations,however, need to be addressed. First, congruent finger movementsalso occurred in the same portion of the space; therefore, spatialcompatibility could explain the results. Second, because we usedonly one finger movement, we could not establish whether theprime representation contained general dynamic information (i.e.,something is moving), which facilitated the detection of move-
ments in subsequent stimuli, or whether it contained specificinformation on the moving finger. Finally, because we did nothave a neutral condition, we could not determine whether thepriming effects were related to facilitation for congruent stimuli orinterference for incongruent stimuli or both. Experiment 2 wasaimed at addressing these issues.
Experiment 2
In Experiment 2, we tested whether visuoperceptual primingeffects could be ascribed solely to action congruence or whethersome contribution of the spatial compatibility between the locationoccupied by the moving finger in the prime and the probe should
Table 1Error Rates (Mean % � Standard Deviation) for Each Condition of the Five Experiments
Experiment 1
Action task:Prime visibility
Form task:Prime visibility
Action congruence Low High Low High
Congruent 7.66 � 4.52 3.61 � 1.63 33.38 � 2.69 20.61 � 12.59Incongruent 7.66 � 3.29 12.44 � 7.32 34.83 � 2.79 21.61 � 1.64
Experiment 2
Perspective
Congruent:Prime visibility
Incongruent:Prime visibility
Action congruence Low High Low High
Congruent 4 � 4.41 2 � 2.35 4 � 3.99 4 � 4.03Incongruent 4 � 2.73 7 � 4.86 4 � 3.97 4 � 4.31Neutral 3 � 3.19 3 � 2.21 4 � 2.52 4 � 4.65
Experiment 3
Effector
Index finger: Probe Little finger: Probe
Prime Static Index finger Little finger Static Index finger Little finger
Static 1 � 2.46 2 � 3.25 8 � 12.02 2 � 3.34 4 � 4.86 1 � 2.69Index finger 4 � 4.67 2 � 4.48 4 � 6.93 5 � 6.90 4 � 7.30 0.1 � 1.74Little finger 7 � 7.76 2 � 3.80 4 � 6.33 4 � 6.19 5 � 6.98 2 � 5.04
Experiment 4
Perspective
Congruent:Prime visibility
Incongruent:Prime visibility
Action congruence Low High Low High
Congruent 0.7 � 1.65 0.7 � 1.33 0.7 � 1.65 0.7 � 1.33Incongruent 0.6 � 1.56 0.9 � 1.97 0.7 � 1.33 0.7 � 1.65
Experiment 5
SOA
Short: Prime visibility Long: Prime visibility
Action congruence Low Periliminal High Low Periliminal High
Congruent 4.2 � 3.2 2.35 � 2.35 2.1 � 2.2 3 � 3.16 2.2 � 2.56 2.9 � 2.2Incongruent 2.4 � 2.01 4 � 3.55 3 � 2.6 2.5 � 2.4 2.7 � 2.1 2.8 � 2.1
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
6 MELE, MATTIASSI, AND URGESI
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

be considered. To this end, we changed the orientation of the handto independently manipulate the compatibility between the identityof the moving finger and its spatial position in the prime and in theprobe stimuli. The prime and the probe hands could be presentedfrom the same perspective or different perspectives (with thefinger pointing upward or downward; Figure 2A), but they alwaysdepicted a right hand, thus ensuring that the two views recruitedthe same hand motor representation (Shmuelof & Zohary, 2006,2008; Vainio & Mustonen, 2011). Additionally, we presented handpictures depicting an index or a little finger abduction (Figure 2B).We used upward and downward hands because the abduction ofthe index finger occurs in the left hemifield for the upward per-spective and in the right hemifield for the downward perspective;the opposite holds true for the little finger abductions. We askedparticipants to report the moving finger by pressing one of twokeys with the left or right thumbs (Figure 2C). Therefore, partic-ipants used different fingers than those that were shown abductedin the stimuli; however, to control for any spatial compatibilityeffects between the side of the moving finger in the probe and theside of response, we counterbalanced between-participants theresponse-key association. In sum, the design was aimed at testingthe relative effects of action and spatial compatibility between theprime and probe hands. Furthermore, using two finger movements,we were able to test whether the priming representations containedspecific information on the action being performed (index vs. littlefinger movements) rather than a general representation of a dy-namic stimulus (i.e., moving vs. not moving hand). Finally,whereas the probe could depict only a hand with the index or littlefinger abducted, the prime could show a hand with congruent orincongruent finger abduction or a still hand, thus allowing us totest both facilitation and interference priming effects with respectto the neutral, still-hand condition. We expected that viewing animplied-action prime should facilitate the discrimination of con-gruent probes and interfere with the discrimination of incongruentones. Furthermore, both facilitation and interference effects shouldoccur irrespective of the visibility of the prime (as in Experiment1) and the spatial compatibility between the side of the spaceoccupied by the two fingers (as in Catmur & Heyes, 2011).
Method
Participants. Twenty-eight students (14 females) of the Uni-versity of Verona (mean age � 26.78 years) participated in theexperiment. All were native Italian speakers of Caucasian ethnic-ity, right-handed, and reported having normal or corrected-to-normal visual acuity. No participants took part in the previousexperiment and recruitment procedures were as in the previousexperiment.
Stimuli. We used the digital pictures depicting the right handof a new group of 8 models (4 females, aged 20–23 years). Themodels’ hands were pictured in three positions: a) with maximumindex finger abduction; b) with maximum little finger abduction;and c) still. To preserve the appearance of naturalistic movement,each picture was taken while the model was actually moving thefinger. The same pictures were used in the two perspectives andupward and downward stimuli were obtained by vertically flippingthe pictures (Figure 2A). Light conditions were kept constantacross the three images of each model and the remaining lumi-nance differences were manually corrected using Corel Paint Shop
Pro X (Corel Inc., Mountain View, CA). Stimuli were presentedwith a 60-Hz CRT monitor (resolution 1024 � 768 pixels) andsubtended a 6° � 8.5° region.
Procedure. Participants were seated at a distance of 57 cmfrom the computer screen while their head was restrained bymeans of a head-and-chin rest. As schematized in Figure 2B, thegeneral procedure involved the presentation of a central luminousdot serving as fixation point for 250 ms; then a picture of a stillhand (sample) appeared on the screen for 250 ms and was followedby the mask consisting of the scrambled version of the prime for100 ms (forward mask); then the prime hand, which could be stillor with the index or little finger abducted, appeared for 17 ms or50 ms and was subsequently masked for 130 ms (backward mask);then, the probe hand, which could show either index or little fingerabduction, was presented for 250 ms, to which a blank screenfollowed and remained on the screen until response or for amaximum of 1,500 ms. Responses were recorded when providedwithin the 1,500-ms period after probe offset. The sample handwas always in the same perspective as the probe hand, whereas theprime hand could be in the same or different upward or downwardperspective with respect to the probe. The maintenance of fixationduring the trial was monitored via an infrared TV camera (ASLEye-Trac, Applied Science Laboratory, Bedford, MA). Partici-pants were asked to identify the finger that was abducted in theprobe hand by pressing one of two keys of a computer keyboardwith the right or left thumbs (Figure 2C). Half of the participantsused the left thumb to report an index finger movement probe andthe right thumb to report a little finger movement; the oppositeresponse-key association was used for the remaining participants.In total we had 768 trials, 384 trials for each prime duration block;in each block, there were 32 repetitions for each of the 12 possiblecombinations between the three prime types, the two probe types,and the two perspectives. The different prime-probe combinationswere randomly presented in each block, whereas the order ofprime-duration blocks was fixed presenting first the low-visibilityblock (17 ms) and then the high-visibility block (50 ms). At thebeginning of each block we administered 16 practice trials. Fur-thermore, at the end of the experimental session, participants wererequested to retrospectively report on the subjective perception ofthe prime in each block; this allowed us to establish whether theparticipants noticed the presentation of the prime. All participantsreported to have not seen the prime in the 17-ms prime durationblock and to have seen it in the 50-ms one.
Data analysis. One subject had two anticipations (RTs � 120ms), and these trials were excluded from the mean calculation ofthe corresponding conditions. ERs were low for all participants.ERs and mean RTs for correct responses were used for the anal-ysis. Preliminary analyses showed that there were no overalldifferences between the trials with upward and downward hands.Thus, data were coded on the basis of the congruence between themovement of the prime and of the probe (congruent index or littlefinger movements; incongruent index or little finger movements;and neutral static primes) and between the perspective of the primeand of the probe (same vs. different upward or downward handdirections). Data were entered into three-way repeated-measuresANOVAs. The study used a 2 � 2 � 3 design with three repeated-measures variables: Prime duration (low-, high-visibility), Per-spective congruence (same or different), and Action congruence(congruent, incongruent and neutral). All pairwise comparisons
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
7UNCONSCIOUS PROCESSING AND ACTION PRIMING
F2
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

Figure 2. Method and results for Experiment 2. A) Examples of the experimental stimuli and factorialmanipulation of the congruence between the prime and probe actions and their spatial perspective. The probe wasalways an implied-action hand with index or little finger abducted, whereas the prime could depict a congruentor incongruent implied-action hand or a still hand. Both the prime and the probe could be depicted with thefingers pointing upward or downward. B) Trial structure: participants viewed a sequence of three hands and wereasked to identify the finger that was abducted in the probe. C) Response configuration: the responses wereprovided by pressing one of two buttons of a computer keyboard with the right or left thumbs. D) Results showedthat, in conditions of low visibility, the presentation of congruent prime-probe pairs facilitated the responses ascompared to both incongruent and neutral pairs, independently of spatial perspective. Conversely, in conditionsof high visibility, both facilitation for congruent and interference for incongruent prime-probe pairs wereobtained as compared to the neutral, still prime trials; however, these effects were dependent on compatiblespatial perspective and no effects were obtained when the prime and probe were viewed from differentperspectives. Asterisks indicate significant comparisons between action congruence conditions at p � .05. Pleasenote that the comparisons highlighted in the low-visibility condition (left panel) refer to the testing of the maineffect of action congruence, collapsing the same and different perspective trials; conversely for the high visibilitycondition (right panel) the comparisons between action congruence conditions refer to only same perspectivetrials.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
8 MELE, MATTIASSI, AND URGESI
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

were performed using the Newman–Keuls post hoc test. Asignificance threshold of p � .05 was set for all statisticalanalyses. Effect sizes were estimated with �p
2. The data arereported as mean � SEM.
Results
Reaction times. The three-way ANOVA on RTs (Figure 2D)revealed a significant main effect of Perspective congruence(F1,27 � 111.480, p � .001, �p
2 � 0.805), because RTs were fasterwhen probes and primes were in the same perspective (602.36 �13 ms) rather than in different perspectives (628.94 � 12.27 ms).The main effect of Action congruence was also significant(F2,54 � 89.422, p � .001, �p
2 � 0.768), revealing that RTs incongruent trials were faster (600.99 � 12.68 ms) than RTs inincongruent (631.85 � 12.33 ms; p � .001) and neutral trials(614.11 � 12.93 ms; p � .001); RTs in incongruent trials werealso slower than RTs in neutral trials (p � .001). The two-wayinteractions Prime duration � Perspective congruence (F1,27 �85.09, p � .001, �p
2 � 0.759), Prime duration � Action congru-ence (F2,54 � 54.635, p � .001, �p
2 � 0.669), and Perspectivecongruence � Action congruence (F2,54 � 45.150, p � .001, �p
2 �0.625) were also significant and were further qualified by a sig-nificant three-way interaction Prime duration � Perspective con-gruence � Action congruence (F2,54 � 37.853, p � .001, �p
2 �0.583). Thus, the congruence between the prime and probe actionsexerted different effects on performance according to the compat-ibility of their perspective and to prime duration.
To explore the significant three-way interaction we ran twoseparate, two-way ANOVAs, directly testing the effects of Per-spective and Action congruence in each visibility condition. TheANOVA on the low-visibility condition revealed a significantmain effect of Action congruence (F2,54 � 6.055, p � .004, �p
2 �0.183), with responses to congruent trials (602.08 � 14.19 ms)being faster than those to incongruent (610.90 � 13.41 ms, p �.004) and neutral trials (608.91 � 14.60 ms, p � .013), which inturn were not different (p � .458). The nonsignificant effects ofPerspective congruence (F1,27 � 1.517, p � .228, �p
2 � 0.053) andof the interaction Perspective congruence � Action congruence(F2,54 � 2.483, p � .093, �p
2 � 0.084) showed that the compati-bility between the viewing perspective of the probe and of theprime had no effect in the condition of low prime visibility.Conversely, considering the high-visibility condition, we found asignificant interaction between Perspective congruence and Actioncongruence (F2,54 � 74.547, p � .001, �p
2 � 0.734). Newman–Keuls post hoc analyses indicated that when the prime and probewere viewed from the same perspective RTs were faster in thecongruent trials (552.09 � 13.46) than in both incongruent (649.91 �13.97; p � .001) and neutral trials (595.26 � 14.11; p � .001);also RTs were faster in neutral than incongruent trials (p � .001).Thus, both facilitation and inhibitory effects were obtained, respec-tively, for congruent and incongruent combinations of prime andprobe hand actions when they were viewed from the same perspec-tive. In contrast, when the prime and probe hands were viewedfrom different perspectives, we did not find any difference be-tween congruent, incongruent, and neutral trials (All p � .09).
Error rates. The three-way ANOVA on ERs (see Table 1)yielded a significant main effect of Action congruence (F2,54 �13.177, p � .001, �p
2 � 0.327), revealing that ERs in incongruent
trials were higher (4.9% � 0.5) than ERs in congruent (3.44% �0.5; p � .001) and neutral trials (3.46% � 0.4; p � .001); ERs incongruent trials were not different from ERs in the neutral condi-tion (p � .95). The main effect of Perspective congruence was notsignificant (F1,27 � 1). The two-way interactions Prime dura-tion � Action congruence (F2,54 � 6.421, p � .003, �p
2 � 0.192)and Perspective congruence � Action congruence (F2,54 � 6.291,p � .003, �p
2 � 0.188) were also significant and were furtherqualified by a significant three-way interaction (F2,54 � 12.826,p � .001, �p
2 � 0.322). To explore the significant three-wayinteraction we ran two separate, two-way ANOVAs, directly test-ing the effects of Perspective and Action congruence in eachvisibility condition. Nonsignificant effects were found in the low-visibility condition (All F � 1), while a significant interactionbetween Perspective congruence and Action congruence was ob-tained for the high-visibility condition (F2,54 � 15.479, p � .001,�p
2 � 0.364). Post hoc analysis indicated that when the prime andprobe were viewed from the same perspective, ERs were higher inthe incongruent trials (7.3% � 0.9) than in both congruent (2.2% �0.4; p � .001) and neutral trials (2.5% � 0.4; p � .001), which inturn did not differ from each other (p � .616). In contrast, whenthe prime and probe hands were viewed from different perspec-tives, we did not find any effects (All p � .72).
Discussion
The results of Experiment 2 showed that the congruence be-tween the prime and the probe actions affected participants’ re-sponses in both low and high prime visibility conditions. Whencompared with neutral, still-hand primes, the observation of in-congruent action primes induced interference (incongruent trialsslower than neutral ones) and the observation of congruent actionprimes induced facilitation of the responses to the probe (congru-ent trials faster than both neutral and incongruent trials). Interfer-ence and facilitation effects, however, were both present onlywhen the prime was consciously perceived (high-visibility condi-tion). When the prime was not consciously perceived (low-visibility condition), we obtained only a facilitation for congruentversus incongruent and neutral primes, but no interferential effectsfor viewing incongruent versus neutral primes. This pattern ofresults was also confirmed by the analysis of ERs, which showedinterference, but not facilitation, effects in the high-visibility con-dition, whereas no effects at all were obtained in the low-visibilitycondition. These results are in line with that of Experiment 1,providing evidence of action specific congruence-related facilita-tion of speed, but not accuracy of responses in the condition oflow-visibility of the prime. Interference effects were found only inthe condition of high visibility of the prime and were not presentwhen the prime duration was too short (17 ms) to allow consciousperception, probably because there was no need to suppress a weakand rapidly decaying representation (see Vainio, 2011).
Experiment 2 also showed that the facilitation effects of viewingcongruent prime-probe combinations are independent of the spatialcompatibility between the positions occupied by the moving fingerin the two stimuli. Indeed, because in the trials with the sameaction but different perspectives the finger movement occurred inopposite hemifields, spatial compatibility cannot explain the ef-fects of action congruence in the low-visibility condition. Actioncongruence effects, however, were affected by the compatibility
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
9UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

between the perspective of the probe and that of the prime inconditions of high prime visibility. Indeed, whereas bidirectionalaction priming effects were found when the prime and probe handswere viewed from the same perspective and action and spatialcongruence effects were in the same direction, no facilitation orinterference effects were found when the prime and the probe haddifferent perspectives and there was a conflict between the direc-tion of the action congruence and that of the perspective congru-ence. In a similar vein, while clear interference on accuracy wasfound for the high-visibility, same-perspective condition, no effectwas obtained for different perspective trials. It is important to note,however, that in no conditions did we find evidence of facilitationor interference of responses according to the spatial compatibilitybetween the side of the moving finger in the prime and in the probe(i.e., inversion of the action congruence effect with different per-spective trials). Thus, spatial compatibility did not induce per sefacilitation or interference effects on the responses to the probe,but attenuated the effect of the action priming, in particular inconditions of high-visibility.
In sum, the results of Experiment 2 provide evidence for gen-uine visuoperceptual action congruence priming effects that areindependent of perceptual awareness. They also suggest that actionspecific representations are affected by the compatibility betweenthe perspective of the prime and probe hands in conditions of high,but not low visibility of the prime.
Experiment 3
The previous experiments demonstrated that masked presenta-tions of implied-action hands facilitate the responses to congruentversus incongruent and neutral action stimuli. This action congru-ence facilitation is independent of perceptual awareness (Experi-ments 1 and 2) and cannot be ascribed to spatial compatibility(Experiment 2). However, the prime stimulus was presented for avery short time (17 ms) and, thus, it was unclear whether thedifference between the low- and high-visibility conditions wasattributable to the short stimulus duration per se or to the fact thatthe mask prevented conscious perception (Vainio, 2011). To fur-ther investigate the effects of unconscious processing of actionprimes, in Experiment 3 we introduced a rotating mask to preventconscious perception of the prime with high presentation times,which were comparable with the high-visibility condition in Ex-periments 1 and 2. Furthermore, we asked our participants not onlyto report their subjective perception of the prime, but also to tryand guess to discriminate the prime in a control task run at the endof the experimental session, after participants were informed of theprime presence. This allowed us to study the visibility of the primeobjectively. Finally, we also tested the interaction between visuo-perceptual and visuomotor priming within the same group ofparticipants and in the same behavioral task directly (Brass et al.,2000; Catmur & Heyes, 2011; Gowen & Poliakoff, 2012). Partic-ipants were required to report the presence of abduction of theindex or little finger in the probe hand by performing, in differentblocks, an abduction movement of the index or little finger and toreport the absence of any finger abduction with the other finger.Therefore, to avoid any explicit response bias (Di Pace, Maran-golo, & Pizzamiglio, 1997; Soutschek, Taylor, Mu, & Schubert,2013), participants were asked to report the presence versus ab-sence of implied action, as in Experiment 1, rather than discrim-
inating the moving finger, as in Experiment 2. In sum, Experiment3 was aimed at testing the effects of the congruence between theprime and the probe (visuoperceptual priming) and between the probeand the manual response (visuomotor priming), when the primeremained unnoticed despite relatively high presentation times.We expected comparable facilitation effects when the prime wascongruent versus incongruent or neutral with respect to the probeand/or the manual response.
Method
Participants. A new group of 27 students (6 men) of theUniversity of Udine (mean age � 22.44 years, standard devia-tion � 3.96 years) participated in the experiment. All participantswere native Italian speakers of Caucasian ethnicity, were right-handed, and reported having normal or corrected-to-normal visualacuity. The recruitment procedures were as in the previous exper-iments.
Stimuli. We used the same hand stimuli as Experiment 2. Inaddition, a rotating mask was prepared by overlapping twoidentical star-like geometrical figures that were textured with ascrambled version of the hand pictures using the Bryce 3Dsoftware (DAZ Productions, Inc, Salt Lake City, UT). To maskany dynamic cues related to the prime, the two mask figuresrotated in opposite directions. We prepared two different ver-sions of the mask that were presented randomly in differenttrials. The foreground figure rotated clockwise while the back-ground figure rotated counterclockwise in one version, and viceversa in the other.
Procedure. Visual stimulation was performed on a com-puter screen (75 Hz CRT), which the participants looked at inbinocular vision from a distance of 57 cm while their head wasrestrained by means of a head-and-chin rest. As schematized inFigure 3A, the general procedure involved the presentation of acentral luminous dot serving as fixation mark for 253 ms; thena picture of a still hand (sample) appeared on the screen for 253ms and was followed by a rotating mask (forward mask). Theduration of the forward mask randomly varied between 106 and265 ms (in 53-ms steps) to prevent predictability of primepresentation. After the forward mask, the prime hand appearedfor 53 ms and was subsequently masked with a rotating mask(backward mask) for 133 ms; then the probe hand was presentedfor 253 ms, to which a blank screen followed until response orfor a maximum of 1,507 ms. Responses were recorded whenprovided within the 1,507-ms period after probe offset. Thesample was always a still hand, whereas both the prime andprobe hands could be still or with the index or little fingerabducted. Participants were required to report the presence orabsence of implied movement in the probe hand using the indexfinger or little finger. The response keys were placed, respec-tively, 2 cm to the left of the index finger and 2 cm to the rightof the little finger of the participant’s right hand (Figure 3B).This configuration forced participants to respond performingabduction movements similar to the ones depicted in the im-plied motion stimuli. This ensured that responses were congru-ent or incongruent with the specific movement shown in theprime and/or probe hand. All participants performed 2 blocks:one block was performed using the index finger to report thepresence of implied-action and the little finger to report its
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
10 MELE, MATTIASSI, AND URGESI
F3
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

Figure 3. Method and results for Experiment 3. A) Trial structure: participants were required to report thepresence or absence of implied movement in the probe hand, which was preceded by the masked presentationof congruent, incongruent, or neutral primes in conditions of low visibility. B) Response configuration: theresponse keys were placed to the left and to the right of the participant’s hand, in a position that forcedparticipants to respond performing abduction movements similar to the ones depicted in the implied-actionstimuli. C) Results showed that the presentation of congruent primes facilitated the responses to the probe withrespect to incongruent and neutral primes (significant Probe � Prime interaction). On the other hand, reportingan implied-action probe was facilitated when participants responded with the same finger movement depicted inthe probe (significant Probe � Effector interaction), whereas no effect of the prime implied-action was obtainedon the execution of different finger movements (nonsignificant Prime � Effector interaction). Asterisks indicatesignificant comparisons at p � .05.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
11UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

absence; the other block was performed with the oppositekey-response association. The order of the two key-responseassociations was varied between participants.
At the end of the two blocks, the participants were interviewedabout their subjective perception of the prime. Additionally, in thepresent experiment, after having been explicitly informed of thepresence of the prime, participants underwent an objective assess-ment of the prime visibility in a forced-choice manual responsetask using the same experimental procedure as the main experi-ment. This allowed us to ensure that participants could not per-ceive consciously the prime action despite relatively high presen-tation times. In this task, participants were asked to indicatewhether the prime showed a still hand or a hand with the indexfinger or little finger abducted; they responded by pressing one ofthree keys on a computer keyboard. They were instructed toanswer “by feeling” and to guess for every trial in which they didnot notice the presence of the prime. The trial structure used wasidentical to that of the experimental session, with the exceptionthat the blank screen was replaced by the request for a response inthe middle of the screen. This ensured that participants were facedwith exactly the same conditions as in the main experiment. Notime limit was given for participants’ responses. The next trialstarted immediately after the response. All 384 trials of the exper-imental session were presented and randomized in 2 blocks of 192trials each. The control task lasted approximately 4 min.
Data analysis. All the participants performed the experimen-tal task without anticipations (�120 ms) and with low ERs. Twoparticipants reported having noticed the prime during the experi-ment and were excluded from the analyses. ERs and mean RTs forcorrect responses of the remaining participants were entered into athree-way, repeated-measures ANOVA. The study used a 2 � 3 �3 design with three repeated-measure variables: Effector (implied-action responses given with the index or the little finger), Probe(index finger abduction, little finger abduction, or still hand), andPrime (index finger abduction, little finger abduction, or stillhand). All pairwise comparisons were performed using theNewman–Keuls post hoc test. A significance threshold of p � .05was set for all statistical analyses. Effect sizes were estimated with�p
2. Data are reported as mean � SEM.
Results
The three-way ANOVA on RTs revealed a significant maineffect of Effector (F1,24 � 10.269, p � .003, �p
2 � 0.299), whichwas further qualified by a significance two-way interaction Effec-tor � Probe (F2,48 � 27.395, p � .001, �p
2 � 0.533), showingautomatic imitation effects (Heyes, 2011) in relation to the probe.Indeed, when participants reported implied-action probes with theindex finger, RTs were slower for the little finger (783.04 � 21.63ms) than index finger (720.22 � 21.88 ms, p � .001) probes. Theopposite pattern was obtained when participants reported the pres-ence of an index or little finger movement with the little finger:RTs were slower for the index finger (744.53 � 23.46 ms) thanlittle finger (679.17 � 20.10 ms; p � .001) probes. Because theresponses to the still probe were given using a different finger fromthose given to implied-action probes, directly comparing the staticand implied motion probes might be affected by intrinsic differ-ences in finger movement speed. However, slower RTs to incon-gruent implied-action than still-hand probes were obtained for both
index (730.44 � 20.53 ms, p � .001) and little finger (706.61 �20.74 ms, p � .018) effector trials. In sum, the results showed clearinterferential visuomotor effects when participants were requestedto report the presence of an implied-action hand by performing afinger movement that was incongruent with the movement de-picted in the observed hand. Such interferential effects for incon-gruent probes were significant when compared with both theresponses to congruent implied-action probes and those to still-hand probes.
Despite a clear visuomotor priming in relation to the probe, novisuomotor priming effect was obtained in relation to the prime,because the two-way interaction between Prime and Effector (F �1) and the three-way interaction of Probe, Prime and Effector(F4,96 � 1.579, p � .186, �p
2 � 0.061) were not significant. Thiscannot be ascribed to the fact that the prime had no effect at all,because the main effect of Prime (F2,48 � 15.707, p � .001, �p
2 �0.395) and the two-way interaction Probe � Prime (F4,96 �40.802, p � .001, �p
2 � 0.629) were significant, showing evidenceof visuoperceptual priming effects in relation to action primes thatwere not consciously perceived. When the probe depicted a statichand, RTs for static prime trials (659.55 � 19.77 ms) were fasterthan RTs for both index (749.08 � 20.97 ms, p � .001) and littlefinger prime trials (746.95 � 20.26 ms, p � .001), which in turndid not differ from each other (p � .804). When the probe depictedan index finger abduction, RTs for index finger prime trials(703.49 � 23.15 ms) were faster than RTs for both still hand prime(735.39 � 20.75 ms, p � .001) and little finger prime trials(758.30 � 23.73 ms, p � .001), which in turn did not differ fromeach other (p � .08). When the probe depicted a little fingerabduction, RTs for little finger prime trials (713.79 � 21.59 ms)were faster than RTs for both still (740.78 � 18 ms, p � .011) andindex finger primes (738.75 � 17.39 ms, p � .012), which, again,did not differ from each other (p � .81). Thus, with respect to theneutral, still hand condition, the results provided evidence offacilitation for congruent prime-probe pairs, but no interference forincongruent prime-probe pairs when the prime was not con-sciously perceived by the observer.
Error rates. The three-way ANOVA on ERs revealed a sig-nificant main effect of Prime (F2,48 � 3.727, p � .031, �p
2 �0.134), because ERs were higher for little finger (4.09 � 0.83%)than still hand primes (2.99 � 0.69%). The two-way interactionsEffector � Probe (F2,48 � 7.779, p � .001, �p
2 � 0.244) andProbe � Prime (F4,96 � 3.17, p � .017, �p
2 � 0.116) weresignificant and were qualified by significant three-way interactionEffector � Probe � Prime (F4,96 � 2.986, p � .022, �p
2 � 0.110).Follow-up Probe � Prime two-way ANOVAs, run on each block,showed that the interaction was significant (F4,96 � 4.39, p �.003, �p
2 � 0.154) only when the participants used the index fingerto report the presence of implied-action probes; in this block, whenthe prime was a still hand, participants committed significantlymore errors in responding to little finger implied-action probesthan to index finger implied-action (p � .007) and still hand probes(p � .004). Conversely, no effects were obtained when the primedepicted an implied-action hand (all p � .1), suggesting that theinteraction between the prime and probe masked the effects of thevisuomotor priming for ERs. Only a significant main effect ofProbe was obtained when participants used the little finger toreport the presence of implied-action hands (F2,48 � 8.35, p �.001, �p
2 � 0.258), with lower ERs for little finger than index
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
12 MELE, MATTIASSI, AND URGESI
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

finger (p � .001) and still hand probes (p � .009). As the numberof errors committed by the participants was very low in thisexperiment (see Table 1), however, any interpretation of theseeffects should be made with caution.
Forced-choice prime discrimination. A 3 � 3, Probe �Prime repeated-measures ANOVA was performed on participants’prime discrimination accuracies in the postexperimental session(see Table 2). The main effects were not significant (All p � .07),whereas the interaction was significant (F(4,96) � 2.733, p � .02;�p
2 � 0.102). Post hoc tests showed that the interaction could beexplained by the fact that participants were more accurate inreporting still-hand than little-finger (p � .02) primes when theprobe depicted a still hand. All other comparisons were not sig-nificant (All p � .15). Furthermore, testing with one-sample t testswhether participants’ prime discrimination performance was betterthan that expected by chance (33%) revealed above-chance levelsonly when both the prime and the probe depicted the same indexfinger abduction (t(24) � 2.482, p � .02) or the same still hand(t(24) � 5.04, p � .001). All together, these results suggest that,even when participants were informed about the presence of theprime and were explicitly focused on its identification, they re-ported the probe and not the prime action and were thus moreaccurate when the prime was congruent with the probe. Theseresults corroborate participants’ subjective reports showing thatthe masked action primes were not consciously perceived andcould not be explicitly discriminated in a forced-choice task.
Discussion
In Experiment 3, we investigated visuoperceptual and visuomo-tor priming when action primes were not consciously perceived.We used forward and backward dynamic masking procedures topresent the prime for a relatively long time (53 ms) but nonetheless
prevent conscious perception by all participants. When we askedour participants whether they saw anything during presentation ofthe rotating mask, most of them (25 of 27) affirmed they had seennothing during mask presentation. This subjective report was con-firmed by the forced discrimination task conducted at the end ofthe main experiment. Indeed, even when participants were explic-itly informed about the presence of the prime and were required todiscriminate whether it depicted a still hand or an index or littlefinger abduction, their discrimination abilities were random andthey responded on the basis of the probe type. This effect isprobably attributable to the masking procedure, because the rotat-ing star-like figure and the following probe hand tended to masknot only the perception of the hand shape but also the flickering ofthe moving finger.
Although the action depicted by the prime could not be con-sciously perceived, it affected the responses to the probe. Wefound, indeed, facilitation effects when the prime was congruentwith the action depicted in the probe. In keeping with the resultsfound in the low-visibility conditions of the previous experiments,no interference effects were found when the masked prime de-picted an incongruent action with respect to a neutral, static hand.Thus, the results showed evidence of visuoperceptual facilitationinduced by action primes even when they were not consciouslyperceived.
The congruence between the effector and the probe affectedRTs, independently of the prime, because responses were inter-fered with when provided with an action that was incongruentversus congruent with the probe. On the other hand, no directeffect of the compatibility between the effector and the prime wasfound, suggesting that the (unconscious) visual representation ofthe prime movement affected the (conscious) visual processing ofthe probe but not the (conscious) motor representation of theresponding finger. In other words, results provide evidence forunconscious visuoperceptual but not visuomotor action primingeffects.
Experiment 4
In Experiment 4 we tested whether the absence of visuomotorpriming for unconsciously perceived primes was attributable to theinterference induced by the simultaneous visuoperceptual primingrelated to the combination of the prime and probe actions. We usedthe same still, index, or little finger action primes of Experiment 3in a standard paradigm for visuomotor priming in which partici-pants respond to symbolic cues. The participants had to respond tocolored frame probes performing abduction movements of theindex or little finger, which were congruent or incongruent withthe prime action. In two consecutive blocks, the prime was pre-sented in conditions of low- or high-visibility. In keeping with theresults of Experiment 3, we expected that the congruence betweenthe prime action and the manual response should affect perfor-mance in conditions of high, but not low, prime visibility.
Method
Participants. A new group of 20 students (6 men) of theUniversity of Udine (mean age � 23.50 years, standard devia-tion � 3.33 years) participated in the experiment. All participantswere native Italian speakers of Caucasian ethnicity, were right-
Table 2Accuracy in the Forced-Choice Prime Discrimination Task(Mean % � SEM) for Experiments 3, 4, and 5
Experiment 3
Prime
Probe Still Index finger Little finger
Still 59.08 � 5.16 43.12 � 5.43 32.96 � 3.75Index finger 36.76 � 5.96 48.16 � 6.10 35.72 � 5.05Little finger 38.64 � 5.81 43.24 � 5.78 40.20 � 5.27
Experiment 4
Prime visibility
Perspective Low High
Congruent 50.45 � 1.31 63.20 � 2.42Incongruent 51.45 � 1.72 62.10 � 2.85
Experiment 5
Prime visibility
SOA Low Periliminal High
Short 50.85 � 1.15 57.85 � 2.15 78.80 � 2.59Long 48.95 � 1.27 60.95 � 2.27 83.45 � 2.78
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
13UNCONSCIOUS PROCESSING AND ACTION PRIMING
T2
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM
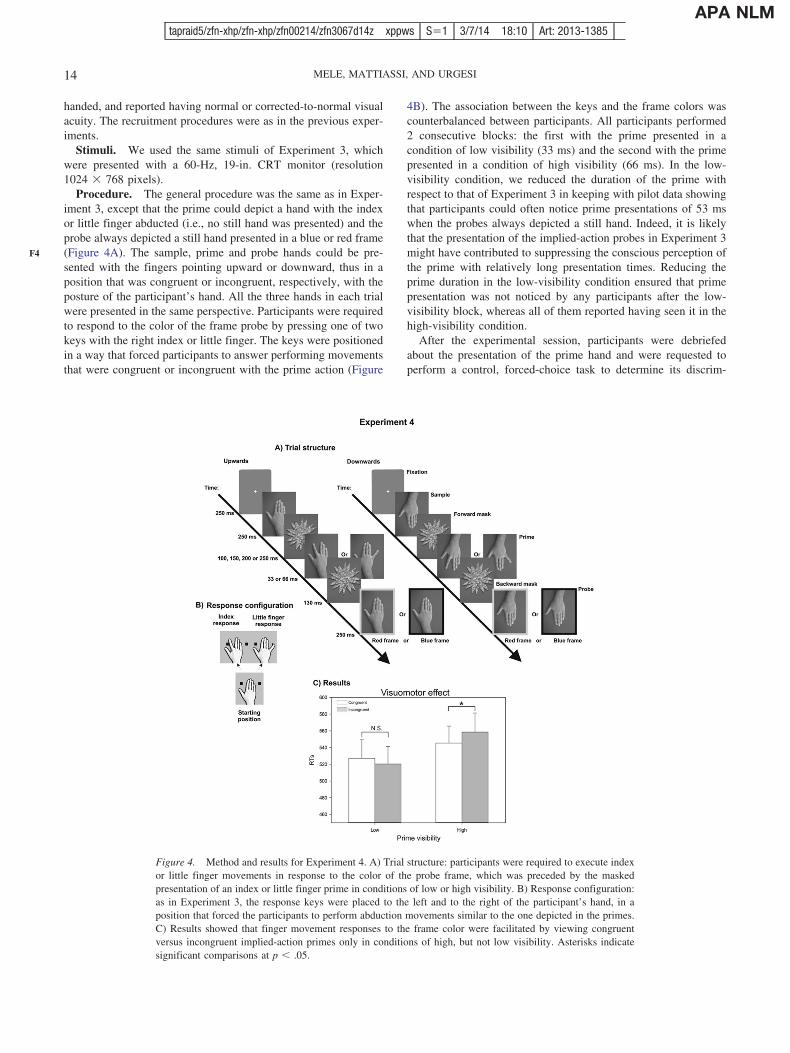
handed, and reported having normal or corrected-to-normal visualacuity. The recruitment procedures were as in the previous exper-iments.
Stimuli. We used the same stimuli of Experiment 3, whichwere presented with a 60-Hz, 19-in. CRT monitor (resolution1024 � 768 pixels).
Procedure. The general procedure was the same as in Exper-iment 3, except that the prime could depict a hand with the indexor little finger abducted (i.e., no still hand was presented) and theprobe always depicted a still hand presented in a blue or red frame(Figure 4A). The sample, prime and probe hands could be pre-sented with the fingers pointing upward or downward, thus in aposition that was congruent or incongruent, respectively, with theposture of the participant’s hand. All the three hands in each trialwere presented in the same perspective. Participants were requiredto respond to the color of the frame probe by pressing one of twokeys with the right index or little finger. The keys were positionedin a way that forced participants to answer performing movementsthat were congruent or incongruent with the prime action (Figure
4B). The association between the keys and the frame colors wascounterbalanced between participants. All participants performed2 consecutive blocks: the first with the prime presented in acondition of low visibility (33 ms) and the second with the primepresented in a condition of high visibility (66 ms). In the low-visibility condition, we reduced the duration of the prime withrespect to that of Experiment 3 in keeping with pilot data showingthat participants could often notice prime presentations of 53 mswhen the probes always depicted a still hand. Indeed, it is likelythat the presentation of the implied-action probes in Experiment 3might have contributed to suppressing the conscious perception ofthe prime with relatively long presentation times. Reducing theprime duration in the low-visibility condition ensured that primepresentation was not noticed by any participants after the low-visibility block, whereas all of them reported having seen it in thehigh-visibility condition.
After the experimental session, participants were debriefedabout the presentation of the prime hand and were requested toperform a control, forced-choice task to determine its discrim-
Figure 4. Method and results for Experiment 4. A) Trial structure: participants were required to execute indexor little finger movements in response to the color of the probe frame, which was preceded by the maskedpresentation of an index or little finger prime in conditions of low or high visibility. B) Response configuration:as in Experiment 3, the response keys were placed to the left and to the right of the participant’s hand, in aposition that forced the participants to perform abduction movements similar to the one depicted in the primes.C) Results showed that finger movement responses to the frame color were facilitated by viewing congruentversus incongruent implied-action primes only in conditions of high, but not low visibility. Asterisks indicatesignificant comparisons at p � .05.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
14 MELE, MATTIASSI, AND URGESI
F4
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

inability. As in Experiment 3, in the forced-choice task participantswere presented with the experimental trials, and after each trial,they were asked to press one of two keys to report whether theprime showed a hand with the index finger or with the little fingerabducted. The trial structure was similar to that used during theexperimental task, with the exception that a response promptappeared in the middle of the screen after the probe. This ensuredthat participants were faced with exactly the same conditions as inthe main experiment. No time limit was given for participants’responses. The next trial started immediately after the response.Two blocks of 136 trials each were presented to the participants insequential order, the first with the prime presented in condition oflow visibility and the second in condition of high visibility. Thecontrol task lasted approximately 8 min.
Data analysis. Two participants had one anticipation(RTs �120 ms) each and the corresponding trial was excludedfrom the computation of the individual ERs and RTs mean. Allparticipants had a good performance in all conditions, withERs �4% (see Table 1). Thus, only mean RTs of correct responseswere used for the analyses. RTs data were entered into a three-way, repeated-measures ANOVA. Considering the congruencybetween the prime implied-action and the finger response, thestudy used a 2 � 2 � 2 design with three repeated-measuresvariables: Prime duration (33 ms, 66 ms), Perspective (upward,downward), and Action congruency (congruent, incongruent). Allpairwise comparisons were performed using the Newman–Keulspost hoc test. A significance threshold of p � .05 was set for allstatistical analyses. Effect sizes were estimated with �p
2. Data arereported as mean � SEM.
Results
Reaction times. The three-way ANOVA on RTs (Figure 4C)revealed a significant main effect of Prime duration (F1,19 �9.724, p � .005, �p
2 � 0.338), because responding in the low-visibility condition was faster (523.71 � 21.73 ms) than respond-ing in the high-visibility condition (551.89 � 21.28 ms). The maineffect of Action congruence was not significant (F1,19 � 1, �p
2 �0.026), but a significant two-way interaction Prime duration �Action congruency (F1,19 � 5.286, p � .033, �p
2 � 0.217) showedthat the visuomotor action priming effect was modulated by primevisibility. Indeed, whereas in the low-visibility condition we didnot obtain any difference between congruent and incongruentprimes (p � .275), in the high-visibility Condition RTs for con-gruent primes (545.35 � 20.24 ms) were faster than RTs forincongruent primes (558.43 � 22.86 ms, p � .046). The perspec-tive of the prime did not affect the visuomotor priming effect, asthe main effect of Perspective and all the two-way (All F1,19 � 1)and three-way (F1,19 � 1.231, p � .280, �p
2 � 0.060) interactionsinvolving the effect of Perspective were not significant.
Forced-choice prime discrimination task. A 2 � 2 repeatedmeasures ANOVA with Prime duration and Perspective as within-subjects variables was performed on participants’ prime discrim-ination accuracies in the forced-choice prime discrimination task(see Table 2). Only the main effect of Prime duration was signif-icant (F(1,19) � 20.927, p � .001; �p
2 � 0.524), with higherdiscrimination performance in the high- than low-visibility condi-tion, whereas all other effects were nonsignificant (All F � 1).Comparing the discrimination performance in each condition
against the chance level (i.e., 50%) showed that only in thehigh-visibility condition did accuracy exceed the chance level forboth upward (t(19) � 5.432, p � .001) and downward (t(19) �4.242, p � .001) perspectives. Conversely, performance was atchance for both perspectives in the low-visibility condition. Theseresults confirm participants’ subjective reports that the prime wasnot consciously perceived in the low-visibility condition and wasconsciously perceived in the high-visibility condition.
Discussion
In Experiment 4 we used a standard visuomotor priming para-digm in which participants executed manual responses in responseto symbolic cues. The manual responses were congruent or incon-gruent with the prime actions, which were task irrelevant and couldbe presented in conditions of low and high visibility. The resultscorroborated those of the Experiment 3 in showing that observedactions affected motor preparation and execution processes onlywhen they were consciously perceived (i.e., the prime in thehigh-visibility condition in Experiment 4 and the probe in Exper-iment 3), but not when they were not consciously perceived (i.e.,the prime in the low-visibility condition in Experiment 4 and inExperiment 3).
Experiment 5
In Experiment 5 we tested whether the absence of visuomotorpriming for unconsciously perceived primes was attributable todifferent durations of the priming effects in the low-visibility andhigh-visibility conditions. Previous studies have shown that visuo-motor priming effects are dependent from the time elapsing be-tween the prime and the response to the probe (Vorberg, Mattler,Heinecke, Schmidt, & Schwarzbach, 2003; Sumner, 2007). Thus,in Experiment 5 we used the same stimuli and procedure ofExperiment 4 in a standard paradigm for visuomotor priming inwhich participants respond to symbolic cues, but we recorded alsothe time to initiate the response rather than only the time tocomplete the response (i.e., reaction time). Furthermore, we mea-sured the response initiation times at two stimulus onset asynchro-nies (SOAs) between prime and probe: one was set at 166 ms(short SOA) and the other one was set at 250 ms (long SOA). Inthree consecutive blocks, the prime was presented in conditions oflow (33 ms), periliminal (66 ms), and high visibility (99 ms). Weincluded the 99-ms prime duration, along with the 33- and 66-msdurations used in Experiment 4, in keeping with pilot data showingthat participants could not notice prime presentations of 66 ms inthe new experimental setting that required recording initiationtimes.
In keeping with the results of Experiment 4, we expected thatthe congruence between the prime action and the manual responseshould affect performance in conditions of high, but not perilimi-nal or low prime visibility.
Method
Participants. A new group of 20 students (4 men) of theUniversity of Udine (mean age � 24.10 years, standard devia-tion � 6.51 years) participated in the experiment. All participantswere native Italian speakers of Caucasian ethnicity, were right-
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
15UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

handed, and reported having normal or corrected-to-normal visualacuity. The recruitment procedures were as in the previous exper-iments.
Stimuli. We used the same stimuli of Experiment 4, whichwere presented with a 60-Hz, 19-in. CRT monitor (resolution1024 � 768 pixels).
Procedure. The general procedure was the same as in Exper-iment 4, except that the probe was a small, colored square appear-ing at the center of the hand image; this ensured that participantswere fixating at the center of the images during prime presentation
rather than paying attention to any part of the image borders. Thetrial structure was similar to that used in Experiment 4, but theduration of the events was adapted according to the manipulationof prime visibility (33 ms, 66 ms, or 99 ms) and SOA (166 ms or250 ms). The forward mask could last 66 ms or 150 ms to preventanticipation of prime presentation, whereas the duration of thebackward mask was dependent from the duration of the previousevents and was varied from 66 ms to 217 ms such that the totalduration of the sequence ‘forward mask–prime–backward mask’was 315 ms in all trials (Figure 5A). A black square was presented
Figure 5. Method and results for Experiment 5. A) Trial structure: participants were required to execute indexor little finger movements in response to the color of a probe square, which was preceded by the maskedpresentation of an index or little finger prime in conditions of low, periliminal or high visibility. The prime couldbe presented for 33 ms, 66 ms, or 99 ms, whereas the stimulus onset asynchrony (SOA) between prime and probecould be 166 ms or 250 ms. The duration of the forward and backward masks was adapted such that the totalduration of the sequence ‘forward mask–prime–backward mask’ was 315 ms in all trials. B) Responseconfiguration: Differently than in Experiment 4, participants had to press two keys at the beginning of the trialand were required to respond to the color of the probe by releasing one of two keys with the right index or littlefinger to press one of two other response keys. The response keys were placed to the left and to the right of theparticipant’s hand, in a position that forced the participants to perform abduction movements similar to the onesdepicted in the primes. C) Results showed that finger movement responses to the probe color were facilitated byviewing congruent versus incongruent implied-action primes only in conditions of high, but not periliminal orlow visibility. Asterisks indicate significant comparisons at p � .05.
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
16 MELE, MATTIASSI, AND URGESI
AQ:9, F5
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

since the beginning of the trial and the probe stimulus corre-sponded to a change of its color to blue or red. In keeping withExperiment 4, participants had to press one of two keys in responseto the probe by performing movements that were congruent orincongruent with the prime action. Differently than in Experiment4, however, at the beginning of each trial participants were re-quired to press with the index and little fingers two start keyspositioned centrally with respect to the response keys; thus, re-sponding to the probe required the release of one of the two startkeys (depending on the response prompted by the probe) and thento press the corresponding response key (see Figure 5B). This way,we were able to record for each trial both the response initiationtime (i.e., the time elapsed between onset of the probe and releaseof the start key) and the response execution time (i.e., the timeelapsed between release of the start key and press of the responsekey). The association between the keys and the frame colors wascounterbalanced between participants. All participants performed3 consecutive blocks: the first with the prime presented in condi-tion of low visibility (33 ms), the second with the prime presentedin condition of periliminal visibility (66 ms), and the third with theprime presented in condition of high visibility (99 ms).
After the experimental session, participants were debriefedabout the presentation of the prime hand and were requested toperform a control, forced-choice task to determine its discrim-inability. As in Experiment 4, in the forced-choice task participantswere presented with the experimental trials, and after each trial,they were asked to press one of two keys to report whether theprime showed a hand with the index finger or with the little fingerabducted. The trial structure was similar to that used during theexperimental task, with the exception that a response promptappeared in the middle of the screen after the probe. This ensuredthat participants were faced with exactly the same conditions as inthe main experiment. No time limit was given for participants’responses. The next trial started immediately after the response.Three blocks of 160 trials each were presented to the participantsin sequential order, the first with the prime presented in conditionof low visibility, the second in condition of periliminal visibility,and the third in condition of high visibility. The control task lastedapproximately 12 min.
Finally, in a stimulus evaluation phase administered at the endof the experimental session we asked participants to evaluate theimplied-motion evoked by each stimulus. Each of the 48 stimuliwas randomly presented in two blocks for a total of 96 trials. Eachtrial started with the presentation of a central fixation point lasting500 ms, followed by the hand image stimulus presented for 2,000ms at the center of the screen. After the stimulus, the question“How much does it imply movement (Quanto implica movimentoin Italian)?” appeared on the screen with a vertical, 10-cm VisualAnalogue Scale (VAS) ranging from “much (molto)” (score �100) to “little (poco)” (score � 0). The top or bottom positions ofthe two extremes were balanced between participants. The partic-ipants were asked to express an implied-motion judgment on thehand stimuli by moving the mouse cursor onto the point of theVAS corresponding to their opinion. The evaluation phase lastedapproximately 5 min.
Data analysis. No participant showed any response anticipa-tion and all had good performance in all conditions, withERs �4.2% (see Table 1). Considered the ceiling effects in accu-racy, we did not analyze further ERs, but mean response initiation
times and response execution times of correct responses were usedfor the analyses. Data were entered into a three-way, repeated-measures ANOVA. Considering the congruency between theprime implied-action and the finger response, the study used a 3 �2 � 2 design with three repeated-measures variables: Prime dura-tion (33 ms, 66 ms, 99 ms), SOA (short, long) and Action con-gruency (congruent, incongruent). All pairwise comparisons wereperformed using the Newman–Keuls post hoc test. A significancethreshold of p � .05 was set for all statistical analyses. Effect sizeswere estimated with �p
2. Data are reported as mean � SEM.
Results
Response initiation times. The three-way ANOVA on re-sponse initiation times revealed a significant main effect of Actioncongruence (F1,19 � 6.431, p � .020, �p
2 � 0.252), becauseresponding in the congruent condition was faster (467.11 � 13.68ms) than responding in the incongruent condition (470.47 � 13.76ms). The main effect of Prime duration was not significant(F2,38 � 1.236, p � .30, �p
2 � 0.061), but a significant two-wayinteraction Prime duration � Action congruency (F2,38 � 5.594,p � .007, �p
2 � 0.227) showed that the visuomotor priming effectwas modulated by prime visibility. Indeed, whereas in the low- andperiliminal-visibility conditions we did not obtain any differencebetween congruent and incongruent primes (all p � .365), in thehigh-visibility Condition RTs for congruent primes (468.20 �13.51 ms) were faster than RTs for incongruent primes (482 �16.36 ms, p � .005). The SOA did not affect the visuomotorpriming effect, as the main effect of SOA and all the two-way (AllF � 1.143, p � .329, �p
2 � 0.056) and three-way (F2,38 � 1)interactions involving the effect of SOA were not significant.
Response execution times. The three-way ANOVA on re-sponse execution times revealed significant main effect of Primeduration (F2,38 � 4.989, p � .011, �p
2 � 0.207), because respond-ing in the high-visibility condition was faster (116.18 � 12.61 ms)than responding in the low-visibility condition (134.15 � 16.50ms, p � .009), but it was not different from responding in theperiliminal-visibility condition (122.99 � 14.14 ms, p � .243);furthermore, responding in the periliminal-visibility conditiontended to be faster than responding in the low-visibility condition(p � .059). No other effects were significant (all F � 2.742, p �.077).
Forced-choice prime discrimination task. A 3 � 2 repeatedmeasures ANOVA with Prime duration and SOA as within-subjects variables was performed on participants’ prime discrim-ination accuracies in the forced-choice prime discrimination task(see Table 2). Only the main effect of Prime duration was signif-icant (F(2,38) � 100.438, p � .001; �p
2 � 0.840), with higherdiscrimination performance in the high- (81.12 � 0.02%) thanboth periliminal- (59.40 � 0.01%, p � .001) and low-visibilityconditions (49.90 � 0.008%, p � .001) and higher discriminationperformance in the perilimininal- than low-visibility condition(p � .001). The main effect of SOA was not significant (F(1,19) �3.490, p � .07; �p
2 � 0.155), whereas the two-way interactionPrime Duration � SOA was significant (F(2,38) � 4.747, p � .014;�p
2 � 0.199). The Post hoc analysis revealed that only in the highvisibility condition the SOA modulated prime discrimination withhigher scores in the long (83.45 � 0.02%) than in the short SOA(78.80 � 0.02%, p � .005). Comparing the discrimination perfor-
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
17UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

mance in each Prime duration against the chance level (i.e., 50%)showed that performance exceeded the chance level in the high-(t(20) � 11.786, p � .001) and periliminal-visibility (t(20) � 4.748,p � .001) conditions, but was at chance for the low-visibilitycondition (t(20) � �0.119, p � .905). These results show thatparticipants did not subjectively report primes presented for 33 msand were not able to discriminate them when their attention wasexplicitly directed to prime presentation; with 66-ms prime dura-tion, participants failed to report prime presentation but, as inExperiment 4, they were able to discriminate above chance theprime type in the forced-choice prime discrimination task; onlywith the 99-ms prime duration did the participants demonstrateconscious prime perception in both the subjective and objectivetests of prime perception.
Stimuli evaluation phase. A 3 � 2 repeated measuresANOVA, with Action posture (index finger abduction, little fingerabduction, still hand) and Perspective (upward, downward) aswithin-subjects variables, was performed on participants’ implied-motion judgments. Only the main effect of Action posture wassignificant (F(2,38) � 108.170, p � .001; �p
2 � 0.850); the post hocanalysis indicated that the VAS implied-motion judgments forboth index finger (57.18 � 4.55 mm) and little finger (59.73 �4.40 mm) abduction stimuli were higher than those for still handstimuli (9.00 � 1.64 mm; all Ps �0.001). No difference wasobserved between the implied motion evoked by index and littlefinger abduction stimuli (p � .515). The main effect of Perspectiveand the two-way interaction were not significant (All Fs �1.71,p � .193). Thus, the results of the stimuli evaluation phaseconfirmed that pictures depicting index or little finger abductionimplied more movement than those depicting a resting relaxedhand.
Discussion
As in Experiment 4, in Experiment 5 we used a standardvisuomotor priming paradigm in which participants executed man-ual responses to symbolic cues. We collected the response initia-tion times and response execution times at two different SOAsbetween prime and probe. The results corroborated those of theExperiment 4 in showing that observed actions affected motorpreparation and initiation processes only when they were con-sciously perceived, but they did not exert any effects when theywere not consciously perceived. Furthermore no visuomotor prim-ing effect was obtained, independently from prime visibility, onthe time needed to execute the movement, suggesting that ob-served actions affect the preparation and initiation phases, but notthe execution phase of motor responses. Finally, the manipulationof the time elapsing between presentation of the action prime andtriggering of the response (i.e., SOA) revealed no effect, suggest-ing that the priming induced by consciously perceived actions wasstill present after 250 ms from the prime onset. On the other hand,no effects were observed for action primes that were not con-sciously perceived, when the priming effects were probed eitherearly on response initiation times or later on RT and longer SOAs.It is also worth noting that in the present experiment we had toincrease the duration of the prime up to 99 ms to ensure allparticipants noticed its presentation. Indeed, at 66 ms duration,which allowed most participants of Experiment 4 to be aware ofthe prime, participants of this experiment failed to notice it. This
may be a result of the experimental setting needed to recordresponse initiation times, because participants had to keep pressingthe start keys during the whole ‘forward-mask–prime–backwardmask’ sequence, which might be expected to be more distractingthan keeping a relaxed hand before responding. This further cor-roborates the claim that observer’s visual awareness of the stimulirather than prime duration or SOA per se may determine whetherobserved actions affects motor preparation and initiation pro-cesses.
General Discussion
In the present study we tested how the effects of action obser-vation on subsequent action perception (visuoperceptual priming)and action preparation (visuomotor priming) are modulated by thereduction of visibility of the prime and by the spatial compatibilitybetween the prime and probe actions. Previous studies have shownthat conscious action observation facilitates the perception(Costantini et al., 2008; Finke & Freyd, 1985) and the execution(Brass et al., 2000; Heyes, 2011; Kilner et al., 2003; Stürmer et al.,2000) of congruent versus incongruent actions, thus providingevidence of visuoperceptual and visuomotor action priming. Theseeffects suggest that viewing an action activates its perceptuo-motorrepresentations, affecting subsequent perceptual and motor pro-cessing. In these previous studies, however, the priming action wasalways presented in conditions of high visibility, and the partici-pants were aware of its presentation. It was thus unclear whetherperceptuo-motor action representations require perceptual aware-ness.
Visuoperceptual Action Representations Do NotRequire Perceptual Awareness
In Experiments 1–3 we tested the modulation of visuoperceptualaction priming according to the visibility of the prime and the typeof task. We found reliable evidence across experiments of actionpriming in conditions of high and low prime visibility, suggestingthat viewing an action activated its perceptual representation andallowed a more efficient perceptual processing of subsequent ac-tions (Finke & Freyd, 1985). This perceptual action representationwas activated automatically, for the participants were not re-quested to process the prime stimulus, and unconsciously, forpriming effects were found even when the participants remainedunaware of prime presentation. Increasing the visibility of theprime only had the effect of increasing the congruence-facilitationeffects, but perceptual awareness was not a condition of theirappearance.
Action primes, independently of their visibility, exerted specificeffects on the processing of the action displayed by the probes,whereas they had no effects on processing the body form cues ofthe same stimuli. This suggests that the prime established action-specific representations. Then again, action primes facilitated ac-tion processing both in an action detection task, when participantswere required to detect the presence of an implied-action handversus a still hand (Experiments 1 and 3), and in an actiondiscrimination task, when participants were requested to discrim-inate between index and little finger abductions (Experiment 2). Inthe latter case, the prime and the probe were related for bothperceptual similarity and association with the same response (i.e.,
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
18 MELE, MATTIASSI, AND URGESI
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

reporting index or little finger movements). Conversely, in theformer case, the two finger abduction stimuli were associated withexactly the same response (i.e., reporting an implied-action hand)and the priming effects were only based on perceptual similarity.In keeping with the perceptual priming effects (Sumner, 2007;Tulving & Schacter, 1990), when compared with the neutral con-dition, unconscious processing of action primes in Experiment 3induced only facilitation of responses to congruent probes, but notinterference of responses to incongruent pairs. In Experiment 2,when both perceptual similarity and motor-response compatibilitywere at hand, we found both facilitation and interference effects.This is in keeping with the notion that the priming related toperceptual similarity induces only facilitation effects, whereas thepriming related to the association with the same motor responseinduces both facilitation and interference effects (Sumner, 2007;Tulving & Schacter, 1990). It is worth noting, however, that theinterference effects in Experiment 2 were only obtained when theprime was presented in conditions of high visibility; conversely,only facilitation for congruent actions was found for low-visibilityprimes. All in all these results suggest that unconscious processingof body actions activates perceptual action representations thatfacilitate the processing of similar actions.
Visuomotor Action Representations RequirePerceptual Awareness
In contrast to the evidence of perceptual facilitation induced byunconscious action processing, we did not find any indication ofvisuomotor priming in conditions of low prime visibility. Thissuggests that the influence of action observation on motor repre-sentations requires perceptual awareness. When the action stimuliwere presented in conditions of high visibility, namely for theprobes in Experiment 3 and the high-visibility primes in Experi-ment 4, the responses provided with incongruent movements wereinterfered with, as compared to those provided with congruentmovements. Thus, in keeping with previous studies on the auto-matic imitation effect (Heyes, 2011), action observation automat-ically triggered the motor representation of the same action, whichconflicted with the execution of incongruent movements. Theeffect was automatic, because it was obtained both when theparticipants responded to the action stimuli (Experiment 3) andwhen they responded to nonspatial symbolic cues (coloredsquares) and had to ignore the hand stimuli (Experiment 4 and 5).Comparing the effects on the time needed to initiate the responseor to execute it in Experiment 5 also revealed that observed actionsaffect motor preparation and initiation processes but not the actualimplementation of the motor program. Furthermore, in keepingwith visuoperceptual priming, visuomotor priming was indepen-dent of the association of the two actions to the same or differenttask responses. Indeed, in Experiment 3, the probes depicting anindex and a little finger abduction were associated with the sameresponse, namely reporting an implied-action hand by abductingthe index or little finger. Nevertheless, index finger movementswere slower in response to little than index finger probes, whereaslittle finger movements were slower in response to index than littlefinger probes. This suggests that the association between thestimulus and the motor responses relied on representations thatpreexisted the experimental task and were likely to have beenestablished after the repeated experience of the visual conse-
quences of one’s own movements (Catmur, Walsh, & Heyes,2009; Heyes, 2011). Although in our experimental sessions par-ticipants were presented with an incongruent association betweenaction observation and a given motor response (e.g., abducting theindex finger to report both index finger and little finger impliedaction probes), which is known to reverse typical automatic imi-tation effects (Heyes, Bird, Johnson, & Haggard, 2005), it wasprobably too short a training to substitute or superimpose the newtask-related association to the long-lasting perceptuo-motor asso-ciations of congruent movements and visual feedbacks.
Several previous studies in a different field of research haveshown visuomotor priming effects with both high- and low-visibility primes when the prime and target were strongly associ-ated to a lateralized motor response (Sumner, 2007; Eimer &Schlaghecken, 2002, 2003). These effects have been found notonly for symbolic cues (e.g., left or right pointing arrows), but alsowhen the prime depicted left or right hands (Vainio, 2009, 2011)or objects with left or right affordance (Vainio, Hammaren,Hausen, Rekolainen, & Riskila, 2011). In these studies, interfer-ence for incongruent versus congruent pairs (positive priming) isfound for high-visibility primes, whereas presentation of low-visibility primes reverses the pattern into a negative priming. Theinterference for incongruent versus congruent motor responsesobtained in Experiments 3 and 4 is in keeping with the positivepriming effects induced by high-visibility primes. On the otherhand, the fact that we did not find any positive or negative primingeffects in condition of low prime visibility cannot be ascribed toinsufficient time for the manifestation of the visuomotor primingeffects (Vorberg et al., 2003), because the SOA was, for allExperiments, in line with the SOA (100–200 ms) at which nega-tive priming effects are typically obtained (Eimer & Schlaghecken,2003). Furthermore, the absence of visuomotor priming effects forlow-visibility primes on both response initiation and completiontimes suggests that the results are unlikely to be ascribed todifferent time courses of the effects of conscious and unconsciousprimes. However, an important factor that must be considered isthat negative visuomotor prime effects strictly depend on theselection of the motor response concurrently required by the ex-plicit task (Eimer & Schlaghecken, 1998, 2003) and seem to reflectthe association of the prime and target with the same or differentlateralized motor responses. In contrast, in our study, the twoaction stimuli were associated to the same motor response inExperiment 3 (i.e., reporting implied-action hands) and were task-independent in Experiment 4 and 5 (i.e., discriminating the framecolor). Thus, visuomotor priming cannot be explained by the sameprocesses as those explaining the visuomotor priming of responsesto spatial cues.
Spatial Compatibility Affects Visuoperceptual, but NotVisuomotor Action Representations
A further aim of the present study was to clarify and dissociatespatial compatibility and action-specific representations in visuo-perceptual and visuomotor action priming. To this end, we pre-sented the prime hand in a perspective that was congruent orincongruent with the perspective of the probe hand (in Experiment2, testing visuoperceptual priming) or with the posture of theparticipant’s responding hand (in Experiment 4, testing visuomotorpriming). Such manipulation changed the side of the space in
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
19UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

which the finger movements occurred, reversing spatial and actioncongruence, without changing the identity of the depicted hand. InExperiment 2, we found that spatial compatibility between theprime and the probe action modulated the visuoperceptual actionpriming in conditions of high but not low prime visibility. Whereasaction-congruence facilitation with low-visibility primes was ob-tained independently of perspective compatibility, facilitation forcongruent prime-probe pairs and interference for incongruentprime-probe pairs (with respect to neutral trials) were obtainedonly when the prime and probe perspectives were compatible. Thenull effect in condition of incompatible perspective might reflectthe interaction between action and spatial congruence, which can-celled each other out when the same moving finger occupiedopposite positions in the prime and in the probe. Such interaction,however, was present only when the prime was consciously per-ceived. This suggests that, whereas perceptual action representa-tions are activated at a completely unconscious processing level,the orienting of spatial attention is not activated by such low-visible primes that could not be detected by the participants evenwhen they were directly prompted to it. This is in keeping withstudies showing no spatial compatibility effects of low-visibilityspatial cues on the detection of visual targets presented 105–150ms later (Ivanoff & Klein, 2003; Mele, Savazzi, Marzi, & Berluc-chi, 2008). Importantly, spatial-compatibility effects were foundfor both low- and high-visibility cues when the cue-target SOAwas 750 ms or longer, suggesting that spatial compatibility, also inconditions of low prime visibility, might affect visuoperceptualaction priming at longer SOAs than those used in this study.
Although spatial compatibility affected visuoperceptual actionpriming in conditions of high prime visibility, no spatial compat-ibility effects were obtained for visuomotor priming in Experiment4. Indeed, no effects were obtained for low-visibility primes,independently from their perspective; conversely, comparablevisuomotor priming effects were obtained for high-visibilityprimes when the perspective of the prime hand was both congruentand incongruent with the responding hand. This is in keeping withprevious findings that automatic imitation is independent of spatialcompatibility (Catmur & Heyes, 2011; Heyes, 2011). Notably,whereas previous studies have controlled spatial compatibilityeffects by presenting left and right hands, we always presentedright hand stimuli viewed with the fingers pointing upward ordownward. This ensured mapping of the same hand actions in theprime and in the response, but varied the egocentric versus allo-centric perspective of the hand. Although previous studies haveshown that motor activation during action observation is higher foractions viewed from a spatial perspective which is compatiblerather than incompatible with the observer’s posture, they havealso consistently shown that the occurrence of such motor activa-tion is independent of the action perspective (Alaerts et al., 2009;Maeda, Kleiner-Fisman, & Pascual-Leone, 2002; Urgesi, Candidiet al., 2006). At the behavioral level, our data are in keeping withprevious evidence (Bortoletto, Mattingley, & Cunnington, 2013)of visuomotor priming effects in both egocentric and allocentricviews and further suggest that they occur even when the observerhas to respond to a symbolic, nonspatial cue and has to ignore theaction stimuli (Experiment 4 and 5). Furthermore, the resultssuggest that visuomotor interaction in representing observed ac-tions, no matter of their viewing perspective, requires perceptualawareness.
Multiple Perceptual and Motor Action Representations
One may argue that our claim that visuomotor interaction re-quires the conscious processing of observed actions is based on anull result, which always leaves open the possibility that othermanipulations of stimuli visibility and task response might lead todifferent results. Obviously, this argument is an inevitable corol-lary of any report of absent modulation following the manipulationof stimulus visibility (or other variables) and would make itvirtually impossible to demonstrate that action observation re-quires perceptual awareness. It should be noted, however, that thesame manipulation of the visibility of action primes did not hindertheir effects on action perception, because visuoperceptual actionpriming was consistently obtained during various action percep-tion tasks (action detection or discrimination) also in low-visibilityconditions. Furthermore, the same task responses were affected byaction stimuli that were consciously perceived independently oftheir role in the task (i.e., the probes in Experiment 3 and thehigh-visibility primes in Experiments 4 and 5). Finally, not onlywere visuoperceptual and visuomotor action-priming effects af-fected differently by the reduction of stimulus visibility, they werealso affected differently by spatial compatibility, with interactingeffects for the former and no modulation for the latter. Thus, themost tenable interpretation of the data is that action observationaffects subsequent perception and execution of actions via differ-ent processes. This does not hamper the general notion ofcommon-coding hypotheses (Hommel et al., 2001; Prinz, 1997)that observed actions are perceived using internal models of thesensory consequences of executed actions, but it argues against acomplete isomorphism between perceptual and motor action rep-resentations. Our results are in keeping with multiple perceptualand motor representations of actions, which only partially overlap,may interact in certain circumstances and not in others, are af-fected differently by perceptual awareness and spatial compatibil-ity and may ultimately serve different functions.
Considering the action priming effects at the functional level, animportant factor that needs to be considered is the level of actionrepresentation that was tested in the present study as in otherautomatic-imitation paradigms (Heyes, 2011). Indeed, the associ-ation between the prime and probe stimuli and between the primeand the manual response concerned the specific kinematics ofintransitive movements. In hierarchical models of action represen-tation (Grafton & Hamilton, 2007; Keele, Cohen, & Ivry, 1990)this corresponds to the first level of movement implementation(e.g., abducting or flexing a specific finger) as compared to that ofaction (e.g., reaching or grasping an object), immediate (e.g.,pressing a button or taking an object) and remote goal (e.g.,switching the light or feeding). Thus, although the present studysuggests that at least partially different processes are involved inthe influences of action observation on action perception andexecution at the movement level, this does not rule out thatcommon perceptual and motor coding is involved in representingactions at more abstract levels.
Previous studies (Costantini et al., 2008) have shown that actionobservation may prime subsequent action perception indepen-dently of the specific effector used (e.g., taking an object with thehands or the feet). In a similar way, the observation of object-directed actions executed with different effectors induced similaractivation of the motor representation of the muscles typically
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
20 MELE, MATTIASSI, AND URGESI
AQ: 4
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

involved in achieving the observed goal (Senna, Bolognini, &Maravita, 2013). Such coding of action end-goals or effects hasbeen also called into action to explain the interaction betweenlanguage and motor processing, as demonstrated for example withthe action-sentence compatibility effect (e.g., Diefenbach, Rieger,Massen, & Prinz, 2013; Ibáñez et al., 2013). In this paradigm,processing sentences that describe motor actions (e.g., moving anobject away from the body) affects the execution of motor re-sponses whose effects are congruent versus incongruent to thedirection of the action effects described in the sentences. Thisaction-sentence compatibility effect is independent from the spe-cific body movements performed to obtain the action effects,suggesting that it involves common-code representation of actiongoals (or effects), rather than of specific motor programs, duringlanguage and motor processing (Diefenbach et al., 2013). Further-more, recording of cortical dynamics during the action-sentencecompatibility task has shown that such language-motor interactionis reflected into modulation of neural activity in both motor- andsemantics-related areas (Aravena et al., 2010; Ibáñez et al., 2013).Overall, these studies point to the existence of an abstract, effec-tor’s independent representation of the goal of observed and exe-cuted actions, which might allow the direct understanding ofothers’ behavior (Rizzolatti & Sinigaglia, 2010). It is unlikely,however, that such abstract goal-representation was activated byobserving the simple, intransitive finger movements used in thepresent study as well as in previous automatic imitation paradigms(Heyes, 2011). Thus, we argue that the visuoperceptual and visuo-motor priming reported here were based on the coding of themovement kinematics of observed actions. At the perceptual level,the activation of movement-specific representations might servethe dynamic integration of different movement phases into aunitary, smooth perception of biological motion, a process thatmay occur at an unconscious level of perception (Güldenpenning,Kunde, Weigelt, & Schack, 2012). At the motor level, the specificrepresentation of the movement kinematics might serve the replicaof the same movements (i.e., imitation), a process that seems toproceed in an automatic manner, independently of the observer’stask, but requires perceptual awareness.
In conclusion, the absence of any effect of the congruencebetween the specific prime action and the executed motor re-sponses in the low-visibility condition of our study suggests thatautomatic imitation effects require perceptual awareness and op-erate at a conscious level of action representation. Such onlyconscious motor processing of others’ actions may allow us tosimultaneously address numerous agents and multiple and possiblyincongruent actions that we must flexibly imitate, complement orreact to (Sartori, Cavallo, Bucchioni, & Castiello, 2012) withoutbeing overloaded by unwanted and uncontrollable automatic imi-tation events.
References
Abernethy, B., Zawi, K., & Jackson, R. C. (2008). Expertise and attun-ement to kinematic constraints. Perception, 37, 931–949. doi:10.1068/p5340
Aglioti, S. M., Cesari, P., Romani, M., & Urgesi, C. (2008). Actionanticipation and motor resonance in elite basketball players. NatureNeuroscience, 11, 1109–1116. doi:10.1038/nn.2182
Alaerts, K., Aggelpoel, T. Van, Swinnen, S. P., & Wenderoth, N. (2009).Observing shadow motions: Resonant activity within the observer’s
motor system? Neuroscience Letters, 461, 240 –244. doi:10.1016/j.neulet.2009.06.055
Aravena, P., Hurtado, E., Riveros, R., Cardona, J. F., Manes, F., & Ibáñez,A. (2010). Applauding with closed hands: Neural signature of action-sentence compatibility effects. PloS one, 5, e11751. doi:10.1371/journal.pone.0011751
Avenanti, A., Candidi, M., & Urgesi, C. (2013). Vicarious motor activationduring action perception: beyond correlational evidence. Frontiers inHuman Neuroscience, 7, 185–192. doi:10.3389/fnhum.2013.00185
Avenanti, A., & Urgesi, C. (2011). Understanding “what” others do: Mirrormechanisms play a crucial role in action perception. Social Cognitiveand Affective Neuroscience, 6, 257–259. doi:10.1093/scan/nsr004
Bertenthal, B. I., Longo, M. R., & Kosobud, A. (2006). Imitative responsetendencies following observation of intransitive actions. Journal ofExperimental Psychology: Human Perception and Performance, 32,210–225. doi:10.1037/0096-1523.32.2.210
Bortoletto, M., Mattingley, J. B., & Cunnington, R. (2013). Effects ofcontext on visuomotor interference depends on the perspective of ob-served actions. PLoS ONE, 8. doi:10.1371/journal.pone.0053248
Brass, M., Bekkering, H., & Prinz, W. (2001). Movement observationaffects movement execution in a simple response task. Acta Psycho-logica, 106, 3–22.
Brass, M., Bekkering, H., Wohlschläger, A., & Prinz, W. (2000). Com-patibility between observed and executed finger movements: Comparingsymbolic, spatial, and imitative cues. Brain and Cognition, 44, 124–143.doi:10.1006/brcg.2000.1225
Briggs, G. G., & Nebes, R. D. (1975). Patterns of hand preference in astudent population. Cortex, 11, 230–238.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., Gallese, V.,. . . Parma, I. (2001). Action observation activates premotor and parietalareas in a somatotopic manner: an fMRI study. European Journal ofNeuroscience, 13, 400–404.
Campana, G., Cowey, A., & Walsh, V. (2002). Priming of motion directionand area V5/MT: A test of perceptual memory. Cerebral Cortex, 12,663–669.
Casile, A., & Giese, M. A. (2006). Nonvisual motor training influencesbiological motion perception. Current Biology, 16, 69–74. doi:10.1016/j.cub.2005.10.071
Catmur, C., & Heyes, C. (2011). Time course analyses confirm indepen-dence of imitative and spatial compatibility. Journal of ExperimentalPsychology: Human Perception and Performance, 37, 409–21. doi:10.1037/a0019325
Catmur, C., Walsh, V., & Heyes, C. (2009). Associative sequence learning:The role of experience in the development of imitation and the mirrorsystem. Philosophical Transactions of the Royal Society of London -Series B: Biological Sciences, 364, 2369–2380. doi:10.1098/rstb.2009.0048
Cattaneo, L., & Barchiesi, G. (2011). Transcranial magnetic mapping ofthe short-latency modulations of corticospinal activity from the ipsilat-eral hemisphere during rest. Frontiers in Neural Circuits, 5, 14. doi:10.3389/fncir.2011.00014
Cattaneo, L., & Rizzolatti, G. (2009). The mirror neuron system. Archivesof Neurology, 66, 557–560. doi:10.1001/archneurol.2009.41
Chong, T. T., Cunnington, R., Williams, M. A., Kanwisher, N., & Mat-tingley, B. (2008). fMRI adaptation reveals mirror neurons in humaninferior parietal cortex. Current Biology, 18, 1576–1580. doi:10.1016/j.cub.2008.08.068
Costantini, M., Committeri, G., & Galati, G. (2008). Effector- and target-independent representation of observed actions: evidence from inciden-tal repetition priming. Experimental Brain Research, 188, 341–351.doi:10.1007/s00221-008-1369-x
Craighero, L., Bello, A., Fadiga, L., & Rizzolatti, G. (2002). Hand actionpreparation influences the responses to hand pictures. Neuropsycholo-gia, 40, 492–502. doi:10.1016/S0028-3932(01)00134-8
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
21UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

Craighero, L., Fadiga, L., Rizzolatti, G., & Umilta, C. a. (1999). Action forperception: A motor-visual attentional effect. Journal of ExperimentalPsychology: Human Perception and Performance, 25, 1673–1692. doi:10.1037/0096-1523.25.6.1673
Craighero, L., Fadiga, L., Umilta, C. a., & Rizzolatti, G. (1996). Evidencefor visuomotor priming effect. Neuroreport, 8, 347–349.
Diefenbach, C., Rieger, M., Massen, C., & Prinz, W. (2013). Action-sentence compatibility: The role of action effects and timing. Frontiersin Psychology, 4, 272. doi:10.3389/fpsyg.2013.00272
Di Pace, E., Marangolo, P., & Pizzamiglio, L. (1997). Response bias incolor priming. Acta Psychologica, 95, 3–14. doi:10.1016/S0001-6918(96)00012-1
Di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G.(1992). Understanding motor events: A neurophysiological study. Ex-perimental Brain Research, 91, 176–180. doi:10.1007/BF00230027
Duncan, D. B. (1955). Multiple range and multiple F tests. Biometrics, 11,1–42. doi:10.2307/3001478
Dunnett, C. W. (1970). Query: Multiple comparison tests. Biometrics, 26,139–141. doi:10.2307/2529050
Eimer, M., & Schlaghecken, F. (1998). Effects of masked stimuli on motoractivation: Behavioral and electrophysiological evidence. Journal ofExperimental Psychology: Human Perception and Performance, 24,1737–1747. doi:10.1037/0096-1523.24.6.1737
Eimer, M., & Schlaghecken, F. (2002). Links between conscious aware-ness and response inhibition: Evidence from masked priming. Psycho-nomic Bulletin and Review, 9, 514–520.
Eimer, M., & Schlaghecken, F. (2003). Response facilitation and inhibitionin subliminal priming. Biological Psychology, 64, 7–26. doi:10.1016/S0301-0511(03)00100-5
Fadiga, L., Craighero, L., & Olivier, E. (2005). Human motor cortexexcitability during the perception of others’ action. Current Opinion inNeurobiology, 15, 213–218. doi:10.1016/j.conb.2005.03.013
Fadiga, L., Fogassi, L., Pavesi, G., & Rizzolatti, G. (1995). Activation inhuman MT/MST by static images with implied motion. Journal ofNeurophysiology, 73, 2608–2611.
Finke, R. A., & Freyd, J. J. (1985). Transformations of visual memoryinduced by implied motions of pattern elements. Journal of Experimen-tal Psychology: Learning Memory and Cognition, 11, 780–794. doi:10.1037/0278-7393.11.1-4.780
Fogassi, L., & Ferrari, P. F. (2011). Mirror systems. Wiley InterdisciplinaryReviews: Cognitive Science, 2, 22–38. doi:10.1002/wcs.89
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Actionrecognition in the premotor cortex. Brain: A Journal of Neurology, 119,593–609.
Gowen, E., & Poliakoff, E. (2012). How does visuomotor priming differfor biological and non-biological stimuli? A review of the evidence.Psychological Research, 76, 407–20. doi:10.1007/s00426-011-0389-5
Grafton, S. T., & Hamilton, A. F. D. C. (2007). Evidence for a distributedhierarchy of action representation in the brain. Human Movement Sci-ence, 26, 590–616.
Güldenpenning, I., Koester, D., Kunde, W., Matthias, W., & Schack, T.(2011). Motor expertise modulates the unconscious processing of humanbody postures. Experimental Brain Research, 213, 383–391. doi:10.1007/s00221-011-2788-7
Güldenpenning, I., Kunde, W., Weigelt, M., & Schack, T. (2012). Primingof future states in complex motor skills. Experimental Psychology, 59,286–294. doi:10.1027/1618-3169/a000156
Hamilton, A. F. D. C., Wolpert, D. M., Frith, U., & Grafton, S. T. (2006).Where does your own action influence your perception of anotherperson’s action in the brain? NeuroImage, 29, 524–535. doi:10.1016/j.neuroimage.2005.07.037
Heyes, C. (2011). Automatic imitation. Psychological Bulletin, 137, 463–83. doi:10.1037/a0022288
Heyes, C., Bird, G., Johnson, H., & Haggard, P. (2005). Experiencemodulates automatic imitation. Cognitive Brain Research, 22, 233–240.
Hogeveen, J., & Obhi, S. S. (2013). Automatic imitation is automatic, butless so for narcissists. Experimental Brain Research, 224, 613–621.doi:10.1007/s00221-012-3339-6
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). TheTheory of Event Coding (TEC): A framework for perception and actionplanning. Behavioral and Brain Sciences, 24, 849–937.
Ibáñez, A., Cardona, J. F., Dos Santos, Y. V., Blenkmann, A., Aravena, P.,Roca, M., . . . Bekinschtein, T. (2013). Motor-language coupling: Directevidence from early Parkinson’s disease and intracranial cortical record-ings. Cortex, 49, 968–984. doi:10.1016/j.cortex.2012.02.014
Ivanoff, J., & Klein, R. M. (2003). Orienting of attention without aware-ness is affected by measurement-induced attentional control settings.Journal of Vision, 3, 32–40.
Jiménez, L., Recio, S., Méndez, A., Lorda, M. J., Permuy, B., & Méndez,C. (2012). Automatic imitation and spatial compatibility in a key-pressing task. Acta Psychologica, 141, 96–103. doi:10.1016/j.actpsy.2012.07.007
Keele, S. W., Cohen, A., & Ivry, R. (1990). Motor programs: Concepts andissues. In M. Jeannerod (Ed.), Attention and performance 13: Motorrepresentation and control (pp. 77–110). Mahwah, NJ: Erlbaum.
Kilner, J. M., Neal, A., Weiskopf, N., Friston, K. J., & Frith, C. D. (2009).Evidence of mirror neurons in human inferior frontal gyrus. The Journalof Neuroscience, 29, 10153–10159. doi:10.1523/JNEUROSCI.2668-09.2009
Kilner, J. M., Paulignan, Y., & Blakemore, S. J. (2003). An interferenceeffect of observed biological movement on action. Current Biology, 13,522–525. doi:10.1016/S0960-9822(03)00165-9
Kristjánsson, A., & Campana, G. (2010). Where perception meets memory:A review of repetition priming in visual search tasks. Attention, Percep-tion, & Psychophysics, 72, 5–18. doi:10.3758/APP.72.1.5
Maeda, F., Kleiner-Fisman, G., & Pascual-Leone, A. (2002). Motor facil-itation while observing hand actions: Specificity of the effect and role ofobserver’s orientation. Journal of Neurophysiology, 87, 1329–1335.
McHugh, M. L. (2011). Multiple comparison analysis testing in ANOVA.Biochemia Medica, 21, 203–209. doi:10.11613/BM.2011.029
Mele, S., Savazzi, S., Marzi, C. A., & Berlucchi, G. (2008). Reaction timeinhibition from subliminal cues: Is it related to inhibition of return?Neuropsychologia, 46, 810–9. doi:10.1016/j.neuropsychologia.2007.11.003
Mukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., & Fried, I. (2011).Single neuron responses in humans during execution and observation ofactions. Current Biology, 20, 750–756. doi:10.1016/j.cub.2010.02.045
Prinz, W. (1997). Perception and action planning. European Journal ofCognitive Psychology, 9, 129–154. doi:10.1080/713752551
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. AnnualReview of Neuroscience, 27, 169–192. doi:10.1146/annurev.neuro.27.070203.144230
Rizzolatti, G., Fadiga, L., Gallese, V., & Fogassi, L. (1996). Premotorcortex and the recognition of motor actions. Brain Research: CognitiveBrain Research, 3, 131–141. doi:10.1016/0926-6410(95)00038-0
Rizzolatti, G., & Sinigaglia, C. (2010). The functional role of the parieto-frontal mirror circuit: Interpretations and misinterpretations. Nature Re-views Neuroscience, 11, 264–274. doi:10.1038/nrn2805
Sandberg, K., Martin, B., Timmermans, B., Cleeremans, A., & Overgaard,M. (2011). Measuring consciousness: Task accuracy and awareness assigmoid functions of stimulus duration. Consciousness and Cognition,20, 1659–1675. doi:10.1016/j.concog.2011.09.002
Sartori, L., Cavallo, A., Bucchioni, G., & Castiello, U. (2012). Fromsimulation to reciprocity: The case of complementary actions. SocialNeuroscience, 7, 146–158. doi:10.1080/17470919.2011.586579
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
22 MELE, MATTIASSI, AND URGESI
AQ: 6
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM

Schütz-Bosbach, S., & Prinz, W. (2007). Perceptual resonance: action-induced modulation of perception. Trends in Cognitive Sciences, 11.doi:10.1016/j.tics.2007.06.005
Senna, I., Bolognini, N., & Maravita, A. (2013). Grasping with the foot:Goal and motor expertise in action observation. Human Brain Mapping.doi:10.1002/hbm.22289
Shmuelof, L., & Zohary, E. (2006). A mirror representation of others’actions in the human anterior parietal cortex. The Journal of Neurosci-ence, 26, 9736–9742. doi:10.1523/JNEUROSCI.1836-06.2006
Shmuelof, L., & Zohary, E. (2008). Mirror-image representation of actionin the anterior parietal cortex. Nature Neuroscience, 11, 1267–1269.doi:10.1038/nn.2196
Silvanto, J., & Soto, D. (2012). Causal evidence for subliminal percept-to-memory interference in early visual cortex. NeuroImage, 59, 840–5.doi:10.1016/j.neuroimage.2011.07.062
Simon, J. R., Hinrichs, J. V., & Craft, J. L. (1970). Auditory S-R compat-ibility: Reaction time as a function of ear-hand correspondence andear-response-location correspondence. Journal of Experimental Psy-chology, 86, 97–102. doi:10.1037/h0029783
Soutschek, A., Taylor, P. C. J., Mu, H. J., & Schubert, T. (2013). Disso-ciable networks control conflict during perception and response selec-tion: A transcranial magnetic stimulation study. The Journal of Neuro-science, 33, 5647–5654. doi:10.1523/JNEUROSCI.4768-12.2013
Stürmer, B., Aschersleben, G., & Prinz, W. (2000). Correspondence effectswith manual gestures and postures: A study of imitation. Journal ofExperimental Psychology: Human Perception and Performance, 26,1746–1759. doi:10.1037//0096-1523.26.6.1746
Sumner, P. (2007). Negative and positive masked-priming – implicationsfor motor inhibition. Advances in Cognitive Psychology, 3, 317–326.
Tulving, E., & Schacter, D. L. (1990). Priming and human memorysystems. Science, 247, 301–306. doi:10.1126/science.2296719
Urgesi, C., Candidi, M., Fabbro, F., Romani, M., & Aglioti, S. M. (2006).Motor facilitation during action observation: Topographic mapping ofthe target muscle and influence of the onlooker’s posture. EuropeanJournal of Neuroscience, 23, 2522–2530. doi:10.1111/j.1460-9568.2006.04772.x
Urgesi, C., Candidi, M., Ionta, S., & Aglioti, S. M. (2007). Representationof body identity and body actions in extrastriate body area and ventralpremotor cortex. Nature Neuroscience, 10, 30–1. doi:10.1038/nn1815
Urgesi, C., Maieron, M., Avenanti, A., Tidoni, E., Fabbro, F., & Aglioti,S. M. (2010). Simulating the future of actions in the human corticospinalsystem. Cerebral Cortex, 20, 2511–2521. doi:10.1093/cercor/bhp292
Urgesi, C., Moro, V., Candidi, M., & Aglioti, S. M. (2006). Mappingimplied body actions in the human motor system. The Journal ofNeuroscience, 26, 7942–7949. doi:10.1523/JNEUROSCI.1289-06.2006
Urgesi, C., Savonitto, M. M., Fabbro, F., & Aglioti, S. M. (2012). Long-and short-term plastic modeling of action prediction abilities in volley-ball. Psychological Research, 76, 542–60. doi:10.1007/s00426-011-0383-y
Vainio, L. (2009). Interrupted object-based updating of reach programleads to a negative compatibility effect. Journal of Motor Behavior, 41,305–316. doi:10.3200/JMBR.41.4.305-316
Vainio, L. (2011). Negative stimulus-response compatibility observed witha briefly displayed image of a hand. Brain and Cognition, 77, 382–390.doi:10.1016/j.bandc.2011.09.007
Vainio, L., Hammaren, L., Hausen, M., Rekolainen, E., & Riskila, S.(2011). Motor inhibition associated with the affordance of briefly dis-played objects. The Quarterly Journal of Experimental Psychology, 64,1094–1110. doi:10.1080/17470218.2010.538221
Vainio, L., & Mustonen, T. (2011). Mapping the identity of a viewed handin the motor system: Evidence from stimulus-response compatibility.Journal of Experimental Psychology: Human Perception and Perfor-mance, 37, 207–221. doi:10.1037/a0021432
Van den Bussche, E., Van den Noortgate, W., & Reynvoet, B. (2009).Mechanisms of masked priming: A meta-analysis. Psychological Bulle-tin, 135, 452–77. doi:10.1037/a0015329
Vogt, S., Taylor, P., & Hopkins, B. (2003). Visuomotor priming bypictures of hand postures: Perspective matters. Neuropsychologia, 41,941–951. doi:10.1016/S0028-3932(02)00319-6
Vorberg, D., Mattler, U., Heinecke, A., Schmidt, T., & Schwarzbach, J.(2003). Different time courses for visual perception and action priming.PNAS Proceedings of the National Academies of Science of the UnitedStates of America, 100, 6275–6280. doi:10.1073/pnas.0931489100
Wiggett, A. J., Downing, P. E., & Tipper, S. P. (2013). Facilitation andinterference in spatial and body reference frames. Experimental BrainResearch, 225, 119–131. doi:10.1007/s00221-012-3353-8
Received August 19, 2013Revision received January 31, 2014
Accepted February 4, 2014 �
Thi
sdo
cum
ent
isco
pyri
ghte
dby
the
Am
eric
anPs
ycho
logi
cal
Ass
ocia
tion
oron
eof
itsal
lied
publ
ishe
rs.
Thi
sar
ticle
isin
tend
edso
lely
for
the
pers
onal
use
ofth
ein
divi
dual
user
and
isno
tto
bedi
ssem
inat
edbr
oadl
y.
23UNCONSCIOUS PROCESSING AND ACTION PRIMING
tapraid5/zfn-xhp/zfn-xhp/zfn00214/zfn3067d14z xppws S�1 3/7/14 18:10 Art: 2013-1385APA NLM




















