
Two Novel Type III-Secreted Proteins of Xanthomonas campestris pv. vesicatoria Are Encoded within...
9
JOURNAL OF BACTERIOLOGY, Mar. 2002, p. 1340–1348 Vol. 184, No. 5 0021-9193/02/$04.000 DOI: 10.1128/JB.184.5.1340–1348.2002 Copyright © 2002, American Society for Microbiology. All Rights Reserved. Two Novel Type III-Secreted Proteins of Xanthomonas campestris pv. vesicatoria Are Encoded within the hrp Pathogenicity Island Laurent Noël, Frank Thieme, Dirk Nennstiel,† and Ulla Bonas* Institut für Genetik, Martin-Luther-Universität Halle-Wittenberg, D-06099 Halle (Saale), Germany Received 24 September 2001/Accepted 4 December 2001 The Hrp type III protein secretion system (TTSS) is essential for pathogenicity of gram-negative plant pathogen Xanthomonas campestris pv. vesicatoria. cDNA-amplified fragment length polymorphism and reverse transcription-PCR analyses identified new genes, regulated by key hrp regulator HrpG, in the regions flanking the hrp gene cluster. Sequence analysis revealed genes encoding HpaG, a predicted leucine-rich repeat- containing protein, the lysozyme-like HpaH protein, and XopA and XopD, which are similar in sequence to Hpa1 from Xanthomonas oryzae pv. oryzae and PsvA from Pseudomonas syringae, respectively. XopA and XopD (Xanthomonas outer proteins) are secreted by the Xanthomonas Hrp TTSS and thus represent putative effector proteins. Mutations in xopA, but not in xopD, resulted in reduced bacterial growth in planta and delayed plant reactions in susceptible and resistant host plants. Since the xopD promoter contains a putative hrp box, which is characteristic of hrpL-regulated genes in P. syringae and Erwinia spp., the gene was probably acquired by horizontal gene transfer. Interestingly, the regions flanking the hrp gene cluster also contain insertion se- quences and genes for a putative transposase and a tRNA Arg . These features suggest that the hrp gene cluster of X. campestris pv. vesicatoria is part of a pathogenicity island. Gram-negative bacterial pathogens of plants and animals have evolved specialized export systems to target virulence factors to their hosts. In Agrobacterium tumefaciens, the causal agent of crown gall, the type IV secretion system mediates transfer of the transferred DNA (T-DNA) and associated pro- teins into plant cells, where proteins encoded by the T-DNA manipulate the plant’s hormone balance and metabolism (20). The type III secretion system (TTSS) allows vectorial secretion of so-called effector proteins across the bacterial envelope into the host, where they modulate host defense responses and physiology (11, 18). In general, mutations in type III or type IV secretion systems strongly affect pathogenicity, highlighting the collective contribution of the translocated effectors to viru- lence. Relatively few effectors transported by the TTSS have been identified in plant pathogens (11). Examples are PopB and PopC from Ralstonia solanacearum (21), DspA from Erwinia amylovora (19), and HopPsyA from Pseudomonas syringae pv. syringae (1), which are encoded in the regions flanking the respective hrp (hypersensitive reaction and pathogenicity) gene clusters. hrp genes encode the TTSS in plant pathogens and are essential for bacterial growth in planta, causing disease in sus- ceptible hosts and eliciting a hypersensitive reaction (HR) in resistant plants (2). The HR is a rapid, localized cell death that is part of the plant’s innate defense responses and that halts pathogen ingress (27). In most cases, specific recognition of the pathogen by the resistant plants is based on a gene-for-gene interaction: bacteria expressing a given avirulence gene (avr) are recognized by plants carrying the corresponding disease resistance gene (46). Activity of all avirulence genes tested so far depends on a functional Hrp TTSS system, and more than 10 avr genes elicit the HR when they are expressed within the resistant-plant cell, indicating that Avr proteins are translo- cated effector proteins (11). Xanthomonas campestris pv. vesicatoria is the causal agent of bacterial spot disease of peppers and tomatoes. The chromo- somal 23-kb hrp gene cluster contains six operons, hrpA to hrpF (9, 15, 16, 24; U. Bonas, unpublished data). Among the more than 20 proteins encoded by the hrp region, nine are highly conserved in plant and animal pathogens. These genes were renamed hrc (hrp conserved) and are likely to form the core of the TTSS (8). Analysis of nonpolar mutants in the hrp operons also identified hpa (hrp-associated) genes, which contribute to, but are not essential for, the interaction with the plant (24; U. Bonas, unpublished data). We recently demonstrated Hrp- dependent secretion of HrpB2 and HrpF, which are encoded within the hrp gene cluster and which are essential for patho- genicity (35). Other secreted proteins that are not essential for pathogenicity are Xanthomonas outer proteins (Xop) such as XopB (34) and avirulence proteins AvrBs3, AvrBs4, AvrRxv, AvrBsT, and AvrBs1 (5, 14, 34, 36). Expression of hrp genes is induced in plant leaves and in minimal medium XVM2 (38, 43) and is controlled by regula- tory genes hrpG and hrpX, located outside of the hrp gene cluster. The HrpG protein belongs to the OmpR family of two-component regulatory systems and regulates the expres- sion of a large regulon, which was recently identified using cDNA-amplified fragment length polymorphism (cDNA- AFLP) analysis. The expression profiles of two isogenic X. campestris pv. vesicatoria strains, 85-10 and 85*, which differ in their hrp gene expression status, were compared (34). Strain * Corresponding author. Mailing address: Institut für Genetik, Mar- tin-Luther-Universität Halle-Wittenberg, D-06099 Halle (Saale), Ger- many. Phone: (49) 345 5526290. Fax: (49) 345 5527277. E-mail: [email protected]. † Present address: Bayer-AG Zentrale Forschung, D-51368 Lever- kusen, Germany. 1340
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Two Novel Type III-Secreted Proteins of Xanthomonas campestris pv. vesicatoria Are Encoded within...

JOURNAL OF BACTERIOLOGY, Mar. 2002, p. 1340–1348 Vol. 184, No. 50021-9193/02/$04.00�0 DOI: 10.1128/JB.184.5.1340–1348.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Two Novel Type III-Secreted Proteins of Xanthomonascampestris pv. vesicatoria Are Encoded within the
hrp Pathogenicity IslandLaurent Noël, Frank Thieme, Dirk Nennstiel,† and Ulla Bonas*
Institut für Genetik, Martin-Luther-Universität Halle-Wittenberg, D-06099 Halle (Saale), Germany
Received 24 September 2001/Accepted 4 December 2001
The Hrp type III protein secretion system (TTSS) is essential for pathogenicity of gram-negative plantpathogen Xanthomonas campestris pv. vesicatoria. cDNA-amplified fragment length polymorphism and reversetranscription-PCR analyses identified new genes, regulated by key hrp regulator HrpG, in the regions flankingthe hrp gene cluster. Sequence analysis revealed genes encoding HpaG, a predicted leucine-rich repeat-containing protein, the lysozyme-like HpaH protein, and XopA and XopD, which are similar in sequence toHpa1 from Xanthomonas oryzae pv. oryzae and PsvA from Pseudomonas syringae, respectively. XopA and XopD(Xanthomonas outer proteins) are secreted by the Xanthomonas Hrp TTSS and thus represent putative effectorproteins. Mutations in xopA, but not in xopD, resulted in reduced bacterial growth in planta and delayed plantreactions in susceptible and resistant host plants. Since the xopD promoter contains a putative hrp box, whichis characteristic of hrpL-regulated genes in P. syringae and Erwinia spp., the gene was probably acquired byhorizontal gene transfer. Interestingly, the regions flanking the hrp gene cluster also contain insertion se-quences and genes for a putative transposase and a tRNAArg. These features suggest that the hrp gene clusterof X. campestris pv. vesicatoria is part of a pathogenicity island.
Gram-negative bacterial pathogens of plants and animalshave evolved specialized export systems to target virulencefactors to their hosts. In Agrobacterium tumefaciens, the causalagent of crown gall, the type IV secretion system mediatestransfer of the transferred DNA (T-DNA) and associated pro-teins into plant cells, where proteins encoded by the T-DNAmanipulate the plant’s hormone balance and metabolism (20).The type III secretion system (TTSS) allows vectorial secretionof so-called effector proteins across the bacterial envelope intothe host, where they modulate host defense responses andphysiology (11, 18). In general, mutations in type III or type IVsecretion systems strongly affect pathogenicity, highlighting thecollective contribution of the translocated effectors to viru-lence.
Relatively few effectors transported by the TTSS have beenidentified in plant pathogens (11). Examples are PopB andPopC from Ralstonia solanacearum (21), DspA from Erwiniaamylovora (19), and HopPsyA from Pseudomonas syringae pv.syringae (1), which are encoded in the regions flanking therespective hrp (hypersensitive reaction and pathogenicity) geneclusters. hrp genes encode the TTSS in plant pathogens and areessential for bacterial growth in planta, causing disease in sus-ceptible hosts and eliciting a hypersensitive reaction (HR) inresistant plants (2). The HR is a rapid, localized cell death thatis part of the plant’s innate defense responses and that haltspathogen ingress (27). In most cases, specific recognition of thepathogen by the resistant plants is based on a gene-for-gene
interaction: bacteria expressing a given avirulence gene (avr)are recognized by plants carrying the corresponding diseaseresistance gene (46). Activity of all avirulence genes tested sofar depends on a functional Hrp TTSS system, and more than10 avr genes elicit the HR when they are expressed within theresistant-plant cell, indicating that Avr proteins are translo-cated effector proteins (11).
Xanthomonas campestris pv. vesicatoria is the causal agent ofbacterial spot disease of peppers and tomatoes. The chromo-somal 23-kb hrp gene cluster contains six operons, hrpA to hrpF(9, 15, 16, 24; U. Bonas, unpublished data). Among the morethan 20 proteins encoded by the hrp region, nine are highlyconserved in plant and animal pathogens. These genes wererenamed hrc (hrp conserved) and are likely to form the core ofthe TTSS (8). Analysis of nonpolar mutants in the hrp operonsalso identified hpa (hrp-associated) genes, which contribute to,but are not essential for, the interaction with the plant (24; U.Bonas, unpublished data). We recently demonstrated Hrp-dependent secretion of HrpB2 and HrpF, which are encodedwithin the hrp gene cluster and which are essential for patho-genicity (35). Other secreted proteins that are not essential forpathogenicity are Xanthomonas outer proteins (Xop) such asXopB (34) and avirulence proteins AvrBs3, AvrBs4, AvrRxv,AvrBsT, and AvrBs1 (5, 14, 34, 36).
Expression of hrp genes is induced in plant leaves and inminimal medium XVM2 (38, 43) and is controlled by regula-tory genes hrpG and hrpX, located outside of the hrp genecluster. The HrpG protein belongs to the OmpR family oftwo-component regulatory systems and regulates the expres-sion of a large regulon, which was recently identified usingcDNA-amplified fragment length polymorphism (cDNA-AFLP) analysis. The expression profiles of two isogenic X.campestris pv. vesicatoria strains, 85-10 and 85*, which differ intheir hrp gene expression status, were compared (34). Strain
* Corresponding author. Mailing address: Institut für Genetik, Mar-tin-Luther-Universität Halle-Wittenberg, D-06099 Halle (Saale), Ger-many. Phone: (49) 345 5526290. Fax: (49) 345 5527277. E-mail:[email protected].
† Present address: Bayer-AG Zentrale Forschung, D-51368 Lever-kusen, Germany.
1340

85* expresses hrpG*, a mutated form of key regulatory genehrpG, which leads to the constitutive expression of hrp genes(45). Expression of the second known regulator, hrpX, is de-pendent on hrpG. HrpX, an AraC-type transcriptional activa-tor, controls expression of operons hrpB to hrpF (43) and isessential for expression of most new members of the hrpGregulon. Among the 35 known genes belonging to the hrpGregulon, seven hrpG-induced (hgi) cDNA fragments map tothe large hrp gene cluster (34) (Fig. 1A). Three of these hgigenes map to the hrp flanking regions, which had not previ-ously been characterized.
Here, we report the detailed analysis of both regions flank-ing the X. campestris pv. vesicatoria hrp gene cluster. The entireregion has features of a pathogenicity island (PAI) and en-codes novel proteins secreted by the Hrp TTSS.
MATERIALS AND METHODS
Bacterial strains, growth conditions, and plasmids. Bacterial strains and plas-mids used in this study are described in Table 1. Escherichia coli cells werecultivated at 37°C in Luria-Bertani medium, and X. campestris pv. vesicatoriastrains were cultivated at 30°C in NYG (12) or in hrp-inducing medium XVM2(43). Plasmids were introduced into E. coli by electroporation and into X.campestris pv. vesicatoria by conjugation, using pRK2013 as a helper plasmid intriparental matings as described previously (13, 17). Antibiotics were added tothe media at the following final concentrations: ampicillin, 100 �g/ml; kanamy-cin, 25 �g/ml; tetracycline, 10 �g/ml; rifampin, 100 �g/ml; spectinomycin, 100�g/ml.
Plant material and plant inoculations. Inoculation of the nearly isogenicpepper cultivars Early Cal Wonder (ECW) and ECW-10R, ECW-20R, andECW-30R, which carry the Bs1, Bs2, and Bs3 resistance genes, respectively, was
performed as described previously (9). Unless stated differently, bacterial sus-pensions were infiltrated into the leaf at 108 CFU/ml in 1 mM MgCl2. In plantagrowth in ECW was determined as described previously (9). Experiments wererepeated at least three times.
DNA and RNA analyses. To sequence the region downstream of hrpA, thesequence of pK2 was completed using an ABI 377 Prism DNA sequencer (Ap-plied Biosystems Inc., Foster City, Calif.). Deletions in both directions of thepBLB1 insert were generated using the Exo mung bean deletion kit (Stratagene,La Jolla, Calif.). A 3-kb HindIII fragment overlapping with pK2 and pBLB1 wascloned from pXV9 (9) into pBluescript II KS (pB-KS), giving pBLH1. Tosequence the 3.1-kb region flanking hrpF, the sequence of pBF (5.4-kb EcoRVfragment encompassing hrpF) was completed. The adjacent 2.3-kb EcoRV frag-ment was subcloned from pXV4 into pB-KS, giving pBLr5, and partially se-quenced.
Sequences were analyzed using the Sequencher program (Gene Codes Corp.,Ann Arbor, Mich.) and the DNASTAR package (DNASTAR Inc., Madison,Wis.). The tRNA gene was identified using the tRNAscan-SE algorithm (31).
RNA extraction, cDNA synthesis, and reverse transcription-PCR (RT-PCR)experiments were performed as described previously (34). Primer sequences areavailable upon request. Molecular biology experiments were performed accord-ing to standard procedures (4).
Generation of mutations in the genes downstream of hrpA. To introduce the�L deletion into strain 85*, suicide construct pOLB (34) was used, creating85*�L (24). A 1.0-kb deletion of ORF1, causing an out-of-frame (oof) mutationat codon 96, was achieved by BstEII digestion and religation of pULB1 (34),giving pU�ORF1. A 4.4-kb BamHI/ApaI fragment was cloned into suicide plas-mid pOK (24), giving pOORF1, which was introduced in strain 85-10, creating85-10�ORF1.
In hpaH, a frameshift mutation was introduced at codon 38 by digestion ofpULB1 by Csp45I, filling in, and religation, giving pUhpaH. The 5.6-kb BamHI/ApaI fragment of pUhpaH was subcloned into pOK, giving pOhpaH, which wasintroduced into strain 85-10, creating 85-10 hpaH-oof.
To delete xopA, the 2.7-kb NheI/ApaI fragment of pULB1 was cloned intopB-SK digested by SpeI/ApaI, creating pBxopA. pBxopA was partially digested
FIG. 1. Map of the large hrp gene cluster. (A) Schematic overview of the hrp operons hrpA to hrpF plus flanking regions. Gray circles, locationsof seven hgi fragments; black dots, PIP boxes. (B) Restriction map of the region downstream of hrpA. Arrows indicate the extent of deletionsgenerated. �, introduced frameshift mutations. (C) ORFs in the region downstream of hrpA. Single-headed arrows, ORFs with high codingprobability (DNAstar; Genequest, Pseudomonas aeruginosa codon preference matrix) and the direction of transcription; double-headed arrow,insertion sequence ISXc 7; �, frameshifts; black dots and open dots, PIP and hrp boxes, respectively; gray circles, hgi fragments. The most distalpart of the sequence does not contain ORFs with high coding probability (see also Table 2). (D) Comparison of the region shown in panel C tothe orthologous sequence in X. oryzae pv. oryzae (Xoo; GenBank accession no. AF026197 and AF232058) (48). Gray areas, noncolinear DNAregions between X. oryzae pv. oryzae and X. campestris pv. vesicatoria. Conserved sequences are more than 80% identical on the DNA level.
VOL. 184, 2002 hrp PATHOGENICITY ISLAND 1341

TABLE 1. Strains and plasmids used in this study
Strain or plasmid Relevant characteristicsa Reference or source
X. campestris pv. vesicatoria85-10 Pepper race 2; wild type; expresses avrBs1 and avrBs2; Rifr 1085-10�hrpG 85-10 derivative; carries a nonpolar hrpG deletion; Rifr 4585-10�R 85-10 derivative; carries a deletion from hpaF to hpaG; Rifr 3485-10�L 85-10 derivative; carries a deletion from ORF1 to xopA; Rifr 3485-10�ORF1 85-10 derivative; carries a deletion in ORF1; Rifr This study85-10�hpaH-oof 85-10 derivative; carries a frameshift mutation in hpaH; Rifr This study85-10�xopA 85-10 derivative; carries a deletion in xopA; Rifr This study85-10�xopD 85-10 derivative; carries a deletion and a frameshift mutation in xopD; Rifr This study85* 85-10 derivative containing the hrpG* mutation; constitutive hrp gene expression; Rifr 4582* 82-8 derivative containing the hrpG* mutation; constitutive hrp gene expression;
pepper race 1; Rifr45
85*�L 85* derivative; carries a deletion from ORF1 to xopA; Rifr This study85*�hrpX 85* derivative with a nonpolar hrpX deletion; Rifr 3485*�hrcV and 82*�hrcV 85* and 82* derivatives with a nonpolar hrcV deletion; nonfunctional TTSS; Rifr 35, 3685*::pICxopA 85* derivative containing pICxopA; expresses c-myc-tagged XopA; Rifr Tcr This study85*::pICxopD 85* derivative containing pICxopD; expresses c-myc-tagged XopD; Rifr Tcr This study85-10::pICxopA 85*::pICxopA derivative; wild-type hrpG restored using pOG; Rifr Tcr This study85-10::pICxopD 85*::pICxopD derivative; wild-type hrpG restored using pOG; Rifr Tcr This study
E. coliDH10B F� mcrA (mrr-hsdRMS-mcrBC) � 80d lacZ�M15 �lacX74 endA1 recA1 deoR D(ara,
leu)7697 ara�139 galU galK ��nupG rpsLBRL, Bethesda, Md.
DH5� F� �80d lacZ�M15 �(lacZYA-argF)U169 endA1 recA1 deoR hdsR17 (rk� mk
�) phoAsupE44 �� thi-1 gyrA96 relA1
BRL
DH5� �pir DH5� derivative, �pir; for replication of pIC1 and pOK derivatives 32
PlasmidspUC118 ColE1 replicon; Apr 42pULB1 pUC118 derivative with 7.5-kb BamHI insert from pXV331; contains ORF1 to xopD 34pB-KS, PB-SK Phagemid, pUC derivative; Apr StratagenepK2 pB-KS derivative containing a 4.5-kb insert of pXV9; carries hrpA and 1.5-kb
downstream region44
pBLB1 pB-KS derivative containing a 7.5-kb BamHI insert from pXV331; carries ORF1 toxopD
34
pBLH1 pB-KS derivative containing a 3.0-kb HindIII insert from pXV331 (xopD) 34pBF pB-KS derivative containing a 5.3-kb EcoRV insert from pXV2 containing hrpF 23pBL5r pB-KS derivative containing a 2.3-kb EcoRV insert from pXV4 containing hpaF and
hpaG23
pRK2013 Helper plasmid; ColE1 replicon TraRK� Mob�; Kmr 17pDS300F pDSK602 derivative carrying avrBs3 tagged with FLAG epitope at the C terminus;
broad host range; Smr41
pLAFR3 RK2 replicon Mob� Tra�; contains plac; Tcr 39pL3hpaFC pLAFR3 derivative carrying hpaF and hpaG; HpaF is c-myc tagged at amino acid
position 66This study
pXV331 pLAFR3 derivative; contains a region from downstream of ORF1 to hrpA from X.campestris pv. vesicatoria strain 75-3
34
pXV4 pLAFR3 derivative; contains a region from hrpC2 to hpaG from X. campestris pv.vesicatoria strain 75-3
43
pXV4::462 Tn 3-gus insertion derivative of pXV4; Tcr Kmr 43pXV9 pLAFR3 derivative; contains region from ISX c7 to hrpE from X. campestris pv.
vesicatoria strain 75-39
pOK Suicide vector; pKNG101 derivative; Smr/Sucs 24pOG pOK derivative; carries wild-type hrpG 34pOLB pOK derivative; carries a 3.7-kb insert with a 3.8-kb deletion from ORF1 to xopA 34pOR pOK derivative; carries a 5.7-kb insert with a 2.5-kb deletion from hpaF to hpaG 34pOORF1 pOK derivative; carries a 4.4-kb insert with a 1.0-kb deletion in ORF1 This studypOhpaH pOK derivative; carries a 5.6-kb insert with an out-of-frame mutation at codon
position 38 of hpaHThis study
pOxopA pOK derivative; carries a 2.4-kb insert with a 0.3-kb deletion in xopA This studypOxopD pOK derivative; carries a 1.85-kb insert with a 0.15-kb deletion in xopD causing an
out-of-frame mutation after codon position 311 of xopDThis study
pIC1 Integrative vector; carries a triple-c-myc epitope followed by a promoterless uidAgene; Tcr
34
pICxopA pIC1 derivative; carries the entire xopA ORF fused in frame to the c-myc epitope This studypICxopD pIC1 derivative; carries the entire xopD ORF fused in frame to the c-myc epitope This study
a Ap, ampicillin; Km, kanamycin; Rif, rifampin; Sm, spectinomycin; Suc, sucrose; Tc, tetracycline; r, resistant; s, sensitive.
1342 NOËL ET AL. J. BACTERIOL.

by ClaI and SalI, filled in, and religated, resulting in pB�xopA. In this construct,xopA (354 bp) contains a 312-bp deletion leaving only the first 11 amino acids ofthe protein unchanged. The 2.4-kb ApaI/XbaI insert was subcloned into pOK,giving pOxopA, which was introduced into 85-10, creating 85-10�xopA.
To mutate xopD, the 2.0-kb ApaI/BamHI fragment of pULB1 was subclonedinto pB-SK, giving pBxopD. pBxopD was digested by EcoRI, filled in, andreligated, giving pB�xopD. The 1.85-kb ApaI/BamHI insert of pB�xopD wascloned into pOK, giving pOxopD, which was introduced into 85-10, resulting in85-10�xopD, which carries an oof mutation after amino acid 311 of XopD.
GUS assays. �-Glucuronidase (GUS) assays were performed with exponen-tially growing X. campestris pv. vesicatoria as described previously (36). OneGUS unit is defined as 1 nmol of 4-methylumbelliferone released per min perbacterium.
Epitope tagging of XopA, XopD, and HpaF. A 0.4-kb fragment containingxopA and a 2.0-kb fragment encompassing xopD were amplified by PCR frompXV331 and cloned into pIC1 in frame with a triple-c-myc epitope, givingpICxopA and pICxopD, respectively. Primer sequences are available upon re-quest.
To tag hpaF, a 2.1-kb fragment starting 100 bp before the putative translationstart codon of hpaF was cloned into pB-KS, giving pBhpaF. A PCR-amplifiedtriple-c-myc tag was cloned in frame into the SalI site, which is situated 165 bpafter the putative translation start codon of hpaF, giving pBhpaFC. ThepBhpaFC insert was cloned into pLAFR3 under the control of the lac promoter,giving pL3hpaFC. Further cloning details are available upon request.
Protein analysis and secretion experiments. Secretion experiments and West-ern blot analyses were performed as described previously (36). The followingprimary antibodies were used: a polyclonal anti-AvrBs3 antibody (28), a mono-clonal anti-c-myc antibody (Roche, Mannheim, Germany; dilution 1:10,000),polyclonal anti-HrcN antiserum (36), and polyclonal anti-HrpF antiserum (D.Büttner and U. Bonas, unpublished data). Horseradish peroxidase-labeled goatanti-mouse or goat anti-rabbit antibodies were used as secondary antibodies.Reactions were visualized by enhanced chemiluminescence (Amersham Phar-macia Biotech, Piscataway, N.J.).
Nucleotide sequence accession numbers. The sequences of the 9.1-kb frag-ment downstream of hrpA and the 3.1-kb sequence downstream of hrpF from X.campestris pv. vesicatoria have been submitted to GenBank (U33548 andAF056246, respectively). The predicted HpaF protein sequence from Xanthomo-nas oryzae pv. oryzae has been added to the original submission (AB045312).
RESULTS
Sequence analysis of the region flanking hrpA. We recentlyidentified two hrpG- and hrpX-regulated genes (hgi27 andhgi81), which hybridized to a 7.5-kb BamHI fragment in theregion downstream of hrpA (34) (Fig. 1B). Sequence analysisof 9.1 kb flanking hrpA (see Materials and Methods) revealed
six open reading frames (ORFs) with high coding probability(Fig. 1C; Table 2).
The sequence containing hpaH (corresponds to hgi81) andxopA (corresponds to hgi27) revealed more than 80% homol-ogy to the orthologous sequence of X. oryzae pv. oryzae (Fig.1C and D). The predicted HpaH protein contains an N-termi-nal signal peptide of 34 amino acids and is almost identical toHpa2 from X. oryzae pv. oryzae (96% identity, 98% similarity)(48). Both proteins belong to the lysozyme-like protein family(33). The predicted XopA protein (Xanthomonas outer pro-tein) shows similarity to Hpa1 from X. oryzae pv. oryzae (46%identity, 59% similarity) (48). Perfect (TTCGC-N15-TTCGC)and imperfect (TTCGC-N15-TTCGT) PIP (plant-induciblepromoter) boxes are located 214 and 130 bp upstream of thepredicted translation start codons of xopA and hpaH, respec-tively. PIP boxes are found in promoter regions of X. campes-tris pv. vesicatoria hrp operons (16, 43).
The three genes located between xopA and hrpA do not have
FIG. 2. Genetic organization of the region downstream of hrpF.(A) Open arrows, ORFs; black arrow, tRNAArg gene; gray circles, hgifragments; triangle, position and orientation of Tn3-gus insertion no.462 (43); double-headed arrow, �R deletion. (B) Schematic drawing ofthe orthologous sequence in X. oryzae pv. oryzae (GenBank accessionno. AB045312). Black dots, PIP boxes (TTCGC-N16-TTCGC in X.campestris pv. vesicatoria and TTCGC-N16-TTCAC in X. oryzae pv.oryzae); gray areas, DNA sequences with more than 90% identity tothe X. campestris pv. vesicatoria region.
TABLE 2. Description of genes in the regions flanking the hrp gene cluster of X. campestris pv. vesicatoria
Gene Closest protein homolog(organism; accession no.)a
Identity/similarityb
(%) Comments
Downstream of hrpAORF1 340 aaf
hpaH Hpa2 (X. oryzae pv. oryzae; AAF61278) 96/98 Imperfect PIP box; hgi8Ig; 157 aaxopA Hpa1 (X. oryzae pv. oryzae; AAC95121) 46/59 PIP box; hgi27g; 50% G�C content; 117 aaORF4 Putative transposase (Y. pestis; CAB54971) 50/78 Conserved frameshifting site (CAAAAAAA)xopD PsvA (P. syringae pv. eriobotryae; BAA87062) 65/74c hrp box; 54% G�C content; 612 aa; NLSh
ORF6 Putative transposase (X. campestris pv.mangiferaeindicae; AF249895)
74/78 Frameshift; part of ISXc 7
Downstream of hrpFhpaF HpaF (X. oryzae pv. oryzae; AB045312) 81/87d PIP box; hgi203g; 197 aa; NLShpaG HpaF (X. oryzae pv. oryzae; AB045312) 87/93e 432 aa; LRRs
a Closest homologous protein using the BlastX algorithm (37) in nonredundant databases.b Identities and similarities between the respective proteins were determined by using the Blast 2 Sequences algorithm (40).c Refers to the last 300 amino acids of XopD and PsvA.d Refers to the first 180 amino acids of HpaF from X. oryzae pv. oryzae.e Refers to the last 430 amino acids of HpaF from X. oryzae pv. oryzae.f aa, amino acids; refers to the predicted size of the encoded protein.g hrpG-induced cDNA fragment; gene identified by cDNA-AFLP (34).h Putative NLS.
VOL. 184, 2002 hrp PATHOGENICITY ISLAND 1343
orthologs in X. oryzae pv. oryzae (Fig. 1; Table 2). Interestingly,the C-terminal portion of XopD contains a putative nuclearlocalization signal (NLS; KKKK) at amino acid position 602and shows 74% similarity to the last 300 amino acids of viru-lence factor PsvA from P. syringae pv. eriobotryae (26). ThexopD promoter does not contain a PIP box, but rather a hrpbox, which is found in all hrpL-dependent promoters in P.syringae and Erwinia spp. (GGAACTNA-N13-CGACNNA;consensus: GGAACcNa-N13/14-cCACNNA; lowercase lettersindicate less-well-conserved bases [25]). ORF4 is predicted toencode a protein with similarity to transposases from X.campestris and Yersinia pestis. However, the sequence conser-vation is restricted to the N-terminal 84 amino acids and thencontinues in the product of another reading frame. Finally,ORF6, which is interrupted by a stop codon at codon position121, encodes a protein highly similar to the putative trans-posase encoded by insertion sequence (IS) IS1595 of X.campestris pv. mangiferaeindicae. Since ORF6 is flanked by23-bp inverted repeats and 2-bp direct repeats, it has featurestypical of an IS element and was designated ISXc 7.
Sequence analysis of the region flanking hrpF. Sequenceanalysis of the 3.1-kb region downstream of hrpF, which con-tains hgi203, revealed two ORFs, hpaF and hpaG, and a puta-tive tRNAArg gene (anticodon CCG; Fig. 2). hrpF and theregion downstream up to the tRNA gene are highly similar tothe orthologous region of X. oryzae pv. oryzae. Curiously, in X.oryzae pv. oryzae, there is only one ORF, which we designatedhpaF. X. campestris pv. vesicatoria cDNA-AFLP fragmenthgi203 was derived from hpaF, which encodes a putative pro-tein of 197 amino acids containing a putative NLS (RPRRR)at amino acid 43. hpaG encodes a putative protein of 432amino acids, which contains six leucine-rich repeats (LRRs;consensus sequence, LXXLXXLXXLXLXXXXXLXXLPX X).HpaF is 87% similar to the N terminus of HpaF from X. oryzae pv.oryzae, and HpaG is 93% similar to its C terminus. The DNAsequences of hpaF and hpaG from X. campestris pv. vesicatoriaand hpaF from X. oryzae pv. oryzae mainly differ by a 7-bp dele-tion in the 3 region of hpaF in X. campestris pv. vesicatoria. Thisdeletion leads to a frameshift in hpaF which is present in all threedifferent X. campestris pv. vesicatoria strains tested (data notshown). An imperfect PIP box (TTCGC-N16-TTCGC) is located82 bp upstream of the predicted translation start codon of hpaF inX. campestris pv. vesicatoria.
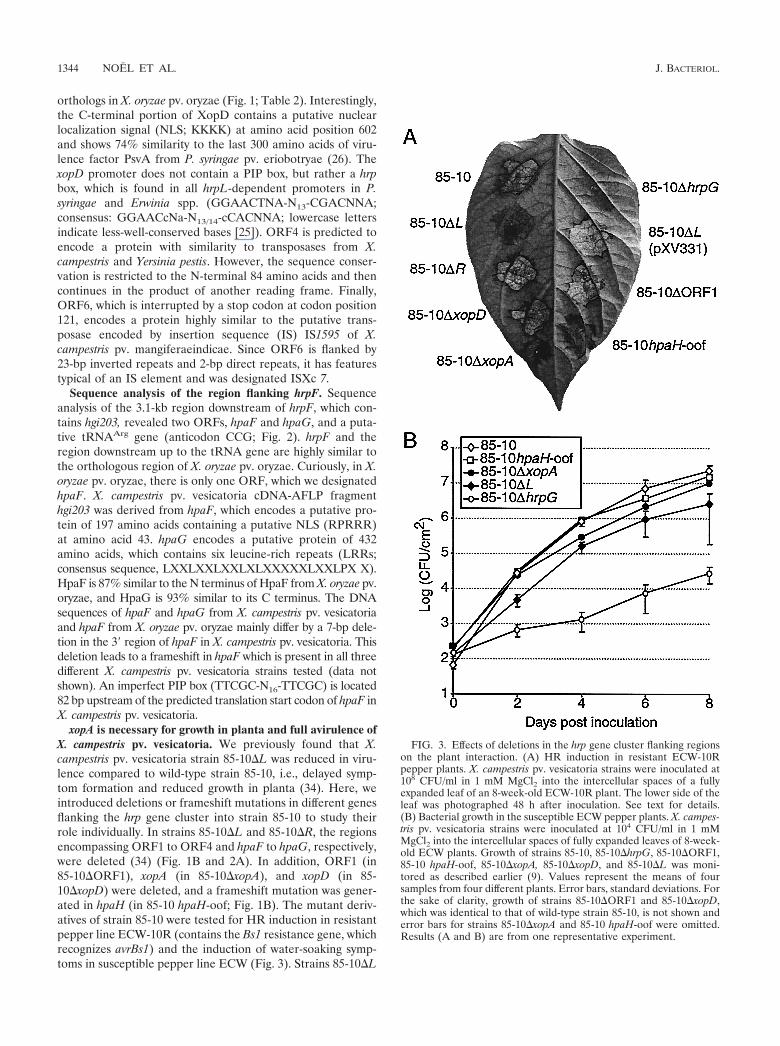
xopA is necessary for growth in planta and full avirulence ofX. campestris pv. vesicatoria. We previously found that X.campestris pv. vesicatoria strain 85-10�L was reduced in viru-lence compared to wild-type strain 85-10, i.e., delayed symp-tom formation and reduced growth in planta (34). Here, weintroduced deletions or frameshift mutations in different genesflanking the hrp gene cluster into strain 85-10 to study theirrole individually. In strains 85-10�L and 85-10�R, the regionsencompassing ORF1 to ORF4 and hpaF to hpaG, respectively,were deleted (34) (Fig. 1B and 2A). In addition, ORF1 (in85-10�ORF1), xopA (in 85-10�xopA), and xopD (in 85-10�xopD) were deleted, and a frameshift mutation was gener-ated in hpaH (in 85-10 hpaH-oof; Fig. 1B). The mutant deriv-atives of strain 85-10 were tested for HR induction in resistantpepper line ECW-10R (contains the Bs1 resistance gene, whichrecognizes avrBs1) and the induction of water-soaking symp-toms in susceptible pepper line ECW (Fig. 3). Strains 85-10�L
FIG. 3. Effects of deletions in the hrp gene cluster flanking regionson the plant interaction. (A) HR induction in resistant ECW-10Rpepper plants. X. campestris pv. vesicatoria strains were inoculated at108 CFU/ml in 1 mM MgCl2 into the intercellular spaces of a fullyexpanded leaf of an 8-week-old ECW-10R plant. The lower side of theleaf was photographed 48 h after inoculation. See text for details.(B) Bacterial growth in the susceptible ECW pepper plants. X. campes-tris pv. vesicatoria strains were inoculated at 104 CFU/ml in 1 mMMgCl2 into the intercellular spaces of fully expanded leaves of 8-week-old ECW plants. Growth of strains 85-10, 85-10�hrpG, 85-10�ORF1,85-10 hpaH-oof, 85-10�xopA, 85-10�xopD, and 85-10�L was moni-tored as described earlier (9). Values represent the means of foursamples from four different plants. Error bars, standard deviations. Forthe sake of clarity, growth of strains 85-10�ORF1 and 85-10�xopD,which was identical to that of wild-type strain 85-10, is not shown anderror bars for strains 85-10�xopA and 85-10 hpaH-oof were omitted.Results (A and B) are from one representative experiment.
1344 NOËL ET AL. J. BACTERIOL.

and 85*�L (85-10�L derivative carrying the hrpG* mutation)both caused clearly delayed water-soaking symptoms in ECWpepper plants and delayed HR in ECW-10R pepper plants (12h to 24 h later than strains 85-10 and 85*, respectively) (Fig.3A; data not shown). While strains 85-10�R, 85-10�ORF1,and 85-10�xopD behaved like wild-type strain 85-10, strain85-10�xopA displayed an intermediate phenotype compared tostrains 85-10�L and 85-10, i.e., slightly delayed water-soakingsymptoms and HR. Strain 85-10 hpaH-oof induced sometimesa slightly delayed HR. The mutant phenotypes of strain 85-10�L in ECW and ECW-10R plants could be complementedby pXV331 (Fig. 3A; data not shown).
The reduced avirulence of strain 85-10�L is not specificallyassociated with the avrBs1-Bs1 interaction and was also ob-served in association with avrBs2 recognition in ECW-20Rplants (contains the Bs2 resistance gene; data not shown).Similarly, strain 85-10�L(pDS300F), which expresses theavrBs3 gene, induced a delayed and partial HR in the corre-sponding resistant ECW-30R plants (contains the Bs3 resis-
tance gene; data not shown). In addition, the necrosis normallyinduced by strain 85* in tobacco was weaker with 85*�L (datanot shown) (45).
To determine which gene is responsible for the reducedgrowth of strain 85-10�L in susceptible pepper line ECW, wedetermined the growth of mutants with mutations in individualgenes. Strain 85-10�ORF1 behaved like the wild type, whereasstrain 85-10�xopA always showed a reduction in growth com-pared to strain 85-10 with levels intermediate between those ofwild-type strain 85-10 and strain 85-10�L (Fig. 3B). Thegrowth of strain 85-10 hpaH-oof was not affected or was weaklyaffected. The mutation in xopD, which was not included in the�L deletion, did not affect growth in planta (data not shown).
In summary, these results indicate that the reduced aviru-lence and growth in planta of strain 85-10�L are essentiallydue to the loss of xopA but partially also due to the loss ofhpaH.
AvrBs3 and HrpF secretion is not affected in strain 85*�L.To test if the reduced-virulence and avirulence phenotypes ofthe deletion mutants are due to reduced secretion of effectorproteins by the Hrp system, we analyzed the secretion of twoTTSS substrates: AvrBs3, which is secreted by the X. campestrispv. vesicatoria TTSS (36), and HrpF, which is an essential,secreted pathogenicity protein that, it has been suggested,plays a role in translocation (35). Levels of in vitro secretion ofboth AvrBs3 and HrpF in strain 85*�L were comparable to thewild-type levels (Fig. 4). Thus, the genes deleted in strain85*�L do not encode components of the type III secretionmachinery.
xopD belongs to the hrpG regulon. We investigated the reg-ulation of gene expression in the two regions flanking the hrpgene cluster using different strains grown in complex mediumNYG or in hrp-inducing medium XVM2 (Fig. 5). The expres-sion of xopA (hgi27), hpaH (hgi81), and hpaF (hgi203) is strictlydependent on hrpG and hrpX and is induced in XVM2 (34);these findings were confirmed here by RT-PCR experiments(Fig. 5). The expression of ORF1 appears to be constitutiveand independent of hrpG and hrpX. Interestingly, xopD expres-sion is induced in a hrpG- and hrpX-dependent manner, indi-cating that xopD also belongs to the hrpG regulon (Fig. 5).Finally, primers that amplify a 520-bp fragment spanning hpaFand hpaG were used to detect a cDNA fragment of the ex-pected size. This shows that hpaG is regulated in the same wayas hpaF and indicates that the two genes are probably in thesame operon (Fig. 5).
FIG. 4. The �L deletion has no effect on type III secretion in vitro.X. campestris pv. vesicatoria strains expressing AvrBs3 from pDS300Fwere incubated in secretion medium. Total protein extracts (lanes 1 to3) and supernatants (lanes 4 to 6) of strains 85* (lanes 1 and 4),85*�hrcV (lanes 2 and 5), and 85*�L (lanes 3 and 6) were separatedby sodium dodecyl sulfate–10% polyacrylamide gel electrophoresisand analyzed by immunoblotting using AvrBs3- and HrpF-specificantibodies. Total protein extracts and supernatants were concentrated10- and 100-fold, respectively. Membranes were reprobed with theantiserum directed against cytoplasmic protein HrcN to assure that nobacterial lysis had occurred during the experiment (data not shown).
FIG. 5. Expression profiles of ORF1, hpaH, xopA, xopD, hpaF, andhpaG. Shown is RT-PCR analysis of X. campestris pv. vesicatoriastrains 85-10, 85*, and 85*�hrpX, grown in NYG, and of strains 85-10and 85-10�hrpG, grown in hrp gene induction medium XVM2. 16SrDNA was used as a standard. The DNA samples were separated on a1.5% agarose gel and stained with ethidium bromide.
TABLE 3. Promoter activities of xopA, xopD, and hpaF inX. campestris pv. vesicatoria strains 85-10 and 85*a
Reporter construct(gene)
Activity (GUS units) SDb in strain: Fold induction
(compared to 85-10)85-10 85*c
pICxopA (xopA) 1.2 0.2 24 3 20pICxopD (xopD) 0.24 0.06 1.9 0.4 8pXV4::462 (hpaF) 0.06 0.02 2.4 0.5 40
a Bacteria were grown in NYG medium.b One unit corresponds to 1 nmol of 4-methylumbelliferone released per min
per bacterium.c Carries the hrpG* mutation, which leads to constitutive hrp gene expression
(45).
VOL. 184, 2002 hrp PATHOGENICITY ISLAND 1345

The findings on transcriptional regulation were confirmed byusing GUS reporter constructs to monitor the promoter activ-ity of xopA, xopD, and hpaF (Table 3). To study hpaF promoteractivity, we used a Tn3-gus insertion (no. 462) in the 3 regionof hpaF, which was obtained in pXV4 (pXV4::462 [43]) andwhich was conjugated in 85-10 and 85*. For xopA and xopD, weused pIC1, a suicide vector, to simultaneously tag the C terminiof the proteins with a triple-c-myc tag (see below) and generatea transcriptional fusion with a promoterless uidA reporter gene(34). The resulting constructs, pICxopA and pICxopD, wereconjugated into 85*. To exclude the possibility that lack ofexpression is due to an aberrant insertion of the pIC con-structs, 85-10::pICxopA and 85-10::pICxopD were generatedby restoring the wild-type hrpG allele in 85*::pICxopA and85*::pICxopD, respectively, using pOG (34). Analysis of GUSactivities showed that xopA, xopD, and hpaF expression was 20,8, and 40 times higher, respectively, in strains expressing hrpG*than in the hrpG background (Table 3). These results confirmthe hrpG-dependent regulation of xopA, xopD, and hpaF (Fig.5).
Secretion of XopA and XopD by the Xanthomonas Hrp TTSS.Because it was proposed that Hpa1, the XopA homolog in X.oryzae pv. oryzae, acts as a harpin (48) (see Discussion) andbecause both XopD and HpaF contain putative NLSs, all threeproteins are possible substrates for the TTSS. To allow detec-tion, the proteins were tagged with a triple-c-myc epitope. Asdescribed above, integrative vector pIC1 allowed C-terminaltagging of XopA and XopD. To detect HpaF, a triple-c-myctag was inserted after amino acid 55 and the gene was clonedinto the pLAFR3 vector under the control of the lac promoter,which is constitutive in X. campestris pv. vesicatoria, givingpL3hpaFC. XopA and XopD expression was analyzed in
strains 85* and 85*�hrcV, a strain, which carries a deletion ina conserved component of the TTSS. For technical reasons,expression studies of HpaF were carried out in 82* and deriv-atives (Table 1).
Western blot analysis of bacterial total protein extracts re-vealed that, as expected from the promoter studies, XopA–c-myc and XopD–c-myc were not detectable in strain 85-10grown in NYG (Fig. 6A). However, a protein of 18 kDa,corresponding to the expected size for XopA–c-myc (13 kDaplus a 5-kDa epitope), was detected in extracts of85*::pICxopA. XopD–c-myc (predicted size: 70 plus 5 kDa)was detected at approximately 110 kDa in 85*::pICxopD.HpaF–c-myc (predicted size: 21 plus 5 kDa) could be visual-ized at approximately 33 kDa in total bacterial protein extracts(data not shown). To test for hrp-dependent secretion, bacteriawere incubated in secretion medium. While HpaF could not bedetected in culture supernatants (data not shown), XopA andXopD were present in culture supernatants of 85* strains (Fig.6B). Since XopA and XopD were undetectable in culture su-pernatants of TTSS mutant 85*�hrcV, we conclude that XopAand XopD are secreted by the Hrp TTSS.
DISCUSSION
In this study we investigated the role of genes induced byhrpG and hrpX in the regions flanking the hrp gene cluster ofX. campestris pv. vesicatoria and identified new Xop proteinsand virulence factors. Nonpolar mutations in four genes re-vealed that xopA and hpaH contribute to bacterial growth andinduction of plant reactions. The predicted HpaH protein be-longs to the family of lysozyme-like proteins (33). The mostinteresting features of members of this protein family, besidesan N-terminal signal peptide, are conserved lytic transglycosy-lase motifs and association with type III or type IV transportmachinery. It is conceivable that the establishment of the typeIII transport machinery requires remodeling of the periplasmicpeptidoglycans and that HpaH plays a role similar to thatproposed for VirB1 from A. tumefaciens and related proteins(33). While analyses of mutants revealed reduced tumorigen-esis by the A. tumefaciens virB1 mutant (7), mutations in hpaHand hpa2 (48) from Xanthomonas had only a weak effect(hpaH) or no effect (hpa2) on the interaction with the host.
The second virulence factor, XopA, from X. campestris pv.vesicatoria is similar only to the predicted Hpa1 protein fromX. oryzae pv. oryzae, which also plays a role in disease insusceptible plants and HR induction in resistant plants (48).Here we show that xopA encodes a protein that is secreted bythe Hrp TTSS. This finding, together with the fact that xopAdeletion mutants are reduced in virulence without affectingtype III-dependent in vitro secretion, suggests that XopAcould be involved in effector protein translocation into the hostcell. Whether XopA plays a role before or after its secretionand whether it is itself translocated into the host cell remain tobe elucidated. Interestingly, both the predicted Hpa1 proteinfrom X. oryzae pv. oryzae and PopA, a secreted harpin from R.solanacearum (3), are very glycine rich (more than 20% of theamino acid residues), a feature that is typical for harpins fromplant pathogenic bacteria (48). Harpins are secreted by theHrp TTSS and trigger an HR-like response when infiltratedinto nonhost plants (11), and recent data suggest an interaction
FIG. 6. XopA and XopD are secreted by the Hrp TTSS. (A) Ex-pression of XopA and XopD is hrpG dependent. pICxopA and pICx-opD were conjugated into 85-10 and 85* to study the expression ofXopA–c-myc and XopD–c-myc, respectively. Total protein extracts ofX. campestris pv. vesicatoria grown in NYG were separated by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE; 15and 8% polyacrylamide for XopA and XopD, respectively) and ana-lyzed by immunoblotting using an anti-c-myc antibody. The approxi-mate sizes of the proteins are shown at the right. (B) Total proteinextracts (10-fold concentrated) and supernatants (200-fold concen-trated) of strains 85*�hrcV and 85*, containing pICxopA or pICxopD,were separated by SDS-PAGE (15 or 8% polyacrylamide for XopAand XopD, respectively) and analyzed by immunoblotting using thec-myc antibody. Membranes were reprobed with antiserum directedagainst cytoplasmic protein HrcN to assure that no bacterial lysis hadoccurred during the experiment (data not shown).
1346 NOËL ET AL. J. BACTERIOL.

of the P. syringae harpin with the plant plasma membrane (29,30). Although XopA (8% glycine) is not homologous toharpins, it shares a lack of cysteine residues and an acidicisoelectric point with PopA and Hpa1 and is secreted by theHrp system, which was also shown for PopA (3). Preliminaryresults indicate that infiltration of an XopA–glutathione S-transferase fusion protein into tobacco leaves did not inducean HR-like response.
Another Xop identified in this study is XopD, which is sim-ilar to PsvA from P. syringae pv. eriobotryae but not to pre-dicted proteins in the sequenced genomes of P. syringae pv.tomato DC3000 and R. solanacearum (http://www.tigr.org/cgi-bin/BlastSearch/blast.cgi?organism�p_syringae and http://sequence.toulouse.inra.fr/ralsto/public/doc/RalstoForm.html). PsvA plays a role in virulence on loquat trees (26),whereas the pathogenicity of the X. campestris pv. vesicatoriaxopD mutant was indistinguishable from that of the wild typeunder the conditions used. Interestingly, both XopD and PsvAcontain a putative NLS, suggesting that they might be translo-cated to the plant nucleus to meet their virulence targets.Surprisingly, the xopD promoter contains an hrp box, which istypical for hrpL-regulated promoters in pathovars of P. syringaeand Erwinia spp. (25, 47) and which is also found in the pre-dicted promoter of the psvA gene from P. syringae pv. eriobot-ryae (GGAACCNA-N13-CCTACTA) 240 bp upstream of thepredicted translation start codon. This is the first description ofan hrp box in the promoter of an X. campestris pv. vesicatoriagene and is indicative of horizontal transfer of xopD fromPseudomonas or Erwinia spp. It would be interesting to knowwhether xanthomonads contain hrpL homologs and whetherxopD could be regulated by hrpL in P. syringae.
On the other side of the hrp gene cluster, downstream ofhrpF, we identified ORFs hpaF and hpaG, which belong to thehrpG regulon. Since deletion mutations have no effect, theirrole for the bacterium-plant interaction is unknown. Bothgenes are homologous to the single hpaF gene in X. oryzae pv.oryzae, which is predicted to encode a protein with nine LRRs,a feature found in many eukaryotic proteins but also in anumber of type III effector proteins, e.g., PopC from R. so-lanacearum (21). The presence of LRRs in the predicted X.oryzae pv. oryzae HpaF protein is suggestive of secretion by theHrp system. However our attempts to detect HpaG-His6 in X.campestris pv. vesicatoria were unsuccessful, suggesting thatthe hpaG RNA is not translated under the conditions used (F.Thieme and U. Bonas, unpublished data).
DNA sequence comparison of the X. campestris pv. vesica-toria and X. oryzae pv. oryzae hrp gene clusters and flankingregions revealed almost identical sequences in the core hrpgene clusters and downstream of hrpF. While the regionsflanking hrpA in X. campestris pv. campestris and X. oryzae pv.oryzae show very similar organizations (M. Arlat, personalcommunication), the orthologous region in X. campestris pv.vesicatoria has a scrambled organization, probably due to de-letions and insertions. The G�C contents of xopA (50%) andxopD (54%) are significantly lower than that of the hrp genecluster (64%), suggesting that xopA and xopD might have beenacquired by horizontal transfer. In addition, the Xanthomonasregions flanking the hrp gene cluster contain genes involved invirulence (xopA and hpaH), a tRNA gene, and mobile geneticelements (ISXc 7, IS1114, transposase coding sequence). Tak-
ing into account that mobility of the hrp gene cluster has beenobserved in X. campestris pv. vesicatoria (6), the region con-taining the hrp genes fulfills the criteria of PAIs (22). Similarconclusions were drawn for P. syringae (1). Sequencing or-thologous regions from different P. syringae pathovars revealedan exchangeable effector locus (EEL) differing in G�C andgene content and containing sequences related to mobile ge-netic elements, whereas the region on the other side of the hrpgene cluster is highly conserved (conserved effector locus;CEL) (1). Genome sequencing of different xanthomonads willshow whether the EEL/CEL concept is also valid for Xan-thomonas spp. and will address questions of diversity and dy-namics of the PAIs.
ACKNOWLEDGMENTS
We are grateful to J. Mansfield, D. Büttner, R. Koebnik, and T.Lahaye for critically reading the manuscript and to M. Arlat for com-municating unpublished sequences. We thank C. Kretschmer and A.Landgraf for excellent technical assistance and E. Huguet for identi-fying hpaG and hpaI.
L.N. was supported in part by the École Normale Supérieure deLyon. This work was funded by a grant from the Deutsche Forschungs-gemeinschaft (SFB 363) to U.B.
REFERENCES
1. Alfano, J. R., A. O. Charkowski, W. L. Deng, J. L. Badel, T. Petnicki-Ocwieja,K. van Dijk, and A. Collmer. 2000. The Pseudomonas syringae Hrp patho-genicity island has a tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector and conserved effectorloci that contribute to parasitic fitness and pathogenicity in plants. Proc. Natl.Acad. Sci. USA 97:4856–4861.
2. Alfano, J. R., and A. Collmer. 1997. The type III (Hrp) secretion pathway ofplant pathogenic bacteria: trafficking harpins, Avr proteins, and death. J.Bacteriol. 179:5655–5662.
3. Arlat, M., F. Van Gijsegem, J. C. Huet, J. C. Pernollet, and C. A. Boucher.1994. PopA1, a protein which induces a hypersensitivity-like response onspecific Petunia genotypes, is secreted via the Hrp pathway of Pseudomonassolanacearum. EMBO J. 13:543–553.
4. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl (ed.). 1996. Current protocols in molecular biology.John Wiley & Sons, Inc., New York, N.Y.
5. Ballvora, A., M. Pierre, G. van den Ackerveken, S. Schornack, O. Rossier, M.Ganal, T. Lahaye, and U. Bonas. 2001. Genetic mapping and functionalanalysis of the tomato Bs4 locus, governing recognition of the Xanthomonascampestris pv. vesicatoria AvrBs4 protein. Mol. Plant-Microbe Interact. 14:629–638.
6. Basim, H., R. E. Stall, G. V. Minsavage, and J. B. Jones. 1999. Chromosomalgene transfer by conjugation in the plant pathogen Xanthomonas axonopodispv. vesicatoria. Phytopathology 89:1044–1049.
7. Berger, B. R., and P. J. Christie. 1994. Genetic complementation analysis ofthe Agrobacterium tumefaciens virB operon: virB2 through virB11 are essen-tial virulence genes. J. Bacteriol. 176:3646–3660.
8. Bogdanove, A., S. V. Beer, U. Bonas, C. A. Boucher, A. Collmer, D. L. Coplin,G. R. Cornelis, H.-C. Huang, S. W. Hutcheson, N. J. Panopoulos, and F. VanGijsegem. 1996. Unified nomenclature for broadly conserved hrp genes ofphytopathogenic bacteria. Mol. Microbiol. 20:681–683.
9. Bonas, U., R. Schulte, S. Fenselau, G. V. Minsavage, B. J. Staskawicz, andR. E. Stall. 1991. Isolation of a gene-cluster from Xanthomonas campestrispv. vesicatoria that determines pathogenicity and the hypersensitive responseon pepper and tomato. Mol. Plant-Microbe Interact. 4:81–88.
10. Bonas, U., R. E. Stall, and B. Staskawicz. 1989. Genetic and structuralcharacterization of the avirulence gene avrBs3 from Xanthomonas campestrispv. vesicatoria. Mol. Gen. Genet. 218:127–136.
11. Cornelis, G. R., and F. Van Gijsegem. 2000. Assembly and function of typeIII secretory systems. Annu. Rev. Microbiol. 54:735–774.
12. Daniels, M. J., C. E. Barber, P. C. Turner, M. K. Sawczyc, R. J. W. Byrde,and A. H. Fielding. 1984. Cloning of genes involved in pathogenicity ofXanthomonas campestris pv. campestris using the broad host range cosmidpLAFR1. EMBO J. 3:3323–3328.
13. Ditta, G., S. Stanfield, D. Corbin, and D. Helinski. 1980. Broad host rangeDNA cloning system for gram-negative bacteria: construction of a gene bankof Rhizobium meliloti. Proc. Natl. Acad. Sci. USA 77:7347–7351.
14. Escolar, L., G. Van den Ackerveken, S. Pieplow, O. Rossier, and U. Bonas.2001. Type III secretion and in planta recognition of the Xanthomonasavirulence proteins AvrBs1 and AvrBsT. Mol. Plant Pathol. 2:287–296.
VOL. 184, 2002 hrp PATHOGENICITY ISLAND 1347

15. Fenselau, S., I. Balbo, and U. Bonas. 1992. Determinants of pathogenicity inXanthomonas campestris pv. vesicatoria are related to proteins involved insecretion in bacterial pathogens of animals. Mol. Plant-Microbe Interact.5:390–396.
16. Fenselau, S., and U. Bonas. 1995. Sequence and expression analysis of thehrpB pathogenicity operon of Xanthomonas campestris pv. vesicatoria whichencodes eight proteins with similarity to components of the Hrp, Ysc, Spa,and Fli secretion systems. Mol. Plant-Microbe Interact. 8:845–854.
17. Figurski, D., and D. R. Helinski. 1979. Replication of an origin-containingderivative of plasmid RK2 dependent on a plasmid function provided intrans. Proc. Natl. Acad. Sci. USA 76:1648–1652.
18. Galán, J. E., and A. Collmer. 1999. Type III secretion machines: bacterialdevices for protein delivery into host cells. Science 284:1322–1328.
19. Gaudriault, S., L. Malandrin, J. P. Paulin, and M. A. Barny. 1997. DspA, anessential pathogenicity factor of Erwinia amylovora showing homology withAvrE of Pseudomonas syringae, is secreted via the Hrp secretion pathway ina DspB-dependent way. Mol. Microbiol. 26:1057–1069.
20. Gelvin, S. B. 2000. Agrobacterium and plant genes involved in T-DNA trans-fer and integration. Annu. Rev. Plant Physiol. Plant Mol. Biol. 51:223–256.
21. Guénéron, M., A. C. Timmers, C. Boucher, and M. Arlat. 2000. Two novelproteins, PopB, which has functional nuclear localization signals, and PopC,which has a large leucine-rich repeat domain, are secreted through theHrp-secretion apparatus of Ralstonia solanacearum. Mol. Microbiol. 36:261–277.
22. Hacker, J., and J. B. Kaper. 2000. Pathogenicity islands and the evolution ofmicrobes. Annu. Rev. Microbiol. 54:641–679.
23. Huguet, E., and U. Bonas. 1997. hrpF of Xanthomonas campestris pv. vesi-catoria encodes an 87-kDa protein with homology to NolX of Rhizobiumfredii. Mol. Plant-Microbe Interact. 10:488–498.
24. Huguet, E., K. Hahn, K. Wengelnik, and U. Bonas. 1998. hpaA mutants ofXanthomonas campestris pv. vesicatoria are affected in pathogenicity butretain the ability to induce host-specific hypersensitive reaction. Mol. Micro-biol. 29:1379–1390.
25. Innes, R. W., A. F. Bent, B. N. Kunkel, S. R. Bisgrove, and B. J. Staskawicz.1993. Molecular analysis of avirulence gene avrRpt2 and identification of aputative regulatory sequence common to all known Pseudomonas syringaeavirulence genes. J. Bacteriol. 175:4859–4869.
26. Kamiunten, H. 1999. Isolation and characterization of virulence gene psvAon a plasmid of Pseudomonas syringae pv. eriobotryae. Ann. Phytopathol.Soc. Jpn. 65:501–509.
27. Klement, Z. 1982. Hypersensitivity, p. 149–177. In M. S. Mount and G. H.Lacy (ed.), Phytopathogenic prokaryotes, vol. 2. Academic Press, New York,N.Y.
28. Knoop, V., B. Staskawicz, and U. Bonas. 1991. Expression of the avirulencegene avrBs3 from Xanthomonas campestris pv. vesicatoria is not under thecontrol of hrp genes and is independent of plant factors. J. Bacteriol. 173:7142–7150.
29. Lee, J., D. F. Klessig, and T. Nürnberger. 2001. A harpin binding site intobacco plasma membranes mediates activation of the pathogenesis-relatedgene hin1 independent of extracellular calcium but dependent on mitogen-activated protein kinase activity. Plant Cell 13:1079–1093.
30. Lee, J., B. Klusener, G. Tsiamis, C. Stevens, C. Neyt, A. P. Tampakaki, N. J.Panopoulos, J. Noller, E. W. Weiler, G. R. Cornelis, J. W. Mansfield, and T.Nürnberger. 2001. HrpZPsph from the plant pathogen Pseudomonas syringaepv. phaseolicola binds to lipid bilayers and forms an ion-conducting pore invitro. Proc. Natl. Acad. Sci. USA 98:289–294.
31. Lowe, T. M., and S. R. Eddy. 1997. tRNAscan-SE: a program for improved
detection of transfer RNA genes in genomic sequence. Nucleic Acids Res.25:955–964.
32. Meinhardt, L. W., H. B. Krishnan, P. A. Balatti, and S. G. Pueppke. 1993.Molecular cloning and characterization of a sym plasmid locus that regulatescultivar-specific nodulation of soybean by Rhizobium fredii USDA257. Mol.Microbiol. 9:17–29.
33. Mushegian, A. R., K. J. Fullner, E. V. Koonin, and E. W. Nester. 1996. Afamily of lysozyme-like virulence factors in bacterial pathogens of plants andanimals. Proc. Natl. Acad. Sci. USA 93:7321–7326.
34. Noël, L., F. Thieme, D. Nennstiel, and U. Bonas. 2001. cDNA-AFLP analysisunravels a genome-wide hrpG-regulon in the plant pathogen Xanthomonascampestris pv. vesicatoria. Mol. Microbiol. 41:1271–1281.
35. Rossier, O., G. Van den Ackerveken, and U. Bonas. 2000. HrpB2 and HrpFfrom Xanthomonas are type III-secreted proteins and essential for pathoge-nicity and recognition by the host plant. Mol. Microbiol. 38:828–838.
36. Rossier, O., K. Wengelnik, K. Hahn, and U. Bonas. 1999. The XanthomonasHrp type III system secretes proteins from plant and mammalian pathogens.Proc. Natl. Acad. Sci. USA 96:9368–9373.
37. Schaffer, A. A., L. Aravind, T. L. Madden, S. Shavirin, J. L. Spouge, Y. I.Wolf, E. V. Koonin, and S. F. Altschul. 2001. Improving the accuracy ofPSI-BLAST protein database searches with composition-based statistics andother refinements. Nucleic Acids Res. 29:2994–3005.
38. Schulte, R., and U. Bonas. 1992. Expression of the Xanthomonas campestrispv. vesicatoria hrp gene cluster, which determines pathogenicity and hyper-sensitivity on pepper and tomato, is plant inducible. J. Bacteriol. 174:815–823.
39. Staskawicz, B. J., D. Dahlbeck, N. Keen, and C. Napoli. 1987. Molecularcharacterization of cloned avirulence genes from race0 and race1 of Pseudo-monas syringae pv. glycinea. J. Bacteriol. 169:5789–5794.
40. Tatusova, T. A., and T. L. Madden. 1999. Blast 2 sequences–-a new tool forcomparing protein and nucleotide sequences. FEMS Microbiol. Lett. 174:247–250.
41. Van den Ackerveken, G., E. Marois, and U. Bonas. 1996. Recognition of thebacterial avirulence protein AvrBs3 occurs inside the host plant cell. Cell87:1307–1316.
42. Vieira, J., and J. Messing. 1987. Production of single-stranded plasmidDNA. Methods Enzymol. 153:3–11.
43. Wengelnik, K., and U. Bonas. 1996. HrpXv, an AraC-type regulator, acti-vates expression of five of the six loci in the hrp cluster of Xanthomonascampestris pv. vesicatoria. J. Bacteriol. 178:3462–3469.
44. Wengelnik, K., C. Marie, M. Russel, and U. Bonas. 1996. Expression andlocalization of HrpA1, a protein of Xanthomonas campestris pv. vesicatoriaessential for pathogenicity and induction of the hypersensitive reaction. J.Bacteriol. 178:1061–1069.
45. Wengelnik, K., O. Rossier, and U. Bonas. 1999. Mutations in the regulatorygene hrpG of Xanthomonas campestris pv. vesicatoria result in constitutiveexpression of all hrp genes. J. Bacteriol. 181:6828–6831.
46. White, F. F., B. Yang, and L. B. Johnson. 2000. Prospects for understandingavirulence gene function. Curr. Opin. Plant Biol. 3:291–298.
47. Xiao, Y., and S. W. Hutcheson. 1994. A single promoter sequence recognizedby a newly identified alternate sigma factor directs expression of pathoge-nicity and host range determinants in Pseudomonas syringae. J. Bacteriol.176:3089–3091. (Erratum, 176:6158.)
48. Zhu, W. G., M. M. Magbanua, and F. F. White. 2000. Identification of twonovel hrp-associated genes in the hrp gene cluster of Xanthomonas oryzae pv.oryzae. J. Bacteriol. 182:1844–1853.
1348 NOËL ET AL. J. BACTERIOL.




















