
Fast, Precise, and Reliable Multiplex Detection of Potato ...
Upload
independentCategory
view
5download
0
Accepted Manuscript
Trypanosoma cruzi: Multiplex PCR to Detect and Classify Strains according to
groups I and II1
Daniel B. Liarte, Silvane M.F. Murta, Mario Steindel, Alvaro J. Romanha
PII: S0014-4894(08)00320-2
DOI: 10.1016/j.exppara.2008.12.005
Reference: YEXPR 5672
To appear in: Experimental Parasitology
Received Date: 27 June 2008
Revised Date: 4 December 2008
Accepted Date: 5 December 2008
Please cite this article as: Liarte, D.B., Murta, S.M.F., Steindel, M., Romanha, A.J., Trypanosoma cruzi: Multiplex
PCR to Detect and Classify Strains according to groups I and II1, Experimental Parasitology (2008), doi: 10.1016/
j.exppara.2008.12.005
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers
we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and
review of the resulting proof before it is published in its final form. Please note that during the production process
errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

ACCEPTED MANUSCRIPT 1
Trypanosoma cruzi: Multiplex PCR to Detect and Classify 1
Strains according to groups I and II12
3
4
5
Daniel B. Liarte a, Silvane M.F. Murta a, Mario Steindel b, Alvaro J. Romanha a6
7
8
a Laboratório de Parasitologia Celular e Molecular, Centro de Pesquisas René Rachou-9
Fiocruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, Minas Gerais, Brazil10
11
b Departamento de Microbiologia e Parasitologia, Universidade Federal de Santa Catarina,12
88040-900 Florianópolis, Santa Catarina, Brazil.13
14
15
Corresponding author: Laboratório de Parasitologia Celular e Molecular, Centro de 16
Pesquisas René Rachou – Fiocruz. Av. Augusto de Lima 1715, 17
30190-002 Belo Horizonte, Minas Gerais, Brazil.18
Tel.: +55 31 3349 7786; fax: +55 31 3295 311519
E-mail address: [email protected] (Daniel B. Liarte)20
21
1The nucleotide sequences reported in this paper are available in the GenBank 22
database under the accession numbers: DQ91442-521.23
24

ACCEPTED MANUSCRIPT 2
Abstract25
A multiplex PCR was developed for simultaneous detection of T. cruzi DNA and 26
classification of the parasite strain into groups I and II. As little as 10 fg of T. cruzi DNA 27
could be detected by multiplex PCR. The technique was shown to be specific for T. cruzi 28
DNA, since no PCR amplification products were obtained with DNA from other29
tripanosomatid species. Multiplex PCR was validated by assaying genomic DNA from 3430
strains of T. cruzi that had been previously characterized; 24 blood samples from31
experimentally-infected mice and non-infected controls; 20 buffy coat samples from 32
patients in the acute phase of Chagas disease and non-infected individuals, and 15 33
samples of feces from naturally infected Triatoma infestans. T. cruzi samples from patients 34
and from Y strain infected mice were classified by multiplex PCR as T. cruzi II and samples 35
from T. infestans and Colombiana strain infected mice as T. cruzi I.36
Index descriptors and abbreviations37
DNA, deoxyribonucleic acid; kDNA, kinetoplast deoxyribonucleic acid; PBS, phosphate 38
buffered saline; PCR, polymerase chain reaction; rDNA, ribosomal DNA; RAPD, random 39
amplification of polymorphic DNA; RFLP, restriction fragment length polymorphism; T. 40
cruzi I, Trypanosoma cruzi group I; T. cruzi II, Trypanosoma cruzi group II; Z1, Z2, Z3, 41
zymodemes 1, 2 and 3; Triatomine; Trypanosoma cruzi; Triatoma infestans; Satellite DNA; 42
Multiplex PCR.43
44
45
46

ACCEPTED MANUSCRIPT 3
1. Introduction47
Chagas disease is caused by the protozoan Trypanosoma cruzi (Family: 48
Trypasonomatidae, Order: Kinetoplastida). In Latin America, some 15 million people are 49
infected with T. cruzi, whilst up to 28 million people are at risk of contracting the disease 50
(World Health Organization, 2007). The extensive variability in the symptoms of Chagas 51
disease may be correlated with specific genetic markers in the host or parasite, although it 52
is likely that both will affect the outcome of the infection (Sturm et al., 2003).53
T. cruzi exhibits a considerable intra-specific diversity that is exemplified by 54
differences in morphology of blood forms, virulence, pathogenicity, susceptibility to 55
chemotherapeutic agents, immunological properties and infectivity in host cells (Murta and56
Romanha, 1999). In an attempt to standardize the nomenclature applied to the parasite, T. 57
cruzi has been separated into two major groups, T. cruzi I and T. cruzi II (Anonymous, 58
1999). This classification has received support from studies using a variety of molecular59
markers including polymorphism of rDNA and mini-exon genes, RAPD and RFLP (Steindel 60
et al., 1993; Souto et al., 1996; Bastrenta et al., 1999; Sturm et al., 2003). Although T. cruzi61
I is associated with the sylvatic cycle of the disease, and T. cruzi II with the domestic cycle, 62
this epidemiologically important association is not totally correct once T. cruzi I and T. cruzi63
II have already been isolated in both sylvatic and domestic cycles (Yeo et al, 2007).64
The total DNA content (nuclear and kinetoplastic) of T. cruzi varies from strain to 65
strain and also amongst clones of the same strain, but is typically in the range 0.12 - 0.33 66
pg per cell (Lanar et al., 1981; Kooy et al., 1989; Thompson and Dvorak, 1989; McDaniel 67
and Dvorak, 1993). The genome of T. cruzi is estimated to contain between 1.8 and 2.5 x 68
108 bp, although more than 50% is apparently associated with repetitive elements, such as 69
transposons and multi-copy genes, according to the recently published genomic sequence 70
of the parasite strain CL-Brener (El-Sayed et al., 2005). Such highly repetitive sequences71
probably play important roles in genomic structure and in gene expression. Since most are 72

ACCEPTED MANUSCRIPT 4
species specific, they may be employed in the diagnosis of Chagas disease. Moreover, the 73
rapid evolutionary change in the interspersion pattern of the repetitive sequences also 74
makes such elements useful for strain-classification studies (Requena et al., 1996).75
A satellite DNA element of 195 bp with 120,000 copies per genome was the first 76
repeated nuclear sequence to be described in T. cruzi (Gonzalez et al., 1984). The high77
abundance and intra-specific diversity of this repeat suggested its possible application to 78
detect T. cruzi infection. Indeed, it is possible to detect minimum amounts of parasite DNA79
in diverse biological samples including infected blood, biopsies of heart tissue and feces of 80
triatomines, through PCR amplification of the 195 bp repeat element (Moser et al., 1989; 81
Olivares-Villagómez et al., 1998; Machado et al., 2000; Virreira et al., 2003). Furthermore, 82
studies carried out by Bastrenta et al. (1999) and Elias et al. (2005) have demonstrated83
that there is an association between the sequences of satellite DNA repeats of T. cruzi and 84
parasite groups.85
Whilst PCR targeting of highly repetitive sequences provides sensitive detection of86
T. cruzi DNA, markers that could distinguish between T. cruzi strains are also important as87
they would allow parasite classification without the need to isolate strains. In the present 88
work we have developed a multiplex PCR that provides simultaneous diagnosis of89
infection with T. cruzi and classification according to T. cruzi groups I and II.90
2. Materials and methods91
The use of human blood samples were approved by the Ethical Committee of the 92
Universidade Federal de Santa Catarina, Brazil (Nº 233/05).93
2.1. Samples analyzed94
Thirty four strains of T. cruzi that had been previously characterized according to 95
the isoenzyme profile, fragment f30 and T. cruzi group (Murta et al., 1998; Toma et al., 96
2007) were used in this study (Table 1). These strains were isolated from patients with 97

ACCEPTED MANUSCRIPT 5
acute and chronic Chagas disease, and from other vertebrate and invertebrate hosts from98
various geographical areas (Filardi and Brener, 1987; Toma et al., 2007). Twelve strains (4 99
strains of each zymodeme, Z1, Z2 and ZB) were selected for sequencing and analysis of 100
the 195 bp satellite repetitive DNA. All the strains used in this study are listed on Table 1.101
For the technical validation trials, DNA samples were obtained from: (i) blood of 102
non-infected mice (10 samples; negative controls), and of mice that had been 103
experimentally infected with T. cruzi strains Colombiana (8 samples) and Y (6 samples),104
both in acute phase of infection, (ii) the Buffy coat of individuals living in the state of Santa 105
Catarina, Brazil (an area non-endemic for Chagas disease) who presented negative 106
serology (indirect immunofluorescence assay) and parasitological examinations (11 107
samples), and of patients in the acute phase of Chagas disease (9 samples) who were 108
infected in a recent outbreak in the same state (Steindel et al., 2005 and 2008), (iii) feces109
of Triatoma infestans captured from Mato Queimado region (Rio Grande do Sul state, 110
Brazil - 15 samples), and (iv) strains of Trypanosoma rangeli, Leishmania braziliensis and 111
Leishmania amazonensis available at the Laboratório de Parasitologia Celular e Molecular 112
of the Centro de Pesquisas René Rachou. All the DNA samples were stored at -20°C. The 113
DNA samples of triatomine feces diluted on PBS 1X and blood of mice (5-25 µl of each114
sample), were first collected on filter paper and dried at room temperature. After this step, 115
each sample was sealed in plastic bags and stored to 4 - 8°C until the extraction of DNA116
using a simple lysis method, as described below. 117
2.2. DNA extraction118
DNA from epimastigote forms of different T. cruzi strains was extracted using the119
standard phenol-chloroform method (Maniatis et al., 1982). A simple lysis method was 120
employed in order to extract DNA from blood samples (5 µl) of mice and from the feces of 121
T. infestans. In each case, the filter paper containing the sample was perforated and 122
placed in a tube together with 100 µl of deionized water, heated at 70ºC for 10 min, 123

ACCEPTED MANUSCRIPT 6
centrifuged at 17,900 x g for 3 min at room temperature, and the supernatant (DNA 124
preparation) used in the PCR amplification experiments.125
2.3. Multiplex PCR126
The primers used to amplify sequences of T. cruzi satellite DNA in the multiplex 127
PCR experiments were sense Diaz 7 (5’ CGCAAACAGATATTGACAGAG 3’) and anti-128
sense Diaz 8 (5’ TGTTCACACACTGGACACCAA 3’), as previously described by Diaz et 129
al. (1992), and sense TcSat 4 (5' GCAGCCGCTCGAAAACTATCC 3'), which was 130
designed on the basis of nucleotide sequences determined in the present study. In each 131
experiment the reaction mixture (10 μl) contained: PCR buffer (50 mM KCl, 20 mM Tris-132
HCl; pH 8.4), 1.9 mM MgCl2, 200 μM of each dNTP (Promega, Madison, WI, USA), 5 pmol133
of each primer; 0.5 units of Taq DNA polymerase (Invitrogen, Carlsbad, CA, USA) and 1 μl 134
of DNA from the T. cruzi DNA preparation. Amplifications were carried out in a Perkin 135
Elmer (CA, USA) Mastercycler® thermocycler and involved an initial denaturation step of 5 136
min at 95°C, followed by 30 cycles of 20 s at 95°C, 10 s at 60°C and 15 s at 72°C, and a137
final extension step of 5 min at 72°C. An aliquot (3 µl) of the PCR product was subjected to 138
electrophoresis on a 6% non-denaturing polyacrylamide gel and the components 139
visualized by silver staining (Sanguinetti et al., 1994).140
2.4. Purification and cloning of satellite DNA141
The PCR-amplified products were purified with Qiagen (Valencia, CA, USA) 142
QIAquick® PCR Purification and Gel Extraction kits, and subsequently cloned into pCR®2.1 143
TOPO vector using a TOPO TA Cloning® kit (Invitrogen), according to the protocols 144
provided by the manufacturers. Recombinant vectors were inserted into chemo competent145
E. coli TOP10 F’ cells and the recombinant plasmids purified using a QIAprep® Spin 146
Miniprep kit (Qiagen) and quantified spectrophotometrically.147

ACCEPTED MANUSCRIPT 7
2.5. DNA sequencing148
Recombinant plasmids were sequenced using an Amersham Bioscience 149
Megabace® 500 capillary sequencer and DYEnamic® ET Terminator sequencing kit.150
Reactions were carried out in 96-well plates and each mixture contained 4 µl of DYEnamic 151
ET Terminator sequencing solution, 5.0 pmoles of M13 forward primer (5’ 152
GTAAAACGACGGCCAG 3’), Diaz 7 or Diaz 8 primer (see item 2.3.), and ca. 150 ng of 153
DNA in a final volume of 10 µl. Samples were submitted to 25 cycles of 20 s at 95°C, 15 s 154
at 50°C and 1 min at 60°C in a Mastercycler® thermocycler, precipitated and electro-155
injected according to the protocols described by the manufacturer.156
2.6. Analysis of sequences157
Vector sequences were masked using the Cross-Match algorithm (Gotoh, 1982) 158
and sequence quality was evaluated with Phred base-calling software (Ewing and Green, 159
1998; Ewing et al., 1998). Only bases with Phred values larger than 20 (> 99% accuracy) 160
were included in the 195 bp repetitive satellite DNA sequences established for strains of T. 161
cruzi. DNA sequences were aligned using the Clustal X program (Thompson et al., 1997),162
and discrepancies between sequences were confirmed by visual inspection of the 163
chromatograms and were edited manually when necessary. Phylogenetic trees were 164
constructed using MEGA 3 (Kumar et al., 2004) and Clustal X programs. Some trees were 165
viewed with the aid of the TreeView program developed by Page (2001).166
167
3. Results168
3.1. Analysis of satellite DNA nucleotide sequences established for strains of T. cruzi169
Samples of DNA from 34 strains of T. cruzi were submitted to PCR analysis using170
the primers Diaz 7 and 8 (Table 1). A 195 bp fragment was detected in all samples171

ACCEPTED MANUSCRIPT 8
analyzed, together with additional bands corresponding to the amplification of tandem172
repeats of this element. The sensitivity of the PCR analysis was 1 fg for Colombiana (T. 173
cruzi I) strain and 0.1 fg for Ernane (T. cruzi II) strain. No amplification products were174
observed when DNA from T. rangeli, L. braziliensis or L. amazonensis was used as 175
template in amounts up to 10 pg per reaction. These results are in accord with the 176
literature and confirm the use of Diaz 7 and 8 primers to amplify sequences of the satellite 177
DNA of T. cruzi.178
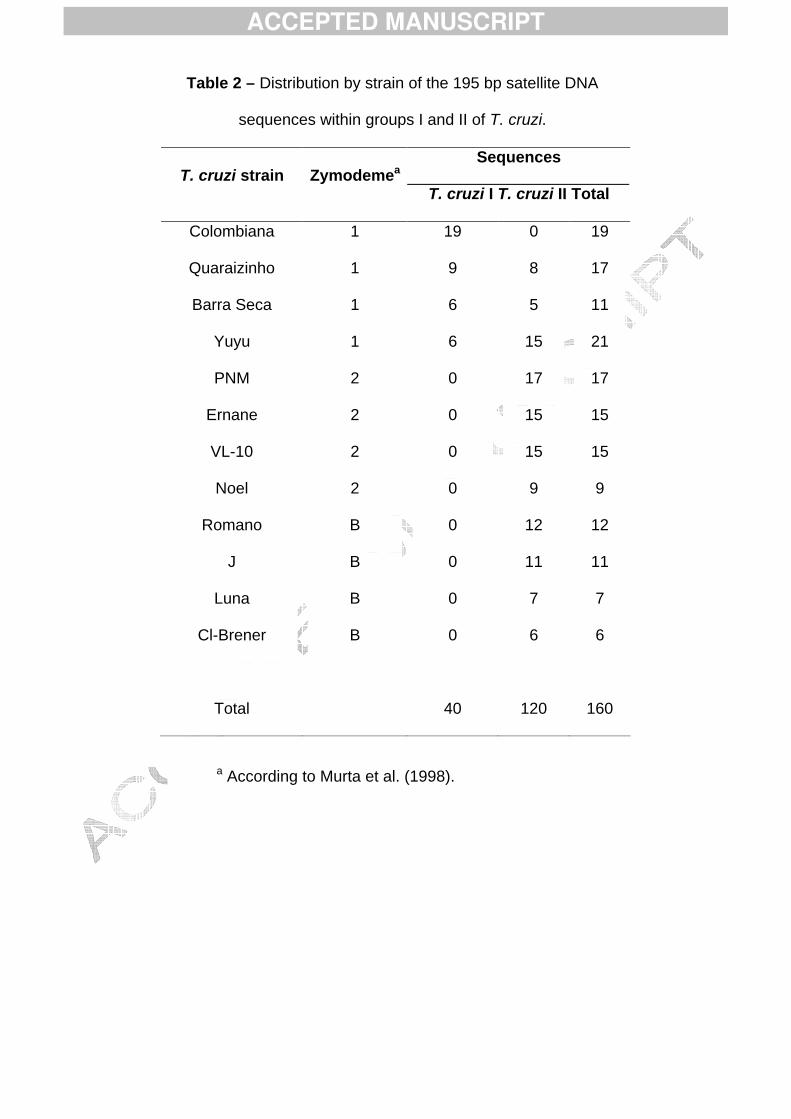
One hundred and sixty nucleotide sequences of satellite DNA derived from 12179
strains of T. cruzi were analyzed and the distribution of these sequences is presented in 180
Table 2. Polymorphisms were detected among the 195 bp sequences of each strain and 181
among strains from different zymodemes. A total of 89 polymorphic sites were observed, 182
but most of them were present in only a small number of sequences and were not 183
representative. Nevertheless, 8 polymorphic sites in satellite DNA were exclusive to strains 184
of T. cruzi I, although no sequence presented all 8 polymorphisms simultaneously.185
Figure 1 show a phylogenetic tree constructed on the basis of the nucleotide 186
sequences determined for T. cruzi satellite DNA. Two main branches were associated with 187
groups I and II of T. cruzi. The minor branch, delimited by a circle, contained exclusively 188
strains of group I (40 out of the 68 Z1 sequences), whilst the major branch contained 189
strains of group II (all 56 Z2 sequences and 36 ZB sequences) and some from group I (the 190
remaining 28 Z1 sequences). Of the four group I strains of T. cruzi analyzed, i.e. 191
Colombiana, Quaraizinho, Barra Seca and Yuyu (Table 2), only sequences of the 192
Colombiana strain were clustered exclusively in the group I branch. In the case of the 193
Quaraizinho strain, 9 out of the 17 sequences determined were clustered in the group I 194
branch, for the Barra Seca strain 6 out of 11 sequences were in the group I branch, and for 195
the Yuyu strain 6 out of 21 sequences were in the group I branch. This heterogeneity 196
suggests that some polymorphisms are exclusive to the satellite DNA of group I strains, 197

ACCEPTED MANUSCRIPT 9
and that these polymorphisms are not present in all of the repeat units from one same198
strain. 199
3.2. Multiplex PCR200
From these results, five primers (two sense and three anti-sense) complementary to 201
the polymorphic 3’ end bases characteristic of T. cruzi I strains were designed. Different 202
primer combinations were tested in PCR amplification reactions with DNA derived from 203
strains Colombiana and Quaraizinho (Z1), Noel and Ernane (Z2), and CL Brener and 204
Romano (ZB). Control PCR amplification reactions were also carried out with Diaz 7 and 8 205
primers. PCR product profiles that allowed the differentiation of T. cruzi strains into groups 206
I and II were obtained when primers TcSat 4, Diaz 7 and 8 were used together. Using this 207
set of primers, a panel of 34 T. cruzi strains consisting of 11 strains of T. cruzi I (Z1), 19 208
strains of T. cruzi II (10 strains of Z2 and 9 of ZB) and 4 strains of zymodeme 3 were 209
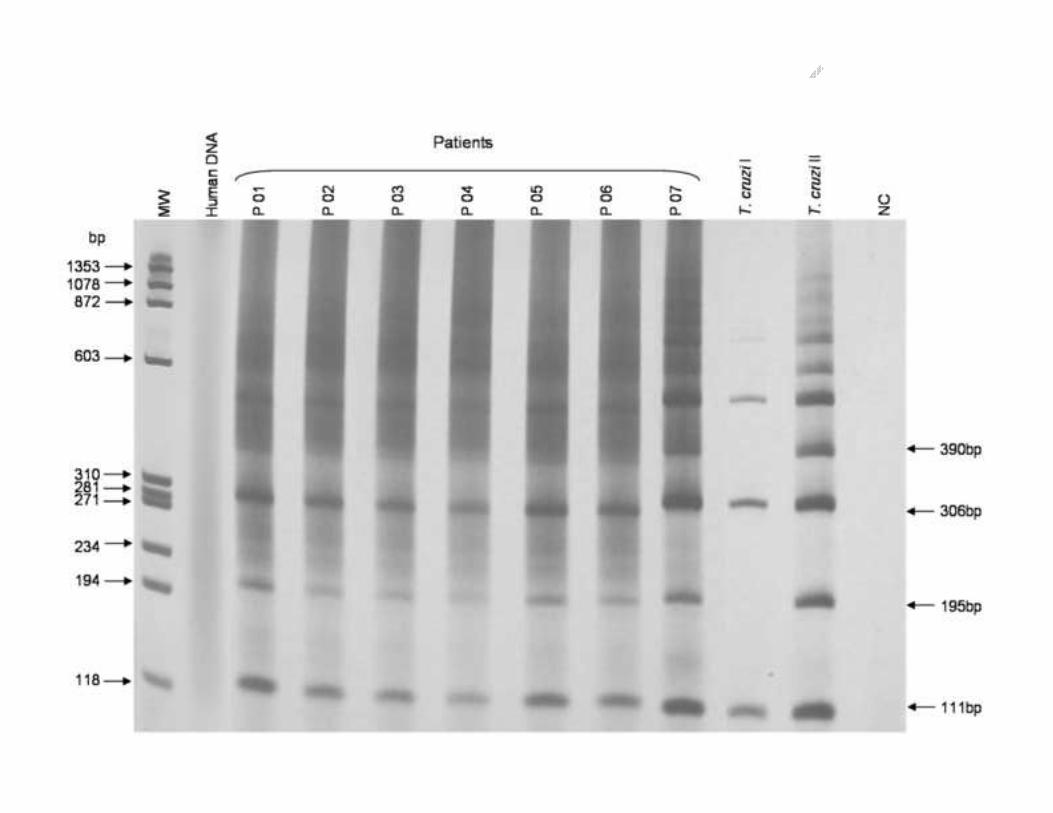
subjected to PCR amplification. Figure 2 shows the expected 111 bp fragment, resulting 210
from PCR amplification using TcSat 4 and Diaz 8 primers. It is present in all T. cruzi strains 211
and was used as a positive control for T. cruzi in DNA amplification reactions. Concerned 212
to, the 195 bp fragment generated by the PCR amplification with Diaz 7 and 8 primers, it is 213
present in all T. cruzi II (Z2 and ZB) and zymodeme 3 strains and absent in T. cruzi I 214
strains. Fragments of 306 and 390 bp can also be observed in the PCR product profile. 215
They resulted from primer annealing to adjacent sequences.216
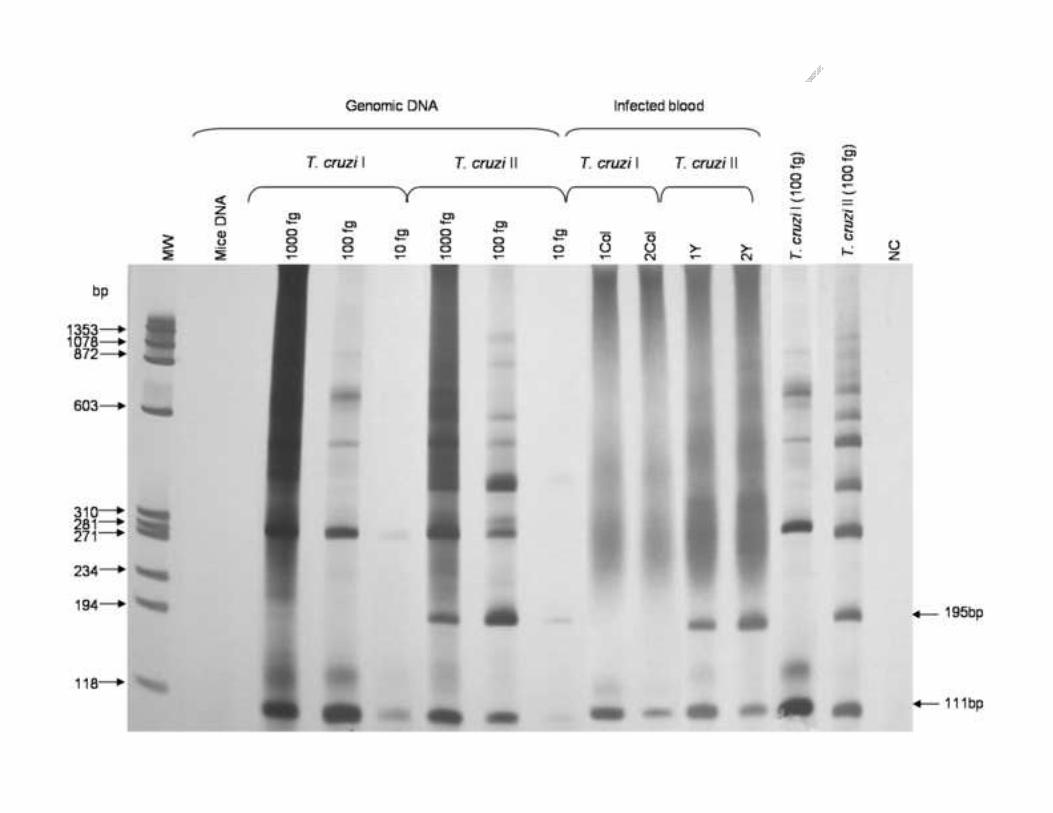
To determine the sensitivity of the method, genomic DNA from strains Colombiana 217
(T. cruzi I) and Ernane (T. cruzi II) was assayed by multiplex PCR using 10 pg to 1 fg. The 218
minimum amount of DNA that allowed the detection of both 111 and 195 bp bands was 10 219
fg for both strains. The specificity of multiplex PCR was evaluated using DNA from T.220
rangeli, L. braziliensis and L. amazonensis. No amplification products were observed when 221
DNA from these phylogenetically related parasites was used in a concentration of up to 10 222
pg, indicating that multiplex PCR is specific to T. cruzi.223

ACCEPTED MANUSCRIPT 10
3.3. Validation of multiplex PCR224
The application of the multiplex PCR technique was evaluated using samples from 225
different sources: (i) blood, collected on filter paper, from mice experimentally infected with 226
T. cruzi and from non-infected controls, (ii) buffy coat DNA from patients in the acute 227
phase of Chagas disease and from individuals that were not infected with the parasite, and 228
(iii) feces from Triatoma infestans naturally infected with T. cruzi, collected on filter paper.229
Fourteen blood samples from Swiss mice infected with Colombiana (T. cruzi I) and 230
Y (T. cruzi II) strains were analyzed by multiplex PCR. The expected 111 bp fragment was 231
detected in all samples, whilst the 195 bp fragment was detected exclusively in samples 232
from animals infected with the Y strain (Figure 3). Furthermore, amplification products 233
were not observed in any of the 10 negative control samples derived from non-infected 234
mice. A uniform smear was, however, observed in all samples and this is probably derived 235
from excess protein that was co-extracted with the DNA. In order to evaluate any possible236
effects of mouse DNA on the multiplex PCR reaction, amplification profiles were obtained 237
using 10 ng of DNA derived from non-infected mice, either alone or mixed with 1 pg to 10 238
fg of genomic DNA from Colombiana and Y strains. No amplification products were 239
observed using mouse DNA alone, and the presence of mouse DNA did not change the 240
sensitivity of the multiplex PCR even when present in an amount that was 106-times higher241
than that of parasite DNA.242
Nine samples of DNA from patients infected with T. cruzi in a recent outbreak of 243
Chagas disease in the state of Santa Catarina, Brazil, were assayed by multiplex PCR. A 244
T. cruzi II profile, with the presence of a 195 bp fragment similar to that of a standard 245
sample from group II, was observed for all patients in the acute phase of the disease. 246
None of the 11 samples derived from non-infected individuals living in the same area 247
presented amplification products. Similarly to mouse DNA, human DNA did not interfere 248

ACCEPTED MANUSCRIPT 11
with multiplex PCR. There was no change in the sensitivity of the PCR even when human 249
DNA was present in an amount 2.4 x 106-times higher than that of parasite DNA (Figure 4).250
Fifteen samples of feces from T. infestans potentially infected with T. cruzi were 251
assayed by multiplex PCR. A previous microscopic examination revealed that 11 of the 252
triatomines were infected with T. cruzi whilst 4 were not. Multiplex PCR confirmed the253
microscopic findings and showed that all the strains were T. cruzi I.254
255
4. Discussion256
Chagas disease is an important public health problem in Latin America. The 257
resurgence of numerous cases of acute infection and the occurrence of outbreaks of oral 258
infection with T. cruzi reinforce the need to implement more sensitive and rapid diagnostic 259
methods. PCR detection of kDNA through amplification of repeated DNA sequences of T. 260
cruzi has been used to diagnose Chagas disease in humans (Galvão et al., 2003; Virreira 261
et al., 2003). However, the sensitivity of the PCR may vary according to the target DNA, 262
the strain of parasite and to the type of clinical sample employed. Moreover, classification 263
of the infecting strain of T. cruzi either by DNA or isoenzyme methods require large 264
amounts of biological material, necessitating parasite isolation and cultivation, which is not 265
always possible. In the present study, we have contributed to the epidemiological studies 266
of Chagas disease by developing a multiplex PCR technique by which diagnosis of the 267
disease and classification of the parasite into groups T. cruzi I and T. cruzi II may be 268
carried out simultaneously.269
The target of the multiplex PCR was nuclear 195 bp satellite DNA of T. cruzi that 270
presents two important features: (i) it is present in a large number of copies in the genome271
of the parasite, thus providing high sensitivity of detection, and (ii) the frequency of 272
polymorphism is high within and between different strains (Gonzalez et al., 1984), thus273
allowing strain classification. A number of studies have shown that this repeat is an 274

ACCEPTED MANUSCRIPT 12
appropriate target for the molecular diagnosis of Chagas disease and also for taxonomic 275
purposes (Gonzalez et al., 1984; Olivares-Villagómez et al., 1998; Bastrenta et al., 1999; 276
Virreira et al., 2003). Furthermore, it has been demonstrated that strains of T. cruzi may be 277
separated into two groups, T. cruzi I and T. cruzi II, by RFLP analysis of the 195 bp 278
satellite DNA (Bastrenta et al., 1999), or by analysis of the nucleotide sequences of the 279
repeats (Elias et al., 2005). However, no methodology has yet been described that would 280
permit the simultaneous diagnosis and classification of T. cruzi strains.281
In the present study, polymorphisms in the sequences of satellite DNA of T. cruzi I282
and T. cruzi II strains have been identified, and primers complementary to these 283
polymorphic sites were designed. Multiplex PCR using a combination of Diaz 7 and 8 and 284
TcSat 4 primers, yielded amplification products that allowed differentiation between T. 285
cruzi strains of groups I and II. Multiplex PCR was then used to analyze 34 strains of T. 286
cruzi isolated from 8 host species from 4 different countries. The PCR product profile of all 287
T. cruzi strains exhibited a 111 bp fragment, which functioned as a positive control for the 288
PCR, and a 195 bp fragment that was characteristic of T. cruzi strains of group II. In T. 289
cruzi group I the 195 bp was absent.290
It is suggested that the different PCR product profiles established in the two groups 291
of T. cruzi could be related to quantitative differences in the satellite DNA present. Elias et 292
al. (2003) and Vargas et al. (2004) reported that the amount of satellite DNA in T. cruzi I293
strains was 4 to 9-times smaller than T. cruzi II. Thus, in T. cruzi II strains, the abundance 294
of satellite DNA may be sufficient to enable both of the sense primers (TcSat 4 and Diaz 7) 295
to bind to their templates, leading to the amplification of 111 and 195 bp fragments. 296
However, when the number of repeats is smaller, as in T. cruzi I strains, competition for 297
complementary sites favor the TcSat 4 primer, leading to amplification of the 111 bp 298
fragment only. An important factor with respect to the preferential binding of the TcSat 4 299
primer is its higher annealing temperature compared with that of Diaz 7, allowing it to 300

ACCEPTED MANUSCRIPT 13
anneal to its respective complementary site earlier and more readily than the Diaz 7 301
primer.302
It’s unclear the classification of the strains of T. cruzi belonging to zymodeme 3 into 303
the two major groups. According to an expert committee, T. cruzi I and T. cruzi II304
correspond, respectively, to zymodemes Z1 and Z2; however the strains of zymodeme 3 305
should not be grouped into any of the major groups until further characterization of these 306
isolates be performed (anonymous, 1999). Later, Brisse et al. (2000) proposed the 307
subdivision of T. cruzi II into five sub-groups (T. cruzi IIa-IIe) and the parasites of 308
zymodeme 3 were distributed into sub-groups IIa and IIc. On the other hand, Freitas et al. 309
(2006) proposed that Z3 stocks correspond to an ancestral lineage T. cruzi III (although 310
some strains, as Can III, cannot be accommodated into this scenario) and Pedroso et al. 311
(2007) suggest that Z3 isolates from the Brazilian Amazon constitute an independent 312
cluster with low intra-group genetic diversity. The multiplex PCR described here showed a 313
profile of T. cruzi II for the strains of zymodeme 3. It’s important to observe that the primers 314
used for multiplex PCR were made for diagnosis and classification of T. cruzi strains only 315
into the groups I and II. Multiplex PCR was validated using reference strains of T. cruzi I 316
(Colombiana strain), T. cruzi II (Y strain), including hybrid groups of T. cruzi IIe (CL-Brener 317
strain) and T. cruzi IIa (Can III strain, hybrid according to Freitas et al., 2006).318
Multiplex PCR is sensitive compared with other techniques employed in molecular 319
diagnosis. The sensitivity of the method when applied to samples of genomic DNA was 10 320
fg, which corresponds to about 1/30th of the DNA of one parasite. Such sensitivity is equal 321
to that achieved by other primers used to diagnose Chagas disease, including S 35 and 36 322
(Kirchhoff et al., 1996), which amplify a kDNA fragment. Although the use of Diaz 7 and 8 323
primers alone provides an even higher PCR sensitivity of ca. 0.1 to 1 fg depending on the 324
strain of T. cruzi, multiplex PCR offers the considerable advantage of being able both to 325
detect and to characterize the parasite with respect to group. In contrast, strain genotyping 326

ACCEPTED MANUSCRIPT 14
using rDNA or mini-exon genes (Murta et al., 1998) demands concentrations of DNA 327
around of 100 pg, i.e. 10,000 times higher than the necessary for the method proposed in 328
this study. Recently, Valadares et al. (2008) described seven polymorphic microsatellite 329
loci that are useful for characterization of parasites directly in infected tissues. For all these 330
loci, the authors designed two pairs of primers for full nested PCR assay. Using this 331
strategy, it’s possible to detect as little as 200 fg of parasite DNA (20X more than multiplex 332
PCR) and to identify strains of T. cruzi groups I, II, III (T. cruzi IIc) and hybrid groups (T. 333
cruzi IId-e). This method however is limited by diverse factors such as the small amount of 334
parasites in the infected tissues and the varying reaction sensitivities of each microsatellite 335
locus. A further important consideration is that the cultivation of parasites required to 336
obtain sufficient DNA for analysis may select for a specific sub-population, thus genotyping 337
may not reflect the heterogeneity of the original parasite population (Veloso et al., 2005). 338
The specificity of multiplex PCR was confirmed by the absence of PCR products when 339
DNA from phylogenetically related parasites (T. rangeli and Leishmania spp.) was 340
amplified. This result is in accordance with previous reports that the 195 bp satellite DNA 341
is exclusive of T. cruzi (Moser et al., 1989; Virreira et al., 2003).342
In order to validate the multiplex PCR method, clinical and field samples were 343
assayed. The collection of samples on filter paper facilitates the handling of field samples 344
and simplifies the extraction method, thus saving time and money. Using this technique, it345
was possible to correctly detect and classify blood samples of mice that had been346
experimentally infected with T. cruzi group I (Colombiana) and group II (Y) strains.347
Samples of DNA, extracted from feces collected on filter paper, of naturally infected T.348
infestans were also tested by multiplex PCR. All of the positive samples showed infection 349
with T. cruzi I strains. Machado et al. (2000) reported that the use of feces collected on 350
filter paper provided a rapid and practical method for the detection of T. cruzi and T. 351
rangeli in triatomines.352

ACCEPTED MANUSCRIPT 15
Multiplex PCR analysis of DNA from buffy coat of individuals in the acute phase of 353
Chagas disease revealed that all T. cruzi strains were of group II. These findings are in 354
accordance with recent studies using rDNA, mini-exon genes and isoenzyme (Steindel et 355
al., 2008). Buffy coat DNA samples from non-infected individuals were all negative for T. 356
cruzi according to multiplex PCR. Neither human nor mouse DNA affected the sensitivity 357
of multiplex PCR, indicating that the technique may be applied to clinical and field samples 358
without interference from the presence of excess host DNA.359
One of difficulties observed in the field research is the identification of mixed 360
infections with different genotypes of T. cruzi. The presence of multiple genotypes was 361
observed in triatomines and mammals (Bosseno et al., 1996; Solari et al., 2001). However, 362
there are few studies about T. cruzi groups I and II mixed infections and their prevalence in 363
field. It is possible that the frequency of mixed infections varies according to the 364
geographical area, the species of reservoirs and vectors and the tool (genetic marker) 365
utilized. Steindel et al. (1995) observed two mixed infections in 59 T. cruzi strains (3.4%)366
isolated from sylvatic reservoirs or naturally infected vectors. In an outbreak of Chagas 367
disease occurred in 2005, Steindel et al (2008) found two T. cruzi isolates obtained from 368
opossums belonged to the T. cruzi I, nine acute chagasic patients belonged to the T. cruzi369
II and two isolates from T. tibiamaculata with mixed infections. Marcet et al. (2006) found 370
only one mixed infection among 28 triatomines (3.6%) naturally infected in the northeast of 371
Argentina. Exceptionally, Bosseno et al. (1996) observed 45.7% of mixed infections in T. 372
infestans from Bolivia. Such data should be taken with caution since the triatomines were 373
captured in the same nest. In the Paraná state, Brazil Spitzner et al. (2007) observed 50% 374
of mixed infections in 18 strains isolated from triatomines. The authors found 6 of 9 375
Triatoma sordida and 3 of 9 P. megistus infected with T. cruzi I and T. cruzi II parasites, 376
simultaneously. Mixed infections were also observed in vertebrate hosts (Pinho et al., 377

ACCEPTED MANUSCRIPT 16
2000; Lisboa et al., 2006; Herrera et al., 2005), however the small number of samples 378
does not permit an evaluation of the frequency of mixed infections.379
The detection of mixed infections will depend on the tool (genetic marker) used. As 380
demonstrated by Steindel et al. (2008) using a phenotypic marker (MLEE) and by Spitzner 381
et al. (2007) with the mitochondrial gene cytochrome oxidase II (COII), it is possible to 382
detect mixed infections in samples of T. cruzi in which rDNA assay, SL gene analyzes and 383
RAPD do not detected. However, they depend of large amount of the parasite. The ability 384
of the multiplex PCR to detect mixed infection was performed using a mix of genomic DNA 385
of the strains Colombiana (T. cruzi I) and Y (T. cruzi II). Unfortunately, the profile observed 386
was similar to T. cruzi II and multiplex PCR couldn’t identify mixed infections into DNA 387
samples. However, the resolution of mixed infections is possible by the isolation of the 388
parasite clones by direct plating on a sensitive solid medium, as described by Yeo et al. 389
(2007). In this case, multiplex PCR could be performed even with a single parasite.390
In summary, the multiplex PCR described here is a sensitive, specific, rapid and 391
practical methodology that may be employed with different sources biological samples,392
containing minimal amounts of DNA, in order to detect and classify T. cruzi. The primers 393
designed were specific for classification into the groups T. cruzi I and T. cruzi II and cannot 394
be used for classification of T. cruzi II sub-group or T. cruzi III (T. cruzi IIc). Our study 395
shows that the 195 bp repetitive nuclear DNA sequences is an ideal target for molecular 396
diagnosis of Chagas disease and simultaneously for classification of the infecting T. cruzi 397
strain.398
Acknowledgements399
The authors wish to thank Danielle Scholl (Laboratório de Protozoologia, 400
Universidade Federal de Santa Catarina, SC, Brazil) and Ivan Sonoda (Laboratório de 401
Triatomíneos e Epidemiologia da Doença de Chagas, CPqRR, Fiocruz, MG, Brazil) for 402
their support in obtaining buffy coat samples from patients and feces from triatomines, 403

ACCEPTED MANUSCRIPT 17
respectively. This work was supported by grants from the Brazilian agencies: Conselho 404
Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação de Amparo 405
à Pesquisa do Estado de Minas Gerais (FAPEMIG).406
References407
Anonymous, 1999. Recommendations from a satellite meeting. International Symposium 408
to commemorate the 90th anniversary of the discovery of Chagas disease. Memórias 409
do Instituto Oswaldo Cruz 94 (Suppl. 1), 429-432.410
Bastrenta, B., Bosseno, M.F., Barnabé, C., Tibayrenc, M., Brenière, S.F., 1999. Restriction 411
fragment length polymorphism of 195 bp repeated satellite DNA of Trypanosoma cruzi412
supports the existence of two phylogenetic groups. Memórias do Instituto Oswaldo 413
Cruz 94, 323-328.414
Bosseno, M.F., Telleria, J., Vargas, F., Yaksic, N., Noireau, F., Morin, A., Breniere, S.F., 415
1996. Trypanosoma cruzi: study of the distribution of two widespread clonal genotypes 416
in Bolivian Triatoma infestans vectors shows a high frequency of mixed infections. 417
Experimental Parasitology. 83, 275-282.418
Brisse, S., Dujardin, J.C., Tibayrenc, M., 2000. Identification of six Trypanosoma cruzi419
phylogenetic lineages by random amplified polymorphic DNA and multilocus enzyme 420
eletrophoresis. International Journal for Parasitology. 30, 35-44.421
Diaz, C., Nussenzweig, V., Gonzalez, A., 1992. An improved polymerase chain reaction 422
assay to detect Trypanosoma cruzi in blood. American Journal of Tropical Medicine 423
and Hygiene. 46, 616-623.424
Elias, M.C., Vargas, N.S., Zingales, B., Schenkman, S., 2003. Organization of satellite 425
DNA in the genome of Trypanosoma cruzi. Molecular and Biochemical Parasitology. 426
129, 1-9.427
Elias, M.C., Vargas, N., Tomazi, L., Pedroso, A., Zingales, B., Schenkman, S., Briones, 428
M.R., 2005. Comparative analysis of genomic sequences suggests that Trypanosoma 429

ACCEPTED MANUSCRIPT 18
cruzi CL Brener contains two sets of non-intercalated repeats of satellite DNA that 430
correspond to T. cruzi I and T. cruzi II types. Molecular and Biochemical Parasitology. 431
140, 221-227.432
El-Sayed, N.M., Myler, P.J., Bartholomeu, D.C., Nilsson, D., Aggarwal, G., Tran, A.N., 433
Ghedin, E., Worthey, E.A., Delcher, A.L., Blandin, G., Westenberger, S.J., Caler, E., 434
Cerqueira, G.C., Branche, C., Haas, B., Anupama, A., Arner, E., Aslund, L., Attipoe, P., 435
Bontempi, E., Bringaud, F., Burton, P., Cadag, E., Campbell, D.A., Carrington, M., 436
Crabtree, J., Darban, H., da Silveira, J.F., de Jong, P., Edwards, K., Englund, P.T., 437
Fazelina, G., Feldblyum, T., Ferella, M., Frasch, A.C., Gull, K., Horn, D., Hou, L., 438
Huang, Y., Kindlund, E., Klingbeil, M., Kluge, S., Koo, H., Lacerda, D., Levin, M.J., 439
Lorenzi, H., Louie, T., Machado, C.R., McCulloch, R., McKenna, A., Mizuno, Y., 440
Mottram, J.C., Nelson, S., Ochaya, S., Osoegawa, K., Pai, G., Parsons, M., Pentony, 441
M., Pettersson, U., Pop, M., Ramirez, J.L., Rinta, J., Robertson, L., Salzberg, S.L., 442
Sanchez, D.O., Seyler, A., Sharma, R., Shetty, J., Simpson, A.J., Sisk, E., Tammi, 443
M.T., Tarleton, R., Teixeira, S., Van Aken, S., Vogt, C., Ward, P.N., Wickstead, B., 444
Wortman, J., White, O., Fraser, C.M., Stuart, K.D., Andersson, B., 2005. The genome 445
sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science 309, 409-446
415.447
Ewing, B., Green, P., 1998. Base-calling of automated sequencer traces using Phred. II. 448
Error probabilities. Genome Research. 8, 186-194.449
Ewing, B., Hillier, L., Wendl, M.C., Green, P., 1998. Base-calling of automated sequencer 450
traces using Phred. I. Accuracy assessment. Genome Research. 8, 175-185.451
Filardi, L.S., Brener, Z., 1987. Susceptibility and natural resistance of Trypanosoma cruzi452
strains to drugs used clinically in Chaga’s disease. Transactions of the Royal Society of 453
Tropical Medicine and Hygiene. 81, 755-759.454

ACCEPTED MANUSCRIPT 19
Freitas, J.M., Augusto-Pinto, L., Pimenta, J.R., Bastos-Rodrigues, L., Gonçalves, V.F., 455
Teixeira, S.M.R., Chiari, E., Junqueira, A.C.V., Fernandes, O., Macedo, A.M., 456
Machado, C.R., Pena, S.D.J., 2006. Ancestral genomes, sex and the population 457
structure of Trypanosoma cruzi. PLoS Pathogens. 2(3), e24.458
Galvão, L.M., Chiari, E., Macedo, A.M., Luquetti, A.O., Silva, S.A., Andrade, A.L., 2003. 459
PCR assay for monitoring Trypanosoma cruzi parasitemia in childhood after specific 460
chemotherapy. Journal of Clinical Microbiology. 41, 5066-5070.461
Gonzalez, A., Prediger, E., Huecas, M.E., Nogueira, N., Lizardi, P., 1984. Mini-462
chromosomal repetitive DNA in Trypanosoma cruzi: its use in a high-sensitivity parasite 463
detection assay. Proceedings of the National Academy of Sciences USA 81, 3356-464
3360.465
Gotoh, O., 1982. An improved algorithm for matching biological sequences. Journal for466
Molecular Biology. 162, 705-708.467
Herrera, L., D'Andrea, P.S., Xavier, S.C., Mangia, R.H., Fernandes, O., Jansen, A.M., 468
2005. Trypanosoma cruzi infection in wild mammals of the National Park 'Serra da 469
Capivara' and its surroundings (Piaui, Brazil), an area endemic for Chagas disease. 470
Transactions of the Royal Society of Tropical Medicine and Hygiene. 99(5), 379-88.471
Kirchhoff, L.V., Votava, J.R., Ochs, D.E., Moser, D.R., 1996. Comparison of PCR and 472
microscopic methods for detecting Trypanosoma cruzi. Journal of Clinical Microbiology. 473
34, 1171-1175. 474
Kooy, R.F., Ashall, F., Van Der Ploeg, M., Overdulve, J.P., 1989. On the DNA content of 475
Trypanosoma cruzi. Molecular and Biochemical Parasitology. 36, 73-76.476
Kumar, S., Tamura, K., Nei, M., 2004. MEGA3. Integrated software for molecular 477
evolutionary genetics analysis and sequence alignment. Briefings Bioinformatics 5, 478
150-163.479

ACCEPTED MANUSCRIPT 20
Lanar, D.E., Levy, S.C., Manning, J.E., 1981. Complexity and content of the DNA and 480
RNA in Trypanosoma cruzi. Molecular and Biochemical Parasitology. 3, 327-341.481
Lisboa, C.V., Mangia, R.H., Luz, S.L., Kluczkovski, A. Jr., Ferreira, L.F., Ribeiro, C.T., 482
Fernandes, O., Jansen, A.M., 2006. Stable infection of primates with Trypanosoma 483
cruzi I and II. Parasitology 133(Pt 5), 603-11.484
Machado, E.M., Alvarenga, N.J., Romanha, A.J., Grisard, E.C., 2000. A simplified method 485
for sample collection and DNA isolation for polymerase chain reaction detection of 486
Trypanosoma rangeli and Trypanosoma cruzi in triatomine vectors. Memórias do 487
Instituto Oswaldo Cruz 95, 863-866.488
Maniatis, T.M., Fritsch, E.F., Sambrook, J., 1982. Molecular Cloning. A Laboratory Manual. 489
Cold Spring Harbor University Laboratory Press, Cold Spring Harbor.490
Marcet, P.L., Duffy, T., Cardinal, M.V., Burgos, J.M., Lauricella, M.A., Levin, M.J., Kitron, 491
U., Gürtler, R.E., Schijman, A.G., 2006 PCR-based screening and lineage identification 492
of Trypanosoma cruzi directly from faecal samples of triatomine bugs from 493
northwestern Argentina. Parasitology 132(Pt 1), 57-65. 494
McDaniel, J.P., Dvorak, J.A., 1993. Identification, isolation and characterization of 495
naturally-occurring Trypanosoma cruzi variants. Molecular and Biochemical 496
Parasitology. 57, 213-222.497
Moser, D.R., Kirchhoff, L.V., Donelson, J.E., 1989. Detection of Trypanosoma cruzi by 498
DNA amplification using the polymerase chain reaction. Journal of Clinical 499
Microbiology. 27, 1477-1482.500
Murta, S.M.F., Romanha, A.J., 1999. Characterization of Trypanosoma cruzi.501
Memórias do Instituto Oswaldo Cruz 94, 177-180.502
Murta, S.M.F., Gazzineli, R.T., Brener, Z., Romanha, A.J., 1998. Molecular 503
characterization of susceptible and naturally resistant strains of Trypanosoma cruzi to 504
benznidazol and nifurtimox. Molecular and Biochemical Parasitology. 93, 203-214. 505

ACCEPTED MANUSCRIPT 21
Olivares-Villagómez, D., Mccurley, T.L., Vnencak-Jones, C.L., Correa-Oliveira, R., Colley, 506
D.G., Carter, C.E., 1998. Polymerase chain reaction amplification of three different 507
Trypanosoma cruzi DNA sequences from human chagasic cardiac tissue. American 508
Journal of Tropical Medicine and Hygiene. 59, 563-570.509
Page, R.D.M., 2001. TreeView – tree drawing software for Apple Macintosh and Windows. 510
Available at http://taxonomy.zoology.gla.ac.ukl/rod/treeview.html. Accessed in August 511
2006512
Pedroso, A., Cupolillo, E., Zingales, B., 2007. Trypanosoma cruzi: exploring the nuclear 513
genome of zymodeme 3 stocks by chromosome size polymorphism. Experimental 514
Parasitology. 116(1), 71-6.515
Pinho, A.P., Cupolillo, E., Mangia, R.H., Fernandes, O., Jansen, A.M., 2000. Trypanosoma 516
cruzi in the sylvatic environment: distinct transmission cycles involving two sympatric 517
marsupials. Transactions of the Royal Society of Tropical Medicine and Hygiene. 94(5), 518
509-14.519
Requena, J.M., Lopez, M.C., Alonso, C., 1996. Genomic repetitive DNA elements of 520
Trypanosoma cruzi. Parasitology Today 12, 279-282.521
Sanguinetti, C.J., Dias Neto, E., Simpson, A.J., 1994. Rapid silver staining and recovery of 522
PCR products separated on polyacrylamide gels. Biotechniques 17, 914-921.523
Solari, A., Campillay, R., Ortíz, S., Wallace, A., 2001. Identification of Trypanosoma cruzi524
genotypes circulating in Chilean chagasic patients. Experimental Parasitology. 97(4), 525
226-33. 526
Souto, R.P., Fernandes, O., Macedo, A.M., Campbell, D.A., Zingales, B., 1996. DNA 527
markers define two major phylogenetic lineages of Trypanosoma cruzi. Molecular and 528
Biochemical Parasitology. 83, 141-152.529
Spitzner, F.L., Freitas, J.M., Macedo, A.M., Toledo, M.J.O., Araújo, S.M., Prioli, A.J., 530
Gomes, M.L., 2007. Trypanosoma cruzi - triatomine associations and the presence of 531

ACCEPTED MANUSCRIPT 22
mixed infections in single triatomine bugs in Paraná state, Brazil. Acta parasitologica. 532
52, 74-81.533
Steindel, M., Dias Neto, E., Menezes, C.L.P., Romanha, A.J., Simpson, A.J., 1993. 534
Random amplified polymorphic DNA analysis of Trypanosoma cruzi strains. Molecular 535
and Biochemical Parasitology. 60, 71-80. 536
Steindel, M., Toma, H.K., Ishida, M.M., Murta, S.M.F., de Carvalho Pinto, C.J., Grisard, 537
E.C., Schlemper Jr, B.R., Ribeiro-Rodrigues, R., Romanha, A.J., 1995. Biological and 538
isoenzymatic characterization of Trypanosoma cruzi strains isolated from sylvatic 539
reservoirs and vectors from the State of Santa Catarina, Southern Brazil. Acta Tropica540
60, 167-177.541
Steindel, M., Dias, J.C.P., Romanha, A.J., 2005. Doença de Chagas: Mal que ainda 542
preocupa. Ciência Hoje 37, 32-38.543
Steindel, M., Kramer Pacheco, L., Scholl, D., Soares, M., de Moraes, M.H., Eger, I., 544
Kosmann, C., Sincero, T.C., Stoco, P.H., Murta, S.M.F., de Carvalho-Pinto, C.J., 545
Grisard, E.C., 2008. Characterization of Trypanosoma cruzi isolated from humans, 546
vectors, and animal reservoirs following an outbreak of acute human Chagas disease 547
in Santa Catarina State, Brazil. Diagnostic Microbiology and Infectious Diseases. 60(1), 548
25-32.549
Sturm, N.R., Vargas, N.S., Westenberger, S.J., Zingales, B., Campbell, D.A., 2003. 550
Evidence for multiple hybrid groups in Trypanosoma cruzi. International Journal for551
Parasitology. 33, 269-279.552
Thompson, C., Dvorak, J.A., 1989. Quantitation of total DNA per cell in an exponentially 553
growing population using the diphenylamine reaction and flow cytometry. Analytical 554
Biochemistry. 177, 353-357.555

ACCEPTED MANUSCRIPT 23
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G., 1997. The 556
Clustal X Windows interface: flexible strategies for multiple sequence alignment aided 557
by quality analysis tools. Nucleic Acids Research. 25, 4876-4882.558
Toma, H.K., Yamada-Ogatta, S.F., Brandão, A., Krieger, M.A., Goldenberg, S., 559
Fernandes, O., 2007. Trypanosoma cruzi: Subtractive hybridization as a molecular 560
strategy to generate new targets to distinguish groups and hybrids. Experimental 561
Parasitology. 117, 178-187.562
Valadares, H.M., Pimenta, J.R., de Freitas, J.M., Duffy, T., Bartholomeu, D.C., Oliveira, R. 563
de P., Chiari, E., Moreira, M. da C., Filho, G.B., Schijman, A.G., Franco, G.R., 564
Machado, C.R., Pena, S.D., Macedo, A.M., 2008. Genetic profiling of Trypanosoma 565
cruzi directly in infected tissues using nested PCR of polymorphic microsatellites. 566
International Journal for Parasitology. 38(7), 839-50.567
Vargas, N., Pedroso, A., Zingales, B., 2004. Chromosomal polymorphism, gene synteny 568
and genome size in T. cruzi I and T. cruzi II groups. Molecular and Biochemical 569
Parasitology. 138, 131-141.570
Veloso, V.M., Romanha, A.J., Lana, M., Murta, S.M., Carneiro, C.M., Alves, C.F., Borges, 571
E.C., Tafuri, W.L., Machado-Coelho, G.L., Chiari, E., Bahia, M.T., 2005. Influence of 572
the long-term Trypanosoma cruzi infection in vertebrate host on the genetic and 573
biological diversity of the parasite. Parasitology Research. 96, 382-389. 574
Virreira, M., Torrico, F., Truyens, C., Alonso-Vega, C., Solano, M., Carlier, Y., Svoboda, 575
M., 2003. Comparison of polymerase chain reaction methods for reliable and easy576
detection of congenital Trypanosoma cruzi infection. American Journal of Tropical 577
Medicine and Hygiene. 68, 574-582.578
Yeo, M., Lewis, M.D., Carrasco, H.J., Costa, N.A., Llewellyn, M., Valente, S.A.S., Valente,579
V.C., Arias, A.R., Miles, M.A., 2007. Resolution of multiclonal infections of 580
Trypanosoma cruzi from naturally infected triatomine bugs and from experimentally 581

ACCEPTED MANUSCRIPT 24
infected mice by direct plating on a sensitive solid medium. Internacional Journal for582
Parasitology. 37(1), 111-120.583
World Health Organization (WHO), 2007. Available at http://www.who.org Accessed in 584
October 2008.585
586
Legends to figures587
Figure 1588
Neighbour-joining phylogenetic tree of representative sequences of satellite DNA from 12 589
T. cruzi strains. The major branch contains sequences of T. cruzi groups I and II. The 590
minor branch, delimited by a circle, contains only sequences of T. cruzi group I. For each 591
strain, the numbers of identical sequences determined are shown in brackets. Strains of T. 592
cruzi: BS - Barra Seca, Col – Colombiana, Qua – Quaraizinho, CLB – CL-Brener, Roma –593
Romano, Ern – Ernane.594
* – Sequences of T. cruzi I strains present in the major branch.595
Figure 2596
Electrophoretic profiles (6% polyacrylamide gel silver stained) of PCR products obtained 597
by amplification of satellite DNA from 16 T. cruzi strains of different zymodemes and 598
groups using TcSat 4, Diaz 7 and 8 primers. 599
MM – Molecular Marker Φ X 174 digested with Hae III enzyme (bp). 600
NC – PCR negative control (without DNA).601
Figure 3602
Electrophoretic profiles (6% polyacrylamide gel silver stained) of the products of multiplex 603
PCR obtained in the presence of excess mouse DNA to which had been added known 604
amounts of T. cruzi DNA and blood samples derived from mice experimentally infected 605

ACCEPTED MANUSCRIPT 25
with T. cruzi. Standard DNA samples of T. cruzi I (Colombiana) and II (Y) strains were 606
used.607
MM – Molecular Marker Φ X 174 digested with Hae III enzyme (bp). 608
NC – PCR negative control (without DNA). 609
Figure 4610
Electrophoretic profiles (6% polyacrylamide gel silver stained) of the products of multiplex 611
PCR obtained by amplification of DNA samples from patients in the acute phase of 612
Chagas disease. 613
MM – Molecular Marker Φ X 174 digested with Hae III enzyme (bp). 614
NC – PCR negative control (without DNA).615

ACCEPTED MANUSCRIPT
Table 1 – Strains of Trypanosoma cruzi analysed by multiplex PCR (n = 34).
T. cruzi
strainOrigina Host Zymodemeb Groupc
Multiplex
PCRd
Barra secae RS Triatoma infestans 1 I I
CA-I Arg Human CPg 1 I I
Colombianae Col Human CPg 1 I I
DM-28 Ven Didelphis marsupialis 1 I I
Quaraizinhoe RS T. infestans 1 I I
SC-14 SC Panstrongylus megistus 1 I I
SC-28 SC D. marsupialis 1 I I
SC-42 SC D. marsupialis 1 I I
SC-51 SC P. megistus 1 I I
Tehuantepec Mex Triatomine 1 I I
Yuyue BA T. infestans 1 I I
Chaninha MG Felis domesticus 2 II II
Exr 424/3 SP Philander opossum 2 II II
Exr 426/1 SP D. marsupialis 2 II II
Ernanee GO Human CPg 2 II II
Noele MG Human APf 2 II II
PNMe MG Human CPg 2 II II
São Felipe BA Human CPg 2 II II
SMJ GO Human CPg 2 II II
VL-10e MG Human CPg 2 II II
Y SP Human APf 2 II II
CL RS T. infestans B II II

ACCEPTED MANUSCRIPT
Cl-Brenere RS T. infestans B II II
FL RS T. infestans B II II
Ibarra Arg Human CPg B II II
Je MG Human APf B II II
JA GO Human CPg B II II
Lunae Arg Human APf B II II
MR RS T. infestans B II II
Romanoe Arg Human APf B II II
3663 AM Panstrongylus geniculatus 3 ND II
4167 AM Rhodnius brethesi 3 ND II
Can III PA Human 3 ND II
JJ AM Human 3 ND II
a Origin of strains: AM – Amazonas, Arg – Argentina, BA – Bahia, Col – Colômbia, GO –
Goiás, Mex – Mexico, MG – Minas Gerais, PA – Pará, RS – Rio Grande do Sul, SC –
Santa Catarina, SP – São Paulo and Ven – Venezuela. AM, BA, GO, MG, PA, RS,
SC and SP are different states from Brazil.
b According to Murta et al. (1998) and Toma et al (2007).
c According to Anonymous, (1999). ND, not determined.
d Typed in this study.
e Strains selected for nucleotide sequence analysis of satellite DNA.
f AP – Acute phase.
g CP – Chronic phase
ACCEPTED MANUSCRIPT
Table 2 – Distribution by strain of the 195 bp satellite DNA
sequences within groups I and II of T. cruzi.
T. cruzi strain ZymodemeaSequences
T. cruzi I T. cruzi II Total
Colombiana 1 19 0 19
Quaraizinho 1 9 8 17
Barra Seca 1 6 5 11
Yuyu 1 6 15 21
PNM 2 0 17 17
Ernane 2 0 15 15
VL-10 2 0 15 15
Noel 2 0 9 9
Romano B 0 12 12
J B 0 11 11
Luna B 0 7 7
Cl-Brener B 0 6 6
Total 40 120 160
a According to Murta et al. (1998).

ACCEPTED MANUSCRIPT

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT
Copyright © 2022 FDOKUMEN




















