
Early Cambrian Brachiopods and Other Shelly Fossils from the Basal Kinzers Formation of Pennsylvania
Upload
jagiellonianCategory
view
1download
0
ELSEVIER Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79-87
Trace fossils indicating bottom aeration changes: Folusz Limestone, Oligocene, Outer Carpathians, Poland
Stanislaw Leszczynski a, Alfred Uchman a, Richard G. Bromley b a Institute of Geological Sciences, Jagiellonian University, ul. Oleandry 2a, Pl-30-063 Krak6w, Poland b Geological Institute, Copenhagen University, Oster Voldgade 10, DK-1350 Kobenhaven K, Denmark
Received 24 November 1994; revised and accepted 24 May 1995
Abstract
Bioturbation sequences in the pelagic Folusz limestone indicate deposition under decreasing bottom aeration, possibly in association with the development of microbial mats. U-shaped burrows Diplocraterion and Arenicolites exclusively were produced in the laminated limestone, and they represent the only colonizers (sessile suspension feeders) of the oxygen-deficient sediment.
1. Introduction
There have been many attempts to employ the degree of bioturbation and the taxonomic com- position of trace fossils for interpreting oxygena- tion at the sediment/water interface and within the sediment (e.g., Savrda and Bottjer, 1986, 1987, 1989; Ekdale and Mason, 1988; Leszczynski, 1991; Savrda et al., 1991; Savrda, 1992; Uchman, 1992). Finely laminated, fine-grained sediments lacking bioturbation generally are referred to anoxic floors, whereas bioturbated sediments are considered indicative of aerated sites. However, many different factors influence bioturbation (e.g., Wetzel, 1991; Leszczynski, 1992). Therefore the trace fossil assemblages of similarly aerated bottoms but of different sedimentary environments or of different geological age need not be comparable. These variables complicate the models of oxygen-related ichnocoenoses.
In this paper, oxygen-related changes in bio-
0031-0182~6/$15.00 © 1996 ElsevierScienceB.V. Allrightsrese~ed SSD! 0031-0182(95)00052-6
turbation intensity and trace fossil assemblage are described from the Oligocene pelagic limestone (the Folusz limestone) of the Carpathian flysch. An unusual case is demonstrated where domichnial burrows are the only biogenic structures in lami- nated pelagic limestone. These burrows represent the work of pioneers in a dysoxic sea floor.
2. Material
The Carpathian flysch (Tithonian-Lower Miocene) comprises a series of imbricated nappes consisting of variable deep-sea deposits mainly of turbidite facies, approximately 6000 m thick. These deposits accumulated in several deep-marine, elon- gate basins of the western Tethys realm. In the Oligocene part of the Menilite-Krosno series, thin layers (usually several centimeters thick) of coccoli- thic limestone occur at several horizons (see Jucha,

80 s. Leszczynski et al./Palaeogeography, Palaeoclimatology. Palaeoecology 121 (1996) 79 87
1957, 1958, 1969; Jucha and Kotlarczyk, 1958, 1961; Koszarski and Zytko, 1959, 1961; Haczewski, 1989). The limestone at certain hori- zons differs in age and sedimentary features. It is mentioned in the older literature as the "Jaslo shale", and recently has been separated into distin- guishable horizons with separate names, i.e., Tylawa, Jaslo, Zagdrz and Folusz limestone (Haczewski, 1984).
The Folusz limestone is poorly exposed at the only locality where it is known, occurring in a tectonically complicated area on the boundary between the Silesian, Jaslo and Magura nappes (Koszarski, 1985), in the village of Folusz, immedi- ately upstream of the bridge across the Klopotnica stream (the road toward a mine; Fig. 1). The limestone is underlain and overlain by breccia and may be involved in it. The sequence is included in the Oligocene, Upper Krosno Beds (cf. Slaczka, 197/).
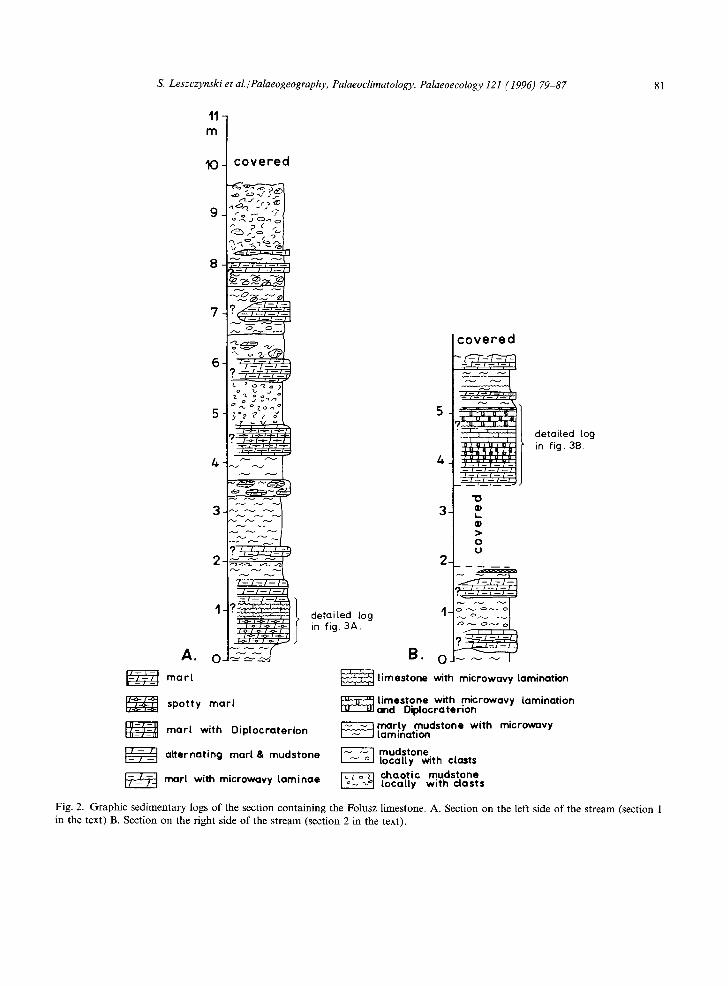
The limestone on either side of the stream differs both in style of bioturbation and in lithologic sequence (Figs. 2, 3). In both sections, the lime-
stone is weakly lithified and has the character of chalk. There are layers of non-laminated, nearly homogeneous limestone and marl up to about 50 cm thick, and thinner layers of finely laminated limestone displaying characteristic "microwavy" lamination of alternating white and dark sediment. The laminae show very irregular, "wavy" contours. Their thickness varies from 0.1 to 1 mm. The limestone is composed chiefly of coccoliths (e.g., Nowak, 1965; Haczewski, 1989) and the tests of planktic Foraminifera are numerous. Haczewski (1989) recognized that (p. 495) "The white lami- nae consist of flattened lenticular micrite nodules, separated by wavy, discontinuous and branching dark partings. The partings are 5-20 gm thick and consist of clay, organic matter and pyrite. The "dark laminae" also consist mainly of micrite nodules, but smaller, so that the dark partings are more densely spaced". According to Haczewski (1989), "...the distinction between the dark part- ings and the calcite nodules is much less pro- nounced" in the Folusz limestone as compared to the Jaslo limestone.
Q c ~ c o w C A R P A T H I A N F O R E D E E P ~ ' ' " I
S,,ESIAN NAPPE X
F M U ORL
. . . . . . . . . ?eo o PK.B ~ @ " ~ " k~..~,FM ~
~ ~ \ MAGURANAPPE ' _ "_% P O D H A L E F L Y S C H ~ x ~ • S L O V A K I A
0 30 km I I I I
SUN - SUB-SILESIAN UNIT
FMU - FORE-MAGURA UNIT
PKB - PIENINY KLIPPEN BELT
GORLICE 18 KM
i MAGURA BEDS (U. EOCENE PASSAGE BEDS (OLIGOCENE GREY THICK-BEDDED SANDY FLYSCH) ~ ] SILTSTONEB ALTERNATED WITH
DARK SHALES)
HIEROGLYPHIC BEDS (M. EOCENE THIN-BEDDED FLYSCH) ~ MENILITE BEDS (OLIGOCENE DARK
BROWN MARLS AND BLACK SHALES
VARIEGATED SHALE (L. EOCENE) / OVERTHRUSTS
KROSONO BEDS (OL[GOCENE SILTSTONES- MUDSTONES WITH OLISTOSTROME, • WHICH INCLUDES THE FOLUSZ LIMESTONE /# MAIN ROADS
Fig. I. Location and stratigraphy of investigated section.
S. Leszczynski et aL /Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79-87 81
11 rn
10 c o v e r e d
e J J -
8
7
/,
detailed log in fig. 3A.
A . o
mart
spotty mar l
mar l with Diplocraterion
alternating marl & mudstone
marl with microwavy laminae
/ ,
_
_
c o v e r e d
"o ® t - ® >. o u
,r / - - / - - I ~ I - - J
I -
o ~ j ~ limestone microwavy lamination with
~ l imestone with microwavy lamination and Dipiocraterion
~ marly mudstone with microwavy lamination
-'] mudstone locally with ctasts
~ chaotic mudstone locally with dasts
detailed log in fig. 3B,
Fig. 2. Graphic sedimentary logs of the section containing the Folusz limestone. A. Section on the left side of the stream (section 1 in the text) B. Section on the right side of the stream (section 2 in the text).

82 S. Leszczynski et aL /Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79 87
S E C T I O N 2 .,,.,ES,ONE TElC,lC,NUS " 1 LAMINATION ~ THALASSINOIDES _ ~ _ ~ 2D
u D I . L O C . ~ E R I O N - - M O T T , I N G L -
I U ARENICOUTES I ~
CHONDRITES
S E C T I O N 1 l - ,
-]-I_-N u UU 2B I
~ O O N I - -
-I - - - I A 20]cm 'IT---I- 2"-- -I/Tl_ "~'--~ 2A 0 ' - [ . . . . . , , , , ~
UNIT 1 2 3 4 S UNIT I 2 3 4 5 INDEX OF B. INDEX OF
A. BIOTURBATION BIOTURBATION
Fig. 3. Graphic sedimentary logs and bioturbation of the Folusz limestone. A. Section on the left side of the stream (section 1 in the text). B. Section on the right side of the stream (section 2 in the text).
The Folusz limestone, like the Jaslo limestone, in the light of Haczewski's (1989) interpretations (p. 498), "...is a product of exceptionally rapid pelagic deposition of calcareous nannoplankton remains... The white laminae record successive blooms of coccolithophores in surface waters". Haczewski interpreted the micronodular texture as resulting from deposition of coccoliths chiefly in faecal pellets of copepods. The mass occurrence of coccoliths, accompanied by globigerinids, indicates warm and oligotrophic surface waters of normal salinity. The lack of benthic fauna, the undisturbed fine lamination, the usual lack of bioturbation and the perfect preservation of fish bodies and plant debris are all considered to indicate oxygen-
deficient to anoxic bottom waters. Episodes of oxygenated bottom water are marked by bioturba- tion as described herein.
The possible occurrence in an olistostrome sug- gests that the Folusz limestone might have been deposited in shallower, more oxygenated water than the other Jaslo-type limestone which occurs in situ. The in situ Jaslo limestone is much thinner than the Folusz limestone, which is at least 3 m thick. Contrary to the Folusz limestone, the Jaslo limestone does not contain totally bioturbated layers. Haczewski (pers. comm.) recorded only Trichichnus. The very fine-grained texture and subtle lamination in the Folusz limestone indicate that it was deposited below storm wave base.

S. Leszczynski et al./Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79-87 83
3. Results
The Folusz limestone was investigated in two sections on either side of the Klopotnica Stream. Section 1 (Fig. 3A) displays two distinct units denoted 1A and lB. Unit 1A consists of light grey, totally bioturbated marl. The ichnofabric contains several types of biogenic structures in the form of irregular, circular or elongate spots of different sizes, outline and distinctiveness occurring in a mott led background (Fig. 4). These structures are referred to Planolites, Teichichnus, Thalassinoides, and very small Chondrites. Teichichnus seems to be more frequent in the upper part of the unit whereas the other trace fossils appear to be equally distributed throughout the unit.
Unit 1B consists of light grey, laminated, unbio- turbated limestone. The limestone displays thin microvavy lamination of alternating white and black laminae, and four white, thicker laminae ( 10-15 ram). A small fold of probable sedimentary origin (slump?) is noted in the finely laminated sediment. The contact between units 1A and 1B is sharp.
Four units, named 2A, 2B, 2C, and 2D, are distinguished in section 2 (Fig. 3B). Unit 2A con- sists of light grey, totally bioturbated marl and marly limestone. Trace fossils and biodeformation structures are similar to those in unit 1A, although
structures of the Teichichnus-type are very rare and poorly preserved in sediments of the unit 2A.
Unit 2B consists of light grey, totally bioturbated marl and marly limestone. Trace fossils are sharply contrasted against the mott led background and they are represented only by the U-shaped forms Diplocraterion and rare Arenicolites (Fig. 5). The latter occurs only in the lower part of unit 2B. Transition between units 2B and 2A is gradual.
Serial sections, sliced about 10 m m apart, paral- lel to bedding, show significant differences that are caused not only by disappearance of some burrows and appearance of others but also by change of burrow position and shape (Fig. 6). Some bur- rows, particularly Arenicolites, appear to twist. In some cases, these largest burrows also appear to interconnect. Diplocraterion displays a spreite chiefly in its lower part. The distinctiveness of the spreite may change vertically. In general, spreiten are most distinctive in Diplocraterion in microwavy laminated limestone. In all cases, the spreite is protrusive.
The fluctuating vertical course of a burrow may result f rom bedding-parallel dislocations at a scale of millimetres, and f rom sediment deformation
Fig. 4. Marl from unit 1A. The mottled background fabric contains a diverse association of poorly visible structures in form of streaks, wisps and various spots slightly differing in colour. Scale bar 1 cm.
Fig. 5. Limestone from unit 2B. Arenicolites (A) and Diplocraterion (D) cut an indistinctly bioturbated background. Burrow in upper-right of photo seems to represent basal portion of Arenicolites. Some specimens of Diplocraterion do not display spreital connection of tubes. Probably only the uppermost portion of these burrows has been sectioned. In some other Diplocraterion, the connection is only poorly visible (centre). Scale bar 1 cm.

84 S. Leszczvnski et al./Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79-87
during diagenesis. Characteristically, the outlines of Arenicolites have a fractured appearance and burrow shape changes vertically, whereas the shapes appear more stable and regular in Diplocraterion. However, in both burrow types, there are cases where at some levels only one shaft is visible. Some burrows disappear at levels where they are cut by a thread-like vein (filled joints). Thus, the analysis of the appearance of these trace fossils at different levels in cross sections allows us to recognize small-scale sediment deformation resulting from dissolution and micro-scale dislocations.
Unit 2C consists of light grey unbioturbated limestone. The limestone has white and black, thin microwavy lamination of the Jaslo limestone type. The contact between units 2C and 2B is sharp.
Unit 2D consists of light grey limestone having lamination as in unit 2C, and distinct burrows of Diplocraterion are present (Figs. 7, 8). The trans- ition from unit 2D to unit 2C is sharp.
4. Discussion
According to Ekdale and Mason (1988), Phanerozoic trace fossils change their ethologic character at distinct oxygenation levels. These authors suggested that, in response to increasing oxygenation, the preserved trace-fossil assemblage
Fig. 6. Limestone from unit 2B. Three horizontal serial sections, each couple 10ram apart. Arenicolites (A) and Diplocraterion (D) display vertical changes in burrow course. Scale bar 1 cm.
Fig. 7. Limestone from unit 2D. Diplocraterion is the only trace fossil occurring in this sediment. The spreiten are visible only in lower portion of the burrow. Scale bar 1 cm.

S. Leszczynski et al./Palaeogeography, Palaeoclimatology, Palaeoecology 121 (1996) 79-87 85
Fig. 8. Limestone from unit 2D in horizontal section. Several individuals of Diplocraterion of different sizes. Only two burrows at left-centre of photo display distinctive spreiten. The other forms have probably been cut in the upper portion where the spreite is lacking. Scale bar 1 cm.
should change from one dominated by subsurface deposit feeding structures (fodinichnia) through one dominated by grazing traces (pascichnia) to one dominated by dwelling structures (domichnia).
Wheatcroft (1989) pointed out that the trend suggested in the Ekdale and Mason model is at variance with the successional paradigm of modern shallow-water marine benthos described by Pearson and Rosenberg (1978), and Rhoads and Boyer (1982). In dysaerobic settings, the first colonists are small, tube-building, surface-deposit feeders or suspension feeders. They produce dom- ichnia of the Skolithos-type. As oxygen appears in the pore-water, the sediment is colonized by larger, mobile, subsurface deposit feeders of the pascichnia- or fodinichnia-type, depending on the distribution of food in the sediment. Hence, according to Wheatcroft (1989), the succession of trace fossil associations under increasing oxygena- tion should change from a dominance of domich- nia to one of fodinichnia, a trend opposite to that proposed by Ekdale and Mason. Admittedly, Ekdale and Mason (1989) recognized that some suspension-feeding organisms may inhabit poorly- oxygenated bottoms. Nevertheless, they stressed that their experience with the trace fossil record suggested that domichnia are rare in sediments that exhibit such direct indicators of oxygen deple-
tion as fine lamination and high organic carbon content.
Trace fossil assemblages of the Folusz limestone seem to be controlled chiefly by sediment oxygena- tion. The completely bioturbated units 1A and 2A appear to have been deposited in oxic conditions. The trace fossils referred to Thalassinoides, Planolites, Teichichnus, and Chondrites represent preservation of only the deepest tiers.
The environmental histories of sections 1 and 2 are different. Unit 1B is presumed to represent anoxic conditions. The composition of the lami- nated limestone of faecal pellets suggests that bacterial mats might have developed on the bottom during sedimentation of this deposit. This is indi- cated by an increase in the amount of organic matter and the presence of pyrite framboids in the dark laminae (Haczewski, 1989). There is no evidence of filamentous structures, commonly regarded as indicators of microbial mats. However, non-filamentous microorganisms, mainly hetero- trophic bacteria, also produce microbial mats, especially below the photic zone.
Such mats would have significantly restricted oxygen exchange between the sediment and overly- ing water mass. The border between units 1A and 1B is sharp, and this feature seems to indicate rather rapid oxygen depletion in the sediment. As a result of the oxygenation decrease, shortening of the benthic boundary layer and an upward shifting of tiers would have occurred. The more abundant occurrence of Teiehichnus in the top of unit 1A is considered to have resulted from such oxygenation change. Moreover, Teichichnus production was favoured by the increasing amount of organic matter in the sediment, which would have followed the decrease in oxygenation.
Teichichnus may represent the work of opportu- nistic deposit feeders able to live in dysoxic condi- tions (Ekdale and Mason, 1988). The mottled structure of the sediment in unit 1A up to the border with the unit 1B indicates that also some sediment mixers still penetrated the bottom, their activity preventing the development of a microbial mat. A rapid development of a dense microbial mat, i.e., appearance of the first biolamina, would have abruptly limited the activity of burrowers as it stiffened the bottom surface and cut off the

86 s. Leszczynski et al./Palaeogeography, Palaeoclimatology, Palaeoecology 121 (19963 79 87
influx of oxygen from the water column into the sediment. The significance of biolamination for sediment oxygenation was discussed by Sageman et al. (1991), Gerdes et al. (1991), and Savrda et al. (1991).
The decrease in oxygenation expressed in sec- tion 2 is slower and more gradual than the former case. This tendency is indicated by the appearance of the U-shaped burrows in the mottled sediment of unit 2B. The trace fossils Diplocraterion and Arenicolites represent the work of surface-detritus or suspension feeders. Their producers occupied deeper tiers than the sediment mottlers as they were able to pump water through the burrows. The almost homogeneously mottled background of unit 2B probably represents the "mixed layer" and resulted from sediment mixing close to the sea floor by animals that inhabited a soupy sediment. Thus, shafts of Arenicolites and Diplocraterion must have been quite resistant to sediment defor- mation by the mixers, unless they were built after sediment mixers had disappeared from this area. The presence of a lining (Fig. 7) suggests a rein- forcement of the burrows.
Subsequent decrease of oxygenation, as inferred for unit 2C, interrupted infaunal activity. Decline of infaunal activity in this case also coincided with the formation of the first biolamina on the sea floor. Slight increase in oxygenation in unit 2D allowed recolonization of the sediment by the Diplocraterion producers, which were able to keep pace with the microbial mats. They pumped oxygenated water into the sediment and disrupted the mats only minimally. Simultaneously, the mats prevented the activity of other sediment endo- benthos. It should be stressed, however, that microbial mats and activity of burrowers mutually affected each other: the lack of deposit feeders allowed the mats to form.
Sea-floor colonization by suspension feeders only is a common event on present day poorly oxygenated bot toms (Rhoads and Boyer, 1982). In fossil examples, the pioneer bo t tom colonization by suspension feeders is usually obliterated by subsequent activity of deposit feeders. Moreover, suspension feeders may be eliminated by deposit feeders owing to trophic group amensalism (Rhoads and Young, 1970). Preservation of activ-
ity of suspension feeders is possible only when their burrows escape destruction by deposit feeder activity. This is possible when activity of deposit feeders decreases, for instance, owing to poor oxygenation. Probably, stationary suspension feed- ers are able to coexist with sediment mixers up to a limited level of activity of the latter. Above that level, suspension feeders are not able to keep pace with deposit feeders. Traces of suspension feeders, including Diplocraterion, may be preserved when they become frozen by rapid deposition, for exam- ple, beneath storm beds (cf. Frey and Goldring, 1992).
Diplocraterion was likely produced by a suspen- sion feeder or a surface-scraping detritus feeder (Ft~rsich, 1974). This is indicated by the good and probably permanent connection with the sea floor. A vertical U-spreite would be ineffective for deposit feeding. Also, the host sediment and spreite show little difference in texture, indicating a lack of the sediment selection that would result from nutrient exploitation.
Thus, microbial mats have presumably inhibited intensive bioturbation of the laminated Folusz and Jaslo limestones. The mats prevented colonization of the bot tom by sediment-mixing deposit feeders. Instead, the bot toms covered with a microbial mat appear to have been initially colonized by suspen- sion feeders.
References
Ekdale, A.A. and Mason, T.R., 1988. Characteristic trace- fossil associations in oxygen-poor sedimentary environments. Geology, 16:720 723.
Frey, R.W. and Goldring, R., 1992. Marine event beds and recolonization surfaces as reveled by trace fossil analysis. Geol. Mag., 129:325 335.
Farsich, F.T., 1974. On DiplocraterionTorrell 1870 and the significance of morphological features in vertical, spreite- bearing, U-shaped trace fossils. J. Paleontol., 48:952 962.
Gerdes, G., Krumbein, W.E. and Reineck, H.-E., 1991. Biolaminations ecological versus depositional dynamics. ln: G. Einsele et al. (Editors), Cycles and Events in Stratigraphy. Springer, Berlin, pp. 592 606.
Haczewski, G., 1984. Correlation of laminae in chronohorizons of Jaslo limestone and Zagdrz limestone (Outer Carpathians). Kwart. Geol., 28: 675-688.
Haczewski, G., 1989. Coccolith limestone horizons in the

S. Leszczynski et al./Palaeogeography, Palaeoclirnatology, Palaeoecology 121 (1996) 79-87 87
Menilite-Krosno series (Oligocene, Carpathians): identifica- tion, correlation and origin. Ann. Soc. Geol. Pol., 59: 493-502.
Jucha, S., 1957. Lupki jasielskie w Karpatach fiiszowych. Prz. Geol., 5: 521-525.
Jucha, S., 1958. Contributions on Jaslo shaly limestones in Polish Carpathians. Bull. Acad. Pol. Sci. S6r. Sci. Chim. G6ol. G6ogr., 6: 681-688.
Jucha, S., 1969. Les schistes de Jaslo, leur importance pour la stratigraphie et la s6dimentologie de la s6rie m6nilitique et des couches de Krosno; Carpathes ftyscheuses. Pr. Geol. Pol. Akad. Nauk Odd. Krakowie, 52, 128 pp.
Jucha, S. and Kotlarczyk, J., 1958. Prdba nowego podzialu stratygraficznego serii menilitowej i warstw krosnienskich. Nafta, 14:205 207.
Jucha, S. and Kotlarczyk, J., 1961. La s6rie des couches ~i menilite et des couches de Krosno dans le flysch des Karpates. Pr. Geol. Pol. Akad. Nauk Odd. Krakowie, 4, 115 pp.
Koszarski, L. (Editor), 1985. Geology of the Middle Carpathians and the Carpathian Foredeep. In: Guide to excursion 3. Carpatho-Balkan Geological Association XIII Congress, Cracow. Geol. Inst., Warszawa, 254 pp.
Koszarski, L. and Zytko, K., 1959. Remarks on the develop- ment and stratigraphy of the Jaslo shales in the Menilitic and Krosno series of the Middle Carpathians. Kwart. Geol., 3:996 1015.
Koszarski, L. and Zytko, K., 1961. Jaslo shales within the Menilite-Krosno series in the Middle Carpathians. Inst. Geol., Biul., 166:87 232.
Leszczynski, S., 1991. Oxygen-related controls on predeposi- tional ichnofacies in turbidites, Guiptizcoan flysch (Albian- Lower Eocene), northern Spain. Palaios, 6: 271-281.
Leszczynski, S., 1992. Controls on trace fossil distribution in flysch deposits. Uniw. Jagiellonski, Rozpr. Habilit., 236, 82 pp.
Nowak, W., 1965., Sur l'origine organique des calcaires de Jaslo des couches m6nilitiques et de Krosno dans des Karpathes ftyscheuses (Oligocene). In: Carpatho-Balkan Geol. Assoc. 7th Congr. Sofia, September 1965, Rep., 2, 1, pp. 287-290.
Rhoads, D.C. and Boyer, L.F., 1982. The effects of marine benthos on physical properties of sediments: a successional perspective. In: P.L. McCall and M.J.S. Tevesz (Editors),
AnimaPSediment Relations. The Biogenic Alteration of Sediments. Topics Geobiol., 2: 3-52.
Rhoads, D.C. and Young, D.K., 1970. The influence of deposit-feeding organisms on sediment stability and com- munity trophic structure. J. Mar. Res., 28:150 178.
Sageman, B.B., Wignall, P.B. and Kauffman, L.G., 1991. Biofacies models for oxygen-deficient facies in epicontinental seas: Tool for paleoenvironmental analysis. In: G. Einsle et al. (Editors), Cycles and Events in Stratigraphy. Springer, Berlin, pp. 542-567.
Savrda, C.S., 1992. Trace fossil and benthic oxygenation. In: C.G. Maples and R.R. West (Editors), Short Courses in Paleontology, 5:172 196.
Savrda, C.S. and Bottjer, D.J., 1986. Trace-fossil model for reconstruction of paleooxygenation in bottom waters. Geology, 14: 3-6.
Savrda, C.S. and Bottjer, D.J., 1987. Trace fossils as indicators of bottom-water redox conditions in ancient marine environ- ments. In: D.J. Bottjer (Editor), New Concepts in the Use of Biogenic Sedimentary Structures for Paleoenvironmental Interpretation. Guidebook. Soc. Econ. Paleontol. Mineral. Pac. Sect., Los Angeles, 52, pp. 3 26.
Savrda, C.S. and Bottjer, D.J., 1989. Trace-fossil model for reconstructing oxygenation histories of ancient marine bottom waters: application to Upper Cretaceous Niobrara Formation, Colorado. Palaeogeogr. Palaeoclimatol. Palaeoecol., 74:49 74.
Savrda, C.S., Bottjer, J.D. and Seilacher, A., 1991. Redox- related benthic events. In: G. Einsle et al. (Editors), Cycles and Events in Stratigraphy. Springer, Berlin, pp. 524-541.
Slaczka, A., 1977. Problem I-A--Rozw6j facjalny i problemy paleogeograficznemlodszego paleogenu jednostki dukiel- skiej. In: A. Slaczka (Editor), Przewodnik XLIX Zjazdu Polskiego Towarzystwa Geologicznego, Krosno, 22 25 wrzesnia 1977. Wydawn. Geol., Warszawa, pp. 73-87.
Uchman, A., 1992. An opportunistic trace fossil assemblage from the flysch of the Inoceramian beds (Campania~ Paleocene), Bystrica Zone of the Magura Nappe, Poland. Cretaceous Res., 13: 539-547.
Wetzel, A., 1991. Ecologic interpretation of deep-sea trace fossil communities. Palaeogeogr. Palaeoclimatol. Palaeoecol., 85: 47-69.
Wheatcroft, R.A., 1989. Comment. In: Comment and Reply on "Characteristic trace-fossil associations in oxygen-poor sedimentary environments". Geology, 17: 674.
Copyright © 2022 FDOKUMEN




















