
To eat or not to eat. The significance of the cutmarks on the bones from wild canids, mustelids and...
96
Transcript of To eat or not to eat. The significance of the cutmarks on the bones from wild canids, mustelids and...


The aim of this dissertation is to discuss the use of wild canids, mustelids and felids at Hjerk Nor,
a Danish Erteb0lle site with an unusually high amount of 'fur animal' bones. The bones of fox
(Vulpes wipes), badger (Meles meles), pine marten (Murtes martes), polecat (Mustela putoris),
otter (Lutra lufra): lynx (Lynx hnx) and wild cat (Felis silvestris) were studied in a microscope
with 6-50 times magnification, in order to detect cutmarks. The placing of the cutmarks on each
species were compared to finds from two Neolithic sites in the Netherlands: Hazendonk and
Swifterbant, and from one Neolithic and three Mesolithic sites in Denmark: Kongemose,
Muldbjerg I, Praestelyng, and Tybrind Vig.
It was concluded that, with the exception of polecat, all 'fur animal' species at Hjerk Nor
were utilized for both skin and meat. Cutmarks deriving from dismembering and filleting were
particularly plentiful on wild cat and otter. Regarding the skinning method, it could be concluded
that otter and wild cat were open skinned, i.e. cut open along the ventral midline. Badger was
likely used for its fat, as well as for its fur. A tame fox brings an interesting aspect to the use of
wild canids during the Mesolithic. It may have been used as a 'fur farm', as a decoy, or as a pet.
The use of meat from wild canids, mustelids and felids seems to be rather common during
the Stone Age. Not all species were used to the same extent. Otter was eaten at all comparative
sites, whereas pine marten and polecat were rarely eaten. This variation in the frequency of
cutmarks related to butchering may indicate that some carnivores were considered more tasty that others. The use of meat from carnivores may also have had medical or ritual purposes.

Lena Strid - To eat or not to eat (MA dissertation)
ERRATA
p. 4, andpassim) "In ca 1937" SHOULD READ "In c. 1937"
p. 6: Table 1.2.) "Badly preserved" SHOULD READ "Poorly preserved"
P 9) "(Andersen 50-5 1)" SHOULD READ "(Andersen 1995:50-51)"
P. 18) "deducted" SHOULD READ "deduced"
P 50) "the snout is left on the carcass" SHOULD READ "the snout was left on the carcass"

1.2. Methods .......................................................................................................................... 2 1.3. Cutmarks ........................................................................................................................ 2 . .
1.3.1. Defimtion of cutmarks ....................................................................................... 2 1.3.2. Interpretation of cutmarks .................................................................................. 3
1.4. Hjerk Nor ...................................................................................................................... 4 1.5. Comparative sites ........................................................................................................... 6
1.5.1. Hazendonk ......................................................................................................... 6 1.5.2. Kongemose ........................................................................................................ 6 1.5.3. Muldbjerg I. ....................................................................................................... 6 1.5.4. Pwstelyng .......................................................................................................... 7 1.5.5. Swifterbant ......................................................................................................... 7
........................................................................................................ 1.5.6. Tybrind Vig 7
2 . Settlement patterns and economy in Southern Scandinavia during the Ertebfille period ............. 8
3 . The environment in Southern Scandinavia during the Ertebglle period ..................................... 10 3.1. The environment in the Limfjord region ....................................................................... 10
4 . The wild canids. mustelids and felids of Hjerk Nor .................................................................. 13 4.1. Fox (Vulpes vulpes) ...................................................................................................... 13 4.2. Badger (Meles meles) ................................................................................................... 14 4.3. Pine marten (Martes martes) ........................................................................................ 15 4.4. Polecat (Mustela putoris) .............................................................................................. 16
........................................................................................................ 4.5. Otter (Lutra lutra) 17 4.6. Lynx (Lynxlynx) ........................................................................................................... 17 4.7. Wild cat (Felis silveshis) .............................................................................................. 18
5 . Cutmarks on the bones of wild canids. mustelids and felids from Hjerk Nor .......................... 20 5.1. Cutmarks on the fox bones ........................................................................................... 20 5.2. Cutmarks on the badger bones ..................................................................................... 20 5.3. Cutmarks on the pine marten bones ............................................................................. 21 5.4. Cutmarks on the polecat bone ....................................................................................... 21
......................................................................................... 5.5. Cutmarks on the otter bones 21 5.6. Cutrnarks on the lynx bones ......................................................................................... 21 5.7. Cutmarks on the wild cat bones .................................................................................... 23
6 . Cutmarks on the bones from the comparative sites ................................................................ 24 6.1. Hazendonk .................................................................................................................... 24
6.1.1. Fox ................................................................................................................... 24 6.1.2. Badger .............................................................................................................. 24 6.1.3. Pine marten ...................................................................................................... 25 6.1.4. Poleca~ ............................................................................................................. 25 6.1.5. Otter ................................................................................................................. 25
6.2. Kongemose .................................................................................................................. 26 6.2.1. Pine marten ...................................................................................................... 26 6.2.2. Otter ................................................................................................................. 26 6.2.3. Wild cat ............................................................................................................ 26
6.3. Muldbjerg L .................................................................................................................. 26 6.3.1. Pinemarten ...................................................................................................... 26 6.3.2. Otter ................................................................................................................. 26

6.4. Pr~stelyng .................................................................................................................... 28 6.4.1. Pine marten ...................................................................................................... 28 6.4.2. Otter ................................................................................................................. 28
6.5. Swifterbant ................................................................................................................... 28 6.5.1. Otter ............... : ................................................................................................. 28
6.6. Tybrind Vig .................................................................................................................. 29 6.6.1. Pine marten ...................................................................................................... 29 6.6.2. Polecat. ............................................................................................................. 29 6.6.3. Otter ................................................................................................................. 30 6.6.4. Wild cat ............................................................................................................ 30
7 . The skinning and butchering experiment ................................................................................... 32 7.1. Method ......................................................................................................................... 32 7.2. Result ............................................................................................................................ 33 7.3. Discussion .................................................................................................................... 36
8 . Discussion ................................................................................................................................. 37 8.1. The individual species ................................................................................................... 37
8.1.1. Fox ................................................................................................................... 37 8.1.2. Badger .............................................................................................................. 39 8.1.3. Pine marten ...................................................................................................... 40 8.1.4. Polecat .............................................................................................................. 40
................................................................................................................. 8.1.5. Otter 43 8.1.6. Lynx ................................................................................................................. 45 8.1.7. Wild cat ............................................................................................................ 46
8.1.7.1. Skinning .......................................................................................... 46 8.1.7.2. Disarticulation ................................................................................. 47 8.1.7.3. Filleting ........................................................................................... 48
................... 8.1.7.4. Comparison with other sites and general interpretation 49
8.2. The overall view ....................................................................... : ................................... 50 8.2.1. Skinning methods ............................................................................................ 50 8.2.2. Wild canids. mustelids and felids as food ........................................................ 51 8.2.3. Emergency food or not? ................................................................................... 51 8.2.4. Other uses ........................................................................................................ 52
9 . Concluding words ..................................................................................................................... 52
Bibliography .................................................................................................................................. 53
Appendix 1 . Map of Southern Scandinavia with al l mentioned sites marked Appendix 2 . Osteological and anatomical terminology Appendix 3 . Measurements Appendix 4 . Cutmarks on individual bones

1. INTRODUCTION
Ever since working on my bachelor essay in Prehistoric archaeology, The hunting offur animals in Southern Scandinavia during the Mesolithic (Strid 1999). I have been interested in the wild
carnivores and rodents found in archaeological assemblages. These animals, i.e. wolf, fox,
badger, wolverine, pine marten, polecat, ermine, weasel, otter, lynx, wild cat, beaver, squirrel, and
sometimes also bear, are usually categorized as fur animals. This categorization is almost always implicit. Here follows one of the few explicit instances: "Regarding the smaller mammals,
beaver, fox otter, pine inarten and wild cat, they are all found in site assemblages from earlier and later periods and can - archaeologically considered - be regarded as 'fur animals' killed with thispurpose." ( M a 1984:49, translation by author).
This dichotomy of animals into meat- and fur animals is an over-simplification. Many
animals are used for both fur and meat. In present day Sweden, beaver and bear are such animals.
In older sources regarding Northern Sweden, we can fmd references of lynx meat being eaten by
'upper class hunters' and the Saami, but not by the local farmers. Lynx meat was said to taste like
prime veal (Ekman 1983:121-122). It is also important to remember that animals are not used for their meat andlor skin alone. The bones can be used for tools, and the tendons for thread.
Intestines can be used as rain garments or as containers. Further, animals can be used for ritual or
medical purposes. Examples of this can be found in ethnographical sources from North America,
where it for instance is mentioned that the shamans of the Attawapiskat indians ate dog in order to
enhance the shaman's ability to locate caribou herds (Honigmann 1981:220).
At Stone Age sites in Scandinavia, a certain blurring of the-meat animal - fur animal
dichotomy can be observed. At several sites, cutmarks indicative of filleting and dismembering have been observed on bones of so called fur animals (NoeNygaard 1995, Trolle-Lassen 1987).
At Ringkloster and Agernss, two Ertebprlle sites in Denmark, an overrepresentation of newborn
red deer and roe deer calves, has been interpreted as the calves being hunted for their spotted skin
(Andersen 1998:51; Noe-Nygaard & Richter 2000).
The aim of this dissertation is to study the Mesolithic people's use of the wild canids, mustelids
and felids at the Danish Erteballe site Hjerk Nor. Were the wild canids, mustelids and felids all
solely used for their fur, or were their use somewhat more diverse?
My study will be carried out by using microscopical analysis of cutmarks on the bones of
wild canids, mustelids and felids from the Danish Ertebprle site Hjerk Nor, and considering these in the context of the assemblage of bones of these animals as a whole.

1.2. Methods
The biological aspects of the animal population, such as age, sex and size, were studied using the data and methods found in Ahnlund (1976), Degerwl (1933), De Marinis (1995), Englund (1970), HabermeN (1983, Heinrich (1991), Kratochvfl (1976), Neal & Cheeseman (1996), Ondrias (1960), Ondrias (1961), Reichstein (1984), Reichstein (1991), Schauenberg (1980),
Stubbet Krapp (1993a1, Stubbe& Krapp (1993b), TroUeLassen (1986), and Zeiler (1987). For estimation of age, tooth eruption and epiphyseal fusion were used.
The bones from the wild canids, mustelids and felids were studied in microscope with 6-50 times magnification. They were measured according to von den Driesch (1976) and De Marinis (1995), and the position of all cutmarks were drawn on a picture of the relevant bone (see appendix 4). Articulated bones were not specifically looked for, due to lack of time.
The results from my study were compared with other cutmark analyses of wild canid, mustelid and felid bones from Mesolithic and Neolithic sites in Nortfi-Western Europe. These
sites are Hazendonk (Zeiler 1987), Kongemose (Noe-Nygaard 1995), Muldbjerg I (ibid), Priestelyng (ibid), Swifterbant (Zeiler 1987) and Tybrind Vig (Trolle-Lassen 1985; TrolleLassen
1987) (see figure 1.2.). The Latin terminology for bone anatomy follow Kompendium i osteologi by Kirsten H.
Schauser and Vibeke Dantzer (Schauser & D a n k r 1995). A short guide of Latin terminology can be found i appendix 2.
1.3. Cutmarks
1.3.1. Defiiition of cutmarks, and of some other man-made marks on bones
In this work, the definition of cutmarks and other man-made marks follows Noe-Nygaard (1995). A cutmark is defined as a more or less straight groove with a V- or U-shaped cross- section (see figure 1.l.a). Cutmarks originating from skinning are usually fairly superficial, whereas cutmarks originating from dismembering are more deep and pronounced. Cutmarks can
also stem from filleting (see chapter 1.3.2.) (NoeNygaard 1995:180-181). Cutmarks are not the only kind of man-made mark that can be found on bone. Other marks
are scrape marks, sawing marks and chopmarks. None of these were, however, found in the Hjerk Nor assemblage. A scrape mark derives from the removal of meat by filleting, and usually
appears as a series of shallow subparallell grooves (see figure 1.l.b). Sawing marks occurs when the flint tool is being sawed on the bone. This may occur at attachment areas with particularly strong tendons, or on bones that have been deliberately prepared to break at a certain point. The cross-section of sawing marks has terraced walls and crossing striae in the uneven bottom (see
figure 1.l.c). Chopmarks are deep grooves that are tilted in the groove direction, and with a well-

Figure 1.1. a) Cross-section of cutmark; b) view of scrapemark; c) cross-section of sawing mark; d) cross-section of
chopmark (Noe-Nygaard1995:180-182).
defined border. A cross-section of a chopmark looks like a tilted 'v', with one smooth side and
one side with an irregular surface and border (see figure 1.l.d) (NoeNygaard 1995:181-182).
In order to discern these traits, one can either use a microscope or a scanning electron
microscope (SEM). The SEM gives a much higher resolution and depth of field, as well as the
possibility of extreme magnification. The drawbacks are, however, that one must use casts of the
bones, as the vacuum chamber of the SEM is small, and that all non-conducting samples must be
coated with gold in order to create conductivity. These procedures make the SEM a rather time
consuming method (Shipman 1981:360-362), and it is rarely used for large quantities of
archaeological samples.
1.3.2. Interpretation of cutmarks
There are two ways by which cutmarks have been interpreted: through ethnografic parallells and
through experiments. Most studies have concerned larger animals, such as reindeer or red deer,
and may therefore occasionally be of limited use in the interpretation of cutmarks on small
animals. The possibly most wideknown ethnographic study of cutmark interpretation is Lewis
Binford's study of the Nunamiut eskirnos in North Central Alaska (Binford 1978; Binford 1981).
During four years (1969-1972) he studied their treatment of butchered caribou, i.e. all stages from
primary butchering to extraction of bone grease. Binford argues that cutmarks are best interpreted
through their placement on the bones (White 1992:145, in Magnell 1996:2). Cutmarks deriving
from skinning are argued thus: "There are actually very few places on the anatomy where the manipulation of the skin brings the butcher in direct contact with bone. The two places where this is most likely are the lower legs and the head." (Binford 1981:106-107). Cutmarks deriving from
dismemberment are associated with points of articulation, such as the atlas-cranium joiut, the
femur-pelvis joint, and the humerus-radius-ulna joint. Cutmarks deriving from filleting are found
at the vertebrae, the pelvis and the leg bones. Such cutmarks are either long ones placed longitudinally on the bone, or shorter ones placed more obliquely. The longitudinally ones ate the
primary cut in order to expose the bone from which the meat is to be removed. The shorter ones
occur when the meat is being removed from the bone (Binford 1981:33,90, 106-107, 126-130).

Binford's detailed descriptions of the placing of cutmarks, as well as breakages occuring thorugh marrow processing, have been much used by osteologists and archaeologists. NoeNygaard describes the use of different methods for cutmark interpretation thus: "Modern hunting analogies, well described by Binford (1978, 1981), are invaluable in the interpretation of the
processes that form the various types of marks. Modern butchering eqeriments on red deer, roe deer, hare, badger, and several species of bird in combination with information from the literalure, make it possible to undertake detaifed mterpretaiions of the origin of the various marks and to aid in distinguishing between systematically placed Upes of marks and ra~ldomly
occurring unimportant marks. (NoeNygaard 1995:243). One important study concerning skinning and butchering experiments on smaller animals
was canied out by Tine TrolleLassen, as part of her Master of Art dissertation at the Institute of
Prehistoric Archaeology at the University of Aarhus, Denmark. She skinned a stone marten, and compared the cutmarks with those found on the pine marten bones at the Danish Mesolithic site Tybrind Vig. As both the recent stone marten and the Mesolithic pine martens showed almost the same placing of cutmarks, i.e. on the pubic bone, on the corpus mandibulae, and on the snout, she could conclude that the pine martens at Tybrind Vig had been skinned using open skinning (see figure 7.1.), and that the nose had not been included in the fur. The difference was the cutmarks on the phalanges. They had not been found in the Tybrind Vig material, and it was thus concluded that the claws had been excluded from the fur. In modem skinning of martens, and as Trolle-Lassen had done, the claws are included in the fur. With this procedure, cutmarks on the phalanges will occur (Trollelassen 1985:Appendix VI).
Troll-Lassen also skinned and butchered an otter, and ate the meat. As with the stone marten, the cutmarks on the otter showed great similarity in placing to the ones found in the Tybrind Vig assemblage. The skinning cutmarks were found at the same places as the ones on the stone marten. Cutmarks derived from dismembering were found at collum femorir, distally on the humerus and the radius, as well as proximally on the radius and the ulna. The dismembered limbs were cooked for 2-3 hours. The meat was described as tender, but somewhat stringy. The taste was considered to be rather bland, and not as gamy flavoured as hare or roe deer (Troll-
Lassen 1985:Appendix VII).
1.4. Hjerk Nor
Hjerk Nor is a kitchen midden situated on the western part of the Limfjord in Jutland, Denmark (see figure 1.2.). In ca 1937, bones and artefacts were collected on a low tide by an amateur archaeologist. No proper excavation was made, and it is not known whether the gathered finds stemmed from a concentrated area of the kitchen midden or if they were scattered. Nor do we know if the collection was a thorough one, or if many finds were left at the midden. The finds
were kept at the local museum in Skive, when zoologist Magnus Degerbal noted them on a visit

to the museum in 1944. Upon his request, the osteological material were sent in the following year to the zoological museum in Copenhagen to be deposited there and analysed. The analysis was only extended to determination of species. In 1973, an article was published in Kaskelot, a danish biology magazine (Hatting et al 1973). Upon reading this article, Skive Museum informed Zoologisk Museum that they had some more bones from Hjerk Nor, which they could deposit at Zoologisk Museum. These bones, called Supplement 1973, were also determined to species. No further analyses have been made on this material. It is typologically dated to the Ertebprlle culture (Hatting et al 1973: Simonsen pers. comm.; excerpt from the records of Zoologisk Museum).
Hjerk Nor has received brief attention in various textbooks on Danish prehistory, due to the abundanceof bones from 'fur animals' (Andersen 1981:95; Hatting 1972:154). Of the 651 bones found, 310 (48 %) came from wild canids, mustelids and felids (see table 1.1.). Such high amount of 'fur animal' bones is only surpassed by one other Danish site: Tybrind Vig (see chapter 1.5.6.). Most Mesolithic sites in Southern Scandinavia contain ca 5 % 'fur animal' bones. A few sites reach ca 20-25 %.
The Hjerk Nor bones of wild canids, mustelids and felids are in a remarkably good condition (see table 1.2.), and are thus suitable for a microscopic analysis. Further, no traces of
burning have been found on the bones, and only one bone displayed gnaw marks. Over two thirds of the bones are complete.
Y - 1 ) Hjerk Nor 2j ~azendonk 3) Kongemose 4) Prrestelyng 5) Muldhjerg I 6) Swifterbant 7) Tybrind Vig
Figure 1.2. Map of Southern Scandinavia and the Netherlands, wifh Werk Nor and the comparative sites marked.

Number of Species Fragments Aurochs 87 Red deer 123 Wild boar 98 b 18 Fox 14 Badger 9 Pine marten 27 Polecat 1 Otter 40 Lynx 12 W~ld cat 207 Whale unspec. 8 Swan unspec. 6 Osprey 1
,Total 651
Whole bone well preserved: Whole bone moderatety well preserved: Whole bone pooriy preserved: Bones predominately well preserved,
but with some parts moderateiy well preserved: Bones predominately well preserved,
but with some parts poorly preserved: 20.0% Bones predominately moderately well preserved,
but with some parts poaiy preserved:
Table 7.2. Condition o f the bones.
Well preserved = c 5 % o f surface spotwise eroded. Moderately well preserved = 5-40 % of the surface
spotwise ercded. Badly preserved = > 40 % of the surface eroded.
Table 7.1. Distribution of species
and number of hagments at Hjerk Nor.
1.5. Comparative sites
1.5.1. Hazendonk
Hazendonk is a Neolithic site in the South-Western part of the Netherlands (see figure 1.2.), then
situated on a river bank. The site has been radiocarbondated to 3400-1700 B.C. It was not a
continuous settlement, but consisted of nine separate phases of habitation. A considerable part of
the bone material consists of 'fur animals', mostly beaver and otter (Zeiler 1987:245-246).
1.5.2. Kongemose
The Kongemose site, the archetype for the Kongemose culture, is situated on North-Western
Sjaelland in Denmark (see figure 1.2.), in the bog Amosen, a former lake. In 1955,350 m2 was
excavated. Kongemose is radiocarbondated to 5600 F 120 b.c. Roe deer, red deer and wild boar strongly dominate the animal material. After the fairly common beaver, other animal species are
only found in a few fragments (J0rgensen 1956:25; NoeiNygaard 1995:68,78).
1.5.3. Muldbjerg I
Muldbjerg I is, like Kongemose, situated in Amosen in Denmark (see figure 1.2.). As one of the aims of the excavation was ta do a total excavation and to reaieve everything connected to the site,
close to 1400 m2 was excavated during 1951-1966. Muldbjerg I is radiocarbondated to 2830 k
6

100 b.c., i.e. Early Neolithic. It is believed that the site was populated one or two summers only, and functioned as a stationary hunting camp. After roe deer and red deer, water vole, beaver and
otter are the most common animals found in the material. It might be tempting to regard the water vole as being an intruding species, whether Neolithic or recent. 'Ibis would however be a mistake, as cuanarks from skinning have been found on 17 water vole bones, showing that at least some
of these rodents wereused by thehumans at the settlement (NoeNygaard 1995:65,76, 143,230- 231).
Virtually nothing has been published about the Praestelyng site. It is a Late Erteballe site, also situated in Amosen in Denmark (see figure 1.2.). It is radiocarbondated to 3100 + 80 bc. The animal remains were in 1969 subject to a prize dissertation by Nanna Noe-Nygaard (Noe- Nygaard 1969). Of the animal bones, roe deer is by far the most common fmd. The bones of 'fur animals' (i.e. wild canids, mustelids, felids, beaver and squirrel) contribute to a mere 1.3 % of all mammal fragments (NosNygaard 1995:77,69).
1.5.5. Swifterbant
The Neolithic site Swifterbant was situated on a low-lying levee close to a creek, in the Central Netherlands (see figure 1.2.). Swifterbant does actually consist of four sites - Swifterbant 2, 3 ,4 and 5 - but no separation of the finds has been made in the articles. The sites were excavated in the 1970s, and are radiocarbondated to 3400-3300 B.C. A considerable part of the bone material
consists of 'fur animals', mostly beaver and otter (Clason & Brinkhuizen 1978:69-75; Zeiler 1987:245-246).
15.6. Tybrind Vig
Only one more Mesolithic site in Southern Scandinavia show a similar high amount of fur animal bones as Hjerk Nor: the Erteballe site Qbrind Vig. Tybrind Vig is a submerged site just outside the west coast of Funen in Denmark (see figure 1.2.). It was excavated in the 1970s and 1980s, and a huge amount of organic material was found, ranging from human and animal bones to wooden boats and paddles. More fragile organic material, such as fish traps and fragments of textiles were also found. Tybrind Vig is radiocarbondated to 3700-3200 b.c. An osteological analysis of the animal bones found between 1978 and 1982 were presented as a MA dissertation
by Tine TrolleLassen in 1985 (TrolleLassen 1985). Her studies emphasized butchering and

cutmarks on red deer and 'fur animals' (Andersen 1985; Trolle-Lassen 1985). The mammalian part of the osteological material is dominated by pine marten, followed by
red deer and wild boar. The bones of pine marten, and of other carnivores, were in almost all cases found in clusters, each representing an individual animal. This procedwe would indicate complete bodies thrown out into the sea (Andersen 1985:57; TrolleLassen 1985:44-128).
2. SETTLEMENT PATTERNS AND ECONOMY IN SOUTHERN SCANDINAVIA DURING THE ERTEBPILLE PERIOD
In Southern Scandinavia, both coastal and inland Ertebplle sites have been discovered. As Denmark is tilting on a NW-SE axis (see figure 2.1.), coastal sites southwest of this line are now submerged. Therefore, they have not been subject for excavation at the same frequency as coastal sites elsewhere in Southern Scandinavia. It is generally believed that during the Mesolithic, especially in the two earlier periods: Maglemose (9500-6800 B.C.) and Kongemose (6800-5500 B.C.), people were migrating seasonally between inland and coastal sites. Specialised hunting sites were also used regularly according to season (Andersen 1995:42; Burenhult 1999:184-187, 199).
This seasonal migration pattern may, however, not be generally true for the Ertebplle period (5500-4100 B.C.). There are basically two facts that imply this hypothesis. One was expressed by Nanna NoeNygaard, in her analysis of 613C tests on canine and human bones from Danish Mesolithic and Neolithic inland and coastal sites. She found that dwing the Mesolithic, human and canine bones from inland sites displayed typical terrestrial 6°C values, whereas bones from coastal sites displayed marine values. During the Neolithic, bones from both inland and coastal sites displayed terrestrial 6'" values (NoeNygmd 1988:88-91). As the turnover ratio of 6I3C in
human bones is 5-20 years (Lidh 1995:16), NoeNygaard's study implies that there was little or no seasonal migration between inland and coastal sites.
Another, more widely recognised indicator of the cessation of seasonal migration, is the emergence of what is assumed to be permanent settlements. Often, the inland settlements tended to become smaller and more seasonal of nature, whereas coastal settlements became larger and more permanent of nature (Larsson 1987:168). It is believed that this development has been connected to an increasing use of a 'delayed return'-system, i.e. the
use of complicated andlor stationary trapping equipment, and possibly also forest clearances. Such work intensive investments increased the confinement to the territory, Figure 2.1. The NW-SF tilting axis of
and in that way led to more permanent settlements Denmark(Aaffi-S~rensen 1988:179).

(Burenhult 1999:218-219). In the Lif jord region, in Northern Jutland, where Hjerk Nor is situated, inland settlements
are not likely to have existed. As the coastline is very jagged, with many long fjords and small
land areas 'untouched' by the se& the region has not enough inland areas to support both a coastal and an inland population. Instead, people lived at the coast, and used inland hunting sites for the
hunting of large forest dwelling game. Smaller coastal sites were also used for the hunting of seals, whales and seabirds (Andersen 50-51).
The most important game animals on virtually all Mesolithic sites in Southern Scandinavia were red deer, wild boar and roe deer. At some sites one of these species dominate completely, but this should not be automatically interpreted as 'specialised' hunting sites. It could simply mean that the environment favoured one species more than the two others. It is likely these three
species, together with fish (see below), that were of most economic importance (Magnell pers. comm.).
Bird bones are found on most sites. The number of fragments varies greatly. Usually, many species are found, but with few bones identified to each species. This may reflect a great taphonomic loss of the fragile bird bones, but it may also suggest birds being of lesser use, and that this use seldom was connected to a certain species. Some sites, such as Aggersund, have a great number of bones from certain species, and have been interpreted as specialised seasonal bid-hunting sites (M0hl1979).
On coastal sites, marine subsistence were of great importance. Unfortunately, as fish remains are easily destroyed, we may not really grasp the importance of this resource. Soren H. Andersen means that the large number of species found at the Late Mesolithic sites is an indication of the great importance of fishing (Andersen 1995:52). Inge Bprdker Enghoff is of the opinion that the large number of species, often coastal species, reflect the fishing methods. On many sites, stationary fish traps have been excavated. Fish hooks are less common. It is therefore likely that fish were mostly caught in stationary fishing equipment, such as fishtraps and nets, that were used close to the coast (Enghoff 19939 16; Enghoff 1995:72).
The small number of seal and whale bones indicate that the hunting of sea mammals are likely to have been of lesser importance than fish and terrestrial animals. The regular frequency of seal, even at small sites, indicates that seals were likely to have been an important resource, not only for meat and skin, but also for blubber and oil. Low, elongated ceramic bowls, so called oil lamps, have been found at several Mesolithic sites. Some have been chemically analysed, and were proved to have contained animal fat (Andersen 1995:53).
Few researchers have to any greater extent discussed the role of vegetables in the Mesolithic
economy. Hazelnuts are found at almost any site containing organic remains, and their contribution to the diet has been a well-known factor since the beginning of the 20th century. At the Danish site Holmegaard, a cache of water-lily seeds was found. The seeds were interpreted as "a very important supplement to the animal diet. " (Jessen 1924:19, translation by author). Such fmds of plants m greater quantities at sites are rare, and this is likely the reason that vegetables are

more or less ignored in analyses of economy of Mesolithic sites. Nevertheless, most
archaeologists do believe that vegetables formed a large part of the human diet during the
Mesolithic. They are, however, unsure of how to prove it.
3. THE ENVIRONMENT IN SOUTHERN SCANDINAVIA DURING THE ERTEB0LLE PERIOD
During the so called Atlantic chronozone (8000-5000 BP), the postglacial climate optimum
occurred. The average temperature in Southern Scandinavia during the Ertebolle period was ca 2 - 4°C higher than the average temperature in the region today. The Ertebolle climate can be
compared with the climate of North-Eastem France, i.e. warm summers and mild winters. Snow
was likely to have been a rare occurrence. Botanical remains indicate that the climate had a maritime character, with fairly high rainfall. (Liljegren & Lager& 1993:32).
The forests were dominated by decidious trees, mostly oak, elm, ash and lime. The forests
were rather thick, and subsequently dark, but mixed with lighter glades where various grasses,
herbs and bushes such as hazel, rowan and rosehip grew. The thickening of the forests led to the decrease of animals that mainly fed on grass, for example aurochs (Lijegren & Lageris 199332-
33). The larger animals that dominated the fauna were red deer, roe deer and wild boar. Seas and
skeams provided ample habitat for beaver and otter, as well as for several kinds of birds (Aaris-
Sorensen 1988:189-190, 196). The marine transgressions created coastlines filled with bays and
lagoons, where the marine fauna flourished (Burenhult 1999:218).
3.1. The environment in the Limfjord region
Little botanical remains have been excavated from the Mesolithic settlements in the Limfjord region. A single charred hazelnut shell was found at Ertebolle. However, analyses of charcoal
found at the sites, indicate a landscape covered with decidious forest, dominated by oak, hazel,
aspen, willow, elm and birch (Andersen 199545; Andersen & Johansen 1987:53). The lack of
pollen analyses renders it difficult to detect other plants than trees. Also, it is not possible to
discern the distribution of forest in relation to open land.
Due to the shortage of botanical analyses, we must therefore also look to the faunal
remains. Seven Ertebolle sites in the Limfjord region contain animal remains: Aasted,
Aggersund, Bjomsholm, Ertebolle, Hjerk Nor, Krabbesholm and Virksund (see figure 3.1.).
Bjomsholm and Ertebglle are considered to be permanent settlements, whereas Aggersund is
interpreted as a swan hunting site, probably occupied at late autumnlwinter (Andersen 1995:51;
Mohl 1979). Aasted, Krabbesholm and Virksund are kitchen middens where bones and artefacts
were collected in the 19th century. They have not been further excavated, and no analyses apart

1) Hjerk Nor 2) Aasted 3) Aggersund 4) Bjarnsholm 5) Ertebnlle 6) Krabbesholm 7) Virksund
Figure 3.1. The Limfjord region during the Erteblle period, with some known Ertebnlle sites marked.
from determination of species have been made (Winge 1904:205). The species found at these
sites are listed in table 3.1., together with the number of fragments of each species.
Of the eighteen species of mammals found, nine are connected to forests. The red deer, roe
deer, elk, wild boar, badger, pine marten, wild cat, lynx and squirrel all live in forests, preferrably
mixed with open terrain. The wolf and fox are more adaptable, and will live in almost any kind of
landscape. The polecat prefers moist lowlands with both forests and open terrain. The otter lives at
water of all kinds: streams, beaches, lakes. The seals and whales live in sea water. Seals live in the
proximity of land, whether archipelagos, beaches or solitary islands, where the cubs are born, and
where they often lie resting during the day. Whales live in open water, but may occasionally swim
fairly close to shores (Bf&vall& Ullstr6m 1995:80, 147, 152,173,178-180, 188, 190, 194, 198, 200-207,214,224,230,238,252,258-283).
A majority of the bird species are connected to the coast. Two, however, live exclusively in forests: the capercaille and the ural owl. Some of the coastal birds, for example the osprey and the
red-necked grebe also live at inland seas (Bmun & Singer 1975:20,22,30,44,46,52-54,62,72, 82,96, 120,122-124, 142-150, 174,262-264).
The fish remains show a similar picture. Of a total 28 identified fish species, five are
freshwater fish and eighteen are seawater fish. Five species can be caught in both fresh- and
seawater (Andersen & Johansen 1986:60; Enghoff 1991:107). Of the above discussed habitat preferences of the animals found at the sites, it can be
deduced that the environment at the Limfjord in the Ertebplle period mostly consisted of
deciduous forest, albeit not a very dense one. mere must have been many open areas within
forest glades and at the forest margin. Rivers ran from some smaller inland lakes and bogs into
the Limfjord. In the many fjords and bays seals and otter could be found, as well as several
species of waterfowl.

Spccles Hiark Nor i Aqrrrrrund ! Bi.rnsholm I Ertcb.lle 1 Aasred 1 Knbberholmi Virksund
9.k ism-& . 87 i.i.i.i.iiiiii.i.i.ii ..i.i.i.i.i.i.i..i. .. - ? L. ..... t ........ Red deer jcem cAphur 123 i 40 ! 103 + i +
. . . .........-.. -kd~.c ;!?PLSE.EW.E . . . . ..-.- - L ......... ? ............ i ! .. . . ....... ....... i .- t Elk : l e e s aECI I f
......... ......... ....... E!!..k:: -SE..'F~~ 98 iiii.....!!. ?. i..i.i.i~B.B.B.BB. orS iCanir hmilkrk 18 i ; 5
.won ............................ :~e.~*.... ................. i ! .? - ........ FQX : V W s w&s 14 i I S i 15
Table 3.1. Number of fragments of species found at six Ertebnlle sites at the Limfjord. + indcates an unknown
number of fragments. According to Bratlund (pers. comm.), the Erteb~lk material mostly consists of red deer, roe
deer and wild boar. Ail other mammals are represented with 1-3 fragmens.
Sources: Hjerk Nor: Hatting et al 1973:17 Aggenund: Mehl1979 Bj~msholm: Bratlund 1993:lOl; Enghoff 1993:107 Ertebelle: Andenen & Johansen 1986:59-60 Aasted, Krabbeshdm and Virksund: DegeM 1933:407,424,435,477, 533, 557, 582; Winge 1904:205

Table 4.7. Distribution of skeletal elements on wild canids, mustelids and fefids. The numbers within parentheses indicate the number of bones with cutrmrks.
Cranium Mandible Teeth Vertebrae Scapula Humerus Radius Ulna Metacarpals Pelvis Femur Tibia Fibula Caicaneus Metatarsals Penis bane
Total
4. THE WILD CANIDS, MUSTELIDS AND FELIDS OF HJERK NOR
Fox Badger Pine marten Polecat Otter Lynx Wild cat 4 3 (2) 1 6 (2) 7 (3)
2 (2) 1 2 (1) 1 (1) 12 (2) 4 1
1 6 (2) 2 5(1) 5 (4) 2 (1) 34(29) 1 2 3 3 (1) 23 (9)
4 (1) 3 (2) 25 (13) 1
1 6 ( 5 ) 12(10) 2 2 9 5 (4) 33 (1 71
3(1) 5 (1) 6 (1) 4 0 ) 2 34 (1 2) 2 (1) 2 (1)
1 13
1
14 9 27 1 40 12 207 -
Twelve animal species were identified in the Hjerk Nor assemblage. Another two were identified
to family or order (see table 1.1.).
As the site was not properly excavated, many smaller bones are missing (see table 4.1.).
This makes it difficult to study whether there was a similarity in the details of the skinning
methods on the different sites. The lack of claws in an assemblage, or the presence of cutmarks
on them, can indicate whether the claws were left on the skinned carcass, or remained as a part of
the fur. At Tybrind Vig, it was found that the claws of wild cat and otter were part of the fur (TrolleLassen 1985:77; TrolleLassen 1987:88), whereas the claws of pine marten were left on
the carcass (see chapter 1.3.2.). At Svaerdborg I, an almost intact wolf skeleton was found - lacking the claws (Aaris-Sorensen 1976:141). However, many other aspects of the utilization of
the animals at Hjerk Nor can be deducted. Such aspects will be discussed in the following
chapters and subchapters.
4.1. Fox (Vulpes vulpes)
The fourteen fox bones all belong to adult foxes, i.e. older than one year [Englund 1970:6,
Habermehl 1985:115-116). A minimum number of four individuals was found.
The sexual dimorphism of foxes show a large overlap (Heinrich 1991:84-85). A comparison with measurements of recent foxes (Reichstein 1984, in Stubbe & Krapp

Figure 4.1. Skull of assumed tame fox from Hjerk Nor (Hattfflg et a1 1973:76).
1993a:153), shows, however, that the
greatest length of humerus, radius and
femur of the Hjerk Nor fox remains are
a l l within the range of male foxes. The
two tibiae are both within the overlap
between male and female (see table 4.2.).
Unfortunately, it has not been possible to
compare measurements from the skulls, in order to determine sex of these individuals.
The above reasoning regarding sex
estimation is of course only valid if the size Table 4.2. Greatest length of fox limb bones from Hjerk
of foxes and the overlap in sexual Nw and comparative materiak. All measurements in mm..
dimorphism have not changed significantly Figures by Reichstein (7984).
since the Mesolithic.
One cranium and two mandibles articulate. This fox's head is of some interest, as it has
been interpreted as a captive fox, due to the lack of sagittal crest, and the heavily worn teeth (see
figure 4.1.) (Hatting et a1 1973:16; cf Degerbldl 1933:433-438). Such heavy wear is normally only
observed in old captive foxes (Jonsson 1988:67). The fox may have been kept as a decoy or as a
pet fox. If it was a female, it may have also been kept for breeding purposes, in order to secure a
steady supply of fox furs. As the fox is likely to have been rather old, it was probably not kept for
its own fur. Similar finds of captive foxes have been noted at the Danish Maglemose site
Holmegaard, and in the Swedish Elteballe site Skateholm (Degerbol 1933:433-438; Jonsson
1988:67). One tibia displayed canine gnawing marks on the proximal and distal ends. It was not
possible to determine wh&er the bone was gnawed by a dog or a fox.

4.2. Badger (Meles meles)
I Nine badger bones were found among , the remains. Age determination of I
badgers using epiphyseal fusion is I
somewhat difficult, as only the fusion of tibial epiphyses have been
correlated to a certain age. The distal epiphysis fuses during the badger's first winter, at Ca ten months of age. Table 4.3. Comparison of measuremerrts of pine marten
The proximal epiphysis starts to fuse limb bones. AN measurements in mm. Figures by
during the badger's second autumn ondrias(1961:zrs-277).
and winter, and the epiphyseal line is still visible during the following spring. A completely fused tibia does therefore belong to a badger of at least two years of age. As the proximal epiphysis of the tibia is the last to fuse of the limb bone epiphyses, any unfused limb bones will derive from badgers younger than two years (Ahnlund 1976:120). Two tibiae derived from badgers under ten months of age, one tibia from a badger of ca two years of age, and two tibiae from badgers over two years of age. One femur was unfused proximally and distally, and therefore estimated to derive from a badger under two years of age. A minimum number of four individuals was found in the assemblage; three adults and
one juvenile. The sexual dimorphism in badgers is rather small, and overlaps between the sexes (Neal &
Cheeseman 1996:20; Ondrias 1960:578). No adequate reference material regarding measurements has been found, and I choose therefore not to make any estimation of sex for the Hjerk Nor badger remains.
4.3. Pine marten (Martes martes)
Of the twenty-seven pine marten bones found in the assemblage, only three belonged to juveniles. When compared to bones of pine martens and stone martens of known age, it was estimated that these juveniles were under four months of age. The adult bones belonged to pine martens older than one year (Trolle-Lassen 1986:120-122). A minimum number of seven individuals was found; four adults and three juveniles.
The pine marten show a sfrong sexual dimorphism (Reichstein 1991:52; Stubbe & Krapp 1993a:394). A comparison of the greatest length of humeri and radii between the Hjerk Nor assemblage and recent Danish pine martens (Ondrias 1961:311, 315, 317), indicate that a majority of the Hjerk Nor pine martens were males (see table 4.3.). Unfortunately, no comparative measurements could be found for femur and tibia. As the sample is rather small, it

must be emphasised that the suggested sex ratio may not be representative of the whole - unexcavated - assemblage.
4.4. Polecat (Mustela putoris)
The polecat is represented by a single adult cranium. The cranial measurements, i.e. condylobasal
length and rostrum widthhpper canine length, suggest the polecat being male (see table 4.4. and figure 4.2.) (De Marinis 1995:6 1; Reichstein 1991:52-53).
It is possible that the cranium was not the remains of a local hunt, but did instead serve as a decoration on clothing or on equipment, that was later discarded on the site. Such decorations - possibly of totemistic significance - have been found on skeletons at the Erteballe gravefield at Skateholm, Sweden (Larsson 1988:139-143).
Table 4.4. Condylobasal length of polecat from Hjerk Nor and compaative materials. All measurements in mm.
Figures by Reichstein (1 99 152-53).
I 1.1 I
12 19 (4 15 16 IT 18 1D
ROSTRUM WIDTH
MALE8 O FEMALES
Hjerk Nor cranium
Figure 4.2. Rostrum width/upper canine length of the Hjerk Nor polecat and recent Italian pokcats.
All measurements in mm (De Marinis 7995:61, with addition by author).

Table 4.5. Greatest length of otter humeri from Hjerk Nor and comparative materials.
All measurements in mm Figures by Ondrias (7961:3 75) and Degerbd ( 1 933584-585).
Table 4.6. Greatest length of otter radii from Hjerk Nor and comparative materials.
AM measurements in mm. Figures by Ondrias (1961:317) and Degerbal(1933:584-585).
45. Otter (Luau lutra)
Of the forty otter bones, thugr-one were from individuals of 2-3 years or older, six were from
individuals of 1-2 years of age, and one came from an animal younger than 1-2 years. Two bones
could not be aged (Habermehl 1985137; Zeiler 1987:250). A minimum number of eight
individuals was found; six adults and two juveniles.
Measurements of humerus and radius indicate an over-representation of males in the
assemblage. Four bones were within the range of males, whereas the remaining one was within
the range of females according to both Ondrias' and Degerbol's studies of Danish otters (Ondrias
1961:311,315,317; Degerbal 1933584-585) (see table 4.5. and 4.6.). The find of a penis bone
confirm the presence of at least one male in the Hjerk Nor assemblage. As with the pine marten
(see chapter 4.3.), the lack of comparative measurements for femur and tibia makes this sex ratio
somewhat uncertain.
4.6. Lynx (Lynx lynx)
Of the twelve lynx bones, four derived from adults, and five from juveniles. Three bones could
not be aged. A minimum number of three individuals was found; one adult and two juveniles of
different ages.

Table 4.7. Greatest length of lynx limb bones from H W Nor and a comparative material.
AN measurements in mm. Figures by Larje (pers. comm.).
Three bones, a humerus, a radius and an ulna, articulate. These three bones were the only
ones where the greatest length could be measured. This lynx was a fairly large individual, which
could indicate it being a male. According to S tubbe and Krapp (1993b:1136), the lynx show great
overlaps in its sexual dimorphism. However, a small sample of Swedish lynx show no overlaps
at all between the sexes (Larje pers. comm.). When compared to this Swedish sample (see table
4.7.), the Hjerk Nor lynx is well within the range of males.
4.7. Wi cat (Felis silvestris)
In the Hjerk Nor assemblage, 207 bones of wild cat was found. Ageing wild cats from
archaeological assemblages are rather difficult, as little detailed information have been published. The dental eruption f ~ s h e s at nine months of age (Stubbe & Krapp 1993b:lll I), and the limb
bones are a l l fused at 18 months of age (Schauenberg 1980:553, in TrolleLassen 1985:75). From
this it can be deducted that in the Hjerk Nor assemblage, 19 crania and mandibles belonged to cats
older than nine months, and that 26 limb bones belonged to cats under 18 months of age. There
were 113 fused or fusing limb bones in the assemblage, which would give a ratio of 14:3,
regarding fused - unfused l i b bones. A minimum number of 19 individuals was found. This
included twelve adults, four juveniles and three individuals of unknown age.
The wild cat, as well as the domestic cat (Felis catus), show a considerable overlap in the
sexual dimorphism (Johansson & Hiister 1987:22-23; Kratochvil 1976:166). Measurements of
the limb bones of the Hjerk Nor wild cats show an ordinary curve (see figure 4.3. and 4.4.),
indicating a normal sex distribution within the population. It has therefore only been possible to
estimate sex for a few bones. As these bones are of a great minority, no sex ratio has been
estimated.
A comparison of measurements of the wild cat bones from Hjerk Nor with wild cat bones
from Erteb0lle and Meilgaard (see figure 4.5.-4.8.), show that there is a great similarity in size

I rnm
IA8.0 110.0 112.0 114.0 116.0 118.0 120.0 122.0 124.0
Figwe 4.3. Greatest length (GL) and greatest distal breadth (Ed) of wild cat humeri from Hjerk Nor.
L A n m 128.0 130.0 132.0 134.0 136.0 138.0 140D 10.0 1M.O 146.0 148.0
Figure 4.4. Greatest length (GL) and greatest distal breadth (Ed) of wild cat tibiae from Hjerk Nor.
I Hjetk Nor 63 Erreballe O Meilgaard
.nm 1W.0-111.9 111.0.ll4.~ 11.0-111.1 11..0.l20l 121,01219 11.0-1-9
I Hjerk Nor kSl Erteballe 0 Meilgaard
Figure 4.5. Greatest length of wild cat humeri F&e 4.6. Greatest drstal breadth of wild cat humeri
from Hjerk Nor and ErtebnIe. from Hjerk Nor, Ertebnlle and Meilgaam!
8 B
Hjerk Nor I 7 I Erteb~lle 69 Erteb~lle 6 D Meilgaard 6
5 5
4 4
3 3
2 2
1 1
0 0 ".m
Figure 4.7. Greatest length of wild cat radii Figure 4.8. Greatest distal breadth of wild cat ndii
from Hjerk Nor and Ertebslle. from Hjerk Nor, Ertebnlk and Meilgaad
19

and robusticity in the populations. This would indicate a fairly homogenous wild cat population in
Northern Jutland. There are, however, few measurements from Meilgaard, and still fewer
complete long bones from ErtebBlle, which adds an uncertainty regarding the correctness of the
homogenity.
5. CUTMARKS ON THE BONES OF WILD CANIDS, MUSTELIDS AND FELIDS FROM HJERK NOR
5.1. Cutmarks on the fox bones
Of the twelve fox bones, three showed cutmarks. These are two mandibles from the same
individual, and one tibia. The skull that fits with the mandibles, showed no cutmarks. The lack of
cutmarks can, however, be due to insufficient magnification during the analysis. As the skull was
so large, it was not possible to reach any depth of field, except on very low magnification. The
distribution of cutmarks can be found in figure 5.l.a-b.
5.2. Cutmarks on the badger bones
Of the nine badger bones, a single tibiia showed cutmarks. The cutmarks were placed on the lateral
distal part of the tibia shaft (see figure 5.2.). They were however regarded as 'somewhat
uncertain'.
Figure 5.1. Cutmarks on fox bones from Hjerk Noc a) mandible, b) tibia.
20
Figure 5.2. Cutmarks on
badger tibia from Hjerk Nor.

5.3. Cutmarks on the pine marten bones
Of the twenty-seven pine marten bones, five showed cutmarks. These bones are two crania, one
mandible, one humems and one tibia. The distribution of cutmarks can be found in figure 5.3.a-d.
5.4. Cutmarks on the polecat bone
No cutmarks were found on the polecat skull. This may be due to insufficient magnification, as
the size of the skull made it impossible to reach any depth of field, except on very low
magnification.
5.5. Cutmarks on the otter bones
Of the forty otter bones, nineteen showed cutmarks. These are two crania, one mandible, four humeri, one ulna, five pelves, four femora, one tibia and one fibula. The cutmarks occurred most
often on the humeri, pelves and femora, where all but one bone of each k i d showed cutmarks.
The distribution of cubnarks can be found in figure 5.4.a-h.
5.6. Cutmarks on the lynx bones
Of the twelve lynx bones, four showed cutmarks; a humerus, a radius and two ulnae. Three of these bones, the humerus, the radius and one of the ulnae, articulate (see chapter 4.6.). The distal
cutmark on the humerus and the proximal one on the ulna do not derive from the same cut, as
they are of different depth. The distribution of cutmarks can be found in figure 5.5.a-c.
Figure 5.1. Cutmarks on pine marten bones from Hjerk Nor: a) cranium, b) mandible, c) humerus, d) tibia.
21

Figure 5.4. Cutmarks on otter bones from Hjerk Nor: a) cranium, b) mandible, c) humerus, d) ulna, e) pelvis,
f ) femur, g) tibia, h) fibula.
Figure 5.5. Cutmarks on lynx bones from Hjerk Nor: a) humerus, b) radius, c) ulna.
22

5.6. Cutmarks on the wild cat bones
Of the 207 bones of wild cat, 98 showed cutmarks. They are found on virtually all skeletal
elements (see table 4.1.). The elements most frequently represented are humerus and pelvis, where over 85 % of the bones show cutmarks. Other elements with a rather high representation of cutmarks are ulna and femur, with 52 % each. The cutmarks are mostly found at the proximal andlor distal ends of bones, which indicate them deriving from dismemberment and/or from fieting (see chapter 1.3.2.). The distribution of cutmarks can be found in figure 5.6.a-j.
Figure 5.6. Cutmarks on wild cat bones from Hjerk Nor: a) cranium, b) mandible,
C) scapula, d) humerus, e) radius, f) ulna.

Figure 5.6. Cutrnarks on wiM cat bones from Hjerk Nor: g) pelvis, h) femur, ij tibia, j ) fibula.
6. CUTMARKS ON THE BONES FROM THE COMPARATIVE SITES
6.1. Hazendonk
6.1.1. Fox
A tibia of a juvenile fox showed cutmarks on the distal end (see figure 6.1.a). Zeiler interprets this
as indications of filleting (Zeiler 1987:260-261).
6.1.2. Badger
A femur of an adult badger showed cutmarks on the proximal anterior part of the shaft (see figure
6.1.b). This femur had been sawn in two at midshaft. The cutmarks are interpreted as probable f111eting marks (Zeiler 1987:261).

6.1.3. Pine marten
Of the four pine marten bones excavated at Hazendonk,
cutmarks were only found on a mandible. The placing
of the cutmarks on the basal part of the corpus mandibulae indicates the place where the fur was cut
loose at skinning (see figure 6.l.c) (Zeiler 1987:260).
6.1.4. Polecat
Two polecat mandibles showed cutmarks on the basal
part of the corpus mandibulae (see figure 6. I .d).
These cuanarks likely originated from the skinning
process (Zeiler 1987:260).
6.1.5. Otter
Cutmarks were only found on a small number of
the otter bones excavated at Hazendonk. The skeletal
elements that display cutmarks are mandible,
humerus and tibia (see figure 6. Leg). The mandible
cutmarks are interpreted as a result of the skinning
process, whereas the humerus cutmarks are interpreted
as filleting cutmarks. The cutmarks on the tibiae are
not interpreted in the text, but as Zeiler interprets the
skinning method to be the same as the Tybrind Vig
one (see chapter 1.3.2.) (Zeiler 1987:254), it can be
assumed that these cutmarks also originate from
filleting. Ca 22 % of a l l otter bones were burnt, and traces
of burning were found on almost every type of skeletal
element. Zeiler concludes that the otters at Hazendonk
were hunted for both their fur and meat (Zeiler 1987:254).
Figure 6.1. Cutmarks on bones from
Hazendonk: a) tibia of fox, b) femur of
badger, c) mandible of pine marten,
d) mandible of polecat, e) mandible of
otter, f) humerus of otter, g) tibia of
otter (Zeiler 1987:254,26 1).

6.2. Kongemose
62.1. Pine marten
Cutmarks were found distally on two tibiae
(Noe-Nygaard 1995:139). No further interpretation
is made.
6.22. Otter Figure 6.2. Cutmarks on pelvis of otter
from Kongemose (Noe-Nygaard 1995:225).
Cutmarks were only found on pelvis (see figure 6.2.a). These cutmarks are interpreted as deriving
from dismembering and from butchering (Noe-Nygaard 1995:137,224-225).
6.23. Wid rat
Of the eight bones of wild cat found at Kongemose, five limb bones - one radius, two femora and
two tibiae - showed cutmarks. The placing of thecutmarks are, however, not mentioned by N o e
Nygaard, nor are they interpreted (Noe-Nygaard 1995:140).
6.3. Muldbjerg I
6.3.1. Pine marten
Two calcanei of pine marten were found at Muldbjerg I. Noe-Nygaard states "Cut marks are fouiuf." (NoeNygaard 1995:139), but does not enter into any detail whether only one or both
calcanei showed cutmarks, or the placing of the cutmarks.
6.3.2. Otter
Of the total 115 fragments of otter, ca 25 showed cutmarks. Figures of the placement of cutmarks
are shown for six skeletal elements: cranium, mandible, scapula, radius, phalanx, and rib (see
figure 6.3.a-f). Cuhnarks were also found on one tibia and an unknown number of vertebrae, but
their placing were not described. The cutmarks on the cranium and mandible are believed to
originate from the skinning process, whereas the cutmarks on the ribs occurred when the ribs were twisted loose from the vertebrae. The cutmarks on the pelvis may derive from skinning, but
26

Figure 6.3. Cutmarks on otter bones from Muldbjerg 1: a) cranium, b) mandible, c) scapula, d) radius, e) phalanx, f ) rib
(Noe-Nygaard 1995:224).
Figure 6.4. Cotmarks on otter bones from Prastelyng: a ) mandible, b) scapula, c) humem, d) ulna, e) pelvis, f ) tibia
(Noe-Nygaard 1995:225).
may also derive from primary butchering. The distally/dorsally placing of the cutmark on the phalanx is the result of the skinning method. None of the other 17 phalanges showed cutmarks (cf chapter 7.3.). The cutmarks on the scapula are likely to have been caused by filleting (Noe- Nygaard 1995:135-136,223-224).

6.4.1. Pine marten
Of the sixteen fragments found, cutmarks are only found on two mandibles and on one rib. The cutmarks on the mandibles are placed "on the keel of corpus mandibularis" (Noe-Nygaard
1995:226), and are interpreted as deriving from the skinning process. The cutmark on the rib is categorized as a scrape mark (see chapter 1.4. for definition) (NoeNygaard 1995:139,226).
6.4.2. Otter
Eight otter bones from Prrestelyng showed cutmarks. The placing of cutmarks on mandible, scapula, humerus, ulna, pelvis and tibia can be found in figure 6.4.a-f. Noe-Nygaard writes: "... both skull and mandible have typical cut m a r k from skinning, and the rest of the postcranial skeleton show clear cut marks from dismembering." (Noe-Nygaard 1995:224). Also, some of the cutmarks on the scapula are interpreted as deriving from filleting, and the cutmarks on the pelvis as deriving from butchering (Noe-Nygaard 1995:136-137,223-224).
6.5. Swifterbant
6.5.1. Otter
The cutmarks on the otter bones at Swifterbant were found on mandible, humerus, ulna and femur (see figure 6.5.a-d). Those on the basal and lateral part of the corpus mandibulae are likely to have been traces of the skinning process. Zeiler is of the opinion that the cutmark on the oral
part of the ramus mandibulae may be due to dismemberment, but as such cutmarks are missing from Trolle-Lassen's experimental work as well as from the Tybrind Vig material, Zeiler refrains from making a definite conclusion regarding the origin of this cutmark. All cutmarks on the humeri are interpreted as having been caused by filleting. The cutmarks on i~tcbura trochlearis on
the ulna, are marks of dismembering (Zeiler 1987:254). Zeiler does not interpret the cutmarks on the femora, or those on the distal part of the ulnae.
Almost 15 % of the otter bones showed traces of burning. This, as well as the cutrnarks, indicates that otters were being eaten at Swifterbant (Zeiler 1987:254).

Figure 6.5. Cutmarks on otter bones from Swifterbant a) mandible, b) humerus, c) ulna, d) femur (Zeiler 1987:253).
6.6. Tybrind Vig
6.6.1. Fox
Cutrnarks were found on the IateraUbasal side of a mandible (see figure 6.6.a). They are believed to derive from skinning (Trolle-Lassen 1985:120).
6.6.2. Pine marten
Cutmarks were found on cranium, mandible and pelvis. These cutmarks derive from the skiining
process (see figure 6.6.b-d). There were also frequent breakages on the pubic bone parallell to the
cutmarks on the pelvis. On several pelves, there were breakages, but no cutmarks. TrolleLassen
interprets these breakages as deriving from the same process as the cutmarks, i.e. the animals
being cut open along the ventral midline when skinning. Often, the zygomatic arches on the
cranium were also broken by the pressure used when skinning. The cutmarks and breakages
indicate that the pine martens were skinned using open skiining, and that their paws were not
included in the fur (Trolle-Lassen 1987:87).
6.63. Polecat
A single pelvis showed cutmarks deriving from skinning (see figure 6.6.e). Another pelvis
displayed breakage of the pubic bone (TrolJeLassen 1987:88).

6.6.4. Otter
Cutmarks deriving from skinning were found on cranium, mandible, pelvis, metapodials and
phalanges (see figure 6.6.f-i). Cutmarks deriving from butchering were found on cranium, atlas,
scapula, humerus, radius, ulna, pelvis, femur and tibia (see figure 6.6.i-0). The s k i g cutmarks
indicate that the otters were skinned usmg open skinning, and that their paws were left in the fur. All limbs were dismembered and filleted. It is, however, not possible to determine whether the
dismembering occured before the fieting or after (TrolleLassen 1987:88).
6.6.5. Wid cat
Cutmarks deriving from skinnuig were found on cranium, mandible, metapodials and phalanges
(see figure 6.6.q-s). Cutmarks deriving from butchering were found on humerus, radius and
femur (see figure 6.6.t-v). The skinning procedure used for wild cat was the same as for otter.
The meat was likely filleted off the limbs after any cooking had taken place. Much fewer bones of
wild car than of otter showed cutmarks. TrolleLassen argues that this may be due to differences
in muscular attachments, or to the somewhat larger proportion of eroded wild cat bones (Trolle-
Lassen 1985:77-79; Trolle-Lassen 1987:87).
Figure 6.6. Cutmarks on bones from Tybrind Vig: a) mandible of fox, b) cranium of pine marten, c) mandible of pine
marten, d) pelvis of pine marten. e) pelvis of polecat (Tmlle-Lassen 1987:95-98).

Figure 6.6. Cutmarks on bones from Tybrind Vig: f) cranium of otter, g) mandible of otter, h) phalanges of otter, i)
pelvis of otter, j) scapula of otter, k) humerus of otter, I) radius of otter, m) ulna of otter, n) femur of otter, o)
tibia of otter, p) mandible of wild cat, q) metapodiak of wild cat, r) phalanges of wild cat, s) humerus of wild cat, t )
radius of wild cat, u) femur of wild cat (Trdle-Lassen 1987:98-102).

7. THE SKINNING AND BUTCHERING EXPERIMENT
The aim of the experiment was to determine placement and distribution of cutmarks deriving
from dismemberment and from fdeting. Previous studies have been carried out on stone marten
and otter (Trolle-Lassen 1985) (see chapter 1.3.2.), but as different species have to an extent
different muscular and skeletal anatomy, cutmarks on bones of different species may vary in
placement and distribution (Trolle-Lassen 1985:78; Trolle-Lassen pers. comm., in Zeiler
1987:247).
7.1. Method
A dead feral cat was acquired from the Technical bureau in Lund, Sweden. It had been shot by
hunters on commission by the city council, and was afterwards put in a freezer, waiting to be
destroyed. On receiving, the cat was put in a fridge to thaw. Blade knives of flint were obtained
from Jenny Eliasson, a local flintknapper.
The cat, an approximately one year old female, was skinned using'case skining (see figure 7.2.), with the exception that the forelimbs were also cut open longitudinally. By using case
skining, any cutmarks relating to skinning would not be found on the pelvis, md thereby not
able to be confused with cutmarks on the pelvis deriving from dismemberment or filleting. Due
to the time limit, the cat was only skinned up to the neck, and the head and torso - with the
exception of the pelvis - were not included in the skeletation process. The left front leg was filleted
and disarticulated at the scapula-humerus joint, humerus-radius-ulna joint, and the carpal joint.
The right hind leg was filleted and disarticulated at the femur-pelvis joint, the femur-tibia joint,
and the tarsal joint. The right front leg and the left hind leg were only filleted. The hind legs were
not filleted very carefully, i.e. close to the bone. The front legs, on the other hand, were cut free of
most flesh during filleting.
Figure 7.1. Open skinning (Andersson Figure 7.2. Case skinning (Andersson & Paulsson 1993:47).
& Paulsson 1993:47).

7.2. Result
Cutmarks were found on all larger bones, except for one scapula, one pelvis and one fibula.
Among the smaller bones, i.e. patellae, calcanei, astragali, carpal and tarsal bones, metapodials
and phalanges, cumarks were only found on one calcaneus, one astragalus, three metacarpals and
seven metatarsals (see figure 7.3.a-w).
Cutmarks resulting from skinning were found on three metacarpals and seven metatarsals.
Cutmarks and breakage of the pubic bone, as TrolleLassen has reported, did not occur, possibly
due to the skinning method being case skinning and not open skinning, or due to not enough
pressure being applied on the pelvis during skinning.
Despite both scapulae being filleted, cutmarks
occured only on the right one. As the right
scapula-humerus joint was not disarticulated, al l cutmarks on the scapula derived from filleting.
The right humerus show cutmarks posteriorly just
below caput humeri. These are likely to have derived
from the filleting of the lateral/posterior humerus muscles,
when the inner muscle layers were cut off at the proximal
end. The outer layers were still connected to the scapula,
and were cut off at the proximal end of the scapula. The
left humerus show some similar cutmarks, as well as
some others on tuberculum minus and crista tuberculi rninoris. These must have derived from the cutting off
of the laterdposterior humerus muscles, as cutting posteriorly towards the anterior side did not lead to any
cutmarks occuring during the disarticulation of the
scapula-humerus joint. There are also several cutmarks
on the distal end of the left humerus. The placement of
cutmarks on the medial ridge that surrounds foramen supracondylare, which is very frequent in the Hjerk Nor
material, do no occur here. Instead, the cutmarks are
found on either side of this ridge, as well as on the
anterior and posterior sides of the distal humerus. The
most proximal of these, and possibly afew medially placed ones, may derive from filleting, when cutting
off the muscle distally and fdeting 'upwards', whereas
it is certain that the others derive from the
dismemberment process. The right ulna show several cutmarks deriving
Fig. 7.3.a) Right scapula Lateral and medial view
!I Fig. 7.3.b.) Right humems Posterior view
Fig. 7.3.c) Left humerus Lateral, medial, anteriol; and posterior view
Figure 7.3. a-c. Cutmarks on bones of
experimentalb butchered cat.

from filleting on the lateral proximal part of the shaft.
Such cutmarks were also found on the left ulna, but a
slight bit further down the shaft, The left ulna has
some cutmarks mediallylposteriorly on olecrarzon
and on the mediaVposterior distal end. The proximal
ones derive from the disarticulation of the
humerus-radius-ulna joint, and the distal ones derive
either from the disarticulation of the carpal joint or
from the initial cut when filleting.
The right radius show no cutmarks, whereas
the left one show several cutmarks laterally and
medially on the distal end. These cutmarks correspond
with the disarticulation of the carpal joint, as the many
tendons placed anteriorly made the dismembering
somewhat difficult, and forced a great deal of sawing
with the blade.
Of the ten metacarpals, only three showed cutmarks. These are the fourth and fifth right
metacarpals, and the third left metacarpal. All cutmarks
are found on the proximal part of the shaft. The
cutmark on the fourth right metacarpal is placed
posteriorly, whereas the cutmarks on the fifth right
metacarpal are placed anteriorlynaterally. The cutmarks
on the third left metacarpal are placed anteriorly. All
cutmarks derive from the skinning process.
The right pelvis show several cutmarks around
the acetabulum, all deriving from the disarticulation
of the femur. This was a rather difficult process, as the
hip socket is deep, and the thick muscle layer made the
actual placing of caput fernoris somewhat uncertain.
Also, there is one cutmark on the lateral part of the ischiurn. This is likely to derive from the skinning
process, as no filleting took place on the ischium. The
left pelvis, on the other hand, show no cutmarks,
despite fdeting on the ilium.
The right femur has one cutmark placed
medially/posteriorly on the mid part of the shaft. This
cutmark is likely to derive from filleting. There are
also several cutmarks on the proximal end of the right
Fig. 7.3.d) Right ulna Medial, anterior, and lateral view
Fig. 7.3.e) Left ulna Lateral, posterior, and medial view
Fig. 7.3.0 Left radius Lateral, and medial view
Fig. 7.3.g) Right metacarpal 4 Medial, and posterior view
Fig. 7.3.h) Right metacarpal 5 Lateral, and anterior view
i! Fig. 7 . 3 4 Left metacarpal 3 Anterior view
Figure 7.3. d-i. Cutmarks on bones of
experimentally butchered cat.

femur. The ones below caput femork, the ones on the
dorsal side of coIlum femoris, and the ones placed
laterally on trochanter major, are believed to derive
from the disarticulation of the femur from the
acetabulum. The cutmarks that are found
anteriorlylmedially just below trochanrer minor, are
assumed to derive from fdleting. The left femur show
two cutmarks placed medially on the lower part of the
shaft. These cutmarks are likely to derive from filleting. The right tibia has two cutmarks placed medially
on the proximal end. These cutmarks must have occured
when the posterior tibia muscle was cut off from the
bone after filleting. Otherwise no cutmarks were found on the right tibia. The left tibia did not display any
cutmarks at all.
A similar distribution is found on the fibulae. No cutmarks were found on the left fibula, whereas the
right one show two cutmarks placed posteriorlyflaterally
on the proximal end. The lateral tibia muscle was not
cut off towards the bone, but outwards, which should
not have caused any cutmarks. The flint knife may have
touched the fibula during that cut, without me noticing,
but this is ratherunlikely. The sound of flint cutting into bone is very distinctive. The cutmarks on the fibula may
have derived from the same cut that caused the cutmarks
on the tibia, although this seems to be rather unlikely.
Of the bones in the tarsal joints, cutmarks are only
found on the right calcaneus and astragalus. The cutmarks
on the calcaneus were found anteriorly on &her calcanei,
and the ones on astragalus were found anteriorly on collum tali. These cutmarks all derive from the
dismemberment of the tarsal joint. Seven of the eight metatarsals showed cutmarks on
the mid- and lower part of shaft. All four right metatarsals
display several cutmarks anteriorly. The second right
metatarsal also has cutmarks on the medial side, and the
fi th has cutmarks on the lateral side. The left metatarsals
display fewer cutmarks on each bone. The second
metatarsal has one cutmark placed medially, and the third
Fig. 7.3.j) Right pelvis Lateral view
Fig. 7.3.k) Right femur Anterior, posterior, medial, and lateral view
Fig. 7.3.1) Left femur Medial view
Fig. 7.3.m) Right tibia Posterior, and medial view
Figure 7.3. j-m. Cutmarks on bones of
experimentally butchered cat.

metatarsal has one placed anteriorly. The f ~ t h
metatarsal has some cutmarks placed posteriorly,
laterally and anteriorly.
7.3. Discussion
The result of the skinning procedure show great
similarity wlth Trolle-Lassen's skinning and
butchering experiments (see chapter 1.3.2.). In all
three experiments. many cutmarks were found
on the phalanges and metapodials, respectively. As these bones are not covered in musculature, the flint knives w~l l come into contact with the
bone. The strong tendons there can make some sawing necessary, resulting in multiple cumarks
on each bone. As the cat's skull was not included
in the skinning, no comparison can be made
regarding cutmarks on the cranium and mandible.
There were also some similarities in the
results of the butchering procedure. In both
Trolle-Lassen's butchering of otter, and my
butchering of cat, the disarticulation of the
femnr-pelvis joint caused cutmarks on caput
femoris, collunt fe~noris, troclzar~ter ntajor and
along the lateral side of acetabulum. The same placing of cutmarks on otter and cat was found at
the humerus-radius-ulna joint, where cutmarks
occured on the medial and lateral ridges of the
distal humerus, anteriorly on the distal humerus,
and around incisura trochlearis on the ulna. The
only difference in the placing of cutmarks at this
joint, was the lack of cutmarks below the proximal
articulation surface of the cat radins. The filleting
of scapula caused cutmarks parallell to spina
scapulae on the lateral side. The direction of the
filleting cutmarks on the medial side are different
on otter and cat, and may be due to the butcherer
fiieting distally-proximally or anteriorly-posteriorly .
Fig. 7.3.0) Right calcaneus Anterior view
$$ Fig. 7:3.p),Right stragalus Antenor vlew
Fig. 7.3.q) Right metatarsal 2 Anterior, and medial view
Fig. 7 . 3 4 Right metatarsal 3 Anterior view
Fig. 7.3.11) Left metatarsal2 Medial view
Fig. 7.3.v) Left metatarsal 3 Anterior view
F i Fig. 7.3.w) Left meratarsal5 Posterior, anterior, and lateral view
Fgure 7.3. n-w. Cutmarks on bones of
experimentally butchered cat.

Otherwise, few filleting traces were found on the otter. Otters have very strong muscle
attachments, and the filleting was claimed to be very difficult. The limbs were not filleted thoroughly, and I believe this may be the reason for the lack of cutmarks (Trolle-Lassen 1985:Appendix VI; TrolleLassen 1985:Appendix VII).
Regarding the cat metatarsals, there was a discrepancy in the number of cutmarks on the left
and right side. This is likely due to inexperience, where the side fust disarticulatedlf111eted received a rougher treatment, and subsequently more cutmarks. As a comparison, it can be mentioned that only one of 17 phalanges of otter at Muldbjerg I displayed cutmarks - clearly a work of experienced skinners (see chapter 6.3.2.).
It is worth noting that filleting does not automatically lead to cutmarks. On several bones, no traces of filleting could be seen. This is especially true for the front legs, as they were filleted
much more thoroughly than the hind legs. It seems that the periosteum is rather resistent to Kit . This is important, as the sound of flint scraping across the periosteum is no different from flint actually cutting into bone. On several occasions I believed that my actions would have caused cutmarks, simply because I heard the flint knife coming into contact with bone. Instead, the flint only touched the periosteum, and no cutmarks occured on the bone.
It seemed easier to- fillet bones if the limb was fust dismembered. This may be due to the smaller size making the separate parts easier to handle.
8. DISCUSSION
8.1. The individual species
8.1.1. Fox
One of the two mandibles displayed a cutmark on the basal part of corpus mandibulae. The
placing of cutrnarks on the basal part of corpus mandibulae is found in almost every other comparative site, and on most species. It is confirmed through ethnographical studies and through
experimental work, that such cutmarks derive from skinning, when the skin is cut off at the snout area.
The cutmark on the rarnus mandibulae is somewhat more difficult to interpret. Similar cutmarks have to my knowledge only been found on e k mandibles from Bokeberg 111, red deer mandibles from Lolliiuse, a dog mandible from Smakkerup Huse, an otter mandible from Swifterbant (see figure 8.1.a-d), and on one of the cat mandibles from Hjerk Nor (Magnell
1996:12, 21; Friborg 1999:82; Hede 199996, Zeiler 1987:253-254). The interpretation of these cutmarks varies. Ola Magnell, Malene Nina asterberg Friborg and 3. T. Zeiler interpret them as butchering cutmarks, whereas Signe Ulfeldt Hede believes them to derive from skinning (ibid). Only Friborg enter into any discussion as of why she makes these interpretations. She refers to

Figure 8.1. a) Elk mandible from B6keberg 111, b) red deer mandible from Lollikhuse, c) dog mandible from Smakkerup
Huse, d) otter mandible from Swifterbant. Note cutmarks on the lateral side of ramus mandibulae (Magnell 1996:lZ;
Friborg 7 999:82; He& 1999:96; Zeiler 7 987:253-254).
Binford's (1981) accounts of disarticulation of cranium and mandible on caribous (Friborg
1999232). However, Binford's descriptions of
the placings of cutmarks caused by the
disarticulation of the mandible do not include
such cutmarks as those found on the fox
mandible at Hjerk Nor (Binford 1981 :109).
Cutmarks caused by disarticulation of the
mandible in a frozen animal or one afflicted
by rigor mortis, as Binford refers to, may
cause cutmarks on the ranius mandibulae, but these cutmarks will likely only mn on the Figure 8.2. Possible way of skinning at the jaw
lateral side, due to the stiffness, and not on the (Anderson & Paukson 1993:63).
oral side, as was found on the fox mandibles
at Hjerk Nor (cf Binford 1981:109). I find it hard to believe Hede's hypothesis of the cutmarks
deriving from skinning, as the skin usually is pulled away from the carcass in the cranial
direction. To loose the skin at the lips of the animal, one can simply let the knife mn in the cranial
direction without making any cuts on the oral side of the ramus mandibulae (see figure 8.2.). I
am therefore more likely to interpret the cutmarks as deriving from disarticulation of the
mandible. The cutmarks on the tibia clearly derive from filleting. There are several parallell cutmarks,
which are concordant with both Binford's description of cutmarks deriving from filleting 38

(Binford 1981:129), and the cutmarks found on the left ulna after my experimental butchering
(see chapter 7.2.). Due to the muscular anatomy of the tibia, it is likely that the initial cut occured
distally and that the flesh was faeted upwards towards the femur-tibii joint. Of the six comparative sites, fox remains were only found at Hazendonk. There, cutmarks
were found on distally on a tibia shaft. As with the ones of Hjerk Nor, they were interpreted as
deriving from filleting. This should not be interpreted as foxes regularly being used as meat
animals on Mesolithic sites. Bones of foxes are fairly common on Mesolithic sites in Southern
Scandinavia, but the lack of cutmark analyses makes any hypothesis regarding foxes as a
common meat source doubtful.
The tame fox, that was briefly discussed in chapter 4.1., adds an interesting aspect to the
utilization of fox at Hjerk Nor. Unfortunately no sexual differential measurements could be taken,
and thus no sex estimation could be made. No sexual estimation has been madeon the two other
known cases of tame foxes at Mesolithic sites in Southern Scandinavia (see chapter 4.1.)
(Degerb011933:433-438; Jonsson 1988:67). It remains to be seen if these tame foxes were of the
same sex, and if it in that case may indicate a concious choice of domestication. An example of
this would be tame females that would mate with wild foxes and thus be used as 'fur farms'. The
use of a fox as a decoy or as a pet, would, on the other hand, not necessarily be dependant of the
animal's sex.
Of the fox remains on the Hjerk Nor assemblage, it can be concluded that foxes wereused
both for their furs and for their meat. It is not known whether the preferred skinning method for
foxes was case skinning or open skinning (see figure 7.1. and 7.2.). The cutmark on the corpus ma~zdibulae indicate that the skin was cut off at the snout, possibly in a circling motion. Due to the
small sample and the few cutmarks, it is not known whether fox meat was a regular component
in the diet, or if it was used as emergency food. In the case of the older tame fox, its part in the
diet may have been viewed as the last use of an animal that would soon die anyway.
8.1.2. Badger
Only one badger bone from the Hjerk Nor assemblage displays a cutmark. As it is placed on the distal part of the tibia shaft it can be deriving from either skinning or filleting. As animals from
virtually all Mesolithic sites in Europe have displayed cutmarks deriving from skinning on the
metapodials or phalanges, i.e. all skin on the legs were used, it would suggest that the same
skinning method was also used at Hjerk Nor. The cutmark may have derived from filleting, when
an initial cut was made on the distal tibia, in order to fillet 'upwards'. Disarticulation is not that
likely, as the cutmark seems to be placed too much proximally on the shaft. The lack of cutmarks
on the other badger bones from Hjerk Nor seems to indicate that the badgers generally were not
butchered. The lack of cutmarks on the mandible can be explained by the skinner not cutting
towards the mandible when finishing the skinning, but instead cutting forwards/outwards.

Another possibilty is that the skin on the jaw were left on the carcass.
Of the comparative sites, bones of badger are only found at Hazendonk. There, cutmarks,
interpreted as deriving from filleting, were found on the upper part of a femur shaft, which also
had been sawn in two (see chapter 6.1.2.). Bones of badgers are fairly common on Mesolithic
sites from Southern Scandinavia. As very few cutmark analyses have been made, it is not
possible to discem whether the filleting of badgers was a common occurrence or if the animals
were mostly only skinned.
Another reason for the utilization of badgers is their fat. During autumn, a badger will on
average increase its weight with 30-50 %. This is mostly stored as fat (Mathiasson & Dalhov
1987:125). The fat could be used as grease for items of leather, such as shoes, clothes and tents.
Also, it could be used in food, and as lamp oil. The use of oil for ceramic lamps have been
verif~ed for the Erteb0lle period, but the oil is mostly assumed to be blubber from sea mammals
(Andersen 199553). Such hunting of mammals with the main reason of utilizating their fat is
known from North America, where the indians in the Shield and Mackenzie borderlands hunted
bear mostly for its fat (Rogers & Smith 1981:132).
According to the placing of the cutmarks - and lack of them - the badgers at Hjerk Nor were
hunted mainly for their fur, and possibly also for their fat. Badger meat seems to have been a rare
occurence. The use of badger meat is confirmed at Hazendonk. On the other comparative sites, no
badger bones were found.
8.1.3. Pine marten
The cutmarks on pine marten bones from Hjerk Nor can be divided into two categories: skinning
related and dismemberment/flleting related. In the first category the two crania and the mandible
are found. The placing of cutmarks on the temporal arch and on the snout or osfrontale, as well
as on the basal part of corpus mandibulae are almost ubiquituous on skulls from skinned
carnivores and other smaller animals from Prehistoric assemblages (cf Desse 1975:206; von den
Driesch & Boessneck 1975:TafelS; Rowley-Conwy 1998:95-96; TrolleLassen 1987:95-101).
The cutmarks are either placed on the snout or on osffontale depending on where the skinner wants the fur to end. This variation is likely to be random or, possibly, due to different skinning
traditions at different sites. The cutmarks that are related to dismemberment andlor filleting are
found on the upper part of the ridge that forms foramen supracondylare on the humerus and
proximally/medially on the proximal part of the tibia shaft. The humerus cutmark may just as
likely be due to dismemberment as f~lleting, whereas the tibia cutmarks derive from fdeting.
The cutmark evidence from the Dutch site Hazendonk, suggests that the pine martens were
skinned, but not butchered. However, any evidence of the pine martens at Hazendonk not being
butchered is vague at best. Only four bones were found: one mandible, two distal par& of a scapula, and one pelvis fragment. Cutmarks were only found on the mandible. The remaining

bones are not necessarily any skeletal parts with a high frequency of cutmarks deriving from
dismemberment and/or filleting. Due to the small size of the pine marten, any filleting is likelier to have taken place on the humerus, the femur and the tibii, than on the scapula and the pelvis.
The cutmarks placed distally on the Kongemose pine marten tibiae may, depending on their placing, have derived from either skinning, dismemberment or filleting. If skinning, this would
indicate a different skinning method than the one confirmed at Tybrind Vig (see chapter 1.3.2.). The skin would instead have been cut off at the tarsal joint, thereby completely disregarding the
skin at the end of the legs. If the cutmarks derive from dismemberment, it may have been a variation of the above mentioned skinning method, where the tarsal joint was cut off, in order to
keep the entire foot in the prepared skin, maybe as a decoration. Any dismemberment traces may
also have been due to the cutting off of the foot, in order to use the foot for decorative or ritual purposes. This latter hypothesis is compatible with the hypothesis regarding the skinning method
mentioned above. The last explanation regards filleting. Cutmarks that have been interpreted as deriving from filleting have been found on a distal fox tibia from Hazendonk (see chapter 6.1.1.)
and on tibiae of fox, badger and wild cat from Hjerk Nor (see chapter 8.1.1 ., 8.1.2. and 8.1.7.). The pine marten tibia may thus have been filleted, but due to information from osteological and
ethnographical studies (see below), it is not the most conceivable interpretation. NoeNygaard states that "the bones [two crania and two tibiae] are well preserved, apart
from lzuman breakage." (NoeNygaard 1995139, with addition by author). She does not explain what she means by 'human breakage', or on which bones it occured on. As nothing is further discussed, I am of the opinion that it may have been broken zygomatic arches. This is a common
phenomenon in archaeological assemblages, and have been interpreted by Trolle-Lassen as an indicator of skinning (see chapter 6.6.2.). One other possible explanation could be heavily fragmented braincases. Such were found on some otter skulls from Muldbjerg I, and were discussed at some length regarding the use of traps, and possible brain consuming (Noe-Nygaard 1995:223). The lack of a similar discussion makes me reject this form of breakage on the Kongemose pine martens. Yet another possibility is the tibiae being fragmented for marrow removal, but as above, I feel that is this were the case, it would have been discussed further, as pine martens are normally not regarded as 'food animals'.
At Muldbjerg I, two calcanei were the only pine marten bones found. Both calcanei showed cutmarks. It may indicate a similar skinning method as the one suggested for Kongemose (see
above), or disarticulation of the tarsal joint (see chapter 7.2.), or the use of pine marten feet for decoration or ritual purposes.
The Przstelyng pine martens show two types of marks: cutmarks deriving from skinning, and a scrapemark. Despite a fair representation of the entire skeleton, cutmarks are only found on
the corpus mandibulae on two mandibles. No cutmarks deriving from dismemberment or f111eting were found. The single scrapemark on the rib is therefore unexpected. Any regular eating
of pine marten would assumably have left traces in the skeleton to a rather high extent. The scrapemark could have happened accidentaly, as of a child playing at butchering, or the rib being

the single remain of a pine marten being used as emergency food.
At Tybrind Vig, several facts indicate that pine martens were not eaten, and k i d only for
their fur. Firstly, almost all pine marten bones were found in clusters, signifying complete
individuals being deposited. Secondly, all cutmarks were found on os frontale, on corpus
mandibulae and the pubic bone, which is indicative of open case skinning (Trolle-Lassen
1987236-87).
According to Noe-Nygaard, the pine martens found at Dyrholrnen may have been treated
differently than pine martens at other Danish sites: " ... Degerb)I (1942) describedpirze marten
bones from the Erteb9lle site Dyrholmen that bore numerous cut marks, and mandibles with fragmentation of the corpus mandibularis". She is of the opinion that this "may indicate tlzatpine
martens were sometimes eaten by man, or the strong sinews in canalis mandibularis have been of
some use e.g. as thread." (Noe-Nygaard 1995:226). This reasoning may, however, be a modified
truth. A subsequent inspection of her source, revealed only a picture of a marrow fractured pine
marten mandible. Nothing in the text indicated any cutmarks, burning, or other breakages of pine
marten bones (Degerbal 1942234). It is possible that the information on cutmarks and frequent
mandible fragmentation were given to Noe-Nygaard orally, and that such reference was later
omitted by mistake. Also, canalis mandibulae do not contain sinew, but a fatty tissue rich in
nerves and bloodvessels (Iregren pers. comm.). It may have been the fat that the Dyrholmen
inhabitants coveted.
In concluding, it seems that pine martens were mostly seen solely as a fur animal. Signs of
flesh utilization are extremely rare, also in later Prehistoric periods (see Clason & Brinkhuizen
1993:65; Desse 1975:206; Ericson et al 1988:85). This is concordant with ethnographical studies,
where pine martens, just as many other smaller carnivores, are rarely claimed to being eaten,
except as emergency food (Eidlitz 1969:18-19, 130; Helm & Sturtevant 1981). Signs of the
eating of pine marten are found in the Hjerk Nor assemblage, where one humerus was
disarticulated at the humerus-radius-ulna joint, and one tibia was filleted. As with fox (see chapter
8.1.1.), the only certainty regarding skinning method, is the cutmarks on the cranium and the
mandible, where the skin was cut off at the snout. It is not known if the pine martens were
skinned using case skinning or open skinning.
8.1.4. Polecat
The lack of cutmarks on the polecat cranium from Hjerk Nor is rather peculiar. On almost all
other crania of wild carnivores found at the comparison sites, cutmarks occur either on osfronmle
or on the snout (Noe-Nygaard 1995:223; Trolle-Lassen 1985:77; TrolleLassen 1987:87-88; see
also chapter 8.1.1.). However, the three crania of polecats from Tybrind Vig do not display
cutmarks. This may be due to only the 'wrong' fragments of the crania being present in the
assemblage. However, it may also signify that the polecat crania were either included in the

prepared skin, or that the skin from the skull were left on the carcass after skinning. It is possible that similar skinning methods were used for polecats at Hjerk Nor. Another possibility is that the polecat cranium were part of a decoration, with or without its skin (see chapter 4.4.).
At the comparative sites, the number of polecat bones are generally few. Polecats have only
been found at three sites: Hazendonk, Swifterbant and Tybrind Vig. At Hazendonk, two mandibles and a calcaneus were found, and at Swifterbant, two mandible fragments. The exception is Tybrind Vig, where 41 bones were found. Most of the skeleton was represented, but still, only one bone displayed cutmarks. The cutmarks on the Hazendonk mandibles and on the
Tybrind Vig pelvis are all indicative of skinning. The lack of cutmarks on the Swifterbant mandibles may be due to the 'wrong' part being present, or to the skin of the jaw being
disregarded in the skinning process (TrolleLassen 1985:55-57; Zeiler 1987:260). The reasoning above, seems to indicate that the polecat was used for its skin at all four sites.
Any other use of polecat is unlikely, unless we are dealing with uses of some ritual andlor medical nature, or extreme starvation. Ethnographical sources are unanimous regarding polecat
meat, as well as meat from other small mustelids such as the ermine: the meat stink, and not even dogs will eat it (Eidlitz 1969:130; Wesenberg-Lund 1927:142, in TroUe-Lassen 1985:58).
8.1.5. Otter
The cutmarks on otter bones from Hjerk Nor indicate both skinning, dismemberment and filleting. The cutmarks deriving from skinning are found on the skull, i.e. on the snout, os frontale, the zygomatic arch and corpus mandibulae. One pelvis shows cutmarks that derives either from skinning or from dismemberment. Other cutrnarks deriving from dismemberment
are found on humerus, ulna, pelvis and femur. Humerus, ulna, pelvis and femur also display cutmarks deriving from filleting (see figure 5.4.a-h). The cutmarks deriving from skinning are similar to the ones found on fox, pine marten and wild cat, i.e. a circular cut around the snoutleye region. The anteriorly-posteriorly placed cutmarks on the ischium (see appendix 4, otter pelvis No. a) may be the f i s t indicators of whether the animals at Hjerk Nor were skinned using open
skinning or case skinning. The cutmarks are similar to the ones found on pine marten, polecat and otter at Tybrind Vig (Trolle-Lassen 1987:95-98), and occur when animals are open skinned (TrolleLassen 1987:87).
The cutmarks deriving from dismemberment show that the scapula-humerus joint, humerus-radius-ulna joint and femur-pelvis joint were frequently disarticulated. Only a few humeri and femora do not show any cutmarks indicative of such treatment. According to the number of cutmarks, it seems that the humerus-radius-ulna joint was more often disarticulated
than the scapula-humerus joint. Such train of thought would, however, be erroneous, as the humerus-radius-ulna joint is a very complex joint, and is hard to disarticulate, whereas the
scapula-humerus joint is relatively easy to disarticulate (cf chapter 7.2.).

Cutmarks indicative of filleting occur on the shafts of three humeri, one ulna and three femora. Three pelves may also have been filleted. Similar cutmarks on ischium have been found on otter bones at Kongemose and Praestelyng, where they have been interpreted as deriving from butchering (Noe-Nygaard 1995:225). Cutmarks placed on the ilium were found on beaver pelves at Hazendonk. They were, however, not interpreted by Zeiler (Zeiler 1987:258-260), and the conclusion of them deriving from filleting is therefore somewhat uncertain. A further discussion of the interpretation of cutmarks found on pelvis can be found in chapter 8.1.7.3. The cutmarks on the humeri varies in number from a few to many. The ulna and the femora, on the other hand, display few cutmarks. The otter is said to be difficult to fiiet, due to strong muscle attachments (Trolle-Lassen 1985:Appendix 7). The variation of the number of cutmarks on the humeri could be caused by butchers of different experience (cf chapter 7.3.), or by a variation in the need for meat. If meat was plentiful, there was no need to remove every scrap.
Otter bones with cutmarks are found on all of the comparative sites. Otters were treated roughly the same on all sites. Frequent cutmarks on corpus mandibulae and on the snout give evidence of skinning. At Tybrind Vig, cutmarks on the pubis indicate the use of open skinning. This skinning method is otherwise not confirmed for the other comparative sites. At Tybrind Vig, cutmarks on the phalanges, as well as absence of the claws, indicate the paws being left in the skin (Trolle-Lassen 1987:88). Dismemberment occured at the pelvis-femur joint and at the humerus- radius-ulna joint at virtually all sites. On Praestelyng, evidence was also found for the dismembering of the humerus-scapula joint and the femur-tibia joint. Cutmarks deriving from filleting occured at almost all limb bones. Regarding the amount of cutmarks on individual bones, a direct comparison to Hjerk Nor is somewhat difficult, as no individual descriptions of the placing of cutmarks can be found for the comparative assemblages. However, in the pichres of the placings of cutmarks, Swifterbant and Tybrind Vig stand out, in that there is a large amount of cutmarks spread all over the bones (Zeiler 1987:253; Trolle-Lassen 1987:99-100). At the other sites, only a few cutmarks are marked on each skeletal element (NoeNygaard 1995:224-226).
The treatment of otter at Hjerk Nor do no stand out when compared to the other sites. At all seven sites, the otter was used for both fur and meat. At both Hjerk Nor and Tybrind Vig, cutmarks and fragmentation patterns indicate that the otters were skinned using open skinning.
As it was possible to carry out an age estimation for the Hjerk Nor otters, an interesting comparison regarding the hunting of otter can be made. In the Hjerk Nor assemblage, there was a large over-representaion of older otters, i.e. over two years (see chapter 4.5.). This is similar to the otters at Hazendonk and Swifterbant, where Zeiler interprets the age estimation as a deliberate hunting of otters of two years of age or more (Zeiler 1987:247-252), and the otters at Tybrind Vig, who were over one year of age at death (TrolleLassen 1987:89).
The estimated sex ratio of otters at Hjerk Nor adds another dimension to the issue of otter hunting. Canadian and Danish studies concerning the sex ratio of trapped and caught otters (Hamilton & Eadie 1964; Jensen 1961; in Zeiler 1987:252), both show a seasonal difference in
sex ratio of otters. During October-February the sex ratio was 1:l in both countries, whereas

during March-September it was 3:2 and 5:2, respectively. Hamilton and Eadie were of the
opinion that the higher number of trappedlcaught males during spring reflected a difference in
activity during the mating and whelping season (ibid). It is very difficult - if not impossible - to
distinguish between male and female otters in the open field. The over-representation of males in
March-September can therefore not be due to sex selective hunting (Zeiler 1987:252). A true
over-representation of male otters in Hjerk Nor (see chapter 4.5.) would indicate otters being
mainly killed during spring and summer. The 'rule' of fur animals always being hunted during
late autumnlearly winter, when the fur is at its best, do not apply for otter, as their fur do not
change dramatically between seasons (Hahr 1882; Jensen 1972; Jensen 1984; in TrolleLassen
1987:89).
The above discussed implications of the age and sex distribution indicate that the otters at
Hjerk Nor were hunted selectively. The selective criterion is likely to be the size of the animal. If
the hunting took place in March-September, it would have mostly been males that were out and
about. Younger, smaller animals would have been neglected by the hunters. The use of traps,
instead of selective hunting, would have given a distribution of age and sex that was equal to the
living population. Traps may have been used, but only to a lesser extent. Comparisons with
ethnographical and historical sources reveal that possible methods for selective hunting of otter
were bow and arrow, spear, and harpoon (Strid 1999:9-19). Arrowheads and harpoons, which of
course would have been used for the hunting of several diiferent species, are common finds at
Mesolithic sites in the region. Spearheads are, on the other hand, rather rare.
8.1.6. Lynx
Cutmarks were found on four bones: a humerus, radius and ulna from one individual, and an ulna
from a second individual. The forelimb of the first individual displays cutmarks at the humerus-
radius-ulna joint, as well as cutmarks on the mid part of the radius shaft, and distally on the ulna.
The cutmarks at the humerus-radius-ulna joint are mostly found on the humerus, but a few are
found on the ulna's dorsal side. The cutmarks found on the humerus' anterior side may derive from filleting, but as such cutmarks were only found on the cat humerus that was disarticulated
during theexperimental butchering (see chapter 7.2.), it is likely thatall cutmarks that were found
at the lynx' humerus-radius-ulna joint derive from disarticulation. The cutmarks on the radius, on
the other hand, show concordance with Binford's description of cutmarks deriving from filleting
(Binford 1981:129). The origin of the cutmarks found distally on the ulna are somewhat unclear.
Similar cutmarks were found on the experimentally butchered cat, but it could not be affirmed
whether they were caused by fieting or by dismemberment (see chapter 7.2.). In the comparative
material, such cutmarks have been found on an otter ulna from Swifterbant (see chapter 6.5.1.).
Zeiler does not, however, interpret these cutmarks (Zeiler 1987:254).
The second ulna derived from a juvenile lynx, where both epiphyses were unfused. It is

therefore possible that more cutmarks occured, but that they either occured on the lost epiphyses or on the cartilage between epiphysis and diaphysis. The placing of the single cutmark found proximally show similarity to the cutmarks found on the left ulna of the experimentally butchered cat (see chapter 7.2., and figure 7.e.). As those cutmarks only occured on the ulna that was dismembered, they are likely to derive from the disarticulation of the humerus-radius-ulna joint.
Unfortunately, no lynx bones were found at the comparative sites. It is therefore difficult to
make a comparison of the utilization of lynx during the Mesolithic and Neolithic in Southern Scandinavia. Bones of lynx have been found at contemporary sites in the Limfjord region (see chapter 3.1.), which would be indicative of the hunting and/or trapping of lynx. However, as no cutmark analyses have been carried out, it is not possible do discern if the lynx was hunted for its meat as well as for its fur.
8.1.7. Wild cat
8.1.7.1. Skinning
Several skeletal elements of wild cat display cutmarks that are relevant for the interpretation of the skinning procedure. Two crania show cutmarks on maxilla, and one cranium shows cutmarks on osfrontale. This would indicate different skinning techniques regarding where the skinning was to be ended. This is further supported by the fact that the other crania in the assemblage - that do not display cutmarks - either lack inailla or osfrontale. As this variation in skinning technique is found on the same site, it may indicate different methods used by different skinners. The few centimetres between maxilla and osfrontale are not likely to have made any greater difference on
the fur, and this varation in skinning should therefore not be overinterpreted. The mandibles also show a variation in the cutmarks. Only one mandible show cutmarks
on corpus mandibulae. As the other 11 mandibles lack cutmarks there, it is possible that the skinners did not cut towards the mandible when cutting off the skin, but rather forward/outward.
Such a cut was tried on a front paw during the experimental skinning of the cat, and did not cause any cutmarks (see chapter 7.2.). Another possibility is that the skin of the jaw was not included in
the fur. As most other sites show frequent cutmarks on corpus mandibulae, I consider this hypothesis less likely than the first one.
One pelvis displays cutmarks on the pubis, which would indicate the use of open skinning. On almost all pelves the pubic bone is broken off. According to Trolle-Lassen, this is also an
indication of open skinning (Trolle-Lassen 1987:87). The lack of cutmarks on the metatarsals show that the skin of the paws was either cut off at
the phalanges, at the tarsal joint, or cut off outwardly, as was discussed regarding the mandibles. As the phalanges are missing, it is not possible to deduce anything regarding whether the claws
were included or excluded from the finished skin.

8.1.7.2. Disarticulation
Cutmarks believed to derive from disarticulation are found on mandible, scapula, humerus,
radius, ulna, pelvis, femur and tibia. The cutmarks on the mandible are similar to the ones found
on the fox mandible. As the interpretation of those cutmarks were discussed in some detail in
chapter 8.1. I., no further discussion will be made here.
The disarticulation-related cutmarks on the scapula are situated on the margin of cavitas
glenoidalis and on processus suprahamatus spinae. Corresponding cutmarks on humerus are
extremely rare, being only found on one humerus (No. 17a). They are situated on sulcus
intertubercularis and on the anterior part of tuberculum majus. The scarcity of cutmarks at the
scapula-humerus joint does not necessarily mean that this joint was not disarticulated. The
disarticulation of the scapula in my butchering experiment did not cause any cutmarks (see
chapter 7.2.). Similarily, Binford writes "As in the case of cut marks on the scapula, few marks on the proximal humerus ... can be related to initial butchering." (Binford 1981:121). However,
for a small mammal like the wild cat, the smaller joint may have been easier to disarticulate than a
caribou's and disarticulation may therefore not necessarily have caused cutmarks at all. Cutmarks
on scapula are rather rare in the comparative material, possibly due to the fragility of this bone.
In contrast to the scapula-hume~s joint, cutmarks at the humerus-radius-ulna joint are
numerous. They are found on almost all humeri and ulnae that display cutmarks. The cutmarks
on the humerus are usually found above, below and on the ridge that forms foramen
supracondylare, whereas the ones on the ulna are mostly found on the anterior side of olecranon, and on either side of incisura trochlearis. The cutmarks on the humerus and ulna of the
experimentally butchered cat (see chapter 7.2.) deviate somewhat from the ones in the Hjerk Nor
assemblage, as no cutmarks are found on the edge of incisura trochlearis. Also, there is a large
amount of cutmarks on the anterior side of the distal humerus, as well as around foramen
supracondylare. This may, however, be related to my inexperience in dismemberment. As some
of the humeri in the Hjerk Nor assemblage also display an unusual amount of cutmarks at this
joint, it may suggest the activity of less experienced butchers.
Cutmarks on the proximal radius that relates to dismembering are very rare. Two radii
display several cutmarks on the lateral side of the proximal articulation surface. Similar cutmarks
were not found on the experimentally butchered cat, but occur on a beaver radius at Muldbjerg I,
where the cutmarks have been interpreted as dismemberment cutmarks (NoeNygaard 1995:227).
The amount of cutmarks at the wild cat radii, as well as them being parallell, seem to indicate
them deriving from disarticulation. I find it unlikely that such repeated cutting would have been
necessary for filleting.
Two radii and two ulnae display cutmarks distally. The cutmarks placed laterally on the
distal radii derive from disarticulation of radius-carpals. Several tendons run here and it is virtually
impossible to cut them off without damaging the bone underneath. The cutmarks on the ulnae do
either derive from such disarticulation or from fdleting (see chapter 7.2.). In those cases where no

cutmarks have been found distally on radii, it is certain that either the cut was placed at the metacarpals or at the carpal bones, or that no disarticulation took place. In the Hjerk Nor case, only a few metatarsals have been found - all without cutmarks - and any tarsal or carpal bones in the kitchen midden were likely neglected by the collector. It is therefore not possible to discern
this any further. Three pelves show cutmarks around the acetabulum, indicative of disarticulation of the
femur. Corresponding cutmarks, placed on collum femoris and trochanter major, are found on six femora. As the pelvis-femur joint is hard to disarticulate, cutmarks are expected to occur as a result of disarticulation. Such cutmarks are found on virtually every species where this kind of disarticulation has been carried out.
Cutmarks deriving from disarticulation of the femur-tibia joint occur on three distal femora and on three proximal tibiae. The relatively small number of cutmarks, compared to the ones at
the femur-pelvis joint, can be explained by the femur-tibia joint being much easier to dismember. This was observed in the experimental butchering of the cat, where the dismembering of the
femur-tibia joint only caused one cutmark on the distal femur. No cutmark relating to this disarticulation was found on the proximal tibia (see chapter 7.2.).
Cutmarks relating to disarticulation of the tarsal joint have been found on one tibia. As some of the distal femora and proximal tibiae show signs of disarticulation, one would expect
more signs of disarticulation at the more complex tarsal joints. The reason for the lack of cutmarks is likely the absence of calcanei and astragali. In the experimental butchering (cf chapter 7.2.) cutmarks derivning from the disarticulation of the tarsal joint were not found on the tibia, but on the calcaneus and the astragalus. The cutmarks on the calcaneus were found anteriorly on tuber
calcanei, and the ones on the astragalus were found anteriorly on collum tali. Another possibility regarding the few cutmarks found at the tarsal joint, is that disarticulations may not have happened
very often. Maybe the tibia was easy enough to fillet without removing the paw Fist. A similar case may be found for the carpal joint, where few cutmarks occured (see above). An answer to these ideas may be found in the cooking methods, which will be discussed in chapter 8.2..
8.1.7.3. Filleting
Cutmarks deriving from filleting can be found on scapula, humerus, radius, ulna, pelvis, femur, tibia and fibula. The filleting cutmarks on the scapula are found on the distal part of the medial side. On the butchered cat (cf chapter 7.2.) and on scapulae from TrolleLassen's butchered otter,
such filleting cutmarks are found on various places on the medial side (Trolle-Lassen 1985:Appendix VII). On nine pelves, cutmarks that may derive from filleting are found. They are
in most cases placed dorsally-ventrally on ilium and ischium. Similar cutmarks are found on pelves of otter at Hjerk Nor (see chapter 8.1.6.), and at Kongemose and Praestelyng, where they
have been interpreted by NoeNygaard as deriving from butchering (NoeNygaard 1995:225). As

Noe-Nygaard claims butchering to be either primary butchering, i.e. dismemberment, or
secondary butchering, which "... ainzs at reducing the size of the dismembered pieces.", such as
removal of less meaty pacts of a bone element (NoeNygaard 1995:185). Secondary butchering is
mostly used for larger animals, and I find it dubious if there was a need to reduce the size of a cat
pelvis or otter pelvis. I find it therefore more likely that these cutmarks derives from filleting
processes. Of the long bones, 21 humeri, 8 radii, 6 ulnae, 12 femora, 7 nbiae and one fibula show
oblique or horisontal cutmarks on the shafts. On one radius, a longitudinal cutmark occur. The
cutmarks vary generally greatly in their position on the bones. The ones on the humeri are found
on all parts of the shaft; from foramen supracondylare to tuberculum mi~zus. The cutmarks on
radius are mostly found at midshaft, but on two radii they occur proximally. On the ulnae,
cutrnarks are often found below the joint. Some are also placed at midshaft and distally. Like the
humerus, the cutmarks on the femora are found all over the shaft. There is however, a slight
dominance for the proximal end of the shaft. On the tibiae, the cutmarks are placed on the
proximal half of the bone. This varation in the placing of filleting cutmarks are likely due to
muscular anatomy. The skeletal elements covered in most meat are humerus, femur and tibia On
the tibia, only the proximal half is to any greater extent covered by meal. This corresponds with
the position of cutmarks on these bones. On the other long bones, i.e. radius, ulna and fibula, the cutmarks are more varied in position.
8.1.7.4. Comparison with other sites and general interpretation
Of the comparative sites, wild cat was found a t~ongemose and Tybrind Vig. At Kongemose,
cutmarks were found on radius, femur and tibia. Judging by the representation of skeletal
elements, I would argue that the cutmarks derive from either disarticulation or filleting. Cutmarks
deriving from skinning could be placed on the distal radius and tibia, if the skin on the paws was
to be disregarded. It is, however, unlikely that cutmarks on the femur would reflect such
procedure. At Tybrind Vig, cutmarks show that both the meat and the fur of wild cat were
utilized. As mentioned earlier in this subchapter, the skinning method employed at Tybrind Vig
was similar to the one used at Hjerk Nor. Cutmarks not deriving from skinning are rather few,
and indicate disarticulation of the proximal humerus, as well as filleting of humerus, radius and
femur (TroUeLassen 1987:89, 102).
When comparing the evidence of the cutmarks on wild cat bones at Hjerk Nor and the
comparative sites, it is made clear that cat meat was eaten on all three sites. Evidence for skinning
is only found at Hjerk Nor and Tybrind Vig, but is assumed to also have occured at Kongemose.
The cutmarks are indicative of filleting and dismemberment. Few individual bones on each site
have a large amount of cutmarks. This may be related to the ease with which a cat can be filleted
(cf chapter 7..2 and 7.3.), but also to whether food was scarce or not. If the wild cats had all been
used as emergency food, there would likely have been more cutmarks, in order to remove the last

scrap, and a higher number of fragmented bones.
As it has not been possible to receive any detailed information regarding the epiphyseal fusion of wild cat, no thorough analysis on hunting and trapping of wild cat can be carried out. However, some deduction regarding this topic is possible. According to Tetley (Tetley 1941:16, in Trolle-Lassen 1985:81), wild cats almost reach their adult size during their first winter. Very few bones in the Hjerk Nor assemblage belong to very young individuals, i.e. with both proximal and distal epiphyses unfused. This might indicate that hunting took place during autumnlwinter. The wild cat can have two litters per year (Bj&vall& Ullstram 1995:194), and the few unfused bones at Hjerk Nor might reflect such late litters.
8.2. The overall view
8.2.1. Skinning methods
Concerning the use of wild canids, mustelids and felids at Hjerk Nor, it can be confirmed that all species were, with the possible exception of polecat, used for their fur. As no cutmarks were found on the polecat cranium, it cannot be claimed for certain that the polecat was skinned (see chapter 8.1.4.). Regarding skinning methods, fox, pine marten, otter and wild cat showed cutmarks on the snout/osfrontale and on the mandible, indicating that the skin was cut off, probably with a circling motion, in front of or above the eyes. This seems to have been the most common method for skinning smaller animals' heads. In contrast to nowadays, the snout is left on the carcass. Cutmarks did not occur on any rnetapodials in the assemblage, but as phalanges
are lacking, the treatment of the fur on the paws cannot be discerned. On some sites, such as Tybrind Vig, claws were part of the furs of otter and wild cat, but were left on the carcasses of
pine marten (TroUe-Lassen 1985235; Trolle-Lassen 1987:88-89). At Segebro, a Kongemose site in Sweden, it was argued that the exceptionally good condition of some metapodials and phalanges in the assemblage, was due to them being part of furs, and therefore better protected against taphonomic processes (Lepiksaar 1982:113). As the metapodials at Hjerk Nor are as well preserved as most other bones of wild canids, mustelids and felids at the site, this hypothesis is not of use for determining details of skinning methods at Hjerk Nor. For otter and wild cat, there
are indications of them being open skinned. This kind of skinning method (see figure 7.1.) is known to have been used at Tybrind Vig. It is possible that open skinning were used for all
mammals in Southern Scandinavia during the Mesolithic, but as few cutmark analyses have been carried out, and as indications of this method only are found on the pubic bone and on ischium on
the pelvis, little can be argued for or against it.

8.2.2. W i d canids, mustelids and felids a s food
On fox, badger, pine marten, otter, lynx and wild cat, the flesh was also utilized. As these animals
constitute a smaller amount of the other species at virtually every site - both in fragments and in meat weight - it is not likely that they were considered important meat animals during the
Mesolithic. However, the cutmarks prove that the meat was used. The wild canids, mustelids and felids seem to have been utilized roughly in the same way.
Traces of dismembering and filleting occur on every species, except polecat. Why were the bones
disarticulated so thoroughly? For larger animals the limbs need to be cut down into smaller bits, that can be handled, but for such small animals as otter and wild cat, this seems not likely to be
the reason. During my butchering experiment, I noticed that it was easier to fillet disarticulated limbs, as they were easier to handle. Another possiblity of the need for disarticulation is cooking. As the bones are not burnt, they were likely not cooked in a 'cooking pit' with direct contact with coal andlor fire. That may have occured at Hazendonk, Swifterbant and Skateholm, where burning traces have been found on bones of otter and wild cat (Zeiler 1987:254; Jonsson 1988:65). In Northern Sweden, there is a tradition in cooking meat wrapped in birch-bark in a 'cooking pit' (Andersson et al 1983). A similar tight-fitting wrapping would have made it impossible for any coal or soot to get in contact with the meat and burn the bone. The lack of burning on the bones can also be explained by the fdleting cutmarks. If all meat was removed, and the bone thrown away, there would not be any burn marks. The meat could have been cooked in a 'cooking pit', or in a ceramic vessel. Ertebglle vessels are found in all sizes (Bergenstr3e pers. comm.), and filleting is, however, not necessary in order to fit the limbs into the pot. Experiments with Ertebglle vessels have shown that reaching high temperatures was possible. Some researchers, though, believe that the vessels were used for fermenting, rather than cooking (Isaksson 1996:3-5).
8.2.3. Emergency meat or not?
The eating of carnivores at Hjerk Nor is beyond doubt. The question remains, though, whether it was to be regarded as emergency food or not. Emergency food is often connected to winter. The sex ratio of otters indicated habitation of the site during spring and summer (see chapter 8.1.5.),
and if this sex ratio is correct, we may want to look for other reasonings. However, fur animals are usually hunted for their fur in autumn/winter, when it is at its finest. The large amount of wild
cat bones seem to indicate that either the sex ratio of otter is not representative, or that Hjerk Nor was inhabited during different seasons of the year.
Usually, an indicator of bad years is phalanges broken for marrow. As I have not studied the other species at the site, I do not know the fragmentation frequency for the larger mammals. As the 'excavation' of Hjerk Nor was not very thorough, I assume that any phalanges of larger

mammals are likely to be missing. As there generally are few cutmarks on each bone in the assemblage and as very few bones are fractured, it would seem likely that the inhabitants at Hjerk Nor were not very desperate for meat and marrow. If bad seasons occured, they were probably not very intense. Maybe the carnivores were used as food when the hunting occasionally failed,
and by luck a cat had been trapped the same day. This idea might explain the bones not being heavily exploited for meat and marrow, as the inhabitants knew that they would likely eat deer the
next day.
8.2.4. Other uses
Animals were of more use than food and fur. As mentioned in chapter 8.1.2., badger was likely
killed for its fat Besides fat, sinew, and other practical uses, animals may have been used for ritual and medicinal purposes. Ritual deposits are known from later periods, but are likely to have
occured during the Mesolithic. The disarticulation of the mandible on wild cat and fox, may be an indication of the use of the skull, maybe for magical purposes. Another 'unusual' use of a wild
animal is the tame fox, that may have been used as a 'fur farm', as a decoy or as a pet None of the bones of wild canids, mustelids and felids from Hjerk Nor show signs of
working. At Ertebolle, three ulnae of wild cats were worked into awls (Winge 1904, in DegerbBl 1933:405). It is not known whether the use of wild cat bones for tools was a common
phenomenon during the Stone Age, as such finds are categorized as artefacts, and not included in the osteological analyses. While they may be discussed in archaeological analyses, they are
usually not determined to species, and therefore they often slip through our examining eyes and disappear into the storage - hopefully not forever, though.
9. CONCLUDING WORDS
The use of wild canids, mustelids and felids for food and fur was widespread in Southern Scandinavia during the Mesolithic. They were likely only limitedly utilized for their flesh, as red
deer, wild boar, aurochs and roe deer gave much more meat. To neglect this aspect of the wild canids, mustelids and felids is in my opinion to do oneself a disservice. It is easier to dichotomize,
but trying to discern a complex world can be far more rewarding, both to ourselves and to our research.

Aaris-S~rensen, K. 1976. A zoological investigation of the bone material from Svaerdborg I - 1943.
A r h o l o g i s k studier, VoL 3, pp. 137-148. - 1988. Dannlarkr forhistorisk dyreverden. Fra istid ti1 vikingatid. Gyldendals bogklub; Copenhagen.
ISBN: 87-00-24356-6. Aaris-S~rensen, K. & Nord Andreasen, T. 1993. Small mammals from Danish Mesolithic sites. Journalof
Danish Archaeology, Vol. 11, pp. 30-38. Ahnlund, H. 1976. Age determination in the European badger, Meles nzeles L. Zeitschrifrfiir Saugeiierkunde,
Vol. 41, pp. 119-125. Andersen, S. H. 1979. Aggersund. En ertebelleboplads ved Limfjorden. Kunzl, 1978. pp. 7-56.
- 1981. Stenalderen I. Jegerstenalderen Danmarkshistorien. Sesam; Copenhagen. ISBN: 87-7324-422-8. - 1985. 'Qbrind Vig. A preliminary report on a submerged Ertebglle settlement on the west coast of
Fyn. Journal of Danbh Archaeology, Vol. 4, pp. 52-69. - 1993. Bjernsholm. A stratified kgkkenmeddiig on the Central Limfjord, North Jutland. burnal of
Danish Archaeology, Vol. 10, 1991, pp. 59-96. - 1995. Coastal adaptation and marine exploitation in Late Mesolithic Denmark - with special
emphasis on the Limfjord region. in Fischer, A. (ed). 1995. Man and sea in the Mesolithic. Oxbow books; Oxford. ISBN: 0-946897-96-4. pp. 41-66.
- 1998. Ringkloster. Ertebelle trappers and wild boar hunters. A survey. Joional of Danish Archaeology, Vol. 12, 1994-1995. pp. 13-59.
Andersen, S. H. & Johansen, E. 1987. Erteb0lle revisited. Journal of Danish Archaeology, Vol. 5, 1986. pp. 31-61.
Andersson, M., Forsberg, L., Frykholrn, G. & Larsson, L. 1983. Nigra gamla mauagningstekniker. Milnniska - Natur - T e h l o g i , Nr. 2 1983. http://www.forntidateknik.z.s~/MNT~l983/maUag.btm
(2000-07-31). Andersson, A. & Paulsson, T. 1993. Jligarens skinn och hudar. Bokf5rlaget Settern; Orkelljunga. ISBN: 91-7586-410-X. Binford, L. R. 1981. Bones. Ancient nten and modern nzyths. Academic Press; New York. London. Toronto,
Sydney, San Fransisco. ISBN: 0-12-100035-4. Bratlund, B. 1993. The bone remains of mammals and buds from the Bjemsholm shell-mound. A preliminary
report. Journal ofDanish Archaeology, Vol. 10, 1991, pp. 97-104. Bmun, B. & Singer, A. 1975. Third revised printing. Alln Europas fdghr i fdrg. Albert Bonniers forlag;
Stockholm. ISBN: 91-0-040558-2. Burenhult, G. 1999. Arkeologi i Norden I . Natur och kuhr; Stockholm. ISBN. 91-27-07345-9. Clason, A. T. & Brinkhuizen, D. C. 1978. Swifterbant, mammals, birds, fishes. A preliminary report. Heliniunz. Revue consacrie b l'archiolagie des Pays-Bas, de la Belgique ef du Grand-duchf de Luxembourg. VoL
28, pp. 69-82. DegerbBI, M. 1933. Danmarlcs pattedyr i fortiden i sammenligning med recente former. Videnskobelige meddelelserfm Damk nufurhistorisk forening i Kgbenhm, Band 96, pp. 357-641.
- 1942. Et knoglemateriale fra Dyrholmbopladsen, en aeldre kekkenmeddiig. Med sredigt henblik paa uroksens kens-diiorphisme og paa kannibalisme i Danmark. in Mathiassen, T., Degerbel, M. & Troels-Smith.
J. 1942. Dyrholmen. En stenaldersbopladr p~ Djursland. Kgbenhavn. pp. 77-136. De Marinis, A. M. 1995. Craniometric variability of polecat Mustela putoris L. 1758 from North-Cenual
Italy. in Prigioni, C. (ed). 1995. Proceedings I1 Italian symposium on carnivores. Biology and conservation of mustelids. Hysnix, Vol. 7. pp. 57-68.

Desse, J. 1975. Vestiges temoignant d'une activitc? de pelleterie sur le chantier neolithique recent d'Auvernier
Brise-Lames. BuIL Soc. neuchhteL Sci. not., VoL 98, pp. 203-208.
van den Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum Bullerin 1. ISBN: 0-87365950-3. von den Driesch, A. & Boessneck, J. 1975. Schnittspuren an neolitischen Tierknochen. Ein Beiuag zur Schlachtiierzerlegung in vorgeschichtlicher Zeit Germania, Vol. 53, pp 1-23.
Eidlitz, K. 1969. Food and emergency food in the circumpolar area Studia Ethnographica Upsaliensia, VoL
32. Ekman, S. 1983. Norrlands jakt ochfske. TvB fbrlZggare bokfiirlag; Ume& ISBN: 91-85920-10-X.
Enghoff, I. B. 1993. Mesolithic eel-fishing at Bj0rnsholm. Denmark, spiced with exotic species. Journal of Danish Archaeology, Vol. 10, 1991, pp. 105-118.
- 1995. Fishing in Denmark during the Mesolithic period. in Fischer, A. (ed). 1995. Man ond sea in the Mesolithic. Oxbow books; Oxford. ISBN: 0-946897-964 pp. 67-74.
Englund, J. 1970. Some aspects of reproduction and mortality rates in Swedish foxes (Vuulpes vulpes), 1961- 63 and 1966-69. Vilnevy. Swedish wildlife, Vol. 8, No. 1. pp. 1-82. Ericson, P. G. P., Iregren, E. & Vretemark M. 1988. Animal exploitation at Birka - a preliminary report Fornviinnen, Vol. 83, pp. 80-88.
Habermehl, K-H. 1985. Altersbestinzn~ung bei Wild- und Pelztiere. 2nd edition. Paul Parey Verlag; Hamburg, Berlin. ISBN: 3-490-06812-2.
Hahr, TH. 1882. Handbokfdr jiigore och jakivdnner. Andra bandet P.A. Nordstedt & stlner; Stockholm. Hatting, T. 1972. JagMdt in Rud, M. (ed). 1972. Ar!uzologiskABC. Hdndbog i danskforhiswrie. Politikens
forlag; Copenhagen. ISBN: 87-567-1635-5. pp. 154-155. Hatting, T., Holm, E. & Rosenlund, K. 1973. En pelsjaegerhoplads fra stenalderen. Kaskelot 1973, pp 13-21. Heinrich, D. 1991. Untersuchungen an Skeletuesten wildlebender Sagetiere aus dem mittelalterlichen
Schleswig. Ausgrabung Schiid 1971-1975. Ausgrabungen in Schleswig. Berichte und Studien 9. Karl Wachholtz Verlag; Neumiinster. ISBN: 3-529-0145.
Helm, J. & Sturtevant, W. C. (eds). 1981. Handbook of North American Indians. Vol. 6. Subarctic. Smithsonian Institution; Washington.
Honigmann, H. H. 1981. West Main Cree. in Helm, 1. & Sturtevant, W. C. (eds). 1981. HandbookofNorth American Indians. Vol. 6. Subarctic. Smithsonian Institution: Washington. pp. 217-230.
Isaksson, K. 1996. En smmesolitisk kastrull? Unpublished Bachelor essay in Prehistoric archaeology. Institute of Archaeology, Lund University. Lund. Jensen, A. 1972. Odder. in Hvass, H. (ed.). 1972. Danmarks dyreverden. Vol. 10. Rosenkide & Bagger;
Copenhagen. pp. 51-70. - 1984. Pelsdyr og pels. Narur og nluseunr 23 (4).
Jessen, K. 1924. De geologiske forhold ved de to bopladser i Holmegaards mose. in Broholm. H. C. 1924. Nye fund fra den aeldste stenalder. Holmegaard- og Svaerdborgfundene. Aarb(ger fra Nordisk oldlqndlighed og historie 1924. pp. 14-27. Johansson, F. & Hiister, H. 1987. Untersuchungen an Skelettresten von Katzen aus Haithahu (Ausgrabung
1966-1969). Bericha iiber die Ausgrabungen in Haithabu. Bericht 24. Karl Wachholtz Verlag; Neumiinster. ISBN: 3-529-01924-0.
Jonsson, L. 1988. The vertebrate faunal remains from the Late Atlantic settlement Skateholm in Scania. South Sweden. in Larsson, L. (ed). 1988. The Skateholm project I. Man and environment. Lund. Kratochvil, Z . 1976. Sex dimorphism of the domestic cat (Felis lybica f. catus L) on the skull and on the mandible. Acta veterinaria Bmo, VoL 45, pp. 159-167.

Larsson, L. 1987. Some aspects of cultural relationship and ecological conditions during the Late Mesolithic and Early Neolithic. in Burenhult, G., Carlsson, A.. Hyenstmnd, A. & Sjgvold, T. (eds). 1987. Theoretical
approaches to artefacts, senlenzent and society. BAR International Series 366 (i). Oxford. ISBN: 0-86054-475-3. Lepiksaar, J. 1982. Djurrester f r h den tidigatlantiska boplatsen vid Segebro ngra Malmo i Skine (Sydsverige). in Larsson, L. 1982. Segebro. En tidigatlantisk boplars via' Sege b nzynning. Malmilfynd 4. Malmil mus&; Malm6. pp. 105-128. L idb , K. 1995. Prehistoric diet hansltions. Theses and papers in scientific archaeology 1. Archaeological
research laboratory, Stockholm university; Stockholm. ISBN: 91-628-1679-9. Liljegren, R. & Lager&, P. 1993. Frdn mammuts~pp till kohage. Djurens historia iSverige. Lund. Magnell, 0. 1996. Mesolitisk slakt. En analys av slokupdr pd det osteologiska materialet av kronhjort, dlg och
rddjur f i n den sydrkandmaviska och senmesolitiska inlondsboplafsen Bbkeberg Ill. Unpublished Master essay
in Prehistoric archaeology. Institute of Archaeology, Lund University. Lund. Mathiasson, S. & Dalhov, G. 1987. V&a vilda djur. Bokfilrlaget Prisma; Stockholm. ISBN: 915182009-9. Mghl, U. 1979. Aggersund-bopladsen zoologisk belyst Svanejagt som &sag ti1 bosienelse? Kuml 1978. pp. 57-73. Neal, E. & Cheeseman C. 1996. Badgers. Poyser Natural History; London. ISBN: 0-85661-082-8. NoeNygasrd, N. 1969. Prmstelyng. En mesolithisk boplads i Amosen. En faunistisk unders@gelse og en
@kologisk rekonstruh?ion. Unpublished prize dissertation. University of Copenhagen. - 1988. ST-values of dog bones reveal the nature of changes in man's food resources at the
Mesolithic-Neolithic transition, Denmark. Chenzichal geology (Isoiope geoscience section), Vol. 73, pp. 87-96. - 1995. Ecological, sedimentary, and geochemical evolution of the late-glacial to postglacial h o s e
lacustrine basin, Denmark. Fossils and sham, NO. 37. Scandinavian University Press; Oslo, Copenhagen, Stockholm. ISBN. 82-00-37656-7. Noe-Nygaard, N. & Richter, J. 2090. A late Mesolithic hunting station at Agernms, Fyn, Denmark
Differentialion and specialisation in the late Ertebmlle culture - heralding infroduction of agriculture? Abstract from The 6th International Conference on the Mesolithic in Europe. 4-8 Sept 2000 in Stockholm, Sweden. hIIp://www.arkeologi.uu.se/meso200O/no~ (2000-08-17). Ondrias, J. 1960. Secondary sexual variation and body skeletal propo~tions in European Mustelidae. Arkvfdr
zoologi, Vol. 12, pp. 577-583. Reichstein, H. 1991. Die wildlebenden Saugetiere von Haithabu (Ausgrabungen 1966-1969 und 1979-1980). Berichre iiber die Ausgrabungen in Haithabu. Bericht 30. Karl Wachholtz Verlag; Neumiinster. ISBN: 3-529-01930-5. Rogers, E. S. & Smith, J. G. E. 1981. Environment and culture in the Shield and Mackenzie borderlands. in Helm, J. & Snulevant, W. C. (eds). 1981. HandbookofNorth American Indians. Vol 6. Subarctic. Smithsonian Institution; Washington. pp. 130-145. Rowley-Conwy, P. 1998. Mear, furs and skins. Mesolithic animal bones from Ringkloster, a seasonal hunting camp in Jutland. Journal ofDanish Archaeology, Vol. 12,1994-95. pp. 87-98. Schauser, K. H. & Dantzer, V. 1995. Konlpendium i osteologi. JordbmgsforlageS Fredriksberg, Copenhagen. ISBN: 87-7432-431-4. Schwartz, J. H. 1995. Skeleton keys. An inwoduction to human skelernl morphology, developntenr. and analysis.
Oxford University Press; New Y or& Oxford. ISBN: 0-19-505638-8. Strid, L. 1999. Pdlsilsdjursjokr i Sydskandinavien under mesolitihurn. Unpublished Bachelor essay in Prehistoric archaeology. Institute of Archaeology, Lund University. Lund. Stubbe, M. & Krapp, F. 1993a. Handbuch der Stlugetiere Europas. Band 5. RaubsZuger - Carnivora. Teil 1.
Aula Verlag GmbH; Wiesbaden. ISBN: 3-89104-029-6.

- 1993b. Handbuch der Sliugetiere Europas. Band 5. Raubs2uger - Carnivora Teil2. Aula Verlag GmbH;
Wiesbaden. ISBN: 3-89104-528-X. Tetley, H. 1941. On the Scottish wild cat Proceedings j?om the Zoological Society, Vol. 111B. pp. 13-23. Trolle-Lassen, T. 1985. En zooarkreologisk analyse af erteb@llebopladsen Tybrind Vig, prinuert baseret pd knogler af pelsdyr og bonhjort. Unpublished Master essay. Department of Prehistoric Archaeology, Aarhus University. Aarhus.
- 1986. Human exploitation of the pine marten (Martes martes (L.)) at the late Mesolithic settlement of Tybring Vig in Western Funen. Striae, Vol. 24, pp. 119-124.
- 1987. Human exploitation of fur animals in Mesolithic Denmark - a case study. Archaeozoologia, Vol.
1 (2), pp. 85-102. - 1990. Butchering of red deer (Cervus elaphus L.) - a case study from the late Mesolithic settlement of
Tybrind Vig. Journal of Danish Archaeology, Vol. 9, pp. 7-37. Zeiler, J. T. 1987. Exploitation of fur animals in neolithic Swifterbant and Hazendonk (Central and Western Netherlands). Paheohistoria. Actn et contmunicationes instituti bio-archoeologici universitntis Groninganae 29. Rotterdam. Brookfield. pp. 245-263.
- 1988. Age determination based on epiphyseal fusion in post-cranial bones and tooth wear in otters (Lutra lutra). Journal of Archaeological science, VoL 15, pp. 555-561.
Wesenberg-Lnnd, E. 1927. Rovdyrene. in Brehm (ed.). 1927. Dyrenes liv, III bind. 2. afsnit White, T. D. 1992. Prehistoric cannibalism at Mancos SMTUMR-2346. Princeton University Press; Princeton. ISBN: 0-691-09467-5. Winge, H. 1904. Om jordfundne partedyr fra Danmark Videnskabelige nteddeleser fra Dansk naturhistorisk forming, VoL 56. pp. 193-304.
PERSONAL COMMENTS
Ingrid Bergenstrihle. Ph.D. student in Prehistoric archaeology. Institute of Archaeology, Lund University. Sweden. Bodil Bratlund. Research assistant at the Archaeo-Osteoiogical Research Laboratory. Institute of Archaeology, Stockholm University. Sweden. Elisabeth Iregren. Ph. D. Lecturer in Historical osteoiogy. Institute of Archaeology, Lund University.
Sweden. Rita Larje. Research assistant Swedish Museum of Natural History. Stockholm, Sweden. Ola Magnell. Ph.D. student in Historical osteology. Institute of Archaeology, Lund University. Sweden. John Simonsen. Archaeologist Skive Museum. Skive, Denmark. Letter 7 Aug. 2000.


APPENDIX 1. - Map of Southern Scandinavia, with all mentioned sites marked.
Aasted Agernres Aggersund Bj0rnsholm Bokeberg IlI Dyrhohen Ertebfille Hjerk Nor Holmegaard Krabbesholm Kongemose
Lollikhuse Meilgaard Muldbjerg I Przestelyng Ringkloster Segebro Skateholm Smakkerup Huse Svaerdborg I Tybrind Vig Virksted
APPENDIX 2. - Osteological and anatomical terminology.
Directions
Dorsal Dorsal A
Oral , . Aboral Cranial + Caudal I I
Basal Ventral
Proximal
Lateral Medial .
Sinister = left
Dexter =right
Anterior Posterior
+ Distal
Pictures from Schwartz (1995:4-6)

APPENDIX 2. - Osteological and anatomical terminology.
Skeletal elements
Costae
Metapodials = Metacarpals and metatarsals
0 s bacculum = penis bone

APPENDIX 2. - Osteological and anatomical terminology.
Bone anatomy
Zygamatic arch 0s fmntaie
Sulcus intatubmularis Tuberculum minus ~bm~umm~[ r -Caputhumeri
Crista hlbaruli minoris
Diaphysis = shaft af bone Epiphysis = end of bonc Periosteum = Ulin membrane surrounding the bonc

APPENDIX 3. - Measurements

APPENDIX 3. - Measurements

APPENDIX 3. - Measurements
i POSTCRANIAL SKELETON 1 i i ..: ......................... .
Sprier i ~ l ~ n x n t j side i ldmtifratbn nulnbcr i Q i GLC 80 1 i ~d i BPC GLP 8 F.X. ..?!?.Eil!*-..i ..... H.Y.ENI ..... i ..... ?..k .... ...: .......... C ": .......... -..I ....... :: ........ : - ........
i HYIMN. i dx i 9 - i . i 21.44 i - _ i . ! 22.70 i - .......................... .i .... .!!!!.Er.Y.? ..... j ......... ; ; ............. : .................... T .......
i Hvmervs i dx i 11 . i . i 20.26 i - ..... ...... ...-... ...... ........ ......... ........ ........................... 1 ..... ..... dx ......................... ..........I : I : ~....zo~B! I. L .-
....................... " ... i Tibi. i *in i i Tihh i sin i
I i Tibin i sin i 13 i 146.21 i - 1 24.74 i i 17.91 i - I - I

APPENDIX 3. - Measurements

APPENDIX 4. - Cutmarks on individual bones.
VULPESVULPES
Tibia c Anterior, and lateral view
MELES MELES
Tibia c Anterior view
Mandibula a Lateral, and oral view
Mandibula b Lateral, and oral view

APPENDIX 4. - Cutmarks on individual bones.
MARTES MARTES
Cranium a Dorsal, and lateral view
- - - - - - - - -*r - -.- - . - - # \ 4 . . .. .
\ s ___----. V _ - - _ I \ I
, I '__ _ ,'! , - ..- , Cranium c ------;+; 4? s : .- - -.=---, Lateral view
Humerus 5 Anterior view
Mandibula b Lateral, and basal view
Tibia 1 Posterior view

APPENDIX 4. - Cutmarks on individual bones.
LUTRA LUTRA
Cranium b Dorsal, and lateral view
Cranium e Dorsal, and lateral view
Mandibula a Lateral view

APPENDIX 4. - Cutmarks on individual bones.
LUTRA LUTRA
Humerus 1 Anterior, and lateral view
Humerus a Anterior, and lateral view
Humerus 2 Anterior, and lateral view
Humerus b Lateral view

APPENDIX 4. - Cutmarks on individual bones.
LUTRA LUTRA
Lateral view II uhab
Pelvis b Lateral view
Pelvis a Lateral view
Pelvis c Lateral view
Pelvis e Lateral view
Pelvis f Lateral view

APPENDIX 4. - Cutrnarks on individual bones.
LUTRA LUTRA
Femur 4 Medial view
Femur 5 Anterior view
Tibia 1 Medial view
Fibula a Lateral view
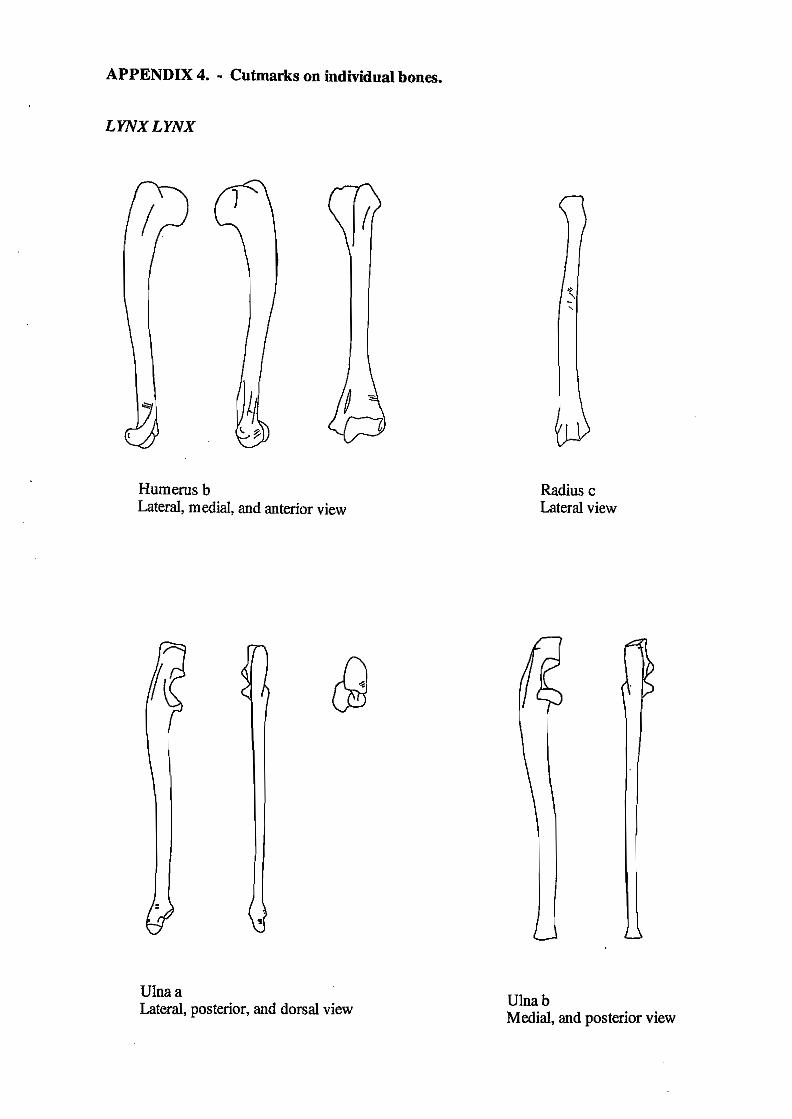
APPENDIX 4. - Cutmarks on individual bones.
LYNX LYNX
Humerus b Lateral, medial, and anterior view
Ulna a Lateral, posterior, and dorsal view
Radius c Lateral view
Ulna b Medial, and posterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Cranium c Dorsal view
Cranium 4 Lateral view
Cranium 2 Lateral view
Mandibula 1 Lateral view
Mandibula 2 Lateral view
Scapula e Lateral, and medial view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRIS
Humerus 1 Medial view
Humerus 2 Lateral, medial, and posterior view
Humerus 3 Posterior, anterior, medial, and lateral view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Humerus 5 Anterior, medial, and lateral view
Humerus 7 Posterior, and medial view
Humerus 6 Anterior view
Humerus 8 Lateral, medial, anterior, and posterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Humerus 9 Posterior, anterior, and medial view
Humerus 11 Anterior, and medial view
Humerus 12 Medial view
Humerus 10 Medial view
Humerus 13 Medial, and lateral view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRIS
Humerus 14 Medial, and anterior view
Humerus 16 Medial view
Humerus 15 Lateral, medial, anterior, and posterior view
Humerus 17a Medial, anterior, and posterior view

APPENDIX 4. - Cutmarks on individual bones.
FELIS SILVESTRIS
Humerus 17b Anterior, and medial view
Humerus 18 Medial view
Humerus 19 Medial view
Humerus 21a Humerus 21b Medial, and posterior view
Humerus 22 Medial view Medial, anterior, and posterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Humerus 24 Anterior, and lateral view
Humerus 25 Medial, anterior, and posterior view
Humerus 30 Humerus 31 Humerus 32 Medial, and posterior view Medial, and posterior view Medial, and anterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTMS
Humerus 33 Lateral, medial, and anterior view
Radius 1 Anterior, and medial view
Radius 5 Lateral view
Humerus a Medial, and anterior view
Radius 7 Lateral view

APPENDIX 4. - cutmarks on individual bones.
FELZS SZL VESTRlS
Radius 8 Anterior, lateral, and medial view Radius 12
Lateral view
Radius 13 Medial, and anterior view Radius 16
Lateral view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Radius b Anterior, lateral, and medial view
Radius c Medial view
Ulna a Anterior, and lateral view
Ulna b Anterior view
Ulna d Anterior

APPENDIX 4. - Cutmarks on individual bones.
FELIS SILVESTRIS
Ulna e Lateral, anterior, and medial view
Ulna g Anterior, and lateral view
Ulna h Ulna k Ulna 1 Anterior view Anterior, posterior, and lateral view Anterior, and posterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Ulna o Ulna p Lateral view Lateral view
Ulna v Ulna z Posterior, and medial view Posterior view
Ulna r Lateral, anterior, and medial view
Pelvis a Lateral view
--;\: . ---... -
. -. - ,' Pelvis c Lateral view

APPENDIX 4. - Cutrnarks on individual bones.
FELZS SZLVESTRZS
Pelvis d Lateral view
Pelvis f Lateral view
Pelvis e Lateral view
Pelvis g Lateral view
. , , . \ . . , ', '--- '. .. ' . r - . - . I
Pelvis i . --
Lateral view Pelvis j Lateral view
. I . <
Pelvis k Lateral view
Pelvis l Lateral, and ventral view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Femur 1 Femur 2 Anterior view Medial, posterior, and anterior view
Femur 7 Anterior view
Femur 8 Anterior view
Femur 4 Medial view
Femur 10 Lateral, posterior, and anterior view

APPENDIX 4. - Cutmarks on individual bones.
FELIS SILVESTRIS
Femur 1 1 Medial, and posterior view
Femur 13 Posterior, and lateral view
Femur 12 Anterior, posterior, medial, and lateral view
Femur 14 Anterior, posterior, lateral, and dorsal view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Femur 16 Anterior, posterior, medial, and lateral view
Femur 18 Anterior, medial, and lateral view
Femur 17 Lateral view
Femur a Medial, and anterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Femur b Anterior view
Tibia 1 Anterior view
Femur h Lateral, and posterior view Femur m
Anterior view
Tibia 3 Medial, posterior, and anterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRIS
Tibia 5 Medial, posterior, and anterior view
Tibia 10 Medial, and anterior view
Tibia 8 Medial, and posterior view
Tibia 12 Medial, posterior, and anterior view

APPENDIX 4. - Cutmarks on individual bones.
FELZS SZLVESTRZS
Tibia 15 Tibia 18 Medial view Anterior view
Tibia 23 Anterior view
Tibia 24 Medial view
Tibia 19 Lateral, medial, and anterior view
Tibia i Fibula b Anterior, lateral, and medial view Lateral view




















